3 Adaptations to Terrestrial Environments
3Adaptations to Terrestrial Environments Adaptations of the dromedary camel. Dromedary camels, such as this one in the Hajar Mountains of the United Arab Emirates, have a wide range of adaptations that allow them to live in hot, dry environments. The Evolution of Camels When you think of camels, you might envision the iconic animals of the African and Asian deserts. The ancestor of all camel species actually originated in North America about 30 million years ago and camels roamed many parts of North America until about 8,000 years ago. Current evidence suggests that some of these ancestors crossed the Bering land bridge from Alaska about 3 million years ago and made their way into Asia and Africa. These individuals evolved into two modern-day species of camels: the endangered Bactrian camel (Camelus bactrianus) and the much more common dromedary camel

(C. dromedarius). Other ancestors moved from North America down to South America and evolved into a second group: guanacos (Lama guanicoe), llamas (L. glama), vicuñas (Vicugna vicugna), and alpacas (V. pacos). All these animals live in dry environments and have evolved a number of adaptations that help them cope with the harsh conditions. For example, the dromedary camel—the most-studied of the group— risks overheating in the very hot deserts where it lives. During the day, the rays of the Sun strike its body and warm its surface. The camel can respond behaviorally by facing into the Sun and presenting a smaller profile for the Sun’s rays to warm. The sandy desert soil is also hot and radiates the heat it absorbs, which makes the camel even hotter. Fortunately, the camel has a very large body relative to its surface area, so these heat inputs raise its body temperature slowly. Although camels, like all mammals, try to maintain a constant body temperature, they can tolerate a rise in body temperature of 6 °C before experiencing any harmful effects. In contrast, most mammals can only tolerate an increase of about 3 °C. At night, as the air and sand cool rapidly, the camel radiates its excess heat into the air or lies down and transfers its excess body heat to the sand. One of the many amazing adaptations of camels and other mammals in hot environments is their ability to cool their brains. Whereas much of the body can tolerate short-term increases in temperature, the brain cannot. The camel has evolved an arrangement of veins and arteries that helps solve this problem. As the animal breathes, veins positioned beside the long nasal cavity are cooled by means of evaporating water vapor. The veins that carry cool blood then travel toward the back of the camel’s head, where they come into close contact with the arteries that supply blood to the brain. Although the blood does not mix between the veins and arteries, heat is exchanged between the two types of vessels. This cools the arterial blood before it travels to the brain and keeps the brain several degrees cooler than the rest of the body. Lack of water is also a challenge in the desert environment. As an adaptation, the dromedary camel can store large amounts of water in its body, most of which resides in its tissues. As water is lost from the
bloodstream, the water from the tissues enters the blood. As much as 30 to 40 percent of the camel’s body mass is water that can be used over several days when it is unable to drink water. In other mammals, a mere 15 percent loss of water can be lethal. Camels also conserve the water that they take in by producing relatively dry feces and urine that is high in waste products and low in water. In addition, while sweating can be an effective way to cool the body through evaporation, camels that are low on water can reduce the amount that they sweat. Collectively, these adaptations allow the camels to survive when water is scarce. “One of the many amazing adaptations of camels and other mammals in hot environments is their ability to cool their brains.” Camels represent just one case in which terrestrial organisms have evolved numerous adaptations to the challenges posed by terrestrial environments. In this chapter, we will explore the challenges of living on land and the adaptations of terrestrial plants and animals that make their lives possible. Guanacos of the Patagonia. Guanacos, from the Patagonian region of Chile, share a common ancestor with Asian camels. As a result of their common ancestry, guanacos and Asian camels also share a number of adaptations for coping with a dry environment.
Cain, J. W., et al. 2006. Mechanisms of thermoregulation and water balance in desert ungulates, Wildlife Society Bulletin 34:570–581. Ouajd, S., and B. Kamel, 2009. Physiological particularities of dromedary (Camelus dromedarius) and experimental implications, Scandinavian Journal of Laboratory Animal Science 36:19–29.
Learning Objectives
After reading this chapter, you should be able to:
3.1 Explain how most terrestrial plants obtain nutrients and water from the soil.
3.2 Illustrate how sunlight provides the energy for photosynthesis.
3.3 Describe the ways in which terrestrial environments pose a challenge for animals to balance water, salt, and nitrogen.
3.4 Understand how adaptations to different temperatures allow terrestrial life to exist around the planet. As we discussed in Chapter 2, life on Earth probably arose in the water. Following these early origins, life forms evolved adaptations that allowed them to live on land. The transition from water to land posed a number of new challenges. Plants evolved to obtain water and nutrients from the soil, and to conduct photosynthesis under hot and dry conditions. Animals evolved to balance water, salts, and wastes, and to adjust to extreme temperatures in terrestrial environments. In this chapter, we will examine the diversity of adaptations that allow plants and animals to live in terrestrial environments.
#### 3.1 Most Terrestrial Plants Obtain Nutrients and Water from the Soil
3.1 Most terrestrial plants obtain nutrients and water from the soil A few unusual plants, such as the epiphytes discussed in Chapter 1 (Figure 1.9), obtain necessary water and nutrients without being rooted in the soil. However, the vast majority of plants obtain nutrients and water from the soil through their root systems. As a result, plants have a number of adaptations that help them perform this task.
Soil Nutrients
In addition to the oxygen, carbon, and hydrogen that plants incorporate into carbohydrates to fuel their survival and growth, plants require many other inorganic nutrients including nitrogen, phosphorus, calcium, and potassium to make proteins, nucleic acids, and other essential organic compounds. Whereas oxygen and carbon are available in the air, other nutrients are obtained as ions dissolved in the water held by the soil around plant roots. Nitrogen exists in soil as ammonium (NH4 +) and nitrate (NO3 –) ions, phosphorus exists as phosphate ions (PO4 3–), and calcium and potassium exist as the elemental ions Ca2+ and K+, respectively. The availability of these and other inorganic nutrients varies with their chemical form in the soil, and with temperature, pH, and the presence of other ions. A scarcity of inorganic nutrients, such as nitrogen, often limits plant production in terrestrial environments. We shall have much more to say about nutrient uptake by plants in later chapters.
Soil Structure and Water-Holding Capacity
To understand how plants obtain water and nutrients, it is first necessary to understand how water behaves in the soil. The movement of water in the soil can be described in terms of its water potential, which is a measure of the water’s potential energy. Water potential affects the movement of water in the soil from one location to another and depends on several factors that include gravity, pressure, osmotic potential (discussed in Chapter 2), and matric potential, so named because the collection of soil particles is known as the soil matrix. The matric (or matrix potential) is the potential energy generated by the attractive forces between water molecules and soil particles.
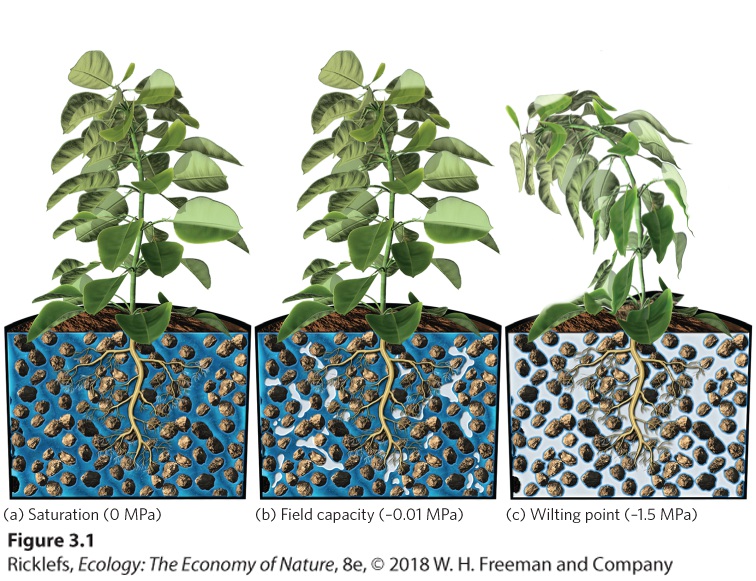
It exists because water molecules, which have electrical charges, are attracted to the surfaces of soil particles, which also have electrical charges. This attraction explains why soil is able to retain water against the downward pull of gravity. Water potential A measure of water’s potential energy. Matric potential The potential energy generated by the attractive forces between water molecules and soil particles. Also known as Matrix potential. Because electrical charges are responsible for the attraction between water molecules and soil particles, those water molecules closest to the surfaces of soil particles adhere the most strongly. When water is plentiful, most of the water molecules are not close to the surfaces of soil particles. As a result, these water molecules are not held tightly and plant roots can easily take up the water. As more of the water is used up, however, the water molecules that remain are positioned close to the soil particles and adhere tightly. As we learned when discussing osmotic pressure in Chapter 2, scientists quantify water potential in units of pressure, called megapascals (MPa). In a soil that is completely saturated with water, as illustrated in Figure 3.1a, the matric potential is 0 MPa. When a saturated soil drains under the force of gravity, the resulting matric potential is about –0.01 MPa. At this point, the force of gravity on the water molecules is equally opposed by the attractive force of soil particles on the water molecules. The maximum amount of water held by soil particles against the force of gravity is called the field capacity of the soil. The field capacity, illustrated in Figure 3.1b, represents the maximum amount of water available to plants.
Figure 3.1 Soil water. (a) Immediately after a rain event, soils can become saturated with water and all spaces between soil particles are filled. (b) The field capacity of soil represents the amount of water remaining after it has been drained by gravity. (c) The wilting point occurs when the opposing attractive forces of the soil particles prevent plants from extracting any more water. Field capacity The maximum amount of water held by soil particles against the force of gravity. As water becomes less abundant, such as when plants take up some of the water from the soil, the matric potential values become more negative. Water always moves from areas of higher potential (less negative values) to areas of lower potential (more negative values). So, for plants to extract water from the soil, they must produce a water potential that is lower than that of the soil. As soils dry out, they hold the remaining water ever more tightly because a greater proportion of that water lies close to the surfaces of soil particles. Most crop plants can extract water from soils with water potentials down to about –1.5 MPa. At lower soil water potentials, these plants wilt, even though some water still remains in the soil, as shown in Figure 3.1c. Scientists refer to a water potential of –1.5 MPa as the wilting point of soil because this is the lowest water potential at which most plants can obtain water from the soil. There are, however, species of drought-adapted plants that can extract
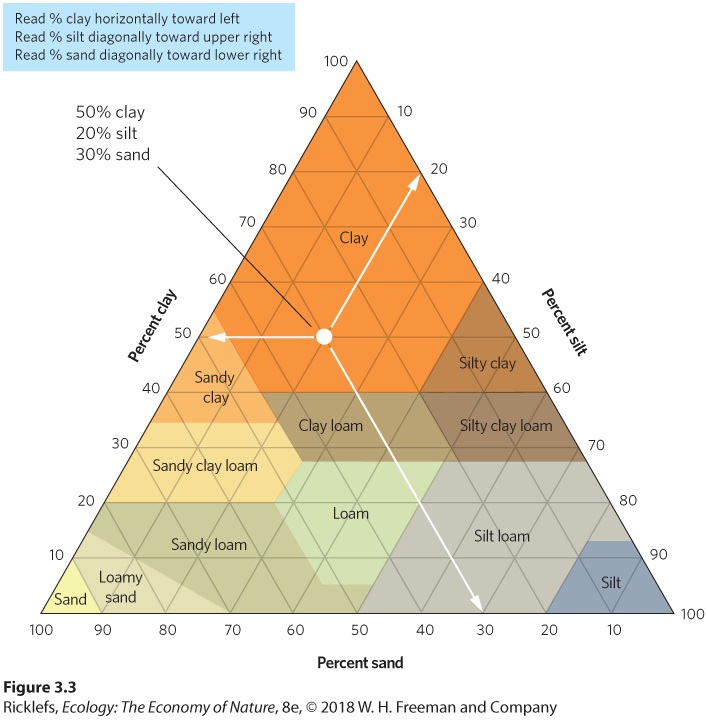
water when the water potential is less than –1.5 MPa. Wilting point The water potential at which most plants can no longer retrieve water from the soil, which is about –1.5 MPa. The amount of water in soil and its availability to plants depend on the physical structure of the soil. It also explains why the amount of water the soil can hold depends on the soil’s surface area; the more surface area a volume of soil has, the more water it can hold. The surface area of soil depends on the sizes of the particles that comprise the soil. Soil particles include sand, silt, and clay, in addition to organic material from decomposing organisms. As shown in Figure 3.2, sand particles are the largest, with diameters exceeding 0.05 mm. Silt particles have diameters of 0.002 to 0.05 mm and clay particles are the smallest, with a diameter of less than 0.002 mm. Rarely is a soil composed of a single particle size. Instead, as illustrated in Figure 3.3, soils are typically composed of mixtures of different ratios of each particle size. For example, a soil composed of 40 percent sand, 40 percent silt, and 20 percent clay is classified as a loam soil. In contrast, a soil containing a higher proportion of silt and a lower proportion of sand is classified as a silt loam soil.
Figure 3.2 Soil particle size. (a) Soil particles are separated into three sizes: clay, silt, and sand. (b) Each soil particle attracts a surface film of water around it. The greater surface area of small clay particles holds a greater total amount of water than the much larger sand particles, which have a much smaller surface area relative to their volume.
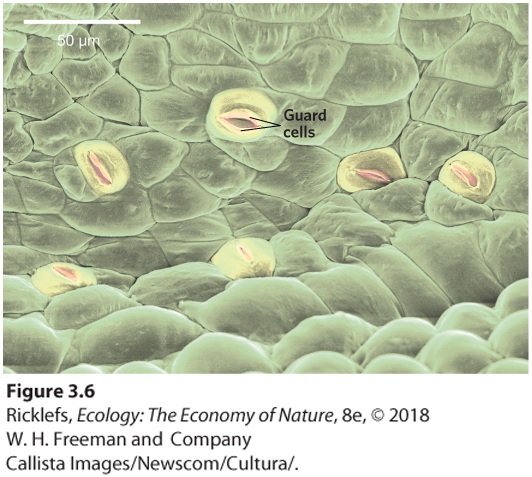
Figure 3.3 Combinations of soil particle sizes used to categorize soils. Most soils are composed of different percentages of sand, silt, and clay. Each name represents a category with a specific composition of the three particle sizes. Smaller particles have a larger surface area, relative to their volume, compared to larger particles. As a result, the total surface area of particles in a given volume of soil increases as particle size decreases. Therefore, soils with a high proportion of clay particles hold more water than soils with a high proportion of silt particles, which hold more water than soils with a high proportion of sand particles. Soils with a high proportion of sand particles tend to dry out because water quickly drains away, leaving many tiny pockets of air between the large sand particles. Clay soils represent the opposite extreme; each tiny particle of clay can attract a thin film of water on its surface, leaving little space for air pockets. Although clay soils retain a lot of water, clay particles can hold water molecules so tightly that it can be difficult for plants to extract the water from the soil.
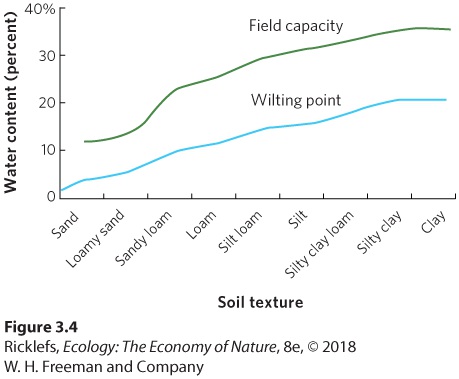
In Figure 3.4, we can see how soil particle size affects the amount of water in the soil, measured in terms of the percent of the soil volume occupied by water. As we move from sand to silt to clay, there is an increase in field capacity. However, there is also an increase in the wilting point. The difference between the field capacity and the wilting point is the amount of water available to plants. Thus, even when precipitation is frequent, sandy soils cannot retain much of the water that enters the soil. At the other extreme, clay soils can retain a lot of water, but if precipitation is not frequent enough for the soil to reach its field capacity, most of the water in the soil will be unavailable. This means that soils high in sand or high in clay are both poor soils for growing many plants, including crops that humans rely on for food. Instead, soils that contain a mixture of clay, silt, and sand particles —such as loam—are some of the best soils for growing plants.
Figure 3.4 Water-holding capacity of different soils. Soils composed of different combinations of sand, silt, and clay differ in their water-holding capacity. Soils containing large amounts of sand have a low field capacity and a low wilting point. In contrast, soils containing large amounts of clay have a high field capacity and a high wilting point.
Osmotic Pressure and Water Uptake
In Chapter 2, we noted that osmotic forces cause water molecules to move from areas of low solute concentration to areas of high solute concentration. At the same time, ions and other solutes diffuse through water from regions of high solute concentration to regions of low solute concentration. In the case of a plant, if a root cell has a higher solute concentration than the soil water, osmotic forces can draw the water into the root. It is this osmotic
potential in plant roots that causes water to enter the roots from the soil against the attractive forces of soil particles and the downward pull of gravity. Without any other adaptations, we would expect the solute concentrations within the root cell and in the soil water to eventually come into equilibrium. At this point, the osmotic potentials of the root cell and its surroundings would be equal, and there would be no net movement of water into the plant. However, root cells possess two adaptations that prevent this equalization. First, semipermeable cell membranes prevent larger solute molecules from leaving the plant’s root. Second, cell membranes can actively transport ions and small molecules against a concentration gradient into the root cells. These two adaptations maintain high solute concentrations inside the roots and allow strong osmotic forces to continue. As noted earlier, plants growing in places with very negative water potentials typically have adaptations to help them extract water beyond –1.5 MPa. Plants living in deserts, for example, can lower the water potential of their roots to as much as –6 MPa, thereby overcoming soil water potentials down to –6 MPa. Plants living in very salty environments can also meet the challenge of extracting water from an environment that contains unusually high concentrations of solutes in the form of salt ions. In both situations, plants have evolved adaptations that allow them to increase the concentrations of amino acids, carbohydrates, or organic acids in their root cells. Maintaining these high concentrations of dissolved substances, however, comes at a high metabolic price for the plants because they must divert some of the energy that would normally be used for growth and use it instead to manufacture these additional organic compounds. Plants lacking the appropriate adaptations grow poorly when exposed to salty conditions. For example, in the deserts of the American Southwest, a large amount of land is irrigated to grow crops, including cotton and orchard trees. Most well water, however, contains small amounts of salt. As this well water is sprayed over the fields, it moves down into the soil and dissolves any salts. If the irrigation system uses large amounts of water, the water can move deeply into the soil and the salts can be flushed away. If irrigation uses smaller amounts of water—just enough to feed the plant roots—the water stays near the soil’s surface. As much of this water is then taken up by plants or evaporates, the salt is left behind on the soil’s surface.
##### Transpiration and the Cohesion–Tension Theory
With repeated irrigation events, the salt concentration of the soil continually increases. After many years, the soil can have such a high solute concentration that many crop plants cannot create a lower water potential than the soil, and therefore cannot obtain sufficient water. The process of repeated irrigation that causes increased soil salinity is known as salinization. High-salt soils occur across 831 million hectares (ha) of land and in 100 countries of the world. It is a particular problem throughout arid areas of the world where irrigation typically is limited to small amounts of water, which concentrates the salts at the soil surface. In 2014, researchers reviewed studies from across the globe and estimated that salinization caused $27 billion in lost crop production around the world. While it is possible to reverse the salinization of soil, it can be costly and take many years. Salinization The process of repeated irrigation, which causes increased soil salinity. TRANSPIRATION AND THE COHESION–TENSION THEORY We have seen how osmotic potential draws water from the soil into the cells of plant roots. How does that water get from the roots to the leaves? Plants conduct water to their leaves through tubular xylem elements, which are the empty remains of xylem cells in the cores of roots and stems, connected to form the equivalent of water pipes. The movement of water through these xylem cells depends on the cohesion of water molecules and differences in water potential between the leaves and roots. The cohesion of water is the result of mutual attraction among water molecules. The attraction of hydrogen bonds causes a water molecule moving up the xylem of a plant to pull other water molecules with it. Cohesion of water helps the entire column of water move through the long vessels of a tall tree. The process, shown in Figure 3.5, begins when osmotic potential in the roots draws water from the soil into the plant and creates a root pressure that forces water into the xylem elements. However, this pressure is counteracted by gravity and the osmotic potential inside living root cells. Because of these two important counteracting forces, root pressure under the best circumstances can raise water to a height of no more than about 20 m, but the tallest trees can achieve heights of more than 100 m.
Figure 3.5 Water movement in plants by cohesion and tension. Differences in water potential, also known as tension, cause water to move from the soil into the roots, from the roots into the stem, and from the stem into the leaves. The cohesion of water causes the water molecules to adhere to each other and to move as a single column of water up the xylem cells. Cohesion The mutual attraction among water molecules. Root pressure When osmotic potential in the roots of a plant draws in water from the soil and forces it into the xylem elements. Fortunately for plants, leaves can also generate water potential as water evaporates from the surfaces of leaf cells inside the leaf and into the tiny air spaces that surround these cells. This evaporated water ultimately moves out
of the leaf and into the air in a process known as transpiration. The column of water in a xylem element is continuous from the roots to the leaves, since it is held together by hydrogen bonds between the water molecules. Thus, low water potentials in leaves can literally draw a column of bonded water molecules upward through the xylem elements against the osmotic potential of the living root cells and the pull of gravity. The water potential is strong enough under most conditions to pull water up through the roots, xylem, and leaves. As a result, the water potential from transpiration creates a continuous gradient from leaf surfaces in contact with the atmosphere down to the surfaces of root hairs in contact with soil water. The movement of water is due to both water cohesion and water tension (which is another name for differences in water potential). This mechanism of water movement from roots to leaves due to water cohesion and water tension is therefore known as the cohesion–tension theory. Transpiration The process by which leaves can generate water potential as water evaporates from the surfaces of leaf cells into the air spaces within the leaves. Cohesion–tension theory The mechanism of water movement from roots to leaves due to water cohesion and water tension. Based on the cohesion–tension theory of water transport in plants, very tall plants should have a more difficult time moving water up their stems because the movement of a tall column of water in the plant is being opposed by the force of gravity. Recent research estimates that this system limits plants to a maximum height of 130 m. In support of this prediction, the tallest tree that has been reliably measured was a 126-m Douglas fir tree. Although transpiration generates a powerful force that moves water through a plant, when the soil reaches the wilting point, there is insufficient water moving into the roots to replace the water lost from the leaves. To prevent further water loss from the leaves under dry conditions, plants have various adaptations for reducing transpiration. Most of the cells on the exterior of a leaf are coated with a waxy cuticle that retards water loss. As a result, gas exchange between the atmosphere and the interior of the leaf primarily occurs through small openings on the surface of the leaves, called stomata (Figure 3.6). The stomata (stoma, singular) are the points of entry
for CO2 and the points of exit for water vapor escaping to the atmosphere by transpiration. When plants experience a scarcity of water, they can reduce water loss to the atmosphere by closing their stomata. As leaf water potential weakens, the guard cells that border a stoma collapse slightly, causing the guard cells to press together and close the stoma. Although closing the stomata provides the important benefit of reducing water loss, it comes at the cost of preventing CO2, needed for photosynthesis, from entering the leaf. As we will soon see, plants living in hot, dry environments have evolved additional adaptations for dealing with this undesirable side effect.
Figure 3.6 Stomata. Stomata are pores in the surfaces of leaves, each bordered by two guard cells. Under conditions of low water availability, the guard cells close the opening and prevent the loss of water from the leaves. Stomata Small openings on the surface of leaves, which serve as the points of entry for CO2 and exit points for water vapor.
Concept Check
1. Explain the relationship between soil particle size and the field capacity of soil. 2. Why is the availability of plant water highest in soils with intermediate particle sizes?
3. How can we be sure that root pressure is not sufficient to explain the movement of water in trees?
#### 3.2 Sunlight Provides the Energy for Photosynthesis
3.2 Sunlight provides the energy for photosynthesis Whether in water or on land, energy from the Sun is essential for the existence of most life on Earth. To understand how this energy is captured, we need to examine the energy that is available, the energy that is absorbed, and how this energy is converted into a usable form by photosynthesis. Plants have evolved several adaptive approaches for conducting photosynthesis in terrestrial environments; these adaptations coincide with the environmental conditions of different regions of the world.
Available and Absorbed Solar Energy
The energy from the Sun, known as electromagnetic radiation, is packaged in small particle-like units called photons. The energy of photons is related positively to their frequency and inversely to their wavelength; the highest energy photons have the highest frequency and the shortest wavelengths. Wavelengths are expressed in units of nanometers (nm), which is onebillionth of a meter. The different wavelengths of light can be separated using a prism. As you can see in Figure 3.7, infrared radiation has long wavelengths, which we know contain lower energy. Short wavelengths, such as ultraviolet radiation, contain higher energy. Between the two extremes of infrared and ultraviolet radiation are wavelengths collectively known as visible light, which are visible to the human eye. Visible light represents only a small part of the spectrum of electromagnetic radiation.
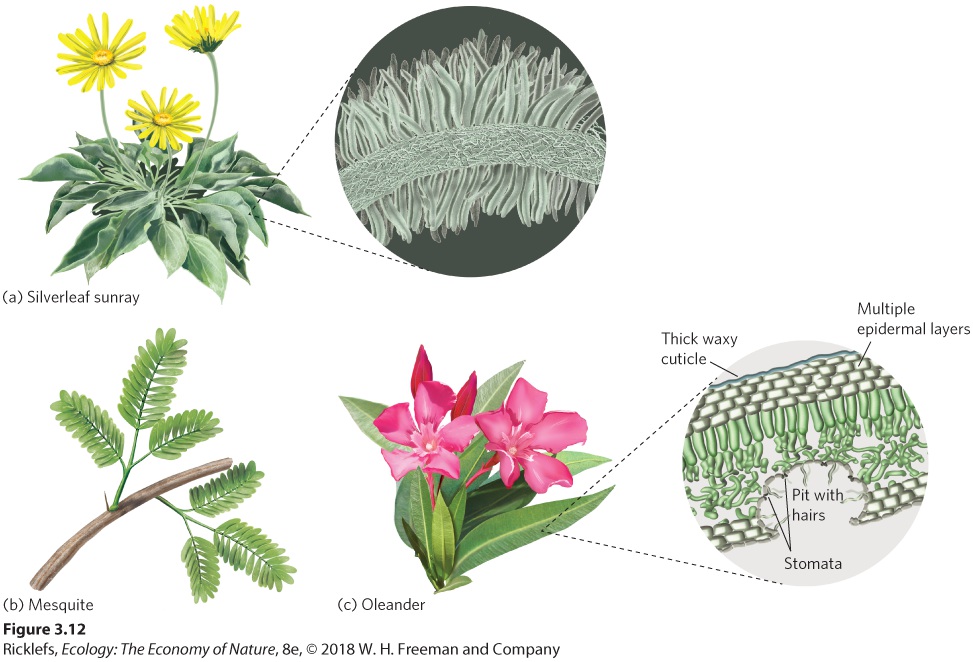
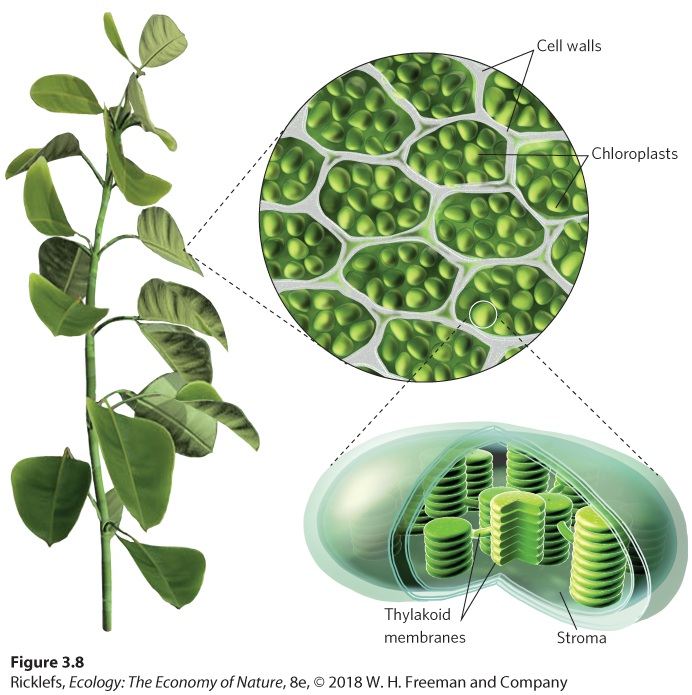
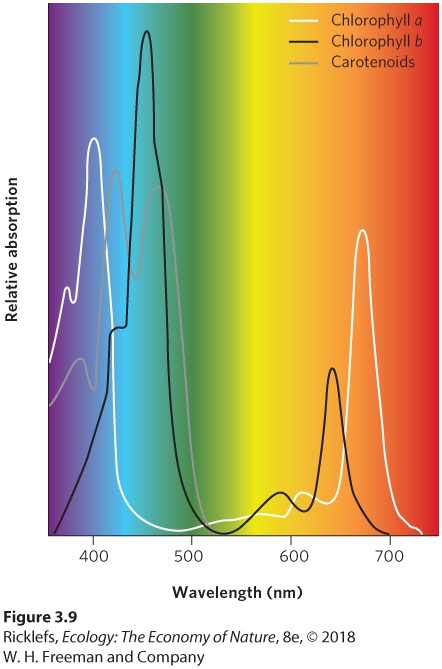
Figure 3.7 The wavelengths of the Sun’s energy. The Sun emits electromagnetic radiation that spans a wide range of energy and wavelengths. Electromagnetic radiation Energy from the Sun, packaged in small particle-like units called photons. Visible light Wavelengths in between infrared and ultraviolet radiation that are visible to the human eye. The visible portion of the spectrum includes the photosynthetically active region, which consists of wavelengths of light that are suitable for photosynthesis. This range of wavelengths falls between about 400 nm (violet) and 700 nm (red). Plants, algae, and some bacteria absorb these wavelengths and assimilate their energy by photosynthesis. These also happen to be the wavelengths of greatest intensity at the Earth’s surface. Photosynthetically active region Wavelengths of light that are suitable for photosynthesis. Eukaryotic photosynthetic organisms contain specialized cell organelles known as chloroplasts. As you can see in Figure 3.8, chloroplasts contain stacks of membranes known as thylakoids and a fluid-filled space surrounding the thylakoids called the stroma. Embedded within the thylakoid membranes are several kinds of pigments that absorb solar radiation,
including chlorophylls and carotenoids. The patterns of absorption for several of these pigments are shown in Figure 3.9.
Figure 3.8 Chloroplasts. Chloroplasts are the site of photosynthesis. The chloroplasts contain stacks of membranes called thylakoids surrounded by a fluid-filled space known as the stroma.
Figure 3.9 Light-absorbing pigments. Photosynthetic organisms contain a variety of photosynthetic pigments including chlorophyll a and b and carotenoids that serve as accessory pigments that capture the Sun’s energy and pass it to the chlorophylls. Chloroplasts Specialized cell organelles found in photosynthetic organisms. Chlorophylls, which are primarily responsible for capturing light energy for photosynthesis, absorb red and violet light. Chlorophylls reflect green and blue light, which is why leaves on most plants are predominantly green in color. Over the past 60 years, scientists identified four types of chlorophyll that differ in the wavelengths that they absorb: chlorophyll a, b, c, and d. Chlorophyll a is found in all photosynthesizing organisms and is responsible for the actual steps in photosynthesis. The other types of chlorophyll act as accessory pigments, meaning that they capture light energy and then pass it on to chlorophyll a. Recently, scientists reported the discovery of a fifth type of chlorophyll that they named chlorophyll f. This pigment, discovered in

algae that live in shallow rocky pools on the coast of Australia, absorbs light at longer wavelengths than the others. Carotenoids are also accessory pigments that absorb primarily blue and green light, thereby complementing the absorption spectrum of chlorophyll. Carotenoids, the pigments that give carrots their orange color, reflect yellow and orange light. By containing carotenoids and several types of chlorophyll, producers can absorb a wider range of solar energy and use it to power photosynthesis.
Photosynthesis
In this section, we review the process of photosynthesis and the different pathways that have evolved under different ecological conditions. As you may recall from your introductory biology course, the process of photosynthesis involves photosynthetic pigments absorbing energy from photons of light. This energy is then converted into chemical energy stored in the high-energy bonds of organic compounds. In its simplest form, photosynthesis is the process of combining CO2, H2O, and solar energy to produce glucose (C6H12O6) and oxygen: 6 CO2+6 H2O+photons→C6H12O6+6 O2 This simple equation summarizes a long chain of complex chemical reactions that takes place in two parts: light reactions and the Calvin cycle. Light reactions depend on light energy from the Sun and include a series of events from the absorption of light to the production of high-energy compounds and oxygen (O2). These high-energy compounds are ATP (adenosine triphosphate) and NADPH (nicotinamide adenine dinucleotide phosphate). The cell uses the energy in these compounds to convert CO2 into glucose in a process known as the Calvin cycle, which takes place in the stroma of the chloroplast. Over evolutionary time, three distinct biochemical pathways have evolved for the Calvin cycle: C3, C4, and CAM photosynthesis. As we will soon see, each of these pathways is suited to particular ecological conditions. C3 Photosynthesis For most plants, photosynthesis begins with a reaction between CO2 and a

five-carbon sugar known as RuBP (ribulose bisphosphate) to produce a sixcarbon compound. This reaction is catalyzed by the enzyme RuBP carboxylase-oxidase (also known as Rubisco). Once the six-carbon compound is created, it immediately splits into two molecules of a threecarbon sugar called G3P (glyceraldehyde 3-phosphate). We can represent this process as CO2+RuBP→2 G3P RuBP carboxylase-oxidase An enzyme involved in photosynthesis that catalyzes the reaction of RuBP and CO2 to form two molecules of glyceraldehyde 3-phosphate (G3P). Also known as Rubisco. This photosynthetic pathway, in which CO2 is initially assimilated into the three-carbon compound (G3P), is known as C3 photosynthesis. The vast majority of plants on Earth use C3 photosynthesis. In most plants, this process occurs in mesophyll cells in leaves. C3 photosynthesis The most common photosynthetic pathway, in which CO2 is initially assimilated into a three-carbon compound, glyceraldehyde 3-phosphate (G3P). One of the challenges for plants using C3 photosynthesis is that Rubisco, the enzyme responsible for joining CO2 and RuBP, has a low affinity for CO2. Consequently, carbon assimilation using Rubisco is quite inefficient at the low concentrations of CO2 found in the mesophyll cells of plant leaves. To achieve high rates of carbon assimilation, plants must therefore pack their mesophyll cells with large amounts of the enzyme Rubisco. In some plant species, Rubisco can constitute up to 30 percent of the dry weight of leaf tissue. Rubisco’s low affinity for CO2 is not the only problem plants face. Under certain conditions—such as when temperatures are high, concentrations of O2 are high, and CO2 concentrations are low—Rubisco preferentially binds to O2 rather than CO2. This occurs when hot, dry conditions cause the leaf stomata to close, preventing new CO2 from entering to replenish the CO2 that
has been consumed by the Calvin cycle. Closed stomata also prevent O2 produced by the light reaction from leaving the leaf. Consequently, hot and dry conditions lead to changes in CO2 and O2 concentrations that cause Rubisco to preferentially bind to O2 rather than CO2. When the Rubisco enzyme binds to O2, it initiates a series of reactions that reverses the outcome of photosynthesis in a process known as photorespiration: 2 G3P→RuBP+CO2 Photorespiration The oxidation of carbohydrates to CO2 and H2O by rubisco, which reverses the light reactions of photosynthesis. The reverse reaction consumes energy and O2, and produces CO2. This reverse reaction in plants is called photorespiration because it resembles the process of respiration. In short, what is accomplished by photosynthesis when Rubisco binds to CO2 is undone by photorespiration when Rubisco binds to O2. The problem of photorespiration is caused, in part, by closed stomata. This leads to high O2 concentrations and low CO2 concentrations in the leaves. One potential solution is to keep the leaf stomata open. This would permit free gas exchange, allowing CO2 to enter the leaves and O2 to exit the leaves. This strategy works as long as plants can replace the water they also lose by transpiration when the stomata are open. However, this solution may be too costly in hot, dry environments where water is scarce. When such costs are too high, natural selection will favor traits that can either reduce the demand for water or reduce the loss of water. C4 Photosynthesis We have seen that hot and dry conditions cause stomata to close, which results in a decrease of CO2, an increase of O2, and an increase in photorespiration. To address this problem, many herbaceous plants, particularly grasses growing in hot climates, have evolved a modified pathway known as C4 photosynthesis. Biologists call this modification C4 photosynthesis because the first step joins CO2 with a three-carbon molecule
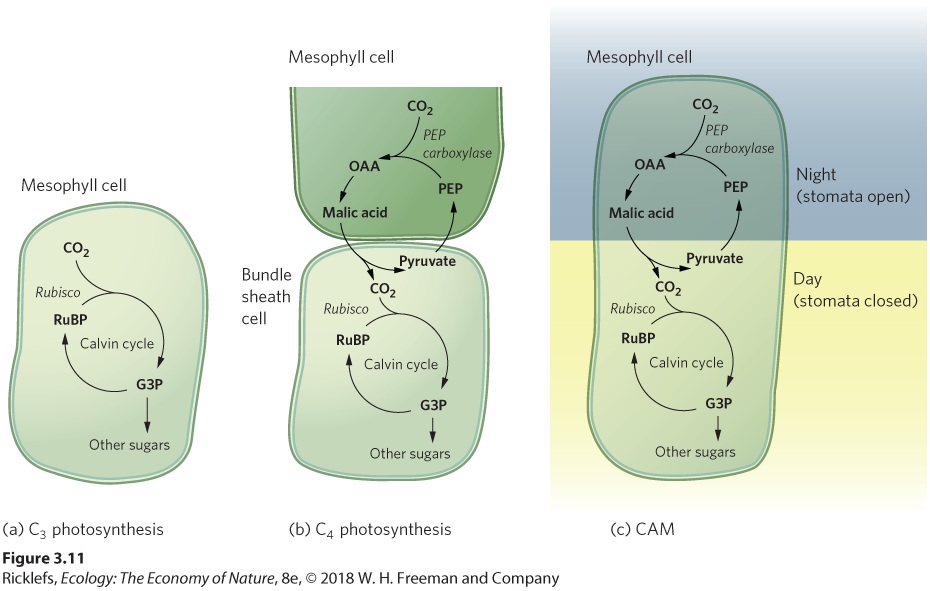
called PEP (phosphoenol pyruvate) to produce a four-carbon molecule called OAA (oxaloacetic acid): CO2+PEP→OAA C4 photosynthesis A photosynthetic pathway in which CO2 is initially assimilated into a four-carbon compound, oxaloacetic acid (OAA). This reaction is the key difference between C3 and C4 photosynthesis. It is catalyzed by the enzyme PEP carboxylase, which has a higher affinity for CO2 than Rubisco. You can see an overview of this process in Figure 3.10.
Figure 3.10 Leaf cell arrangements for C3 versus C4 plants. (a) C3 plants conduct all steps of photosynthesis in the chloroplasts of the mesophyll cells. (b) In C4 plants, the initial step of carbon assimilation and the light reactions take place in the chloroplasts in mesophyll cells. However, the CO2 that has been assimilated is then transported to the bundle sheath cells, where the Calvin cycle occurs. This additional assimilation step occurs in the mesophyll cells of the leaf, which is also the site of the light reaction. In most C4 plants, however, the Calvin cycle takes place in the bundle sheath cells that surround the leaf veins. This means that the plant must move the CO2 that has been assimilated in the mesophyll cells over to the bundle sheath cells. To do this, the plant converts OAA into malic acid, which then diffuses into the bundle sheath cells, where another enzyme breaks it down to produce CO2 and pyruvate, a three-carbon compound. In the bundle sheath cells, the chloroplasts use the
CO2 that is brought over from the mesophyll cells for the Calvin cycle. To complete the cycle, pyruvate moves back into the mesophyll cells, where it is converted back to PEP to be used once again. In short, the C4 pathway is an adaptation that adds a step to the initial assimilation of CO2 to make it more efficient when CO2 is present at low concentrations. This strategy solves the problem of hich reverses the light reactions of photorespiration by creating concentrations of CO2 in the bundle sheath cells that are three to eight times higher than is available to C3 plants. It helps that there are multiple mesophyll cells for each bundle sheath cell, thereby providing a large number of sites for CO2 assimilation that can provide CO2 to each bundle sheath cell. At this higher CO2 concentration, the Calvin cycle operates more efficiently. In addition, because the enzyme PEP carboxylase has a high affinity for CO2, it can bind CO2 at a lower concentration in the cell. This pathway allows the stomata to remain partially or completely closed for longer periods of time, which reduces water loss. However, C4 photosynthesis has two disadvantages that reduce its efficiency: Less leaf tissue is devoted to photosynthesis, and some of the energy produced by the light reactions is used in the initial C4 carbon assimilation step. Whereas C3 plants are favored in cool, moist climates, C4 plants are favored in climates that are hot or have less abundant water. When water is abundant, the C4 pathway does not present a distinct advantage because the costs of the C3 pathway are relatively low. When water is less abundant, however, the C4 pathway has an advantage. C4 photosynthesis has evolved at least 45 times during the past 30 million years in at least 19 different families of angiosperms. However, only about 4 percent of all plant species on Earth are C4 plants and they are primarily found in two types of nonwoody plants: grasses and sedges. C4 plants dominate tropical and subtropical grasslands and are important components of the plant communities found in arid regions of the world, including the Great Plains of North America. Plants using the C4 pathway also include many of our most important crop plants such as corn (maize), sorghum, and sugarcane. These plants are highly productive during hot growing seasons.
They also account for 20 to 30 percent of all CO2 fixation and 30 percent of all grain production. As a result, C4 plants can play substantial roles in the ecosystems in which they live. CAM Photosynthesis Certain succulent plants that inhabit water-stressed environments, for example, cacti and pineapple plants, use the same biochemical pathways as C4 plants. However, rather than separating CO2 assimilation and the Calvin cycle spatially, using mesophyll and bundle sheath cells, these succulent plants separate the steps in time. Plants that follow this pathway, known as crassulacean acid metabolism, or CAM, open their stomata for gas exchange during the cool night, when transpiration is minimal, and then conduct photosynthesis during the hot day. The discovery of this arrangement was first made in plants of the family Crassulaceae (the stonecrop family)— which includes the jade plant (Crassula ovata). Crassulacean acid metabolism (CAM) A photosynthetic pathway in which the initial assimilation of carbon into a four-carbon compound occurs at night. Like the C4 plants, CAM plants use an initial step of assimilating CO2 into OAA, which is then converted to malic acid and stored at high concentrations in vacuoles within the mesophyll cells of the leaf. The enzyme responsible for the assimilation of CO2 works best at the cool temperatures that occur at night, when the stomata are open. During the day, the stomata close, and the stored organic acids are gradually broken down to release CO2 to the Calvin cycle. A different enzyme with a higher optimum temperature, geared to promote daytime photosynthesis, regulates the regeneration of PEP from pyruvate following the release of CO2. Because CAM plants can conduct gas exchange during the night when the air is cooler and more humid, a plant using CAM photosynthesis reduces its water loss. Thus, CAM photosynthesis is an adaptation that results in extremely high water use efficiencies and enables plants that use this pathway to live in very hot and dry regions of the world. While CAM allows photosynthesis to occur in water-limited conditions, it happens at a relatively slow rate. As a result, CAM plants
typically grow much more slowly than C3 or C4 plants. Figure 3.11 compares the three alternative photosynthetic pathways.
Figure 3.11 Alternative photosynthetic pathways. (a) C3 plants conduct photosynthesis in mesophyll cells during the day. (b) C4 plants separate the steps of photosynthesis in space. The initial CO2 assimilation step occurs in the mesophyll cells and the remaining steps occur in the bundle sheath cells. (c) CAM plants separate the steps of photosynthesis in time. Assimilation of CO2 happens at night and the remaining steps occur during the day. Plants possessing the C3 pathway are better adapted to cool, wet conditions, whereas plants possessing the C4 and CAM pathways are better adapted to warm and arid conditions. However, there is not a clear distinction in where these different types of plants grow. For example, regions that are hot and dry during the summer can be cool and moist during the winter and spring. As a result, such an area can be dominated by short-lived C4 plants in the summer and short-lived C3 plants in the winter and spring. Moreover, different photosynthetic pathways represent just one of several adaptations that plants have evolved to handle hot temperatures and a scarcity of water. As we will discuss in the next section, many plants also have evolved structural adaptations.
Structural Adaptations to Water Stress
Heat- and drought-adapted plants have anatomical and physiological

modifications that improve water uptake and retention, reduce transpiration, and reduce the buildup of heat in their tissues. These adaptations include roots that can take advantage of different water sources, resistance to heat buildup, vein configurations that protect against air blockages, and morphological adaptations in leaves. Plants living in arid regions often have very shallow or very deep roots, which represent two different adaptive strategies. Plants with very shallow roots, such as many species of cacti, are able to rapidly uptake water from brief rain events in which the rain does not penetrate very far into the soil. Cacti often pair this adaptation with thick, succulent tissues that can hold a great deal of water whenever it becomes available. In contrast to the cacti, some perennial shrubs, such as mesquite, have roots that can extend several meters down into the soil, allowing the shrubs to access water that is very deep below the surface. Another strategy to combat the effects of heat and drought is to protect plant surfaces from direct sunlight with leaf resins, waxy cuticles, spines, and hairs. You can see many of these adaptations illustrated in Figure 3.12. Resins help seal off much of the leaf from water loss, whereas waxy cuticles help make the surfaces of the plant more resistant to losing water. Spines and hairs produce a boundary layer of still air that traps moisture and reduces evaporation. In some cases, to reduce water loss, the stomata are recessed into deep pits that contain hairs. Because thick boundary layers can also retard heat loss, hair-covered surfaces are also prevalent in arid environments that are cool. Long spines can also serve as structures that dissipate excess heat away from the plant.
Figure 3.12 Structural adaptations of plants against heat and drought. (a) Spines and hairs on leaf surfaces, such as in the silverleaf sunray plant, shade the plant from direct sunlight and reduce evaporation. (b) Finely divided leaves, as found in the mesquite plant, help dissipate any built-up heat. (c) Stomata that are recessed into deep pits containing hairs, as in the oleander plant, slow the evaporation rate of water out of the leaf. Some adaptations reduce heat buildup. Plants can reduce their heat loads by producing finely subdivided leaves with a large ratio of edge to surface area. This large amount of leaf edge breaks up the boundary layer surrounding the leaf, which helps dissipate heat from the leaf. Some desert plants have no leaves at all. Many cacti rely entirely on their stems for photosynthesis; their leaves are modified into thorns for protection. For a long time, scientists observed that plants in habitats with high temperatures generally had smaller leaves than plants in habitats with abundant water. Scientists hypothesized that the smaller leaves represented an adaptation that permitted the dissipation of heat. However, small leaves are not only found in plants from hot, dry places, but also in plants from cool, dry places. Smaller leaves also contain a higher density of large veins that transport and distribute water to the many small veins. In 2011, an international team of scientists discovered that having small leaves with a high density of large veins is actually an adaptation to overcome the problem of air bubbles, known as embolisms, which can form in large veins. Under severe drought stress, air can move into the stomata and up into the large veins, causing a bubble to form in the vein and blocking water movement. A
large density of veins allows the plant to get around this problem by sending water through adjacent veins. This suggests that the small leaf size is actually an adaptation to scarce water in both hot and cool environments, and the fact that small leaves can better dissipate heat in the hot environments may be a valuable secondary benefit. Collectively, these structural adaptations help make it possible for plants to live in regions of the world that have high temperatures or scarce water.
Concept Check
1. Explain how light serves as the ultimate source of energy for a meat-eating animal. 2. Why is C3 photosynthesis inefficient when the concentration of CO2 in a leaf is low? 3. Explain how plants use structural adaptations to reduce water loss.

#### 3.3 Terrestrial Environments Pose a Challenge for Animals to Balance Water, Salt, and Nitrogen
3.3 Terrestrial environments pose a challenge for animals to balance water, salt, and nitrogen While plants have a variety of adaptations for conducting photosynthesis on land, terrestrial life presents many additional challenges, including the need to maintain a balance of water, salt, and nitrogen. This balance is known as homeostasis. Homeostasis is an organism’s ability to maintain constant internal conditions in the face of a varying external environment. All organisms exhibit homeostasis to some degree—for example, the balance of water and salt, or the regulation of body temperature. Homeostasis An organism’s ability to maintain constant internal conditions in the face of a varying external environment. Although the occurrence and effectiveness of homeostatic mechanisms vary, all homeostatic systems exhibit negative feedbacks, meaning that when the system deviates from its desired state, or set point, internal response mechanisms act to restore that desired state. An example that might be familiar to you is the presence of negative feedbacks in the regulation of body temperature. Figure 3.13 shows how this works in mammals. The hypothalamus—a gland in the brain—determines whether the body temperature is above or below the desired set point, which differs among species of mammals. If body temperature drops below this set point, the hypothalamus uses neural and hormonal signals to trigger the body to generate more heat. When the body temperature reaches the set point, the hypothalamus triggers the body to stop producing heat. If the body temperature greatly exceeds the set point, the hypothalamus triggers the body to start using cooling mechanisms, including sweating and panting. Negative feedbacks The action of internal response mechanisms that restores a system to a desired state, or set point, when the system deviates from that state.
Figure 3.13 Negative feedback for regulating body temperature. In mammals, the hypothalamus acts like a thermostat. When the desired set point differs from the current body temperature, the hypothalamus signals the body to return its temperature to the desired set point. In Chapter 2, we saw that to maintain the proper amounts of water and dissolved substances in their bodies, aquatic organisms must balance their gains and losses. This is equally true for organisms on land. Organisms often take in water with a solute concentration that differs from that of their bodies, so they must either acquire additional solutes to make up the deficit or rid themselves of excess solutes. If they do not balance the concentration of solutes, many of their physiological functions will not operate correctly. When water evaporates from the surfaces of terrestrial organisms into the atmosphere, solutes are left behind and their concentration in the body tends to increase. Under such circumstances, organisms must excrete excess salts to maintain the proper concentrations in their bodies.
Water and Salt Balance in Animals
Water is as important to terrestrial animals as it is to terrestrial plants. Terrestrial animals, with their internalized gas exchange surfaces, are less vulnerable to respiratory water loss than plants. Moreover, because terrestrial animals are not immersed continuously in water, they have little trouble retaining ions. They acquire the mineral ions they need in the water they drink and the food they eat, and they use urine to eliminate excess salts in their bodies. Where fresh water abounds, animals can drink large quantities of water to flush out salts that would otherwise accumulate in the body. Where water is scarce, however, animals make use of adaptations to conserve water. As one would expect, desert animals have evolved a number of adaptations in response to water scarcity. Kangaroo rats, for example, are a group of small rodents that live in the dry regions of North America (Figure 3.14). Behavioral and physiological adaptations allow them to live in these places. The kangaroo rat conserves water by hunting for food during the nights and staying in a cool and humid underground burrow during the hot days—a valuable behavioral adaptation. For both the kangaroo rat and the camel, the kidneys provide an additional physiological adaptation to extreme heat and water scarcity. In all mammals, the kidneys are responsible for removing salts and nitrogenous wastes from the blood. These solutes are dissolved in water, but because water is valuable, a structure known as the loop of Henle helps recover some of it before the mixture is excreted. Kangaroo rats and camels possess an unusually long loop of Henle, which provides a greater length over which a kidney can recover water from urine prior to excretion. Whereas human kidneys concentrate most solutes in urine to about four times the blood concentration level, the kidneys of kangaroo rats produce urine with solute concentrations as much as 14 times greater.
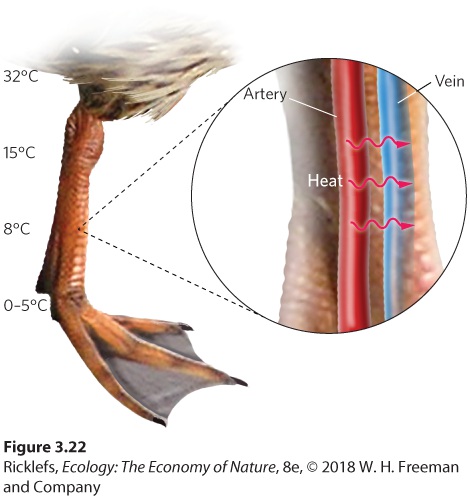
Figure 3.14 Animal adaptations to conserve water. The Ord’s kangaroo rat (Dipodomys ordii) lives in dry, desert environments in North America. These environments favor adaptations that allow the kangaroo rat to conserve water, including feeding at night and having large, efficient kidneys. Although the loop of Henle plays a large role in concentrating the urine of mammals, additional adaptations help conserve water. The efficiency of water use is best determined by the overall size of the kidney relative to the size of a mammal’s body. One way to assess the potential importance of relative kidney size as an adaptation for conserving water is to examine how relative kidney size differs among closely related species of rodents that live in habitats with differing amounts of precipitation. Figure 3.15 shows this relationship in a group of South American rodents. Rodents living in habitats with the lowest amount of precipitation have relatively large kidneys, whereas rodents living in habitats with the greatest amount of precipitation have relatively small kidneys.
Figure 3.15 Kidney size in South American rodents. Species of South American rodents that live in relatively drier habitats have relatively larger, more efficient kidneys. [After G. B. Diaz et al., Renal morphology, phylogenetic history and desert adaptation of South American hystricognath rodents, Functional Ecology 20 (2006): 609–620.] Because sodium and chloride ions participate in the water conservation mechanism of kidneys, the kidneys do not excrete these ions efficiently. Many animals that lack ready access to fresh water have specialized saltsecreting organs that work on a different principle from the kidneys. In fact, these secretory organs are more like the salt glands of mangrove plants. For example, salt glands of birds and reptiles, which are particularly well developed in marine species, are actually modified tear glands located in the orbit of the eye that are capable of secreting a concentrated salt solution. These adaptations help animals to balance their salt and water budget on land (Figure 3.16).
Figure 3.16 Animal adaptations to expel salt. Many animals that live on land and forage in salt water have evolved specialized glands to expel excess salt. The white crust on top of the head of this marine iguana (Amblyrhynchus cristatus) from the Galápagos Islands is salt that was produced in these glands and expelled through the nostrils.
Water and Nitrogen Balance in Animals
Most carnivores, whether they eat crustaceans, fish, insects, or mammals, consume excess nitrogen from the proteins and nucleic acids in their diet. When these compounds are metabolized, the excess nitrogen must be eliminated from the body. Most aquatic animals excrete excess nitrogen in the form of ammonia (NH3) because it is a simple metabolic by-product of nitrogen metabolism. Although ammonia is mildly poisonous to tissues, aquatic animals eliminate it rapidly either in copious dilute urine or directly across the body surface before it reaches a dangerous concentration within the body. Terrestrial animals, however, rarely have access to large quantities of water to excrete excess nitrogen. Instead, they excrete ammonia in forms that are less toxic than ammonia. This allows them to accumulate higher concentrations of metabolic by-products in their blood and urine without any harmful side effects. Mammals excrete nitrogen in the form of urea (CO(NH2)2), which is the same substance that we learned sharks produce and retain to achieve osmotic balance in marine environments. Because urea dissolves in water, excreting urea still causes the loss of some water, although the amount depends on the concentrating power of the kidneys. Birds and reptiles excrete nitrogen in the form of uric acid (C5H4N4O3). Uric acid
##### Analyzing Ecology: Understanding the Different Types of Variables
requires even less water and crystallizes out of solution. As a result, it can be excreted as a highly concentrated paste in the urine.
Analyzing Ecology
Understanding the Different Types of Variables When we think about testing hypotheses in ecology, we often collect and analyze data to determine whether the hypothesis is supported or refuted. However, before we collect and analyze the data, we have to consider what types of data we are collecting. The first distinction is between independent variables and dependent variables. Independent variables are factors that are presumed to cause other variables to change. Dependent variables are factors that are being changed. For example, in the examination of the relative kidney size of South American rodents (Figure 3.15), the hypothesis was that differences in precipitation have caused the evolution of different kidney sizes. In this case, precipitation is the independent variable and kidney size is the dependent variable. Independent variable A factor that causes other variables to change. Dependent variable A factor that is being changed. A second distinction is whether a variable is continuous or categorical. Continuous variables can take on any numeric value, including values that are not whole numbers. In the case of the rodents, for example, precipitation represents a continuous variable because it can take on any value (Figure 3.16). Other continuous variables include temperature, salinity, and light. In contrast, categorical variables, also known as nominal variables, fall into distinct groupings or categories. For example, if we wanted to know how the solute concentration of a kangaroo rat’s urine was affected by its diet, we might provide kangaroo
rats with one of three different species of seeds and then measure their urine concentrations. In this case, the diet is a categorical variable because the different diets fall into three distinct categories. Other categorical variables include sex (e.g., males and females) and species (e.g., dromedary camels, Bactrian camels, and guanacos). Continuous variable A variable that can take on any numeric value, including values that are not whole numbers. Categorical variable A variable that falls into a distinct category or grouping. Also known as Nominal variable. As we will see in later chapters, these distinctions between dependent versus independent variables and continuous versus categorical variables are important in the statistical analysis of ecological data. YOUR TURN If you were to conduct an experiment that examined how the water holding capacity differed among soil types, as in Figure 3.4, which would be the independent variable and which would be the dependent variable? If you were only to compare the water holding capacity of soils that contained 100 percent sand, 100 percent silt, or 100 percent clay, would this soil variable be considered continuous or categorical? Although excreting urea and uric acid conserves water, it is costly in terms of the energy needed to form these compounds. One method that scientists use to quantify energy costs is to determine the amount of organic carbon consumed to produce the energy necessary for excretion. For example, for each atom of nitrogen excreted in the form of ammonia, no atoms of organic carbon are used. In contrast, to excrete nitrogen in the form of urea requires 0.5 atoms of organic carbon, and uric acid uses 1.25 atoms of organic carbon.
1. Why does homeostasis require negative feedbacks?
2. Describe the costs and benefits associated with the different nitrogen products excreted by fish, mammals, and birds. 3. Contrast the concepts of dependent and independent variables.
#### 3.4 Adaptations to Different Temperatures Allow Terrestrial Life to Exist Around the Planet
3.4 Adaptations to different temperatures allow terrestrial life to exist around the planet On Earth, land temperatures can reach as high as 58 °C in northern Africa and as low as –89 °C in Antarctica. These extremes can limit the occurrence of life. To understand how organisms are affected by temperature and the adaptations that have evolved to deal with different temperatures, we first need to examine how they gain and lose heat.
Sources of Heat Gain and Loss
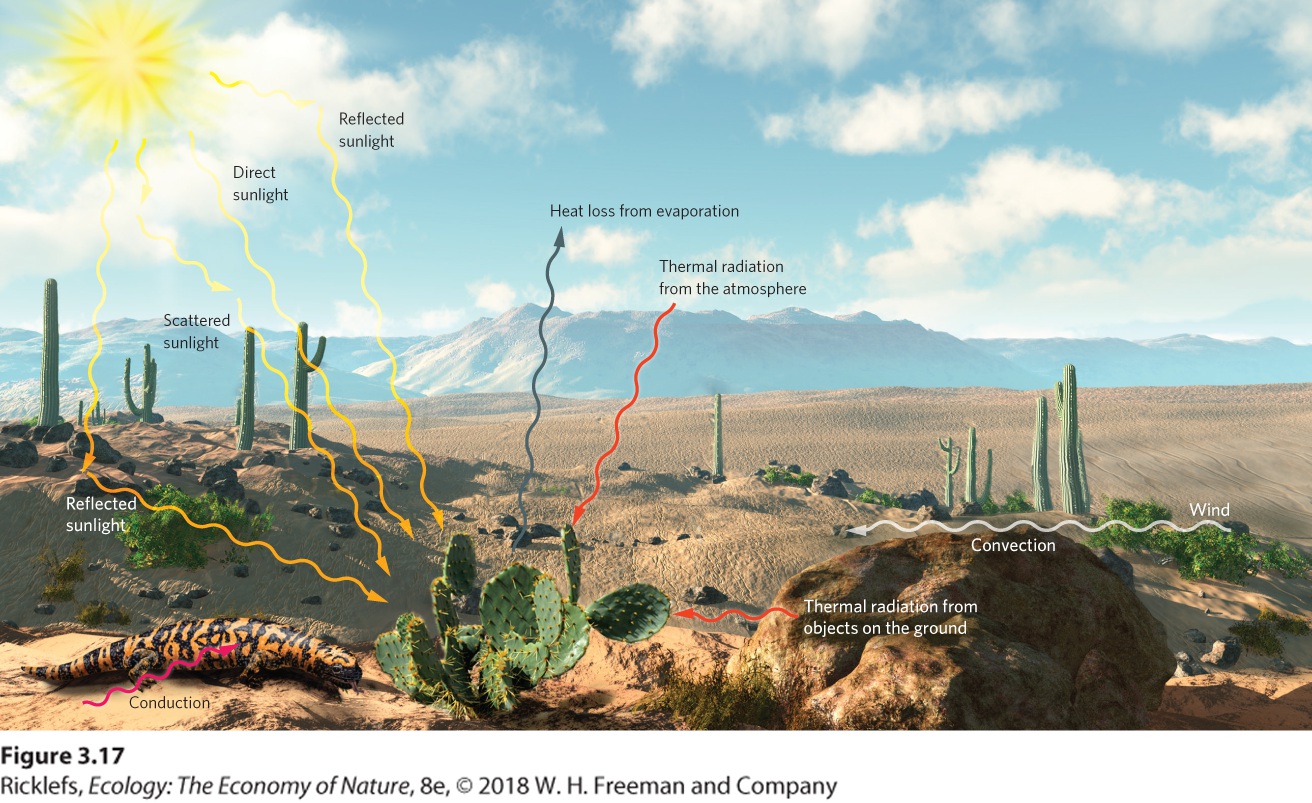
Because body temperature impacts physiological functions, organisms must manage heat gain and heat loss carefully. The ultimate source of heat at the surface of Earth is sunlight, most of which is absorbed by water, soil, plants, and animals and converted to heat. Objects and organisms continuously exchange heat with their surroundings. When the temperature of the environment exceeds the temperature of an organism, the organism gains heat and becomes warmer. When the environment is cooler than the organism, the organism loses heat to the environment and cools. As illustrated in Figure 3.17, this exchange of heat can occur through four processes: radiation, conduction, convection, and evaporation.
Figure 3.17 Sources of heat gain and loss. The Sun is the original source of almost all heat. The heat from the Sun is exchanged among objects across the landscape. In the case of the cactus, heat is gained by direct sunlight, scattered sunlight, and reflected sunlight. Heat can
be lost by evaporation of water vapor to the atmosphere. Heat can also be gained or lost by radiation from surrounding objects such as rocks, by conduction where the cactus comes into contact with the soil, and by convection as winds move hot or cold air over the surface of the cactus and disrupt its boundary layer. Radiation Radiation is the emission of electromagnetic energy by a surface. The primary source of radiation in the environment is the Sun. As objects in the landscape are warmed by solar radiation, they emit more lower-energy radiation in the form of infrared light. The temperature of the radiating surface determines how rapidly an object loses energy by radiation to colder parts of the environment. We measure in units of Kelvins (K), also known as absolute temperature, where 0 °C = 273°K. The amount of heat radiation increases with the fourth power of absolute temperature. For example, we can consider the heat radiation of two small animals such as a mouse and a lizard. If the mammal has a skin temperature of 37 °C (310°K) and the lizard has a skin temperature of 17 °C (290°K), the difference in heat radiation between the mammal and the lizard is 3104÷2904=130% This means that by having a 20 °C higher body temperature, the mammal radiates 30 percent more heat than the lizard. Radiation The emission of electromagnetic energy by a surface. The relatively high amount of heat radiation produced by animals with a higher body temperature than their external environment has been used by ecologists in a variety of research endeavors, including estimates of population sizes. When biologists need to count the number of moose living in remote regions of Alaska, for example, planes equipped with infrared cameras fly over these regions in the winter and the warm bodies of the moose stand out as a bright signal of infrared radiation against their cold, snowy background. Similar efforts have also been conducted to detect warm birds against a cold background environment, such as the penguins shown in
Figure 3.18. Infrared images. Thermal cameras can detect warm animals radiating heat against a cold background, such as these king penguins (Aptenodytes patagonicus) in the Blijdrop Zoo, Rotterdam, Netherlands. Conduction Conduction is the transfer of the kinetic energy of heat between substances that are in contact with one another. For example, lizards often lie flat on hot rocks to warm their bodies by conduction. Water, because it is so much denser than air, conducts heat more than 20 times faster. As a result, you would lose body heat much faster if you stood in 10 °C water than if you stood in 10 °C air. Conduction The transfer of the kinetic energy of heat between substances that are in contact with one another. The rate at which heat moves by conduction between an organism and its surroundings depends on three factors: its surface area, its resistance to heat transfer, and the temperature difference between the organism and its surroundings. An organism’s surface area helps determine its rate of heat conduction because a greater amount of exposed surface allows a greater surface for the energy transfer to take place. This is why many animals curl up in a ball to lessen the amount of exposed surface when they are trying to stay warm on a cold night. An organism’s resistance to heat transfer is just another way of saying how
much insulation the organism has. Thick layers of fat, fur, or feathers have a high resistance to heat transfer and therefore slow the rate of heat loss due to conductance. Indeed, this is why you choose to wear insulated boots rather than walking barefoot in snow. Finally, the rate of heat loss is higher when there are large differences between the temperature of the organism and that of the environment. As we will see in Chapter 4, this feature of conductance is why some hibernating animals lower their body temperatures during the winter. A lower body temperature results in less heat loss to the cold external environment. Convection Convection is the transfer of heat by the movement of liquids and gases: Molecules of air or water next to a warm surface gain energy and move away from the surface. In still air, a boundary layer of air forms over a surface of organisms. Having a thicker boundary layer will tend to slow heat transfer between an organism and its environment. When the environment is colder than the organism, the organism tends to warm this boundary layer, which in turn reduces the animal’s heat loss. If there is a current of air passing by the animal, the current disrupts the boundary layer and heat can be carried away from the body by convection. This convection of heat away from the body surface is the basis of the wind chill factor we hear about in winter on the evening weather report. Wind on a cold day makes you colder. For example, wind blowing at 32 km per hour in an air temperature of –7 °C has the cooling power of still air at –23 °C. Convection The transfer of heat by the movement of liquids and gases. In the same way that air movement can remove heat from a warm organism, air movement can add heat to an organism if the boundary layer is cooler than the surrounding air. If you were to stand in a hot desert, and your boundary layer were cooler than the air, for example, a hot wind would disrupt the boundary layer between your skin and the air and make your body even hotter. Evaporation
Evaporation is the transformation of water from a liquid to a gaseous state with the input of heat energy. Because evaporation removes heat from a surface, it has a cooling effect on an organism. As plants transpire and animals breathe, water evaporates from their exposed gas exchange surfaces, especially at higher temperatures. In dry air, the rate of evaporation nearly doubles with each 10 °C increase in temperature. Evaporation The transformation of water from a liquid to a gaseous state with the input of heat energy. As summarized in Figure 3.18, all these sources of heat gain and loss can occur simultaneously. Radiation from the Sun can occur as direct sunlight, as well as sunlight that has been scattered as it interacts with gas molecules in the atmosphere or is reflected from clouds and the ground. Plants and animals in contact with rocks, soil, and each other can conduct heat to or from these objects, depending on whether their body temperatures are warmer or colder than the surrounding objects. As winds move the air past the organisms, there can be an additional exchange of heat, depending again on the temperature of the air compared to the temperature of the organism. Finally, organisms that experience evaporation can lose heat because evaporation requires heat energy.
Body Size and Thermal Inertia
Most exchanges of energy and materials between an organism and its environment occur across body surfaces. Therefore, the volume and surface of an organism affect the rate of these exchanges. As an example, let’s consider the differences between the body sizes of a mouse and an elephant. The elephant obviously has a much larger volume and it takes much more energy to meet its metabolic needs each day. However, relative to its volume, the elephant has a smaller surface area than the mouse. This relationship becomes more apparent if we make the simplifying assumption that all organisms are shaped like a box with sides of equal length. In this case, the surface area (SA) of an organism increases as the square of its length (L), but the volume (V) of an organism increases as the cube of its length: SA=L2 V=L3
In short, as an organism grows larger, its volume grows faster than its surface area. Of course, organisms are not shaped like boxes, but the same principles apply to the shapes that organisms do have. Because an organism’s metabolic needs are related to its volume and volume increases faster than surface area, an organism’s metabolic needs increase faster than the surface area that exchanges energy and materials between the organism and its environment. The relationship between surface area and volume is particularly relevant when considering heat exchange. Because large organisms have a low surface-to-volume ratio, larger individuals lose and gain heat across their surfaces less rapidly than smaller individuals. In general, larger sizes and lower surface-to-volume ratios make it easier for organisms to maintain constant internal temperatures in the face of varying external temperatures. The resistance to a change in temperature due to a large body volume is known as thermal inertia. Although thermal inertia can be an important advantage in cold environments, in hot environments it causes moderately large individuals to have a harder time ridding themselves of excess heat. For this reason, large individuals run a greater risk of overheating. However, very large animals can benefit from thermal inertia under hot environmental conditions because their bodies heat up more slowly. We saw an example of this in the case of the dromedary camels, whose very large bodies slowly added heat during the day but then released this heat during the night. Thermal inertia The resistance to a change in temperature due to a large body volume.
Thermoregulation
The ability of an organism to control the temperature of its body is known as thermoregulation. Some organisms, known as homeotherms, maintain constant temperature conditions within the cells. Maintaining a constant internal body temperature allows an organism to adjust its biochemical reactions to work most efficiently. In contrast, poikilotherms do not have constant body temperatures. These terms tell us whether an organism’s temperature is constant or variable, not whether the temperature changes of the body are controlled internally or externally.
Thermoregulation The ability of an organism to control the temperature of its body. Homeotherm An organism that maintains constant temperature conditions within its cells. Poikilotherm An organism that does not have constant body temperatures.
Ectotherms
Ectotherms have body temperatures that are largely determined by their external environment. Ectotherms tend to be organisms with low metabolic rates—such as reptiles, amphibians, and plants—or small body sizes—such as insects—that cannot generate or retain sufficient heat to offset heat losses from their bodies. Ectotherm An organism with a body temperature that is largely determined by its external environment. Although ectotherms have body temperatures that match the temperature of their environment, they are not powerless to alter their body temperature. Indeed, many species of ectotherms adjust their heat balance behaviorally by moving into or out of shade, by changing their orientation with respect to the Sun, or by adjusting their contact with warm substrates. When horned lizards are hot, for example, they decrease their exposure to the ground surface by standing erect on their legs. When they are cold, they lie flat against the ground and gain heat both by conduction from the ground and from direct solar radiation. This behavior, known as basking, is common among reptiles and insects (Figure 3.19). Animals that bask in the radiation of the Sun can effectively regulate their body temperatures. Indeed, their temperatures may rise considerably above that of the surrounding air, well into the range of birds and mammals. Some larger species of ectotherms, such as tuna, can generate a substantial amount of heat by exercising their massive muscles. This flexing of large muscles allows such fish to stay warmer than their external environment, making it possible for them to swim and feed in relatively cold waters.
Figure 3.19 Basking. Ectotherms such as these painted turtles (Chrysemys picta) commonly lie in the sun to increase their body temperature. Some plants can occasionally generate enough heat to make their tissues substantially warmer than the external environment. The skunk cabbage (Symplocarpus foetidus), for example, is a foul-smelling plant that lives in wet soils in eastern North America (Figure 3.20). The odor attracts insect pollinators such as flies that typically feed on dead, rotting organisms. The skunk cabbage sprouts new leaves in early spring, even when snow still covers the ground. The plant’s mitochondria generate enough metabolic heat in its tissues to raise its temperature more than 10 °C above the external environment. This incredible achievement requires a great deal of energy, but it provides a number of substantial benefits, including earlier flowering in the spring, more rapid development of flowers, and protection from freezing temperatures. In one species of skunk cabbage, scientists have discovered that generating heat also improves the rate of pollen germination and pollen tube growth in the flowers. The heat also benefits the pollinators, which can absorb some of the heat produced by the plant. Collectively, heat generation in plants can be very beneficial to both the plants and their pollinators.
Figure 3.20 Skunk cabbage. Using mitochondria to generate heat, the skunk cabbage can elevate its temperature by more than 10 °C above the environmental temperature. The elevated temperature melts a path through the snow in early spring, making the skunk cabbage one of the first plants to sprout and attract pollinators to its flowers. In considering the various adaptations of ectotherms, we should note that the internal temperatures of some species may not vary a great deal. This can occur when the environmental temperature is not highly variable. For example, fish living in the polar oceans experience very cold waters that have little temperature variation. These fish are ectotherms because their body temperatures are determined by the surrounding environment, but their body temperatures are nearly homeothermic.
Endotherms
Endotherms are organisms that can generate sufficient metabolic heat to raise body temperature higher than the external environment. Most mammals and birds maintain their body temperatures between 36 °C and 41 °C, even though the temperature of their surroundings may vary from –50 °C to +50 °C. Maintaining a higher body temperature provides the benefit of accelerated biological activity in colder climates, allowing endotherms an improved ability to find food and escape predators. Endotherm An organism that can generate sufficient metabolic heat to raise its body temperature higher than the external environment.
Sustaining internal conditions that differ significantly from conditions in the external environment requires a lot of work and energy. Consider the costs to birds and mammals of maintaining constant high body temperatures in cold environments. As air temperature decreases, the difference between the internal and external environments increases. Recall that heat is lost across body surfaces in direct proportion to this temperature difference. Consider, for example, an animal that maintains its body temperature at 40 °C. At an outside temperature of 20 °C, it loses heat much faster than it does at an outside temperature of 30 °C. The greater the difference between an animal’s body temperature and outside temperatures, the greater the heat loss. To maintain a constant body temperature, endothermic organisms must replace heat lost to their environment either by generating metabolic heat or by gaining heat through other means such as solar radiation, conduction, or convection. The rate of metabolism required to maintain a particular body temperature increases in direct proportion to the difference between the temperature of the body and the temperature of the environment.
Adaptations of the Circulatory System
You have probably noticed that when walking on a cold day, your hands and feet are the first body parts to become cold. Similarly, because the legs and feet of most birds do not have feathers, these extremities would be major potential sources of heat loss in cold regions if they were held at the same temperature as the rest of the body. Exposed extremities lose heat rapidly, due to their high ratio of surface area to volume. The conduction of heat, particularly from exposed extremities, works against the maintenance of a constant warm body temperature. Ectotherms and endotherms have evolved a number of adaptations to minimize the impact of chilled extremities and thereby to help maintain a warm temperature in the core of the body where many vital organs are located. One prominent adaptation is blood shunting. Blood shunting occurs when specific blood vessels can be shut off—at locations called precapillary sphincters—so that less of the animals’ warm blood flows out to the cold extremities such as the forelimbs and hindlimbs. Instead, much of this blood is redirected into the veins before it ever reaches the extremities. From the veins, it returns to the heart, as shown in Figure 3.21. By sending less blood
to nonvital areas such as the limbs, the blood experiences less cooling and allows the core of the animal’s body to maintain a constant internal temperature while expending less energy. Blood shunting An adaptation that allows specific blood vessels to shut off so less of an animal’s warm blood flows to the cold extremities.
Figure 3.21 Blood shunting. In cold environments, some animals can close certain blood vessels at their precapillary sphincters. This reduces the circular flow of blood from the arteries to the extremities and back to the veins, which limits the amount of chilled blood that returns to the heart. Another adaptation to cold extremities is countercurrent circulation. In
Chapter 2, we saw that fish maximize oxygen uptake by this mechanism; the blood in their gills flows in the opposite direction from water. A similar arrangement occurs with the position of veins and arteries in the extremities of many animals; arteries that carry warm blood away from the heart and toward the extremities are positioned alongside veins that carry chilled blood from the extremities back to the heart. Figure 3.22 shows an example. When a gull stands on ice or swims with its feet in frigid water, it conserves heat by using countercurrent circulation in its legs. Warm blood in arteries leading to the feet cools as it passes close to veins that return cold blood to
the body. Rather than being lost to the environment, heat is transferred from the blood in the arteries to the blood in the veins. The feet themselves are kept only slightly above freezing, which minimizes heat transfer to the environment. The muscles used in swimming and walking are in the upper part of the leg, insulated by feathers that keep the upper legs close to the core body temperature.
Figure 3.22 Countercurrent blood circulation. The arteries in a gull’s leg that carry warm blood from the heart to the feet are positioned next to the veins that carry chilled blood away from the feet and back to the heart. This positioning of the arteries and veins allows the birds to transfer heat from arteries to the veins. The diverse range of adaptations for life on land is a fascinating testament to the ability of natural selection to favor those traits that improve the fitness of organisms. Whether we consider the adaptation of plants to obtain water and nutrients, the variety of scenarios for conducting photosynthesis under different environmental conditions, or the ability of animals to balance water, salt, nitrogen, and heat, these adaptations have evolved to help make the transition from life in water to life on land. Throughout this book, we will return to many of these adaptations as we seek to understand the ecology of communities and ecosystems.
Concept Check
1. If a snake is lying on a rock in the desert sun, how is the snake’s body temperature affected by radiation, conduction, convection, and evaporation? 2. Why does an animal’s surface area increase more slowly than its volume? 3. Under what conditions could an ectotherm be homeothermic?
Concepts
The Challenge of Growing Cotton Cotton plants. Cotton has numerous adaptations that help it cope with hot and dry conditions. Throughout this chapter, we have examined the challenges of the terrestrial environment and the adaptations that organisms have evolved to deal with these challenges. For plants, there are the challenges of extracting water and nutrients from the soil and then photosynthesizing at the
cost of water lost due to transpiration. At the same time, plants need to balance their water and salts while also frequently coping with heat gains and losses that can place organisms at the limits of their temperature tolerance. To understand how an organism handles all these challenges, we turn to the common cotton plant. Cotton is a major crop throughout the world because of its many uses that range from clothing to cottonseed oil. Cotton is typically grown in regions of the world that have warm temperatures with either high or low amounts of precipitation. In the United States, cotton is a $5 billion crop and nearly half of the cotton is grown in the panhandle of Texas. Like many arid regions, the Texas Panhandle experiences droughts every few years. In 2011–2016, Texas experienced one of the worst long-term droughts ever recorded, and this made it a tough place to grow cotton. Although C3 plants like cotton are typically associated with moister habitats, cotton can also grow well in relatively dry places. However, extended droughts often cause farmers to irrigate cotton fields because young cotton plants have relatively small and shallow root systems that have a difficult time acquiring water. As we saw with cacti and other succulent plants, shallow root systems can be effective at taking up water from brief rain events, but only if a plant can store excess water in its tissues. However, cotton has a limited ability to store water. The soils in the Texas Panhandle range from loam to clay. Recall from our earlier discussion and Figure 3.4 that such soils have a moderate to high field capacity. However, they also have a moderate to high wilting point,
which makes it hard for cotton to extract water from these soils. In addition, shallow watering in arid regions can lead to soil salinization. In Texas, cotton is traditionally planted in early May and harvested in July or August. Because July and August can be very dry months, farmers have been experimenting with earlier planting dates. When cotton is planted in April, the cotton plants bloom in late June and produce a larger crop, in part because the plants avoid the drier months of July and August. Plants that are stressed by a lack of water can abort the development of their flowers, and flowers are the source of the cotton fibers. In addition, cotton planted earlier matures in June, when days are longer and the plants have more daylight hours to conduct photosynthesis. In 2010, researchers reported that while an earlier planting date is effective, it only works if the seedling plants are irrigated to supplement scarce natural rainwater. Without irrigation, these seedling plants that have a small root system cannot survive. To help cotton farmers achieve larger crop yields, scientists have conducted a great deal of research on how to make cotton more drought resistant. Researchers have recently found that when cytokinin, a natural plant hormone, is sprayed onto young cotton plants, it induces the plants to grow larger root systems. By growing more roots that can penetrate deeper into the soil, the cotton is less prone to experience the effects of a water shortage. Cytokinin also stimulates the cotton plants to build a waxy outer coating, which we know makes plants less susceptible to water loss. These two responses provide a 5 to 10 percent increase in crop yields under drought conditions.
Cotton plants also have to deal with hot temperatures. Researchers have found that the ideal temperature for growing cotton is a daytime high of 28 °C. However, temperatures where cotton is grown can easily exceed 38 °C. At these higher temperatures, the enzyme Rubisco does not work as well. As we discussed, Rubisco is a key enzyme for C3 photosynthesis; poorly performing Rubisco results in a lower rate of photosynthesis and, in turn, lower cotton yields. Plant breeding efforts have developed varieties of heat-tolerant cotton with variations of the Rubisco enzyme that perform well under higher temperatures. Other varieties of cotton have been bred to transpire higher amounts of water vapor out of their stomata, which improves the plant’s ability to cool itself through the process of evaporation. These varieties of cotton can only transpire more water vapor out of their stomata if they have an abundant supply of water coming in through their roots and must therefore be grown in soils that are either naturally moist or irrigated. While cotton has been cultivated for more than 6,000 years and bred for desirable qualities for at least 3,000 years, modern science is offering new opportunities to improve the plant’s growth and crop yield even more. In 2012 and 2014, researchers in China were able to sequence the genome of two species of cotton (Gossypium raimondii and G. arboretum). Having the genome should ultimately offer the opportunity to identify the genes responsible for characteristics such as tolerance to drought, heat, and salt. The story of cotton illustrates how adaptations allow a
species to live under a range of challenging environmental terrestrial conditions. It also illustrates how a knowledge of these adaptations can help farmers adjust their practices and aid scientists in cultivating varieties of plants that can better perform in the face of these challenges and produce higher crop yields. However, not all challenges faced by agricultural crops can be solved through plant breeding. When this happens, we are left to plant the crops in regions of the world where their adaptations are better suited to the environmental conditions. SOURCES: Fuguang, L., et al. 2014. Genome sequence of the cultivated cotton Gossypium arboretum. Nature Genetics 46:567–572. Pettigrew, W. T. 2010. Impact of varying planting dates and irrigation regimes on cotton growth and lint yield production. Agronomy Journal 102:1379–1387. Salvucci, M. E., and S. J. Crafts-Brander. 2004. Inhibition of photosynthesis by heat stress: The activation state of Rubisco as a limiting factor in photosynthesis. Physiologia Plantarum 120:179–186.
Summary of Learning Objectives
3.1 Most terrestrial plants obtain nutrients and water from the soil. Soil nutrients include nitrogen, phosphorus, calcium, and potassium. The water-holding capacity of soil, known as its field capacity, depends on the particle sizes in the soil. The ability of plants to absorb this water requires that the osmotic potential of the roots be stronger than the matrix potential of the soil. This water moves up the plant stem to its leaves through a combination of osmotic pressure, the cohesion of water molecules, and the force of transpiration. Key Terms: Water potential, Matric potential, Field capacity, Wilting point, Salinization, Cohesion, Root pressure, Transpiration, Cohesion–tension theory, Stomata
3.2 Sunlight provides the energy for photosynthesis. Within the full range of electromagnetic radiation produced by the Sun, only a narrow range of wavelengths is used by the photosynthetic pigments of plants. This solar energy is used to power the process of photosynthesis by splitting water molecules and producing molecular oxygen and sugar. There are three pathways of photosynthesis: C3, C4, and CAM. Each differs in how it captures CO2 and each works best in particular environmental conditions. These different pathways are often associated with structural adaptations that help plants from arid regions conserve water. Key Terms: Electromagnetic radiation, Visible light, Photosynthetically active region, Chloroplasts, RuBP carboxylase-oxidase, C3 photosynthesis, Photorespiration, C4 photosynthesis, Crassulacean acid metabolism (CAM)
3.3 Terrestrial environments pose a challenge for animals to balance water, salt, and nitrogen. Organisms attempt to achieve homeostasis in all these compounds, typically through the use of
negative feedbacks. Plants and animals both possess a number of adaptations to balance their concentrations of salt, water, and nitrogenous wastes. Key Terms: Homeostasis, Negative feedbacks, Independent variable, Dependent variable, Continuous variable, Categorical variable
3.4 Adaptations to different temperatures allow terrestrial life to exist around the planet. Organisms can gain and lose heat through radiation, conduction, convection, and evaporation. These processes combine to form an individual’s heat budget. Temperature can be regulated to different degrees by animals through the process of thermoregulation. Poikilotherms have variable body temperatures, whereas homeotherms have relatively constant body temperatures. The body temperatures of ectotherms are largely determined by their external environment, whereas endotherms can raise their body temperatures to be higher than the external environment. Additional adaptations to assist in thermoregulation include the shunting of blood and countercurrent circulation. Key Terms: Radiation, Conduction, Convection, Evaporation, Thermal inertia, Thermoregulation, Homeotherm, Poikilotherm, Ectotherm, Endotherm, Blood shunting
Critical Thinking Questions
1. How do salty soils affect the matric potential and, in turn, affect root pressure and overall water uptake? 2. Based on your knowledge of the relationship between volume and surface area, why do clay soils hold more water than sandy soils? 3. If you were breeding cotton plants to have increased water uptake in dry soils, you could breed plants that contain more amino acids and carbohydrates in the root cells. Why would this be effective and why might crop yield be negatively affected? 4. How do CAM plants solve the problem of obtaining CO2 for photosynthesis while minimizing water loss? 5. How might the fitness cost of photorespiration in C3 plants favor the evolution of C4 plants? 6. How do leaf hairs that surround guard cells reduce water loss through the process of forming a boundary layer? 7. When animals hibernate, they lower their temperature. How would this reduce the rate of heat lost through conduction? 8. In addition to having highly efficient kidneys, what behaviors could you imagine desert animals using to reduce water loss? 9. If you designed an experiment to determine how temperature and the species composition of seeds affect the growth and reproduction of seed-eating birds, what would be the independent and dependent variables? 10. If you were to stand in a river with fast-moving water, which heat exchange processes might occur and why?
#### Graphing the Data: Relating Mass to Surface Area and Volume
GRAPHING THE DATA Relating Mass to Surface Area and Volume In this chapter, we saw that the surface area of an organism increases approximately as the square of the organism’s length, whereas the volume increases approximately as the cube of the organism’s length. Using the following data, graph the relationship between length and surface area and the relationship between length and volume. Based on these two graphs, notice how increases in body length affect volume much more rapidly than surface area. Length (cm) Surface area (cm2) Volume (cm3) 1,000 8,000 27,000 1,600 64,000 2,500 125,000