12 Population Growth and Regulation
12Population Growth and Regulation Animal overpopulation. With the decline of top predators and an increase in suitable habitats, many large herbivores have dramatically increased in abundance to the point that it negatively affects native plants, other wildlife, and humans. In this photo, an overabundance of kangaroos graze on a golf course in Australia. Putting Nature on Birth Control We often hear about the decline of species around the world due to human activities, but some species are doing extraordinarily well. In particular, the populations of many large herbivores have increased to unprecedented numbers. Hundreds of years ago, white-tailed deer lived at relatively low densities throughout the eastern United States when most of the habitat was forested and top predators, such as wolves and mountain lions, roamed throughout the region. Over the past two centuries, much of the forested habitat gave way to farms,
and many of these farms have been subsequently converted into housing developments with a variety of vegetation that deer like to eat. At the same time, there was a push to exterminate the top predators throughout most of the eastern United States. With more food and less predation, deer today live at densities that are approximately 20 times higher than before European colonists arrived. Such high densities cause an over-grazing of plants, increased consumption of agricultural crops, and 1.5 million vehicle collisions and 150 human deaths each year. The story of the white-tailed deer is repeated around the world in many other species. For example, sika deer (Cervus nippon) in Nara, Japan, live in a region that is protected as a World War II monument. Tourists feed the deer, the native wolf is extinct, and the deer have experienced a population explosion to the point where they are stripping the bark off spruce trees and destroying the forest. In Australia, eastern gray kangaroos (Macropus giganteus) have also experienced a dramatic population increase with the extinction of the Tasmanian wolf (Thylacinus cynocephalus) and they are now grazing on agricultural crops and golf courses. In some cases, the growing populations of large herbivores are not native, such as the wild horses (Equus ferus) that were introduced to the American West, and the European and Russian hogs (Sus scrufa) that were introduced to the southern United States as domesticated hogs. In the case of the hogs, they have been spreading rapidly into 39 states over the past 30 years, and their rooting of soil and consumption of crops are causing an estimated $1.5 billion in damage. In all these cases, the current challenge is how to control populations that are experiencing unnatural population explosions and negatively impacting ecosystems and human activities. Several different strategies have been proposed for controlling overabundant wildlife populations. In localized areas, it can help to eliminate the feeding of animals. In large regions, some of the overabundant species are hunted in an effort to replace the historic role of top predators. However, in many regions of the world, the number of hunters has declined over the decades. In addition, some members of the public do not support hunting as a population control
method. As a result, it would be more palatable to the public if we developed methods of animal contraception. “The current challenge is how to control populations that are experiencing unnatural population explosions and negatively impacting ecosystems and human activities.” Research on wildlife birth control has been of interest for decades, but only recently has it become a more viable option. In the case of cats and dogs, there is a history of surgically sterilizing the animals, but this is rarely an option for wild animals in nature, so researchers have focused on nonsurgical contraception. Current approaches for controlling kangaroos are focusing on treating animals with hormones to trick the females into not ovulating. For deer, horses, and elephants, researchers have focused on injecting females with a protein from pig ovaries (porcine zona pellucida, or PZP) that cause the females to make antibodies that stick to an egg, thereby preventing sperm from fertilizing the egg. The researchers used dart guns to inject PZP into the animals and found that a high percentage of the females did not conceive in the subsequent year. At first glance, using birth control to control wild animal populations seems to be an excellent option given the lack of public support for culling animal populations through hunting. However, some of the contraceptives remain effective for only one year, finding and injecting individuals can cost $200 to $1,000 per animal, and the number of animals that must be treated to reduce a regional population can be thousands of individuals. Currently, researchers are working to develop longer-lasting contraceptives and new ways to deliver them, including the use of feeders connected to image-recognition systems to avoid giving drugs to the wrong animal. These research efforts are increasingly providing ways to reduce the problem of animal overpopulation.
Klein, A. 2016. Cont-roo-ception: Hormone implants bring kangaroos under control. New Scientist, June 20. https://www.newscientist.com/article/2094401-cont-roo-ception- hormone-implants-bring-kangaroos-under-control/. Nordstrum, A. 2014. Can wild pigs ravaging the U.S. be stopped? Scientific American, October 21. https://www.scientificamerican.com/article/can-wild-pigs-ravaging-the-u-s-be- stopped/. Gammon, K. 2011. Approved for use: The first birth control for wildlife. Popular Science, September 22. http://www.popsci.com/science/article/2011-08/birth-control-wildlife.
Learning Objectives
12.1 Explain why populations can grow rapidly under ideal conditions.
12.3 Describe how the factors of age, size, and life-history influence population growth. In the previous chapter, we examined the spatial distribution of populations. In this chapter, we will examine changes in population size, known as population dynamics. We will look at how populations grow, and we will explore the factors that regulate their growth. Historically, studies of population growth focused on human populations. Although estimates are understandably crude, experts believe that a million years ago, the ancestors of modern humans numbered a million individuals. With the advent of agriculture 10,000 years ago, greater food production allowed further population growth so that, by the year 1700, the human population had grown to about 600 million people. During the Industrial Revolution, increased wealth brought better nutrition, sanitation, and improvements in medical treatments. As a result, the mortality rate of children fell and the life span of adults increased. An exponential increase in population growth followed these developments. According to the United Nations, the world population hit 1 billion in 1804. Although it took more than 1 million years to reach this mark, it only took 123 years to reach the 2 billion mark in 1927. In recent decades, a billion people have been added to
the planet every 12 to 13 years, and the population has doubled in just 40 years. By 2017, our planet held 7.5 billion people. The rapid growth of the human population in the past has led many scientists to predict population sizes in the future. In the eighteenth century, the British economist Thomas Malthus examined data on human population growth and concluded that the rapid rate of growth would cause humans to outstrip the capacity of the food supply. This insight influenced nineteenthcentury scientists, including Charles Darwin, who realized that this reasoning could apply to every organism on Earth. Understanding population growth and regulation is important because it allows us to predict future population growth. This predictive ability helps us manage the population sizes of species that are harvested by humans, species that are declining and need to be saved from extinction, and pest species that invade new regions and need to be controlled. The study of populations is known as demography. In this chapter, we will explore key demographic models of how populations grow. Demography The study of populations.
#### 12.1 Populations Can Grow Rapidly under Ideal Conditions
12.1 Populations can grow rapidly under ideal conditions Populations of any species can grow at an incredibly rapid rate, given the right conditions. In this section, we will explore two mathematical models of population growth.
The Exponential Growth Model
To understand how populations grow, we must first understand the concept of population growth rate. The growth rate of a population is the number of new individuals that are produced in a given amount of time minus the number of individuals that die. We typically consider the growth rate on an individual, or per capita, basis. Under ideal conditions, such as abundant resources, available mates, and favorable abiotic conditions, individuals can have their maximum reproductive rates and minimum death rates. When this happens, a population achieves its highest possible per capita growth rate, which is called the intrinsic growth rate, denoted as r. For example, abundant resources allow a white-tailed deer to have twins, a western mosquito (Aedes sierrensis) to lay up to 200 eggs in a single clutch, and a white oak tree (Quercus alba) to produce more than 20,000 acorns in one reproductive season. Ideal conditions also lower death rates because stresses, such as hunger and disease, are decreased. Growth rate In a population, the number of new individuals that are produced in a given amount of time minus the number of individuals that die. Intrinsic growth rate (r) The highest possible per capita growth rate for a population. Once we know the intrinsic growth rate for a population, we can estimate how a population will grow over time under ideal conditions. To do this, we use the exponential growth model, so named because it assumes that growth is exponential under ideal conditions: Nt=N0ert Exponential growth model A model of population growth in which the population increases continuously at an exponential rate.
where e is the base of the natural log (e = 2.72). This equation tells us that when conditions are ideal, the size of the population in the future (Nt) depends on the current size of the population (N0), the population’s intrinsic growth rate (r), and the amount of time over which the population grows (t). Populations with higher intrinsic growth rates or a larger number of reproductive individuals will experience a greater rate of increase in population size. We can see how the exponential model behaves with an example using mice. If we begin with 100 mice and their intrinsic rate of growth is 0.4, we can forecast that the mouse population will increase to 739 mice in 5 years: Nt=N0ertN5=100e0.4×5N5=100e2N5=100×7.39N5=739 The exponential growth model produces a J-shaped curve, as shown in
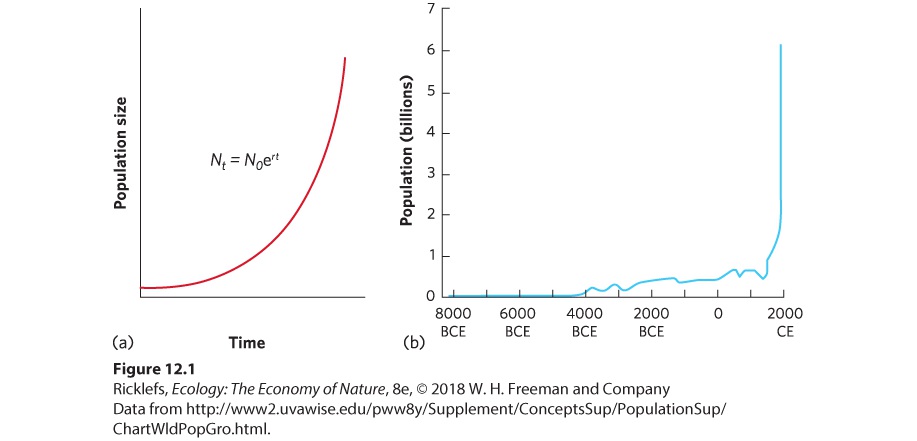
Figure 12.1a. As an example, Figure 12.1b illustrates that the human population has been growing exponentially during the past 300 years.
Figure 12.1 The exponential growth model. (a) Over time, a population living under ideal conditions can experience a rapid increase in population size. This produces a J-shaped curve. (b) Human population growth is an excellent example of a J-shaped curve. J-shaped curve The shape of exponential growth when graphed. Using the exponential model, we can also determine the rate of growth at
any point in time by taking the derivative of the exponential growth equation, as shown in the following equation: dNdt=rN where dNdt represents the change in population size per unit of time. In words, this equation tells us that the rate of change in population size at any particular point in time depends on the population’s intrinsic growth rate and the population’s size at that point in time. Another way to think about this equation is that it tells us the slope of the line relating population size to time at any given point. Looking at the graphs in Figure 12.1, for example, the slope of the line is very shallow early in time and becomes very steep later in time. This confirms that the population initially grows more slowly because there are a small number of reproductive individuals, but then grows much faster as the number of reproductive individuals increases. The exponential growth model for a population resembles the way money grows when it is earning interest in a bank account. Imagine that you have $1,000 in your account at 5 percent annual interest, that you don’t add or withdraw money from the account, and that any interest earned is deposited directly into the account at the end of the year. After 1 year, the balance would be $1,050, which represents an annual growth of $50. The balance after the second year would be $1,102.50—an annual growth of $52.50. In the tenth year, the annual growth would be $77.57, and in the twentieth year the annual growth would be $126.35. As you can see, a constant interest rate results in an ever-increasing balance. Similarly, a constant intrinsic growth rate results in ever-increasing numbers of individuals in a population and the population experiences exponential growth.
The Geometric Growth Model
The exponential growth model applies to species, such as humans, that reproduce throughout the year. However, most species of plants and animals have limited breeding seasons. For example, most birds and mammals reproduce in the spring and summer when there are abundant resources available for their offspring. As an example, let’s look at the California quail (Callipepla californica), a bird from western North America that lays one or two clutches of eggs in the spring. The quail population experiences a large

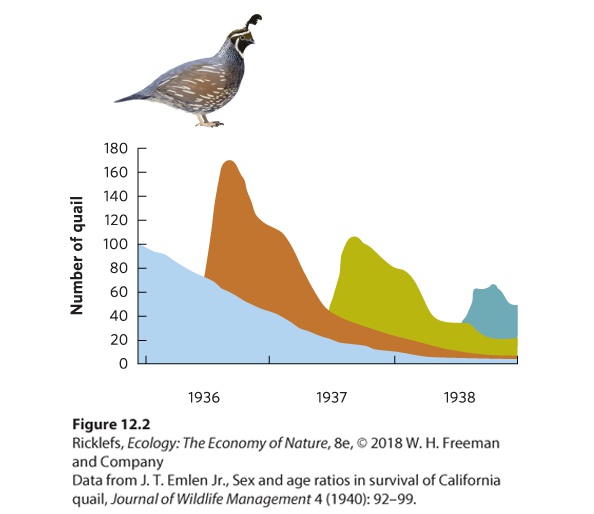
boost in its population size in the spring but then the population slowly declines over the summer, fall, and winter due to deaths. You can see the changes in population sizes in Figure 12.2, where each color in the graph represents the new generation of young quail that are produced each spring. When we examine the pattern of population abundance over several years, we can see that the exponential growth model, which assumes continuous births and deaths throughout the year, does not describe animals such as quail that have a distinct breeding period. For these species, ecologists use the geometric growth model because it compares population sizes at regular time intervals. In the case of the California quail, for instance, the geometric growth model allows us to compare population sizes at yearly intervals.
Figure 12.2 The discrete breeding events of the California quail. Each generation of new quail produced every spring is represented by a different color. Because births only happen in the spring, the growth of the California quail is better modeled by a geometric growth model than by an exponential growth model. Geometric growth model A model of population growth that compares population sizes at regular time intervals.
##### Comparing the Exponential and Geometric Growth Models
The geometric growth model is expressed as a ratio of a population’s size in 1 year to its size in the preceding year (or some other time interval). This ratio is assigned the symbol λ, which is the lowercase Greek letter lambda. A λ value greater than 1 means the population size has increased from 1 year to the next because there have been more births than deaths. When λ is less than 1, the population size has decreased from 1 year to the next because there have been fewer births than deaths. Because there cannot be a negative number of individuals, the value of λ is always positive. If N0 is the size of a population at time 0, then its size one time interval later would be N1=N0λ Next we can examine how we predict population size after more than one time interval. For example, if we wanted to estimate population size after two time intervals: N2=(N0λ)λ which can be rearranged to N2=N0λ2 In more general terms then, we can express the geometric growth model as Nt=N0λt where t equals time. Imagine that we start with a population of 100 quail and we have an annual growth rate of λ = 1.5. After 5 years, the size of the population is N5=N0×λ5N5=100×1.55N5=759
Comparing the Exponential and Geometric Growth
Notice that the equation for exponential growth is identical to the equation for geometric growth, except that er takes the place of λ. Thus, geometric and exponential growth are related by λ=er
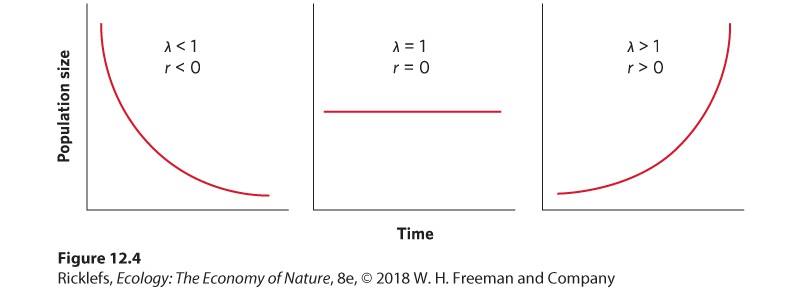
which can be rearranged to logeλ=r From this relationship, we can see that λ and r are directly related to each other. Indeed, if we were to graph the growth of populations over time using both models and setting λ = er, we would find identical growth curves. We see this comparison in Figure 12.3. Although the exponential model has continuous data points whereas the geometric model has discrete data points, the two models show the same pattern in population growth. Figure 12.4 compares the relationships between λ and r by examining the values of λ and r when populations are decreasing, constant, or increasing. When a population is decreasing, λ < 1 and r < 0. When a population is constant, λ = 1 and r = 0. When a population is increasing, λ > 1 and r > 0.
Figure 12.3 Comparing the growth of populations using the exponential and geometric growth models. The exponential model uses continuous data, whereas the geometric model uses discrete data that are calculated at each time point. However, the two models reveal the same increase in population size over time.
Figure 12.4 A comparison of λ and r values when populations are decreasing, constant, or increasing. When the population is decreasing, λ < 1 and r < 0. When the population is constant, λ = 1 and r = 0. When the population is increasing, λ > 1 and r > 0.
Population Doubling Time
We can appreciate the capacity of a population for growth by observing the rapid increase of organisms introduced into a new region with a suitable environment with abundant resources. In 1937, for example, two male and six female ring-necked pheasants were released on Protection Island, Washington. Within 5 years, the population increased to 1,325 adult birds, which means it experienced an annual growth rate of λ = 2.78. In other words, the population almost tripled, on average, each year. Populations maintained under optimal conditions in laboratories can have very high growth rates. Under ideal conditions, the value of λ can be as high as 24 for field voles (Microtus agrestis), a small mouse-like mammal, 10 billion (1010) for flour beetles, and 1030 for water fleas. One way of appreciating the potential growth rate of populations is to estimate the time required for a population to double in abundance, known as the doubling time. To understand how we determine doubling time, we can start by rearranging the exponential growth equation: Nt=N0ertert=Nt÷N0 Doubling time The time required for a population to double in size. When a population doubles, its size is twice its original size at time 0. As a result, we can replace (Nt÷N0) with the value 2 and determine the time required (t2) for a population to double in size:
ert=2rt=loge2t=loge2÷r For the geometric model, the equation is nearly the same, except r is replaced by loge λ: t2=loge2÷logeλ Given that the value of loge 2 is 0.69, the doubling time for the ring-necked pheasant, which has an annual growth rate of λ = 2.78, can be calculated as t2=0.69÷loge2.78t2=0.67 yearst2=246 days The same calculation gives doubling times of 79 days for the field vole, 11 days for the flour beetle, and just 3.6 days for the water flea. Using this calculation, demographers have determined that the current doubling time for the human population is 40 years. The exponential and geometric growth models are excellent starting points for understanding how populations grow. There is abundant evidence that real populations do initially grow rapidly, just as these models suggest. As we will see in the next section, however, no population can sustain exponential growth indefinitely. As populations become more abundant, they are limited by other factors such as competition, predation, and pathogens.
Concept Check
1. Why does the graph of population increase produce a J-shaped curve even though the intrinsic growth rate is a constant? 2. In what situations should we use the geometric population growth model? 3. Using the exponential growth model, what equation can we use to estimate the population doubling time?
12.2 Populations have growth limits In nature, we commonly observe that there are limits on how large a population can grow. Ecologists categorize these limits as being either density independent or density dependent. In this section, we will review these two types of limits on population growth and discuss how we can incorporate them into population growth models.
Density-Independent Factors
As the name suggests, density-independent factors limit population size regardless of the population’s density. Common density-independent factors include natural disasters such as tornadoes, hurricanes, floods, and fires. However, other less dramatic changes in the environment, including extreme temperatures and droughts, can also limit populations. In all these cases, the impact on the population is not related to the number of individuals in the population. Consider, for example, what happens when a hurricane hits a coastal forest. The number of trees that survive the hurricane is independent of the number of trees that were present before the hurricane arrived. Density independent Factors that limit population size regardless of the population’s density. A classic study on density-independent factors was conducted by James Davidson and Herbert Andrewartha in Australia. The apple thrip (Thrips imaginis) was a common insect pest in Australia that would occasionally increase to very large numbers and devastate apple trees and rose bushes throughout large regions of the country. From 1932 to 1946, the researchers surveyed thrip populations on rose bush flowers in an attempt to understand the causes of the large variations in population size. They suspected that these population changes were the result of density-independent factors, such as seasonal variation in temperature and rainfall, so they included these factors in their population growth model. Figure 12.5 shows the population sizes the researchers predicted and the population sizes they observed. As you can see, the predicted changes in thrip abundance closely matched the actual number of thrips they counted in their surveys.
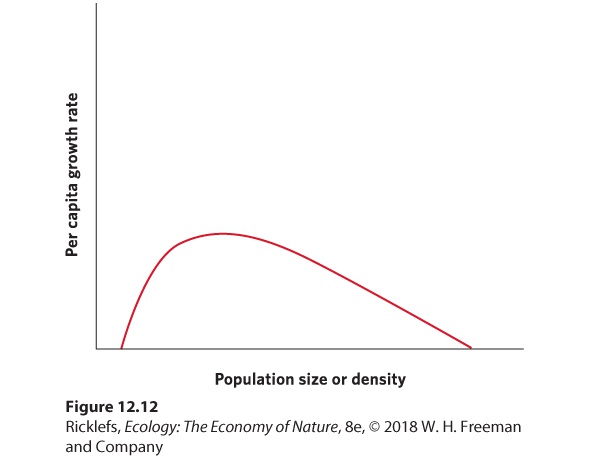
Figure 12.5 Predicting density-independent growth of the apple thrip. Scientists surveyed thrip populations on the flowers of rose bushes from 1932 to 1946. The dashed line represents the predicted number of thrips each year based on a model that included temperature and rainfall variables that cause density-independent effects on thrips. The bars represent the number of thrips actually observed each year. Understanding the role of density-independent factors remains an important goal of ecologists today, particularly because of the impact of global change. For example, in 2011, researchers from the U.S. Forest Service and the Canadian Forest Service reported on the impacts of bark beetles in western North America. Bark beetles are native insects that consume the phloem tissues of trees, causing them to die. During the past several decades, bark beetles have killed billions of coniferous trees covering millions of hectares from Mexico to Alaska (Figure 12.6). Under warmer temperatures, bark beetles develop faster and have more generations per year, but they can die from unusually cold temperatures. As a result, temperature is a major determinant of population size and its effects on the beetle population are density independent. Concerned about the possible effect of climate change on the population of bark beetles, researchers applied expectations of future climate warming and their knowledge of bark beetle ecology to project future growth. They concluded that bark beetle populations are likely to increase dramatically and
Figure 12.6 Bark beetles. Many of these lodgepole pines in central British Columbia, Canada, have been severely damaged by bark beetles, which consume the phloem tissues of the pine trees, causing them to turn brown and die. The mountain pine beetle (Dendroctonus ponderosae) is one of several species of bark beetles that is predicted to become more abundant with warmer global temperatures in the future.
Density-Dependent Factors
Density-dependent factors affect population size in relation to the population’s density. It is useful to break density-dependent factors down into two categories. Negative density dependence occurs when the rate of population growth decreases as population density increases. Positive density dependence (also known as inverse density dependence) occurs when the rate of population growth increases as population density increases. Because positive density dependence was first proposed by the ecologist Warder Allee in 1931, it is also known as the Allee effect. Density dependent Factors that affect population size in relation to the population’s density.
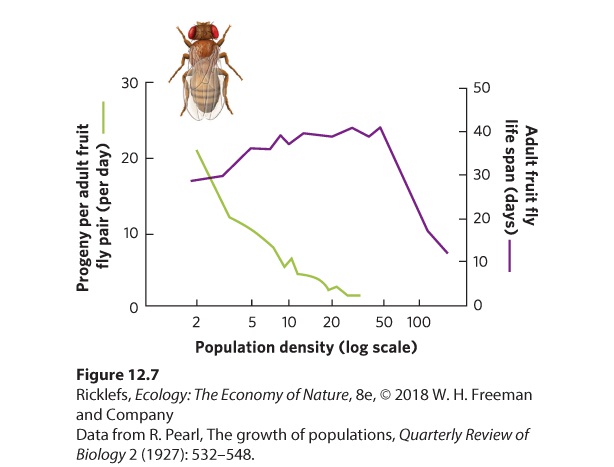
Negative density dependence When the rate of population growth decreases as population density increases. Positive density dependence When the rate of population growth increases as population density increases. Also known as Inverse density dependence or the Allee effect. Some of the most common negative density-dependent factors include a limited supply of resources, such as food, nesting sites, and physical space. When a population is small, there is an abundance of resources for all individuals, but as the population increases, such resources are divided among more individuals. As a result, the per capita amount of resources declines and at some point it reaches a level at which individuals find it difficult to grow, reproduce, and survive. This was the concern Malthus expressed in the eighteenth century about the rapid growth of the human population. Crowded populations also can have higher levels of stress, transmit diseases at a greater rate, and attract the attention of predators. All these factors contribute to slowing, and finally halting, population growth. Negative Density Dependence in Animals In a classic investigation of negative density dependence, Raymond Pearl raised breeding pairs of fruit flies (Drosophila melanogaster) at different densities in laboratory bottles containing identical amounts of food. With an increased number of adults, competition for food was more intense and the daily number of progeny produced per pair of adult flies decreased, as shown in Figure 12.7. In addition, the life span of the adults also sharply declined.
Figure 12.7 Limits on population growth. Larger populations cause increased competition. This graph shows that as competition among fruit flies increases, they experience a decline in the number of progeny produced per adult breeding pair per day. Increased competition also leads to a decline in the life span of adult flies. Although negative density dependence is easily demonstrated in the laboratory, a number of field experiments confirm that negative density dependence occurs in nature. For example, the common tern (Sterna hirundo) is a bird that nests on beaches. In the 1970s, the tern population on the eastern coast of North America began to expand into an area known as Buzzards Bay in Massachusetts. It first colonized Bird Island, where its numbers quickly grew from about 200 to 1,800 individuals by 1990, as shown in Figure 12.8. For these terns, the factor limiting population growth appears to be the availability of suitable nesting sites. By 1991, most suitable breeding sites on Bird Island were occupied and the population leveled off. The following year, birds began to colonize Ram Island, where the population increased to just over 2,000 birds and then leveled off by the year 2000. Around this time, the terns began to colonize Penikese Island. These data demonstrate that although the terns have a high intrinsic growth rate, a limited number of nesting sites ultimately limits the population size.
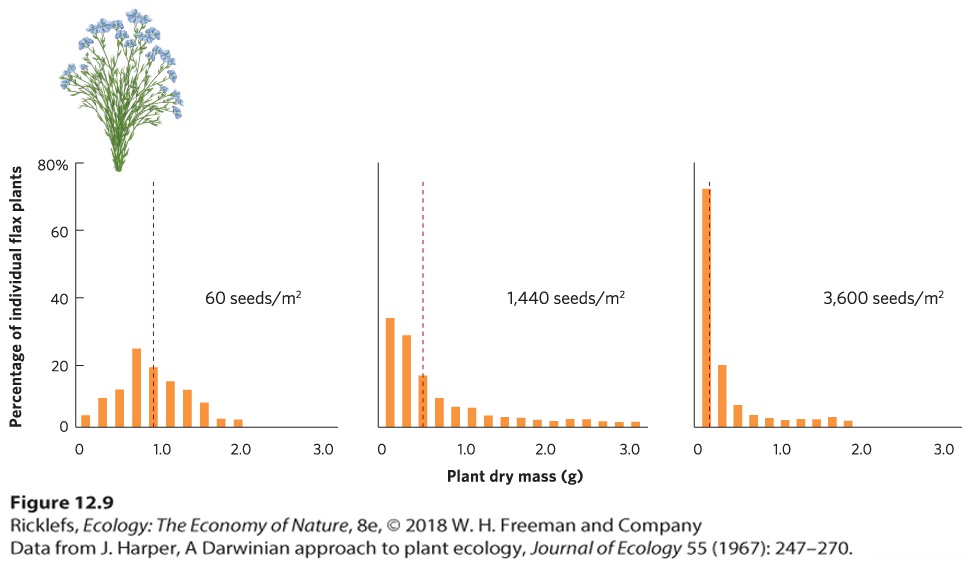
Figure 12.8 Negative density dependence in terns. As the tern population expanded in Buzzards Bay, Massachusetts, it colonized Bird Island. Rapid growth of the population filled most of the available nesting sites and the birds then colonized Ram Island. Once again, the population grew and occupied most of the available nesting sites. The terns then colonized Penikese Island. Negative Density Dependence in Plants The survival, growth, and reproduction of plants are also limited at high population densities. When plants are grown at high densities, each plant has access to fewer resources, such as sunlight, water, and soil nutrients. We can see this outcome in a study of flax plants (Linum usitatissimum) that were grown at a wide range of densities and then dried to determine their mass. The data from this experiment are shown in Figure 12.9. When seeds were sown at a density of 60 seeds per square meter, the average dry weight of individuals was between 0.5 and 1 g. When seeds were sown at densities of 1,440 and 3,600 per square meter, most of the individuals weighed less than 0.5 g. Plants with reduced growth typically experience reduced fecundity. As a result, the competition caused by high plant densities will cause a population of flax plants to grow more slowly.
Figure 12.9 Negative density dependence in flax. When flax seeds are sown at higher densities, the average plant is smaller. Smaller plants are less fecund, so high densities cause plant populations to increase at a slower rate. The dashed red line in each panel represents the mean dry mass. Under very high densities, competition among conspecifics can cause plants to die. In an experiment using horseweed (Erigeron canadensis), seeds were sown at a single extremely high density of 100,000 per square meter. Over time, competition among the tiny seedlings became intense, as you can see in Figure 12.10a, and over an 8-month period, most of them died. The two y axes in the figure reveal that there was a hundredfold decrease in population density over time, but a thousandfold increase in the average mass of the surviving individuals. As a result, the total mass of all surviving plants increased tenfold over time.
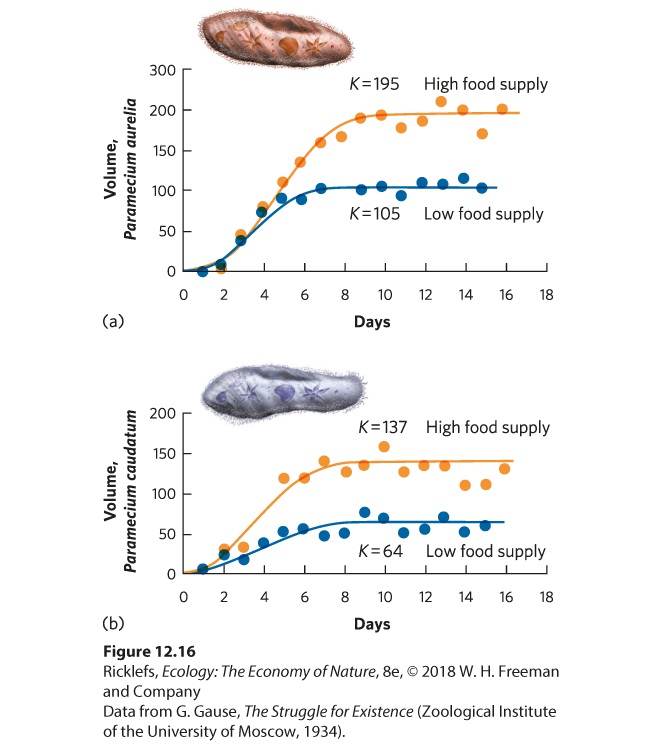
Figure 12.10 Self-thinning curve. Horseweed seeds were sown at a density of 100,000 per square meter. (a) Over time, the number of survivors declined by 100, while the average mass of the surviving plants increased by 1,000. (b) When we plot plant density against average mass per plant on log scales, we see it follows a line with a negative slope. Data from J. Harper, A Darwinian approach to plant ecology, Journal of Ecology 55 (1967): 247–270.
Figure 12.10b, shows the same data, but now with the changing density of surviving plants over time versus the change in average dry mass per plant over time, using a log scale. When we graph these data, they fall on a line with a negative slope. This is called a self-thinning curve, which is a graphical relationship that shows how decreases in population density over
time lead to increases in the mass of each individual in the population. The self-thinning phenomenon has been observed across a wide variety of species. The insights that emerge from the self-thinning curve have a number of practical uses. For example, the curve can be used to predict survival and growth of crop plants that might be sown at different densities in agricultural fields or the growth and survival of tree seedlings that might be planted at different densities in forest plantations. Self-thinning curve A graphical relationship that shows how decreases in population density over time lead to increases in the mass of each individual in the population. In plants, animals, and other taxonomic groups, negative density dependence tends to bring populations under control and to maintain their abundances at a level close to the maximum number that can be supported by the environment. In many cases, populations are affected by densityindependent and density-dependent factors. Next we consider the fascinating process of positive density dependence.
Positive Density Dependence
Whereas negative density dependence causes population growth to decrease as population density increases, positive density dependence causes population growth to increase as population density increases. Positive density dependence typically occurs when population density is very low and it can be caused by several different factors. For example, very low densities make it hard for individuals to find mates or, in the case of flowering plants, to obtain pollen, which can reduce reproductive success and slow population growth. As a result, a small increase in population density can have a positive effect on the population’s growth. Very low densities can also lead to the harmful effects of inbreeding, as discussed in Chapter 4. There can also be problems associated with small population sizes that, by chance, have uneven sex ratios. Small populations with a low proportion of females can suffer low population growth rates. Finally, individuals living in smaller populations can face a higher predation risk than those living in large populations, as we discussed in Chapter 10. In short, while negative density dependence causes slow population growth due to overcrowding, positive density dependence causes slow population growth due to undercrowding.
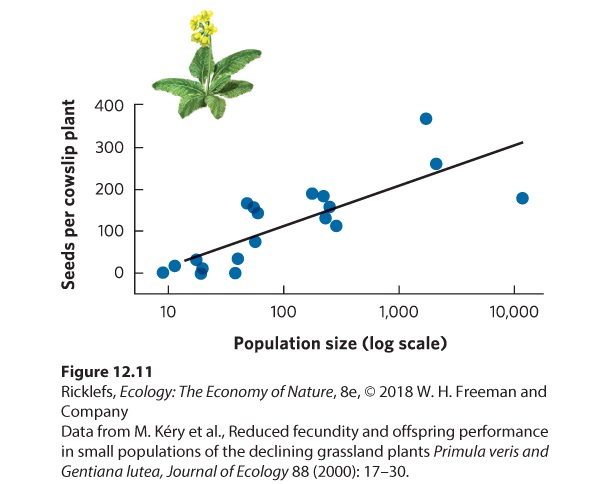
Positive density dependence has been demonstrated in a wide variety of species. Many plant species, for example, avoid the costs of inbreeding through a number of mechanisms that prevent self-fertilization. They depend on receiving pollen from other plants, but this can be difficult when individuals live at low densities and are widely spread over an area. In a study of cowslip (Primula veris), a plant of European grasslands, researchers were interested in determining why many of the smaller populations were declining. When they looked at reproduction in populations of different sizes, they found that populations of fewer than 100 individuals produced fewer seeds per plant, as shown in Figure 12.11. This probably occurred because the small populations were not as good at attracting pollinators. As a result, plants in low-density populations received less pollen and therefore had fewer fertilized flowers and produced fewer seeds.
Figure 12.11 Positive density dependence in cowslip plants. At low densities, plants appear to be pollen-limited, which causes them to produce fewer seeds. At higher densities, each plant produces many more seeds. Positive density dependence also occurs in parasites. In 2012, researchers in British Columbia reported on reproduction in salmon lice (Lepeophtheirus salmonis), which feed on the skin of salmon. When breeding, male and
female lice form a mating pair. The research team counted the number of mating pairs on each fish and found that the probability of an individual forming a mating pair is positively correlated with the number of lice of the opposite sex. Individuals living at low densities have a harder time finding a mate than individuals living at higher densities; therefore, the lice experienced positive density dependence. Although we often think of positive and negative density dependence as isolated phenomena, populations can be regulated by both processes, as illustrated in Figure 12.12. As we move from low densities to intermediate densities, the effects of positive dependence can play an important role. Increased densities provide more individuals for breeding and the growth rate of the population can improve. Above some intermediate density, resources start to become limiting and negative density dependence begins to play a role. As densities continue to increase, negative density dependence continues to slow population growth and, eventually, the population growth rate falls to zero.
Figure 12.12 Positive and negative density dependence. When populations are very small, an increase in density can cause an increase in the population’s per capita growth rate. However, when populations are large and resources start to become limiting, further increases in density can cause a decrease in the population’s per capita growth rate.
An example of the dynamic between positive and negative effects of density on population growth can be found in a population of herring (Clupea harengus) that spawns near Iceland. As you can see in Figure 12.13, the herring experience positive density dependence when the population is low in abundance, but negative density dependence when the populations is high in abundance The existence of positive density dependence in the herring population concerns fisheries managers because it indicates that if the herring were driven to low densities by fishing, it would be difficult for the population to bounce back. In fact, the populations could experience negative growth rates and become extinct. Fortunately, most fish populations do not appear to experience positive density dependence.
Figure 12.13 Positive and negative density dependence in herring. The highest population growth is measured as a ratio between the number of young fish and the number of adult fish. The highest growth rate of the population occurs at intermediate densities. There
is positive density dependence at low herring abundance but negative density dependence at high herring abundance. The herring study highlights why those who manage populations in the wild need to understand positive density dependence. For example, when a species has declined to very low numbers and we wish to improve its numbers, management strategies need to consider how to avoid inbreeding and how to ensure that each female can encounter a sufficient number of mates to fertilize all her eggs. The concept of positive density dependence also offers an opportunity for ecologists to help control undesirable pest species. For example, several pest insect populations have been controlled by releasing sterile males into the population. This skews the sex ratio and causes the females to breed with sterile males, leading to a lower growth rate in the pest population. In other cases, such as in pest control using pesticides, researchers simply reduce the size of an undesirable population to the point that individuals have a hard time finding mates.
The Logistic Growth Model
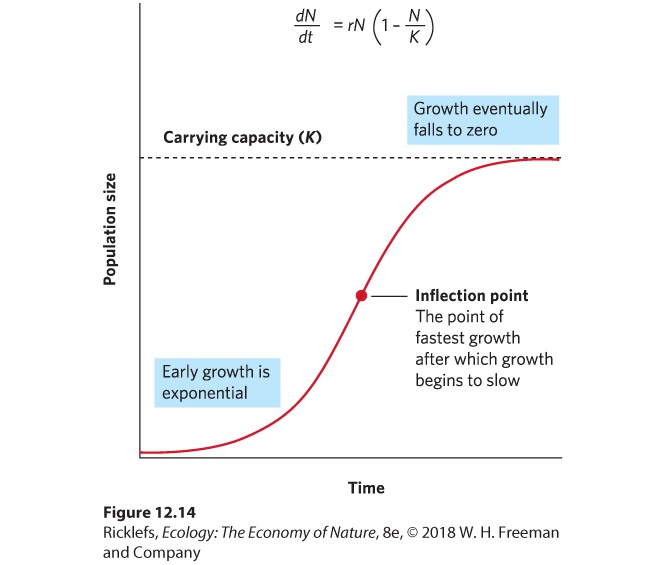
Although positive and negative density dependence both occur in nature, population modelers have focused more on the effects of negative density dependence. As a result, they have developed growth models that mimic the behavior of many natural populations: rapid initial growth followed by slower growth as populations grow toward their maximum size. The maximum population size that can be supported by the environment is called the carrying capacity of the population, denoted as K. Carrying capacity (K) The maximum population size that can be supported by the environment. To model the slowing growth of populations at high densities, we use the logistic growth model. The logistic growth model builds on the exponential growth model, but adds a term, (1 – N/K), that accounts for a decline in growth rate as the population approaches its carrying capacity: dNdt=rN(1-NK) Logistic growth model A growth model that describes slowing growth of populations at high densities.
When the number of individuals in the population is small relative to the carrying capacity, the fraction NK is close to 0 and the term inside the parentheses approaches 1. When this happens, the equation becomes nearly identical to the exponential growth model. However, when the number of individuals in the population approaches the carrying capacity, the term inside the parentheses approaches 0. As a result, the population’s rate of growth approaches 0. When we plot the growth of a population over time using the logistic growth curve, we obtain a sigmoidal, or S-shaped curve, which is shown in Figure 12.14. The middle point on the curve, where the population experiences its highest growth rate, is known as the inflection point. Above the inflection point, the population growth begins to slow.
Figure 12.14 The logistic growth model. When the population is small, growth is exponential. When the population is large, growth slows. The inflection point indicates the point at which the population experiences its fastest rate of growth. Above the inflection point, growth begins to slow. When the population reaches its carrying capacity, K, growth is zero. As a result of this growth pattern, the logistic growth model is represented by an S-shaped growth curve. S-shaped curve
The shape of the curve when a population is graphed over time using the logistic growth model. Inflection point The point on a sigmoidal growth curve at which the population achieves its highest growth rate. We can further understand how the logistic growth model behaves by examining the effect of population size on the rate of increase of the population (dNdt) . As shown in Figure 12.15a, as the population increases from a very small size, the rate of increase grows because the number of reproductive individuals increases. After reaching one-half of the carrying capacity, which corresponds to the inflection point of the S-shaped curve, the rate of increase begins to slow because the reproductive individuals are each obtaining fewer resources.
Figure 12.15 The effect of population size on the rate of increase and the per capita rate of increase. (a) As populations grow from a small number to their carrying capacity, the rate of population growth increases until it reaches the inflection point, after which it decreases. (b) On a per capita basis, the rate of increase continually declines. We can also examine how population size affects the rate of increase on a per capita basis, which is (1N)(dNdt) As you can see in Figure 12.15b, individuals in the population continually decline in their ability to contribute to the growth of the population. Therefore, the logistic growth curve shows an initial rapid increase in growth due to the increasing number of individuals in the population, followed by a
slowing rate of growth as per capita resources become limited. The logistic equation describes the growth of many populations. One of the classic studies was conducted by Russian biologist Georgyi Gause in 1934. Gause raised two species of protists, Paramecium aurelia and Paramecium caudatum, in test tubes and added a fixed amount of food each day. Both populations initially grew exponentially, and the growth then slowed until they reached carrying capacity. Because the two species differ in many ways, their populations stabilized at different carrying capacities, which are illustrated as blue lines in Figure 12.16. Gause suspected that the cause of this maximum population size was the amount of available food. To test this hypothesis, he did the experiment again, but this time doubled the amount of food. As you can see from the orange line in each graph, the two species once again experienced rapid initial growth followed by slower growth that stabilized. With twice as much food available, however, the two species stabilized at population sizes that were twice as large as in the first experiment. This early experiment confirmed that logistic growth can happen in real organisms and that the increased availability of a limiting resource can increase the carrying capacity of a population.
##### Predicting Human Population Growth with the Logistic Equation
Figure 12.16 Logistic growth in two species of Paramecium. (a) When Paramecium aurelia was raised under low or high food supply, the population size—measured in terms of the total volume of protist cells—grew rapidly at first and then slowed as they reached their carrying capacity. When twice as much food was provided, the species achieved a population size that was twice as large. (b) A similar pattern was found for another species of protist, Paramecium caudatum.
Logistic Equation
The logistic growth model was first developed in 1838 by the Belgian mathematician Pierre François Verhulst. Verhulst had read Thomas Malthus’s 1798 essay and sought to formulate a natural law governing the growth of populations. Nearly a century later, in 1920, Raymond Pearl and Lowell Reed independently confirmed the logistic growth model as it applies to human population growth.
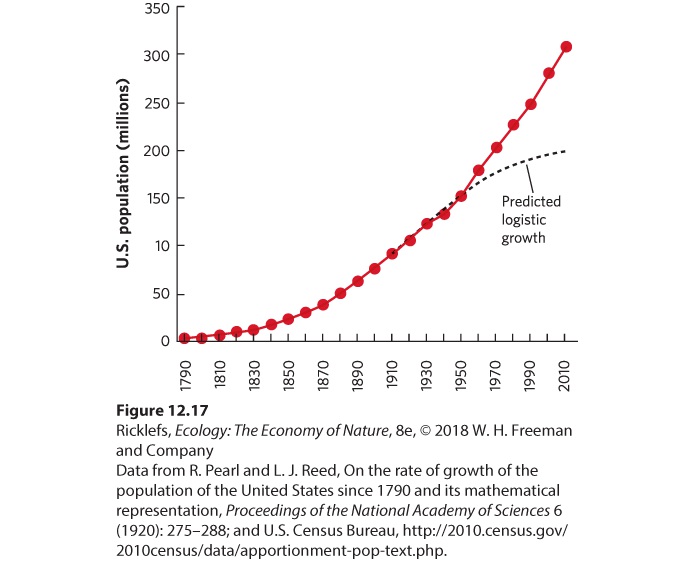
Using U.S. Census Bureau data that had been collected every 10 y Pearl and Reed confirmed the observation by Malthus that the population in the United States initially grew exponentially from 1790 to 1910. They noticed, however, that by 1910 this growth began to slow, as illustrated in Figure 12.17, and that the rate of growth appeared to be declining over time. Pearl and Reed applied their logistic growth equation to the census data and predicted that the population of the United States, which was 91 million in 1910, had a carrying capacity of 197 million. However, they were careful to note that this prediction depended on the population supporting itself using only the land of the United States.
Figure 12.17 The growing human population in the United States. A census conducted every 10 years from 1790 to 1910 indicated that the U.S. population was continuing to increase. However, the rate of increase started to decline by 1910. Based on these data, Pearl and Reed predicted that the population would reach a maximum of 197 million people. In reality, the U.S. population has continued to grow. As you can see in the figure, the U.S. population has greatly exceeded this prediction; by 2017, the population had grown to more than 325 million people. There are several reasons for this. One important reason is that technological advances have allowed farmers in the United States to produce
food much more efficiently. Moreover, a substantial amount of the food consumed by the U.S. population comes from other regions of the world. In addition to the increases in available food, we have had major improvements in public health and medical treatment that have increased survival rates substantially, particularly for infants and children. Finally, the logistic equation did not incorporate the millions of immigrants who came to the United States after 1910.
Concept Check
1. How do density independent effects alter population sizes? 2. What is the connection between negative density dependence and the self-thinning rule in plants? 3. Under what range of densities do we typically observe positive density dependence?
#### 12.3 Population Growth Rate Is Influenced by the Proportions of Individuals in Different Age, Size, and Life History Classes
12.3 Population growth rate is influenced by the proportions of individuals in different age, size, and life history classes In our discussion of population models so far, we have assumed that all individuals in the population have an identical intrinsic growth rate—that is, they have the same birth rates and death rates. While this assumption is helpful to generate simple models of population growth, we know that survival and fecundity vary with an individual’s age, size, and life history stage. In terms of age, individuals cannot reproduce until they have achieved reproductive maturity. In terms of individual size, individuals with greater mass typically have higher fecundity (see Chapter 8). In terms of life history stages, we know that different organisms experience different fecundity rates during each life stage. For example, tadpoles experience low survival and no fecundity, whereas later in life, as frogs, they experience high survival and high fecundity. Many perennial plants alternate between years in which they are reproductive and years in which they are not. In this section, we will examine how changes in survival and fecundity among different classes in a population affect population growth. In doing so, we will focus on age classes while recognizing that the same principles apply to size classes and life history classes.
Age Structure
The age structure of a population is the proportion of individuals that occurs in different age classes. A population’s age structure can tell us a great deal about its past growth and its potential for future growth. Consider the different patterns of age structure among human populations in 2017 that are displayed in Figure 12.18. In these figures, known as age structure pyramids, we see that the nations of India, the United States, and Germany all have declining numbers of people after 50 years of age, due to senescence. But each has a very different pyramid shape below 50 years of age. Age structure pyramids with broad bases reflect growing populations, pyramids with straight sides reflect stable populations, and pyramids with narrow bases reflect declining populations.
Figure 12.18 Age structure for human populations in India, the United States, and Germany in 2017. The age structure of India has a high proportion of young people, which indicates a growing population. The age structure of the United States has very similar numbers of people from 0 to 50 years of age, which indicates a stable population. Germany’s age structure has fewer young people than middle-aged people, an indication that its population is declining. Age structure In a population, the proportion of individuals that occurs in different age classes. In the case of India, for example, people in the younger age classes far outnumber those in the middle age classes. If the young age classes experience good nutrition and health care, they will survive well and the
number of reproductive individuals 2 decades from now will be much greater than it is today, which will cause the population to grow. We can contrast this with the age structure pyramid of the United States. The number of people in the younger age classes is very similar to those in the middle age classes. In this case, we would predict that 2 decades from now, we would have a similar number of reproductive individuals, so the population should remain relatively stable. The other extreme is represented by Germany. In Germany, the current reproductive age classes have experienced reduced reproduction such that the number of people in the younger age classes is less than the number in the middle age classes. If we project ahead 2 decades, we would predict that there will be fewer reproductive individuals in the future and, as a result, the population in Germany should decline.
Survivorship Curves
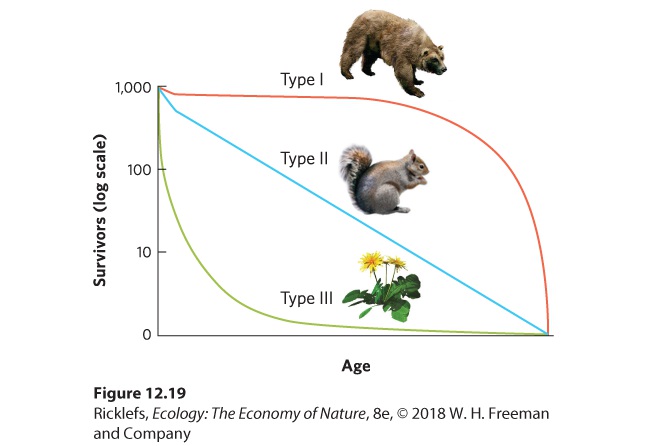
To understand the differences in survival among different age classes, we can graph survival over time, as illustrated in Figure 12.19. We can then categorize survivorship curves as type I, type II, or type III. A type I survivorship curve depicts a population with low mortality early in life and high mortality late in life. Examples include humans, elephants, and whales. A type II survivorship curve occurs when a population experiences relatively constant mortality throughout its life span. Organisms with type II curves include squirrels, corals, and some species of songbirds. A type III survivorship curve depicts a population with high mortality early in life and high survival later in life. This is a common pattern in many species of insects and in plants such as dandelions and oak trees that produce hundreds or thousands of seeds. In reality, most populations have survivorship curves that combine features of type I and type III curves, with high mortality early and late in life.
Figure 12.19 Survivorship curves. Individuals that have a type I curve experience high survival until later in life. Individuals with a type II curve experience a steady decline in survival throughout life. Individuals that have a type III curve experience low survival when young and high survival until later in life.
Life Tables
Because both birth and death rates can differ among age, size, and life history classes, we need a way to incorporate this information into our estimates of population growth rates. In this way, we can better predict how a population will change in abundance over time, which is important for managing populations that humans consume, populations of species that we wish to conserve, and populations of pests that we wish to control. We can achieve this goal if we know the proportions of individuals in different classes and the birth and death rates of each age class. Collectively, such data can tell us whether a population is expected to increase, decrease, or remain unchanged. To determine how age, size, or life history classes affect the growth of a population, we use life tables, which compile class-specific survival and fecundity data. Because it can be hard to ascertain paternity in many species, life tables are typically based on females and fecundity is defined as the number of female offspring per reproductive female. For some populations with highly skewed sex ratios or unusual mating systems (see Chapter 9), using only females can pose problems, but in most cases a female-based life table provides a useful model of population growth.
Life tables Tables that contain class-specific survival and fecundity data. To demonstrate the use of life tables, let’s consider a hypothetical population composed of 100 individuals distributed into different age classes, as illustrated in Table 12.1. In this table, the age classes are denoted by x and the number of individuals in each age class is denoted by nx. The value of nx represents the number of individuals present immediately after the population has produced offspring. In the table, survival rate from one age class to the next age class is denoted as sx. (You can think of “s” for survival.) In our example, the column labeled as sx indicates that the new offspring have a 50 percent survival rate, 1-year-olds have an 80 percent survival rate, 2-yearolds have a 50 percent survival rate, and no 3-year-olds survive to become 4year-olds. The fecundity of each age class is denoted by bx. (You can think of “b” for birth.) In the table, the column labeled as bx indicates that the new offspring cannot reproduce, but 1-year-olds can each produce one offspring, 2-year-olds can each produce three offspring, and 3-year-olds can each produce two offspring.
TABLE 12.1 A Hypothetical Population of 100 Individuals with Age-Specific Rates of Survival and Fecundity Age (x) Number of individuals (nx) Survival rate (sx) Fecundity (bx) 0.5 0.8 0.5 0.0 — — Total The data in this life table allow us to calculate the expected size of the population after one year. Table 12.2 shows us the calculations for the population in Table 12.1. To calculate the number of individuals that survive to the next age class, we multiply the number of individuals in an age class by the annual survival rate of that age class, as shown in the red-shaded region of the table. For example, if we start with 20-year-olds and their
survival rate is 50 percent, then we can calculate the number of 1-year-olds that we will have in the following year:
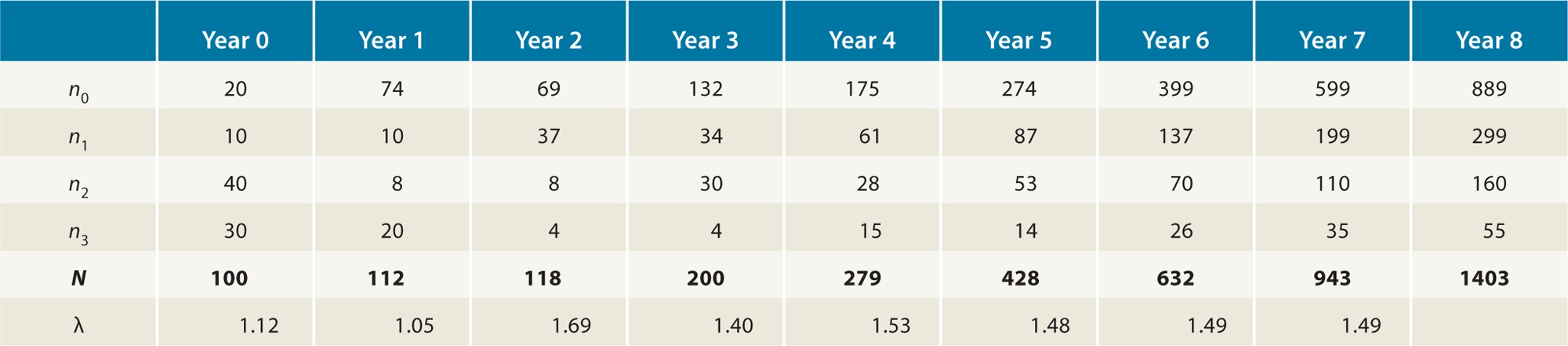
TABLE 12.2 The Number of Survivors and New Offspring After 1 Year for the Hypothetical Population in Table 12.1 To determine the number of new offspring that will be produced next year, we multiply the number of individuals that survive to the next age class—as shown in the orange-shaded region—by the fecundity of that age class. For example, if 10 individuals will survive to be 1-year-olds and each 1-year-old can produce one offspring, then we can calculate the number of new offspring that this age class will produce, as shown in the yellow-shaded region of the table: Number of new offspring produced=(nx)×(sx)×(bx)Number of new offspring produced=20×0.5×1Number of new offspring produced=10 If we do these calculations for every age class, we find that next year we will have a total of 74 new offspring. In addition, we will have 10 individuals that are now 1 year old, 24 individuals that are 2 years old, and 40 individuals that are now 3 years old. You can see a summary of this in Table 12.3.
TABLE 12.3 Population Size Initially and 1 Year Later Based on the Hypothetical Population in Table 12.1 Age (x) Number of individuals (nx) Number of individuals 1 year later (nx + 1)
Total N0 = 100 N1 = 112 When we add up all the age classes, we find that these survival and fecundity rates have caused the population to grow from a total size of 100 to a new total size of 112 individuals 1 year later. As a result, the geometric growth rate of the population is λ=N1N0=112100=1.12 We can continue these calculations to estimate the size of the population for many years into the future. Table 12.4, for example, shows the results of these same calculations for 8 years into the future. We can also continue to calculate the geometric rate of growth of the population for each year. If we examine these values of λ, we can see that they initially fluctuate widely between 1.05 and 1.69. As the years pass, however, the values of λ settle down to 1.49. Provided that the survival and fecundity of each class stay constant over time, λ will stabilize at a single value and the proportion of individuals in each age class also will stabilize. When the proportion of individuals in each age class does not change over time, we say it has a stable age distribution. However, if the survival and fecundity rates were to change, the value of λ and the proportion of individuals in each age class would be altered as well. Stable age distribution When the age structure of a population does not change over time. Calculating Survivorship We have examined life tables using the survival rate (sx) from one age class to the next age class. Another useful measure of survival is the probability of surviving from birth to any later age class, which we call survivorship and
denote as lx. (You can think of “l” for living.) Survivorship in the first age class is always set at 1 because all individuals in the population are initially alive. Survivorship at any given age class is the product of the prior year’s survivorship and the prior year’s survival rate. For example, survivorship to the second year is calculated as l2=l1s1 and survivorship to the third year is calculated as l3=l2s2 The calculated survivorships in our hypothetical population are shown in
TABLE 12.5 Calculating the Survivorship (lx) of the Hypothetical Population in Table 12.1 Age (x) Number of individuals (nx) Survival rate (sx) Survivorship (lx) 0.5 1.0 0.8 0.5 0.5 0.4 0.0 0.2 — 0.0 Total Calculating the Net Reproductive Rate The net reproductive rate is the total number of female offspring that we expect an average female to produce over the course of her life. If this value is greater than 1, then each female more than replaces herself in the population and the population will grow. The net reproductive rate, denoted as R0, is calculated in two steps. First, we multiply the probability of living to each age class (lx) by the fecundity of the respective age classes (bx). Second, we sum these products, as shown in the following equation: R0=∑ lxbx Net reproductive rate The total number of female offspring that we expect an average female to produce over
the course of her life. An example is shown in Table 12.6 for our hypothetical population. In this case, the net reproductive rate is 2.1, which means that each female is producing 2.1 daughters, on average, so the population will grow.
TABLE 12.6 Calculating the Net Reproductive Rate Based on the Hypothetical Population in Table 12.1 Age (x) Survival rate (sx) Survivorship (lx) Fecundity (bx) (lx) × (bx) 0.5 1.0 0.8 0.5 0.5 0.5 0.4 1.2 0.0 0.2 0.4 — 0.0 — 0.0 NET REPRODUCTIVE RATE (R0) = Σ lx bx = 2.1 Calculating the Generation Time We can also use life tables to calculate the generation time (T) of a population, which is the average time between the birth of an individual and the birth of its offspring. To calculate generation time, we must make three sets of calculations, as shown in Table 12.7. First, within each age class, we multiply the age (x), survivorship (lx), and fecundity (bx). Second, we take the sum of these products. This provides us with the expected number of births for a female, weighted by the ages at which she produced the offspring. Generation time (T) The average time between the birth of an individual and the birth of its offspring.
TABLE 12.7 Calculating the Generation Time, T, from the Hypothetical Population in Table 12.1 Age (x) Survival rate (sx) Survivorship (lx) Fecundity (bx) (lx) × (bx) (x) × (lx) × (bx) 0.5 1.0 0.0 0.8 0.5 0.5 0.5 0.5 0.4 1.2 2.4 0.0 0.2 0.4 1.2 — 0.0 — 0.0 0.0
(R0) = Σ lx bx = 2.1 Σ xlx bx = 4.1 Using these calculations, we can now calculate generation time as T=∑ xlxbxlxbxT=4.12.1T=1.95 years This tells us that in our hypothetical population, the average time between the birth of an individual and the birth of its offspring is about 2 years. Calculating the Intrinsic Rate of Increase We can make connections between life tables and our earlier population growth models by using life table data to estimate the population’s intrinsic rate of increase (λ or r). When an intrinsic rate of increase is estimated from a life table, we assume that the life table has a stable age distribution. However, stable age distributions rarely occur in nature because the environment varies from year to year in ways that can affect survival and fecundity. As a result, any approximation of λ or r is necessarily restricted to the set of environmental conditions that the population experiences. There are complicated equations to estimate λ and r. However, we can provide close approximations, denoted as λa and ra (where the letter “a” indicates an approximation), based on our estimates of net reproductive rate (R0) and generation time (T): λa=R01T which given our earlier calculations simplifies to λa=2.111.95λa=1.46 Note that this value of 1.46 is close to our observed λ of about 1.49 after the population achieved a stable age distribution (see Table 12.3). If we wanted to calculate ra, we could do so using the following equation:
##### Analyzing Ecology: Calculating Life Table Values
ra=logeR0Tra=loge2.11.95ra=0.38 You can see that a population’s intrinsic rate of increase depends on both the net reproductive rate (R0) and the generation time (T). The greater the net reproductive rate and the shorter the generation time, the higher will be the intrinsic rate of population increase. A population grows when R0 exceeds 1, which is the replacement level of reproduction for a population. In contrast, a population declines when R0 < 1. The rate at which a population can change increases with shorter generation times.
Collecting Data for Life Tables
To determine the age structure of a population and predict future population growth, we need to collect data or organisms from different ages. This objective can be met by constructing either a cohort life table or a static life table. A cohort life table follows a group of individuals born at the same time from birth to the death of the last individual. In contrast, a static life table quantifies the survival and fecundity of all individuals in a population during a single time interval. As we will see, the two types of life tables are used in different situations and have different advantages and disadvantages. Cohort life table A life table that follows a group of individuals born at the same time from birth to the death of the last individual. Static life table A life table that quantifies the survival and fecundity of all individuals in a population during a single time interval.
Analyzing Ecology
Calculating Life Table Values Life table data are collected for many different species. For example, a group of researchers spent a decade following a population of gray squirrels (Sciurus
carolinensis) in North Carolina to quantify the survival and fecundity of the squirrels. Below is a table of the age-specific survival and fecundity. YOUR TURN Using the data in the table, calculate survivorship (lx), net reproductive rate (R0), generation time (T), and the intrinsic rate of increase (λ).
Gray Squirrel Life Table
(x) (nx) (lx) (bx) (lx) × (bx) (x) × (lx) × (bx) 1.000 0.05 0.253 1.28 0.116 2.28 0.089 2.28 0.058 2.28 0.039 2.28 0.025 2.28 0.022 2.28 SOURCE: Data from F. S. Barkalow, The vital statistics of an unexploited gray squirrel population, Journal of Wildlife Management 34 (1970): 489–500. Cohort Life Tables Cohort life tables are readily applied to populations of plants and sessile animals in which marked individuals can be continually tracked over the course of their entire life. The cohort life table does not work well for species that are highly mobile or for species with very long life spans, such as trees. One of the problems in using a cohort life table is that a change in the environment during one year can affect survival and fecundity of the cohort that year. This makes it difficult to disentangle the effects of age from the effects of changing environmental conditions. For example, imagine that you mark a population of plants that lives for several years. However, in the middle of this life span, the population experiences a severe drought that causes sharp reductions in survival and fecundity. The following year is wet, which causes high survival and fecundity. When you construct the life table,
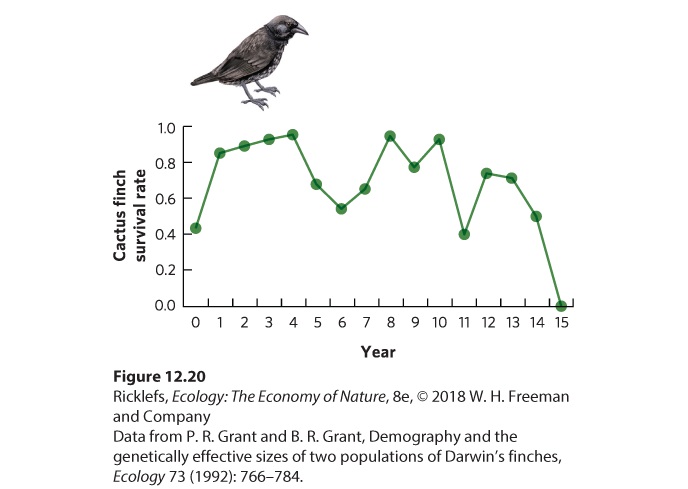
you cannot determine whether the age-specific changes in survival and fecundity are due to a change in age or a change in the environment. An excellent example of a cohort life table comes from Peter and Rosemary Grant, who have studied several species of ground finches in the Galápagos Islands off the coast of Ecuador. On the small island of Daphne Major, the Grants were able to capture all the birds on the island and mark them with uniquely colored plastic leg bands. Because this island is so isolated, few birds left the island or arrived from elsewhere. Using 210 cactus finches (Geospiza scandens) that fledged in 1978, the researchers constructed a cohort life table for the population. In Figure 12.20, you can see the annual survival rates for these birds. As is the case for many species, survival rate was low in the population’s first year and then remained high for several subsequent years. However, survival was quite variable throughout the life of the birds. This variation in survival reflects variation in the environment due to climatic changes during El Niño years (see Chapter 5). Because El Niño years are wet years, the vegetation produces abundant food for the finches and this causes high survival. Following El Niño years are periods of several dry years. During this time, food becomes scarce for the finches and this causes low survival. These data emphasize a disadvantage of cohort life tables; it is difficult to determine whether low survival at a particular age is due to the age of the individuals or due to the environmental conditions that occur during that year.
Figure 12.20 Annual survival rate of cactus finches. Using a cohort of cactus finches, researchers were able to calculate the annual survival over a 15-year period. Static Life Tables Using a static life table avoids many of the problems of the cohort life table. By considering the survival and fecundity of individuals of all ages during a single time interval, differences among the age classes are quantified under the same environmental conditions, so age is not confounded with time. In addition, static life tables allow us to look at the survival and fecundity of all individuals during a snapshot in time, which means we can examine species that are highly mobile as well as species that have long life spans. To construct a static life table, you must be able to assign ages to all individuals since they are not marked at birth. A number of techniques are available to do this. For example, counting annual growth rings in the trunks of trees, in the teeth of mammals, and in the ear bones of fish. The one major concern when using static life tables is that the age-specific data on survival and fecundity only apply to the environmental conditions that existed at the time the data were collected. Because the life table may not be representative of years in which the environmental conditions are quite different, it is helpful to construct static life tables for multiple years to assess how much environmental variation affects the predicted population growth. A classic study using a static life table was conducted by Olaus Murie,
who examined the survival of Dall mountain sheep (Ovis dalli) at a site in Alaska during the 1930s. Murie knew that the horns of the sheep contained annual growth rings that could be used to age the sheep. He also knew that following a cohort of highly mobile sheep for 15 years in Alaska was not feasible, so he took a static approach by conducting a search of all sheep skeletons in the area. He found a total of 608 skeletons from sheep that had died in recent years and assigned an age to each individual based on its horns. Using these estimates, he determined how many of the original 608 sheep died in each age class. His results, depicted in Figure 12.21, show a low survival rate during the first year, followed by high survival for the next 7 years. After 7 years of age, the annual survival rate began to drop, with the exception of those at 12 years of age. However, only four animals made it to 12 years, which gives us a poor estimate of the typical survival rate for 12year-old sheep. Because these data were taken from the skeletons of sheep that presumably died over a relatively short period, we do not see the large variation in survival rate that we saw in the cohort life table of the cactus finch.
Figure 12.21 A static life table and survival rate curve for Dall sheep. Using the ages of Dall sheep skeletons found in Alaska, researchers calculated the survival and survivorship of different age classes. When graphed, these data show that the survival rate of the sheep remains quite high for the first 7 years and then rapidly declines. There were only four animals in the 12-year age class, and this resulted in an unreliable high estimate of survival rate. Today, researchers continue to examine life tables for species, and some
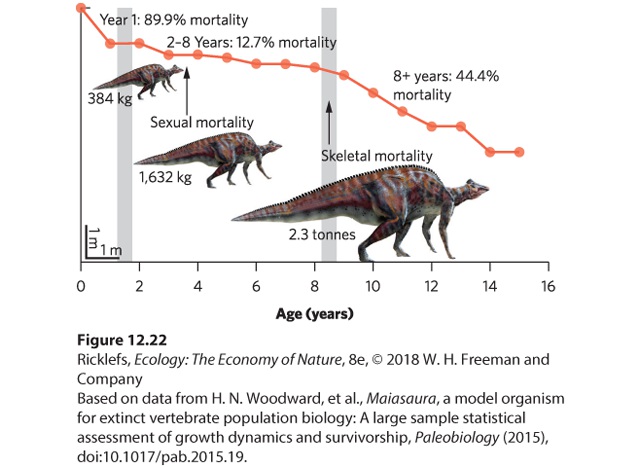
data come from unexpected sources. For example, for decades, paleontologists have been excavating fossil bones of dinosaurs in Montana, including the tibiae, or shin bones, of a single species known as the “good mother lizard” (Maiasaura peeblesorum). Finding the fossilized bones of a single individual of any dinosaur species is an unusual event, but these researchers found an amazing 50 fossils of a single species. They also recognized that they could age the fossil bones—based on the size and structures found in the tibiae—and this would provide data for a static life table. In 2015, they published their data as a survival curve, which is shown in Figure 12.22. Resembling a type I survivorship curve, the youngest age class individuals had high death rates, the middle-aged individuals had high survival rates, and the oldest age classes had high death rates.
Figure 12.22 A survival rate curve for the “good mother dinosaur.” Based on the tibia sizes and characteristics of 50 fossils, research estimated how many individuals were in each age class. From these static life table data, they produced a survivorship curve for the dinosaur. Throughout this chapter, we have examined the mathematical models that mimic natural patterns of population growth. Although populations commonly grow rapidly when at low densities, they become limited as the populations grow larger. The simplest population models are helpful starting
points, but they are not sufficient for most species that have survival and fecundity rates that vary with age, size, or life history stage. Using life tables helps to incorporate this complexity. As we will see below in “Ecology Today: Applying the Concepts,” the analysis of life tables can be very helpful in setting priorities for management strategies to save species from extinction.
Concept Check
1. Why might a model that contains age structure better reflect the reality of population growth than a model that lacks age structure? 2. What are the patterns of survival with age in the three types of survivorship curves? 3. How would you describe the generation time of a population?
Concepts
Saving the Sea Turtles Under the cover of darkness, the swimmers come to shore. After spending a year out in the ocean, female sea turtles crawl up the shore and dig a nest to lay their eggs. After about 2 months, these eggs will hatch and the hatchlings will scramble toward the ocean, where they will face a gauntlet of predators. If they survive, they will need 2 or 3 decades to become reproductively mature. They may live for 80 years. Not long ago, many more females came to shore. Of the
six species of sea turtles, all have declined over the past several centuries. For example, scientists estimate that there were once 91 million green turtles (Chelonia mydas) in the Caribbean; today, there are about 300,000, which is a decline of more than 99 percent. Sea turtles, which live in temperate and tropical waters, are declining worldwide. Historically, people hunted them for food and used turtle shells for decorations; more recently, land development has caused the loss of beach habitat for egg laying. Commercial fishing operations accidentally catch and kill turtles. Across all six species, more than 300,000 were caught along the U.S. coast in the 1980s, and more than 70,000 of these died before being returned to water. About 98 percent of these accidental catches were from shrimp trawlers pulling large nets through the water. Attempts to reverse the decline of the sea turtles have been ongoing for many years. One attempt was to protect nesting areas and to incubate large numbers of eggs artificially. Then hatchlings were released into the oceans to improve their probability of survival. But after spending a great deal of time and money on this effort, population modelers had an interesting insight. When they began constructing life tables for turtles—a real challenge, given that the turtles spend nearly all their life at sea—they discovered that improving the survival of the hatchlings would have little positive impact on the growth of the population. Because so few hatchlings survive to adulthood naturally, adding even thousands more would have only a small positive impact on the turtle population. Rather, the life tables indicated that improving the survivorship of the adult turtles was the key to growing the population.
Sea turtles. Population models have identified the stages in life that need the most protection to allow turtle populations to recover. These models are relevant to all species of sea turtles, including this green sea turtle (Chelonia mydas) off the coast of Hawaii. This insight completely changed the emphasis of sea turtle conservation. The focus shifted to the commercial fishing operations that were accidentally killing adult sea turtles. Pressure mounted for commercial shrimp fishermen to install turtle excluder devices on their trawling nets. These devices work like a sieve; small shrimp pass through the sieve and get captured in the net, while sea turtles, which are too big to pass through the sieve, are shunted out of the net. The installation of these devices has been a tremendous success. In 2011, researchers reported that the number of turtles accidentally netted had been reduced by 60 percent and the number of turtles killed had declined by 94 percent. Turtle researchers continue to explore ways to reduce accidental captures in fishing nets. In 2016, researchers reported an experiment in which they hung inexpensive green lights on 500-m fishing nets to see if it would make
the nets more visible to turtles so they would veer away. They deployed pairs of nets. Within each pair, one net had green lights attached, while the other did not. After several trials, the researchers found that nets with the lights caught the same number of fish, but half as many sea turtles. Given this encouraging result, their follow-up research will examine different colors of lights to see when colors are the most effective at deterring the turtles. Scientists continue to work on obtaining age-specific survival rates on the six species of sea turtles, which is a challenge since the animals can migrate thousands of miles and males do not return to nesting beaches. Indeed, the most recent population models indicate that improving the survival of the hatchlings does have a small positive effect on the population, although improving the survival of older animals continues to have a much greater effect. The story of the sea turtle makes it clear that being able to model population growth can be a critical step in bringing back species from the brink of extinction. SOURCES: Ortiz, N., et al. 2016. Reducing green turtle by-catch in small-scale fisheries using illuminated gillnets: The cost of saving a sea turtle. Marine Ecology Progress Series 545: 251-259. Finkbeiner, E. M., et al. 2011. Cumulative estimates of sea turtle bycatch and mortality in USA fisheries between 1990 and 2007. Biological Conservation 144: 2719–2727. Marzaris, A. D., et al. 2006. An individual based model of a sea turtle population to analyze effects of age dependent mortality. Ecological Modelling 198: 174–182.
Summary of Chapter Concepts
12.1 Populations can grow rapidly under ideal conditions. When populations grow at their intrinsic growth rate, they can initially increase at exponential rates, which can be modeled using either the exponential growth model or the geometric growth model. Key Terms: Demography, Growth rate, Intrinsic growth rate (r), Exponential growth model, J-shaped curve, Geometric growth model, Doubling time
12.2 Populations have growth limits. The limits can be due to density-independent factors, which regulate population sizes regardless of the population’s density. The limits can also be due to density-dependent factors, which affect population growth in a way that is related to the population’s density. Negative density dependence causes populations to grow more slowly as they become larger, whereas positive density dependence causes populations to grow faster as they become larger. Ecologists use the logistic growth model to demonstrate negative density dependence. The logistic model mimics rapid population growth when populations are small and slow population growth when populations approach their carrying capacity. The logistic growth model has been used to predict human population growth, but human populations have exceeded these predictions due to improvements in food production, international trade, and public health. Key Terms: Density independent, Density dependent, Negative density dependence, Positive density dependence, Self-thinning curve, Carrying capacity (K), Logistic growth model, S-shaped curve, Inflection point
12.3 Population growth rate is influenced by the proportions of individuals in different age, size, and life history classes. Most organisms have rates of survival and fecundity that change over their
lifetime, as illustrated by survivorship curves. Life tables were developed to incorporate age-, size-, or life history-specific rates of survival and fecundity. Using life tables, we can determine survivorship (lx), net reproductive rates (R0), generation times (T), and approximations of the intrinsic growth rates (ra and λa). The data needed for life tables can be collected by following a cohort and building a cohort life table or by examining all individuals during a snapshot in time and developing static life tables. Key Terms: Age structure, Life tables, Stable age distribution, Net reproductive rate, Generation time (T), Cohort life table, Static life table
Critical Thinking Questions
1. Compare and contrast the approaches of the geometric growth model and the exponential growth model. 2. Given the relationship between λ and r in the geometric and exponential growth equations, can you demonstrate mathematically why λ must be 1 when r is 0? 3. Given that white-tailed deer give birth to fawns each spring, which population growth model would be the most appropriate and why? 4. Contrast the concepts of a stable population versus a stable age distribution. 5. If a life table projects a population size of 100 females and the sex ratio of the population is one-to-one, how large is the entire population? 6. In a life table, what is the fundamental difference between survival rate (sx) and survivorship (lx) in words and in terms of calculations? 7. Compare and contrast a cohort life table and a static life table. 8. What is the relationship between generation time and the rate of population growth? 9. When using the logistic population growth model, what are the different causes of slow population growth at low population sizes versus high population sizes? 10. What evidence would you need to determine whether a population experiences negative density dependence or positive density dependence?
Graphing the Data Survivorship Curves
Survivorship data are collected for a wide variety of organisms. At a U.S. research site in the state of Georgia, researchers examined the survivorship of two plants: elf orpine (Sedum smallii) and oneflower stitchwort (Minuartia uniflora). For the following survivorship data, plot the survivorship curves using a log-scale y axis.
Survivorship Data in Two Species of Plants
Month Elf orpine Oneflower stitchwort 1.00 1.00 0.81 0.37 0.09 0.05 0.03 0.02 0.02 0.01 0.01 0.01 0.00 0.00 Source: Data estimated from R. R. Sharitz and J. F. McCormick, Population dynamics of two competing annual plant species, Ecology 54 (1973): 723–740.