18 Community Structure
18Community Structure Cacao trees produce fruits that contain the cocoa beans. When old fruit husks are placed around the trees, they provide a habitat for the eggs and larvae of midges, which later turn into adult flies that pollinate the flowers of the cacao tree. Pollinating the “Food of the Gods” Chocolate is one of the most popular food ingredients in modern cuisines. It comes from the beans of the cacao tree (Theobroma cacao) that originated in South and Central America. Given that ancient people used it in religious ceremonies as gifts to their gods, the genus of the plants, Theobroma, translates to “the food of the gods.” Today, the cacao tree is grown in tropical regions around the world, including the Americas, Africa, and Asia. However, there is a widespread problem; only about 10% of cacao flowers get pollinated on modern farms. This results in fewer cocoa beans produced on each plant and less cocoa production for farmers and consumers. As a result, farmers have to hand-pollinate the flowers to improve their

crop yield. Farmers and ecologists have wondered why cacao plants experience such low pollination success. Modern plantations keep the ground cleared around cacao trees so that there are no decomposing old fruit husks. These decomposing husks can attract a fungus that can be harmful to the cacao tree, so removing the old pods reduces the fungus. However, the decomposing plant matter is also a habitat for the larval stage of the main pollinator of cacao trees, which are small flies known as midges. By cleaning up the ground around cacao trees, farmers have reduced the problem of the harmful fungus, but they may have also reduced the abundance of the pollinators. To test this hypothesis, researchers in Australia conducted an experiment in which they manipulated the presence or absence of old cacao fruit husks around groups of eight cacao trees. Within each group of eight trees, they also manipulated the presence or absence of hand pollination. Over a 7-month period, the researchers documented the number of flowers produced, the number of fruits produced, and the mass of fruits produced on each tree. In addition, they documented the presence of any predatory spiders or skinks (a type of lizard) that might be attracted to the insects living in the decomposing husks. In 2017, the researchers reported that the addition of old husks resulted in cacao trees producing more flowers, likely as a result of the decomposing husks providing more nutrients or water to the trees. After adjusting for this difference in total flower number, they discovered that adding old husks dramatically increased the number of flowers that were pollinated by the midges. When they examined the fruits at maturity, they found that the increase in the number of flowers pollinated by adding old husks was nearly as much as the increase observed when they used hand pollination. The total mass of the fruit on each tree followed a similar pattern. In addition, they found that adding the husks attracted more of the predatory spiders and skinks, but these predators did not appear to harm the midges. Collectively, these results demonstrate that decomposing fruit husks of the cacao tree provide essential habitat for the eggs and larvae of the midge and other insects, which in turn improves the pollination of
cacao flowers and provides insect prey for multiple species of predators. Because these decomposing husks can also serve as a habitat for fungal pathogens that are harmful to cacao trees, the researchers suggest that other sources of decomposing plant matter, such as old banana stems, might be a better way of providing midge habitat while reducing the prevalence of the fungal pathogen. This research illuminates how ecological communities are made up of interconnected species and that these connections have a major impact on natural and agricultural communities. “By cleaning up the ground around cacao trees, farmers have reduced the problem of the harmful fungus but they may have also reduced the abundance of the pollinators.”
Forbes, J. F., and T. D. Northfield. 2017. Increased pollinator habitat enhances cacao fruit set and predator conservation. Ecological Applications 27: 887–899.
Learning Objectives
After reading this chapter, you should be able to:
18.1 Illustrate how communities can have distinct or gradual boundaries.
18.2 Explain why the diversity of a community incorporates both the number and relative abundance of species.
18.3 Describe the ways in which species diversity is affected by resource availability, habitat diversity, keystone species, and disturbances.
18.4 Explain how communities are organized into food webs.

18.5 Describe how communities respond to disturbances with resistance, resilience, or switching among alternative stable states. In Chapter 1, we defined a community as an assemblage of species living together in a particular area. We also discussed how ecologists working at the community level focus on a multitude of species interactions, and how these interactions affect the number of species and the relative population size of each species. In Chapters 14 through 17, we discussed the major types of species interactions. We are now ready to expand our understanding of communities in which these interactions take place. In this first of two chapters on ecological communities, we will examine community structure, which includes the composition of species in a community, the relative abundance of each species, and the relationships among the species. We will start by looking at a classic debate over whether or not communities are distinct entities. We will then examine how ecologists quantify patterns of species diversity in communities based on the number of species and the relative abundance of each species in the community. All these species function as producers and consumers in a food web, which helps to determine the number of species, the relative abundance of each species, and the stability of the community.
#### 18.1 Communities Can Have Distinct or Gradual Boundaries
18.1 Communities can have distinct or gradual boundaries Our first step is to consider how communities change across the landscape and how they are categorized. As we will see, because many species move between communities, drawing boundaries around a community can be difficult. For example, many birds migrate every spring and fall, and amphibians spend their larval life in aquatic communities and their adult life in terrestrial communities. In this section, we will discuss community boundaries and investigate how communities are affected when community boundaries are either distinct or gradual.
Community Zonation
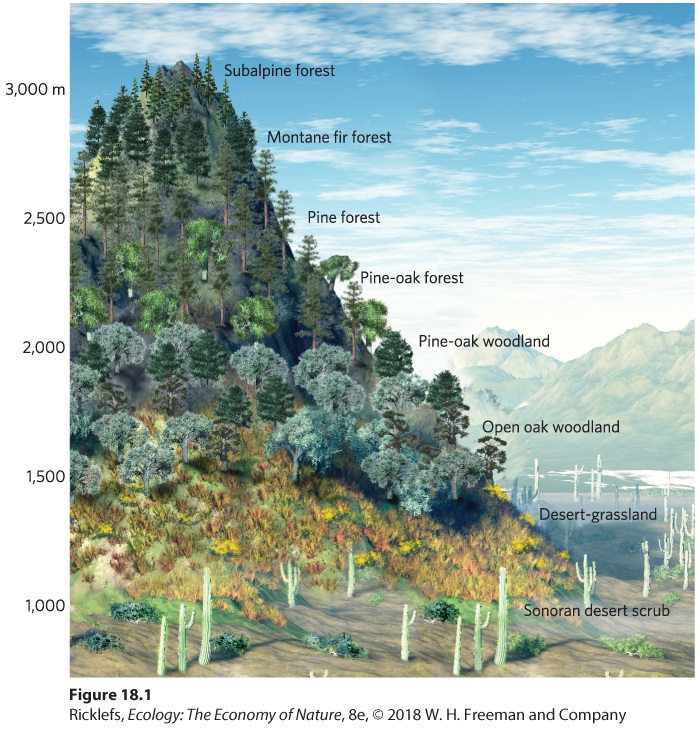
One of the most noticeable features of communities is that the species composition of the community changes as one moves across the landscape. With the change in environmental conditions, some species become better able to survive and compete. For example, if you were to walk from the base to the top of a mountain in the Santa Catalina Mountains in Arizona, you would observe striking changes in the vegetation, as illustrated in Figure 18.1. At the base of the mountains, you would first encounter desert-adapted plants, including rabbit bush (Franseria deltoidea) and creosote bush. As you ascend the mountain, grasses, some shrubs such as narrowleaf goldenbush (Haplopappus laricifolius), and a few scattered oaks predominate. Approaching the top of the mountain, you reach a forest of ponderosa pines and southwestern white pine (Pinus strobiformis), followed by a forest containing Engelmann spruce (Picea engelmanni), white fir (Abies concolor), and subalpine fir. The zones in which each species flourishes reflect different tolerance ranges for temperature and moisture availability, as well as different abilities to compete with other species for resources. There are similar changes in the animal species that live at different elevations on the mountains. The changes in plants and animals at different elevations create continuous changes in community composition from the base of the mountains to the highest peaks.
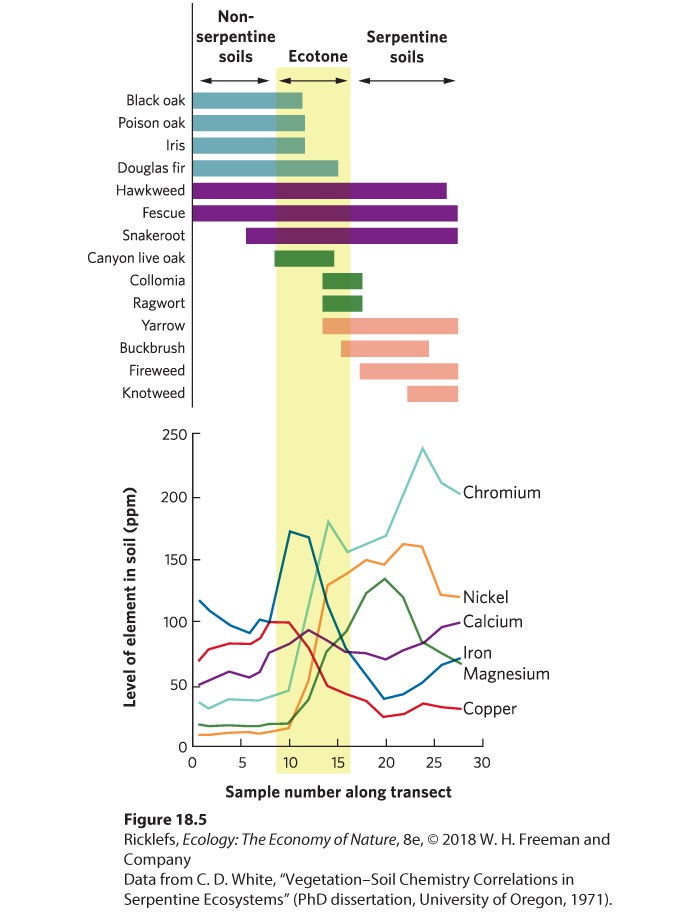
Figure 18.1 Zones in terrestrial communities. As one climbs the Santa Catalina Mountains in Arizona, the vegetation changes from desert shrubs to large trees. Zonation also occurs in aquatic communities, as depicted in Figure 18.2. We saw an example of this in Chapter 16 when we discussed competition between species of barnacles on rocky coasts of Great Britain. Due to both competition for space and the need to resist desiccation, stellate barnacles live in the upper intertidal zone, whereas the rock barnacles live in the lower intertidal zone. As another example, we find kelp forests below the intertidal zone, and a variety of seaweeds, mussels, anemones, rock barnacles, and hermit crabs occupy the lower and middle intertidal zones. In the upper intertidal zone, we find limpets, which are a type of gastropod, and stellate barnacles. Higher up on the shore, an area known as the splash zone is home to more limpets and a type of snail known as the periwinkle (Littorina littorea). The distribution of species in the different zones of the coastline reflects a combination of tolerance to changing abiotic conditions and the
outcome of species interactions that include competition, predation, and herbivory.
Figure 18.2 Zonation of communities along ocean shorelines. As one moves from deep to shallow water in the rocky shoreline of northeastern England, the species composition of the communities changes dramatically.
Categorizing Communities
We characterize communities as we characterize biomes—either by their dominant organisms or by the physical conditions that affect the distribution of species. In North America, for example, we might examine a beech-maple forest community in Ontario, a pine savanna community in Virginia, or a sagebrush community in Wyoming. Each of these communities is named for the dominant plants that are present. In aquatic systems, we focus on physical characteristics—such as a stream community, lake community, or wetland community—or on the dominant group of organisms in that system, such as a coral reef community. Because it is not realistic to study every species in the community, which could number in the hundreds or thousands, we typically focus on a subset of species that live in an area. For example, a study of a wetland community might focus only on bacteria, algae, snails, crustaceans, insects, amphibians, or fish. Several of these groups might be studied simultaneously.
Ecotones
When we consider the zonation of communities, it is clear that some have distinct boundaries that are either natural or constructed by humans. For example, the natural boundary between a lake community and a forest community can be clearly drawn at the water’s edge. This boundary exists because there is a sharp change in the environmental conditions as we leave one community and enter the adjacent community. Similar sharp changes in environmental conditions occur, for example, when there is an abrupt change in the type of soil due to the underlying geology of an area or when an individual moves from north-facing to south-facing slopes of mountains that have large differences in temperatures and moisture. Humans can also create distinct community boundaries. For example, an area of forest that has been cleared for agriculture creates a clear boundary between the field community and the forest community. Sharp changes in environmental conditions over a relatively short distance, accompanied by a major change in the composition of species, creates a boundary known as an ecotone (Figure 18.3). Although some species move between the adjacent communities that come together to form the ecotone, most species live in one of the communities and spread into the ecotone. As a result, ecotones typically support a large number of species, including those from each of the adjoining habitats and species adapted to the ecotone’s special conditions.
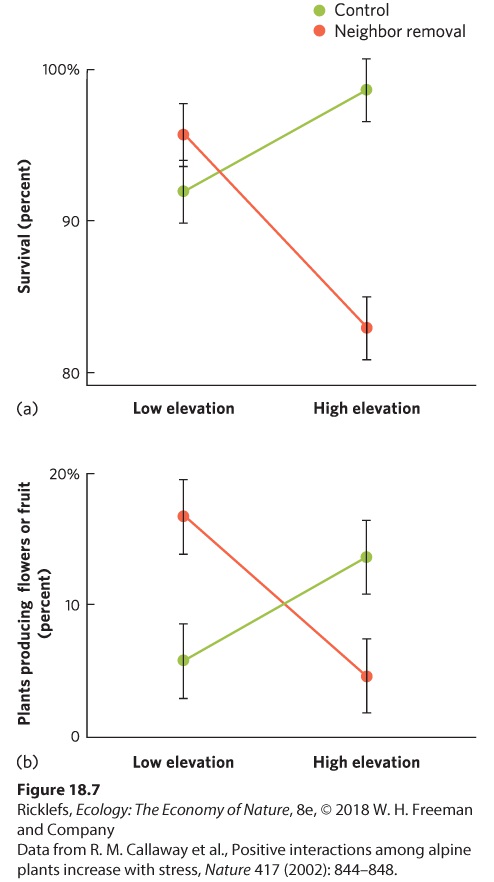
Figure 18.3 Ecotones. Ecotones are regions where two communities come together with a relatively sharp boundary, indicated by a rapid replacement of species. This ecotone, in Greater Sudbury, Ontario, Canada, occurs where a forest and a field come together. Ecotone A boundary created by sharp changes in environmental conditions over a relatively short distance, accompanied by a major change in the composition of species. One way to document the existence of an ecotone is to use a line-transect survey (see Chapter 11) to determine the abundances of different species along an environmental gradient. When an ecotone is present, we expect to observe sharp changes in the distributions of most species as we leave one community and enter the adjacent community. An example can be found in plant communities on serpentine soils. These soils are derived from underlying rock that contains heavy metals, including nickel, chromium, and magnesium. These heavy metals are toxic to many plants, and the soils are also typically low in nutrients such as nitrogen and phosphorus. Serpentine soils exist in small patches across the landscape in many different parts of the world. Because of the harsh conditions, most plant species cannot live in serpentine soils. Yet some species of plants have evolved an ability to tolerate them and a small portion of the species from adjacent communities also inhabit the ecotone (Figure 18.4).
Figure 18.4 Serpentine soils. In areas where serpentine rocks emerge to the soil surface, the soils contain low concentrations of nutrients and high concentrations of metals such as nickel, chromium, and magnesium. At this site in Northern California, the forest abruptly ends where the serpentine soils begin because the trees are not able to survive in the serpentine soils. In one study in southern Oregon, researchers conducted a line-transect survey that stretched from nonserpentine soils to serpentine soils. They measured the concentrations of several metals and quantified the presence of numerous plant species. Concentrations of chromium, nickel, and magnesium all increased in the shift from nonserpentine soils to serpentine soils, as you can see in the bottom half of Figure 18.5. The top of Figure 18.5 shows that species such as black oak (Quercus kelloggii) and poison oak (Rhus diversiloba) did not grow on the serpentine soils, whereas canyon live oak (Quercus chrysolepis) and ragwort (Senecio integerrimus) were found almost entirely in the ecotone where the two soils come together, and species such as fireweed (Epilobium minutum) and knotweed (Polygonum douglasii) were found only in serpentine soils. These species show distinct boundaries as one moves across the gradient of nonserpentine to serpentine soils. As you can also see in the transect data, a few species such as hawkweed (Hieracium albiflorum) and fescue (Festuca californica) were found across the entire
gradient. From these data, we can conclude that while some species are largely restricted to either serpentine or nonserpentine soils, the highest number of species occurs within the ecotone.
Figure 18.5 A line transect of serpentine and nonserpentine soils. As one moves along the transect, the concentration of metals in the soil exhibits a sharp increase midway along the transect. The presence of plant species along the gradient, as indicated by the bars, demonstrates that some species primarily live in nonserpentine soils (shown in blue), a few species primarily live in the ecotone (shown in green), and several species primarily live in the serpentine soils (shown in pink). Three of the species are able to live across the entire transect (shown in purple).
##### Communities with Interdependent versus Independent Species Distributions
Independent Species Distributions
As we have noted, a community is often described by the dominant species that live within it. Although we know that species interact with each other, for many years ecologists questioned whether species in a community are found together because they depended on each other or simply because they have similar habitat needs. Interdependent communities are those in which species depend on each other to exist. In the early twentieth century, plant ecologist Frederic Clements proposed that most communities function as interdependent communities and that they act as superorganisms. He compared individual species to the different parts of an organism’s body, all of which require each other for survival. Interdependent communities Communities in which species depend on each other to exist. Independent communities are those in which the species do not depend on each other to exist. Independent communities are composed of species that live in the same place because they have similar adaptations and habitat requirements. Independent communities Communities in which species do not depend on each other to exist. Although each species has a somewhat different range of conditions under which it can live, both interdependent and dependent communities reflect the overlapping ranges of the species that exist within it. In other words, similar habitat requirements happen to put various species in the same place at the same time. Plant ecologist Henry Gleason rejected Clements’s superorganism metaphor and proposed that most communities consist of species with independent distributions. Differentiating Between Interdependent and Independent Species Distributions How can we determine whether a community is made up of an interdependent or an independent group of species? One approach has been
the use of line-transect studies. If species are interdependent, we should observe suites of species appearing together and disappearing together as we move along a line transect. If species distributions are independent, we should be able to observe gradual changes in species composition as we move along a line transect. Each species will appear and disappear at different points along the line because of the unique habitat requirements of each species. A classic study using this approach was conducted during the 1950s by Robert Whittaker who surveyed the distributions of plant species in the Great Smoky Mountains on the border between Tennessee and North Carolina (Figure 18.6a). At different elevations that varied in temperature and moisture, he calculated the number of stems present for each plant species. As you can see in Figure 18.6b, different species appeared and disappeared at different points along the moisture gradient, and each species achieved its peak abundance at different points. This study and other studies by Whittaker provided strong evidence that when environmental conditions change gradually along a gradient, species in the community commonly show gradual changes in abundance that are independent of each other.
Figure 18.6 Independent species distribution. (a) The Great Smoky Mountains exhibit change in plant species composition with changes in elevation and soil moisture. (b) When a tree survey was conducted across a moisture gradient within an elevation of 1,067 to 1,372 m, researchers found that each species has its greatest abundance at different points along the moisture gradient. Moreover, each species appears and disappears at different points along the moisture gradient. This led researchers to reject the hypothesis that plant communities are distinct entities in which the existence of one species depends on the presence of one or more other species. Observing species distributions along environmental gradients is one way to test the independence of species distributions. However, even when we see abrupt changes in species distributions, we cannot be sure whether species in
a community are independent or interdependent. An abrupt transition may reflect an abrupt change in abiotic conditions whereby many species simply cannot exist under the changed conditions. For example, fish and other aquatic organisms are not restricted to live in a lake because they are necessarily interdependent; they are restricted to the lake because they lack the adaptations to live on land. Interdependency can be determined by monitoring the results of removing one or more species from the community. If species rely on each other to persist, then removing a species from a community should cause other species in the community to decline. If they don’t need each other to persist, then removing a species should leave other species in the community unaffected or even improve their fitness if they happen to compete with the species that is removed. Interdependent Species Under Harsh Environmental Conditions Recent studies have found that some species do rely on each other to persist, particularly in communities that experience harsh environmental conditions, such as extreme high or low temperatures or low moisture. For example, a large experiment conducted in 11 locations around the world examined how 115 plant species growing at either low or high elevation in alpine tundra biomes responded when a neighboring species was removed. The researchers measured the percentage of plants that survived as well as the percentage of plants that produced flowers. Removing neighboring plants at low-elevation sites caused an increase in the survival of the remaining plants, as shown in
Figure 18.7a. This also caused an increase in the percentage of plants that produced flowers or fruit, which you can see in Figure 18.7b. In contrast, removing neighboring plants at high-elevation sites caused a decrease both in the survival of the remaining plants and in the percentage of plants that produced flowers. Plants living under the harsher conditions of high elevations were helped by neighboring species because those species reduced the harsh winds, provided shade, or offered protection from herbivores. Such experiments teach us that while most communities appear to be composed of species with independent distributions, species living under harsh environmental conditions frequently depend on each other.
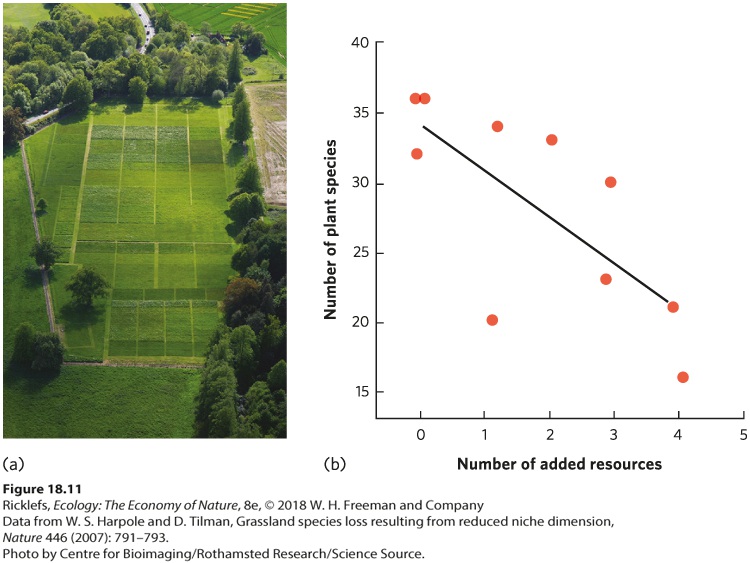
Figure 18.7 Interdependence of species under extreme environmental conditions. In alpine tundra biomes around the world, researchers measured how plants at low and high elevations responded to having neighboring plants removed in terms of (a) survival and (b) the percentage of plants that produced flowers. At low elevations, the removal of neighbors caused plants to survive better and produce more flowers or fruit, which suggests that the plants are competing. At high elevations, removal of neighbors caused the plants to experience lower survival and produce fewer flowers, which suggests the plants in the community facilitate each other. Error bars are standard errors.
Concept Check
1. Why do we often observe more species living at an ecotone, than within either of the adjacent communities? 2. What are the underlying causes of community zonation of
species distributions? 3. How do we determine whether the composition of species in a community is interdependent or independent?
#### 18.2 Community Diversity Incorporates Both the Number and Relative Abundance of Species
18.2 Community diversity incorporates both the number and relative abundance of species To understand the processes that influence the structure and functioning of communities, we need to quantify how communities differ from place to place. In this section, we will examine patterns in species richness, which refers to the number of species in a community. We will also examine patterns in the abundance of individuals for each species and quantifying the diversity of species in a community in terms of richness and abundance. This is an important issue because ecologists often want to compare the diversity of species among communities or assess the effects of human activities on individual abundance and the species diversity of a community. Species richness The number of species in a community.
Patterns of Abundance Among Species
Abundance can be examined in both absolute and relative terms. Relative abundance is the proportion of individuals in a community represented by each species. When ecologists count the number of individuals of each species in a community, they frequently find that only a few species have low or high abundance, whereas most species have intermediate abundance. In a series of classic papers, Frank Preston developed a way to visualize this pattern of variation in abundance of different species. Preston examined the abundance of different bird species near Westerville, Ohio, and plotted their abundances, as shown in Figure 18.8a. He found that only a few species of birds in the community had fewer than 2 individuals or more than 100 individuals; most had 4 to 64 individuals.
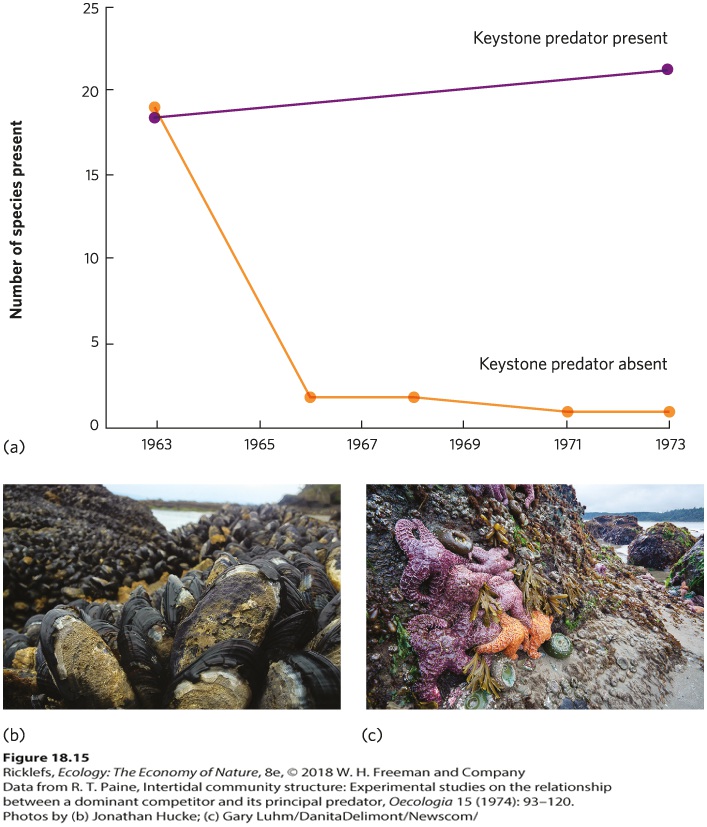
Figure 18.8 Log-normal distributions of species abundance. Log-normal distributions are found by plotting the abundance categories of each species on a log2 scale and then plotting the number of species that contain each abundance category. (a) A 10-year survey of birds in Westerville, Ohio, found that few species had extremely low or high abundances; most bird species had intermediate abundances. (b) A survey of desert plants in Arizona measured abundance using the percent cover of each species, rather than the number of individuals of each species, and found a similar log-normal distribution. Relative abundance The proportion of individuals in a community represented by each species. To plot the data, Preston used the y-axis to represent the number of species and the x-axis to represent the number of individuals that comprise each species. The key to visualizing the patterns of abundance in communities was to use categories of abundance, for example, <2, 2 to <4, 4 to <8, 8 to <16, and so on. When these categories are plotted on a log2 scale, the first value in each category translates into 0, 1, 2, 3, and so on. Data plotted in this way produce a normal, or bell-shaped, distribution such that a few species have high abundance, many species have moderate abundance, and a few species
have low abundance. A normal, or bell-shaped, distribution that uses a logarithmic scale of the x-axis is a log-normal distribution. Log-normal distribution A normal, or bell-shaped, distribution that uses a logarithmic scale on the x-axis. Log-normal distributions of species abundances can be found across a wide variety of communities and taxonomic groups. For instance, Robert Whittaker surveyed desert plants and measured the abundance of each species by quantifying the percent of vegetation cover that each species provided. When he plotted percent cover on a log2 scale, which you can view in Figure 18.8b, he also found a log-normal distribution of abundance. Many studies since Preston’s classical work have also observed log-normal distributions of species abundances.
Rank-Abundance Curves
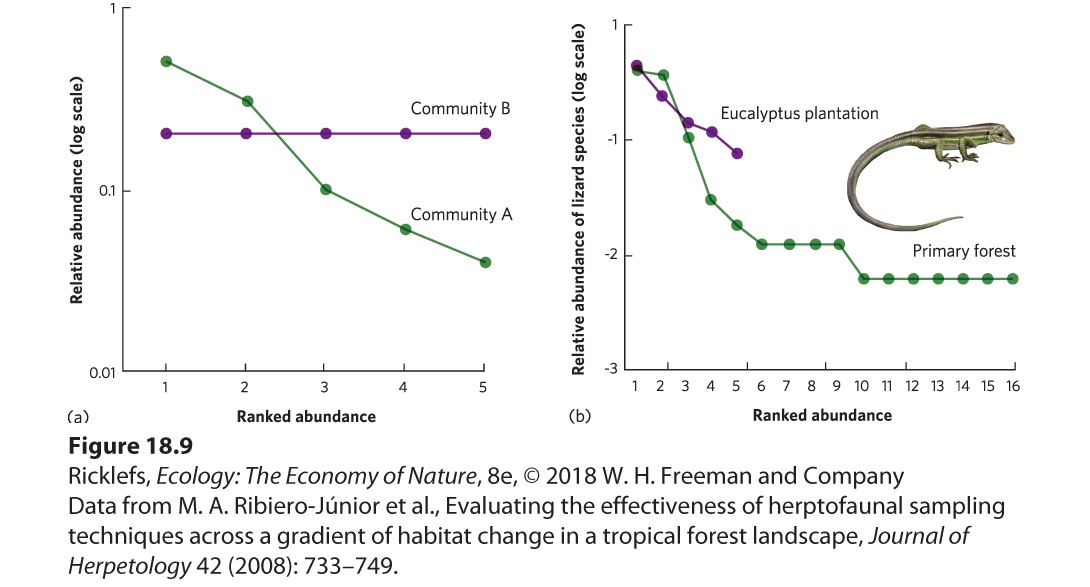
Another way to visualize the relationship between the number of species and the relative abundance of each species is to use rank-abundance curves. Rank-abundance curves plot the relative abundance of each species in a community in rank order from the most abundant to the least abundant. Rankabundance curves are particularly good for illustrating how communities differ in species richness and species evenness. Species evenness is a comparison of the relative abundance of each species in a community. The greatest evenness occurs when all species in a community have equal abundances, and the lowest evenness occurs when one species is abundant and the remaining species are rare. To plot a rank-abundance curve, we rank each species in terms of its abundance; the most abundant species receives a rank of 1, the next most abundant species receives a rank of 2, and so on. Rank-abundance curve A curve that plots the relative abundance of each species in a community in rank order from the most abundant species to the least abundant species. Species evenness A comparison of the relative abundance of each species in a community. Consider two hypothetical communities. Community A has five species with relative abundances of 0.5, 0.3, 0.1, 0.06, and 0.04, and Community B
has five species with relative abundances of 0.2, 0.2, 0.2, 0.2, and 0.2. From these numbers, we can see that both communities have the same species richness, but Community A has lower species evenness than Community B. The rank-abundance curves of these data are shown in Figure 18.9a. Both curves extend equally far to the right, which confirms that they have the same species richness. However, the slope of Community A is considerably steeper than the slope of Community B because the five species in Community A range from a very high relative abundance to a very low relative abundance. In contrast, all five species in Community B have equal relative abundances, which, when plotted, produce a flat line. Rank-abundance curves allow us to determine quickly which communities have greater species richness and evenness.
Figure 18.9 Rank-abundance curves. (a) These rank-abundance curves represent two hypothetical communities with identical species richness of five species. However, the two communities differ in species evenness. Community A has one species with a high relative abundance and other species that have moderate to low abundance, which means this community has low species evenness. In contrast, Community B contains five species that are all equally abundant so there is high evenness. (b) These rank-abundance curves for lizards living in Brazil show that the primary forest has a higher species richness and a lower species evenness than the eucalyptus plantation. Rank-abundance curves are good indicators of how communities differ in richness and evenness. For example, researchers in Brazil recently examined rank-abundance curves for lizards across different forested habitats. They determined the abundance of every lizard species they could find during daytime searches. They ranked each lizard species based on abundance and
plotted the ranks against the relative abundance of each species, as shown in
Figure 18.9b. As you can see in this figure, the rank-abundance curve for the primary forest extends farther to the right than the curve for lizards in the eucalyptus plantation, which indicates that the primary forest contains many more species of lizards than does the eucalyptus plantation. Moreover, if we consider the slopes of the curves across the ranks that the two communities have in common—from ranks 1 to 5—we see that the primary forest has a steeper slope than the curve for the eucalyptus plantation. This indicates that the lizard community in the primary forest has a lower evenness than the community in the eucalyptus plantation.
1. What does the log-normal distribution of species distributions tell us about the abundance of the full suite of species in a community? 2. What does the slope of a rank-abundance curve tell us about the relative abundance of species in a community? 3. What two measures of species diversity are incorporated when calculating indices of species diversity?
#### 18.3 Species Diversity Is Affected by Resources, Habitat Diversity, Keystone Species, and Disturbance
18.3 Species diversity is affected by resources, habitat diversity, keystone species, and disturbance We have seen that communities differ in the number of species they contain, but we have not yet addressed the more fundamental question of why they differ. On a global scale, there is great variation in the number of species found in different locations. For example, naturalists have known for centuries that more species live in tropical regions than in temperate or boreal zones. We will discuss global patterns of species richness in later chapters and consider the effects of long-term processes such as continental drift, the time that has passed since glaciation, the emergence of new islands, the dispersal of species, and the evolution of new species. In this section, we will focus on how species richness of communities within any given region of the world is affected by the resources available, the diversity of the habitat, the presence of keystone species, and the frequency and magnitude of disturbances.
Resources
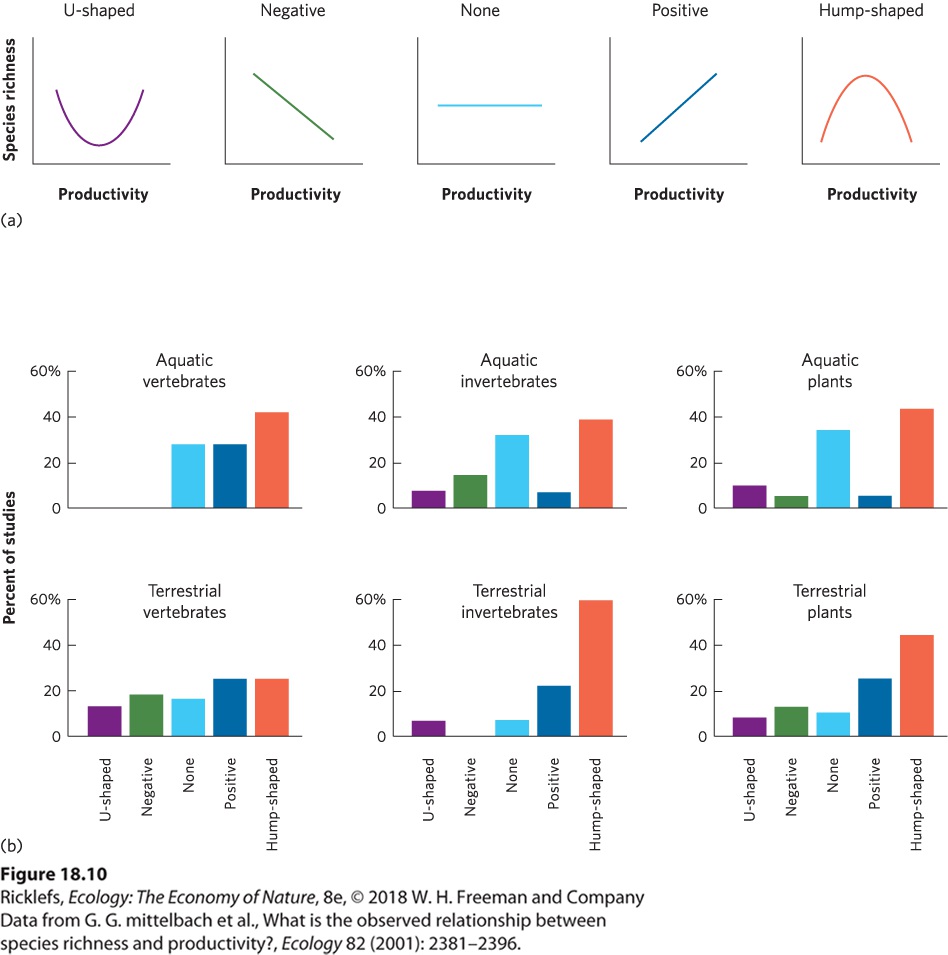
The number of species in a community can be affected by the amount of available resources. Researchers have examined the effects of resources on species diversity by determining correlations between productivity and species richness in nature. They have also looked at how species richness changes when the productivity of a community is experimentally manipulated. Natural Patterns of Productivity and Species Richness For decades, ecologists have examined how the amount of biological productivity correlates with the number of species in communities. To do this, they commonly measure productivity in terms of the biomass of producers or consumers that is generated over time. Among hundreds of studies that have been conducted on animals and plants in both aquatic and terrestrial environments, researchers have found a wide range of patterns, as illustrated in Figure 18.10a. In rare cases, researchers have observed a Ushaped curve in which increased productivity is associated with an initial decrease in species richness that is followed by an increase in species
##### Analyzing Ecology: Calculating Species Diversity
richness. In some studies, conducted in different biomes and in different parts of the world, diversity decreases with increasing productivity. In other cases, the correlation is positive; increases in productivity are associated with increases in species richness. In still other studies, there is no relationship between productivity and species richness. Finally, in some studies, the relationship is best described by a hump-shaped curve; initial increases in productivity are associated with an increase in species richness, but further increases in productivity are associated with a decrease in species richness.
Figure 18.10 Natural patterns of productivity and species richness. (a) The relationship between productivity and species richness can be described by one of five types of curves. (b) Among six categories of organisms, a hump-shaped curve is the most commonly observed relationship between productivity and species richness for aquatic vertebrates, aquatic and terrestrial invertebrates, and aquatic and terrestrial plants.
Analyzing Ecology
Calculating Species Diversity Ecologists agree that communities with more species and greater evenness have higher species diversity. Although rank-abundance curves are a helpful way to visualize differences in richness and evenness among communities, they do not provide a specific value for the species diversity within a community. Over the years, numerous indices of species diversity have been created. We will consider the two that are most common and equally valid: Simpson’s index and Shannon’s index. Both indices incorporate species richness, which we abbreviate as S, and evenness. However, they do so in different ways. To see how we calculate species diversity with either index, we can begin with data from three communities for which we have the absolute abundance for each of the species. From these data, we can then calculate relative abundance for each of the five species in the community, which is denoted as pi. With these relative abundance data, we can calculate both Simpson’s index and Shannon’s index. Simpson’s index, a measurement of species diversity, is given by the following formula: Simpson’s index A measurement of species diversity, given by the following formula: 1∑i=12(pi)2
1∑i=1s(pi)2 In words, this formula means that we square each of the relative abundance values, sum these squared values, and then take the inverse of this sum. For example, for Community A, Simpson’s index of species diversity is 1(0.24)2+(0.162)+(0.08)2+(0.34)2+(0.18)2=1.24=4.21 Simpson’s index can range from a minimum value of 1, which occurs when a community only contains one species, to a maximum value equal to the number of species in the community. This maximum value only occurs when all the species in the community have equal abundances. Shannon’s index (H'), also known as the Shannon–Wiener index, is another measurement of species diversity given by the following formula: Shannon’s index (H') A measurement of species diversity, given by the following formula: H′=-∑i=1s(pi)(lnpi) Also known as the Shannon–Wiener index. H′=-∑i=1s(pi)(lnpi) In words, this formula means that we multiply each of the relative abundance values by the natural log of the relative abundance values, sum these products, and then take the negative of this sum. For example, in Community A, Shannon’s index is -[ (0.24)(ln 0.24)+(0.16)(ln 0.16)+(0.08)(ln 0.08)+(0.34)(ln 0.34)+(0.18) (ln 0.18) ]-[ (-0.34)+(-0.29)+(-0.20)+(-0.37)+(-0.31) ]=1.51
Shannon’s index can range from a minimum value of 0, which represents a community that contains only one species, to a maximum value that is the natural log of the number of species in the community. As we saw with Simpson’s index, the maximum species diversity value occurs when all species in the community have the same relative abundances. YOUR TURN Calculate Simpson’s index and Shannon’s index for Community B and Community C. Based on your calculations, how does species evenness and species richness affect the values of each index? To gain a better sense of the relationship between productivity and species richness, a group of researchers compiled data from the hundreds of studies that have been conducted to determine the most common type of correlations. You can view their results in Figure 18.10b. Among studies of vertebrates, a hump-shaped curve was the most commonly observed relationship in aquatic systems, although hump-shaped and positive relationships were observed with similar frequency in terrestrial systems. For aquatic and terrestrial invertebrates and plants, the most commonly observed relationship was a hump-shaped curve. This reflects the fact that the most productive communities are often dominated by a small number of dominant competitors. In short, although a variety of relationships have been observed in individual studies, the most common relationship across all studies is a hump-shaped curve. This means that a site with medium productivity has a higher species richness than sites with either low or high productivity. Manipulations of Productivity and Species Richness Since most communities exhibit a hump-shaped relationship between productivity and species richness, we ought to be able to predict how species richness will be affected if communities experience increased productivity. Numerous experiments have been conducted in communities of plants in which productivity has been manipulated by adding soil nutrients such as nitrogen and phosphorus. As predicted by the hump-shaped curve, the most common observation in such experiments is that species diversity declines over time at high levels of productivity. In a classic study that took place in England, known as the Park Grass Experiment, researchers in 1856 began
adding different types of fertilizers to several plots of grass each year while leaving other plots as unfertilized controls (Figure 18.11a). Within 2 years of starting the experiment, species richness in the fertilized plots began to decline. This decline continued for several decades and researchers determined that the amount of decline was related to the number of different nutrients that were added, as shown in Figure 18.11b. The annual addition of nutrients continues today, over 150 years later, which makes the Park Grass Experiment one of the longest-running experiments in the world. Although the original experiment contained treatments that were not replicated, subsequent experiments with replicated treatments have supported the conclusions of the Park Grass Experiment.
Figure 18.11 Fertility effects on species richness. (a) Researchers of the Park Grass experiment have manipulated the addition of different types of nutrients in grassland plots over time and observed changes in species richness. (b) Plots that received more types of resources—such as nitrogen, phosphorus, and micronutrients—experienced a greater decline in species richness. The Park Grass Experiment and others like it have demonstrated that added fertility commonly causes a decline in the species richness of producers such as plants and algae. Typically, the total biomass of producers increases when a fertilizer is added, but this

added fertilizer causes a few species to dominate the community, while rare species—which are often competitively inferior—begin to decline until they eventually disappear from the community. This observation has important global ramifications because humans continue to add nutrients inadvertently to aquatic and terrestrial communities through fertilizer runoff and air pollution. Although we know that species richness declines with increased habitat fertility, the reasons have been unclear. In the case of plant communities, ecologists have hypothesized that the increase in nutrients caused competitively dominant plants to cast more shade on the competitively inferior plants. In one experiment, four manipulations were used, as illustrated in Figure 18.12: a control community, a community that received added nutrients, a community that received added light in the plant understory, and a community that received added nutrients and added light in the plant understory. As Figure 18.12 shows, while adding nutrients alone caused a decline in species richness, as expected from past experiments, adding light alone had no effect. However, adding both nutrients to the soils and light to the understory reversed the decline in species richness that was seen with the addition of nutrients alone. These results confirm that soil fertility drives down species richness because it promotes the growth of taller, competitively superior plants that cast shade over less-competitive plants.
Figure 18.12 Reversing the effects of added nutrients on species richness. When fertilizer alone is added to a grassland community, species richness declines. When light is added under the canopy of the grasses, species richness is not affected. However, when fertilizer is added in combination with light under the canopy, there is no decline in richness. These results suggest that adding fertilizer causes richness to decline because increased fertility favors the tall and competitively dominant plants, which in turn shade the competitively inferior plants. Error bars are 95 percent confidence intervals.
Habitat Diversity
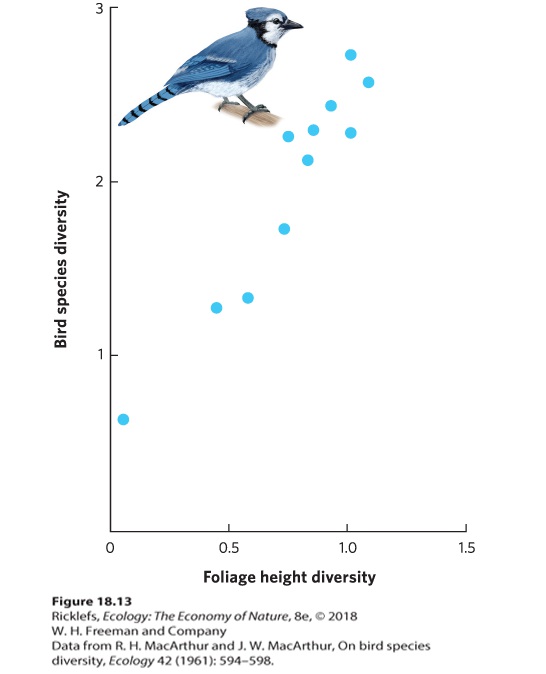
The number of species in a community can also be affected by the diversity of the habitat. Because different habitats provide places to feed and breed for different species, it seems reasonable that communities with a higher diversity of habitats—which should offer more potential niches—will also have a higher diversity of species. In a classic study of the role of habitat diversity, Robert and John MacArthur investigated whether there was a relationship between the diversity of habitats among different regions of the United States and Panama and the diversity of birds in each location. To accomplish this, they measured the density of foliage at different heights above the ground in these different regions, from 0.2 to 18.3 m, and calculated the diversity of foliage heights using the Shannon index. Then they surveyed the number of breeding bird species in each area and calculated the diversity of birds, again using the Shannon index. When they graphed the relationship between the two sets of
data, as shown in Figure 18.13, they found that habitats with greater foliage height diversity supported higher diversity of bird species.
Figure 18.13 Habitat diversity and species diversity. Using the Shannon index, researchers quantified the diversity of foliage height and the diversity of bird species in different locations around the United States and Panama. They found that more diverse habitats contained a greater diversity of birds.
Keystone Species
All species play a role in a community, but some species, known as keystone species, substantially affect the structure of communities despite the fact that individuals of that species may not be particularly numerous. The concept of
the keystone species is a metaphor that comes from the field of architecture. In arches that are built of stone, the center stone is known as the keystone (Figure 18.14). The keystone does not carry much of the arch’s mass, but without the keystone the arch would collapse. Similarly, removing a keystone species can cause a community to collapse.
Figure 18.14 Keystone. A keystone in an arch is the final stone that is inserted to prevent the arch from collapsing. Similarly, a keystone species is one whose removal causes a dramatic change in the community, despite not being particularly abundant.
Keystone species A species that substantially affects the structure of communities despite the fact that individuals of the species might not be particularly numerous. Keystone species affect communities in a wide variety of ways, some in their roles as predators, parasites, herbivores, or competitors. We saw an example of herbivores acting as a keystone species in Chapter 16 when we discussed an experiment in which researchers sprayed several field plots with an insecticide and left other plots unsprayed (see Figure 16.13). In the unsprayed plots, beetles consumed the competitively superior goldenrod and the competitively inferior plants survived and grew better. In the sprayed plots, the beetles remained rare, and this allowed the goldenrod plants to dominate the community. In this community, the beetle was a keystone species because its presence completely altered the structure of the field community. A similar scenario exists in the intertidal communities along the coast of Washington State. In a classic experiment, Robert Paine built cages in the intertidal zone to exclude predatory sea stars (Pisaster ochraceus) from feeding on a variety of herbivores, including mussels, barnacles, limpets, and snails. Adjacent plots of similar size were left as uncaged controls. As you can see in Figure 18.15a, the control plots experienced little change in species composition from 1963 to 1973. In the caged plots, however, nearly 20 species declined to the point that they were eventually eliminated from the area. In their place, a single species of mussel (Mytilus californianus) came to dominate the rock surface. Because the mussels are the superior competitors for space on the rocks, mussels dominate the community when sea stars are absent (Figure 18.15b). When the sea stars are present, they consume large numbers of mussels, and this creates open areas of rock that many species of inferior competitors can colonize (Figure 18.15c). In short, the sea star is a keystone species in the intertidal community.
Figure 18.15 Sea stars as a keystone species. (a) Although they are not very abundant, sea stars act as keystone species in intertidal communities along the coast of Washington State because they remove competitively dominant mussels. (b) In areas where sea stars were removed, the competitively superior mussels came to dominate the community and caused a decline in the diversity of the competitively inferior species. (c) In areas where the sea stars remained, the community retained a high diversity of intertidal species. Keystone species can also affect communities by influencing the structure of a habitat. In such cases, keystone species are sometimes called ecosystem engineers. One of the best-known ecosystem engineers is the beaver. Beavers build dams in streams that block the flow of water and cause large ponds to develop (Figure 18.16). Because the flowing stream is converted into a nonflowing pond, a different community of plants and animals colonize and persist in beaver ponds than in streams. Similarly, alligators create numerous
large depressions in wetlands, known as gator holes, that many other species use, including fish, insects, crustaceans, birds, and mammals. Although alligators and beavers are not particularly numerous, each of these species has a major effect on its community, which makes it a keystone species.
Figure 18.16 Ecosystem engineers. While beavers are not very abundant, the dams they build, like this one in Grand Teton National Park, Wyoming, have a major effect on community structure. Beaver dams flood terrestrial communities, which kills most terrestrial plants. In addition, the aquatic community changes from species that live in flowing streams to those that live in nonflowing ponds.
Disturbances
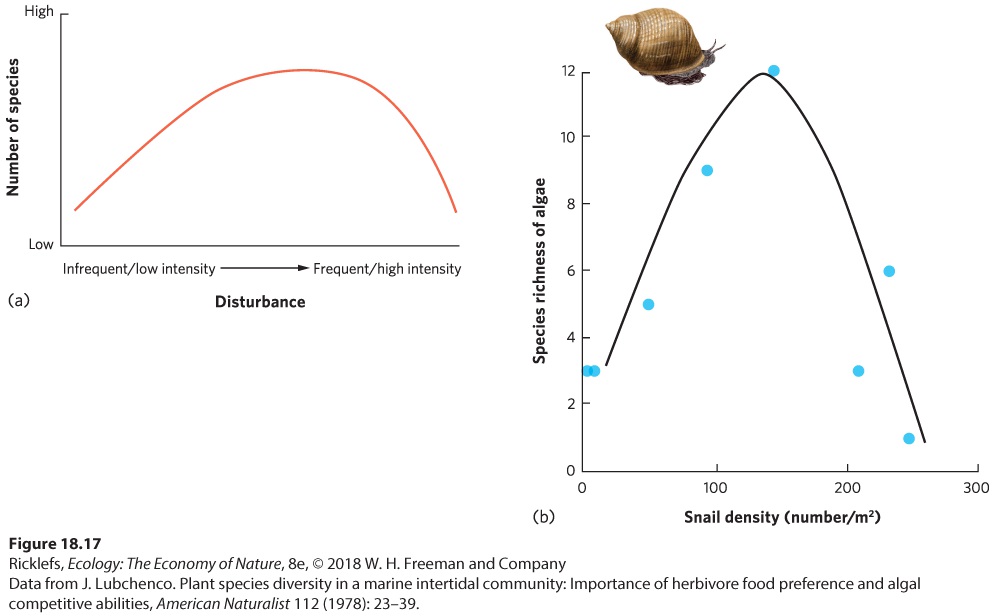
As we discussed in previous chapters, some species are well adapted to environments that experience frequent and large disturbances, such as hurricanes, fires, or intense herbivory. When environments are rarely disturbed or the disturbances have a low intensity, populations can continue to grow, resources become less abundant, and the ability to compete becomes more important for the persistence of a species. Conversely, frequently disturbed habitats typically support species that are adapted to disturbances. However, when habitats experience disturbances at some intermediate frequency, both types of species can persist and the total number of species can be higher than it would be at either extreme. The intermediate disturbance hypothesis tells us that more species are present in a community that experiences occasional disturbances than in a community that experiences frequent or rare disturbances. As depicted in Figure 18.17a, when disturbances in a community are of low frequency or intensity, species richness is relatively low. However, when disturbances are moderate in
frequency or intensity, species richness is relatively high. When disturbances are high in frequency or intensity, it declines.
Figure 18.17 The intermediate disturbance hypothesis. (a) When disturbances are frequent or of high intensity, only those species that are adapted to these conditions can persist. When disturbances are rare or of low intensity, competition becomes more intense and only those species that are well adapted to compete can persist. At intermediate levels of disturbance, species from both extremes can persist, which results in a higher species richness. (b) In a study that manipulated snail density to create different intensities of disturbance from herbivory, there was strong support for the intermediate disturbance hypothesis. Intermediate disturbance hypothesis The hypothesis that more species are present in a community that occasionally experiences disturbances than in a community that experiences frequent or rare disturbances. The intermediate disturbance hypothesis has been observed in many different communities. One classic study was conducted by Jane Lubchenco, who manipulated the density of herbivorous periwinkle snails in tidal pools to determine how an increase in the magnitude of an herbivory disturbance would affect the species richness of algae in the pools. Her data, shown in
Figure 18.17b, reveal a low richness of algal species at low snail densities, a high richness of algal species at moderate snail densities, and a low richness of algal species at high snail densities. Many similar types of studies have been conducted over several decades, and a 2012 analysis of these studies
found strong support for the intermediate disturbance hypothesis with respect to species richness.
Concept Check
1. Why does an increase in nutrients commonly lead to a decline in plant species diversity? 2. What is the relationship between habitat diversity and species diversity? 3. Why are sea stars considered a keystone species in intertidal communities?
#### 18.4 Communities Are Organized into Food Webs
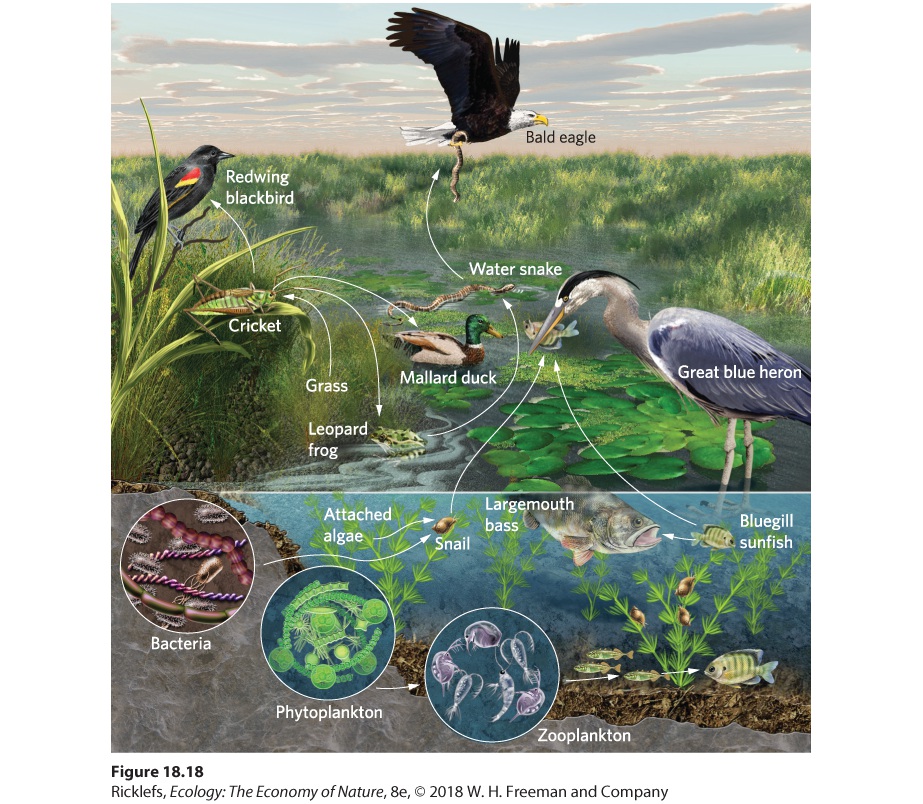
18.4 Communities are organized into food webs Even the simplest communities are composed of a large number of species. To understand the relationships among the species, it is helpful to categorize species in a community into food chains and food webs. Food chains are linear representations of how species in a community consume each other and therefore how they transfer energy and nutrients from one group to another in an ecosystem. Food chains greatly simplify species interactions in a community. In contrast, food webs are complex and realistic representations of how species feed on each other in a community and include links among many species of producers, consumers, detritivores, scavengers, and decomposers. We saw an example of this in our opening story of decomposing cacao husks, pollinators, predators, and cacao trees. In constructing food webs, arrows are used to indicate consumption patterns— and therefore the movement of energy and nutrients—between groups. You can see an example of a food web in Figure 18.18.
Figure 18.18 Food webs. Food webs capture the full diversity of species and species interactions in the community. In this North American lake, the major producers are the plants
along the shoreline and the phytoplankton in the water. These producers are eaten by a variety of primary consumers, as indicated by the arrows that illustrate the flow of energy from an organism to the species consuming it. This food web also includes secondary consumers, such as bluegill sunfish; tertiary consumers, such as the great blue heron; and decomposers, such as bacteria. Food chain A linear representation of how different species in a community feed on each other. Food web A complex and realistic representation of how species feed on each other in a community. Food webs describe the feeding relationships of ecological communities, which is important because feeding relationships help determine whether a species can exist in a community and whether it will be rare or abundant. The species richness in a food web can often be quite high, which makes it challenging to comprehend how the abundance of one species is affected by other species in the community. To simplify this challenge, ecologists categorize species into trophic levels.
Trophic Levels
You may recall the concepts of producer and consumer from the overview of basic ecological concepts in Chapter 1. These represent broad categories of trophic levels, which are the levels in a food chain or food web of an ecosystem. All organisms at a particular trophic level obtain their energy in a similar way. Producers are the autotrophs, including algae such as phytoplankton, and plants that convert light energy and CO2 into carbohydrates through photosynthesis. Producers form the first trophic level of a food web. Consumers can be subdivided into primary consumers, secondary consumers, and tertiary consumers. Primary consumers are those species that eat producers. In the aquatic food web portrayed in Figure 18.18, the primary consumers include zooplankton that eat algae and snails that eat plants. Secondary consumers are those species that eat primary consumers. In the lake food web, these secondary consumers include small fish that eat the zooplankton and ducks that eat the snails. Some communities support tertiary consumers, which eat secondary consumers; for example, large fish in a lake are tertiary consumers because they consume small fish. In addition to these producers and consumers, food webs also include consumers of dead
organic matter, such as scavengers, detritivores, and decomposers. One challenge to placing species into specific trophic levels is that many species are omnivores, meaning that they can feed at several trophic levels. For example, crayfish consume algae, which would make them primary consumers, but they also feed on insects and detritus, which would make them secondary consumers and detritivores. Trophic level A level in a food chain or food web of an ecosystem. Primary consumer A species that eats producers. Secondary consumer A species that eats primary consumers. Tertiary consumer A species that eats secondary consumers. Omnivore A species that feeds at several trophic levels. Within a given trophic level, we can group species that feed on similar items into guilds. For example, a grassland might contain a wide variety of primary consumers that feed on plants in different ways; these consumers can be categorized into guilds of leaf eaters, stem borers, root chewers, nectar sippers, or bud nippers. Members of a guild feed on similar items, but they need not be closely related. In Chapter 16, for example, we discussed how ants and rodents in the deserts of the southwestern United States compete for seeds of the same size. Though ants and rodents are not closely related, their competition for the same seeds would place them in the same guild. Guild Within a given trophic level, a group of species that feeds on similar items.
Direct Versus Indirect Effects
As we have seen, a change in the abundance of any one species can affect the abundance of the other species. The simplest way that one species can affect the abundance of another is through a direct effect, which occurs when two species interact without involving other species. We have discussed a variety of direct effects in our chapters on predation, parasitism, competition, and
mutualism. Direct effect An interaction between two species that does not involve other species. Because so many species are interconnected in a food web, the direct effects of one species on another often set off a chain of events that affects still other species in the community. When two species interact in a way that involves one or more intermediate species, it is called an indirect effect. When indirect effects are initiated by a predator, it is called a trophic cascade. Indirect effects are widespread in ecological communities. For example, in Chapter 14, we discussed how the Myxoma virus was introduced onto Macquarie Island to help reduce a population of European rabbits that had been introduced by sailors in the 1800s. As the virus killed the rabbits, there were fewer rabbits remaining to consume the vegetation, which resulted in an increase in the island’s vegetation. In short, the virus caused a positive indirect effect on the vegetation. Indirect effect An interaction between two species that involves one or more intermediate species. Trophic cascade Indirect effects in a community that are initiated by a predator. Exploitative competition between two animals might at first glance seem like a direct effect because each species has a negative effect on the other. However, it is actually an indirect effect because the two competitors are interacting with each other by feeding on a common resource. In the case of ants and rodents in the southwestern United States, for instance, the presence of ants causes a reduction in available seeds, and this reduction causes the rodent population to decline. In short, the negative effect of one competitor on another is mediated through the abundance of a third species, which is a shared resource. As a result, exploitative competition is a negative indirect effect. Indirect effects are common within communities, but sometimes they can occur between adjacent communities. For example, fish prey on aquatic insects, including the aquatic larvae of dragonflies. As a result, ponds containing fish typically have fewer dragonfly larvae. Dragonfly larvae
eventually metamorphose into dragonfly adults that consume bees and flies, which are common plant pollinators. You can see this food web in Figure 18.19a. When researchers examined the dragonfly populations of ponds with fish and ponds without fish, they confirmed that the ponds without fish had more larval and adult dragonflies. Because of this higher abundance of dragonflies, the ponds without fish also had fewer bees and flies visiting flowering plants along the shoreline, as shown in Figure 18.19b. The scarcity of bees and flies reduced potential seed production in at least one plant species. Thus, a negative direct effect between fish and dragonflies caused a cascade of negative and positive indirect effects on the surrounding terrestrial community.
Figure 18.19 Indirect effects between communities. (a) A food web of species from the aquatic and terrestrial communities includes fish that consume larval dragonflies, adult dragonflies that consume pollinators, and pollinators that pollinate plants. Solid lines represent direct effects. Dashed lines with a minus sign represent a negative indirect effect, and dashed lines with a plus sign represent a positive indirect effect. (b) Researchers found that ponds without fish had higher dragonfly populations, which reduced the numbers of pollinators available to visit flowering plants on the surrounding shoreline. Error bars are standard errors. Ecologists have traditionally assumed that indirect effects occurred only
when changes in one species altered the density of another species, such as a trophic cascade caused by predators. However, more recent research has shown that trophic cascades can also be initiated when one species causes changes in the traits of another species. We now look at a variety of indirect effects in more detail. Density-Mediated Indirect Effects Indirect effects that are caused by changes in the density of an intermediate species are called density-mediated indirect effects. For example, increased densities of sea stars in intertidal communities cause a decline in mussels, which allows other species such as snails to occupy the limited open space on the rocks. The positive indirect effect of sea stars on snails occurs because the sea stars reduce the density of mussels. Similarly, the introduction of Myxoma virus reduced the density of rabbits on Macquarie Island, which allowed an increase in the growth of the plants that rabbits eat. Density-mediated indirect effect An indirect effect caused by changes in the density of an intermediate species. Trait-Mediated Indirect Effects Communities can also experience trait-mediated indirect effects, which are indirect effects that are caused by changes in the traits of an intermediate species. This commonly happens when a predator causes its prey to change its feeding behavior, which in turn alters the amount of food consumed by the prey. For example, the reintroduction of wolves into Yellowstone National Park has caused elk to feed in more protected areas. This behavior alters the amount of plant consumption that occurs in these areas, not because the density of elk has changed but because the behavior of the elk has changed. Trait-mediated indirect effect An indirect effect caused by changes in the traits of an intermediate species. A similar example can be found in a community of spiders, grasshoppers, and grasses. Grasshoppers are insects that readily consume grasses and serve as prey for some species of spiders. There is more grass present when spiders are present in the community. The increase in grass could occur because spiders eat the grasshoppers and therefore lower the grasshopper’s density, or
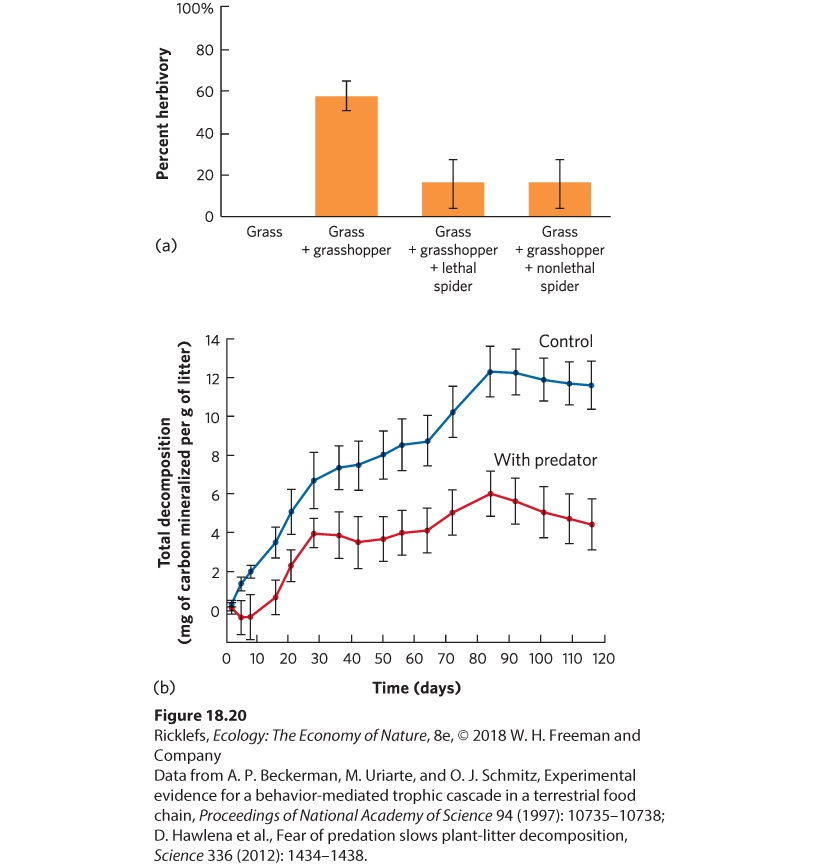
because the spiders scare the grasshoppers so that they hide more and spend less time feeding. To determine which process is operating, researchers conducted an experiment in which they placed cages around small areas of grass in a field and then conducted one of four manipulations by adding: (1) no animals (the control), (2) grasshoppers, (3) grasshoppers and lethal spiders, and (4) grasshoppers and nonlethal spiders. Although the nonlethal spiders could not kill the grasshoppers because their mouthparts had been glued shut, the grasshoppers would still recognize them as potential predators, which should cause grasshoppers to reduce their feeding time. When the lethal spiders were added, there was less herbivory, as you can see in Figure 18.20a. At first glance, this appears to be a density-mediated indirect effect because spiders kill grasshoppers. However, the density of grasshoppers was not affected by the presence of lethal spiders. This is probably because grasshopper reproduction offset spider predation. When the researchers added nonlethal spiders with glued mouthparts, the number of grasshoppers was identical to when lethal spiders were added and there was still reduced herbivory. In short, nonlethal spiders, which can alter the behavioral traits of the grasshoppers but not grasshopper density, had the same positive indirect effect on the grass as lethal spiders, which alter both the density and the traits of grasshoppers. These results illustrate that the mere presence of spiders can alter the behavioral traits of grasshoppers and initiate a trait-mediated indirect effect.
Figure 18.20 Trait-mediated indirect effects. (a) When researchers created simplified communities with only grass and grasshoppers, the amount of grass damage was relatively high. When lethal spiders were added, the grasshoppers caused less damage to the grass, which indicates that spiders had an indirect positive effect on the grass. When spiders with glued mouthparts were added, there was a similar reduction in herbivory because the grasshoppers spent less time feeding. These data confirm that the indirect effect is not mediated by reductions in grasshopper density, but by changes in grasshopper traits. (b) After the grasshoppers died, the bodies of grasshoppers raised with predators were lower in nitrogen, which caused the decomposition of the surrounding dead grass to occur at a slower rate. Error bars are standard errors. Trait-mediated effects can be far-reaching. For example, grasshoppers that are exposed to the threat of spider predation not only feed less, but their bodies also contain less nitrogen. Because nitrogen is typically a limited resource in soils, as we discussed in Chapter 3, researchers suspected that the amount of nitrogen in grasshopper bodies would affect the rate of soil decomposition when grasshoppers die and become part of the detritus. To test
this hypothesis, researchers again raised grasshoppers with no spiders and spiders with glued mouthparts. After being exposed to the two treatments, the grasshoppers were killed and their carcasses were mixed with dead grass. When researchers measured the rate of grass decomposition, they found that it was approximately three times faster with grasshoppers that had been reared without spiders than grasshoppers that had been reared with spiders, as shown in Figure 18.20b. These results demonstrate that trait-mediated effects can have far-reaching consequences throughout communities.
Top-Down and Bottom-Up Effects
The availability of resources as well as the amount of predation and parasitism a species experiences can affect the abundance of species. The same can be said for entire trophic groups. When the abundances of trophic groups in a community are determined by the amount of energy available from the producers in that community, it is referred to as bottom-up control. When the abundance of trophic groups is determined by the existence of predators at the top of the food web, it is called top-down control. You can see both of these concepts illustrated in Figure 18.21. If we return to our lake example, we can start with four trophic groups that consist of large fish, small fish, zooplankton, and phytoplankton. If an increase in phytoplankton causes an increase in the zooplankton, small fish, and large fish, the abundance of the trophic groups experiences bottom-up control. If an increase in the abundance of large fish causes a decrease in the small fish, an increase in the zooplankton upon which the small fish feed, and a decrease in phytoplankton, the abundance of the trophic groups experiences top-down control.
Figure 18.21 Top-down and bottom-up control of communities. When communities are controlled from the bottom up, an increase in the abundance of producers results in an increase in the abundance of higher trophic levels. When communities are controlled from the top down, a trophic cascade occurs from the top down: The trophic level just below the top predator decreases in abundance and the next lower trophic level increases in abundance. Bottom-up control When the abundances of trophic groups in nature are determined by the amount of energy available from the producers in a community. Top-down control When the abundance of trophic groups is determined by the existence of predators at the top of the food web. For many years, ecologists debated whether communities are more commonly under top-down or bottom-up control. If we think of food webs as having three broad trophic levels, top-down control by predators would reduce the abundance of herbivores and this would result in an abundance of vegetation. In a classic paper published in 1960, Nelson Hairston, Frederick Smith, and Lawrence Slobodkin suggested that because most communities contain an abundance of vegetation, trophic groups must be controlled from the top of the food web. Their hypothesis caused a great deal of debate among ecologists and inspired many studies of top-down and bottom-up effects. For example, researchers surveyed the abundance of zooplankton and phytoplankton and found that ponds with more phytoplankton also had more zooplankton, which suggested that zooplankton abundance is controlled from the bottom up.
However, when they conducted manipulative experiments in which small fish that eat zooplankton were added to the pond, the abundance of zooplankton decreased, which caused a trophic cascade that allowed the phytoplankton to increase. This suggested that the abundance of species in the community was controlled from the top down. Over the past 2 decades, it has become clear that many communities are simultaneously controlled both from the top down by predators and from the bottom up by resources. For example, the number of zooplankton in a lake can be influenced by both the amount of phytoplankton available for consumption and by the number of small fish available to consume the zooplankton.
Concept Check
1. What are the trophic levels of communities? 2. Why is exploitative competition considered to be an indirect effect between two competing species? 3. How can predators cause trait-mediated indirect effects?
#### 18.5 Communities Respond to Disturbances with Resistance, with Resilience, or by Switching Among Alternative Stable States
18.5 Communities respond to disturbances with resistance, with resilience, or by switching among alternative stable states Communities commonly experience disturbances from both natural and human causes. Whatever the reason behind the disturbance, the key question is whether the community will be affected and, if it is, whether it will bounce back or become a substantially different community. To address this question, we need to consider the concepts of community stability and alternative stable states.
Community Stability
As we have seen throughout this chapter, communities can experience large changes when the abundances of particular species change. Given enough time, most altered communities can rebound and resemble their original structure of species richness, composition, and relative abundance. The ability of a community to maintain a particular structure is known as community stability. There are two aspects of community stability, community resistance and community resilience. Community resistance is a measure of how much a community changes when acted upon by some disturbance, such as the addition or removal of a species. For example, we might measure community resistance after a predator is removed by looking at how much the abundance of herbivores increases. When the impact only causes a small change in the community, we say that the community is resistant to the disturbance. Community resilience is the time it takes after a disturbance for a community to return to its original state. Community stability The ability of a community to maintain a particular structure. Community resistance The amount that a community changes when acted upon by some disturbance, such as the addition or removal of a species. Community resilience The time it takes after a disturbance for a community to return to its original state. A question of practical importance is whether having a greater variety of
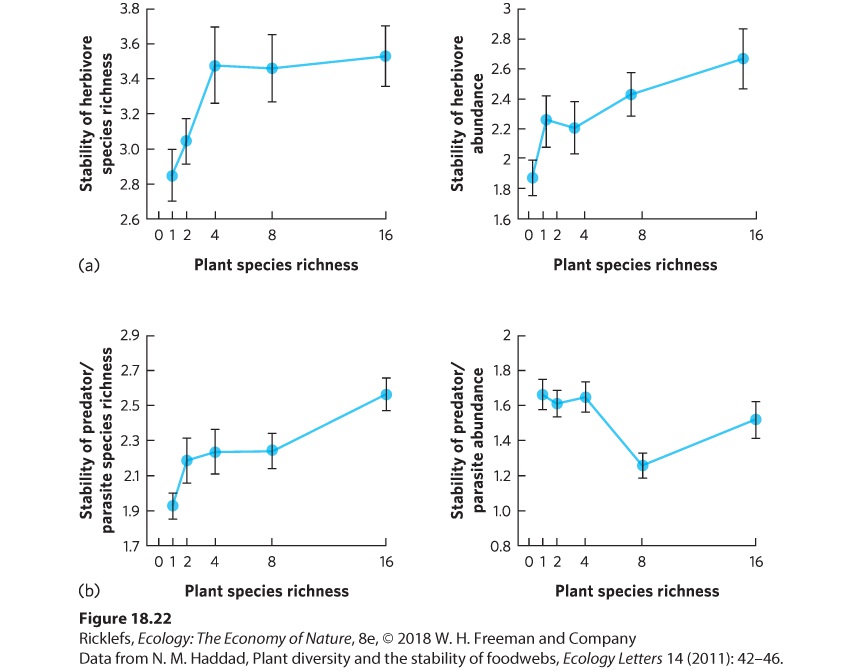
species helps communities bounce back faster from disturbances. A study to address this question was conducted in grasslands where researchers manipulated the diversity of plant species in a Minnesota prairie ecosystem by setting up plots containing 1, 2, 4, 8, or 16 species. For 11 years, the abundances of more than 700 species of invertebrate herbivores and invertebrate predators and parasitoids were monitored. During this time, the environment varied considerably, allowing researchers to ask whether increased plant diversity provided greater community stability. In this study, community stability was defined as the amount of year-to-year variation in the abundance and species richness of the herbivores and predators. The researchers discovered that increasing the number of plant species increased the stability of the communities, as shown in Figure 18.22. When they examined the herbivores, shown in Figure 18.22a, they found that an increase in the number of plant species caused an increase in the stability of herbivore richness and herbivore abundance. When they examined the predators and parasitoids, shown in Figure 18.22b, they found that an increase in the number of plant species caused an increase in the stability of predator and parasitoid, but there was no effect on the stability of predator and parasitoid abundance. The underlying reason that plant diversity led to increased stability of higher trophic levels was because communities with high plant diversity provided more consistently available food and habitat for the herbivores, predators, and parasitoids.
Figure 18.22 Species richness and community stability. Researchers manipulated the species richness of plants and then monitored the stability of the abundance and species richness of invertebrate herbivores, predators, and parasitoids. (a) An increase in the diversity of plants caused greater stability of richness and abundance of herbivores. (b) An increase in the diversity of plants also caused greater stability of richness of predators and parasitoids but had no effect on the stability of abundance. In all cases, stability is defined as the inverse of the coefficient of variation, which is the standard deviation divided by the mean. Error bars are standard errors.
Alternative Stable States
Sometimes when a stable community is strongly perturbed, it does not bounce back to its original state. Rather, it is disturbed so much that the species composition and relative abundance of populations in the community change. The new community structure is called the alternative stable state. For a community to move from one stable state to an alternative stable state typically requires a large disturbance, such as the removal of a keystone species or a dramatic change in the environment. Alternative stable state When a community is disturbed so much that the species composition and relative abundance of populations in the community change, and the new community structure is resistant to further change. Moving between alternative stable states happens commonly where the
prairie and forest biomes come together in the midwestern United States. During years of abundant rain, fire is suppressed and trees spread out into the prairie. Once trees have become established, they shade out many of the grasses and the grass species have little chance of returning. Moreover, the shade of the trees keeps the ground moist, which further reduces the likelihood of a future fire. During years of drought, however, fires are more common and large fires can kill trees, which favors the spread of grasses where trees once lived. Once grasses establish, it is difficult for trees to move into the prairie because grasses are well adapted to resprouting after fires, whereas trees are not. Alternative stable states also occur in aquatic environments. For example, intertidal communities on the coast of Maine are typically dominated by the brown rockweed algae (Ascophyllum nodosum). During the winter, however, the scouring action of ice can scrape areas free of the rockweed algae, and these areas become dominated by another brown algae known as the bladder wrack (Fucus vesiculosus). In this case, the largest disturbances occur when the largest areas are scraped free of organisms by the ice scouring. To determine whether scraping larger areas leads to communities moving to an alternative stable state, researchers simulated ice scouring on the coast by clearing areas of 1, 2, 4, and 8 m2. As shown in Figure 18.23, small scoured areas did not cause the communities to be dominated by bladder wrack, but large scoured areas did become dominated by bladder wrack. Moreover, the bladder wrack continued to dominate sites for at least 20 years. The switch to an alternative state occurred because the brown rockweed algae is a poor disperser, so it is slow to reach newly scoured sites compared to bladder wrack. However, brown rockweed algae is a superior competitor for space, so over time it will dominate the disturbed community.
Figure 18.23 Alternative stable states. Intertidal communities of Maine are commonly dominated by brown rockweed algae. When researchers scoured areas of different sizes in 1996 and then monitored the colonization of the scoured areas, they found that the largest scoured areas switched to an alternative stable state in which the community came to be dominated by a different species of brown algae known as bladder wrack. Bladder wrack is a superior disperser to new scoured sites, but it is an inferior competitor once the brown rockweed algae eventually arrives. In this chapter, we have seen that communities are composed of large numbers of interacting species that operate within food webs. The interconnected nature of food webs means that changes in the abundance of one species can potentially affect many other species in the community. In “Ecology Today: Connecting the Concepts,” we will see that understanding the connectedness of species was a key to discovering the effects of pollution on amphibians.
###### Lethal Effects of Pesticides at Nonlethal Concentrations
1. Why does higher plant species diversity stabilize the number of herbivore species over time? 2. How is community resilience different from community resistance? 3. How do fires occurring at the transition of grasslands and forests promote alternative stable states?
Concepts
Lethal Effects of Pesticides at Nonlethal Concentrations Leopard frog. Low concentrations of an insecticide that cannot kill the tadpoles of the leopard frog can kill other species in the community. This sets off a chain of events that alters the food web and indirectly prevents the leopard frogs from achieving metamorphosis. Appreciating the connectedness of species in a food web can help us understand some of the effects our activities have on ecological
communities. Pesticides, for example, provide important benefits in protecting crops and improving human health, and their widespread use has made them common in ecological communities. Maintaining pesticide concentrations below levels lethal to nonpest organisms is the key to ensuring that pesticides harm only pests and not other species. Pesticides are tested on a range of species in the laboratory to determine concentrations that cause death. However, laboratory tests do not consider the fact that species are part of a food web. Therefore, the question is if a pesticide that is not directly lethal to a species can nonetheless have indirect impacts that cause mortality. Some of the best-studied pesticides are those used to control terrestrial insects that damage crops, as well as insects such as mosquitoes that carry infectious diseases. Of those insecticides, the most commonly applied is malathion, which impairs the nervous system of animals and is highly lethal to insects and other invertebrates. Based on single-species laboratory tests, scientists thought that concentrations found in nature were not lethal to vertebrate animals. Researchers examined the effects of malathion in large outdoor tanks that contained many of the components of natural wetlands, including algae, zooplankton, and tadpoles (see Figure 1.18b). It had been long known that insecticides such as malathion were highly toxic to zooplankton, which are tiny crustaceans, but little was known about how large reductions in zooplankton might affect other species in the food web. In one such experiment, researchers added tadpoles of wood frogs and leopard frogs (R. pipiens) to tanks and then manipulated malathion in one of four ways: no
malathion to serve as a control, 50 parts per billion (ppb) at the beginning of the experiment, 250 ppb at the beginning of the experiment, or 10 ppb once per week. These manipulations allowed the researchers to ask whether single, large applications and multiple, small applications could alter the food web in ways that could affect the amphibians. Soon after malathion was applied, the zooplankton populations declined to very low numbers in all the tanks treated with the insecticide. This decline was not surprising since zooplankton were known to be quite sensitive to insecticides, but it set off a chain of events that ultimately affected the amphibians. Zooplankton feed on phytoplankton. When malathion was added and the zooplankton were killed, the phytoplankton populations rapidly grew over the next few weeks. This rapid growth, known as an algal bloom, caused the water to turn pea green. When the water turned green with phytoplankton, sunlight could no longer reach the bottom of the water column, where another group of algae, known as periphyton, grows attached to objects such as soil and leaves. The periphyton could not grow very well with less available sunlight, so within a few weeks after the start of phytoplankton bloom, the periphyton declined in abundance, especially in those communities that received 10 ppb of malathion every week. In this treatment, the decimated zooplankton populations could never exhibit resilience because the weekly additions of malathion continually reduced the rebounding populations.
Effects of an insecticide on an aquatic community. (a) In the absence of an insecticide, zooplankton are abundant and they consume phytoplankton. This consumption keeps the water relatively clear and allows sunlight to reach the bottom of a wetland where periphyton grow and serve as food for tadpoles. (b) When an insecticide is added, the zooplankton populations are dramatically reduced, which allows an increase in phytoplankton. Abundant phytoplankton cause the water to turn green and prevent sunlight from reaching the periphyton at the bottom of the wetland. With less light available, the periphyton grow poorly, which limits tadpole growth. The number of individuals pictured in each group indicates the relative change in biomass. Periphyton is the primary source of food for amphibians; without it, they cannot grow and metamorphose. Fortunately, the wood frogs develop rapidly. In the experiment, wood frogs metamorphosed after only 5 weeks, which meant that they were able to metamorphose before the bloom of phytoplankton caused a decline in their periphyton food source. As a result, wood frog growth and survival were not affected by the insecticide. The leopard frogs, however, were not so fortunate. Because leopard frogs need 7 to 10 weeks of growth as tadpoles before they can metamorphose, they experienced
the full effect of the declining periphyton when the insecticide was present. In fact, the leopard frogs had such stunted growth in the tanks with the weekly addition of 10 ppb of malathion that almost half of them failed to metamorphose before their aquatic environment dried in late summer. This research demonstrated for the first time that a low concentration of an insecticide that is not directly lethal to amphibians can be indirectly lethal. The insecticide initiated a chain of events that moved through the food web and caused nearly half of the leopard frogs to die. Moreover, the smallest application of the insecticide, which was applied each week, had a much larger effect on the leopard frogs than single applications containing 25 times more insecticide. Quantifying the insecticide’s direct effects on the amphibians suggested that the animals would not be harmed, but tests that incorporated a food web approach with indirect effects found that the insecticide could kill nearly half of the animals. Subsequent experiments have demonstrated that these outcomes are common across a wide range of wetland communities containing zooplankton, algae, and amphibians. However, a new insight arrived when researchers also incorporated aquatic plants into the experiments. Aquatic plants are present in many wetlands, although not in forested wetlands that receive little sunlight due to shading by overhead trees. When researchers examined the effect of adding aquatic plants, they discovered that plants can make malathion-contaminated water safe for zooplankton. The plants accomplish this by elevating the pH of the water (via photosynthesis), which causes the pesticide to break down much more rapidly
(i.e., within hours rather than days). As a result, the zooplankton do not die and the lethal chain of events through the food webs no longer occurs. These results underscore the importance of understanding ecology at the community level. By understanding direct and indirect effects in food webs, we can better understand and predict the impacts of human activities on ecological communities. SOURCES: Brogan III, W. R., and R. A. Relyea. 2017. Multiple mitigation mechanisms: Effects of submerged plants on the toxicity of nine insecticides to aquatic animals macrophytes. Environmental Pollution 220: 688–695. Hua, J., and R. A. Relyea. 2012. East Coast versus West Coast: Effects of an insecticide in communities containing different amphibian assemblages. Freshwater Science 21: 787–799. Relyea, R. A., and N. Diecks. 2008. An unexpected chain of events: Lethal effects of pesticides on frogs at sublethal concentrations. Ecological Applications 18: 1728–1742.
Summary of Learning Objectives
18.1 Communities can have distinct or gradual boundaries. Communities frequently exist in zones that reflect changes in biotic and abiotic conditions. Based on the aggregations of species, ecologists categorize communities in terms of the dominant biological or physical conditions. Although some communities have distinct boundaries as a result of sharp changes in environmental conditions, most communities contain species with geographic ranges that are independent of each other, which produces gradual changes in the composition of communities as one moves across the landscape. Key Terms: Ecotone, Interdependent communities, Independent communities
18.2 The diversity of a community incorporates both the number and relative abundance of species. Species richness is the number of species in a community, whereas species evenness is the similarity of relative abundance among species in a community. Rankabundance curves are graphical representations of richness and evenness. When we plot categories of individual abundance against the number of species that fall within each category, communities commonly exhibit a log-normal distribution, which means that few species have either few or many individuals and that most species have an intermediate number of individuals. Key Terms: Species richness, Relative abundance, Log-normal distribution, Rank-abundance curve, Species evenness
18.3 Species diversity is affected by resources, habitat diversity, keystone species, and disturbance. Increases in the abundance of resources and in the number of different resources can alter species diversity. Increases in habitat diversity offer a greater diversity of niches, which favors higher species diversity. Keystone species can alter the composition of species in a community because they have
large effects, even though they are not particularly numerous. Disturbances also affect the species diversity of communities, with the greatest diversity occurring in communities experiencing disturbances that are intermediate in frequency or intensity. Key Terms: Simpson’s index, Shannon’s index, Keystone species, Intermediate disturbance hypothesis
18.4 Communities are organized into food webs. Food chains are linear representations of how different species in a community feed on each other, whereas food webs are more complex and realistic representations. Species in a food web can be categorized into trophic levels and guilds according to how they obtain their energy as producers, primary consumers, secondary consumers, and tertiary consumers. Because species exist within food webs, the abundance of each species is affected not only by direct effects but also by density- and trait-mediated indirect effects. Because of these indirect effects, communities can experience trophic cascades from the top of the food chain down, and from the bottom up. Key Terms: Food chain, Food web, Trophic level, Primary consumer, Secondary consumer, Tertiary consumer, Omnivore, Guild, Direct effect, Indirect effect, Trophic cascade, Densitymediated indirect effect, Trait-mediated indirect effect, Bottom-up control, Top-down control
18.5 Communities respond to disturbances with resistance, resilience, or switching among alternative stable states. Resistant communities show little or no response to a disturbance. Resilient communities can be affected by a disturbance, but they bounce back to their original states relatively quickly. Some communities respond to large disturbances by moving to alternative stable states in which the community persists with a different composition of species for a relatively long time. Key Terms: Community stability, Community resistance,
Critical Thinking Questions
1. In a desert biome, how could you determine if the distribution of plant species indicates whether the community is independent or interdependent? 2. Given that the distributions of many animals are determined by the species composition of the plant community, what might you predict about the diversity of animals in an ecotone, compared to each adjacent habitat? 3. If you observe zones of different plant species as you move uphill from the edge of a pond, what ecological processes would you hypothesize might underlie these plant distributions? 4. Why do ecologists consider both species richness and species evenness when they quantify species diversity? 5. Compare and contrast log-normal abundance distributions and rank-abundance curves. 6. Why might an increase in total resources or number of added resources lead to declines in species diversity in a lake? 7. In areas experiencing intermediate disturbance, how might species richness and species evenness compare to areas experiencing no disturbances? 8. Rabbits in Australia were once overabundant and decimated the vegetation. Why are the rabbits in Australia not an example of a keystone species? 9. Compare and contrast density- and trait-mediated indirect effects. 10. What evidence would convince you that a grassland was experiencing top-down control versus bottom-up control?
#### Graphing the Data: Log-Normal Distributions and Rank-Abundance Curves
GRAPHING THE DATA Log-Normal Distributions and Rank-Abundance Curves You have been asked to assess the diversity of species in a grassland plot and you find 16 species that vary in relative abundance. Using the data in the table, create a rank-abundance curve and a lognormal distribution. Species Rank Absolute Abundance Relative Abundance Log-Normal Category 0.385 ≥128 0.151 64 to <128 0.081 64 to <128 0.078 32 to <64 0.064 32 to <64 0.051 32 to <64 0.039 16 to <32 0.038 16 to <32 0.034 16 to <32 0.023 16 to <32 0.019 4 to <8 0.016 4 to <8 0.010 4 to <8 0.006 2 to <4 0.005 2 to <4 0.001 <2 TOTAL =