20 Movement of Energy in Ecosystems
20Movement of Energy in Ecosystems The red-backed salamander. The introduction of earthworms from Europe and Asia to northern forests that historically lacked earthworms has caused a decline in leaf litter and soil insects. This has led to a sharp decline in the abundance of red-backed salamanders. Worming Your Way into an Ecosystem Earthworms play an important role in terrestrial ecosystems. As they burrow in the ground to consume detritus, they aerate compact soils, which allows water to better percolate into the ground. Earthworms seem to be everywhere in North America; you may have seen them on sidewalks and roads during rainy days or in the soil if you have ever worked in a garden. Therefore, it is surprising that many common species of earthworms are not native to North America but were introduced from Europe and Asia in the eighteenth century. The northern temperate and boreal forests of North America did not

previously have any earthworms, so these forests are not evolved to the altered environmental conditions caused by earthworms. As a result, the introduced worms are now having a profound effect on the movement of energy in these northern ecosystems. Scientists hypothesize that northern forests completely lack native species of earthworms because the glaciers that advanced over the region during past ice ages eliminated all life. Since the ice receded 10,000 years ago, many other animal species have since returned to the area. However, the native earthworms that survived in the southern United States have a very slow rate of dispersal and have not yet arrived in many habitats in the northern United States. When there are no worms present, the leaf litter in these forests is decomposed primarily by soil fungi and microbes. However, this changed when European colonists inadvertently introduced European and Asian worms to the northern regions of North America. These worms have a high rate of dispersal and they tolerate a wide range of ecological conditions. “Scientists hypothesize that northern forests completely lack native species of earthworms because the glaciers that advanced over the region during past ice ages eliminated all life.” The introduced earthworms are still in the process of spreading into northern forests, assisted by the construction of logging roads and anglers who use the worms as bait. As consumers of dead leaves, the invasive worms can consume much of the energy available in the detritus and move many of the nutrients deeper into the soil, where young plants cannot access them. The worms also leave much less energy for other organisms, such as the soil fungi that specialize on decomposing dead leaves. This is a major shift in the movement of energy. In addition to moving nutrients away from the forest floor, earthworm activity leaves a thinner layer of leaves and other organic
matter, which leads to drier soil conditions. These changes in abiotic conditions dramatically alter the soil food web. For example, researchers recently reported that the recent arrival of introduced worms into the forests of New York and Pennsylvania caused a nearly 50 percent reduction in leaf litter. This, in turn, resulted in a substantial decline in the abundance of soil insects, such as springtails, ants, and beetles. The decline in the insects also meant that there was less energy available for predators of the insects, such as the red-backed salamander (Plethodoncinereus). In areas with high densities of introduced earthworms, the decline in leaf litter and soil insects resulted in an 80 percent reduction in the number of salamanders. While adult salamanders can consume earthworms, young salamanders consume the much smaller soil insects. Introduced earthworms. Northern temperate and boreal forests have lacked native earthworms since glaciers eliminated them during past ice ages. Today, these forests are being invaded by species of earthworms that have been inadvertently introduced from Europe and Asia. Salamanders aren’t the only consumer affected by the earthworm invasion. In 2012, researchers reported that a forest-dwelling ovenbird (Seiurusaurocapilla) was declining in areas of Wisconsin and Minnesota that the introduced earthworms had invaded. While the exact mechanisms were not clear in this case, the researchers suspected that the decline in the ovenbirds occurred, in part, because earthworms reduced the availability of insects on the forest floor, something similar to what caused the decline of the red-backed salamander. In addition to affecting the animals of the forest, earthworms can
also affect the plants. In 2017, researchers brought together all the studies of non-native earthworm effects on North American plants, and what they found was striking. While there was no overall effect of species richness or evenness, a decrease in the abundance of native plants and an increase in the abundance of non-native plants took place. The worms also favored an increase in grasses, but not an increase in herbs, shrubs, or trees. The reason is likely because grasses are especially good at rapidly absorbing available nutrients and tolerating dry summer conditions. Some species of introduced earthworms may also consume small seeds and seedlings of the particular plants. The story of the invasive earthworms demonstrates that species depend on energy flowing between producers, detritivores, and consumers. Changes in energy flow within and between trophic groups can have major impacts on the species that inhabit an ecosystem. In this chapter, we will explore the flow of energy through food webs and the dynamics of energy movement through the ecosystem.
Craven, D., et al. 2017. The unseen invaders: Introduced earthworms as drivers of change in North American forests (a meta-analysis). Global Change Biology 23: 1065–1074. Loss, S. R., et al. 2012. Invasions of non-native earthworms related to population declines of ground-nesting songbirds across a regional extent in northern hardwood forests of North America. Landscape Ecology 27: 683–696. Maerz, J. C., et al. 2009. Declines in woodland salamander abundance associated with nonnative earthworm and plant invasions. Conservation Biology 23: 975–981.
Learning Objectives
After reading this chapter, you should be able to:
20.1 Describe how primary productivity provides energy to the ecosystem.
20.2 Compare the net primary productivity among different
20.3 Explain how the movement of energy depends on the efficiency of energy flow. In Chapter 1, we noted that the ecosystem approach to ecology focuses on the transfer of energy and matter among living and nonliving components within and between ecosystems. The amount of energy that fuels ecosystems and the efficiency with which energy is transferred through trophic levels determine the number of trophic levels in communities and ecosystems. The amount of energy available and the efficiency of its transfer determine the biomass of organisms that exist at each trophic level and the amount of energy that is left behind for scavengers, detritivores, and decomposers. In this chapter, we will discuss the movement of energy in ecosystems, including the importance of primary producers and the flow of energy throughout the food web. In the next chapter, we will concentrate on how matter, in the form of key chemical elements, cycles around ecosystems.
#### 20.1 Primary Productivity Provides Energy to the Ecosystem

20.1 Primary productivity provides energy to the ecosystem The vast majority of all energy that moves through ecosystems originates as solar energy that powers the photosynthesis of producers. As we saw in
Chapter 1, some communities—such as those that form around thermal vents in the deep ocean—rely on chemosynthesis as their source of energy. Regardless of origin, producers harness energy and form the basis of food webs. Producers use this energy for respiration, growth, and reproduction; the amount used for growth and reproduction represents the energy that is available to consumers. In this section, we will examine how to quantify the amount of energy that producers use for different functions, how to measure the energy of producers in different types of ecosystems, and how the amount of energy available in producers affects the growth and reproduction of consumers.
Primary Productivity
Primary productivity is the rate at which solar or chemical energy is captured and converted into chemical bonds by photosynthesis or chemosynthesis. Primary productivity tells us how much energy is available in an ecosystem. A related concept is the standing crop of an ecosystem, which is the biomass of producers present in the ecosystem in a given area at a given time. For example, the standing crop of a forest is the total mass of trees, shrubs, herbs, and grasses that is present in an area of the forest on a particular day. Ecosystems with high primary productivity may or may not have a high standing mass. In lakes where algae experience high productivity, consumers often eat it nearly as quickly as it grows, and so the standing crop of algae remains low. Primary productivity The rate at which solar or chemical energy is captured and converted into chemical bonds by photosynthesis or chemosynthesis. Standing crop The biomass of producers present in a given area of an ecosystem at a particular moment in time.

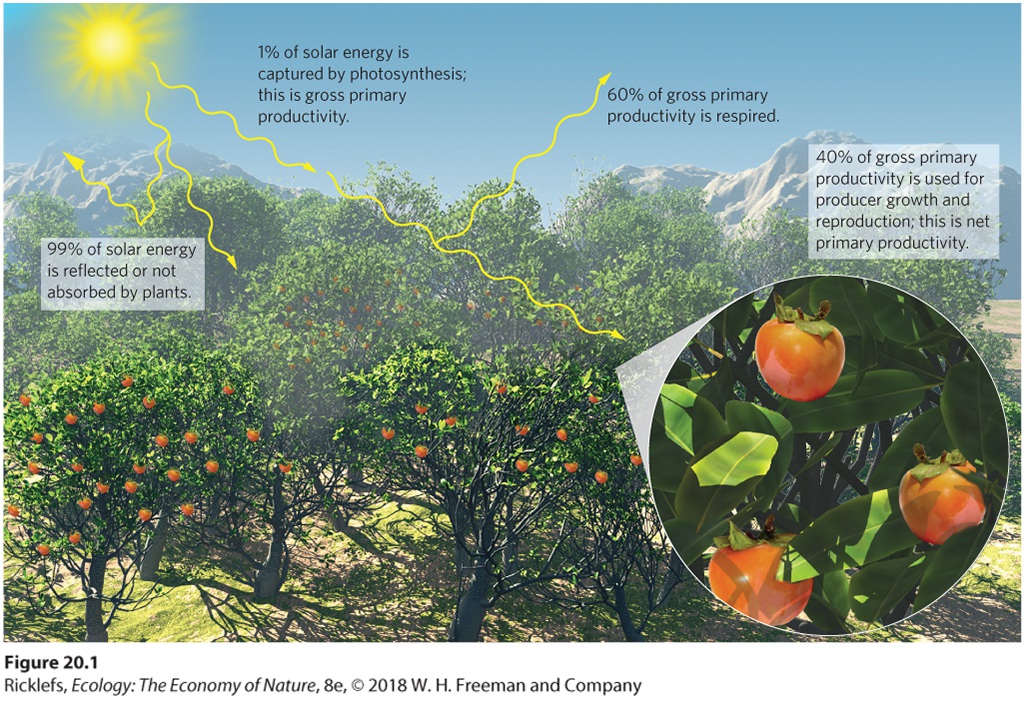
Ecologists identify two types of primary productivity: gross primary productivity and net primary productivity. Gross primary productivity (GPP) is the rate at which energy is captured and assimilated by producers in a given area. We often express productivity in units of Joules (J) or kilo Joules (kJ) per square meter per year. From this total, producers use some of the assimilated energy for their metabolism, which is measured in terms of the amount of respiration. The rest of the assimilated energy is converted into their biomass, which includes growth and reproduction. The rate at which energy is assimilated by producers and converted into producer biomass in a given area is net primary productivity (NPP). We can also show this in the form of an equation: NPP=GPP-Respiration Gross primary productivity (GPP) The rate at which energy is captured and assimilated by producers in a given area. Net primary productivity (NPP) The rate at which energy is assimilated by producers and converted into producer biomass in a given area. Considering the rate at which the Sun provides energy, photosynthesis is not a very efficient process. As illustrated in Figure 20.1, approximately 99 percent of all solar energy that is available to producers either reflects off them or passes through their tissues without being absorbed. Of the 1 percent of the solar energy that producers absorb and use for photosynthesis—that is, gross primary productivity—approximately 60 percent is used for respiration. This means that only 40 percent of absorbed solar energy is used for net primary productivity, which represents the growth and reproduction of producers. For example, if we measure the productivity of a forest in North America in units of kilograms of carbon per square meter per year, such a forest might have a GPP of 2.5 kg C/m2/year. Of this total, the forest will use about 1.5 kg C/m2/year for respiration, leaving 1.0 kg C/m2/year for growth and reproduction, which represents the forest’s NPP.
Figure 20.1 Gross primary productivity and net primary productivity. About 99 percent of solar energy is reflected by producers or passes through their tissues without being absorbed. Only about 1 percent is captured by photosynthesis for gross primary productivity (GPP). Of the total GPP, about 60 percent is used by producers for respiration. The remaining 40 percent is available to producers for growth and reproduction, which is known as net primary productivity (NPP).
Measuring Primary Productivity
Primary productivity is the foundation for the flow of energy through food webs and ecosystems. Being able to measure primary productivity allows us to track how it changes in an ecosystem over time and how it varies in different ecosystems around the globe. Since primary productivity is a rate that is achieved by the benefits of respiration minus the costs of respiration, we can measure it by quantifying the change in the biomass of producers over time, the movement of carbon dioxide over time, or the movement of oxygen over time. In terrestrial ecosystems, we commonly measure the productivity of just the plants, but in aquatic systems, we might measure the productivity of plants, large kelp, or tiny species of algae. As we will see, the choice of how to measure primary productivity depends on the particular ecosystem being studied. When measuring primary productivity, it is critical to determine whether our measurements represent gross or net primary productivity. In some cases, we can measure net primary productivity and respiration separately and then use these values to estimate gross primary productivity. As we will see in the
next section, there is a variety of methods for measuring primary productivity in terrestrial and aquatic ecosystems. Measuring Changes in Producer Biomass One of the simplest ways to measure NPP is by measuring producer biomass in an area at the beginning and at the end of a growing season (Figure 20.2). For example, in prairie ecosystems, researchers commonly measure the amount of new plant growth that has accumulated by the end of the summer growing season, while in aquatic ecosystems, ecologists measure the biomass of large aquatic producers such as kelps. A season’s growth can be harvested, dried to eliminate any water, and then weighed to determine how much growth occurred during the season. When a harvest is measured to determine biomass, the assumption has to be made that no substantial herbivory or tissue mortality has occurred during the period of producer growth. Alternatively, the amount of biomass lost to herbivory or tissue mortality can be estimated and included in the estimate of net primary productivity.
Figure 20.2 Harvesting primary productivity. Net primary productivity is commonly measured by harvesting a defined area of an ecosystem at the end of the growing season and determining the accumulated biomass of producers. This technique can be used on (a) small scales, such as by these students, who are measuring the aboveground biomass of plants, or on (b) large scales, such as the harvesting of crops in a field. In some NPP studies, only the part of the plant that lives aboveground is harvested. However, the amount of biomass belowground for some plants can be substantial. For example, perennial grasses with extensive root systems have twice as much belowground biomass as aboveground biomass. In contrast, most trees have about five times more aboveground biomass than
belowground biomass. Measuring underground biomass is quite a challenge. Retrieving the rhizomes, tubers, and roots of plants from the terrestrial soils or the aquatic benthos is often not feasible because they are either very deep or the root systems are composed of many fine roots that break off when harvested. In many plants, these fine roots die and are replaced with new fine roots, which makes it difficult to estimate how much biomass a group of producers has accumulated. It is also difficult to account for plants in mutualistic relationships with mycorrhizal fungi. These plants may provide carbohydrates to the fungi, but because these carbohydrates are no longer a part of the plant, it is hard to get an accurate estimate of the plant’s productivity. Therefore, we have to be cautious when making conclusions about differences in productivity when we measure only changes in the aboveground biomass to estimate net primary productivity. Measuring CO2 Uptake and Release Because producers take up CO2 during photosynthesis and produce CO2 during respiration, we can measure primary productivity in terrestrial ecosystems by quantifying the uptake and release of CO2 by plants. One way to measure these changes in CO2 is by placing a small plant or a leaf into a sealed container with a highly sensitive CO2 sensor, as illustrated in Figure 20.3. When the container is placed in front of a light that simulates sunlight, the plant consumes CO2 as it conducts photosynthesis. The plant simultaneously produces CO2 as it metabolizes some of its carbohydrates through respiration. Since CO2 uptake from photosynthesis exceeds CO2 release from respiration, the net uptake of CO2 represents net primary productivity.
Figure 20.3 Measuring CO2 uptake and release. (a) In a sealed container illuminated by a light, sensors can detect a decline in CO2 in the air as a leaf conducts photosynthesis. This decline occurs because plant photosynthesis consumes CO2 at a faster rate than plant respiration produces CO2. (b) When the light is turned off, sensors detect an increase in CO2 in the air because the leaf only undergoes respiration. We can also estimate GPP using this technique. Since GPP is the sum of net primary productivity and respiration, we can determine GPP by combining our estimate of NPP with an estimate of how much respiration is occurring in a plant. We can do this by placing leaves or whole plants in chambers without sunlight. If we measure the plant’s rate of respiration in the dark and add it to the plant’s NPP, we can estimate the gross primary productivity using the following rearranged equation: GPP=NPP+Respiration Because these experiments are conducted using some chambers placed in the light and other chambers placed in the dark, they are sometimes referred to as light-dark bottle experiments. Several other techniques can also be used to measure CO2 uptake and release. In one technique, a plant or a leaf is placed in a sealed container and then CO2 containing a rare isotope of carbon, such as 14C, is added to the container. The net movement of 14C from the air to the plant tissues during photosynthesis and from the plant tissues back to the air during respiration can be tracked. This net movement of 14C is a measure of net primary productivity.
On a larger scale, such as a grassland or forest, CO2 uptake and release can be measured using towers that sample the concentration of CO2 at different heights above the ground. The concentration of CO2 moving up from the vegetation, compared to the concentration of CO2 in the atmosphere, gives a measure of the rates of photosynthesis and respiration that are occurring in an area. Measuring O2 Uptake and Release In aquatic ecosystems, the dominant producers are typically algal cells. Because most algae are tiny and rapidly consumed, it is not always feasible to harvest and measure their biomass as a way to measure net primary productivity. Quantifying changes in CO2 concentration is also not a viable way to measure primary productivity because CO2 dissolved in water rapidly converts into bicarbonate ions, as we discussed in Chapter 2. However, because producers release O2 during photosynthesis and take up O2 during respiration, we can estimate NPP and GPP by measuring the uptake and release of O2. To estimate primary productivity using the O2 concentration in the water, we can use a process that is similar to estimating primary production based on CO2. That is, we can conduct a light-dark bottle experiment. We begin by immersing two bottles under the surface of the water to collect the algae. One bottle is clear, allowing sunlight to penetrate. In this bottle, we measure the net increase in O2 production that occurs with the combined effects of photosynthesis and respiration by the algae. The other bottle is opaque, so sunlight cannot penetrate. In this bottle, the algae cannot photosynthesize but can only respire and drive down the concentration of O2. Because the clear bottle measures NPP and the opaque bottle measures only respiration, we can estimate GPP by adding the values obtained from each. Remote Sensing The techniques we have discussed measure gross and net primary productivity on relatively small spatial scales, ranging from a single leaf to a small area of land to a small volume of water. What if we want to assess productivity on very large spatial scales, including changes in productivity
across continents or oceans? One solution to this challenge is remote sensing. Remote sensing is a technique that measures conditions on Earth from a distant location, typically using satellites or airplanes that take photographs of large areas of the globe, as shown in Figure 20.4. These images reveal how different wavelengths of light are reflected or absorbed. As we discussed in
Chapter 3, chlorophyll pigments absorb wavelengths in the red and blue range but reflect wavelengths in the green range. Therefore, satellite images of ecosystems that show a pattern of high absorption of blue and red light and high reflectance of green light indicate ecosystems with high standing crops of producers. Changes in producer biomass over time can then be used to estimate net primary productivity.
Figure 20.4 Remote sensing of primary productivity. Using aerial images from planes or satellites, scientists can determine the wavelengths of light that are absorbed and reflected by the chlorophyll pigments of algal blooms. Here we see satellite images of an algal bloom in Lake Erie. Remote sensing A technique that measures conditions on Earth from a distant location, typically using satellites or airplanes that take photographs of large areas of the globe.
Secondary Production
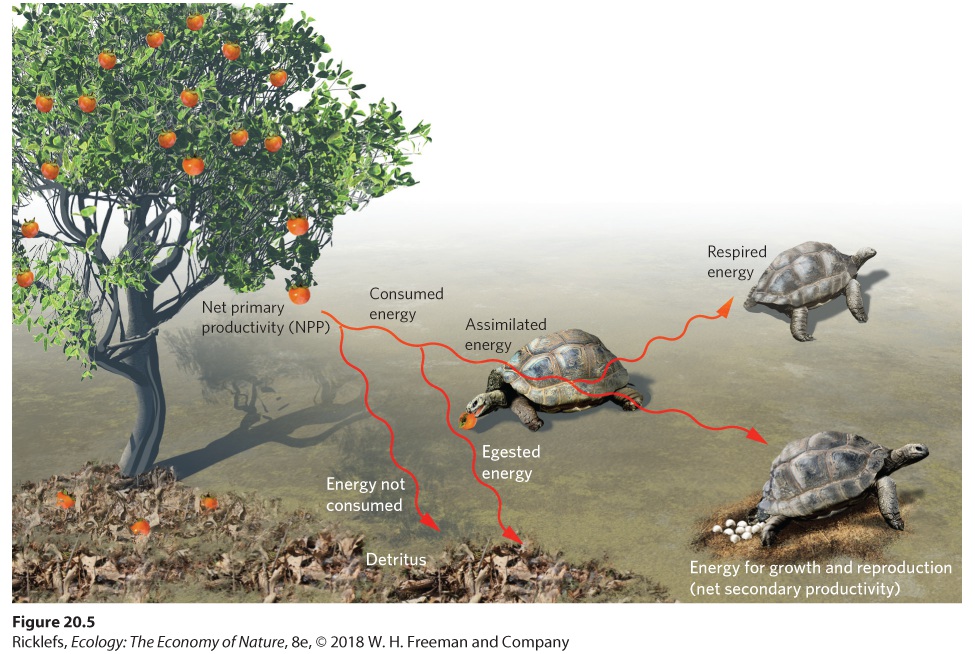
Primary productivity is the foundation of the food web because it represents the source of energy for herbivores. To understand how energy moves from producers to consumers, we need to consider a number of different pathways, shown in Figure 20.5. We begin with producers that herbivores consume. Herbivores consume only a small fraction of the total amount of producer biomass available, and they can only digest a portion of the energy they consume. For example, many fruits contain hard seeds that herbivores cannot digest, so the seeds are excreted whole. We saw an example of this at the end of Chapter 17 when we discussed Aldabra tortoises that had been introduced to an island in the Republic of Mauritius. These tortoises consume fruits of the ebony tree and disperse the seeds when they defecate. The portion of consumed energy that is excreted or regurgitated is known as egested energy. In contrast, the portion of energy that a consumer digests and absorbs is known as assimilated energy.
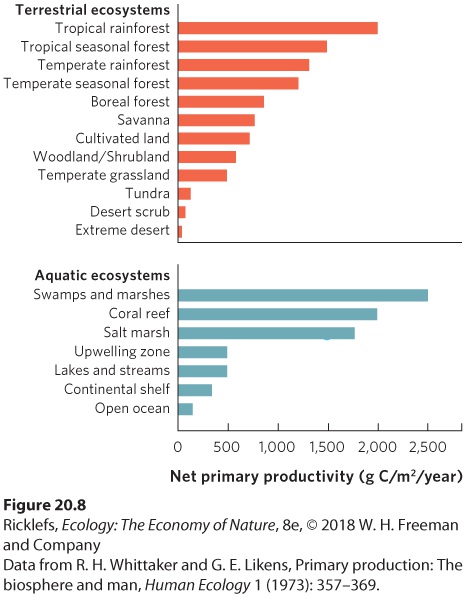
Figure 20.5 The path from primary to secondary productivity. Herbivores gather energy by ingesting producers. Of the energy gained by ingestion, some is lost in the form of egested energy, which represents the indigestible tissues of the producers. The rest of the energy is assimilated. Of the assimilated energy, some is used for respiration. The rest is used for growth and reproduction, which is secondary productivity. Egested energy The portion of consumed energy that is excreted or regurgitated.
Assimilated energy The portion of energy that a consumer digests and absorbs. Of the energy assimilated by a consumer, the portion used for respiration is known as respired energy. The rest can be used for growth and reproduction. For example, consumers such as fruit-eating birds need a great deal of energy for respiration to maintain a constant body temperature, whereas fruit-eating tortoises of the same mass need much less energy for respiration because they are ectotherms. As a result, birds devote more of their assimilated energy to respiration than tortoises do. Because the rate of assimilated energy of consumers is the amount of energy that is used for respiration, growth, or reproduction, it is analogous to the concept of GPP for producers. Respired energy The portion of assimilated energy a consumer uses for respiration. If we consider the assimilated energy of consumers and subtract the energy used for respiration, we are left with the energy used for biomass accumulation; the rate of biomass accumulation of consumers in a given area is called net secondary productivity. Because net secondary productivity depends on primary productivity as its source of energy, we might expect that an increase in net primary productivity would cause an increase in net secondary productivity. Researchers have compiled net primary and net secondary productivity estimates from a wide range of terrestrial biomes. As shown in Figure 20.6a, increases in net primary productivity are positively correlated with increases in net secondary productivity. In aquatic ecosystems, researchers have also found that increases in the productivity of phytoplankton—a dominant producer in many aquatic biomes—are positively correlated to the productivity of the herbivorous zooplankton that eat the algae, which you can see in Figure 20.6b.
Figure 20.6 Secondary productivity. (a) In terrestrial ecosystems, an increase in primary productivity is positively correlated to an increase in secondary productivity. (b) In aquatic ecosystems, a similar relationship is found between the primary productivity of phytoplankton and the secondary productivity of the herbivorous zooplankton that consume the phytoplankton. Net secondary productivity The rate of consumer biomass accumulation in a given area. Quantifying net secondary productivity has many of the same challenges as quantifying net primary productivity, including the need to account for herbivores that are removed from the ecosystem by predators or diseases. In addition, the positive correlation between net primary and secondary productivity suggests an important role of bottom-up control of communities, a concept we discussed in Chapter 18. However, we know that the top-down effects of predators also affect some communities.
Concept Check
1. Why do algae commonly experience a high productivity but a low standing crop? 2. Gross primary production can be used to provide energy for what two processes? 3. What are some key assumptions when measuring the net primary productivity of terrestrial plants?
#### 20.2 Net Primary Productivity Differs Among Ecosystems
20.2 Net primary productivity differs among ecosystems Now that we understand net primary productivity and how it affects the herbivores that consume it, we can examine how net primary productivity varies among ecosystems around the world. Understanding these patterns is important because ecosystems with greater primary productivity should generally support greater secondary productivity, which means that the most productive places will likely have either a high abundance or a high diversity of consumers. In this section, we will investigate patterns of primary productivity in different ecosystems and discuss the abiotic factors that determine these patterns of net primary productivity.
Primary Productivity Around the World
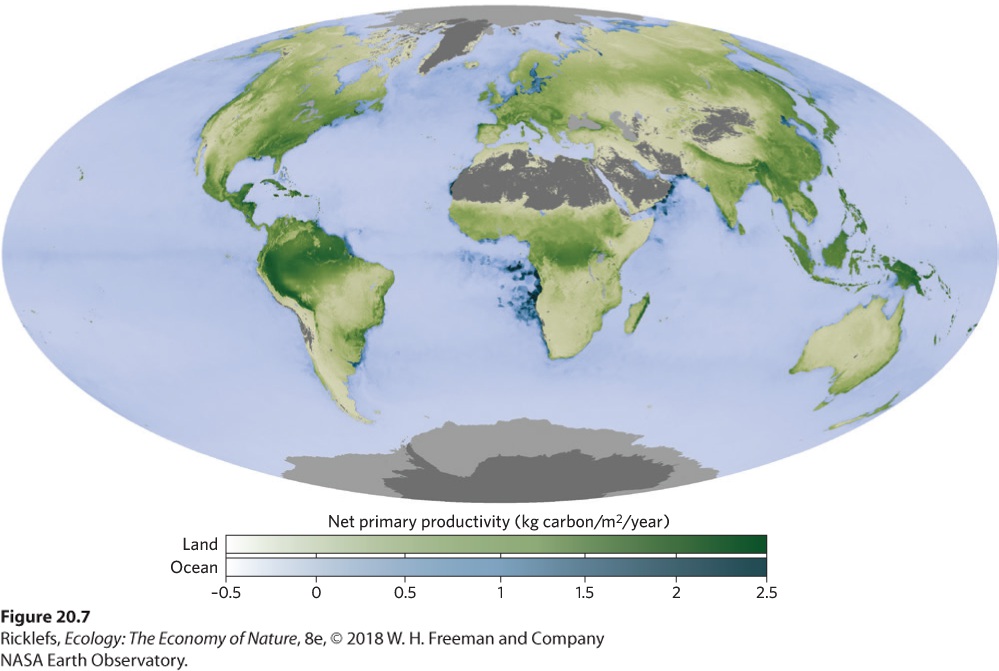
When we look at patterns of NPP around the world in Figure 20.7, we see that net primary productivity varies with latitude. The most productive terrestrial ecosystems occur in the tropics, and productivity declines as we move to temperate and polar regions. The most productive ocean ecosystems are found along coasts, while primary productivity is low in the open ocean.
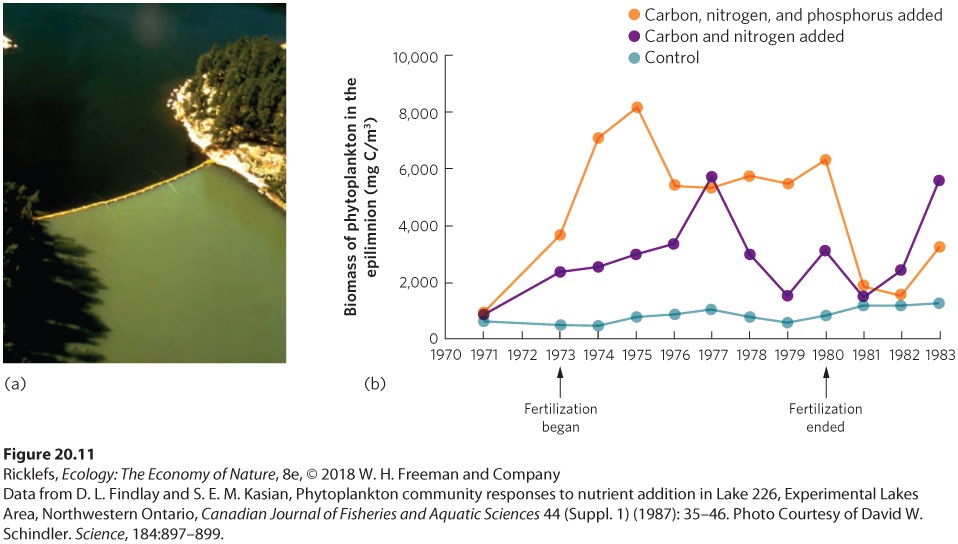
Figure 20.7 Global patterns of net primary productivity. Among terrestrial ecosystems, the highest productivity occurs in the tropics, which experience warm temperatures throughout the year and have abundant rainfall. The most productive aquatic ecosystems include the shallow water near the edges of continents and large islands.
We can also consider the differences in NPP among various aquatic and terrestrial ecosystems. These ecosystem categories coincide well with the biomes described in Chapter 6. Figure 20.8 shows that tropical rainforests are the most productive terrestrial ecosystems. The least productive ecosystems include those that are very cold, such as tundra, and those that are very dry, such as deserts. Among the freshwater ecosystems, lakes and streams, on average, are on the low end of NPP, although NPP can vary widely in these systems. In contrast, marine ecosystems exhibit a wide range of productivity, ranging from high productivity in coral reefs and salt marshes to low productivity in the open ocean. Many abiotic factors are responsible for these patterns of productivity.
Figure 20.8 The net primary productivity of different ecosystems around the world. Among terrestrial ecosystems, tropical rainforests are the most productive, whereas deserts are the least productive. Among aquatic ecosystems, swamps, marshes, and coral reefs are the most productive, whereas the open ocean is the least productive.
Drivers of Productivity in Terrestrial Ecosystems
As you may recall from our discussion of terrestrial biomes in Chapter 6, dominant plant forms are determined by patterns of annual temperature and

precipitation. Similarly, temperature and precipitation are major drivers of NPP. The most productive terrestrial ecosystems occur in tropical areas because those areas have the most intense sunlight, warm temperatures throughout the year, high amounts of precipitation, and rapidly recycled nutrients that support growth. At higher latitudes, such as in temperate and polar regions, productivity is much lower due to shorter periods of sunlight and lower temperatures during the winter. In the deserts that occur at 30° N and 30° S latitude, productivity is primarily constrained by the lack of precipitation. One way to examine the effects of temperature and precipitation on NPP is by looking at a large number of studies that have measured NPP in different parts of the world. For example, Figure 20.9 displays the results of studies from 96 locations. In Figure 20.9a, you can see that an increase in mean annual temperature is positively correlated with an increase in NPP. This reflects the fact that lower latitudes with warmer temperatures favor plant growth and have a longer growing season. In Figure 20.9b, you can see that an increase in mean annual precipitation shows a positive correlation with NPP until 3 m of annual precipitation is reached. Ecosystems that receive 3 m or more of precipitation experience a decline in NPP because nutrients leach away from the soil, and the rates of decomposition are reduced because of waterlogged soils. When organic matter is broken down more slowly, fewer nutrients are available in the soil for primary productivity. In short, while temperature and precipitation are the primary drivers of NPP in terrestrial ecosystems, they also influence the availability of nutrients in ways that affect NPP.
Figure 20.9 The effects of temperature and precipitation on the net primary productivity of terrestrial ecosystems. (a) Locations on Earth that have higher mean annual temperatures have higher NPP. (b) Locations that have higher mean annual precipitation have higher NPP but at locations with the highest amounts of precipitation, increased leaching and reduced decomposition of organic matter cause NPP to decline. Given that nutrients affect NPP in terrestrial ecosystems, which nutrients
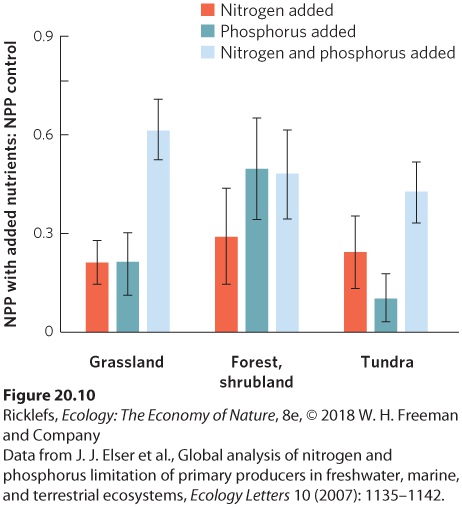
are the most important? For decades, ecologists believed that nitrogen was the most important element that constrained NPP in terrestrial ecosystems. However, ecologists began to discover that some terrestrial ecosystems are also limited by phosphorus or a combination of nitrogen and phosphorus. To obtain a more accurate overview of whether nitrogen or phosphorus limited NPP, researchers compiled data from 141 separate terrestrial experiments that had manipulated nitrogen, phosphorus, or both. For each experiment, they determined the ratio of NPP in treatments with added nutrients, compared to NPP in the control, which had no added nutrients. With a response ratio for each study, the researchers could then determine the average response for all studies in three categories of terrestrial ecosystems: grasslands, forests and shrublands, and tundra. As you can see in Figure 20.10, all three categories of terrestrial ecosystems experienced increases in NPP with the addition of nitrogen or phosphorus. In grasslands and tundra, adding both nitrogen and phosphorus caused a greater increase in NPP than either alone, suggesting that adding more of one nutrient causes the growth of plants to soon be limited by the other nutrient. From these data, we can see that nitrogen and phosphorus are both important nutrients that constrain NPP.
Figure 20.10 Increases in net primary productivity when nutrients are added to terrestrial ecosystems. Based on 141 experiments conducted around the world, researchers examined how NPP responds to different nutrient additions. To quantify the change in NPP,
they determined the ratio of NPP in treatments with added nutrients, compared to NPP in the control, which had no added nutrients. The addition of nitrogen and phosphorus increased the NPP of grassland, forest, shrublands, and tundra ecosystems. When both nutrients were added, the response ratio of NPP was often larger than when either nutrient was added separately. Error bars are standard errors.
Drivers of Productivity in Aquatic Ecosystems
We have seen that terrestrial ecosystems are primarily constrained by temperature, precipitation, and nutrients. In addition to these factors, aquatic ecosystems are also constrained by light since the transmission of light down through the water is required for photosynthesis. Indeed, abundant light is one reason that coral reefs, which exist in shallow tropical waters, are such productive ecosystems. However, within aquatic ecosystems that have similar temperatures and similar light levels, the most important driver of NPP is the amount of nutrients. The limiting role of nutrients can be seen in a variety of aquatic ecosystems. For example, in the open ocean, the remains of dead animals sink to the bottom, where they decompose and release nutrients. Because this regeneration of nutrients is far below the ocean’s surface, the surface of the open ocean experiences low NPP. Small streams also are typically low in nutrients and experience low NPP. Moreover, if the small streams are in a forest, they receive little sun because of the shade from trees, and this restricts their productivity. As we discussed in Chapter 6, a large fraction of the energy and nutrients that exist in small streams enters the stream in the form of allochthonous inputs, such as dead leaves, that drop in from the surrounding terrestrial environment. In contrast, estuaries and coral reefs receive abundant nutrients in the form of runoff from rivers and the adjacent land, which allows these ecosystems to have very high primary productivity. Among all aquatic ecosystems, NPP is most commonly constrained by the availability of phosphorus and nitrogen, although silicon and iron can be limiting in some areas of the open ocean. Limitation by Phosphorus and Nitrogen For many years, it was thought that phosphorus is the most important nutrient that limits the NPP of aquatic ecosystems. For example, in a classic experiment, David Schindler and his colleagues selected an hourglass-shaped lake in Ontario and placed a plastic curtain at the constriction that divided the

lake into two halves (Figure 20.11a). On one side carbon and nitrogen were added, and on the other side carbon, nitrogen, and phosphorus were added. These additions were continued from 1973 to 1980 as the researchers monitored the two sides of the lake. They also monitored a second lake that had an initial fertility similar to the divided lake to serve as a control. In the half of the lake that received carbon and nitrogen, there was a modest increase in NPP compared to the control lake, measured by the growth of cyanobacteria. However, in the half of the lake that received carbon, nitrogen, and phosphorus, there was a large increase in NPP, as shown in Figure 20.11b. After the treatments were terminated in 1980, the NPP of the side of the lake with added phosphorus rapidly declined. This experiment confirmed that human inputs of excess phosphorus—in the form of fertilizers that wash off farms and various household detergents—can have a major effect on the productivity of aquatic ecosystems.
Figure 20.11 Adding phosphorus to half of a lake. (a) Researchers split a Canadian lake in half and then added carbon and nitrogen to one side and carbon, nitrogen, and phosphorus to the other side. (b) Compared to a reference lake that received no nutrient additions, the side that received additions of carbon and nitrogen from 1973 to 1980 experienced a modest increase in NPP, as measured by the biomass of phytoplankton in the epilimnion. The side of the lake that received additions of carbon, nitrogen, and phosphorus experienced a large increase in NPP. When the addition of phosphorus stopped in 1980, NPP in the fertilized side of the lake declined to levels that were similar to those of the control. To gain a broader perspective on how nitrogen and phosphorus affect aquatic ecosystems, let’s revisit the average NPP responses from experiments conducted around the world. As part of the same study discussed earlier,
researchers compiled data from 928 experiments that manipulated the addition of nitrogen, phosphorus, or both in freshwater and marine ecosystems. Among freshwater ecosystems, as shown in Figure 20.12a, nitrogen and phosphorus both caused an increase in NPP, although phosphorus has a much larger effect on the benthic ecosystem of lakes than does nitrogen. Among marine ecosystems, shown in Figure 20.12b, nitrogen and phosphorus additions had similar effects on NPP in ocean ecosystems with soft bottoms, such as estuaries made up of seagrass and algae. However, the addition of nitrogen had a much larger effect on NPP than that of phosphorus in ecosystems with hard bottoms, such as coral reefs and rocky intertidal biomes, and in the open water of oceans. Across all freshwater and marine ecosystems, adding nitrogen and phosphorus together often caused an NPP response that was much larger than adding either nutrient separately. Once again, this suggests that adding more of one nutrient causes the growth of producers to soon be limited by the other nutrient. The results of this research suggest that the availability of nitrogen and phosphorus can constrain the NPP of aquatic ecosystems, as well as that of terrestrial ecosystems.
Figure 20.12 Increases in net primary productivity when nutrients are added to aquatic

ecosystems. Using data from 928 experiments conducted around the world, researchers determined how NPP responds to different nutrient additions. NPP was examined as a ratio between productivity when nutrients were added and a control treatment in which nutrients were not added. In (a) freshwater and (b) marine ecosystems, adding nitrogen or phosphorus caused an increase in NPP, although nitrogen has a larger effect than phosphorus in the softbottom-ocean ecosystems and the ocean–water ecosystems. Error bars are standard errors. Limitation by Silicon and Iron in the Ocean While the primary productivity of aquatic ecosystems is typically limited by the availability of nitrogen and phosphorus, primary productivity remains low in approximately 20 percent of the open ocean even though nitrogen and phosphorus are abundant there. This suggests that productivity in these areas is limited by other nutrients that are unusually scarce, such as silicon and iron. Silicon is the raw material for the silicate shells of diatoms (see Figure 2.5), which comprise most of the phytoplankton in some regions of the ocean. Silicon is lost from the surface waters when diatoms die and their dense shells fall to the ocean bottom. For example, the area west of southern South America, between 40° S and 50° S latitude, appears to have too little silicon, probably because silicon-containing particles sink below the photic zone more rapidly than do nitrogen and phosphorus across this long stretch of the southern Pacific Ocean. Iron is an important component in many metabolic pathways, but it is lost from the ocean’s surface when it combines with phosphorus and precipitates. Rivers are a source of iron, which explains the low concentrations of iron in regions of the ocean that are far from continents. In these areas, the only iron inputs occur in the form of windblown dust. If iron limits the growth of phytoplankton in the ocean, then additions of iron should act as a fertilizer and cause large increases in primary productivity. In a 1993 experiment, 450 kg of iron—roughly the amount in an automobile—was added to over 64 km2 of ocean off the west coast of South America. The treatment caused a 100-fold increase in the concentration of iron and, within just a few days, the concentration of phytoplankton tripled. Although this increase in phytoplankton was of a short duration, the experiment confirmed that a lack of iron limited the producers in this area of the ocean. In subsequent experiments conducted off the coast of Antarctica, similar increases in productivity were observed (Figure 20.13).
Figure 20.13 Fertilizing the oceans. In the Southern Ocean near Antarctica, researchers lower water-sampling equipment to measure the effects of adding iron to the surface waters. Fertilizing the ocean with iron has implications beyond confirming that iron is the limiting nutrient in these areas. If primary productivity in the ocean can be increased, it has the potential to draw down the amount of CO2 in the water. Because CO2 is exchanged between the water and the air, reducing CO2 in the water would subsequently reduce the amount of CO2 in the atmosphere. Since CO2 is a greenhouse gas, some scientists have hypothesized that fertilizing the oceans with iron could be a way to counteract global warming. Although adding iron increased primary production, in some cases the large increase in phytoplankton caused a subsequent large increase in the zooplankton populations that consume phytoplankton. When this happened, the zooplankton that was produced caused an increase in the amount of CO2 due to increased respiration, thereby counteracting the beneficial effects of the phytoplankton. When researchers conducted experiments that monitored the amount of carbon that precipitated out of the water column during iron additions, they found that there was only a small increase in the amount of carbon that precipitated out of the water column. Other researchers have examined historic variation in iron inputs on ocean
productivity. During five major glacial periods, about 2.5 times more iron dust traveled through the atmosphere and arrived in the Pacific Ocean near the equator. In 2016, researchers reported their results of examining cores from the ocean floor in this region of the Pacific, which consists of layers of sediments deposited over centuries. They found no increase in productivity during past periods of increased iron availability due to increased iron dust. Based on the observations from experiments and historic data, the long-term effects of iron enrichment on CO2 remain uncertain, as does the question of whether large-scale applications of iron in the ocean could have potential adverse effects on the ocean ecosystem.
Concept Check
1. Which ecosystems have the highest NPP? 2. Why does adding a combination of nitrogen and phosphorus often result in higher producer growth than adding either nutrient alone? 3. Why have researchers hypothesized that adding iron to certain areas of the ocean might help offset elevated CO2 concentrations in the atmosphere?

#### 20.3 the Movement of Energy Depends on the Efficiency of Energy Flow
20.3 The movement of energy depends on the efficiency of energy flow As we have discussed, the organisms in each trophic level must obtain energy, assimilate some fraction of the energy they secure, and then use some of the assimilated energy for respiration. Whatever remains goes to growth and reproduction. At each step, different organisms vary in the efficiency with which they obtain and retain energy. These efficiencies, combined with trophic interactions that affect the abundance of other trophic levels, affect the amount of energy and biomass that can be supported in a given trophic level. These factors also affect how many trophic levels can exist in an ecosystem. In this section, we will explore patterns in the amounts of energy and biomass that exist in different trophic levels. We will then examine the various efficiencies of energy transfer in trophic groups and how these differences affect the number of trophic levels found in an ecosystem.
Trophic Pyramids
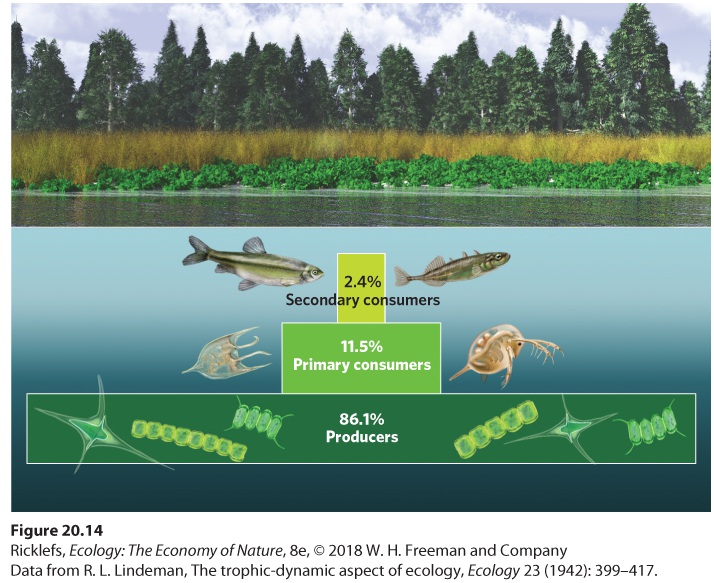
A useful way to think about the distribution of energy or biomass among the trophic groups in an ecosystem is by drawing a trophic pyramid, which is a chart composed of stacked rectangles representing the relative amount of energy or biomass in each trophic group. The first person to consider the flow of energy between trophic levels was Raymond Lindeman, an ecologist who conducted his doctoral dissertation on Cedar Bog Lake in Minnesota during the 1930s. As Lindeman collected data on the energy and biomass produced by different trophic levels in the lake, he concluded that energy must be lost as it moves from one trophic level to the next. He demonstrated that this was the case when he created a trophic pyramid that displayed the percentage of the total energy existing at each trophic level, known as a pyramid of energy. As you can see from his data in Figure 20.14, each trophic level in the lake contained less energy than the trophic level below it.

Figure 20.14 The energy pyramid of Cedar Bog Lake. Most of the energy assimilated in this lake ecosystem is found in producers such as algae. Because energy is lost as it is converted from one trophic level to the next, considerably less energy is found in primary consumers such as zooplankton and secondary consumers such as fish. Trophic pyramid A chart composed of stacked rectangles representing the amount of energy or biomass in each trophic group. Pyramid of energy A trophic pyramid that displays the total energy existing at each trophic level. Pyramids of energy are just one way of representing the distribution of organisms in an ecosystem. We can also create a trophic pyramid that represents the standing crop of organisms present in different trophic groups, which is known as a pyramid of biomass. In terrestrial ecosystems, the distribution of biomass among trophic levels looks quite similar to the pyramid of energy. As Figure 20.15a shows, the greatest amount of biomass occurs in producers, with less biomass in primary and secondary consumers.
Figure 20.15 Pyramids of energy and biomass. (a) In terrestrial ecosystems, pyramids of energy and biomass have similar shapes because most of the energy and most of the standing biomass are found in the producers. (b) In aquatic ecosystems, most of the energy is still found in the producers, but these producers are primarily tiny algae that do not live very long because they are rapidly consumed by herbivores. This continual rapid consumption results in a large biomass of consumers in these systems. Pyramid of biomass A trophic pyramid that represents the standing crop of organisms present in different trophic groups. For example, consider the biomass in a forest ecosystem. The greatest portion of the biomass occurs in producers, which include trees, shrubs, and wildflowers. There is considerably less biomass in primary consumers, which include herbivorous birds, mammals, and insects. There is even less biomass in secondary consumers, which include hawks, owls, and carnivorous mammals. A similar scenario exists in the grassland ecosystem of Africa. All the grass in Africa piled together would dwarf a mound of all the grasshoppers, gazelles, zebras, wildebeests, and other animals that eat grass. That mound of herbivores, in turn, would dwarf the relatively tiny mound of all the lions, hyenas, and other carnivores that feed on them. In aquatic ecosystems, the pyramid of biomass has a very different shape. In these ecosystems, the major producers are the phytoplankton, the tiny
algae that float or swim through the water. Unlike trees and shrubs, algae have short lives with rapid reproduction, and they are consumed in large numbers. As a result, although the productivity of algae is much higher than the consumers of algae, the standing biomass of algae is often much lower than the standing biomass of the consumers of algae. This creates an inverted pyramid, as illustrated in Figure 20.15b.
The Efficiencies of Energy Transfers
As we discussed, the amount of energy that moves from one trophic level to another determines how much energy or biomass can exist at each trophic level. The amount of energy that is transferred between adjacent trophic levels depends on several steps that occur within each trophic level and include consumption, assimilation, and production. When we consider the transfer of energy from one trophic level to another, we can quantify it as the percentage of available energy that is transferred, which is a measure of efficiency. Consumption Efficiency Turning back to Figure 20.4, we see that the first step in the transfer of energy from one trophic level to another is the consumption of energy from the lower trophic level. Some of the total amount of energy available in the lower trophic level is consumed and the rest is left to become dead organic matter. The percentage of energy or biomass in a trophic level that is consumed by the next higher trophic level is known as the consumption efficiency. Consumption efficiency is calculated using the following equation: Consumption efficiency=Consumed energy(J)Net production energy of thenext lower trophic level(J) Consumption efficiency The percentage of energy or biomass in a trophic level that is consumed by the next higher trophic level. For example, there might be 10 J of energy available in a field of wildflowers, but the herbivores might consume only 1 J of that energy
because many of the plant species possess anti-herbivore defenses. In this case, the consumption efficiency of the herbivores would be 10 percent. In ecosystems containing producers with fewer defenses against herbivores, the consumption efficiency would be much higher and more energy would enter the consumer trophic level. Assimilation Efficiency As we have discussed, energy that is consumed is subsequently either assimilated or egested. In the case of plants, especially terrestrial plants, many components such as cellulose and lignin are not easily digested. Similarly, the feathers, bones, exoskeletons, and hair of animals are not easily digestible by the predators that consume them. Owls provide an excellent example. All species of owls are predators that commonly feed on small mammals. They swallow their prey whole, digest it, and then regurgitate the hair and small bones in the form of compact pellets (Figure 20.16).
Figure 20.16 Undigested energy. (a) Owls such as this barn owl (Tytoalba) consume small rodents but they do not digest the hair and bones. (b) Instead, owls regurgitate a pellet that contains these indigestible materials. Consumers that eat large amounts of indigestible materials have low assimilation efficiencies. The percentage of consumed energy that is assimilated is known as the assimilation efficiency and is calculated using the following equation: Assimilation efficiency=Assimilated energy(J)Consumed energy(J) Assimilation efficiency The percentage of consumed energy that is assimilated.
Assimilation efficiency varies a great deal from one trophic level to another. For example, herbivores that feed on seeds, such as many species of birds and rodents, have assimilation efficiencies as high as 80 percent. In contrast, herbivores, such as horses that feed on grasses and woody vegetation, have assimilation efficiencies of only 30 to 40 percent. Secondary consumers, which are predators on the herbivores, typically have high assimilation efficiencies, ranging from 60 to 90 percent. These high assimilation efficiencies occur because consumed animal tissues are generally more digestible than consumed plant tissues. Net Production Efficiency Ultimately, we want to know how much assimilated energy is converted into the growth and reproduction of the organisms in a trophic level. The percentage of assimilated energy that is used for growth and reproduction is the net production efficiency, which is calculated using the following equation: Net production efficiency=Net production energy(J)Assimilated energy(J) Net production efficiency The percentage of assimilated energy that is used for growth and reproduction. Put another way, net production efficiency is the percentage of assimilated energy that remains after respiration. For active homeothermic animals that must spend a large fraction of their energy to maintain a constant body temperature, move about, circulate their blood, and balance their salts, net production efficiency can be as low as 1 percent. In contrast, sedentary poikilothermic animals, particularly aquatic species, channel as much as 75 percent of their assimilated energy into growth and reproduction. Understanding net production efficiency also has practical applications. For example, livestock producers understand that if they keep their animals indoors at high densities during the winter, the animals will spend less of their assimilated energy on respiration to maintain a constant body temperature. As a result, the animals can devote more of their assimilated energy to growth, and faster-growing livestock result in more income for the
livestock producer. Ecological Efficiency Now that we understand the three efficiencies that occur within each trophic level, we can explore the efficiency of energy transfer between adjacent trophic levels. Ecological efficiency, also known as food chain efficiency, is the percentage of net production from one trophic level, compared to the next lower trophic level: Ecological efficiency=Net production energy of a trophic level (J)Net production energy of the next lower trophic level (J) Ecological efficiency The percentage of net production from one trophic level, compared to the next lower trophic level. Also known as Food chain efficiency. When we consider all the efficiencies in the chain of events that starts with consumption and ends with net production, we can see that each plays a role in determining the ecological efficiency between trophic levels. To confirm that this is true, we can multiply the efficiencies in each step of the process to produce the equation for ecological efficiency: Ecological efficiency=Consumed energy (J)×Assimilation efficiency×Net production efficiencyEcological efficiency=Consumed energy (J)Net production energy of thenext lower trophic level (J)×Assimilated energy (J)Consumed energy (J)×Net production energy (J)Assimilated energy (J)Ecological efficiency=Net production energy (J)Net production energy of thenext lower trophic level (J) Because energy is lost at each of these steps, ecological efficiencies are typically quite low and range from 5 to 20 percent. Given this range, ecologists often use 10 percent as a rule of thumb. Such low ecological
efficiencies help us understand why each trophic level in the pyramid of energy becomes much smaller as we move from producers to primary consumers to secondary consumers. An ecological efficiency of 10 percent between adjacent trophic levels means that only 10 percent of the total energy present in the producer trophic level will be found in primary consumers, and only 1 percent will be found in secondary consumers. As you can see, an ecological efficiency of 10 percent makes it difficult to have long food chains in an ecosystem because there is not enough energy to support additional higher trophic levels. The only way to support additional trophic levels is by increasing the amount of energy at the producer level or by increasing the ecological efficiencies moving between adjacent trophic levels. A 10 percent ecological efficiency also applies to humans and where we feed along a food chain. For example, when we act as a primary consumer and eat plants, we can assimilate 10 percent of the energy available from plants. However, when we act as a secondary consumer and consume herbivores, we assimilate 1 percent of the original energy that was in the plants. As a result, we would expect a diet composed of more plants and less meat to dramatically increase the amount of food available to humans. Ecological Efficiency and the Number of Trophic Levels Aquatic ecosystems typically have more trophic levels than terrestrial ecosystems, in part because of differences in ecological efficiencies. In terrestrial ecosystems, producers are composed primarily of plants that range in size from wildflowers to trees. In many of these plants, a large proportion of tissue is dedicated to deterring herbivores from consuming them. Other plants contain an enormous proportion of biomass that herbivores cannot consume, such as the wood of trees. As a result, there is low consumption efficiency in terrestrial ecosystems, so a large fraction of the producer biomass ultimately becomes detritus. In contrast, aquatic ecosystems are composed primarily of unicellular algae that contain fewer defenses and are relatively easy for herbivores to digest. Therefore, algae provide higher consumption and assimilation efficiencies to their herbivores, which leads to higher ecological efficiencies. These higher ecological efficiencies mean that a higher fraction of the ecosystem’s energy can move up the food chain and
support additional trophic levels in aquatic ecosystems, compared to terrestrial ecosystems. In addition to having higher ecological efficiencies, aquatic ecosystems also contain herbivores that are generally quite small, such as the tiny zooplankton that consume algae. These zooplankton are consumed by secondary consumers, such as small species of fish (Figure 20.17). These small fish are consumed by medium fish, which in turn are consumed by large fish. Because the producers are mostly single-cell algae, and because each successive consumer is only a bit larger than the diet it consumes, aquatic ecosystems can often contain five trophic levels.
Figure 20.17 Aquatic ecosystems begin with small herbivores, which are consumed by small fish that are consumed by larger fish. As a result, aquatic ecosystems can often have more trophic levels than terrestrial ecosystems. We can contrast this with the situation that occurs in terrestrial ecosystems. In terrestrial ecosystems, the producers are relatively large, ranging in size from wildflowers to large trees. In addition, many of the major herbivores, such as deer and antelope, are quite large and can be consumed only by very large secondary consumers, such as wolves and lions. Because terrestrial ecosystems contain many large producers and large herbivores, they are less likely to have a fourth and fifth trophic level. In short, the low ecological efficiency of terrestrial ecosystems combined with the large size of many
##### Analyzing Ecology: Quantifying Trophic Efficiencies
producers and herbivores results in terrestrial ecosystems that commonly contain only three or four trophic levels.
Analyzing Ecology
Quantifying Trophic Efficiencies Trophic efficiency calculations derive from data collected on consumption efficiency, assimilation efficiency, net production efficiency, and ecological efficiency. Energy Terrestrial Ecosystem Lake Ecosystem Stream Ecosystem Net Production Available In Lower Trophic Level (J) 1,000 1,000 Consumed Energy (J) Assimilated Energy (J) Net Production Energy (J) We can calculate these efficiencies using the data from the terrestrial ecosystem. Consumption efficiency=Consumed energy (J)Net production energy of thenext lower trophic level (J)Consumption efficiency=250 J1,000 J=25%Assimilation efficiency=Assimilated energy (J)Consumed energy (J)Assimilation efficiency=120 J250 J=48%Net production efficiency=Net production energy (J)Assimilated energy (J)Net production efficiency=50 J120 J=42%Ecological efficiency=Net production energyof a trophic level (J)Net production energy ofnext lower trophic level (J)Ecological efficiency=50 J1,000 J=5%
YOUR TURN Use the data for the lake and stream ecosystems to calculate the four different efficiencies for each ecosystem. Based on these calculations, why do the two aquatic ecosystems have higher ecological efficiencies than the terrestrial ecosystem? Which efficiencies cause the stream ecosystem to have lower net production energy than the lake ecosystem?
Residence Times
Ecological efficiency tells us the proportion of energy that moves from producers into the higher trophic levels. We also want to examine the rate of energy movement between trophic levels, which tells us how long energy remains in a given trophic level and, therefore, how much energy can accumulate at that level. We can define the length of time that energy spends in a given trophic level as the energy residence time. Energy residence time is directly related to the amount of energy that exists in a trophic level; the longer the residence time, the greater the accumulation of energy in that trophic level. The average residence time of energy at a particular trophic level equals the energy present in the tissues of organisms divided by the rate at which energy is converted into biomass, or net productivity: Energy residence time(years)=Energy present in atrophic level (J/m2)Net productivity(J/m2/year)
Energy residence time The length of time that energy remains in a given trophic level. If we substitute biomass for energy in this equation, we can determine biomass residence time, which is the length of time that biomass spends in a given trophic level: Biomass residence time(years)=Biomass present in atrophic level(kg/m2)Net productivity(kg/m2/year) Biomass residence time The length of time that biomass remains in a given trophic level. For example, plants in humid tropical forests produce dry matter at an average rate of 1.8 kg/m2/year and have an average living biomass of 42 kg/m2. Inserting these values into the preceding equation gives a biomass residence time of 23 years. Average biomass residence times for primary producers range from more than 20 years in forest ecosystems to less than 20 days in aquatic phytoplankton-based ecosystems. The much shorter biomass residence time in aquatic ecosystems is the reason these ecosystems frequently have inverted pyramids of biomass, like the one shown in Figure 20.15b; the biomass produced by algae is rapidly consumed by zooplankton. These residence times track the movement of energy that is consumed from one trophic level to another by primary, secondary, and tertiary consumers, but these estimates do not take into account the residence time of the dead organic matter that is consumed by scavengers, detritivores, and decomposers. We calculate the residence time in dead organic matter by using a variation of the equation for energy residence time: Dead organic matter residence time(years)=Dead organic matter presentin a trophic level(kg/m2)Dead organic matterproductivity(kg/m2/year)
For example, the residence time of dead leaf litter is 3 months in humid tropical ecosystems, 1 to 2 years in dry tropical ecosystems, 4 to 16 years in temperate forest ecosystems in the southeastern United States, and more than 100 years in temperate mountains and boreal ecosystems (Figure 20.18). As you can see from the preceding equation, these differences in the residence times of leaf litter are a function of how much litter falls each year and how rapidly decomposition can occur. As we discussed in our tour of the biomes in Chapter 6, warm temperatures and abundant moisture allow the rapid decomposition of litter in lowland tropical regions, while colder and drier conditions cause slow decomposition and litter accumulation in temperate and boreal ecosystems.
Figure 20.18. Residency times for dead organic matter. (a) In tropical forests, such as this site in Thailand, warm temperatures and high precipitation cause a rapid breakdown in dead organic matter. (b) In temperate forests, such as this site in Ohio, the colder temperatures cause a slower breakdown of dead organic matter and therefore a longer residency time.
Stoichiometry
In addition to obtaining energy, organisms also must have the correct balance of nutrients to grow and reproduce. Ideally, the ratio of nutrients an organism needs must match the ratio of nutrients it consumes, but sometimes this is a challenge. The study of the balance of nutrients in ecological interactions, such as between an herbivore and a plant, is called ecological stoichiometry. Understanding ecological stoichiometry is useful in explaining variation in the ecological efficiencies that we have just discussed. Ecological stoichiometry The study of the balance of nutrients in ecological interactions, such as between an herbivore and a plant.
The balance of nutrients required by different species depends on their biology. For example, the composition of an organism’s body can affect the types of nutrients it needs to obtain. Diatoms have high requirements for silicon because they produce glass shells for protection (see Figure 2.5), whereas vertebrates require large amounts of calcium and phosphorus for growing bones and scales. In the case of birds and mammals that primarily consume fruits, which are typically low in calcium and phosphorus, they often must supplement their diets with snail shells or bits of limestone to take in sufficient amounts of calcium and phosphorus. Growth rates and other life-history traits also can influence the nutrient composition of organisms. For instance, if we compare two types of zooplankton, we find that slowly growing marine copepods (Figure 20.19a) have nitrogen-to-phosphorus ratios as high as 50:1, whereas rapidly growing freshwater water fleas (Figure 20.19b) have ratios below 15:1. The fastergrowing water fleas have a lower ratio because they must maintain a higher concentration of phosphorus in their tissues to synthesize the large amounts of proteins necessary for rapid growth.
Figure 20.19 Effects of growth rate on stoichiometry. (a) Slow-growing organisms, such as this marine copepod, have tissues with a high ratio of nitrogen to phosphorus of approximately 50:1. (b) Fast-growing organisms, such as this water flea (Daphnia magna), require much more phosphorus so that they can produce nucleic acids. As a result, their tissues have a lower ratio of nitrogen to phosphorus, approximately 15:1. To understand how stoichiometry affects the efficiencies of energy transfer between adjacent trophic groups, consider again the water flea that has tissues containing a 15:1 ratio of nitrogen and phosphorus. This becomes challenging when the ratio of ingested nutrients in the algae does not match the ratio of nutrients needed by the water fleas. When this occurs, consumers must process larger amounts of food to obtain sufficient amounts of the most
limiting nutrient. If the water flea consumed algae that contained a 30:1 ratio of nitrogen and phosphorus, it would have to consume twice as much algae to meet its phosphorus need. Moreover, when it consumed algae containing a 30:1 ratio, it would have to excrete the excess nitrogen. In this example, the assimilation efficiency of the water flea, in terms of nitrogen, declines and therefore the ecological efficiency of the water flea’s trophic level declines. As you can see, tracking the ecological stoichiometry helps us understand why there can be a low ecological efficiency whenever a nutrient-poor producer is consumed by an herbivore that requires a nutrient-rich diet. In short, understanding ecological stoichiometry helps us explain why ecological efficiencies can vary between trophic levels. In this chapter, we have explored how energy moves through ecosystems. We have seen that this process begins with producers capturing the energy of the Sun and converting it into producer biomass. The rate of primary production influences the rate of production for higher trophic levels. We have seen that the amount of primary productivity differs among ecosystems around the world because of variations in temperature, precipitation, light, and nutrients. The amount of energy that passes through the trophic levels of ecosystems depends on the efficiency of each step in the chain, with all the steps producing an overall ecological efficiency between adjacent trophic levels and different residence times of energy, biomass, and dead organic matter.
Concept Check
1. Why does the energy pyramid exhibit large reductions in energy level as we move up through the trophic levels? 2. What are the two possible fates of assimilated energy in a primary consumer? 3. Why does organic matter accumulate in cold, dry biomes?
Concepts
Feeding an Ocean of Whales Humpback whales. Researchers have determined the number of whales living off the West Coast of the United States, such as this humpback whale. When whale numbers were combined with data on the diet of each species and the ecological efficiencies between adjacent trophic levels, the researchers estimated that whales consume approximately 12 percent of the NPP of the ocean off the West Coast of the United States. The ocean contains a diversity of species feeding at different trophic levels, and whales sit at or near the top trophic level. Although whales are a top predator, it has been a long-standing challenge for us to understand how much NPP is required to provide the energy that whale populations need. Knowing the NPP requirement would tell us how much energy passes through this trophic level. It would also allow us to estimate how much NPP whales used prior to the commercial hunting that has caused many whale populations to decline during the past century. The
reduction in consumed NPP could potentially be consumed by other species in the ecosystem. Given that most whales live in the open ocean, how does one begin to estimate the percentage of NPP that whales consume? A group of researchers decided to tackle this challenge by starting with information about the whales and working down to the producers. First, they settled on a particular study area: the region of ocean within 550 km from the western coast of the United States. Next, they estimated how many whales lived in this area. Fortunately, line transects had been conducted in this region of the ocean for 15 years and these transects provided abundance estimates for 21 species of whales that included dolphins, porpoises, sperm whales (Physetermacrocephalus), and humpback whales (Megapteranovaeangliea). By knowing the typical mass of an individual whale for each species and how much energy whales of different sizes consume, assuming an assimilation efficiency of 80 percent, the researchers could estimate the total energy consumed annually by each species of whale. The next step was to determine the diet of each whale species. Each species has a unique diet of prey made up of different proportions of primary consumers, such as krill, and secondary consumers, such as fish. Using existing data on whale diets, the researchers knew the proportion of primary and secondary consumers that each whale species eats. They then used the ecologist’s rule of thumb that adjacent trophic levels passed along energy with an ecological efficiency of 10 percent. Based on this rule of thumb, they could determine how much NPP was required to produce the prey consumed by the whales.
The final step was to determine how much NPP was available in this region of the ocean. They measured NPP using satellites and the remote sensing techniques for detecting chlorophyll concentrations that we discussed earlier in the chapter. Once NPP was quantified, they determined that whales consumed about 12 percent of the ocean’s NPP. For comparison, this is about half of the NPP required to support fish populations that are commercially harvested. The percentage of NPP consumed by this group of mammals is likely to increase dramatically in the coming decades. For example, during the 1980s and early 1990s, many dolphins were accidentally killed by commercial fishing operations. Because this source of mortality has declined in recent years, we anticipate an increase in the number of some types of whales, such as dolphins. This may cause a higher proportion of the NPP to be consumed. Moreover, because many of the larger species of whales are now protected around the world, their populations are rising. Scientists expect populations of some of the larger species to eventually triple or quadruple, which will result in an even greater consumption of the ecosystem’s NPP. A current challenge is estimating how this large increase in the biomass of top predators will affect the ocean’s distribution of energy among the trophic levels. An insight on this question came in 2014 when researchers reported the results of their studies on whales transporting nutrients. For many years, the paradigm had been that whales consume a great deal of primary productivity and, as a result, may be competing with recreational anglers and commercial fishing operations. By radio-tagging whales to
see where they travel, the researchers discovered that many whales are hunting for food in deep waters and then defecating when they come up to the surface. As a result, the whales are transporting nutrients from the deep waters to the surface waters, where their egested food can provide nutrients for the algae. The researchers noted that this “whale pump” of nutrients might provide a large benefit to the surface-water food web and help feed the fish that humans catch and pursue. As whale populations continue to recover, these plumes of egested nutrients should be even more abundant, and we will continue to learn about the key role that whales play in ocean ecosystems. SOURCES: Barlow, J., et al. 2008. Cetacean biomass, prey consumption, and primary production requirements in the California Current ecosystem. Marine Biology Progress Series 371: 285–295. Roman, J., et al. 2014. Whales as ecosystem engineers. Frontiers in Ecology and the Environment 12: 377–385.
Summary of Learning Objectives
20.1 Primary productivity provides energy to the ecosystem. Primary productivity is the process of capturing solar or chemical energy and converting it into chemical bonds by photosynthesis or chemosynthesis over a given amount of time. We can distinguish between gross primary productivity, which is the total amount of energy assimilated, and net primary productivity, which is the assimilated energy that is converted into producer biomass. Net primary productivity can be measured in numerous ways, including measuring plant biomass, measuring CO2 uptake and release in terrestrial ecosystems, measuring O2 uptake and release in aquatic ecosystems, and using remote sensing. Across all ecosystems, the amount of net primary productivity has a direct positive relationship with the amount of net secondary productivity. Key Terms: Primary productivity, Standing crop, Gross primary productivity (GPP), Net primary productivity (NPP), Remote sensing, Egested energy, Assimilated energy, Respired energy, Net secondary productivity
20.2 Net primary productivity differs among ecosystems. Net primary productivity differs a great deal among ecosystems around the world. In terrestrial ecosystems, major drivers of this productivity include temperature, precipitation, nitrogen, and phosphorus. In aquatic ecosystems, major drivers include temperature, light, nitrogen, and phosphorus.
20.3 The movement of energy depends on the efficiency of energy flow. The energy of ecosystems exists in different trophic levels and moves between these trophic levels with different efficiencies. Pyramids of energy exhibit similar distributions among ecosystems, with producers having the most energy and each higher trophic group possessing less energy. Pyramids of biomass show a
similar trend in terrestrial ecosystems, but they are often inverted in aquatic ecosystems. To understand how energy moves between trophic levels, we can calculate the consumption efficiency, assimilation efficiency, and net production efficiency, all of which can be multiplied to determine the overall ecological efficiency of transferring energy between adjacent trophic groups. The efficiencies can be affected by the stoichiometry of consumer tissues relative to the consumer’s diet and can affect the number of links in an ecosystem’s food chain. Efficiencies can also affect the residence time of energy and biomass in ecosystems. Key Terms: Trophic pyramid, Pyramid of energy, Pyramid of biomass, Consumption efficiency, Assimilation efficiency, Production efficiency, Ecological efficiency, Energy residence time, Biomass residence time, Ecological stoichiometry
Critical Thinking Questions
1. Why is the efficiency of energy transfer between grasses and gazelles quite low? 2. How would you distinguish between gross primary productivity and net primary productivity in a desert ecosystem? 3. Compare and contrast the measurement of primary productivity in terrestrial versus aquatic ecosystems. 4. Compare and contrast the factors that limit the net primary productivity in terrestrial versus aquatic ecosystems. 5. How might remote sensing be used to track changes in the standing crop of aquatic ecosystems in response to global warming? 6. Why might assimilation efficiencies be much higher for herbivores eating seeds than for herbivores eating leaves? 7. What is the likely shape of the pyramid of biomass in a lake? 8. Why would you expect larger changes in stoichiometry between predators and prey to alter assimilation efficiencies? 9. Why are residence times much longer in forest ecosystems than in aquatic phytoplankton-based ecosystems? 10. How might the carrying capacity for people in the United States change if we ate more plant products than animal products?
#### Graphing the Data: NPP Versus the Total Primary Productivity of Ecosystems
GRAPHING THE DATA NPP Versus the Total Primary Productivity of Ecosystems
Figure 20.7 shows estimates of NPP for various ecosystems around the world. However, these data tell us only the amount of primary production per square meter. We can also examine the amount of primary production in terms of the total amount produced in a year among the different ecosystems. Using the data on NPP and area in the table, calculate total production for each ecosystem by multiplying NPP and area. Then plot the total production values using a bar graph. Discuss how the NPP and the area of the different ecosystems affect total production. SOURCE: Data from Whittaker, R. H., and G. E. Likens. 1973. Primary production: The biosphere and man. Human Ecology 1: 357–369. Terrestrial Ecosystems Npp (g/m2/year) Area (106 km2) Total Production (1012 kg/year) Tropical Rainforest 2,000 17.0 Tropical Seasonal Forest 1,500 7.5 Temperate Rainforest 1,300 5.0 Temperate Seasonal Forest 1,200 7.0 Boreal Forest 12.0 Savanna 15.0 Cultivated Land 14.0 Woodland/Shrubland 8.0 Temperate Grassland 9.0 Tundra 8.0 Desert Shrub 18.0
Aquatic Ecosystems Npp (g/m2/year) Area (106 km2) Total Production (1012 kg/year) Swamp and Marsh 2,500 2.0 Coral Reef 2,000 0.6 Salt Marsh 1,800 1.4 Upwelling Zones 0.4 Lake and Stream 2.5 Continental Shelf 26.6 Open Ocean 332.0