22 Landscape Ecology and Global Biodiversity
22Landscape Ecology and Global Biodiversity Species diversity of flowering plants in California. While California has experienced 28 extinctions of native plants, they have also experienced introductions of more than 1,000 nonnative plants from around the world. Can We Have Too Much Biodiversity? The word biodiversity prompts several thoughts. We think of the diversity of evolved species that play unique roles in nature and collectively contribute to the proper functioning of ecosystems. We also may think about the many species around the world that are declining in abundance and some that are even going extinct. What is less appreciated is that while many places on Earth are losing species, these same places are simultaneously experiencing large increases in biodiversity. The reason for this apparent contradiction is that the number of extinctions is often far

outweighed by the number of introduced, non-native species. Given that islands have discrete boundaries, they are a good place to examine extinctions of native species and introductions of nonnative species that have become naturalized over time (i.e., they have sustained populations in nature). As we have discussed in previous chapters, species on islands can be highly susceptible to extinctions due to relatively small population sizes, overharvesting by humans, and a lack of long-term coexistence with introduced competitors, predators, and pathogens. In one study, the change in island biodiversity over time was quantified by examining the number of birds and vascular plants living on islands that are either native and still present, native but extinct, or non-native. The results showed that across multiple islands around the world, the number of bird species that have gone extinct since humans arrived is similar to the number of birds that have arrived and become naturalized, resulting in a new composition of species, but no net change in biodiversity. In contrast, the number of native plants that have gone extinct since humans arrived is far outweighed by the number of non-native plants that have arrived and become naturalized. While larger islands contained more native species and more naturalized species, the average island has about twice as many plant species today as it once had. For example, New Zealand has about 2,000 species of native plants, but today it has another 2,000 species of non-native, naturalized plants. It had no native land mammals except for bats, but today has several species of introduced land mammals. “While many places on Earth are losing species, these same places are simultaneously experiencing large increases in biodiversity.” The observations from islands also pertain to many other regions of the world. For example, California has experienced about 28
extinctions of native plant species, but more than 1,000 non-native plant species have become naturalized. Collectively, these results set up a paradox between the benefits of having high biodiversity in an ecosystem versus the desire to preserve the native species that live in each ecosystem. Put another way, the activities of humans, including the movements of plant and animal species around the world, may be causing the global decline in species at the global scale due to extinctions, but it can often cause no change or an increase in species richness at the local scale in many parts of the world—such as islands or the state of California—due to the large number of introduced species to these places. As a result, we need to consider not only the number of species and how they affect proper ecosystem function, but also the particular community composition in terms of native versus non-native species. Moreover, we need to understand how local and global biodiversity will change in future decades, including whether the naturalized species might eventually drive native species to extinction. The patterns of biodiversity across the globe highlight the need to understand the processes that produce the historical causes of species richness and the fact that we sometimes need to think about ecology at large spatial scales to understand the distribution and composition of species on Earth. In this chapter, we will explore ecology at large spatial scales to examine the distribution of species both across landscapes and around the world. SOURCES: Sax, D. F., et al. 2002. Species invasions exceed extinctions on islands worldwide: A comparative study of plants and birds. The American Naturalist 160: 766–783. Vellend, M. 2017. The biodiversity conservation paradox. American Scientist 105: 94–101.
Learning Objectives
After reading this chapter, you should be able to:

22.1 Discuss how landscape ecology examines ecological patterns and processes at large spatial scales.
22.2 Explain why the number of species increases with geographic area.
22.3 Describe the equilibrium theory of island biogeography.
22.4 Highlight the causes for biodiversity being highest near the equator and lowest toward the poles.
22.5 Explain how the distribution of species around the world is affected by Earth’s history. Throughout this book, we have considered the role of physical conditions and of species interactions at particular locations. We have seen how these factors can affect the ecology of individuals, populations, communities, and ecosystems. When we explored these topics, we often found it helpful to focus on relatively homogeneous areas of land or water. However, as we moved across landscapes and across continents, we observed that terrestrial and aquatic ecosystems vary from place to place. In this chapter, we will consider much larger areas, from landscapes that include a variety of habitats to entire continents that contain a range of climates. In taking this large-scale approach, our goal is to understand why we find different numbers of species and often very different types of species in different places around the globe and why we sometimes find very similar types of species on widely separated continents. Once we understand the patterns of biodiversity, we can understand the processes that affect diversity and develop plans to conserve it, which is the topic of the next chapter.
#### 22.1 Landscape Ecology Examines Ecological Patterns and Processes at Large Spatial Scales
22.1 Landscape ecology examines ecological patterns and processes at large spatial scales If you were to look across an expansive landscape from the window of a plane, you would undoubtedly see a diversity of terrestrial and aquatic habitats comprising a wide range of sizes and shapes (see Figure 4.2). The field of landscape ecology considers the spatial arrangement of habitats at different scales and examines how they influence individuals, populations, communities, and ecosystems. In this section, we will discuss the sources of habitat heterogeneity and how this heterogeneity generates local and regional biodiversity. Landscape ecology The field of study that considers the spatial arrangement of habitats at different scales and examines how they influence individuals, populations, communities, and ecosystems.
Causes of Habitat Heterogeneity
The current heterogeneity of habitats reflects recent and historical events caused by both natural forces and human activities. Historical processes that have long-lasting influences on the current ecology of an area are known as legacy effects. An interesting legacy effect of glaciers that you can observe today is the presence of eskers, which are the remnants of long, winding streams of water that once flowed inside or under the glaciers. Over time, these glacial streams deposited soil and rock in their paths. After glaciers melt away, these old stream beds appear as long and winding hills (Figure 22.1). These hills harbor different microhabitats that favor unique plant and animal communities.
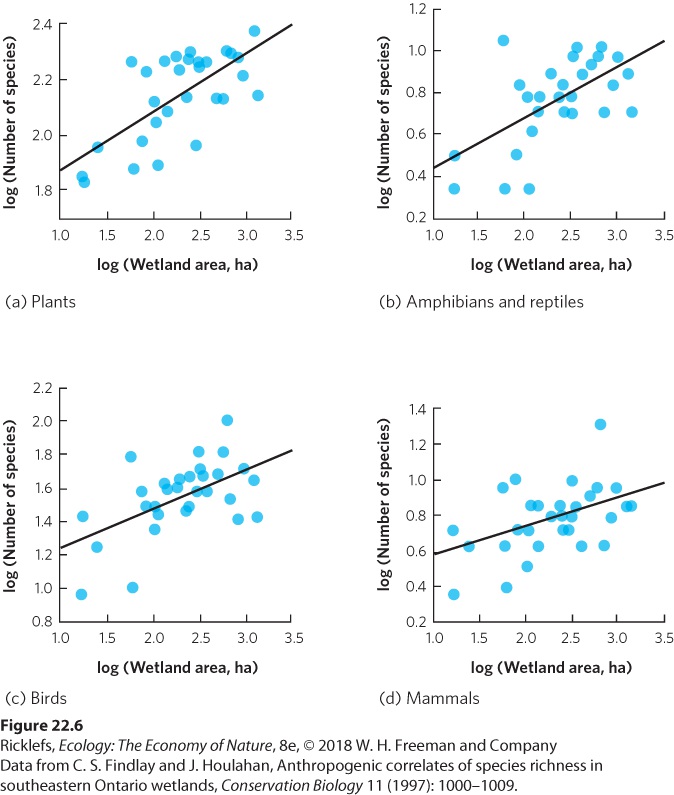
Figure 22.1 A natural legacy effect of the landscape. This long, winding esker near Whitefish Lake, Northwest Territories, Canada, was formed by a stream that once ran through a glacier thousands of meters thick. Particles of soil settled into the bed of the stream, which today forms the long ridge of land that is the esker. Legacy effect A long-lasting influence of historical processes on the current ecology of an area. Natural forces continue to cause habitat heterogeneity in modern times. At both local and regional scales, catastrophes such as tornadoes, hurricanes, floods, mudslides, and fires can alter vegetation structure, which causes changes in populations and communities that depend on it. Although catastrophic events have always occurred naturally, human activity has influenced them. For example, natural fires in Yellowstone National Park were largely suppressed through much of the twentieth century. During the summer of 1988, however, hundreds of fires were ignited both by human activities and by natural causes such as lightning strikes, and these fires were exacerbated by windy conditions and a summer-long drought. Most fires burned relatively small areas of less than 40 ha, but a few of the fires burned much larger areas. In total, nearly 500,000 ha burned, leaving a mosaic of burned and unburned patches across the greater Yellowstone landscape (Figure 22.2).
Figure 22.2 Habitat heterogeneity after a fire. In Yellowstone National Park, hundreds of fires burned patches of the park and left behind a heterogeneous mix of burned and unburned areas. As we saw in Chapter 18, some animals—for example, beavers and alligators—are ecosystem engineers and can alter the habitats in a landscape. Humans are the most extensive ecosystem engineers; they build homes, offices, and factories, construct dams and irrigation channels, channelize waterways for improved navigation, and clear forests for lumber, paper, and agriculture. Logging is a particularly clear example of a human activity that produces a mosaic of habitat types across the landscape. In the western United States, it is common practice to log medium-sized swaths of forest scattered throughout the landscape. This practice helps minimize soil erosion and other damaging effects of large-scale clear-cutting. It also produces a mosaic of forest patches of different ages that persists for many years. Human habitations can also cause habitat heterogeneity. For instance, during the first century ce, the Romans built small villages and farms in France. These farms were abandoned by the fourth century and the land reverted to forest (Figure 22.3a). Researchers studied the soil conditions and plant species located between 0 and 500 m from the former Roman settlements. They discovered that sites closer to the settlements had higher soil pH, more available phosphorus, and greater plant species richness, including many weedy species. These legacy effects, which are shown in
Figure 22.3b, were attributed to two causes: the slow breakdown of the
ancient building materials contributed calcium and phosphorus to the soil; and the Romans introduced numerous plant species to the area. In short, human habitation from 1,600 years ago continues to have strong legacy effects on the modern forest.
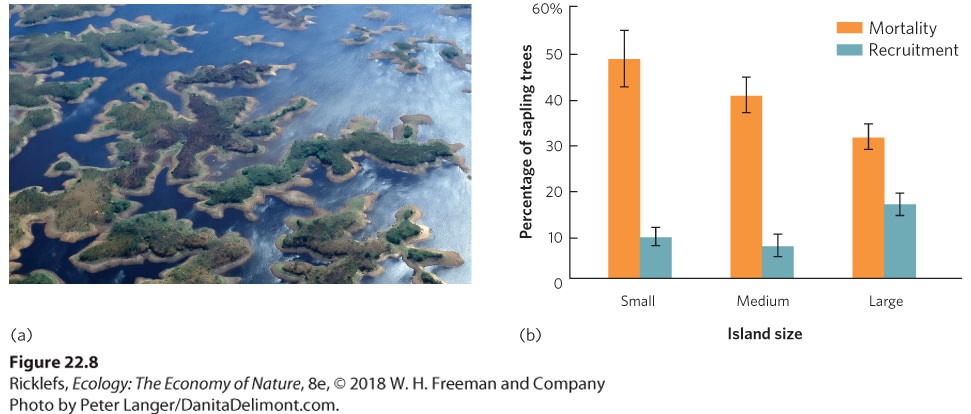
Figure 22.3 Legacy effects of humans on the landscape. (a) Ruins of Roman farming villages are present throughout northern France. These Gallo-Roman ruins are in the forest of Tronçais at the site of “Petit-Jardins” in Isle-et-Bardais, Allier. As the stone building materials weathered, they likely contributed calcium and phosphorus to the soil and altered the abiotic conditions. In addition, the Romans introduced numerous plant species. (b) Collectively, these changes created a legacy of more plant species living close to the old settlements with fewer
##### Relationships between Habitat Heterogeneity and Species Diversity
species living 500 m away. The number of plant species represents the difference from the mean number of plant species found far from the settlements. The dashed line represents the changing mean species richness as one moves away from the center of the settlement.
Species Diversity
When we quantify the number of species in a given area, we include a greater variety of habitats at the landscape level than at the local scale and therefore generally observe a larger number of species. For example, ecologists in Vermont recently surveyed bird species along 27 streams that feed into Lake Champlain. For each stream, they measured the physical characteristics of the stream itself, such as depth and width, and the types of habitats in the riparian areas along the stream banks. While each stream supported an average of only 17 bird species, the entire landscape of streams contained 101 bird species. The number increased significantly because different bird groups preferred different habitat characteristics. For example, waterbirds had their highest species richness and abundance in shallow streams, whereas fisheating birds were more abundant in larger streams with little agriculture along the stream bank because such streams contain more fish. In contrast, the richness and abundance of insect-eating birds were highest in areas containing a variety of habitat types, including shallow streams, meadows, and broadleaf forests. This mixture of meadow, forest, and stream habitats allows a greater variety of insect-eating shore birds and forest birds to exist and feed on the insects that emerge from the stream. Because the heterogeneity of habitat types across the landscape of streams supports a higher richness of bird species than any single habitat, conserving a variety of habitats over a large area is critical to the conservation of high bird species diversity.
Local and Regional Species Diversity
The study of birds in the streams of Vermont highlights the fact that species diversity can be measured at different spatial scales. For example, if we consider the number of species in a relatively small area of homogeneous habitat, such as a stream, we are looking at local diversity or alpha diversity. If we consider the number of species in all the habitats that comprise a large geographic area, we would be looking at regional diversity

or gamma diversity. In the Vermont stream study, the regional diversity would be the entire list of 101 birds that the researchers identified across all 27 streams. Local diversity The number of species in a relatively small area of homogeneous habitat, such as a stream. Also known as Alpha diversity. Regional diversity The number of species in all the habitats that comprise a large geographic area. Also known as Gamma diversity. If each species occurred in all habitats within a region, then the species diversity at the local and regional scales would be identical. However, as we saw in the case of the stream-dwelling birds of Vermont, different species prefer different habitats. Therefore, the number of species at a local scale is less than the number of species at the regional scale. Moreover, the list of species in each local habitat is different from each other. Ecologists refer to the number of species that differ in occurrence between two habitats as beta diversity. For example, imagine two streams in Vermont: Stream A contains five species not found in Stream B and Stream B contains three species not found in Stream A. Because the two streams differ by a total of eight species, the beta diversity is eight. The greater the difference in species between two habitats, the greater the beta diversity. Beta diversity The number of species that differ in occurrence between two habitats. The collection of species that occurs within a region is called the regional species pool. The actual species that live in each local site depend on the species that exist in the regional pool and how well biotic and abiotic conditions at the local scale match the niche requirements of species in the regional pool. Therefore, species in the regional species pool are sorted among localities according to their adaptations and interactions, a process called species sorting. Regional species pool The collection of species that occurs within a region. Species sorting
The process of sorting species in the regional pool among localities according to their adaptations and interactions. An example of species sorting can be seen in an experiment in which researchers set up artificial wetlands and manipulated a variety of conditions, including the fertility of the soil and the amount of flooding the soil experienced. The researchers then sowed the seeds of 20 wetland plant species into each wetland to see which plants would germinate and persist over the next 5 years. Of the original 20 species, one species failed to germinate in any of the wetlands and five others were unable to persist. Of the remaining 14 species, each wetland contained only three to five species by the end of the experiment. Moreover, there were particular combinations of plant species that survived under each flooding and fertility condition. These results confirmed that differences in local conditions cause the sorting of species from the regional species pool.
Concept Check
1. What are two sources of habitat heterogeneity? 2. Why is there a positive relationship between habitat heterogeneity and species richness? 3. Why do habitat preferences of species typically cause alpha diversity to be lower than gamma diversity?

#### 22.2 the Number of Species Increases with Increased Area
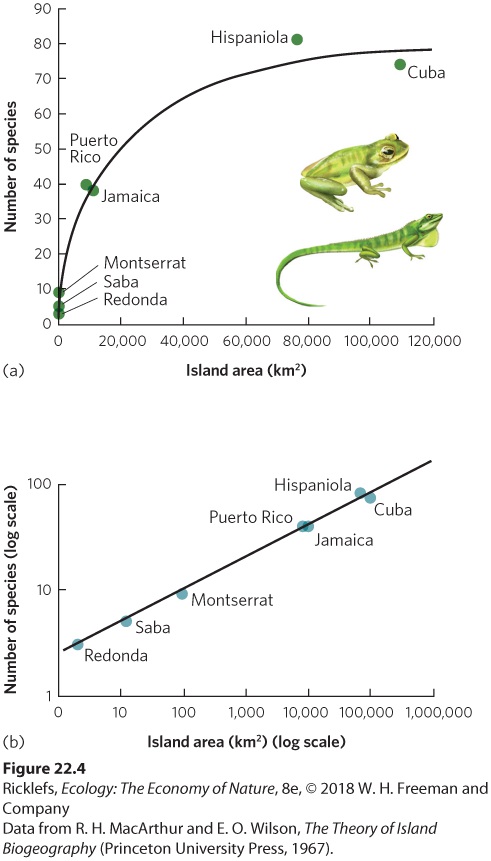
22.2 The number of species increases with increased area When we consider landscape ecology, we see that larger aquatic or terrestrial areas contain a greater number of species. In this section, we discuss examples of this phenomenon from a variety of systems and then explore the mathematical relationship between area size and species richness. We will then consider the underlying processes that cause the positive relationship between area size and species richness. SPECIES–AREA RELATIONSHIPS In a classic study of species diversity, Robert MacArthur and E. O. Wilson examined the patterns of species richness in taxonomic groups living on islands of different sizes. When they plotted the species richness of different islands against the size of the islands, they found that larger islands had a greater species richness. For example, as you can see in Figure 22.4a, the number of amphibian and reptile species living on islands in the West Indies increases with island size.
Figure 22.4 A species–area curve for amphibians and reptiles. On the islands of the West Indies, larger islands contain more species of amphibians and reptiles, such as the Jamaican green tree frog and the Jamaican anole. (a) When plotted as untransformed data, the relationship is a plateauing curve. (b) When plotted on a log scale, the relationship is a straight line. The observation that larger areas tend to contain more species has led to the concept of the species–area curve. The species–area curve is a graphical relationship in which increases in area (A) are associated with increases in the number of species (S). This curve can be described by the following equation: S=cAz Species–area curve A graphical relationship in which increases in area (A) are associated with increases in the
number of species (S). where c and z are constants fitted to the data. To make it easier to work with this equation in the form of a graph, we can take the logarithm of both sides: log S=log c+z log A This is the equation for a straight line with a y intercept of log c and a slope of z. For example, Figure 22.4b plots the same amphibian and reptile data using axes on a log scale. A similar linear relationship can be seen in another collection of data by MacArthur and Wilson in which they examined the number of bird species living on the Sunda Islands in Malaysia, the Philippines, and New Guinea. As you can see in Figure 22.5, when the data are plotted on a log scale, the number of bird species linearly increases with island size.
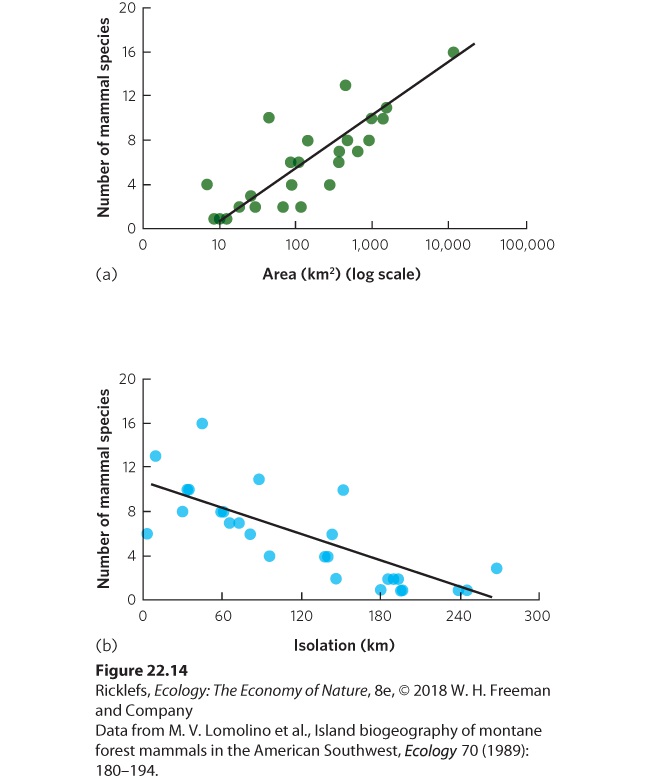
Figure 22.5 A species–area curve for birds. Among the Sunda Islands in Malaysia, Philippines, and New Guinea, islands with a larger area contain more species of birds, such as the Philippines Eagle, when both variables are plotted on a log scale. Across many different groups of organisms, the relationship between log A and log S typically has a slope within the range of z = 0.20 to 0.35 across a wide range of areas, from 1 m2 to a country of modest size. This relatively narrow range of slope values across different studies and different taxa suggests that the relationships between species richness and island area reflect similar processes. Oceanic islands provide good examples of the species–area relationship,
but the pattern can also be observed in a wide range of ecosystems. For example, researchers in Ontario, Canada, surveyed the species living in 30 wetlands of different sizes. As shown in Figure 22.6, they found that as wetland area increased, there was a corresponding increase in the number of species of plants, amphibians, reptiles, birds, and mammals. As you can see, the positive correlation between area and species richness is common in nature.
Figure 22.6 Species richness increases with habitat area. In a survey of 30 wetlands in Ontario, Canada, an increase in wetland area was found to be associated with an increase in species richness in (a) plants, (b) amphibians and reptiles, (c) birds, and (d) mammals.
Habitat Fragmentation
Habitat islands can form when natural processes such as fires and hurricanes cause habitats to become fragmented. Similarly, human activities have caused widespread fragmentation of large habitats throughout the world. For example, in the Central American nation of Costa Rica, the forests in 1940 covered much of the country and existed as a large, continuous habitat. As human populations and their associated activities increased, the forests were cleared on a continuing basis, as shown in Figure 22.7. By 2005, much of the forested area had been cleared and the forests that remained existed in many small fragments, which were essentially islands of forested habitat within a matrix of cleared lands.
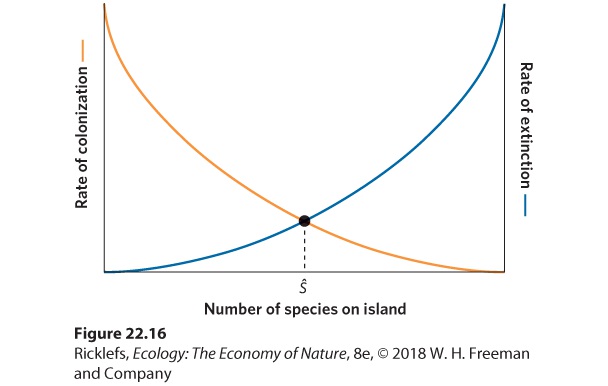
Figure 22.7 The fragmentation of a contiguous forests into islands of forested habitat. In Costa Rica, the forest—shown in dark green—was once largely continuous. Over time, it was cleared by human activities, and today exists only in a number of smaller fragments. Fragmentation of a large contiguous habitat creates several effects: the total amount of habitat decreases, the number of habitat patches increases, the average patch size decreases, the amount of edge habitat increases, and patch isolation increases. Conversely, in the habitat matrix between the fragments, such as cleared fields between forest fragments, the total area increases and the matrix becomes more continuous. Of particular interest is how isolated
habitat patches with different sizes and shapes influence biodiversity, and how habitat corridors and the quality of the matrix between habitat fragments affect the rate of species turnover. Effects of Fragment Size As we saw in the case of oceanic islands, the reduction in habitat size that comes with fragmentation commonly causes a decline in species diversity. This happens because each fragment supports smaller populations of species than existed in the original larger habitat and, as discussed in Chapter 13, smaller populations experience higher rates of extinction (see Figure 13.13). For example, in eastern Venezuela a large river was dammed to create a 4,300-km2 lake known as Lago Guri. The region had been composed of a mixture of grazed fields and tropical forests, but after the dam caused the region to flood, hundreds of high points in the landscape became islands in the newly formed lake, as shown in Figure 22.8a. The smallest islands did not contain enough prey to sustain the large vertebrate predators, so predators on the smallest islands went extinct. These extinctions caused a chain of events that affected the species that remained on the islands. The extinction of the large predators allowed the herbivores on the islands, such as leaf-cutter ants, iguana lizards, and howler monkeys (Alouatta seniculus), to increase. The increase in herbivores then consumed more of the island plants. As you can see in Figure 22.8b, greater abundance of herbivores on smaller islands caused higher mortality in existing sapling trees and reduced recruitment of new trees into the sapling stage.
Figure 22.8 Fragmentation of terrestrial habitats from the flooding of Lago Guri. In eastern Venezuela, a dam was constructed to form Lago Guri in 1978 and the area behind the dam was flooded. (a) Flooding caused the hills of the region to become islands in the lake. (b)
On the smallest islands, large predators were no longer present, so the populations of herbivores increased, with a major effect on the vegetation. Sapling trees on small islands experienced the highest mortality and the lowest recruitment. Error bars are standard errors. Effects of Fragment Edges As you may recall from Chapter 18, when habitats are fragmented, they produce ecotones, which are regions with distinct environmental conditions and unique species compositions over a relatively short distance. Increases in fragmentation cause an increase in the amount of edge habitat compared to the amount of edge present in the original unfragmented habitat. For example, consider a single square habitat with an area of 1 ha compared to the same total area divided into 16 smaller square habitats. As illustrated in
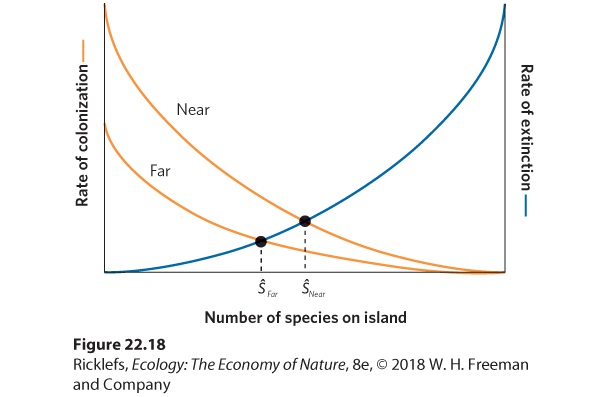
Figure 22.9, the single large habitat would have 400 m of edge, whereas the 16 smaller habitats would have 1,600 m of edge. In regard to fragment shape, round habitats have the lowest ratio of edge to area, while long, skinny ovals or rectangles have much higher edge-to-area ratios.
Figure 22.9 The effect of habitat fragmentation on edge habitat. If we have two habitats of the same total area, the habitat that is fragmented has much more edge. An increase in edge habitat changes both the abiotic conditions and the species composition of a habitat. In a fragmented forest, for instance, the edge of a newly created fragment experiences greater sunlight, warmer temperatures in the summer, and higher rates of evaporation. These changes can make the edge of the forest less suitable to many forest species and more suitable to others. Given that fragmentation increases the amount of habitat edge, it also increases the abundance of those species that prefer edge habitat, and this can affect other species living in the fragment. For
##### Analyzing Ecology: Estimating the Number of Species in an Area
example, in the southern United States, the population of bronzed cowbirds (Molothrus aeneus) has increased with habitat fragmentation. This bird is considered a nest parasite because it reproduces by placing its eggs into the nests of other bird species, which serve as hosts and take care of the cowbird chicks (Figure 22.10). This allows the cowbird to produce offspring without providing time-consuming parental care, but reduces the number of offspring the host bird can raise. While cowbirds spend much of their time living in fields, they enter the forest edge to find nests of forest birds in which they deposit their eggs. Researchers have found that as forest fragmentation has created more edge habitat, more nests are parasitized by the cowbirds. Subsequently, the reproduction and population sizes of the host species of songbirds have declined.

Figure 22.10 Edge habitat and nest parasitism. Bronzed cowbirds deposit their eggs in the nests of forest songbirds along the edges of habitat fragments, such as this nest of a Bewick’s wren in south Texas. The much larger cowbird chick receives a disproportionate amount of food from host parents, and this causes a reduction in the number of host offspring that survive.
Analyzing Ecology
As we have seen, ecologists often need to estimate how exist in an area. Since counting every individual is rarely possible, we must take a sample of an area to estimate how many species are present. One way to estimate the number of species in a sample is to graph the number of species that we observe as we increase our sample size. One would expect that the more we sample, the closer we will get to knowing the actual number of species in an area. Eventually, the curve will reach a plateau so that any additional sampling will not discover additional species. A graph of the number of species observed in relation to the number of individuals sampled is known as a species accumulation curve. Species accumulation curve A graph of the number of species observed in relation to the number of individuals sampled. A second way to estimate the number of species in an area is based on the number of rare species that are detected. When we sample many individuals in a community, we typically end up with a high number of individuals for each of the common species but we may detect only one or two individuals of rare species. We can estimate the number of species actually living in an area (S) by sampling the community to determine how many species are observed (Sobs), how many species are represented by two individuals (f2), and how many are represented by one individual (f1). In essence, species represented by two individuals are likely to be more common in the community than species represented by single individuals. We can therefore estimate the number of actual species that live in a community by starting with how many species we have observed and multiplying this by a ratio that incorporates the number of species represented by one versus two individuals: S=Sobs+f1(f1-1)2(f2+1) As you can see in this equation, the estimated number of species in a community increases with an increase in the number of species
represented by a single individual and decreases with an increase in the number of species represented by two individuals. For example, consider the adjacent table that lists the number of individuals observed for 12 species sampled from three pond communities. Using the data for Community A, we can estimate the actual number of species. In this community, we have observed 12 species. Two individuals were observed for each of two species, whereas one individual was observed for each of three species. Using our equation above, we get S=Sobs+f1(f1-1)2(f2+1)S=12+3(3-1)2(2+1)S=12+66S=13 Species Community A Community B Community C
Dragonflies
Based on these results, we conclude that although we observed 12 species, we estimate that the community actually contains 13 species. YOUR TURN Using the equation, estimate the number of species that are present in Community B and Community C. Based on your results, how
does the number of species represented by single individuals affect the estimated number of species in the community? Fragmentation can also have consequences for human health. You may recall from Chapter 15 that human exposure to Lyme disease depends on a complex food web that starts with mice and chipmunks serving as hosts for newly hatched ticks that carry the pathogenic bacteria. In forest fragments in the northeastern United States, the abundance of many vertebrate animals has declined, but the abundance of the white-footed mouse has increased, probably because most of the mouse’s competitors and predators cannot live in the smallest forest fragments. Surveys of mouse populations in forest fragments of different sizes show that the smallest forest fragments have the highest mouse densities, and thus the highest tick densities. It was also found that mice living in the smallest fragments have the highest proportion of ticks infected with the Lyme disease bacterium. In short, the fragmentation of forests due to human activities has created a landscape that makes us more likely to be exposed to Lyme disease. Corridors, Connectivity, and Conservation In Chapters 11 and 12, we touched on the importance of considering geographic scales larger than the local area. In our discussion of metapopulations, we emphasized that many populations are divided into habitat fragments and that the regional populations persist because each patch is connected by the occasional dispersal of individuals between patches. We saw an example of this in Chapter 11 when we discussed the recovery of the collared lizard. This lizard depended on patches of open habitat, known as glades, as well as corridors between the glades through which they could disperse. Corridors facilitate movement, which can save declining populations of many species from local extinction. They also increase gene flow and genetic diversity within populations, which counteract the negative effects of genetic bottlenecks and genetic drift. Corridors can simply be pieces of preserved habitat or they can be constructed, such as corridors built to let animals cross a highway (Figure 22.11).
Figure 22.11 Building corridors. Road construction through forests is one way that humans create fragmented large habitats. To help wildlife move between forest fragments, some roads, such as this one in Alberta, Canada, include overhead corridors with surfaces of soil and natural vegetation. Although corridors can rescue declining populations by adding new colonists that bring genetic variation, they can also have unintended downsides. For example, corridors built to help conservation of a particular species can facilitate the movement of predators (including poachers), competitors, and pathogens that are harmful to conservation efforts. Therefore, resource managers must carefully consider the costs and benefits of developing corridors among habitat before spending time and money to implement this strategy. The importance of corridors is greatest for those organisms that require a continuous connection to move between habitat fragments. However, organisms such as birds and flying insects can pass over stretches of inhospitable habitat matrix and therefore may not need a continuously connected corridor. Instead, these species can move between large patches of favorable habitat if they have access to small intervening patches where they can stop to rest or forage. These small intervening patches that dispersing organisms can use to move between large favorable habitats are known as stepping stones. Stepping stones Small intervening habitat patches that dispersing organisms can use to move between large favorable habitats.
The role of habitat corridors and stepping stones has spurred major efforts at preservation. India, for example, is home to nearly 60 percent of all Asian elephants (Elephas maximus), which live in several national parks and protected areas. These areas are the fragmented remains of a much larger and contiguous habitat. The World Land Trust and the Wildlife Trust of India are working together to protect important corridors between the protected habitat fragments to ensure the long-term persistence of the elephants (Figure 22.12). Although elephants are charismatic animals that can draw attention to conservation needs, these corridors are likely to assist in the conservation of other charismatic species including tigers (Panthera tigris) and Himalayan black bears (Ursus thibetanus), as well as many less-charismatic species.
Figure 22.12 Wildlife corridors in India. The original habitat of Asian elephants is now fragmented into several protected areas. Conservation groups are currently trying to preserve strips of land between these protected areas to serve as corridors through which elephants and other species can move. The Effects of Matrix Conditions on Dispersal As you may recall from our discussion of metapopulations in Chapter 11, the quality of the matrix that lies between favorable habitat fragments is a key factor that helps determine whether organisms can move between these favorable fragments. The matrix may contain favorable conditions for the movement of organisms or it may be inhospitable. For example, researchers in Colorado mapped the locations of meadows that exist in a matrix of willow thickets and coniferous forests and then studied the movements of more than
6,000 individual butterflies from six taxonomic groups. Butterflies feed in the meadows, but when moving from one meadow to another, they must pass through the matrix. After capturing the butterflies in meadows and writing a unique number on their wings, the researchers released the butterflies. They then recaptured the butterflies to determine whether the butterflies exhibited a preference for flying through the willow-thicket matrix or through the coniferous-forest matrix. In two of the six taxonomic groups, there was no preference; one group rarely left its meadow, while the other group was composed of excellent fliers that easily navigated both matrix habitats. In contrast, individuals from the other four taxonomic groups were 3 to 12 times more likely to move between meadows through the willow-thicket matrix than through coniferous-forest matrix. This confirms that some species have distinct preferences for dispersing through particular matrices, and these preferences are related to their ability to disperse. From this example, we can see that the matrix may affect the movements of organisms among fragments.
Concept Check
1. What does a species–area curve represent? 2. Why does habitat fragmentation lead to an increased abundance of species that prefer ecotones? 3. What is the relationship between increased habitat fragmentation and habitat edge?

#### 22.3 the Equilibrium Theory of Island Biogeography Incorporates Habitat Area and Isolation
22.3 The equilibrium theory of island biogeography incorporates habitat area and isolation When Robert MacArthur and E. O. Wilson examined the relationship between island size and species richness, they noticed that although the size of the island was strongly associated with the number of species, there was a large amount of variation around the species–area curve that suggested other processes were also affecting the number of species living on the island. They considered the locations of the islands from which they collected data and observed that the distance from a source of colonizing species also affects the number of species living on an island. Islands closer to a source of colonizing species—for example, a mainland—appeared to receive more colonizing species. In this section, we will examine the combined effects of island area and isolation on species richness and then explore a graphical model that helps us understand these patterns. Finally, we will explore how this information can be used to design nature reserves.
The Evidence
To test their hypothesis that species richness is determined by both island area and isolation, MacArthur and Wilson collected data on different types of organisms from groups of islands throughout the world. For example, they examined the number of bird species that existed on 25 islands in the South Pacific. The nearest source of new species for these islands was the large island of New Guinea, and these islands could be categorized as being near, intermediate, or far from New Guinea. When the researchers plotted the number of bird species for each island as a function of island area, shown in
Figure 22.13, they found that both island area and the distance from New Guinea affected the number of species living on the island. For islands of a given distance from the mainland, large islands contain more species than small islands. For islands of a given area, near islands contain more bird species than far islands.
Figure 22.13 Effects of island area and isolation on bird species richness. In the South Pacific, researchers examined the number of birds living on islands that were various distances from New Guinea. On islands of a similar distance from New Guinea, large islands contained more bird species than small islands. On islands of a similar area, islands near to New Guinea contained more bird species than islands far from New Guinea. Although oceanic islands provide a good test of how colonization and extinction affect the number of species, we can also consider other types of distinct habitats that exist on continents. For example, James Brown and his colleagues examined the number of mammal species living on mountaintops in the southwestern United States. These mountaintops include alpine tundra and conifer forest habitats that are surrounded by a matrix of low-elevation habitats, including woodlands, grasslands, and desert scrub. Twenty-six species of mammals in the region prefer to live in the mountaintop habitats. Researchers asked whether the number of mammal species on each mountaintop was affected by the area of the mountaintop or the isolation of the mountaintop from two sites containing sources of colonizing species: the southern Rocky Mountains and the Mogollon Rim, a mountain range that cuts across northern Arizona. As you can see in Figure 22.14, the researchers discovered more species of mammals on mountaintops with the largest areas and fewer species of mammals on mountaintops that were farthest away from the two sources of colonizers.
Figure 22.14 Effects of area and isolation on mountaintop mammals. For mammals that live on mountaintop habitats in the southwestern United States, researchers found (a) more mammal species living on mountaintops of greater area and (b) fewer mammal species living on mountaintops that are more distant from a source of colonists. The observations on oceanic islands and mountaintop islands suggest that island area and isolation are important factors in determining species richness. However, a manipulative experiment was needed to demonstrate that these processes do, in fact, cause the observed pattern. In a classic experiment, E. O. Wilson and his graduate student Daniel Simberloff worked on a set of tiny islands in the Florida Keys. These islands typically contain a single mangrove tree and no animals other than insects, spiders, and other arthropods. Prior to the experiment, the team documented that islands located
closer to sources of colonizing species contained more species than islands located far away from sources of colonizing species. Tents were built over selected islands, as shown in Figure 22.15a, and then the islands were fumigated with an insecticide that killed nearly every arthropod. After removing the tents, the researchers returned to the islands every few weeks for a year to track how many species recolonized the islands from large islands nearby. As you can see in Figure 22.15b, each island was rapidly recolonized by arthropods. Moreover, the final number of species was close to the original number of species prior to the fumigation and the more isolated islands once again contained fewer species of arthropods. The actual list of species was different, but the richness was similar. This experiment was important because it demonstrated that isolation was a key factor in determining how many species could live on an island.
Figure 22.15 An experimental test of island biogeography theory. (a) Researchers in the Florida Keys constructed a scaffold frame around islands and covered the scaffold with tarps to act as a tent when fumigating the islands. Fumigating the islands removed most of the arthropods. (b) Over the course of a year, researchers returned to determine how many species of arthropods had recolonized. By the end of the experiment, the islands had nearly recovered their original number of species, as indicated by the dashed lines, with the near islands containing a higher number of species than the far islands.
The Theory
Based on repeated observations that both island size and island isolation affect the number of species living on an island, Robert MacArthur and E. O. Wilson developed the equilibrium theory of island biogeography, which states that the number of species on an island reflects a balance between the
colonization of new species and the extinction of existing species. Equilibrium theory of island biogeography A theory stating that the number of species on an island reflects a balance between the colonization of new species and the extinction of existing species. To understand the equilibrium theory of island biogeography, we need first to understand the factors that affect the colonization of species onto an island and the extinction of species from an island. As we discussed in Chapter 11, species differ in their ability to disperse depending on their overall size and mode of locomotion. So now imagine that we have an island that is completely uninhabited by any species. With no species on the island, a large number of species on the nearby mainland could potentially colonize it. As a growing number of species colonize the island, the pool of species that have not yet colonized the island shrinks and those that remain on the mainland are probably not very good at dispersing. Therefore, the rate of new species colonizing an island declines as a function of how many species have already colonized the island. If the island contained every species that is found on the nearby mainland, the rate of new colonizing species would fall to zero. In
Figure 22.16, the relationship between species richness and rate of colonization by new species is shown as an orange line.
Figure 22.16 Achieving an equilibrium number of species on an island. As the number of species living on an island increases, the rate of colonization by new species from the regional pool declines. At the same time, the rate of extinctions of species living on the island
increases. The equilibrium number of species, Ŝ, occurs where the two curves cross and the opposing processes are balanced. Next we need to consider the factors that affect the rate of extinction on an island. Since the extinction rate is expressed as the number of species that will go extinct over a period of time, the extinction rate must be affected by how many species are present. In the simplest case, when there are no species, there can be no extinctions. As the island begins to be colonized, it holds a few species that could potentially go extinct. As more species live on the island, more species are subject to possible extinction, and so the extinction rate increases. The extinction rate should also be affected by harmful species interactions on the island. For example, competition, predation, and parasitism are all more likely to increase as the total number of species increases. The blue line in Figure 22.16 shows that the extinction rate ranges from zero when no species are present on the island to a maximum rate of extinction when the island contains every species that it can possibly support. Given that the island continues to experience colonization of new species and extinction of existing species, these two opposing forces will eventually reach an equilibrium point where the rate of colonization is equal to the rate of extinction. On our graphical model, equilibrium occurs where the two curves cross. Following the dashed line down from the equilibrium point, we see that the number of species on the island at equilibrium is designated as Ŝ on the x axis. It is important to remember that the model predicts the number of species present at equilibrium, but not a particular composition of species at equilibrium. At equilibrium, there is a continuous turnover of species on the island; while new species colonize the island, others go extinct. We saw an example of this in the islands in the Florida Keys that were fumigated by Simberloff and Wilson. Over time, the species richness of the islands came to equilibrium, though the composition of species changed as the islands continued to experience extinctions and colonizations of invertebrate species. Using the graphical model in Figure 22.16 as a foundation, we can make predictions about the combined effects of island size and isolation on the number of species on an island when it is at equilibrium. Let’s first consider the effects of island size. We would expect smaller islands to sustain smaller populations of each species. As we have discussed in several previous
chapters, smaller populations typically have higher extinction rates. Therefore, smaller islands should experience higher rates of extinction, as shown by the curve labeled “Small” in Figure 22.17. When an island is at equilibrium, extinction rates balance colonization rates. Therefore, the number of species on a small island at equilibrium should be lower than the number of species on a large island at equilibrium. If we follow the two equilibrium points on the figure down to the x axis, we can see that the number of species at equilibrium is lower for small islands than for large islands.
Figure 22.17 Effects of island size on the equilibrium number of species. Because smaller islands support smaller populations that are more prone to extinction, smaller islands have steeper extinction curves. As a result, smaller islands contain fewer species at equilibrium (ŜSmall) than larger islands (ŜLarge). Next we can consider the effects of island isolation. An island that is near a continent should experience higher rates of colonization by new species than an island that is far from a continent. Once again using Figure 22.16 as our starting point, we can make predictions about how island isolation should affect the number of species at equilibrium. In Figure 22.18, we can see that the far island has a lower colonization curve than the near island. This creates two equilibrium points between colonization and extinction rates. If we follow the dashed lines down from each equilibrium point, we see that the
number of species at equilibrium is lower for far islands than for near islands.
Figure 22.18 Effects of island isolation on the equilibrium number of species. Islands that are far from a source of new colonizing species experience lower rates of colonization than islands that are close. As a result, farther islands contain fewer species at equilibrium (ŜFar) than near islands (ŜNear). When we combine the effects of island size and island isolation from a continent, we arrive at several predictions. Looking at Figure 22.19, we see that the combinations of large versus small islands and near versus far islands create four different possible equilibrium points. Once again, we can follow the dashed line down from each equilibrium point to determine how many species are predicted to be on a particular type of island when colonization and extinction rates are at equilibrium. Small islands that are far from a major source of new species should contain the fewest species. In contrast, large islands that are near a major source of new species should contain the most species. In short, the combination of island size and isolation determines the number of species that the island can hold.
##### Applying the Theory to the Design of Nature Reserves
Figure 22.19 The combined effects of island size and isolation on species richness. When we simultaneously consider island size and isolation, we find that the small islands that are far from a continent should have the lowest number of species at equilibrium (ŜSF), whereas large islands that are close to a continent should have the highest number of species (ŜLN).
Reserves
Understanding the effects of island size, shape, and distance from sources of colonizing species has helped scientists to design nature reserves to protect biodiversity, as depicted in Figure 22.20. For example, we know that large areas typically contain more species than small areas because they contain a greater diversity of habitats. Large areas can support larger populations of each species, which lowers the rate of species extinction. Therefore, setting aside large nature reserves will better protect biodiversity than will small nature reserves. Similarly, a single large reserve is the better option to preserve biodiversity than are several small reserves of the same total area. However, a single habitat poses several risks. When a species of conservation concern is located in a single large habitat, it is more likely that the species will be destroyed by natural disaster or disease.
Figure 22.20 Nature reserve design. The number, shape, size, and proximity of reserves affect the probable success of preserving a region’s biodiversity. As noted in this figure, some designs are likely to be better than others for preserving biodiversity. When we are faced with creating multiple reserves, we need to consider the proximity of the reserves to each other. Reserves need to be close enough for organisms to disperse between them but far enough away to reduce the ability for predators and diseases to move between them. Setting this distance is a compromise between these positive and negative outcomes, and the best solution will differ among organisms with different dispersal abilities. Sometimes the ideal distance will be set for the species of greatest conservation interest. A similar challenge exists when reserves are joined by corridors that provide strips of hospitable habitat between adjacent reserves. Because we know that increased edge can have detrimental effects on species that prefer to live in the interior of habitats, it is also important that we consider reserve shape. Round reserves have the lowest ratio of edge to area, while long, skinny ovals or rectangles have much higher edge to area ratios that will favor edge species. Although reserve size, number, shape, and proximity are all important considerations, often the ideal scenario is not
possible; nature reserves typically represent a compromise between the areas that are desirable and the areas that are available.
Concept Check
1. Why do mountaintop habitats function similarly to oceanic islands in regard to the theory of island biogeography? 2. Using the theory of island biogeography, why do we expect the number of species on an island to reach equilibrium? 3. What combination of island size and distance to the mainland would you expect to result in the greatest species richness?
#### 22.4 Biodiversity Is Highest Near the Equator and Declines Toward the Poles
22.4 Biodiversity is highest near the equator and declines toward the poles We have examined the local and regional processes that can affect the number of species in a given location on Earth. However, patterns of biodiversity also exist at the global scale. One of the most striking patterns is that species richness of all taxa combined is highest near the tropics and declines toward the poles. For example, a hectare of forest typically has fewer than 5 species of trees in boreal regions, 10 to 30 species of trees in temperate regions, and up to 300 species of trees in tropical regions. These latitudinal trends in diversity are pervasive and extend even to the oceans. In this section, we will explore the patterns of diversity within and across latitudes. We will then discuss the two general hypotheses for these patterns of biodiversity and explore three important processes.
Patterns of Diversity
In the Northern Hemisphere, the number of species in most groups of animals and plants increases from north to south. For example, when a researcher in North America counted the number of mammal species living in blocks of habitat that were 241 km on a side, he found that there were fewer than 20 mammal species per block in northern Canada but more than 50 species per block in the southern United States. You can view these data in Figure 22.21a.
Figure 22.21 Patterns in North American diversity. When a researcher examined the number of species living in the United States and Canada, he found strong patterns in species richness. The contour lines on the map indicate the numbers of species found in blocks of land that were 241 km on a side. (a) Mammals increase in species richness from north to south and from east to west. (b) Reptiles increase in species richness from north to south. (c, d) For both trees and amphibians, the highest number of species occurs in the southeast, where there is a combination of high precipitation and warm temperatures. The number of mammals also increases as we move from east to west in North America. For instance, there are typically 50 to 75 species per sample block in the east, whereas there are 90 to 120 species per block in the west. Such a pattern is likely due to the greater amount of habitat heterogeneity in the extensive mountain ranges of western North America. This greater heterogeneity of environments in the west apparently provides suitable conditions for a greater number of species. The pattern of species richness for birds in North America resembles the pattern for mammals, but the patterns for reptiles, trees, and amphibians are strikingly different, as you can see in Figures 22.21b–d. The richness of reptile species declines fairly uniformly as temperature decreases toward the north. However, trees and amphibians are more diverse in the moister eastern half of North America than in the drier, more mountainous western regions.
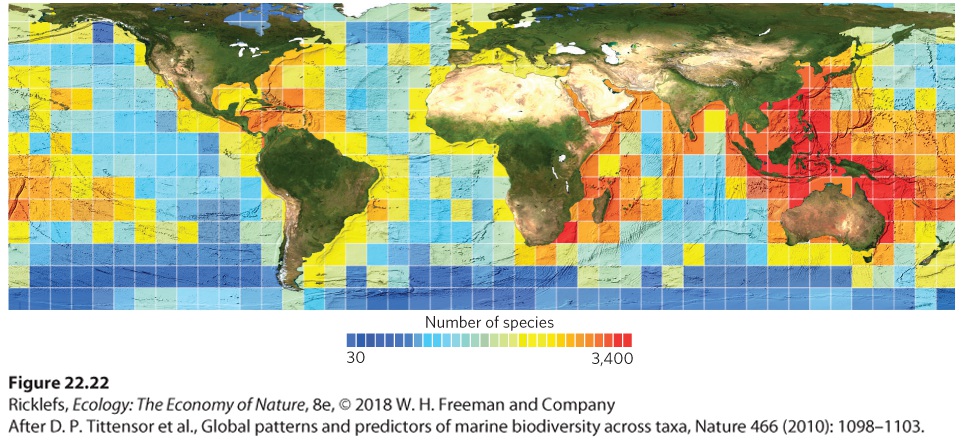
The general pattern of increased species richness as we move closer to the equator also holds true for the oceans. For example, in 2010 researchers compiled millions of records of marine organisms around the world, including whales, dolphins, fish, and corals. Using sample blocks of 880 km on a side, as shown in Figure 22.22, they found that the highest diversity occurred in the tropics including Central America and Southeast Asia, whereas the lowest diversity occurred near the poles where temperatures are colder. At a given latitude, they also found a higher diversity near the coasts and lower diversity in the open ocean.
Figure 22.22 Patterns of marine biodiversity. In the ocean, the greatest number of species exists in the tropics and declines toward the poles. At a given latitude, more species exist near the coasts and fewer species exist in the open ocean. The number of species is calculated using sample blocks of 880 km per side.
Processes That Underlie Patterns of Diversity
Historically, ecologists have considered two general hypotheses for the decline in species richness as we move from the equator to the poles. According to one hypothesis, species are continually created over time and without limit. Because the world’s current temperate and polar regions experienced repeated advances and retreats of glaciers during the Ice Age, species in these regions have been eliminated or driven toward refuges closer to the equator. In contrast, because the world’s tropical regions did not experience glaciation, habitats in these areas have remained stable much longer and so have had more time to accumulate species. The second hypothesis proposes that the number of species reflects an equilibrium between the processes that create new species and the processes that drive species extinct, similar to the state of equilibrium described by the
theory of island biogeography. According to this hypothesis, the higher number of species in the tropics is the result of higher rates of speciation or lower rates of extinction, compared to the temperate and polar regions. Similarly, variation in the number of species across a given latitude should also reflect an equilibrium between processes that create new species and destroy existing species. We will now consider three processes that play substantial roles in determining the number of species that occur in an area: ecological heterogeneity, solar energy and precipitation on land, and water temperatures in the oceans. The Role of Ecological Heterogeneity At any given latitude, we find more species in areas where there is greater ecological heterogeneity, such as heterogeneity in the soils and plant life. For example, grasslands contain vegetation that is less heterogeneous in growth form than shrublands or deciduous forests. As a result of these differences in habitat heterogeneity, surveys of breeding birds in North America find an average of 6 species in grasslands, 14 species in shrublands, and 24 species in floodplain deciduous forests. You may recall that we discussed a similar phenomenon in Chapter 18 when we looked at the positive relationship between the diversity of vegetation height and the diversity of birds (see
Figure 18.13). This same pattern has also been observed in lizards of the southwestern United States, where lizard diversity is associated with vegetation diversity, and in the plants of South Africa, where higher plant diversity is associated with areas with more heterogenous soils. Although more productive terrestrial habitats tend to have a greater number of species, habitat heterogeneity also plays a role in determining species richness when two habitats have similar levels of productivity. For instance, habitats with less variation in vegetation growth form, such as grasslands, have fewer animal species than habitats with similar productivity but more variation in vegetation. This principle also applies to plants. Marshes are highly productive but have a relatively uniform landscape and thus contain relatively few species of plants. While desert vegetation is less productive than marsh vegetation, the greater heterogeneity of the desert landscape provides room for more species of plants (Figure 22.23).
Figure 22.23 Habitat heterogeneity. Although marshes are more productive than deserts, the relative uniformity of the marsh environment has produced a low diversity of species, whereas the heterogeneous landscape of deserts has produced a higher diversity of species. Pictured here are (a) the White River Marsh Wildlife Area in Wisconsin and (b) the Sabino Canyon Recreation Area in Arizona. The Role of Solar Energy and Precipitation on Land The number of species found at any location as one moves from the poles to the tropics is positively correlated to the amount of solar energy and precipitation at that location. Together, solar energy and precipitation can be measured as potential evapotranspiration (PET). PET is the amount of water that could be evaporated from the soil and transpired by plants, if water
was abundant, given the average temperature and humidity of that location on Earth. Because PET integrates temperature and solar radiation, it provides an index to the overall energy input into the environment. As you can see in
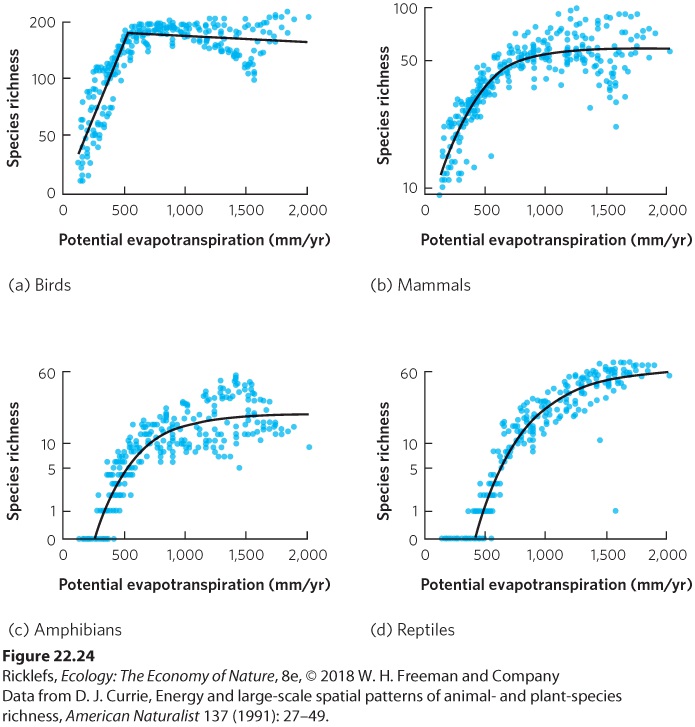
Figure 22.24, PET correlates with the species richness in North American vertebrates relatively well. In each group of vertebrates, the initial rise in PET is associated with a rise in species richness, which reflects increasing diversity from north to south within the continent. However, species richness eventually plateaus at very high levels of PET. These high PET levels occur as we move from east to west at middle latitudes where the increasing temperature fails to improve the capacity of the environment to support additional vertebrate species. In the arid western parts of the continent, for instance, increasing temperature eventually becomes a stressor.
Figure 22.24 The relationship between species diversity and potential evapotranspiration. For all four groups of vertebrates, the potential evapotranspiration of a sample is strongly correlated to the number of species found in the sample block.
Potential evapotranspiration (PET) The amount of water that could be evaporated from the soil and transpired by plants, given the average temperature and humidity. The correlations between PET and species richness for terrestrial vertebrates have given rise to the idea that this correlation may actually represent a causal relationship between the two variables. This hypothesized causal relationship is known as the energy–diversity hypothesis, which states that sites with higher amounts of energy are able to support more species. Higher amounts of energy would also support higher abundances of individuals from each species, which should reduce the rate of extinction. Moreover, higher energy input might accelerate the rate of evolutionary change and thereby increase the rate of speciation. While these ideas are attractive, none of these mechanisms has yet been verified. Energy–diversity hypothesis A hypothesis that sites with higher amounts of energy are able to support more species. The Role of Water Temperature in the Oceans As we have seen, biodiversity in marine environments is greater in the tropics than it is at higher latitudes. However, this pattern does not appear to be driven by greater productivity in the tropics. Though marine productivity is greatest in temperate latitudes (see Figure 20.6), the high productivity in those regions is seasonal; temperature differences and stratification of the water in temperate zones make nutrients readily available during the seasonal mixing of the water column but scarce during periods when the water is stratified. In contrast, tropical marine environments experience relatively stable temperatures that lead to relatively small nutrient fluctuations and low but steady productivity. Researchers have tested whether the patterns in species diversity across the oceans were better explained by marine productivity, mean water temperature, or variation in water temperature. They discovered that the only significant predictor of marine biodiversity across latitudes was the mean temperature of the sea surface. Because a higher mean temperature is a measure of greater total energy, this pattern further supports the energy–diversity hypothesis. The patterns of species richness across and within latitudes are potentially
affected by all three processes of habitat heterogeneity, temperature and precipitation on land, and mean temperature in the oceans. In all three cases, the mechanisms involved suggest that the global distribution of species richness is the outcome of an equilibrium between the processes that create new species and the processes that cause the extinction of existing species.
1. What is the geographic pattern of species richness for mammals in North America? 2. Why would increasing vegetation heterogeneity lead to increased species richness in birds? 3. Why might we expect potential evapotranspiration to be correlated to terrestrial species richness across latitudes?
#### 22.5 the Distribution of Species Around the World Is Also Affected by Earth’s History
22.5 The distribution of species around the world is also affected by Earth’s history When we consider the patterns of species diversity on Earth, we need to remember that Earth was formed 4.5 billion years ago and life arose during the first billion years. This means that the species on Earth today are the result of billions of years of evolution and that current diversity arose in response to past environmental conditions. In this section, we will examine how the movement of continents and historic changes in climate affected the distribution of species.
Continental Drift
During Earth’s history, the continents have repeatedly come together and drifted apart in a process called continental drift. Continental drift occurs because the continents are essentially giant islands of low-density rock that move by underlying convection currents of semi-molten material. About 250 Mya, all of Earth’s landmasses were joined together as a single landmass, named Pangaea, illustrated in Figure 22.25. By 150 Mya, Pangaea had separated into a northern landmass, known as Laurasia, and a southern landmass, known as Gondwana. Laurasia subsequently split into North America, Europe, and Asia, while Gondwana split into South America, Africa, Antarctica, Australia, and India. Ultimately, India collided into Asia, which caused the land to rise and created the Himalayas. In the Northern Hemisphere, a widening Atlantic Ocean separated Europe from North America, but a land bridge had already formed on the other side of the world between North America and Asia. More recently, Europe and Africa joined about 17 Mya and a land bridge formed between North and South America at the Isthmus of Panama 3 to 6 Mya.
Figure 22.25 Continental drift. About 250 Mya, all of Earth’s landmasses were joined together into Pangaea. Pangaea later split into Gondwana and Laurasia, which, in turn, split into modern-day continents. Many of these landmasses subsequently joined together in new configurations. Continental drift The movement of landmasses across the surface of Earth. Pangaea The single landmass that existed on Earth about 250 Mya and subsequently split into Laurasia and Gondwana. Laurasia The northern landmass that separated from Pangaea about 150 Mya and subsequently split into North America, Europe, and Asia. Gondwana The southern landmass that separated from Pangaea about 150 Mya and subsequently split into South America, Africa, Antarctica, Australia, and India. An important consequence of continental drift was the changing opportunities for dispersal among continents. Once separated, continents could independently evolve species in different regions of Earth. For instance, because Australia has experienced a long period of isolation from other continents, it has evolved many unique groups of species that include a wide variety of marsupial animals, such as koalas and kangaroos, and plants, such as eucalyptus trees. When continents later joined together, groups of organisms that were unique to one landmass were able to move into new areas. You may recall from Chapter 3 that camels originated in North America about 30 Mya. When North America became connected to Asia at the Bering Land Bridge in Alaska, ancestral camels dispersed into Asia and then moved into Africa, where they diversified into the modern-day Bactrian
camels and dromedary camels. At about the same time, other ancestral camels dispersed into South America, where they further diversified into llamas, guanacos, vicuñas, and alpacas. In short, the history of continents separating and coming together continues to affect the distributions of present-day animals and plants.
Biogeographic Regions
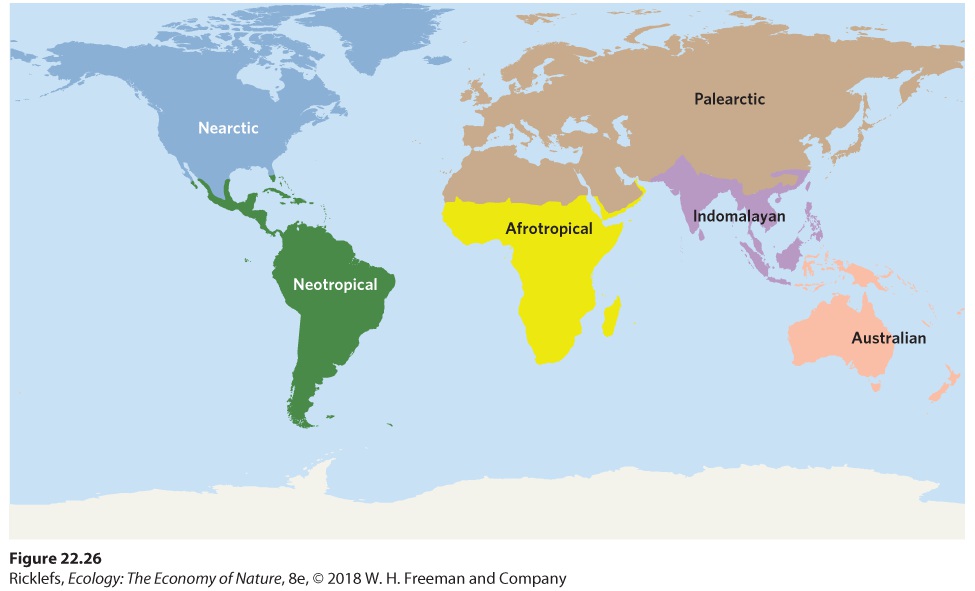
Because continental drift has allowed individual continents independently to evolve groups of organisms for long periods, we can see patterns of species distributions on each continent. Alfred Wallace, a contemporary of Charles Darwin and co-discoverer of evolution by natural selection, first described these patterns and delineated six major biogeographic regions that are based on the distributions of animals. These regions, illustrated in Figure 22.26, are still recognized today. Botanists also recognize six major biogeographic regions, based on plant distributions, with boundaries that closely coincide with those regions based on animal distribution.
Figure 22.26 Biogeographic regions. Terrestrial regions of the world can be categorized in terms of distinctive groups of plants and animals that largely reflect the history of continental drift. The Northern Hemisphere is divided into the Nearctic region, which roughly corresponds to North America, and the Palearctic region, which corresponds to Eurasia. Through most of the past 100 million years, the continents of these regions maintained connections across what is now
Greenland between North America and Eurasia and across the Bering Strait between Alaska and Russia. Consequently, these two regions share many groups of animals and plants. European forests seem familiar to tourists from North America, and vice versa; although few species are the same, both regions have representatives of many of the same genera and families. Nearctic region The biogeographic region of the Northern Hemisphere that roughly corresponds to North America. Palearctic region The biogeographic region of the Northern Hemisphere that corresponds to Eurasia. The Southern Hemisphere is divided into four biogeographic regions. The Neotropical region corresponds to Central and South America and the Afrotropical region, also known as the Ethiopian region, corresponds to most of Africa. Farther to the east is the Indomalayan region, also known as the Oriental region, which includes India and Southeast Asia. The final biogeographic region is the Australasian region, which includes Australia, New Zealand, and New Guinea. In Wallace’s delineation, the continent of Antarctica was not included. Neotropical region The biogeographic region of the Southern Hemisphere that corresponds to South America. Afrotropical region The biogeographic region of the Southern Hemisphere that corresponds to most of Africa. Also known as Ethiopian region. Indomalayan region The biogeographic region of the Southern Hemisphere that corresponds to India and Southeast Asia. Also known as Oriental region. Australasian region The biogeographic region of the Southern Hemisphere that corresponds to Australia, New Zealand, and New Guinea. Each of these regions contains unique groups of species that reflect their long histories of isolation from the rest of the terrestrial world and the subsequent exchange of species after they were joined together. For example, when the Isthmus of Panama joined the Nearctic and Neotropical regions, there was a large, though uneven, exchange of mammals. Many North
American lineages moved into South America and caused the extinction of most of the marsupial animals that had come to South America during an earlier connection with Australia. In contrast, only a few mammals moved into North America from South America; the most prominent are the opossums, which is the only marsupial species in the United States and Canada.
Historic Climate Change
As we saw in Chapter 5, the position of the continents and the circulation of water around the continents influence the variation in climates around the world. Because the continents were drifting over the past 250 million years, it is not surprising that the climates of the world experienced dramatic changes. We know from fossil evidence that large portions of North America and Europe once had tropical climates. Tropical forests reached into Russia and Canada and warm temperate forests covered the Bering Land Bridge from Alaska to Asia. Moreover, the Antarctic land connection between South America and Australia supported temperate vegetation and abundant animal life. However, as Antarctica drifted over the South Pole and as North America and Eurasia gradually encircled the northern polar ocean, the Arctic Ocean became largely enclosed between North America and Eurasia. This created a circumpolar ocean current around Antarctica, which caused cooler temperatures at high latitudes. As a result, Earth’s climates became more strongly differentiated from the equator to the poles. Tropical environments contracted into a narrow zone near the equator, and temperate and boreal climate zones expanded. These climate changes had profound effects on the geographic distributions of plants and animals. About 2 Mya, the gradual cooling of the planet gave way to a series of dramatic oscillations in climate known as the Ice Age. Climate changes during this time had dramatic effects on habitats and organisms in most parts of the world. Alternating periods of cooling and warming caused the advance and retreat of ice sheets at high latitudes over much of the Northern Hemisphere and cycles of cool, dry climates and warm, wet climates in the tropics. Ice sheets came as far south as Ohio and Pennsylvania in North America and covered much of northern Europe, driving vegetation zones southward, possibly restricting tropical forests to isolated refuges where conditions remained moist.
A striking example of this disruption is the migration of forest trees in eastern North America and Europe. At the peak of the most recent glacial period, many tree species were restricted to southern refuges, but about 18,000 years ago after the ice began to retreat and the forests started to spread north again. Pollen grains, which are deposited in the lakes and bogs left by retreating glaciers, record the coming and going of plant species. These records show that the composition of plant associations changed as species migrated back north over different routes across the landscape. The migrations of some representative tree species from their southern refuges are mapped in Figure 22.27. As you can see in this figure, spruce trees shifted northward just behind the retreating glaciers. Oaks expanded out of their southern refuges to cover most of the eastern part of temperate North America, from southern Canada to the Gulf Coast. Hemlocks expanded into the Great Lakes Region and into central Canada.
Figure 22.27 Changing tree distribution following the retreat of glaciers. As the glaciers began retreating in North America 18,000 years ago, the trees began moving north to colonize
the newly exposed habitats. The forests of Europe suffered even more extinctions from the spread of glaciers than those in North America because populations were blocked from shifting southward by the Alps and the Mediterranean Sea. Several northern European tree species went extinct. Many species that survived were restricted to refuges in southern Europe, from which they expanded after the glaciers receded beginning about 18,000 years ago. Recent research suggests that many tree species in Europe have not yet expanded fully into their potential ranges, which suggests that the European flora has not returned to an equilibrium state. As we will see in the next chapter, the ability of species to respond to changing climates raises questions about how species will respond to global warming. Throughout this chapter, we have seen that studying the distribution of species must be done at both the landscape scale and the global scale. In doing so, we see that biodiversity is maintained by large-scale factors including area, isolation, and historical events that continue to show their effects today. In “Ecology Today: Applying the Concepts,” we will see that this knowledge has helped scientists design a massive nature reserve to help protect the biodiversity of Florida.
Concept Check
1. How has continental drift affected the modern distribution of species on Earth? 2. How are biogeographic regions delineated? 3. How do scientists know that tree species have been moving north since the most recent ice age?
Concepts
Taking a Long Walk for Conservation The Florida panther. Many species, such as this Florida panther— an endangered subspecies of mountain lion—are expected to benefit from the Florida Wildlife Corridor project that will link together large areas of protected habitats. As we have seen throughout this chapter, the distribution and persistence of biodiversity depend on a variety of factors. Knowing the factors responsible for maintaining diversity helps when designing nature reserves to protect biodiversity. Although we have a good idea about the features of an ideal nature reserve, in reality, the design of nature reserves and the corridors that connect them is largely determined by existing habitats, their configurations, and land ownership. An excellent example of this can be found in a conservation effort known as the Florida Wildlife Corridor, a visionary plan to create a continuous connection of protected habitat from the southern tip of Florida all the way up to Georgia. This would provide a large number of species, including endangered species such as the Florida panther, with a tremendous amount of land to inhabit for a long time to come. The ultimate goal is to connect a number of current
federal nature reserves, including the Everglades National Park, the Big Cypress National Preserve, the Ocala National Forest, and the Okefenokee National Wildlife Refuge, with a variety of properties owned by the state of Florida, conservation organizations such as The Nature Conservancy, and private landowners including large cattle ranches. The Florida Wildlife Corridor. Conservationists hope to link public and private land through the state of Florida in a way that promotes the movement of wild organisms between habitats and facilitates their long-term persistence. With such a multitude of landowners, the Florida Wildlife Corridor is an excellent case study of the challenges
presented by the design of nature reserves. It is as much a sociological problem as an ecological problem, so the promoters of the corridor had to think about a public relations campaign that was as big as the 1,600-km swath of land down the middle of the state. In 2012, four leaders of the project decided to travel the entire length of the proposed corridor in what they referred to as “1,000 miles in 100 days.” Moving on foot and in kayaks, the conservationists traversed swamps, grasslands, and forests. They called attention to their effort by using daily photo and video updates on their websites. Along the way, they were joined by state and federal biologists, conservationists, politicians, and private ranch owners. At every stop, they emphasized that preserving this swath of land through the middle of Florida provided benefits not only to the organisms that lived in undeveloped lands, but also to the residents of Florida who rely on tourism as well as the water and agriculture that come out of this region. On Earth Day 2012, the 100-day expedition was completed at the Stephen C. Foster State Park in southern Georgia. In 2015, the group did a second walk for conservation, from the Everglades to the western edge of the Florida Panhandle. On this trip, they observed a great deal of fragmented habitat, but it was clear that the possibility of protecting key habitats and corridors along this region of Florida still existed. Collectively, the Florida Wildlife Corridor is made up of 6.4 million ha, which is home to 42 federally endangered species. About 60 percent of the land is currently protected, and the group’s goal is to protect an additional 120,000 ha of key habitats along the corridor by 2020. The effort to protect the land in a north–south corridor
through Florida continues today, with a growing momentum to find a way to protect the lands that lie between the state and federal reserves, while at the same time retaining the long-held traditions of agriculture and ranching on private lands. If such protection can be achieved, then the state will have a total reserve that is both large and interconnected, and this should result in the long-term persistence of species in the region. SOURCE: The Florida Wildlife Corridor (http://www.floridawildlifecorridor.org/about).
Summary of Learning Objectives
22.1 Landscape ecology examines ecological patterns and processes at large spatial scales. Habitat heterogeneity exists across the landscape as a result of both natural processes and human activities, both in the past and in modern times. This heterogeneity allows more species to exist at the landscape scale than at local scales. Species diversity can change with spatial scale; we quantify alpha diversity at the local scale, gamma diversity at the regional scale, and beta diversity across habitats. The process of species sorting determines which species from the regional species pool are found in a local community. Key Terms: Landscape ecology, Legacy effect, Local diversity, Regional diversity, Beta diversity, Regional species pool, Species sorting
22.2 The number of species increases with increased area. The number of species found in areas of different sizes can be described by the species–area curve. As the area sampled grows, we see an increase in species richness because we begin to sample new habitats that contain different species. Habitat fragmentation divides a large habitat into several small habitats and thereby increases the amount of habitat edge. These effects cause the decline in the abundance of many species, but favor those species that prefer habitat edges. Key Terms: Species–area curve, Species accumulation curve, Stepping stones
22.3 The equilibrium theory of island biogeography incorporates both habitat area and isolation. Species diversity is highest on large islands that are close to a source of colonizing species because these islands have a low rate of species extinction and a high rate of colonization by new species. In contrast, species diversity is lowest on
small islands that are far from a source of colonizing species because these islands have a high rate of species extinction and a low rate of colonization by new species. Key Term: Equilibrium theory of island biogeography
22.4 Biodiversity is highest near the equator and declines toward the poles. Species diversity can also vary within a given latitude belt as a result of habitat heterogeneity, temperature, and precipitation. In terrestrial ecosystems, the best predictor of species diversity is potential evapotranspiration. The energy–diversity hypothesis states that sites with higher amounts of energy are able to support more species. In marine ecosystems, the best predictor of species diversity is the temperature at the surface of the sea. Key Terms: Potential evapotranspiration (PET), Energy– diversity hypothesis
22.5 The distribution of species around the world is also affected by Earth’s history. Continental drift is a major factor in determining the distribution of species. As continents have come together and separated, they have experienced unique evolutionary lineages that today are recognized as distinct biogeographic regions. The drift of the continents has also affected historic climate changes, including widespread changes in the distribution of tropical, temperate, and polar regions around the world. Key Terms: Continental drift, Pangaea, Laurasia, Gondwana, Nearctic region, Palearctic region, Neotropical region, Afrotropical region, Indomalayan region, Australasian region
Critical Thinking Questions
1. How can the fragmentation of a landscape have both positive and negative effects on biodiversity? 2. Why would it be important to consider the quality of the matrix that exists between habitat fragments when considering the movement of amphibian species between ponds? 3. How do local diversity and regional diversity affect the slope and intercept of the species–area relationship? 4. Compare and contrast alpha, gamma, and beta diversity. 5. Why might we expect that regions with high species diversity will also exhibit high niche diversity? 6. Why should small islands that are distant from a mainland have fewer plant species than large islands that are close to a mainland? 7. When North America and South America came together by continental drift, what may have caused the North American mammals that moved into South America to drive many South American mammals extinct? 8. In what ways might the age and area of a region affect its species richness? 9. How does knowledge of historic climatic patterns affect our interpretation of present-day patterns of species diversity? 10. What is the likely relationship between the length of time the modern continents have been connected and the similarity among the families and genera on each continent?
#### Graphing the Data: Species Accumulation Curves
GRAPHING THE DATA Species Accumulation Curves Ecologists often need to determine the number of species in an area they are sampling. As we have discussed in this chapter, they can use an equation to estimate the total number of species or they can graph a species accumulation curve that plots the sample size along the x axis and the number of species observed along the y axis. Using the data in the table, graph species accumulation curves for forest communities A and B. Based on your graph, which community has been sampled sufficiently to provide a good estimate of the species richness that exists and which community requires more sampling? Explain your answer. Number of Individuals Sampled Number of Species Observed in Community A Number of Species Observed in Community B