Chapter 12: Genetics and Evolution

Chapter 12: Genetics and Evolution
SCIENCE MASTERY ASSESSMENT
Every pre-med knows this feeling: there is so much content I have to know for the MCAT! How do I know what to do first or what's important?
While the high-yield badges throughout this book will help you identify the most important topics, this Science Mastery Assessment is another tool in your MCAT prep arsenal. This quiz (which can also be taken in your online resources) and the guidance below will help ensure that you are spending the appropriate amount of time on this chapter based on your personal strengths and weaknesses. Don't worry though— skipping something now does not mean you'll never study it. Later on in your prep, as you complete full-length tests, you'll uncover specific pieces of content that you need to review and can come back to these chapters as appropriate.
How to Use This Assessment
If you answer 0–7 questions correctly:
Spend about 1 hour to read this chapter in full and take limited notes throughout. Follow up by reviewing all quiz questions to ensure that you now understand how to solve each one.
If you answer 8–11 questions correctly:
Spend 20–40 minutes reviewing the quiz questions. Beginning with the questions you missed, read and take notes on the corresponding subchapters. For questions you answered correctly, ensure your thinking matches that of the explanation and you understand why each choice was correct or incorrect.
If you answer 12–15 questions correctly:
Spend less than 20 minutes reviewing all questions from the quiz. If you missed any, then include a quick read-through of the corresponding subchapters, or even just the relevant content within a subchapter, as part of your question review. For questions you answered correctly, ensure your thinking matches that of the explanation and review the Concept Summary at the end of the chapter.
- What is the gene order of linked genes M, N, O, and P, given the following recombination frequencies?
MN: 6% NO: 18%
MO: 12% NP: 1%
MP: 5% OP: 17%
- MOPN
- NPMO
- ONPM
- PNMO
- Suppose that in a mammalian species, the allele for black hair (B) is dominant to the allele for brown hair (b), and the allele for curly hair (C) is dominant to the allele for straight hair (c). When an organism of unknown genotype is crossed against one with straight, brown hair, the phenotypic ratio is as follows:
- 25% curly black hair
- 25% straight black hair
- 25% curly brown hair
- 25% straight brown hair
What is the genotype of the unknown parent?
- BbCC
- bbCc
- Bbcc
- BbCc
- If a genotypical male who has hemophilia (XhY) is crossed with a genotypical female carrier of both color blindness and hemophilia (XcXh), what is the probability that a genotypically female child would be born without either condition?
- 0%
- 25%
- 50%
- 100%
- If a test cross on a species of plant reveals the appearance of a recessive phenotype in the offspring, what must be true of the phenotypically dominant parent?
- It must be genotypically heterozygous.
- It must be genotypically homozygous.
- It could be either genotypically heterozygous or homozygous.
- It must have the same genotype as the test cross control parent.
- Which of the following definitions is FALSE?
- Penetrance—the percentage of individuals in the population carrying the allele who actually express the phenotype associated with it
- Expressivity—the percentage of individuals in the population carrying the allele who do not express the phenotype associated with it
- Incomplete dominance—occurs when the phenotype of the heterozygote is an intermediate of the phenotypes of the homozygotes
- Codominance—occurs when multiple alleles exist for a given gene and more than one of them is dominant
- In a species of plant, a homozygous red flower (RR) is crossed with a homozygous yellow flower (rr). If the F1 generation is self-crossed and the F2 generation has a phenotypic ratio of red:orange:yellow of 1:2:1, which characteristic accounts for these results?
- Codominance
- Incomplete dominance
- Penetrance
- Expressivity
- Which of the following statements is INCORRECT regarding inheritance of traits according to the modern synthesis model?
- A mutation due to excessive amounts of ultraviolet light occurs in an unfertilized egg; this will affect the child who is born from that egg.
- The muscular strength gained by a weight lifter over a lifetime is inherited by the weight lifter’s children.
- A green-feathered bird that survived all of the predators in the forest will pass on the green feather genes to its offspring.
- A flower with a tasty nectar eaten by a butterfly is more likely to pass on its genes through the pollen spread by the butterfly than a flower with less desirable nectar.
- Which of the following statements is FALSE based on Darwin’s theory of natural selection?
- Natural selection is the driving force for evolution.
- Favorable genetic variations become more and more common in individuals throughout their lives.
- Natural selection can drive organisms living in groups to ultimately become distinct species.
- Fitness is measured by reproductive success.
- Which of the following is NOT a necessary condition for Hardy–Weinberg equilibrium?
- Large population size
- No mutations
- Monogamous mating partners
- No migration into or out of the population
- As the climate became colder during the Ice Age, a particular species of mammal evolved a thicker layer of fur. What kind of selection occurred in this population?
- Stabilizing selection
- Directional selection
- Disruptive selection
- Speciation selection
- At what point are two populations descended from the same ancestral stock considered to be separate species?
- When they can no longer produce viable, fertile offspring
- When they look significantly different from each other
- When they can interbreed successfully and produce offspring
- When their habitats are separated by a significantly large distance so that they cannot meet
- In a nonevolving population, there are two alleles, R and r, which code for the same trait. The frequency of R is 30 percent. What are the frequencies of all the possible genotypes?
- 49% RR, 42% Rr, 9% rr
- 30% RR, 21% Rr, 49% rr
- 0.09% RR, 0.42% Rr, 0.49% rr
- 9% RR, 42% Rr, 49% rr
- In a particular Hardy–Weinberg population, there are only two eye colors: brown and blue. Of the population, 36% have blue eyes, the recessive trait. What percentage of the population is heterozygous?
- 24%
- 48%
- 60%
- 64%
- Tay-Sachs disease is an autosomal recessive disorder characterized by defective hexosaminidase A. Proteomic analysis of a cell affected by this condition revealed a shortened protein with an abnormal amino acid sequence at the C-terminus. Based on this information, which mutation is mostly likely?
- Silent mutation
- Missense mutation
- Nonsense mutation
- Frameshift mutation
- A child is born with a number of rare phenotypic features and genetic testing is performed. The child is determined to have partial trisomy 21, with three copies of some segments of DNA from chromosome 21, and partial monosomy 4, with only one copy of some segments of DNA from chromosome 4. Which of the following mutations could have occurred in one of the parental gametes during development to explain both findings?
- Deletion
- Insertion
- Translocation
- Inversion
Answer Key
- B
- D
- C
- A
- B
- B
- B
- B
- C
- B
- A
- D
- B
- D
- C
Chapter 12: Genetics and Evolution
CHAPTER 12
GENETICS AND EVOLUTION
In This Chapter
12.1 Fundamental Concepts of Genetics
Patterns of Dominance
Penetrance and Expressivity
Mendelian Concepts
DNA as Genetic Material
12.2 Changes in the Gene Pool
Mutations
Leakage
Genetic Drift
12.3 Analytical Approaches in Genetics
Punnett Squares
Gene Mapping
Hardy–Weinberg Principle
12.4 Evolution
Natural Selection
Speciation
Measuring Evolutionary Time
Concept Summary
CHAPTER PROFILE
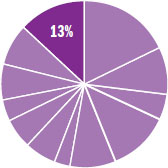
The content in this chapter should be relevant to about 13% of all questions about biology on the MCAT.
This chapter covers material from the following AAMC content category:
1C: Transmission of heritable information from generation to generation and the processes that increase genetic diversity
Introduction
For generations, European royal families practiced what is known as royal intermarriage. For purposes of establishing or continuing political alliances, maintaining bloodline purity, or smoothing out diplomatic relations, marriages between royal families were arranged, resulting in such an interweaving of bloodlines that eventually much of European royalty was—and still is—genetically related.
Such marriage unions led to rather severe restrictions on the gene pool, or all the alleles represented in the royal family lines. Offspring of parents who were also related to each other through blood lineage (consanguinity) came to have greater similarities in their genotypes, and certain alleles became so frequent that their phenotypic expression became almost a hallmark of royal descent. The House of Habsburg, which ruled a number of European kingdoms from the 11th century until the late 18th century, was perhaps the most infamous for its inbreeding practices. Members of this royal family bore the unmistakable mark of their restricted genes through a jaw malformation that even came to be known as the Habsburg lip. Medically termed prognathism (Greek: “forward jaw”), the condition is a misalignment of the mandible and maxilla. The Habsburg family portraits present individuals with prominent, forward-thrusting lower jaws and chins characteristic of mandibular prognathism. The genetic condition has more than just aesthetic implications; it can lead to serious disfigurement and disability. Charles II of Spain suffered from the worst case of the Habsburg lip on record—his lower teeth protruded so much farther than his upper teeth that he was not able to chew his food.
In this chapter, we will explore the concepts of classical genetics, which were originally described in the mid-1800s. Consider this chapter in tandem with the discussions of molecular genetics in Chapters 6 and 7 of MCAT Biochemistry Review. Then, we will explore the changes in the gene pool that occur over time with a discussion of evolution. We’ll also quantify the genetics of populations that are not undergoing evolution with the Hardy–Weinberg principle.
12.1 Fundamental Concepts of Genetics
LEARNING OBJECTIVES
After Chapter 12.1, you will be able to:
- Recall what traits make alleles dominant or recessive
- Differentiate between homozygous, heterozygous, and hemizygous genotypes
- Compare and contrast complete dominance, codominance, and incomplete dominance
- Explain the difference between penetrance and expressivity
- Connect Mendel’s laws with the phase of meiosis to which they are most closely correlated
The physical and biochemical characteristics of every living organism are determined by genes, which are DNA sequences that code for heritable traits that can be passed from one generation to the next. Taken together, all genes (as well as a large supply of noncoding DNA) are organized into chromosomes to ensure that genetic material is passed easily to daughter cells during mitosis and meiosis. Each gene may have alternative forms called alleles. We’ve already explored the ABO blood antigens as an example of three alleles for the same gene (IA, IB, and i). The genetic combination possessed by an individual is known as a genotype, and the manifestation of a given genotype as an observable trait is known as a phenotype.
BRIDGE
Blood type A individuals carry the A antigen on their erythrocytes and have circulating anti-B antibodies. Blood type B individuals carry the B antigen on their erythrocytes and have circulating anti-A antibodies. Those with type AB have both antigens and neither antibody; those with type O have neither antigen and both antibodies. That makes type O individuals universal donors and type AB individuals universal recipients. Blood typing is discussed in Chapter 7 of MCAT Biology Review.
Human beings typically possess two copies of each chromosome, called homologues, with the exception of the sex chromosomes of genotypical males, who have one X chromosome and one Y chromosome. Each gene has a particular locus, or location on a specific chromosome. The normal locus of a particular gene is consistent among human beings—a gene can be described by its location. Because each chromosome is part of a homologous pair, a person will inherit two alleles for all genes (again, except for male sex chromosomes). Alleles can be categorized based on their expression. If only one copy of an allele is needed to express a given phenotype, the allele is said to be dominant and is usually represented with a capital letter. If two copies are needed, the allele is said to be recessive and is usually represented with a lowercase letter. If both alleles are the same for a given gene, the individual is said to have a homozygous genotype. If the alleles are different, the individual has a heterozygous genotype. A hemizygous genotype describes a situation in which only one allele is present for a given gene, as is the case for parts of the X chromosome in genotypical males.
Patterns of Dominance
When only one dominant and one recessive allele exist for a given gene, there is said to be complete dominance. In this case, the presence of one dominant allele will mask the recessive allele, if present. When more than one dominant allele exists for a given gene, there is codominance. For example, a person with one allele for the A blood antigen and one allele for the B blood antigen will express both antigens simultaneously. Finally, incomplete dominance occurs when a heterozygote expresses a phenotype that is intermediate between the two homozygous genotypes. A classic example of incomplete dominance is the mating of certain flowers, in which a red flower crossed with a white flower results in pink flowers, as shown in Figure 12.1.
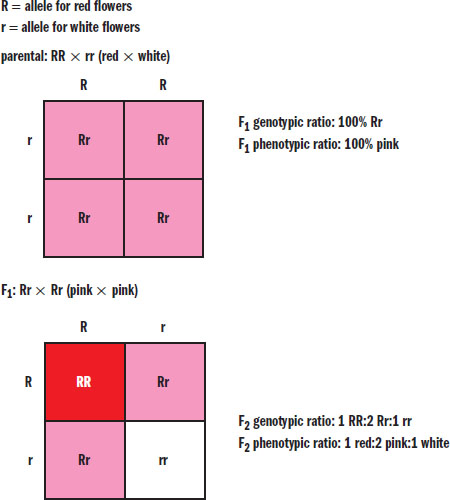
Figure 12.1. Incomplete Dominance Snapdragons display incomplete dominance, in which neither allele is dominant and the heterozygous phenotype is a mixture of the two homozygous phenotypes.
Penetrance and Expressivity
Penetrance and expressivity both reveal the complex interplay between genes and the environment. Penetrance is a population measure defined as the proportion of individuals in the population carrying the allele who actually express the phenotype. In other words, it is the probability that, given a particular genotype, a person will express the phenotype. Alleles can be classified by their degree of penetrance; Huntington’s disease, caused by an expansion of a repetitive sequence in the huntingtin gene, is a classic example. Individuals with more than 40 sequence repeats have full penetrance—100 percent of individuals with this allele show symptoms of Huntington’s disease. Individuals with fewer sequence repeats show high penetrance, wherein most (but not all) of those with the allele show symptoms of the disease. With fewer sequence repeats, the gene comes to have reduced penetrance, low penetrance, or even nonpenetrance.
A related but distinct concept is expressivity, which is defined as varying phenotypes despite identical genotypes. If expressivity is constant, then all individuals with a given genotype express the same phenotype. However, if expressivity is variable, then individuals with the same genotype may have different phenotypes. Whereas penetrance is a population parameter (what percentage of individuals with a given genotype express the phenotype?), expressivity reflects the gray area in expression and is more commonly considered at the individual level. For example, the disease neurofibromatosis type II is an autosomal dominant disease that results from a mutation of the gene NF2 (merlin). Interestingly, a range of phenotypes is associated with carrying the defective allele. Many patients have debilitating tumors of the vestibulocochlear nerve, which is needed for hearing and balance. Some have cataracts, while others have tumors in the skin called neuromas; still others have spinal lesions. A small proportion of the population is nonpenetrant. The disease shows variable expressivity because presentations range from no clinical effect to severe disability.
KEY CONCEPT
- Penetrance—the proportion of the population with a given genotype who actually express the phenotype
- Expressivity—the different manifestations of the same genotype across the population
Mendelian Concepts
Gregor Mendel, an Augustinian friar, developed several of the tenets of genetics in the 1860s based on his work with pea plants. While the study of genetics has come a long way, from pedigree analysis to DNA probes to whole-genome sequencing, many of Mendel’s original ideas still hold.
Mendel’s First Law: Law of Segregation
There are four basic tenets of the modern interpretation of Mendel’s first law (of segregation), some which have already been discussed:
- Genes exist in alternative forms (alleles).
- An organism has two alleles for each gene—one inherited from each parent.
- The two alleles segregate during meiosis, resulting in gametes that carry only one allele for any inherited trait.
- If two alleles of an organism are different, only one will be fully expressed and the other will be silent. The expressed allele is said to be dominant, while the silent allele is recessive. (Keep in mind that codominance and incomplete dominance are exceptions to this rule.)
The key cellular correlate to draw here is the separation of homologous chromosomes during anaphase I of meiosis. By separating—segregating—these chromosomes into different cells, each gamete carries only one allele for any given trait.
Mendel’s Second Law: Law of Independent Assortment
Mendel’s second law (of independent assortment) states that the inheritance of one gene does not affect the inheritance of another gene. Remember from Chapter 2 of MCAT Biology Review that spermatogonia and oogonia undergo genome replication before meiosis I. The daughter DNA strand is held to the parent strand at the centromere. Together, these DNA strands are known as sister chromatids. During prophase I of meiosis, homologous chromosomes pair up to form tetrads, which derive their name from the four chromatids involved (two chromatids in each of two homologous chromosomes). Small segments of genetic material are swapped between chromatids in homologous chromosomes, resulting in novel combinations of alleles that were not present in the original chromosomes (recombination). This allows the inheritance of one gene to be independent of the inheritance of all others.
Mendel’s second law has been complicated by the discovery of linked genes. We will discuss nonindependent assortment and linkage later in this chapter in the section on analytical approaches in genetics.
Both segregation of homologous chromosomes and independent assortment of alleles increase the genetic diversity of gametes and, subsequently, the genetic diversity of offspring. This has been demonstrated to improve the ability of a species to evolve and adapt to environmental stresses.
KEY CONCEPT
Segregation of chromosomes and independent assortment of alleles allow for greater genetic diversity in the offspring.
BRIDGE
The transforming principle described in Griffith’s experiment is the same as bacterial transformation discussed in Chapter 1 of MCAT Biology Review. Remember that transformation is one of three main ways bacteria increase genetic variability; the other two are conjugation and transduction.
DNA as Genetic Material
While Mendel noticed there were certain patterns of inheritance, what he did not know was that DNA was the genetic material transferred to offspring and that genes were made of DNA. In fact, the scientific community rejected Mendel’s initial papers on inheritance. It wasn’t until the early 1900s that his work was rediscovered. In the early to mid-1900s, it was largely believed that protein was the heritable material. In the mid-1900s, there were three experiments conducted that largely pointed to DNA’s role in genetic inheritance.
In the 1920s, Frederick Griffith was a scientist working for the British government, studying Streptococcus pneumoniae, a bacteria that causes pneumonia. Two strains of S. pneumoniae were identified: a virulent (disease-causing) strain and a nonvirulent strain. In successive trials, Griffith exposed mice to these strains of bacteria under different conditions and observed whether the mice lived or died, as shown in Figure 12.2. The virulent S. pneumonia has a smooth capsule that helps the bacterium evade the immune system and cause disease. He injected this strain into mice, which resulted in death of the mice. Naturally, if the virulent bacteria were killed prior to injection, no disease resulted. Likewise, exposure of the mice to the nonvirulent strain, which has a rough capsule, did not cause disease. However, when both dead virulent bacteria and live nonvirulent bacteria were injected into the mouse, the mouse died and live bacteria with smooth capsules could be found in the mice. He theorized that the live, nonvirulent bacteria must have acquired the ability to form smooth capsules from the dead virulent bacteria. This was known as the transforming principle.
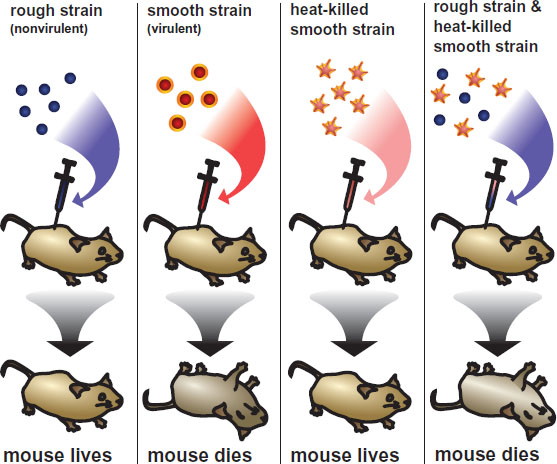
Figure 12.2. The Griffith Experiment
Researchers at the Rockefeller Institute confirmed the transformation principle. Three American scientists—Oswald Avery, Colin MacLeod, and Maclyn McCarty—were attempting to identify the exact material underlying the transformation principle. These scientists purified a very large quantity of heat-killed virulent S. pneumoniae bacteria and separated the subcellular components of the bacteria into different extracts. It was noted that the addition of one particular extract to nonvirulent S. pneumoniae transformed the bacteria and enabled them to kill the mouse when injected. When this substance was treated with enzymes known to degrade DNA, the bacteria were not transformed and the mice lived. However, when the substance was treated with enzymes known to degrade proteins, the bacteria were still transformed and the mice died. Thus, the group concluded that the transforming substance must be DNA.
In 1952 (one year before the description of the Watson–Crick model), Alfred Hershey and Martha Chase worked to confirm the idea that DNA could independently carry genetic information. These scientists created bacteriophages with radiolabeled DNA and protein. One group of bacteriophages contained radiolabeled sulfur, which is found in protein but not in DNA. Another group contained radiolabeled phosphorus, which is found in DNA but not in protein. Each of these bacteriophages was permitted to infect a group of nonlabeled bacteria. Recall from Chapter 1 of MCAT Biology Review that when bacteriophages infect a bacterium, they inject their genetic material into the cell and leave their capsid outside. After the phages and bacteria were incubated, the sample was centrifuged to separate the material that remained outside the cell from the bacterial cells themselves. It was determined that while no radiolabeled protein entered the cells, radiolabeled DNA had. It was known that viruses must enter a cell to cause disease and replicate, so this experiment once again helped confirm that DNA was the heritable genetic material.
BRIDGE
Radiolabeling sulfur was an appropriate choice to tag proteins in the Hershey–Chase experiment. Recall from Chapter 1 of MCAT Biochemistry Review that two amino acids—cysteine and methionine—contain sulfur in their R group, while no nucleotides contain any sulfur.
Epigenetics
Epigenetics is a general term for changes in DNA that do not involve an alteration to the nucleotide sequence. The prefix epi– is Greek for “over” or “above,” so the name suggests regulatory mechanisms that are in addition to the traditional features of inheritance. Epigenetic modifications can include the covalent attachment of different chemical groups to nucleotides and histone proteins, including but not limited to methylation or acetylation. These modifications can be temporary and function to increase or decrease the expression of specific genes. In general, modifications like methylation of DNA promoter regions tends to decrease the expression of specific genes, whereas modifications of lysines and/or arginines on histones have variable effects on the expression of genes.
Because almost all cells in the body contain the same genetic code, epigenetics allows cells to undergo differentiation, where different cells are able to express certain genes while silencing others. One prominent example of this is X-inactivation in cells that have two X chromosomes, in which one of the two X chromosomes is transcriptionally silenced via methylation; the inactive X chromosome is called a Barr body.
Epigenetic changes can have significant clinical impact. Hypermethylation of tumor suppressor genes and hypomethylation of oncogenes have been implicated in some forms of cancer. Furthermore, studies have shown that epigenetic changes can be a mechanism by which organisms’ responses to their changing environment can be passed down to their progeny. Indeed, the Överkalix study found that the grandchildren of those who had experienced food scarcity had different health outcomes compared to the grandchildren of those who had not experienced famine.
Another important concept is imprinting, an epigenetic process in which gene expression is determined by the contributing parent. There are certain genes that are differentially methylated when inherited from one parent; one of the most well known examples of imprinting comes from a region on the long arm of chromosome 15. If the deletion of this region of the chromosome occurs on the paternal chromosome and the maternal copy is heavily methylated, it leads to a developmental disorder called Prader-Willi syndrome. Deletion of the same genomic region on the maternal chromosome and paternal methylation leads to a different outcome, Angelman syndrome.
BRIDGE
Methylation of DNA is commonly used for transcriptional regulation. Recall from Chapter 7 of the MCAT Biochemistry Review that the addition of methyl groups to adenine and cytosine is linked to gene silencing.
MCAT CONCEPT CHECK 12.1
Before you move on, assess your understanding of the material with these questions.
- What does it mean for an allele to be dominant? Recessive?
- Dominant:
- Recessive:
- What does it mean for a genotype to be homozygous? Heterozygous? Hemizygous?
- Homozygous:
- Heterozygous:
- Hemizygous:
- What is the difference between complete dominance, codominance, and incomplete dominance?
- Complete dominance:
- Codominance:
- Incomplete dominance:
- What is the difference between penetrance and expressivity?
- Penetrance:
- Expressivity:
- With which phase of meiosis does each of Mendel’s laws most closely correlate?
- Mendel’s first law:
- Mendel’s second law:
12.2 Changes in the Gene Pool
LEARNING OBJECTIVES
After Chapter 12.2, you will be able to:
- Identify the three main types of point mutations and the genetic changes they are associated with
- Recall the two main types of frameshift mutation
- Explain why genetic leakage would increase within the last century
- Describe the relationship between genetic drift, small population, and the founder effect
- Identify the main types of chromosomal mutation and the impact they have at the chromosomal level:
All of the alleles that exist within a species are known as the gene pool. When mutations or genetic leakage occur, new genes are introduced into the gene pool. Genetic variability is essential for the survival of a species because it allows it to evolve to adapt to changing environmental stresses. Certain traits may be more desirable than others and confer a selective advantage that allows for an individual to produce more viable, fertile offspring. In this section, we will consider genetic diversity and mutations, leakage, and genetic drift, which cause changes to the alleles or their frequency in the gene pool.
Mutations
A mutation is a change in DNA sequence, and it results in a mutant allele. Mutant alleles can be contrasted with their wild-type counterparts, which are alleles that are considered “normal” or “natural” and are ubiquitous in the study population. New mutations may be introduced in a variety of ways. Ionizing radiation, such as ultraviolet rays from the sun, and chemical exposures can damage DNA; substances that can cause mutations are called mutagens. DNA polymerase is subject to making mistakes during DNA replication, albeit at a very low rate; proofreading mechanisms help prevent mutations from occurring through this mechanism. Elements known as transposons can insert and remove themselves from the genome. If a transposon inserts in the middle of a coding sequence, the mutation will disrupt the gene.
Flawed proteins can arise in other ways without an underlying change in DNA sequence. Incorrect pairing of nucleotides during transcription or translation, or a tRNA molecule charged with the incorrect amino acid for its anticodon, can result in derangements of the normal amino acid sequence.
The major types of nucleotide-level mutations are discussed in great detail in Chapter 7 of MCAT Biochemistry Review, so we offer just a brief overview here of each type.
Nucleotide-Level Mutations
Many mutations occur at the level of a single nucleotide (or a very small number of nucleotides). These mutations are shown in Figure 12.3 and are summarized below.
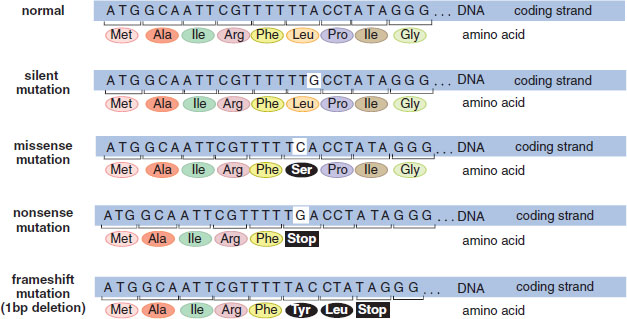
Figure 12.3. Common Nucleotide-Level Mutations
Point mutations occur when one nucleotide in DNA (A, C, T, or G) is swapped for another. These can be subcategorized as silent, missense, or nonsense mutations:
- Silent mutations occur when the change in nucleotide has no effect on the final protein synthesized from the gene. This most commonly occurs when the changed nucleotide is transcribed to be the third nucleotide in a codon because there is degeneracy (wobble) in the genetic code.
- Missense mutations occur when the change in nucleotide results in substituting one amino acid for another in the final protein.
- Nonsense mutations occur when the change in nucleotide results in substituting a stop codon for an amino acid in the final protein.
Frameshift mutations occur when nucleotides are inserted into or deleted from the genome. Because mRNA transcribed from DNA is always read in three-letter sequences called codons, insertion or deletion of nucleotides can shift the reading frame, usually resulting in either changes in the amino acid sequence or premature truncation of the protein (due to the generation of a nonsense mutation). These can be subcategorized as insertion or deletion mutations.
Chromosomal Mutations
Chromosomal mutations are larger-scale mutations in which large segments of DNA are affected, as demonstrated in Figure 12.4 and summarized below.
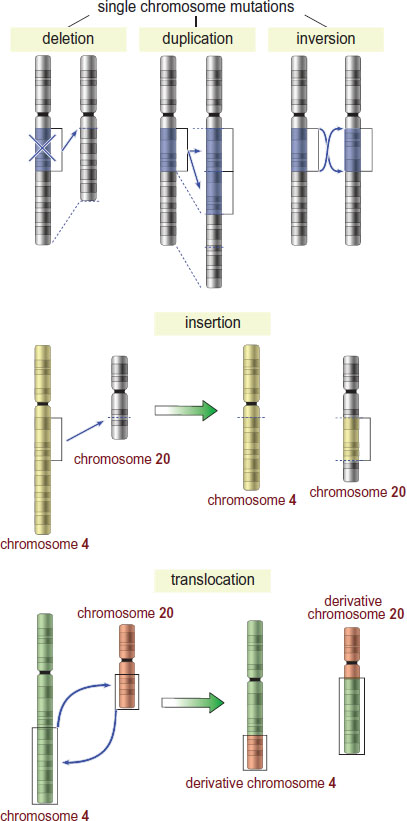
Figure 12.4. Common Chromosomal Mutations
- Deletion mutations occur when a large segment of DNA is lost from a chromosome. Small deletion mutations are considered frameshift mutations, as described previously.
- Duplication mutations occur when a segment of DNA is copied multiple times in the genome.
- Inversion mutations occur when a segment of DNA is reversed within the chromosome.
- Insertion mutations occur when a segment of DNA is moved from one chromosome to another. Small insertion mutations (including those where the inserted DNA is not from another chromosome) are considered frameshift mutations, as described previously.
- Translocation mutations occur when a segment of DNA from one chromosome is swapped with a segment of DNA from another chromosome.
Consequences of Mutations
Mutations can have many different consequences. Some mutations can be advantageous, conferring a positive selective advantage that may allow the organism to produce fitter offspring. For example, sickle cell disease is a single nucleotide mutation that causes sickled hemoglobin. While the disease itself is detrimental to life, heterozygotes for sickle cell disease usually have minor symptoms, if any, and have natural resistance to malaria because their red blood cells have a slightly shorter lifespan—just short enough that the parasitic Plasmodium species that causes malaria cannot reproduce in them. Thus, heterozygotes for sickle cell disease have a selective advantage because they are less likely to die from malaria.
On the other hand, some mutations can be detrimental or deleterious. For example, xeroderma pigmentosum (XP) is an inherited defect in the nucleotide excision repair mechanism. In patients with XP, DNA that has been damaged by ultraviolet radiation cannot be repaired appropriately. Ultraviolet radiation can introduce cancer-causing mutations; since they lack a repair mechanism, patients with XP are frequently diagnosed with malignancies, especially of the skin.
One important class of deleterious mutations is known as inborn errors of metabolism. These are deficiencies in genes required for metabolism. Children born with these deficient genes often require very early intervention in order to prevent permanent damage from the buildup of metabolites in various pathways. For example, in phenylketonuria (PKU), the enzyme phenylalanine hydrolase, which completes the metabolism of the amino acid phenylalanine, is defective. In the absence of this enzyme, toxic metabolites of phenylalanine accumulate, causing seizures, impairment of cerebral function, and learning disabilities, as well as a musty odor to bodily secretions. However, if the disease is discovered shortly after birth, then dietary phenylalanine can be eliminated and treatments can be administered to aid in metabolizing any remaining phenylalanine.
Leakage
Genetic leakage is a flow of genes between species. In some cases, individuals from different (but closely related) species can mate to produce hybrid offspring. Many hybrid offspring, such as the mule (hybrid of a male horse and a female donkey), are not able to reproduce because they have odd numbers of chromosomes—horses have 64 chromosomes and donkeys have 62, so mules, with 63 chromosomes, cannot undergo normal homologous pairing in meiosis and cannot form gametes. In some cases, however, a hybrid can reproduce with members of one species or the other, such as the beefalo (a cross between cattle and American bison). The hybrid carries genes from both parent species, so this can result in a net flow of genes from one species to the other.
Genetic Drift
Genetic drift refers to changes in the composition of the gene pool due to chance. Genetic drift tends to be more pronounced in small populations. The founder effect is a more extreme case of genetic drift in which a small population of a species finds itself in reproductive isolation from other populations as a result of natural barriers, catastrophic events, or other bottlenecks that drastically and suddenly reduce the size of the population available for breeding. Because the breeding group is small, inbreeding, or mating between two genetically related individuals, may occur in later generations. Inbreeding encourages homozygosity, which increases the prevalence of both homozygous dominant and recessive genotypes. Ultimately, genetic drift, the founder effect, and inbreeding cause a reduction in genetic diversity, which is often the reason why a small population may have increased prevalence of certain traits and diseases. For example, branched-chain ketoacid dehydrogenase deficiency (also called maple syrup urine disease) is especially common in Mennonite communities; this implies a common origin of the mutation, which may have been in a very small original population.
This loss of genetic variation may cause reduced fitness of the population, a condition known as inbreeding depression. On the opposite end of the spectrum, outbreeding or outcrossing is the introduction of unrelated individuals into a breeding group. Theoretically, this could result in increased variation within a gene pool and increased fitness of the population.
MCAT CONCEPT CHECK 12.2
Before you move on, assess your understanding of the material with these questions.
- What are the three main types of point mutations? What change occurs in each?
-
-
-
- What are the two main types of frameshift mutations?
-
-
- What are the three main types of chromosomal mutations that do NOT share their name with a type of frameshift mutation? What change occurs in each?
-
-
-
- Why would genetic leakage in animals be rare prior to the last century?
-
- Why is genetic drift more common in small populations? What relationship does this have to the founder effect?
12.3 Analytical Approaches in Genetics
LEARNING OBJECTIVES
After Chapter 12.3, you will be able to:
- Predict the phenotype ratio in the offspring of a cross, such as AaBB x AAbb
- Order genes on a chromosome given their recombination frequencies
- Recall the five criteria of the Hardy–Weinberg principle and the underlying population characteristic each one implies
- Solve calculation problems requiring use of the Hardy–Weinberg equations: p + q = 1, and p2 + 2pq + q2 = 1
Genetics is a field in which a number of biometric techniques, or quantitative approaches to biological data, have been developed. These range from pedigrees and the Punnett square to mapping of chromosomes with recombinant frequencies to Hardy–Weinberg equilibrium.
Pedigrees
Pedigree charts are diagrams that show relationships between people within families and indicate the presence or absence of a trait of interest. On the MCAT, only a basic understanding of pedigree charts is required: squares represent biological males, circles represent biological females, empty shapes represent the absence of a trait, and colored-in shapes represent the presence of a trait. Figure 12.5 shows an example of a pedigree chart showing three generations of a family (A, B, and C). In generation A, an unaffected male (A1) and female (A2) had two unaffected children (B1 and B2), while a female with albinism (A3) had two unaffected children (B3 and B4) with an unaffected male (A4). In generation C, two of the three children (C1 and C2) of B2 and B3 also had albinism, while the third female child (C3) was unaffected.
MCAT EXPERTISE
The MCAT will only test a fundamental understanding of how to read pedigree charts. The symbology of pedigree analysis is complex and intricate, and beyond the scope of the MCAT.
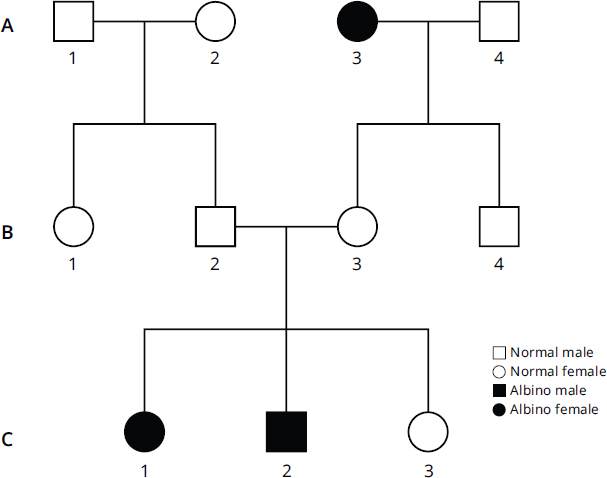
Figure 12.5. Example of a Pedigree Chart The black square represents a male with albinism, while the black circles represent females with albinism.
Commonly, these types of charts are used to help determine the inheritance pattern of the trait of interest by looking at its inheritance pattern among family generations. The types of inheritance patterns most commonly tested on the MCAT include autosomal dominant, autosomal recessive, X-linked recessive, mitochondrial, and Y-linked.
In an autosomal dominant pattern of inheritance, the trait is encoded by a dominant gene on an autosomal chromosome. A tell-tale sign of an autosomal dominant pattern of inheritance is the trait never skips a generation: In other words, an affected child must have at least one affected parent, as demonstrated in Figure 12.6. However, affected parents can have unaffected children if the affected parent is heterozygous and carries one allele that codes for the trait and one allele that does not. Therefore, another indication a pedigree is showing an autosomal dominant pattern of inheritance is two affected parents having an unaffected child.
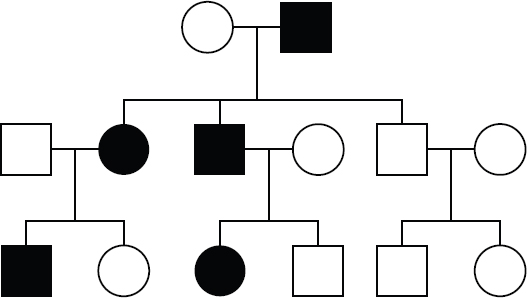
Figure 12.6. An Autosomal Dominant Pattern of Inheritance
In contrast, a pedigree like the ones shown in Figures 12.7 and 12.8, where unaffected individuals have affected children, is indicative of a trait encoded by a recessive allele. In an autosomal recessive pattern of inheritance, where the trait is only expressed if an individual has two copies of the recessive allele, both males and females are equally likely to have the trait. Another indication of a recessive trait is when two affected parents only have affected children.
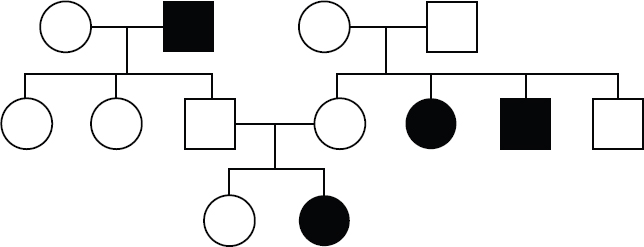
Figure 12.7. An Autosomal Recessive Pattern of Inheritance Multiple affected members of this family have unaffected parents, suggesting autosomal recessive inheritance.
In an X-linked recessive pattern of inheritance, where the trait is encoded by a gene located on the X chromosome, males are disproportionately affected. This is due to the fact that males have only one copy of the X chromosome, while females have two and require both genes to be affected to express the trait. X-linked recessive and autosomal recessive patterns of inheritance can look similar, but there are certain indicators that can favor one inheritance pattern over the other. For example, in X-linked recessive patterns of inheritance, all male children of an affected female will express the trait.
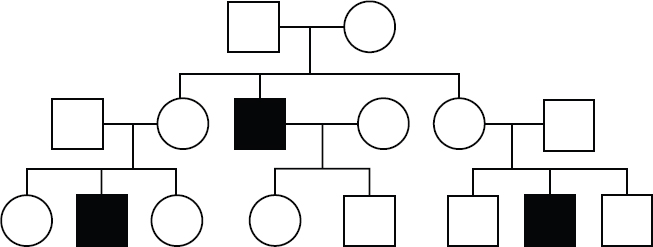
Figure 12.8. An X-Linked Recessive Pattern of Inheritance Males are disproportionately affected and can have unaffected parents.
Mitochondrial inheritance occurs when the mutated allele is located on mitochondrial DNA rather than nuclear DNA. While both males and females are equally affected by these types of mutations, mitochondrial DNA is only passed down from the mother in a phenomenon known as maternal inheritance. Therefore, all children of an affected mother will also have the trait, as demonstrated by the pedigree shown in Figure 12.9. On the flip side, no child of an affected male and unaffected female will inherit the trait.
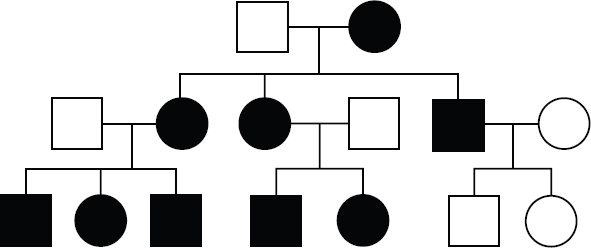
Figure 12.9. A Mitochondrial Pattern of Inheritance All children of affected females also express the trait.
A Y-linked pattern of inheritance is less frequently tested on the MCAT, but refers to an inheritance pattern where a trait is carried on the Y chromosome. Because only males have Y chromosomes, only males are affected and females are unable to pass on the trait. Furthermore, any son of any affected male will also be affected.
Punnett Squares
Punnett squares are diagrams that predict the relative genotypic and phenotypic frequencies that will result from the crossing of two individuals. The alleles of the two parents are arranged on the top and side of the square, with the genotypes of the progeny represented at the intersections of these alleles. The genotypes of the progeny will be the product of the two parental alleles.
Monohybrid Cross
In genetics problems, including those on the MCAT, dominant alleles are assigned capital letters and recessive alleles are assigned lowercase letters. If both copies of the allele are the same, that individual is said to be homozygous; if they are different, the individual is heterozygous.
A cross in which only one trait is being studied is said to be monohybrid. The parent or P generation refers to the individuals being crossed; the offspring are the filial or F generation. Multiple generations can be denoted F generations by using numeric subscripts. If you think of your grandparents as the P generation, then your parents are in the F1 generation, and you are in the F2 generation.
Mendel worked with pea plants that had either purple or white flowers. Before crossing the different plants, each group contained homozygotes; subsequent experimentation revealed that the allele for purple color was dominant (P) and the allele for white color was recessive (p). Thus, crossing a homozygous purple flower with a white flower would be crossing PP with pp, resulting in an F1 generation that contained 100 percent Pp or heterozygotes, as shown in Figure 12.10. All of the flowers in this generation would be purple because P is a dominant allele.
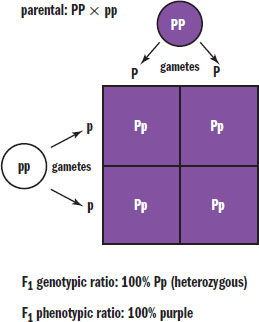
Figure 12.10. Punnett Square of Homozygous Parents
If two members of the F1 generation were crossed, the resulting offspring in the F2 generation would be more genotypically and phenotypically diverse than their parents. Crossing two plants with the genotype Pp would result in 25 percent PP, 50 percent Pp, and 25 percent pp offspring, as shown in Figure 12.11. Phenotypically, this would be a 3:1 distribution because both the homozygous dominant and heterozygous dominant offspring would be purple-flowering plants. Thus, crossing two heterozygotes in a case of complete dominance will result in a 1:2:1 distribution of genotypes (homozygous dominant:heterozygous dominant:homozygous recessive) and a 3:1 distribution of phenotypes (dominant:recessive). These ratios are, of course, theoretical probabilities and will not always hold true—especially in a small population of offspring. Usually, the more offspring parents have, the closer their phenotypic ratios will be to the expected ratios.
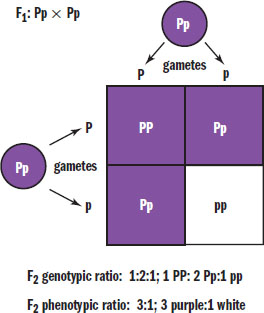
Figure 12.11. Punnett Square of Heterozygous Parents
KEY CONCEPT
Crossing two heterozygotes for a trait with complete dominance results in a 1:2:1 ratio of genotypes and a 3:1 ratio of phenotypes. Know these ratios cold for Test Day!
MCAT EXPERTISE
The ability to create and read a Punnett square quickly on Test Day is one of the most useful skills for questions involving Mendelian inheritance. Often, an entire passage in the Biological and Biochemical Foundations of Living Systems section will be devoted to classical and molecular genetics and will require the use of at least one Punnett square.
Test Cross
A test cross is used to determine an unknown genotype, as shown in Figure 12.12. In a test cross, the organism with an unknown genotype is crossed with an organism known to be homozygous recessive. If all of the offspring (100 percent) are of the dominant phenotype, then the unknown genotype is likely to be homozygous dominant. If there is a 1:1 distribution of dominant to recessive phenotypes, then the unknown genotype is likely to be heterozygous. Because a test cross is used to determine the genotype of the parent based on the phenotypes of its offspring, test crosses are sometimes called back crosses.
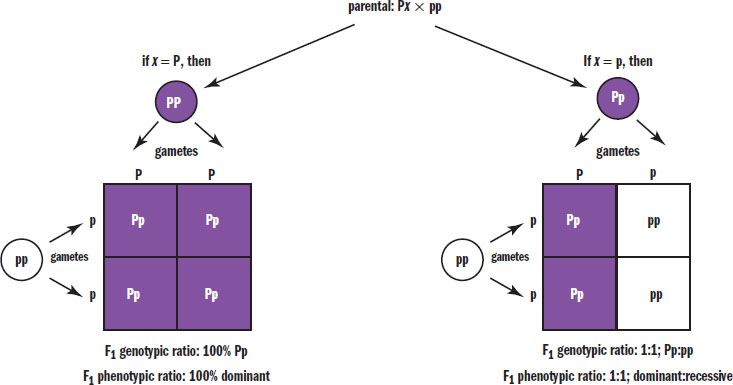
Figure 12.12. Test Cross An organism with an unknown genotype is crossed with a homozygous recessive organism to identify the unknown genotype using the phenotypes of the resulting offspring.
Dihybrid Cross
We can extend a Punnett square to account for the inheritance of two different genes using a dihybrid cross. Remember, according to Mendel’s second law (of independent assortment) the inheritance of one gene is independent of the inheritance of the other. This will hold true for unlinked genes, although it will be more complicated for linked genes, as described later in this chapter.
If we expand the previous crosses to consider not only flower color, but also plant height, then we can create a 4 × 4 Punnett square as shown in Figure 12.13. Remember that purple is dominant (P) and white is recessive (p); similarly, tall is dominant (T) and short or dwarf is recessive (t). If we cross two plants that are heterozygous for both traits, then the offspring have a phenotypic ratio of 9:3:3:1 (9 tall and purple:3 tall and white:3 dwarf and purple:1 dwarf and white). Note that the 3:1 phenotypic ratio still holds for each trait (12 tall:4 dwarf and 12 purple:4 white), reflecting Mendel’s second law.
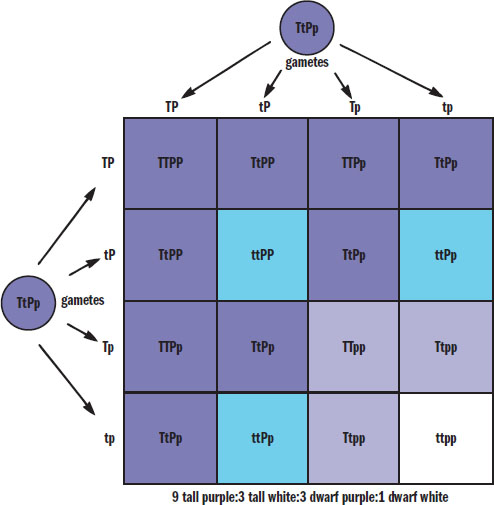
Figure 12.13. Dihybrid Cross
MCAT EXPERTISE
Just as with the 3:1 phenotypic ratio of a monohybrid cross, it is worth memorizing the 9:3:3:1 distribution for dihybrid crosses between two heterozygotes with complete dominance.
Sex-Linked Crosses
When considering sex-linked (X-linked) traits, a slightly different system is used to symbolize the various alleles because genotypical females have two X chromosomes and thus may be homozygous or heterozygous for a condition carried on the X chromosome. Genotypical males have only one X chromosome (and one Y chromosome) and are hemizygous for many genes carried on the X chromosome. This is why sex-linked traits are much more common in genotypical males; having only one recessive allele is sufficient for expression of the recessive phenotype.
When writing genotypes for sex-linked traits, we use X and Y to symbolize normal X and Y chromosomes. An X chromosome carrying a defective allele is commonly given a subscript, such as Xh, to indicate the presence of the disease-carrying allele. Hemophilia is a particularly common example of a sex-linked trait; Punnett squares for a heterozygous (carrier) female and both an unaffected (normal) male and affected (hemophiliac) male are shown in Figure 12.14.
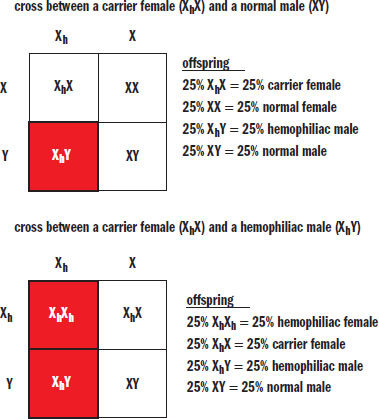
Figure 12.14. Sex-Linked Cross Unless stated otherwise, assume that all sex-linked traits on the MCAT are X-linked recessive.
KEY CONCEPT
Because an egg necessarily carries an X chromosome, the sperm determines the chromosomal or genotypical sex of a child. It follows that, for genotypical males with a sex-linked trait, genotypically female offspring will either carry the trait or express it, but genotypically male offspring will do neither (unless the X chromosome in the egg contains the affected allele, in which case they will express it).
Gene Mapping
Genes are organized in a linear fashion on chromosomes. As discussed earlier, crossing over during prophase I of meiosis causes alleles to be swapped between homologous chromosomes, supporting Mendel’s second law (of independent assortment). However, genes that are located very close together on a chromosome are less likely to be separated from each other during crossing over. In other words, the further apart two genes are, the more likely it is that there will be a point of crossing over, called a chiasma, between them. The likelihood that two alleles are separated from each other during crossing over, called the recombination frequency (θ), is roughly proportional to the distance between the genes on the chromosome. We can also describe the strength of linkage between genes based on the recombination frequency: tightly linked genes have recombination frequencies close to 0 percent; weakly linked genes have recombination frequencies approaching 50 percent, as expected from independent assortment.
By analyzing recombination frequencies, a genetic map representing the relative distance between genes on a chromosome can be constructed. By convention, one map unit or centimorgan corresponds to a 1 percent chance of recombination occurring between two genes. Thus, if two genes were 25 map units apart, we would expect 25 percent of the total gametes examined to show recombination somewhere between these two genes. Recombination frequencies can be added in a crude approximation to determine the order of genes in the chromosome, as shown in Figure 12.15.
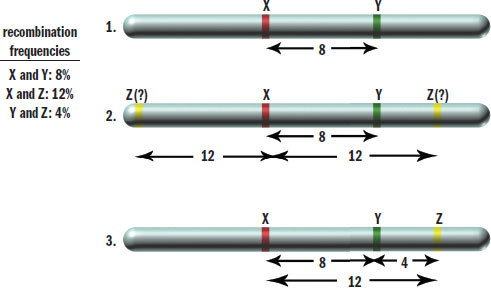
Figure 12.15. Genetic Maps from Recombination Frequencies If the recombination frequencies are known, one can deduce the order of genes on the chromosome because map units are roughly additive.
Hardy–Weinberg Principle
How often an allele appears in a population is known as its allele frequency. For example, if we took a one-cell sample from 50 of Mendel’s plants, we could collect 100 copies of alleles for flower color (two from each cell). If 75 of these alleles were the dominant allele, we could say that the allele frequency of P is 75 ÷ 100 = 0.75. Note that this does not indicate which flowers contain the allele or if those flowers are homozygous or heterozygous; it only tells us the representation of the allele across all chromosomes in the population. Evolution results from changes in these gene frequencies in reproducing populations over time. However, when the gene frequencies of a population are not changing, the gene pool is stable and evolution is ostensibly not occurring. The following five criteria are mandatory for this to be possible:
- The population is very large (no genetic drift).
- There are no mutations that affect the gene pool.
- Mating between individuals in the population is random (no sexual selection).
- There is no migration of individuals into or out of the population.
- The genes in the population are all equally successful at being reproduced.
Provided that all of these conditions are met, the population is said to be in Hardy–Weinberg equilibrium, and a pair of equations can be used to predict the allelic and phenotypic frequencies.
Let us define a particular gene as having only two possible alleles, T and t. We will define p to be the frequency of the dominant allele T and q to be the frequency of the recessive allele t. Because there are only these two choices at the same gene locus, p + q = 1. That is, the combined allele frequencies of T and t must equal 100 percent. If we square both sides of the equation, we get:
(p + q)2 = 12 p2 + 2pq + q2 = 1
where p2 is the frequency of the TT (homozygous dominant) genotype, 2pq is the frequency of the Tt (heterozygous dominant) genotype, and q2 is the frequency of the tt (homozygous recessive) genotype. Note that the sum p2 + 2pq would represent the frequency of the dominant phenotype (both homozygous and heterozygous dominant genotypes).
KEY CONCEPT
All you need to know to solve any MCAT Hardy–Weinberg problem is the value of p (or p2) or q (or q2). From there, you can calculate everything else using p + q = 1 and p2 + 2pq + q2 = 1.
For Test Day, you should know the two key Hardy–Weinberg equations demonstrated above:
p + q = 1 p2 + 2pq + q2 = 1
Equation 12.1
Each equation provides us with different information. The first tells us about the frequency of alleles in the population, whereas the second provides information about the frequency of genotypes and phenotypes in the population.
KEY CONCEPT
The Hardy–Weinberg equations allow you to find two pieces of information: first, the relative frequency of alleles in a population, and second, the frequency of a given genotype or phenotype in the population. Remember that there will be twice as many alleles as individuals in a population because each individual has two autosomal copies of each gene.
These equations can also be used to demonstrate that evolution is not occurring in a population. Assuming that the conditions listed earlier are met, the allele frequencies will remain constant from generation to generation. For example, imagine that we have a population of Mendel’s pea plants in which the frequency of the tall allele, T, is 0.80. This value is represented by p. This means that q (the short allele, t) is 0.20 by subtraction. Setting up our F1 cross for two heterozygotes, we can see the results of such a mating:

We see that the filial generation contains 64 percent homozygous tall, 32 percent heterozygous tall, and 4 percent homozygous short plants. These are the genotypic frequencies. We can determine the allele frequencies in this generation as follows:
64 % TT = 64 % T and 0 % t 32 % Tt = 16 % T and 16 % t 4 % tt = 0 % T and 4 % t allele frequencies = 80 % T and 20 % t
Notice that the allele frequencies are unchanged compared to the parent generation. T is still 0.80 and t is still 0.20. Populations in Hardy–Weinberg equilibrium will exhibit this property.
MCAT CONCEPT CHECK 12.3
Before you move on, assess your understanding of the material with these questions.
- For each of the crosses below, what is the phenotypic ratio seen in the offspring?
Cross Phenotypic Ratio
Bb × Bb Aa × aa DdEe × ddEE XqX × XY XrX × XrY
- If genes Q and R have a recombination frequency of 2%, genes R and S have a recombination frequency of 6%, genes S and T have a recombination frequency of 23%, and genes Q and T have a recombination frequency of 19%, then what is the order of these four genes in the chromosome?
- All five criteria of the Hardy–Weinberg principle are required to imply what characteristic of the study population?
- Assume that a population is in Hardy–Weinberg equilibrium. If 9% of the population is homozygous dominant, then solve for the following:
- The frequency of the dominant allele:
- The frequency of the recessive allele:
- The portion of the population that is heterozygous:
- The portion of the population with a homozygous recessive genotype:
- The portion of the population with a dominant phenotype:
12.4 Evolution
LEARNING OBJECTIVES
After Chapter 12.4, you will be able to:
- Describe the key tenets of the major theories of evolution, including punctuated equilibrium, natural selection, inclusive fitness, and the modern synthesis model
- Identify the three patterns of selection and the changes they create in the population phenotype
- Recall the three patterns of evolution between species and the outcome of each
- Recall the biological definition of a species
Evolutionary thought has a relatively short history; the first theories suggesting that new species may arise from older ones were proposed in the 19th century. Significant alterations to these initial theories have been made since then.
Natural Selection
Natural selection, sometimes called survival of the fittest, is the theory that certain characteristics or traits possessed by individuals within a species may help those individuals have greater reproductive success, thus passing on those traits to offspring. This theory was originally proposed by Charles Darwin in his 1859 publication On the Origin of Species. His theory was built on several basic tenets:
- Organisms produce offspring, few of which survive to reproductive maturity.
- Chance variations within individuals in a population may be heritable. If these variations give an organism even a slight survival advantage, the variation is termed favorable.
- Individuals with a greater preponderance of these favorable variations are more likely to survive to reproductive age and produce offspring; the overall result will be an increase in these traits in future generations. This level of reproductive success is termed fitness, and an organism’s fitness is directly related to the relative genetic contribution of this individual to the next generation.
Darwin’s theory was ultimately proven to be correct in many ways—although not completely. In the 20th century, modern genetics led to the development of the currently accepted theory.
KEY CONCEPT
Evolution is not equivalent to natural selection. The MCAT likes to test your ability to understand that natural selection is simply a mechanism for evolution. Natural selection is, however, equivalent to survival of the fittest.
Modern Theories
The modern synthesis model, sometimes called neo-Darwinism, adds knowledge of genetic inheritance and changes in the gene pool to Darwin’s original theory. Once scientists showed that inheritance occurs through the passing of genes from parent to child and that genes ultimately change due to mutation or recombination, Darwin’s theory was updated to its current form: when mutation or recombination results in a change that is favorable to the organism’s reproductive success, that change is more likely to pass on to the next generation. The opposite is also true. This process is termed differential reproduction. Over time, those traits passed on by the more successful organisms will become ubiquitous in the gene pool. Because the gene pool changes over time, it is important to note that populations evolve, not individuals.
Also germane to the modernization of Darwin’s theory is a shift in scope to focus on inclusive fitness over the fitness of an individual organism. Inclusive fitness is a measure of an organism’s success in the population, based on the number of offspring, success in supporting offspring, and the ability of the offspring to then support others. Early descriptions of evolutionary success, like those of Darwin, were based solely on the number of viable offspring of an organism. However, contemporary theories take into account the benefits of certain behaviors on the population at large. For example, the existence of altruism could be supported by the observation that close relatives of an individual will share many of the same genes; thus, promoting the reproduction and survival of related or similar individuals can also lead to genetic success. Other species show examples of inclusive fitness by protecting the offspring of the group at large. By endangering themselves to protect the young, these organisms ensure the passing of genes to future generations. Inclusive fitness therefore promotes the idea that altruistic behavior can improve the fitness and success of a species as a whole.
One final theory to consider was proposed as a result of research into the fossil record. Upon examination, it was discovered that little evolution would occur within a lineage of related lifeforms for long periods of time, followed by an explosion in evolutionary change. Niles Eldredge and Stephen Jay Gould proposed the theory of punctuated equilibrium to explain this in 1972. In contrast to Darwin’s theory, punctuated equilibrium suggests that changes in some species occur in rapid bursts rather than evenly over time.
Modes of Natural Selection
Natural selection may occur as stabilizing selection, directional selection, or disruptive selection, as shown in Figure 12.16.
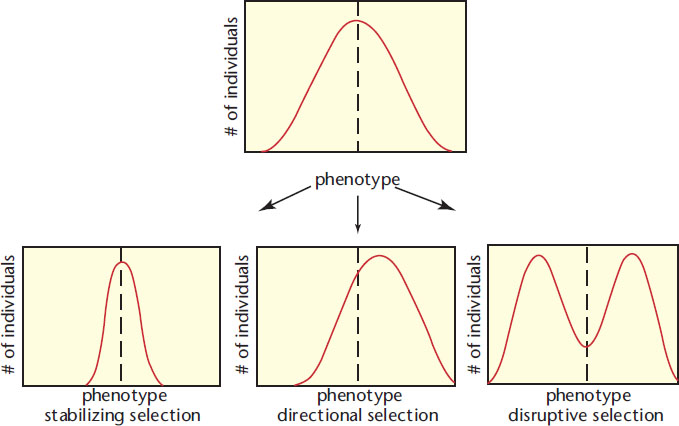
Figure 12.16. Modes of Natural Selection
Stabilizing selection keeps phenotypes within a specific range by selecting against extremes. For instance, human birth weight is maintained within a narrow band by stabilizing selection. Fetuses that weigh too little may not be healthy enough to survive, and fetuses that weigh too much can experience trauma during delivery through the relatively narrow birth canal. In addition, the larger the fetus, the more maternal resources it requires. For all of these reasons, there is a fitness advantage to keeping birth weights within a narrow range.
Adaptive pressure can lead to the emergence and dominance of an initially extreme phenotype through directional selection. For example, if we have a heterogeneous plate of bacteria, very few may have resistance to antibiotics. If the plate is then treated with ampicillin (an antibiotic), only those colonies that exhibit resistance to this antibiotic will survive. A new standard phenotype emerges as a result of differential survivorship. Natural selection is the history of differential survivorship over time. The emergence of mosquitoes resistant to dichlorodiphenyltrichloroethane (DDT), a type of pesticide, is attributed to directional selection.
In disruptive selection, two extreme phenotypes are selected over the norm. When Darwin studied finches on the Galapagos Islands, he noted that although there were many species, all the species arguably had a common ancestor, given their similar appearances. However, when he compared beak sizes they were all either large or small, as shown in Figure 12.17. No animals exhibited the intermediate phenotype of medium-size beaks. Darwin hypothesized that the sizes of seeds on the island (the finches’ food) led to this effect. Seeds were either quite large or fairly small, requiring a large or small beak, respectively. Thus, if the original ancestor had a medium-size beak, over time the animals with slightly larger or smaller beaks would be selected for. Disruptive selection is facilitated by the existence of polymorphisms—naturally occurring differences in form between members of the same population, such as light and dark coloration in the same species of butterfly. Adaptive radiation is a related concept that describes the rapid rise of a number of different species from a common ancestor. The benefit of adaptive radiation is that it allows for various species to occupy different niches. A niche is a specific environment, including habitat, available resources, and predators, for which a species is specifically adapted. Adaptive radiation is favored by environmental changes or isolation of small groups of the ancestral species.
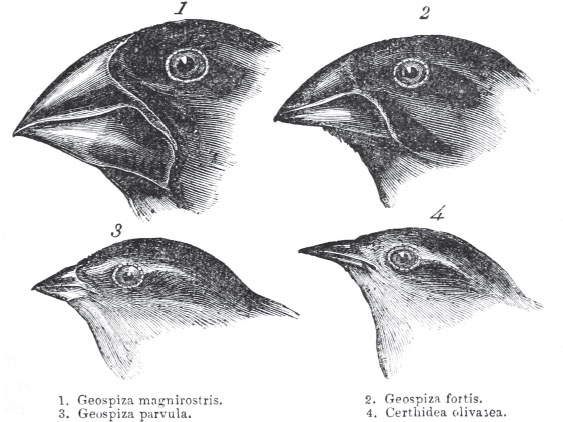
Figure 12.17. Darwin’s Finches Image of Darwin’s finches as drawn by John Gould
Speciation
A species is defined as the largest group of organisms capable of breeding to form fertile offspring; the formation of a new species through evolution is called speciation. If we took two populations from the same species and separated them geographically for a long period of time, different evolutionary pressures would lead to different adaptive changes. If enough time passed, the changes would be sufficient to lead to isolation, which means the progeny of these populations could no longer freely interbreed. We would now consider the two groups separate species.
Reproductive isolation may occur either prezygotically or postzygotically. Prezygotic mechanisms prevent formation of the zygote completely; postzygotic mechanisms allow for gamete fusion but yield either nonviable or sterile offspring. Examples of prezygotic mechanisms include temporal isolation (breeding at different times), ecological isolation (living in different niches within the same territory), behavioral isolation (a lack of attraction between members of the two species due to differences in pheromones, courtship displays, and so on), reproductive isolation (incompatibility of reproductive anatomy), or gametic isolation (intercourse can occur, but fertilization cannot). Postzygotic mechanisms include hybrid inviability (formation of a zygote that cannot develop to term), hybrid sterility (forming hybrid offspring that cannot reproduce), and hybrid breakdown (forming first-generation hybrid offspring that are viable and fertile, but second-generation hybrid offspring that are inviable or infertile). As described earlier in this chapter, mules are an example of postzygotic hybrid sterility. Although a horse and donkey can produce a viable mule, the mule will be sterile and thus unable to establish a self-perpetuating mule lineage.
Patterns of Evolution
When we look at similarities between two species, we must be careful to determine whether those similarities are due to sharing a common ancestor or sharing a common environment with the same evolutionary pressures. When analyzing species this way, three patterns of evolution emerge: divergent evolution, parallel evolution, and convergent evolution, as shown in Figure 12.18.
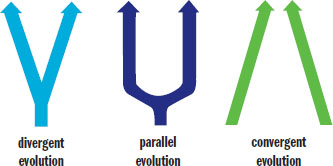
Figure 12.18. Patterns of Evolution
Divergent evolution refers to the independent development of dissimilar characteristics in two or more lineages sharing a common ancestor. For example, seals and cats are both mammals in the order Carnivora, yet they differ markedly in general appearance. These two species live in very different environments and adapted to different selection pressures while evolving.
Parallel evolution refers to the process whereby related species evolve in similar ways for a long period of time in response to analogous environmental selection pressures.
Convergent evolution refers to the independent development of similar characteristics in two or more lineages not sharing a recent common ancestor. For example, fish and dolphins have come to resemble one another physically, though they belong to different classes of vertebrates. They evolved certain similar features in adapting to the conditions of aquatic life.
Measuring Evolutionary Time
Evolution is a slow process, featuring changes in the environment and subsequent changes in genotypes and phenotypes of a population over time. The rate of evolution is measured by the rate of change of a genotype over a period of time and is related to the severity of the evolutionary pressures on the species. In other words, if a species is already perfectly suited to its habitat and there are no changes to the conditions in which it lives, the rate of evolution will be exceedingly slow—although there will still be some small base rate of genetic mutation. On the other hand, if an organism lives in a rapidly changing environment, the rate of evolution will be greater, as selection for and against certain traits will be actively occurring within that population.
By comparing DNA sequences between different species, scientists can quantify the degree of similarity between two organisms. For example, chimpanzees share over 95 percent of their genome with humans, whereas mice share only about 70 percent. As species become more taxonomically distant, the proportion of the shared genome will decrease. Molecular evolutionists correlate the degree of genomic similarity with the amount of time since two species split off from the same common ancestor; the more similar the genomes, the more recently the two species separated from each other. This is sometimes called the molecular clock model.
MCAT CONCEPT CHECK 12.4
Before you move on, assess your understanding of the material with these questions.
- What are the key tenets of each of the following theories of evolution?
- Natural selection:
- Modern synthesis model:
- Inclusive fitness:
- Punctuated equilibrium:
- What are the three patterns of selection? What changes would each create to the population phenotype?
Pattern of Selection Change to Population Phenotype
- What are the three patterns of evolution between species? What is the outcome of each one?
Pattern of Evolution Outcome
- What is the biological definition of a species?
BIOLOGY GUIDED EXAMPLE WITH EXPERT THINKING
While the genus Saccharomyces is best known for the model and industrial yeast S. cerevisiae, it also includes eight closely related additional species. The repeated isolation of S. cerevisiae from wine, beer, and other fermented beverages, and the difficulty in finding its truly natural habitats, has led to the common view that this species was a product of domestication. However, the unusual degree of shared physiological characteristics among Saccharomyces species along with the recent isolation of S. cerevisiae from a natural environment suggests S. cerevisiae may be a result of natural evolution. In the present work we used a comparative genomics approach and publicly available complete genome sequences of five Saccharomyces species to search for proteins exhibiting molecular patterns of evolution. We used the dN/dS ratio [rate of nonsynonymous substitutions per nonsynonymous site (dN)/rate of synonymous substitutions per synonymous site (dS)] of the complete Saccharomyces ORFeomes to measure protein divergence and corrected for phylogenetic distance. From this data, we created the phylogenetic tree shown in Figure 1 and searched for genes associated with divergence. Some of the identified genes had been previously associated with adaptation to growth at suboptimal temperatures. This prompted us to examine catabolic fluxes at different temperatures in Saccharomyces species with different growth temperature preferences. Figure 1 depicts schematically the phylogenetic relationships of the species in the genus Saccharomyces and assigns them to the thermotolerant or cryotolerant groups. Saccharomyces has 9 related species Issue: Is the formation of S. cervevisiae due to domestication or natural evolution? There's a lot of technical jargon here, so to rephrase, the key takeaway is that the researchers used comparative genomics to create the phylogenic tree in Figure 1 After creating the tree, the researchers saw that optimal growth temperature was a pattern that corresponded to the phylogeny
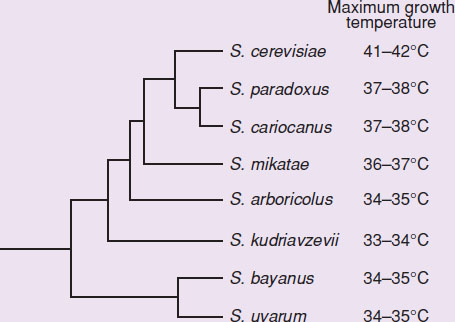
Figure 1
Trend: maximum growth temperature is highest at the top of the tree, and generally descends in order
Adapted from Gonçalves, P., Valério, E., Correia, C., de Almeida, J. M., & Sampaio, J. P. (2011). Evidence for divergent evolution of growth temperature preference in sympatric Saccharomyces species. PloS One, 6(6), e20739.
**Based on the information provided, could temperature have provided the selective pressure for the divergent evolution of Saccharomyces species?**
The question is asking us if there is support in the passage and figure for the hypothesis that temperature can explain the divergence of the various Saccharomyces species in the phylogenetic tree. Notice the phrasing of the question—it’s not asking if we have proof that the evolution was due to temperature selection. This analysis is retrospective, so we would have no way to verify whether or not the relationship between growth and temperature leading up to speciation was causal.
In paragraph 2, the researchers used fairly complex techniques to analyze the genomes of Saccharomyces species and arranged them in the phylogenetic tree. We don’t need to understand all of the details behind this method. The takeaway is that the phylogeny was generated prior to the analysis of optimal growth temperatures. The researchers then searched for genes associated with divergence and noticed that some of those genes corresponded to adaptations for growth at nonideal temperatures. Finally, an analysis of the optimal growth temperature for the listed Saccharomyces species gave us a workable pattern—species that are close together on the tree also have very similar optimal growth temperatures. We know from content background that changes in environment can definitely apply selection pressures strong enough to lead to speciation, or the formation of new species. Recall also that divergent evolution is the development of species with different characteristics despite a shared common ancestor.
Since temperature is an environmental factor, we can say that based on the given information, there is plausible evidence to support that temperature could have provided the selection pressure for divergent evolution of Saccharomyces species.
Conclusion
Genetics and the mechanisms of evolution are becoming increasingly important in medicine, as we unintentionally breed strains of highly resistant bacteria. Antibiotic stewardship, or the use of the appropriate antibiotics only as necessary, is very important as the medical community seeks to preserve the effectiveness of antibiotics. In order to understand and apply the concepts of antibiotic stewardship, one must understand that creating environmental pressures leads to directional selection in microorganisms which can increase the frequency of the resistant phenotype. In this chapter, we covered genetics and mutations, as well as evolution. We also gave you some tools to analyze the biometric (statistical) side of genetics through the use of Punnett squares, recombinant frequencies, and the Hardy–Weinberg equations.
It seems fitting to complete this book with a discussion of evolution. You’ve spent hundreds of pages (and hours!) preparing for the MCAT, learning the basics of cell biology, embryogenesis and development, anatomy and physiology, genetics, and evolution. Our understanding of these topics relies on generations and generations of scientists who came before us, who passed down their knowledge through books, letters, articles, lectures, and—more recently—television, film, and popular media. But science is a field that is constantly evolving itself. At the beginning of medical school, students are often told that no more than 25 percent of what they learn during the first year will remain true by the time they enter practice. We’re not sure if this statistic actually holds, but it does speak to the importance of staying on top of the latest research—not only as a medical student, but also as a practitioner. Every day, new discoveries about the human body and the practice of medicine are being made—soon, you’ll be one of those making these very discoveries and bringing them to your practice, improving your patients' lives. And at the end of it all, as a provider, an attending physician, or a researcher, you too will pass on your knowledge to future generations of physicians who will also help medical science to evolve and improve. The human body is astoundingly complex. Take a moment to genuinely think about that—the human body is astoundingly complex. There’s so much more to learn. Medical school and your future awaits!
GO ONLINE
You've reviewed the content, now test your knowledge and critical thinking skills by completing a test-like passage set in your online resources!
CONCEPT SUMMARY
Fundamental Concepts of Genetics
- Chromosomes contain genes in a linear sequence.
- Alleles are alternative forms of a gene.
- A dominant allele requires only one copy to be expressed.
- A recessive allele requires two copies to be expressed.
- A genotype is the combination of alleles one has at a given genetic locus.
- Having two of the same allele is termed homozygous.
- Having two different alleles is termed heterozygous.
- Having only one allele is termed hemizygous (such as in male sex chromosomes).
- A phenotype is the observable manifestation of a genotype.
- There are different patterns of dominance.
- Complete dominance occurs when the effect of one allele completely masks the effect of another.
- Codominance has more than one dominant allele.
- Incomplete dominance has no dominant alleles; heterozygotes have intermediate phenotypes.
- Penetrance is the proportion of a population with a given genotype who express the phenotype.
- Expressivity refers to the varying phenotypic manifestations of a given genotype.
- The modern interpretations of Mendel’s laws help explain the inheritance of genes from parent to offspring.
- Mendel’s first law (of segregation) states that an organism has two alleles for each gene, which segregate during meiosis, resulting in gametes carrying only one allele for a trait.
- Mendel’s second law (of independent assortment) states that the inheritance of one allele does not influence the probability of inheriting an allele for a different trait.
- Support for DNA as genetic material came through a number of experiments.
- The Griffith experiment demonstrated the transforming principle, converting nonvirulent live bacteria into virulent bacteria by exposure to heat-killed virulent bacteria.
- The Avery–MacLeod–McCarty experiment demonstrated that DNA is the genetic material because degradation of DNA led to a cessation of bacterial transformation.
- The Hershey–Chase experiment confirmed that DNA is the genetic material because only radiolabeled DNA could be found in bacteriophage-infected bacteria.
Changes in the Gene Pool
- All of the alleles in a given population constitute the gene pool.
- Mutations are changes in DNA sequence.
- Nucleotide mutations include point mutations (the substituting of one nucleotide for another) and frameshift mutations (moving the three-letter transcriptional reading frame).
- A silent mutation has no effect on the protein.
- A missense mutation results in the substitution of one amino acid for another.
- A nonsense mutation results in the substitution of a stop codon for an amino acid.
- Insertions and deletions result in a shift in the reading frame, leading to changes for all downstream amino acids.
- Chromosomal mutations include larger-scale mutations affecting whole segments of DNA.
- Deletion mutations occur when a large segment of DNA is lost.
- Duplication mutations occur when a segment of DNA is copied multiple times.
- Inversion mutations occur when a segment of DNA is reversed.
- Insertion mutations occur when a segment of DNA is moved from one chromosome to another.
- Translocation mutations occur when a segment of DNA is swapped with a segment of DNA from another chromosome.
- Genetic leakage is a flow of genes between species through hybrid offspring.
- Genetic drift occurs when the composition of the gene pool changes as a result of chance.
- The founder effect results from bottlenecks that suddenly isolate a small population, leading to inbreeding and increased prevalence of certain homozygous genotypes.
Analytical Approaches in Genetics
- Pedigree charts track the presence or absence of specific traits among family members, thereby illustrating inheritance patterns.
- Autosomal dominant traits never skip generations and affect males and females equally.
- Autosomal recessive traits also affect males and females equally, but can skip generations; all offspring of two affected parents will be affected.
- X-linked recessive traits can skip generations like autosomal recessive, but disproportionately affect males who have only one X chromosome.
- Mitochondrial inheritance is evident when traits are passed down exclusively through the maternal line: all and only children (both male and female) of affected females are affected.
- Y-linked inheritance is uncommon on the MCAT, but evident when traits only affect males who have Y chromosomes, with all and only sons of affected males also being affected.
- Punnett squares visually represent the crossing of gametes from parents to show relative genotypic and phenotypic frequencies.
- The parent generation is represented by P; filial (offspring) generations are represented by F1, F2, and so on in sequence.
- A monohybrid cross accounts for one gene; a dihybrid cross accounts for two genes.
- In sex-linked crosses, sex chromosomes are usually used to indicate sex as well as genotype.
- The recombination frequency (θ) is the likelihood of two alleles being separated during crossing over in meiosis. Genetic maps can be made using recombination frequency as the scale in centimorgans.
- The Hardy–Weinberg principle states that if a population meets certain criteria (aimed at a lack of evolution), then the allele frequencies will remain constant (Hardy–Weinberg equilibrium).
Evolution
- Natural selection states that chance variations exist between individuals and that advantageous variations—those that increase an individual’s fitness for survival or adaptation to the environment—afford the most opportunities for reproductive success.
- The modern synthesis model (neo-Darwinism) accounts for mutation and recombination as mechanisms of variation and considers differential reproduction to be the mechanism for reproductive success.
- Inclusive fitness considers an organism’s success to be based on the number of offspring, success in supporting offspring, and the ability of the offspring to then support others; survival of offspring or relatives ensures appearance of genes in subsequent generations.
- Punctuated equilibrium considers evolution to be a very slow process with intermittent rapid bursts of evolutionary activity.
- Different types of selection lead to changes in phenotypes.
- Stabilizing selection keeps phenotypes in a narrow range, excluding extremes.
- Directional selection moves the average phenotype toward one extreme.
- Disruptive selection moves the population toward two different phenotypes at the extremes and can lead to speciation.
- Adaptive radiation is the rapid emergence of multiple species from a common ancestor, each of which occupies its own ecological niche.
- A species is the largest group of organisms capable of breeding to form fertile offspring. Species are reproductively isolated from each other by pre- or postzygotic mechanisms.
- Two species can evolve with different relationship patterns.
- Divergent evolution occurs when two species sharing a common ancestor become more different.
- Parallel evolution occurs when two species sharing a common ancestor evolve in similar ways due to analogous selection pressures.
- Convergent evolution occurs when two species not sharing a recent ancestor evolve to become more similar due to analogous selection pressures.
- According to the molecular clock model, the degree of difference in the genome between two species is related to the amount of time since the two species broke off from a common ancestor.
ANSWERS TO CONCEPT CHECKS
**12.1**
- A dominant allele is one that requires only one copy for expression. A recessive allele requires two copies for expression.
- A homozygous genotype is one in which the two alleles are the same. A heterozygous genotype is one in which the two alleles are different. A hemizygous genotype is one in which only one allele is present for a given gene (such as parts of the X chromosome in males).
- Complete dominance occurs when one allele (the dominant one) completely masks the expression of the other (the recessive one). Codominance occurs when a gene has more than one dominant allele, and two different dominant alleles can be expressed simultaneously. Incomplete dominance occurs when a gene has no dominant alleles, and heterozygotes have phenotypes that are intermediate between homozygotes.
- Penetrance describes the proportion of the population that expresses a phenotype, given a particular genotype. Expressivity describes the differences in expression (severity, location, and so on) of a phenotype in individuals.
- Mendel’s first law (of segregation) most aligns with anaphase I of meiosis. Mendel’s second law (of independent assortment) most aligns with prophase I of meiosis.
**12.2**
- Silent point mutations occur when one nucleotide is changed for another, but there is no change in the protein coded for by this DNA sequence (due to redundancy in the genetic code). Missense mutations occur when one nucleotide is changed for another, and one amino acid is substituted for another in the final protein. Nonsense mutations occur when one nucleotide is changed for another, and a stop codon substitutes for an amino acid in the final protein.
- The two types of frameshift mutations are insertion and deletion mutations.
- Duplication mutations occur when a segment of DNA is copied multiple times in the genome. Inversion mutations occur when a segment of DNA is reversed in the genome. Translocation mutations occur when a segment of DNA from one chromosome is swapped with a segment of DNA from another chromosome.
- Genetic leakage requires the formation of a hybrid organism that can then mate with members of one or the other parent species. While hybrids existed historically (especially mules), fertile hybrids were certainly rare before a more modern understanding of genetics (and before a commercial, financial, or academic impetus existed to create these organisms).
- Genetic drift occurs due to chance, so its effects will be more pronounced with a smaller sample size (in smaller populations). The founder effect occurs when a small group is reproductively isolated from the larger population, allowing certain alleles to take on a higher prevalence in the group than in the rest of the population.
**12.3**
-
Cross Phenotypic Ratio
Bb × Bb 3 dominant:1 recessive
Aa × aa 1 dominant:1 recessive
DdEe × ddEE 1 dominant (for D)/dominant (for E): 1 recessive (for D)/dominant (for E)
XqX × XY Female: all unaffected; male: 1 unaffected:1 affected
XrX × XrY Both male and female: 1 unaffected:1 affected
- The genes must be in the order SQRT:
S ↔ 4 % Q ↔ 2 % R ↔ 17 % T
- The criteria for the Hardy–Weinberg principle all imply that the study population is notundergoing evolution; thus, the allele frequencies will remain stable over time.
- The frequency of the dominant allele (p) is 0.3. The frequency of the recessive allele (q) is 0.7. The fraction of the population with a heterozygous genotype (2pq) is 2 × 0.3 × 0.7 = 0.42 (42%). The fraction of the population with a homozygous recessive genotype (q2) is (0.7)2 = 0.49 (49%). The fraction of the population with a dominant phenotype (p2+ 2pq) is 0.09 + 0.42 = 0.51 = 51%.
**12.4**
- Natural selection states that certain traits that arise from chance are more favorable for reproductive success in a given environment, and that those traits will be passed on to future generations. The modern synthesis model takes natural selection and explains that selection is for specific alleles, which are passed on to future generations through formation of gametes; the alleles for these favorable traits arise from mutations. Inclusive fitness explains that the reproductive success of an organism is not only due to the number of offspring it creates, but also the ability to care for young (that can then care for others); it explains changes not only at the individual level, but changes based on the survival of the species (and that individual’s alleles within the species, including in other related individuals). Punctuated equilibrium states that for some species, little evolution occurs for a long period, which is interrupted by rapid bursts of evolutionary change.
-
Pattern of Selection Change to Population Phenotype
Stabilizing Loss of extremes, maintenance of phenotype in a small window
Directional Movement toward one extreme or the other
Disruptive Movement toward both extremes with loss of the norm; speciation may occur
-
Pattern of Evolution Outcome Divergent Two species with a common ancestor become less similar because of different evolutionary pressures
Parallel Two species with a common ancestor remain similar because of similar evolutionary pressures
Convergent Two species with no recent common ancestor become more similar because of similar evolutionary pressures
- A species is defined as the largest group of organisms capable of breeding to form fertile offspring.
SCIENCE MASTERY ASSESSMENT EXPLANATIONS
1. B
This is a gene-mapping problem. Because there is a correlation between the frequency of recombination and the distance between genes on a chromosome, if we are given the frequencies, we can determine gene order. Remember that one map unit equals 1 percent recombination frequency. The easiest way to begin is to determine the two genes that are farthest apart; in this case, N and O recombine with a frequency of 18%, so they are 18 map units apart on the chromosome:
N ↔ 18 % O
N and P recombine with 1% frequency, and P and O recombine with 17% frequency, so P must be between N and O:
N ↔ 1 % P ↔ 17 % O
Finally, M and P recombine with 5% frequency, and M and O recombine with 12% frequency, so M must be between P and O:
N ↔ 1 % P ↔ 5 % M ↔ 12 % O
2. D
In this dihybrid problem, a doubly recessive individual is crossed with an individual of unknown genotype; this is known as a test cross. The straight- and brown-haired organism has the genotype bbcc and can thus only produce gametes carrying bc. Looking at the F1 offspring, there is a 1:1:1:1 phenotypic ratio. The fact that both the dominant and recessive traits are present in the offspring means that the unknown parental genotype must contain both dominant and recessive alleles for each trait. The unknown parental genotype must therefore be BbCc. If you want to double-check the answer, you can work out the Punnett square for the cross BbCc × bbcc:
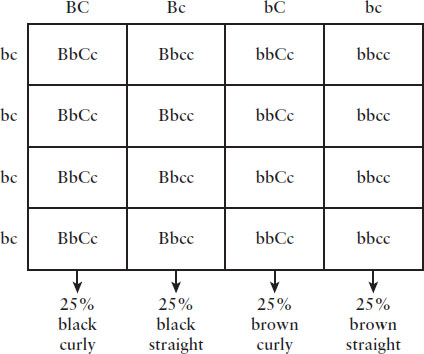
3. C
The female in this example is a carrier of two sex-linked traits; based on the genotype, the affected alleles are found on different X chromosomes. Drawing out a Punnett square, we see that 25% of the offspring will be female hemophiliacs (XhXh) and 25% will be female carriers of both alleles (XcXh). This question is asking what percentage of females will have a phenotype with no hemophilia or color blindness, which would be half of the females (those who are carriers for both traits).

4. A
The control parent in a test cross is always recessive. Therefore, if the test parent is phenotypically dominant, yet can provide a recessive allele (as evidenced by the presence of recessive children), then the parent must have both a dominant and recessive allele. Therefore, this test parent must be heterozygous.
5.B
The definition given here for expressivity is a much better match for defining penetrance (or, really, one minus the penetrance). Expressivity refers to the variable manifestations of a given genotype as different phenotypes; the degree to which various phenotypes are expressed. All of the other definitions given are accurate.
6. B
Some progeny in the second generation are apparently blends of the parental phenotypes. The orange color is the result of the combined effects of the red and yellow alleles. An allele is incompletely dominant if the phenotype of the heterozygotes is an intermediate of the phenotypes of the homozygotes.
7. B
To find the correct answer, we have to read each choice and eliminate the ones that fit with the modern-day theories of inheritance, which state that the genes that make an organism most fit for its environment will be passed to offspring. This can be seen in (C) and (D), both of which demonstrate that an organism with improved fitness will pass those genes to offspring. (A) mentions a gamete being exposed to mutagens; a zygote created from this gamete would contain any mutations that were present in the egg and would be affected by them. Thus, (B) must be the correct answer: indeed, acquired characteristics not encoded in the genome should not be passed to offspring according to the modern synthesis model.
8. B
Darwin’s theory of natural selection argues that chance variations between organisms can help certain organisms survive to reproductive age and produce many offspring, transmitting their variations to the next generation. Thus, natural selection would drive the process of evolution forward, enabling the persistence of characteristics that impart an advantage in the environment, eliminating (A). In Darwin’s theory, fitness is measured in terms of reproductive success, as (D) states. Through natural selection, organisms may be separated into groups depending on environmental pressures, and these groups can eventually separate to the point of becoming distinct species, eliminating (C). (B) is the correct answer because the theory of natural selection applies to a population of organisms, not to a particular individual. As such, favorable genetic variations become more and more common from generation to generation, not during the lifetime of an individual.
9. C
Hardy–Weinberg equilibrium exists under certain ideal conditions that, when satisfied, allow one to calculate the gene frequencies within a population. The Hardy–Weinberg equation can be applied only under these five conditions: (1) the population is very large; (2) there are no mutations that affect the gene pool; (3) mating between individuals in the population is random; (4) there is no migration of individuals into or out of the population; (5) the genes in the population are all equally successful at being reproduced. Thus, from the given choices, only (C) is false: monogamy is not a necessary condition for Hardy–Weinberg equilibrium.
10. B
The situation described in the question stem is an example of directional selection. In directional selection, the phenotypic norm of a particular species shifts toward an extreme to adapt to a selective pressure, such as an increasingly colder environment. Only those individuals with a thicker layer of fur were able to survive during the Ice Age, thus shifting the phenotypic norm.
11. A
A species is defined as the largest group of organisms that can interbreed to produce viable, fertile offspring. Therefore, two populations are considered separate species when they can no longer do so.
12. D
Let’s use the information provided by the question stem to set up our equations. We are told that the frequency of R equals 30%, and as such, p = 0.30. The frequency of the recessive gene is r = 100% – 30% = 70%; thus, q = 0.70. The frequency of the genotypes, according to the Hardy–Weinberg equilibrium, are given by p2 = RR, 2pq = Rr, and q2 = rr. Therefore, the frequency of the genotypes are (0.3)2 = 0.09 = 9% RR, 2 × 0.3 × 0.7 = 0.42 = 42% Rr, and (0.7)2 = 0.49 = 49% rr.
13. B
Using the information given in the question stem, we can determine that the percentage of the population with blue eyes (genotype bb) = 36% = 0.36 = q2; therefore, q = 0.6. Because this is a Hardy–Weinberg population, we can assume that p + q = 1, so p = 1 – 0.6 = 0.4. The frequency of heterozygous brown eyes is therefore 2pq = 2 × 0.4 × 0.6 = 0.48 = 48%.
14. D
Frameshift mutations include insertions and deletions of nucleotides that change the triplet reading frame. In such a mutation, the amino acid sequence preceding the mutation is normal, yielding a normal N terminal end. However, the triplet reading frame after the mutation is changed, yielding a drastically different C terminal sequence that often includes a premature stop codon. These factors together support (D) as the correct answer. Note that nonsense mutations also introduce premature stop codons. However, (C) can be eliminated since a nonsense mutation will create a stop codon directly and will not change the identity of the amino acids before the stop codon.
15. C
This scenario—a deletion of some DNA and a duplication of other DNA—would be consistent with a translocation between chromosomes 4 and 21 during development of an egg or sperm. If part of chromosome 21 was swapped with part of chromosome 4, then a gamete resulting from meiosis in this cell would result in a daughter cell with two copies of some of the DNA from 21 and no copies of some of the DNA from 4. Therefore, after fertilization, there would be partial trisomy 21 and partial monosomy 4. While a deletion or insertion could explain one of the findings, it cannot explain both, eliminating (A) and (B). An inversion should not lead to partial trisomy or partial monosomy because the DNA is simply reversed, eliminating (D).
GO ONLINE
Consult your online resources for additional practice.
EQUATIONS TO REMEMBER
(12.1) Hardy-Weinberg equations:
p+q=1p2+2pq+q2=1
SHARED CONCEPTS
Behavioral Sciences Chapter 10
Social Thinking
Biochemistry Chapter 6
DNA and Biotechnology
Biochemistry Chapter 7
RNA and the Genetic Code
Biology Chapter 1
The Cell
Physics and Math Chapter 11
Reasoning About the Design and Execution of Research
Physics and Math Chapter 12
Data-Based and Statistical Reasoning