The Auditory and Vestibular Systems
BOX 11.1 OF SPECIAL INTEREST: Ultrasound and Infrasound
BOX 11.2 OF SPECIAL INTEREST: The Deaf Shall Hear: Cochlear Implants
BOX 11.3 OF SPECIAL INTEREST: Hearing with Noisy Ears
Response Properties of Neurons in the Auditory Pathway
BOX 11.4 PATH OF DISCOVERY: Capturing the Beat, by Donata Oertel
The Sensitivity of Binaural Neurons to Sound Location
BOX 11.5 OF SPECIAL INTEREST: How Does Auditory Cortex Work? Ask a Specialist
The Effects of Auditory Cortical Lesions and Ablation
BOX 11.6 OF SPECIAL INTEREST: Auditory Disorders and Their Treatments
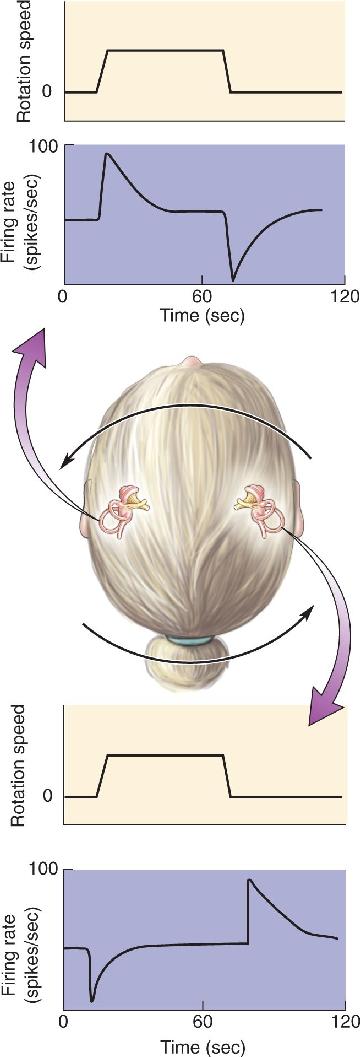
Central Vestibular Pathways and Vestibular Reflexes
In this chapter, we’ll explore two sensory systems that have very different functions but surprising similarities of structure and mechanism: the sense of hearing, also known as audition, and the sense of balance, regulated by the vestibular system. Hearing is a vivid part of our conscious lives, while balance is something we experience all day but rarely think about.
When we cannot see something or someone, we can often detect its presence, identify its origin, and even receive a message from it just by hearing its sounds. Anyone who has ever hiked through the forest in an area where there are bears or snakes knows that the sound of rustling leaves can be a powerful attention–grabber. Aside from the ability to detect and locate sound, we can perceive and interpret its nuances. We can immediately distinguish the bark of a dog, the voice of a particular friend, the crash of an ocean wave. Because humans are able to produce a wide variety of sounds as well as hear them, spoken language and its reception via the auditory system have become an extremely important means of communication. Audition in humans has even evolved beyond the strictly utilitarian functions of communication and survival; musicians, for example, explore the sensations and emotions evoked by sound.
In contrast to hearing, the sense of balance is strictly a personal, internalized process. The vestibular system informs our nervous system where our head and body are and how they are moving. This information is used, without conscious effort, to control muscular contractions that keep or put our body where we want it to be, to reorient ourselves when something moves us, and to move our eyes so that our visual world stays fixed on our retinas even when our head is bouncing around.
Here, we will explore the mechanisms within the ear and brain that translate the sounds in our environment into meaningful neural signals and the movements of our head into a sense of where we are. We will find that these transformations are carried out in stages rather than all at once. Within the inner ear, neural responses are generated by auditory receptors from the mechanical energy in sound and by vestibular receptors from the tilts and rotations of the head. At subsequent stages in the brain stem and thalamus, signals from the receptors are integrated before they ultimately reach auditory and vestibular cortex. By looking at the response properties of neurons at various points in the system, we begin to understand the relationship between neural activity and our perception of sound and balance.
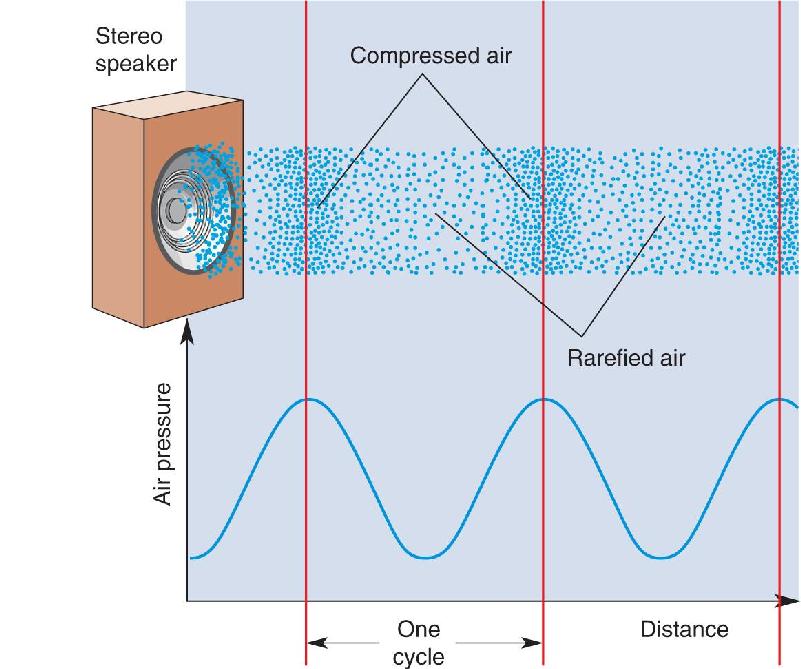
Sounds are audible variations in air pressure. Almost anything that can move air molecules can generate a sound, including the vocal cords of the human larynx, the vibration of a string on a guitar, and the explosion of a firecracker. When an object moves toward a patch of air, it compresses the air, increasing the density of the molecules. Conversely, the air is rarefied (made less dense) when an object moves away. This is particularly easy to visualize in the case of a stereo speaker, in which a paper cone attached to a magnet vibrates in and out, alternately rarefying and compressing the air (Figure 11.1). These changes in air pressure are transferred away from the speaker at the speed of sound, which is about 343 m/sec (767 mph) for air at room temperature.

FIGURE 11.1 The production of sound by variations in air pressure. When the paper cone of a stereo speaker pushes out, it compresses the air; when the cone pulls in, it rarefies the air. If the push and pull are rhythmic, there will also be a rhythmic variation in the air pressure, as shown in the graph. The distance between successive compressed (high-pressure) patches of air is one cycle of the sound (indicated by the vertical lines). The sound wave propagates away from the speaker at the speed of sound. The blue line is a graph of air pressure versus distance. Description
Many sources of sound, such as vibrating strings or a stereo speaker reproducing the sound of a stringed instrument, produce variations in air pressure that are rhythmic. The frequency of the sound is the number of compressed or rarefied patches of air that pass by our ears each second. One cycle of the sound is the distance between successive compressed patches; the sound frequency, expressed in units called hertz (Hz), is the number of cycles per second. Because sound waves all propagate at the same speed, high-frequency sound waves have more compressed and rarefied regions packed into the same space than low-frequency waves (Figure 11.2a).

FIGURE 11.2 The frequency and intensity of sound waves. Each graph plots air pressure versus distance for a sound of constant frequency and intensity. Note that the X axis also represents time because the velocity of sound is constant. (a) Frequency is the number of sound waves per unit of time or distance. We perceive high-frequency waves as having a higher pitch. (b) Intensity is the air pressure difference between peaks and troughs of the sound waves. We perceive high-intensity waves as louder. Description
Our auditory system can respond to pressure waves over the remarkable range of 20 Hz to 20,000 Hz (although this audible range decreases significantly, especially at the high-frequency end, as we age and expose ourselves to loud noise). Whether a sound is perceived to have a high or low tone, or pitch, is determined by the frequency. In order to understand frequency, realize that a room-shaking low note on an organ is about 20 Hz and an ear-piercing high note on a piccolo is about 10,000 Hz. Although humans can hear a great range of frequencies, there are high and low sound wave frequencies our ears cannot hear, just as there are electromagnetic waves of light our eyes cannot see (Box 11.1).
Most people are familiar with ultrasound (sound above the 20 kHz upper limit of our hearing) because it has everyday applications, from ultrasonic cleaners to medical imaging. Many animals can hear these high frequencies. For instance, ultrasonic dog whistles work because dogs can hear up to about 45 kHz. Some bats vocalize at frequencies up to 100 kHz, then listen to the echoes of their calls in order to locate objects (see Box 11.5). Some fish in the shad and herring family can detect sounds as high as 180 kHz, thereby enabling them to hear the echolocating ultrasound generated by dolphins that prey on them. Needless to say, the dolphins can hear their own ultrasonic calls. Similarly, nocturnal moths listen for the ultrasound of hungry bats, so they can evade these predators.
Infrasound is sound at frequencies lower than humans can hear, below about 20 Hz. Some animals can hear infrasonic frequencies; one is the elephant, which can detect 15 Hz tones inaudible to humans. Whales produce low-frequency sounds, which are thought to be a means of communication over distances of many kilometers. The earth also produces low-frequency vibrations, and some animals may sense an impending earthquake by hearing such sounds.
Even though we usually cannot hear very low frequencies with our ears, they are present in the environment and we can sometimes feel them as vibrations with our somatosensory system (see Chapter 12). Infrasound is produced by such devices as air conditioners, boilers, aircraft, and automobiles, and it can have unpleasant subconscious effects. Although even intense infrasound from these machines does not cause hearing loss, it can cause dizziness, nausea, and headache. Many cars produce low-frequency sound when they’re moving at highway speeds, making sensitive people carsick. At very high levels, low-frequency sound may also produce resonances in body cavities such as the chest and stomach, which can damage internal organs.
In addition to mechanical equipment, our own bodies generate inaudible low-frequency sound. When muscle changes length, individual fibers vibrate, producing low-intensity sound at about 25 Hz. While we cannot normally hear these sounds, you can demonstrate them to yourself by carefully putting your thumbs in your ears and making a fist with each hand. As you tighten your fist, you can hear a low rumbling sound produced by the contraction of your forearm muscles. Other muscles, including your heart, produce inaudible sound at frequencies near 20 Hz.
Another important property of a sound wave is its intensity, or amplitude, which is the difference in pressure between compressed and rarefied patches of air (Figure 11.2b). Sound intensity determines the loudness we perceive, loud sounds having higher intensity. The range of intensities to which the human ear is sensitive is astonishing: The intensity of the loudest sound that doesn’t damage our ears is about a trillion times greater than the intensity of the faintest sound that can be heard. If our auditory system were much more sensitive, we would hear a constant roar from the random movement of air molecules.
Real-world sounds rarely consist of simple rhythmic sound waves at one frequency and intensity. It is the simultaneous combination of different frequency waves at different intensities that gives different musical instruments and human voices their unique tonal qualities.
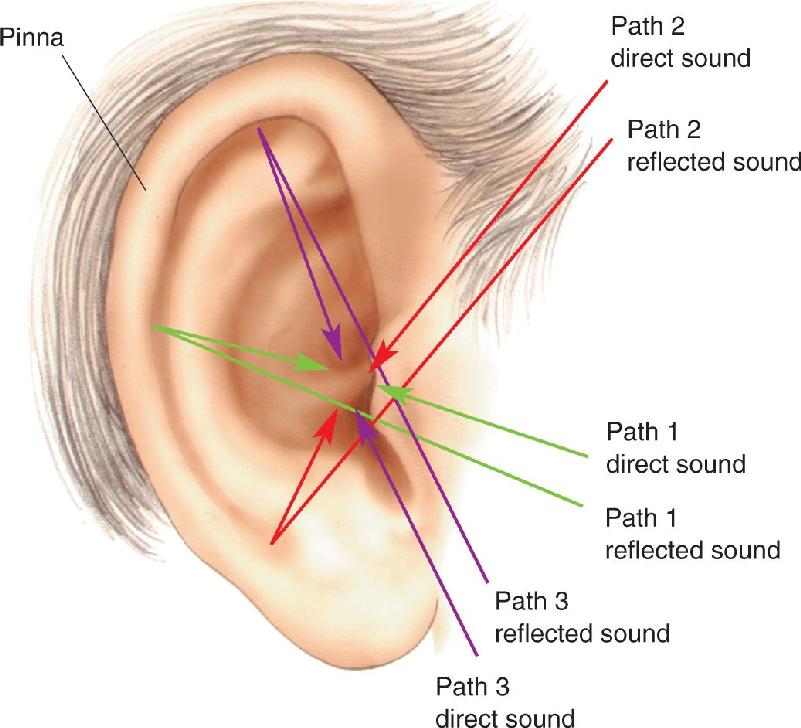
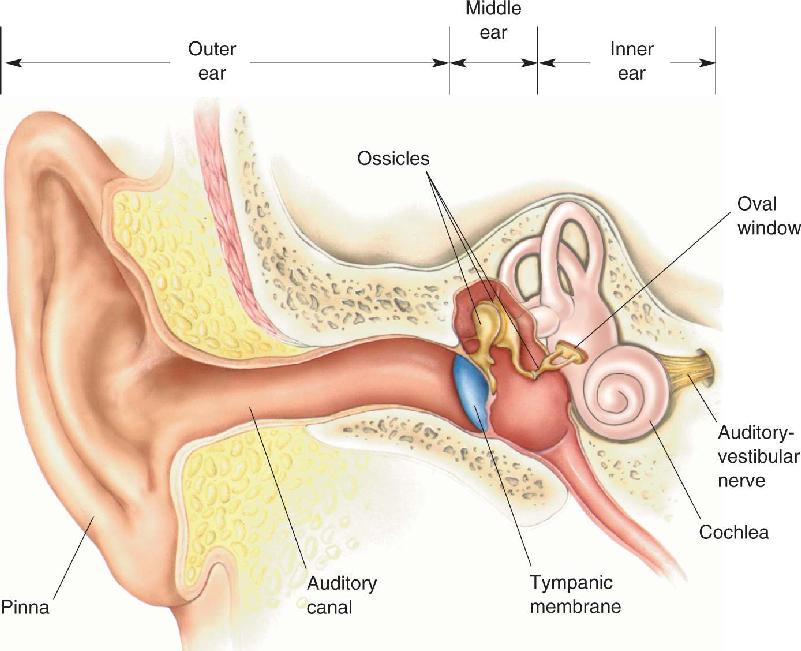
Before exploring how variations in air pressure are translated into neural activity, let’s quickly survey the structure of the auditory system. The components of the ear are shown in Figure 11.3. The visible portion of the ear consists primarily of cartilage covered by skin, forming a sort of funnel called the pinna (from the Latin for “wing”), which helps collect sounds from a wide area. The shape of the pinna makes us more sensitive to sounds coming from ahead than from behind. The convolutions in the pinna play a role in localizing sounds, as we’ll discuss later on. In humans, the pinna is more or less fixed in position, but animals such as cats and horses have considerable muscular control over the position of their pinna and can orient it toward a source of sound.

The entrance to the internal ear is called the auditory canal, which extends about 2.5 cm (1 inch) inside the skull before it ends at the tympanic membrane, also known as the eardrum. Connected to the medial surface of the tympanic membrane is a series of bones called ossicles (from the Latin for “little bones”; the ossicles are indeed the smallest bones in the body). Located in a small air-filled chamber, the ossicles transfer movements of the tympanic membrane into movements of a second membrane covering a hole in the bone of the skull called the oval window. Behind the oval window is the fluid-filled cochlea, which contains the apparatus for transforming the physical motion of the oval window membrane into a neuronal response. Thus, the first stages of the basic auditory pathway look like this:
Motion at the oval window moves fluid in the cochlea. →
Movement of fluid in the cochlea causes a response in sensory neurons.
All the structures from the pinna inward are considered components of the ear, conventionally referenced in three main divisions. The structures from the pinna to the tympanic membrane make up the outer ear, the tympanic membrane and the ossicles constitute the middle ear, and the apparatus medial to the oval window is the inner ear.
Once a neural response to sound is generated in the inner ear, the signal is transferred to and processed by a series of nuclei in the brain stem. Output from these nuclei is sent to a relay in the thalamus, the medial geniculate nucleus (MGN). Finally, the MGN projects to primary auditory cortex, or A1, located in the temporal lobe. In one sense, the auditory pathway is more complex than the visual pathway because there are more intermediate stages between the sensory receptors and cortex. However, the systems have analogous components. Each starts with sensory receptors, which connect to early integration stages (located in the retina for vision and the brain stem for audition), then to a thalamic relay, and then to sensory cortex (Figure 11.4).

FIGURE 11.4 Auditory and visual pathways compared. Following the sensory receptors, both systems have early integration stages, a thalamic relay, and a projection to sensory cortex. Description
The outer ear funnels sound to the middle ear, an air-filled cavity containing the first elements that move in response to sound. In the middle ear, variations in air pressure are converted into movements of the ossicles. In this section, we’ll explore how the middle ear performs an essential transformation of sound energy.
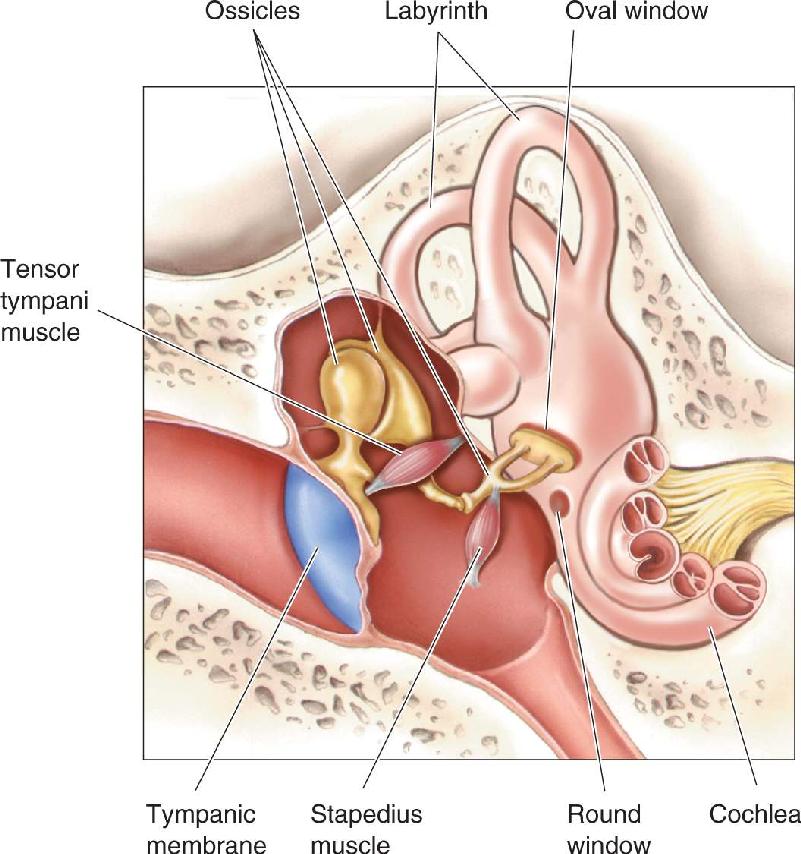
The structures within the middle ear are the tympanic membrane, the ossicles, and two tiny muscles that attach to the ossicles. The tympanic membrane is somewhat conical in shape, with the point of the cone extending into the cavity of the middle ear. There are three ossicles, each named (from the Latin) after an object it slightly resembles (Figure 11.5). The ossicle attached to the tympanic membrane is the malleus (“hammer”), which forms a rigid connection with the incus (“anvil”). The incus forms a flexible connection with the stapes (“stirrup”). The flat bottom portion of the stapes, the footplate, moves in and out like a piston at the oval window. The movements of the footplate transmit sound vibrations to the fluids of the cochlea in the inner ear.

FIGURE 11.5 The middle ear. As the arrows indicate, when air pressure pushes the tympanic membrane, the bottom of the malleus is pushed inward and the lever action of the ossicles makes the footplate of the stapes push inward at the oval window. The pressure pushing at the oval window is greater than that at the tympanic membrane, in part because the surface area of the footplate of the stapes is smaller than the surface area of the tympanic membrane.
The air in the middle ear is continuous with the air in the nasal cavities via the Eustachian tube, although a valve usually keeps this tube closed. When you’re in an ascending airplane or a car heading up a mountain, the pressure of the surrounding air decreases. However, as long as the valve on the Eustachian tube is closed, the air in the middle ear stays at the pressure of the air before you started to climb. Because the pressure inside the middle ear is higher than the air pressure outside, the tympanic membrane bulges out, and you experience unpleasant pressure or pain in the ear. Yawning or swallowing, both of which open the Eustachian tube, can usually equalize the air pressure in the middle ear with the ambient air pressure and so relieve the pain. The opposite can happen as you descend. The air pressure outside is then higher than the pressure inside the middle ear, and opening the Eustachian tube again can relieve the discomfort you may feel.
Sound waves move the tympanic membrane, and the ossicles move another membrane at the oval window. Why isn’t the ear arranged so that sound waves simply directly move the membrane at the oval window? The problem is that the cochlea is filled with fluid, not air. If sound waves impinged directly on the oval window, the membrane would barely move and all but about 0.1% of the sound energy would be reflected away because of the pressure the cochlear fluid exerts at the back of the oval window. If you’ve ever noticed how quiet it is under water, you know how well water reflects sound coming from above. The fluid in the inner ear resists being moved much more than air does (i.e., fluid has greater inertia), so more pressure is needed to vibrate the fluid than the air can provide. The ossicles provide this necessary amplification in pressure.
To understand the process, consider the definition of pressure. The pressure on a membrane is defined as the force pushing it divided by its surface area. The pressure at the oval window will become greater than the pressure at the tympanic membrane if (1) the force on the oval window membrane is greater than that on the tympanic membrane, or (2) the surface area of the oval window is smaller than the area of the tympanic membrane. The middle ear uses both mechanisms. It increases pressure at the oval window by altering both the force and the surface area. The force at the oval window is greater because the ossicles act like levers. Sound causes large movements of the tympanic membrane, which are transformed into smaller but stronger vibrations of the oval window. And the surface area of the oval window is much smaller than that of the tympanic membrane. These factors combine to make the pressure at the oval window about 20 times greater than at the tympanic membrane, and this increase is sufficient to move the fluid in the inner ear.
Two muscles attached to the ossicles have a significant effect on sound transmission to the inner ear. The tensor tympani muscle is anchored to bone in the cavity of the middle ear at one end and attaches to the malleus at the other end (Figure 11.6). The stapedius muscle also extends from a fixed anchor of bone and attaches to the stapes. When these muscles contract, the chain of ossicles becomes much more rigid, and sound conduction to the inner ear is greatly diminished. The onset of a loud sound triggers a neural response that causes these muscles to contract, a response called the attenuation reflex. Sound attenuation is much greater at low frequencies than at high frequencies.

FIGURE 11.6 The middle and inner ear. The stapedius muscle and the tensor tympani muscle are both attached to the wall of the middle ear at one end, and to the ossicles at the other ends.
A number of functions have been proposed for this reflex. One function may be to adapt the ear to continuous sound at high intensities. Loud sounds that would otherwise saturate the response of the receptors in the inner ear could be reduced to a level below saturation by the attenuation reflex, thus increasing the dynamic range we can hear. The attenuation reflex also protects the inner ear from loud sounds that would otherwise damage it. Unfortunately, the reflex has a delay of 50–100 msec from the time that sound reaches the ear, so it doesn’t offer much protection from very sudden loud sounds; damage might already be done by the time the muscles contract. This is why, despite the best efforts of your attenuation reflex, a loud explosion can still damage your cochlea. Because the attenuation reflex suppresses low frequencies more than high frequencies, it tends to make high-frequency sounds easier to discern in an environment with a lot of low-frequency noise. This capability enables us to understand speech more easily in a noisy environment than we could without the reflex. It is thought that the attenuation reflex is also activated when we speak, so we don’t hear our own voices as loudly as we otherwise would.
Although considered part of the ear, not all of the inner ear is concerned with hearing. The inner ear consists of the cochlea, which is part of the auditory system, and the labyrinth, which is not. The labyrinth is an important part of the vestibular system, which helps maintain the body’s equilibrium. The vestibular system is discussed later in the chapter. Here, we are concerned only with the cochlea and the role it plays in transforming sound into a neural signal.
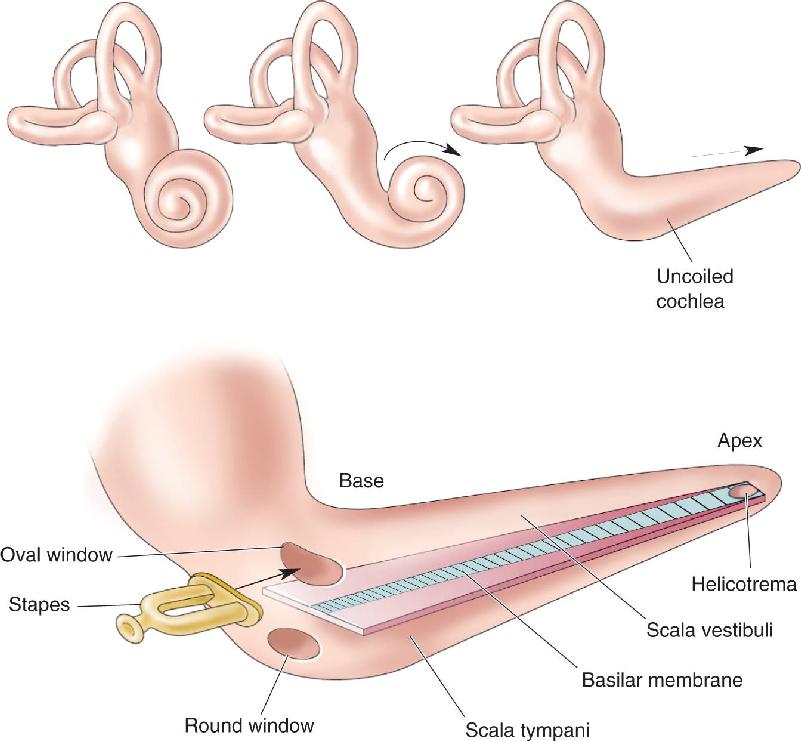
The cochlea (from the Latin for “snail”) has a spiral shape resembling a snail’s shell. Figure 11.6 shows the cochlea cut in half. The structure of the cochlea is similar to a drinking straw wrapped two and a half to three times around the sharpened tip of a pencil. In the cochlea, the hollow tube (represented by the straw) has walls made of bone. The central pillar of the cochlea (represented by the pencil) is a conical bony structure. The actual dimensions are much smaller than the straw-and-pencil model, the cochlea’s hollow tube being about 32 mm long and 2 mm in diameter. Rolled up, the human cochlea is about the size of a pea. At the base of the cochlea are two membrane-covered holes: the oval window, which is below the footplate of the stapes, as we have seen, and the round window.
If the cochlea is cut in cross section, we can see that the tube is divided into three fluid-filled chambers: the scala vestibuli, the scala media, and the scala tympani (Figure 11.7). The three scalae wrap around inside the cochlea like a spiral staircase (scala is from the Latin for “stairway”). Reissner’s membrane separates the scala vestibuli from the scala media, and the basilar membrane separates the scala tympani from the scala media. Sitting upon the basilar membrane is the organ of Corti, which contains auditory receptor neurons; hanging over this organ is the tectorial membrane. At the apex of the cochlea, the scala media is closed off, and the scala tympani becomes continuous with the scala vestibuli at a hole in the membranes called the helicotrema (Figure 11.8). At the base of the cochlea, the scala vestibuli meets the oval window and the scala tympani meets the round window.

FIGURE 11.7 The three scalae of the cochlea. Viewed in cross section, the cochlea contains three small parallel chambers. These chambers, the scalae, are separated by Reissner’s membrane and the basilar membrane. The organ of Corti contains the auditory receptors; it sits upon the basilar membrane and is covered by the tectorial membrane.

FIGURE 11.8 The basilar membrane in an uncoiled cochlea. Although the cochlea narrows from base to apex, the basilar membrane widens toward the apex. Notice that the basilar membrane is the narrow blue band only. The helicotrema is a hole at the apex of the basilar membrane, which connects the scala vestibuli and scala tympani.
The fluid in the scala vestibuli and scala tympani, called perilymph, has an ionic content similar to that of cerebrospinal fluid: relatively low K+ (7 mM) and high Na+ (140 mM) concentrations. The scala media is filled with endolymph, which is an unusual extracellular fluid in that it has ionic concentrations similar to intracellular fluid, high K+ (150 mM) and low Na+ (1 mM). This difference in ion content is generated by active transport processes taking place at the stria vascularis, the endothelium lining one wall of the scala media and contacting the endolymph (see Figure 11.7). The stria vascularis absorbs sodium from, and secretes potassium into, the endolymph. Because of the ionic concentration differences and the permeability of Reissner’s membrane, the endolymph has an electrical potential that is about 80 mV more positive than that of the perilymph; this is called the endocochlear potential. We shall see that the endocochlear potential is vital because it enhances auditory transduction.
The structure of the cochlea is complex, but its basic operation is fairly simple. Look at Figure 11.8 and imagine what happens when the ossicles move the membrane that covers the oval window. They work like a tiny piston. Inward motion at the oval window pushes perilymph into the scala vestibuli. If the membranes inside the cochlea were completely rigid, then the increase in fluid pressure at the oval window would reach up the scala vestibuli, through the helicotrema, and back down the scala tympani to the round window. Because the fluid pressure has nowhere else to escape, the membrane at the round window would bulge out in response to the inward movement of the membrane at the oval window. Any motion at the oval window must be accompanied by a complementary motion at the round window. Such movement must occur because the cochlea is filled with incompressible fluid held in a solid bony container. The consequence of pushing in at the oval window is a bit like pushing in one end of a tubular water balloon: The other end has to bulge out.
This simple description of the events in the cochlea is complicated by one additional fact: Some structures inside the cochlea are not rigid. Most importantly, the basilar membrane is flexible and bends in response to sound.
The Response of the Basilar Membrane to Sound. The basilar membrane has two structural properties that determine the way it responds to sound. First, the membrane is wider at the apex than at the base by a factor of about 5. Second, the stiffness of the membrane decreases from base to apex, the base being about 100 times stiffer. Think of it as a flipper of the sort used for swimming, with a narrow, stiff base and a wide, floppy apex. When sound pushes the footplate of the stapes at the oval window, perilymph is displaced within the scala vestibuli, and endolymph is displaced within the scala media because Reissner’s membrane is very flexible. Sound can also pull the footplate, reversing the pressure gradient. Sound causes a continual push–pull motion of the footplate; again, think of a tiny piston.
We owe much of our understanding of the response of the basilar membrane to the research of Hungarian–American biophysicist Georg von Békésy. Von Békésy determined that the movement of the endolymph makes the basilar membrane bend near its base, starting a wave that propagates toward the apex. The wave that travels up the basilar membrane is similar to the wave that runs along a rope if you hold one end in your hand and give it a snap (Figure 11.9). The distance the wave travels up the basilar membrane depends on the frequency of the sound. If the frequency is high, the stiffer base of the membrane will vibrate a good deal, dissipating most of the energy, and the wave will not propagate very far (Figure 11.10a). However, low-frequency sounds generate waves that travel all the way up to the floppy apex of the membrane before most of the energy is dissipated (Figure 10.10b). The response of the basilar membrane establishes a place code in which different locations of membrane are maximally deformed at different sound frequencies (Figure 11.10c). Systematic organization of sound frequency within an auditory structure is called tonotopy, analogous to retinotopy in the visual system. Tonotopic maps exist on the basilar membrane and within each of the auditory relay nuclei, the MGN, and auditory cortex.

FIGURE 11.9 A traveling wave in the basilar membrane. As the stapes moves in and out, it causes perilymph to flow, as shown by the arrows. This generates a traveling wave in the basilar membrane. (The size of the wave is magnified about 1 million times in this illustration.) At this frequency, 3000 Hz, the fluid and membrane movement end abruptly about halfway between the base and apex. Note that the scala media is not illustrated here. (Source: Adapted from Nobili, Mammano, and Ashmore, 1998, Fig. 1.)

FIGURE 11.10 The response of the basilar membrane to sound. The cochlea is again shown uncoiled. (a) High-frequency sound produces a traveling wave, which dissipates near the narrow and stiff base of the basilar membrane. (b) Low-frequency sound produces a wave that propagates all the way to the apex of the basilar membrane before dissipating. (The flexing of the basilar membrane is greatly exaggerated for the purpose of illustration.) (c) There is a place code on the basilar membrane for the frequency that produces the maximum amplitude deflection. Description
As we shall see, the differences in the traveling waves produced by different sound frequencies are responsible for the neural coding of pitch.
The Organ of Corti and Associated Structures. Everything we have discussed to this point involves the mechanical transformations of sound energy that occur in the middle and inner ear. Now, we come to the point in the system where neurons are first involved. The auditory receptor cells, which convert mechanical energy into a change in membrane polarization, are located in the organ of Corti (named for the Italian anatomist who first identified it). The organ of Corti consists of hair cells, the rods of Corti, and various supporting cells.
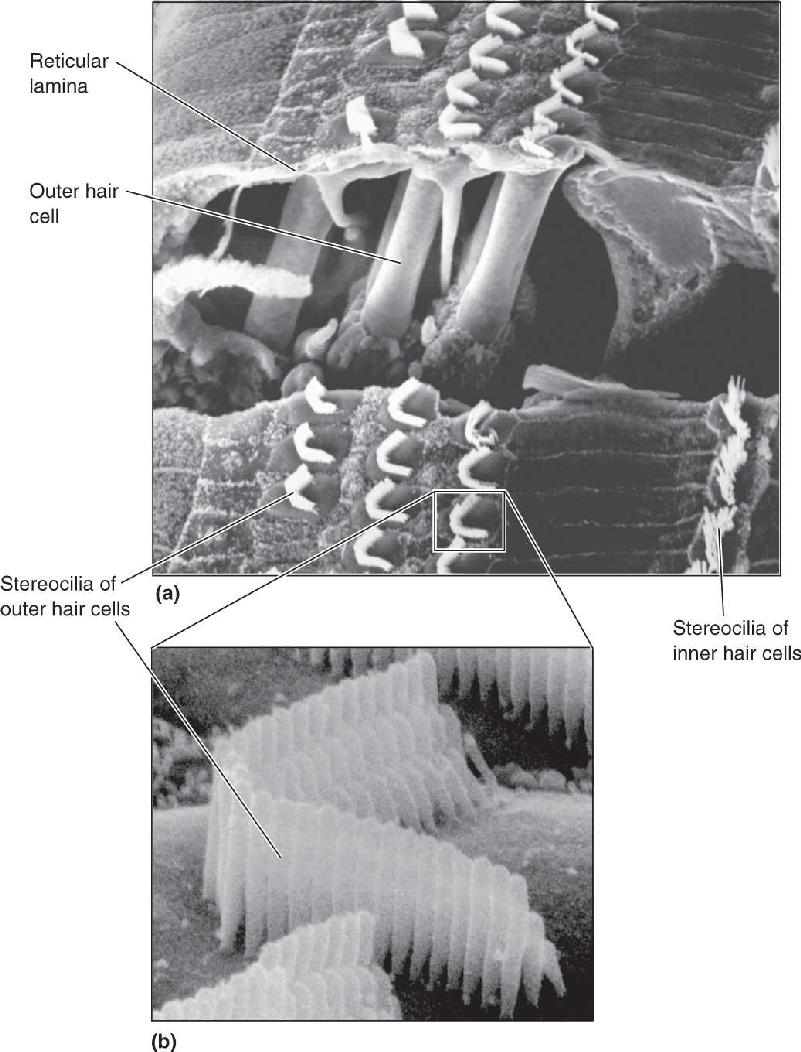
The auditory receptors are called hair cells because each one has 10–300 hairy-looking stereocilia extending from its top. Hair cells are not neurons. They lack axons, and in mammals, they do not generate action potentials. Hair cells are actually specialized epithelial cells. The hair cells and stereocilia are shown in Figure 11.11 as they appear when viewed with a scanning electron microscope. The critical event in the transduction of sound into a neural signal is the bending of these cilia. For this reason, we will examine the organ of Corti in more detail to see how flexing of the basilar membrane leads to bending of the stereocilia.

FIGURE 11.11 Hair cells viewed through the scanning electron microscope. (a) Hair cells and their stereocilia. (b) A higher resolution view of the stereocilia on an outer hair cell. The stereocilia are approximately 5 μm in length. (Source: Courtesy of I. Hunter-Duvar and R. Harrison, The Hospital for Sick Children, Toronto, Ontario, Canada.)
The hair cells are sandwiched between the basilar membrane and a thin sheet of tissue called the reticular lamina (Figure 11.12). The rods of Corti span these two membranes and provide structural support. Hair cells between the modiolus and the rods of Corti are called inner hair cells (about 4500 form a single row), and cells farther out than the rods of Corti are called outer hair cells (in humans, there are about 12,000–20,000 arranged in three rows). The stereocilia at the tops of the hair cells extend above the reticular lamina into the endolymph, and their tips end either in the gelatinous substance of the tectorial membrane (the outer hair cells) or just below the tectorial membrane (the inner hair cells). To keep the membranes within the organ of Corti straight in your mind, remember that the basilar is at the base of the organ of Corti, the tectorial forms a roof over the structure, and the reticular is in the middle, holding onto the hair cells.

FIGURE 11.12 The organ of Corti. The basilar membrane supports tissue that includes the inner and outer hair cells and the stiff rods of Corti. The tectorial membrane extends from the bony modiolus to cover the stereocilia that protrude from the tops of the hair cells.
Hair cells form synapses on neurons whose cell bodies are located in the spiral ganglion within the modiolus. Spiral ganglion cells are bipolar, with neurites extending to the bases and sides of the hair cells, where they receive synaptic input. Axons from the spiral ganglion enter the auditory nerve, a branch of the auditory–vestibular nerve (cranial nerve VIII), which projects to the cochlear nuclei in the medulla. It is possible to treat certain forms of deafness by using electronic devices to bypass the middle ear and the hair cells, and activate the auditory nerve axons directly (Box 11.2).
Hair cell damage or death is the most common cause of human deafness (see Box 11.6). In most cases, the auditory nerve remains intact, making it possible to restore some hearing with a cochlear implant—essentially an artificial, electronic cochlea. The roots of this technology can be traced back two centuries to the pioneering work of the Italian physicist Alessandro Volta (after whom the electrical unit called the volt was named). In 1800, shortly after he invented the battery, Volta gamely (some might say foolishly) inserted the two contacts from a 50 volt battery into his ears. Here’s how he described the result:
At the moment when the circuit was completed, I received a shock in the head, and some moments after I began to hear a sound, or rather noise in the ears, which I cannot well define: it was a kind of crackling with shocks, as if some paste or tenacious matter had been boiling. . . The disagreeable sensation, which I believe might be dangerous because of the shock in the brain, prevented me from repeating this experiment. . . . 1
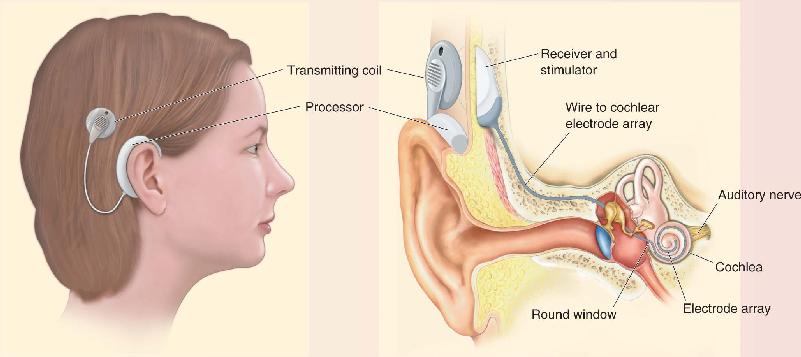
The art of electrically stimulating the ears has improved considerably since Volta’s singular experiment. In fact, in recent years, cochlear implant systems have revolutionized the treatment of inner ear damage for many people. Most of the system is actually external to the body (Figure A). It starts with a headpiece containing a microphone, which receives sound and converts it to an electrical signal. This signal is sent to a battery-powered digital processor. A small radio transmitter placed over the scalp transmits the digital code to a receiver that has been surgically implanted beneath the skin, in the mastoid bone behind the ear. The transmitter and receiver are held close to one another with magnets, and no wires penetrate the skin.

Figure A The headpiece of a cochlear implant, positioned behind the ear.
Figure B A cochlear implant in place inside the ear. Description
The receiver translates the code into a series of electrical impulses that it sends to the cochlear implant itself—a very thin, flexible bundle of wires that has been threaded through a tiny hole and into the cochlea (Figure B). The cochlear electrode array has about 22 separate stimulation sites that allow it to activate the auditory nerve at various places along the cochlea, from the base toward the apex. The most clever feature of the cochlear implant is that it takes advantage of the tonotopic arrangement of auditory nerve fibers; stimulation near the base of the cochlea evokes a perception of high-frequency sounds, and stimulation toward the apex evokes low-frequency sounds.
By 2012, there were more than 340,000 cochlear implant users in the world, and the popularity of the devices is increasing. About 38,000 children have implants in the U.S. alone. Unfortunately, they are very expensive.
Cochlear implants can provide an extraordinary hearing capacity for many previously deaf people. With several months of training, people can achieve a remarkably good understanding of conversational speech, even when listening on a telephone. Most can understand more than 90% of spoken words when listening in a quiet room. The success of the implants varies widely, for reasons that are often unclear. Researchers are working hard to improve the technology of cochlear implants, to reduce their size, and to determine the best ways to train patients in their use.
The best candidates for cochlear implants are young children (optimally as young as one year), and older children or adults whose deafness was acquired after they learned some speech. For adults whose deafness preceded any experience with speech, on the other hand, cochlear implants seem to provide only a crude perception of sounds. It seems that the auditory system, like other sensory systems in the brain, needs to experience normal inputs at a young age in order to develop properly. If it is deprived of exposure to sounds early in life, the auditory system can never develop completely normal function even if hearing is restored later. The concept of critical periods in brain development is described in Chapter 23.
1 Quoted in Zeng F-G. 2004. Trends in cochlear implants. Trends in Amplification 8:1–34.
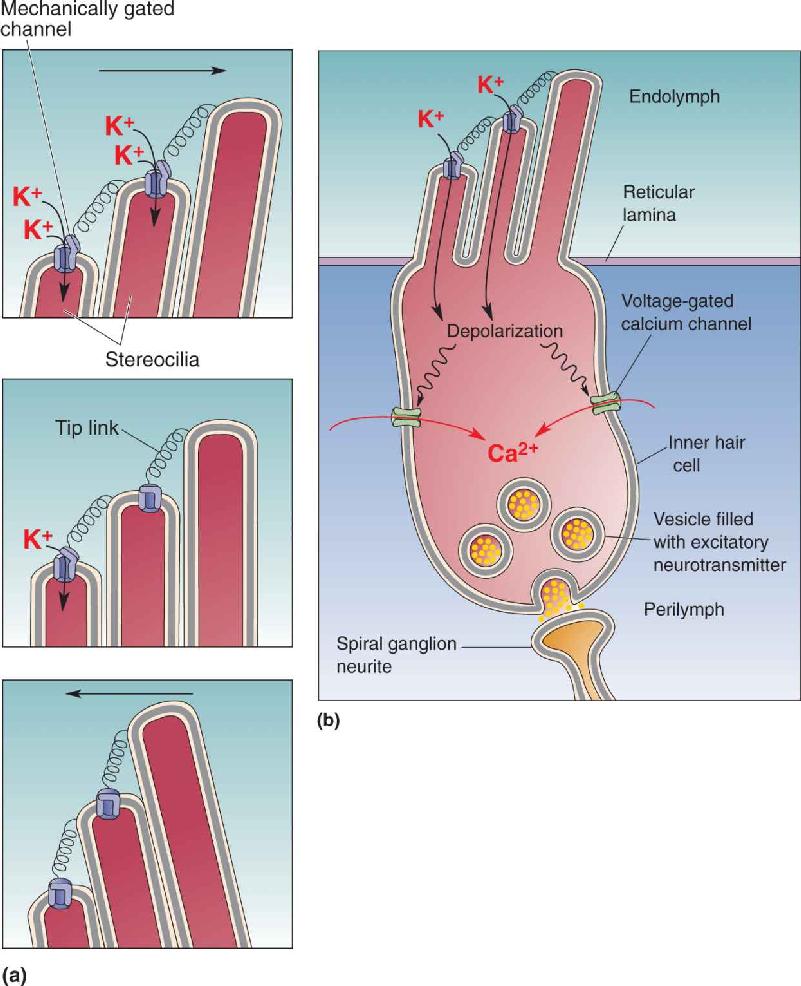
Transduction by Hair Cells. When the basilar membrane moves in response to a motion at the stapes, the entire foundation supporting the hair cells moves because the basilar membrane, rods of Corti, reticular lamina, and hair cells are all rigidly connected. These structures move as a unit, pivoting up toward the tectorial membrane or away from it (Figure 11.13). When the basilar membrane moves up, the reticular lamina moves up and in toward the modiolus. Conversely, downward motion of the basilar membrane causes the reticular lamina to move down and away from the modiolus. When the reticular lamina moves inward or outward relative to the modiolus, it also moves in or out with respect to the tectorial membrane. Because the tectorial membrane holds the tips of the outer hair cell stereocilia, the lateral motion of the reticular lamina relative to the tectorial membrane bends the stereocilia on the outer hair cells one way or the other. The tips of stereocilia from inner hair cells are also bent, probably because they are pushed by moving endolymph. Aligned actin filaments make stereocilia rigid rods, and they bend only at the base, where they attach to the top of the hair cell. Cross-link filaments make the stereocilia stick to one another, so all the cilia move as a unit. Now imagine a sound wave making the basilar membrane jiggle between the two positions shown in parts a and b of Figure 11.13, and it will be clear how the hair cell cilia are bent back and forth by the tectorial membrane.

FIGURE 11.13 The bending of stereocilia produced by the upward motion of the basilar membrane. (a) At rest, the hair cells are held between the reticular lamina and the basilar membrane, and the tips of the outer hair cell stereocilia are attached to the tectorial membrane. (b) When sound causes the basilar membrane to deflect upward, the reticular lamina moves up and inward toward the modiolus, causing the stereocilia to bend outward. Description
Determining how hair cells convert the bending of stereocilia into neural signals was a very challenging problem. Because the cochlea is encased in bone, it is difficult to record from the hair cells. In the 1980s, A. J. Hudspeth and his colleagues, then at the California Institute of Technology, pioneered a new approach in which hair cells are isolated from the inner ear and studied in vitro. The in vitro technique has revealed much about the transduction mechanism. Recordings from hair cells indicate that when the stereocilia bend in one direction, the hair cell depolarizes, and when they bend in the other direction, the cell hyperpolarizes (Figure 11.14a). When a sound wave causes the stereocilia to bend back and forth, the hair cell generates a receptor potential that alternately hyperpolarizes and depolarizes from the resting potential of –70 mV (Figure 11.14b).

FIGURE 11.14 Hair cell receptor potentials. (a) The hair cell depolarizes or hyperpolarizes, depending on the direction in which the stereocilia bend. (b) The hair cell receptor potential closely follows the air pressure changes during a low-frequency sound. Description
To appreciate just how efficiently the ear works, take a moment to notice the scale on the X axis of Figure 11.14a. Its unit is nm; recall that 1 nm equals 10–9 m. The graph shows that the receptor potential of the hair cell is saturated by the time the tips of its stereocilia have moved about 20 nm to the side; this is what an extremely loud sound might do. But the softest sound you can hear moves the stereocilia only 0.3 nm to each side, which is an astoundingly small distance—about the diameter of a large atom! Since each stereocilium is about 500 nm (or 0.5 μm) in diameter, a very soft sound needs to wiggle the stereocilia only about 1/1000 of their diameter in order to produce a perceptible sound. How does the hair cell transduce such infinitesimally small amounts of sound energy?
The tip of each stereocilium has a special type of ion channel that is induced to open and close by the bending of stereocilia. When these mechanosensitive transduction channels are open, an inward ionic current flows and generates the hair cell receptor potential. Despite considerable research effort, the molecular identity of the channels is still uncertain. One reason the channels have been so difficult to identify is that there are so few of them; the tip of each stereocilium has only one or two such channels, and an entire hair cell may have only 100. Some recent experiments suggest that the hair cell transduction channels belong to the transmembrane protein-like (TMC) family of proteins, but other studies challenge this conclusion. All we know for sure is that energetic research on this issue will continue.
Figure 11.15 shows how the transduction channels are believed to function. A stiff filament called a tip link connects each channel to the upper wall of the adjacent cilium. When the cilia are pointing straight up, the tension on the tip link causes the channel to spend part of the time in the opened state, allowing a small amount of K+ to move from the endolymph into the hair cell. Displacement of the cilia in one direction increases tension on the tip link, increasing the rate of channel openings and the amount of inward K+ current. Displacement in the opposite direction relieves tension on the tip link, thereby causing the channel to spend more time closed, reducing inward K+ movement. The entry of K+ into the hair cell causes a depolarization, which in turn activates voltage-gated calcium channels (Figure 11.15b). The entry of Ca2+ triggers the release of the neurotransmitter glutamate, which activates the spiral ganglion fibers lying postsynaptic to the hair cell.

FIGURE 11.15 Depolarization of a hair cell. (a) Ion channels on stereocilia tips are opened when the tip links joining the stereocilia are stretched. (b) The entry of K+ depolarizes the hair cell, which opens voltage-gated calcium channels. Incoming Ca–+ leads to the release of neurotransmitter from synaptic vesicles, which then diffuses to the postsynaptic neurite from the spiral ganglion. Description
It is interesting that the opening of K+ channels produces a depolarization of the hair cell, whereas the opening of K+ channels hyperpolarizes most neurons. The reason that hair cells respond differently from neurons is the unusually high K+ concentration in endolymph, which yields a K+ equilibrium potential of 0 mV, compared to the equilibrium potential of –80 mV in typical neurons. Another reason that K+ is driven into hair cells is the 80 mV endocochlear potential, which helps create a 125 mV gradient across the stereocilia membranes.
Hair Cells and the Axons of the Auditory Nerve. The auditory nerve consists of the axons of neurons whose cell bodies are located in the spiral ganglion. Thus, the spiral ganglion neurons, which are the first in the auditory pathway to fire action potentials, provide all the auditory information sent to the brain. Curiously, the numbers of auditory axons contacting the inner and outer hair cells are enormously different. There are about 35,000–50,000 neurons altogether in the spiral ganglion. Despite the fact that inner hair cells are outnumbered by outer hair cells by a factor of 3 to 1, more than 95% of the spiral ganglion neurons communicate with the relatively small number of inner hair cells, and less than 5% receive synaptic input from the more numerous outer hair cells (Figure 11.16). Consequently, one spiral ganglion fiber receives input from only one inner hair cell; moreover, each inner hair cell feeds about 10 spiral ganglion neurites. The situation is the opposite with outer hair cells. Because they outnumber their spiral ganglion cells, one spiral ganglion fiber synapses with numerous outer hair cells.

FIGURE 11.16 The innervation of hair cells by neurons from the spiral ganglion. Description
Simply based on these numbers, we can infer that the vast majority of the information leaving the cochlea comes from inner hair cells. If it’s true that the brain is paying very little attention to the outer hair cells, then what are the outer hair cells for?
Amplification by Outer Hair Cells. Given that outer hair cells far outnumber inner hair cells, it seems paradoxical that most of the cochlear output is derived from inner hair cells. However, ongoing research suggests that outer hair cells play a critical role in sound transduction. Ironically, one clue to the nature of this role was the discovery that the ear not only transduces sound; it can create it, too (Box 11.3).
Sensory systems are supposed to detect stimulus energy in the environment, not generate it. Can you imagine eyes glowing in the dark, or noses smelling like roses? How about ears buzzing loudly? The truth is that retinas don’t radiate light, and olfactory receptors don’t emit odors, but some ears can definitely generate sounds loud enough for a bystander to hear! Such sounds are called otoacoustic emissions. In one early description, a man sitting next to his dog realized the animal was humming; after some anxious investigation, he discovered that the sound came from one of the dog’s ears.
The ears of all vertebrates, including humans, can emit sounds. Presenting a short sound stimulus, such as a click, to a normal human ear causes an “echo” that can be picked up with a sensitive microphone in the auditory canal. We don’t usually notice such echoes because they are too faint to be heard over other sounds in the environment.
Ears that emit relatively loud sounds spontaneously, in the absence of any incoming sound, have usually sustained cochlear damage as the result of exposure to extremely loud sounds (from explosions, machines, rock bands), drugs, or disease. If spontaneous otoacoustic emissions are loud enough, they may cause one form of tinnitus—a ringing in the ears (see Box 11.6).
The mechanism that causes the ear to generate its own sounds—the cochlear amplifier—is the same one that functions to improve its detection of environmental sounds, but operating in reverse. Normal outer hair cells, stimulated with a click, react with a quick movement that drives the cochlear fluids and membranes, which move the ossicles, and ultimately vibrate the tympanic membrane to produce sound in the outside air (the echo). Spontaneous emissions occur because the sensitivity of the cochlear amplifier is very high. Most people with normal hearing can perceive them in an exceptionally quiet environment.
Damaged regions of the cochlea can somehow facilitate the spontaneous movement of some outer hair cells, so that they vibrate all the time. Strangely enough, most people are unaware that their ears are broadcasting sounds. Apparently, their central auditory neurons recognize the spontaneous cochlear activity as noise and suppress the perception of it. The benefit is that they are spared from an otherwise maddening tinnitus, but the cost is a partial hearing loss in the affected frequency range.
Because otoacoustic emissions are a normal attribute of ears, they can be used as a quick and easy test of ear function. A series of sounds is played into the ears, and the echoes they evoke are recorded and analyzed. The characteristics of the echoes can tell us a lot about the function of the middle and inner ears. This is especially useful for testing people who are unable to tell the examiner whether or not they have heard test sounds—newborn babies, for example.
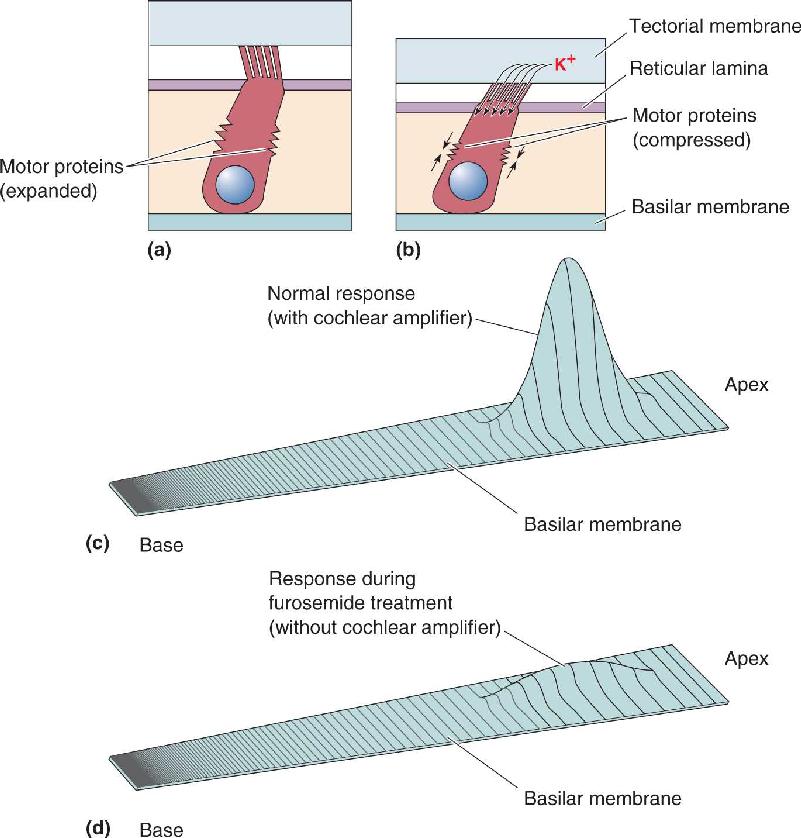
Outer hair cells seem to act like tiny motors that amplify the movement of the basilar membrane during low-intensity sound stimuli. This action of the outer hair cells on the basilar membrane is called the cochlear amplifier. There are two molecular mechanisms that probably contribute to this amplifier. The first, and best understood, mechanism involves special motor proteins found only in the membranes of outer hair cells (Figure 11.17a). Motor proteins can change the length of outer hair cells, and outer hair cells respond to sound with both a receptor potential and a change in length (Figure 11.17b). The motor proteins do not resemble any other system of cellular movement. The hair cells’ motor is driven by the receptor potential, and it does not use adenosine triphosphate (ATP) as an energy source. It is also extremely fast, as it must be able to keep up with the movements induced by high-frequency sounds. The hair cell’s primary motor is an unusual protein called prestin (from the musical notation presto, meaning “fast”). Prestin molecules are tightly packed into the membranes of the outer hair cell bodies, and they are required for outer hair cells to move in response to sound. A second possible molecular mechanism of the cochlear amplifier is located right in the hair bundles. A special type of the contractile protein myosin is attached to the upper end of the tip links. Myosin and other tip-link proteins may somehow rapidly enhance the movement of hairs in response to weak sounds; this idea is controversial.

FIGURE 11.17 Amplification by outer hair cells. (a) Motor proteins in the membranes of outer hair cells. (b) Bending of the stereocilia causes potassium to enter the hair cell, depolarizing it, and triggering motor proteins to activate and shorten the hair cell. (c) The shortening and lengthening of the hair cell increase the flexing of the basilar membrane. (d) Furosemide decreases hair cell transduction, consequently reducing the flexing of the basilar membrane. (Source: Adapted from Ashmore and Kolston, 1994, Figs. 2, 3.) Description
Because outer hair cells are attached to the basilar membrane and reticular lamina, when motor proteins change the length of the hair cell, the basilar membrane is pulled toward or pushed away from the reticular lamina and tectorial membrane. This is why the word “motor” is used; the outer hair cells actively change the physical relationship between the cochlear membranes.
The motor effect of outer hair cells makes a significant contribution to the traveling wave that propagates down the basilar membrane. This was first demonstrated in 1991 by Mario Ruggero and Nola Rich at the University of Minnesota, who administered the chemical furosemide into experimental animals. Furosemide temporarily decreases the transduction that normally results from the bending of stereocilia on hair cells, and it was found to significantly reduce the movement of the basilar membrane in response to sound (Figure 11.17c, d). This effect of furosemide is believed to result from inactivation of the outer hair cell motor proteins and loss of the cochlear amplifier. When the outer hair cells amplify the response of the basilar membrane, the stereocilia on the inner hair cells bend more, and the increased transduction process in the inner hair cells produces a greater response in the auditory nerve. Through this feedback system, therefore, outer hair cells contribute significantly to the output of the cochlea. Without the cochlear amplifier, the peak movement of the basilar membrane would be about 100-fold smaller.
The effect of outer hair cells on the response of inner hair cells can be modified by neurons outside the cochlea. In addition to the spiral ganglion afferents that project from the cochlea to the brain stem, there are also about 1000 efferent fibers projecting from the brain stem toward the cochlea. These efferents diverge widely, synapsing onto outer hair cells and releasing acetylcholine. Stimulation of these efferents changes the shape of the outer hair cells, thereby affecting the responses of inner hair cells. In this way, descending input from the brain to the cochlea can regulate auditory sensitivity.
The amplifying effect of outer hair cells explains how certain antibiotics (e.g., kanamycin) that damage hair cells can lead to deafness. After excessive exposure to antibiotics, many inner hair cells are less sensitive to sound. However, the antibiotic almost exclusively damages outer hair cells, not inner hair cells. For this reason, deafness produced by antibiotics is thought to be a consequence of damage to the cochlear amplifier (i.e., outer hair cells), demonstrating just how essential a role the amplifier plays.
Prestin, the protein that is essential for the outer hair cells’ motor, is also necessary for the cochlear amplifier to function. When the gene that encodes prestin is eliminated from mice, the animals are nearly deaf; their ears are more than 100-fold less sensitive to sound than normal.
The auditory pathway appears more complex than the visual pathway because there are more nuclei intermediate between the sensory organ and the cortex. Also, in contrast to the visual system, there are many more alternative pathways by which signals can travel from one nucleus to the next. Nonetheless, the amount of information processing in the two systems is similar when you consider that the cells and synapses of the auditory system in the brain stem are analogous to interactions in the layers of the retina. We will now look at auditory circuitry, focusing on the transformations of auditory information that occur along the way.
Afferents from the spiral ganglion enter the brain stem in the auditory–vestibular nerve. At the level of the medulla, the axons innervate the dorsal cochlear nucleus and ventral cochlear nucleus ipsilateral to the cochlea where the axons originated. Each axon branches so that it synapses on neurons in both cochlear nuclei. From this point on, the system gets more complicated, and the connections are less well understood, because there are multiple parallel pathways. Rather than trying to describe all of these connections, we will follow one particularly important pathway from the cochlear nuclei to auditory cortex (Figure 11.18). Cells in the ventral cochlear nucleus send axons that project to the superior olive (also called the superior olivary nucleus) on both sides of the brain stem. Axons of the olivary neurons ascend in the lateral lemniscus (a lemniscus is a collection of axons) and innervate the inferior colliculus of the midbrain. Many efferents of the dorsal cochlear nucleus follow a route similar to the pathway from the ventral cochlear nucleus, but the dorsal path bypasses the superior olive. Although there are other routes from the cochlear nuclei to the inferior colliculus, with additional intermediate relays, all ascending auditory pathways converge onto the inferior colliculus. The neurons in the inferior colliculus send axons to the medial geniculate nucleus (MGN) of the thalamus, which in turn projects to auditory cortex.

FIGURE 11.18 Auditory pathways. Neural signals can travel from the spiral ganglion to auditory cortex via numerous pathways. Here, a primary pathway is shown schematically (at left) and through brain stem cross sections. Notice that only the connections from one side are illustrated. Description
Before moving on to the response properties of auditory neurons, we should make several points:
- Projections and brain stem nuclei other than the ones described contribute to the auditory pathways. For instance, the inferior colliculus sends axons not only to the MGN but also to the superior colliculus (where the integration of auditory and visual information occurs) and to the cerebellum.
- There is extensive feedback in the auditory pathways. For instance, brain stem neurons send axons that contact outer hair cells, and auditory cortex sends axons to the MGN and inferior colliculus.
- Each cochlear nucleus receives input from just the one ear on the ipsilateral side; all other auditory nuclei in the brain stem receive input from both ears. This explains the clinically important fact that the only way by which brain stem damage can produce deafness in one ear is if a cochlear nucleus (or auditory nerve) on one side is destroyed.
Response Properties of Neurons in the Auditory Pathway
To understand the transformations of auditory signals that occur in the brain stem, we must first consider the nature of the input from the neurons in the spiral ganglion of the cochlea. Because most spiral ganglion cells receive input from a single inner hair cell at a particular location on the basilar membrane, they fire action potentials only in response to sound within a limited frequency range. After all, hair cells are excited by deformations of the basilar membrane, and each portion of the membrane is maximally sensitive to a particular range of frequencies.
Figure 11.19 shows the results of an experiment in which action potentials were recorded from a single auditory nerve fiber (i.e., the axon of a spiral ganglion cell). The graph represents the firing rate in response to sounds at different frequencies. The neuron is most responsive to sound at one frequency, called the neuron’s characteristic frequency, and it is less responsive at neighboring frequencies. This type of frequency tuning is seen in many neurons in each of the relays from cochlea to cortex.

FIGURE 11.19 The response of an auditory nerve fiber to different sound frequencies. This neuron is frequency-tuned and has its greatest response at the characteristic frequency. (Source: Adapted from Rose, Hind, Anderson, and Brugge, 1971, Fig. 2.) Description
As one ascends, the auditory pathway in the brain stem, the response properties of the cells become more diverse and complex, just as in the visual pathway. For instance, some cells in the cochlear nuclei are especially sensitive to sounds varying in frequency over time (think of the sound of a trombone as it slides from a low note to a high note). In the MGN, there are cells that respond to fairly complex sounds such as vocalizations, as well as other cells that show simple frequency selectivity, as in the auditory nerve. An important development in the superior olive is that cells receive input from cochlear nuclei on both sides of the brain stem. As discussed later, such binaural neurons are probably important for sound localization.
If you stop reading this book for a moment, you can focus on the many sounds around you. You can probably hear sounds you have been ignoring, and you can selectively pay attention to different sounds occurring at the same time. We are usually bathed in an amazing diversity of sounds—from chattering people to cars to electrical noises to sounds generated within our own bodies—and our brain must be able to analyze just the important sounds while ignoring the noise. We cannot yet account for the perception of each of these sounds by pointing to particular neurons in the brain. However, most sounds have certain features in common, including intensity, frequency, and the location from which they emanate. Each of these features is represented differently in the auditory pathway.
Information about sound intensity is coded in two interrelated ways: the firing rates of neurons and the number of active neurons. As a stimulus gets more intense, the basilar membrane vibrates with greater amplitude, causing the membrane potential of the activated hair cells to be more depolarized or hyperpolarized. As a result, the nerve fibers with which the hair cells synapse fire action potentials at greater rates. In Figure 11.19, the auditory nerve fiber fires faster to the same sound frequencies when the intensity is increased. In addition, more intense stimuli produce movements of the basilar membrane over a greater distance, which leads to the activation of more hair cells. In a single auditory nerve fiber, this increase in the number of activated hair cells causes a broadening of the frequency range to which the fiber responds. The loudness we perceive is correlated with the number of active neurons in the auditory nerve (and throughout the auditory pathway) and with their firing rates.
From the hair cells in the cochlea through the various nuclei leading to auditory cortex, most neurons are sensitive to stimulus frequency. They are most sensitive at their characteristic frequency. How is frequency represented in the central nervous system?
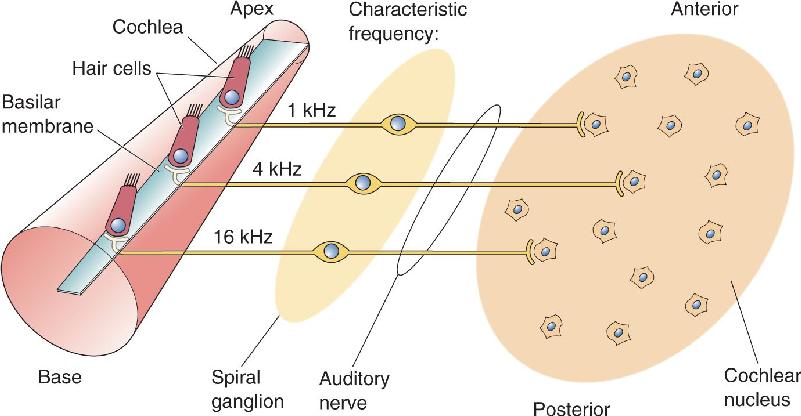
Tonotopy. Frequency sensitivity is largely a consequence of the mechanics of the basilar membrane because different portions of the membrane are maximally deformed by sound of different frequencies. Moving from the base to the apex of the cochlea, a progressive decrease occurs in the frequency that produces a maximal deformation of the basilar membrane. This is an example of tonotopy, as we discussed earlier. There is a corresponding tonotopic representation in the auditory nerve; auditory nerve fibers connected to hair cells near the apical basilar membrane have low characteristic frequencies, and those connected to hair cells near the basal basilar membrane have high characteristic frequencies (Figure 11.20). When auditory axons in the auditory–vestibular nerve synapse in the cochlear nuclei, they do so in an organized pattern based on characteristic frequency. Nearby neurons have similar characteristic frequencies, and a systematic relationship exists between position in the cochlear nucleus and characteristic frequency. In other words, there is a map of the basilar membrane within the cochlear nuclei.

FIGURE 11.20 Tonotopic maps on the basilar membrane and cochlear nucleus. From the base to the apex of the cochlea, the basilar membrane resonates with increasingly lower frequencies. This tonotopy is preserved in the auditory nerve and cochlear nucleus. In the cochlear nucleus, there are bands of cells with similar characteristic frequencies; characteristic frequencies increase progressively from anterior to posterior. Description
Because of the tonotopy present throughout the auditory system, the location of active neurons in auditory nuclei is one indication of the frequency of the sound. However, frequency must be coded in some way other than the site of maximal activation in tonotopic maps, for two reasons. One reason is that these maps do not contain neurons with very low characteristic frequencies, below about 200 Hz. As a result, the site of maximal activation might be the same for a 50 Hz tone as for a 200 Hz tone, so there must be some other way to distinguish them. The second reason that something other than tonotopy is needed is that the region of the basilar membrane maximally displaced by a sound depends on its intensity in addition to its frequency (see Figure 11.19). At a fixed frequency, a more intense sound will produce a maximal deformation at a point further up the basilar membrane than a less intense sound.
Phase Locking. The timing of neural firing provides an additional type of information about sound frequency that complements information derived from tonotopic maps. Recordings made from neurons in the auditory nerve show phase locking, the consistent firing of a cell at the same phase of a sound wave (Figure 11.21). If you think of a sound wave as a sinusoidal variation in air pressure, a phase-locked neuron would fire action potentials at the peaks, or the troughs, or some other constant location on the wave. At low frequencies, some neurons fire action potentials every time the sound is in a particular phase (Figure 11.21a). This makes it easy to determine the frequency of the sound; it is the same as the frequency of the neuron’s action potentials.

FIGURE 11.21 Phase locking in the response of auditory nerve fibers. Sound at a low frequency can elicit a phase-locked response, either (a) on every cycle of the stimulus or (b) on some fraction of the cycles. (c) At high frequencies, the response does not have a fixed phase relationship to the stimulus. Description
Phase locking can still occur even if an action potential has not fired on every cycle (Figure 11.21b). For instance, a neuron may respond to a 1000 Hz sound with an action potential on only perhaps 25% of the cycles of the input, but those action potentials still always occur at the same phase of the sound. If you have a group of such neurons, each responding to different cycles of the input signal, it is possible to have a response to every cycle (by some member of the group) and thus a measure of sound frequency. It is likely that intermediate sound frequencies are represented by the pooled activity of a number of neurons, each of which fires in a phase-locked manner; this is called the volley principle. Phase locking occurs with sound waves up to about 5 kHz. Above this point, the action potentials fired by a neuron occur at random phases of the sound wave (Figure 11.21c) because the intrinsic variability in the timing of the action potential becomes comparable to the time interval between successive cycles of the sound. In other words, the sound waves cycle too fast for the action potentials of single neurons to accurately represent their timing. Above 5 kHz, frequencies are represented by tonotopy alone.
Many auditory neurons in the brainstem have peculiar membrane properties that make them uniquely sensitive to the precise timing of their synaptic inputs. Adaptations for precise timing are particularly impressive in neurons of the cochlear nuclei, as research by Donata Oertel and her colleagues at the University of Wisconsin has demonstrated (Box 11.4).
While the timing of action potentials carries information everywhere in the brain, the time scales on which neurons work vary dramatically. Neurons in the auditory nuclei of the brain stem can fire with a temporal precision better than 200 μsec. Contrast this with cortical neurons, whose responses to identical stimuli are a hundred-fold less precise. In the auditory system, the timing of firing conveys important sensory information about the pitch of sounds and whether sounds come from the right or left.
In the mid-1960s, computers made possible detailed analyses of the relationship between the waveforms of sounds and the firing of neurons. These studies, some done by my colleagues at the University of Wisconsin, revealed that auditory neurons encode the frequency of sounds not only by their position in the tonotopic map but also by firing in phase with the sounds, by phase locking. This temporal code breaks down at sound frequencies higher than 5 kHz because the firing of neurons is not precise enough to resolve periods shorter than about 200 μsec.
Phase locking at low frequencies is valuable to human beings. First, our impressive ability to distinguish tones as similar as 1000 Hz and 1002 Hz seems to depend on phase locking by neurons in the brain stem. Phase-locked neurons also detect the relative time of arrival of a sound at the two ears with every cycle of the sound, a mechanism important for localizing sounds in the horizontal plane.
How can auditory neurons convey information with a temporal precision of 200 μsec across multiple synapses—hair cells to spiral ganglion cells, to neurons of the cochlear nuclear nucleus, to neurons in the superior olive—using synaptic potentials and action potentials whose durations are in the millisecond range? To achieve this, the firing of postsynaptic neurons must follow the firing of the presynaptic neurons rapidly and with an unvarying delay.
In 1979, Bill Rhode, Phil Smith, and I began to address these issues by making intracellular recordings in anesthetized cats, but those experiments were terribly difficult. One of the problems was that auditory brain stem nuclei are difficult to reach, being surrounded by cerebellum, the inner ear, and the jaw. Another was that blood pulsations and respiratory movements made the microelectrodes unstable. In 1980, I realized I could eliminate these difficulties by making recordings in brain slices, a technique then being used to study the rodent hippocampus and the chicken brain stem. I developed a slice preparation of the mouse cochlear nuclei. There were lots of details to work out. I had to learn how to remove the brain stem without stretching the auditory nerve, to optimize the properties of the saline that bathed the tissue, to generate a flow of saline fast enough to promote efficient gas exchange but not so turbulent that it yanked the electrodes out of cells. One of the first things I discovered is that some auditory neurons have exceptionally low membrane resistance and fast time constants. These properties help them sharpen and convey precise timing information. Auditory neurons are specialized for precision in other ways as well. Their synapses deliver exceptionally large currents using the speediest subtypes of the glutamate receptors to depolarize cells with low resistances rapidly.
A critical function of some auditory neurons is coincidence detection—detecting when two inputs arrive at the same time. Two groups of auditory neurons, octopus cells of the cochlear nuclei and principal cells of the superior olivary nucleus, are truly exquisite coincidence detectors. Nace Golding, Ramazan Bal, and Michael Ferragamo, working in my lab, demonstrated that octopus cells have exceptionally large, mutually opposing types of voltage-sensitive ion channels. These give the cells short time constants and allow them to detect coincidence in the submillisecond time range.
Nace and I puzzled over the fact that synaptic potentials sum over a fraction of one millisecond, yet auditory nerve inputs to octopus cells are activated by a traveling wave that sweeps down the cochlea over multiple milliseconds. Matthew McGinley helped to resolve that puzzle. Octopus cells earn their name by extending dendrites in only one direction; cell bodies send dendrites across the tonotopic array of auditory nerve fibers so that the earliest synaptic inputs (tuned to high frequencies) impinge on the tips of dendrites while the latest synaptic inputs (tuned to low frequencies) terminate near the cell body. EPSPs take time to propagate along the dendrites of octopus cells; different dendritic delays thus compensate for the traveling wave delays in the cochlea, allowing octopus cells to signal the occurrence of clicks and onsets of complex sounds with a single, sharply timed action potential. When Nace established his own lab, he studied principal cells of the medial superior olive, which compare the timing of inputs to the two ears. He found that they were in many ways similar to the octopus cells he had worked on as a graduate student. Both cells have specialized ion channels that make them fast, both use their dendrites for detecting coincident EPSPs, and both balance their need to fire action potentials with their need to integrate inputs without the interference of action potentials. And both are still devilishly difficult to record from in vivo!
To summarize, here is how different sound frequencies are represented by brain stem neurons. At very low frequencies, phase locking is used; at intermediate frequencies, both phase locking and tonotopy are useful; and at high frequencies, tonotopy must be relied on to indicate sound frequency.
While the use of frequency information is essential for interpreting sounds in our environment, sound localization can be of critical importance for survival. If a predator is about to eat you, finding the source of a sudden sound and running away are much more important than analyzing the subtleties of the sound. Wild animals do not eat humans very often anymore, but there are other situations in which sound localization can be helpful. If you are carelessly crossing the street, your localization of a car’s horn may be all that saves you. Our current understanding of the mechanisms underlying sound localization suggests that we use different techniques for locating sources in the horizontal plane (left–right) and vertical plane (up–down).
If you close your eyes and plug one ear, you can locate a bird singing as it flies overhead almost as well as with both ears open. But if you try to locate the horizontal position of a duck quacking as it swims across a pond, you’ll find that you’re much less able using only one ear. Thus, good horizontal localization requires a comparison of the sounds reaching the two ears, whereas good vertical localization does not.
An obvious cue to the location of a sound source is the time at which the sound arrives at each ear. If we aren’t facing a sound’s source directly, it takes the sound longer to reach one ear than the other. For instance, if a sudden noise comes at you from the right, it will reach your right ear first (Figure 11.22a); it will arrive at your left ear later, after what is known as an interaural time delay. If the distance between your ears is 20 cm, sound coming from the right, perpendicular to your head, will reach your left ear 0.6 msec after reaching your right ear. If the sound comes from straight ahead, there is no interaural delay; and at angles between straight ahead and perpendicular, the delay will be between 0 and 0.6 msec (Figure 11.22b). Sounds from the left side yield delays opposite to those on the right. Thus, there is a simple relationship between location and interaural delay. Detected by specialized neurons in the brain stem, the delay enables us to locate the source of the sound in the horizontal plane. The interaural delays we can detect are impressively brief. People can discriminate the direction of a sound source in the horizontal plane with a precision of about 2°. This demands that they discriminate the 11 μsec difference between the times it takes a sound to reach their two ears.

FIGURE 11.22 Interaural time delay as a cue to the location of sound. (a) Sound waves coming from the right side will reach the right ear first, and there will be a large interaural delay before the sound propagates to the left ear. (b) If the sound comes from straight ahead, there is no interaural delay. Delays for three different sound directions are shown. Description
If we don’t hear the onset of a sound because it is a continuous tone rather than a sudden noise, however, we cannot know the initial arrival times of the sound at the two ears. Thus, continuous tones pose more of a problem for sound localization because they are always present at both ears. However, we can still use arrival time to localize the sound but in a slightly different manner from localizing a sudden sound. The only thing that can be compared with continuous tones is the time at which the same phase of the sound wave reaches each ear. Imagine you are exposed to a 200 Hz sound coming from the right. At this frequency, one cycle of the sound covers 172 cm, which is much more than the 20 cm distance between your ears. After a peak in the sound pressure wave passes the right ear, it takes 0.6 msec, the time for sound to travel 20 cm, before you detect the peak at the left ear. Of course, if the sound is straight ahead, peaks in the continuous tone will reach the ears simultaneously. Because the sound wave is much longer than the distance between the ears, we can reliably use the interaural delay of the peak in the wave to determine sound location.
Things are more complicated with continuous tones at high frequencies. Suppose that the sound coming from the right now has a frequency of 20,000 Hz, which means that one cycle of the sound covers 1.7 cm. After a peak reaches the right ear, does it still take 0.6 msec before a peak arrives at the left ear? No! It takes a much shorter time because many peaks of such a high-frequency wave will fit between your ears. No longer is there a simple relationship between the direction the sound comes from and the arrival times of the peaks at the two ears. Interaural arrival time is simply not useful for locating continuous sounds with frequencies so high that one cycle of the sound wave is smaller than the distance between your ears (i.e., greater than about 2000 Hz).
Fortunately, the brain has another process for sound localization at high frequencies. An interaural intensity difference exists between the two ears because your head effectively casts a sound shadow (Figure 11.23). There is a direct relationship between the direction the sound comes from and the extent to which your head shadows the sound to one ear. If sound comes directly from the right, the left ear will hear a significantly lower intensity (Figure 11.23a). With sound coming from straight ahead, the same intensity reaches the two ears (Figure 11.23b), and with sound coming from intermediate directions, there are intermediate intensity differences (Figure 11.23c). Neurons sensitive to differences in intensity can use this information to locate the sound. Intensity information cannot be used to locate sounds at lower frequencies because sound waves at these frequencies diffract around the head, and the intensities at the two ears are roughly equivalent. There is no sound shadow at low frequencies.

FIGURE 11.23 Interaural intensity difference as a cue to sound location. (a) With high-frequency sound, the head will cast a sound shadow to the left, when sound waves come from the right. Lower intensity sound in the left ear is a cue that the sound came from the right. (b) If the sound comes from straight ahead, a sound shadow is cast behind the head, but the sound reaches the two ears with the same intensity. (c) Sound coming from an oblique angle will partially shadow the left ear. Description
Let’s summarize the two processes for localizing sound in the horizontal plane. With sounds in the range of 20–2000 Hz, the process involves interaural time delay. From 2000–20,000 Hz, interaural intensity difference is used. Together these two processes constitute the duplex theory of sound localization.
The Sensitivity of Binaural Neurons to Sound Location. From our discussion of the auditory pathway, recall that neurons in the cochlear nuclei receive afferents only from the ipsilateral auditory–vestibular nerve. Thus, all of these cells are monaural neurons, meaning that they only respond to sound presented to one ear. At all later stages of processing in the auditory system, however, there are binaural neurons whose responses are influenced by sound at both ears. The response properties of binaural neurons imply that they play an important role in sound localization in the horizontal plane.
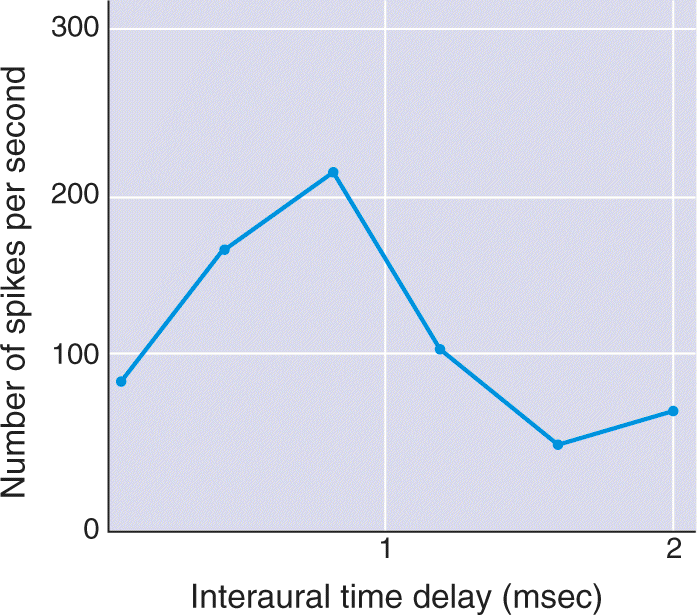
The first structure where binaural neurons are present is the superior olive. Although some controversy exists about the relationship between the activity of such neurons and the behavioral localization of sound, there are several compelling correlations. Neurons in the superior olive receive input from cochlear nuclei on both sides of the brain stem (see Figure 11.18). Cells in the cochlear nuclei that project to the superior olive typically have responses phase locked to lower frequency sound input. Consequently, an olivary neuron receiving spikes from the left and right cochlear nuclei can compute interaural time delay. Recordings made in the superior olive show that each neuron typically gives its greatest response to a particular interaural delay (Figure 11.24). Because interaural delay varies with sound location, each of these neurons may be encoding a particular position in the horizontal plane.

FIGURE 11.24 Responses of a neuron in the superior olive sensitive to interaural time delay. This neuron has an optimal delay of about 1 msec. Description
How can a neural circuit produce neurons sensitive to interaural delay? One possibility is to use axons as delay lines, and to measure small time differences precisely. A sound hitting the left ear triggers action potentials in the left cochlear nucleus, which propagate along afferent axons into the superior olive (Figure 11.25). Within 0.6 msec of hitting the left ear, that sound reaches the right ear (assuming the sound comes directly from the left) and triggers action potentials in axons from the right cochlear nucleus. However, because of the way the axons and neurons are arranged in the olive, the action potentials from each side take different lengths of time to arrive at the various postsynaptic neurons in the olive. For example, the axon from the left cochlear nucleus has a longer path to travel to neuron 3 in Figure 11.25 than the axon from the right cochlear nucleus; therefore, the arrival of the spike from the left side is delayed just enough that it coincides with the arrival of the spike from the right side. By arriving at precisely the same time, action potentials from the two sides produce excitatory postsynaptic potentials (EPSPs) that summate, yielding a larger EPSP that more strongly excites olivary neuron 3 than an EPSP from each ear could alone. When an interaural delay is more or less than 0.6 msec, the spikes do not arrive together, and thus the EPSPs they trigger do not summate as much.

FIGURE 11.25 Delay lines and neuronal sensitivity to interaural delay. Description
Other neurons in the superior olive are tuned to other interaural times because of systematic differences in the arrangement of axonal delay lines. To measure timing differences as accurately as possible, many neurons and synapses of the auditory system are specially adapted for rapid operation; their action potentials and EPSPs are much faster than those of most other neurons in the brain. There are limits to auditory time measurement of this type, however. Phase locking is essential for a precise comparison of the timing of inputs, and because phase locking occurs only at relatively low frequencies, it makes sense that interaural delays are useful only for localizing sounds of relatively low frequency.
The mechanism described in Figure 11.25 is clearly present in bird brains, but it is unlikely that mammals calculate interaural delays exactly this way. Studies on gerbils have suggested that synaptic inhibition, rather than axonal delay lines, generates the sensitivity of superior olivary neurons to interaural delay. It is possible that inhibition and axonal delay lines work together for this purpose.
In addition to their sensitivity to interaural delay, neurons in the superior olive are sensitive to the other sound location cue, interaural intensity. One type of neuron is moderately excited by sound presented to either ear but only gives a maximal response when both ears are stimulated. The other type of neuron is excited by sound in one ear but inhibited by sound in the other ear. Presumably, both types of neurons contribute to horizontal localization of high-frequency sound by encoding differences in interaural intensity.
Comparing inputs to both ears is not very useful for localizing sounds in the vertical plane because as a sound source moves up and down, neither the interaural delay nor the interaural intensity changes. This is why, as discussed earlier, localizing sounds in the vertical plane is much less affected by plugging one ear than localizing sounds in the horizontal plane. In order to seriously impair vertical sound localization, one must place a tube into the auditory canal to bypass the pinna. The sweeping curves of the outer ear are essential for assessing the elevation of a source of sound. The bumps and ridges apparently produce reflections of the entering sound. The delays between the direct path and the reflected path change as a sound source moves vertically (Figure 11.26). The combined sound, direct and reflected, is subtly different when it comes from above or below. In addition, the outer ear allows higher frequency sounds to enter the auditory canal more effectively when they come from an elevated source. Vertical localization of sound is seriously impaired if the convolutions of the pinna are covered.

FIGURE 11.26 Vertical sound localization based on reflections from the pinna.
Some animals are extremely good at vertical sound localization even though they do not have a pinna. For example, a barn owl can swoop down on a squeaking mouse in the dark, locating accurately by sound, not sight. Although owls do not have a pinna, they can use the same techniques we use for horizontal localization (interaural differences) because their two ears are at different heights on their head. Some animals have a more “active” system for sound localization than humans and owls. Certain bats emit sounds that are reflected off objects, and these echoes are used to locate objects without sight. Many bats detect and capture insects using reflected sound, analogous to the sonar used by ships. In 1989, James Simmons at Brown University made the startling discovery that bats can discriminate time delays that differ by as little as 0.00001 msec. This finding challenges our understanding of how the nervous system, using action potentials lasting almost a millisecond, can perform such fine temporal discriminations.
Axons leaving the MGN project to auditory cortex via the internal capsule in an array called the acoustic radiation. Primary auditory cortex (A1) corresponds to Brodmann’s area 41 in the temporal lobe (Figure 11.27a). The structure of A1 and the secondary auditory areas is in many ways similar to corresponding areas of the visual cortex. Layer I contains few cell bodies, and layers II and III contain mostly small pyramidal cells. Layer IV, where the medial geniculate axons terminate, is composed of densely packed granule cells. Layers V and VI contain mostly pyramidal cells that tend to be larger than those in the superficial layers. Let’s look at how these cortical neurons respond to sound.

FIGURE 11.27 Primary auditory cortex. (a) Primary auditory cortex (purple) and secondary auditory areas (green) on the superior temporal lobe. (b) Tonotopic organization within primary auditory cortex. The numbers are characteristic frequencies. Description
In general, neurons in monkey (and presumably human) A1 are relatively sharply tuned for sound frequency and possess characteristic frequencies covering the audible spectrum of sound. In electrode penetrations made perpendicular to the cortical surface in monkeys, the cells encountered tend to have similar characteristic frequencies, suggesting a columnar organization on the basis of frequency. In the tonotopic representation in A1, low frequencies are represented rostrally and laterally, whereas high frequencies are represented caudally and medially (Figure 11.27b). Roughly speaking, there are isofrequency bands running mediolaterally across A1. In other words, strips of neurons running across A1 contain neurons that have fairly similar characteristic frequencies.
In the visual system, it is possible to describe large numbers of cortical neurons as having some variation on a general receptive field that is either simple or complex. So far, it has not been possible to place the diverse auditory receptive fields into a similarly small number of categories. As they do at earlier stages in the auditory pathway, cortical neurons have different temporal response patterns; some have a transient response to a brief sound, and others have a sustained response.
In addition to the frequency tuning that occurs in most cells, some neurons are intensity-tuned, giving a peak response to a particular sound intensity. Even within a vertical column perpendicular to the cortical surface, considerable diversity can exist in the degree of tuning to sound frequency. Some neurons are sharply tuned for frequency, and others are barely tuned at all; the degree of tuning does not seem to correlate well with cortical layers. Other sounds that produce responses in cortical neurons include clicks, bursts of noise, frequency-modulated sounds, and animal vocalizations. Attempting to understand the role of these neurons that respond to seemingly complex stimuli is one of the challenges researchers currently face (Box 11.5).
The function of an animal’s brain is to help it stay alive and reproduce. Different species have vastly different habits and needs, and some animals have evolved a sensory system specialized for processing its favorite stimuli. The exaggerated systems of the sensory specialists, such as owls and bats, can help us understand how we sensory generalists work.
Barn owls find their prey (a scurrying mouse, for example) in the dark by listening very carefully. They are particularly adept at identifying and localizing faint sounds, and some of the neural mechanisms of sound localization were first understood in owls. Bats have a more unique and active auditory technique. They find their food (a fluttering moth, for example) by echolocating it. A bat emits brief calls and listens for the faint echoes reflected from the target. Bats require their cortex for proper echolocation. Studying bat cortex can certainly provide insight into how auditory cortex works in bats, but it may also enlighten us about the human cortex.
The most interesting stimuli for echolocating bats are their own calls and echoes. A bat’s language is very limited. To echolocate, most bats scream loudly at ultrasonic frequencies (20–100 kHz) using essentially a one-word vocabulary. The call of the mustached bat (Pteronotus parnellii) is very brief, no more than 20 msec long, and consists of a steady constant frequency (CF) part followed by a sweep of descending frequency—the frequency modulated (FM) part. Figure A is a graph of the bat’s call and echo, showing the frequencies of the sounds plotted against time. As it flies, the bat rapidly and continually repeats the call. By listening to its own calls and their echoes, and carefully comparing them in many ways, the bat builds a remarkably detailed auditory image of the nearby world. For example, the delay between the call and its echo depends on the distance to a reflecting target (1 msec of delay for each 17 cm of distance). If the target is moving toward or away from the bat, relative to the bat’s own movement, the frequency of the echo is Doppler-shifted higher or lower (think of the shifting pitch of an ambulance siren as it passes by you; a 1 kHz shift corresponds to a speed of about 3 m/s). A moth’s beating wings cause a rhythm in the echoes, and that helps the bat know there’s a particular kind of insect in front of it rather than something less edible. Many other subtle changes in the echo’s frequency, timing, loudness, and pattern tell the bat about other features of targets.

Figure A A bat’s call and echo. (Source: Adapted from Suga, 1995, p. 302.)
The processing of call-echo information by the auditory cortex of the mustached bat has been studied in great detail by Nobuo Suga at Washington University. Suga found that the bat’s auditory cortex is a mixture of distinct auditory areas. Many are specialized for detecting particular features important for echolocating, and others seem more generalized. For example, a large region is devoted to processing Doppler shifts of echoes right around 60 kHz, the CF part of the bat’s call; this area processes information about target velocity and location. Three separate areas detect call-echo delays and yield information about target distance.
The basic features of a bat’s calls and a human’s spoken words are similar, although human speech is much slower and lower in pitch. Human language syllables consist of particular combinations of CF periods, FM sweeps, brief pauses, and bursts of noise. For example, the syllable “ka” differs from “pa” because their initial FM sweeps bend in different directions (Figure B). The long “a” and the long “i” sound different because each one uses different combinations of CFs. It is very likely that the neuronal circuits that process speech sounds in human auditory cortex use principles very similar to those in the bat’s cortex. Interpreting those speech sounds as words, and understanding the concepts they imply, is the realm of language; the brain mechanisms of language will be discussed in Chapter 20.

Figure B A human’s spoken words. (Source: Adapted from Suga, 1995, p. 296.)
Given the wide variety of response types that neurophysiologists encounter in studying auditory cortex, you can understand why it is reassuring to see some sort of organization or unifying principle. One organizational principle already discussed is the tonotopic representation in many auditory areas. A second organizational principle is the presence in auditory cortex of columns of cells with similar binaural interaction. As at lower levels in the auditory system, one can distinguish cells that respond more to stimulation of both ears than to either ear separately, as well as cells that are inhibited if both ears are stimulated. As we discussed regarding the superior olive, neurons sensitive to interaural time delays and interaural intensity differences probably play a role in sound localization.
In addition to A1, other cortical areas located on the superior surface of the temporal lobe respond to auditory stimuli. Some of these higher auditory areas are tonotopically organized, and others do not seem to be. As in visual cortex, the stimuli that evoke the strongest responses in higher auditory areas tend to be more complex than those that best excite neurons at lower levels in the system. An example of specialization is Wernicke’s area, which we will discuss in Chapter 20. Destruction of this area does not interfere with the sensation of sound, but it seriously impairs the ability to interpret spoken language.
The Effects of Auditory Cortical Lesions and Ablation
Bilateral ablation of auditory cortex leads to deafness, but deafness is more often the consequence of damage to the ears (Box 11.6). A surprising degree of normal auditory function is retained after unilateral lesions in auditory cortex. This outcome is in marked contrast to that of the visual system, in which a unilateral cortical lesion of striate cortex leads to complete blindness in one visual hemifield. The reason for greater preservation of function after lesions in auditory cortex is that both ears send output to cortex in both hemispheres. In humans, the primary deficit that results from a unilateral loss of A1 is the inability to localize the source of a sound. It may be possible to determine which side of the head a sound comes from, but there is little ability to locate the sound more precisely. Performance on such tasks as frequency or intensity discrimination is near normal.
Although the effects of cortical lesions provide important information about the role of auditory cortex in perception, the perceptual deficit we all associate with the auditory system, deafness, usually results from problems in or near the cochlea. Deafness is conventionally considered in two categories: conduction deafness and nerve deafness.
Hearing loss caused by a disturbance in the conduction of sound from the outer ear to the cochlea is called conduction deafness. Causes of this sensory deficit range from something as simple as excessive wax in the ear to more serious problems such as rupture of the tympanic membrane or pathology of the ossicles. A number of diseases cause binding of the ossicles to the bone of the middle ear, impairing the transfer of sound. Fortunately, most of the mechanical problems in the middle ear that interfere with sound conduction can be treated surgically.
Nerve deafness is deafness associated with the loss of either neurons in the auditory nerve or hair cells in the cochlea. Nerve deafness sometimes results from tumors affecting the inner ear. It also can be caused by drugs that are toxic to hair cells, such as quinine and certain antibiotics, or exposure to loud sounds, such as explosions and loud music. Depending on the degree of cell loss, different treatments are possible. If the cochlea or auditory nerve on one side is completely destroyed, deafness in that ear will be absolute. However, a partial loss of hair cells is more common. In these cases, a hearing aid can be used to amplify the sound for the remaining hair cells. In more serious cases where hearing loss is bilateral and the auditory nerve is intact, cochlear implants are an important option (see Box 11.2).
With deafness, a person hears less sound than normal. With a hearing disorder called tinnitus, a person hears noises in the ears even in the absence of any sound stimulus. The subjective sensation can take many forms, including buzzing, humming, and whistling. You may have experienced a mild, temporary form of tinnitus after being at a party with really loud music; your brain may have had fun, but your hair cells were in shock! Tinnitus is a relatively common disorder that can seriously interfere with concentration and work if it persists. You can imagine how distracting it would be if you constantly heard whispering or humming or the crinkling of paper.
Tinnitus can be a symptom of a number of neurological problems. Although it frequently accompanies diseases involving the cochlea or auditory nerve, it may also result from exposure to loud sounds, abnormal vasculature of the neck, or simple aging. It now seems that many of the phantom sounds of tinnitus are caused by changes in central auditory structures, including auditory cortex. Damage to the cochlea or auditory nerve may induce alterations in the brain, such as a downregulation of synaptic inhibition. Although clinical treatment of tinnitus is often only partially successful, using a device that produces a constant sound in the affected ear(s) can often lessen the annoyance of the noise. For unknown reasons, the constant real sound is less annoying than the sound of the tinnitus that gets blocked.
Studies in experimental animals indicate that smaller lesions can produce rather specific localization deficits. Because of the tonotopic organization of A1, it is possible to make a restricted cortical lesion that destroys neurons with characteristic frequencies within a limited frequency range. Interestingly, there is a localization deficit only for sounds roughly corresponding to the characteristic frequencies of the missing cells. This finding reinforces the idea that information in different frequency bands may be processed in parallel by tonotopically organized structures.
Strangely enough, listening to music and balancing on a bicycle both involve sensations that are transduced by hair cells. The vestibular system monitors the position and movement of the head, gives us our sense of balance and equilibrium, and helps coordinate movements of the head and eyes, as well as adjustments to body posture. When the vestibular system operates normally, we are usually unaware of it. When its function is disrupted, however, the results can include the unpleasant, stomach-turning feelings we usually associate with motion sickness—vertigo and nausea, plus a sense of disequilibrium and uncontrollable eye movements.
The vestibular and auditory systems both use hair cells to transduce movements. Common biological structures often have common origins. In this case, the organs of mammalian balance and hearing both evolved from the lateral line organs present in aquatic vertebrates, including fish and some amphibians. Lateral line organs are small pits or tubes along an animal’s sides. Each pit contains clusters of hairlike sensory cells whose cilia project into a gelatinous substance that is open to the water in which the animal swims. The purpose of lateral line organs in many animals is to sense vibrations or pressure changes in the water. In some cases, they are also sensitive to temperature or electrical fields. Lateral line organs were lost as reptiles evolved, but the exquisite mechanical sensitivity of hair cells was adopted and adapted for use in the structures of the inner ear that derived from the lateral line.
In mammals, all hair cells are contained within sets of interconnected chambers called the vestibular labyrinth (Figure 11.28a). We have already discussed the auditory portion of the labyrinth, the spiraling cochlea (see Figure 11.6). The vestibular labyrinth includes two types of structures with different functions: the otolith organs, which detect the force of gravity and tilts of the head, and the semicircular canals, which are sensitive to head rotation. The ultimate purpose of each structure is to transmit mechanical energy, derived from head movement, to its hair cells. Each is sensitive to different kinds of movement not because their hair cells differ but because of the specialized structures within which the hair cells reside.

FIGURE 11.28 The vestibular labyrinth. (a) Locations of the otolith organs (utricle and saccule) and semicircular canals. (b) A vestibular labyrinth resides on each side of the head, with the semicircular canals arranged in parallel planes. Description
The paired otolith organs, called the saccule and the utricle, are relatively large chambers near the center of the labyrinth. The semicircular canals are the three arcing structures of the labyrinth. They lie in approximately orthogonal planes, which means there is an angle of about 90° between any pair of them (Figure 11.28b). A set of vestibular organs resides on each side of the head, mirror images of each other.
Each hair cell of the vestibular organs makes an excitatory synapse with the end of a sensory axon from the vestibular nerve, a branch of the auditory–vestibular nerve (cranial nerve VIII). There are about 20,000 vestibular nerve axons on each side of the head, and their cell bodies lie in Scarpa’s ganglion.
The saccule and utricle detect changes of head angle, as well as linear acceleration of the head. When you tilt your head, the angle between your otolith organs and the direction of the force of gravity changes. Linear acceleration also generates force, in proportion to the mass of an object. Forces due to linear acceleration are the sort you encounter when you ride in an elevator or a car as it starts or stops. In contrast, when a car or elevator moves smoothly at constant velocity, acceleration is zero, so there is no force (apart from gravitational force). That’s why you can fly steadily at 600 mph in a jet yet feel perfectly still; the sudden bouncing you experience during air turbulence, however, is another good example of the forces generated by linear acceleration, and of movements detected by your otolith organs.
Each otolith organ contains a sensory epithelium called a macula, which is vertically oriented within the saccule and horizontally oriented within the utricle when the head is upright. (Note that the vestibular macula and the retinal macula are entirely different structures.) The vestibular macula contains hair cells, which lie among a bed of supporting cells with their cilia projecting into a gelatinous cap (Figure 11.29). Movements are transduced by hair cells in the maculae when the hair bundles are deflected. The unique feature of the otolith organs is the tiny crystals of calcium carbonate called otoconia, 1–5 μm in diameter. (The word is Greek for “ear stone.”) Otoconia encrust the surface of the macula’s gelatinous cap, near the tips of the hair bundles, and they are the key to the tilt sensitivity of the macula. The otoconia have a higher density than the endolymph that surrounds them.

FIGURE 11.29 Macular hair cells responding to tilt. When the utricular macula is level (the head is straight), the cilia from the hair cells also stand straight. When the head and macula are tilted, gravity pulls the otoconia, which deform the gelatinous cap, and the cilia bend. Description
When the angle of the head changes, or when the head accelerates, a force is exerted on the otoconia; this exerts a force in the same direction on the gelatinous cap, which moves slightly, and the cilia of the hair cells bend. Not just any deflection will do, however. Each hair cell has one especially tall cilium, called the kinocilium. The bending of hairs toward the kinocilium results in a depolarizing, excitatory receptor potential. Bending the hairs away from the kinocilium hyperpolarizes and inhibits the cell. The cell is exquisitely direction-selective. If the hairs are bent perpendicular to their preferred direction, they barely respond. The transduction mechanism of vestibular hair cells is essentially the same as that in auditory hair cells (see Figure 11.15). As with auditory hair cells, only tiny hair movements are needed. The response saturates when the hairs are bent less than 0.5 μm, about the diameter of one cilium.
The head can tilt and move in any direction, but the hair cells of the utricle and saccule are oriented to transduce all of them effectively. The saccular maculae are oriented more or less vertically, while the utricular maculae are mostly horizontal (Figure 11.30). On each macula, the direction preferences of the hair cells vary in a systematic way. There are enough hair cells in each macula to cover a full range of directions. Because of the mirror-image orientation of the saccule and utricle on each side of the head, when a given head movement excites hair cells on one side, it will tend to inhibit hair cells in the corresponding location on the other. Thus, any tilt or acceleration of the head will excite some hair cells, inhibit others, and have no effect on the rest. The central nervous system, by simultaneously using the information encoded by the full population of otolithic hair cells, can unambiguously interpret all possible linear movements.

FIGURE 11.30 Macular orientation. (a) The macula in the utricle is horizontal. (b) The macula in the saccule is vertical. The arrows on each macula represent how the sheets of hair cells are polarized. Hair cells in the vicinity of an arrow are all polarized the same way; their stereocilia are all oriented so that bending them in the direction of the arrow depolarizes them. Description
The semicircular canals detect turning movements of the head, such as shaking your head from side to side or nodding up and down. Like the otolith organs, the semicircular canals also sense acceleration, but of a different kind. Angular acceleration is generated by sudden rotational movements, and it is the primary stimulus for the semicircular canals.
The hair cells of the semicircular canals are clustered within a sheet of cells, the crista, located within a bulge along the canal called the ampulla (Figure 11.31a). The cilia project into the gelatinous cupula, which spans the lumen of the canal within the ampulla. All the hair cells in an ampulla have their kinocilia oriented in the same direction, which means that they all get excited or inhibited together. The semicircular canals are filled with endolymph, the same fluid that fills the scala media of the cochlea. Bending of the cilia occurs when the canal is suddenly rotated about its axis like a wheel; as the wall of the canal and the cupula begin to spin, the endolymph tends to stay behind because of inertia. The sluggish endolymph exerts a force upon the cupula, much like wind upon a sail (Figure 11.31b). This force bows the cupula, which bends the cilia, which (depending on the direction of the rotation) either excites or inhibits the release of neurotransmitter from the hair cells onto the vestibular nerve axons.

FIGURE 11.31 A cross section through the ampulla of a semicircular canal. (a) The cilia of hair cells penetrate into the gelatinous cupula, which is bathed in the endolymph that fills the canals. (b) When the canal rotates leftward, the endolymph lags behind and applies force to the cupula, bending the cilia within it. Description
If head rotation is maintained at a constant velocity, the friction of endolymph with the canal walls eventually makes the two move together, thereby reducing and then eliminating the bending of the cupula after 15–30 seconds. Such adaptation to rotation can be seen clearly in the firing rates of vestibular axons from the canals (Figure 11.32). (This sort of prolonged head rotation is not something you encounter very often, unless you have a taste for certain amusement park rides.) When rotation of the head (and its canals) is finally stopped, the inertia of the endolymph causes the cupula to bend in the other direction, generating an opposite response from the hair cells and a temporary sensation of counter-rotation. This mechanism explains why you felt dizzy and unbalanced whenever, as a child, you stopped spinning your body like a top; your semicircular canals were temporarily sending the message that your body was still spinning, but in the opposite direction.

FIGURE 11.32 Push–pull activation of the semicircular canals. Head rotation causes the excitation of hair cells in one horizontal semicircular canal and the inhibition of hair cells in the other. The graphs show that long-lasting head rotation leads to adaptation of the firing in vestibular axons. When rotation is stopped, the vestibular axons from each side begin firing again, but with opposite patterns of excitation and inhibition. Description
Together, the three semicircular canals on one side of the head help sense all possible head rotation angles. This is further ensured because each canal is paired with another on the opposite side of the head (see Figure 11.28b). Each member of a pair sits within the same orientation plane as its partner and responds to rotation about the same axis. However, while rotation excites the hair cells of one canal, it inhibits the hair cells of its contralateral partner’s canal. Vestibular axons fire at high rates even at rest, so their activity can be driven either up or down depending on the direction of rotation. This “push–pull” arrangement—each rotation causing excitation on one side and inhibition on the other (see Figure 11.32)—optimizes the ability of the brain to detect rotational movements.
Central Vestibular Pathways and Vestibular Reflexes
The central vestibular pathways coordinate and integrate information about head and body movement and use it to control the output of motor neurons that adjust head, eye, and body positions. Primary vestibular axons from cranial nerve VIII make direct connections to the medial and lateral vestibular nuclei on the same side of the brain stem, as well as to the cerebellum (Figure 11.33). The vestibular nuclei also receive inputs from other parts of the brain, including the cerebellum, and the visual and somatic sensory systems, thereby combining incoming vestibular information with data about the motor system and other sensory modalities.

FIGURE 11.33 A summary of the central vestibular connections from one side. Description
The vestibular nuclei in turn project to a variety of targets above them in the brain stem, and below them into the spinal cord (see Figure 11.33). For example, axons from the otolith organs project to the lateral vestibular nucleus, which then projects via the vestibulospinal tract to excite spinal motor neurons controlling muscles in the legs that help maintain posture (see Chapter 14). This pathway helps the body stay upright even on the rolling deck of a boat. Axons from the semicircular canals project to the medial vestibular nucleus, which sends axons via the medial longitudinal fasciculus to excite motor neurons of trunk and neck muscles that orient the head. This pathway helps the head stay straight even as the body cavorts around below it.
Similar to the other sensory systems, the vestibular system makes connections to the thalamus and then to the neocortex. The vestibular nuclei send axons into the ventral posterior (VP) nucleus of the thalamus, which projects to regions close to the representation of the face in the primary somatosensory and primary motor areas of cortex (see Chapters 12 and 14). At the cortical level, there is considerable integration of information about movements of the body, the eyes, and the visual scene. It is likely that the cortex continually maintains a representation of body position and orientation in space, which is essential for our perception of equilibrium and for planning and executing complex, coordinated movements.
The Vestibulo-Ocular Reflex (VOR). One very important function of the central vestibular system is to keep your eyes pointed in a particular direction, even while you are dancing like a fool. The vestibulo-ocular reflex (VOR) performs this function. Recall that accurate vision requires the image to remain stable on the retinas despite movement of the head (see Chapter 9). Each eye can be moved by a set of six extraocular muscles. The VOR works by sensing rotations of the head and immediately commanding a compensatory movement of the eyes in the opposite direction. The movement helps keep your line of sight tightly fixed on a visual target. Because the VOR is a reflex triggered by vestibular input rather than visual input, it works amazingly well even in the dark or when your eyes are closed.
Imagine driving down a very bumpy road. From constant adjustments by the VOR, your view of the world ahead is quite stable because each bump, and its consequent movement of your head, is compensated by an eye movement. To appreciate how effective your VOR is, compare the stability of a passing object during the bumpy drive as you look at it first with your eyes alone, and then with the viewfinder of a simple camera. You’ll find that your camera view jumps around hopelessly because your arms are not nearly quick or accurate enough to move the camera with each bump. Many cameras now have an electromechanical equivalent of a VOR that stabilizes an image even when the camera or the photographer holding it is bumping around.
The effectiveness of the VOR depends on complicated connections from the semicircular canals to the vestibular nucleus to the cranial nerve nuclei that excite the extraocular muscles. Figure 11.34, which shows only half of the horizontal component of this circuit, illustrates what happens when the head turns to the left and the VOR induces both eyes to turn right. Axons from the left horizontal canal innervate the left vestibular nucleus, which sends excitatory axons to the contralateral (right) cranial nerve VI nucleus (abducens nucleus). Motor axons from the abducens nucleus in turn excite the lateral rectus muscle of the right eye. Another excitatory projection from the abducens crosses the midline, back to the left side, and ascends (via the medial longitudinal fasciculus) to excite the left cranial nerve III nucleus (oculomotor nucleus), which excites the medial rectus muscle of the left eye.

FIGURE 11.34 Vestibular connections mediating horizontal eye movements during the VOR. These pathways are active when the head suddenly turns to the left, causing the eyes to turn to the right. Excitatory connections are denoted by green plus signs; the inhibitory connection is denoted by a red minus sign. Description
Mission accomplished, so it would seem: Both eyes are turning right. However, to further ensure speedy operation, the left medial rectus muscle also gets excited via a projection from the vestibular nucleus directly to the left oculomotor nucleus. Speed is also maximized by activating inhibitory connections to the muscles that oppose this movement (the left eye’s lateral rectus and right eye’s medial rectus, in this case). In order to respond to head rotations in any direction, the complete VOR circuit includes similar connections between the right horizontal canal, the other semicircular canals, and the other extraocular muscles that control eye movements.
The vestibular system can be damaged in a variety of ways; for example, high doses of antibiotics such as streptomycin can be toxic to hair cells. People with bilateral lesions of the vestibular labyrinths have enormous trouble fixating on visual targets as they move about. Even the minute head pulsations from the blood pressure surges of heartbeats can be disturbing in some cases. When people with vestibular disturbances cannot stabilize an image on their moving retinas, they may also experience the disconcerting feeling that the world is constantly moving around them. The sensation can make walking and standing difficult. Compensatory adjustments come with time, as the brain learns to substitute more visual and proprioceptive cues to help guide smooth and accurate movements.
Hearing and balance begin with nearly identical sensory receptors, the hair cells, which are exquisitely sensitive to deflections of their stereocilia. These movement detectors are surrounded by three sets of inner ear structures that give them selectivity for three different kinds of mechanical energy: periodic waves of air pressure (sound), rotational forces (head turns), and linear forces (head tilt or acceleration). Except for the similarity in transduction, and the fact that the hair cells of both systems are located in the inner ear, the auditory and vestibular systems are quite different. The sound sensed by audition comes mainly from the external environment, while the vestibular system senses only the movements of itself. Auditory and vestibular pathways are entirely separate except perhaps at the highest levels of the cortex. Auditory information is often at the forefront of our consciousness, while vestibular sensation usually operates unnoticed to coordinate and calibrate our every movement.
We have followed the auditory pathways from the ear to cerebral cortex and seen the ways in which information about sound is transformed. Variations in the density of air are converted to movements of the mechanical components of the middle and inner ear, which are transduced into neural responses. The structures of the ear and cochlea are highly specialized for the transduction of sound. However, this fact should not blind us to the considerable similarities between the organization of the auditory system and that of other sensory systems. Many analogies can be made between the auditory and visual systems. In the sensory receptors of both systems, a spatial code is established. In the visual system, the code in the photoreceptors is retinotopic; the activity of a given photoreceptor indicates light at a particular location. The receptors in the auditory system establish a spatial code that is tonotopic because of the unique properties of the cochlea. In each system, the retinotopy or tonotopy is preserved as signals are processed in secondary neurons, the thalamus, and finally in sensory cortex.
The convergence of inputs from lower levels produces neurons at higher levels that have more complex response properties. Combinations of LGN inputs give rise to simple and complex receptive fields in visual cortex; similarly in the auditory system, the integration of inputs tuned to different sound frequencies yields higher level neurons that respond to complex combinations of frequencies. Another example of increasing visual complexity is the convergence of inputs from the two eyes, which yields binocular neurons that are important for depth perception. Analogously, in the auditory system, input from the two ears is combined to create binaural neurons, which are used for horizontal sound localization. These are just a few of the many similarities in the two systems. Principles governing one system can often help us understand other systems. Keep this in mind while reading about the somatic sensory system in the next chapter, and you’ll be able to predict some features of cortical organization based on the types of sensory receptors.
1. How is the conduction of sound to the cochlea facilitated by the ossicles of the middle ear?
2. Why is the round window crucial for the function of the cochlea? What would happen to hearing if it suddenly didn’t exist?
3. Why is it impossible to predict the frequency of a sound simply by looking at which portion of the basilar membrane is the most deformed?
4. Why would the transduction process in hair cells fail if the stereocilia as well as the hair cell bodies were surrounded by perilymph?
5. If inner hair cells are primarily responsible for hearing, what is the function of outer hair cells?
6. Why doesn’t unilateral damage to the inferior colliculus or MGN lead to deafness in one ear?
7. What mechanisms function to localize sounds in the horizontal and vertical planes?
8. What symptoms would you expect to see in a person who had recently had a stroke affecting A1 unilaterally? How does the severity of these symptoms compare with the effects of a unilateral stroke involving V1?
9. What is the difference between nerve deafness and conduction deafness?
10. Each macula contains hair cells with kinocilia arranged in all directions. What is the advantage of this arrangement compared to an arrangement of all the cells in the same direction?
11. Imagine a semicircular canal rotating in two different ways: around its axis (like a rolling coin) or end over end (like a flipped coin). How well would its hair cells respond in each case, and why?
12. How would you expect the functions of the otolith organs and the semicircular canals to change in the weightless environment of space?
Ashida G, Carr CE. 2011. Sound localization: Jeffress and beyond. Current Opinion in Neurobiology 21:745–751.
Cullen KE. 2012. The vestibular system: multimodal integration and encoding of self-motion for motor control. Trends in Neurosciences 35:185–196.
Guinan JJ Jr, Salt A, Cheatham MA. 2012. Progress in cochlear physiology after Békésy. Hearing Research 293:12–20.
Holt JR, Pan B, Koussa MA, Asai Y. 2014. TMC function in hair cell transduction. Hearing Research 311:17–24.
Kazmierczak P, Müller U. 2012. Sensing sound: molecules that orchestrate mechanotransduction by hair cells. Trends in Neurosciences 35:220–229.
Oertel D, Doupe AJ. 2013. The auditory central nervous system. In Principles of Neural Science, 5th ed., ed. Kandel ER, Schwartz JH, Jessell TM, Siegelbaum SA, Hudspeth AJ. New York: McGraw-Hill Companies, Inc., 682–711.
Additional figures