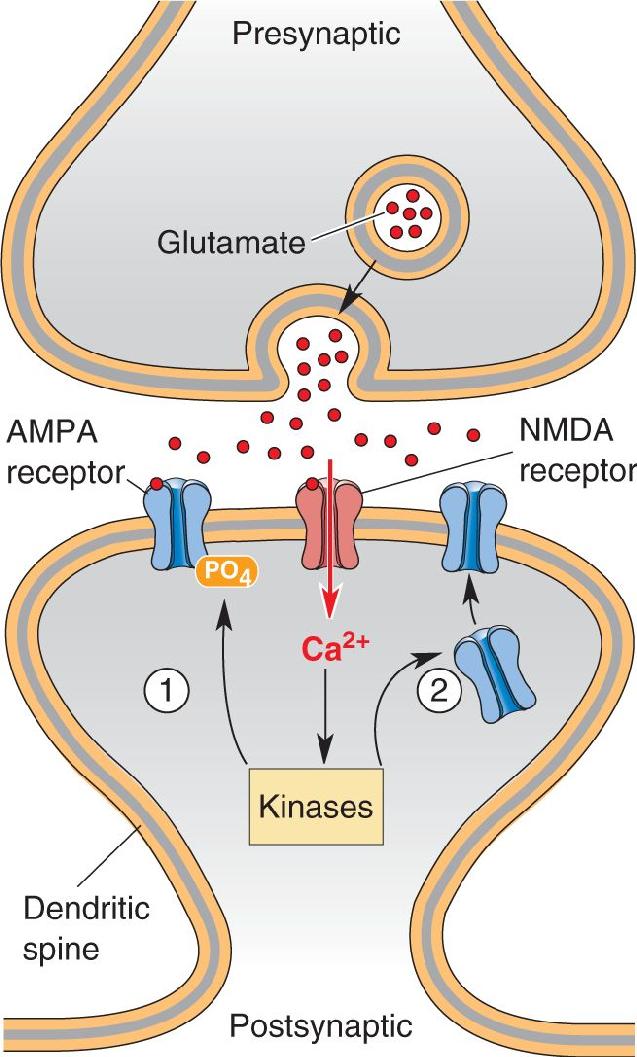
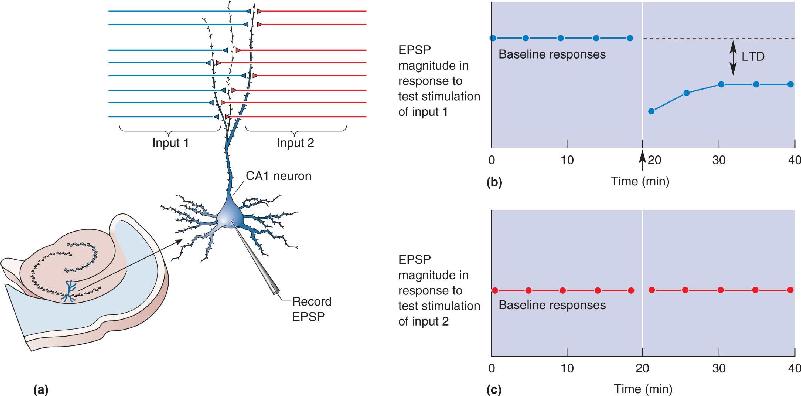
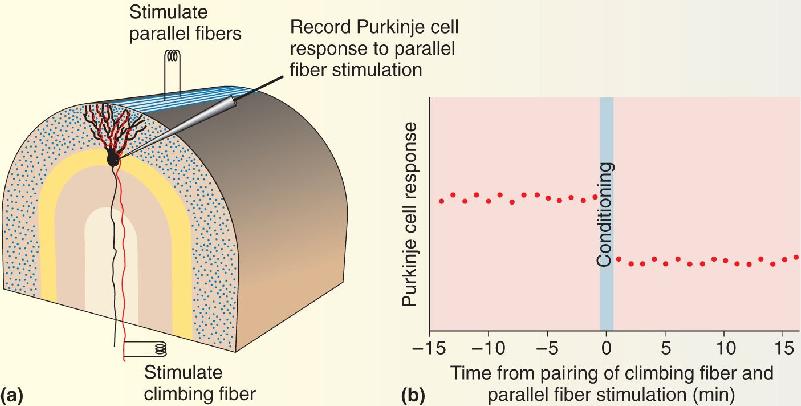
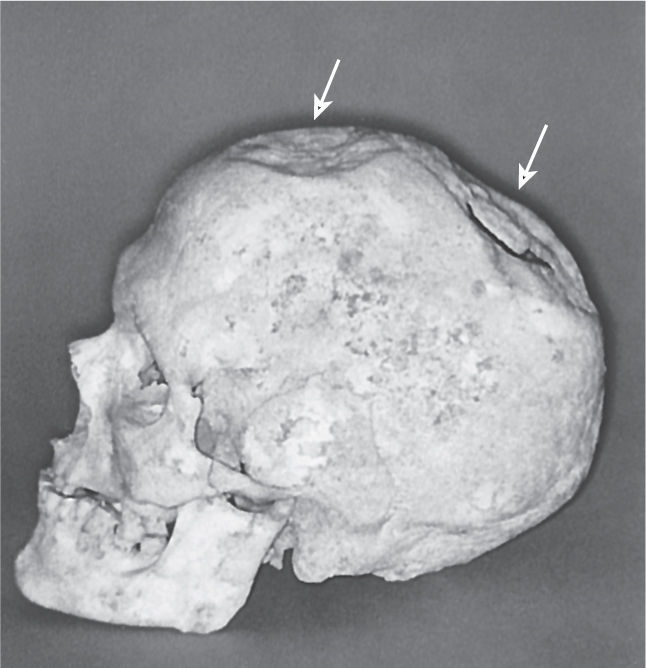
Ch 1 Neuroscience: Past, Present, and FutureCh 2 Neurons and GliaCh 3 The Neuronal Membrane at RestCh 4 The Action PotentialCh 5 Synaptic TransmissionCh 6 Neurotransmitter SystemsCh 7 The Structure of the Nervous SystemCh 8 The Chemical SensesCh 9 The EyeCh 10 The Central Visual SystemCh 11 The Auditory and Vestibular SystemsCh 12 The Somatic Sensory SystemCh 13 Spinal Control of MovementCh 14 Brain Control of MovementCh 15 Chemical Control of the Brain and BehaviorCh 16 MotivationCh 17 Sex and the BrainCh 18 Brain Mechanisms of EmotionCh 19 Brain Rhythms and SleepCh 20 LanguageCh 21 -One The Resting Brain, Attention, and ConsciousnessCh 22 -Two Mental IllnessCh 23 -Three Wiring the BrainCh 24 -Four Memory SystemsCh 25 -five Molecular Mechanisms of Learning and Memory
Ch 1Neuroscience: Past, Present, and FutureRead full chapter →
Sections in this chapter
- Neuroscience: Past, Present, and Future
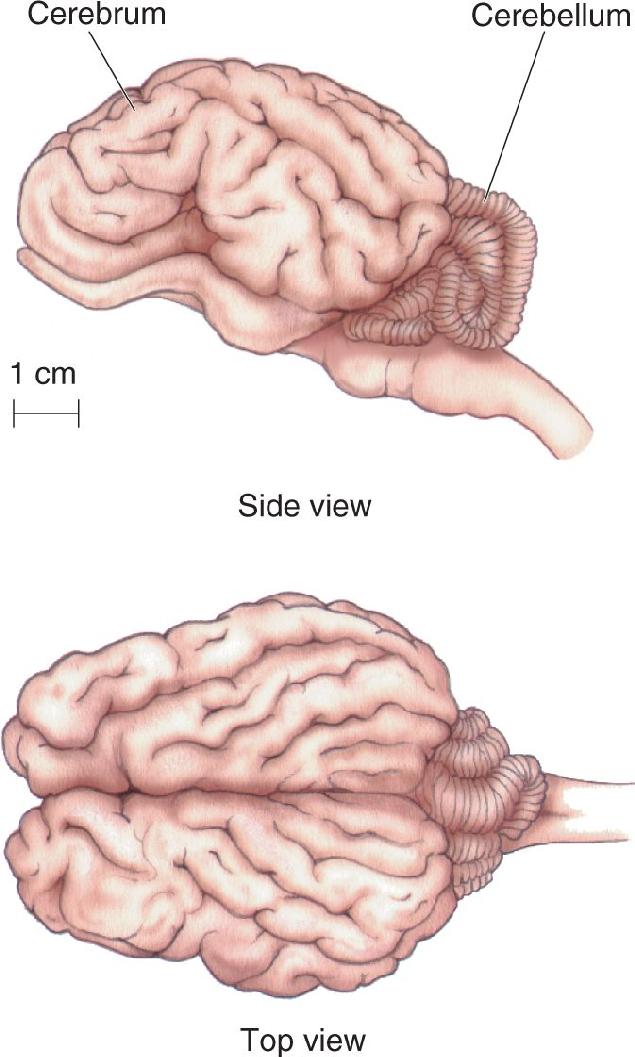
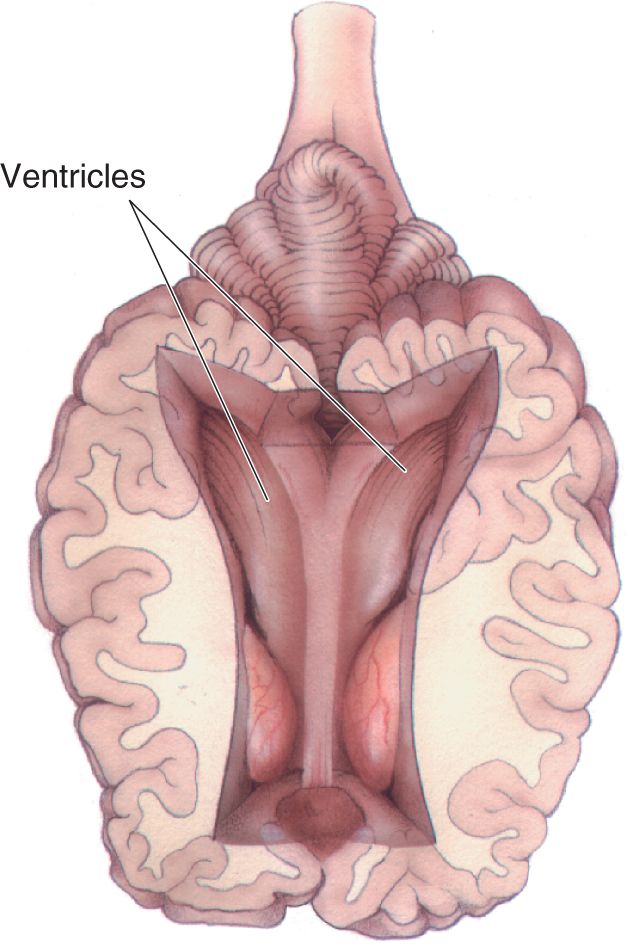
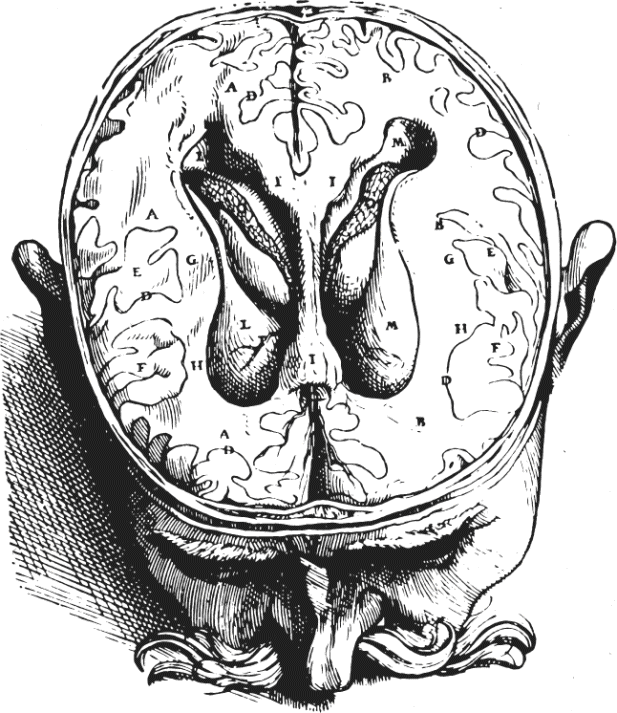
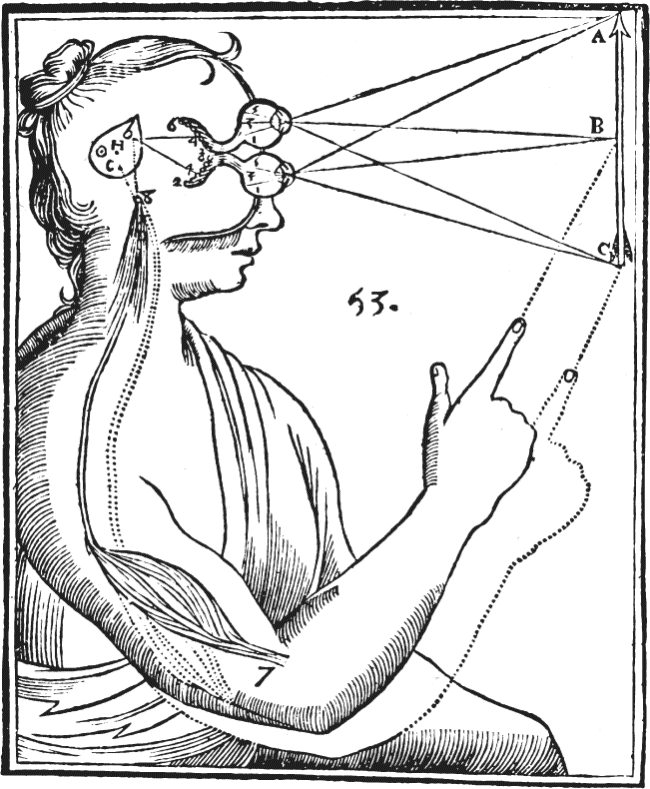
Ch 2Neurons and GliaRead full chapter →
Sections in this chapter
- Neurons and Glia
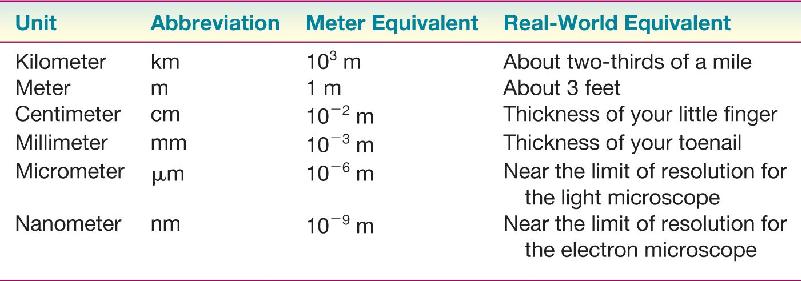


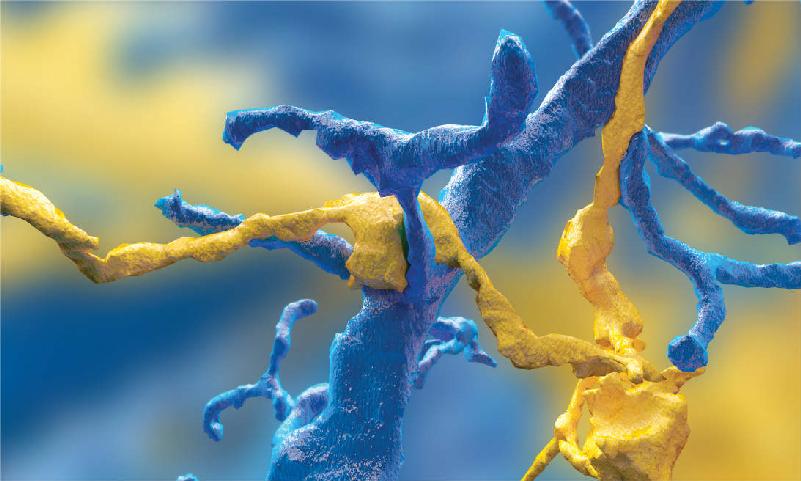
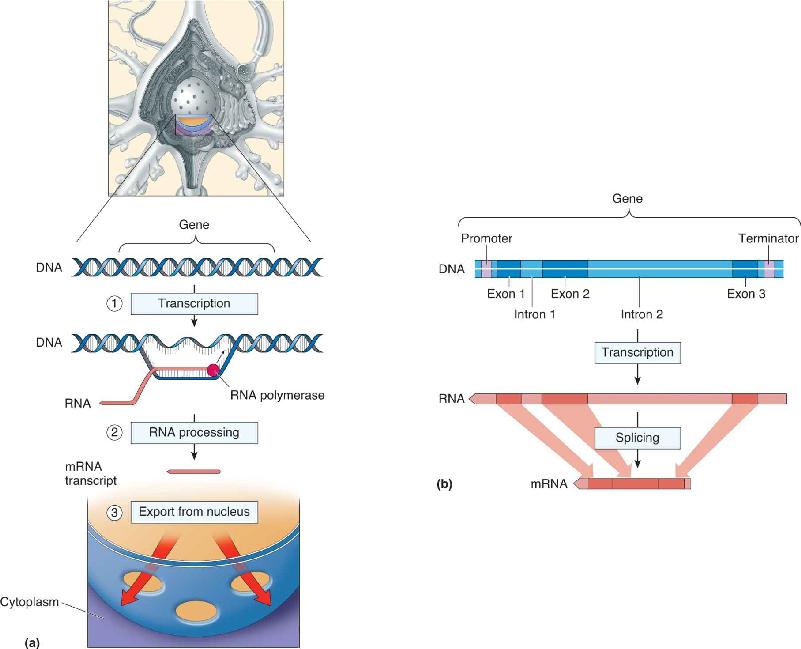
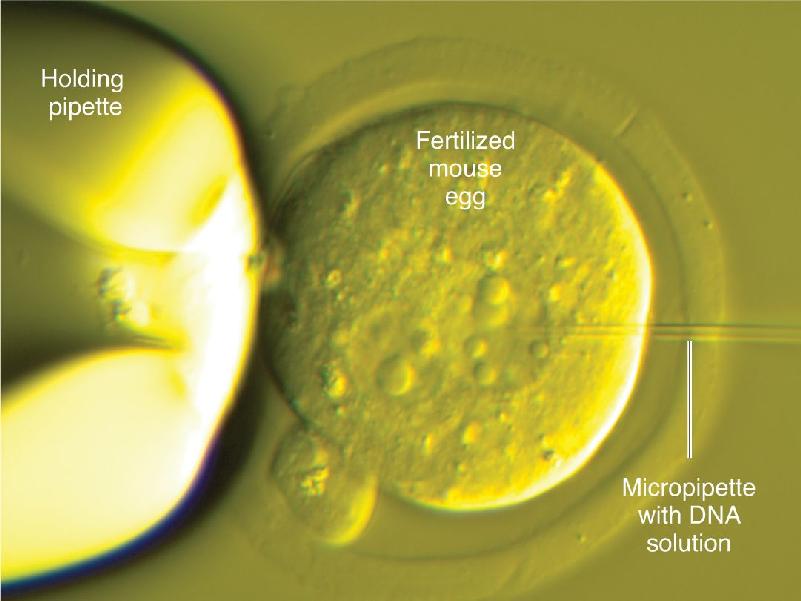
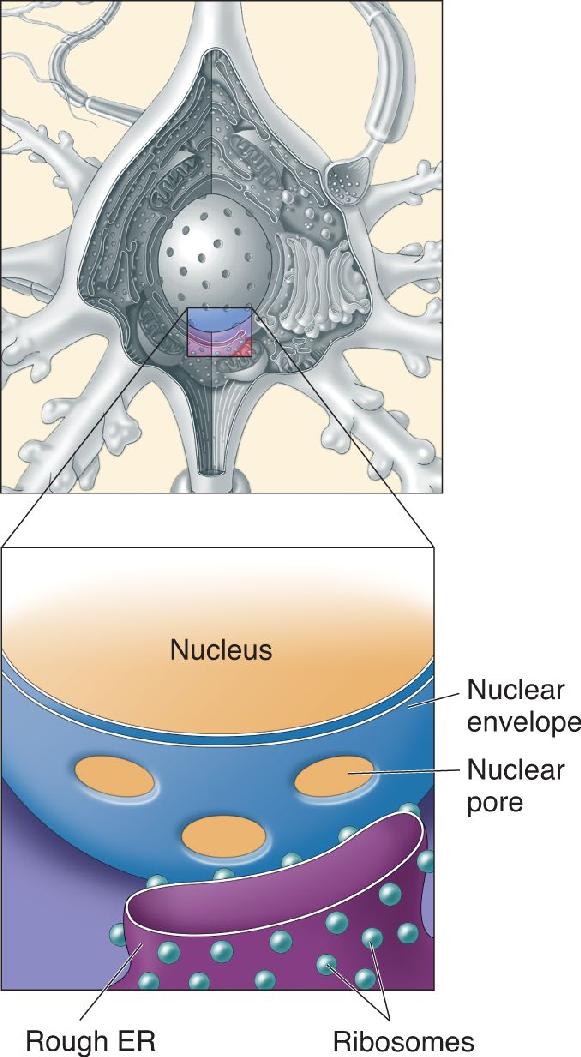
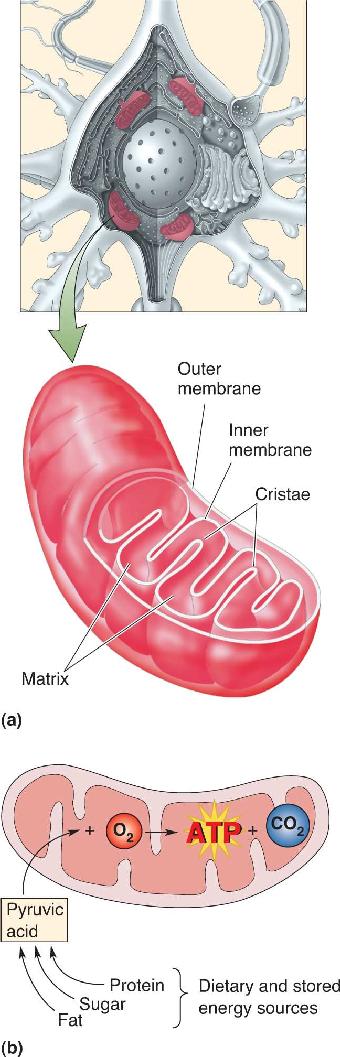
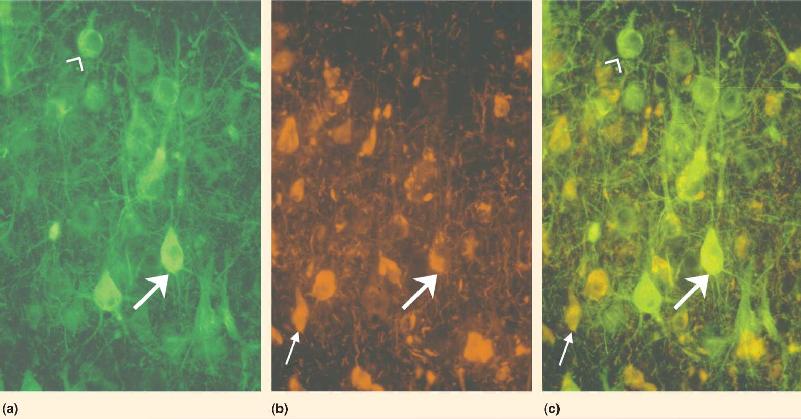
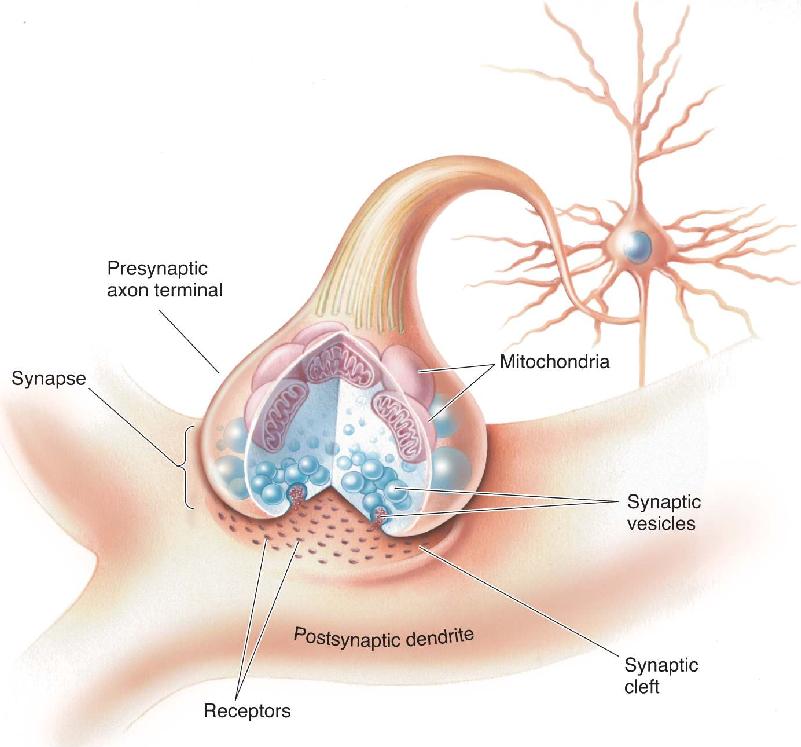
Ch 3The Neuronal Membrane at RestRead full chapter →
Sections in this chapter
- The Neuronal Membrane at Rest
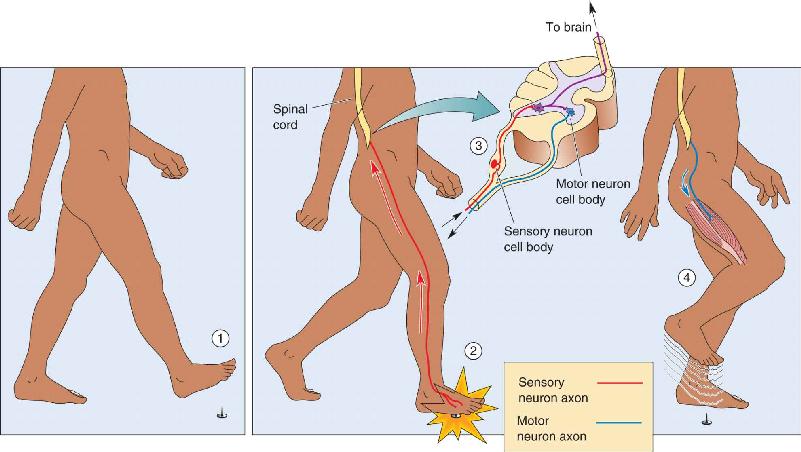
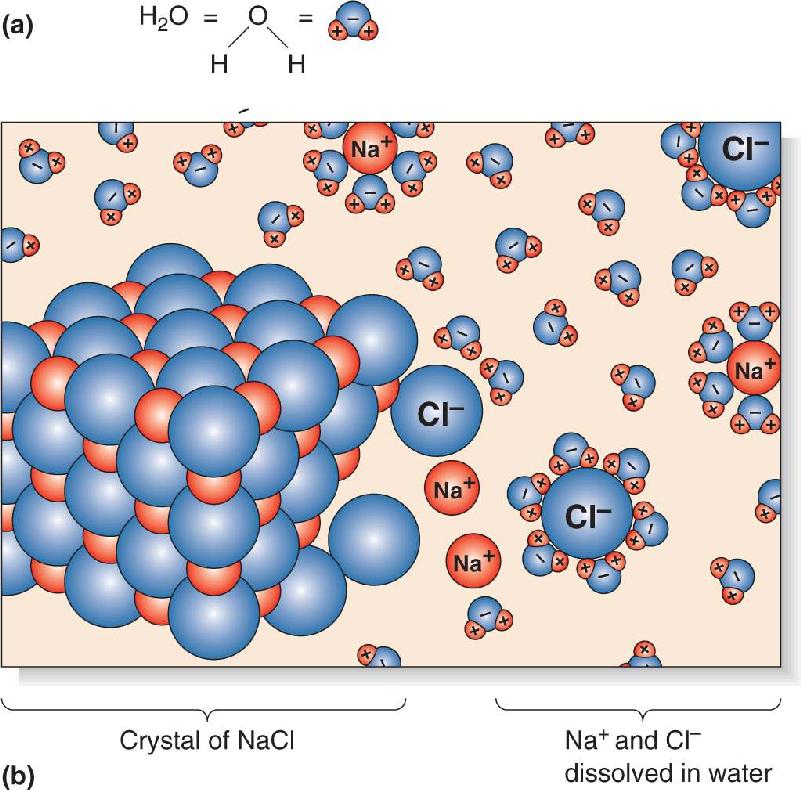
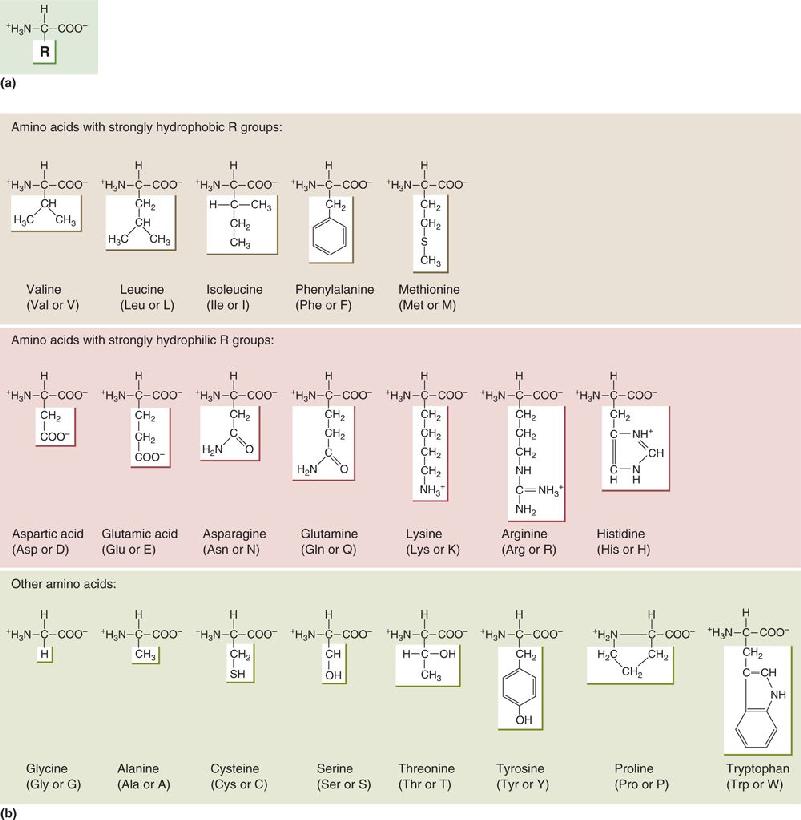
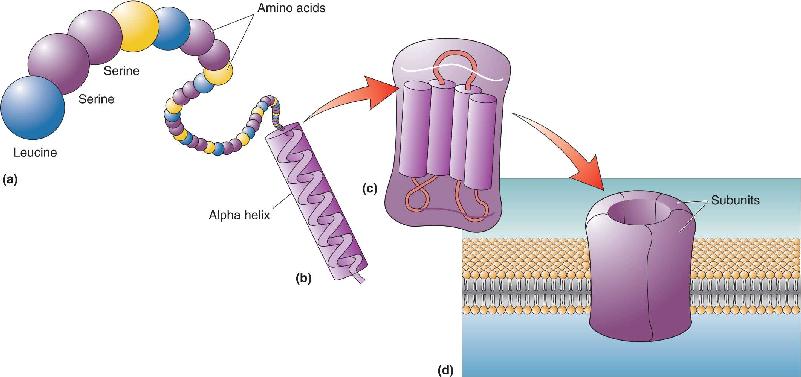
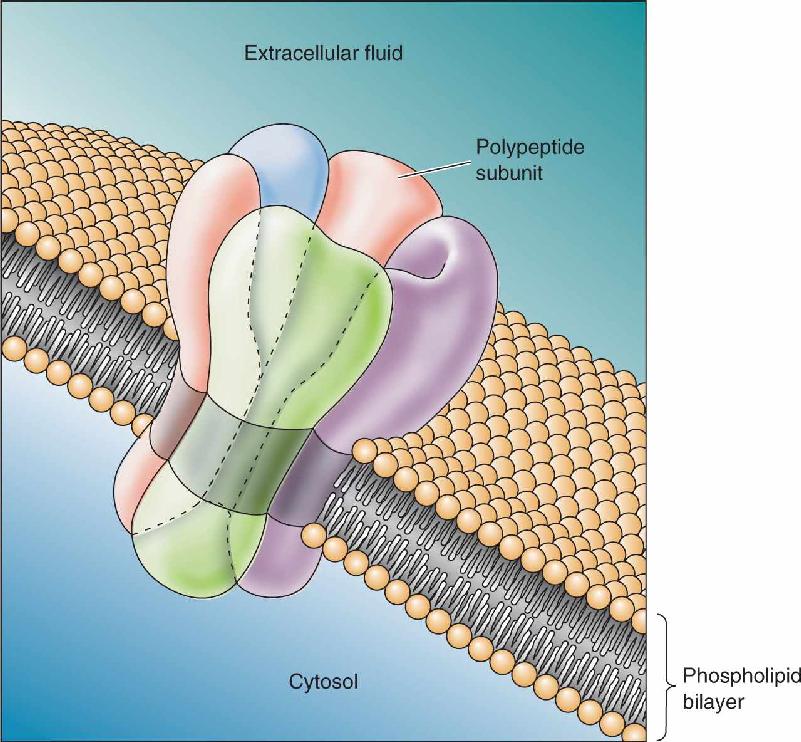
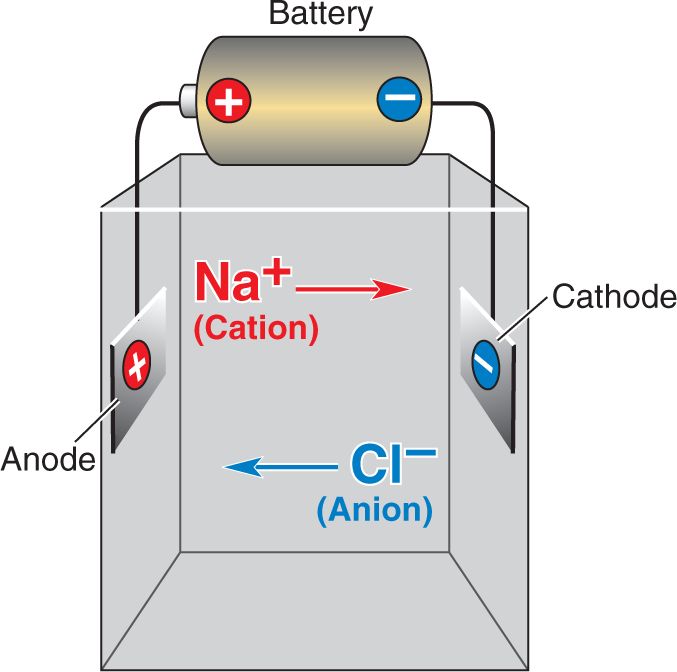
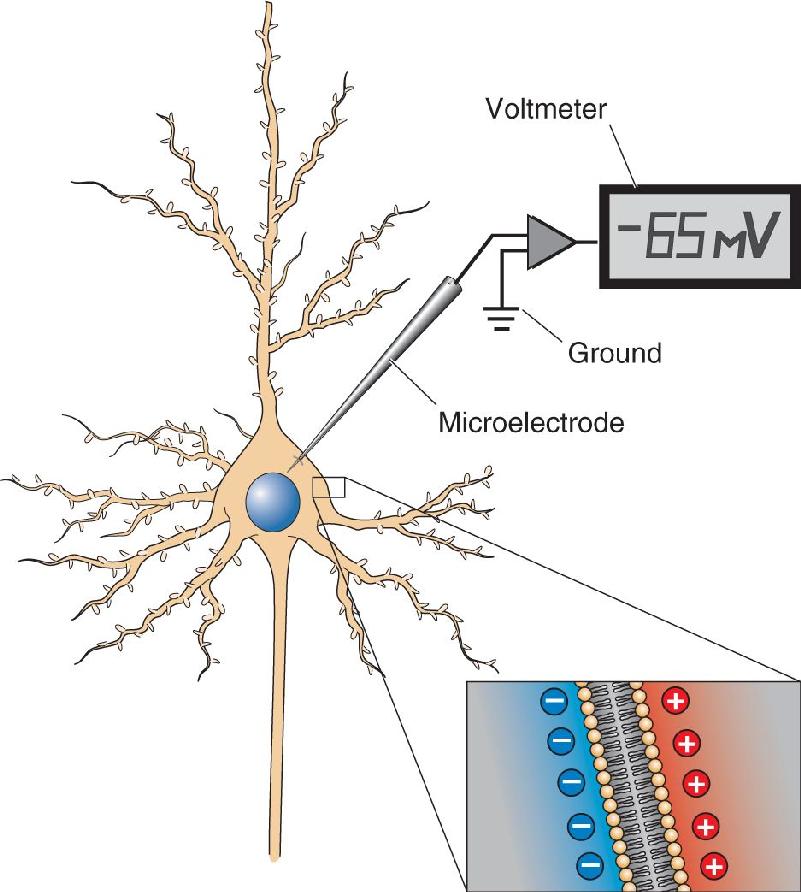
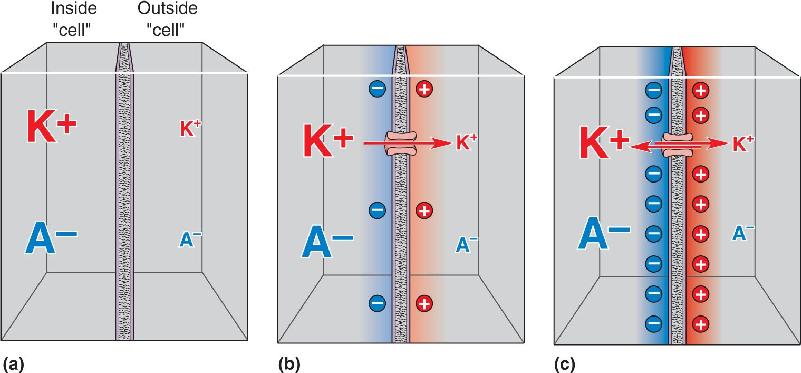
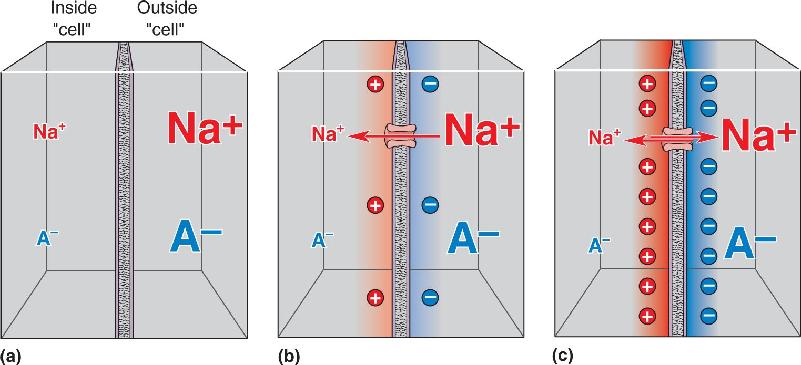
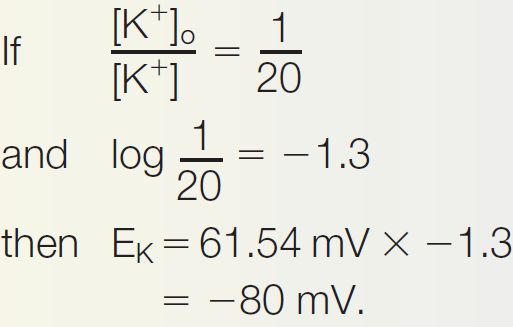
Ch 4The Action PotentialRead full chapter →
Sections in this chapter
- The Action Potential
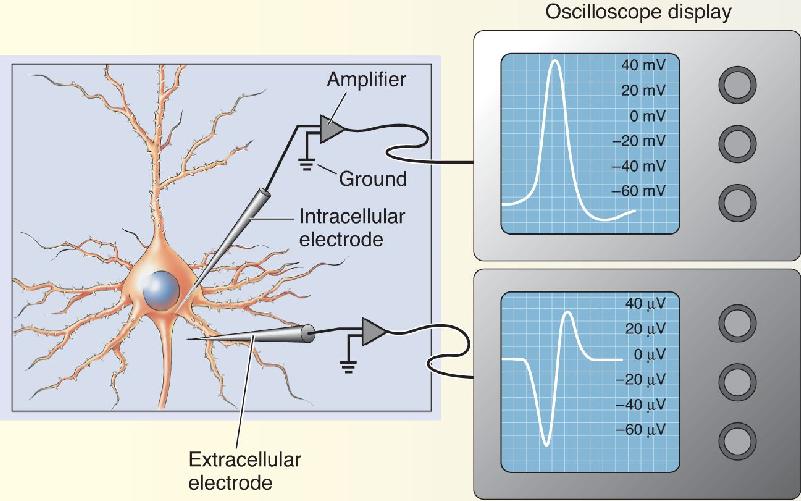
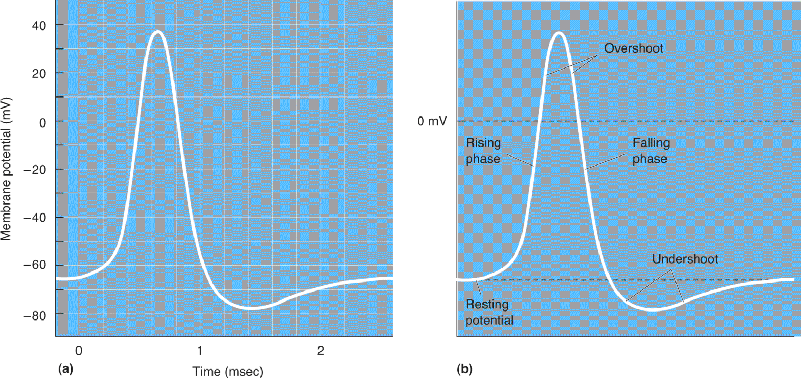
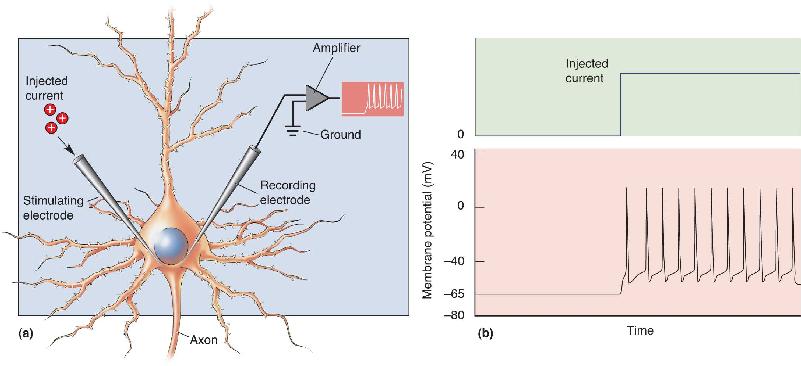

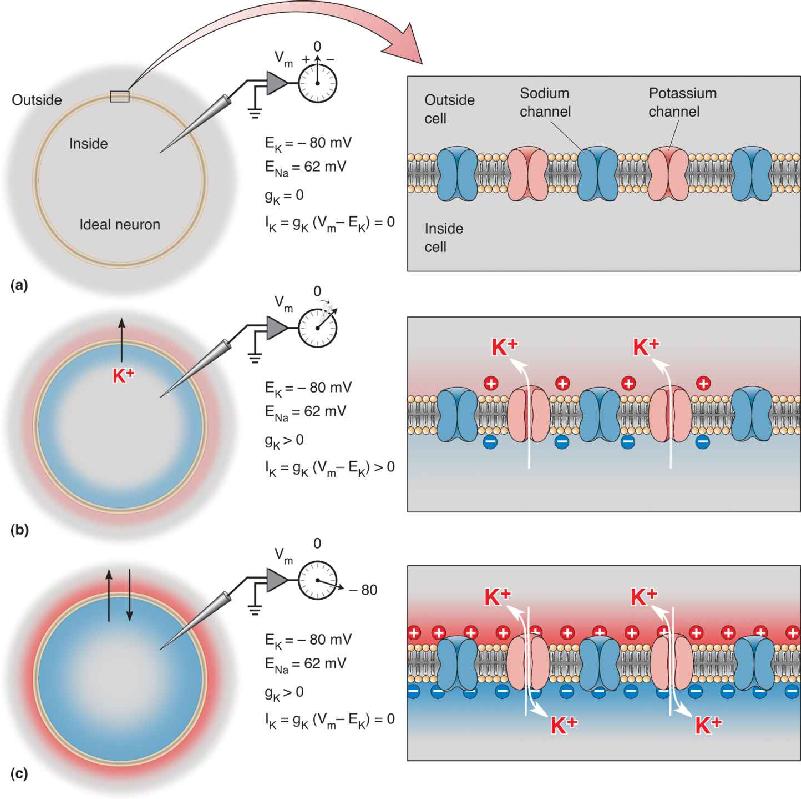
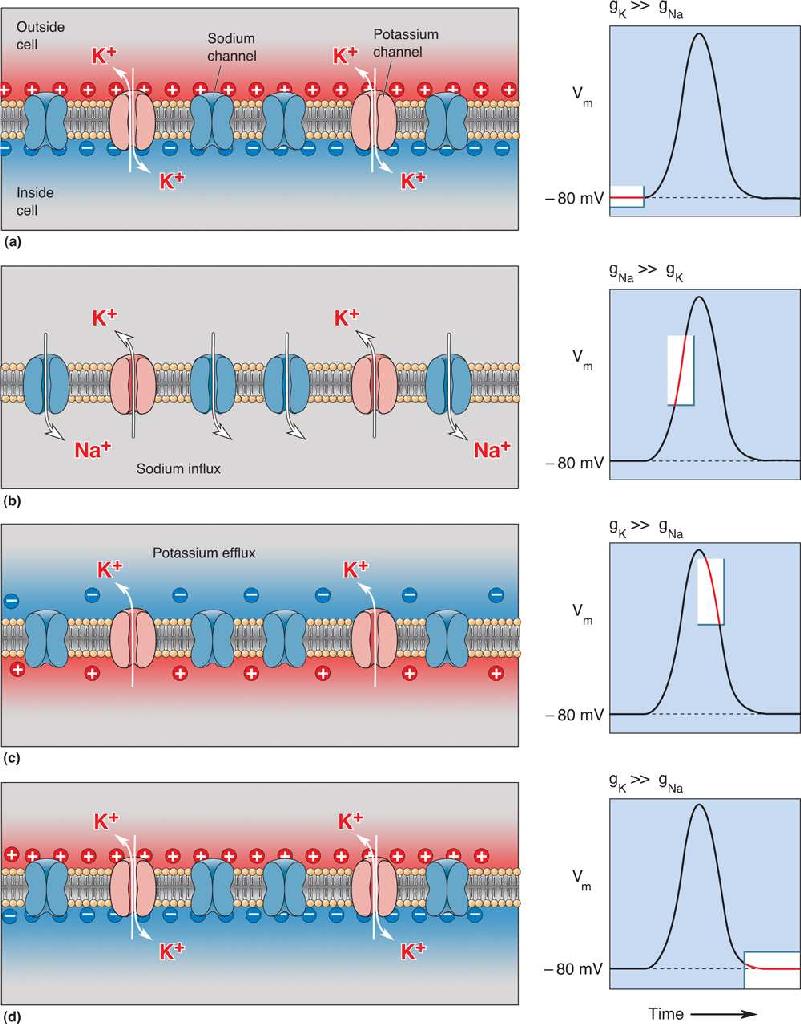
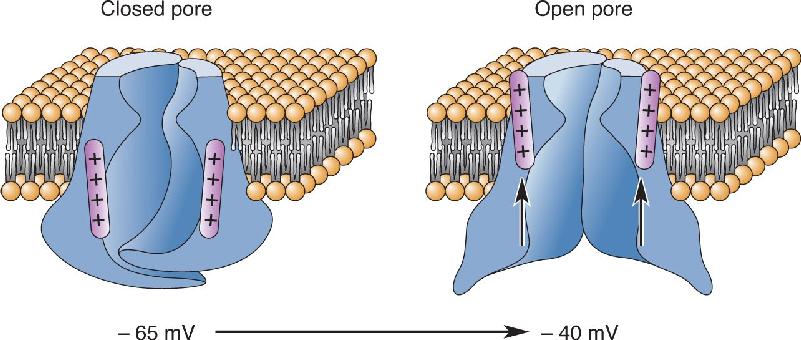
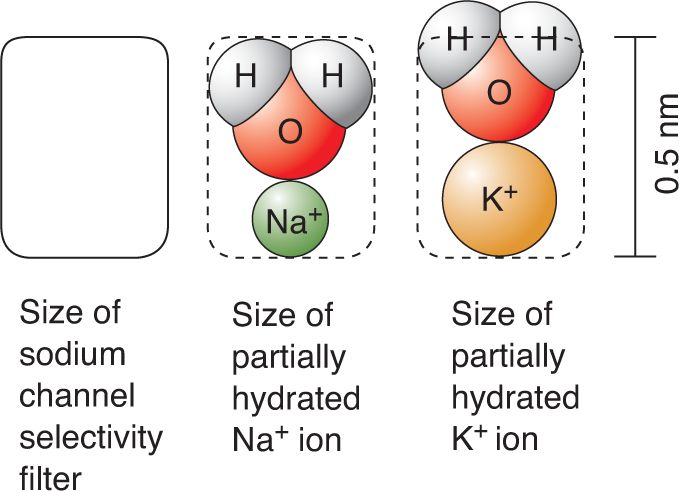
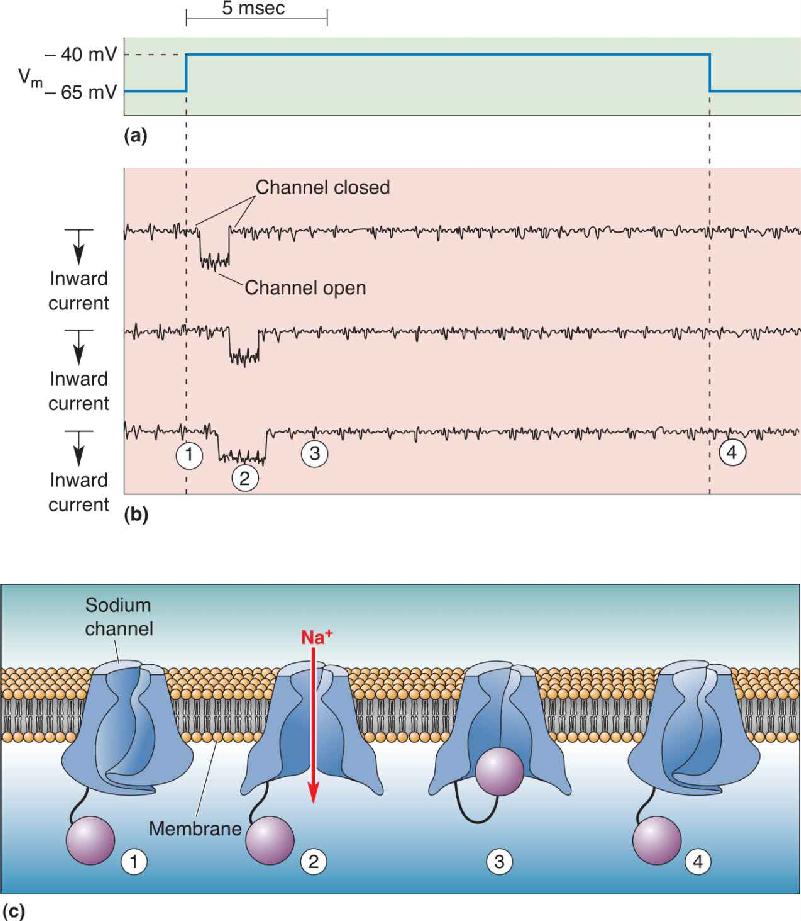
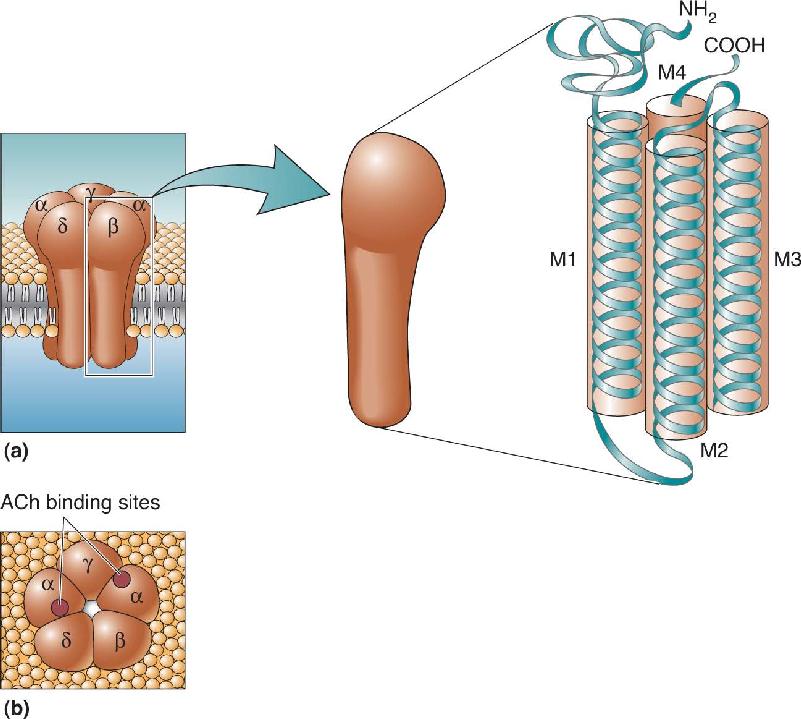
Ch 5Synaptic TransmissionRead full chapter →
Sections in this chapter
- Synaptic Transmission
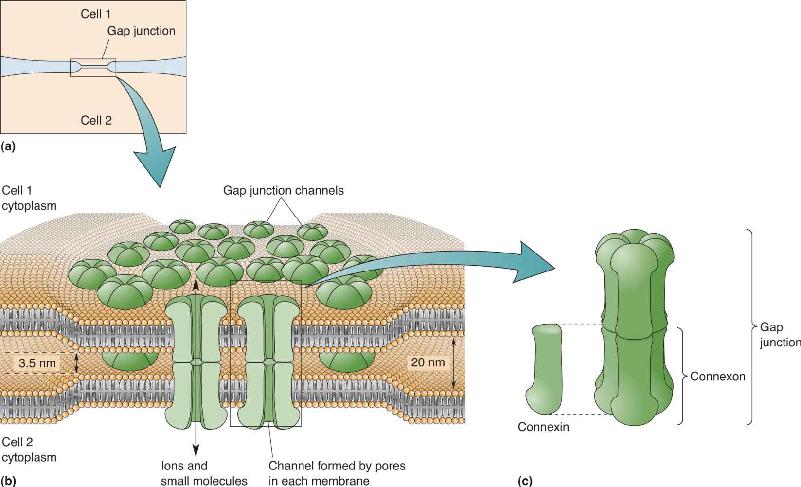
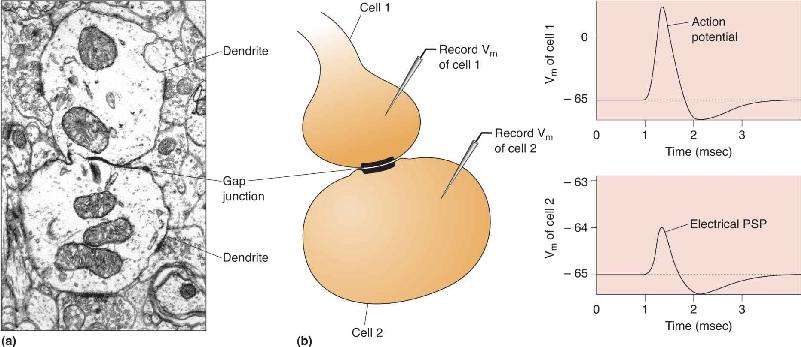
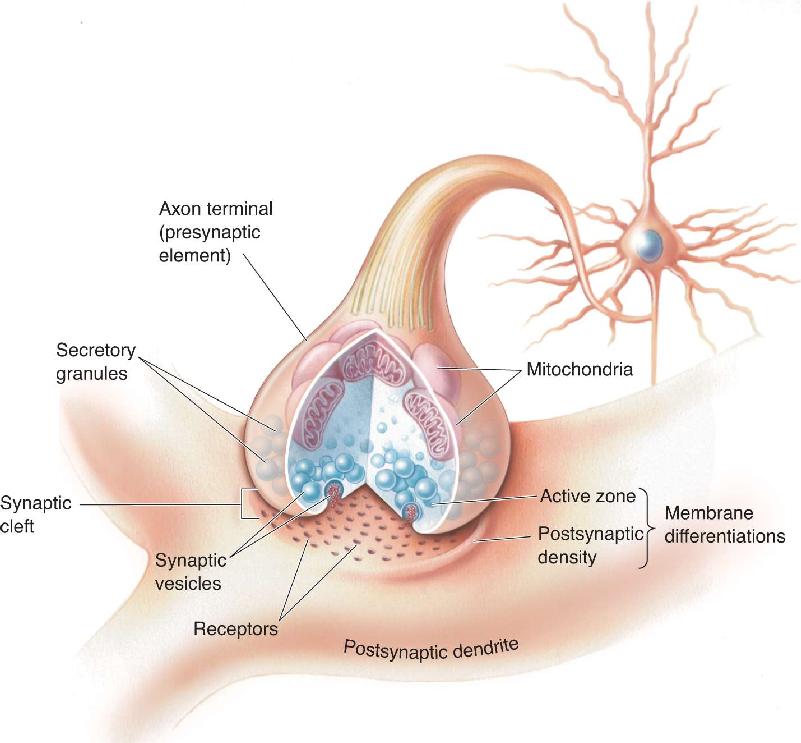
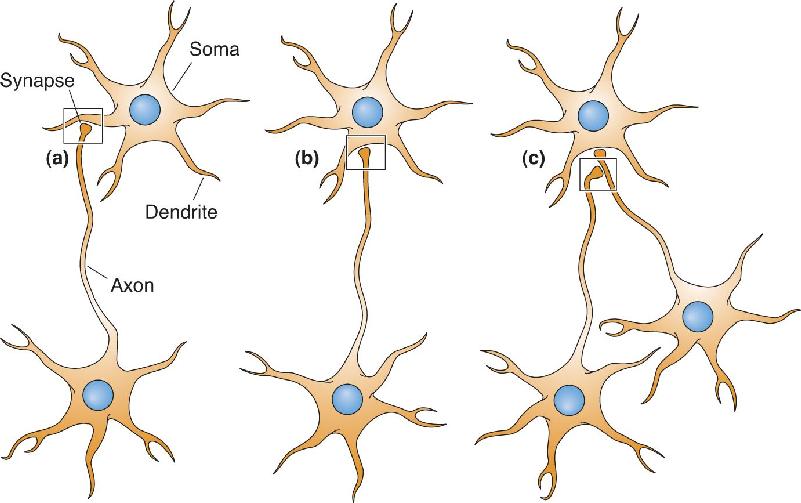

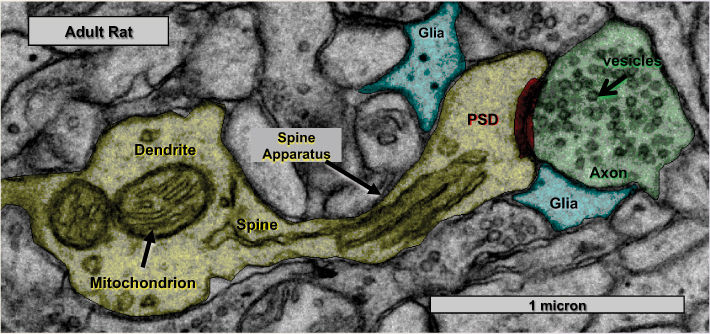
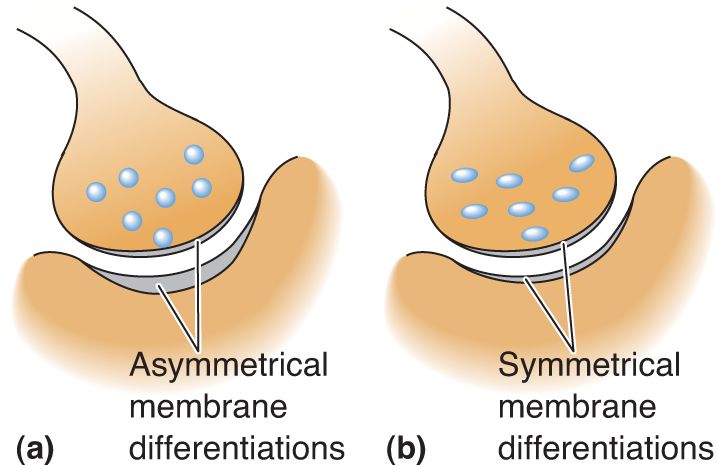
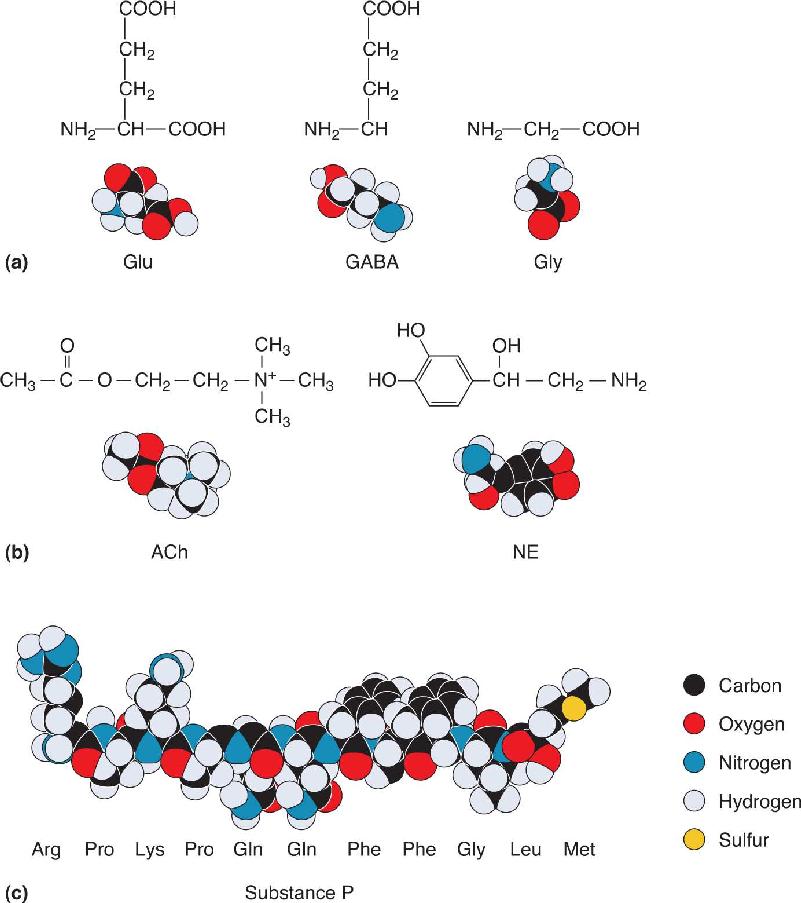
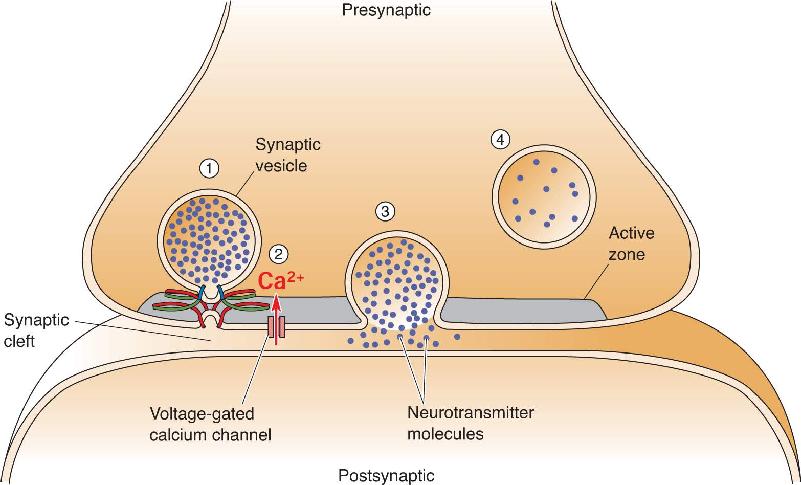
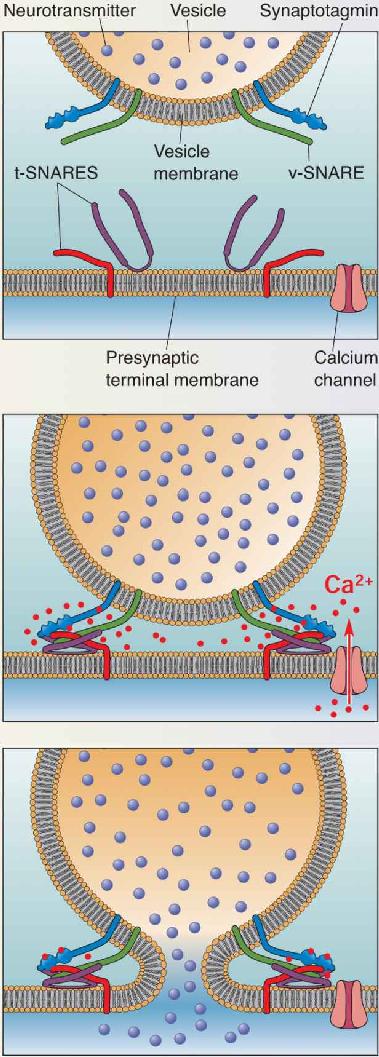
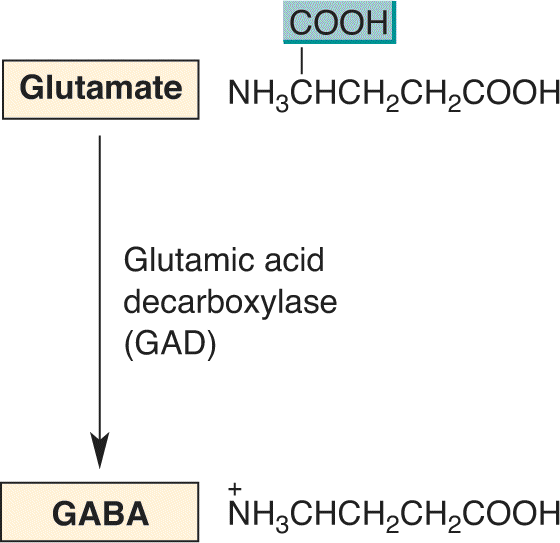
Ch 6Neurotransmitter SystemsRead full chapter →
Sections in this chapter
- Neurotransmitter Systems
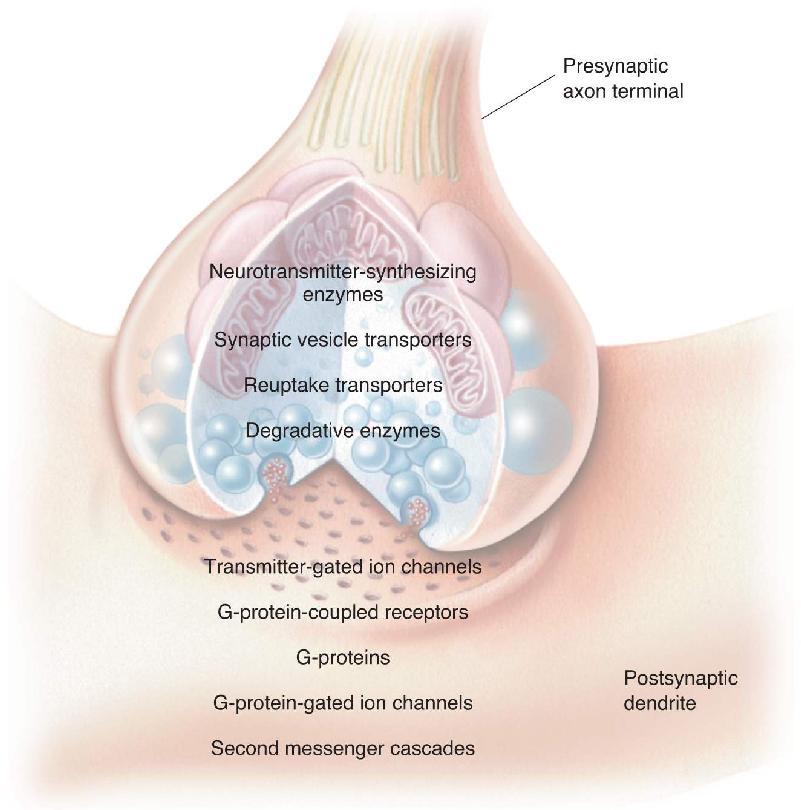
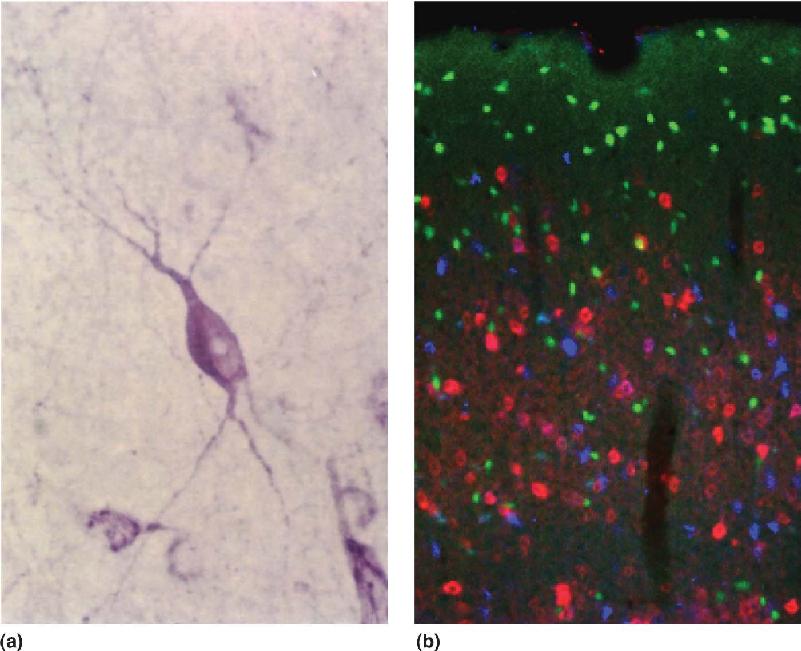
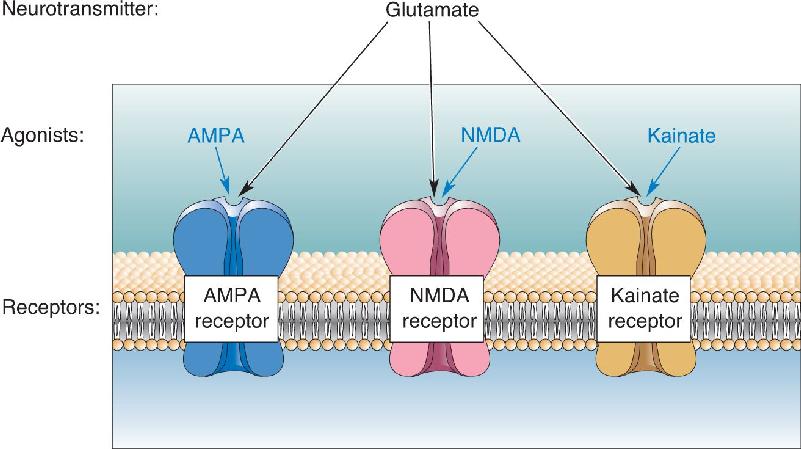

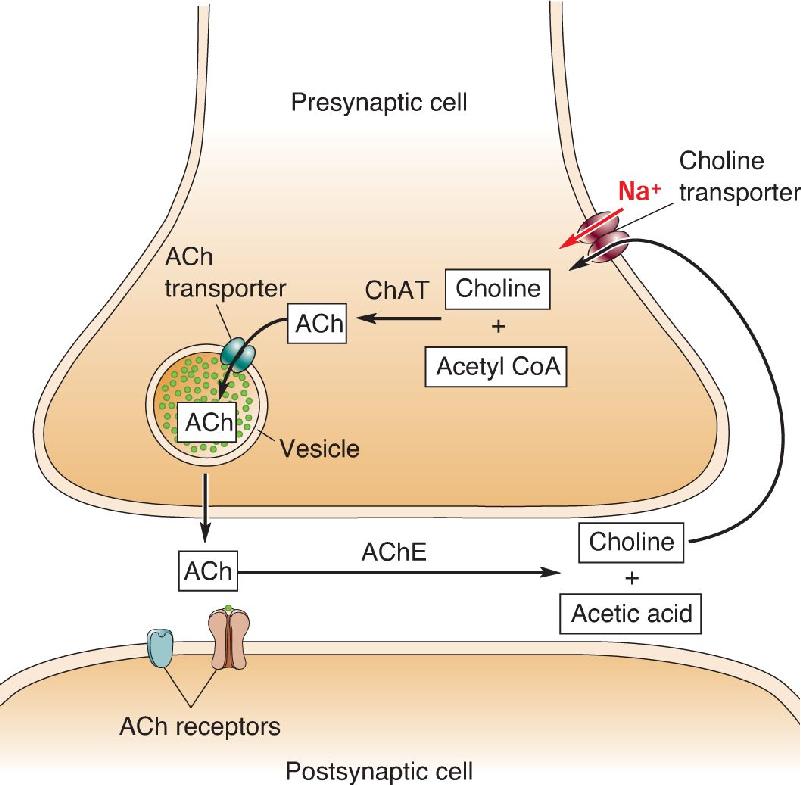
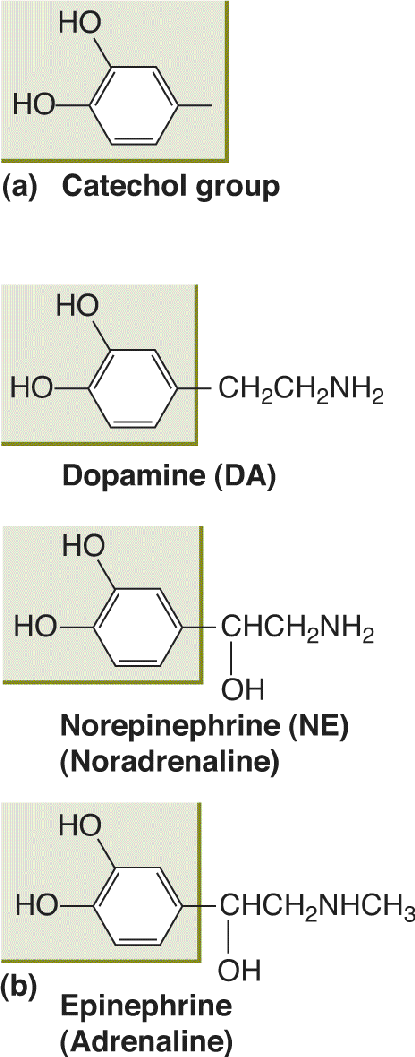
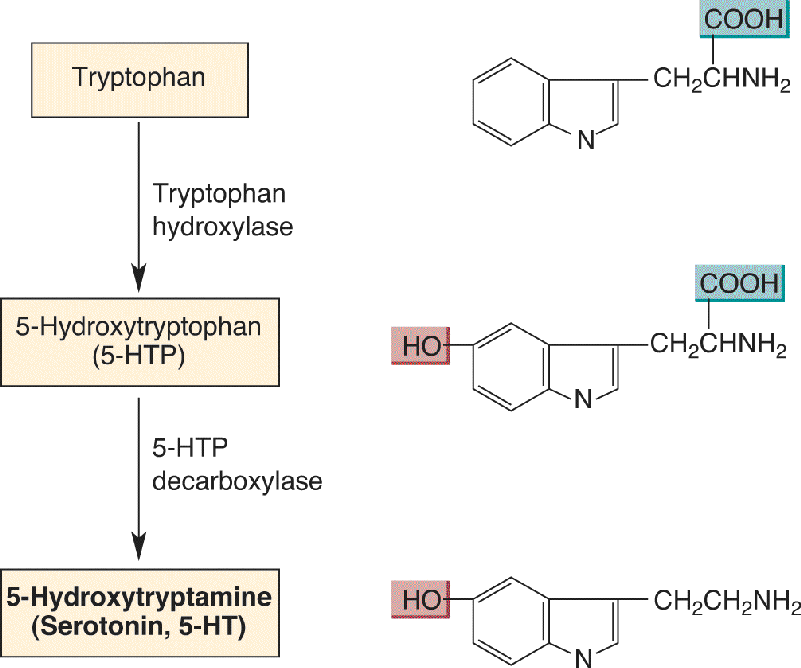
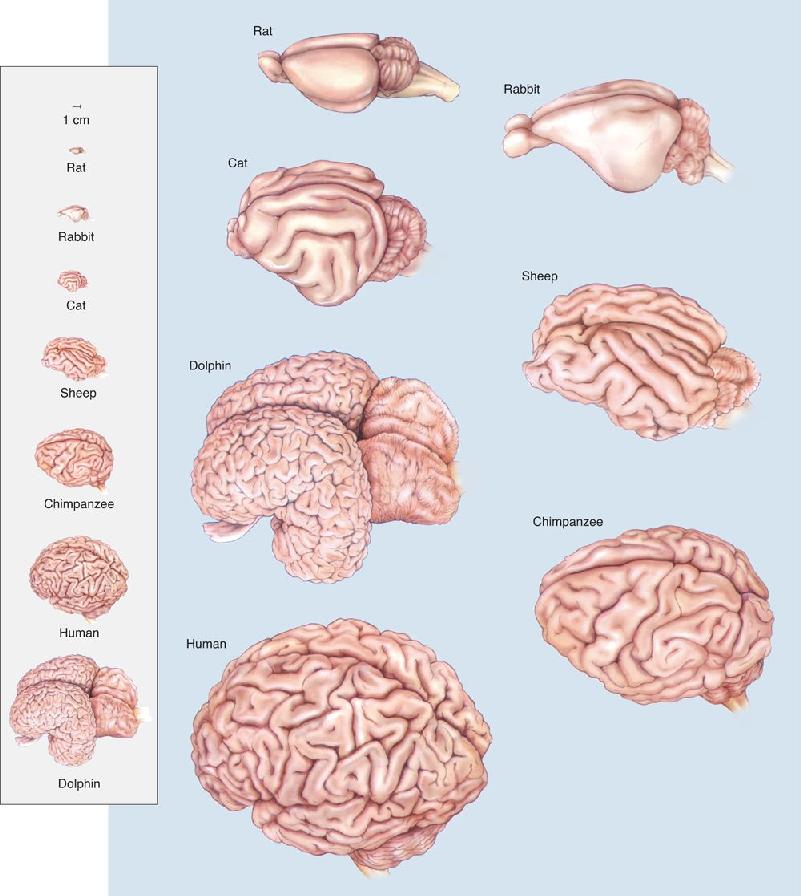
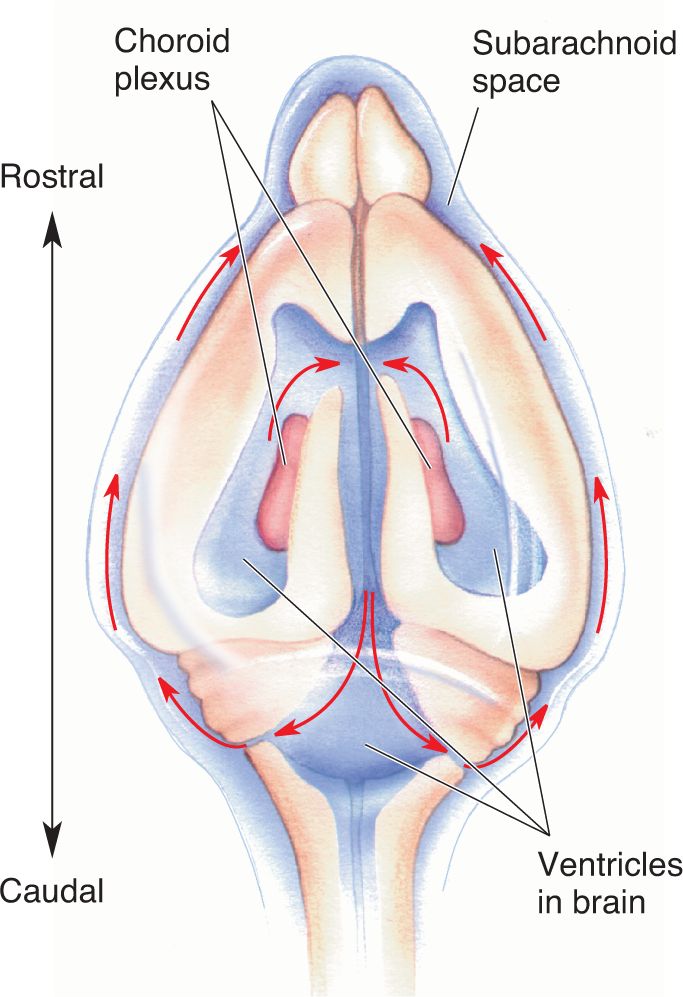
Ch 7The Structure of the Nervous SystemRead full chapter →
Sections in this chapter
- The Structure of the Nervous System
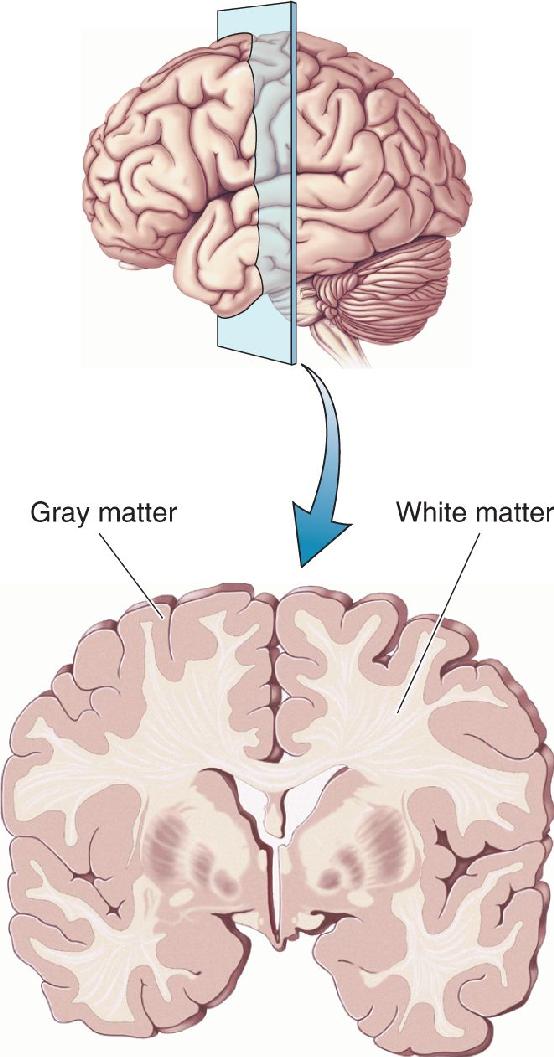
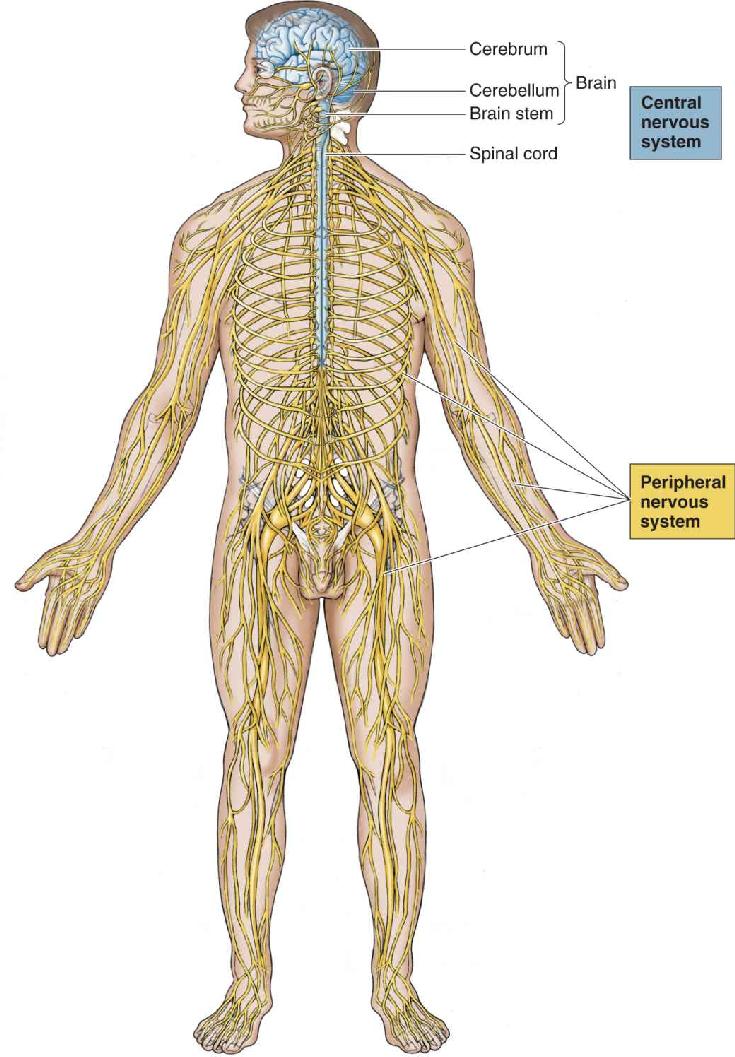
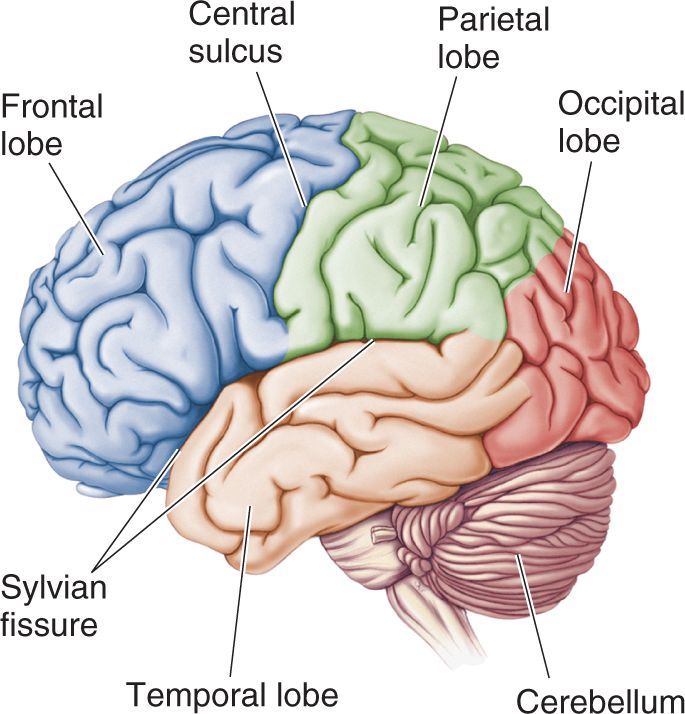
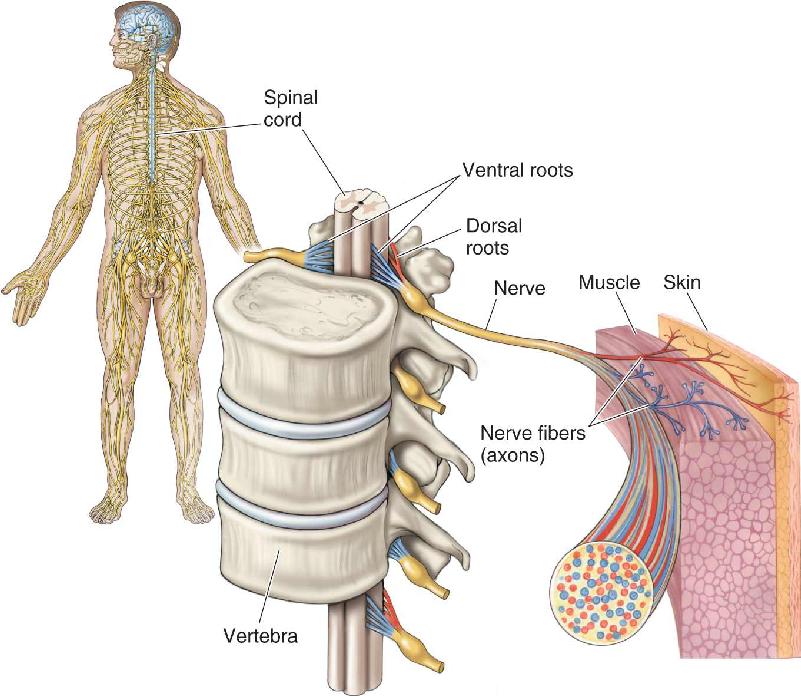
- An Illustrated Guide to Human Neuroanatomy
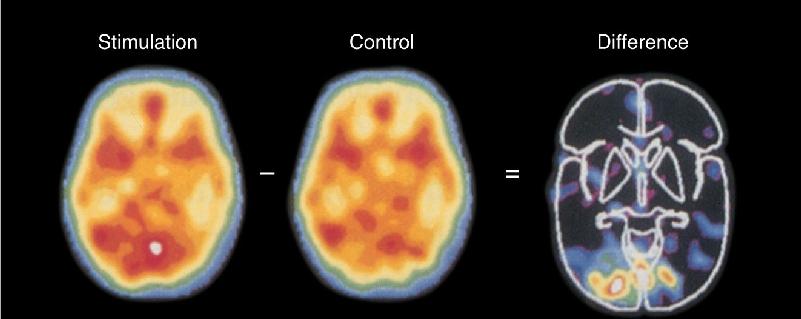
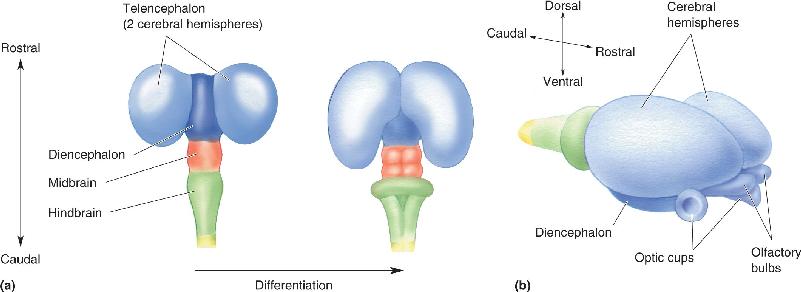
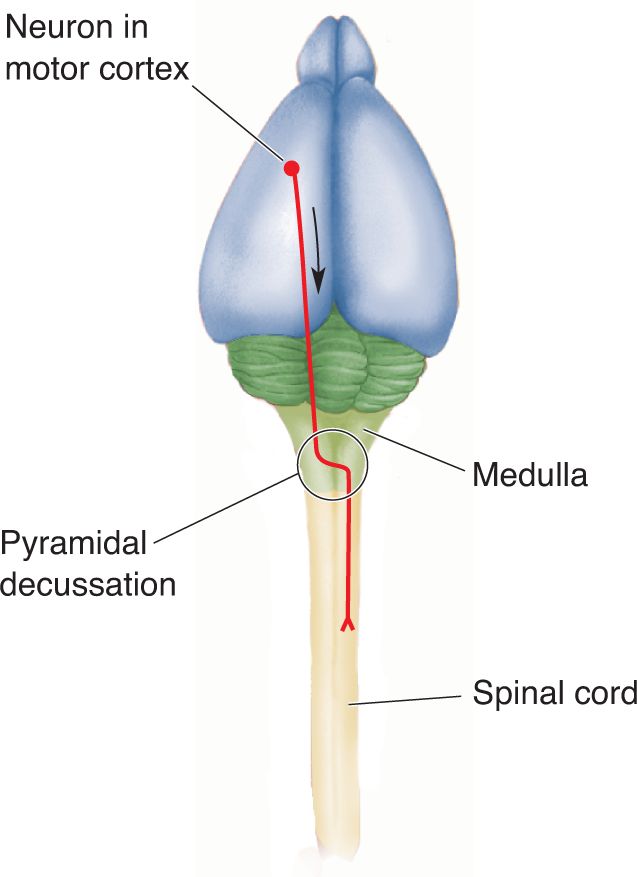
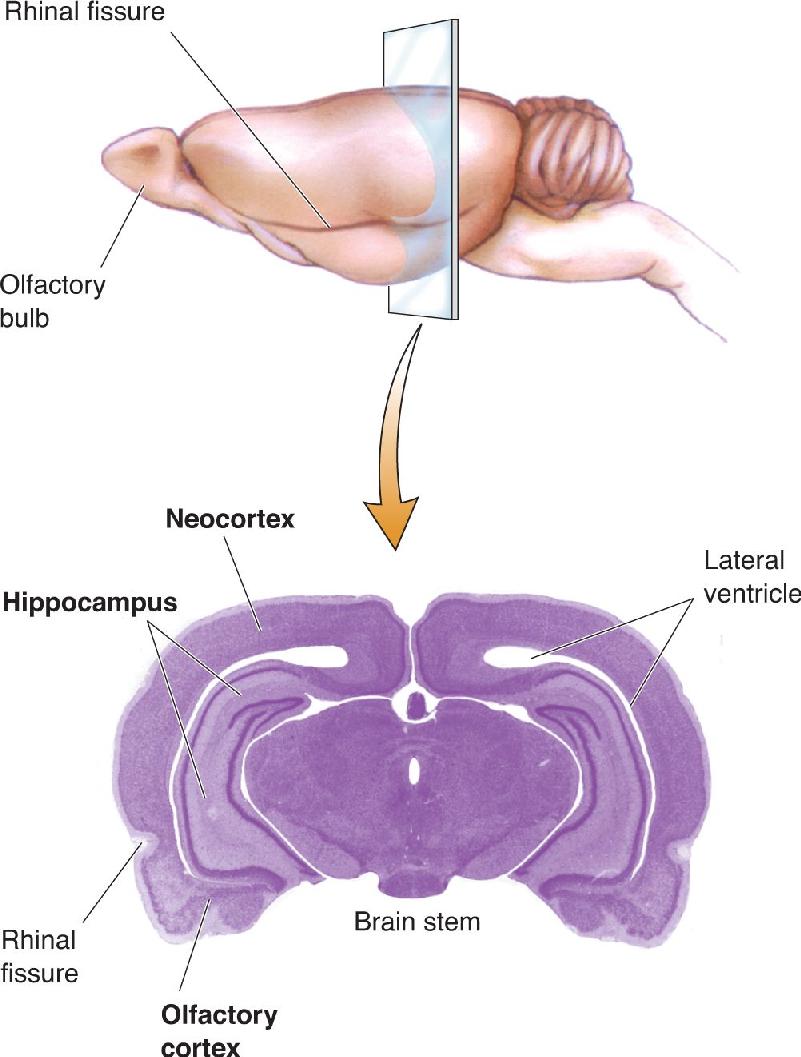
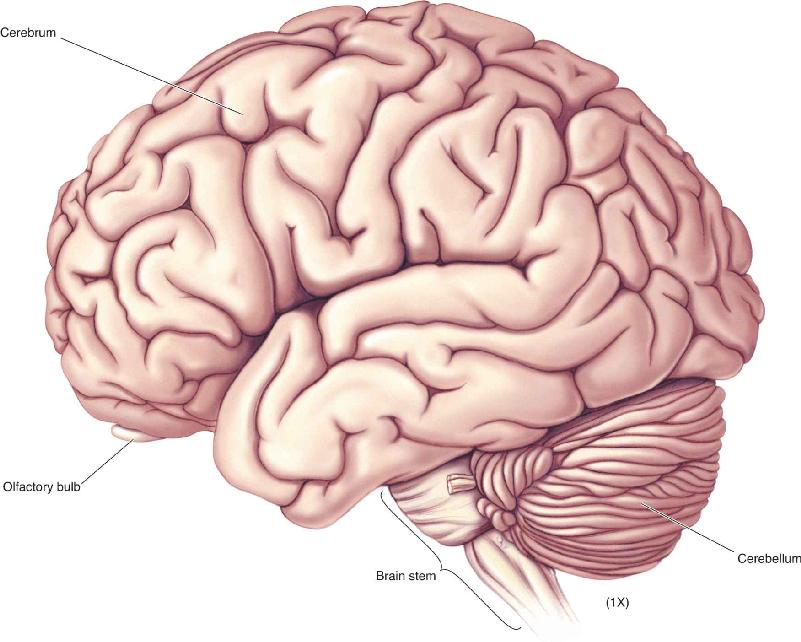
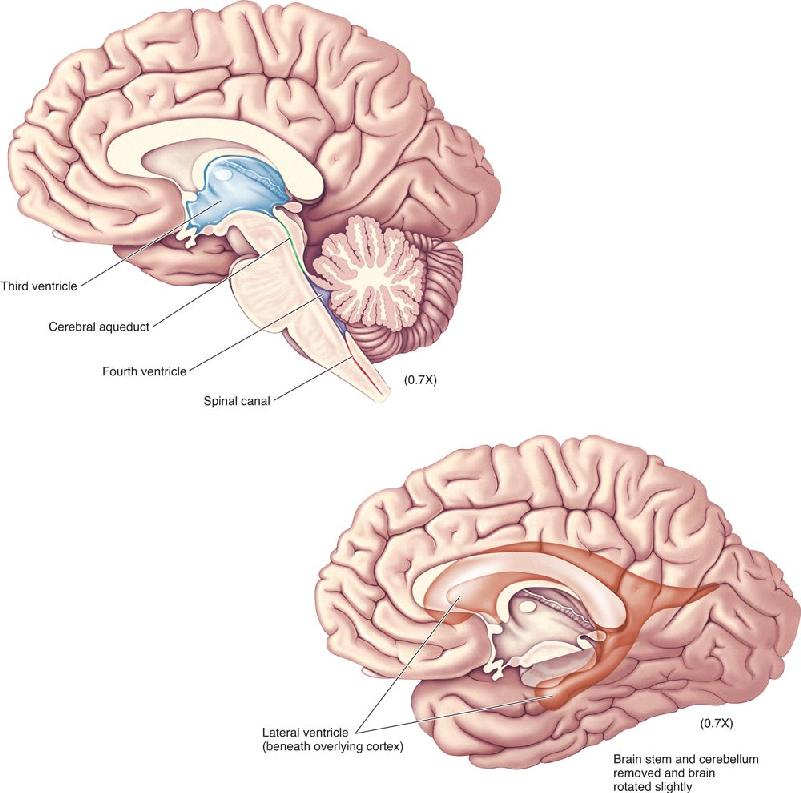
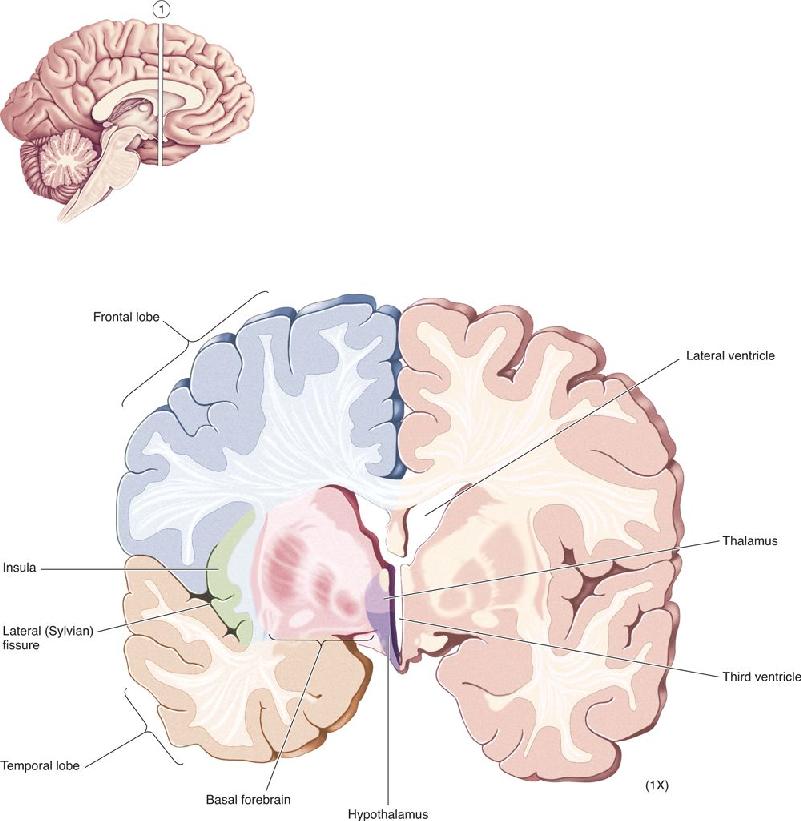
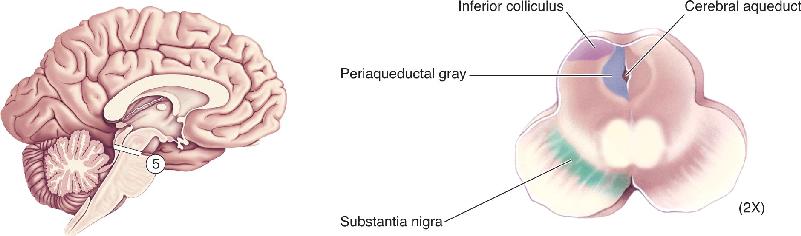
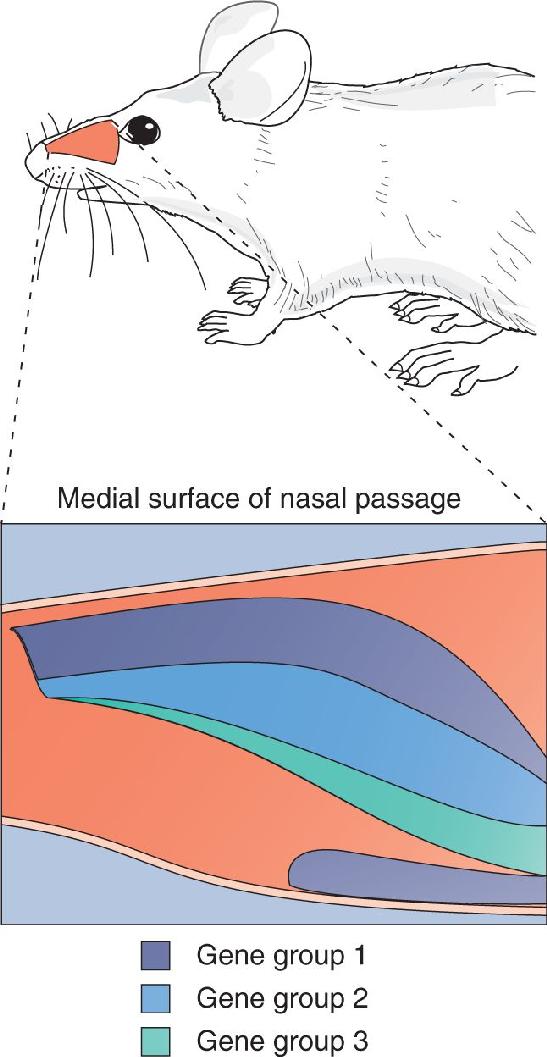
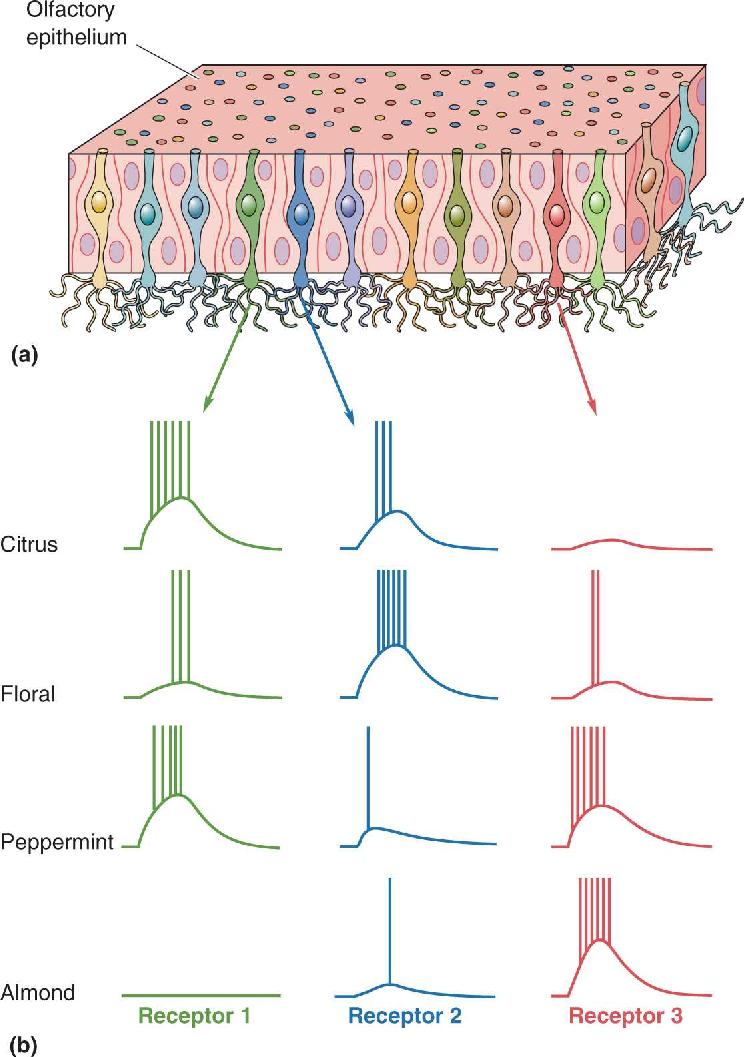
Ch 8The Chemical SensesRead full chapter →
Sections in this chapter
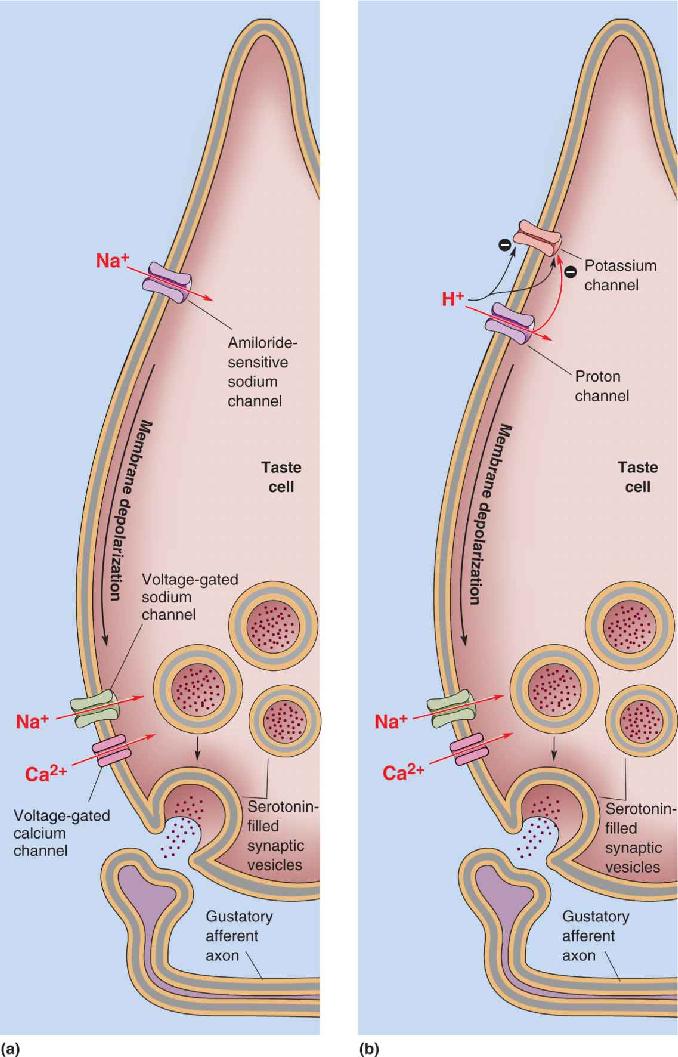
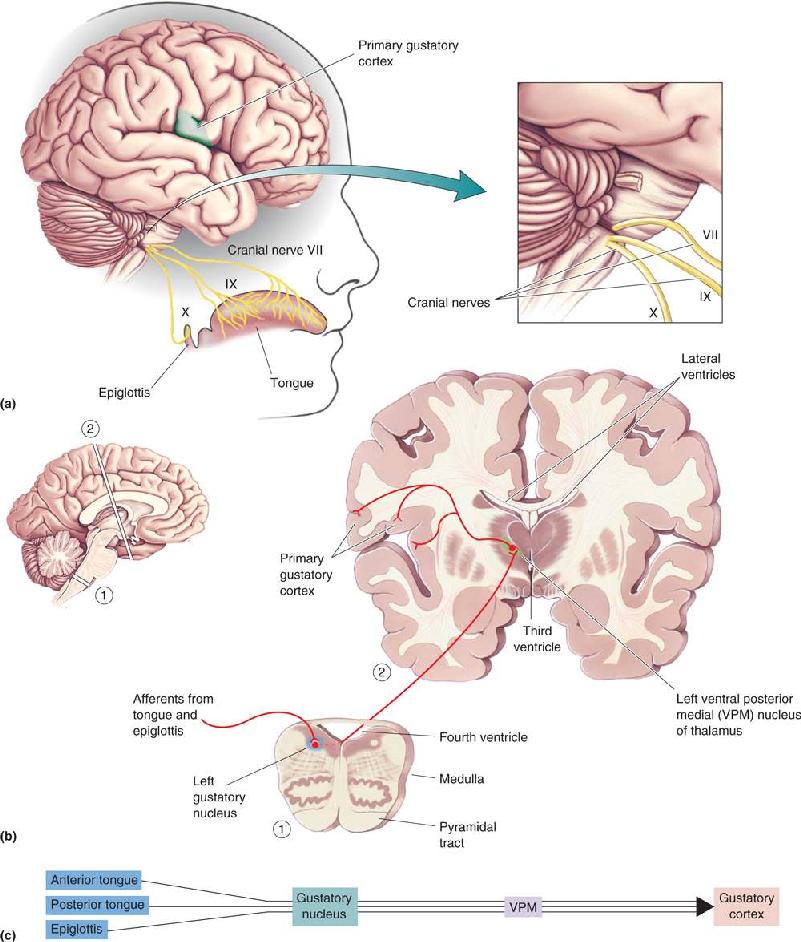
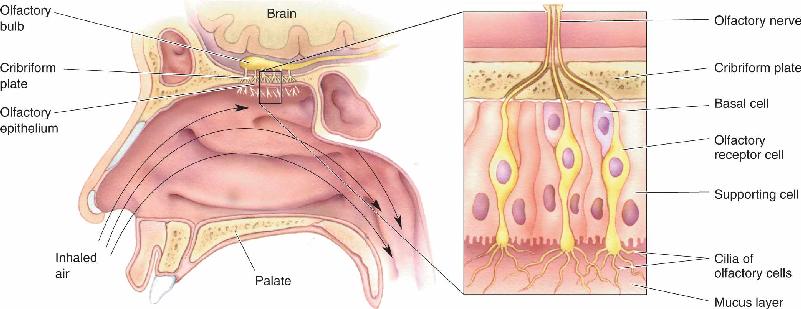
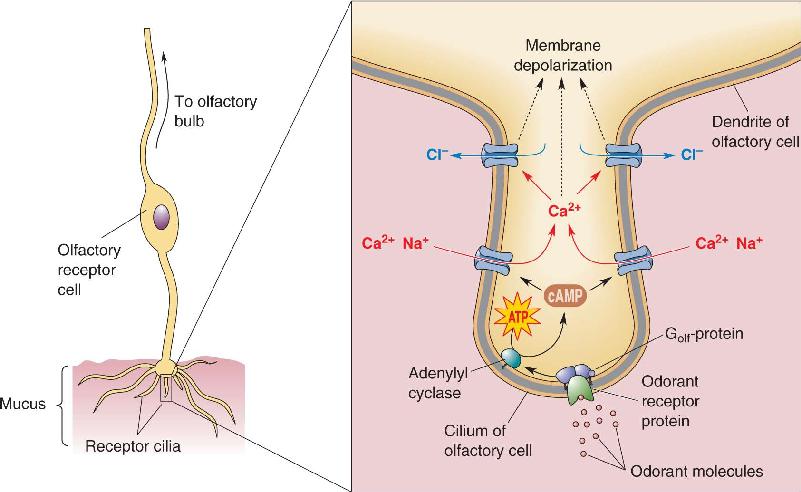
- The Chemical Senses
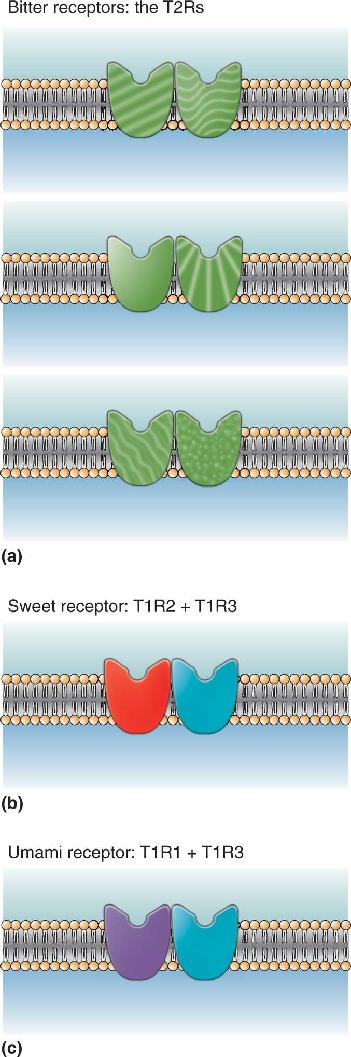
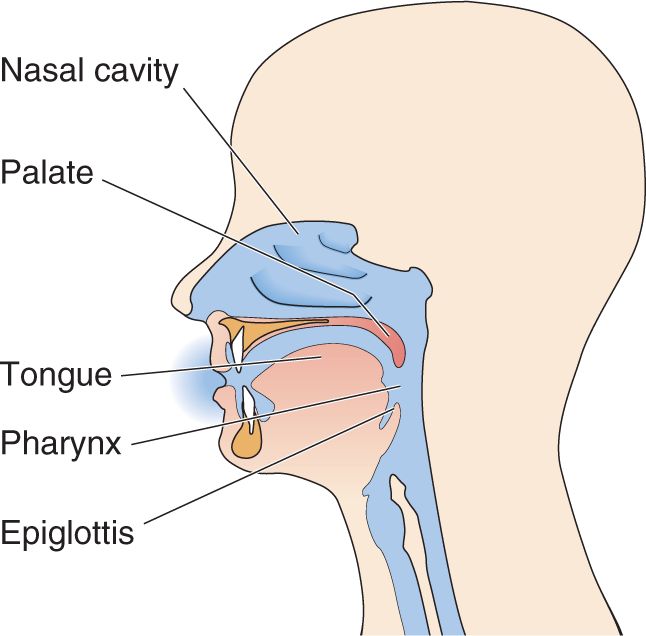
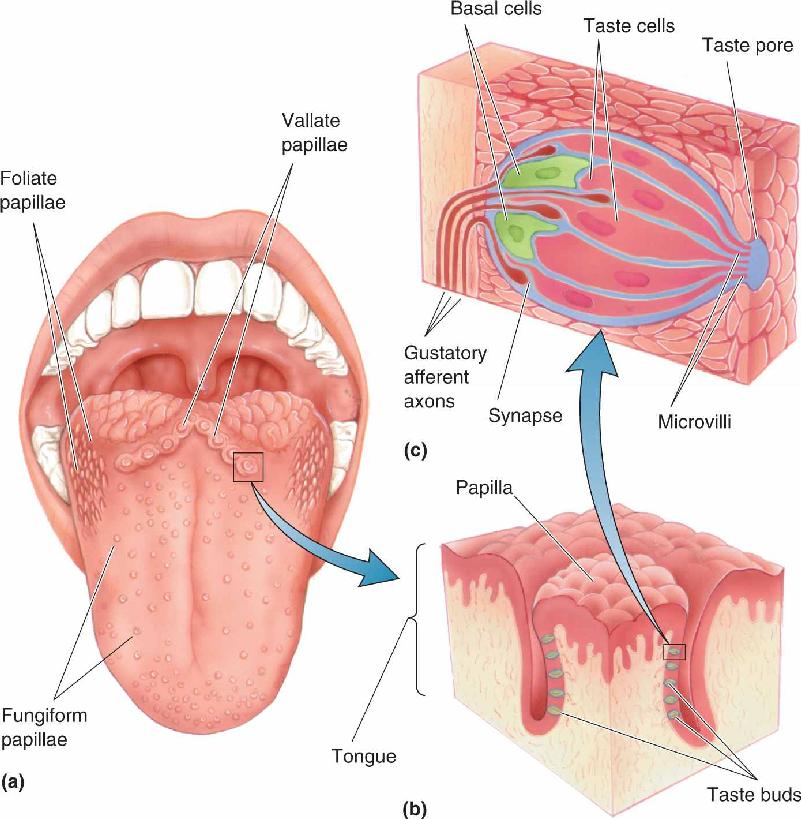
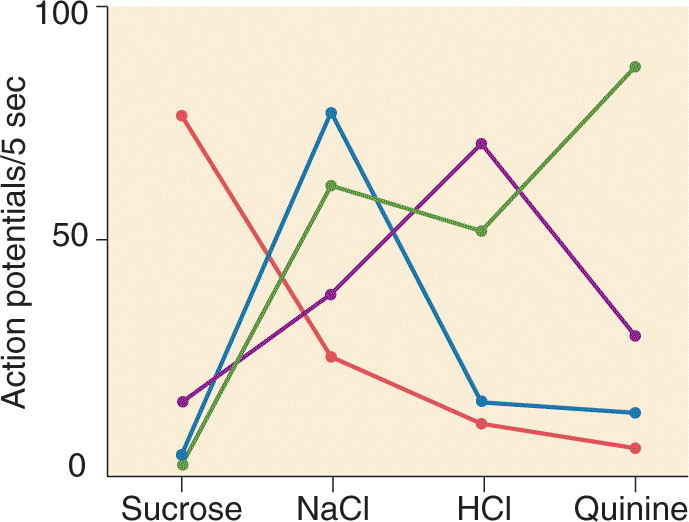
Ch 9The EyeRead full chapter →
Sections in this chapter
- The Eye
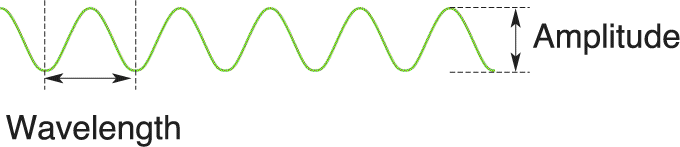
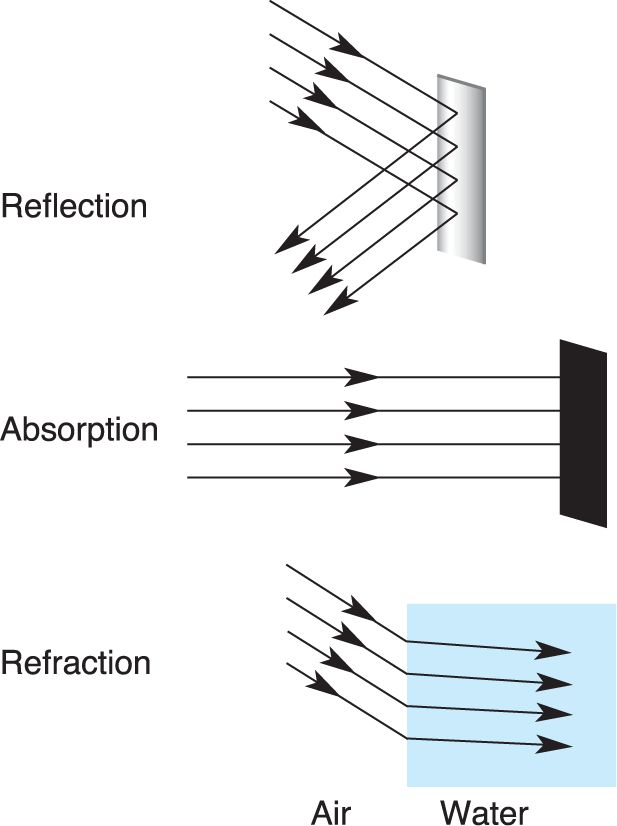
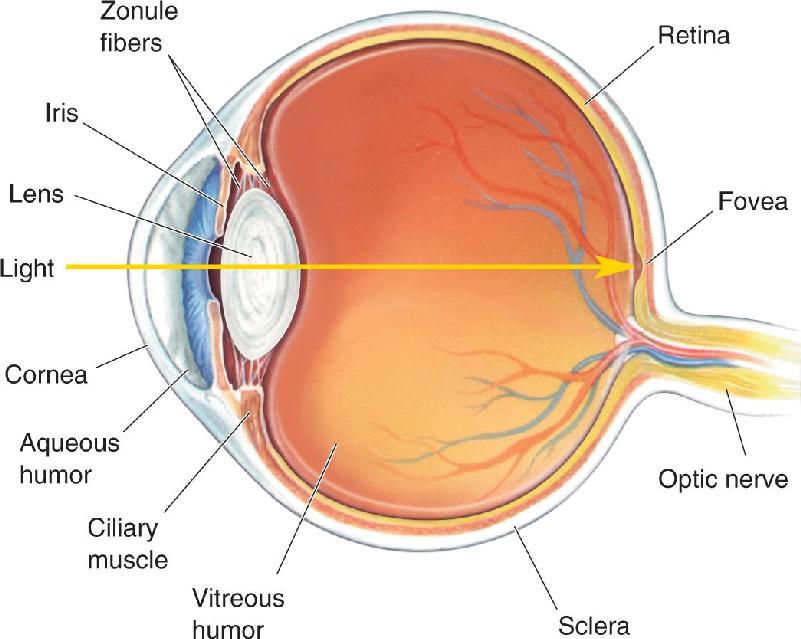
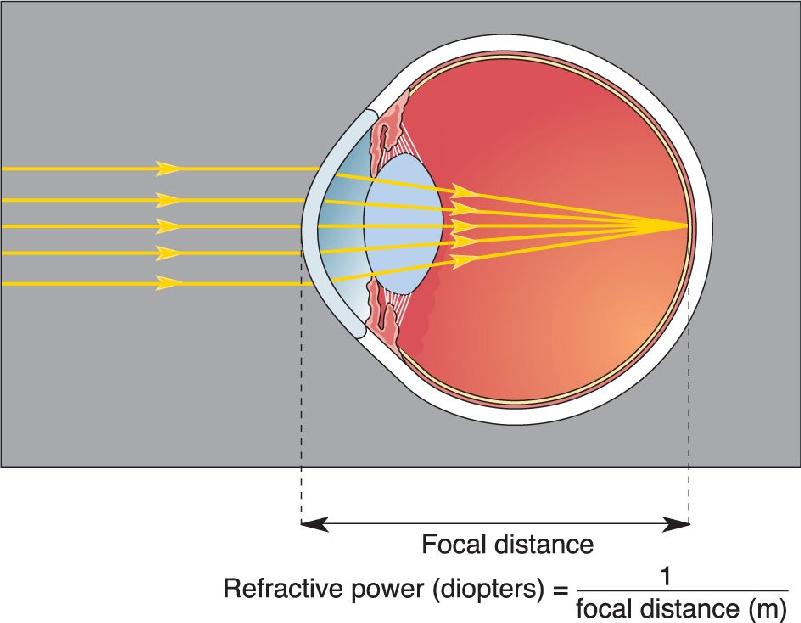
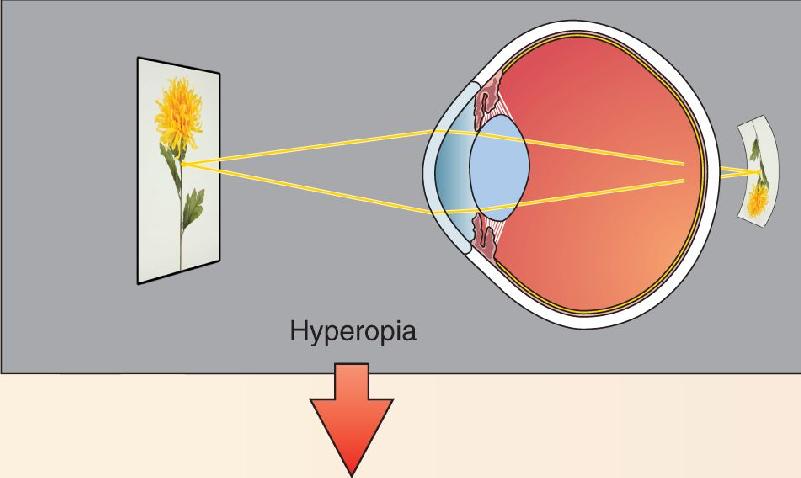
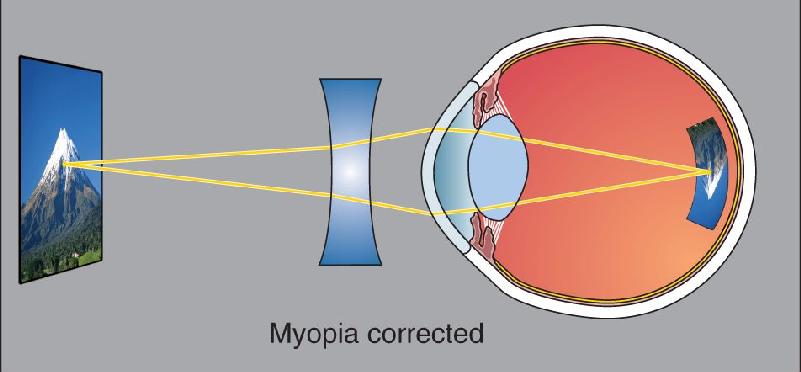
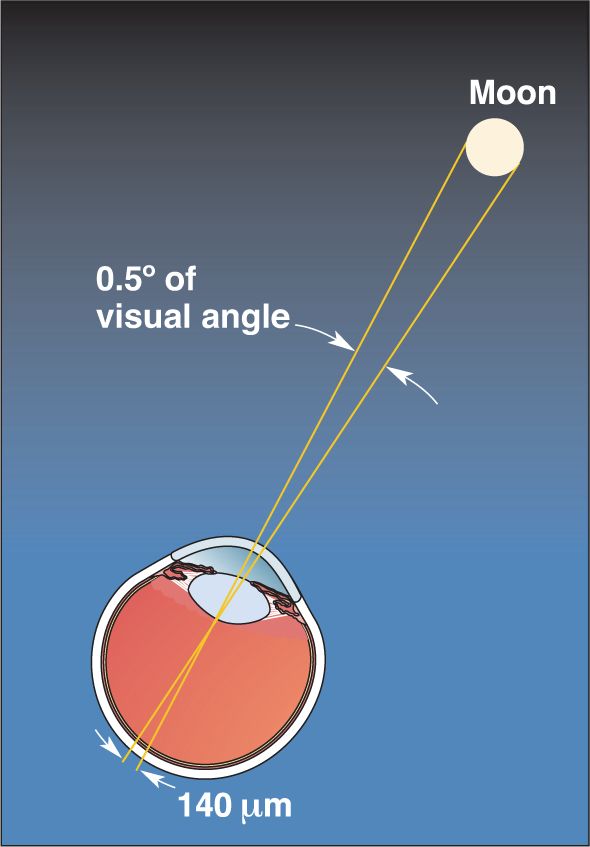


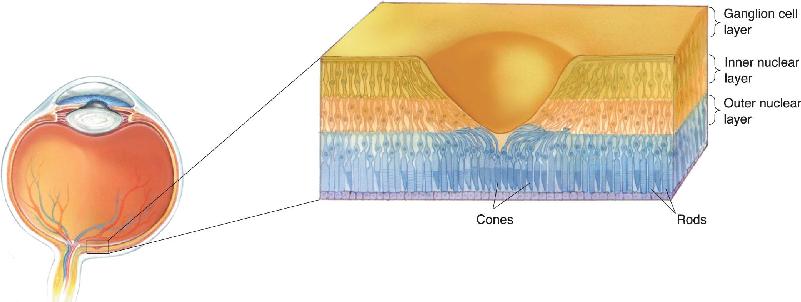
Ch 10The Central Visual SystemRead full chapter →
Sections in this chapter
- The Central Visual System
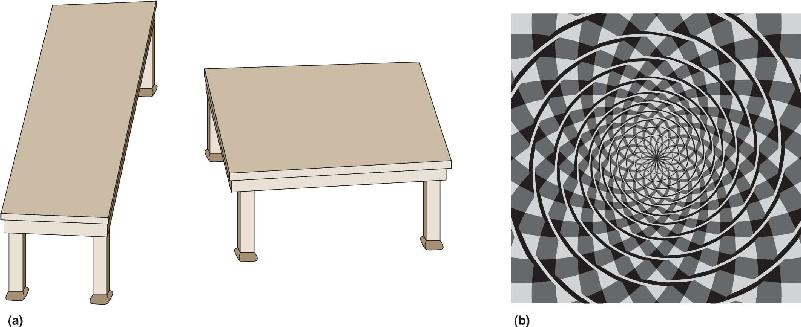
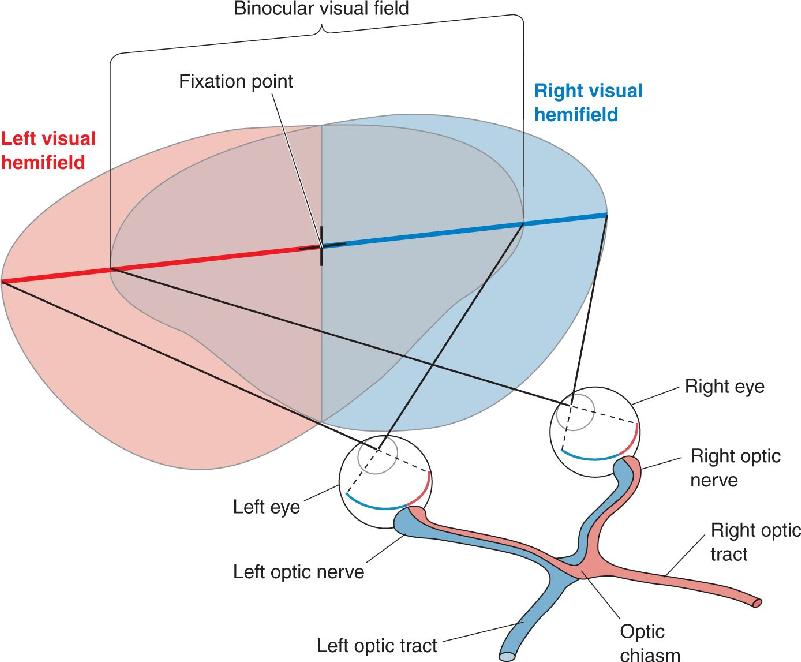
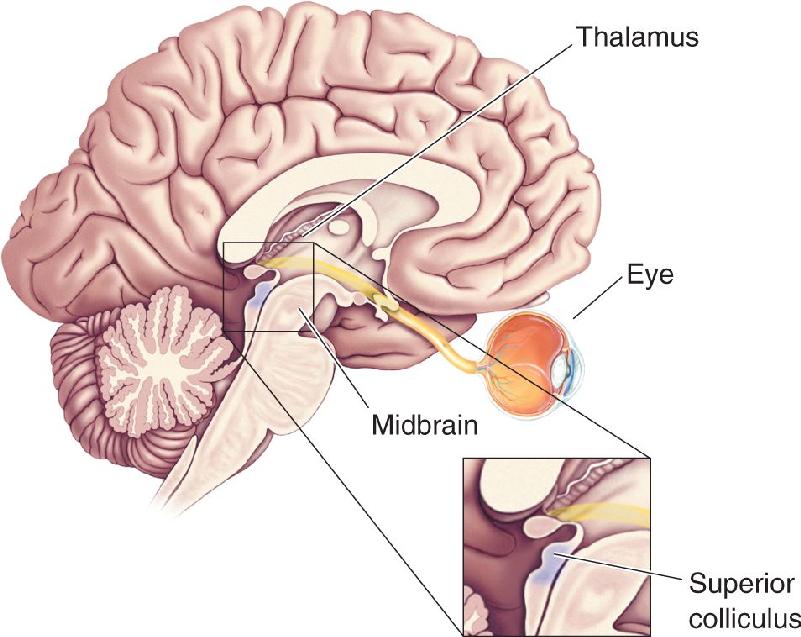
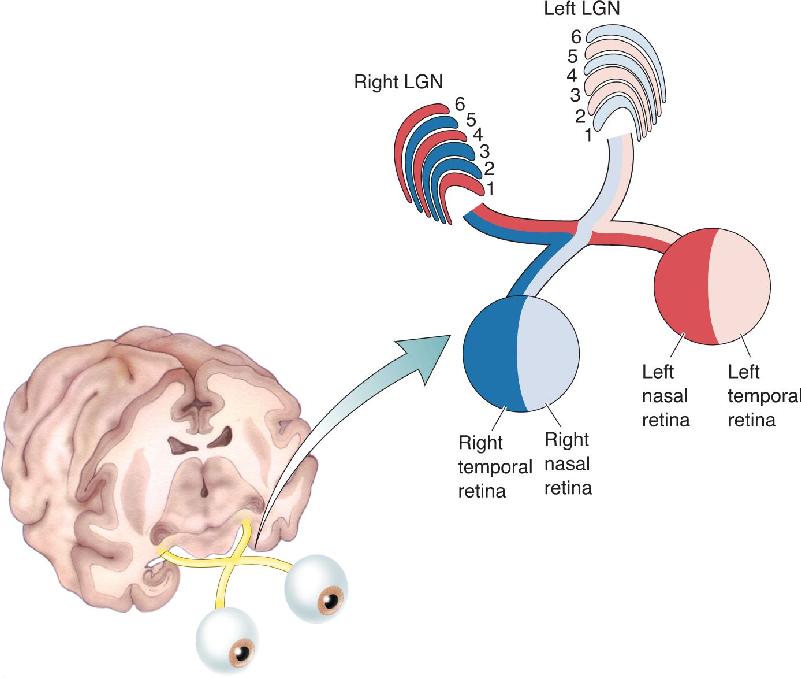
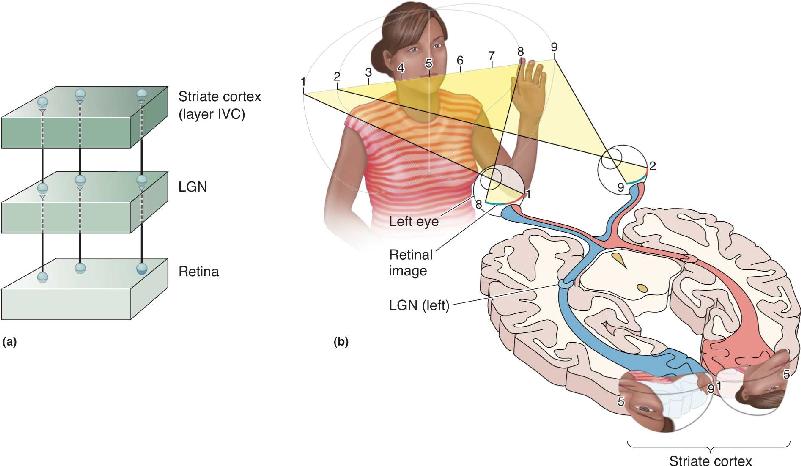
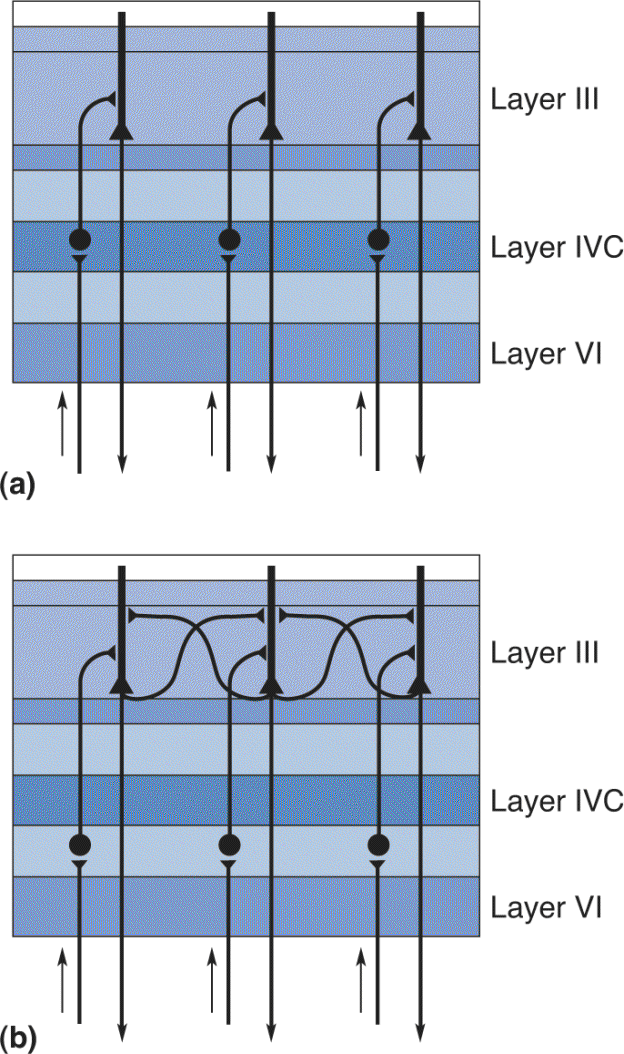

Ch 11The Auditory and Vestibular SystemsRead full chapter →
Sections in this chapter
- The Auditory and Vestibular Systems
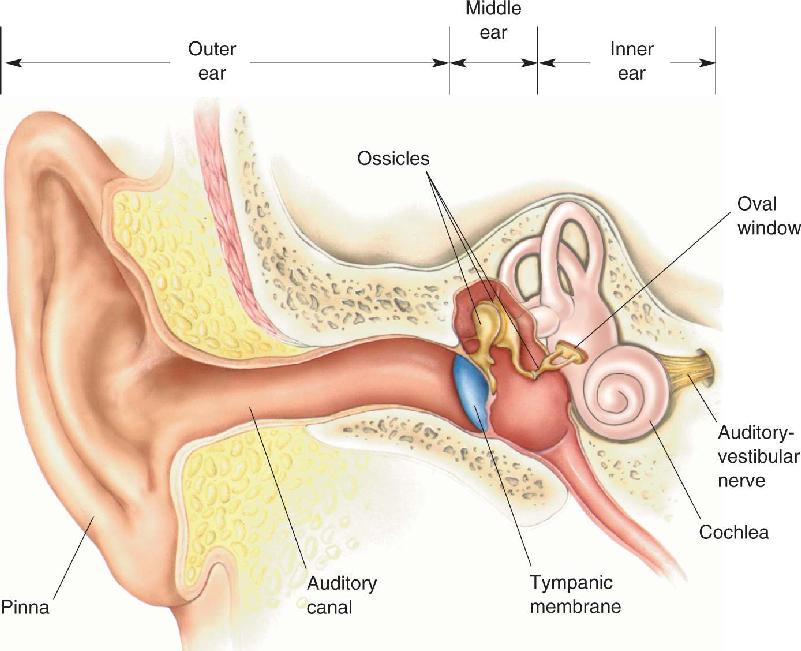
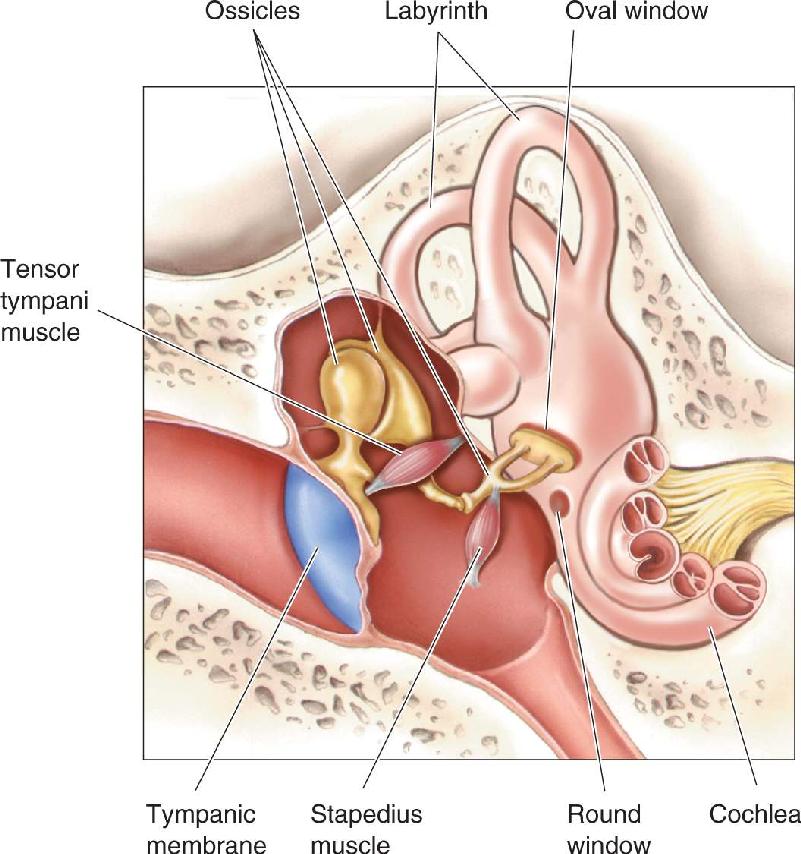
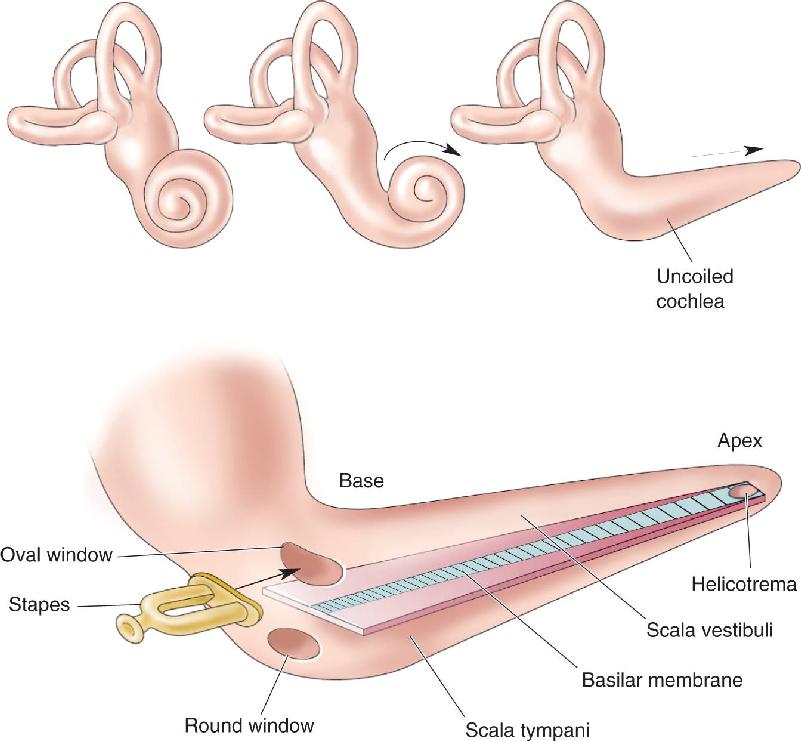
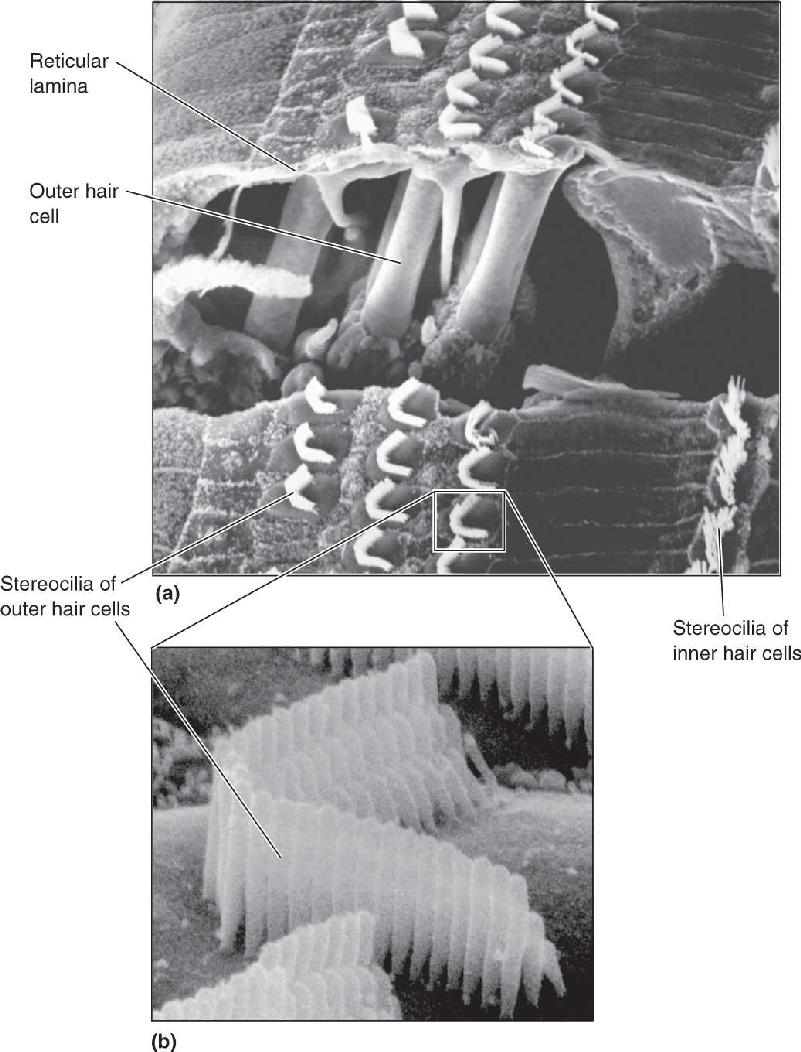

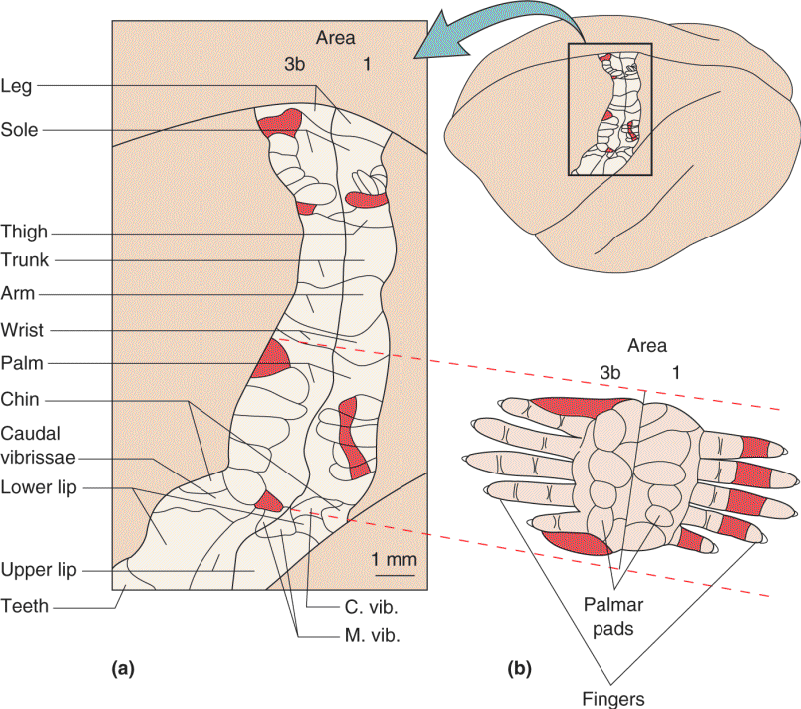
Ch 12The Somatic Sensory SystemRead full chapter →
Sections in this chapter
- The Somatic Sensory System
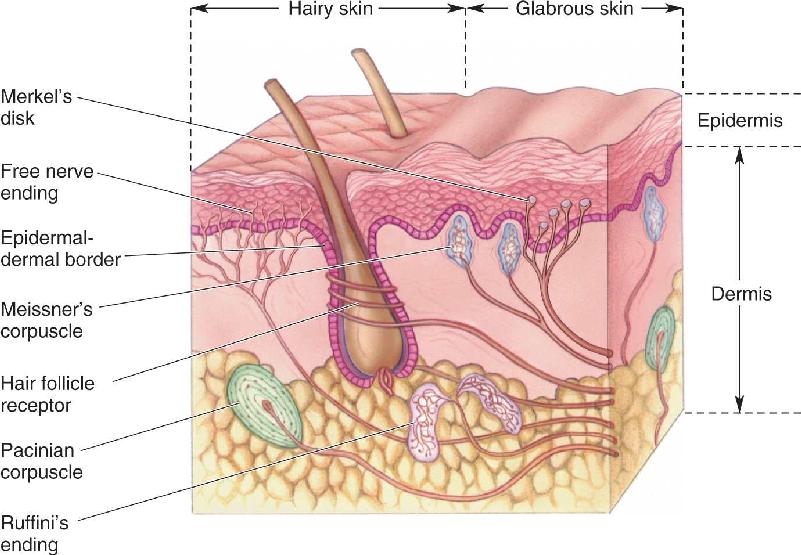
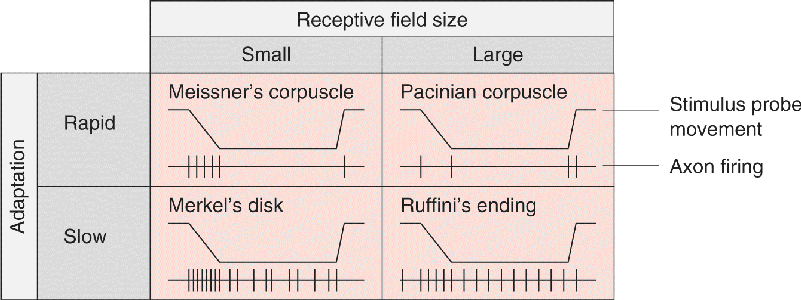
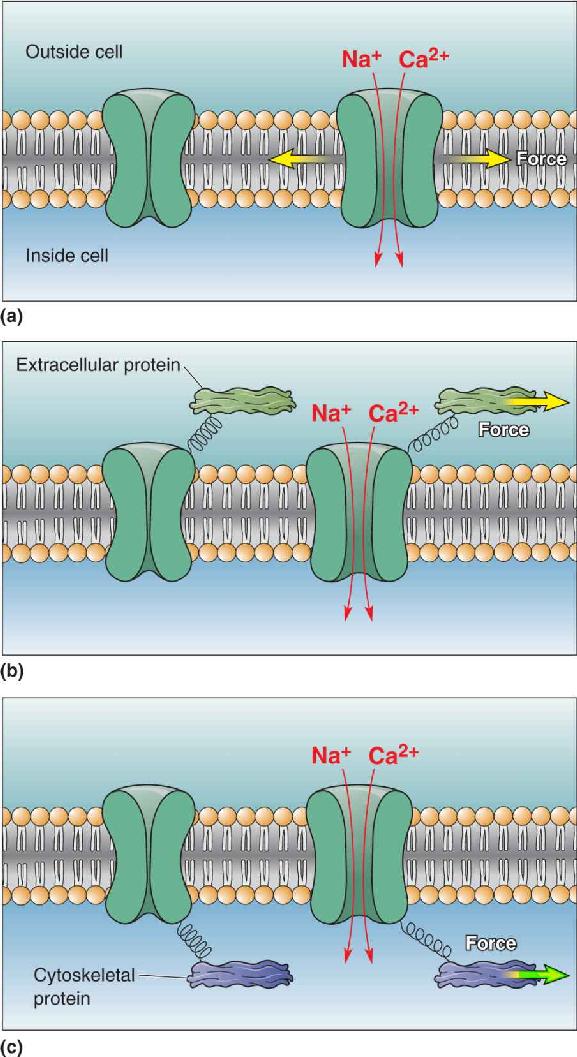
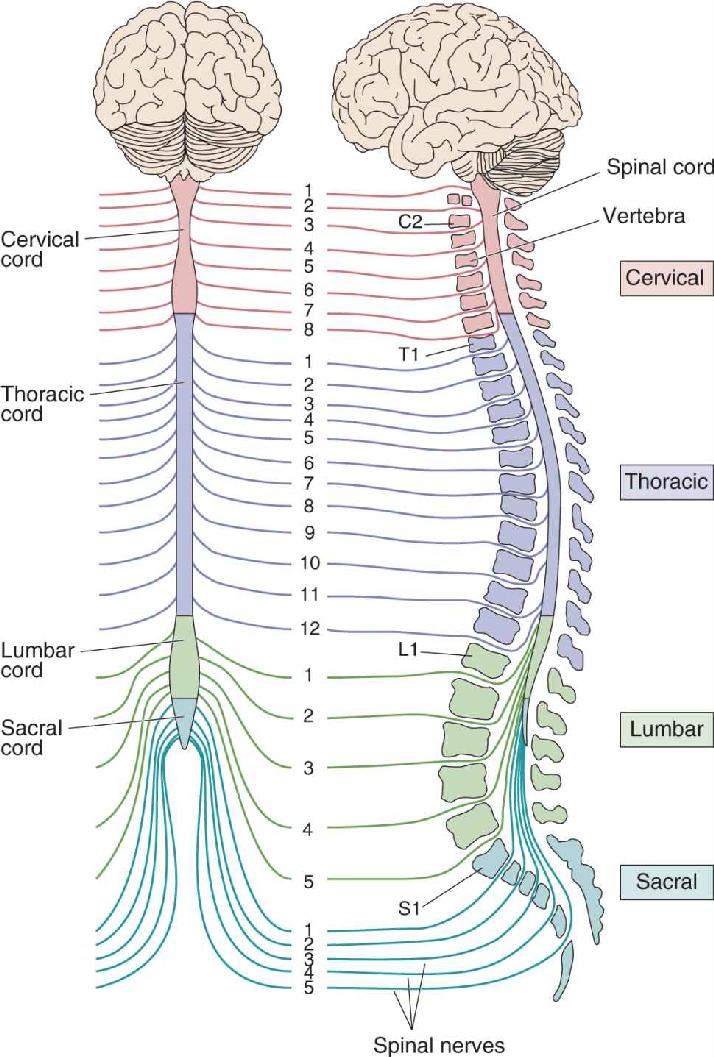
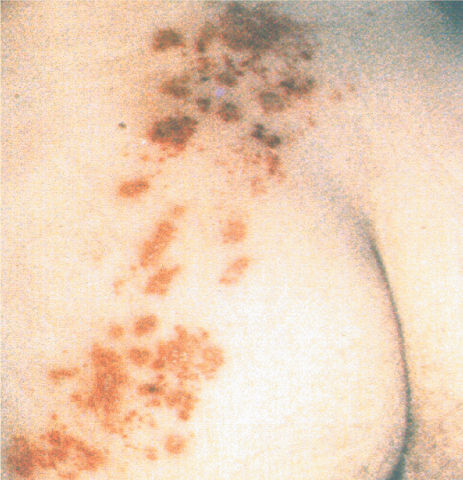
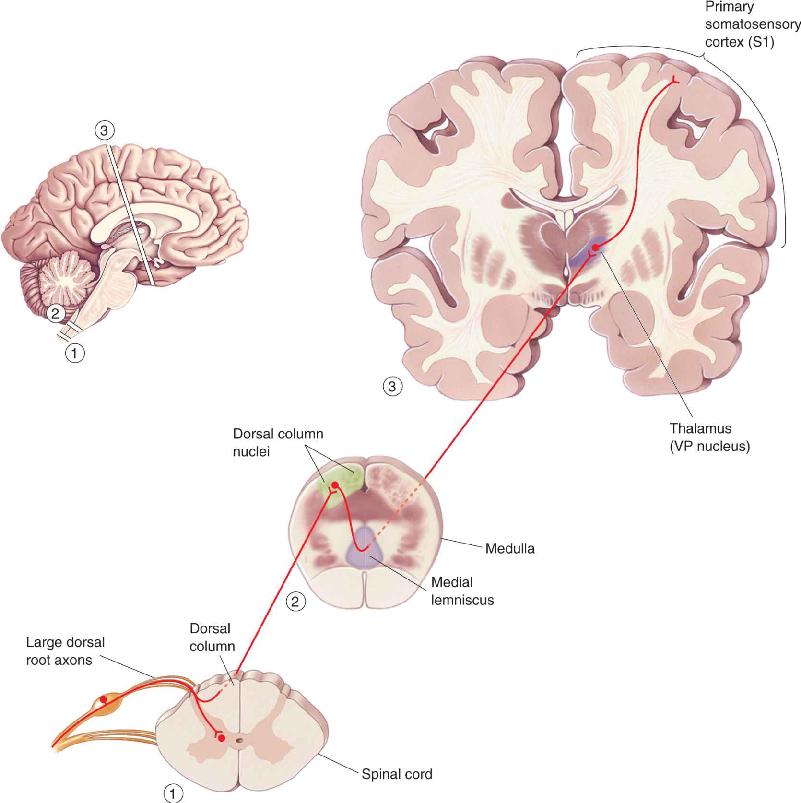
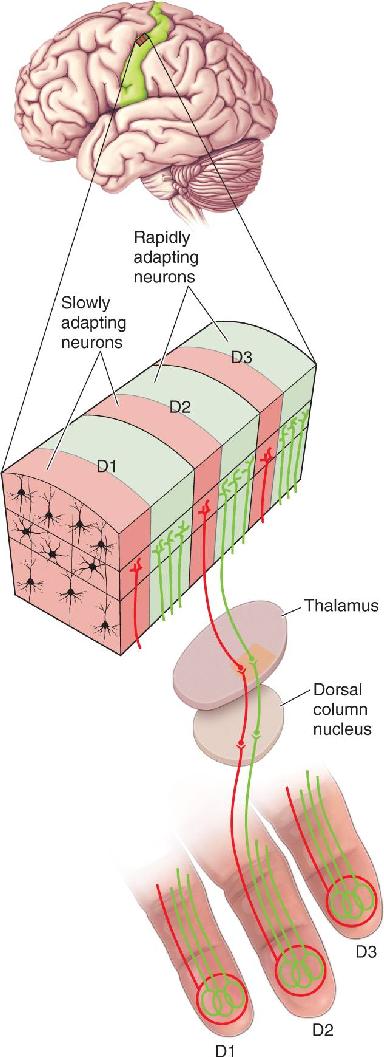
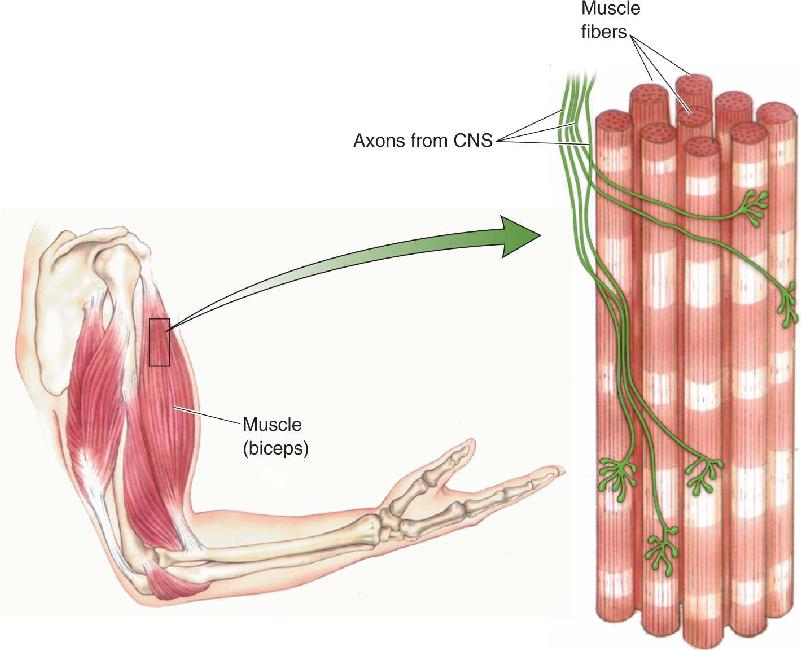
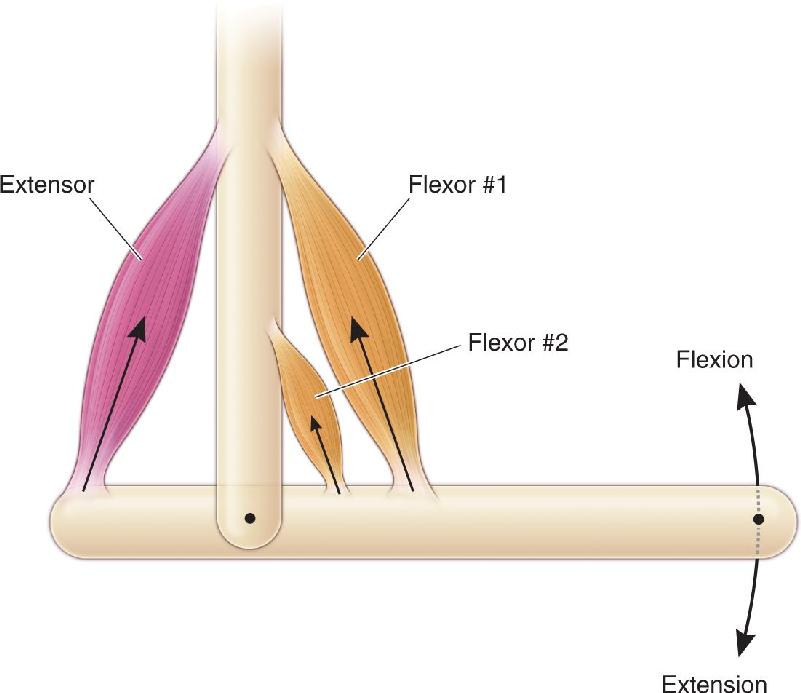
Ch 13Spinal Control of MovementRead full chapter →
Sections in this chapter
- Spinal Control of Movement
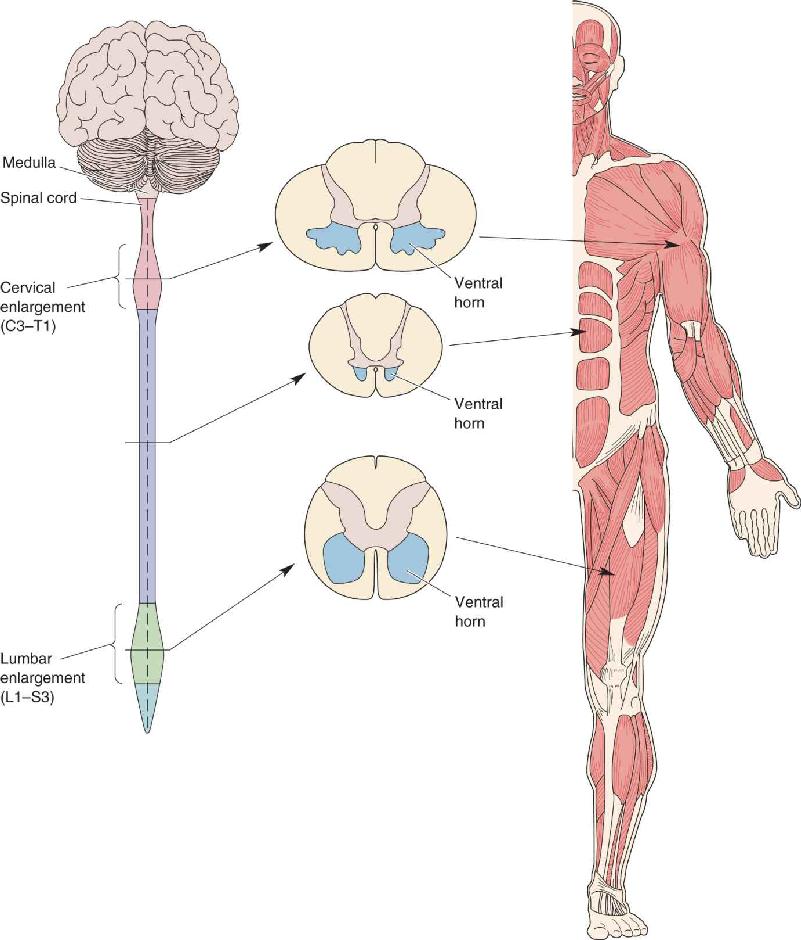
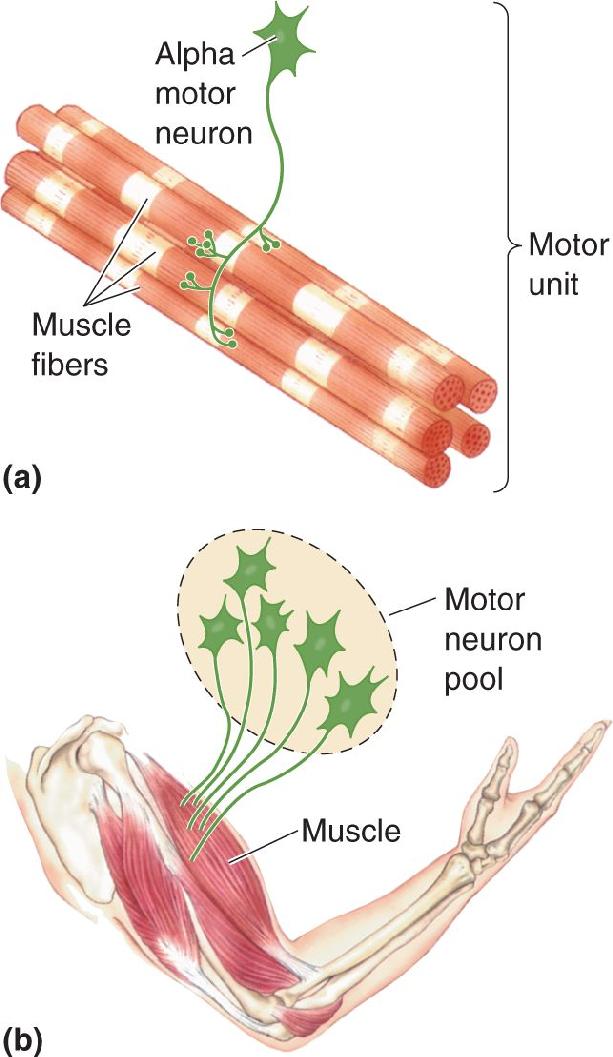
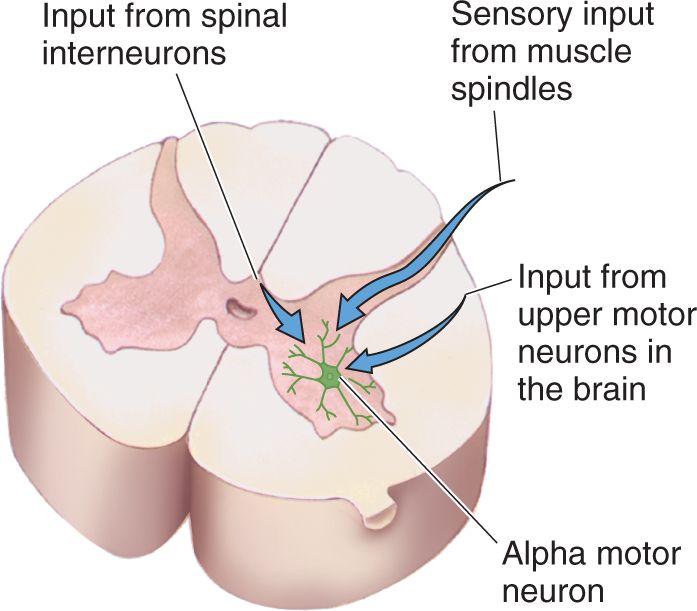
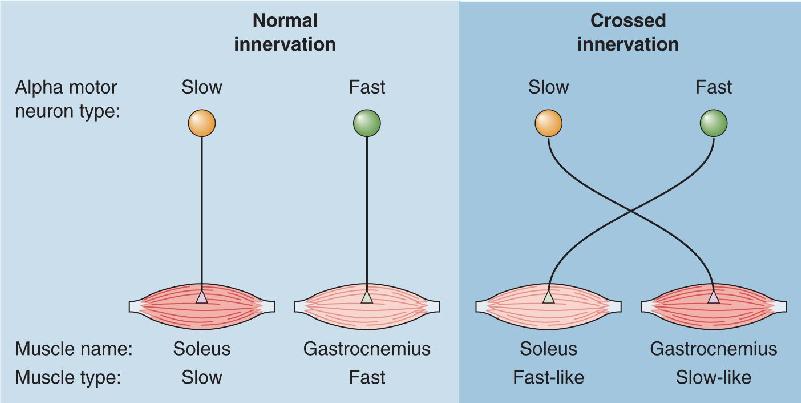
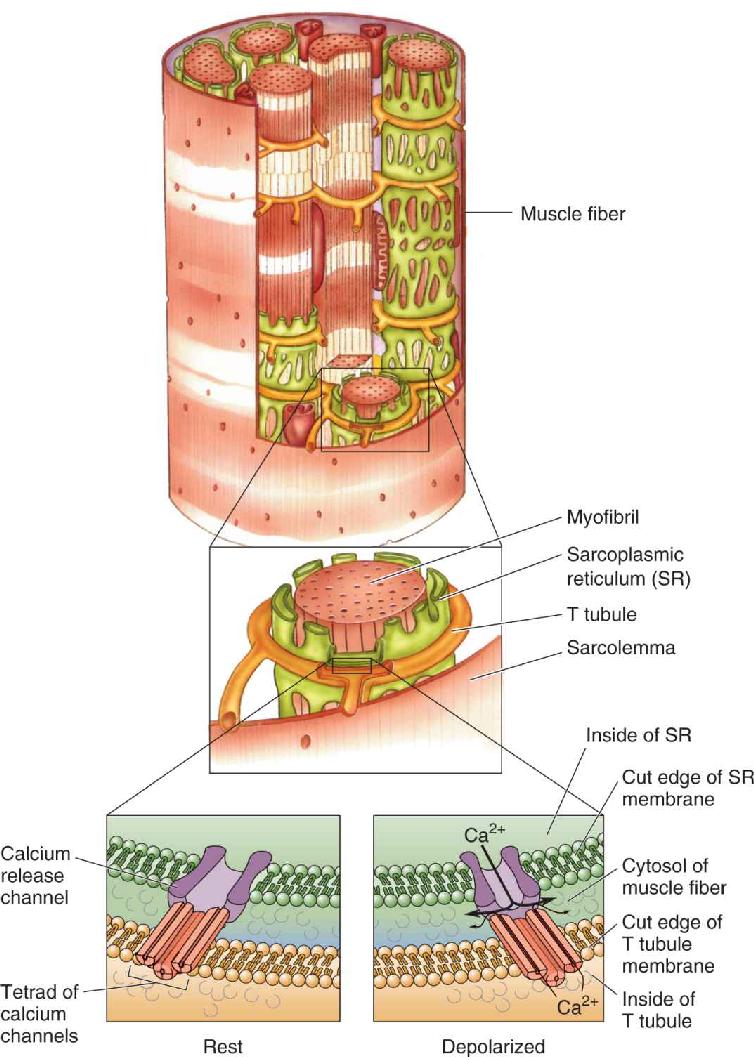
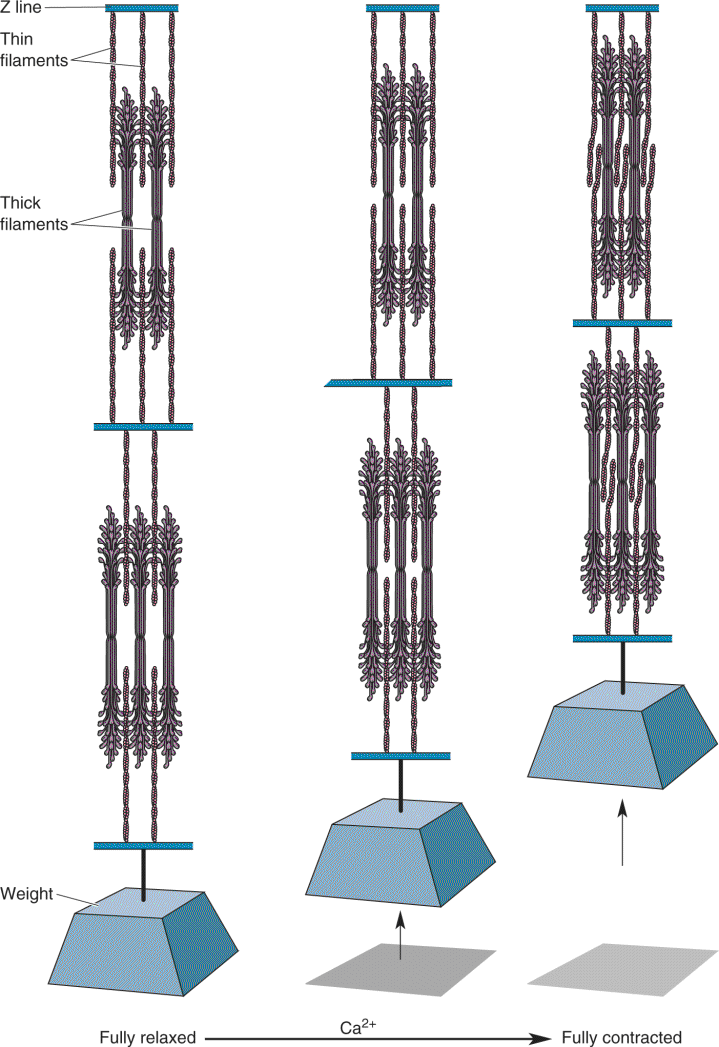
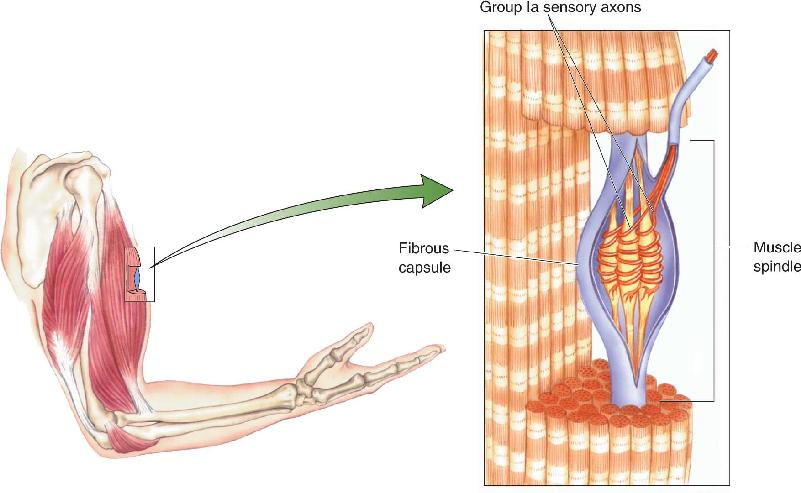
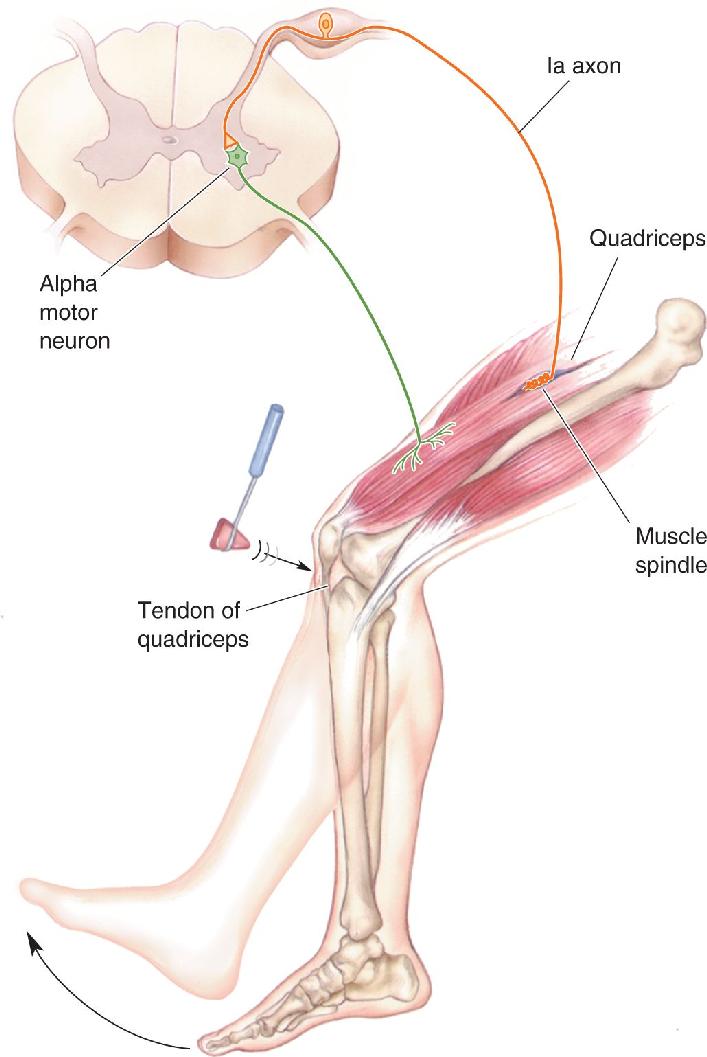
Ch 14Brain Control of MovementRead full chapter →
Sections in this chapter
- Brain Control of Movement

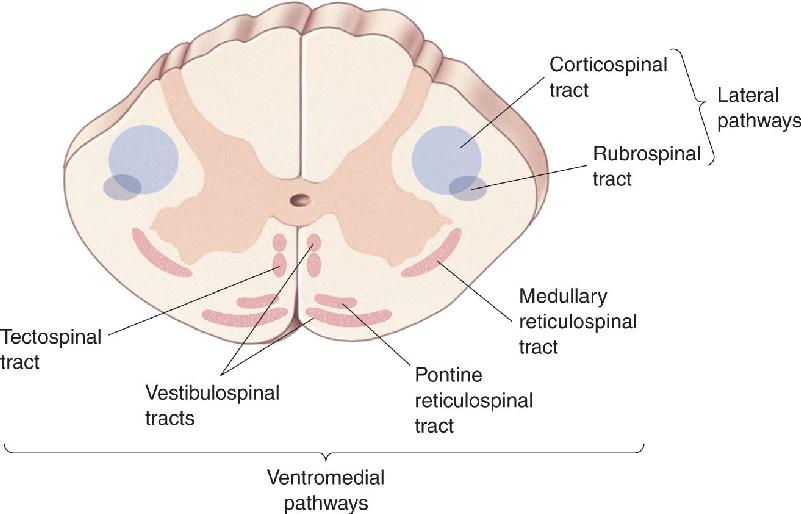
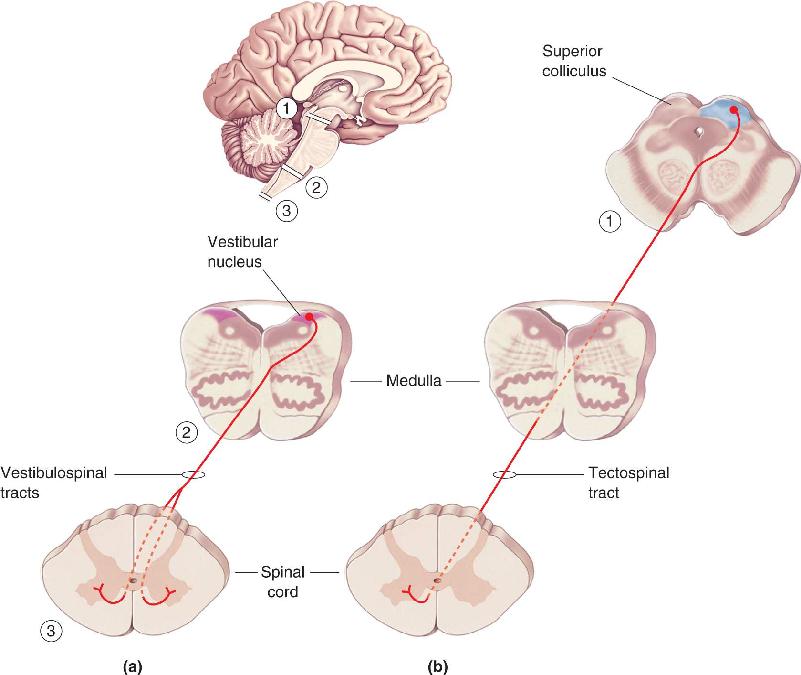
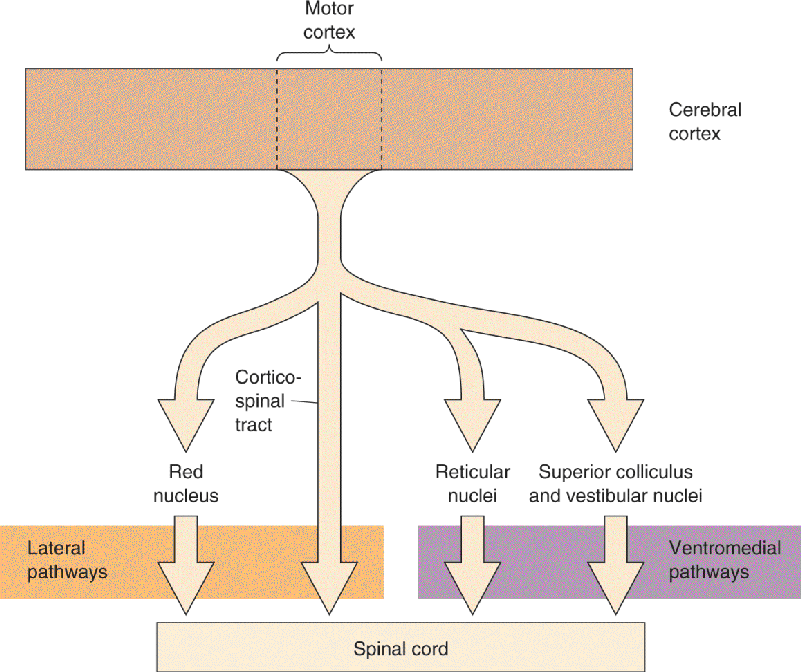
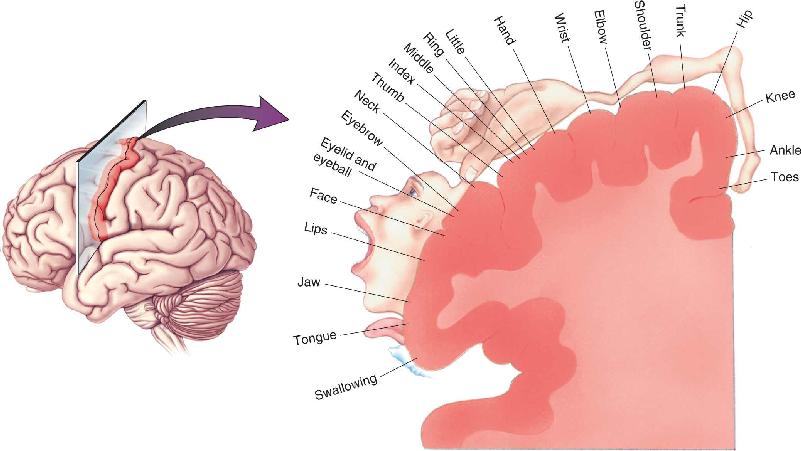
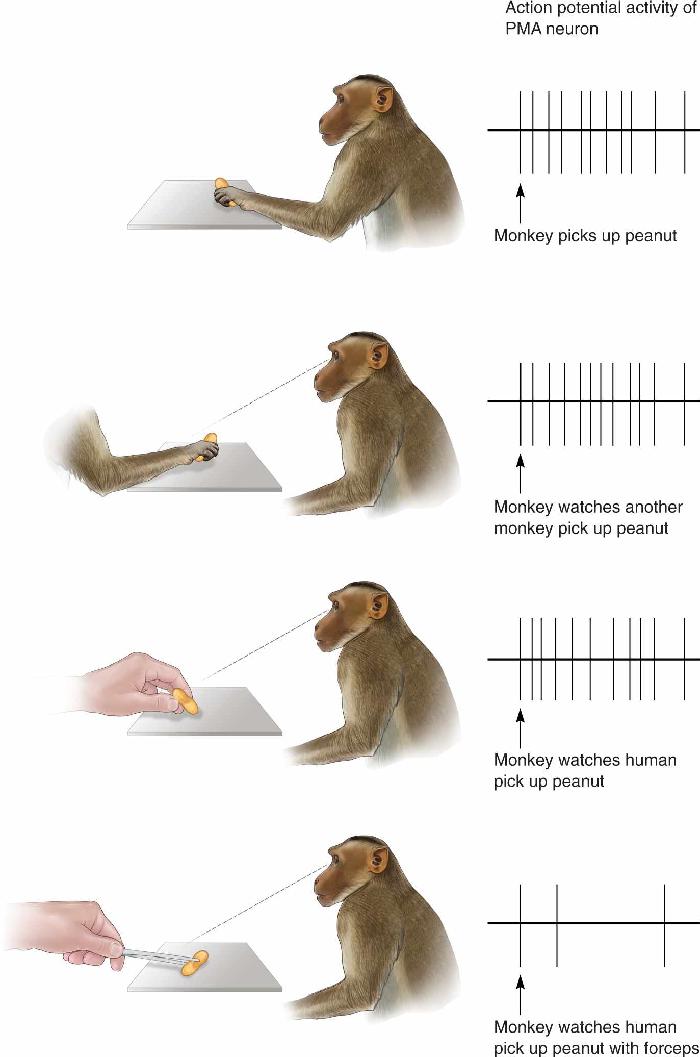
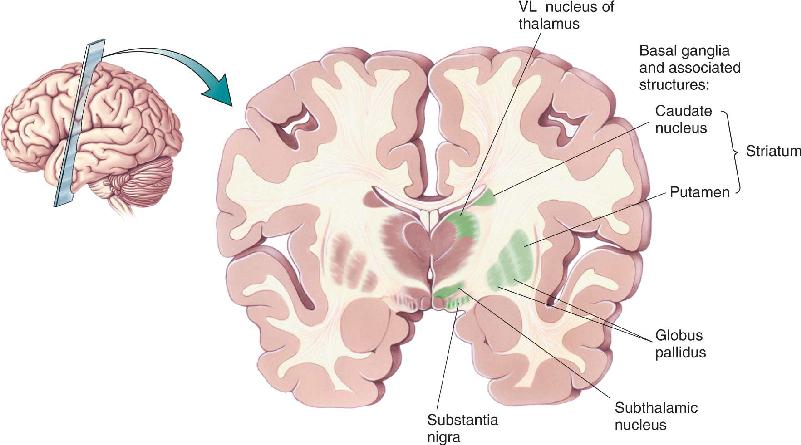
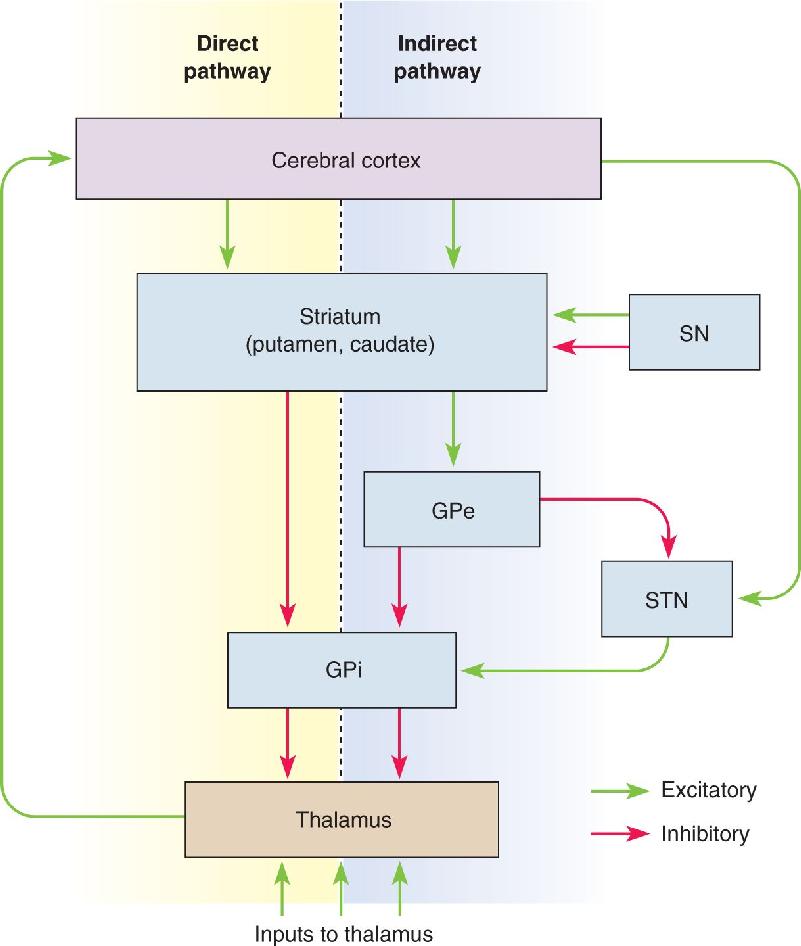
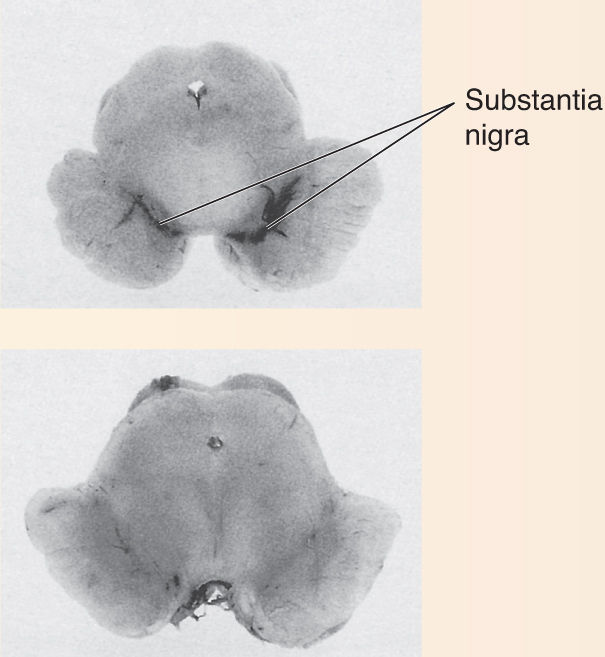
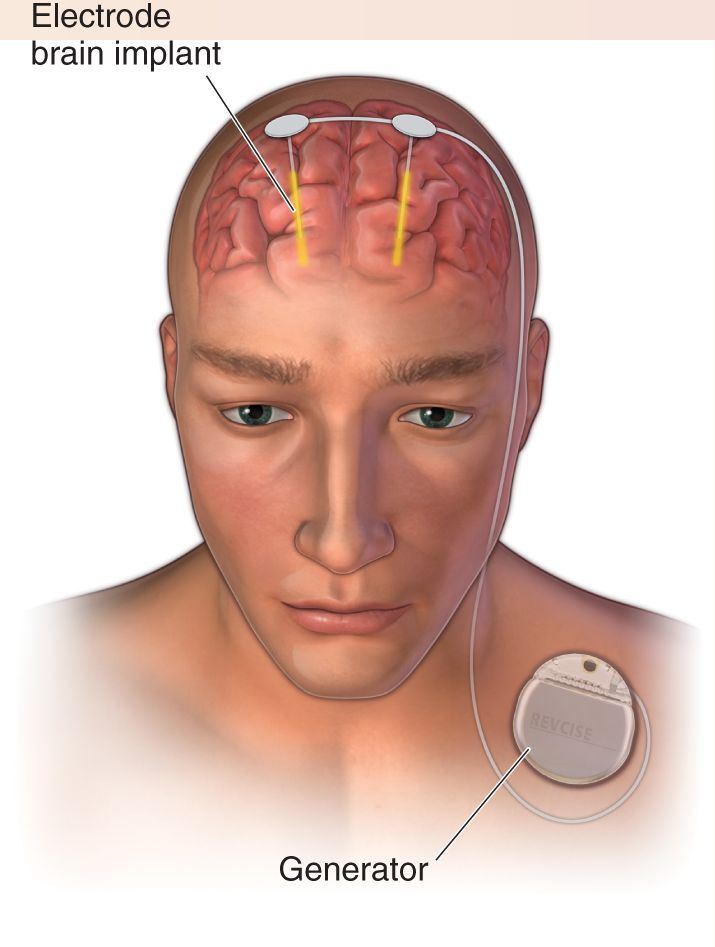
Ch 15Chemical Control of the Brain and BehaviorRead full chapter →
Sections in this chapter
- Chemical Control of the Brain and Behavior
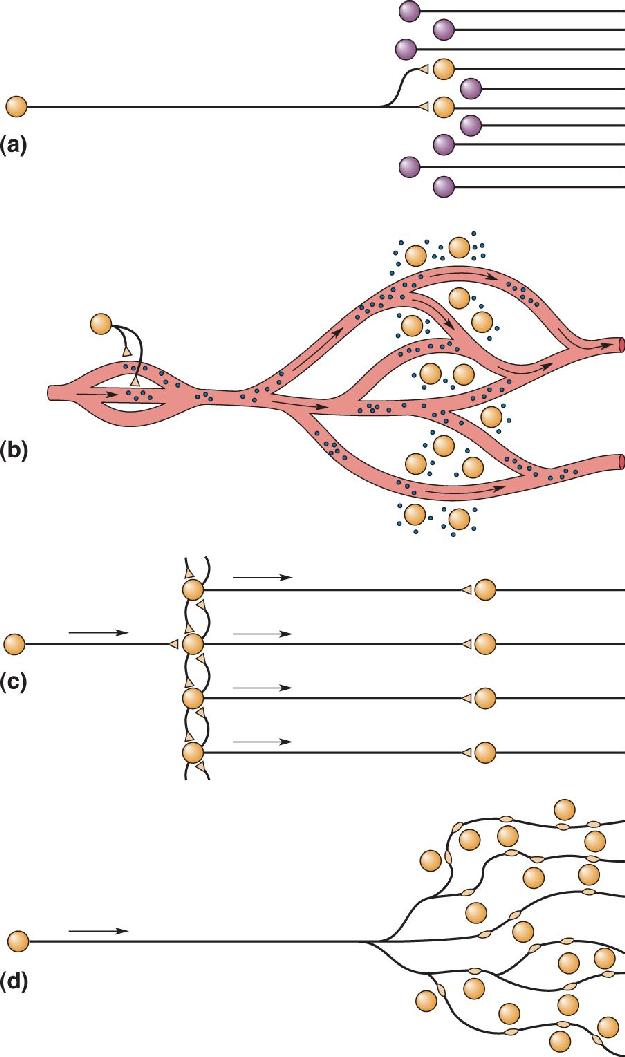
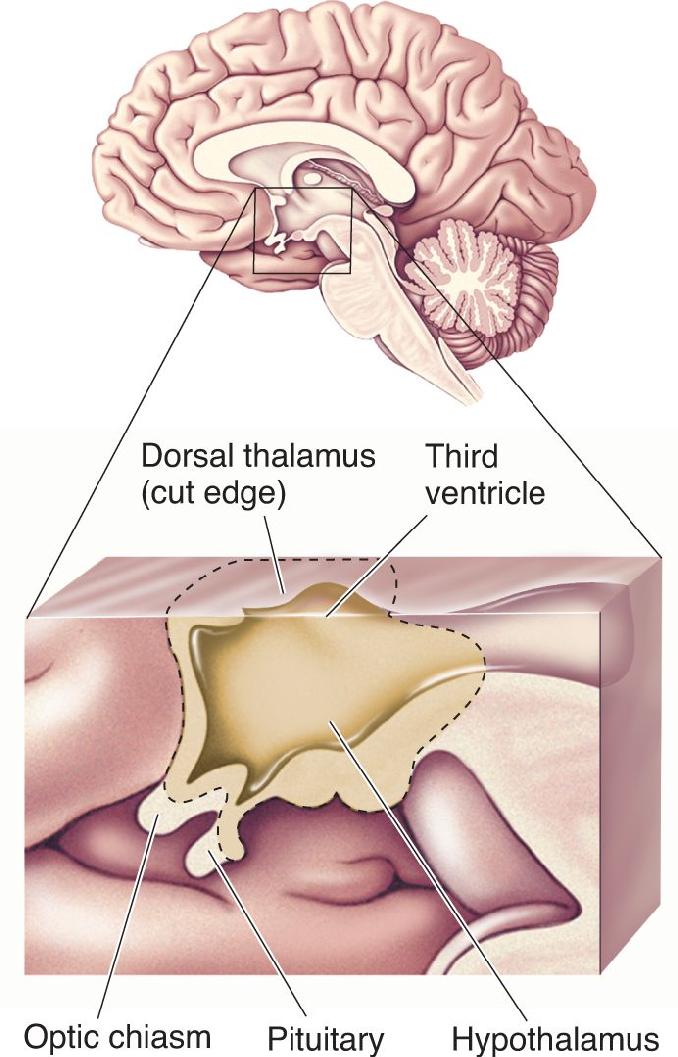
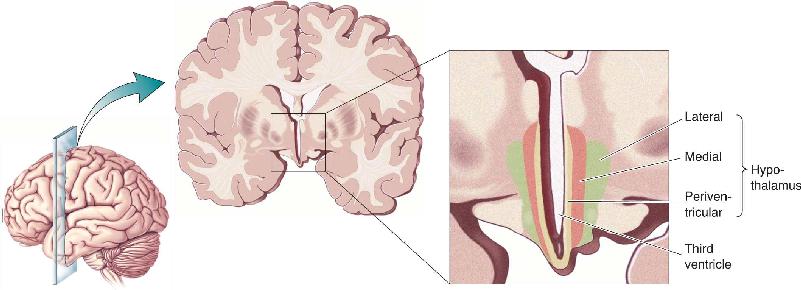
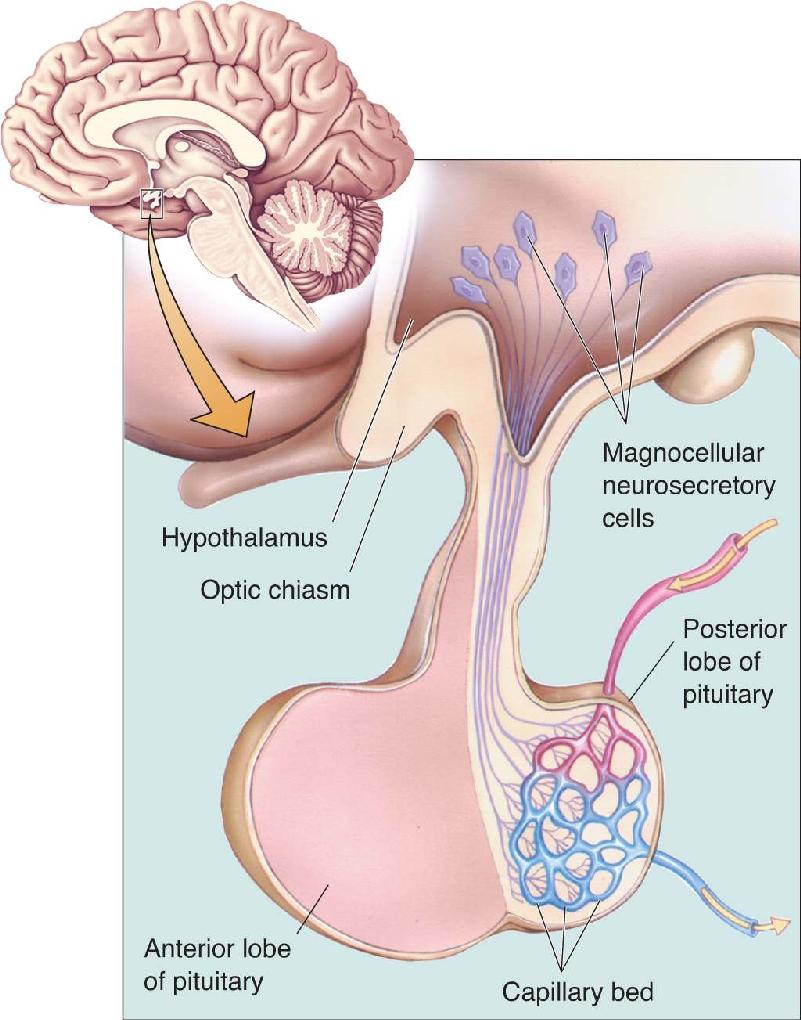
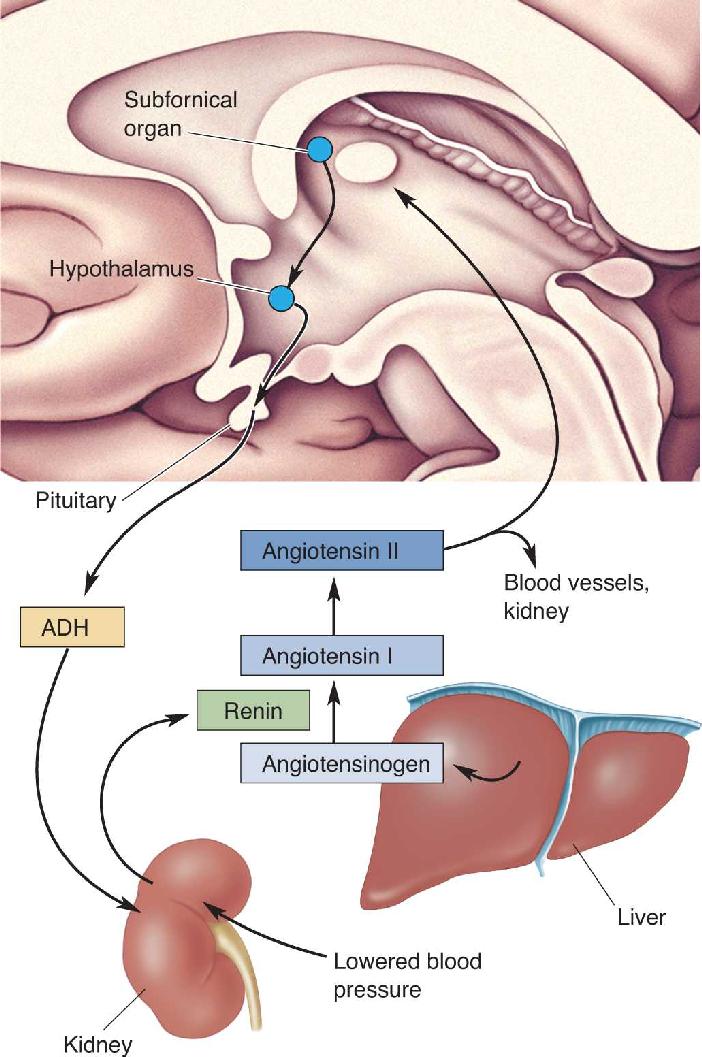
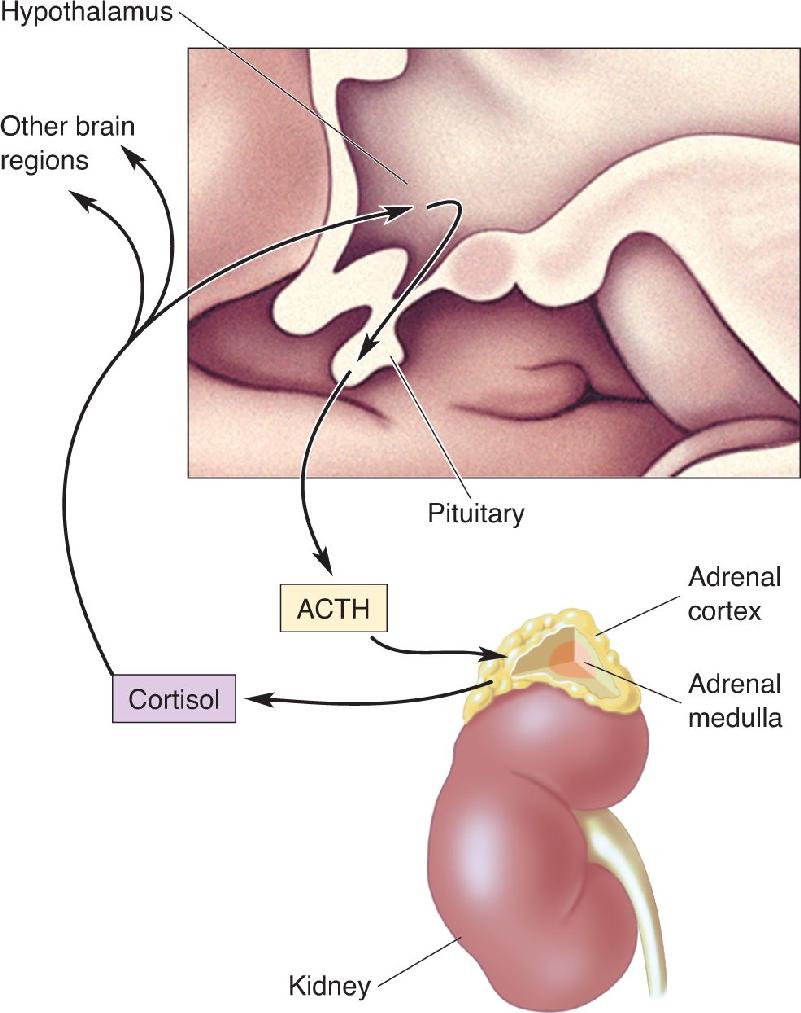
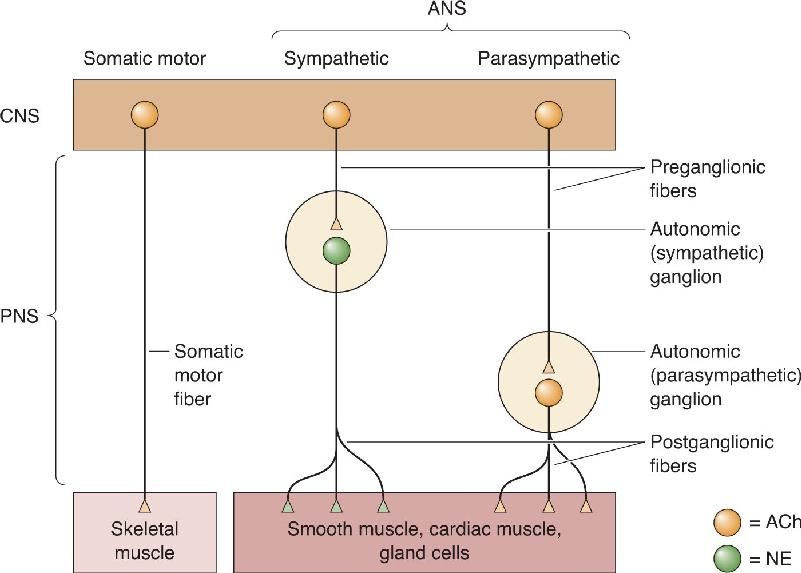
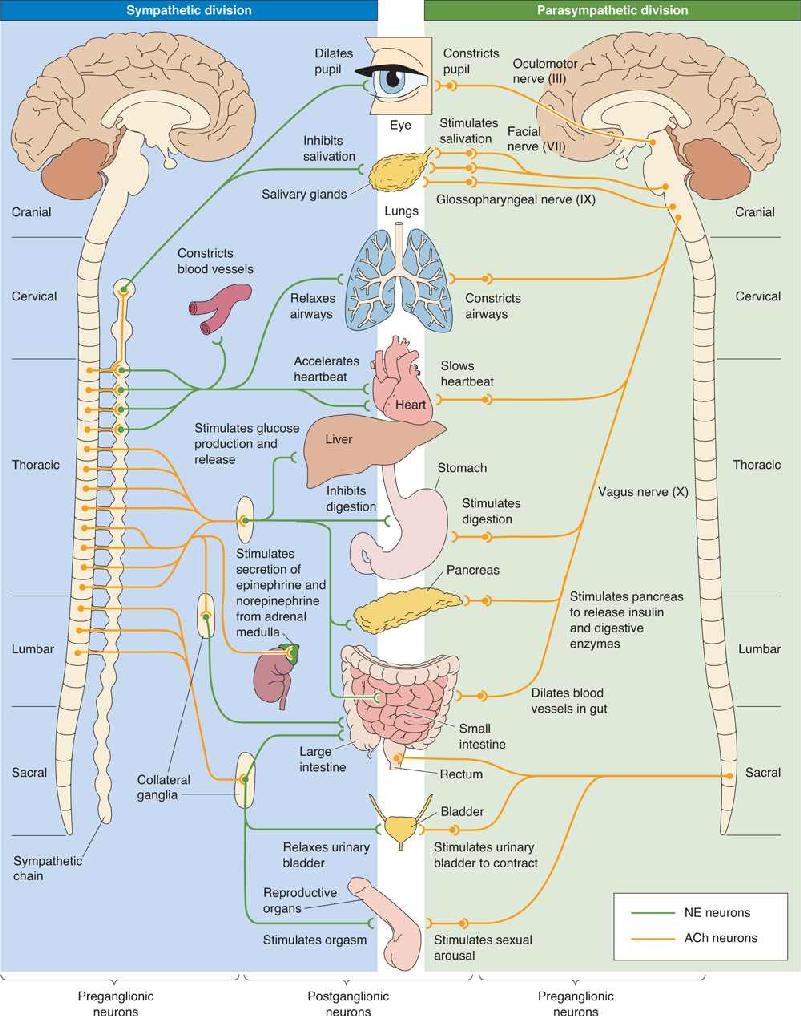
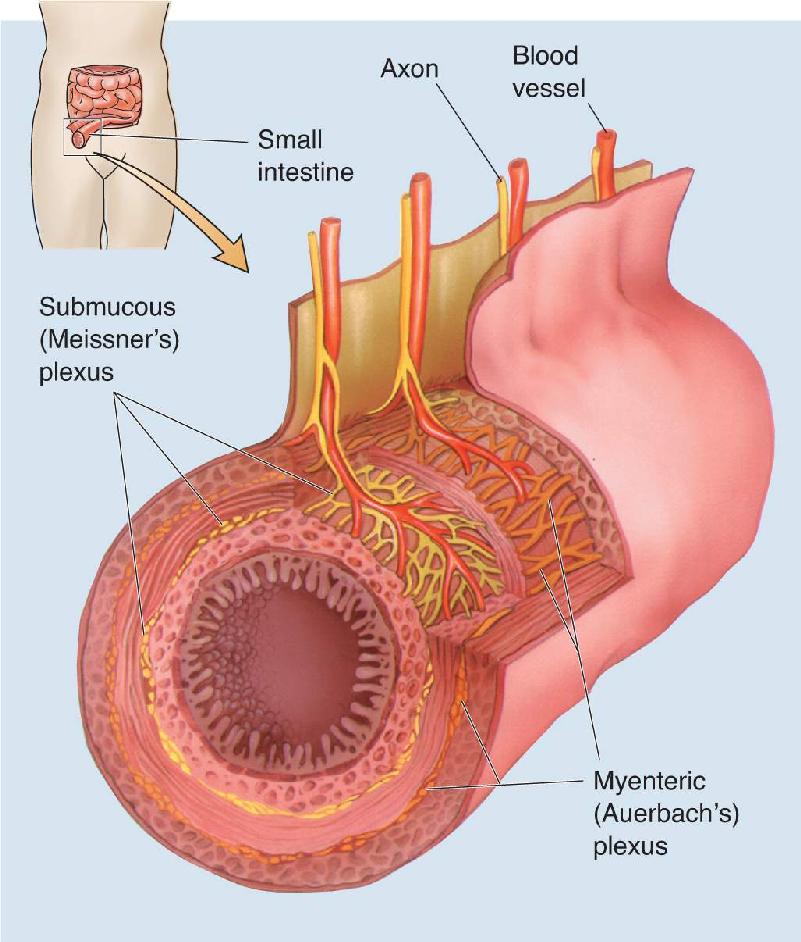
Ch 16MotivationRead full chapter →
Sections in this chapter
- Motivation
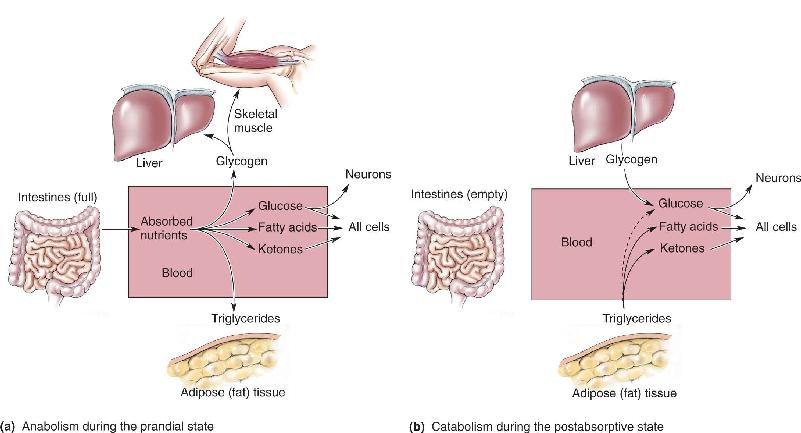
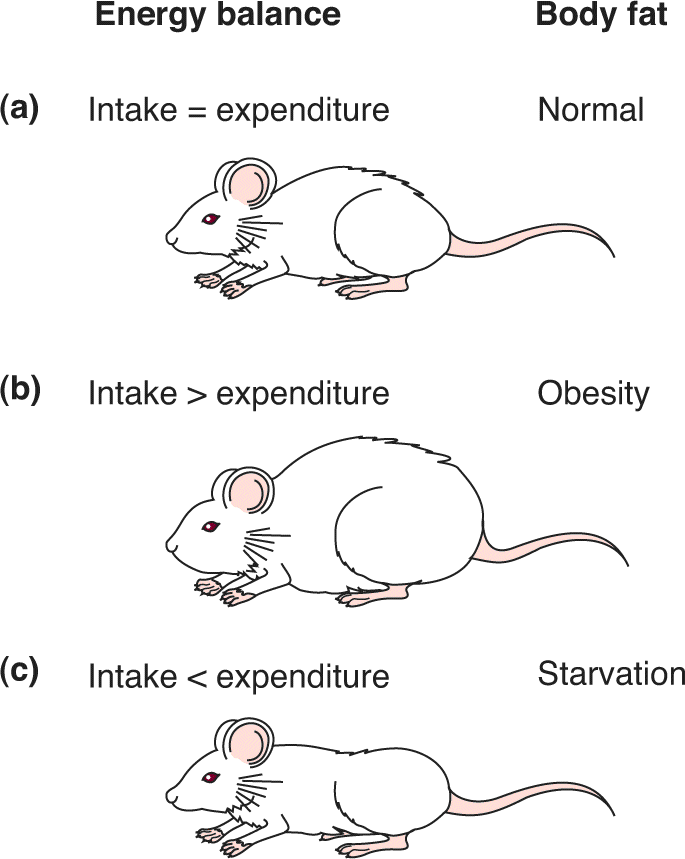
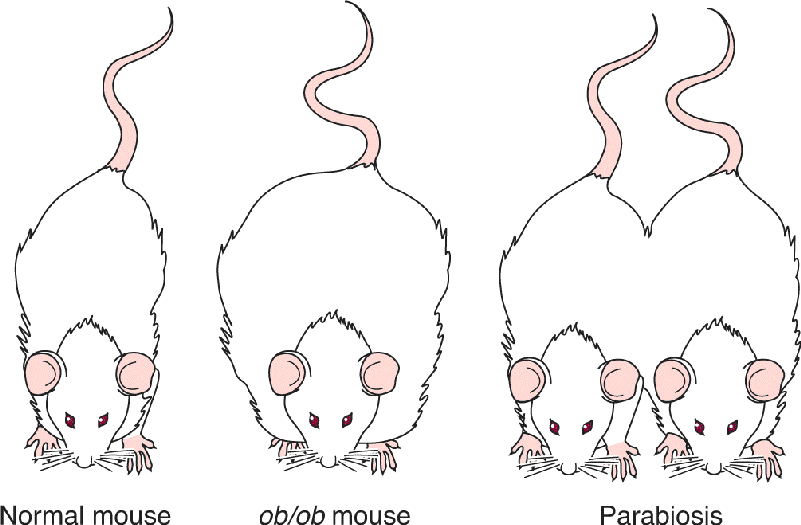
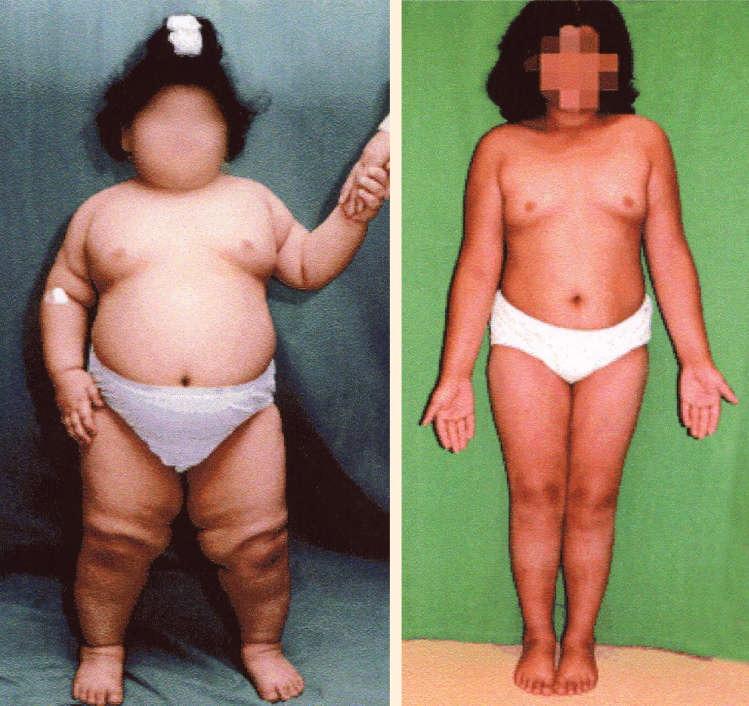
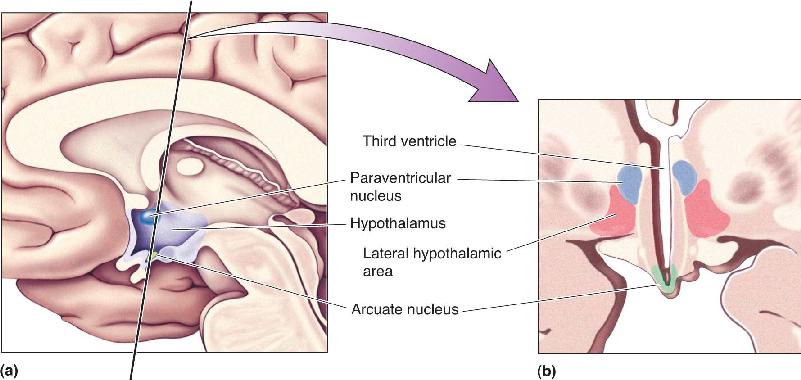
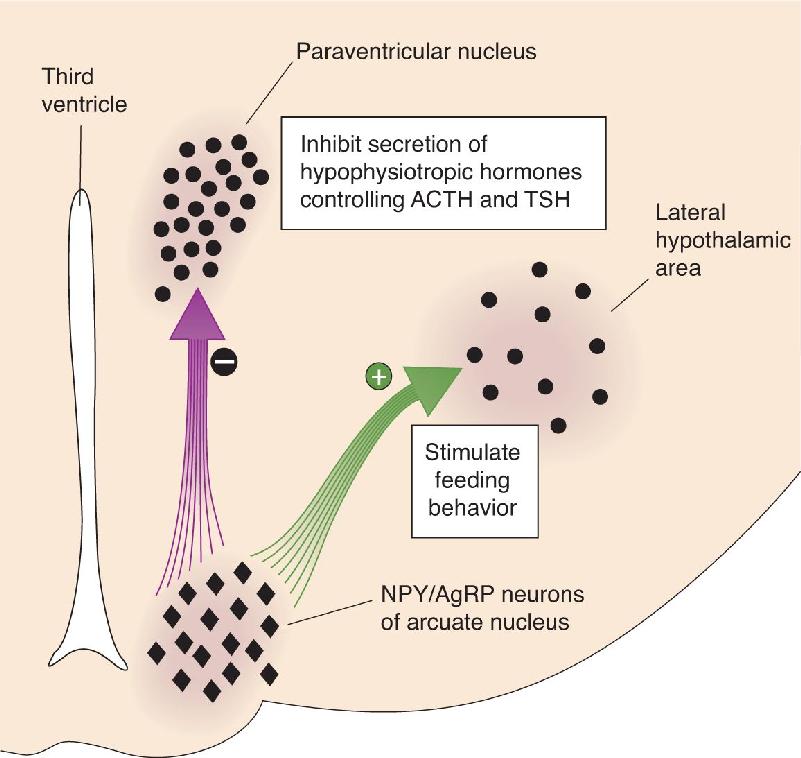
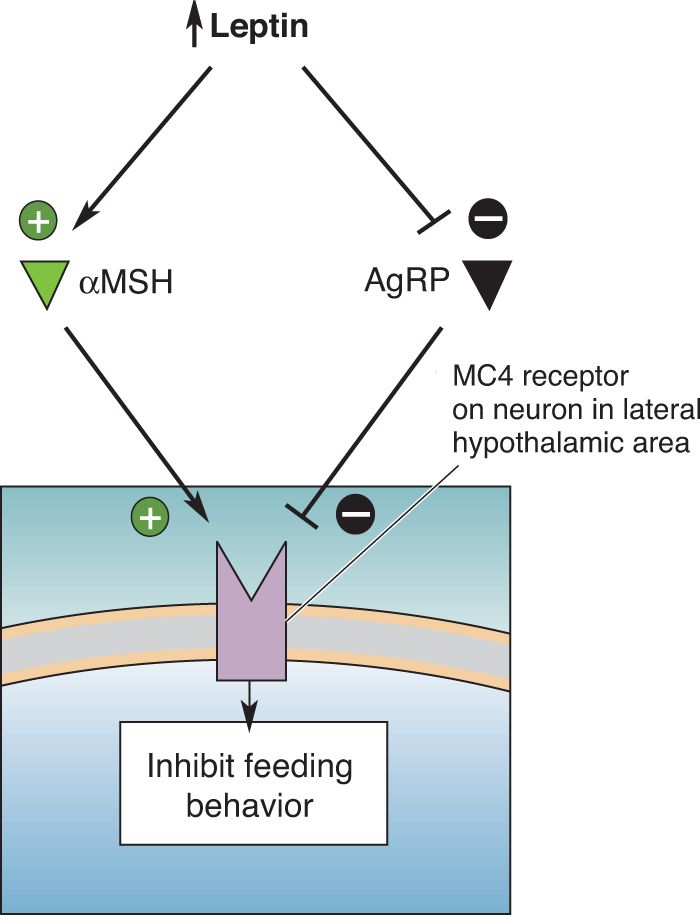
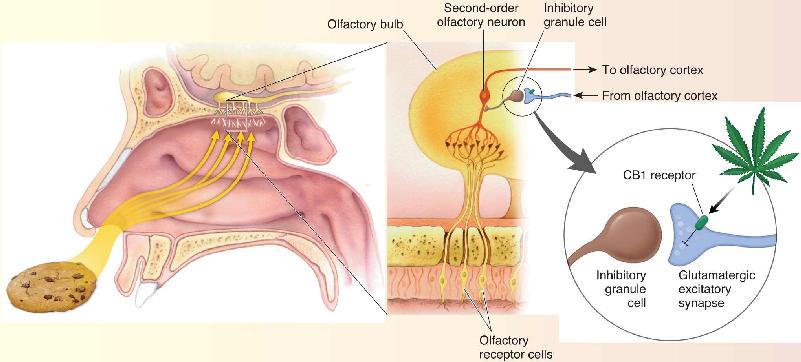
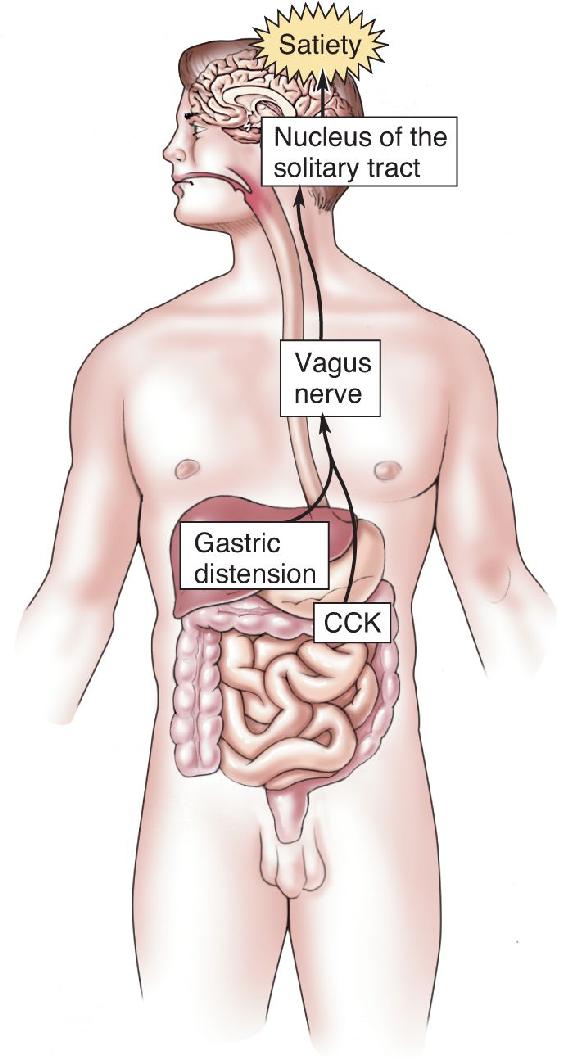
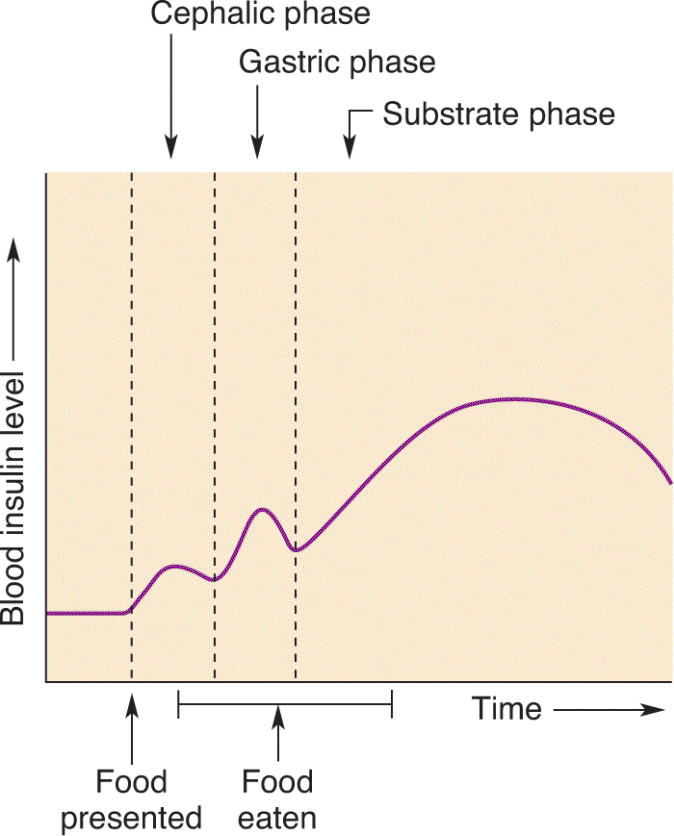
Ch 17Sex and the BrainRead full chapter →
Sections in this chapter
- Sex and the Brain

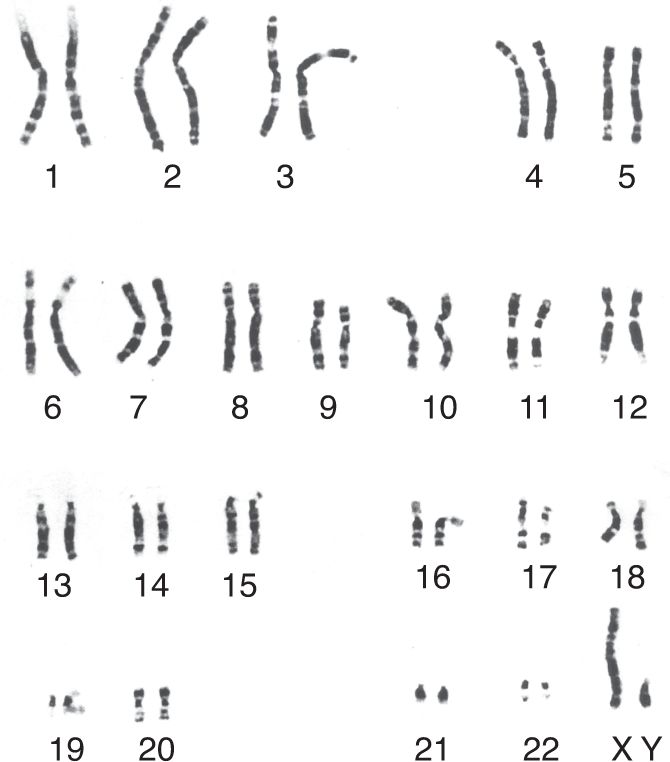
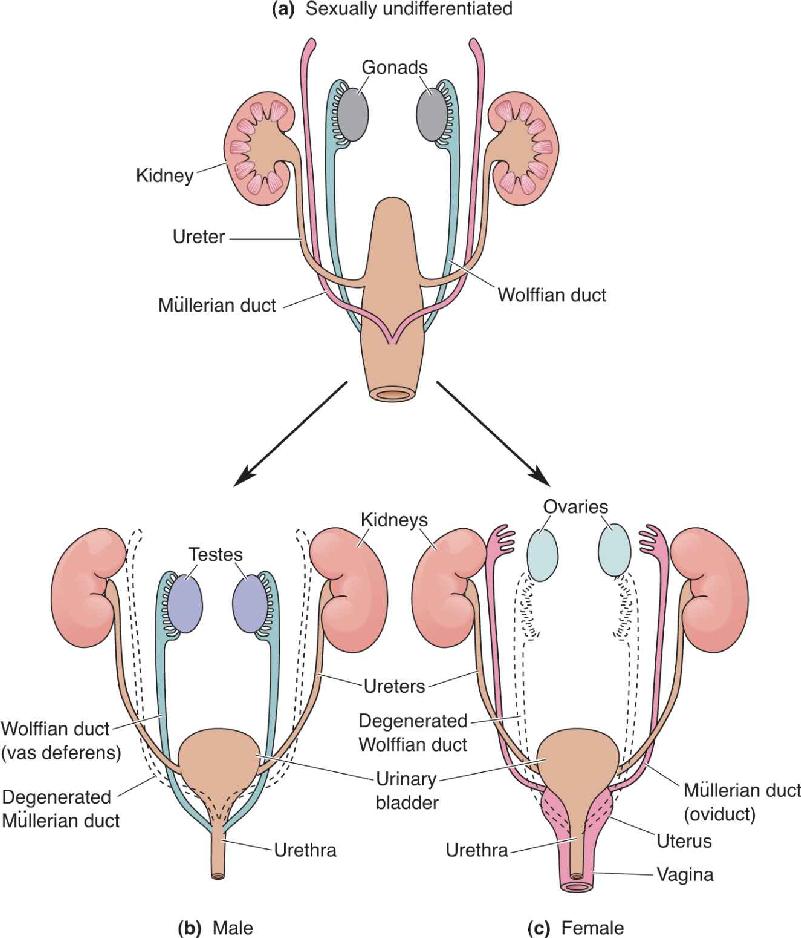
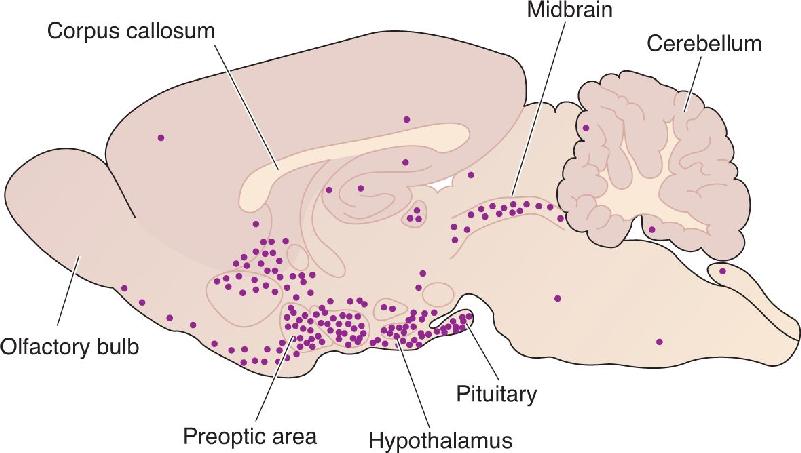
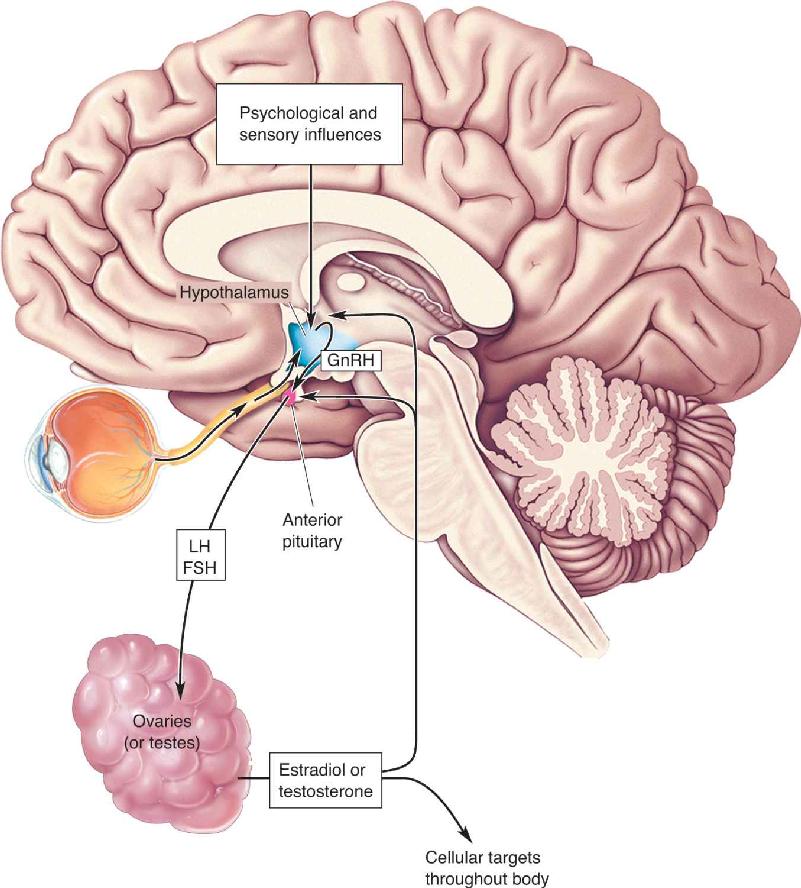


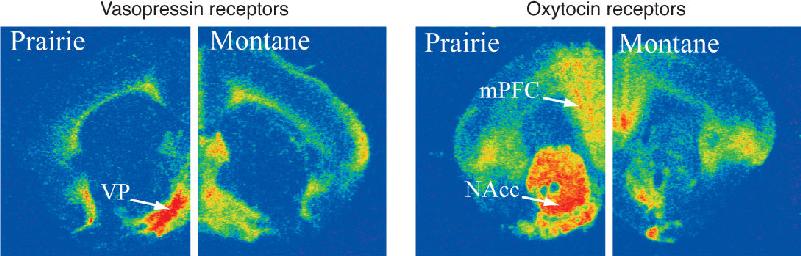
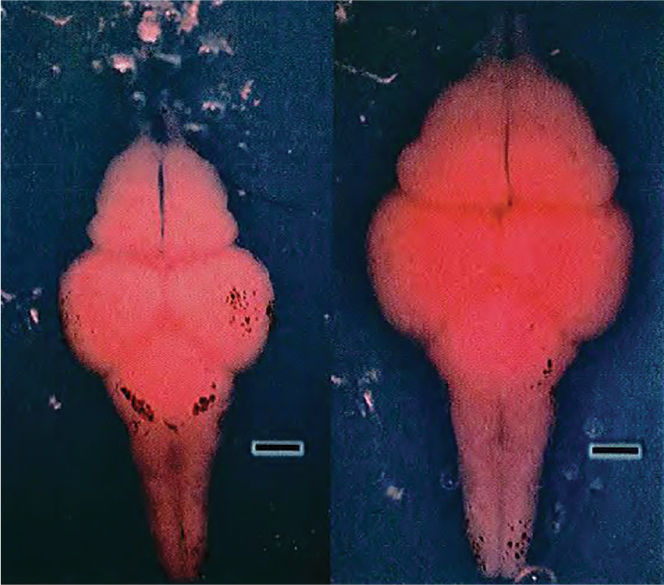
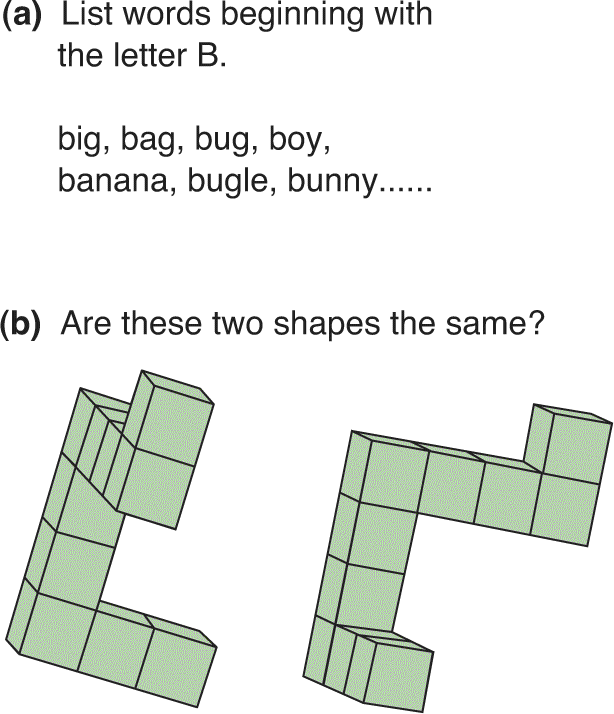
Ch 18Brain Mechanisms of EmotionRead full chapter →
Sections in this chapter
- Brain Mechanisms of Emotion

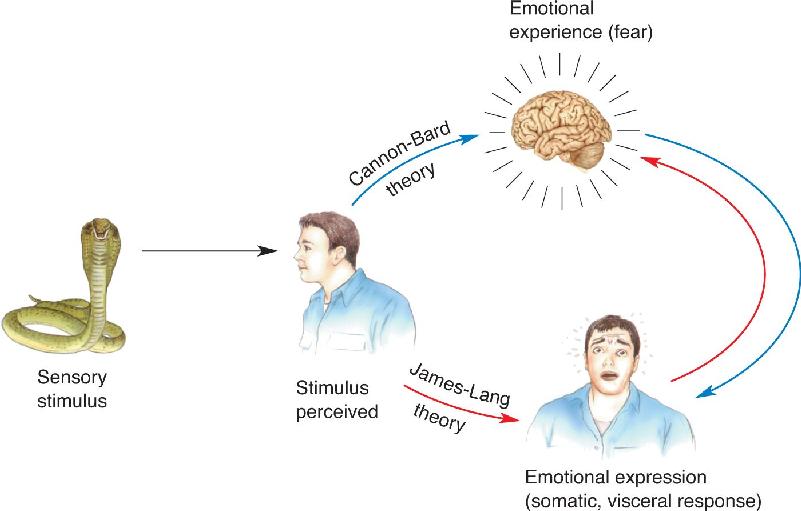
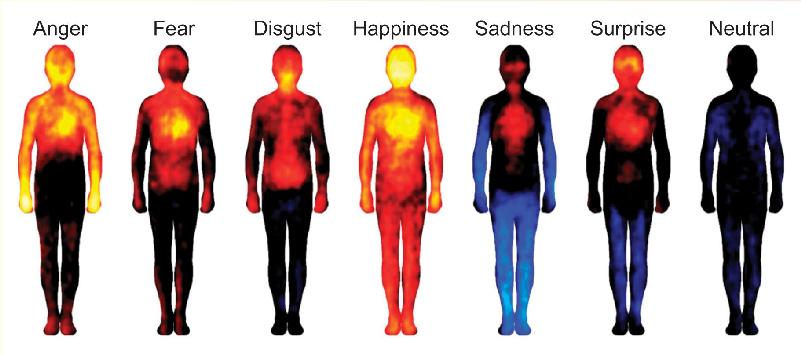
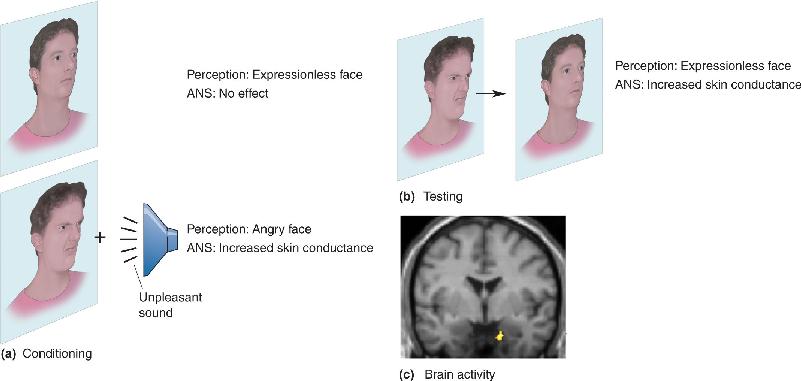
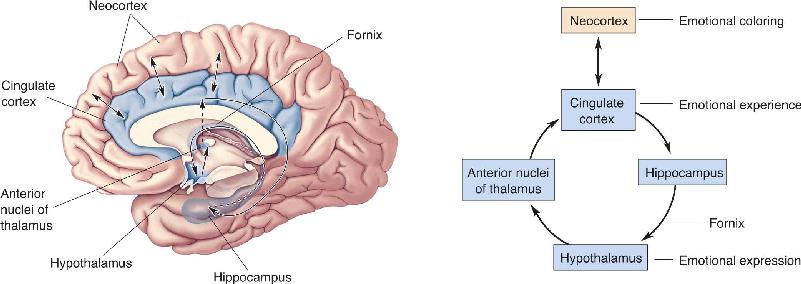


Ch 19Brain Rhythms and SleepRead full chapter →
Sections in this chapter
- Brain Rhythms and Sleep
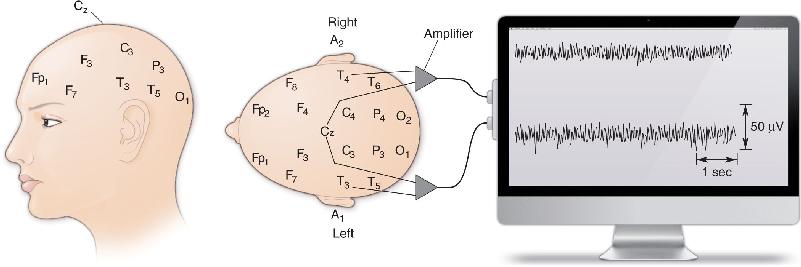
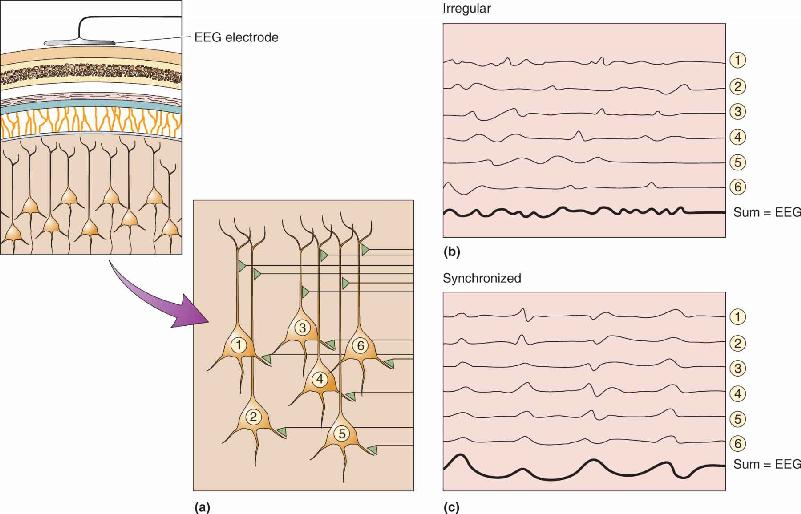

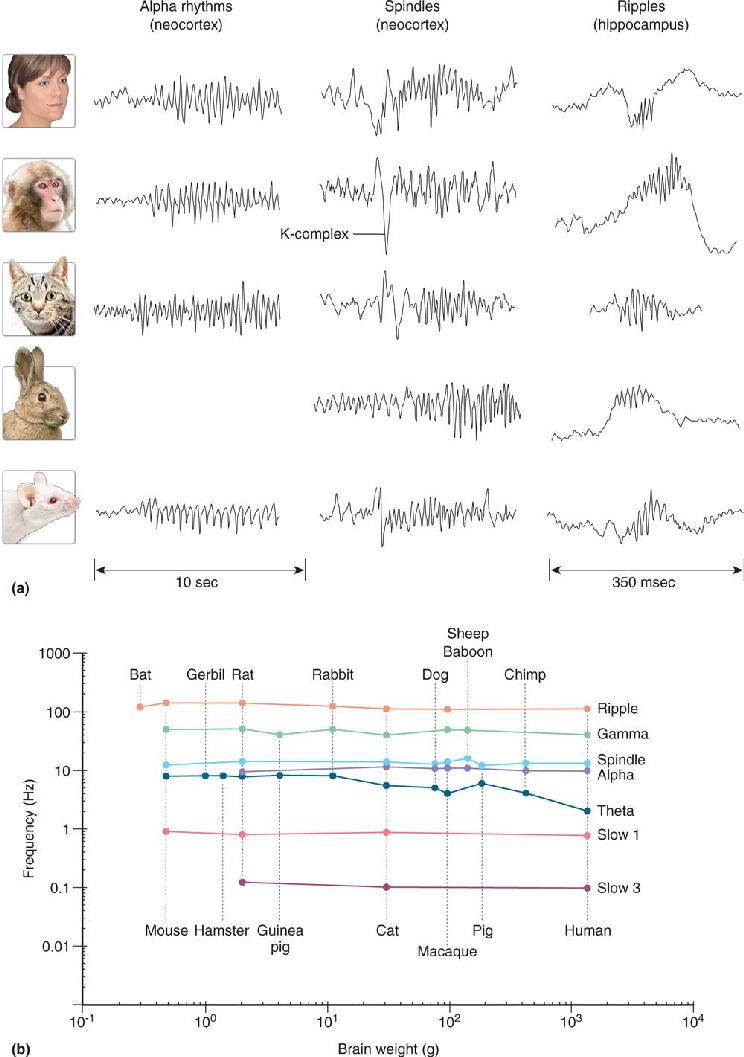
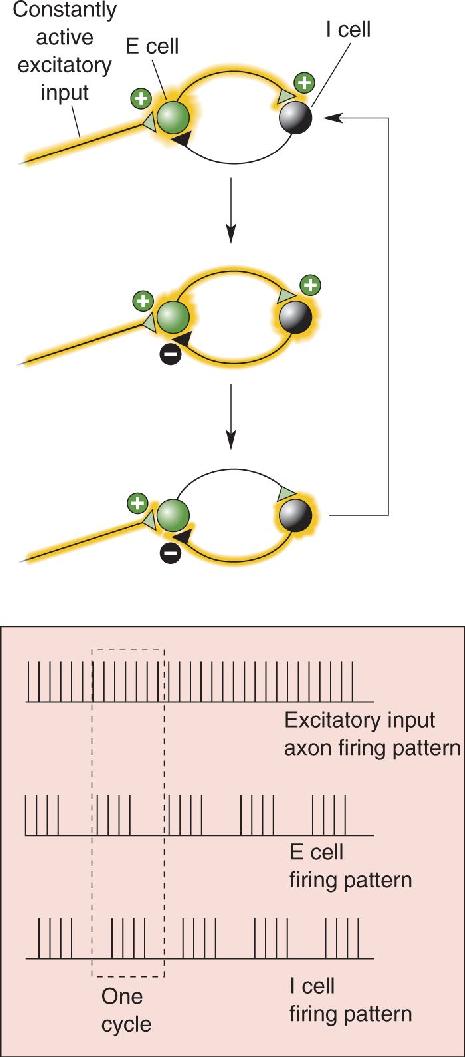
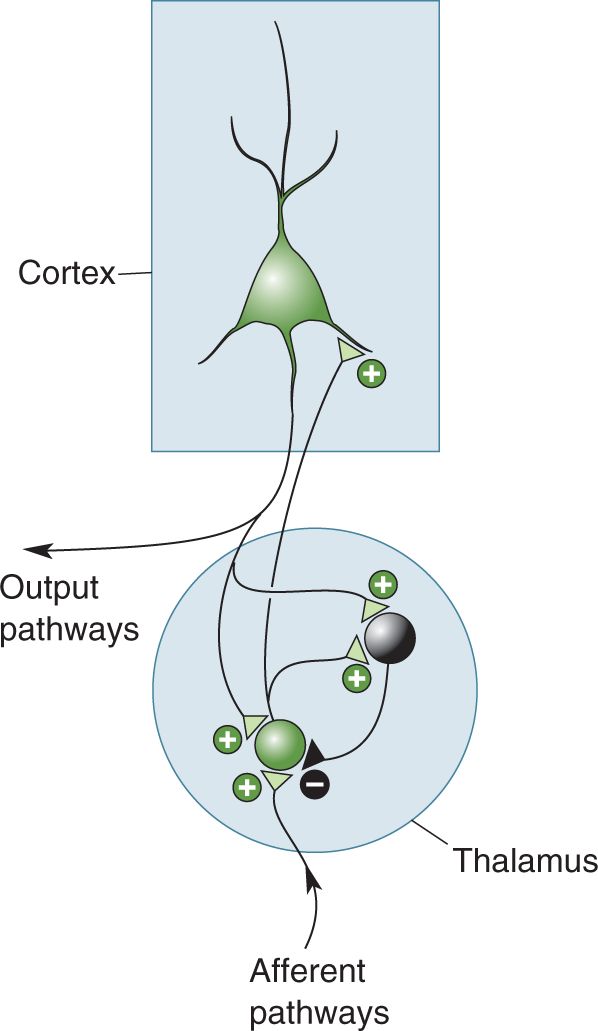
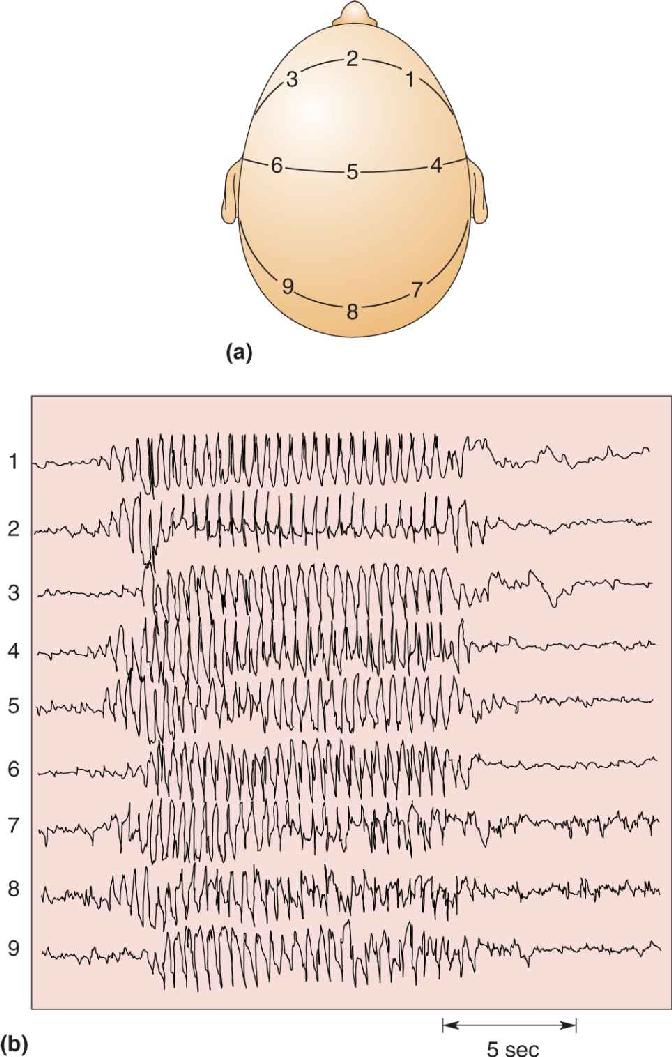
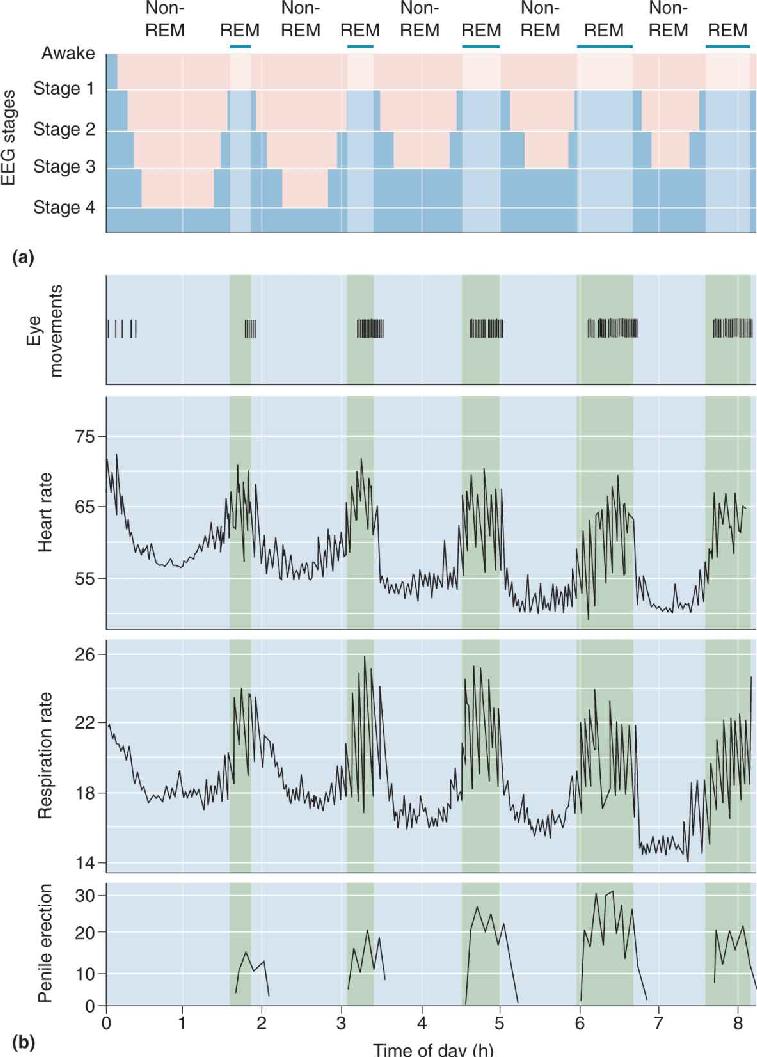
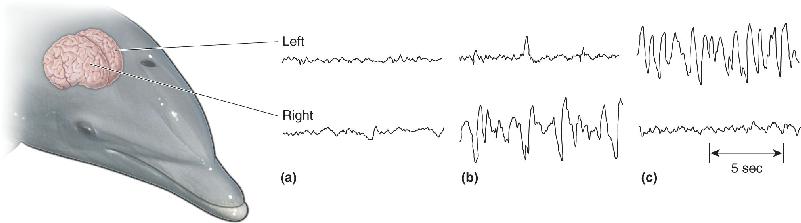
Ch 20LanguageRead full chapter →
Sections in this chapter
- Language
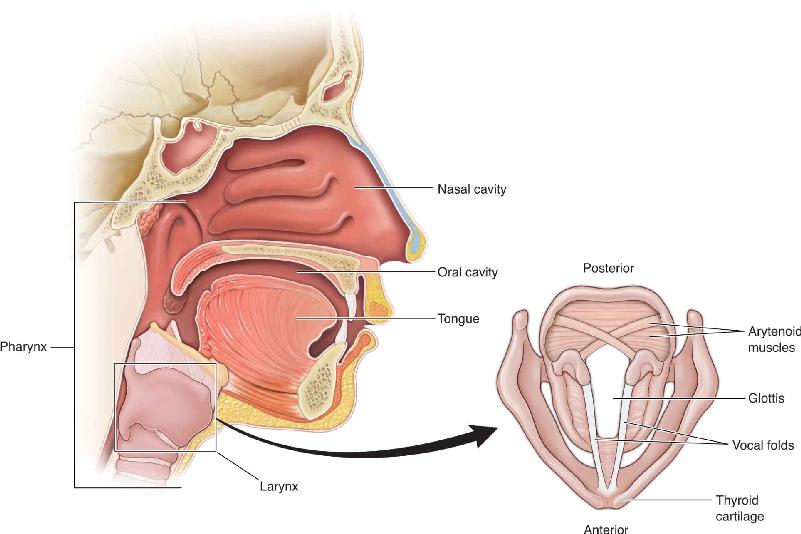

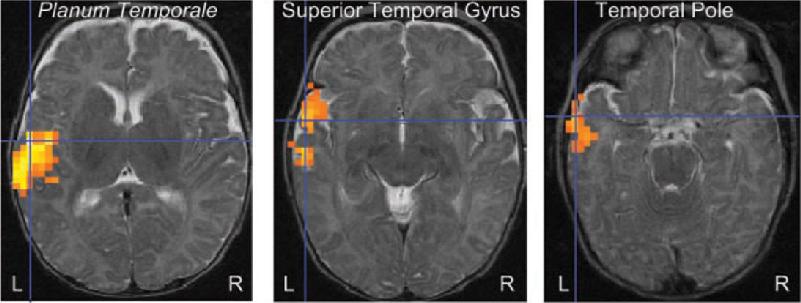
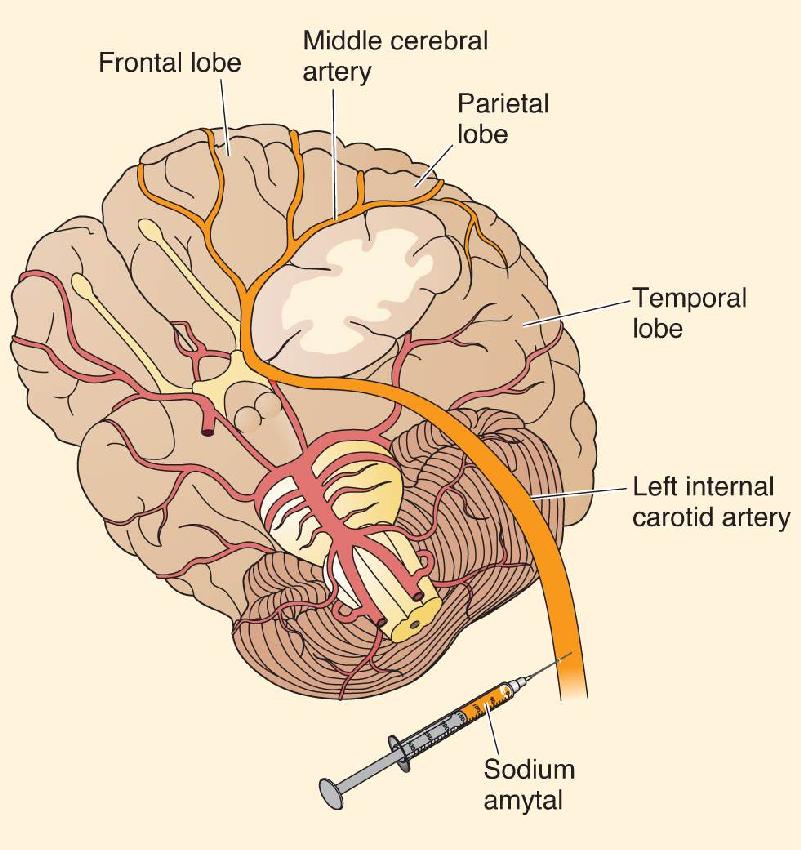
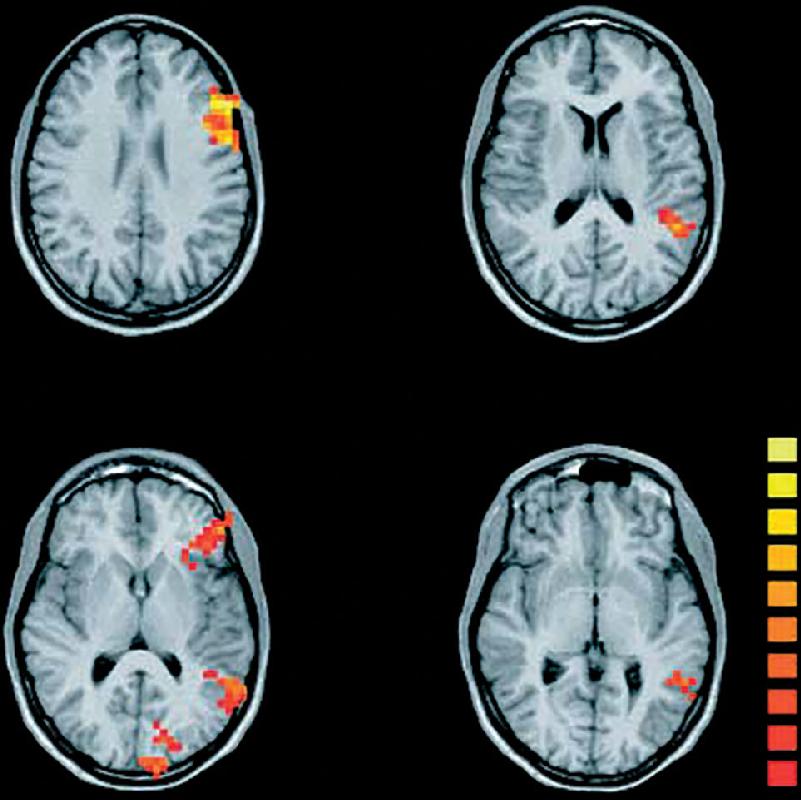
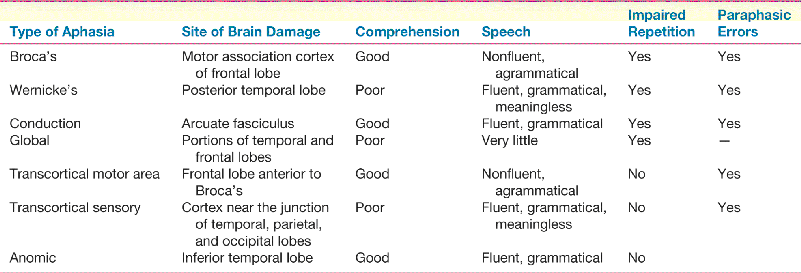
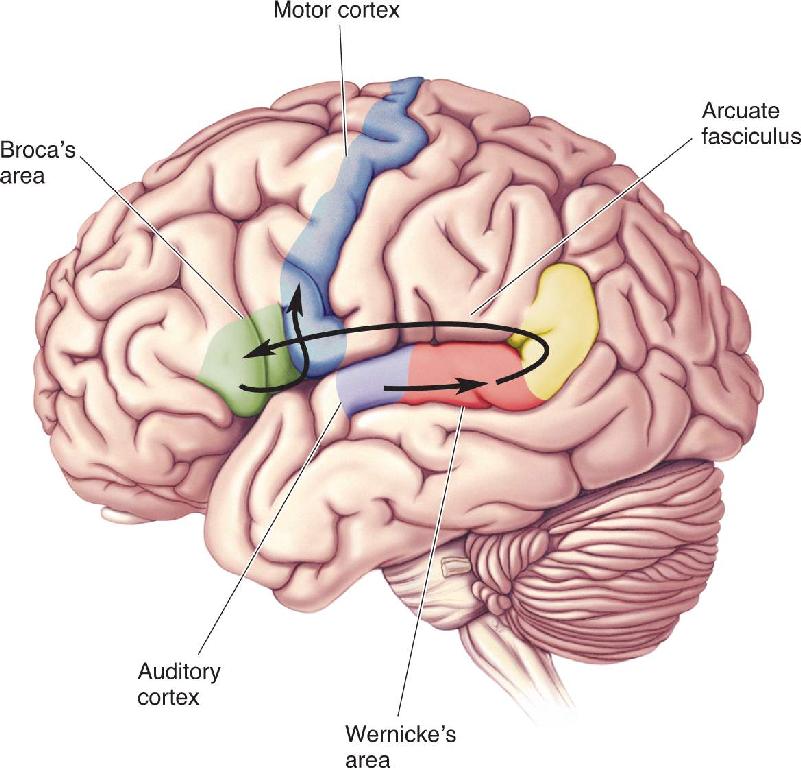
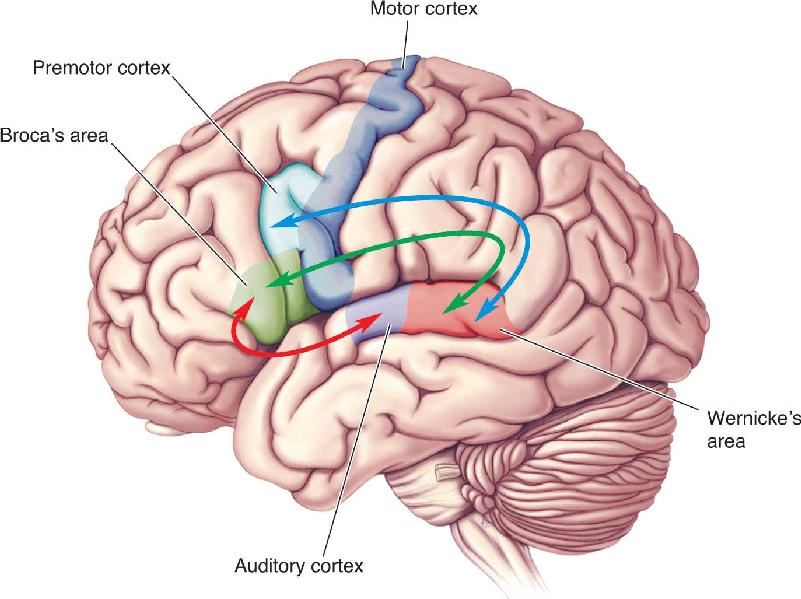
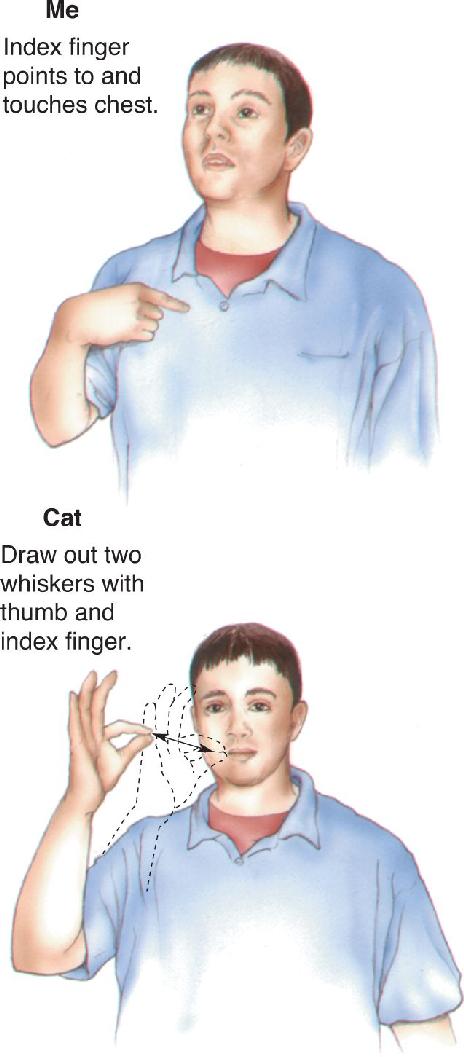
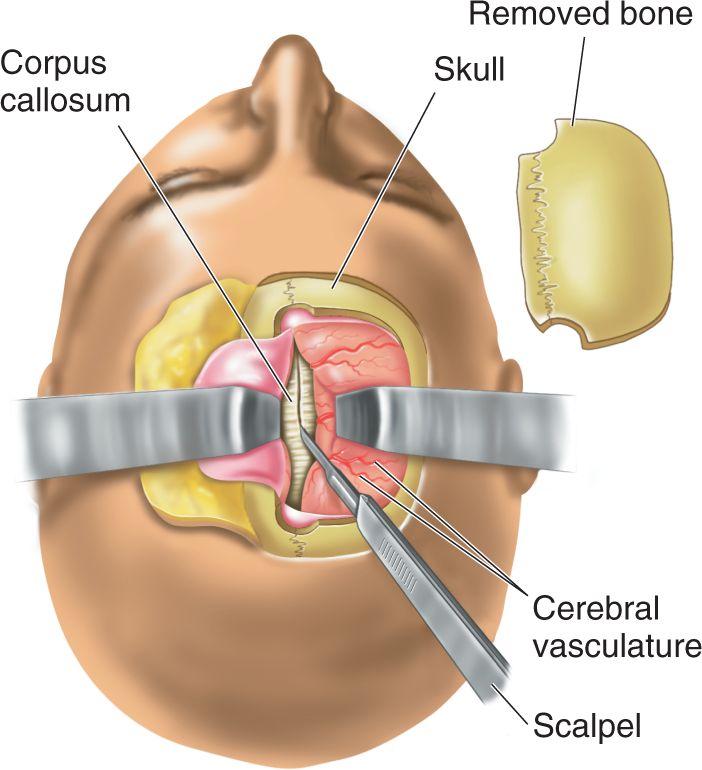
Ch 21-One The Resting Brain, Attention, and ConsciousnessRead full chapter →
Sections in this chapter
- The Resting Brain, Attention, and Consciousness
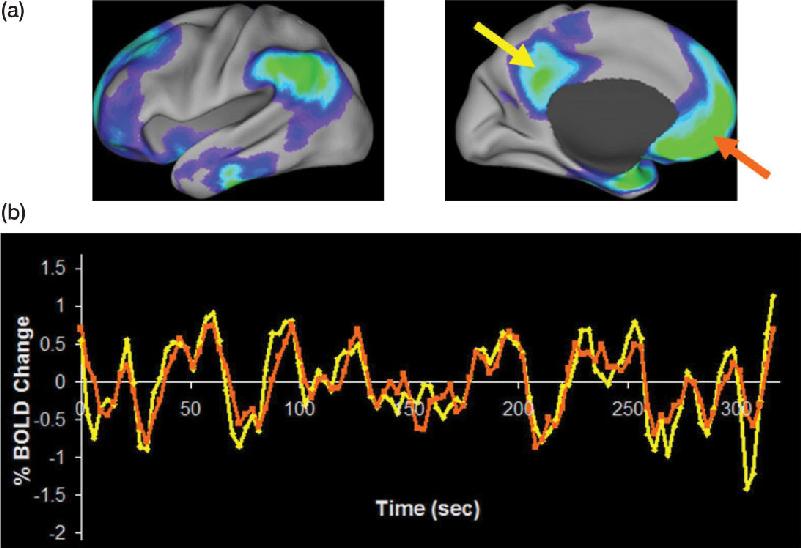
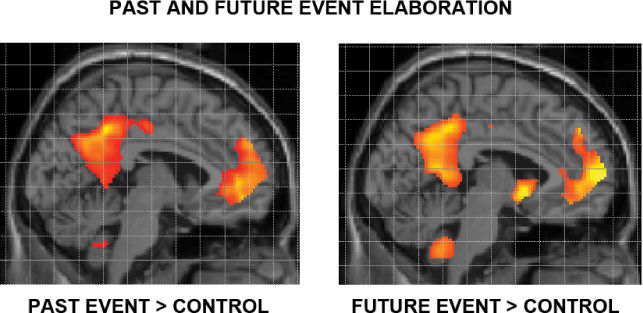
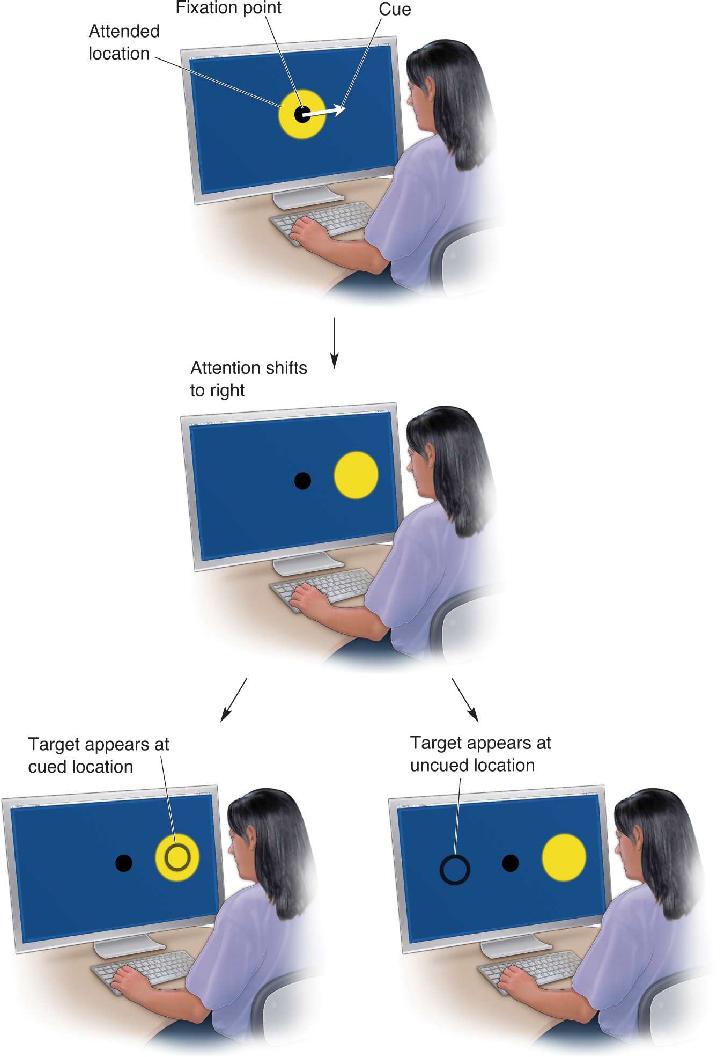
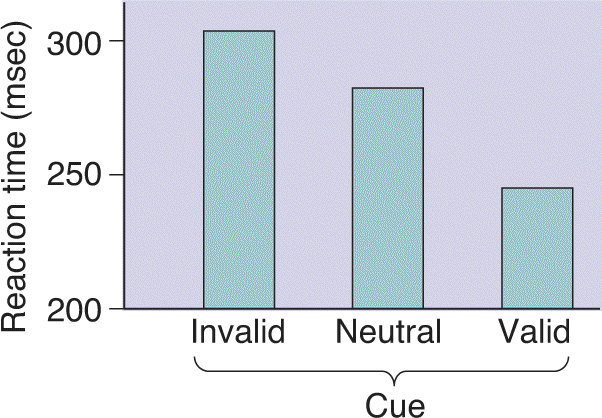
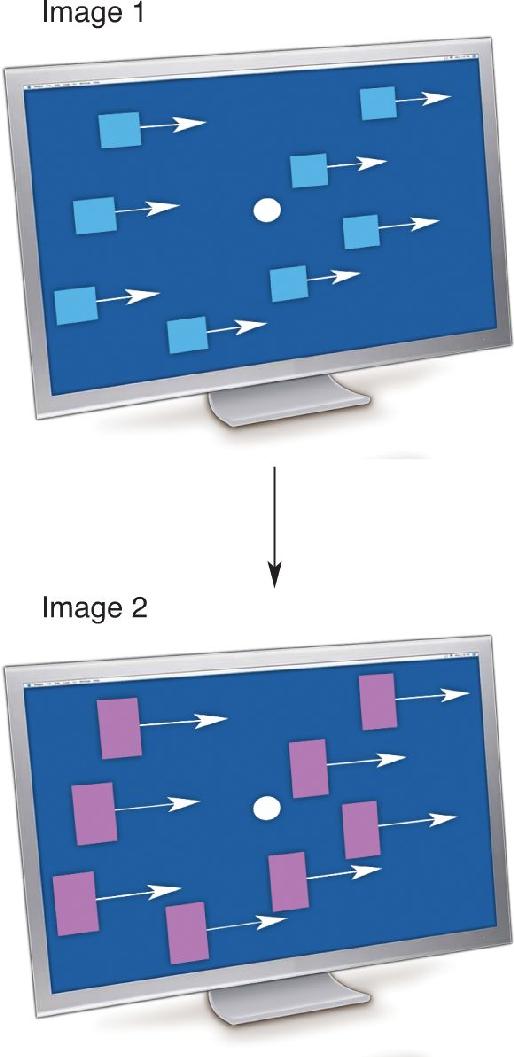
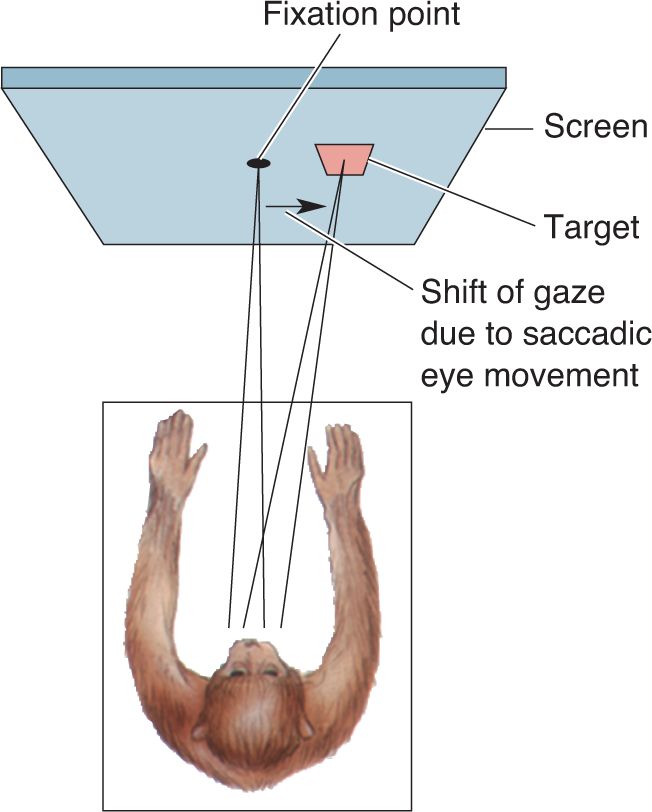
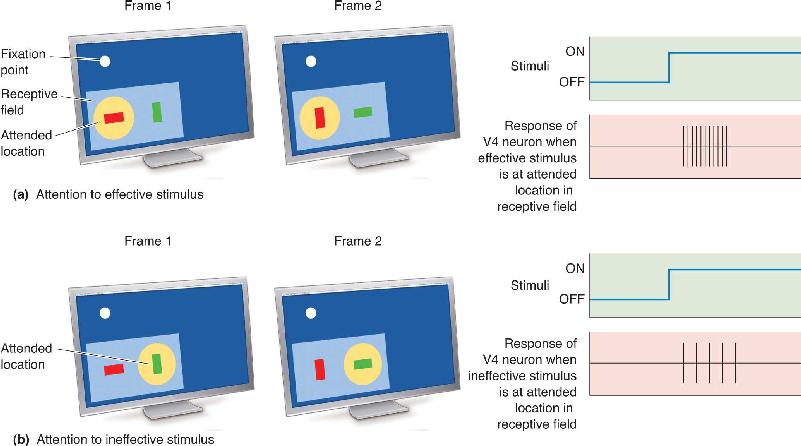
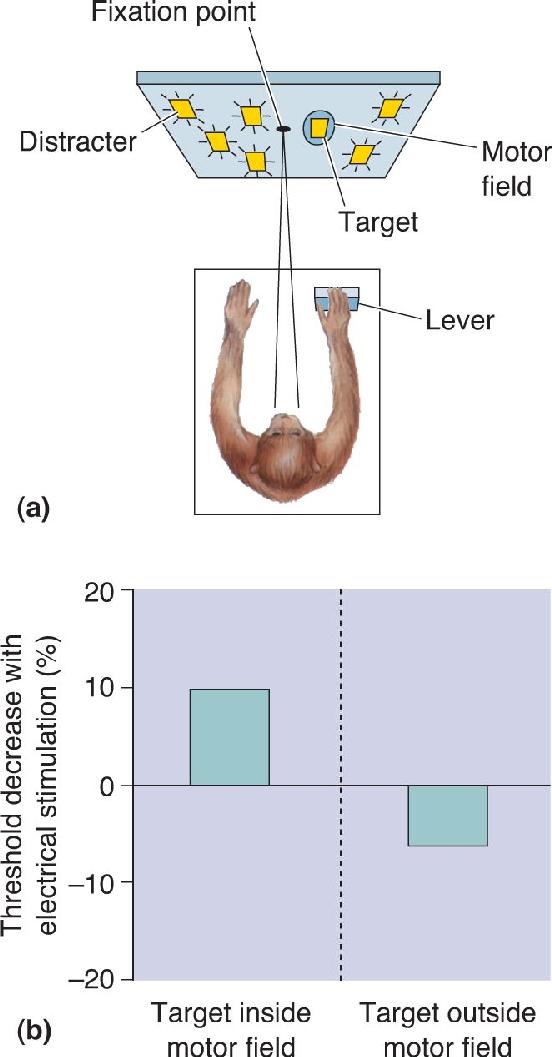
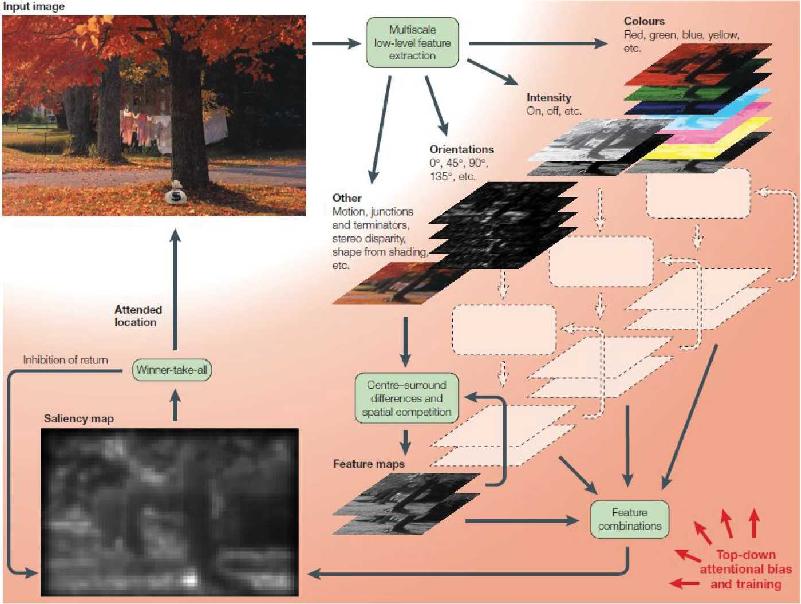
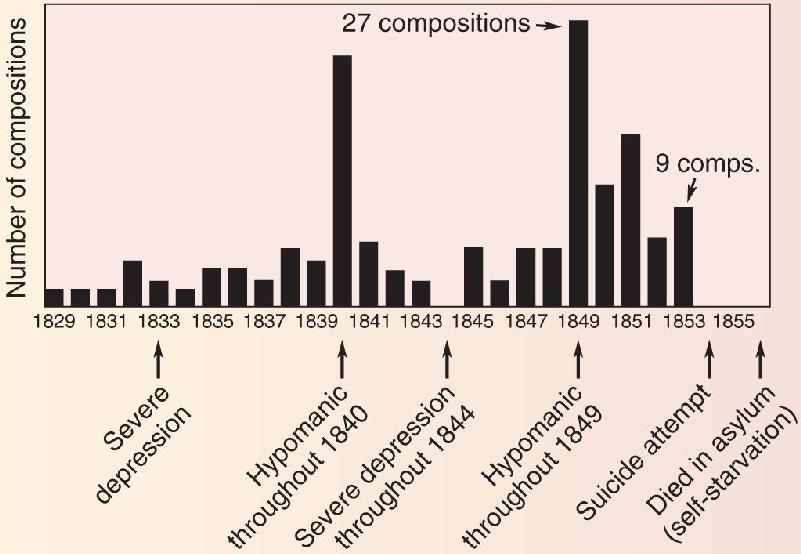
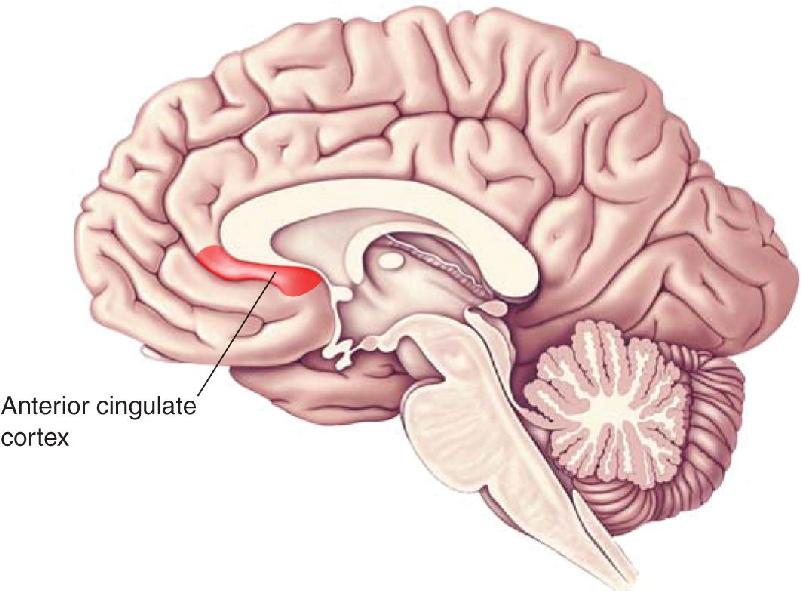
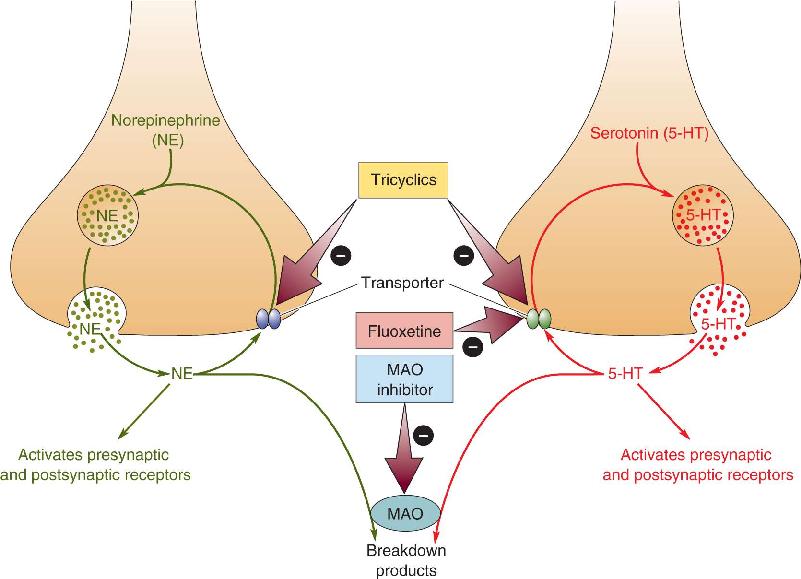
Ch 22-Two Mental IllnessRead full chapter →
Sections in this chapter
- Mental Illness

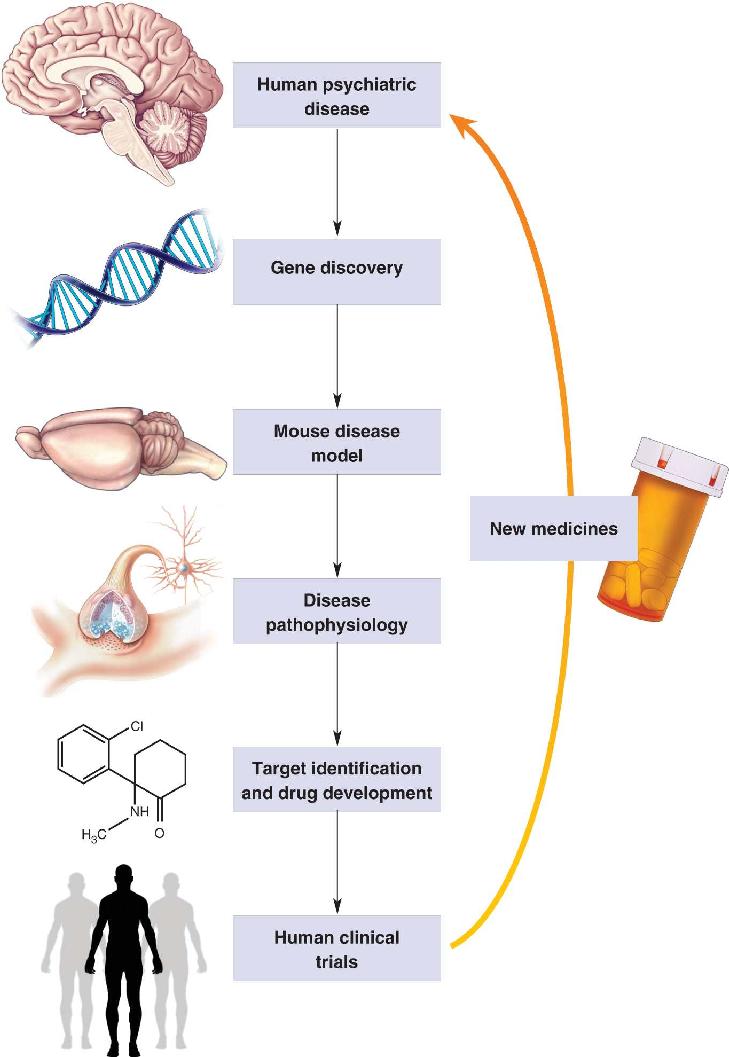
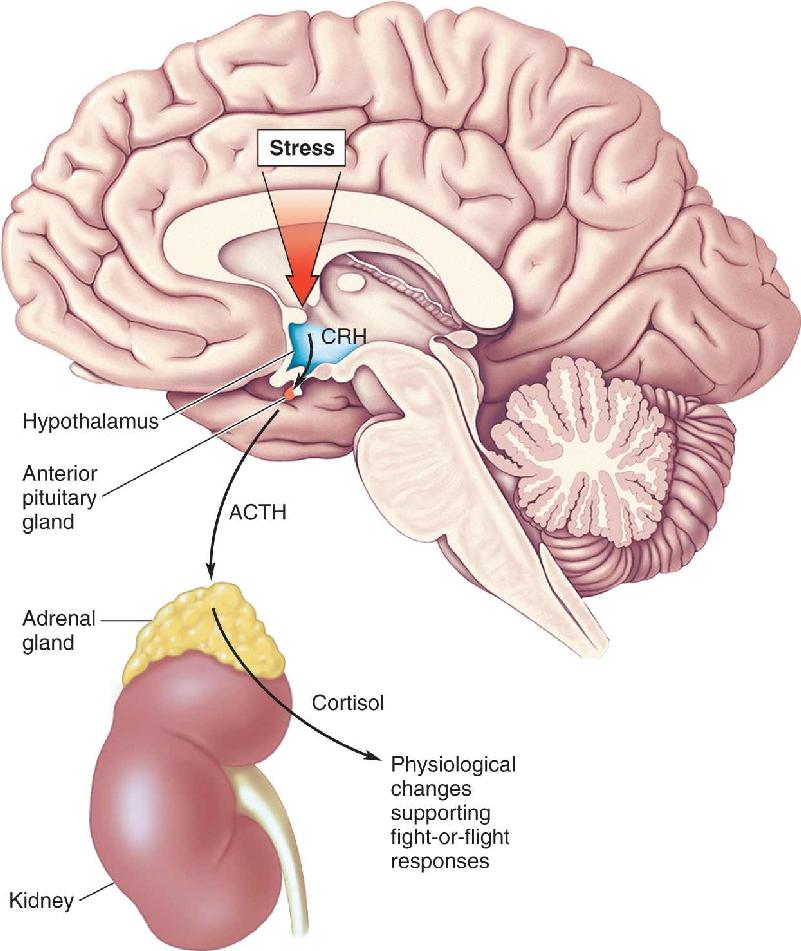
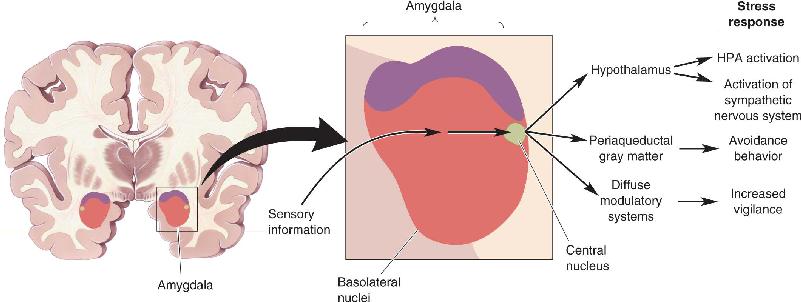
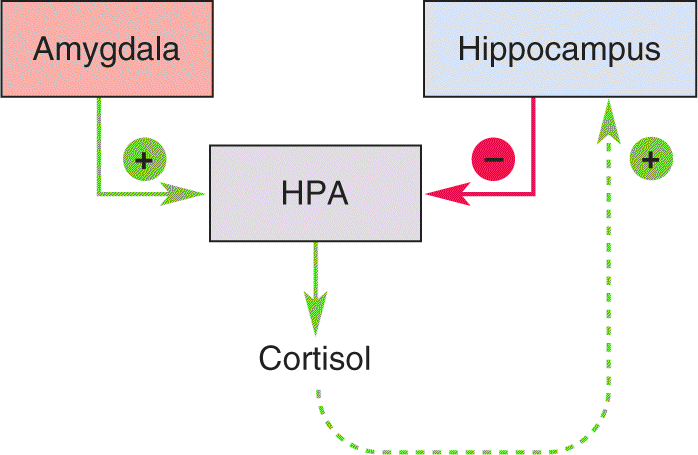
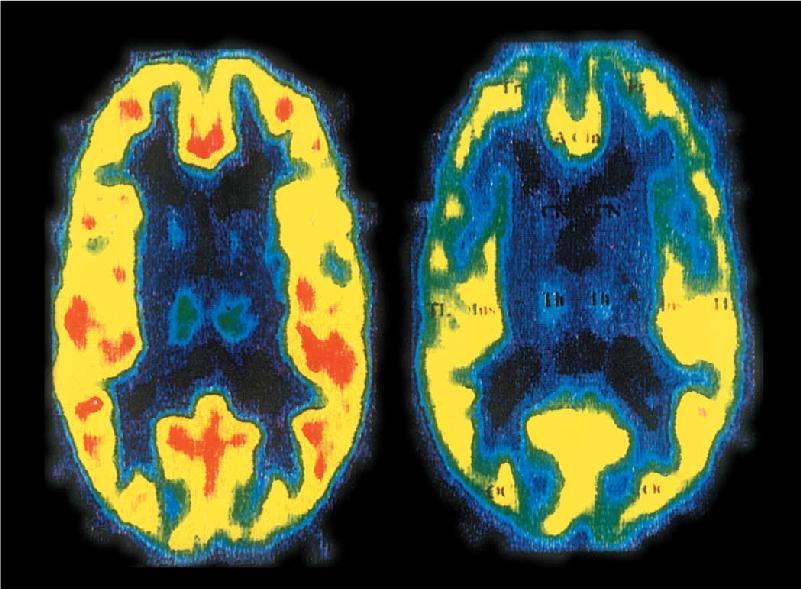

Ch 23-Three Wiring the BrainRead full chapter →
Sections in this chapter
- Wiring the Brain
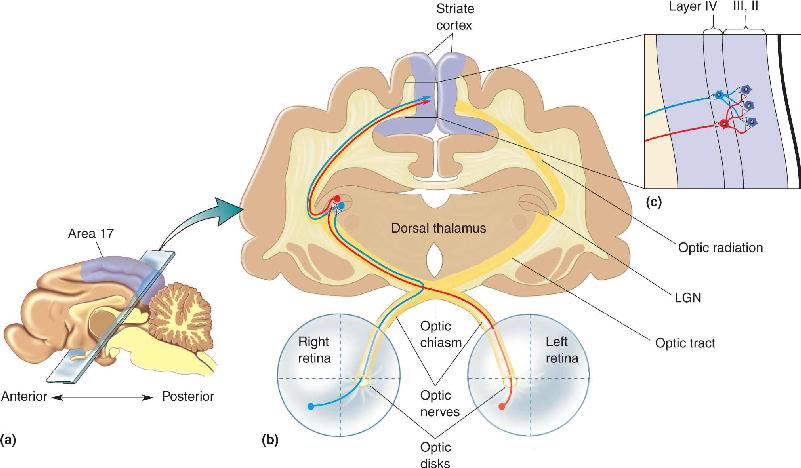
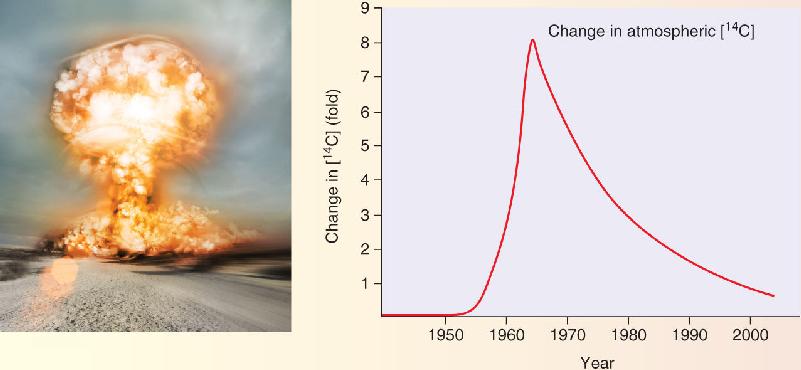
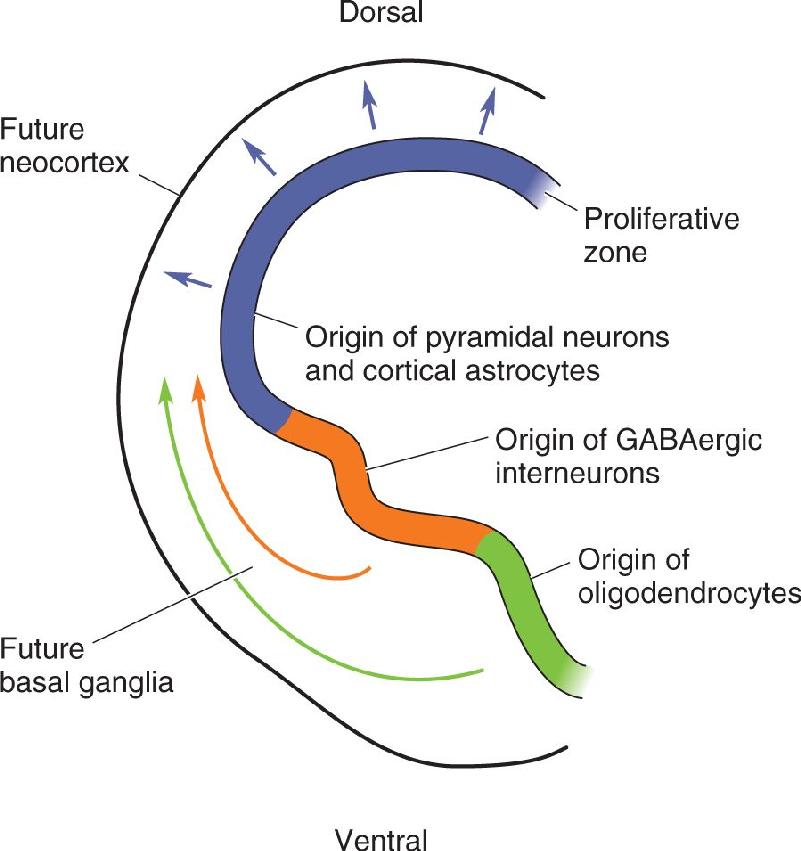
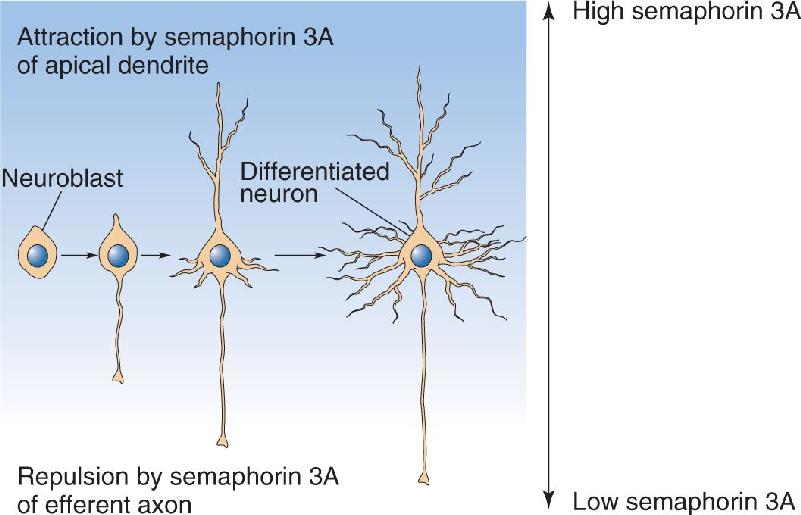
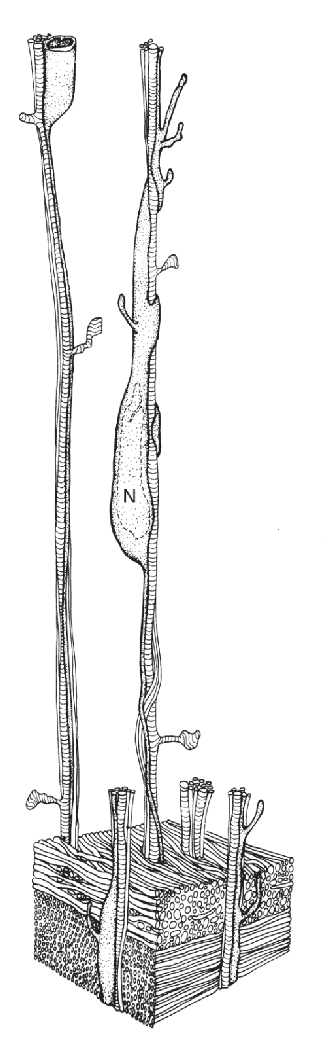
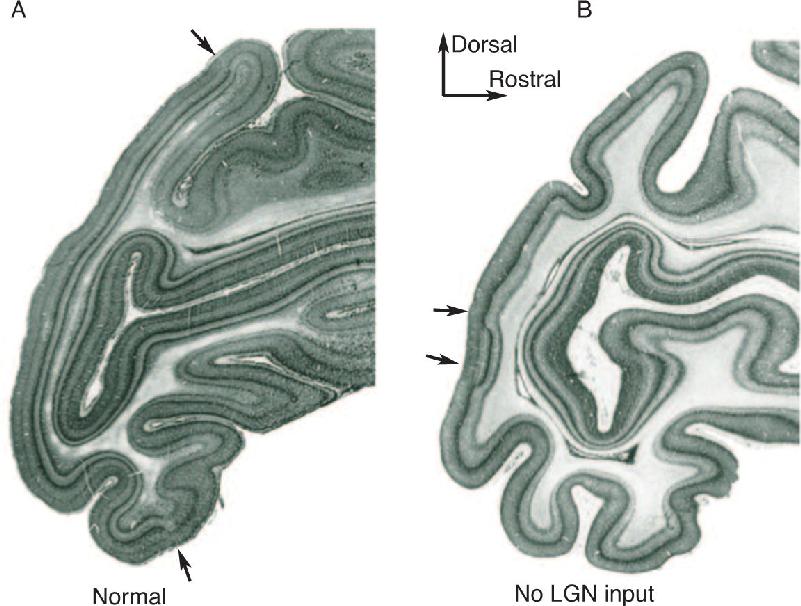
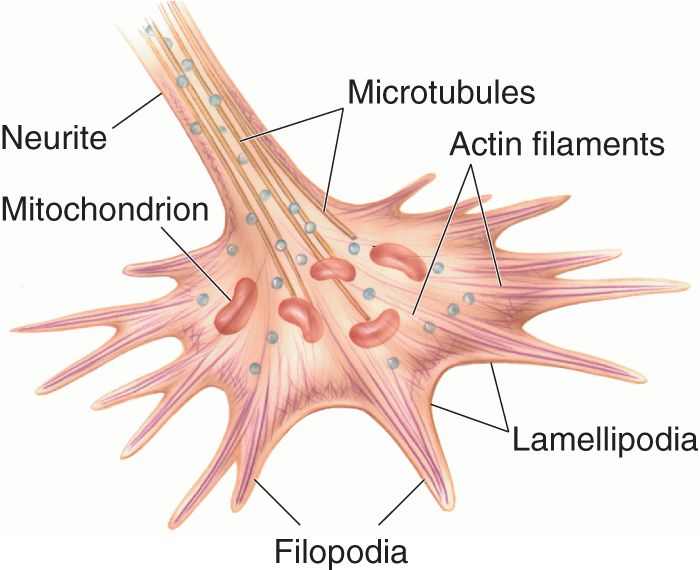
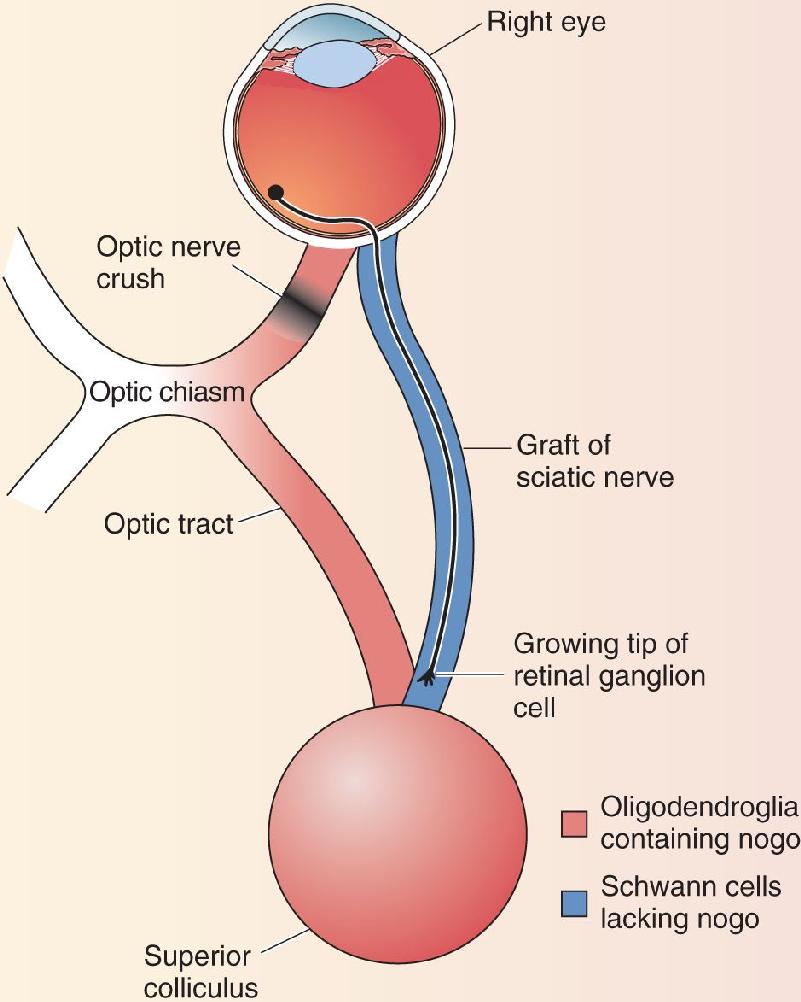
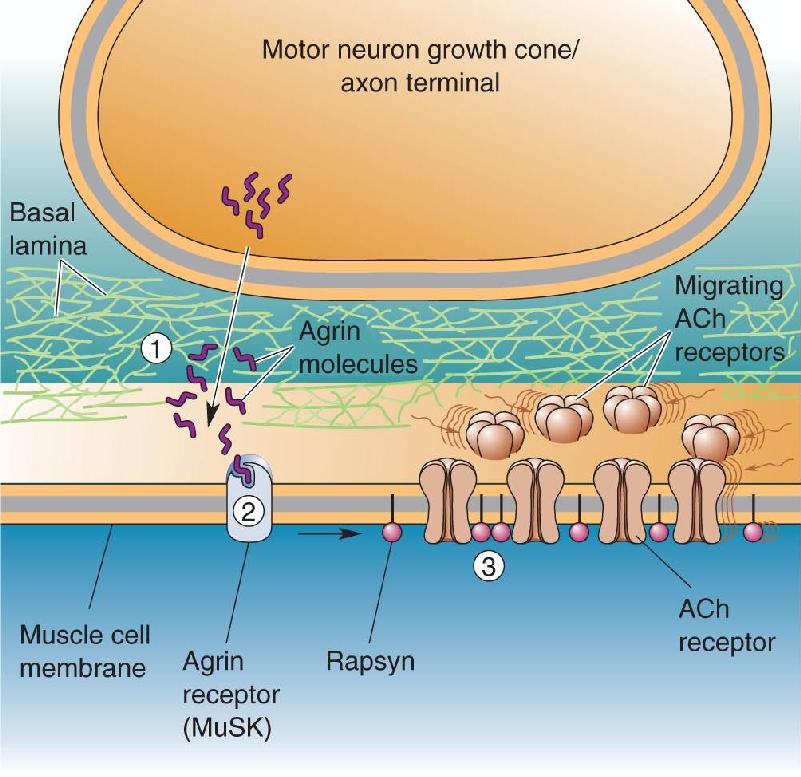
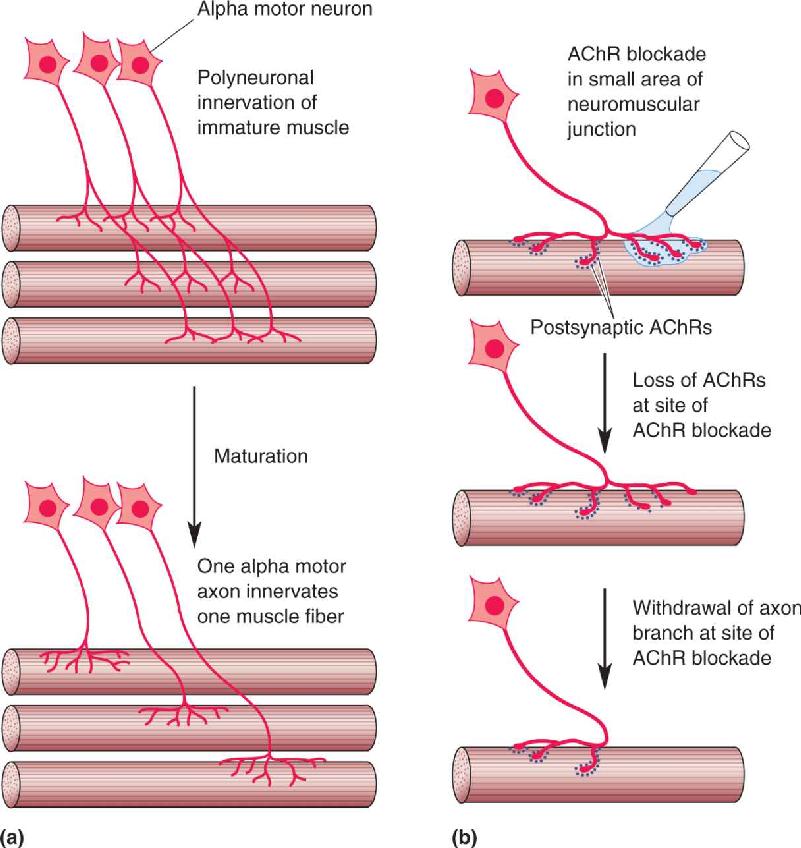
Ch 24-Four Memory SystemsRead full chapter →
Sections in this chapter
- Memory Systems
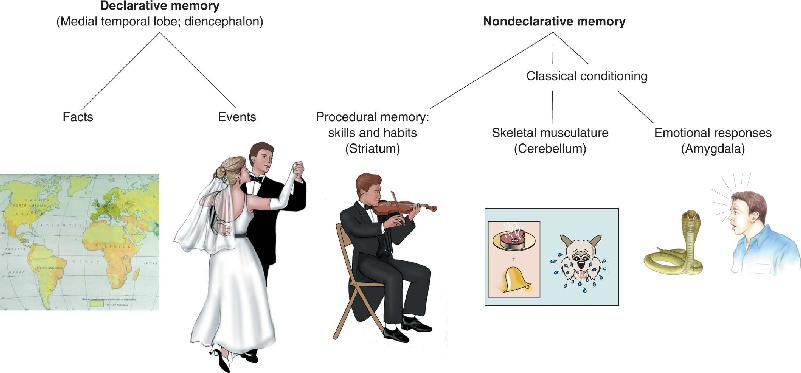
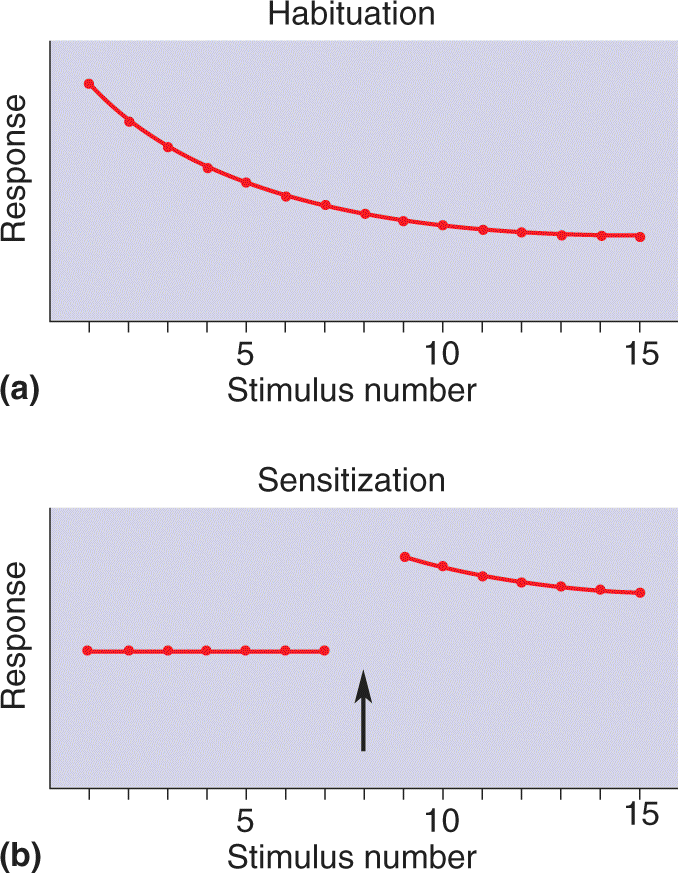
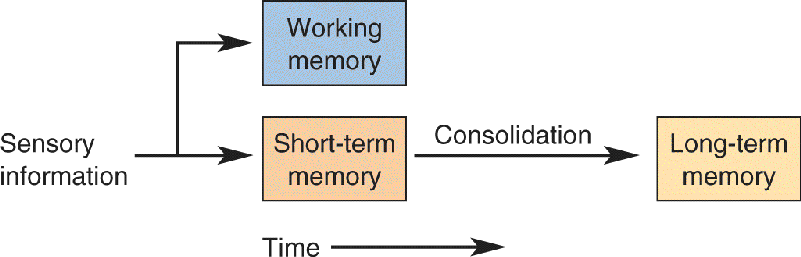
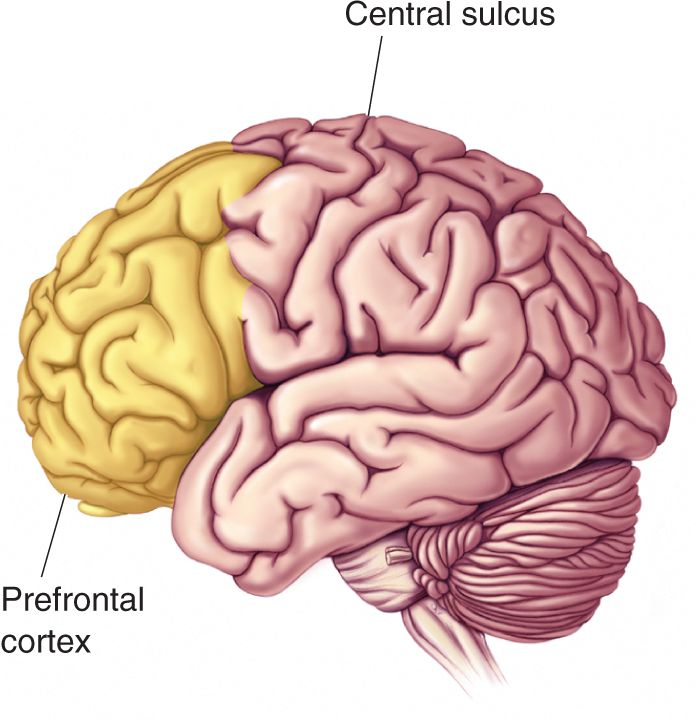
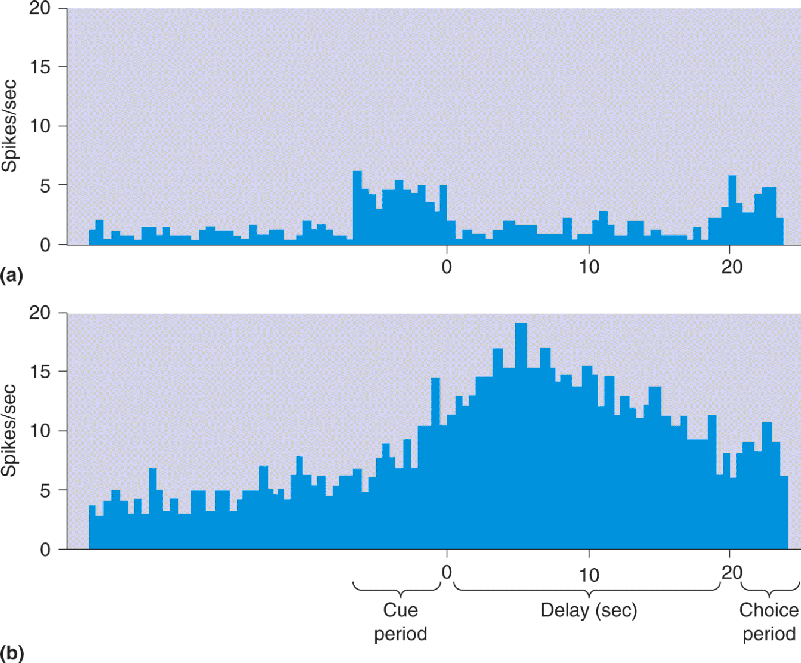
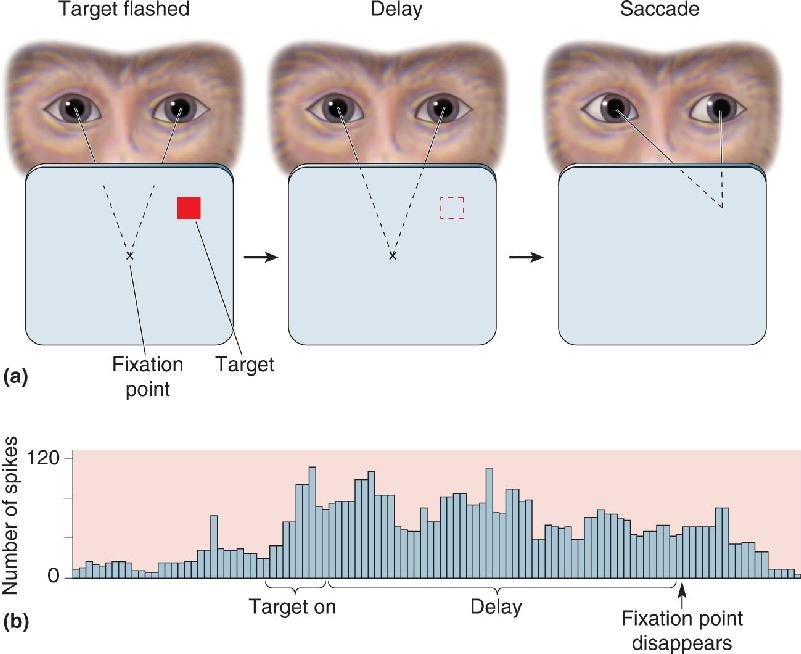
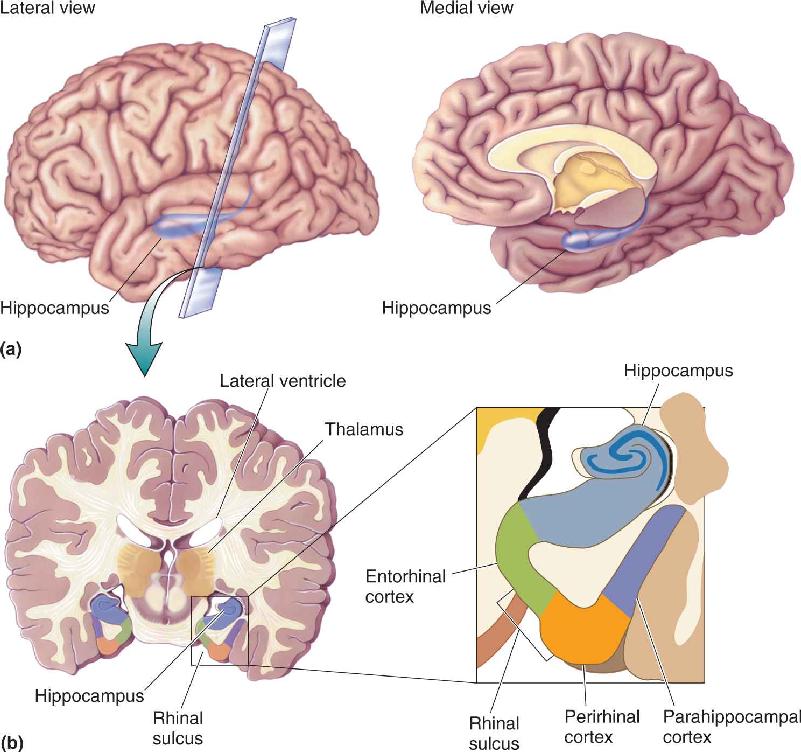
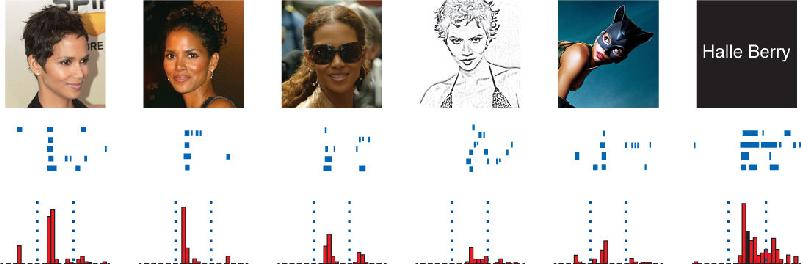
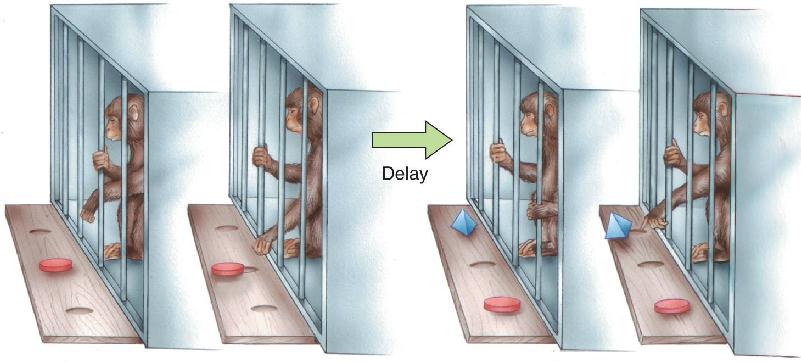
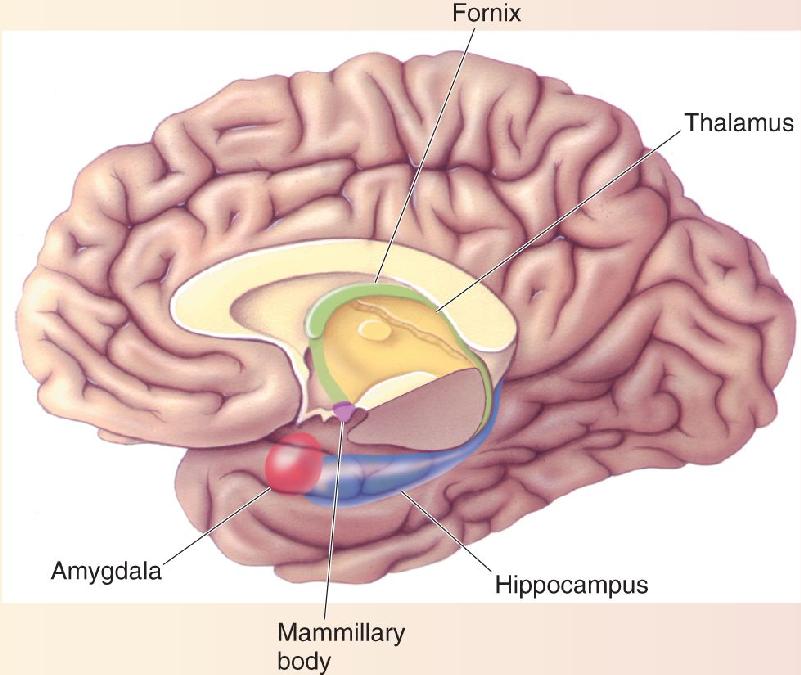
Ch 25-five Molecular Mechanisms of Learning and MemoryRead full chapter →
Sections in this chapter
- Molecular Mechanisms of Learning and Memory