Introduction
Chapter 1 Evolution: Molecules, Genes, Cells, and Organisms Tardigrades are tiny -long invertebrates that have evolved to survive exposures to extreme conditions that kill other life forms; they are the only metazoan organisms known to survive exposure to the deadly combination of low pressure and intense radiation in outer space. [Schokraie E., Warnken U., Hotz-Wagenblatt A., Grohme M. A., Hengherr S., et al. 2012. Comparative proteome analysis of Milnesium tardigradum in early embryonic state versus

adults in active and anhydrobiotic state. PLoS ONE 7(9): e45682. https://doi.org./10.1371/journal.pone.0045682]
1.4 Unicellular Eukaryotic Organisms Widely Used in Cell Biology Research
1.5 Metazoan Structure, Function, Evolution, and Differentiation
1.6 Metazoan Organisms Widely Used in Cell Biology Research Nothing in biology makes sense except in the light of evolution. — Theodosius Dobzhansky, 1973, essay in American Biology Teacher 35:125–129 Biology is a science fundamentally different from physics or chemistry, which deal with unchanging properties of matter that can be described by mathematical equations. Biological systems, of course, follow the rules of chemistry and physics, but biology is a historical science, as the forms and structures of the living world today are the results of billions of years of evolution. Through evolution, all organisms both living and extinct are related in an evolutionary family tree extending from primitive singlecelled organisms that lived in the distant past to the diverse plants, animals, and microorganisms of the present era (Figure 1-1, Table 1-1).

The great insight of Charles Darwin was the principle of natural selection: organisms vary randomly and compete within their environment for resources. Only those that survive and reproduce are able to pass down their genetic traits (Figure 1-2).
FIGURE 1-1 All living organisms descended from a common ancestral cell. All organisms, from simple bacteria to complex mammals, probably evolved from a common single-celled ancestor. This evolutionary tree depicts the relationships among the three major lineages of organisms. The structure of the tree was initially ascertained from morphological criteria: creatures that look alike were put close together. More recently, the sequences of DNA and proteins found in organisms have provided more information-rich criteria for assigning relationships. The greater the similarities in these macromolecular sequences, the more closely related organisms are thought to be. The trees based on morphological comparisons and the fossil record generally agree well with those based on molecular data. This evolutionary tree has branches. Organisms like bacteria exchange pieces of their DNA genome with one another, and mitochondria and chloroplasts are thought to derive from primitive bacteria that took up residence inside the organisms that gave rise to present eukaryotes.
[Data from J. R. Brown, 2005, “Universal Tree of Life,” in Encyclopedia of Life Sciences, Wiley InterScience (online).] Description The tree shows three main branches. The common ancestor for all three groups is presumed to be the last common ancestor of all extant organisms. The common ancestor for Archaea and Eukaryota is presumed to be the last common ancestor of eukaryotes and archaebacteria. The sub-branches from the first main branch correspond to the following names: Aquifex, Thermus, Cyanobacteria, Bacteroides, Flexibacter, Fusobacteria, Spirochaetes, gamma slash beta purples, alpha purples, delta slash epsilon purples, high G plus C gram positives, low G plus C gram positives and Thermotogales. All these are collectively labeled Eubacteria. The sub-branches from the second main branch correspond to the following names: Korarchaeota, Euryarchaeota, and Crenarchaeota. All these are collectively labeled Archaea. The sub-branches from the third main branch correspond to the following names: Metamonda (example- Giardia), Parabasalia (example- Trichomonas), Kinetoplasta (exampleTrypanosoma), Euglena, Apicomplexa (example- Plasmodium), Slime molds, Entamoeba, plants, animals, fungi, and Microsporidia. All these are collectively labeled Eukaryota. A curved arrow extending from alpha purples of Eubacteria, passes through Archaea, and ends at Metamonda of Eukaryota that reads, Mitochondria. A dashed arrow pointing from cyanobacteria to a node that branches into Euglena and Kinetoplasta reads, Chloroplasts.
TABLE 1-1 • Timeline for Evolution of Life on Earth, as Determined from the Fossil Record and from Recent DNA Sequence Analyses
million years ago The planet Earth forms from material revolving around the young sun. million years ago Cells resembling prokaryotes appear. These first organisms are chemoautotrophs: they use carbon dioxide as a carbon source and oxidize inorganic materials to extract energy. million years ago Photosynthesizing cyanobacteria evolve; they use water as a reducing agent, thereby producing oxygen as a waste product. million years ago Unicellular eukaryotic cells appear. million years ago Simple multicellular organisms evolve, mostly consisting of cell colonies of limited complexity. 580–500 million years ago Most modern phyla of animals begin to appear in the fossil record during the Cambrian explosion. million years ago Major diversification of living things in the oceans: chordates, arthropods (e.g., trilobites, crustaceans), echinoderms, mollusks, brachiopods, foraminifers, radiolarians, etc. First vertebrates with true bones (jawless fishes) evolve.
million years ago million years ago First primitive plants arise on land. million years ago Earliest dinosaurs (prosauropods) and teleost fishes appear. million years ago Gymnosperm forests dominate the land; herbivores grow to huge sizes. million years ago First mammals evolve. 65.5 million years ago The Cretaceous-Tertiary extinction event eradicates about half of all animal species, including all of the dinosaurs. 6.5 million years ago First hominids evolve. million First members of the genus Homo appear in the fossil record.
years ago thousand years ago Neanderthals appear. thousand years ago Anatomically modern humans appear in Africa. thousand years ago Extinction of Neanderthals.
FIGURE 1-2 The first evolutionary tree in Charles Darwin’s notebook. One year after his epic voyage on HMS Beagle, Darwin had already begun formulating his concept of natural selection, which would be published in his On the Origin of Species (1859). Shown here is page 36 of his 1837 “B” notebook on Transmutation of Species; he wrote “I think” above his first evolutionary tree. In On the Origin of Species he describes in greater detail the evolutionary history of organisms both living and extinct and includes a more elaborate hypothetical evolutionary tree. [Page 36 of Charles Darwin’s 1837 “B” notebook on Transmutation of Species.] Description The evolutionary tree contains a root and numerous branches. Some ends of branches are labeled with capital letters. The handwritten text from Darwin describes the distance on the tree between pairs of letters, which represent genera or species. At first glance, the biological universe does appear amazingly diverse — from tiny ferns to tall fir trees, from single-celled bacteria and protozoans visible only under a microscope to multicellular animals of all kinds. The discovery that all living organisms are made up of cells was one of the most remarkable discoveries in biology. Indeed, cells come in an astonishing variety of sizes and shapes (Figure 1-3). Some move rapidly and have fast-changing structures, as we can see in movies of amoebae and rotifers. Others are largely stationary and structurally stable. Oxygen kills some cells but is an absolute requirement for others. Most cells in multicellular organisms are intimately involved with other cells. Although some unicellular organisms live in isolation (Figure 1-3a), others form colonies or live in close association with other types of organisms (Figure 1-3b, d), such as the bacteria that help plants extract nitrogen from the air or the bacteria that live in our intestines and help us digest food.
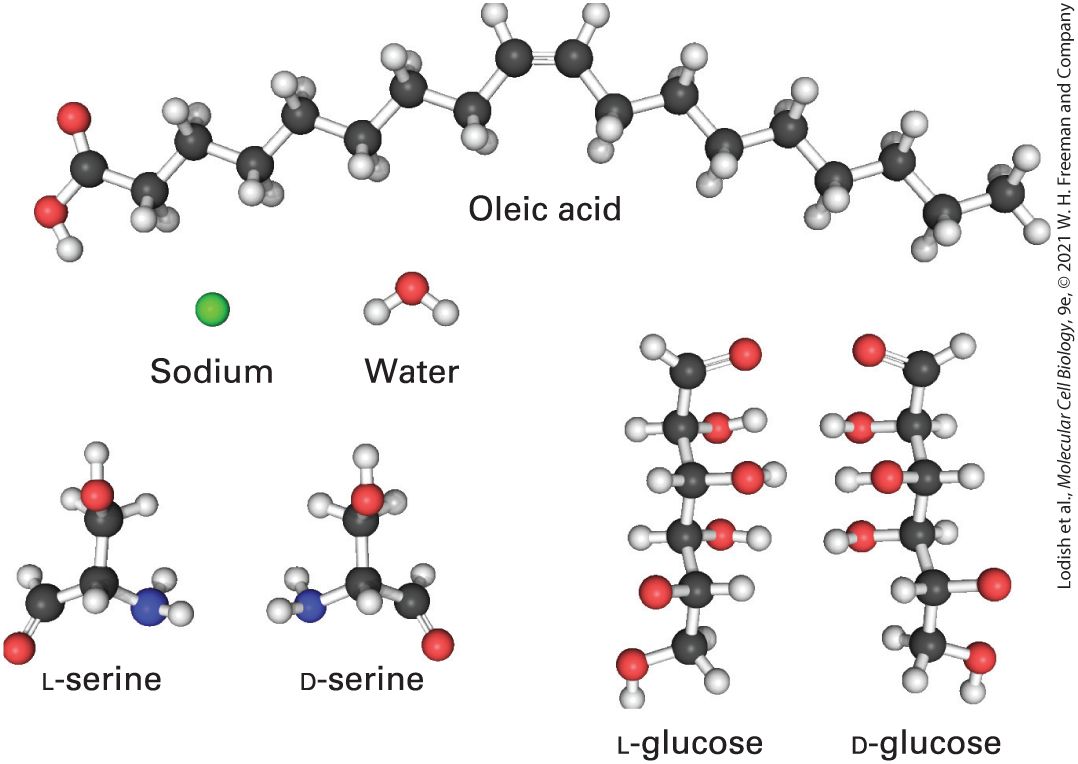
FIGURE 1-3 Cells come in an astounding assortment of shapes and sizes. Some of the morphological variety of cells is illustrated in these photographs. In addition to morphology, cells differ in their ability to move, internal organization (prokaryotic versus eukaryotic cells), and metabolic activities. (a) Eubacteria: Lactococcus lactis, which are used to produce cheese such as Roquefort, Brie, and Camembert. Note the dividing cells. (b) A mass of archaeans (Methanosarcina) that produce their energy by converting carbon dioxide and hydrogen gas to methane. Some species that live in the rumens of cattle give rise to of methane gas each day. (c) Human blood cells, shown in false color. The red cells are oxygen-bearing erythrocytes, the white cells (leukocytes) are part of the immune system and fight infection, and the green false-colored cell fragments are platelets that plug wounds and contain substances to initiate blood clotting. (d) A colonial singlecelled green alga, Volvox aureus. The surfaces of the large spheres contain thousands of individual cells, visible as blue or green dots. The yellow masses inside are daughter colonies, each made up of many cells. (e) A single Purkinje neuron of the cerebellum, which can form more than a hundred thousand connections with other cells through its branched network of dendrites. The cell was made visible by introduction of a geneencoding green fluorescent protein; the cell body with its nucleus is the bulb at the upper right. (f) Plant cells in vascular plants are fixed firmly in place, supported by a rigid
cellulose skeleton. Spaces between the cells are joined into tubes for transport of water and food. Description The six micrographs labeled A through F are as follows: A. Eubacteria: The micrograph shows clusters of oval-shaped structures on a scale of 1 micrometer. B. Archaeans: The micrograph shows several clusters of rounded structures on a scale of 1 micrometer. C. Human blood cells: The micrograph shows three different cells; erythrocytes, red biconcave disc-shaped cells; leucocytes, white sphere-shaped cells, and platelets, green false-colored irregular shaped cells. The scale reads 10 micrometers. D. Colonies of green alga: The micrograph shows several rounded shapes, each containing clusters of rounded structures within. The scale reads 100 micrometers. E. A single Purkinje neuron: The micrograph shows a roughly rounded cell body that tapers to form a narrow stem that forms a network of tiny branches. The scale reads 20 micrometers. F: Plant cells: The micrograph shows a mesh-like configuration with tubular structures between them on a scale of 20 micrometers. Yet the bewildering array of outward biological forms overlies a powerful uniformity: thanks to our common ancestry, all biological systems are composed of cells containing the same types of chemical molecules and employing similar principles of organization and function at the cellular level. Although the basic kinds of biological molecules have been conserved during the billions of years of evolution, the patterns in which
they are assembled to form functioning cells and organisms have undergone considerable change. We now know that genes, which chemically are composed of deoxyribonucleic acid (DNA), ultimately define biological structure and maintain the integration of cellular function. Many genes are copied into messenger RNA (mRNA) that in turn directs the synthesis of specific proteins, the primary molecules that make up cell structures and carry out virtually all cellular activities. Alterations in the structure and organization of genes, or mutations, provide the random variation that can alter biological structure and function. While the vast majority of random mutations have no observable effect on a gene’s or protein’s function, many are deleterious, and only a few confer an evolutionary advantage on the organism. In all organisms, mutations in DNA are constantly occurring, allowing over time the small alterations in cellular structures and functions that may prove to be advantageous. Entirely new cellular structures are rarely created; more often, existing cellular structures undergo changes that better adapt the organism to new circumstances. Slight changes in a protein can cause important changes in its function or abolish its function entirely. For instance, in a particular organism, one gene may randomly become duplicated, after which one copy of the gene and its encoded protein retain their original function while, over time, the second copy of the gene mutates such that its protein takes on a slightly different or even a totally new function. During the evolution of some organisms, the entire genome became duplicated, allowing the second copies of many genes to undergo
mutations and acquire new functions. The cellular organization of organisms plays a fundamental role in this process because it allows these changes to come about by small alterations in previously evolved cells, giving them new abilities. The result is that closely related organisms have very similar genes and proteins as well as similar cellular and tissue organizations. Multicellular organisms, including the human body, consist of such closely interrelated elements that no single element can be fully appreciated in isolation from the others. Organisms contain organs, organs are composed of tissues, tissues consist of cells, and cells are formed from molecules (Figure 1-4). The unity of living systems is coordinated by many levels of interrelationship: molecules carry messages from organ to organ and cell to cell, and tissues are delineated and integrated with other tissues by molecules secreted by cells. Generally all the levels into which we fragment biological systems interconnect.
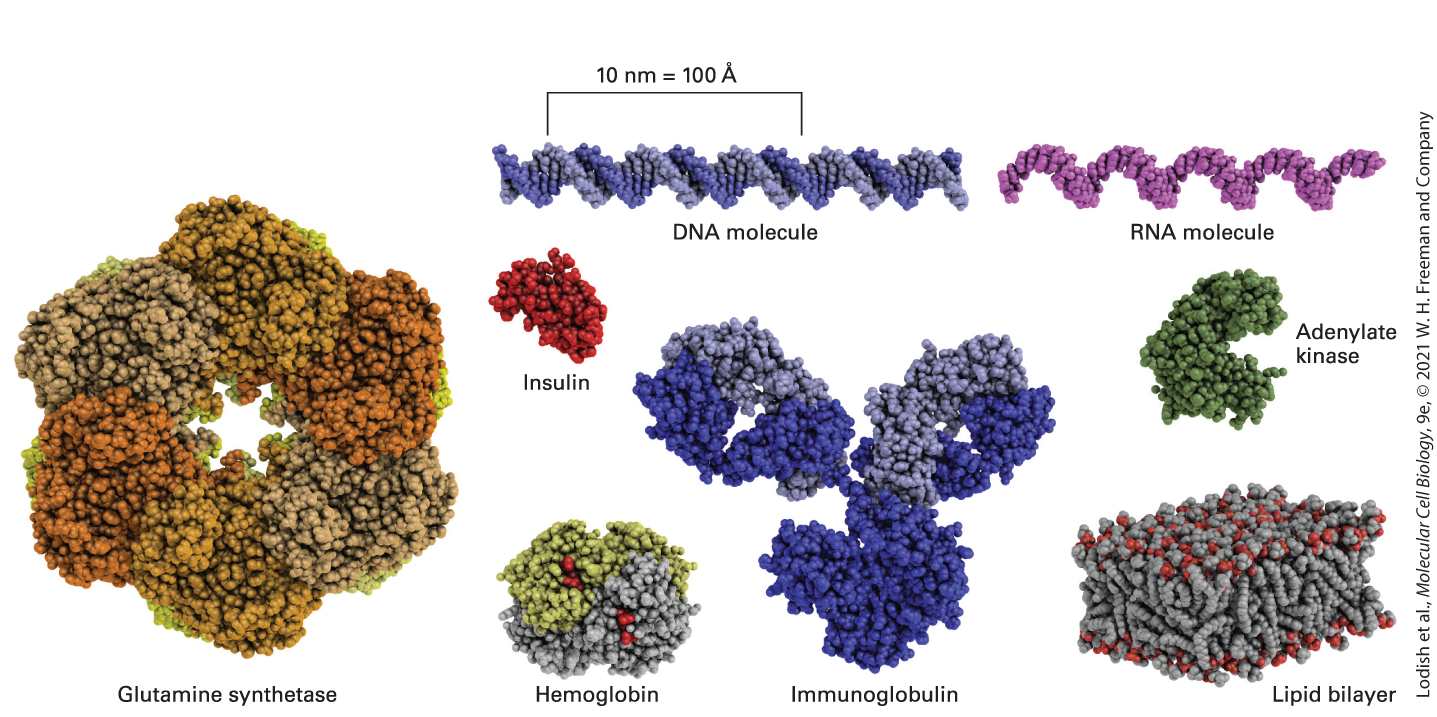
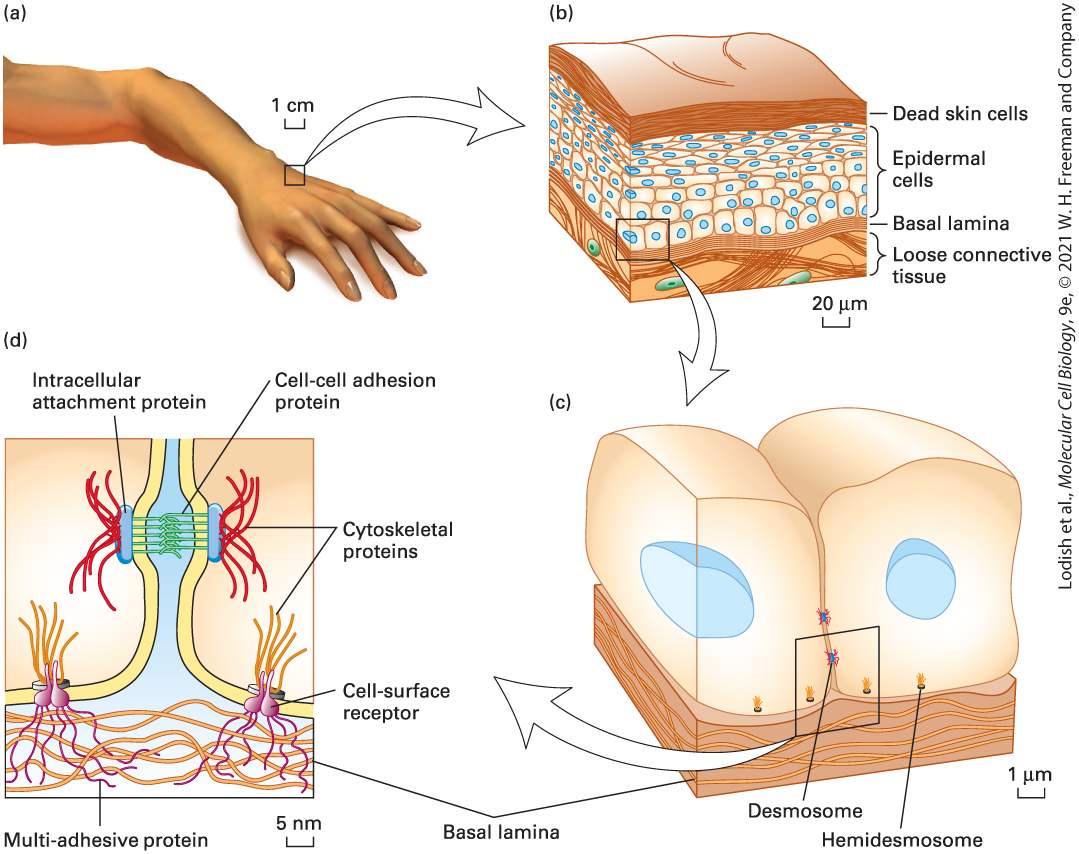
FIGURE 1-4 Living systems such as the human body consist of closely interrelated elements. (a) The surface of the hand is covered by a living organ, skin, that is composed of several layers of tissue. (b) An outer covering of hard, dead skin cells protects the body from injury, infection, and dehydration. This layer is constantly renewed by living epidermal cells, which also give rise to hair and fur in animals. Deeper layers of muscle and connective tissue give skin its tone and firmness. (c) Tissues are formed through subcellular adhesion structures (desmosomes and hemidesmosomes) that join cells to one another and to an underlying layer of supporting fibers. (d) At the heart of cell-cell adhesion are its structural components: phospholipid molecules that make up the cell-surface membrane, and large protein molecules. Protein molecules that traverse the cell membrane often form strong bonds with internal and external fibers made of multiple proteins. Description
The first part shows a human hand. An one centimeter portion is highlighted and enlarged in the second part. The second part shows a lateral sectional view of the skin with the following layers labeled from the top: Dead skin cells, epidermal cells, basal lamina and loose connective tissue. The scale reads 20 micrometers. A zoomed out view of the epidermal cell is shown in the third part. The third part shows two epidermal cells in a section cut through the cells with the nucleus and the basal lamina at the base of the cells. Several small structures labeled hemidesmosomes line the basal lamina. Two structures labeled desmosomes are present at the point of contact between the two cells. The scale reads, 1 micrometer. A section from the point of contact between the two cells, with the hemidesmosomes and the desmosomes are magnified in the fourth part. The fourth part shows a close up view of the desmosome that attaches two cells by cellto-cell adhesion proteins between the cells, and intracellular attachment proteins on the insides of the cells. The hemidesmosomes have a similar structure, with cytoskeletal proteins extending in the cell body, and cell-surface receptors extending into the basal lamina. The scale reads, 5 nanometers. To learn about biological systems, however, we must examine one small portion of a living system at a time. The biology of cells is a logical starting point because an organism can be viewed as consisting of interacting cells, which are the closest thing to autonomous biological units that exist. The last common ancestor of all life on Earth was a single cell (see Figure 1-1), and at the cellular level all life is remarkably similar. All cells use the same molecular building blocks; similar methods for the storage, maintenance, and expression of genetic information; and similar processes of energy metabolism, molecular transport, signaling, development, and structure.
In this chapter, we introduce the common features of cells. We begin with a brief discussion of the principal small molecules and macromolecules found in biological systems. Next we discuss the fundamental aspects of cell structure and function that are conserved in present-day organisms, focusing first on prokaryotic organisms — single-celled organisms without a nucleus — and their uses in studying the basic molecules of life. Then we discuss the structure and function of eukaryotic cells — cells with a defined nucleus — focusing on their many organelles. This discussion is followed by a section describing the use of unicellular eukaryotic organisms in investigations of basic aspects of molecular and cellular biology, focusing on yeasts and the ability to generate and analyze mutations in genes that are essential for life. We also discuss the parasite that causes malaria, focusing on the several dramatic transformations this cell undergoes within its human and mosquito hosts. We then turn to the structure and function of metazoans — multicellular animals — focusing on cell–cell interactions that allow tissues and organs to form. We now have the complete sequences of the genomes of many metazoans, as well as that of several thousand prokaryotic organisms and single-celled eukaryotes; these sequences have provided considerable insight into the evolution of genes and organisms. We discuss how this sequence data has led to the development of many new data-driven and computer-based techniques — the fields of genomics and bioinformatics — that have greatly informed our knowledge about biology. Analysis of the complete sequences of thousands of human DNAs has led, for example, to identification of mutations in many genes that cause human diseases. Analysis of ancient DNA samples has led to an understanding of
the relationships between modern humans and extinct Neanderthals and the finding that many humans today have up to 2 percent of their DNA genome derived from Neanderthals as a result of matings thousands of years ago (Figure 1-5). Such DNA studies have also elucidated the migratory pathways taken by ancient humans in and from Africa to their present locations. Analysis of messenger RNAs present in individual human cells has led to the discovery of totally new types of cells in the human body. Continuing advances in genomics and bioinformatics, along with developments in artificial intelligence and machine learning over the coming years will certainly illuminate many other aspects of human biology and disease.
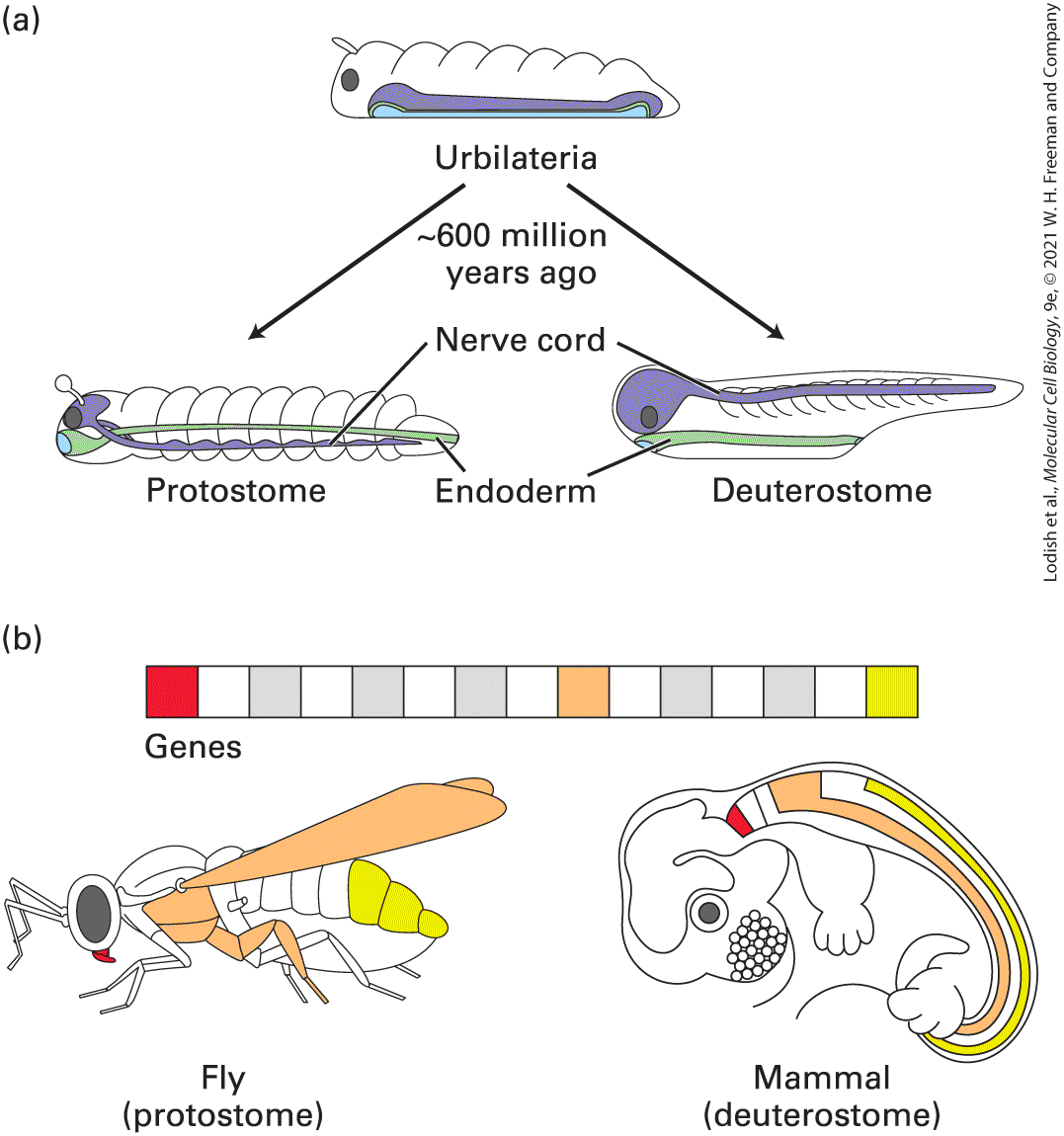
FIGURE 1-5 Approximate proportions of Neanderthal DNA found in present day humans. Humans bred with Neanderthals in the distant past, most likely in present-day Europe, and up to 2 percent of the DNA in present-day humans has a sequence derived from Neanderthal DNA. Plotted in this graph is the amount of Neanderthal DNA in humans in various locations, relative to the maximum observed of 2 percent. The entire Neanderthal genome was first sequenced from several -year-old bones found in a cave in Croatia; clearly Neanderthals mated with forerunners of modern humans in Eurasia, and we see different segments of Neanderthal DNA remaining in different modern non-African people. Neanderthal DNA is nearly absent from indigenous people living in South Africa, presumably because the great majority of their ancestors never encountered Neanderthals. [Data from: “Map” by Oliver Uberti, copyright © 2018 by David Reich & Eugenie Reich; from Who We Are and How We Got Here: Ancient DNA and the New Science of the Human Past by David Reich, Pantheon Books.] Description The map shows bars indicating the percentage of Neanderthal D N A and other ancestral D N A for populations in each location. The charts are in red for Neanderthal D N A and in white for other ancestral D N A. Text on the map reads, “Bar charts show the Neanderthal ancestry in each population as a fraction of the maximum of approximately 2 percent today.” The data from the map are as follows: Most of Africa: mostly ancestral D N A. Europe, the Middle East, and Central Asia: Three-fourths Neanderthal D N A and onefourth ancestral D N A. The remaining parts of Asia and Australia: Mostly or entirely Neanderthal D N A. In the final section of this chapter we introduce several widely studied metazoans — the fruit fly Drosophila melanogaster, the roundworm Caenorhabditis elegans, planaria, and the zebrafish Brachydanio rerio — that have been especially useful in uncovering the conserved roles of many
genes in metazoan development and function. We show how the accumulating amount of genetic data is and will continue to be analyzed to refine the evolutionary relationships among organisms as well as our understanding of human development, function, and disease. Indeed, biologists use evolution as a research tool: if a gene and its protein have been conserved in all metazoans but are not found in unicellular organisms, the protein probably has an important function in all metazoans and thus can be studied in whatever metazoan organism is most suitable for the investigation. Because the structure and function of many types of metazoan cells are also conserved, we now understand the structure and function of many cell types in considerable detail, including muscle and liver cells and the sheets of epithelial cells that line the intestine and form our skin. But many other cells — especially the multiple types that form our nervous and immune systems — still retain mysteries we need to solve; much important cell biological experimentation is needed on these and other cell systems and organs that form our bodies.
1.1 The Molecules of Life
1.1 The Molecules of Life While large polymers are the focus of molecular and cellular biology, small molecules are the stage on which all cellular processes are set. Water, inorganic ions, and a wide array of relatively small organic molecules (Figure 1-6) account for 75 to 80 percent of living matter by weight, and water accounts for about 75 percent of a cell’s volume. These small molecules serve as substrates for many of the chemical reactions that take place inside the cell, including the reactions of energy metabolism and cell signaling. Cells acquire these small molecules in different ways. Ions, water, and many small organic molecules are imported into the cell (see Chapter 11); other small molecules are synthesized within the cell, often by a series of chemical reactions (see
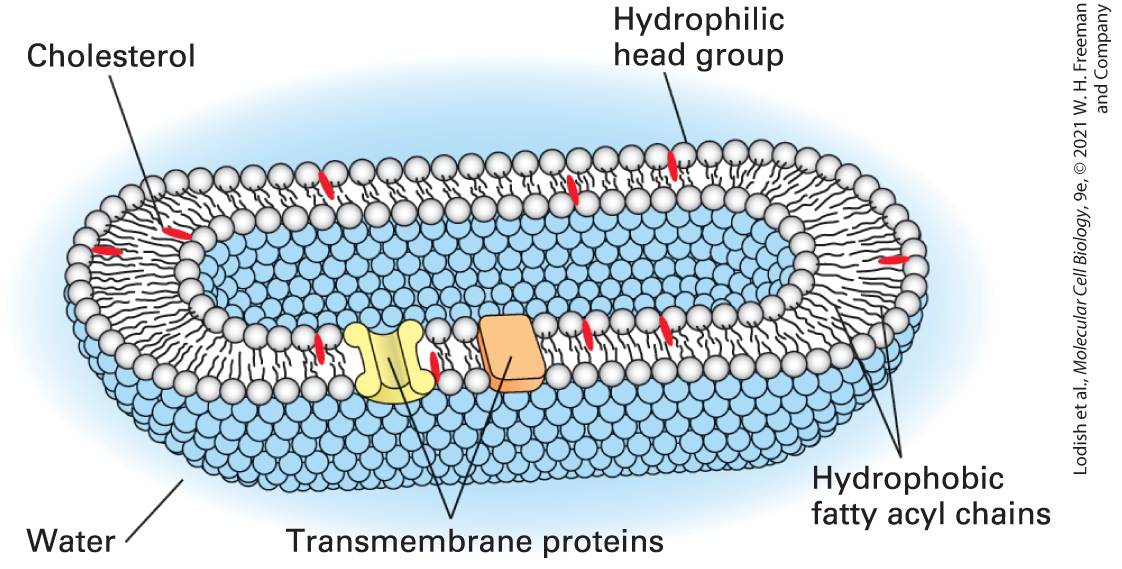
FIGURE 1-6 Some of the many small molecules found in cells. Only the L-forms of amino acids, such as serine depicted here, are found in cells and incorporated into proteins, not their D-mirror images. Only the D-form of glucose, not its L-mirror image, is found in nature and can be metabolized by cells to carbon dioxide and water. Description Sodium is represented by a single sphere. Water comprises of one large sphere (oxygen) with two small spheres (hydrogen) attached to it. L and D serines are made of three carbon, three oxygen, one nitrogen, and seven hydrogen atoms and are mirror images of each other. L and D glucose are made of six carbon, twelve hydrogen, and six oxygen atoms and are mirror images of each other. Oleic acid is an 18-carbon chain comprising two oxygen and 34 hydrogen atoms. Even in the structures of many small molecules, such as sugars, vitamins, and amino acids, we see the footprint of evolution. For example, all amino
acids except glycine have an asymmetric carbon atom, yet only the Lstereoisomer, never the D-stereoisomer, is incorporated into proteins. Similarly, only the D-stereoisomer of glucose is found in organisms, never the mirror-image L-stereoisomer (see Figure 1-6). At an early stage of biological evolution, our common cellular ancestor evolved the ability to catalyze chemical reactions that act on one stereoisomer instead of the other. How these selections happened is unknown, but now these choices are locked in place. The small molecules in cells have a variety of functions, For example, the universally conserved small molecule adenosine triphosphate (ATP) stores readily available chemical energy in two of its chemical bonds (Figure 1-7). When one of these energy-rich bonds in ATP is broken, forming ADP (adenosine diphosphate) and inorganic phosphate, , often abbreviated , the released energy can be harnessed to power energy-requiring processes such as muscle contraction or protein biosynthesis. To obtain energy for making ATP, many types of cells break down food molecules. For instance, when sugar is degraded to carbon dioxide and water, the energy stored in the sugar molecule’s chemical bonds is released, and much of it can be captured in the energy-rich bonds in ATP (see Chapter 12). Bacterial, plant, and animal cells can all make ATP by this process. In addition, plants and a few other organisms can harvest energy from sunlight to form ATP in photosynthesis. Some prokaryotes that live in extreme environments produce ATP by catalyzing reactions with reduced compounds like hydrogen sulfide .
FIGURE 1-7 Adenosine triphosphate (ATP) is the most common molecule used by cells to capture, store, and transfer energy. ATP is formed from adenosine diphosphate (ADP) and inorganic phosphate by photosynthesis in plants and photosynthetic algae and by the breakdown of sugars and fats in most cells. The energy released by the splitting (hydrolysis) of from ATP drives many cellular processes. Description In A T P, two high-energy bonds between the phosphates are identified with arrows. In A D P, one high-energy bond is identified with an arrow. The cycle indicates that A D P plus inorganic phosphate are combined by the input of energy from the sun via photosynthesis, or from high-energy compounds via respiration. Several uses of the energy bound in A T P are listed, including: 1. Synthesis of cellular macromolecules, such as D N A, R N A, proteins, and polysaccharides. 2. Synthesis of other cellular constituents, such as membrane phospholipids and certain required metabolites.
3. Cellular movements, including muscle contraction, crawling movement of entire cells, and movement of chromosomes during mitosis. 4. Transport of molecules against a concentration gradient. 5. Generation of an electric potential across a membrane, which is important for nerve function. 6. Generation of heat. Other small molecules (e.g., hormones such as adrenaline and histamine) act as signals that direct the activities of cells (see Chapters 15 and 16), and neurons (nerve cells) communicate with one another by releasing and sensing certain small signaling molecules such as serotonin (see Chapter 23). The powerful physiological effects of a frightening event, for example, come from the instantaneous flooding of the body with the small-molecule hormone adrenaline, which mobilizes the fight-or-flight response (see Chapter 15). Certain small molecules (monomers) can be joined to form polymers (also called macromolecules) through repetition of a single type of covalent chemical-linkage reaction. Cells produce three types of large macromolecules: polysaccharides, proteins, and nucleic acids. Sugars, for example, are the monomers used to form polysaccharides. Different polymers of D-glucose form cellulose, an important component of plant cell walls (see Chapter 20), and glycogen, a storage form of glucose found in liver and muscle (see Chapter 15). The cell is careful to provide the appropriate mix of small molecules needed as precursors for synthesis of macromolecules.
Proteins Give Cells Structure and Perform Most Cellular Tasks
Proteins Give Cells Structure and Perform Most Cellular Tasks Proteins, the workhorses of the cell, are the most abundant and functionally versatile of the cellular macromolecules. Cells string together 20 different amino acids (see Figure 2-14) in linear chains, each with a defined sequence, to form proteins, which commonly range in length from 100 to 1000 amino acids. During or just after its polymerization, a linear chain of amino acids folds into a complex shape, conferring a distinctive three-dimensional structure and function on the protein (Figure 1-8). Humans obtain amino acids either by synthesizing them from other molecules or by breaking down proteins that we eat.
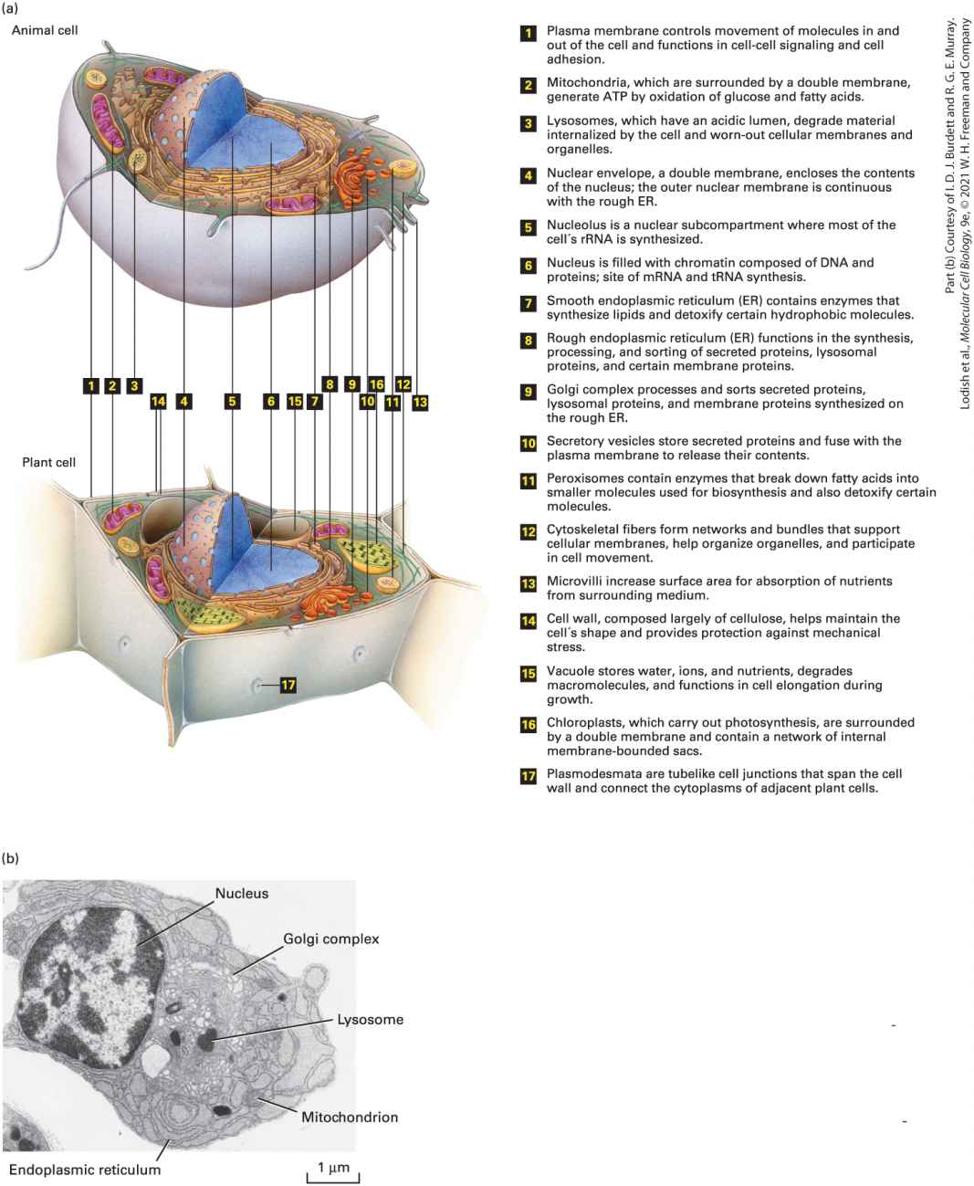
FIGURE 1-8 Models of some representative proteins drawn to a common scale and compared with a small portion of a lipid bilayer, a double-stranded DNA molecule, and a single-stranded RNA molecule. Each protein has a defined three-dimensional shape held together by numerous covalent and noncovalent chemical bonds. The illustrated proteins
include enzymes (glutamine synthetase and adenylate kinase), an antibody (immunoglobulin), a hormone (insulin), and the oxygen carrier (hemoglobin) in red blood cells. [Glutamine synthetase data from H. S. Gill and D. Eisenberg, 2001, Biochemistry 40:1903– 1912, PDB ID 1fpy. Insulin data from E. N. Baker et al., 1988, Phil. Trans. R. Soc. Lond. B Biol. Sci. 319:369–456, PDB ID 4ins. Hemoglobin data from G. Fermi et al., 1984, J. Mol. Biol. 175:159–174, PDB ID 2hhb. Immunoglobulin data from L. J. Harris et al., 1998, J. Mol. Biol. 275:861–872, PDB ID 1igy. Adenylate kinase data from G. Bunkoczi et al., PDB ID 2c9y.] Description The illustration shows proteins that include enzymes, antibody, hormone, and oxygen carrier. The enzymes are glutamine synthetase (composed of 12 identical subunits separated into two face-to-face rings) and adenylate kinase (represented in C shaped quaternary structure). An antibody (Y shaped immunoglobulin), a hormone (insulin represented by an irregular quaternary structure), and the oxygen carrier (hemoglobin represented by a tetrahedral structure with heme groups) in red blood cells. The proteins are compared with double-stranded D N A, single-stranded R N A, and a lipid bilayer represented by two layers of phospholipids. Proteins have a variety of functions in the cell. Many proteins are enzymes, which accelerate (catalyze) chemical reactions involving small molecules or macromolecules (see Chapter 3). Certain proteins catalyze steps in the synthesis of all proteins; others catalyze synthesis of macromolecules such as DNA and RNA (see Chapter 5). Other proteins form long fibers that serve as structural components of a cell, for example, by forming an internal skeleton termed the cytoskeleton. Other proteins associated with the cytoskeleton power the movement of subcellular structures such as chromosomes, and even of whole cells, by using energy stored in the chemical bonds of ATP (see Chapters 17 and 18). Still other
Nucleic Acids Carry Coded Information for Making Proteins at the Right Time and Place
proteins bind adjacent cells together or form parts of the extracellular matrix (see Figure 1-4). Proteins can be sensors that change shape as temperature, ion concentrations, or other properties of the cell change. Many proteins that are embedded in the cell-surface (plasma) membrane import and export a variety of small molecules and ions (see Chapter 11). Some proteins, such as insulin, are hormones; others are hormone receptors that bind their target protein or small molecule and then generate a signal that regulates a specific aspect of cell function (see Chapters 15 and 16). Other important classes of proteins bind to specific segments of DNA, turning genes on or off (see Chapter 8). In fact, much of molecular cell biology consists of studying the function of specific proteins in specific cell types. Nucleic Acids Carry Coded Information for Making Proteins at the Right Time and Place The macromolecule that garners the most public attention is deoxyribonucleic acid (DNA), whose functional properties make it the cell’s master molecule. The three-dimensional structure of DNA was first proposed by James D. Watson and Francis H. C. Crick in 1953 based on the x-ray crystal analysis of DNA by Rosalind Franklin. The DNA molecule consists of two long helical strands that are coiled around a common axis to form a double helix (Figure 1-9). The double-helical structure of DNA, one of nature’s most magnificent constructions, is
critical to the phenomenon of heredity, the transfer of genetically determined characteristics from one generation to the next.
FIGURE 1-9 DNA consists of two complementary strands wound around each other to form a double helix. The double helix is stabilized by weak hydrogen bonds between the A and T bases and between the C and G bases as well as by interactions between the stacked bases. During replication, the two strands are unwound and used as templates to produce complementary strands. The outcome is two identical copies of the original double helix, each containing one of the original strands and one new daughter (complementary) strand. DNA strands are composed of monomers called nucleotides; these monomers are often referred to as bases because they contain cyclic organic bases (see Chapter 5). Four different nucleotides, abbreviated A, T, C, and G, are joined to form a DNA strand, with the base parts projecting inward from the backbone of the strand. Two strands bind together via hydrogen bonding between the bases in the strands and twist to form a double helix. Each DNA double helix has a simple construction: wherever one strand has an A, the other strand has a T, and each C is matched with a G (see Figure 1-9). This complementary matching of the two strands is so strong that if complementary strands are separated under the right salt
concentration and temperature conditions, they will spontaneously zip back together. This property is critical for DNA replication and inheritance, as we will learn in Chapter 5, and also underlies many of the techniques for studying DNA molecules that are detailed in Chapter 6. The genetic information carried by DNA resides in its sequence, the linear order of nucleotides along a strand. Specific segments of DNA, termed genes, carry instructions for making specific proteins. Most bacteria have a few thousand protein-coding genes; yeasts and other unicellular eukaryotes have about 5000. Humans and other metazoans have between 13,000 and 23,000, while many plants have more (Table 1-2).
TABLE 1-2 • Genome Sizes of Organisms Used in Molecular Cell Biology Research That Have Been Completely Sequenced Base Pairs (Millions) Approximate Number of Encoded Proteins Chromosomes Reference Eubacteria Mycoplasma genitalium 0.58 a Helicobacter pylori 1.67 1,500 a Haemophilus influenzae 1.83 1,600 a Escherichia coli 4.64 4,100 a Bacillus subtilis 4.22 4,200 a i ii
Archaea Methanococcus jannaschii 1.74 1,800 a Sulfolobus solfataricus 2.99 3,000 a Single-Celled Eukaryotes Saccharomyces cerevisiae 12.16 6,700 b Chlamydomonas reinhardtii 120.4 14,400 b Plasmodium falciparum 23.26 5,400 b Multicellular Eukaryotes (Metazoans) Drosophila melanogaster 168.74 13,900 b Caenorhabditis elegans 100.29 20,500 b Schmidtea mediterranea (planarian) c Danio rerio (zebrafish) 1412.46 26,500 b Gallus gallus (chicken) 1072.54 15,500 b Mus musculus 3480.96 23,100 b iii
(mouse) Homo sapiens (human) 3326.74 20,800 b Arabidopsis thaliana 135.67 27,400 b iNumbers of encoded proteins are current estimates rounded to the nearest 100 based on genome DNA sequences. They will most likely change slightly in Eubacteria and Archaea because of the inclusion of newly discovered genes that code for very small proteins, and modestly in eukaryotes because of newly discovered small genes and because of pseudogenes that are not expressed. Only nuclear chromosomes are counted in eukaryotes, including distinct sex chromosomes in metazoans. Predicted value. SOURCE: Table courtesy of Dr. Juan Alvarez-Dominguez. References: a, http://www.ncbi.nlm.nih.gov/genome/; b, http://ensemblgenomes.org/; c, http://www.genome.gov/12512286. As we discuss later in this chapter, many of the genes in bacteria specify the sequences of proteins that catalyze reactions that occur universally, such as the metabolism of glucose and the synthesis of nucleic acids and proteins. These genes, and the proteins encoded by them, are conserved throughout all living organisms, and thus studies on the functions of the genes and proteins in bacterial cells have yielded profound insights into basic life processes. Similarly, many genes in unicellular eukaryotes, such as yeasts, encode proteins that are conserved throughout all eukaryotes but are not found in prokaryotes; we will see how yeasts have been used in ii iii
studies of processes such as cell division that have yielded profound insights into human diseases such as cancer. How is information stored in the DNA sequences converted into proteins? Cells use two processes in series (Figure 1-10). In the first process, called transcription, the protein-coding region of a gene is copied into a singlestranded ribonucleic acid (RNA) whose sequence is the same as one of the two strands of the double-stranded DNA. A large enzyme, RNA polymerase, catalyzes the linkage of nucleotides into an RNA chain using DNA as a template. In eukaryotic cells, the initial RNA product is processed into a smaller messenger RNA molecule, which moves out of the nucleus to the cytoplasm, the region of the cell outside of the nucleus. Here the ribosome, an enormously complex molecular machine composed of both RNA and proteins, carries out the second process, called translation. During translation, the ribosome assembles and links together amino acids in the precise order dictated by the mRNA sequence according to the nearly universal genetic code. We examine the cell components that carry out transcription and translation in detail in Chapter 5.
FIGURE 1-10 In eukaryotic cells the information encoded in DNA is converted into the amino acid sequences of proteins by a multistep process. Step 1 : Transcription factors and other proteins bind to the regulatory regions of the specific genes they control to activate those genes. Step 2 : RNA polymerase begins transcription of an activated gene at
a specific location, the start site. The polymerase moves along the DNA, linking nucleotides into a single-stranded pre-mRNA transcript using one of the DNA strands as a template. Step 3 : The transcript, whose nucleotide sequence is identical to that of the nontemplate strand, is processed to remove noncoding intron sequences. Step 4 : The mature mRNA moves to the cytoplasm, where it is bound by ribosomes that read its sequence and assemble a protein by chemically linking amino acids one at a time into a linear chain. Description The process of conversion of the information in D N A to proteins involves four numbered steps. Step 1. activation; step 2. transcription; step 3. processing, which occurs in the nucleus; and step 4. translation, which occurs in the cytoplasm. The steps are as follows: Step 1: A transcription factor binds to a D N A double helix. Step 2: The transcription factor marks the site where transcription starts, and R N A polymerase moves along the D N A strand, producing a pre-memory R N A containing protein coding and non-protein coding R N A as it moves toward the right of the double helix. Step 3: The pre-m-R N A is treated to remove non-coding regions of R N A. Step 4: The mature m-R N A is transported from the nucleus to the cytoplasm, where ribosomes translate the R N A into an amino acid chain. In addition to its role in transferring information from nucleus to cytoplasm, RNA can serve as a framework for building a molecular machine. The ribosome, for example, is built of four RNA chains that bind to more than 50 proteins to make a remarkably precise and efficient mRNA reader and protein synthesizer. While most chemical reactions in cells are catalyzed by proteins, a few, such as the formation by ribosomes
of the peptide bonds that connect amino acids in proteins, are catalyzed by RNA molecules. Well before the entire human genome was sequenced, it was apparent that only about 10 percent of human DNA consists of protein-coding genes, and for many years the remaining 90 percent was considered “junk DNA”! In recent years, we’ve learned that much of the so-called junk DNA is actually copied into thousands of RNA molecules that, though they do not encode proteins, serve equally important purposes in the cell (see Chapter 9). At present, however, we know the function of only a very few of these abundant noncoding RNAs. Like enzymes, certain RNA molecules, termed ribozymes, catalyze chemical reactions, as exemplified by the RNA inside a ribosome. Many scientists support the RNA world hypothesis, which proposes that RNA molecules that could replicate themselves were the precursors of current life forms; billions of years ago, the RNA world gradually evolved into the DNA, RNA, and protein world of today’s organisms. All organisms must control when and where their genes are transcribed. Nearly all the cells in our bodies contain the full set of human genes, but in each cell type only some of these genes are active, or turned on, and used to make proteins. For instance, liver cells produce some proteins that are not produced by muscle cells, and vice versa. Moreover, many cells respond to external signals or changes in external conditions by turning specific genes on or off, thereby adapting their repertoire of proteins to meet current needs. Commonly, protein-coding genes contain two parts:
Phospholipids Are the Conserved Building Blocks of All Cellular Membranes
the coding region specifies the amino acid sequence of a protein, and the regulatory region binds specific proteins called transcription factors, which act as switches, either activating or repressing transcription of particular genes, as discussed in Chapter 8. By this means, the regulatory region controls when and in which cells the gene’s encoded protein is made. Phospholipids Are the Conserved Building Blocks of All Cellular Membranes Life depends on cells being enclosed by a surface membrane that forms the interface of a cell with its environment. In eukaryotes, membranes also surround all of the internal compartments, termed organelles. Small molecules called phospholipids spontaneously pack together in such a way that they form the basic structure of these membrane sheets. Each of these bipartite molecules has a “water-loving” (hydrophilic) “head” and a “water-hating” (hydrophobic) “tail” composed of fatty acid chains. Cellular membranes are composed primarily of a bilayer (two layers) of phospholipid molecules. The two phospholipid layers of a membrane are oriented with all the hydrophilic heads directed toward the inner or outer surfaces of the membrane and the hydrophobic tails buried within its interior (Figure 1-11). Smaller amounts of other lipids, such as cholesterol, are inserted into this phospholipid framework. Cellular membranes are extremely thin relative to the size of a cell. If you magnify
a bacterium or yeast cell about 10,000 times to the size of a soccer ball, the plasma membrane is about as thick as a sheet of paper!
FIGURE 1-11 The watery interior of cells is surrounded by the plasma membrane, a two-layered shell of phospholipids. The phospholipid molecules are oriented with their hydrophobic fatty acyl chains (black squiggly lines) facing inward and their hydrophilic head groups (spheres) facing outward. Thus both sides of the membrane are lined by head groups, mainly charged phosphates, adjacent to the watery spaces inside and outside the cell. All biological membranes have the same basic phospholipid bilayer structure. Cholesterol (red) and various proteins are embedded in the bilayer. The interior space is vastly larger relative to the volume of the plasma membrane than is depicted here. Description The phospholipid molecules are oriented with their hydrophobic fatty acyl chains indicated by black squiggly lines that face inward. Spheres that face outward indicate their hydrophilic head groups. Both sides of the membrane are lined by head groups, mainly charged phosphates, adjacent to the watery spaces inside and outside the cell. Transmembrane proteins are on the outer membranes, and cholesterols represented by a red line are embedded in the bilayer.
Quality Control of All Cellular Macromolecules Is Essential for Life
Phospholipid membranes are impermeable to water, all ions, and virtually all hydrophilic small molecules. Thus each membrane in each cell also contains groups of proteins that allow specific ions and small molecules to cross. Other membrane proteins serve to attach the cell to other cells or to polymers that surround it (see Figure 1-4); still others give the cell its shape or allow its shape to change. We will learn more about membranes and how molecules cross them in Chapters 10 and 11. New cells are always derived from parental cells by cell division. We’ve seen that the two strands of the parental DNA are templates for the synthesis of new DNA molecules such that each daughter DNA molecule has the same sequence as the parental one. In parallel, new membranes are made by incorporation of phospholipids and proteins into existing membranes in the parental cell and divided between daughter cells by fission. Thus membrane synthesis, like DNA synthesis, is templated by a parental structure. Quality Control of All Cellular Macromolecules Is Essential for Life All cells are constantly bombarded by potentially hazardous chemicals and radiation that can damage the macromolecules within, and cells expend a significant amount of their energy to ensure that damaged molecules — especially DNA — are repaired or degraded. Proteins function only when they are folded into their proper three-dimensional structure (see Figure 18). Proteins that fail to fold, or become unfolded as a result of stresses
such as an elevated temperature, are usually rapidly degraded by enzymes within the cell, releasing their constituent amino acids, which are reused for making new proteins. Occasionally, though, misfolded proteins aggregate together inside the cell, causing damage to the cell. As discussed in Chapter 3, such protein aggregates that form over time in brain nerve cells are thought to be the cause of Alzheimer’s and other neurodegenerative diseases. Ultraviolet and gamma radiation, as well as many chemicals, can damage DNA, causing changes in the normal DNA sequence. Such mutations can cause errors in DNA replication and lead to inability of that sequence to direct the synthesis of the correct protein. Cells have several mechanisms, discussed in Chapter 5, to repair such damage to DNA, converting the mutation back to the normal sequence. Occasionally mutations are not repaired and cause death of the cell. Alternatively, unrepaired mutations can cause proteins to be made in the wrong cell or at the wrong time and lead to uncontrolled growth of the cell. Chapter 25 details how many types of mutations that occur over an individual’s life cause cancer.
1.2 Prokaryotic Cell Structure and Function
1.2 Prokaryotic Cell Structure and Function The biological universe consists of two types of cells: prokaryotic and eukaryotic. Prokaryotic cells such as bacteria consist of a single closed compartment that is surrounded by a plasma membrane, lack a defined nucleus, and have a relatively simple internal organization (Figure 1-12). Eukaryotic cells contain a defined membrane-bounded nucleus and extensive internal membranes that enclose the organelles (Figure 1-13).
FIGURE 1-12 Prokaryotic cells have a relatively simple structure. (a) Electron micrograph of a thin section of Escherichia coli, a common intestinal bacterium. The nucleoid, consisting of the bacterial DNA, is not enclosed within a membrane. E. coli and other gram-negative bacteria are surrounded by two membranes separated by the periplasmic space. The thin cell wall is adjacent to the inner membrane. (b) This artist’s drawing shows the nucleoid (blue) and a magnification of the layers that surround the cytoplasm in a gram-negative bacterium. Most of the cell is composed of water, proteins, ions, and other molecules that are too small to be depicted at the scale of this drawing.
Description In transmission electron micrograph of the rod-shaped prokaryotic cell the following parts are labeled: The periplasmic space and cell wall, outer membrane, inner (plasma) membrane, cytoplasm, and the nucleoid. The scale reads, 0.5 micrometers. The schematic illustration shows a rod-shaped double-walled cell containing a mesh-like structure labeled nucleoid in the cytoplasm. A zoomed-out view of the double wall surrounding the cytoplasm shows an outer membrane, followed by a periplasmic membrane, the cell wall, and the inner (plasma) membrane.
FIGURE 1-13 Subcellular organization of eukaryotic cells. (a) Schematic overview of a typical animal cell (top) and plant cell (bottom) and their major substructures. Not every cell type will contain all the organelles, granules, and fibrous structures shown here, and other substructures can be present in some cell types. Cells also differ considerably in shape and in the prominence of various organelles and substructures. (b) Electron micrograph of a
plasma cell, a type of white blood cell that secretes antibodies, showing some of the larger organelles. Not all of the organelles are labeled. Description The labeled parts that are common in both the animal and plant cells are as follows: 1. Plasma membrane controls movement of molecules in and out of the cell and functions in cell-cell signaling and cell adhesion. 2. Mitochondria, which are surrounded by a double membrane, generate ATP by oxidation of glucose and fatty acids. 3. Lysosomes, which have an acidic lumen, degrade material internalized by the cell and worn-out cellular membranes and organelles. 4. Nuclear envelope, a double membrane, encloses the contents of the nucleus; the outer nuclear membrane is continuous with the rough E R. 5. Nucleolus is a nuclear sub-compartment where most of the cell´s r R N A is synthesized. 6. Nucleus is filled with chromatin composed of D N A and proteins; site of mRNA and t R N A synthesis 7. Smooth endoplasmic reticulum (E R) contains enzymes that synthesize lipids and detoxify certain hydrophobic molecules. 8. Rough endoplasmic reticulum (E R) functions in the synthesis, processing, and sorting of secreted proteins, lysosomal proteins, and certain membrane proteins. 9. Golgi complex processes and sorts secreted proteins, lysosomal proteins, and membrane proteins synthesized on the rough E R. 10. Secretory vesicles store secreted proteins and fuse with the plasma membrane to release their contents. 11. Peroxisomes contain enzymes that break down fatty acids into smaller molecules used for biosynthesis and also detoxify certain molecules.
Prokaryotes Comprise Two Kingdoms: Archaea and Eubacteria
12. Cytoskeletal fibers form networks and bundles that support cellular membranes, help organize organelles, and participate in cell movement. 13. Microvilli increase surface area for absorption of nutrients from surrounding medium. 14. Cell wall, composed largely of cellulose, helps maintain the cell´s shape and provides protection against mechanical stress. 15. Vacuole stores water, ions, and nutrients, degrades macromolecules, and functions in cell elongation during growth. 16. Chloroplasts, which carry out photosynthesis, are surrounded by a double membrane and contain a network of internal membrane-bounded sacs. 17. Plasmodesmata are tube like cell junctions that span the cell wall and connect the cytoplasms of adjacent plant cells. The electron micrograph shows an irregular plasma cell and the parts labeled are as follows: Nucleus, golgi complex, lysosome, mitochondrion, and the endoplasmic reticulum. The scale reads, 1 micrometer. Prokaryotes Comprise Two Kingdoms: Archaea and Eubacteria In recent years, detailed analysis of DNA sequences from a variety of prokaryotic organisms has revealed two distinct kingdoms: the Eubacteria, often simply called “bacteria,” and the Archaea (see Figure 1-1). Eubacteria are single-celled organisms; they include the photosynthetic cyanobacteria, or “blue-green algae.” Figure 1-12 illustrates the general structure of a typical eubacterial cell; archaeal cells have a similar structure. Bacterial cells are commonly in size and consist of a
single closed compartment containing the cytoplasm and bounded by the plasma membrane. The genome is composed of a single circular, doublestranded DNA molecule; many prokaryotes contain additional small circular DNA molecules called plasmids. Although bacterial cells do not have a defined nucleus, the DNA is extensively folded into the central region of the cell, called the nucleoid. Many proteins are precisely localized within the cytosol or nucleoid or in the plasma membrane or cell wall, indicating the presence of an elaborate structural and functional organization. Most ribosomes are found in the cytoplasm. Unlike mRNAs in eukaryotes (see Figure 1-10), bacterial mRNAs undergo limited if any processing. Because there is no membrane barrier between bacterial DNA and cytoplasm, ribosomes are able to bind to an mRNA as soon as part of it has been synthesized by RNA polymerase; thus in prokaryotes, transcription and translation occur contemporaneously. Bacterial cells possess a cell wall, which lies adjacent to the external side of the plasma membrane. The cell wall is composed of layers of peptidoglycan, a complex of proteins and oligosaccharides; it helps protect the cell and maintain its shape. Some bacteria (e.g., E. coli) have a thin inner cell wall and an outer membrane separated from the inner cell wall by the periplasmic space. Such bacteria are not stained by the Gram technique and thus are classified as gram-negative. Other bacteria (e.g., Bacillus polymyxa) that have a thicker cell wall and no outer membrane take the Gram stain and thus are classified as gram-positive.
Many Bacteria Including Escherichia coli Are Widely Used in Biological Research
In addition to DNA sequence distinctions that separate them from Eubacteria, Archaea have cell membranes that differ dramatically in composition from those of Eubacteria and eukaryotes. Many archaeans grow in unusual, often extreme, environments that may resemble the ancient conditions that existed when life first appeared on Earth. For instance, halophiles (“salt lovers”) require high concentrations of salt to survive, and thermoacidophiles (“heat and acid lovers”) grow in hot sulfur springs, where a of less than 2 is common. Many Bacteria Including Escherichia coli Are Widely Used in Biological Research The bacterial lineage includes Escherichia coli, a favorite experimental organism, which in nature is common in soil and in animal intestines. E. coli and several other bacteria have a number of advantages as experimental organisms. They grow rapidly in a simple and inexpensive medium containing glucose and salts, in which they can synthesize all necessary amino acids, lipids, vitamins, and other essential small molecules. Like all bacteria, E. coli possesses elegant mechanisms for controlling gene activity that are now well understood. Over time, researchers have developed powerful systems for genetic analysis of this organism. These systems are facilitated by the small size of bacterial genomes, the ease of obtaining mutants, the availability of techniques for transferring genes into bacteria, an enormous wealth of knowledge about
bacterial gene control and protein functions, and the relative simplicity of mapping genes relative to one another in the bacterial genome. In Chapter 6, we see that E. coli is widely used today in multiple types of recombinant DNA research. Bacteria such as E. coli that grow in environments as diverse as the soil and the human gut have about 4000 genes, encoding about the same number of proteins (see Table 1-2). Certain disease-causing bacteria that only grow inside human cells, such as Chlamydia, have only genes; they acquire amino acids and other nutrients from their host cells and thus have evolved to lack genes for enzymes that catalyze reactions in the synthesis of amino acids and certain lipids. Other parasitic bacteria such as Mycoplasma species have even fewer genes. Many bacterial genes encoding proteins essential for DNA, RNA, and protein synthesis, and for membrane function are conserved in all organisms, and much of our knowledge of these important cellular processes was uncovered first by studies in E. coli and other bacteria. For example, certain E. coli membrane proteins that import amino acids across the plasma membrane are closely related in sequence, structure, and function to membrane proteins in mammalian brain cells that import small nerve-to-nerve signaling molecules called neurotransmitters (see Chapters 11 and 23). Throughout this chapter, we will encounter eukaryotic organisms that have been chosen for study because, like E. coli, they are easy to grow and study. Of course, many bacteria cause serious diseases. Research on
disease-causing bacteria is often focused on understanding their unique biology and on discovering antibiotics that selectively kill them but not their human or animal hosts — for instance, drugs that block formation of parts of the cell wall.
1.3 Eukaryotic Cell Structure and Function
1.3 Eukaryotic Cell Structure and Function Eukaryotes comprise all members of the plant and animal kingdoms as well as protozoans (proto, “primitive”; zoan, “animal”), which are exclusively unicellular and include fungi and amoebae. Eukaryotic cells are commonly about across, generally much larger than bacteria. A typical human fibroblast, a connective tissue cell, is about across, with a volume and dry weight some thousands of times those of an E. coli cell. An amoeba, a single-celled protozoan, can have a cell diameter of approximately , more than 30 times that of a fibroblast. Eukaryotic cells, like prokaryotic cells, are surrounded by a plasma membrane. However, unlike prokaryotic cells, all eukaryotic cells (the human red blood cell is an exception) also contain extensive internal membranes that enclose specific subcellular compartments, the organelles (see Figure 1-13). The cytosol, the organelle-free part of the cytoplasm, contains water, dissolved ions, small molecules, and many types of proteins. Plant cells and most fungal cells are surrounded by a cell wall that gives the cell a rigid shape and also allows for rapid cell expansion. All eukaryotic cells have many of the same organelles and other subcellular structures, attesting to their common evolutionary origin.
The Cytoskeleton Has Many Important Functions
Many organelles are surrounded by a single phospholipid membrane, but the nucleus, mitochondrion, and chloroplast are enclosed by two membranes. Each organelle membrane and each space in the interior of an organelle has a unique and specific set of proteins that enable each type of organelle to have a distinct physiological function — for example, production of ATP from ADP and occurs in mitochondria and chloroplasts; and many lipids, membrane proteins, and proteins to be secreted from cells are synthesized in the rough endoplasmic reticulum. Many outworn or misfolded proteins are degraded in lysosomes and their constituent amino acids recycled. Much like a factory building automobiles or computers, such compartmentalization of functions allows the cell as a unit to operate efficiently. The membranes defining these subcellular compartments contain proteins that control their internal ionic composition so that the internal composition of each compartment generally differs from that of the surrounding cytosol as well as compartments in the other organelles. Here we describe the organelles common to all eukaryotic cells as well as several that are found only in certain types of eukaryotes. We begin with the proteins that give eukaryotic cells their shapes and organize the organelles. The Cytoskeleton Has Many Important Functions
The cytoplasm contains an array of fibrous proteins collectively called the cytoskeleton (see Chapters 17 and 18). Three classes of fibers compose the cytoskeleton: microtubules ( in diameter), built of polymers of the protein tubulin; microfilaments ( in diameter), built of the protein actin; and intermediate filaments ( in diameter). All of these fibers are long chains of multiple copies of one or more small protein subunits (Figure 1-14).
FIGURE 1-14 The three types of cytoskeletal filaments have characteristic distributions within mammalian cells. Three views of the same cell. A cultured fibroblast was treated with a detergent to make it permeable to large molecules and then treated with three different antibody preparations. Each antibody binds specifically to the protein monomers forming one type of filament and is chemically linked to a differently colored fluorescent dye (green, blue, or red). Visualization of the stained cell in a fluorescence microscope reveals the locations of filaments bound to a particular dye-antibody preparation. In this case, microtubules are stained blue; actin microfilaments, red; and intermediate filaments, green. All three fiber systems contribute to the shape and movements of cells. The cytoskeleton has many crucial functions. It gives the cell strength and rigidity, thereby helping to maintain its shape; this is perhaps most obvious with neurons, in which microtubules and other fibers allow the formation of the long, slim protuberances — the axons and dendrites (see
Figure 1-3e and Chapter 23) — that emanate from the cell body and allow
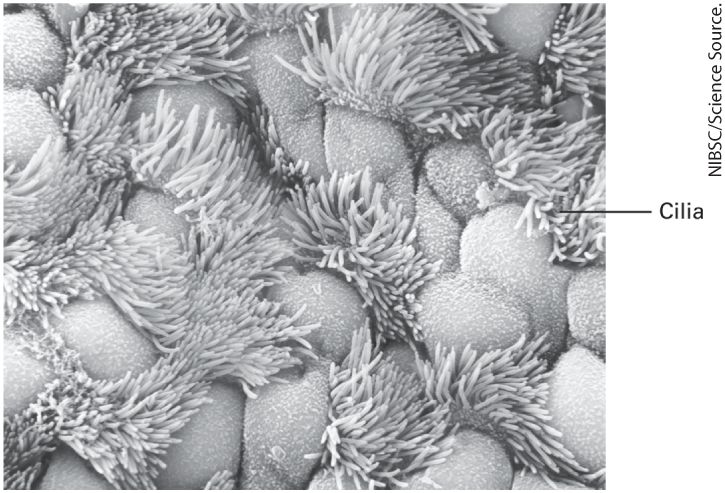
each neuron to carry out its specialized functions. Certain cytoskeletal fibers provide the tracks along which many organelles move, thus ensuring that they are localized to the correct place in the cytosol. Other fibers play key roles in cell motility and, in muscle cells, the ability to contract. Perhaps most important, cell division and the segregation of chromosomes and organelles into the two daughter cells could not occur without the organizational framework provided by the microtubule cytoskeleton and its associated proteins (Chapter 19). Cilia and flagella are similar fingerlike extensions of the cell surface. Surrounded by the plasma membrane, they contain a bundle of microtubules that gives them their shape and, acting together with motor proteins, allows them to beat rhythmically. They propel materials across epithelial surfaces (Figure 1-15), enable sperm to swim, and push eggs through the oviduct (see Chapter 18). As detailed in Chapter 16, most vertebrate cells contain at least one cilium that plays a key role in cell-cell signaling.
The Nucleus Contains the DNA Genome, Apparatuses for Synthesis of DNA and RNA, and a Fibrous Matrix
FIGURE 1-15 Surface of the ciliated epithelium lining a mammalian trachea viewed in a scanning electron microscope. Beating cilia, which have a core of microtubules, propel mucus and foreign particles out of the respiratory tract, keeping the lungs and airways clear. The Nucleus Contains the DNA Genome, Apparatuses for Synthesis of DNA and RNA, and a Fibrous Matrix In a growing or differentiating cell, the nucleus, the largest organelle in animal cells, is the site of DNA replication and the synthesis of ribosomal RNA, mRNA, and a large variety of noncoding RNAs (see Chapters 5 and 8). The nucleus is surrounded by two membranes, each one a phospholipid bilayer containing many different types of proteins (Figure 1-16). The
inner nuclear membrane defines the nucleus itself. In most cells, the outer nuclear membrane is continuous with the endoplasmic reticulum, and the space between the inner and outer nuclear membranes is continuous with the lumen — the central cavity — of the endoplasmic reticulum (Figure 116a). The two nuclear membranes appear to come together at nuclear pore complexes, ringlike structures composed of specific membrane proteins embedded in the inner and outer membranes. RNAs and proteins move between the nucleus and the cytosol through these nuclear pores (red arrow in Figure 1-16a). The structure of the nuclear pores and the regulated transport of material through them are detailed in Chapters 9 and 13. Intermediate-filament proteins called lamins form a two-dimensional network, called the nuclear lamina, that adheres to the inner surface of the inner membrane, giving it shape and rigidity. The lamina breaks down early in cell division and is reassembled after the two daughter cells have separated, as we detail in Chapter 19.
FIGURE 1-16 Structure of the nucleus. (a) Schematic diagram of the structure of a typical cell nucleus and the connection of the outer nuclear membrane with the rough endoplasmic reticulum. The small black dots attached to the membrane of the rough endoplasmic reticulum are ribosomes that are synthesizing membrane and secreted proteins. The red arrow depicts the bidirectional movement of molecules between the nucleus and cytosol through the nuclear pores. For simplicity, many of the organelles in the cytoplasm (see
Figure 1-13) are not shown. (b) Electron micrograph of a pancreatic acinar cell from the bat Myotis lucifugus. The nucleolus is a subcompartment of the nucleus and is not surrounded by a membrane; ribosomal RNA is produced in the nucleolus and ribosomes are assembled there from ribosomal RNA and ribosomal proteins. Darkly staining areas in the nucleus outside the nucleolus are regions of heterochromatin, regions of chromatin that are compact and unable to be transcribed into messenger RNA. Description The illustration shows the structure of the nucleus. The nucleus is present within a double wall labeled plasma membrane. The nucleus is enveloped in an outer and inner nuclear membrane. The openings in the membranes are labeled nuclear pore complex. Along the inside of the inner nuclear membrane are many laminar structures. At the center of the nucleus, a dense region of genetic material is labeled the nucleolus. Throughout the nucleus, many strands of chromatin are present. Forming a complex network of channels, the rough endoplasmic reticulum extends outward from the outer nuclear membrane. The outer nuclear membrane and endoplasmic reticulum are studded with ribosomes. The electron micrograph of the pancreatic cell shows a dark nucleolus within a circular nucleus. Small apertures in the inner and outer nuclear membranes are labeled as nuclear pores. Darker regions in the nucleus are labeled as condensed heterochromatin. Outside of the nucleus, the endoplasmic reticulum is seen as a dense pattern of folded membranes. The total DNA in an organism is referred to as its genome. In prokaryotic cells, most or all of the genetic information resides in a single circular DNA molecule about a millimeter in length; this molecule lies, folded
back on itself many times, in the central region of the micrometer-sized cell (see Figure 1-12 and Table 1-2). In contrast, DNA in the nuclei of eukaryotic cells is distributed among multiple long linear structures called chromosomes. The length and number of chromosomes are the same in all cells of a particular species, but vary among different species (see Table 12). As detailed in Chapter 7, each chromosome comprises a single DNA molecule associated with numerous histones and other proteins. In a nucleus that is not dividing, the chromosomes are dispersed and are not dense enough to be observed in the light microscope. Typical nuclei contain several subcompartments, not bound by membranes, that have specific functions. The largest, the nucleolus, gathers together the parts of several different chromosomes containing the genes that direct the synthesis of ribosomal RNA and is the site of assembly of the ribosomal RNAs and ribosomal proteins into ribosomes. When nondividing cells are visualized in an electron microscope, the nonnucleolar regions of the nucleus, called the nucleoplasm, can be seen to have dark- and light-staining areas. The dark areas, which are often closely associated with the nuclear membrane, contain condensed, concentrated DNA that cannot be transcribed into RNA, called heterochromatin (see
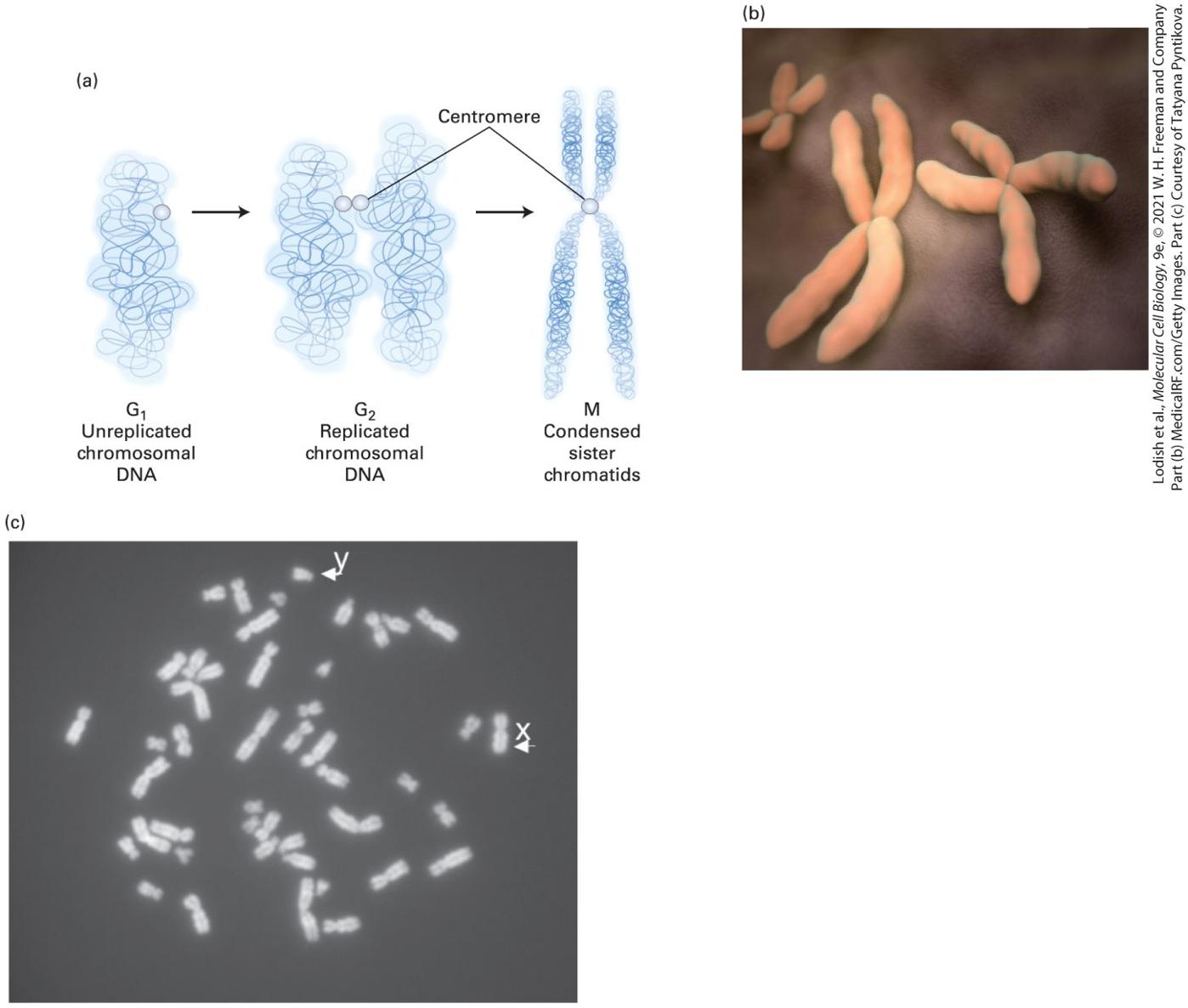
Figure 1-16b). Chromosomes, which stain intensely with basic dyes, are visible in light and electron microscopes only during cell division, when the DNA becomes tightly compacted (Figure 1-17). Although the large genomic DNA molecule in prokaryotes is also associated with proteins, the
arrangement of DNA within a bacterial chromosome differs greatly from that within the linear chromosomes of eukaryotic cells; bacterial chromosomes are circular and are associated with different types of proteins than are eukaryotic chromosomes.
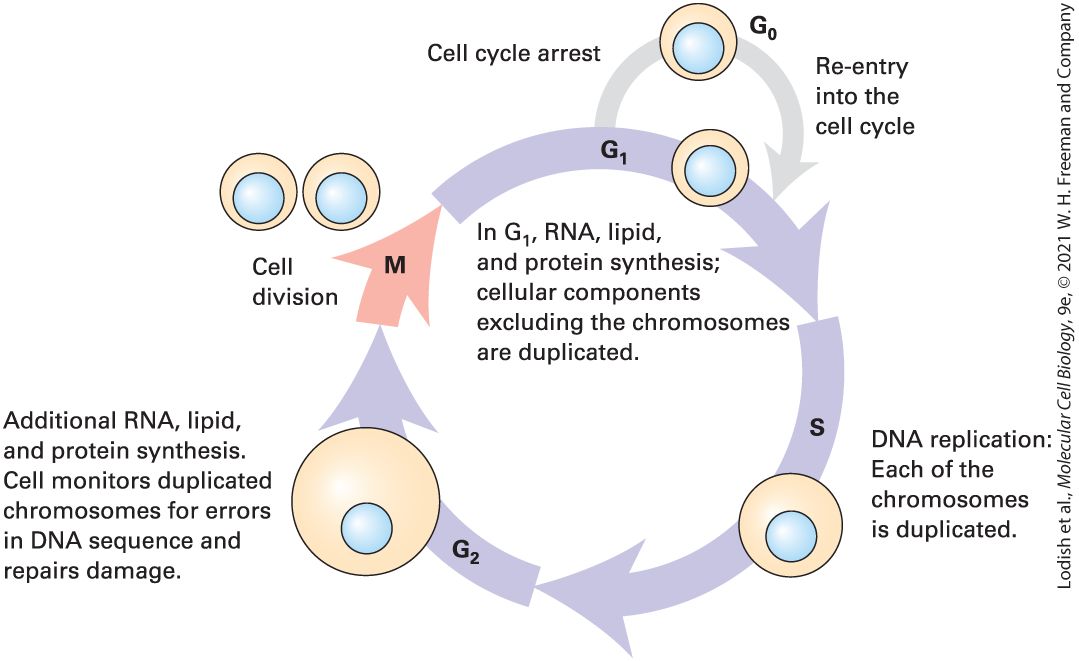
FIGURE 1-17 Individual chromosomes can be seen in cells during cell division (mitosis). (a) During the S phase of the cell cycle (see Figure 1-22) the chromosomal DNA together with its attached proteins (not shown) is duplicated, and the daughter sister chromatids, each with a complete copy of the chromosomal DNA, remain attached at the centromere. (b) During the actual cell division process (mitosis), the chromosomal DNA becomes highly compacted, and the pairs of sister chromatids can be seen in the electron microscope, as depicted here. (c) Light-microscope image of a chromosomal spread from a cultured human
The Endoplasmic Reticulum Is the Site of Synthesis of Most Membrane and Secreted Proteins as Well as Many Lipids
male lymphoid cell arrested in the metaphase stage of mitosis by treatment with the microtubule-depolymerizing drug colcemid. There is a single copy of the duplicated X and Y chromosomes and two copies of each of the others. Description The illustration labeled A shows the following events that occur during cell division: G subscript 1- unreplicated chromosomal D N A leads to G subscript 2 to form replicated chromosomal D N A which further leads to M and forms condensed sister chromatids. The centromere is labeled in G subscript 2 and M. The micrograph labeled B shows pairs of X shaped sister chromatids. The micrograph labeled C shows tiny fragments of chromosomes. The Endoplasmic Reticulum Is the Site of Synthesis of Most Membrane and Secreted Proteins as Well as Many Lipids We noted earlier that each type of organelle membrane, and the interior space of each organelle, contains a discrete set of proteins that enable it to carry out its specific function. Indeed, about a third of the proteins in a typical eukaryotic cell are found in the plasma membrane, in a specific organelle membrane, or in the interior space of an organelle. Thus there are a large number of proteins (and lipids) that have to be synthesized and sorted to different locations — about one-third of the proteome, the totality of proteins made by a eukaryotic cell. The synthesis of lipids, secreted proteins, and many types of membrane proteins occurs on the
organelle with the largest membrane surface in a eukaryotic cell, the endoplasmic reticulum (ER). The ER is an extensive network of closed, flattened, membrane-bounded sacs called cisternae (Figure 1-18; see also Figure 1-16a), and is generally subdivided into regions termed the smooth and rough endoplasmic reticulum. The smooth ER is smooth because it lacks ribosomes; it is the site of synthesis of fatty acids and phospholipids. In contrast, the cytosolic side of the rough endoplasmic reticulum is studded with ribosomes; these ribosomes synthesize certain membrane and organelle proteins and virtually all proteins that are to be secreted from the cell (see Chapter 13). Newly made membrane proteins become associated with the rough ER membrane, and proteins to be secreted accumulate in the lumen, the aqueous interior of the organelle.
The Golgi Complex Sorts Secreted Proteins and Many Membrane Proteins to Their Final Destinations in the Cell
FIGURE 1-18 The Golgi complex and rough endoplasmic reticulum. An electron micrograph of a section of a human liver cell shows the abundant ribosome-studded rough endoplasmic reticulum and the Golgi complex, as well as many ribosomes free in the cytosol. Description The lumen of rough endoplasmic reticulum, Lumen of Golgi vesicle, and the Vesicles moving proteins from the rough endoplasmic reticulum to the Golgi complex are labeled. The scale reads, 200 nanometers. The Golgi Complex Sorts Secreted Proteins and Many Membrane
Endosomes Bring Proteins and Particles from the Outside into Cells
Proteins to Their Final Destinations in the Cell Several minutes after secreted proteins and membrane proteins are synthesized in the rough ER, most of them leave the organelle within small, membrane-bounded transport vesicles about 50 nanometers in diameter. These vesicles, which bud from regions of the rough ER not coated with ribosomes, carry the proteins to another membrane-bounded organelle, the Golgi complex (see Figure 1-18). As detailed in Chapter 14, secreted proteins and membrane proteins undergo a series of enzyme-catalyzed chemical modifications in the Golgi complex that are essential for these proteins to function normally. After these proteins are modified in the Golgi complex, they are transported out of the complex by a second set of small, diameter vesicles, which bud from one side of the Golgi complex. These vesicles carry proteins to the plasma membrane or to lysosomes or to other organelles, or they release proteins from the cell into the extracellular space. How intracellular transport vesicles are formed, how they target specific proteins to specific membranes, and how they fuse with these membranes to deliver their contents is also discussed in
Chapter 14. Endosomes Bring Proteins and Particles from the Outside into Cells
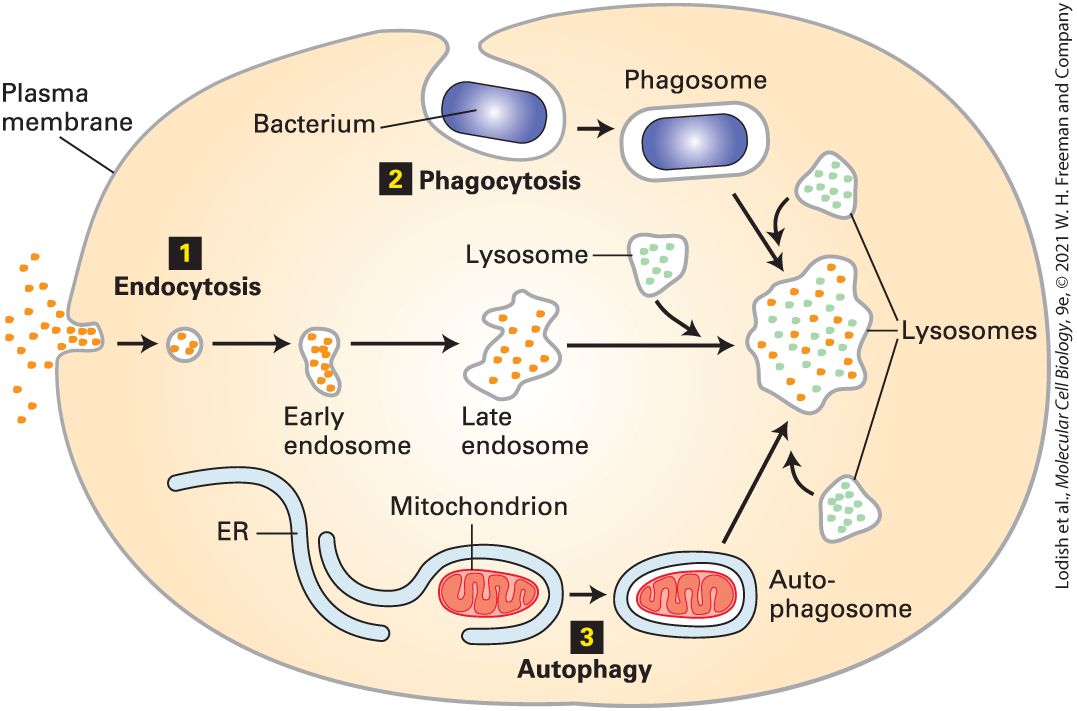
Although transport proteins in the plasma membrane mediate the movement of ions and small molecules into the cell across the phospholipid bilayer, proteins, particles such as viruses, and some other soluble macromolecules in the extracellular milieu are brought into the cell by endocytosis. In this process, a segment of the plasma membrane buds inward — a process termed invagination — into a coated pit, whose cytosolic face is lined by a specific set of proteins that cause these vesicles to form. The pit pinches from the membrane into a small, - diameter membrane-bounded vesicle that contains the extracellular material. The vesicle is delivered to and fuses with an endosome, a sorting station formed of a vesicle and membrane-lined tubules (Figure 1-19). From this compartment, some membrane proteins are recycled back to the plasma membrane; other membrane proteins are transported in vesicles that eventually fuse with lysosomes for degradation. The entire endocytic pathway is described in detail in Chapter 14.
FIGURE 1-19 Endosomes and other cellular structures deliver materials to lysosomes. Schematic overview of three pathways by which materials are moved to lysosomes. Within the acidic lumen of a lysosome, hydrolytic enzymes degrade proteins, nucleic acids, lipids, and other large molecules. 1 Soluble macromolecules and molecules bound to proteins on the cell surface are taken into the cell by invagination of segments of the plasma membrane and delivered to lysosomes through the endocytic pathway; endosomes fuse with lysosomes, intermixing their contents. 2 Whole cells and other large, insoluble particles move from the cell surface to lysosomes through the phagocytic pathway; phagosomes fuse with lysosomes, intermixing their contents. 3 Worn-out organelles and bulk cytoplasm are delivered to lysosomes through the autophagic pathway. Description Three numbered pathways are depicted. The information presented is as follows:
Lysosomes Are Cellular Recycling Centers
1. In endocytosis, the plasma membrane encapsulates extra-cellular material, bringing these into the cell. This encapsulated early endosome matures to the late endosome, after which it merges with lysosomes, digesting the contents. 2. Phagocytosis. The plasma membrane encapsulates a bacterium, which is transported in a phagosome to the lysosomes. 3. Autophagy. The endoplasmic reticulum encapsulates a mitochondrion within the cell and transports it to the lysosomes. Lysosomes Are Cellular Recycling Centers Lysosomes contain a group of enzymes that degrade polymers into their monomeric subunits. Found exclusively in animal cells, lysosomes are responsible for degrading many components that have become obsolete for the cell or organism. As examples, nucleases degrade RNA and DNA into their mononucleotide building blocks; proteases degrade a variety of proteins and peptides into amino acids; phosphatases remove phosphate groups from mononucleotides, phospholipids, and other compounds; still other enzymes degrade complex polysaccharides and lipids into smaller units. The small molecules so generated are usually reutilized by the cell to make new proteins, nucleic acids, and carbohydrates. All of these lysosomal enzymes, collectively termed acid hydrolases, work most efficiently at acidic values. The acidic found within lysosomes helps to denature proteins, making them accessible to the action of the lysosomal hydrolases. These enzymes are less active at the neutral of cells and most extracellular fluids. Thus if a lysosome
Plant Vacuoles Store Water, Ions, and Small-Molecule Nutrients Such as Sugars and Amino Acids
releases its enzymes into the cytosol, where the is between 7.0 and 7.3, they cause little degradation of cytosolic components. The low of lysosomes is generated by proteins in the lysosomal membrane that pump protons ( ions) from the cytosol into the lumen of the lysosome, as detailed in Chapter 11. Lysosomes provide an excellent example of the ability of intracellular membranes to form closed compartments in which the composition of the lumen (the aqueous interior of the compartment) differs substantially from that of the surrounding cytosol. Aged organelles are degraded in a lysosome by a process called autophagy (“eating oneself”). Materials taken into a cell by endocytosis or phagocytosis may also be degraded in lysosomes (see Figure 1-19). In phagocytosis, large, insoluble particles (e.g., bacteria) are enveloped by the plasma membrane and internalized, and the particles are quickly delivered to lysosomes for degradation. Cytosolic and nuclear proteins generally are not degraded in lysosomes, but rather in proteasomes, large multiprotein complexes in the cytosol (see Chapter 3). Plant Vacuoles Store Water, Ions, and Small-Molecule Nutrients Such as Sugars and Amino Acids Most plant cells contain at least one membrane-bounded vacuole. A variety of membrane proteins in the vacuolar membrane transport water, ions, and small-molecule nutrients such as sugars and amino acids from the cytosol into the vacuole lumen. The number and size of vacuoles
depend on both the type of cell and its stage of development; a single vacuole may occupy as much as 80 percent of a mature plant cell (Figure 1-20). Like that of a lysosome, the lumen of a vacuole contains a battery of degradative enzymes and has an acidic , which is maintained by similar transport proteins in the vacuolar membrane. Thus plant vacuoles may also have a degradative function similar to that of lysosomes in animal cells. Similar storage vacuoles are found in green algae and in many microorganisms such as fungi.
FIGURE 1-20 Electron micrograph of a thin section of a leaf cell. In this cell, a single large vacuole occupies much of the cell volume. Parts of five chloroplasts and the cell wall are also visible. Note the internal subcompartments in the chloroplasts. Description The cell is surrounded by a thick cell wall. The large space within the cell is labeled vacuole. Several large, round structures are labeled chloroplasts. Within the
Peroxisomes and Plant Glyoxisomes Metabolize Fatty Acids and Other Small Molecules Without Producing ATP from ADP and Pi
chloroplasts, many dark, stacked structures are labeled granum. The colorless fluid surrounding the grana are labeled stroma. The single unstacked dark structure inside the chloroplast is labeled thylakoid membrane. The scale bar reads, 2 micrometers. Peroxisomes and Plant Glyoxisomes Metabolize Fatty Acids and Other Small Molecules Without Producing ATP from ADP and P All animal cells (except erythrocytes) and many plant and fungal cells contain peroxisomes, a class of roughly spherical organelles in diameter. A major function of peroxisomes is to catabolize (degrade) long-chain fatty acids to . As discussed in Chapter 12, mitochondria also contain enzymes that degrade fatty acids to , but in contrast to mitochondria, peroxisomes do not synthesize ATP from ADP and . Several of the enzymes in peroxisomes are oxidases that use molecular oxygen to add oxygen atoms — a process termed oxidation — to a variety of organic substances and in the process form hydrogen peroxide , a corrosive substance. Peroxisomes also contain copious amounts of the enzyme catalase, which degrades hydrogen peroxide to yield water and oxygen (see Chapter 12). Plant seeds contain glyoxisomes, small organelles that oxidize stored lipids as a source of carbon and energy for growth. They are similar to i
Mitochondria Are the Principal Sites of ATP Production in Aerobic Cells
peroxisomes and contain many of the same types of enzymes as well as additional ones used to convert fatty acids into glucose precursors. Mitochondria Are the Principal Sites of ATP Production in Aerobic Cells The activities of all parts of a cell are powered by the energy stored in ATP molecules. The main sites of ATP production during aerobic metabolism (metabolism requiring oxygen) are the mitochondria (Figure 1-21), which can occupy up to 25 percent of the volume of the cytoplasm. These complex organelles are generally exceeded in size only by the nucleus, vacuoles, and chloroplasts. The two membranes that are found in all mitochondria differ in composition and function. The outer mitochondrial membrane contains proteins that allow many molecules to move from the cytosol to the intermembrane space between the inner and outer membrane. The inner mitochondrial membrane, which is much less permeable, is about 20 percent lipid and 80 percent protein — a proportion of protein that is higher than those in other cellular membranes. The surface area of the inner membrane is greatly increased by a large number of infoldings, or cristae, that protrude into the matrix, the central aqueous space inside the inner membrane. The large number of proteins in the inner membrane are needed to complete the aerobic degradation of fuels to form ATP from ADP and inorganic phosphate (see Figure 1-7 and
FIGURE 1-21 Electron micrograph of a mitochondrion in a pancreas cell. The smooth outer membrane forms the outside boundary of the mitochondrion. The inner membrane is distinct from the outer membrane and is highly invaginated to form sheets and tubes called cristae; ATP is produced by proteins embedded in the membranes of the cristae. The aqueous space between the inner and outer membranes (the intermembrane space) and the space inside the inner membrane (the matrix) each contain specific proteins important for the metabolism of sugars, lipids, and other molecules. Description The mitochondrion has a double-walled outer membrane. The space between the inner and outer membranes is labeled intermembrane space. The inner membrane folds over many times to form the cristae. Several dark spots in the mitochondrion are labeled
Chloroplasts Contain Internal Compartments in Which Photosynthesis Takes Place
matrix granules. The interior of the mitochondrion is labeled the matrix. The scale reads, 3 micrometers. In non-photosynthetic cells, the principal fuels for ATP synthesis are fatty acids and glucose. The complete aerobic degradation of 1 molecule of glucose to carbon dioxide and water is coupled to the synthesis of as many as 30 molecules of ATP from ADP and inorganic phosphate (see Figure 17). In eukaryotic cells, the initial stages of glucose degradation take place in the cytosol, where 2 ATP molecules per glucose molecule are generated. The terminal stages of oxidation and ATP synthesis are carried out by enzymes in the mitochondrial matrix and inner membrane (see Chapter 12); as many as 28 ATP molecules per glucose molecule are generated in mitochondria. Similarly, virtually all the ATP formed in the oxidation of fatty acids to carbon dioxide is generated in mitochondria. Thus mitochondria can be regarded as the power plants of the cell. As discussed in Chapter 12, the popular endosymbiont hypothesis postulates that mitochondria originated by endocytosis of an ancient aerobic bacterium by the precursor of a eukaryotic cell; the bacterial plasma membrane evolved to become the inner mitochondrial membrane. Indeed, mitochondria contain small DNA molecules that encode a small number of mitochondrial proteins and that are thought to have descended from the DNA genome of this ancient bacterium. Chloroplasts Contain Internal Compartments in Which
Photosynthesis Takes Place Except for vacuoles, chloroplasts are the largest and the most characteristic organelles in the cells of plants and green algae (see Figure 1-20). The endosymbiont hypothesis (see Chapter 12) posits that these organelles originated by endocytosis of a primitive photosynthetic bacterium. Chloroplasts can be as long as and are typically wide, but they vary in size and shape in different cells, especially among the algae. In addition to the inner and outer membranes that bound a chloroplast, this organelle also contains an extensive internal system of interconnected membrane-bounded vesicles called thylakoids, which are flattened to form disks. Thylakoids often form stacks called grana and are embedded in an aqueous matrix termed the stroma. The thylakoid membranes contain green pigments (chlorophylls) and other pigments that absorb light, as well as enzymes that generate ATP from ADP and inorganic phosphate during photosynthesis. Some of the ATP is used to convert carbon dioxide into three-carbon intermediates by enzymes located in the stroma; the intermediates are then exported to the cytosol and converted into sugars. The molecular mechanisms by which ATP is formed in mitochondria and chloroplasts are very similar, as explained in Chapter 12. Besides being surrounded by two membranes, chloroplasts and mitochondria have other features in common: both often migrate from place to place within cells, and both contain their own DNA, which encodes some of the key organelle proteins (see Chapter 12). The proteins encoded by mitochondrial or
Many Organelle-Like Structures Are Unbounded by a Membrane
chloroplast DNA are synthesized on ribosomes within the organelles. However, most of the proteins in each organelle are encoded in nuclear DNA and are synthesized in the cytosol; these proteins are then incorporated into the organelles and directed to the correct membrane or space by processes described in Chapter 13. Many Organelle-Like Structures Are Unbounded by a Membrane In addition to membrane-enveloped organelles, eukaryotic cells contain a variety of substructures in which important biochemical functions are concentrated but that are not surrounded by a membrane. These membraneless organelles range in size from 0.1–3 micrometers in diameter. One ubiquitous example is the nucleolus, a subcompartment of the nucleus where ribosomal RNA is synthesized and ribosomes are assembled (see Figure 1-16b and Chapter 9). Other subcompartments of the nucleus have recently been made visible by the new microscopic techniques detailed in Chapter 4. These include nuclear speckles, sites where RNA polymerase and other gene regulatory proteins are concentrated and where messenger RNAs are synthesized and processed. Subcompartments of the cytosol include P bodies, sites where many messenger RNAs are degraded, and cytosolic stress granules, membraneless organelles where some messenger RNAs are stored awaiting a signal to be translated into a protein.
All Eukaryotic Cells Use a Similar Cycle to Regulate Their Division
Although much remains to be learned about the organization of these membraneless organelles, recent evidence suggests that they are formed by many weak but specific interactions between proteins and nucleic acids that exclude the surrounding aqueous phase. Their properties are characteristic of liquid droplets suspended in another, chemically distinct liquid, and they are often referred to as biomolecular condensates (see
Chapter 3). Their structures are highly dynamic, in that proteins can diffuse into and out of these condensates. Clearly much needs to be learned about their formation, structure, and biological functions. All Eukaryotic Cells Use a Similar Cycle to Regulate Their Division Unicellular eukaryotes, animals, and plants all use essentially the same cell cycle, the series of events that prepares a cell to divide, and the same actual division process, called mitosis. The eukaryotic cell cycle is commonly divided into four phases (Figure 1-22). The chromosomes and the DNA they carry are duplicated during the S (synthesis) phase. The replicated chromosomes condense (see Figure 1-17) and separate during the M (mitotic) phase during which the cell divides, and each daughter cell gets one copy of each chromosome. The M and S phases are separated by two gap phases, the phase and the phase, during which mRNAs, proteins, lipids, and other cell constituents are made and the cell increases in size.
FIGURE 1-22 During growth, all eukaryotic cells continually progress through the four phases of the cell cycle. In proliferating cells, the four phases of the cell cycle proceed successively. In humans, the cycle takes from 10 to 20 hours depending on cell type and developmental state. Yeasts divide much faster. During interphase, which consists of the , , and phases, the cell roughly doubles its mass. Replication of DNA during the S phase leaves the cell with two copies of each type of chromosome (see Figure 1-17c). In the mitotic (M) phase, the chromosomes are evenly partitioned into two daughter cells, and in most cases the cytoplasm divides roughly in half by division along the midline. Under certain conditions, such as starvation or when a tissue has reached its final size, cells will stop cycling and remain in a waiting state called . Some types of cells in can reenter the cell cycle if conditions change. Description The different phases are labeled G subscript 1, S, G subscript 2, and M. M leads back to G subscript 1 and the cycle continues. The different activities during each phase are as follows: G subscript 1: R N A, lipid, and protein synthesis; cellular components excluding the chromosomes are duplicated.
S phase: D N A replication. Each of the chromosomes is duplicated. G subscript 2 phase: Additional R N A, lipid, S and protein synthesis. Cell monitors duplicated chromosomes for errors in D N A sequence and repairs damage. M phase: cell division takes place. M phase leads to G subscript 1 phase where cell cycle arrest takes place which leads to G subscript 0, followed by re-entry into the cell cycle. Under optimal conditions, some bacteria, such as E. coli, can divide to form two daughter cells once every 20 minutes. Most eukaryotic cells take considerably longer to grow and divide, generally several hours. Moreover, the cell cycle in eukaryotes is highly regulated (see Chapter 19). This tight control prevents imbalanced, excessive growth of cells and tissues if essential nutrients or certain hormonal signals are lacking. Some highly specialized cells in adult animals, such as neurons and heart muscle cells, divide rarely, if at all. However, an organism usually replaces wornout cells or makes more cells in response to a new need — for example, new muscle cells are generated from undifferentiated stem cells in response to exercise or damage. Another example is the formation of additional red blood cells when a person ascends to a higher altitude and needs more capacity to capture oxygen. The fundamental defect in cancer is loss of the ability to control the growth and division of cells. In Chapter 25, we examine the molecular and cellular events that lead to inappropriate, uncontrolled proliferation of cells.
1.4 Unicellular Eukaryotic Organisms Widely Used in Cell Biology Research
1.4 Unicellular Eukaryotic Organisms Widely Used in Cell Biology Research Our current understanding of the molecular functioning of eukaryotic cells largely rests on studies of just a few types of organisms, often termed model organisms (Figure 1-23). Because of the evolutionary conservation of genes, proteins, organelles, cell types, and so forth, discoveries about biological structures and functions obtained with one experimental organism often apply to others. Thus researchers generally conduct studies with the organism that is most suitable for rapidly and completely answering the question being posed, knowing that the results obtained in one organism are likely to be broadly applicable.
FIGURE 1-23 Many eukaryotic organisms used in cell biology research have advantages for certain types of studies. The yeast Saccharomyces cerevisiae (a) has the cellular organization of a eukaryote but is a relatively simple single-celled organism that is easy to grow and to manipulate genetically. The green alga Chlamydomonas reinhardtii (b) is widely used to study photosynthesis and the structure and function of flagella. In the roundworm Caenorhabditis elegans (c), which has a small number of cells arranged in a nearly identical way in every worm, the formation of each individual cell can be traced. The fruit fly Drosophila melanogaster (d), first used to discover the properties of chromosomes, has been especially valuable in identifying genes that control embryonic development. Many of these genes are evolutionarily conserved in humans. Planaria (e) are flatworms that can regenerate any part of the body that is cut off, including the head and the photoreceptors. The stem cells that give rise to their new cells and tissues are widely
studied. The zebrafish Danio rerio (f) is used for rapid genetic screens to identify genes that control vertebrate development and organogenesis. Of the experimental animal systems, mice (Mus musculus) (g) are evolutionarily the closest to humans and have thus provided models for studying numerous human genetic and infectious diseases. The mustard-family weed Arabidopsis thaliana (h) has been used for genetic screens to identify genes involved in nearly every aspect of plant life. Description The information presented is as follows: A. Yeast (Saccharomyces cerevisiae): Control of cell cycle and cell division. Protein secretion and membrane biogenesis. Function of the cytoskeleton- Cell differentiation, Aging Gene regulation, and chromosome structure. B. Alga (Chlamydomonas reinhardtii): Structure and function of flagella; Chloroplasts and photosynthesis; Organelle movement; Phototaxis. C. Roundworm (Caenorhabditis elegans): Development of the body plan; Cell lineage; Formation and function of the nervous system; Control of programmed cell death; Cell proliferation and cancer genes; Aging; Behavior; Gene regulation and chromosome; Structure. D. Fruit fly (Drosophila melanogaster): Development of the body plan; Generation of differentiated cell lineages; Formation of the nervous system and musculature; Programmed cell death; Genetic control of behavior; Cancer genes and control of cell proliferation; Control of cell polarization; Effects of drugs, alcohol, pesticides. E. Planarian ((Schmidtea mediterranea): Stem cells; Turnover of adult tissues; Wound healing; Regeneration. The pharynx and the photoreceptors are labeled. F. Zebrafish (Danio rerio): Development of vertebrate body tissues; Formation and function of brain and nervous system; Birth defects; Cancer. G. Mouse (Mus musculus), including cultured cells: Development of body tissues; Function of mammalian immune System; Formation and function of brain and nervous system; Models of cancers and other human diseases; Gene regulation and inheritance; Infectious disease; Behavior.
Yeasts Are Used to Study Fundamental Aspects of Eukaryotic Cell Structure and Function
H. Plant (Arabidopsis thaliana): Development and patterning of tissues; Agricultural applications; Physiology; Gene regulation; Immunity; Infectious disease. As we have seen, bacteria are excellent experimental systems for studies of several cellular functions, but they lack the organelles found in eukaryotes. Unicellular eukaryotes such as yeasts are used to study many fundamental aspects of eukaryotic cell structure and function. Metazoan organisms such as the roundworm, fruit fly, and mouse are required to study more complex tissue and organ systems and development. As we will see in this section and the next, these and other metazoans are widely used to understand complex cell systems and mechanisms. Yeasts Are Used to Study Fundamental Aspects of Eukaryotic Cell Structure and Function One group of single-celled eukaryotes, the yeasts, has proven exceptionally useful in molecular and genetic analysis of eukaryotic cell formation and function. Yeasts and their multicellular cousins, the molds, are fungi. Fungi have important ecological roles in breaking down plant and animal remains for reuse. They also make numerous antibiotics and are used in the manufacture of bread, beer, and wine. The common yeast used to make bread and beer, Saccharomyces cerevisiae (Figure 1-24a), appears frequently in this book because it has proved to be an extremely useful experimental organism. S. cerevisiae is a
spherical cell that divides by budding of a daughter cell from a side of the mother cell, eventually breaking free (Figure 1-24a, b). Homologs of many of the approximately 6600 different proteins expressed in an S. cerevisiae cell (see Table 1-2) are found in most, if not all, eukaryotes and are important for cell division or for the functioning of individual eukaryotic organelles. Much of what we know of the proteins in the endoplasmic reticulum and Golgi complex that promote protein secretion was elucidated first in genetic studies of yeasts (see Chapter 14). Yeasts were also essential for the identification of many proteins that regulate the cell cycle and catalyze DNA replication and transcription.
FIGURE 1-24 The yeasts Saccharomyces cerevisiae and Schizosaccharomyces pombe can be haploid or diploid and can reproduce sexually or asexually. (a) Scanning electron micrograph of the budding yeast Saccharomyces cerevisiae. These cells grow by an unusual type of mitosis termed Smitotic budding. One daughter nucleus remains in the mother cell; the other daughter nucleus is transported into the bud, which grows in size and soon is released as a new cell. After each bud cell breaks free, a scar is left at the budding site, so the number of previous buds on the parent cell can be counted. The orange-colored cells are bacteria. (b) Haploid S. cerevisiae cells can have different mating types, called a
(blue) and (orange). Both types contain a single copy of each yeast chromosome, half the usual number, and grow by mitotic budding. Two haploid cells that differ in mating type, one a and one , can fuse together to form an diploid cell. An a haploid cell secretes a pheromone protein termed the a mating factor that binds to a receptor protein on the surface of an cell, and conversely an cell secretes the factor protein that binds to a different surface receptor on the a cell. Thus each type of cell recognizes the mating factor produced by the opposite type, and these hormones trigger fusion of the a and cell. The diploid cell contains two copies of each chromosome; diploid cells can multiply by mitotic budding in a manner similar to that of the haploid cells. Under starvation conditions, a diploid cell can undergo meiosis, a special type of cell division, to form four haploid ascospores. Rupture of an ascus releases four haploid spores, which can germinate into haploid a and cells. These cells can also multiply asexually. (c) Both haploid and diploid Schizosaccharomyces pombe cells have a mitotic division cycle different from that of S. cerevisiae, one that more resembles that of human cells than that of S. cerevisiae. Bracket indicates a cell undergoing mitotic division. Description The micrograph labeled A shows the ovoid-shaped Saccharomyces cerevisiae with budding scars and bacteria in the background. The illustration labeled B shows the steps involved in the life cycle of yeast cells. The steps involved are as follows: Step 1. Mating between haploid cell or opposite mating type, for example, a and alpha, forming diploid cells. Step 2. Vegetative growth of diploid cells; cells bud to form daughter diploid cells. Step 3. Starvation causes ascus formation and induces meiosis. Four haploid ascopores are contained with the ascus. Step 4. The ascus ruptures and the spores germinate. Step 5. Vegetative growth of haploid cells. Step 5 leads back to step 1 and thus the cycle continues.
The micrograph labeled C shows a cell undergoing mitotic division. The yeast species Schizosaccharomyces pombe diverged from S. cerevisiae between 300 and 600 million years ago and is also widely used in research. Unlike S. cerevisiae, S. pombe is a rod-shaped cell that grows by elongation at the ends; like human cells it divides by division at the cell midpoint (Figure 1-24c; see also Figure 1-22). Like human cells, but unlike S. cerevisiae, most genes in S. pombe have introns that are removed from RNA transcripts by processing in the nucleus (see Figure 1-10). S. cerevisiae, S. pombe, and other yeasts offer many advantages to molecular and cellular biologists: Vast numbers of yeast cells can be grown easily and cheaply in culture from a single cell; the cells in such clones are genetically identical and have the same biochemical properties. Individual proteins or multiprotein complexes can be purified from large amounts of cells and then studied in detail. Yeast cells may be either haploid (containing one copy of each chromosome) or diploid (containing two copies of each chromosome), and both forms can divide by mitosis; this ability makes isolating and characterizing mutations in genes encoding essential yeast cell proteins relatively straightforward, as we see in the next section. Yeasts, like many organisms, have a sexual cycle that allows exchange of genes between cells. Under starvation conditions, diploid cells undergo meiosis (see Chapter 19) to form haploid daughter cells. Haploid S. cerevisiae cells are of two types, a and cells. If haploid a and cells come close to one another, they secrete
Mutations in Yeast Led to the Identification of Key Cell Cycle Proteins
hormones that induce the two cells to fuse, forming an diploid cell that contains two copies of each chromosome, one from each parent cell (Figure 1-24b). With the use of a single species such as S. cerevisiae as an experimental organism, results from studies carried out by tens of thousands of scientists worldwide, using multiple investigational techniques, can be combined to yield a deeper level of understanding of a single type of cell. As we will see many times in this book, conclusions based on studies of S. cerevisiae and S. pombe have often proved true for all eukaryotes. These conclusions have provided a basis for exploring the evolution of more complex processes in multicellular animals and plants. Mutations in Yeast Led to the Identification of Key Cell Cycle Proteins Biochemical studies can tell us much about an individual protein, but they cannot prove that it is required for cell division or any other cell process. The importance of a protein is demonstrated most firmly if a mutation that prevents its synthesis or makes it nonfunctional adversely affects the process under study. In a classical genetics approach, scientists isolate and characterize mutants that lack the ability to do something a normal organism can do. Often large genetic screens are done to look for many different mutant
individuals (e.g., fruit flies, yeast cells) that are unable to complete a certain process, such as cell division or muscle formation. Mutations are usually produced by treatment with a mutagen, a chemical or physical agent that promotes mutations in a largely random fashion. However, inactivating genes that encode proteins for an essential cell process will kill the organism. How then can we isolate and maintain mutant organisms or cells that are defective in some process, such as cell division or protein secretion, that is essential for survival? One way is to isolate organisms with a temperature-sensitive mutation. These mutants are able to grow at the permissive temperature, but not at another, usually higher temperature, the nonpermissive temperature. Normal cells can grow at either temperature. In most cases, a temperaturesensitive mutant produces an altered protein that works at the permissive temperature but unfolds and is nonfunctional at the nonpermissive temperature. Screens for temperature-sensitive mutations are most readily done with haploid organisms such as yeasts because they have only one copy of each gene, and thus a mutation in it will immediately have a consequence. By analyzing the effects of numerous different temperature-sensitive mutations that altered the division of haploid yeast cells, geneticists discovered most of the genes necessary for cell growth and division without knowing anything, initially, about which proteins they encode or how these proteins participate in the process. In general, the great power of genetics is to reveal the existence and relevance of all proteins required for a particular cell function without prior knowledge of their biochemical
identity or molecular function. These mutation-defined genes can be isolated using recombinant DNA techniques discussed in Chapter 6. With the isolated genes in hand, the encoded proteins can be produced in a test tube or in engineered bacteria or cultured cells. In this way, biochemists can investigate whether the genes necessary for cell division encode proteins that associate with other proteins or DNA or catalyze particular chemical reactions during cell division (see Chapter 19). Most of these yeast cell cycle genes are found in human cells as well, and the encoded proteins have similar amino acid sequences. Proteins from different organisms, but with similar amino acid sequences, are said to be homologous; such proteins often have the same or similar functions. Remarkably, it has been shown that a human protein that regulates the cell cycle, when expressed in a mutant yeast defective in the homologous yeast protein, is able to rescue the defect of the mutant yeast (i.e., to allow the cell to grow normally), thus demonstrating the protein’s ability to function in a very different type of eukaryotic cell. This experimental result, which together with identification and characterization of many other cell cycle genes garnered a Nobel Prize for Paul Nurse, was especially notable because the common ancestor of present-day yeasts, plants, and humans is thought to have lived over a billion years ago. Clearly the eukaryotic cell cycle and many of the genes and proteins that catalyze and regulate it evolved early in biological evolution and have remained quite constant over a very long period of evolutionary time. Subsequent studies showed that mutations in many yeast cell cycle proteins that allow uncontrolled cell growth also frequently occur in human cancers (see Chapter 25), again
Studies in the Alga Chlamydomonas reinhardtii Led to the Development of a Powerful Technique to Study Brain Function
attesting to the important conserved functions of these proteins in all eukaryotes. Studies in the Alga Chlamydomonas reinhardtii Led to the Development of a Powerful Technique to Study Brain Function The green unicellular alga Chlamydomonas reinhardtii (see Figure 1-23b), which swims using its two long flagella, is widely used in studies of the structure, function, and assembly of this organelle. In part because of the powerful genetic techniques now available, Chlamydomonas is also used in studies of chloroplast formation and photosynthesis. The Chlamydomonas genome (see Table 1-2) encodes many more proteins than do those of yeasts, including flagellar proteins and proteins needed to build a chloroplast, organelles not found in yeasts. One important outcome of the use of this experimental organism came from studies of phototaxis, the behavior in which an organism moves toward or away from a source of light. Chlamydomonas needs to move toward light to undergo photosynthesis and thus generate the energy it needs to grow and divide, but light that is too intense repels it, as it causes damage to the chloroplast. Studies of Chlamydomonas phototaxis led to the discovery of two proteins in its plasma membrane that, when they absorb light, open a channel in the membrane that allows ions such as
The Parasite That Causes Malaria Has Novel Organelles That Allow It to Undergo a Remarkable Life Cycle
to flow from the extracellular medium into the cytosol, triggering phototactic responses. As detailed in Chapter 23, recombinant DNA techniques have been used to express one such protein in specific neurons in the mouse brain, allowing investigators to activate just one or a few cells in the brain using a point source of light. Thus studies on this humble alga have led to the development of an important experimental system — optogenetics — for the study of brain function. The Parasite That Causes Malaria Has Novel Organelles That Allow It to Undergo a Remarkable Life Cycle Whereas yeasts are used in the manufacture of bread, beer, wine, and cheese, some unicellular eukaryotes cause major human diseases and are widely studied in an attempt to develop vaccines or drugs that will kill them but not injure their human host. Entamoeba histolytica causes dysentery; Trichomonas vaginalis, vaginitis; and Trypanosoma brucei, sleeping sickness. Each year the worst of these protozoans, Plasmodium falciparum and related species, cause more than 200 million new cases of malaria, a disease that kills 1.5 million to 3 million people annually. These protozoans inhabit mammals and mosquitoes alternately, changing their morphology and behavior in response to signals in each of these environments. The complex life cycle of Plasmodium dramatically illustrates how a single cell can adapt to multiple environments (Figure 1-25a).
Additionally, the merozoite form that infects human red blood cells contains several organelles, not found in most eukaryotes, that enable the parasite to invade a red blood cell, including the rhoptries, polar ring, and micronemes, as well as a fuzzy surface coat on the outside of the cell (Figure 1-25b, c). Entry of the parasite into a red blood cell is initiated by binding of certain parasite cell-surface proteins to proteins on the red blood cell surface, accompanied by the secretion of proteins stored in the micronemes and rhoptries. A tight junction is formed between the two plasma membranes, and the fuzzy coat is lost as parasites enter the host cell (Figure 1-25c).
FIGURE 1-25 Plasmodium species, the parasites that cause malaria, are single-celled protozoans with a remarkable life cycle. Many Plasmodium species are known, and they can infect a variety of animals, cycling between insect and vertebrate hosts. The four species that cause malaria in humans undergo several dramatic transformations within their human and mosquito hosts. (a) Diagram of the life cycle. Like many yeasts, Plasmodium spends most of its life cycle as a haploid organism, with a single copy of most genes. Step 1 : Haploid sporozoites enter a human host when an infected Anopheles mosquito bites a person. Step 2 : They migrate to the liver, where they develop into merozoites, which are released into the blood. Merozoites differ substantially from sporozoites, so this transformation is a metamorphosis (Greek, “to transform” or “many shapes”). Step 3 : Circulating merozoites invade red blood cells (RBCs) and reproduce within them. Proteins produced by some Plasmodium species move to the surface of infected RBCs, causing the cells to adhere to the walls of blood vessels. This prevents infected RBCs from circulating to the spleen, where cells of the immune system would destroy the RBCs and the Plasmodium organisms they harbor. Step 4 : After growing and reproducing in RBCs for a period of time characteristic of each Plasmodium species, the merozoites suddenly burst forth in synchrony from large numbers of infected cells. It is this event that brings on the fevers and shaking chills that are the well-known symptoms of malaria. Some of the released merozoites infect additional RBCs, creating a cycle of reproduction and infection. Step 5 : Eventually, some merozoites develop into male and female gametocytes, another metamorphosis. These cells do not replicate further, waiting to be transferred to an Anopheles mosquito during a bloodmeal. Step 6 : In the mosquito’s stomach, the gametocytes are transformed into microgametocytes (sperm) or macrogametocytes (eggs), yet another metamorphosis marked by development of slender motile sperm. Step 7 : Fusion of sperm and eggs generates the diploid zygotes, which implant into the lining of the stomach and grow into oocysts. Within the oocysts, parasites undergo meiosis, returning to a haploid chromosome content, followed by several rounds of mitosis that essentially are factories for producing sporozoites. Step 8 : Rupture of an oocyst releases thousands of sporozoites, which migrate to the salivary glands, setting the stage for infection of another human host. (b) Organelles of the Plasmodium vivax merozoite. Some of these organelles are found only in Plasmodium and related eukaryotic parasitic microorganisms. (c) Section of a Plasmodium vivax merozoite invading a human red blood cell. See A. Cowman and B. Crabb, 2006, Cell 124:755–766.
Description The steps in the life cycle of the plasmodium parasite are as follows: Step 1. Sporozoites enter human tissue by the bite of an infected mosquito. The sporozoites travel to the liver. Step 2. Formation of merozoites. Step 3. Infection and growth within red blood cells. Step 4. Sporulation and rupture, leading to release of merozoites. These can reinfect red blood cells or move to the next step. Step 5. The formation of gametocytes. Step 6. Gametocytes mature into macrogametocytes (eggs) and microgametocytes (sperm), which infect a mosquito upon feeding. Step 7. A zygote forms inside the mosquito, leading to: Step 8. The formation of an oocyst. The oocyst eventually ruptures, and the release of sporozoites restarts the cycle at step 1. The schematic shows a teardrop-shaped, single-celled organism. Several organelles are highlighted: the nucleus, ribosomes, dense granules; the plasma membrane containing the organelles; the exterior surface coat, plastids, mitochondria, rhoptry, microneme, and the polar rings. The micrograph shows a merozoite entering a red blood cell. The merozoite has partially invaded the cell, and tight junctions grip the exterior surface of the merozoite. The rhoptry is labeled within the head of the merozoite. All the cellular transformations that occur during the Plasmodium life cycle are governed by instructions encoded in the genetic material of this parasite (see Table 1-2). The Plasmodium genome has about the same number of protein-coding genes as the yeast S. cerevisiae, but about two-
thirds of the Plasmodium genes appear to be unique to this and related parasites. Their many unique genes attest to the great evolutionary distance between these parasites, members of the genus Apicomplexa (see
Figure 1-1), and most other eukaryotes as well as the presence of unusual organelles required for their complex life cycles.
Multicellularity Requires Cell-Cell and Cell-Matrix Adhesions
1.5 Metazoan Structure, Function, Evolution, and Differentiation The evolution of multicellular organisms most likely began when cells remained associated in small colonies after division instead of separating into individual cells. A few prokaryotes and the colonial algae Volvox (see
Figure 1-3d), as well as many fungi and slime molds, exhibit such rudimentary social behavior. The full flowering of multicellularity, however, occurred in the eukaryotic organisms called metazoans whose cells became differentiated and organized into groups, or tissues, in which the different cells performed specialized functions. In this section, we describe some of the principal features of metazoans: adhesion of cells to each other, communication between cells, and regulation of development such that the correct cells are produced at the right time in the right place in the organism. Multicellularity Requires Cell-Cell and Cell-Matrix Adhesions The cells of higher plants are encased in a network of chambers formed by the interlocking cell walls surrounding the cells and are connected by cytoplasmic bridges called plasmodesmata (see Figure 1-13a). Animal cells are often “glued” together into a chain, a ball, or a sheet by celladhesion proteins on their surfaces, often called cell-adhesion molecules,
Epithelia Originated Early in Evolution
or CAMs (see Figure 1-4d and Chapter 20). Some CAMs bind cells to one another; other types bind cells to the fibrous proteins and sugar polymers (polysaccharides) that form the extracellular matrix, forming a cohesive unit. In animals, the matrix cushions cells and allows nutrients to diffuse toward them and waste products to diffuse away. A specialized, especially tough matrix called the basal lamina, made up of polysaccharides and multiple proteins such as collagen, forms a supporting layer underlying cell sheets and prevents the cell aggregates from ripping apart (see Figure 1-4). Many CAMs and extracellular-matrix proteins found in humans also occur in invertebrates, indicating their importance during metazoan evolution. Similarly, many of the proteins and small molecules used by metazoans as signaling molecules are conserved in humans and many invertebrates, as are their receptors, the cellular proteins that bind to these signaling molecules and trigger an effect in the receiving cell. Epithelia Originated Early in Evolution Metazoans, which are thought to have evolved in an ocean-like, saline environment, had to solve a fundamental problem: separating the inside of the organism from the outside. The external surfaces of all metazoan animals, as well as the surfaces of their internal organs, are covered by a sheet-like layer of tissue called an epithelium. Epithelia commonly serve as barriers and protective surfaces, as exemplified by the sheets of
Cells Are Organized into Tissues and Tissues into Organs
epidermal cells that form the skin (see Figure 1-4). Other epithelia are one cell-layer thick and line internal organs such as the small intestine, where they are crucial for uptake and transport of the products of digestion (e.g., glucose and amino acids) into the blood (see Chapter 11). As discussed in
Chapter 20, epithelia in different body locations have characteristic morphologies and functions. Cells that form epithelial tissues are said to be polarized because their plasma membranes are organized into at least two discrete regions. Typically, the distinct surfaces of a polarized epithelial cell are the apical surface — the “top” of the cell facing the external world — and the basal and lateral (collectively, basolateral) surfaces that face the organism’s interior. As shown in Figure 1-4, the basal surface usually contacts an underlying extracellular matrix, the basal lamina. As discussed in Chapter 20, specialized junction proteins in the basolateral plasma membrane link adjacent cells together and also bind the cells to the basal lamina. Cells Are Organized into Tissues and Tissues into Organs Cells in metazoans do not work in isolation; specialized groups of differentiated cells often form tissues, which are themselves the major components of organs. For example, a blood vessel is composed of layers of different types of tissue. The lumen of a small blood vessel is lined with a sheet-like layer of endothelial cells, or endothelium, which prevents blood cells from leaking out (Figure 1-26). A layer of smooth muscle
tissue encircles the endothelium and basal lamina and contracts to limit blood flow. During times of fright, constriction of smaller peripheral vessels forces more blood to the vital organs. The muscle layer of a blood vessel is wrapped in an outer layer of connective tissue, a network of fibers and cells that encases the vessel walls and protects them from stretching and rupture.
FIGURE 1-26 All organs are organized arrangements of various tissues, as illustrated in this cross section of a small artery (arteriole). Blood flows through the vessel lumen, which is lined by a thin sheet of endothelial cells forming the endothelium and by the underlying basal lamina. This tissue adheres to the overlying layer of smooth muscle tissue;
Genomics Has Revealed Important Aspects of Metazoan Evolution and Cell Function
contraction of the muscle layer controls blood flow through the vessel. A fibrillar layer of connective tissue surrounds the vessel and connects it to other tissues. This hierarchy of tissues is copied in other blood vessels, which differ mainly in the thickness of the layers. The wall of a major artery must withstand much stress and is therefore thicker than that of a minor vessel. The strategy of grouping and layering different tissues is used to build other complex organs as well. In each case, the function of the organ is determined by the specific functions of its component tissues, and each type of cell in a tissue produces the specific groups of proteins that enable the tissue to carry out its functions. Genomics Has Revealed Important Aspects of Metazoan Evolution and Cell Function Metazoans — be they invertebrates such as the fruit fly D. melanogaster and the roundworm C. elegans, or vertebrates such as mice and humans — contain between 13,000 and 26,000 protein-coding genes, about two to four times as many as a yeast (see Table 1-2). Sequencing of entire genomes has shown that many of these genes are conserved among the metazoans, and genetic studies have shown that many of them are essential for the formation and function of specific tissues and organs. Thus many of the organisms listed in Table 1-2 are used to study the roles of these conserved proteins in cell development and function.
The human and mouse genomes encode about the same number of proteins as those of the roundworm C. elegans, frogs, and fish, yet mammalian cells contain about 30 times the DNA of a roundworm and two to three times the DNA of frogs and fish. Only about 10 percent of human DNA encodes proteins. We now know that much of the remaining 90 percent has important functions. As detailed in Chapter 8, many DNA segments, termed enhancers, bind proteins that regulate expression of nearby genes. The expression of many genes is controlled by large numbers of such regulatory sequences; control by multiple enhancers allows each mammalian gene to make the precise amount of mRNA and protein needed in each of many different types of cells. Other segments of DNA are used to synthesize the thousands of RNA molecules that do not direct the synthesis of proteins and whose function in regulating gene expression is only now being uncovered. As an example, hundreds of different kinds of micro-RNAs, 20 to 25 nucleotides long, are abundant in metazoan cells. Each microRNA binds to and represses the activity of a specific set of target mRNAs, either by inhibiting the ability of these mRNAs to be translated into proteins or by triggering their degradation (see Chapter 9). By this means, each of these small RNAs can indirectly regulate the activity of many genes. Some of this nonprotein-coding DNA probably regulates expression of genes that make us uniquely human. Indeed, fish and humans have about the same number of protein-coding genes — about 20,000 — yet as noted above, the human genome is over twice the size of that in fish (see Table 1-2). The human brain can perform complex mental processes such as
reading and writing a textbook. Somehow these 20,000 human genes are exquisitely regulated such that humans produce a brain with about 100,000,000,000 neurons, which communicate with one another at about 100,000,000,000,000 interaction sites termed synapses. Genomics — the study of the entire DNA sequences of organisms — has shown us how close humans really are to Neanderthals and to our nearest nonhominid relatives, the great apes (Figure 1-27). Human DNA is 99 percent identical in sequence to that of chimpanzees and bonobos; the 1 percent difference is about 3,000,000 base pairs, but it somehow explains the obvious differences between our species, such as the evolution of human brains during the past ∼5,000,000 years since we last shared a common ancestor.
FIGURE 1-27 Evolutionary tree connecting monkeys, apes, and humans. This evolutionary tree of humans, Neanderthals, great apes, a small ape, and an Old World monkey was estimated from the divergence among their genomic DNA sequences. Wholegenome DNA sequences were aligned, and the average nucleotide divergence in unique
DNA sequences was estimated. Estimates of the times the different species diverged from each other, indicated at each node, were calculated in millions of years (Myr) based on DNA sequence identity; implies approximately or less. [Data from D. P. Locke et al., 2011, Nature 469:529–533 and K. Prüfer et al., 2014, Nature 505:43.] Description From the base of the tree, the information corresponding to each node is as follows: 25 to 33 million years ago, 0.949- Cercopithecidae Old World monkeys, Rhesus macaque, Macaca mulatta. 18 to 20 million years ago, 0.971- Hylobatidae, Small apes, Gibbon Nomascus leucogenys. 12 to 16 million years ago, 0.974- Sumatran orangutan, Pongo abelii. Node unnamed- Gorilla, Gorilla gorilla. 6 to 8 million years ago, 0.984- Human, Homo sapiens. 0.5 to 0.8 million years ago, 0.9986- Neanderthal. 4.5 to 6 million years ago, 0.990: this node leads to another node which branches into two. Approximately one million year ago, 0.996- Bonobo, Pan paniscus and finally Chimpanzee, Pan troglodytes. The branches from 16 million years ago to 1 million years ago are grouped as "the hominidae (great apes)." Genomics coupled with paleontological findings indicates that humans and mice descended from a common mammalian ancestor that probably lived about 75 million years ago. Nonetheless, both organisms contain about the same number of genes, and about 99 percent of mouse protein-
coding genes have homologs in humans, and vice versa. Over 90 percent of mouse and human genomes can be partitioned into regions of synteny — that is, DNA segments that have the same order of unique DNA sequences and genes along a segment of a chromosome. This observation suggests that much of the gene order in the most recent common ancestor of humans and mice has been conserved in both species (Figure 1-28). Of course, mice are not people; relative to humans, mice have expanded families of genes related to immunity, reproduction, and olfaction, probably reflecting the differences between the human and mouse lifestyles.
FIGURE 1-28 Conservation of synteny between human and mouse. Shown is a 510,000base-pair (bp) segment of mouse chromosome 12 that shares common ancestry with a 600,000-bp section of human chromosome 14. Pink lines connect the conserved unique DNA sequences in the two genomes. Mb, 1 million base pairs. [Data from Mouse Genome Sequencing Consortium, 2002, Nature 420:520.] It’s not only human evolution that interests us! Polar bears live in the Arctic and eat a high-fat diet, mostly composed of seals. Recent genome sequencing allowed researchers to conclude that the most recent common ancestor of polar bears and their brown bear relatives, which live in temperate climates, was present about 500,000 years — or only about
Development Uses a Conserved Set of Master Transcription Factors and Involves Epigenetic Modifications to DNA and Its Associated Histone Proteins
20,000 bear generations — ago. During that rather short evolutionary period the polar bear genome acquired changes in many genes regulating cardiovascular function, fat metabolism, and heart development, allowing it to consume a diet very rich in fats. Development Uses a Conserved Set of Master Transcription Factors and Involves Epigenetic Modifications to DNA and Its Associated Histone Proteins The astute reader will note a paradox in the previous discussion: if indeed most human protein-coding genes are shared with apes and mice, and many with flies and worms, how is it that these organisms look and function so differently? The answer to this question resides in the way genes are regulated during the development of all metazoans from a single cell, the fertilized egg. As we learn in Chapters 7 and 8, each proteincoding gene is associated with regulatory DNA sequences that differ in different organisms. Many of these regulatory sequences bind proteins that direct the expression of the gene, and thus the amount of a protein it makes, in specific types of cells. Some of these proteins are termed master transcription factors; these proteins bind to regulatory DNA sequences, are conserved throughout evolution, and control the development of specific types of cells by activating or repressing groups of genes, often at different or successive stages of development.
During development, many segments of each chromosome undergo enzyme-catalyzed chemical modifications — termed epigenetic modifications — that affect the ability of genes within these segments to be transcribed into RNA. As one example, addition of methyl groups to multiple cytosine bases in a segment of DNA triggers the formation of heterochromatin — condensed, concentrated DNA that cannot be transcribed into RNA. Histone proteins bound to DNA can undergo multiple kinds of enzyme-catalyzed modifications, some enhancing and others inhibiting the ability of RNA polymerase to transcribe the adjacent gene into RNA. As detailed in Chapter 8, many of these modifications are replicated and passed on to daughter cells, a type of cell heredity that does not involve changes in the primary nucleotide sequence of the DNA genome. The early stages in the development of a human embryo are similar to those in the mouse. They are characterized by rapid cell divisions (Figure 1-29) followed by the differentiation of cells into tissues. In all organisms, the embryonic body plan — the spatial pattern of cell types (tissues) and body parts — emerges from two influences: a program of genes that specifies the pattern of the body, and local cell interactions that induce different parts of the program.
FIGURE 1-29 The first few cell divisions of a fertilized egg set the stage for all subsequent development. A developing mouse embryo is shown at the (a) two-cell, (b) four-cell, and (c) eight-cell stages. The embryo is surrounded by supporting membranes. The corresponding steps in human development occur during the first few days after fertilization. With only a few exceptions, animals display axial symmetry; that is, their left and right sides mirror each other. This most basic of patterns is encoded in the genome. Developmental biologists have divided bilaterally symmetric animal phyla into two large groups depending on where the mouth and anus form in the early embryo. Protostomes develop a mouth close to a transient opening in the early embryo (the blastopore) and have a ventral nerve cord; protostomes include all worms, insects, and mollusks. Deuterostomes develop an anus close to this transient opening in the embryo and have a dorsal central nervous system; they include echinoderms (such as sea stars and sea urchins) and vertebrates. The bodies of both protostomes and deuterostomes are divided into discrete segments that form early in embryonic development. Protostomes and deuterostomes probably evolved from a common ancestor, termed Urbilateria, that lived approximately 600 million years ago (Figure 130a).
FIGURE 1-30 Similar master transcription factors, conserved during evolution, regulate early developmental processes in diverse animals. (a) Urbilateria is the presumed ancestor of all protostomes and deuterostomes that existed about 600 million years ago. The positions of its nerve cord (violet), surface ectoderm (mainly skin; white), and endoderm (mainly digestive tract and internal organs; light green) are shown. (b) Highly conserved master transcription factors called Hox proteins, which determine the identity of body segments during embryonic development, are found in both protostomes and deuterostomes. Hox genes are found in clusters on the chromosomes of most or all animals,
and they encode related master transcription factors that control the activities of other genes. In many animals, different Hox genes direct the development of different segments along the head-to-tail axis, as indicated by corresponding colors. Each Hox gene is activated (transcriptionally) in a specific region along the head-to-tail axis and controls the growth and development of tissues in that segment. For example, in the mouse, a deuterostome, certain Hox genes are responsible for the distinctive shapes of vertebrae. Mutations affecting Hox genes in the fruit fly, a protostome, cause body parts to form in the wrong locations, such as legs in lieu of antennae on the head. In both organisms, these genes provide a head-to-tail address and serve to direct the formation of structures in the appropriate places. Description Protostomes and Deuterostomes evolve from Urbilateria, about 600 million years ago. The nerve cord lies along the lower part of the body in Protostomes and it lies along the upper part of the body in Deuterostomes. The nerve cord lies along the lower part of the body in Urbilateria. The second part shows a horizontal bar with red, white, orange, gray, and yellow-colored segments, representing genes. The parts of the body of the fly and the mammal are color-coded. The fly has head color-coded with red, white, and gray Hox genes, thorax color-coded with white, and orange, and the abdomen color code with white and yellow. In mammals, Hox genes play a vital role in the formation of the distinct shape of the vertebrae and color-coded with red, white, orange, and yellow. Many patterning genes encode master transcription factors that control expression of other genes and specify the general organization of an organism, beginning with the major body axes — anterior-posterior (headto-tail), dorsal-ventral (back-to-belly), and left-right — and ending with body segments such as the head, chest, abdomen, and tail. The conservation of axial symmetry from the simplest worms to mammals is explained by the presence of conserved patterning genes in their genomes. Other patterning genes encode proteins that are important in cell adhesion
or in cell signaling. This broad repertoire of patterning genes permits the integration and coordination of events in different parts of the developing embryo and gives each segment in the body its unique identity. Remarkably, many patterning genes encoding master transcription factors are highly conserved in both protostomes and deuterostomes (Figure 130b). This conservation of body plan reflects evolutionary pressure to preserve the commonalities in the molecular and cellular mechanisms controlling development in different organisms.
1.6 Metazoan Organisms Widely Used in Cell Biology Research
1.6 Metazoan Organisms Widely Used in Cell Biology Research Many invertebrate and vertebrate organisms have been and continue to be immensely valuable for cell biology research. Invertebrate organisms with large-celled embryos that develop outside the mother’s body (e.g., sea urchins and sea stars) are extremely useful for tracing the fates of cells as they form different tissues, as well as for making extracts for biochemical studies. For instance, a key protein in regulating cell division in all eukaryotes, including humans, was first identified in studies with sea star and sea urchin embryos and subsequently purified from extracts prepared from these embryos (see Chapter 19). Other organisms are investigated because of their adaptation to specific ecological niches. An example is the tardigrades, tiny -long, 8-legged invertebrates (see the photo on the first page of the chapter). Tardigrades are widely studied because of their ability to survive exposures to extreme temperatures, high and low pressures, dehydration, and starvation. Indeed, they are the only organisms known to survive exposure to the deadly combination of low pressure and intense radiation of outer space. A few invertebrates — notably the roundworm Caenorhabditis elegans, the fruit fly Drosophila melanogaster, and the planarian Schmidtea mediterranea — are especially amenable to a variety of experimental approaches. These invertebrates have been extensively studied by
Drosophila melanogaster and Caenorhabditis elegans Are Used to Identify Genes That Regulate Animal Development
thousands of research groups using multiple experimental technologies. Studies on these and other invertebrates led to important insights into animal development and cell function, and also to understanding of many human diseases. Drosophila melanogaster and Caenorhabditis elegans Are Used to Identify Genes That Regulate Animal Development D. melanogaster has been used as an experimental organism since 1910, when Thomas Hunt Morgan established the Fly Room laboratory at Columbia University. It is still extensively used in research because it can be readily reared in the laboratory, has only four pairs of chromosomes, breeds quickly, and lays many eggs (see Figure 1-23d). Each step in the development of this organism — from fertilized egg through the embryonic larval and pupal stages to the mature adult — is well understood. Most importantly, mutations in many genes important for development have been characterized in detail and these have provided important information about the functions of the encoded proteins. As an example, many genes important for development of the Drosophila eye have been identified; one of them, eyeless, is the master transcription factor that initiates eye development. Fly eyes and human eyes are very different in their structure, function, and nerve connections. Nonetheless, eyeless in the fly and Pax6 in the human are highly related proteins that
regulate the activities of other genes and are descended from the same ancestral gene. Mutations in both the eyeless and the Pax6 genes cause major defects in eye formation (Figure 1-31).
FIGURE 1-31 Homologous genes regulate eye development in Drosophila and humans. (a) Development of the large compound eyes in fruit flies requires a gene called eyeless (named for the mutant phenotype). (b) Flies with inactivated eyeless genes lack eyes. (c) Normal human eyes require the gene Pax6, the homolog of eyeless. (d) People lacking adequate Pax6 function have the genetic disease aniridia, a lack of irises in the eyes. Eyeless and Pax6, which encode highly related master transcription factors that regulate the
Planaria Are Used to Study Stem Cells and Tissue Regeneration
activities of other genes, are homologs and presumably descended from the same ancestral gene. The roundworm Caenorhabditis elegans was developed as an experimental organism by Sydney Brenner only in the 1960s. The adult worm (see
Figure 1-23c) — about long and in diameter — can be either a hermaphrodite (a worm with both male and female organs that can fertilize itself and produce viable offspring) or a male (which can fertilize a hermaphrodite); this unusual reproductive behavior has many advantages for the rapid isolation and characterization of mutations affecting development or cell function. The hermaphrodite has precisely 959 somatic (non-germ) cells, whereas the male has 1031 (see Figure 22-26d). These small numbers of cells are arranged in a nearly identical way in every worm, and scientists have traced the lineage of each somatic cell in C. elegans from the fertilized egg to the mature worm using microscopy to follow the development of live worms (see Figure 22-26c). As we learn in
Chapter 22, many of the cells born during development of C. elegans are destined to die by a process termed programmed cell death, or apoptosis. A similar process utilizing similar proteins occurs in many human cells, and the absence of programmed cell death contributes to the development of many human cancers. Planaria Are Used to Study Stem Cells and Tissue Regeneration
In single-celled organisms, both daughter cells usually (though not always) resemble the parent cell. Similarly, in multicellular organisms, when many types of cells divide, the daughter cells look a lot like the parent cell — liver cells, for instance, divide to generate liver cells with the same characteristics and functions as their parent, as do insulinproducing cells in the pancreas. In contrast, stem cells and certain other undifferentiated cells can generate multiple types of differentiated descendant cells; these cells often divide in such a way that the two daughter cells are different. Such asymmetric cell division is characteristic of stem cells and is critical to the generation of different cell types in the body (see Chapter 22). Often one daughter cell resembles its parent in that it remains undifferentiated and retains its ability to give rise to multiple types of differentiated cells. The other daughter cell divides many times, and each of its daughter cells differentiates into one or several specific types of cells. The planarian Schmidtea mediterranea is best known for its capacity to regenerate complete individuals — with a normal head — from minuscule body parts formed by dissection (see Figure 1-23e). Planaria contain stem cells, termed neoblasts, that replace cells lost to normal turnover. In portions of a dissected animal, neoblasts will, after several cell divisions, generate any cell type needed during regeneration. These stem cells have served as a potent experimental system to discover how heads and tails, each built of many types of cells, are formed (see Chapters 16 and 22). The hormones that instruct stem cells in different parts of the body to generate specific types of cells are similar to those used in mammals, including humans, in development (see Chapter 16), and thus future
Studies on Fish, Mice, and Other Vertebrate Organisms Inform the Study of Human Development and Disease
studies on planarian regeneration may inform scientists how to regenerate human body parts such as a hand or an eye. Studies on Fish, Mice, and Other Vertebrate Organisms Inform the Study of Human Development and Disease The zebrafish Danio rerio (see Figure 1-23f) is widely used to study the development and functions of organs not found in invertebrates, especially the brain and the central nervous system. Its generation time — from fertilized egg to daughter fertilized eggs — is only 3 months, short for a vertebrate. The fertilized egg rapidly becomes transparent, allowing visualization of developing organs, and the embryo develops rapidly, such that precursors to all major organs appear within 36 hours of fertilization. Mice (see Figure 1-23g) have one enormous advantage over other experimental organisms: they are the most closely related to humans of any animal for which powerful genetic approaches have been available for many years. Mice and humans have shared living structures for millennia, have similar nervous systems, have similar immune systems, and are subject to infection by many of the same pathogens. As noted, both organisms contain about the same number of genes, and about 99 percent of mouse protein-coding genes have homologs in the human genome, and vice versa.
Human Genetic Diseases Elucidate Important Aspects of Cell Function
Using the Cas9 and other experimental systems described in Chapter 6, researchers can inactivate any desired gene and thus abolish production of its encoded protein. This approach is being used extensively to produce animal versions of human genetic diseases, in mice as well as in other animals. As an example, people with autism spectrum disorder often have mutations in specific protein-coding genes that regulate development of specific types of nerve cells. To understand the role of these mutations, these genes have been inactivated in mice and more recently in macaque monkeys. These animals often exhibit symptoms of the human disease, including repetitive actions such as excessive grooming, strongly suggesting that the human mutation indeed has a role in triggering the disorder. Such approaches can be useful in uncovering the role of specific genes in higher order brain tasks such as learning and memory, or in studies of viruses that infect only humans and nonhuman primates. Once animal models of a human disease are available, investigators can perform further studies on the molecular defects causing the disease and can test new treatments, thereby minimizing the testing of new drugs on humans. Human Genetic Diseases Elucidate Important Aspects of Cell Function We have emphasized how analysis of mutant organisms missing a specific protein has informed the normal function of that gene and encoded protein. Although we cannot induce mutations in humans as we might in other organisms, we can study naturally occurring mutations in humans. Many genetic diseases are caused by mutations in a single protein; thus
Unbiased Single Cell Sequencing Experiments Identify Altogether New Cell Types
studies on humans with these diseases also can shed light on the normal function of those proteins. As an example, consider Duchenne muscular dystrophy (DMD), the most common of the hereditary muscle-wasting diseases, collectively called muscular dystrophies. DMD is a disorder caused by mutations in the dystrophin gene localized on the X chromosome. It affects 1 in 3300 boys and results in cardiac or respiratory failure and death, usually in the late teens or early twenties. The first clue to understanding the molecular basis of this disease came from the cloning of the dystrophin gene. As detailed in Chapters 17 and 20, the very large protein encoded by this gene binds to other proteins in muscle cells to form a large multiprotein complex. One end of the long dystrophin molecule binds to actin microfilaments that are a component of the contractile apparatus (see Figure 1-14); the other binds to a complex of proteins in the muscle plasma membrane (see Figure 20-41). The resulting large multiprotein assemblage, the dystrophin glycoprotein complex (DGC), links the extracellular matrix protein laminin to the cytoskeleton within muscle cells and prevents the muscle cell from ripping apart during contractions. Mutations in dystrophin or other DGC components can disrupt the DGC-mediated link between the exterior and interior of muscle cells and cause muscle weakness and eventual muscle deterioration and death. The first step in identifying the entire dystrophin glycoprotein complex was to clone the dystrophin-encoding gene using DNA from normal individuals and from patients with Duchenne.
Unbiased Single Cell Sequencing Experiments Identify Altogether New Cell Types How many different types of cells make up a mouse or a human body? Certain types of cells may carry out very important functions, but because they are present in miniscule amounts in a specific organ or tissue their very existence may have gone unnoticed! The new technique of single cell RNA sequencing (Chapter 6) allows researchers to identify cell types present in only small amounts. The technique identifies the types of messenger RNAs that are expressed in the largest numbers in a single isolated cell. If researchers apply the technique to many individual cells isolated from a tissue, they can identify new types of cells whose components of messenger RNAs, and thus of the proteins they encode, are different from those of known types of cells. In a recent study using single-cell RNA sequencing, researchers analyzed tens of thousands of cells from mouse and human airways, mapping the physical locations of cell types and creating a cellular atlas of the tissue. Using a computer program to analyze the overall similarity between the types of mRNAs expressed in each cell, the researchers easily found six groups of cells that each expressed a similar population of mRNAs. These six groups of cells corresponded to the six well-known cell types present in the trachea — the abundant basal, club, and ciliated cells and the rarer
tuft, neuroendocrine, and goblet cells. One extremely rare cell type, making up roughly 1 percent of the cell population, had a profile of mRNAs radically different from the 6 other known cells. The team named this new cell the pulmonary ionocyte. Strikingly, pulmonary ionocytes were found to express the CFTR gene at levels higher than any other cell type. The CFTR protein is critical for airway function; mutations in the CFTR gene cause cystic fibrosis in humans. For decades, researchers and clinicians assumed that the CFTR gene is frequently expressed at low levels in ciliated cells, a cell type spread throughout the entire airway. According to the new data, the majority of CFTR is expressed in only a few cells (Figure 1-32), which researchers didn’t even know existed until now.
The Following Chapters Present Many Experimental Techniques and Much Experimental Data That Explains How We Know What We Know About Cell Structure and Function
FIGURE 1-32 A new cell type in the mouse airway identified by unbiased single-cell sequencing of messenger RNAs. Ionocytes, a minor cell type in the upper respiratory tract of the mouse that express the CFTR protein, are stained green with an antibody that detects a protein expressed only in these cells. All cell nuclei are stained with a blue dye. By counting the number of blue nuclei in the tissue section, it can be determined that ionocytes comprise about 1 percent of the cells lining the mouse trachea (airway). These cells extend through the layer of neighboring epithelial cells to the airway surface. See text for an explanation of how these cells were identified. [Republished with permission of Nature Publishing Group, from D. Montoro et. al., 2018. A revised airway epithelial hierarchy includes CFTR-expressing ionocytes. Nature 560:319; permission conveyed through Copyright Clearance Center, Inc.] The Following Chapters Present Many Experimental Techniques and Much Experimental Data That Explains How We Know What We Know About Cell Structure and Function In subsequent chapters of this book, we discuss cellular processes in much greater detail. We begin (in Chapter 2) with a discussion of the chemical nature of the building blocks of cells and the basic chemical processes required to understand the macromolecular processes discussed in subsequent chapters. We go on to discuss the structure and function of proteins (in Chapter 3). Chapter 4 discusses many of the techniques biologists use to culture and fractionate cells and to visualize specific
proteins and structures within cells. Chapter 5 describes how DNA is replicated, how segments of DNA are copied into RNA, and how proteins are synthesized on ribosomes. Chapter 6 describes many of the techniques used to study genes, gene expression, and protein function, including the generation of animals with specific genetic mutations. Gene and chromosome structure and the regulation of gene expression are covered in Chapters 7, 8, and 9. Biomembrane structure is the topic of Chapter 10. The transport of ions and small molecules across membranes is covered in
Chapter 11, and Chapter 12 discusses cellular energetics and the functions of mitochondria and chloroplasts. Membrane biogenesis, protein secretion, and protein trafficking — the directing of proteins to their correct subcellular destinations — are the topics of Chapters 13 and 14. Chapters 15 and 16 discuss the many types of signals and signal receptors used by cells to communicate and regulate their activities. The cytoskeleton and cell movements are discussed in Chapters 17 and 18. Chapter 19 discusses the cell cycle and how cell division is regulated. The interactions among cells, and between cells and the extracellular matrix, that enable formation of tissues and organs are detailed in Chapter 20. A chapter new to this edition, Chapter 21, details how cells respond to changes in their environment — specifically, to changes in oxygen levels, temperature, day and night, and nutrient levels. Later chapters of the book discuss important types of specialized cells — stem cells (Chapter 22), neurons (Chapter 23), and cells of the immune system (Chapter 24). Chapter 25 discusses cancer and the multiple ways in which cell growth and differentiation can be altered by mutations.
OCR Supplement (Image-Only Pages)
_The following pages contained text rendered as part of a flattened image; their content was recovered by tesseract OCR (300 dpi)._