2 Adaptations to Aquatic Environments
2Adaptations to Aquatic Environments Sperm whale. Modern whales, such as these sperm whales (Physeter macrocephalus) swimming off the coast of Portugal, have a number of adaptations that enable them to live in an aquatic environment. The Evolution of Whales Life on Earth began in the water. Of the many species that live in water, some of the most fascinating are the whales—a group that is particularly well suited to aquatic life. Surprisingly, the ancestor of modern whales can be traced back to a terrestrial mammal related to cattle, pigs, and hippos. Scientists first proposed an evolutionary relationship between whales and this group of land mammals in 1883, based on observations of similarities in their skeletons. In the 1990s, DNA technology confirmed that the groups are also related genetically. In 2007, scientists discovered a critical link between

hippos and whales: the fossilized bones of a previously unknown, large terrestrial mammal, in the genus Indohyus, that may have spent at least some of its time in the water. Scientists speculate that over the subsequent 50 to 60 million years, selection imposed by the challenges of living in an aquatic environment led to the evolution of whales as we know them today. Modern whales have evolved a wide range of adaptations for aquatic life. One of the most obvious challenges is the ability to swim efficiently. For example, the killer whale is capable of swimming up to 48 km per hour. Such speeds are possible only with a highly streamlined body. Over evolutionary time, natural selection would have favored any individuals that had a more streamlined body, including bodies with reduced hindlimbs. In modern whales, tiny remnants of the hindlimb bones can be found entirely within the whale’s body. During the course of the whale’s evolutionary history, whale ears became internalized. Whether this was due to selection for a streamlined body or for some other reason is not known, but the outcome made for a more streamlined body. Obtaining oxygen is another challenge for whales because they need to dive for long periods in search of food. Over time, there was an evolutionary change in the location of the nostrils from the front of the head to the top. While scientists cannot be certain of the selective forces that caused this change, one hypothesis is that changes in nostril position may have been favored over time to the point that the modern whale can more easily grab a breath of air when surfacing. In addition, whales can hold their breath for long periods. Sperm whales, for example, dive to depths of 500 m and may stay below the surface for more than an hour as they search for fish, squid, and other food. During a dive, the sperm whale relies on oxygen stored in its body. It might surprise you to learn that very little of this oxygen resides in the lungs; most is bound to hemoglobin in the blood or to a similar oxygen storage molecule, myoglobin, in the muscles. Under water, whales slow their metabolism by reducing blood flow to nonvital organs such as the skin, intestines, lungs, and kidneys, while blood flow to the brain and heart continues. Consequently, during a dive the temperature of all but a few key organs drops, the heart rate slows,
and demand for oxygen is reduced. “Surprisingly, the ancestor of modern whales can be traced back to a terrestrial mammal related to cattle, pigs, and hippos.” Whale ancestor. The ancestor of modern whales, in the genus Indohyus, was a terrestrial animal that spent part of its time in the water. Over time, the descendants of this animal developed numerous adaptations for living in water that are found in modern-day whales. Regulating body temperature is yet another challenge. Because heat loss occurs much faster in water than in air, a thick layer of fat under the skin insulates most oceanic mammals living in cold waters. Like a warm coat, this insulation slows the loss of heat generated by its internal organs. Whales also maintain a higher metabolic rate than land mammals of similar size, and this helps generate extra heat. The whale’s vascular structure also helps maintain heat; the veins and arteries of a whale’s flippers and tail are next to each other. This allows the warm arterial blood that travels away from the heart to transfer heat to cooler blood in adjacent veins as it returns from a whale’s extremities to the heart. A final challenge is how to find food in the water, and two different strategies evolved long ago: Baleen whales have long plates in their mouths to filter tiny prey out of the water, whereas toothed whales have teeth in their mouth to grab prey. Modern toothed whales also
use echolocation, which involves emitting a sound into the water and then listening for the sound to bounce back as an “echo” when it intercepts objects such as food. Since all toothed whales use echolocation, but baleen whales do not, scientists suspected that echolocation must have evolved long ago in an ancestor of toothed whales. In 2016, researchers reported that they examined a whale skull that was found in South Carolina and estimated to be 27 million years old. Their striking discovery was that the skeleton contained a specially shaped inner ear, which indicated the whale was capable of echolocation, confirming that this adaptation occurred very early in the evolution of toothed whales. The evolution of whales took place over a period of 50 million years. While the selective forces occurring over this time cannot be known with certainty, the fossils found to date suggest that an ancient terrestrial mammal ultimately evolved a number of adaptations that gave rise to modern-day whales. In this chapter, we examine how the challenges posed by living in an aquatic environment have caused a wide variety of organisms to evolve adaptations. SOURCES: New fossil evidence suggests echolocation evolved early in whales, Science News, August 5, 2016, https://www.sciencenews.org/article/new-fossil-suggests-echolocation-evolved-early- whales. Valley of the whales, National Geographic Magazine, August 2010, http://ngm.nationalgeographic.com/2010/08/whale-evolution/mueller-text. Whales descended from tiny deer-like ancestors, Science Daily, December 21, 2007, http://www.sciencedaily.com/releases/2007/12/071220220241.htm.
Learning Objectives
After reading this chapter, you should be able to:

2.1 Describe some of the many properties of water that make it favorable to life.
2.2 Explain the challenges faced by aquatic animals and plants of
maintaining an internal balance of water and salt concentrations.
2.3 Describe how the uptake of gases from water is limited by diffusion.
2.4 Explain how temperature limits the occurrence of aquatic life. Scientists generally agree that life began in the ocean and that the first forms of life were simple bacteria. Over millions of years, these bacteria gave rise to an incredible diversity of organisms, many of which still live in the water. Other species, as we saw in the case of the whale, evolved terrestrial forms that later evolved to return to life in the ocean. In this chapter, we begin an exploration of ecology at the individual level by examining the ways in which the properties of water support and constrain aquatic organisms and direct the evolution of adaptations.
#### 2.1 Water Has Many Properties Favorable to Life
2.1 Water has many properties favorable to life Water is abundant over most of Earth’s surface. Because water has an immense capacity to dissolve inorganic compounds, it is an excellent medium for the chemical processes of living systems. In fact, it is hard to imagine a form of life that could exist without water. In this section, we will look at how water makes life possible, including water’s thermal properties, its density and viscosity, and its function as a solvent for inorganic nutrients.
Thermal Properties of Water
On Earth, water can be found as a solid (ice), as a liquid, and as a gas (water vapor). Pure water—water not containing any dissolved minerals or other compounds—becomes a solid below 0 °C and becomes a gas above 100 °C at sea level. At higher elevations, the freezing point of water changes very little, but the boiling point can be several degrees lower. Within the temperature range organisms usually encounter, it is present in liquid form. When water contains dissolved compounds, such as salts, its freezing temperature drops below 0 °C. This is why road salts are applied to ice- and snow-covered roads; the salts allow the ice and snow to melt at a lower temperature than they otherwise would. Dissolved compounds also raise the boiling point of water above 100 °C. The temperature of water remains relatively steady even when heat is removed or added rapidly, as can happen at the air–water interface or at an organism’s surface, such as on the surface of a whale in the ocean. This is because water has a high specific heat, which is the amount of heat required to increase its temperature by 1 °C. In addition, water transfers heat rapidly, causing heat to spread evenly throughout a body of water, which also slows localized changes in temperature. Water also resists changing from one state to another. For example, raising the temperature of 1 kg of liquid water by 1 °C requires the addition of 1 Calorie of heat. However, converting 1 kg of liquid water into water vapor requires the addition of 540 Calories of heat. Similarly, lowering the temperature of 1 kg of liquid water by 1 °C requires the removal of 1 Calorie of heat, but converting that amount of liquid water to ice requires the removal of 80 Calories of heat. In short, liquid water is very resistant to changing

states. This resistance helps prevent large bodies of water from freezing solid during winter. Another curious, yet fortunate, thermal property of water is the way it changes density with changes in temperature. You may recall from previous courses in biology or chemistry that most compounds become denser under cooler temperatures. However, water achieves its highest density—that is, its molecules are the most packed together—at 4 °C. Above and below 4 °C, the water molecules are less tightly packed, so the water becomes less dense. Below 0 °C, pure water is converted into ice, which is less dense than liquid water, as you can see in Figure 2.1. As a result of its lower density, ice floats on the surface of liquid water. This means that lakes experiencing cold winters will generally have a layer of 4 °C water at the bottom. Above this layer will be water that is cooler than 4 °C and on top of that a layer of ice.
Figure 2.1 The density of water. As water cools, the molecules contract and become more dense. Below 4 °C, they begin to expand and become less dense. Below 0 °C, pure water is converted into ice that is even less dense. As a result of its lower density, ice floats on the surface of liquid water. Water’s unusual thermal properties are especially important to aquatic plants and animals. In large bodies of fresh water, such as lakes, the bottom
of the lake does not freeze, in part because of the insulation that ice provides from very cold air temperatures. The salt in ocean water lowers the freezing point of the water to well below 0 °C, which helps prevent most oceans from freezing. In both cases, the available liquid water provides a refuge for organisms during periods of cold temperatures.
Density and Viscosity of Water
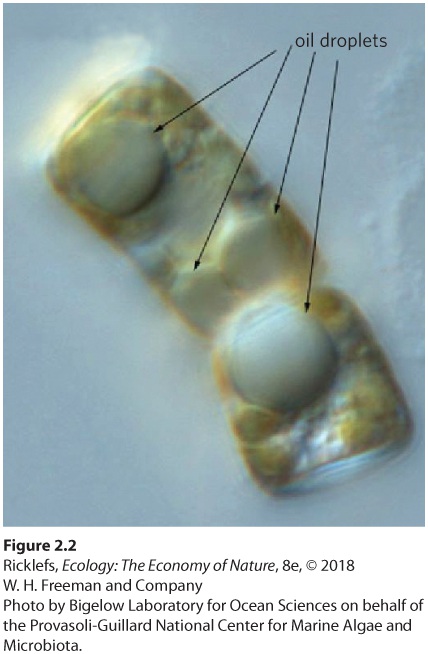
The adaptations of aquatic organisms often exploit the density and viscosity of water. For example, animals and plants are made up of bone, proteins, and other materials that are somewhat denser than salt water, and much denser than fresh water. However, organisms can also contain fats and oils that are less dense than water. In some cases, they also possess pockets of air, such as the lungs of the whales that were described at the beginning of this chapter. The combination of the materials that compose an animal’s body and the presence of air pockets determine whether an organism will float or sink in water. For those organisms that are denser than their surrounding water, a variety of adaptations can either reduce an organism’s density or retard its rate of sinking. For example, many fish species have a gas-filled swim bladder that can adjust in size to make the density of the fish’s body equal to that of the surrounding water. Human divers use this same concept when they wear airfilled inflatable vests to help match the density of the water. When the vest is full of air, divers float on the surface of the water. As they release some of the air, they begin to sink. By adjusting the amount of air in the vest, divers can make their density equal to the water at a particular depth, making it easier to swim because they are not spending energy resisting their body’s attempt to float or sink. Some large kelps, like those we saw in Figure 1.8, have gasfilled bulbs that cause their leaflike blades to float up into the sunlit surface waters. The whales discussed earlier become buoyant when they take a breath of air, but a slow release of air bubbles will help them sink to a particular depth. At the other end of the size spectrum, many of the microscopic unicellular algae that float in great numbers in the surface waters of lakes and oceans use droplets of oil as flotation devices (Figure 2.2). Because oils are less dense than water, the algae can use the oil droplets to help offset their natural tendency to sink.
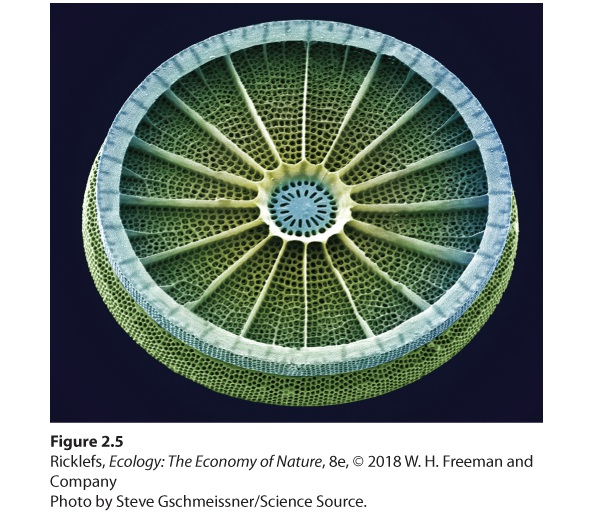
Figure 2.2 Adaptation to water density. These algae (Cyclotella cryptica) are able to float near the water’s surface by using oil droplets that have a lower density than water. Aquatic organisms also possess adaptations to deal with the high viscosity of water. We can think of viscosity as the thickness of a fluid that causes objects to encounter resistance as they move through it. In response to living in water, fast-moving aquatic animals such as fish, penguins, and whales have evolved highly streamlined shapes that reduce the drag caused by the high viscosity of water (Figure 2.3). The viscosity of water is higher in cold water than in warm water, which can make swimming in cold water more difficult. Movement in water is even more difficult for smaller animals. However, the same high viscosity that impedes the progress of tiny organisms as they swim in the water also impedes them from sinking. Because these organisms are slightly denser than water, they are prone to sinking due to the force of gravity. To take advantage of water’s viscosity, many tiny marine animals have evolved long, filamentous appendages that
cause greater drag in the water. The appendages function like a parachute slowing the fall of a body through air (Figure 2.4).
Figure 2.3 Streamlined shape. Large, fast-moving aquatic organisms like the barracuda (Sphyraena barracuda) have evolved highly streamlined shapes to help them move through the highly viscous water.
Figure 2.4 Adaptation to water viscosity. Some small aquatic organisms exploit the high viscosity of water by evolving large appendages, such as the antenna and feathery projections of this tiny marine crustacean. These appendages help slow down movement through the viscous water and thereby retard sinking. Viscosity The thickness of a fluid that causes objects to encounter resistance as they move through it.
Dissolved Inorganic Nutrients
Both aquatic and terrestrial organisms require a variety of nutrients to build necessary biological structures and maintain life processes. Large amounts of hydrogen, carbon, and oxygen are necessary for building most compounds found in organisms. Nitrogen, phosphorus, and sulfur are used in building proteins, nucleic acids, phospholipids, and bones. Other major nutrients— including potassium, calcium, magnesium, and iron—play important roles as solutes and as structural components of bones, woody plant cells, enzymes, and chlorophyll. Certain organisms need additional minor nutrients. For example, diatoms are a group of algae that need silica to construct their glassy shells (Figure 2.5). Similarly, some species of bacteria require the element molybdenum, which makes up part of the enzyme they use to convert nitrogen from the atmosphere (N2) into ammonia (NH3).
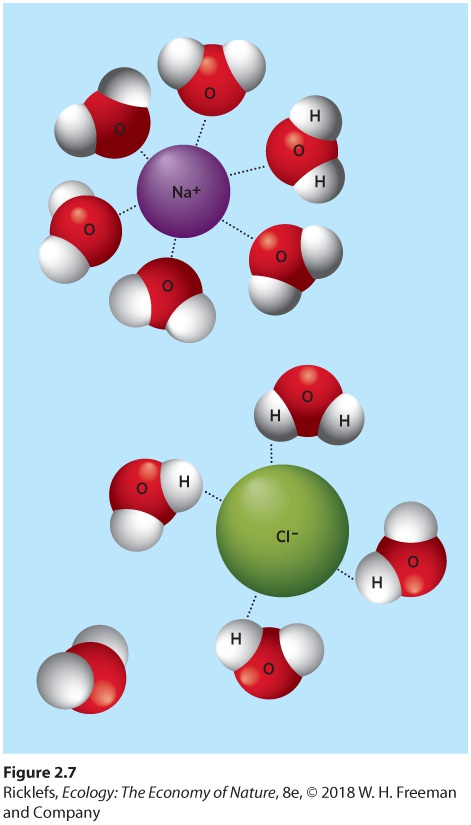
Figure 2.5 Use of inorganic nutrients. Diatoms, such as this species of Arachnoidiscus, are a type of algae that require minor nutrients such as silica to build a hard, glassy shell. Image is magnified 175 times. The Solubility of Minerals Water is a powerful solvent with an impressive capacity to dissolve substances, which makes them accessible to living systems. Because of this property, water also provides a medium in which substances can react chemically to form new compounds. Water acts as a solvent because of its molecular structure. As you can see in Figure 2.6, water molecules consist of an oxygen atom in the middle and two hydrogen atoms connected in a V-shaped arrangement. This arrangement results in an unequal sharing of electrons: The oxygen end of the water molecule has a slightly negative charge and the hydrogen end has a slightly positive charge. When the two ends of the molecule possess opposite charges, we say that the molecule is polar. Water is a polar molecule: The negative oxygen end of one water molecule is strongly attracted to the positive hydrogen end of another. These forces of attraction are known as hydrogen bonds.
Figure 2.6 Water molecules. Because of the configuration of water molecules, they are negatively charged on the oxygen end and positively charged on the hydrogen end. The attractive forces of these opposite charges, known as hydrogen bonds, allow water molecules to be attracted to each other and to the charged ions of other compounds such as salts and sugars. The polar nature of water molecules also allows them to be attracted to other polar compounds. Some solid compounds consist of electrically charged atoms, or groups of atoms, called ions. For example, common table salt—sodium chloride (NaCl)—contains positively charged sodium ions (Na+) and negatively charged chloride ions (Cl−). In their solid form, these ions are arranged in a crystal lattice. In water, however, the charged sodium and chloride ions are attracted by the charges of the water molecules. As shown in Figure 2.7, the attraction of these ions to water molecules is stronger than the attraction that holds the crystal together. As a result, when sodium chloride is added to water, its crystal lattice breaks apart, and water molecules surround the salt ions. In other words, when you put salt in water, it dissolves. This solvent ability of water is not restricted to ionic compounds
such as salts; it occurs with any polar compound, including the various types of sugars that organisms commonly use. In contrast, water is not a good solvent for oils and fats because they are nonpolar compounds.
Figure 2.7 Dissolving ions in water. Because water molecules have negative and positive ends, they attract the negatively and positively charged ions, such as the sodium and chlorine ions found in sodium chloride. The forces of attraction to water molecules are stronger than the forces of attraction within the crystal, so the ions separate and become surrounded by water molecules. Ions Atoms or groups of atoms that carry an electric charge. The solvent properties of water explain the presence of minerals in
streams, rivers, lakes, and oceans. When water vapor in the atmosphere condenses to form clouds, the condensed water in the atmosphere is nearly pure. However, as it falls back to Earth as rain or snow, water acquires some minerals from dust particles in the atmosphere. Precipitation that hits land comes into contact with rocks and soils, and it dissolves some of their minerals. These dissolved minerals are carried toward the ocean with the rainwater. Water in most lakes and rivers contains a dissolved mineral concentration of 0.01 to 0.02 percent, whereas ocean water contains a dissolved mineral concentration of 3.4 percent. Oceans have much higher concentrations of dissolved minerals because mineral-laden water continuously flows in from streams and rivers. The constant evaporation from the ocean’s surface removes pure water and leaves the minerals behind. Over billions of years, this process has caused an increase in the concentrations of minerals in the oceans. Every mineral has an upper limit of solubility in water, known as saturation. This limit generally increases with higher temperatures. After a mineral achieves saturation, water cannot hold any more of the mineral and it begins to precipitate out of solution. For some minerals, such as sodium, ocean concentrations are far below saturation. Most of the sodium that is washed into ocean basins remains dissolved and its concentration in seawater continues to increase over time. In contrast, the concentrations of other minerals in the oceans commonly exceed their saturation concentrations. For example, calcium ions (Ca2+) in the water readily combine with dissolved CO2 to form calcium carbonate (CaCO3), which has a low solubility in ocean water. Over millions of years, the excess calcium carbonate that has washed into the oceans from streams and rivers has subsequently precipitated out of the water. This precipitated calcium carbonate, combined with the calcium carbonate from the bodies of countless tiny marine organisms, has resulted in massive limestone sediments (Figure 2.8). Today, these sediments are important sources of limestone for construction applications such as stone blocks and concrete, for agricultural uses such as fertilizer, and for numerous industrial processes.
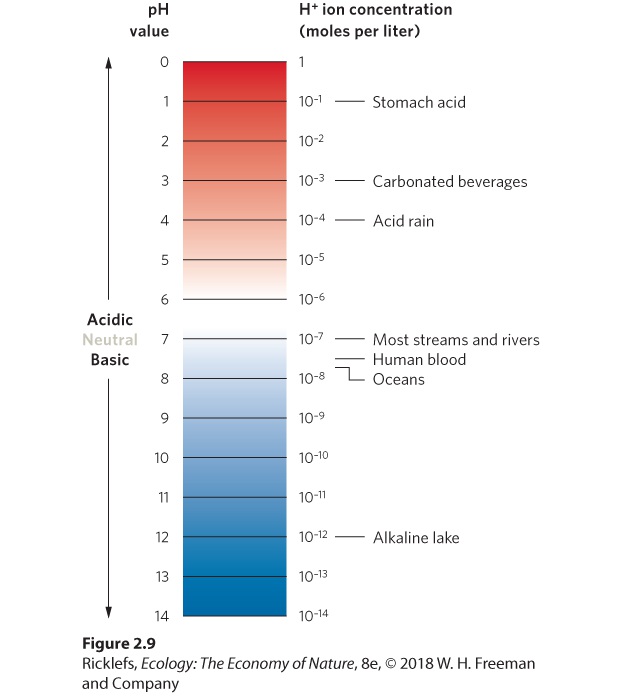
Figure 2.8 The formation of limestone sediments. The continuous addition of calcium minerals into oceans from streams and rivers causes calcium to combine with CO2 to become calcium carbonate. Because calcium carbonate is not very soluble in water, it precipitates out of the water to form massive sediments over millions of years. This site of limestone sediments located in Victoria, Australia, was once under water but is now above the water due to changes in ocean depth. Saturation The upper limit of solubility in water. Hydrogen Ions Among dissolved substances in water, hydrogen ions (H+) deserve special mention because they are extremely reactive with other compounds. In pure water, a small fraction of the water molecules (H2O) break apart into their hydrogen (H+) and hydroxide (OH−) ions. The concentration of hydrogen ions in a solution is referred to as its acidity. Acidity is commonly measured as pH, which is defined as the negative logarithm of hydrogen ion concentration (as measured in moles/L, where 1 mole = 6.02 × 1023 molecules): pH=-log[H+]
Acidity The concentration of hydrogen ions in a solution. pH A measure of acidity or alkalinity; defined as pH = −log [H+]. As you can see in the pH scale shown in Figure 2.9, water containing a high concentration of hydrogen ions has a low pH value, whereas water containing a low concentration of hydrogen ions has a high pH value. Therefore, we categorize water with low pH values as acidic, water with a mid-range value of 7 as neutral, and water with a high pH value as basic or alkaline. As we will see, natural rain or snow can vary a great deal in pH, depending on the presence of different chemical compounds in the atmosphere that affect the pH value.
Figure 2.9 The relationship between pH and hydrogen ion concentration in water. The pH scale of hydrogen ion concentration extends from 0 (highly acidic) to 14 (highly alkaline). The pH of rainfall can vary a great deal around the world. Hydrogen ions, because of their high reactivity, dissolve minerals from rocks and soils, enhancing the natural solvent properties of water. For example, in the presence of hydrogen ions, the calcium carbonate found in limestone dissolves readily, according to the following chemical equation: H++CaCO3→Ca2++HCO3Calcium ions are important to life processes, and their presence is vital to organisms such as snails, mussels, and clams that form shells made of calcium carbonate. As a result, these animals are less abundant in streams and lakes that are low in calcium. Therefore, hydrogen ions are essential for making certain nutrients available for life processes. At high concentrations, however, hydrogen ions negatively affect the activities of most enzymes. In addition, high concentrations of hydrogen ions cause many heavy metals to begin dissolving in water. These heavy metals, including arsenic, cadmium, and mercury, are highly toxic to most aquatic organisms. The normal range of pH of lakes, streams, and wetlands is between 5 and 9. However, some bodies of water can have even lower pH values. Sometimes lower pH conditions have a natural cause. Bogs, for example, are aquatic habitats with vegetation such as sphagnum mosses that release H+ ions into the water and thereby make the water more acidic and unsuitable for many other species of plants. Other bodies of water have low pH as a result of human influences. For example, the release of sulfur dioxide (SO2) and nitrogen dioxide (NO2) from coal-powered industrial plants became a major environmental issue in the 1960s. At that time, ecologists in Russia, China, northern Europe, the United States, and Canada began to notice that many bodies of water were becoming more acidic and less hospitable to numerous species of fish and other aquatic organisms. They also noticed that trees were dying, particularly in the spruce-fir forests that existed at high elevations in these regions of the world. It turned out that the areas with more acidic bodies of water and dying trees were all far downwind of industrial areas with coal-powered factories

that had tall smokestacks. Years of data collection revealed that the sulfur dioxide and nitrogen dioxide emitted into the atmosphere by these smokestacks were converted to sulfuric acid and nitric acid in the atmosphere. These acids were transported downwind and then came back down to Earth as acid deposition, also known as acid rain. The acid deposition occurred in two forms: as gases and particles that stuck to plants and soil, a form called dry acid deposition, and as rain and snow, a form called wet acid deposition. Acid deposition lowered the pH of precipitation and, as a result, water of unusually low pH was entering streams, lakes, and forests. Most aquatic species cannot tolerate water with a pH lower than 4, so these bodies of water became toxic to many aquatic organisms, which included insects and fish. Acid deposition Acids deposited as rain and snow or as gases and particles that attach to the surfaces of plants, soil, and water. Also known as Acid rain. In forests, acid deposition has several effects. First, it leaches the calcium out of the needles of conifer trees such as spruce. It also causes increased leaching of soil nutrients that trees require, including calcium, magnesium, and potassium. Finally, acid deposition causes aluminum to dissolve in the water. Although aluminum naturally occurs in the soil, it is typically not in a form that is available to plants. Dissolved aluminum can negatively affect a plant’s ability to take up nutrients. Collectively, these effects of acid deposition make trees more susceptible to the harmful effects of natural stressors that include drought, diseases, and extreme temperatures. In short, while the trees do not die directly from acid deposition, they become more susceptible to other causes of death. Because acid deposition interacts with so many other natural stressors, scientists recognize that acid deposition has contributed to the death of trees in North America, Europe, and Asia. However, the complexity of the interactive effects has made it difficult to accurately estimate how much of the observed tree death is directly attributable to acid deposition. Once scientists understood the causes and consequences of acid deposition, they began to explore solutions. In the United States, legislation required the installation of smokestack scrubbers that force smokestack gases through a
slurry of limestone and water, which remove the gases. The use of these scrubbers has resulted in a major reduction in the amount of acidic compounds going into the atmosphere. The U.S. Environmental Protection Agency (EPA) reports that from 1980 to 2015, emissions of sulfur dioxide declined by 84 percent. At the end of this chapter, in “Connecting the Concepts: The Decline of Coral Reefs,” we discuss another example of how understanding environmental problems related to pH can help us develop effective solutions.
Concept Check
1. What is unique about water with regard to how temperature affects its density? 2. How can the viscosity of water both hinder and facilitate movement in aquatic animals? 3. Describe the changes in mineral content of water as it moves from rainwater to lake water and, eventually, to ocean water.
#### 2.2 Aquatic Animals and Plants Face the Challenge of Water and Salt Balance
2.2 Aquatic animals and plants face the challenge of water and salt balance
The Challenge of Salt and Water Balance
It might surprise you to learn that organisms in an aquatic environment need specialized mechanisms to maintain the proper amount of water in their bodies. To understand why, we need to recognize that the water surrounding an aquatic organism and the water inside its body contain dissolved substances, known as solutes. These solutes affect the movement of water in and out of an organism. The movement of water occurs at the cellular level, where water passes across cell membranes from regions of low solute concentration to regions of high solute concentration. At the same time, solutes attempt to move across membranes to equalize the concentration of solutes. Cell membranes typically do not allow the free movement of large solute molecules such as carbohydrates and most proteins. Membranes that allow only particular molecules to pass through—for example, water and small solute ions and molecules—are known as semipermeable membranes. Solute A dissolved substance. Semipermeable membrane A membrane that allows only particular molecules to pass through. Solutes can move across semipermeable membranes using either passive or active transport. Passive transport occurs when ions and small molecules move through a membrane along a concentration gradient, from a location with many solutes to a location with few solutes. In contrast, active transport occurs when cells transport ions and small molecules through a membrane against a concentration gradient to maintain their concentrations. Active transport expends energy because it requires the cell to work against the concentration gradient of solutes. Passive transport The movement of ions and small molecules through a membrane along a concentration gradient, from a location with many solutes to a location with few solutes. Active transport
##### Adaptations for Osmoregulation in Freshwater Animals
The movement of molecules or ions through a membrane against a concentration gradient. When the water inside a cell has a higher concentration of solutes than the water outside, water tries to move into the cell from the surrounding environment. In contrast, when the water inside a cell has a lower concentration of solutes than the water outside, water tries to move out of the cell. The movement of water across a semipermeable membrane is called osmosis. The force with which an aqueous solution attracts water by osmosis is known as its osmotic potential, which is expressed in megapascals (MPa), a unit of pressure. The osmotic potential generated by an aqueous solution depends on its solute concentration, which is measured as the number of solute molecules in a given volume of water. Osmosis The movement of water across a semipermeable membrane. Osmotic potential The force with which an aqueous solution attracts water by osmosis. The challenge for most aquatic organisms is that they live in water with a solute concentration that differs from the solute concentration of their bodies. This difference causes water and solutes to attempt to move in or out of the organism’s body, which makes it difficult for organisms to maintain the proper amount of water and solutes in their bodies. Maintaining a particular solute concentration in the body is important because solute concentrations affect the way proteins interact with other molecules. In short, altered solute concentrations can disrupt cell functions. If they take in too many solutes, they must rid themselves of the excess. If they take in too few solutes, they must make up the deficit. Because solutes determine the osmotic potential of body fluids, the mechanisms that organisms use to maintain a proper solute balance are referred to as osmoregulation. Osmoregulation The mechanisms that organisms use to maintain proper solute balance.
Adaptations for Osmoregulation in Freshwater
We have seen that the water balance of aquatic animals is closely tied to the concentrations of salts and other solutes in their body tissues and in the environment. Freshwater animals, such as bass and trout, that have higher solute concentrations in their tissues than the surrounding water are said to be hyperosmotic compared to their freshwater environment. Hyperosmotic When an organism has a higher solute concentration in its tissues than the surrounding water. Because water and solutes move in the direction that will equalize the concentrations of solutes on both sides of a membrane, a hyperosmotic organism faces a constant challenge: Water attempts to enter its body while solutes attempt to leave. Figure 2.10a shows how hyperosmotic fish regulate the balance of solutes in their tissues. Freshwater fish continuously gain water both when they consume food and when osmosis occurs across the mouth and gills, which are the most permeable tissues exposed to the water. Fish respond to this influx of water by eliminating the excess water through their urine. They add solutes to their bloodstream by using their gill cells to actively transport solutes into the body from the water. In addition, their kidneys remove ions from their urine.
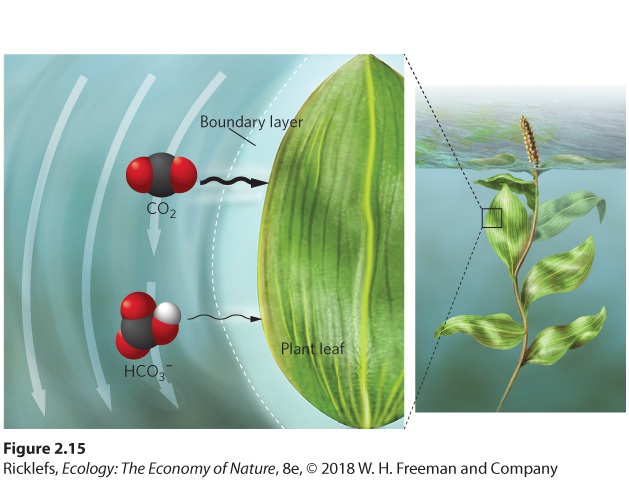
Figure 2.10 Osmoregulation in fish. Exchanges of water and solutes differ between freshwater and marine fish. (a) Freshwater fish are hyperosmotic: They have a higher salt
##### Adaptations for Osmoregulation in Saltwater Animals
concentration in their body than exists in the surrounding water. To maintain salt balance, they must excrete large amounts of water and use their gills and kidneys to actively retain solutes. (b) Saltwater fish are hyposmotic: They have a lower salt concentration in their body than exists in the surrounding water. To maintain salt balance, they must excrete large amounts of solutes, so their gills and kidneys actively exclude solutes.
Adaptations for Osmoregulation in Saltwater
Saltwater animals—such as whales, sardines, and plankton—have lower solute concentrations in their tissues than exist in the surrounding water. Such organisms are said to be hyposmotic compared to their saltwater environment. Hyposmotic organisms face a constant challenge to maintain the balance of water and solutes in their tissues. As shown in Figure 2.10b, water tries to leave their bodies and solutes try to enter. To replace the loss of water, saltwater animals drink large amounts of saltwater and release only small amounts of urine. To counteract the accompanying influx of solutes, the excess solutes are actively excreted out of the body using the kidneys and, in the case of fish, the gills. Hyposmotic When an organism has a lower solute concentration in its tissues than the surrounding water. Some sharks and rays have evolved a unique adaptation to the challenge of water balance in a saltwater environment. Like all vertebrate animals, when sharks and rays digest proteins, they produce ammonia as a by-product, as shown in Figure 2.11. Aquatic vertebrates excrete ammonia in their urine, whereas terrestrial vertebrates generally convert this ammonia to urea, which they then excrete at high concentrations in their urine. Interestingly, sharks and rays convert ammonia into urea, too, but they do not excrete all of it. Unlike terrestrial vertebrates, which maintain concentrations of urea below 0.03 percent in their bloodstream, sharks and rays actively retain urea and allow its concentration to increase to 2.5 percent in their bloodstream. Retaining more than 80 times more urea raises the osmotic potential of their blood to that of seawater without any increase in the concentrations of sodium and chloride ions. Consequently, the movement of water across the animal’s body surface becomes balanced in relation to the surrounding saltwater, with neither gain nor loss. This adaptation frees sharks and rays

from having to drink salt-laden water to replace water lost by osmosis.
Figure 2.11 Unique adaptations to saltwater. When aquatic vertebrates digest proteins, they produce ammonia as a by-product. (a) Most aquatic organisms, such as dolphins, excrete this ammonia in their urine. (b) Sharks and rays convert this ammonia into urea and then retain some of the urea in their bloodstream. The result is a higher solute concentration that helps offset the challenge of being hyposmotic in saltwater. Most vertebrates do not retain much urea in their bloodstream because urea impairs protein function. However, sharks, rays, and many other marine organisms that use urea to maintain their water balance have an additional adaptation: They accumulate high concentrations of a compound called trimethylamine oxide to protect proteins from the harmful effects of urea. Freshwater species of rays do not accumulate urea in their blood, though they are similar to saltwater rays in other respects. This confirms the importance of urea for osmoregulation for species of sharks and rays that live in the ocean. Certain environments pose unusual osmotic challenges. For instance, the salt concentrations in some landlocked bodies of water greatly exceed those of the ocean. This is particularly common in arid regions where, because
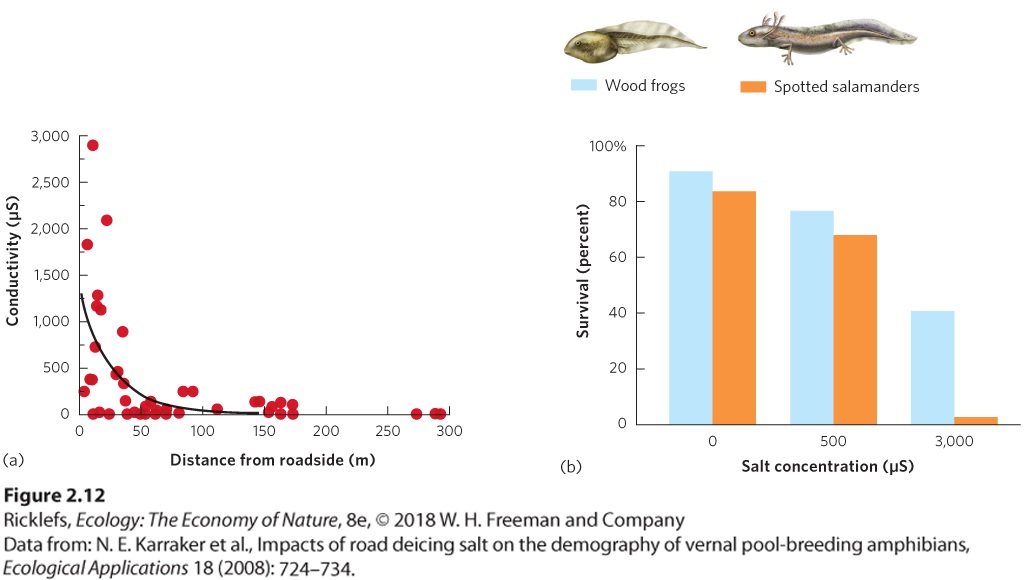
evaporation outpaces precipitation, very high concentrations of solutes accumulate in the water. The Great Salt Lake in Utah, for example, contains 5 to 27 percent salt, depending on the water level. This is up to eight times more salt than seawater. The osmotic potential of the water in the Great Salt Lake would cause most organisms to shrivel. However, a few aquatic creatures, such as brine shrimp, have adaptations that allow them to thrive in these conditions by excreting salt at a very high rate. This level of excretion comes at a high energetic cost, which they meet by feeding on the abundant photosynthetic bacteria that live in the lake. The ability of an organism to cope with the osmotic potential of its environment reflects the outcome of evolutionary processes. As we will discuss in Chapter 4, some environments naturally experience large fluctuations in osmotic potential and the organisms living in these environments for long periods of time have evolved ways to adjust to these fluctuations. However, when changes in osmotic concentrations are not within the natural range that organisms have experienced over evolutionary time, individuals typically lack appropriate adaptations and can be harmed. For example, in the northern United States, various mixtures of salts are spread on roads to melt the ice and snow to provide safer driving conditions during winter weather. With the arrival of warmer spring weather, however, all of this salt has to go somewhere. In 2008, researchers reported on the concentrations of road salt in ponds inhabited by amphibians at different distances from salt-treated roads. Because salt ions allow electricity to be conducted through water, the concentration of salt can be measured in units of microsiemens (μS). As you can see in Figure 2.12a, ponds close to roads had salt concentrations up to 3,000 μS, which corresponds to 0.12 percent salt. Ponds at least 200 m from a road had essentially 0 μS. The researchers then conducted experiments in which they exposed larval wood frogs (Rana sylvatica) and spotted salamanders (Ambystoma maculatum) to a range of relevant salt concentrations, from 0 μS to 3,000 μS. The data, shown in Figure 2.12b, revealed that increases in salt concentration caused the larvae of both species to die in high numbers. Not having been exposed to high salt concentrations during their evolutionary history, these freshwater organisms are not adapted to these stressful conditions and are unable to survive in them.
Figure 2.12 Effect of salt concentration on amphibians. (a) Based on a sample of ponds in the Adirondack Mountains of New York State, ponds closer to roads had higher conductivity, a measure of salt concentration. (b) At this range of salt concentrations, spotted salamanders and wood frogs experienced a decline in survival. The continued contamination of freshwater habitats by road salt has raised the possibility that some species might be able to evolve increased tolerance to salt over multiple generations. In 2017, researchers reported a case of evolving higher salt tolerance in zooplankton, which are tiny crustaceans that live in freshwater lakes. When a particular species of zooplankton, known as water fleas (Daphnia pulex), was exposed to a range of increased salt concentrations for several generations, those raised under high salt concentrations were subsequently more tolerant to salt. While such studies offer hope that some species can rapidly evolve salt tolerance, we do not know how common this evolutionary ability is among the wide diversity of animals that live in freshwater habitats.
Adaptations for Osmoregulation in Aquatic Plants
Some aquatic plants also face major challenges of salt balance. Mangrove trees, for example, grow on coastal mudflats that are inundated by saltwater during high tides (Figure 2.13a). Not only does this habitat impose a high salt load on the trees, but the high osmotic potential of the saltwater environment also makes it difficult for the roots to take up water. To counter these osmotic problems, many mangroves maintain high concentrations of organic solutes —various amino acids and small sugar molecules—in their roots and leaves to increase their osmotic potential so that water will diffuse into the plant’s
##### Analyzing Ecology: Standard Deviation and Standard Error
tissues. In addition, mangroves possess salt glands in their leaves that can secrete salt by active transport to the exterior surface of the leaf (Figure 2.13b). Many mangrove species also exclude salts from their roots by active transport. Because relatively few species of terrestrial plants have evolved this adaptation, mangrove forests do not contain numerous species of plants.
Figure 2.13 Salt balance in mangrove trees. (a) The roots of mangrove trees are frequently submerged in saltwater at high tide. These trees are from Palau, an island in the South Pacific. (b) Specialized salt glands in the leaves of the button mangrove (Conocarpus recta) excrete a salty solution. As the solution evaporates, it leaves behind salt crystals on the outer surface of the leaves.
Analyzing Ecology
Standard Deviation and Standard Error When the researchers tested the effects of road salt on larval amphibians, they exposed groups of larvae to three salt concentrations and replicated the experiment five times. In the previous chapter, we discussed how ecologists use data from such manipulative experiments to determine how different factors affect the means and variance of the variables that are measured. While the variance is a useful measure of how consistent measurements are among replicates, ecologists also use several other related measures of variation, including the sample standard deviation and the standard

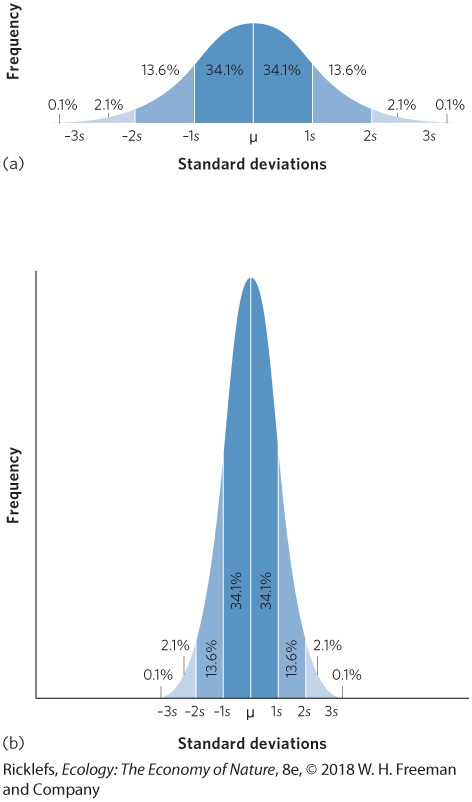
error of the mean. Each of these can be calculated from a measure of sample variance (s2), as we discussed in Chapter 1. When data are collected from limited samples of a much larger distribution of data, we can gain additional information about the data by calculating a sample standard deviation. The sample standard deviation gives us a standardized way of measuring how widely our data are spread from the mean. Large sample standard deviations indicate that many of our data are spread far from the mean. Small sample standard deviations indicate that most of our data points are close to the mean value. Sample standard deviation A statistic that provides a standardized way of measuring how widely data are spread from the mean. If data are normally distributed—that is, if they follow a bell-shaped curve like the one in the figure at the right—then about 68 percent of our data will fall within 1 standard deviation of the mean. Moreover, about 95 percent of our data will fall within 2 standard deviations of the mean and 99.7 percent of our data will fall within 3 standard deviations of the mean. For data that have a wide distribution of frequencies, as in the upper figure (a), the standard deviation value will be large. For data that have a narrow distribution of frequencies, as in the lower figure (b), the standard deviation value will be small. The sample standard deviation, denoted as s, is defined as the square root of the sample variance: s=s2 The standard error of the mean is a useful measurement of variation in our data because it takes into account the number of replicates that were used to measure the standard deviation. The higher the number of replicates, the more precise the estimate we should have of the mean. As a result, an increase in the number of replicates in a given experiment produces a decrease in the standard error of the mean. As we will see in later chapters, the standard error of the mean is commonly used to determine whether two means are significantly different from each other.
The normal distribution. In a normal distribution, the most frequent observations fall near the mean and the least frequent observations occur farther away from the mean. The dark blue areas represent 1 standard deviation from the mean and account for 68.3 percent of all data. The medium and dark blue areas combined represent 2 standard deviations from the mean and account for 95.5 percent of all data. The light, medium, and dark blue areas combined represent 3 standard deviations from the mean and account for 99.7 percent of all data. The remaining small amount of data falls outside of 3 standard deviations. (a) Data that have a wide distribution have a large standard deviation. (b) Data that have a narrow distribution have a small standard deviation. Standard error of the mean A measurement of variation in data that takes into account the number of replicates that were used to measure the standard deviation. The standard error of the mean (denoted as SE) is defined as the sample standard deviation divided by the square root of the number of observations or replicates (denoted as n):
SE=s÷n Consider the following set of observations on the percentage of surviving wood frog tadpoles that were exposed to salt concentrations of 0 µS or 3,000 µS: Replicate 0 µS 3,000 µS Mean Variance 5.0 48.5 Using the data from the five replicates that exposed wood frog tadpoles to 0 µS, we see that the mean survival is 91 percent and the sample variance is 5 percent. Using this sample variance, we can calculate the sample standard deviation as s=s2=5=2.2 and we can calculate the standard error of the mean as SE=s÷n=2.2÷5=1.0 YOUR TURN Use the data collected from the five replicates that exposed wood frog tadpoles to 3,000 µS and calculate the standard deviation of the mean and the standard error of the mean. Explain why the values of s and SE differ.
Concept Check
1. What is the difference between passive and active transport of solutes? 2. Compare and contrast the terms hyperosmotic and hyposmotic.

3. Describe one adaptation for osmoregulation in freshwater animals, saltwater animals, and saltwater plants.
#### 2.3 the Uptake of Gases from Water Is Limited by Diffusion
2.3 The uptake of gases from water is limited by diffusion Almost 21 percent of Earth’s atmosphere is oxygen, but because we live on land, we rarely think about the task of obtaining this required element. Aquatic organisms also require oxygen to support their metabolism, but obtaining a sufficient supply can be a problem for them because of the limited solubility of oxygen in water. The same is true of the CO2 required by aquatic plants for photosynthesis. Organisms obtain sufficient quantities of these necessary gases in the aquatic environment through several adaptations that we examine in this section.
Carbon Dioxide
Getting enough CO2 for photosynthesis is a particular challenge for aquatic plants and algae. The solubility of CO2 in fresh water is about 0.0003 liters of gas per liter of water, which is 0.03 percent by volume, which is about the same as its concentration in the atmosphere. The problem for aquatic plants is that CO2 diffuses very slowly through water, and plants can use the CO2 close to the surface of their leaves faster than it arrives by diffusion. When CO2 dissolves in water, most of the molecules combine with water and are quickly converted to a compound called carbonic acid (H2CO3): CO2+H2O→H2CO3 As shown in Figure 2.14, carbonic acid can increase to high concentrations and provide a reservoir of carbon required for photosynthesis. Depending on the acidity of the water, carbonic acid molecules can release hydrogen ions (H+) to form either bicarbonate ions (HCO3 −) or carbonate ions (CO3 2−):
Figure 2.14 An equilibrium reaction for carbon in water. The reaction converting CO2 into bicarbonate ions (HCO3−) is an equilibrium reaction. When photosynthetic organisms preferentially use CO2, because it is used more efficiently, the amount of dissolved CO2 in the water declines. As the amount of CO2 declines, some of the bicarbonate ions are converted to CO2 to replenish the supply. The size of each circle represents the relative size of the carbon pool. H2CO3→H++HCO3-→2H++CO32Bicarbonate ion (HCO3−) An anion formed by the dissociation of carbonic acid. Carbonate ion (CO32−) An anion formed by the dissociation of carbonic acid. Although some of the carbonate ions can combine with calcium ions to form calcium carbonate, as we saw in Figure 2.8, bicarbonate ions dissolve readily in water, so bicarbonate ion is the most common form of inorganic carbon in aquatic habitats. The result, as shown in Figure 2.14, is a concentration of bicarbonate ions equivalent to 0.03 to 0.06 L of CO2 gas per liter of water (3 to 6 percent)—more than 100 times the concentration of CO2 in air. In short, this means that most of the CO2 that is dissolved into water is rapidly converted to bicarbonate ions in aquatic systems. Dissolved CO2 and bicarbonate ions are in a chemical equilibrium, which represents the balance achieved between H+ and HCO3 − on one hand, and
CO2 and H2O on the other. When CO2 is removed from the water by plants and algae that are performing photosynthesis, some of the abundant bicarbonate ions combine with hydrogen ions to produce more CO2 and H2O. In essence, the reaction now moves in the reverse direction: H++HCO3−→CO2+H2O Although the bicarbonate ion is the most common form of inorganic carbon under moderate pH conditions, CO2 is the most common form under more acidic conditions, such as in bogs. This may be why some species of plants and algae that live in aquatic habitats with low pH can only use CO2 for photosynthesis. In contrast, because CO2 and bicarbonate ions are both abundant under moderate pH conditions (pH = 5 to 9), many species of aquatic plants and algae can use both CO2 and bicarbonate ions for photosynthesis. They can either directly uptake the bicarbonate ion or they can use adaptations to convert the bicarbonate ion to CO2. One way to do this is by secreting an enzyme into the water that is highly effective at converting bicarbonate ions into CO2, which can then be taken up by the organism. Plants and algae can also obtain CO2 by secreting hydrogen ions into the surrounding water. This helps drive the chemical equilibrium in a direction that converts more of the bicarbonate ions into CO2 that can then be taken up by the organism. Even when CO2 and bicarbonate ions are abundant in water, the slow rate at which these carbon sources diffuse through the water prevents organisms from getting access to them. Indeed, carbon dioxide diffuses through unstirred water about 10,000 times more slowly than through air, and diffusion of HCO3 − is even slower because larger molecules diffuse at a slower rate. Compounding this slow rate of diffusion is the fact that every surface of an aquatic plant, alga, or microbe is surrounded by a boundary layer. A boundary layer is a region of unstirred air or water that surrounds the surface of an object. In the water, the boundary layer ranges from as little as 10 micrometers (10 μm, or 0.01 mm) for single-celled algae in turbulent waters to 500 μm (0.5 mm) for a large aquatic plant in stagnant water. As
Figure 2.15 illustrates, because this boundary layer is composed of unstirred
water, CO2 and HCO3 − can be depleted within the boundary layer by uptake —especially in the region closest to the photosynthesizing organism—but the removed gases are slow to be replaced from the surrounding water. Without a boundary layer, the moving water in the surrounding environment would continually provide the plant with a supply of CO2 and HCO3 −. So, despite the generally high concentration of bicarbonate ions in the water, photosynthesis may still be limited by carbon availability within the boundary layer.
Figure 2.15 Boundary layer. A thin boundary of unstirred water along the surface of photosynthesizing organisms slows the rate of gas diffusion through the water. Smaller molecules, such as CO2, diffuse faster than larger molecules, such as bicarbonate ions. Boundary layer A region of unstirred air or water that surrounds the surface of an object.
Oxygen in the atmosphere has a concentration of 0.21 L per liter of air (21 percent by volume). In water, however, the maximum solubility of oxygen is
0.01 L per liter of water (1 percent), under the conditions of fresh water at 0 °C. Oxygen’s low solubility in water can limit the metabolism of organisms
in aquatic habitats. For marine mammals such as whales, this is not a problem because they obtain O2 from the air and store copious amounts in their hemoglobin and myoglobin. For organisms that obtain O2 from the water, however, the problem of low O2 concentration is compounded by its slow diffusion in water, similar to that of CO2. Oxygen is in even shorter supply in waters that cannot support photosynthesis—and therefore do not receive the O2 produced by photosynthesis—including in deep water that does not receive sunlight and in waterlogged sediments and soils. These habitats can become severely depleted of dissolved O2, making them challenging environments for animals and microbes that use aerobic respiration. One important adaptation that allows aquatic animals to deal with a limited amount of oxygen involves the direction of blood flow in the gills. Many aquatic animals have gills to extract oxygen from the water. When water passes over the gills, oxygen diffuses across the membranes of the gill cells and enters the capillaries, which are part of the bloodstream. The key to extracting the most oxygen from the water lies in the use of countercurrent circulation. In countercurrent circulation, two fluids move in opposite directions on either side of a barrier and heat or materials are exchanged. In contrast, concurrent circulation involves two fluids moving in the same direction on either side of a barrier and heat or materials are exchanged. Countercurrent circulation Movement of two fluids in opposite directions on either side of a barrier through which heat or dissolved substances are exchanged. Concurrent circulation Movement of two fluids in the same direction on either side of a barrier through which heat or dissolved substances are exchanged. As illustrated in Figure 2.16, if blood and water were to flow in the same direction, the concentration of oxygen would quickly come to an intermediate equilibrium. After this region of contact, there is no net movement of oxygen across the membrane. In contrast, when blood and water flow in opposite directions, the concentration of oxygen in the water exceeds the concentration in the blood throughout most of the region of contact. This happens because even as the capillaries begin to develop high concentrations of oxygen, the adjacent water still has a higher concentration of oxygen. As a result, the
oxygen continues to diffuse into the gill capillaries. Thus, countercurrent blood flow in animal gills allows much more oxygen to move from the water to the gills.
Figure 2.16 Concurrent versus countercurrent circulation. (a) Concurrent flow in aquatic animals would cause oxygen to diffuse from the water and into the capillaries of the gills. Once 50 percent of the oxygen was transferred, the two flows would come into equilibrium. (b) When animals use countercurrent circulation, more oxygen can be transferred into the gill capillaries because even if the capillaries contain a high amount of oxygen, the oxygen content of the water is higher, allowing oxygen diffusion to continue. Species of animals that live in habitats with low amounts of oxygen have evolved a number of additional adaptations. In the deep oceans, many organisms have very low rates of activity, thereby reducing their need for oxygen. Many species of zooplankton, a group of tiny crustaceans, can increase the amount of hemoglobin in their bodies to the point that their normally transparent bodies turn red. Other animals, such as tadpoles and fish that live in oxygen-depleted swamps, swim to the surface and take gulps of air. Many tadpoles can use this air because they possess primitive lungs in addition to gills. Fish store this air in a swim bladder from which they extract the oxygen into their bloodstream.
One of the most surprising animal adaptations for obtaining oxygen was recently discovered in a species of North American salamander. For more than a century, it was known that eggs of the spotted salamander, which are usually attached to sticks that are submerged in water, have a mutualistic relationship with a species of algae (Oophila amblystomatis). The algae obtain a place to live and photosynthesize while the developing embryo obtains oxygen from the photosynthesizing algae (Figure 2.17). This oxygen benefit is important because it allows the salamander embryos to have a higher survival rate and to hatch earlier and larger. In 2011, however, scientists reported that this relationship was much closer than they had recognized. The algae not only live in the fluid of the egg surrounding the embryo, but also move into the embryo, positioning themselves in between the developing embryo’s cells. This was the first discovery of an algae living within the tissues of a vertebrate animal. Two years later, in 2013, scientists discovered that the algae not only provide oxygen to the developing embryos but also provide sugars produced by photosynthesis. Together, these two beneficial effects of the algae allow faster growth of the salamander embryos.
Figure 2.17 Salamander embryos and algae. The embryos of the spotted salamander contain algae that live in the egg sac and within the cells of the embryo. The algae provide extra oxygen to the embryos, which improves embryo survival and growth. When an environment becomes completely devoid of oxygen, it is referred
to as anaerobic or anoxic. Anaerobic conditions pose problems for terrestrial plants rooted in waterlogged soils, such as the many species of mangrove trees that live along coastal mudflats. The roots of these trees need oxygen for respiration, so the plants have evolved special air-filled tissues extending from the roots that rise above the waterlogged soils and exchange gases directly with the atmosphere (see Figure 2.13). Anaerobic Without oxygen. Also known as Anoxic. Many microbes are able to live in environments without oxygen because they use anaerobic respiration. A common product of anaerobic respiration by bacteria living in anoxic soils is hydrogen sulfide gas (H2S). This gas is the cause of the rotten-egg smell that occurs when soils saturated with water become anaerobic.
Concept Check
1. Why does the boundary layer surrounding a photosynthetic organism make it more difficult to exchange CO2 and O2? 2. What is the equilibrium reaction that illustrates the conversion of CO2 to bicarbonate? 3. Why are deep ocean waters typically low in oxygen?
#### 2.4 Temperature Limits the Occurrence of Aquatic Life
2.4 Temperature limits the occurrence of aquatic life At the beginning of this chapter, we saw that whales possess a number of adaptations that allow them to deal with the cold ocean temperatures, including thick layers of fat, high metabolism, and countercurrent circulation that exchanges heat between the warm arteries and cold veins. For aquatic organisms, most physiological processes occur only within the range of temperatures at which water is liquid. Relatively few organisms can survive temperatures above 45 °C, which is the upper limit of the physiological range for most eukaryotic organisms. In this section, we will look at how organisms have adapted to hot and cold temperatures.
Heat and Biological Molecules
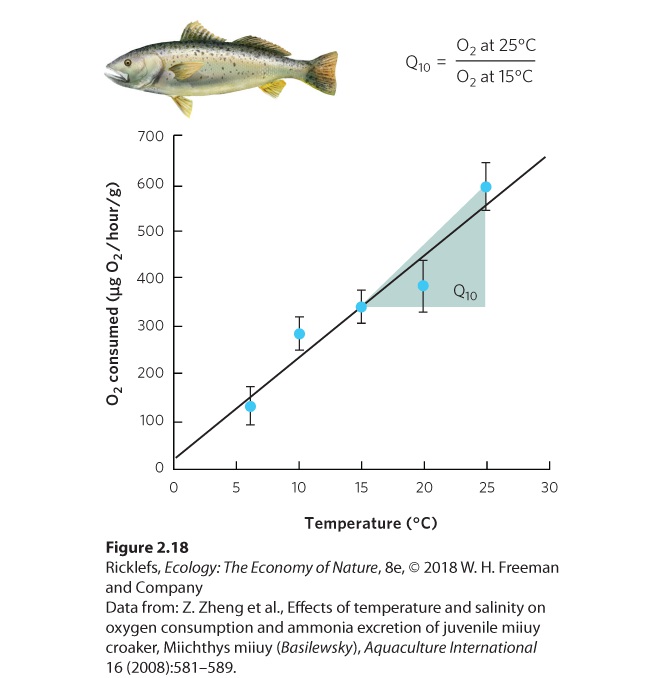
Temperature influences physiological processes because of the way in which heat affects organic molecules. Heat imparts kinetic energy to living systems, causing biological molecules to change shape. Heat also accelerates chemical reactions by increasing the rate of molecule movement. In fact, the rate of most biological processes increases two to four times for each 10 °C rise in temperature. We can see this when we examine the data for a fish known as the miiuy croaker (Miichthys miiuy), shown in Figure 2.18. The fish’s rate of oxygen consumption approximately doubles when the environmental temperature increases from 15 °C to 25 °C. To understand the relationship between temperature and physiological process, we calculate the ratio of the physiological rate at one temperature to the physiological rate when the temperature is 10 °C cooler, a ratio that is referred to as the Q10 of the physiological process. By knowing the Q10 values of different physiological processes, we can better understand which processes are more sensitive to a change in temperature.
Figure 2.18 Oxygen consumption as a function of temperature. For the miiuy croaker, the amount of oxygen it consumes increases as the temperature increases. By dividing the rate of oxygen consumption at 25 °C by the rate of oxygen consumption at 15 °C, one can arrive at the Q10 value for this physiological function. Error bars are standard deviations. Q10 The ratio of the rate of a physiological process at one temperature to its rate at a temperature 10 °C cooler. Higher temperatures allow organisms to do many things more rapidly. They can swim, run, and fly faster. They can also digest and assimilate more food and, as a result, grow and develop faster. Beyond a certain point, however, high temperatures can depress life processes. In particular, proteins and other biological molecules become less stable at higher temperatures, and may not function properly or may lose their structure. The molecular motion caused by heat tends to open up, or denature, the structure of these
molecules. Given that proteins denature at high temperatures, scientists have been intrigued by the fact that some organisms, such as thermophilic (heat-loving) bacteria, can live at very high temperatures. For example, some photosynthetic bacteria can tolerate temperatures as high as 75 °C and some archaebacteria can live in hot springs at temperatures up to 110 °C (Figure 2.19). The chemosynthetic bacteria that live near deep-sea vents are a group of thermophiles that we discussed in Chapter 1. Researchers have discovered that the proteins of thermophilic bacteria have higher proportions of particular amino acids that form stronger bonds than the proteins of other, heat-intolerant species. These strong forces of attraction within and between molecules prevent them from being shaken apart under high temperatures, so the proteins do not denature.
Figure 2.19 Thermophiles. Some species of bacteria and archaebacteria can live under very hot conditions, such as these hot springs in Fly Geyser, Nevada. Thermophiles often come in a variety of bright colors. Thermophilic Heat-loving.
Temperature also affects other biological compounds. For instance, the physical properties of fats and oils, which are major components of cell membranes and constitute the energy reserves of animals, depend on temperature. When fats are cold, they become stiff; when they are warm, they become fluid. Organisms are negatively affected when exposed to temperatures that range above or below those to which they are adapted. Water used to keep nuclear power plants cool provides an example of this problem. Many nuclear power plants extract water from nearby lakes, rivers, or oceans for cooling the power plants and then return the water—which has become much warmer in the process—to its source. But the organisms in these bodies of water have no evolutionary history of being exposed to such high water temperatures and many of them die. In Ohio, for example, a coal-burning power plant raised the temperature of an adjacent stream up to 42 °C, which is much warmer than would naturally occur, and warmer than the maximum of 32 °C set by the U.S. EPA. Surveys found numerous dead fish in the stream and low numbers of fish downstream where the stream emptied into the Ohio River. Discharging water that is too hot to sustain aquatic species is known as thermal pollution. Understanding the thermal optima for aquatic species has led to regulations that restrict how much a discharging power plant can raise the temperature of a lake or river. Thermal pollution Discharging water that is too hot to sustain aquatic species.
Cold Temperatures and Freezing
Temperatures on Earth’s surface rarely exceed 50 °C, except in hot springs and at the soil surface in hot deserts. However, temperatures below the freezing point of water are common, particularly on land and in small ponds, which may freeze solid during winter. When living cells freeze, the crystal structure of ice disrupts most life processes and may damage delicate cell structures, eventually causing death. Many organisms successfully cope with freezing temperatures either by maintaining their body temperatures above the freezing point of water, or by activating chemical pathways that enable them to resist freezing or to tolerate its effects. It might surprise you to learn that marine vertebrates are susceptible to
freezing in cold seawater. How can blood and body tissues freeze solid while immersed in liquid water? The answer is that dissolved substances lower the temperature at which water freezes. While pure water freezes at 0 °C, seawater, which contains about 3.5 percent dissolved salts, freezes at −1.9 °C. Because the blood and body tissues of most vertebrates contain about half the salt content of seawater, the animals can freeze solid before the surrounding seawater does. Marine animals have evolved a number of adaptations to combat the problem of freezing in water. We know that high salt concentrations interfere with many biochemical processes, so raising the concentration of solutes in blood and tissues is not a viable option. Instead, some Antarctic fish prevent freezing by raising their blood and tissue concentrations of nonsalt compounds such as glycerol. Glycerol is a chemical that prevents the hydrogen bonds of water from coming together to form ice unless the temperatures are well below freezing. A 10 percent glycerol solution in the body lowers the freezing point of water to about −2.3 °C without severely disrupting biochemical processes. This is enough to lower the freezing point of the body below the freezing point of seawater. Glycoproteins are another group of compounds that can be used to lower the freezing temperature of water. Glycerol and glycoproteins act as antifreeze compounds, similar to the antifreeze used in automobiles, and allow fish such as the Arctic cod (Boreogadus saida) to remain active in seawater that is cold enough to cause most fish to freeze solid (Figure 2.20). Some terrestrial invertebrates also use the antifreeze approach; their body fluids may contain up to 30 percent glycerol as winter approaches.
Figure 2.20 Adaptation to different water. The Arctic cod, which is commonly harvested in Russia for human consumption, can live in water that is less than 4 °C. Antifreeze compounds in its tissues prevent it from freezing solid.
Glycerol A chemical that prevents the hydrogen bonds of water from coming together to form ice unless the temperatures are well below freezing. Glycoproteins A group of compounds that can be used to lower the freezing temperature of water. Supercooling provides a second physical solution to the problem of freezing. Under certain circumstances, liquids can cool below the freezing point without developing ice crystals. Ice generally forms around an object, called a seed, which can be a small ice crystal or other particle. In supercooling, however, glycoproteins in the blood impede ice formation by coating any ice crystals that begin to form. In the absence of ice seeds, pure water may cool to more than −20 °C without freezing. Such supercooling has been recorded down to −8 °C in reptiles and −18 °C in invertebrates. Supercooling A process in which glycoproteins in the blood impede ice formation by coating any ice crystals that begin to form.
Thermal Optima
Every organism is best suited to a narrow range of environmental conditions; this range defines the optimum environmental conditions. In terms of temperature, most organisms have a thermal optimum, meaning the range of temperatures in which they perform best. The thermal optimum is determined by the properties of enzymes and lipids, the structures of cells and tissues, body form, and other characteristics that influence the ability of an organism to function well under the particular conditions of its environment. Returning to the example of fish in the ocean waters of the Antarctic, many species swim actively and consume oxygen at a rate comparable to fish living in much warmer regions near the equator. However, if you put a tropical fish in cold water, it becomes sluggish and soon dies; conversely, Antarctic fish cannot tolerate temperatures warmer than 5 °C to 10 °C. Optimum environmental conditions The narrow range of environmental conditions to which an organism is best suited. Thermal optimum The range of temperatures within which organisms perform best.
Certain adaptations allow fish in cold oceans to swim as actively as fish in warm oceans. Swimming involves a series of biochemical reactions, most of which depend on enzymes. These reactions generally proceed more rapidly at high temperatures, so cold-adapted organisms must either have more of the substrate for a biochemical reaction, more of the enzyme that catalyzes the reaction, or a different version of the enzyme that operates better under colder temperatures. Different forms of an enzyme that catalyze a given reaction are called isozymes. Isozymes Different forms of an enzyme that catalyze a given reaction. Consider the case of the rainbow trout (Oncorhynchus mykiss), a fish that lives in cold streams throughout much of North America. Seasonal changes in temperature are predictable for the trout: During winter, water temperatures may drop close to the freezing point, while summer temperatures can become very warm. In response to these seasonal temperature changes, the trout has evolved the ability to produce different isozymes in winter and in summer. One of these enzymes is acetylcholinesterase, which plays an important role in ensuring proper functioning of the nervous system by binding with the neurotransmitter acetylcholine. To understand how well different isozymes function at different temperatures, we examine the rate of the chemical reaction between acetylcholine and acetylcholinesterase, a measure known as enzyme-substrate affinity. The winter isozyme, shown as the blue line in Figure 2.21, best catalyzes the reaction at 0 °C to 10 °C. This affinity drops rapidly at higher temperatures. In contrast, the summer isozyme, shown as the orange line, has a weak affinity for acetylcholine at 10 °C and catalyzes reactions the best at 10 °C to 20 °C, though the affinity drops slowly at higher temperatures. As you might predict, the particular isozyme that a trout produces depends on the temperature of the water it lives in. When trout live at 2 °C, they produce the winter isozyme, whereas when they live at 17 °C, they produce the summer isozyme.
Figure 2.21 Using isozymes as adaptations to changing water temperatures. In winter, the rainbow trout produces a form of the enzyme acetylcholinesterase that has a high substrate affinity at 0 °C to 10 °C, but a lower substrate affinity at warmer temperatures. In summer, the trout produces a different form of the enzyme, which has a high substrate affinity at 10 °C to 20 °C, but a lower substrate affinity at colder temperatures.
Concept Check
1. Explain the adaptation that allows thermophiles to survive in very high temperatures. 2. Describe the adaptations that enable fish to survive in very cold seawater. 3. How do isozymes help organisms function in habitats with a wide range of temperatures?
Concepts
The Decline of Coral Reefs Coral reef diversity. Coral reefs, such as this one off Beqa Island, Fiji, in the South Pacific, are among the most species-diverse places on Earth. Coral reefs are some of the most beautiful places on Earth and are home to an incredible diversity of species. For instance, the Great Barrier Reef off the eastern coast of Australia contains more than 400 species of coral, 200 species of birds, and 1,500 species of fish. For these reasons, many people become concerned when the biodiversity of coral reefs is threatened by human activity. We have long understood that overfishing and pollution have affected the species that inhabit coral reefs. In the past 20 years, however, scientists have discovered that changes in the abiotic aquatic environment—including changes in temperature, pH, and salinity—are damaging coral reef ecosystems. Corals are a group of tiny animals that secrete hard exoskeletons made of limestone (calcium carbonate). They
can be found worldwide in relatively shallow ocean waters that are low in nutrients and food. Although each individual coral is small—only a few millimeters in size—the limestone bodies of dead corals accumulate over hundreds or thousands of years to form massive coral reefs that can exceed 300,000 km2. The corals survive in these lownutrient waters by living in symbiotic relationships with several species of photosynthetic algae known as zooxanthallae. Corals have tubular bodies with tentacles that stick out and catch bits of food and detritus that go by. Their digestion produces CO2, which can be used by symbiotic algae during photosynthesis. As we learned earlier in this chapter, CO2 can often be difficult for aquatic producers to obtain. In exchange, the algae produce O2 and sugars, some of which can be passed to the coral. In short, the algae get a safe place to live and a steady supply of CO2 for photosynthesis, while the corals get a source of energy in the form of sugars and a steady supply of O2 for respiration.
Coral reef decline. Numerous changes in the ocean environment are causing declines in the numbers of live corals. In the Caribbean, for example, a sharp drop in the number of live corals occurred from 1963 to 2012. After J. Jackson et al., Tropical Americas Coral Reef Resilience Workshop, http://cmsdata.iucn.org/downloads/caribbean_coral_report_jbcj_0309 During the past two decades, scientists have learned that the symbiotic relationship between the corals and algae is very sensitive to environmental changes. When corals are experiencing stress in their environment, they expel the symbiotic algae from their bodies. Because corals obtain their bright colors from the symbiotic algae, corals that expel their algae often look white and are said to experience coral bleaching. Coral bleaching Loss of color in corals as a result of the corals expelling their symbiotic algae.
Coral bleaching is associated with unusually high ocean temperatures. As we discussed in this chapter, while increases in water temperature can increase the rate of chemical reactions, temperatures that exceed the thermal optima can be detrimental. Bleaching can begin if summer ocean temperatures are even just 1 °C higher than the average maximum. If the temperature rise is brief—a few days or weeks—the algae can recolonize the corals. However, the corals will experience slower growth and reduced reproduction. However, if the temperatures are 2 °C to 3 °C higher than the average maximum for longer periods of time, corals can die. The rise in ocean temperatures also appears to stress corals in a way that makes them more susceptible to pathogens that can kill them. During the past two decades, scientists have witnessed major bleaching events worldwide: in 1998, 2003, 2005, 2010, and 2016. During the 2016 event, for example, areas in the Great Barrier Reef off the coast of Australia experienced up to 67 percent declines in the survival of shallow-water corals. Fortunately, other regions around Australia experienced much lower death rates. With continuing increases in global temperatures, a topic we will cover in detail in later chapters, temperature-induced coral bleaching is expected to continue. Changes in salt concentration are also an issue for corals. High ocean temperatures increase the evaporation of water from the ocean, which increases the ocean’s salt concentration. In the case of corals, the stress of increased salt concentrations combined with the stress of high temperatures makes them increasingly vulnerable to coral bleaching and coral death. Another source of decline in corals is a decrease in the
pH of seawater. Because atmospheric CO2 is in equilibrium with dissolved CO2 in the ocean, the recent increase in atmospheric CO2 has caused an increase in dissolved CO2. This, in turn, has caused an increase in carbonic acid, and a decline in the pH of the oceans. As we have seen, carbonic acid (H2CO3) disassociates to bicarbonate ions (HCO3 −) and hydrogen ions (H+). The hydrogen ions can then combine with other carbonate ions to produce bicarbonate, making carbonate ions less available for the corals to create their exoskeleton of calcium carbonate. This decline in pH can also dissolve the calcium carbonate skeletons of the coral. The decline in coral reefs has become a serious problem. In the Caribbean, for example, the percent of live corals has declined from more than 50 percent in 1977 to less than 10 percent in 2012. Only 20 years ago, scientists debated whether abiotic changes were playing a role in the decline of corals. As more data have been collected, there is now general agreement that changes in temperature, pH, salinity, and pollutants are all contributing. Given that these impacts are expected to rise even more in future decades, scientists currently predict that many species of coral will continue to decline. However, some species of coral may have sufficient genetic variation to allow evolutionary changes that permit them to adapt to the changing environmental conditions and persist. SOURCES: McGuirk, R. Great Barrier Reef sees record coral deaths this year, Associated Press, November 29, 2016, http://www.nytimes.com/aponline/2016/11/28/world/asia/ap-as-
australia-dying-coral.html. Renema, W., et al. 2016. Are coral reefs victims of their own past success?, Science Advances 2: e1500850, doi:10.1126/sciadv.1500850. Carpenter, K. E., et al. 2008. One-third of reef-building corals face elevated extinction risk from climate change and local impacts, Science 321:560–563. Hoegh-Guldberg, O., et al. 2007.Coral reefs under rapid climate change and ocean acidification, Science 318:1737–1742.
Summary of Learning Objectives
2.1 Water has many properties favorable to life. These include thermal resistance to changes in temperature, a density and viscosity that select for adaptations for movement, and the ability to dissolve many elements and compounds that are essential to life. Key Terms: Viscosity, Ions, Saturation, Acidity, pH, Acid deposition
2.2 Aquatic animals and plants face the challenge of water and salt balance. This challenge occurs because animal tissues are generally hyperosmotic or hyposmotic compared to the solute concentration of the surrounding aquatic environment. Key Terms: Solute, Semipermeable membrane, Passive transport, Active transport, Osmosis, Osmotic potential, Osmoregulation, Hyperosmotic, Hyposmotic, Sample standard deviation, Standard error of the mean
2.3 The uptake of gases from water is limited by diffusion. This limitation can make it difficult for organisms to exchange gases such as CO2 and O2. Diffusion is slowed because organisms are surrounded by a thin boundary layer of unstirred water. Although CO2 can be abundant in water—either as a dissolved gas or in the form of bicarbonate ions—oxygen’s low solubility in water makes it less abundant. As a result, many animals have respiratory structures with large surface areas and countercurrent circulation to obtain oxygen from the water. Key Terms: Bicarbonate ion (HCO3 −), Carbonate ion (CO3 2−), Boundary layer, Countercurrent circulation, Concurrent circulation, Anaerobic
2.4 Temperature limits the occurrence of aquatic life. Although warmer temperatures increase the rate of chemical reactions, excessively high temperatures cause proteins and other important molecules to become unstable and denature. Cold temperatures also pose a challenge, and many organisms living under near-freezing conditions have evolved adaptations that include the use of glycoproteins and supercooling to prevent the harmful effects of ice crystals growing inside their cells. Organisms that live under a wide range of temperatures often use isozymes to permit proper physiological function at each temperature. Key Terms: Q10, Thermophilic, Thermal pollution, Glycerol, Glycoproteins, Supercooling, Optimum, Thermal optimum, Isozymes, Coral bleaching
Critical Thinking Questions
1. As water cools below 4 °C, it expands and becomes less dense. Why is this beneficial to organisms living in a lake during a cold winter? 2. Given that heat readily transfers between an organism’s body and water, what would you predict about the temperature of the boundary layer around an animal’s body compared to the temperature of the surrounding water in cold weather? 3. Based on the properties of water, why do non-polar compounds such as fats and oils not dissolve well in water? 4. In lakes around the world, some have thick deposits of limestone, which is made up of calcium carbonate, and are resistant to the effects of acid rain. What parallels can you draw between these lakes and the smokestack scrubbers being used in coal-powered factories? 5. Both saltwater and freshwater fish have adaptations to control the movement of water and salts across their external surfaces. Describe what would happen without these adaptations. 6. How would the process of natural selection favor the evolution of increased salt tolerance during the decades that cold regions of the world have increased the use of road salt, which ultimately finds its way into streams and lakes? 7. Using your knowledge of how CO2 dissolves and reacts in water, explain why global increases in atmospheric CO2 should cause a decrease in the pH of ocean water. 8. How would counter-current selection in fish gills be beneficial for an animal trying to get rid of CO2? 9. How might the use of isozymes permit an aquatic organism to persist with a gradual increase in thermal pollution but not a rapid increase in thermal pollution?
10. If the algae that live in corals need CO2 as a carbon source for photosynthesis, why do scientists argue that global increases in CO2 are playing a role in the decline in coral reefs around the world?
#### Graphing the Data: Determining Q10 Values in Salmon
GRAPHING THE DATA Determining Q10 Values in Salmon Salmon researchers measured the oxygen demands of fish across a range of different water temperatures. Using their data in the following table, and with reference to Figure 2.18, create a graph that demonstrates how oxygen demand changes with temperature. Based on this graph, calculate the Q10 value for the salmon between 5 and 15 °C. Next, calculate the Q10 value for the salmon between 10 and 20 °C and compare your two answers. Temperature (°C) Oxygen demand (mg O2/kg/minute) 2.0 2.7 4.0 5.6