4 Adaptations to Variable Environments
4Adaptations to Variable Environments Gray treefrog tadpoles. Tadpoles of the gray treefrog that live without predators exhibit high activity and develop relatively small tails that are drab. In contrast, tadpoles raised with predators exhibit low activity and develop large, red tails. The Fine-Tuned Phenotypes of Frogs Every spring the female gray treefrog (Hyla versicolor) must choose where she will lay her eggs. The treefrog is a medium-sized frog that lives throughout much of eastern North America and through the central United States to the Gulf Coast of Texas. As adults, they spend most of their time in forests, feeding on insects in trees. However, in the spring the male and female frogs move to the water to breed. Under ideal conditions, females lay their eggs in ponds that remain free of predators throughout the two months it takes for them to hatch into tadpoles and then metamorphose into frogs.

Unfortunately, the females have no way to predict whether a pond will contain predators in the weeks ahead. Their offspring, however, have evolved an amazing ability to adjust to a wide range of different predator environments. After a female frog lays her eggs, the embryos experience rapid growth and development; in just a few days, they are ready to hatch. The timing, however, can change depending on the presence of predators, such as crayfish, that commonly consume frog eggs. Embryos of gray treefrogs, like many species of frogs, can detect the presence of predators by sensing chemical cues that the predators produce. When the embryo detects a nearby predator, development accelerates and it hatches into a tadpole sooner than it would normally, thereby reducing the risk of predation as an embryo. Although it survives the egg predator, it emerges smaller than it would have been if it had remained an embryo for a longer period, so it is more vulnerable to tadpole predators. The gray treefrog has also evolved an ability to respond to changing environmental conditions after eggs hatch into tadpoles. Like the embryos, gray treefrog tadpoles can sense predators in the water through chemical cues. When they detect the presence of a predator, tadpoles hide at the bottom of the pond, become less active, and start to change shape. Within a few days, the tadpoles develop big red tails. While the reason for the red color remains a mystery, the large tails improve a tadpole’s ability to escape from predators because they serve as a large sacrificial target that can be lost to a predator and regrown. However, the energy required to grow (or regrow) a large tail is so great that other body parts cannot grow as fast. Consequently, tadpoles with large tails have smaller mouths and shorter digestive tracts, which limit their ability to eat and grow. In short, the presence or absence of predators influences the tadpole’s phenotype. In an environment without predators, tadpoles become highly active, small-tailed, and fast growing. In the presence of predators, tadpoles become inactive, large-tailed, and slow growing. But the flexibility does not end there. Tadpoles not only detect the presence of predators but also distinguish different species of predators. This allows them to distribute the use of their energy
according to the level of risk; they adjust their defenses most strongly to the most dangerous predators, and produce more modest defenses against less dangerous predators. This strategy has the advantage of allowing the tadpole to use its energy where it will make the most difference to its survival. “Tadpoles can even detect what a predator had for lunch.” Additional studies have also shown that tadpoles can even detect what a predator had for lunch. When a predator habitually feeds on tadpoles, the treefrog tadpoles spend more time hiding and undergo changes in shape as we have noted. However, if the predator is feeding on something else, such as snails, the treefrog tadpoles spend less time hiding and only undergo small shape changes. In essence, the tadpoles detect that they are in more danger from predators feeding on tadpoles than predators feeding on other prey, and defend themselves accordingly. The chemical cues emitted by predators feeding on tadpoles could be produced when predators attack and chew the tadpoles or when predators digest the tadpoles. In 2016, researchers isolated each of these possibilities and discovered that chemical cues are emitted during both stages in the predation event. Chemical cues from predators who are only chewing or only digesting tadpoles induce moderate changes in tadpole traits, but predators who are chewing and digesting induce a large change in tadpole traits. Gray treefrog tadpoles also respond to other environmental conditions, including the presence of intraspecific and interspecific competitors. In ponds without predators, many tadpoles survive and compete for algae, the food on which they depend. In response to the relative scarcity of food, tadpoles develop larger mouths and longer intestines. The larger mouths contain wider rows of toothlike projections, and these improve the ability to scrape algae from rocks
and leaves. Longer intestines enable a more efficient extraction of energy from the limited amount of available algae. To grow a large body, however, the tadpole must divert energy away from its tail. As a result, tadpoles that live in an environment with high competition have smaller tails. There is a cost associated with the phenotypic adaptation best suited to survive high competition; if a predator arrives, the tadpoles with smaller tails will be more vulnerable because they do not have the anti-predator phenotype. The story of the gray treefrog represents a situation in which a species can experience a tremendous amount of environmental variation within and across generations. In response to this variation, the gray treefrog has evolved a wide range of strategies that help to improve its fitness. The responsiveness of the gray treefrog tadpole represents just one example of how organisms have evolved to quickly respond to variation in their environment. In this chapter, we will explore the wide range of environmental variation and look at how species have evolved the ability to alter phenotype in response to changing environments. SOURCES: Shaffery, H. M., and R. A. Relyea. 2016. Dissecting the smell of fear: Investigating the processes that induce anti-predator defenses in larval amphibians, Oecologia 180:55-65. Schoeppner, N. M., and R. A. Relyea. 2005. Damage, digestion, and defense: The roles of alarm cues and kairomones for inducing prey defenses, Ecology Letters 8:505–512.
Learning Objectives
After reading this chapter, you should be able to:
4.1 Illustrate how variable environments favor the evolution of variable phenotypes.
4.2 Explain the commonly evolved adaptations in response to enemies, competitors, and mates.
4.3 Understand commonly evolved adaptations in response to variable
4.4 Describe why migration, storage, and dormancy are strategies to survive extreme environmental variation.
4.5 Explain how variation in food quality and quantity is the basis of optimal foraging theory. In Chapters 2 and 3, we discussed the range of environmental conditions in aquatic and terrestrial environments and the many adaptations that organisms have evolved to handle these conditions. However, the environmental conditions an organism faces can also vary considerably over time and among different places. In this chapter, we will look at environmental variation and the adaptations that organisms have evolved to respond to these changing environments.
#### 4.1 Environmental Variation Favors the Evolution of Variable Phenotypes
4.1 Environmental variation favors the evolution of variable phenotypes Most properties of the environment change over time and across space, and they can change at different rates. For example, air temperature can plunge dramatically in a matter of hours as a cold front passes through a region. On the other hand, ocean water may require weeks or months to cool the same amount. In this section, we will look at temporal and spatial variation in the environment, and then examine how such variation favors the evolution of variable phenotypes.
Temporal Environmental Variation
Temporal environmental variation describes how environmental conditions change over time. Some temporal variation is predictable, including alternation of day and night and seasonal changes in temperature and precipitation. Superimposed on these predictable cycles are irregular and unpredictable variations, including weather and climate. Weather refers to the variation in temperature and precipitation over periods of hours or days. Climate refers to the typical atmospheric conditions that occur throughout the year, measured over many years. For example, the climate of Wyoming is typically cold and snowy in the winter but hot and dry in the summer. However, the weather on any particular day cannot be predicted very far in advance; it can vary over intervals of a few hours or days with the movement of cold and warm air masses. Although climate describes the average conditions of a given location throughout the year, climate also can vary over long periods of time. For example, a location might experience a string of years that are much wetter or drier than the average year. Temporal environmental variation The description of how environmental conditions change over time. Weather The variation in temperature and precipitation over periods of hours or days. Climate The typical atmospheric conditions that occur throughout the year, measured over many years.

Some types of temporal variation can cause large impacts on ecosystems but occur infrequently in a particular place. For example, droughts, fires, tornadoes, and tsunamis can cause major changes in the landscape, yet their frequency at a particular location is rare. Other sources of temporal variation, such as the current warming of Earth, occur very slowly over decades and centuries. How organisms and populations respond to temporal variation in their environment depends on the severity of the change and how often it occurs. In general, the more extreme the event, the less frequently it occurs.
Spatial Environmental Variation
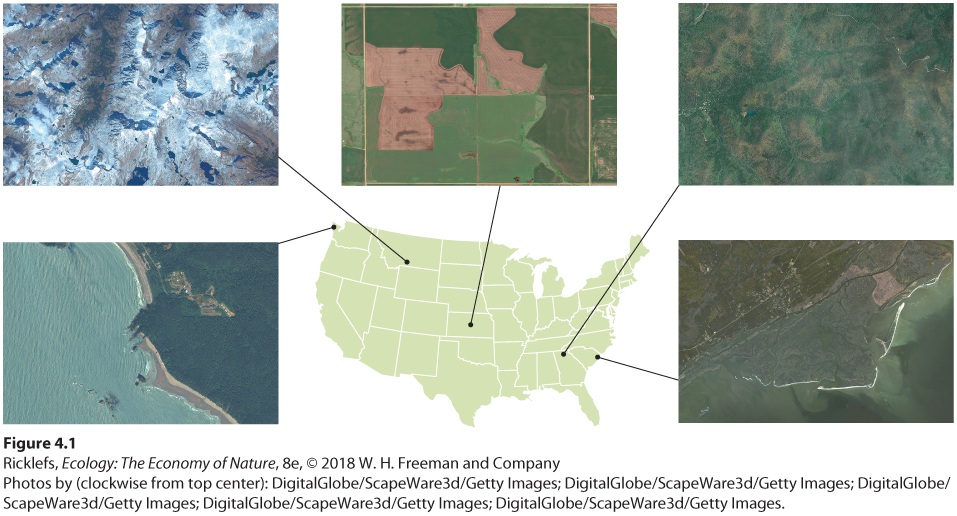
Environmental variation also occurs from place to place due to large-scale variation in climate, topography, and soil type (Figure 4.1). If you were to fly across the United States from Oregon to South Carolina, for instance, you would observe a series of major, large-scale environmental changes along the trip: the rocky western coast, northwestern forests, western rangelands, midwestern agricultural fields, eastern forests, and coastal beaches. At smaller scales, environmental variation is generated by the structures of plants, the activities of animals, the composition of soil, and the activities of humans. As with temporal variation, a particular scale of spatial variation may be important to one organism but not to another. The difference between the top and the underside of a leaf, for example, is important to an insect, but not to a moose, which happily eats the whole leaf, insect and all.
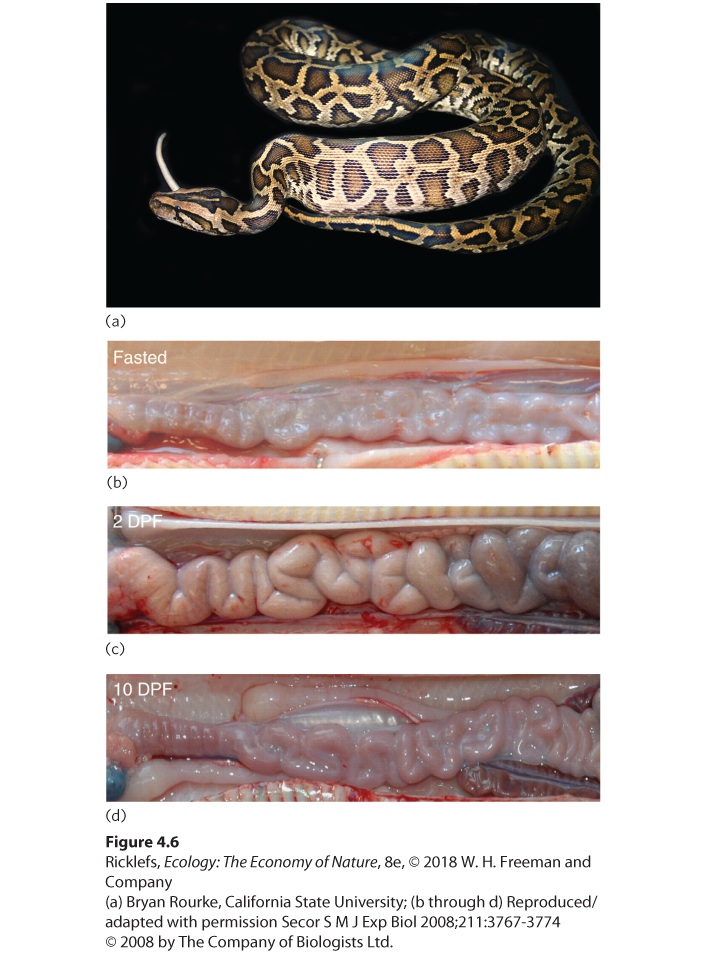
Figure 4.1 Spatial variation in the environment. As one travels from the West Coast to the East Coast of the United States, there is tremendous variation in the natural and humanaltered habitats.
Moving through environments that vary in space, an individual experiences environmental variation as a sequence in time. In other words, a moving individual perceives spatial variation as temporal variation. The faster an individual moves and the smaller the scale of spatial variation, the more quickly the individual encounters new environmental conditions and the shorter the temporal scale of the variation. This principle applies to plants as well as to animals. For example, as plant roots grow, they push their way through the soil, and soils commonly contain small-scale variation in moisture and nutrients. If a plant’s roots grow quickly in soil with a fine scale of variation, the plant roots will frequently encounter new soil environments. Similarly, wind and animals disperse the seeds of plants. The variety of habitats in which the seeds might land depends on the distance the seeds travel and the scale of spatial variation in the habitat.
Phenotypic Trade-Offs
In Chapter 2, we saw that rainbow trout that express cold-water isozymes in their tissues perform well in cold water but poorly in warm water. In contrast, trout expressing warm-water isozymes perform well in warm water but poorly in cold water (Figure 2.21). Throughout the natural world, we commonly see that phenotypes well suited to one environment may be poorly suited to other environments.
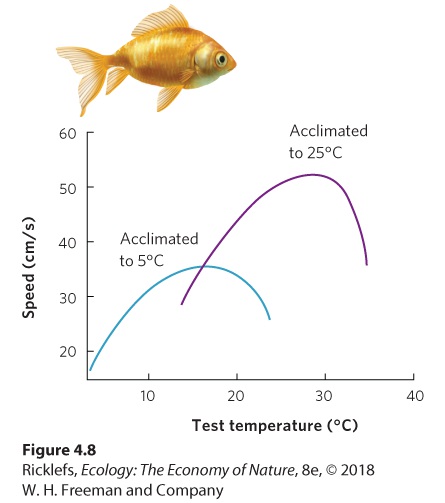
Figure 4.2 illustrates phenotypic fitness in relation to different environments. In Figure 4.2a, an individual possessing phenotype X is well suited to environment X and therefore experiences high fitness. In environment Y, however, the phenotype is no longer well suited to the environment and it therefore experiences reduced fitness. In contrast, an individual that possesses phenotype Y is well suited to environment Y and experiences high fitness in environment Y. However, it is poorly suited to environment X, so it experiences reduced fitness in environment X. When a given phenotype experiences higher fitness in one environment whereas other phenotypes experience higher fitness in other environments, we say that there is a phenotypic trade-off, meaning that neither phenotype does well in both environments.
Figure 4.2 Environments, phenotypes, and fitness. Different environments cause phenotypes to experience different amounts of fitness. (a) Phenotypic plasticity evolves because a phenotype has high fitness in one environment and low fitness in another environment. (b) Given this trade-off, nonplastic genotypes have high fitness in one environment but low fitness in other environments. In contrast, a plastic genotype can have high fitness in both environments. (c) If we consider the average fitness across both environments, we see that the plastic genotype experiences higher average fitness than either of the two nonplastic genotypes. Phenotypic trade-off A situation in which a given phenotype experiences higher fitness in one environment, whereas other phenotypes experience higher fitness in other environments. But what if an individual could produce a range of phenotypes and each phenotype could perform well in a specific environment? Individuals with mutations that allow them to produce multiple phenotypes that are uniquely suited to each environment would experience relatively high fitness in both environments and therefore be favored by natural selection. The ability of a single genotype to produce multiple phenotypes is called phenotypic plasticity. Phenotypic plasticity is a widespread phenomenon in nature; nearly every organism—bacteria, protists, plants, fungi, and animals— possesses phenotypically plastic traits. Different traits can change at different rates and these environmentally induced traits can be either reversible or irreversible. By changing its traits, an individual often maintains a high level
of performance when the environment changes. This means that phenotypically plastic traits often are a mechanism of achieving homeostasis, a concept we discussed in Chapter 3. Phenotypic plasticity The ability of a single genotype to produce multiple phenotypes.
Figure 4.2b shows the advantage of being phenotypically plastic. In contrast to the two nonplastic genotypes, labeled as genotype X and genotype Y, the plastic genotype, labeled as genotype Z, has a relatively high fitness in both environments because it can produce a phenotype that is nearly as fit as genotype X in environment X and a phenotype that is nearly as fit as genotype Y in environment Y. If we examine the average fitness of the three different genotypes in Figure 4.2c, we see that it is higher for the plastic genotype. Whenever environmental variation coincides with phenotypic trade-offs across different environments, natural selection will favor the evolution of phenotypic plasticity. For a long time, scientists applied the concept of phenotypic plasticity to only certain types of traits, such as changes in morphology or physiology. However, we now recognize that many other types of traits—for example, behavior, growth, development, and reproduction—frequently represent phenotypes that can change under different environmental conditions. As a result, the conceptual framework of phenotypic plasticity has expanded in recent years to consider all these types of traits. We can see the advantage of phenotypic plasticity in the example of the gray treefrog tadpoles discussed at the beginning of this chapter. In environments with predators, the tadpoles produce a phenotype that is well suited to escaping detection and capture. In predator-free environments, the tadpoles produce a phenotype that is well suited for faster growth. If tadpoles only had a single phenotype, they would perform poorly whenever the environment changed. In contrast, a tadpole that can change its behavior and body shape performs relatively well when the environment changes. The fitness advantage of phenotypic plasticity occurs whenever environmental variation in space or time occurs frequently. If environmental conditions frequently change, then the phenotype favored by natural selection also changes frequently and this gives the plastic genotype a higher average
fitness than the nonplastic genotype. If spatial or temporal variation is not common, a single phenotype will be favored; the phenotype with the highest fitness in the stable environment will be favored. As we have noted in earlier chapters, all phenotypes are the product of genes interacting with environments. As a result, environmentally induced traits have a genetic basis, but reflect the ability of the environment to turn certain genes on or off, which causes different phenotypes to develop. The environments that induce these changes may change so rapidly that they occur within a generation or they may be somewhat slower and vary across generations. The experience of the tree frog is a good example; predators and competitors can differ substantially from pond to pond in a particular year, from year to year in a particular pond, and even from week to week during the time it takes for a generation of tadpoles to metamorphose and leave the pond as frogs.
Environmental Cues
For an organism to alter its phenotype in an adaptive way, it must first be able to sense its environmental conditions. For example, the gray treefrog tadpole first senses whether the pond contains predators or competitors and then alters its phenotype accordingly to improve its fitness. As we will see throughout the rest of this chapter, environmental cues can take many forms, including smells, sights, sounds, and changes in abiotic conditions. Of the numerous potential cues an organism might use, the best are those that offer the most reliable information about the environment. For example, an animal that requires a reliable cue about competition for food could use the presence of a large number of conspecifics—members of its own species—that will be eating the same thing. But if ample food is available, even a large number of conspecifics would not result in competition for food, so the number of conspecifics might be a poor indicator of the level of competition. If this is the case, then a better environmental cue to indicate high competition for food might be the amount of food that an individual can acquire each day. In such a case where a species has multiple possible environmental cues that it could use, we might expect the species to evolve to use the most reliable cue. When organisms have very reliable cues, they can more accurately produce a phenotype that is well suited to the environment.
Response Speed and Reversibility
Phenotypically plastic traits respond to changes in the environment at different rates. Some of the trait changes are irreversible. The most rapid responses are typically behavioral traits, which can be altered in seconds. For example, most prey rapidly respond to a predator’s pursuit; often it takes less than 1 second for the prey to flee. Physiological plasticity, which is an environmentally induced change in an individual’s physiology—sometimes referred to as acclimation—can also be relatively rapid. Consider the time it takes humans to acclimate to the low-oxygen conditions that are caused by lower air pressures at high altitudes, or the time required for human skin to tan. Both of these physiological changes can be accomplished in just a few days. In contrast to these behavioral and physiological changes, changes in morphology—including changes in body shape and the size of internal organs—and changes in life history—including time to sexual maturity and number of progeny produced—can take weeks, months, or years. Acclimation An environmentally induced change in an individual’s physiology. Differences in response speed have implications for the reversibility of the induced traits. Behavioral traits that are induced by a change in the environment typically can be rapidly reversed if the environment reverts to its original condition. For example, an animal can quickly and easily adjust its food intake as food conditions change over time. Induced changes in morphology and life history are more difficult to reverse. For many organisms such as plants, changes in morphology are difficult or impossible to undo. For example, plants commonly respond to low light conditions by growing taller in an attempt to rise above neighboring plants that are casting shade. If the environment suddenly becomes sunny, a plant cannot make itself shorter. Even less reversible are life history decisions such as those related to the timing of reproductive maturity and the number of offspring produced. Once sexual maturity has been achieved, an organism cannot become sexually immature, although it can refrain from reproducing. The differences in the speed of phenotypic changes, and the ability to reverse phenotypic changes, influence which traits are favored by natural selection. When environments fluctuate rapidly relative to the length of an
individual’s lifetime, selection should favor plastic behavioral and physiological traits because these traits can often respond rapidly and reverse rapidly. When environments fluctuate more slowly, selection can favor many more types of traits, including morphological and life history traits that are slow to respond and are often much less reversible.
Concept Check
1. What is the distinction between weather versus climate? 2. How do phenotypic trade-offs favor the evolution of phenotypic plasticity? 3. Why do phenotypically plastic responses depend on reliable environmental cues?

#### 4.2 Many Organisms Have Evolved Adaptations to Variation in Enemies, Competitors, and Mates
4.2 Many organisms have evolved adaptations to variation in enemies, competitors, and mates Many types of environmental variation can induce phenotypic plasticity. Among biotic environments, three of the best-studied types of environmental variation involve the occurrence of enemies, competitors, and mates.
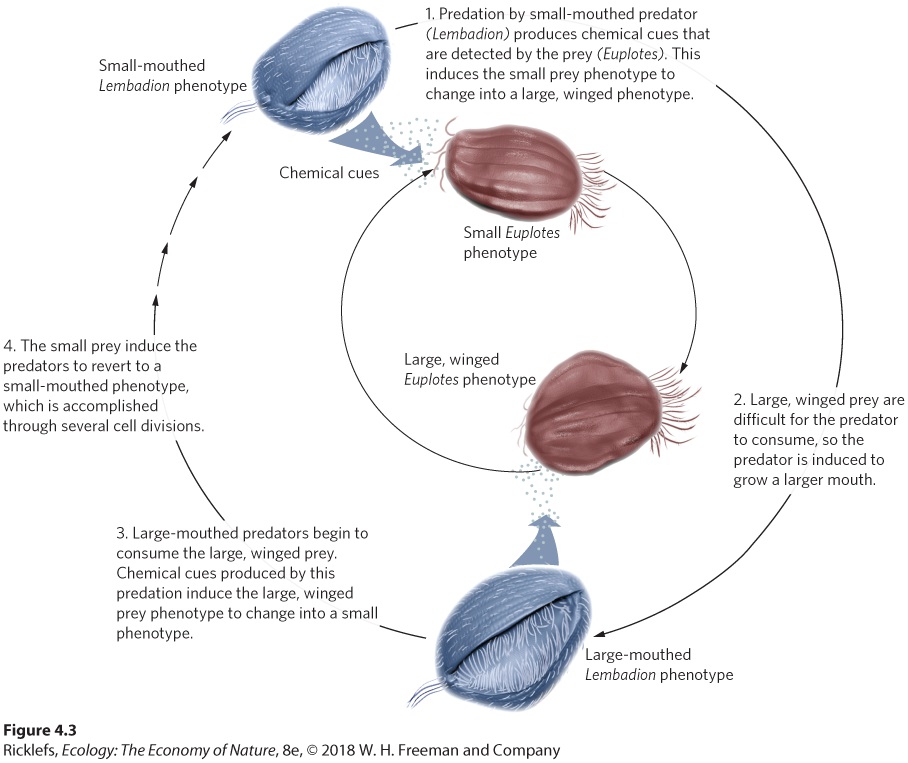
Because enemies—including predators, herbivores, parasites, and pathogens —pose a major risk to organisms that are consumed, we would expect that many organisms have evolved defenses against their enemies. Like the treefrog tadpoles, many aquatic animals including fish, salamanders, insects, zooplankton, and protists alter their growth and change shape in response to predators in their environment. These changes might improve a prey’s ability to escape, make the prey difficult to fit in the predator’s mouth, or deter consumption by producing sharp spines. Ciliates in the genus Euplotes are tiny protists that live in lakes and streams. As you can see in Figure 4.3, these tiny organisms can sense chemical cues emitted by predators and, within hours, respond by growing “wings” and numerous other projections that make them up to 60 percent larger. The larger size makes it difficult for a predator to fit Euplotes into its mouth, so the winged phenotype suffers less predation. However, because of the considerable amount of energy required to grow these projections, this phenotype takes 20 percent more time to develop. Thus, in a predator-free environment, the superior strategy for Euplotes is to have the nonwinged phenotype.
Figure 4.3 Inducible defenses and offenses. The ciliate Euplotes serves as prey for a larger ciliate, Lembadion. When Euplotes detects the predatory Lembadion in the water via chemical cues, it changes shape from an uninduced small phenotype to a predator-induced phenotype that has large “wings” and other projections that make it too large to fit in the predator’s mouth. When the predator begins to encounter the large, winged prey, mechanical cues induce it to increase the size of its mouth. This larger mouth can capture the large, winged prey but it is poor at capturing the small prey, which develop in response to chemical cues emitted by large-mouthed predators. As small prey become more abundant, the predatory Lembadion rapidly divides several times to once again have a small mouth. After H. W. Kuhlmann and K. Heckmann, Hydrobiologica 284 (1994): 219–227; M. Kopp and R. Tollrian, Ecology 84 (2003): 641–651; M. Kopp and R. Tollrian, Ecology Letters 6 (2003): 742–748. When prey make themselves more difficult to catch or consume, how might predators respond? It turns out that predators can have plastic abilities as well. For example, when the predatory ciliate Lembadion is surrounded by the predator-induced, winged Euplotes, Lembadion grows a larger body with a bigger mouth that can engulf the winged prey. In doing so, Lembadion benefits by consuming the winged prey. However, once most of the winged prey have been consumed and only small prey remain, the large Lembadion is poorly suited to eat the small prey. As a result, the large Lembadion experiences lower fitness than the small Lembadion. When this occurs, a large Lembadion can go through several cell divisions and revert back to

having a smaller phenotype with the smaller mouth. Prey also use behavioral defenses against predators. They may move away from areas containing predators or become less active to avoid detection. Some animals also congregate in refuges, which are locations that are safe from predation. For example, newly hatched fish take refuge in the dense weeds along a lake shoreline to hide from large predatory fish. Although these behaviors usually reduce the risk of predation, increased safety comes at a price. When prey become less active or congregate in refuges, they spend less time feeding. Also, the supply of food in and around crowded refuges can be depleted quickly. As a result, behavioral defenses commonly come at the cost of slower growth, development, or reproduction. In the absence of predators, prey become more active and leave the refuges to find more food; this change in behavior allows more rapid growth. Plants also have the ability to respond to the presence of organisms that consume them. The Virginia pepperweed, a member of the mustard family, is eaten by several species of herbivores, including caterpillars and aphids. As
Figure 4.4a shows, when an herbivore chews on the leaves of pepperweed, the plant rapidly develops extra leaf hairs, called trichomes, that make it harder for the leaves to be consumed. It also increases its production of glucosinolates, a group of chemicals that gives mustard its strong flavor and functions as a natural insecticide. If previously attacked plants and nonattacked plants are subsequently placed in a garden, the attacked plants, which have more trichomes and more glucosinolates, attract fewer herbivores and exhibit better survival, as shown in Figure 4.4b.
Figure 4.4 Herbivore-induced responses by plants. (a) Virginia pepperweed (Lepidium virginicum) responds to herbivore attacks by growing more leaf hairs, known as trichomes, and more types of defensive chemicals, called glucosinolates. (b) Plants attacked by herbivores in the past developed more trichomes and glucosinolates and have fewer aphids on them than plants that were not previously attacked by herbivores. Error bars are standard errors. After A. Agrawal, Benefits and costs of induced plant defense for Lepidium virginicum
Competition for Scarce Resources
Most organisms face the challenge of scarce resources, which leads to competition. However, the intensity of competition varies across and within habitats. As a result, organisms have evolved a variety of phenotypically plastic strategies for high and low competition. As we would expect, responses to high competition often exhibit phenotypic trade-offs that favor the evolution of phenotypically plastic responses. Jewelweed (Impatiens capensis), a flowering plant with beautiful orange flowers, is found in moist habitats throughout much of North America (Figure 4.5). In nature, it can grow in clumps that are either very sparse or very dense. The range of densities has an effect on the intensity of competition among the plants for sunlight. When jewelweed is shaded by other plants, it responds by elongating its stems, which allows the plant to become taller and to rise above the competing plants. If competition for light causes the jewelweed to grow taller, we might wonder why the plant doesn’t always grow tall. Researchers have discovered that elongated jewelweeds experience greater fitness in high-competition environments, whereas short jewelweeds experience greater fitness in low-competition environments. Although the researchers were not able to identify the reasons why, it is clear that different phenotypes perform better in different environments, so a plastic phenotype is an effective way to gain high fitness when the intensity of competition varies over time and space.
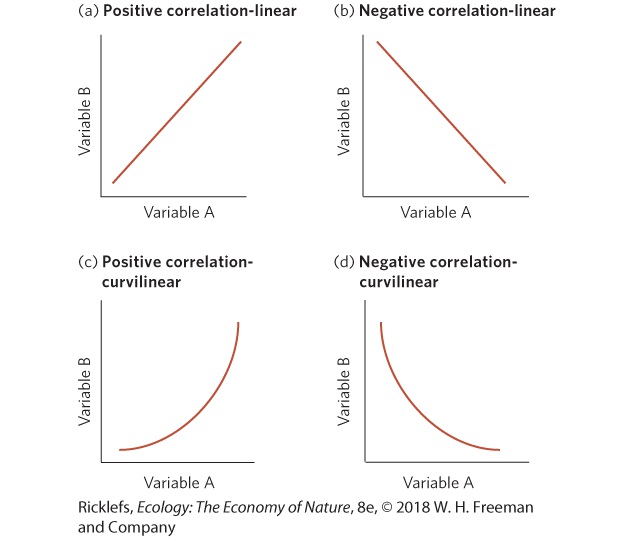
Figure 4.5 Jewelweed. When jewelweed is shaded by other plants, it elongates its stems to become taller and rise above the competing plants. Elongated jewelweeds experience greater fitness in high-competition environments, whereas short jewelweeds experience greater fitness in low-competition environments. Animals also respond to competition in a number of fascinating ways. For example, we might expect animals to spend more time searching for food when it is rare than when it is abundant. However, a second option is to find ways to extract more nutrients from the food that is available. One way to do this is to alter the size of the digestive tract, as we saw in the example of gray treefrogs, which respond to competition by increasing the length of their intestine. When the intestine is longer, food spends more time traveling through it and the organism can extract more nutrients from it. A striking example of a plastic response to variation in food availability can be found in the digestive tracts of snakes. The Burmese python (Python bivittatus), for example, can consume a large rodent that is 25 percent of its own body weight, but it may find such a prey item only once a month (Figure 4.6). Between meals, the snake’s stomach and intestine shrink a great deal, which reduces its weight and the associated carrying costs. Once a prey is consumed, however, the python can enlarge its cells and double the length of its digestive tract in just 24 hours. This dramatically increases the surface
area of the intestine, which allows the snake to absorb more energy from the digested prey. The snake also sends 10 times more blood to the intestines to assist in the absorption of nutrients into the bloodstream. To handle this increase in blood flow, the snake increases the size of its heart by 40 percent, a remarkable achievement given that it occurs in 2 days.
Figure 4.6 The plasticity of a python. (a) A Burmese python can consume large prey, but may do so only once a month. (b) A fasting animal has a shrunken intestine. (c) Within 2 days of eating, the intestine doubles in length and swells in diameter. (Note that DPF means days post feeding.) (d) Ten days later, digestion is complete and the intestine shrinks again.
When mates are rare, reproduction can be a challenge. Consider the situation of flowering plants that depend on pollinators to deliver the pollen that contain a male’s sperm. In this situation, the probability of being pollinated can be highly variable, which means that the probability of finding a mate is also highly variable. There is a solution for flowering plants that are hermaphrodites, which are individuals that produce both male and female gametes. Such individuals are often self-compatible, that is, they are capable of fertilizing their eggs with their own sperm. This process, known as selffertilization, often carries the potential cost of inbreeding depression, in which offspring can experience reduced fitness when deleterious alleles are inherited from both the egg and the sperm. Because mate availability can be quite variable and self-fertilization can have a substantial cost, the organism must develop one of two life history choices: wait to find a mate and enjoy higher fitness, or self-fertilize. In some cases, this choice is made at the level of the entire organism. In other cases, as in many hermaphroditic species of flowering plants, the choice is made by each flower. If a flower is not pollinated within a given period of time, the flower will self-fertilize. Although self-fertilizing may result in lower fitness than breeding with a mate, low fitness is better than the zero fitness the organism would obtain by avoiding self-fertilization altogether. Hermaphrodite An individual that produces both male and female gametes. Inbreeding depression The decrease in fitness caused by matings between close relatives due to offspring inheriting deleterious alleles from both the egg and the sperm. Reproductive adaptations to variation in mate availability have been studied in a variety of hermaphroditic plants and animals. For example, the common pond snail (Physa acuta) is a hermaphrodite that experiences large variation in population sizes, which means that it also experiences large variation in the availability of mates. When potential mates are abundant, a snail will normally mate with another individual; when potential mates are rare, it can fertilize its eggs with its own sperm. In an experiment designed to measure the fitness effects of the two alternative strategies, researchers assigned snails to one of two groups: snails living with mates or snails living
without mates. The researchers then observed the time it took for each group to begin reproduction and the total number of eggs that each group of snails laid. As you can see in Figure 4.7a, snails living without mates delayed their reproduction for 2 weeks before they used the alternative strategy of selffertilization. As predicted, this choice came at a cost. Looking at Figure 4.7b, we see that self-fertilizing snails laid many fewer eggs than snails that had mates available. However, the self-fertilizing snails did gain some fitness, which is better than forgoing reproduction. The good news for the pond snail, and many other hermaphroditic organisms, is that by evolving multiple strategies they can experience higher fitness over time than would be possible without a phenotypically plastic strategy.
Figure 4.7 nail responses to variation in mates. In the common pond snail, mate availability can vary a great deal. (a) Snails raised without mates wait nearly 2 weeks before self-fertilizing their eggs. (b) Snails without mates produce fewer progeny compared to snails with mates, but this strategy is more fit than forgoing all reproduction. Error bars are standard errors. After A. Tsitrone et al., Delayed selfing and resource reallocations in relation to mate
availability in the freshwater snail Physa acuta, American Naturalist 162 (2003): 474–488.
Concept Check
1. How might predators and prey both evolve phenotypically plastic strategies to improve their fitness? 2. How is the phenotypic plasticity of intestines an adaptation to variable food availability? 3. If self-fertilization causes lower fitness than cross-fertilization, under what environmental conditions would self-fertilization be the superior strategy?
#### 4.3 Many Organisms Have Evolved Adaptations to Variable Abiotic Conditions
4.3 Many organisms have evolved adaptations to variable abiotic conditions We have seen that variation in biotic conditions, including enemies, competitors, and mates, can be quite high. Abiotic conditions, including temperature, water availability, salinity, and oxygen, also vary. Faced with this abiotic variation, many species have evolved phenotypically plastic traits that allow them to improve their fitness.
Temperature
Organisms have evolved a number of plastic responses to temperature variation. As we saw in Chapter 2, isozymes in rainbow trout allow the fish to have proper nerve transmission in cold winter water and warm summer water. Isozymes are actually a form of phenotypic plasticity with a rapid response time, in some cases a matter of hours or days. For example, goldfish (Carassius auratus) can be held at either 5 °C or 25 °C for a few days and then tested to determine how fast they can swim in a variety of temperatures. As shown in Figure 4.8, fish acclimated to 5 °C swim faster at low temperatures but swim poorly at high temperatures. In contrast, fish acclimated to 25 °C swim faster at warm temperatures but swim poorly at low temperatures. This demonstrates that goldfish can adjust their physiology to maintain relatively high swimming speeds across different environmental temperatures.
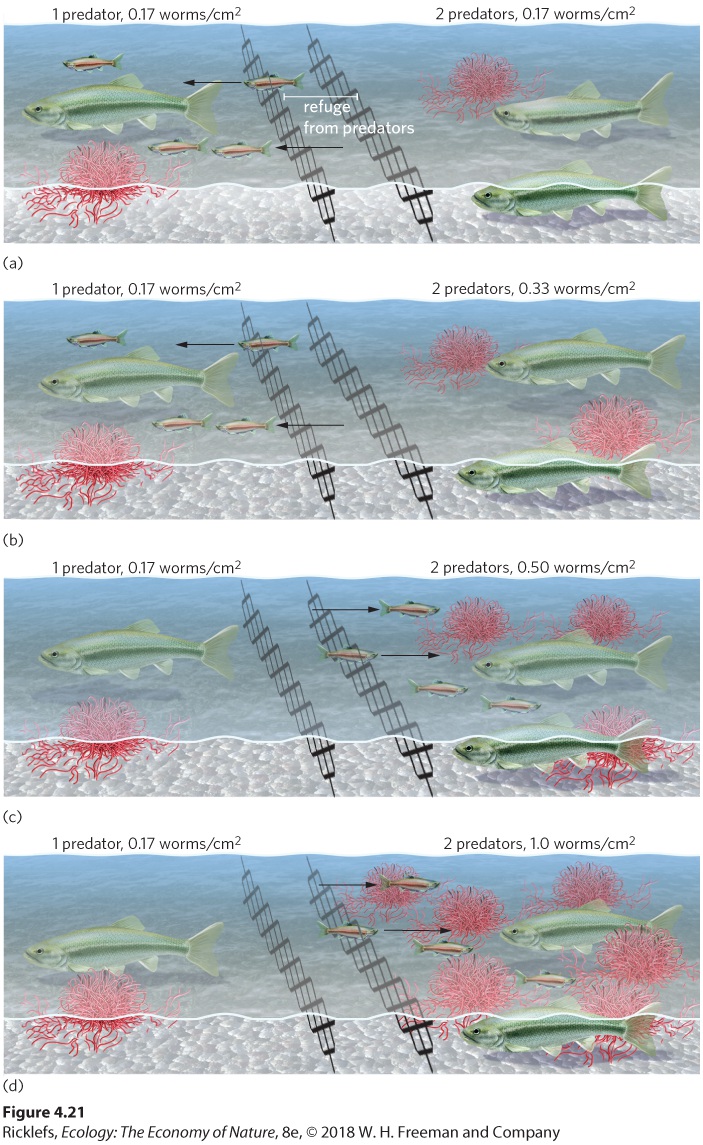
Figure 4.8 Acclimation to different temperatures. Goldfish raised at low temperatures swim faster in cold water and more slowly in warmer water. Individuals raised at high temperatures swim faster in warmer water than in cold water. After F. E. J. Fry and J. S. Hart, Cruising speed of goldfish in relation to water temperature, Journal of the Fisheries Research Board of Canada 7 (1948): 169–174. Many animals respond to changing temperatures by moving to habitats with more favorable temperatures. Migrating birds present an extreme example; they fly every autumn to warmer latitudes. Not all animals travel a long distance. Some move to a specific location within a habitat, called a microhabitat, that contains more favorable abiotic conditions. Microhabitat A specific location within a habitat that typically differs in environmental conditions from other parts of the habitat. The use of microhabitats can be illustrated by the daily behavior cycle of the desert iguana (Dipsosaurus dorsalis), shown in Figure 4.9. Although the desert iguana cannot regulate its temperature by generating heat metabolically, it can take advantage of sunny and shady microhabitats to alter its temperature. The iguana has a preferred temperature range of 39 °C to 43 °C. In the southwestern United States, the air temperature can reach 45 °C. As temperatures rise during the day, the lizard first moves to the shade of
plants or rocks and then to a cooler underground burrow. If temperatures begin to cool, the iguana can move out of its burrow and bask in the sun to raise its temperature to its preferred range. Such behavioral plasticity allows the iguana to remain within its preferred temperature range for a greater proportion of the day. In effect, the plasticity of the iguana’s behavior permits homeostasis in its body temperature.
Figure 4.9 Selecting microhabitats. The desert iguana regulates its body temperature by selecting microhabitats that contain favorable abiotic conditions. When the lizard is cool, it can bask in the sun to increase its internal temperature. As the temperature becomes hot during the day, the lizard can seek shade or move into a burrow to lower its temperature.
Water Availability
When faced with changes in water availability, most animals can move among different microhabitats. However, plants are typically rooted in a single location and therefore face a tremendous challenge in locating water. As a result, plants have a number of phenotypically plastic adaptations for coping with water variability. Closing the stomata is one of the most common adaptations. As we saw in Chapter 3, when water is plentiful, the guard cells in a plant’s leaves open and transpiration occurs through the stomata. However, this process causes water loss. When water is scarce, these cells change shape and the stomata close to conserve water. In this way, the plant can transpire when water is abundant but stop transpiring when water is scarce. Other plant strategies in response to a lack of water are even more dramatic. For example, plants living on coastal dunes in Europe commonly

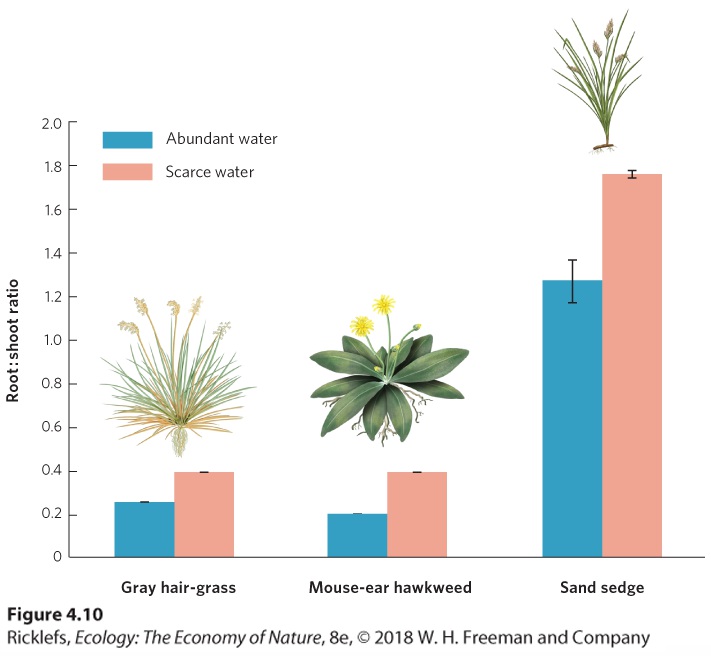
experience drought because water drains quickly through the sandy substrate. Three common plants of these coastal dunes—gray hair-grass (Corynephorus canescens), mouse-ear hawkweed (Hieracium pilosella), and sand sedge (Carex arenaria)—demonstrate how plants adjust their relative allocations of energy and materials to grow either roots or shoots. To demonstrate this phenomenon, researchers grew each of the three plants under conditions of abundant versus scarce water. After 5 months of growth, the researchers measured the ratio of root growth to shoot growth and the results are shown in Figure 4.10. When water was abundant, the plants devoted more energy to the growth of shoots, which function primarily to photosynthesize. When water was scarce, the plants devoted more energy to the growth of roots, which expanded their ability to capture what little water was available. Given that these plants experience variation in water availability, it is clear that no single allocation strategy would be as beneficial as the phenotypically plastic strategy that they exhibit.
Figure 4.10 Morphological plasticity in response to water. Plants living under scarce water conditions have a higher root-to-shoot ratio. By devoting more growth into the roots than
into the shoots, these plants can better obtain water when it is scarce. Error bars are standard errors. After A. Weigelt et al., Competition among three dune species: The impact of water availability on below-ground processes, Plant Ecology 176 (2005): 57–68.
Salinity
In Chapter 2, we learned that freshwater and saltwater organisms have evolved numerous adaptations to handle their aquatic environments. However, some organisms live in aquatic environments characterized by solute concentrations that fluctuate widely over short periods of time. To survive, these organisms must have the ability to make rapid physiological adjustments. For example, the copepod Tigriopus lives along rocky Pacific coasts in pools that receive sea water infrequently, from the splash of high waves (Figure 4.11). As water in the pools evaporates, the salt concentration rises to high levels, but a heavy rainfall can lower the salt concentration—a rapid reversal of environmental conditions for the copepod.
Figure 4.11 Adaptations to fluctuating salt environments. (a) Pools of water on the rocky coasts of La Jolla Cove in San Diego, California, are filled with a mixture of rain water and sea water from waves. Evaporation can cause high concentrations of salts and other solutes. (b) The tiny copepod Tigriopus, shown here carrying eggs, is able to handle the widely fluctuating solute concentrations in its environment by adjusting its solute concentrations through the production of amino acids. Like sharks and rays, Tigriopus manages its water balance by changing the
osmotic potential of its body fluids. When the salt concentration in a pool is high, individuals synthesize large quantities of certain amino acids such as alanine and proline. These small molecules increase the osmotic potential of the body fluids to match that of the environment without the deleterious physiological consequences that come with high levels of salts or urea. However, this response to excess salts in the environment is costly in terms of the energy it requires. When individual Tigriopus are switched from 50 percent sea water to 100 percent sea water, the respiration rate of the copepods initially declines, owing to salt stress, and then increases as they synthesize alanine and proline to restore their water balance. When switched from 100 percent sea water back to 50 percent sea water, the copepods’ respiration rate immediately increases as they rapidly degrade and metabolize excess free amino acids to reduce their osmotic difference to be more in line with their new environment.
If you have ever been to a location at a high elevation, you probably experienced the challenge of low oxygen. As you continue to rise above sea level, the air pressure becomes lower, which reduces the amount of available oxygen. At thousands of meters above sea level, breathing becomes labored and physical activity is very difficult. Animals that travel up and down mountains as part of their daily or seasonal movements, for example, the llamas of South America, are able to adjust their physiology to this variation in oxygen concentration. Similarly, human mountaineers, who tackle Mount Everest, experience an extreme oxygen challenge for which their bodies can only partially adjust. At its peak, the mountain is 8,848 m above sea level and the oxygen pressure is only one-third of that found at sea level. To acclimate to the low oxygen conditions, climbers stop periodically for several days along the way. Initial changes include more rapid breathing and an increased heart rate. After a week or two, additional changes improve the body’s ability to carry oxygen, including an increase in the number of red blood cells and an increased concentration of hemoglobin in the red blood cells. At the highest altitudes, where humans rarely spend much time, the body is not able to fully adjust to such low oxygen concentrations; this has contributed to the deaths of many climbers at high altitudes. When climbers later return to low altitudes, their physiological changes slowly revert to their original state.
Concept Check
1. How does the function of isozymes represent an example of phenotypic plasticity? 2. Why do plants alter their root-to-shoot ratios? 3. What are three adaptations humans undergo in response to decreased oxygen at high elevations?
#### 4.4 Migration, Storage, and Dormancy Are Strategies to Survive Extreme Environmental Variation
4.4 Migration, storage, and dormancy are strategies to survive extreme environmental variation In many parts of the world, extremes of temperature, drought, darkness, and other adverse conditions are so severe that individuals either cannot change enough to maintain their normal activities, or the change required would not be worth the cost. Under such conditions, organisms resort to a number of extreme phenotypically plastic responses. These include migration, storage, and dormancy.
Migration
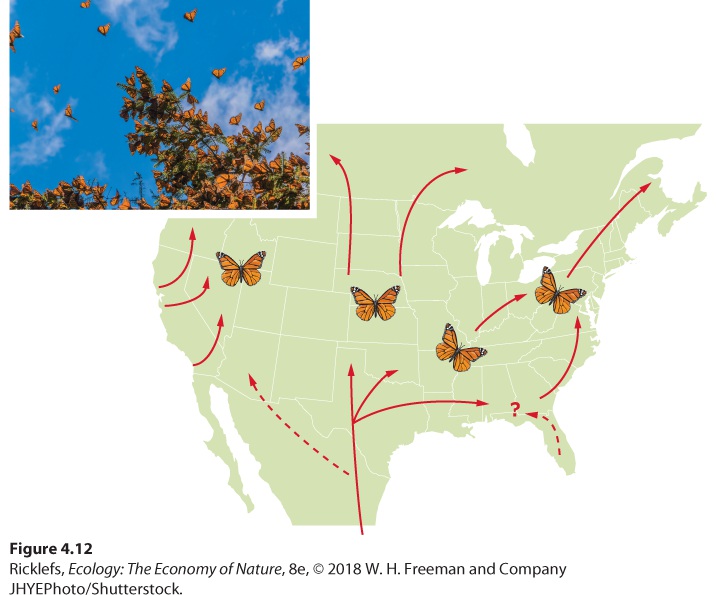
Migration is the seasonal movement of animals from one region to another. In this case, the phenotype is the behavior of living in a particular location and the plasticity is displayed in the act of migrating, which allows the animal to express the alternative phenotypes of living in multiple locations. Each fall, hundreds of species of land birds leave temperate and Arctic North America, Europe, and Asia for the south, in anticipation of cold winter weather and dwindling food supplies. In east Africa, many large herbivores, such as wildebeests (Connochaetes taurinus), migrate long distances, following the geographic pattern of seasonal rainfall and fresh vegetation. Some insects also migrate. Monarch butterflies offer a fascinating example of insect migration, as shown in Figure 4.12. The adult butterflies living in the northern United States and southern Canada migrate to wintering areas in the southern United States and Mexico. Here, they hibernate through the winter and then begin heading back north. On their way north, they breed and produce a second generation of butterflies. This second generation completes the migration back to the summer breeding areas. In all cases, the decision to migrate is a plastic behavior in response to environmental changes including lower temperature and shorter day length.
Figure 4.12 Monarch butterfly migration. The monarch butterfly population follows a variety of migration routes from its overwintering areas in the southern United States and Mexico to more northern regions where it spends the summer. Migration The seasonal movement of animals from one region to another. Some migratory movements occur in response to reductions in local food supplies. For example, consider locusts, which are species of short-horned grasshoppers. Locust migrations occur when the insects leave an area where they have a large population and a depleted food supply (Figure 4.13). The migrations can include billions of locusts and, when they pass through an area, these herbivores can cause extensive crop damage over wide areas. As a result, their arrival can cause major damage to human food supplies. This migratory behavior in locusts is the result of a number of behavioral changes. In sparse populations, adult locusts are solitary and sedentary. In dense populations, however, frequent contact with other locusts stimulates young individuals to develop gregarious, highly mobile behavior, which can grow into a mass migration.
Figure 4.13 Locust migration. When locusts experience a reduced food supply, the normally solitary grasshoppers produce offspring that are highly gregarious and mobile. These offspring move across the landscape in huge swarms and cause major damage by consuming crops, such as this example in North Africa.
Where environmental variation shifts the food supply from feast to famine and migration is not a possibility, storing resources can be an adaptive strategy. For example, during infrequent rainy periods, desert cacti swell with water stored in their succulent stems, as discussed in Chapter 3. In habitats that frequently burn—such as the chaparral of southern California—perennial plants store food reserves in fire-resistant root crowns, shown in Figure 4.14. The surviving root crowns send up new shoots shortly after a fire has passed.
Figure 4.14 Energy storage. (a) Some species of plants, including one known as death camas due to the toxic chemicals it produces, possess root crowns that are resistant to fire and store large amounts of energy. (b) This stored energy can be used by the plant to quickly re-sprout after a fire. When fire burns an area, such as this site in Angeles National Forest, California, the death camas plant can quickly re-sprout. Many temperate and Arctic animals accumulate fat during mild weather as a reserve of energy for periods of harsher weather when snow and ice make food inaccessible. However, fat reserves can make an animal slower and less agile, and therefore more likely to be caught by predators. One way to avoid this problem is to store the food before consuming it. Some mammals and birds that are active during the winter—for example, beavers, squirrels, pikas, acorn woodpeckers, and jays—cache food supplies underground or under the bark of trees for later retrieval. These caches can be large enough to sustain
Dormancy
Environments sometimes become so cold, dry, or depleted of nutrients that organisms can no longer function normally. Some species that do not migrate have evolved a strategy of dormancy, a condition in which they dramatically reduce their metabolic processes. One of the most obvious forms of dormancy occurs when many temperate and Arctic trees shed their leaves in the fall before the onset of winter frost and long nights. Similarly, many tropical and subtropical trees shed their leaves during seasonal periods of drought. Plant seeds and spores of bacteria and fungi also exhibit dormancy. Indeed, there are many cases in which researchers have sprouted seeds recovered from archeological excavations where the seeds have been dormant for hundreds of years. Whereas many plants experience dormancy, in animals we distinguish among four specific types of dormancy: diapause, hibernation, torpor, and aestivation. Dormancy A condition in which organisms dramatically reduce their metabolic processes. In most species, worsening environmental conditions are anticipated and individuals proceed through a series of physiological changes that prepare them for a partial or complete physiological shutdown. Diapause is a type of dormancy that is common in insects in response to unfavorable environmental conditions. For example, as winter approaches, insects shut down their metabolism to barely detectable levels. In doing so, the insects need to either reduce the quantity of water in their bodies or chemically bind the water in their bodies to prevent freezing. Similarly, insects facing drought conditions can enter a summer diapause by dehydrating themselves. They either tolerate the desiccated condition of their bodies or secrete an impermeable outer covering to prevent further drying. Diapause A type of dormancy in insects that is associated with a period of unfavorable environmental conditions. During hibernation, a less extreme type of dormancy that occurs in
mammals, animals reduce the energetic costs of being active by lowering their heart rate and decreasing their body temperature. Many mammals, including ground squirrels and bats, hibernate during seasons when they are unable to find food (Figure 4.15). Prior to hibernation, the animal consumes enough food to produce a thick layer of fat that provides the energy necessary to survive the hibernation period without eating.
Figure 4.15 Hibernation. Some mammals, such as this chipmunk, spend the winter in a deep sleep. During this time, the chipmunk slows its breathing and heart rate and reduces its temperature close to 0 °C to help conserve energy through the winter. Hibernation A type of dormancy that occurs in mammals in which individuals reduce the energetic costs of being active by lowering their heart rate and decreasing their body temperature. Some types of dormancy occur over short periods of time to deal with cold temperatures. At low air temperatures, some birds and mammals are not able to maintain a high body temperature. Doing so would require that the animal burn up its stored energy faster than it can consume and digest the food needed to replace the energy lost in creating body heat. In this situation, the animal might go into a brief period of dormancy, known as torpor, during which the animal reduces its activity and its body temperature decreases. During torpor, reduced activity and reduced body temperature help conserve energy. Torpor may last as little as a few hours or extend for a few days and
is a voluntary, reversible condition. Torpor A brief period of dormancy that occurs in birds and mammals in which individuals reduce their activity and their body temperature. Many small birds and mammals use torpor. Hummingbirds, a group of tiny birds with body lengths of 7.5 to 13 cm, provide a good example. These small birds have a high surface-to-volume ratio. This causes a rapid loss of heat across the body surface relative to the volume of body that can produce heat. As the air temperature declines, hummingbirds must metabolize increasing amounts of energy to maintain a resting body temperature near 40 °C. Figure 4.16 shows the relationship between air temperature and energy needs for the West Indian hummingbird (Eulampis jugularis). Its metabolic rate is measured as the amount of oxygen consumed as it converts its stored energy into body heat. When the bird enters torpor, it reduces its resting body temperature by 18 °C to 20 °C. If the air temperature drops to 20 °C, torpor allows the bird to stop burning extra energy to generate body heat and thereby conserve its energy reserves. Torpor does not mean that the animal ceases to regulate its body temperature; it merely changes the set point on its thermostat to reduce the difference between ambient and body temperature, and thereby reduces the energy expenditure needed to maintain its temperature at the set point.
Figure 4.16 Torpor. The tiny West Indian hummingbird has a high surface-to-volume ratio, causing it to lose much of the heat it generates during periods of cold air temperatures. To save energy, measured as the amount of energy consumed to fuel the metabolism of its food, the bird enters torpor when it is resting at night. Data from F. R. Hainsworth and L. L. Wolf, Regulation of oxygen consumption and body temperature during torpor in a hummingbird, Eulampis jugularis, Science 168 (1970): 368–369.
Analyzing Ecology
Correlations In the hummingbird example, we saw that as air temperature decreased, the bird’s consumption of oxygen increased (Figure 4.16). This is an example of a statistical correlation. A correlation is a statistical description of how one variable changes in relation to another variable. For example, at the beginning of this chapter, we observed that when tadpoles faced more dangerous predators, they exhibited larger phenotypic changes. Correlation
A statistical description of how one variable changes in relation to another variable. Two variables can be related to each other in a variety of ways, shown in the graphs below. A positive correlation (a) indicates that as one variable increases in value, the second variable also increases. A negative correlation (b) indicates that as one variable increases in value, the second variable decreases. Such increases or decreases may be linear, meaning that the data fall along a straight line, as shown in the example. They may also be curvilinear, as shown in (c) and (d), meaning that the data follow a curved line. Correlations do not tell us anything about causation. For example, the positive correlation between variable A and variable B in panel (a) might occur because a change in variable A causes the change in variable B. But it could also be that a change in variable B causes the change in variable A. Alternatively, a third, unmeasured variable might cause both A and B to change. Consider the case of humans climbing Mount Everest. As a person ascends the mountain over several weeks, the temperature continually decreases and the climber’s efficiency in obtaining oxygen from the air increases. This is a correlation, but it is not causation. Lower temperatures do not cause a person to acquire oxygen more efficiently. We know that a third variable, the declining oxygen pressure at high altitudes, is the real cause of the increased efficiency in climbers obtaining oxygen from the air during the time that they spend climbing. YOUR TURN For the following set of data, construct a graph that demonstrates the relationship between the number of predators in a pond and the activity level of gray treefrog tadpoles (i.e., the proportion of their time spent moving). After constructing the graph, determine (a) whether it is a positive or negative correlation and (b) whether it is linear or curvilinear. Number of predators Activity level
Correlations. (a) When an increase in one variable is associated with a linear increase in another variable, it is a positive linear correlation. (b) When an increase in one variable is associated with a linear decrease in another variable, it is a negative linear correlation. Correlations also can be (c) positive and curvilinear or (d) negative and curvilinear. A fourth type of dormancy is aestivation, which is the shutting down of metabolic processes during the summer in response to hot or dry conditions. Well-known aestivating animals include snails, desert tortoises, and crocodiles. Aestivation The shutting down of metabolic processes during the summer in response to hot or dry conditions. By whatever mechanism it occurs, dormancy reduces exchange between organisms and their environments, enabling animals and plants to survive in unfavorable conditions.
Adaptations to Prevent Freezing
In Chapter 2, we discussed the adaptations that some aquatic animals have, to avoid the damaging effects that freezing can cause to their tissues. In a similar fashion, some terrestrial animals survive very cold weather on land by using special adaptations, including the production of antifreeze chemicals that prevent or control the formation of ice crystals. For example, in cold northern climates, many insects spend the winter living just beneath the bark of trees. This bark helps insulate insects to some degree, but they still experience temperatures below the freezing point of water. Similarly, many species of amphibians spend the winter buried a short distance below the soil surface. These animals can freeze solid underground in a state that requires very little metabolic activity (Figure 4.17). Two strategies—the use of antifreeze chemicals and forming ice crystals between rather than within cells —help insects and amphibians avoid tissue damage. As temperatures warm in the spring, the animals slowly thaw out and resume their normal activities.
Figure 4.17 Freezing solid. Many species of frogs, such as this gray treefrog, can become dormant during cold winter months and then thaw out when spring arrives. By producing antifreeze and forming ice crystals between their cells rather than within their cells, the frogs can dramatically reduce their metabolic activity through the winter. Migration, storage, and dormancy represent phenotypically plastic
strategies that allow organisms to cope with extreme changes in their environment. Such behavioral and physiological flexibility provides a substantial selective advantage.
Concept Check
1. Explain how migration and dormancy are both examples of phenotypic plasticity. 2. Why would it be adaptive for subtropical trees to shed their leaves during seasonal periods of drought? 3. Explain the differences between the four types of dormancy in animals.
#### 4.5 Variation in Food Quality and Quantity Is the Basis of Optimal Foraging Theory
4.5 Variation in food quality and quantity is the basis of optimal foraging theory As we discussed earlier in this chapter, animal behavior is a form of phenotypic plasticity. Foraging is one of many important behaviors for animals and a great deal of research has been conducted on how animals search for food and select from a diversity of food choices. Because abundance of food items varies over space and time, no single feeding strategy can maximize an animal’s fitness. Hence, feeding decisions represent phenotypically plastic behavior because different feeding strategies represent different behavioral phenotypes. These phenotypes are induced by unique environmental cues and each alternative feeding behavior is well suited to a particular environment but not well suited to other environments. Therefore, the alternative behavioral phenotypes experience fitness trade-offs. Animals must determine where to forage, how long to feed in a certain habitat, and which types of food to eat. Ecologists predict how animals should make foraging decisions based on estimated costs and benefits of feeding in particular environmental situations. They then compare these predictions to observations of foraging animals to see which strategy provides the highest fitness. Although it would be ideal to measure costs and benefits in terms of survival and reproduction, these components of evolutionary fitness can be difficult to measure. Consequently, ecologists usually look at factors correlated with fitness, such as foraging efficiency. This is based on an assumption that animals able to gather more food in less time should be more successful at survival and reproduction. The idea that animals should strive for the best balance between the costs and benefits of different foraging strategies is known as optimal foraging theory. Optimal foraging theory A model describing foraging behavior that provides the best balance between the costs and benefits of different foraging strategies. Animals have four responses to food variation in space and time: central place foraging, risk-sensitive foraging, optimal diet composition, and diet mixing.
Central Place Foraging
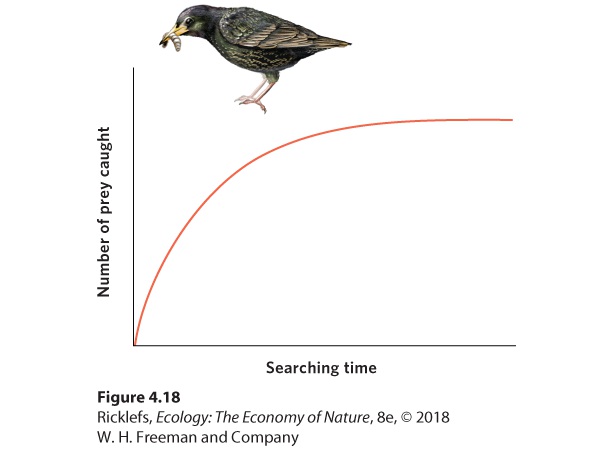
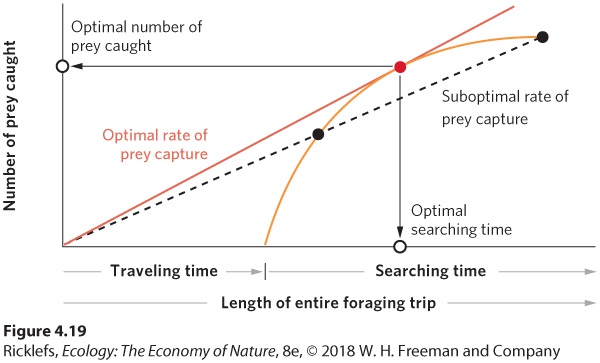
When birds feed their offspring in a nest, the chicks are tied to a single location, while the parents are free to search for food at a distance. This situation is known as central place foraging because acquired food is brought to a central place, such as a nest with young. As the parent ranges farther from the nest, it finds a greater amount of potential food sources. However, traveling a longer distance increases time, energy costs, and exposure to risk. The animal must choose the amount of time spent gathering food before returning to the nest as well as how much food to bring back with each trip. Central place foraging Foraging behavior in which acquired food is brought to a central place, such as a nest with young. Researchers have used these choices to investigate the feeding behavior of European starlings (Sturnus vulgaris). During the summer, starlings typically forage on lawns and pastures for the larvae of craneflies, called leatherjackets. Starlings feed by thrusting their bills into the soft ground and spreading their beak to expose the prey. When they are gathering food for their young, they hold captured leatherjackets in the base of their bill. Researchers predicted that as a starling continued to capture more leatherjackets and to hold them in its bill, grabbing the next one would become more difficult This is analogous to shopping for items in a grocery store without a cart or a basket; the more items you hold, the harder it is to add another. As a result, the number of prey caught by the starlings should slow down over time. The prediction, shown in Figure 4.18, was supported by the data. The shape of the curve shows that the rate of food gathering rises rapidly at first and then, as the starling fills its beak, begins to slow down. We say that the starling experiences diminishing benefits over time.
Figure 4.18 Diminishing benefits over time. The rate of food gathering for the European starling is rapid at first but, as time passes, it experiences diminishing benefits because the amount of prey gathered per unit of time decreases. The rate at which the parent bird brings food back to its offspring is a function of how much food it obtains and how much total time it takes to obtain the food. The total time it takes to obtain the food depends on the time needed to fly round-trip to the site that contains the food, known as the traveling time, plus the time spent obtaining the food once the bird arrives at the site, known as the searching time. Figure 4.19 shows a graphical model of how an animal should make decisions as an optimal forager. The diminishing benefits line (from Figure 4.18) is shown in orange. To this, we can add a fixed traveling time, which is the amount of time the bird needs to get to the feeding area. We can then draw a red line from the origin of the trip to intersect the benefits curve. If we draw the red line at the steepest slope that intersects the orange benefits curve, the two lines cross at the red dot on the figure. This intersection point—which is drawn tangent to the orange benefits curve—represents the highest rate of food capture the bird can obtain, including traveling time. If the starling expressed any alternative behavioral phenotypes—for example, if it stayed at the feeding site for longer or shorter periods indicated by the black dots in the figure—it would have a lower rate of food acquisition. You can see this lower rate of food acquisition as the
Figure 4.19 Optimal foraging. The optimal rate of foraging for an animal that leaves its nest to find food depends on the time needed to travel to a location that contains food and the time spent feeding once it has arrived. For a given benefits curve (the orange line), the optimal rate of prey capture is found by drawing a straight line from the origin of the trip tangent to the benefits curve. The point of tangency indicates the optimal time that the animal should spend searching and the optimal amount of food it should bring back. Spending more or less time feeding in the location, as indicated by black dots, results in suboptimal amounts of food obtained per unit of time. Given our understanding of how the starling should forage when the feeding location is at a fixed distance from its nest, how should the bird’s behavior change when the food source is closer or farther away? At sites that are farther away, the bird should spend more time searching for food and bring back more food to help offset the extra travel time. In contrast, as travel time decreases for sites that are closer to the nest, the bird should spend less time searching the site for food and bring back less food. Recall the example of the grocery store. If the store happened to be across the street from your house, you would probably make frequent trips, spend a short amount of time searching for food, and bring back a few items on each trip. If the store were an hour’s drive away, you would likely make fewer trips, spend a longer time searching for food, and bring back an armload of items on each trip. As with the starling, these decisions improve your efficiency in bringing back food. To what extent do organisms actually forage optimally? Researchers addressed this question in a clever experiment. They trained starlings to visit
feeding tables that offered mealworms through a plastic tube at precisely timed intervals. A starling would arrive at the table, pick up the first mealworm, and then wait for the next mealworm to be delivered. Each successive mealworm was presented at progressively longer intervals, mimicking the longer intervals at which a starling would catch leatherjackets as its beak became increasingly full. The researchers set up feeding tables at different distances from the starling nests and observed how many mealworms a starling picked up before it departed back to its nest. The predicted number of prey brought back to the bird’s nest, shown as a blue line in Figure 4.20, agrees with the actual number observed in the experiment, represented by the red data points.
Figure 4.20 Predicted versus observed prey capture for a central-place forager. Based on an optimal foraging model, researchers predicted that longer travel times would cause starlings to return to their nests with a larger number of mealworms. The researchers offered mealworms to starlings on tables that were located at different distances from their nests. The observed number of mealworms brought back to the nests show agreement with the predictions. After A. Kacelnik, Central place foraging in starlings (Sturnus vulgaris), I. Patch residence time, Journal of Animal Ecology 53 (1984): 283–299.
Risk-Sensitive Foraging
Our predictions of how animals should forage have assumed that animals are
simply maximizing their rate of energy gain. However, most animals have other considerations, including predators. Because the act of feeding puts most animals at risk for predation, they must consider this danger when making their foraging decisions. Animals that incorporate the risk of predation into their foraging decisions are said to practice risk-sensitive foraging. Risk-sensitive foraging Foraging behavior that is influenced by the presence of predators. The creek chub is a fish that faces the common challenge of finding its lunch rather than becoming lunch for a predator. Small creek chubs feed on sludge worms (Tubifex spp.) that live in the mud of streams; understandably, the fish prefer to feed in locations that have more worms. But what if locations containing more worms also contain more predators including cannibalistic larger creek chubs? How much food would it take to entice the small creek chubs to feed in the riskier location? To address this question, researchers placed small creek chubs into an artificial stream that contained a refuge from predators in the middle section of the stream. On one end of the stream, the researchers placed a large creek chub and a low density of worms. On the other end of the stream, they placed two large creek chubs and manipulated different densities of worms. As you can see in Figure 4.21, when the opposite end of the stream had two large creek chubs, the small creek chubs would not move to the side containing two predators until that side contained at least three times as many worms to feed on. In short, the small creek chubs balance the risk of predation against the benefits of food availability and adjust their behavioral phenotype accordingly.
Figure 4.21 Risk-sensitive foraging. The sensitivity of young creek chubs (Semotilus atromaculatus) to food density and predators was tested in artificial streams. All streams had one side that contained one predatory adult creek chub and a low density of food (0.17 worms/cm2). (a) When the right end of the stream contained two predators and the same low density of food, the young chubs moved to the left side. (b) When the right end contained two
predators and a medium food density (0.17 worms/cm2), the young chubs still moved to the left side. Only when the side with two predators contained either a (c) high food density (0.50 worms/cm2) or (d) very high food density (1.0 worms/cm2) did the young chubs move to the right side of the stream. After J. F. Gilliam and D. F. Fraser, Habitat selection under predation hazard: Test of a model with foraging minnows, Ecology 68 (1987): 1856–1862.
Optimal Diet Composition
Most animals do not consume a single food item but choose from a range of food items. For example, consider the food choices of coyotes living in the western United States. In Idaho, the coyote can consume a variety of prey species, including small prey such as voles (Microtus montanus), medium prey such as cottontail rabbits (Sylvilagus nuttallii), and large prey such as jackrabbits (Lepus californicus). Larger prey species provide larger energy benefits to the coyote, but require more time and energy to subdue and consume. Given these options, the coyote has to decide which prey species it should pursue and which prey species it should ignore. To determine the optimal food decision, we must balance the energy obtained from the prey and the handling time—the time required to subdue and consume the prey. Assuming the same amount of handling time for each choice, the optimal decision for the predator will depend on the energy obtained from each prey and the abundance of the prey. The optimal decision can change if handling time is not equal. In this situation, we need to consider the amount of energy gained per unit time for each prey species. We can do this by dividing the energy benefit of a prey item by its handling time. When we do this, we sometimes find that the smallest prey should be consumed because, although they provide a smaller energy benefit than larger prey, their low handling time can provide the predator with the highest energy gain per unit time. In the case of the coyotes, researchers have found that although jackrabbits require more effort to catch and consume than cottontail rabbits or voles, jackrabbits also provide a larger energy benefit, so the coyotes should rank jackrabbits as the most profitable food item, followed by cottontail rabbits and then voles. Handling time The amount of time that a predator takes to consume a captured prey. Once we know how different food items compare in terms of energy
gained per unit of handling time, we can make a number of predictions. For example, the predator should always eat the prey species that provides the highest energy benefit; if this prey is abundant, it is the only prey that the predator should consume. This strategy maximizes the animal’s energy gain. However, if this highest-energy prey is rare and the predator’s energy needs are not met, the animal should include less profitable items in its diet. Prey species of very low energy value should never be included in the diet unless all higher-energy prey are scarce. In the case of the coyote, researchers have found that the coyotes appear to be making optimal diet choices. The coyotes always consumed the jackrabbits regardless of their abundance. However, when jackrabbits became less abundant, the coyotes would increase their consumption of cottontail rabbits and voles.
Diet Mixing
Some foragers consume a varied diet because one type of food might not provide all the necessary nutrients. Humans, for example, can synthesize many amino acids, but essential amino acids can only be obtained from one’s diet. A diet of only rice or only beans does not possess the complete set of essential amino acids needed by humans. However, a diet that combines rice and beans contains all the required essential amino acids because each contains the essential amino acids that are missing in the other. The benefits of diet mixing have been demonstrated using nymphs (immature stages) of the American grasshopper (Schistocerca americana). As you can see in Figure 4.22a, grasshopper nymphs grew faster when fed a mixture of kale, cotton, and basil than when they were offered any one of these food plants alone. The effect was even more pronounced on lowerquality, natural food plants, such as mesquite and mulberry: Nymphs with mixed diets grew almost twice as fast as those feeding on either one of these plant species alone, shown in Figure 4.22b. Based on these data, we might predict that when given a choice, these grasshoppers would decide to forage on a mixed diet to improve their fitness.
Figure 4.22 Mixed diets. Young grasshoppers grow faster on mixed diets than on any single diet, regardless of whether comparisons were made using (a) crop plants or (b) natural plants. In general, mixed diets supply a more complete range of nutrients needed by animals than single diets. Error bars are standard errors. After E. A. Bernays et al., Dietary mixing in a generalist herbivore: Tests of two hypotheses, Ecology 75 (1994): 1997–2006. Throughout this chapter, we have seen that organisms commonly experience spatial and temporal variation in their environment. In response to this variation, many have evolved the ability to produce multiple phenotypes from a single genotype. The strategy of using multiple phenotypes— including changes in morphology, physiology, or behavior—is effective when there are trade-offs such that no single phenotype performs well in all environments. The evolution of phenotypic plasticity is common among all groups of organisms on Earth, wherever there are reliable environmental cues.
Concept Check
1. Why is optimal foraging an example of phenotypic plasticity? 2. Why does central place foraging cause animals that travel farther to bring back larger amounts of food? 3. What are the costs and benefits that animals must consider during risk-sensitive foraging?
###### Responding to Novel Environmental Variation
Concepts
Responding to Novel Environmental Variation Elevated CO2 experiment. Tall towers at the Duke University Forest have been emitting CO2 into the atmosphere for several years and researchers have been tracking the effects on the plants Ecologists have a good understanding of phenotypically plastic adaptations to environmental variation that has been present for hundreds of thousands of generations, long enough to evolve an appropriate phenotypic response mechanism. But how do organisms respond to more recent environmental variation? One of the most profound changes in our environment has been the global increase in atmospheric CO2. In 1958, Charles Keeling began recording atmospheric CO2 concentrations atop 3,400-m-high Mauna Loa on the island of Hawaii. Keeling wanted to determine whether anthropogenic emissions were increasing the
concentration of CO2 in the atmosphere. At the time he began his study, scientists had no accurate long-term measurements of atmospheric CO2 concentrations. In 1958, the CO2 concentration was about 316 parts per million (ppm; 316 CO2 molecules per million molecules of air, mostly nitrogen and oxygen). During the subsequent decades, the concentration of CO2 in the atmosphere has increased dramatically, rising to 354 ppm by 1990 and 404 ppm by the end of 2016, with no sign of leveling off. Other research indicates that this concentration of CO2 has not been present on Earth for at least the past 10,000 years. As demand for energy and agricultural land increases, the concentration of CO2 is expected to reach 500 to 1,000 ppm by the year 2100. How will organisms respond to such a change in their environment given that they have not experienced such high concentrations of CO2 in the past 10,000 years? To address this question, scientists began conducting large outdoor experiments in the1990s in which tall towers emit CO2 gas over forests, deserts, wetlands, and croplands to cause an elevation in CO2 ranging from 475 to 600 ppm. Many of these experiments continue today and, averaged across several such experiments, researchers have found that elevated CO2 causes an increase in the rate of photosynthesis by 40 percent. In addition, because plants open their stomata to obtain CO2, the elevated CO2 concentrations allow plants to keep their stomata closed more often, resulting in a 22 percent reduction in transpiration of water. All this translates into improved plant growth. Plants experiencing elevated CO2 had a 17 percent
increase in the growth of their shoots and a 30 percent increase in the growth of their roots. These growth responses represent averages across a variety of species, but not all species responded in the same manner. For example, growth improved in C3 plants but not in C4 plants. Researchers hypothesize that because the C4 pathway of photosynthesis already pumps high concentrations of CO2 into the bundle sheath cells of the leaf, higher atmospheric concentration of CO2 has little additional effect. Given that most plant species use the C3 pathway, most plants will experience higher growth unless other nutrients become limiting or if herbivory of the plants also increases and causes an increased loss of plant tissues. On the other hand, C4 plants, which include corn, sugarcane, and many other important crops, are not expected to grow any faster as humans continue to elevate the concentration of CO2 in the atmosphere. Changes in atmospheric CO2 over time. Measurements on the island of Hawaii have shown that CO2 concentrations have continuously increased during the past 50 years. After
http://www.esrl.noaa.gov/gmd/ccgg/trends/. In 2016, researchers announced that they were planning to begin the first CO2 emission experiments in a tropical rainforest in which they will expose small areas of the forest to 600 ppb of CO2. This is important because it has been hypothesized that tropical rainforests may absorb a lot of atmospheric CO2. The project, which will be based in Manaus, Brazil, faces a number of challenges, including raising tens of millions of dollars to fund the research project and the practical problem of how to transport thousands of metric tons of CO2 along a 34-km treacherous dirt road in the Amazon. The project is expected to last at least 12 years and provide a number of insights regarding how much of the elevated CO2 will be absorbed by the plants and whether the forest will respond with increased growth rates. The change in CO2 concentrations is just one example of the many anthropogenic changes occurring on Earth today. Most organisms have flexible phenotypes that have been shaped by natural selection in response to past environmental variation. Organisms facing novel environmental variation from anthropogenic causes may be able to make use of these existing adaptations and they may also experience continued evolution for new types of flexible phenotypes. However, many other types of anthropogenic impacts—such as air and water pollution— may be far outside the range of historic environmental variation for a population. As a result, populations may not possess phenotypically plastic strategies that will allow them to perform well when facing these types of
anthropogenic impacts. SOURCES: Grossman, D. 2016. Amazon rainforest to get a growth check. Science 353:635–636. Jaub, D. 2010. Effects of rising atmospheric concentrations of carbon dioxide on plants. Nature Education Knowledge 1:21. Ainsworth, E. A., and S. P. Long. 2005. What have we learned from 15 years of free-air CO2 enrichment (FACE)? A meta-analytic review of the responses of photosynthesis, canopy properties and plant production to rising CO2. New Phytologist 165:351–372.
Summary of Chapter Concepts
4.1 Environmental variation favors the evolution of variable phenotypes. Temporal variation occurs across a range of hours to years, with the most extreme variation being the least frequent. Spatial variation also exists due to differences in climate, topography, and soils. Phenotypic plasticity, the ability to produce alternative phenotypes, is favored when organisms experience environmental variation, when reliable cues indicate the current state of the environment, and when no single phenotype is superior in all environments. Phenotypically plastic traits include behavior, physiology, morphology, and life history. Each type of trait differs in how fast it can respond to environmental change and whether the responses are reversible. Key Terms: Temporal environmental variation, Weather, Climate, Phenotypic trade-off, Phenotypic plasticity, Acclimation
4.2 Many organisms have evolved adaptations to variation in enemies, competitors, and mates. Responses to enemies include changes in behavior that make individuals harder to detect, morphological defense that makes prey harder to capture, and chemical defense that makes prey less palatable. Responses to competitors include morphological changes in plants to make them better able to obtain resources, morphological changes in animals that make them better able to consume and digest scarce food, and behavioral strategies in animals that improve their ability to find scarce food. Organisms generally favor breeding with another individual, but a scarcity of mates can make self-fertilization a viable alternative for some species. Key Terms: Hermaphrodite, Inbreeding depression
4.3 Many organisms have evolved adaptations to variable abiotic conditions. Variation in temperature has favored the evolution of
isozymes and switching between microhabitats. Variation in water availability has favored plants that can open and close their stomata and alter their root-to-shoot ratios. Variation in salinity has favored the evolution of novel ways of adjusting solute concentrations to minimize the cost of osmoregulation. Variation in oxygen can cause adaptive increases in red blood cells and hemoglobin to improve the uptake of oxygen at high altitudes. Key Term: Microhabitat
4.4 Migration, storage, and dormancy are strategies used to survive extreme environmental variation. Migration allows organisms to leave areas with degrading environments, storage allows organisms to have an extra supply of energy to make it through periods of degrading environments, and dormancy allows organisms to shut down their metabolism until the harmful environmental conditions have passed. Key Terms: Migration, Dormancy, Diapause, Hibernation, Torpor, Correlation, Aestivation
4.5 Variation in food quality and quantity is the basis of optimal foraging theory. Central place foraging predicts that the amount of time spent foraging at a site and the amount of food brought back to a central nest will depend on the benefits gained over time at the site and the round-trip travel time. Risk-sensitive foragers consider not only the energy to be gained, but also the predation risk posed. Many animals must also consider a range of alternative food items, the energy and abundance of each food item, and whether they should consume a mixture of food items to meet all their nutritional needs. Key Terms: Optimal foraging theory, Central place foraging, Risk-sensitive foraging, Handling time
Critical Thinking Questions
1. What is the difference between weather and climate in terms of temporal variation? 2. How could spatial heterogeneity be perceived by an organism as temporal heterogeneity? 3. Why do we have to consider the mean fitness across all environments when evaluating whether evolving a phenotypically plastic genotype will be favored over a non-plastic genotype? 4. How would the presence of unreliable environmental cues affect the evolution of phenotypically plastic responses to environmental variation? 5. Would the frequency of experiencing predator versus no-predator environments affect the evolution of phenotypically plastic traits? 6. If predators reduce prey abundance to very low levels, how might variable predator environments affect the evolution of flexible mating strategies? 7. If a plant can improve its ability to obtain water by growing more roots, why shouldn’t the plant always grow more roots? 8. How could you experimentally determine whether migrating birds use day length or temperature as an environmental cue for migration? 9. What is the relationship between correlation and causation? 10. When determining whether to feed on small, medium, or large prey, why should predators evaluate both the energy obtained from each prey item, the abundance of each prey item, and the handling time of each prey item?
#### Graphing the Data: the Foraging Behavior of American Robins
GRAPHING THE DATA The Foraging Behavior of American Robins The following data were collected by a scientist observing the number of earthworms that American robins (Turdus migratorius) can hold in their beaks as they searched a lawn after a summer rain. Graph the data and describe the relationship between time and the number of collected earthworms. Does this correlation between time and the number of worms collected represent both a correlation and causation? Time (min) Average number of earthworms collected 1.0 2.0 2.8 3.4 3.7 3.9 4.0 4.0