7 Evolution and Adaptation
7Evolution and Adaptation Ratite birds. Two-wattled Cassowary (Casuarius casuarius) female on a forest edge in Australia, Queensland, Moresby Range National Park. Favoring Flightless Birds One of the distinguishing characteristics of birds is that they possess feathers that help them fly. However, not all birds have the ability to fly, including penguins and a group of birds known as ratites. Ratites are a group of well-known species, such as the ostriches of Africa (Struthio spp.), the emus and cassowaries of Australia (Dromaius spp., Casuarius spp.), the kiwis of New Zealand (Apteryx spp.), and the rheas of South America (Rhea spp.). For decades, scientists have assumed that the ratites all shared a common ancestor that was flightless and, following the breakup of the continents, the isolated populations of this ancestor gradually evolved into the distinct species that we see today. While this interpretation is an attractive story due to the simple explanation, research during the past decade has
demonstrated that this evolution of flightless ratites was much more complex. Using DNA from the various species of ratites and other birds has allowed researchers to build a family tree to determine which species are closely versus distantly related to each other. It turns out that some species of ratites are more closely related to other species of flying birds than to other species of ratites. This means that the ancestor of the ratites was likely a species that could fly and that today’s flightless ratites actually became flightless through many independent evolutionary events rather than flightlessness evolving first and then the ratites evolving into separate species. In many cases, the evolution of flightless birds occurs on islands where predators are few or completely lacking. Predator-free islands should reduce the benefits of flying, since the birds would not need flight as a means of escape. Although flight certainly serves many other purposes, it leads one to hypothesize that birds living on islands, even if they have not evolved to be flightless, may evolve a reduction in the investment of their flight machinery. To test this hypothesis, researchers examined the size of the flight muscles and the length of the legs in bird species living on islands versus related species that live on the mainland continents. In 2016, they reported their discovery that flying species of birds that live on islands have evolved smaller flight muscles and longer legs. Moreover, the magnitude of this effect was greatest on those islands containing the fewest predators. The researchers interpreted the evolution of longer legs as permitting a slower, more energyefficient takeoff for the birds by jumping higher, since the birds no longer needed to take off to avoid predators rapidly by primarily using their flight muscles. Such trait changes may represent the intermediate evolutionary steps that the ratite birds went through during their evolution. “Flying species of birds that live on islands have evolved smaller flight muscles and longer legs.”
The evolution of reduced flight muscles on predator-free islands also means that these birds are more susceptible to any predators that are introduced to the islands. As we will see in later chapters, the unintentional introduction of non-native bird predators, such as snakes, has decimated the bird fauna of many islands. The research on bird flight is just one example of how scientists can use the theory of evolution by natural selection to obtain tremendous insight into how natural selection operates in nature. By studying how evolution occurs in wild populations, we can see how natural selection can alter the traits of species and populations over time and, in turn, affects species interactions. In this chapter, we will explore the ways in which evolution causes populations to become genetically distinct and how this leads to the origin of new species. SOURCES: Wright N. A., et al. 2016. Predictable evolution toward flightlessness in Volant island birds. PNAS 113: 4765–4770. Harshman J., et al. 2008. Phylogenomic evidence for multiple losses of flight in ratite birds. PNAS 105: 13462–13467.
Learning Objectives
After reading this chapter, you should be able to:

7.1 Describe how the process of evolution depends on genetic variation.
7.2 Explain how evolution can occur through random processes or through selection.
7.3 Illustrate how microevolution operates at the population level.
7.4 Describe the way that macroevolution operates at the species level and higher levels of taxonomic organization.

The story of evolving flightlessness demonstrates that evolution shapes the form and function of organisms according to properties of their environments. Evolution depends on genetic variation, and this variation can arise from a number of processes. Over time, populations and species can evolve changes in traits, such as the size of bird flight muscles. These changes can evolve due to random processes or due to the nonrandom process of selection. Some of the most important sources of natural selection include differences in physical conditions, food resources, and interactions with competitors, predators, pathogens, and individuals of the same species. In this chapter, we will examine such processes and explore how genes and the environment come together to cause the evolution of populations and new species.
#### 7.1 the Process of Evolution Depends on Genetic Variation
7.1 The process of evolution depends on genetic variation In Chapter 4, we discussed how the traits expressed by an individual are the outcome of genotypes and the environment interacting. When genetic variation is present, it allows evolution by natural selection. In this section, we review the structure of DNA, the process of how genes help determine the phenotypes of organisms, and the process by which variation in genes is produced.
The Structure of Dna
Genetic information is contained in the molecule deoxyribonucleic acid, also known as DNA—a molecule composed of two strands that are wound together into a shape known as a double helix. Each strand is composed of subunits called nucleotides and each nucleotide is composed of a sugar, a phosphate group, and one of four different nitrogenous bases: adenine (A), thymine (T), cytosine (C), and guanine (G). Just as a sequence of letters signifies a particular meaning, or word, genetic information is coded in the particular order of the different nitrogenous bases. Long strands of DNA are wound around proteins into compact structures called chromosomes. Deoxyribonucleic acid (DNA) A molecule composed of two strands of nucleotides that are wound together into a shape known as a double helix. Chromosomes Compact structures consisting of long strands of DNA that are wound around proteins.
Genes and Alleles
Genes are regions of DNA that code for particular proteins, which in turn affect particular traits. Different forms of a particular gene are referred to as alleles. In diploid organisms—those having two sets of chromosomes—one allele comes from the mother’s gamete and the other comes from the father’s gamete. As you may recall, each gamete is haploid, meaning that it has just one set of chromosomes. Alleles

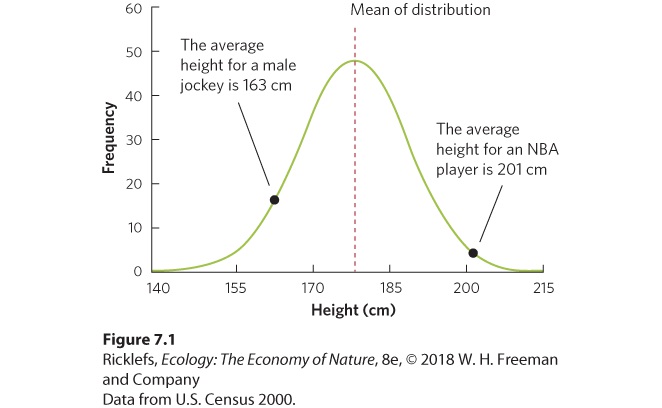
Different forms of a particular gene. In many cases, a change in alleles can create differences in an organism’s phenotype. The ABO blood groups in humans, for example, are determined by which allele a person inherits from each parent—A, B, or O. The allele is responsible for production of the antigens A and B, which are molecules on the surface of our red blood cells that interact with the immune system. (Note that the O allele does not produce an antigen.) Individuals with blood type A have AA or AO genotypes; individuals with blood type B have BB or BO genotypes. All remaining individuals have either AB or OO genotypes. In this case, the link between the genotype and the phenotype is direct, and the pattern of inheritance is straightforward. For example, children of an AA father and a BB mother will all have the AB genotype. Whereas blood type is determined by different alleles on a single gene, polygenic traits reflect the effects of alleles from several genes. For example, eye color in humans is determined by at least three genes that control pigments in different parts of the iris of the eye. Patterns of phenotype inheritance that depend on interactions among multiple alleles can be quite complex. Polygenic When a single trait is affected by several genes. Many phenotypes in a population can span a range of values because they are polygenic traits. Body size is a good example. Most populations exhibit a normal, or bell-shaped, distribution of body sizes, as shown in Figure 7.1. In this distribution, most individuals cluster near the middle of the range with progressively fewer located toward the upper and lower extremes. Part of this continuous variation might be due to environmental differences, such as the amount of resources available. However, much of the variation can be attributed to the actions of many genes, each with a relatively small influence on the value of the trait. If several genes influence body size, an individual’s size will depend on the mix of alleles for all of these genes. The tendency for individuals to be concentrated toward the center of the distribution reflects the relative improbability of an individual inheriting mostly alleles that code for large body size or mostly alleles that code for small body size. Think of
this in terms of flipping a coin. The chance of getting 10 tails in a row (about 1 in 1,000) is much more remote than the chance of getting 5 heads and 5 tails (about 1 in 4).
Figure 7.1 Frequency distribution of polygenic traits. When a continuous trait is determined by many genes, the distribution follows a bell-shaped curve. For the heights of adult males in the United States, for example, there is a symmetrical distribution around the mean. Most individuals express an intermediate trait value, whereas only a few individuals, such as professional basketball players and jockeys, express extreme trait values. Whereas some genes only affect a single trait, such as size, other genes affect multiple traits, an effect referred to as pleiotropy. For example, chickens have a gene—known as the frizzle gene—which causes feathers to curl outward rather than to lay flat along the body. However, this gene causes other variations, including faster metabolism, slower digestion, and less frequent egg laying. When a gene has pleiotropic effects, any changes in the gene can have far-reaching effects on the traits of organisms. Pleiotropy When a single gene affects multiple traits. In some cases, the expression of one gene can be controlled by other genes. This is known as epistasis. In the case of mouse hair color, for example, there is a gene that determines whether a mouse will make black or brown hair pigments. However, there is a second gene that determines whether the hair will receive any pigments at all. If alleles in the second gene prevent
pigments from being deposited in the hair, the alleles of the first gene become irrelevant and the mouse will have a white coat. Epistasis When the expression of one gene is controlled by another gene.
Dominant and Recessive Alleles
Every individual has two copies of each gene, one inherited from its mother and one from its father. Exceptions to this rule include genes located on sex chromosomes, the genes of organisms that reproduce by self-fertilization, haploid organisms, and organisms such as plants that alternate between haploid and diploid generations as part of their normal life cycle. An individual with two different alleles of a particular gene is said to be heterozygous for that gene, as in the case of a person with the AB blood type. An individual with two identical alleles is said to be homozygous, for example, a person with AA blood type. When an individual is heterozygous, the two different alleles may produce an intermediate phenotype, as in the case of a person with the AB blood type that expresses both alleles. When both alleles contribute to the phenotype, the alleles are said to be codominant. Codominance is also found in the flower color of several species of plants (Figure 7.2). Alternatively, one allele may mask the expression of the other. In this case, the allele that is expressed is called dominant and the allele that is not expressed is called recessive. In domestic pigs, for example, the allele for white coat color is dominant and the allele for black coat color is recessive.
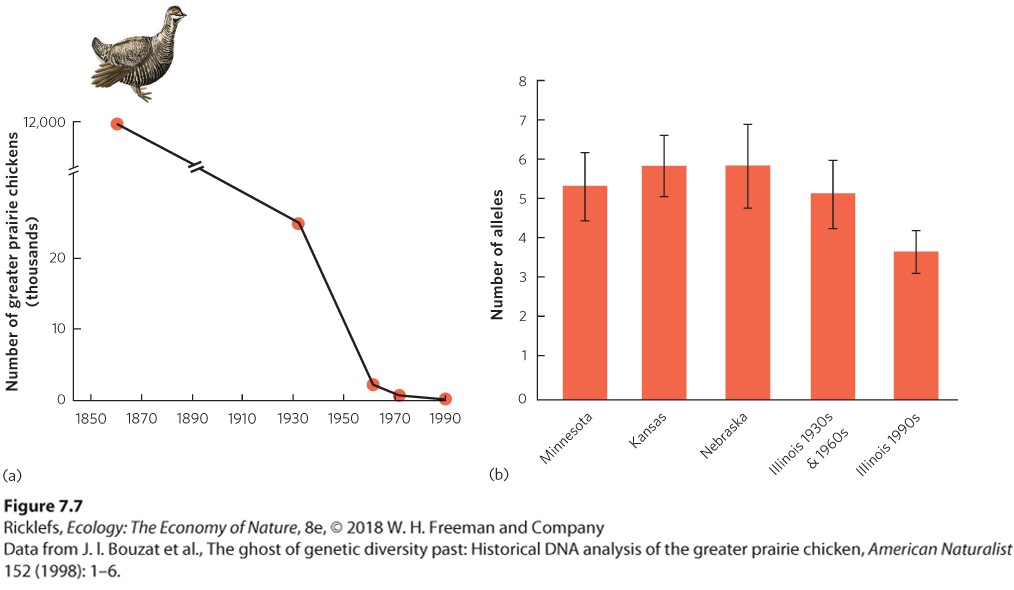
Figure 7.2 Codominance. In snapdragons, the red and white flowers are homozygous genotypes. The pink flower gets its color from one red gene and one white gene that are codominant. Heterozygous When an individual has two different alleles of a particular gene. Homozygous When an individual has two identical alleles of a particular gene. Codominant When two alleles both contribute to the phenotype. Dominant allele An allele that masks the expression of the other allele of a given gene. Recessive An allele whose expression is masked by the presence of another allele. Fortunately, most harmful alleles are recessive and are therefore not expressed in a heterozygous individual. Any dominant harmful alleles that might arise are expressed when homozygous or heterozygous. Because they reduce fitness, dominant harmful alleles are strongly selected against and removed from the population over time. In contrast, recessive harmful alleles are expressed when homozygous but not when heterozygous, so they can
persist in a population because they are not selected against when they occur in heterozygous individuals. Examples of recessive harmful alleles in humans include those that cause sickle cell anemia and cystic fibrosis. A gene pool consists of the alleles from all of the genes of every individual in a population. The gene pools of most populations that reproduce sexually contain substantial genetic variation. With the ABO blood type gene, for example, the human population of the United States includes 61 percent O alleles, 30 percent A alleles, and 9 percent B alleles. The proportions of these alleles vary among populations. Populations of Asian descent, for instance, tend to have higher frequencies of B alleles, while populations of Irish descent have higher frequencies of O alleles. Gene pool The collection of alleles from all individuals in a population.
Sources of Genetic Variation
Now that we understand the role of genes and alleles, we need to review how we obtain genetic variation in the traits of organisms. One of the most common ways to generate variation is through sexual reproduction. By combining a haploid sex cell of one parent with a haploid sex cell of another parent, new combinations of alleles can be produced in the offspring across many different chromosomes. The chromosomes in a haploid gamete are a random assortment of the chromosomes in the parent’s diploid cells, meaning that they can be any combination of chromosomes that the parent received from its mother and father. When an individual produces an egg cell, for example, some chromosomes in the egg will have come from the individual’s father, whereas other chromosomes in the egg will have come from the individual’s mother. As we will see in Chapter 9, the creation of new gene combinations through sexual reproduction represents a major strategy for species to create offspring that are resistant to rapidly evolving parasites and pathogens. Random assortment The process of making haploid gametes in which the combination of alleles that are placed into a given gamete could be any combination of those possessed by the diploid parent.
Two additional ways genetic variation arises are through mutation and recombination. Mutation is a random change in the sequence of nucleotides in regions of DNA that either comprise a gene or control the expression of a gene. Mutations can occur anywhere along the chromosomes, although some regions of the chromosome can experience higher frequencies of mutation than others. Many mutations have no detectable effect and are referred to as silent, or synonymous, mutations. Other mutations may simply alter the appearance, physiology, or behavior of the individual. When phenotypic changes are better suited to the environment, these phenotypes will be favored by natural selection. Some mutations, however, can cause drastic, often lethal, changes in the phenotype. Many human genetic disorders, such as sickle cell anemia, Tay-Sachs disease, cystic fibrosis, and albinism, as well as tendencies to develop certain cancers and Alzheimer’s disease, are caused by single-nucleotide mutations of individual genes. Mutation A random change in the sequence of nucleotides in regions of DNA that either comprise a gene or control the expression of a gene. Genetic recombination is the reshuffling of genes that can occur as DNA is copied during meiosis, the process that creates haploid gametes from diploid parent cells. During meiosis, pairs of homologous chromosomes—one member of which is inherited from each parent—line up next to each other. When the two chromosomes in the pair do not exchange any DNA, we end up with haploid cells that contain unaltered chromosomes. However, sometimes the two chromosomes in the pair do exchange DNA, in a process known as crossing over, as shown in Figure 7.3. In some cases, crossing over can also occur between nonhomologous chromosomes. In either case, new genes are not being created, but new combinations of alleles are produced that have the potential to produce new phenotypes.
Figure 7.3 Genetic variation through recombination. During meiosis in eukaryotic organisms, pairs of homologous chromosomes line up together. When the chromosomes exhibit crossing over, they exchange DNA and each chromosome contains a new combination of genes. Recombination The reshuffling of genes that can occur as DNA is copied during meiosis and chromosomes exchange genetic material. One of the best-known examples of recombination involves the immune system of vertebrates. Vertebrates face a diversity of pathogens that continually evolve so that they become better at attacking their hosts. To combat these ever-changing pathogens, vertebrates need an ever-changing immune system that can identify and destroy the pathogens. Recombination provides the mechanism to create the high genetic variation in the immune system that vertebrates need to match the rapid evolution of their parasites. We will talk much more about this issue in Chapter 9.
Concept Check
1. What is the difference between genes and alleles? 2. Why is it essential that traits be inherited for evolution to occur? 3. What are the three primary sources of genetic variation?
#### 7.2 Evolution Can Occur Through Random Processes or Through Selection
7.2 Evolution can occur through random processes or through selection In western New York State, there is a herd of white-tailed deer that looks very different from most white-tailed deer. Many of the deer living on the 4,300-ha Seneca Army Depot do not have typical reddish brown coats. Rather, their coats are white (Figure 7.4). The white coat phenotype is due to a rare mutation that occurs in white-tailed deer populations. Because a white coat can frequently make a deer more visible to predators, it does not provide any fitness benefits and we would not expect it to persist. So why is there a high frequency of white deer on the Seneca Army Depot? It turns out that when the army depot was built in 1941, the 4,300-ha area was fenced and several dozen deer were trapped inside. A few years later, 2 white deer were observed. Given that white deer were such an unusual sight, authorities at the depot banned hunters from killing the deer with the white phenotype. Over time, the deer population grew larger and the white phenotype became more frequent. Today, of the nearly 800 deer living on the property, about 200 are white. In recent years, the U.S. Corps of Engineers has been trying to sell the land since it was no longer needed by the federal government. This made the future of the white deer uncertain, particularly if the ban on hunting white deer was lifted. In 2016, the land was sold by to a company that pledged to develop some of the land for industry while still protecting the deer herd with the rare white phenotype. As a result, the rare phenotype that emerged due to random genetic processes will continue to persist.
Figure 7.4 A mutant white-tailed deer. At the Seneca Army Depot in western New York State, a mutation for white hair appeared in the 1940s. Since that time, the white phenotype was protected while the normal brown phenotype was hunted. Over the subsequent 70 years, the white phenotype has come to compose about 25 percent of the population. The story of the white deer demonstrates how evolution often happens through multiple processes. Random events, such as mutations, may confer no fitness advantage when they first appear. Such is the case for most deer populations in which the white mutation occasionally arises but rarely becomes frequent in a population. However, if selection for the mutant phenotype occurs after the mutant appears, as happened at the Seneca Army Depot with reduced hunting pressure on deer with the white phenotype, this nonrandom process can cause the mutant to become more frequent in the population.
Evolution Through Random Processes
Random processes can facilitate evolutionary change in a population. In addition to mutation, random processes include genetic drift, bottleneck effects, and founder effects. Mutation We have seen that mutation is an important way in which genetic variation can arise in a population. Because genes often code for functions that are vital to performance and fitness, mutations that negatively impact these functions
are not favored by selection. However, a small fraction of mutations can be beneficial. For example, Figure 7.5 illustrates what happened in the deer herd of the Seneca Army Depot. In a herd of deer, a mutation for the white coat appeared, which added genetic variation to the population. After the mutation appeared, the ban on hunting white deer caused selection for the white phenotype and the frequency of white deer increased.
Figure 7.5 Evolution by mutation. Mutations, such as white coat in white-tailed deer, can arise in populations. If the mutation confers a fitness benefit, the mutation can increase in frequency in the population over multiple generations. Mutation rates vary a great deal in different groups of organisms, but among genes that are expressed and can be observed as altered phenotypes, mutation rates range from 1 in 100 to 1 in 1,000,000 per gene per generation. The more genes that a species carries, the higher the probability that at least one gene will experience a mutation. Similarly, the larger the size of the population, the higher the probability that an individual in the population will carry a mutation. Genetic Drift Genetic drift, another random process, occurs when genetic variation is lost because of random variation in mating, mortality, fecundity, or inheritance. Genetic drift is more common in small populations because random events can have a disproportionately large effect on the frequencies of genes in the population. But how does one determine whether an evolved phenotype is the outcome of drift versus some other process, such as natural selection? Research on the Mexican cavefish (Astyanax mexicanus) provides an answer. Genetic drift A process that occurs when genetic variation is lost because of random variation in mating, mortality, fecundity, and inheritance.
The Mexican cavefish is a species composed of some populations that live in cave streams and other populations that live in surface streams. Although the populations can interbreed, they look quite different. As is the case with many cave-adapted animals, the cave populations have very reduced eyes and reduced pigmentation (Figure 7.6a). In contrast, populations living in surface streams have normal eyes and dark pigmentation. To determine whether these changes were due to natural selection or genetic drift, researchers raised individuals from the cave population, individuals from the surface population, and hybrid offspring made by interbreeding the cave and surface populations. The researchers then examined regions of the fish DNA that coded for eye size and pigmentation, which could contain one or more genes. In 2007, they reported that the 12 regions of DNA that coded for large eyes in the surface population all coded for small eyes in the cave population. The results are shown in Figure 7.6b. This suggests that natural selection favored all of the eye genes to evolve in a similar direction to produce smaller eyes. In contrast, when they examined the 13 regions of DNA that coded for pigmentation, they found that 5 of the regions coded for increased pigmentation in the cave populations and 8 of the regions coded for decreased pigmentation, as shown in Figure 7.6c. The lack of a consistent pattern among the 13 regions of DNA suggests that natural selection was not involved. Instead, the differences in pigmentation in the cavefish populations were likely produced by genetic drift. Given that small populations tend to experience more genetic drift than large populations, it may be the case that the cave population was initially very small.
Figure 7.6 Evolution by genetic drift. (a) Mexican cavefish populations that live in surface streams have large eyes and dark pigmentation, whereas populations that live in caves have small eyes and reduced pigmentation. (b) When researchers compared how different regions of DNA that code for eye size changed between surface and cave populations, they found that all 12 regions coded for smaller eyes in the cave population than in the surface population. Because all 12 regions changed in the same direction, this suggests that natural selection selected for smaller eyes. (c) When researchers looked at changes in regions of DNA that code for pigmentation, they found 5 regions coded for increased pigmentation, while 8 regions coded for decreased pigmentation. Because the 13 regions did not change in the same direction, this suggests that changes in pigmentation were due to genetic drift. Bottleneck Effects A reduction in genetic variation can also occur because of a severe reduction in population size, known as the bottleneck effect. When a population experiences a large reduction in the number of individuals, the survivors carry only a fraction of the genetic diversity that was present in the original, larger population. Moreover, after being reduced to a small population by the bottleneck effect, the small population can then experience genetic drift. Population reductions can result from natural causes, for example, a drought that reduces the abundance of food, or anthropogenic causes, such as loss of habitat due to the construction of homes or factories. Bottleneck effect A reduction of genetic diversity in a population due to a large reduction in population size. One example of a bottleneck effect is the greater prairie chicken (Tympanuchus cupido), which is a grassland bird that historically lived throughout much of the middle United States, including Minnesota, Kansas, Nebraska, and Illinois. While prairie chickens have remained abundant in many neighboring states, the population in Illinois declined from approximately 12 million in 1860 to only 72 birds by 1990, as illustrated in
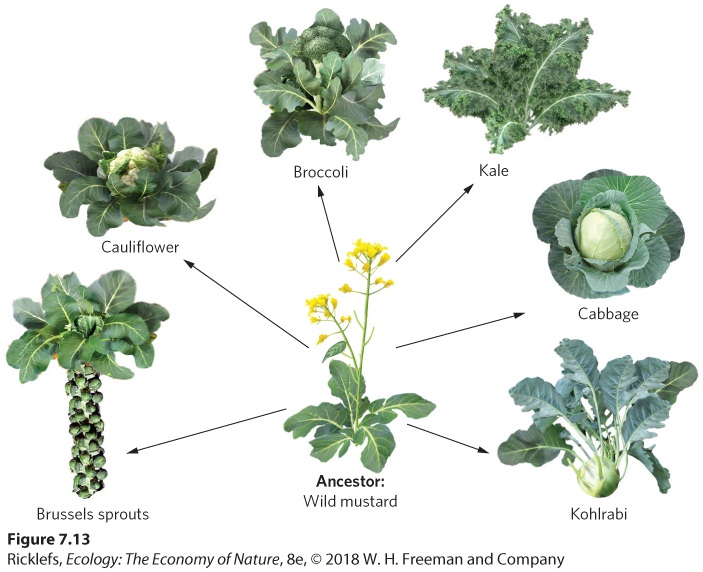
Figure 7.7a. To determine whether this dramatic decline in population size was associated with a decline in genetic diversity, researchers collected DNA samples from museum specimens of prairie chickens from the 1930s, when the population was 25,000, and from the 1960s, when the population was 2,000. They defined this time period from the 1930s to the 1960s as “prebottleneck.” They also compared the genetic diversity of the Illinois birds before and after the bottleneck to the genetic diversity in the present-day populations of prairie chickens in Minnesota, Kansas, and Nebraska. In all
cases, they examined the number of alleles that a population contained for each of six different genes. As shown in Figure 7.7b, the large populations of the neighboring states and the large historic population in Illinois all contained a high number of alleles. The current population in Illinois, however, has a lower number of alleles, which reflects the genetic bottleneck effect. The state of Illinois has since purchased more prairie habitat and introduced hundreds of prairie chickens from neighboring states to help bolster the Illinois population and increase its genetic diversity.
Figure 7.7 Evolution by the bottleneck effect. (a) The greater prairie chicken population in Illinois declined from approximately 12 million in the 1860s to 72 birds in 1990. (b) Averaged across six different genes, the mean number of alleles is high for birds in neighboring states that still have large populations and for the larger, historic population in Illinois that existed in the 1930s and 1960s. In the current population in Illinois, however, the small population is experiencing a population bottleneck and has a lower mean number of alleles. Error bars are standard errors. The bottleneck effect is of particular interest because the subsequent reduction in genetic diversity may prevent the population from adapting to future environmental changes. This is especially true for organisms that face deadly pathogens. An inability to evolve against new strains of a pathogen could lead to the extinction of the host organism. For example, the African cheetah (Acinonyx jubatus) experienced a population bottleneck approximately 10,000 years ago. Although the cause of that bottleneck is unknown, the current population contains very little genetic variation. This low genetic variation makes cheetahs more vulnerable to pathogens,
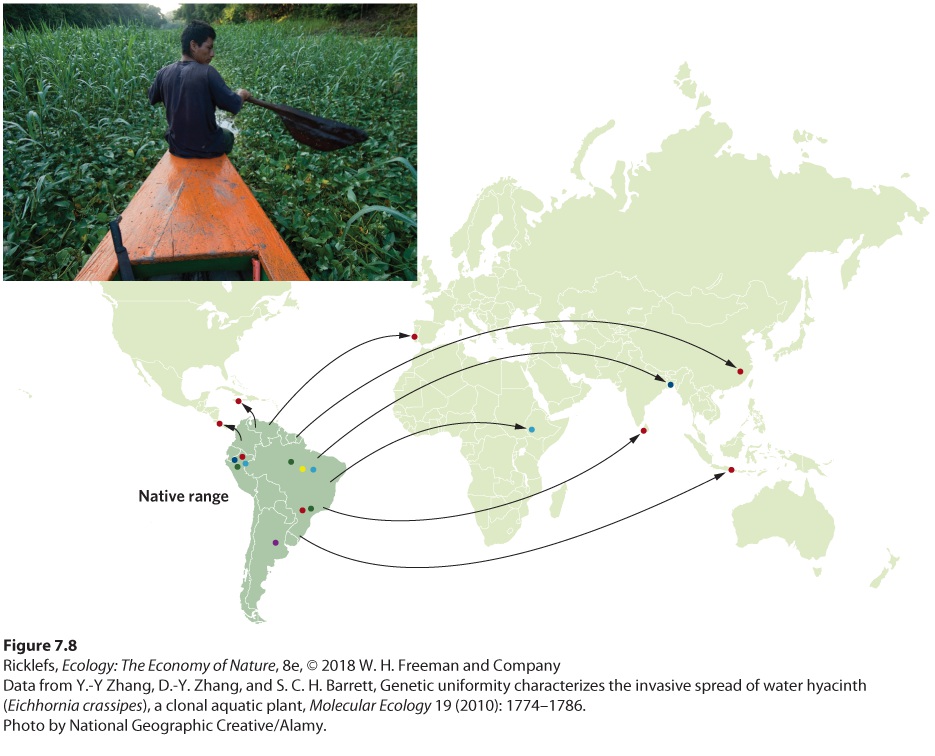
including a deadly pathogen that causes the disorder known as AA amyloidosis and kills up to 70 percent of cheetahs held in captivity. Founder Effects The founder effect occurs when a small number of individuals leave a large population to colonize a new area and bring with them only a small amount of genetic variation. Following the founding by this small population, genetic drift can cause additional reductions in genetic variation. The genetic variation remains low until enough time has passed to accumulate new mutations. The water hyacinth (Eichhornia crassipes), which has been introduced by humans to many parts of the world, provides an example of the founder effect. Water hyacinth is an aquatic plant that is native to South America. During the past 150 years, the plant has been either accidentally or intentionally introduced to many other parts of the world. Once introduced, it grows and spreads quite rapidly, dominating shallow-water areas and displacing native plants. Today, water hyacinth has become one of the most invasive plants in the world. Founder effect When a small number of individuals leave a large population to colonize a new area and bring with them only a small amount of genetic variation. Because most water hyacinth introductions are thought to have happened with just a few individuals, researchers wondered if the plant would show indications of founder effects in those parts of the world where it was not historically present. They sampled 1,140 plants from around the world and determined their genotypes. In 2010, they reported a single widespread genotype occurred in 71 percent of the sampled plants, and this genotype dominated 75 percent of all populations that were outside the plant’s native range, as shown in Figure 7.8. Moreover, 80 percent of all populations outside the native range were composed of a single genotype, whereas populations in the native range of South America had up to five different genotypes. This pattern suggests that there were few founders in the invaded regions of the world and that they carried with them only a small proportion of the genetic diversity of native populations in South America.
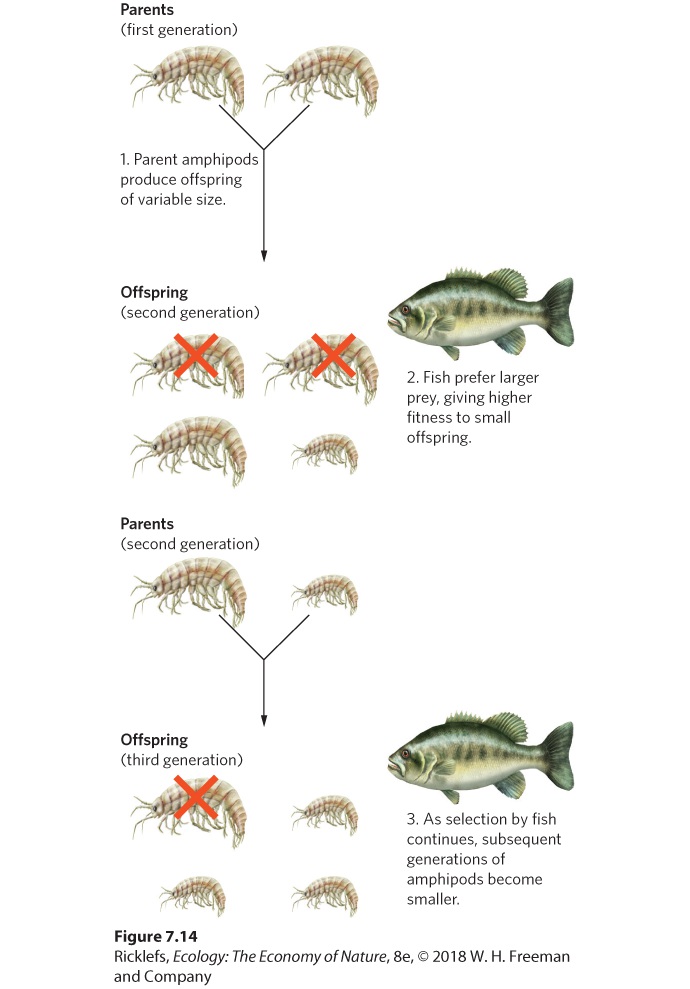
Figure 7.8 Evolution by the founder effect. Water hyacinth is an aquatic plant that was native to South America, where many different genotypes exist, as indicated by the differentcolored dots. Introductions around the world are thought to have occurred with low numbers of founders. Today, most populations existing outside of South America are represented by a single genotype.
Evolution Through Selection, A Nonrandom Process
The nonrandom process of selection also plays a substantial role in evolution. Selection is the process by which certain phenotypes are favored to survive and reproduce over other phenotypes. As we saw in the story of island birds at the beginning of this chapter, selection is a powerful force that can change the phenotypes (and therefore the gene frequencies) of a population. Depending on how the environment varies over time and space, selection can influence the distribution of traits in a population in three ways: stabilizing selection, directional selection, and disruptive selection. Selection The process by which certain phenotypes are favored to survive and reproduce over other phenotypes. Stabilizing Selection
When individuals with intermediate phenotypes have higher survival and reproductive success than those with extreme phenotypes, we call it stabilizing selection. As shown in Figure 7.9a, stabilizing selection begins with a relatively wide distribution of phenotypes, as illustrated by the orange line. After stabilizing selects for parents who possess intermediate phenotypes, their progeny have a more narrow distribution of phenotypes, as illustrated by the blue line. In doing so, it performs genetic housekeeping for a population, sweeping away harmful genetic variation. An example of stabilizing selection can be seen in selection for body mass in a species of bird from South Africa called the sociable weaver (Philetairus socius). Over an 8-year period, researchers marked nearly 1,000 adult birds and examined how a bird’s mass was related to its subsequent survival. As you can see in
Figure 7.9b, the mass of the adults in the study follows a normal distribution with a mean of approximately 29 g. The researchers then asked how well birds of different mass survive. When mass was graphed against survival, as shown in Figure 7.9c, they found that the smallest and largest birds survived poorly, whereas birds of intermediate mass survived the best. That is, selection favored the intermediate phenotype. When the environment of a population is relatively unchanging, stabilizing selection is the dominant type of selection. Because the average phenotype does not change, little evolutionary change takes place.
Figure 7.9 Stabilizing selection. (a) Stabilizing selection favors intermediate phenotypes and selects against both extremes. (b) In the sociable weaver bird, body size has a normal distribution. (c) The bird experiences stabilizing selection on body size because birds with
intermediate body sizes experience high survival, while birds with low and high body sizes have low survival. This selection for intermediate phenotypes would cause the next generation of birds to have a narrower distribution of phenotypes. Stabilizing selection When individuals with intermediate phenotypes have higher survival and reproductive success than those with extreme phenotypes. Directional Selection Directional selection, which occurs when an extreme phenotype experiences higher fitness than the average phenotype of the population, as shown in
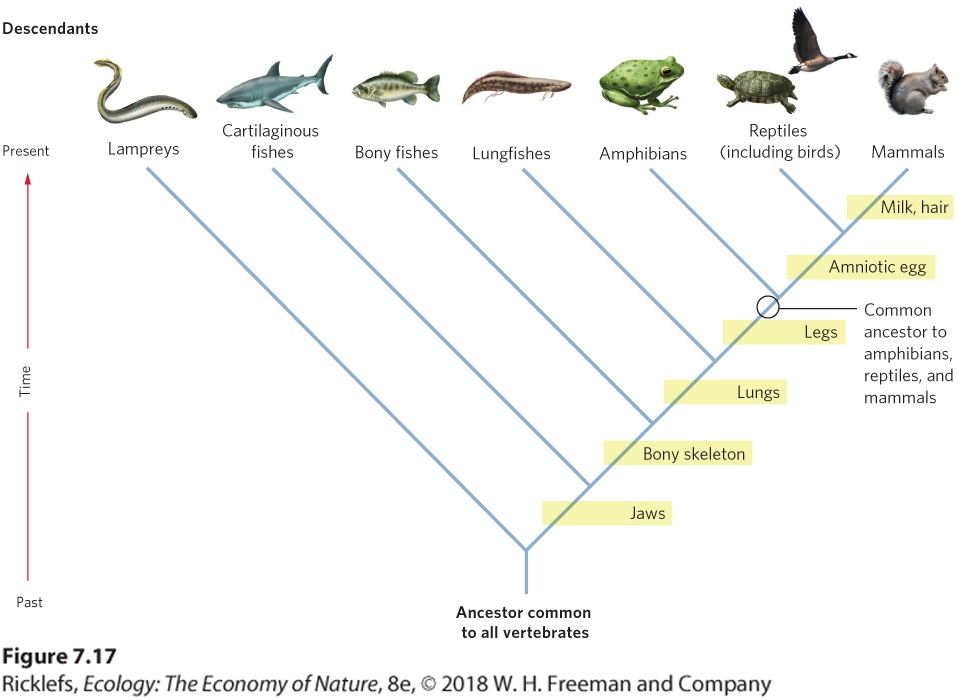
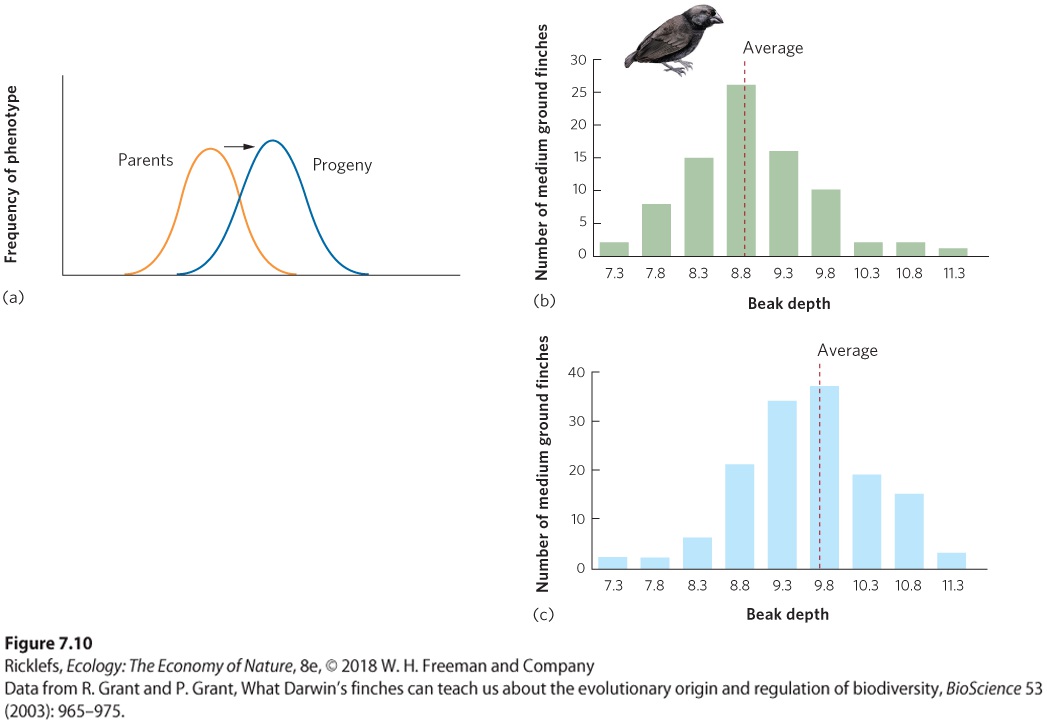
Figure 7.10a. In the medium ground finch (Geospiza fortis), for example, Peter and Rosemary Grant quantified the distribution of beak sizes in offspring hatched in 1976, which was just prior to a drought. As you can see in Figure 7.10b, the beak sizes of these offspring followed a normal distribution with a mean beak depth of 8.9 mm. Individuals with large beaks can generate the force needed to crack the largest seeds, whereas individuals with small beaks are better at handling the smallest seeds. When a drought came, although all seeds became less abundant, there were proportionately more large seeds remaining. The large seeds are harder to crack, so birds with deeper beaks were better able to feed on hard seeds and they had better survival. Because beak depth is a heritable trait, the offspring that were hatched in 1978 had deeper beaks, as shown in Figure 7.10c.
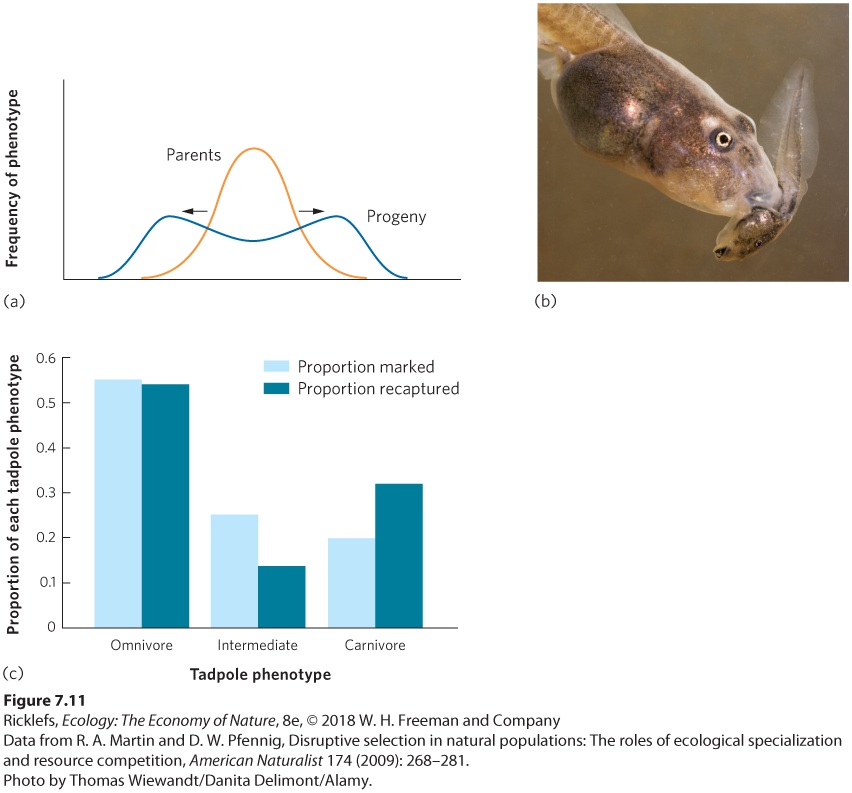
Figure 7.10 Directional selection. (a) Directional selection favors phenotypes of one extreme and selects against phenotypes of the other extreme. (b) Prior to a drought in 1976, the beak size in offspring of the medium ground finch had a mean depth of 8.9 mm, as indicated by the dashed red line. During the drought, when mostly large seeds were available, birds with larger beaks survived better. (c) Two years later, finch offspring had a mean beak depth of 9.7 mm, confirming that larger seeds caused directional selection for larger beaks. Directional selection When individuals with an extreme phenotype experience higher fitness than the average phenotype of the population. Disruptive Selection Under some circumstances, we see another type of selection known as disruptive selection. In disruptive selection, individuals with extreme phenotypes at either end of the distribution can have higher fitness than individuals with intermediate phenotypes. Disruptive selection is illustrated in Figure 7.11a. For example, tadpoles of the Mexican spadefoot toad (Spea multiplicata) can express a range of possible phenotypes that are related to what they eat. At one extreme is the omnivorous phenotype that has small jaw muscles, numerous little teeth, and a long intestine that makes it well suited for feeding on detritus. At the other extreme is the carnivorous phenotype that has large jaw muscles, a notched mouthpart, and a short intestine that makes it well suited for feeding on freshwater shrimp and cannibalizing conspecifics. Intermediate phenotypes are not well suited to
feed on either of the two food types. To test whether the tadpoles experienced disruptive selection, researchers collected more than 500 tadpoles from a desert pond, marked them to indicate their phenotype, and then returned them back to their pond. They sampled the pond again 8 days later to determine the survival of the three phenotypes. As you can see in Figure 7.11b, the omnivorous and carnivorous phenotypes survived relatively well, but the intermediate phenotypes—which had intermediate jaw muscles, tooth number, and intestine length—survived poorly. Because disruptive selection removes the intermediate phenotypes, it increases genetic and phenotypic variation within a population. In doing so, it creates a distribution of phenotypes with peaks toward both ends of the original distribution.
Figure 7.11 Disruptive selection. (a) Disruptive selection favors both extreme phenotypes and selects against intermediate phenotypes. (b) In the tadpoles of the spadefoot toad, an individual can have specialized mouthparts for carnivory and cannibalism (shown in this photo), omnivory, or an intermediate phenotype. (c) When more than 500 tadpoles of each phenotype were marked and released, about 10 percent were recovered. Of those recovered, the omnivores and carnivores experienced relatively high survival, while the intermediate phenotypes experienced lower survival.
##### Analyzing Ecology: Strength of Selection, Heritability, and Response to Selection
Disruptive selection When individuals with either extreme phenotype experience higher fitness than individuals with an intermediate phenotype.
Analyzing Ecology
Strength of Selection, Heritability, and Response to Selection Researchers often wish to know exactly how much selection will move the mean phenotype in a population. For example, if a plant breeder selects for larger tomato plants, she might want to know how much larger the next generation will be. Similarly, a government agency that regulates fishing might want to know if harvesting only the largest individuals might cause the population to evolve to be smaller in the next generation. Let’s consider the case of directional selection in which one end of the phenotypic distribution is favored. If there is selection for more extreme phenotypes and the phenotype has a genetic basis, directional selection will cause the mean phenotype to change. Can we determine exactly how much the mean phenotype will change in the next generation? To answer this question, we need to know both the strength of selection on the phenotype and the heritability of the phenotype. The strength of selection is the difference between the mean of the phenotypic distribution before selection and the mean after selection, measured in units of standard deviations (see Chapter 2). For example, imagine that we wanted to select for larger tomatoes. The phenotype (tomato mass) follows a normal distribution with a mean of 100 g and a standard deviation of 10 g. Now imagine that we select the upper end of the distribution and use these individuals to breed the next generation of tomatoes. If this selected group has a mean of 115 g, our selected group has a mean that is 1.5 standard deviations away from the mean of the entire population. Thus, the strength of selection is 1.5. Strength of selection
The difference between the mean of the phenotypic distribution before selection and the mean after selection, measured in units of standard deviations. We also know that phenotypes are the products of genes and the environment. If we wish to know how much the mean phenotype will change, we have to determine the proportion of the total phenotypic variation caused by genes, which is called the heritability. Heritability values can range between 0 and 1. If all of the phenotypic variation that we see in a normal distribution is due to the environment, the heritability is 0. If all of the phenotypic variation is due to genetic variation, the heritability is 1. By convention, the symbol for heritability is h2. (This notation can be confusing because nothing is being squared.) Heritability The proportion of the total phenotypic variation that is caused by genetic variation. Using the concepts of strength of selection and heritability, we can build an equation that describes how much a population will respond to selection in the next generation. Because a population’s response to selection is a function of the strength of selection and the heritability of the phenotype, R=S×h2 where R is the response to selection, S is the strength of selection, and h2 is the heritability. Using our tomato example, we can calculate how much larger the tomatoes should be in the next generation. If we select parents that are 1.5 standard deviations above the population mean and if the heritability is 0.33, then R=1.5×0.33=0.5 which means that the mean phenotype of the next generation of tomatoes will be 0.5 standard deviations—or 5 g—greater than that of the parent’s generation. YOUR TURN Given the following values for the strength of selection and heritability on tomato mass, calculate the expected response to selection
in units of standard deviations and grams: S h2 0.5 0.7 1.0 0.7 1.5 0.7 2.0 0.9 2.0 0.6 2.0 0.3 2.0 0.0 Based on your calculations, how is the response to selection affected by the strength of selection and heritability?
Concept Check
1. Compare and contrast the processes of bottleneck effects and founder effects. 2. How should stabilizing selection and disruptive selection affect the magnitude of phenotypic variation from one generation to the next? 3. How does heritability affect the response to selection?

#### 7.3 Microevolution Operates at the Population Level
7.3 Microevolution operates at the population level The random and nonrandom processes that cause evolution can operate at a variety of levels. The evolution of populations is known as microevolution, and it is pervasive. It is the process responsible for producing different breeds of cats, cattle, and dogs, and for producing distinct populations of wild organisms, including salmon, bears, and the flu virus. Microevolution is affected by both random process and selection. Selection at the microevolution level can be further divided into artificial selection and natural selection. Microevolution Evolution at the level of populations.
Artificial Selection
In his book On the Origin of Species, Charles Darwin discussed the wide variety of domesticated animals that humans have bred to produce particular suites of traits. In the case of dogs, for example, humans began by domesticating gray wolves. Over time they bred individuals that possessed particular characteristics, such as body size, coat color, and hunting ability. As shown in Figure 7.12, just a few centuries of breeding have created dog breeds with widely divergent phenotypes—from Saint Bernards to Chihuahuas. All of these dogs belong to the same species as the wolf and could potentially interbreed. Thanks to dog breeders, there is an excellent paper trail of exactly how the various breeds have been produced. This is an example of artificial selection, in which humans decide which individuals will breed, and the breeding is done with a preconceived goal for the traits desired in the population. Similar artificial selection has occurred to create numerous breeds of other domesticated animals, including cattle, sheep, pigs, and chickens.
Figure 7.12 Breeds of domestic dogs. Beginning with domesticated individuals of the gray wolf, humans have created a wide diversity of dog breeds through the process of artificial selection. Artificial selection Selection in which humans decide which individuals will breed and the breeding is done with a preconceived goal for the traits of the population. Artificial selection also has been applied to plants. One of the best-known examples is the artificial breeding of wild mustard (Brassica oleracea). As you can see in Figure 7.13, the wild mustard has been bred into a diversity of vegetables by selecting for unique stem, leaf, and flower traits. Today, the wild mustard can be consumed as cabbage, brussels sprouts, cauliflower,
Figure 7.13 Artificial selection on wild mustard. Over the years, plant breeders have produced a variety of common vegetables through artificially selecting for different traits of the wild mustard. Human practices can also cause artificial selection unintentionally, sometimes with detrimental consequences. For example, the widespread application of pesticides has caused resistance in more than 500 species of pests that harm food production and human health. Similarly, extensive use of antibiotics has caused many harmful human pathogens to evolve resistance to antibiotics, as described in “Drug-Resistant Tuberculosis” at the end of this chapter. In these cases, the role of evolutionary mechanisms is clear. When pesticides or antibiotics are targeted at millions of organisms, a small number of individuals commonly carry a mutation that confers resistance. Since only the mutants survive and the mutation is heritable, the next generation becomes more resistant. Darwin’s case for evolution by natural selection was strengthened by his observations of how artificial selection worked. He argued that if humans could produce such a wide variety of animal and plant breeds in only a few centuries through artificial selection, natural selection could certainly have
Natural Selection
A person conducting artificial selection typically has a particular suite of traits in mind, for example, increased milk production in cattle. This is not the case for natural selection, which favors any trait combination that provides higher fitness to an individual. Artificial and natural selection both operate by favoring certain traits over others. Both select on traits that are heritable; the difference lies in how traits are selected. There can be multiple ways to improve an individual’s fitness, and all of them are favored by natural selection, regardless of the resulting phenotype. For example, a prey organism might reduce its probability of being eaten either by hiding from predators so it is not detected or by growing spines so that it cannot be eaten. Both strategies are effective at improving the prey’s fitness, and both traits could be favored by natural selection. In artificial selection, humans determine the fitness of traits, and they often select attributes for particular purposes that would actually reduce the fitness of individuals if they lived in the natural environment. Most evolutionary biologists agree that the diversification of organisms over the long history of life on Earth has occurred primarily by natural selection. Natural selection is an ecological process: It occurs because of differences in reproductive success among individuals endowed with different form or function in a particular environment. That is, as individuals interact with their environment—including physical conditions, food resources, predators, other individuals of the same species, and so on—traits that lead to greater fitness in that environment are passed on. Evolution by natural selection is a common phenomenon in populations. For example, many predators cause selection on the traits of their prey. We can see this process in Figure 7.14 for fish that prey on amphipods, a tiny crustacean species. The parent amphipods produce an abundance of offspring that vary in size. However, fish prefer to consume the largest amphipods because they offer the highest amount of energy per unit effort. Smaller amphipods are more likely to survive, and since body size is a heritable trait in amphipods, subsequent generations evolve smaller body sizes.
Figure 7.14 Natural selection by predators on prey. The amphipod, a tiny crustacean, produces an abundance of offspring that vary in size. Predatory fish prefer to eat the largest amphipods, and this causes selection for small body size. One of the most striking demonstrations of microevolution is the case of the peppered moth (Biston betularia). During the early nineteenth century in England, most individuals of this moth were white with black spots, but occasionally there would be a dark, or melanistic, individual (Figure 7.15a).
Over the next hundred years, dark individuals became more common in forests near heavily industrialized regions, a phenomenon often referred to as industrial melanism. In regions that were not industrialized, the light phenotype still prevailed.
Figure 7.15 Selection by bird predation for different moth phenotypes. (a) In unpolluted forests, trees have light-colored bark and moths with the light phenotype are better camouflaged. (b) In polluted forests, trees have dark-colored bark and moths with the dark phenotype are better camouflaged. (c) When researchers placed both moth phenotypes on trees in polluted and unpolluted forests fewer light-colored moths were consumed by birds on unpolluted trees but fewer dark moths were consumed on polluted trees. Industrial melanism A phenomenon in which industrial activities cause habitats to become darker due to
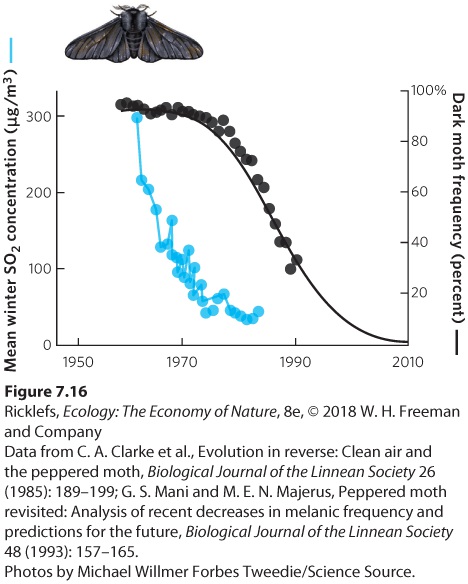
pollution and, as a result, individuals possessing darker phenotypes are favored by selection. Since melanism is an inherited trait, it seemed reasonable to suppose that the environment must have been altered to give dark forms a survival advantage over light forms. The specific agent of selection was easily identified. Peppered moths rest on trees during the day. Scientists observed that air pollution in industrial areas had darkened the trees with soot, as shown in Figure 7.15b, so they suspected that the predatory birds could see the lighter moths more easily. Because trees in nonpolluted regions were much lighter in color, the dark moths in these regions would be more visible. To test these hypotheses, equal numbers of light and dark moths were placed on tree trunks in polluted and unpolluted woods. As you can see in Figure 7.15c, when both types of moths were attached to the light-colored trees in unpolluted regions, birds consumed more dark moths. When both types of moths were attached to the dark-colored trees in polluted regions, birds consumed more light moths. This confirmed that the change in phenotypes observed over time in England reflected evolution of the population in response to changing environmental conditions. In recent years, as pollution control programs reduced the amount of soot in the air and improved the conditions of the forests, the frequencies of melanistic moths have decreased, as we would expect. Figure 7.16 shows data for the area around the industrial center of Kirby in northwestern England. As the amount of pollution declined—measured in terms of sulfur dioxide and shown as a blue line—the bark of the trees began to turn lighter. After two decades of decreasing pollution, the trees became lighter and the frequency of the dark form of the moth decreased from more than 90 percent of the population in 1970 to about 30 percent by 1990, as shown by the black line. The story of the melanistic moths demonstrates that microevolution can occur in a relatively short time.
Figure 7.16 Reversing the effects of pollution. As industries around Kirby, England, reduced the amount of sulfur dioxide pollution they emitted into the air, the color of the trees became lighter. After a decade of declining pollution, the frequency of the dark form of the peppered moth began to decline rapidly.
1. How is the domestication of wild mustard an example of artificial selection? 2. What traits are favored by natural selection? 3. What are the four required conditions for natural selection to occur?
#### 7.4 Macroevolution Operates at the Species Level and Higher Levels of Taxonomic Organization
7.4 Macroevolution operates at the species level and higher levels of taxonomic organization Whereas microevolution is a process that occurs at the level of the population, macroevolution is a process that occurs at higher levels of organization, including species, genera, families, orders, and phyla. For our purposes, we will restrict our discussion of macroevolution to the evolution of new species, a process known as speciation. The pattern of speciation over time can be illustrated using phylogenetic trees and speciation can occur in one of two ways: allopatric speciation or sympatric speciation. Macroevolution Evolution at higher levels of organization, including species, genera, families, orders, and phyla. Speciation The evolution of new species.
Phylogenetic Trees
Scientists can often document microevolution because it can happen in a relatively short time. In some cases, scientists have monitored wild populations over time to track the evolutionary process. In other cases, we have historical documents that trace the development of domesticated plants and animals. For example, most modern dog breeds are the result of artificial selection over the last three centuries and records show the older breeds that gave rise to newer breeds. Understanding how macroevolution has occurred, however, is a much more daunting challenge. Because we cannot travel back in time and there are no written records from millions of years ago, the true patterns of evolution can never be known for certain, although fossils can be helpful when examining the evolution of morphological traits. In the absence of more direct evidence, scientists work from the premise that species with the greatest number of traits in common are the most closely related. These traits can include shapes and sizes of structures of living and fossilized organisms as well as the ordering of nitrogenous bases in the DNA of different organisms. To map these relationships, scientists use phylogenetic trees, which are hypothesized patterns of relatedness among different groups such as populations, species, or genera. In essence, phylogenetic trees are
attempts to understand the order in which groups evolved from other groups.
Figure 7.17 shows a phylogenetic tree for several major groups of vertebrates. From this tree, you can see that all vertebrates share a common ancestor. Over time, this ancestor has given rise to fish, amphibians, mammals, and reptiles (including birds).
Figure 7.17 Phylogenetic tree. By looking for similarity in phenotypes and DNA, scientists can develop hypotheses about the relatedness of different groups of organisms. In this phylogenetic tree of the major vertebrate groups, the yellow boxes indicate points in time at which important new traits evolved. Phylogenetic trees Hypothesized patterns of relatedness among different groups such as populations, species, or genera.
Allopatric Speciation
Allopatric speciation is the evolution of new species through the process of geographic isolation. Imagine that we start with a single large population of a field mouse, shown as the first stage in Figure 7.18. At some point in time, a portion of the population is separated from the rest. This could occur because some individuals colonize a new island, such as the first finches to arrive in the Galápagos Islands from South America, which Charles Darwin described
in his theory of evolution by natural selection. Alternatively, the population could be divided by a geographic barrier, such as a new river that cuts through the middle of the terrestrial habitat, a rising mountain range that cannot be crossed, or a large lake that becomes two smaller lakes. In each case, the two populations are isolated from each other, as shown in Step 2 of the figure. They are no longer able to interbreed because of physical separation, so each population evolves independently. If one or both of the populations has few individuals, founder effects and genetic drift can strongly influence the direction in which that population evolves. When ecological conditions differ in the two isolated locations, natural selection will cause each population to evolve adaptations that improve fitness under local environmental conditions. Over time, as shown in Steps 3–5, the populations can become so different that they are no longer capable of interbreeding, even if they are brought back together. At this point, the two populations have evolved into different species.
Figure 7.18 Allopatric speciation. When geographic barriers divide populations, each evolves independently. Over time, the two populations can become so different that they are not able to interbreed. At this point, they have become two distinct species. Allopatric speciation
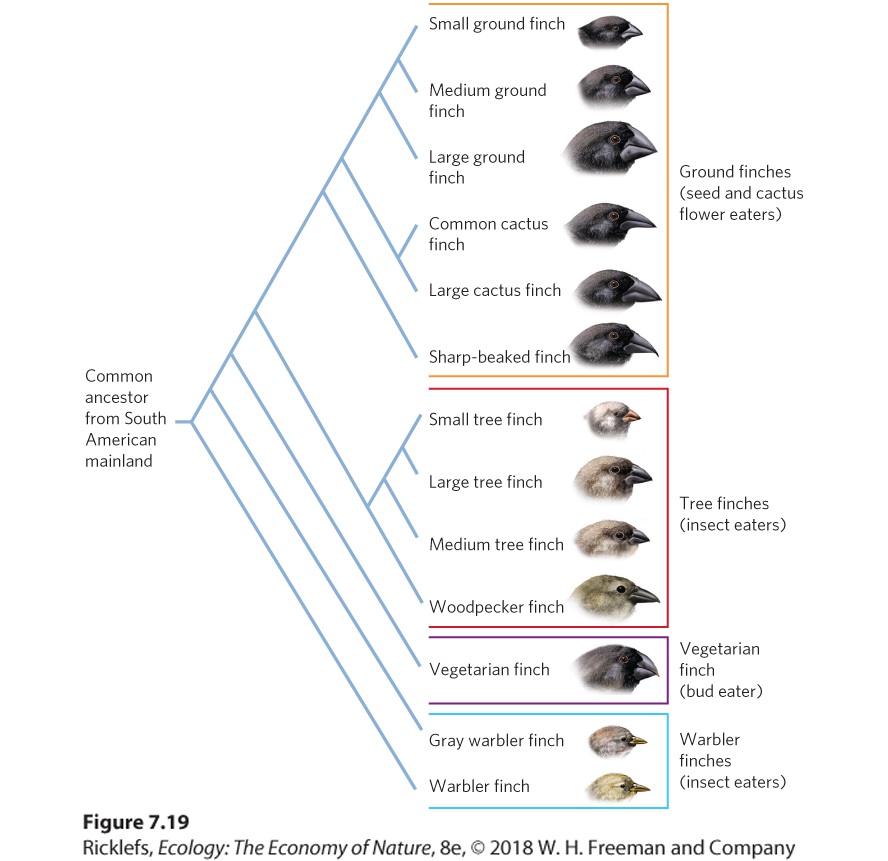
The evolution of new species through the process of geographic isolation. Allopatric speciation is thought to be the most common mechanism of speciation. Figure 7.19 shows the process for Darwin’s finches using a phylogenetic tree. Darwin hypothesized that the ancestor species of the finches he saw probably came from the mainland of South America. Once the ancestral finch species arrived in the Galápagos Islands, the population grew and eventually colonized many of the islands in the archipelago. The isolation and unique ecological conditions present on each island favored the process of allopatric speciation. Researchers hypothesize that these conditions gave rise to some of the 14 currently recognized species of finches in the Galápagos Islands, although other species of finches on the islands appear to have evolved through a different process known as sympatric speciation.
Figure 7.19 Allopatric speciation in Darwin’s finches. Through allopatric speciation, a single ancestor species from the South American mainland has evolved into 14 different species of finches on the Galápagos Islands. (Note that the phylogenetic study was done on
Sympatric Speciation
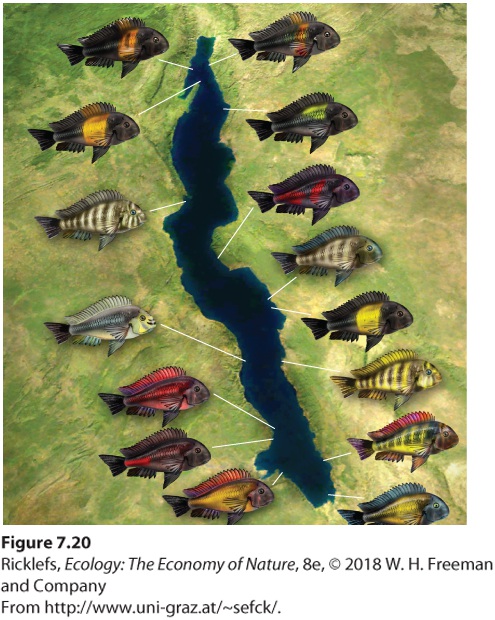
In contrast to allopatric speciation, sympatric speciation gives rise to new species without geographic isolation. In some cases, species evolve into a diversity of new species within a given location. An example of this is the group of cichlid fish species that live in Lake Tanganyika in eastern Africa. Over millions of years, a single ancestor fish has given rise to more than 200 unique species of fish, including insect eaters, fish eaters, and mollusk eaters (Figure 7.20). This massive amount of speciation appears to have been facilitated by the presence of many distinct habitats throughout the lake, such as rocky versus sandy shorelines. This small-scale habitat variation may have favored the evolution of different phenotypes that then led to the evolution of new species.
Figure 7.20 Sympatric speciation. More than 200 species of cichlid fishes of Lake Tanganyika have evolved from a single ancestor. Sympatric speciation The evolution of new species without geographic isolation.
A common way that sympatric speciation can occur in some types of organisms is through polyploidy. Polyploid species, which contain three or more sets of chromosomes, arise when homologous chromosomes fail to separate properly during meiosis, with the result that gametes are diploid rather than haploid. If a diploid egg, for example, is fertilized by a haploid sperm, the resulting zygote will contain three sets of chromosomes. At this point, the organism is a polyploid. Because it now has more than two sets of chromosomes, it is unable to breed with any diploid individuals. Thus, when a polyploid is formed, it instantly becomes a species that is genetically distinct from its parents. Several species of insects, snails, and salamanders are polyploid, as are 15 percent of all flowering plant species. Polyploid A species that contains three or more sets of chromosomes. An interesting example of polyploidy can be found in a group of salamanders. The blue-spotted salamander (Ambystoma laterale) and the Jefferson salamander (A. jeffersonianum) are both diploid species. As Figure 7.21 illustrates, at some point in the past, a blue-spotted salamander experienced incomplete meiosis and accidentally produced a diploid gamete. It then mated with a Jefferson salamander, which produced a normal haploid gamete. The resulting offspring was a triploid salamander that instantly became a distinct species known as Tremblay’s salamander (A. tremblayi). The Tremblay’s salamander is an all-female species that produces clonal daughters. These salamanders will breed with another salamander species to stimulate their own reproduction, but they can make daughters without incorporating DNA from any of the other species. If they do incorporate the haploid sperm from a male of the other species, their offspring can then carry four sets of chromosomes, which would make them tetraploids.
Figure 7.21 Polyploidy in salamanders. Triploid species can occur when one individual that experiences incomplete meiosis and produces a diploid gamete mates with another individual that experiences normal meiosis and produces a haploid gamete. The Tremblay’s salamander is an all-female, triploid species that arose through sympatric speciation from the mating of the blue-spotted salamander and the Jefferson salamander. Plant breeders have developed techniques to intentionally cause polyploidy to produce more desirable characteristics in plants. This is a form of artificial selection at the species level. By exposing plants to sudden cold temperatures at the time of reproduction, they can increase the chances of a plant producing gametes that are diploid rather than haploid. Polyploid plants tend to be larger and produce bigger flowers and fruits. Many beautiful flowers you see in a florist shop are the product of human-induced polyploidy. Many crops are also polyploids, including watermelons, bananas, strawberries, and wheat. As you can see in Figure 7.22, plant breeders used a species of wheat that has two sets of chromosomes to develop new species that have four or six chromosomes. The more sets of chromosomes in wheat, the larger the plant and the larger the seeds.
Figure 7.22 Polyploidy in wheat. Starting with a diploid species of wheat, plant breeders have bred new species that contain four or six sets of chromosomes. (a) The ancestral einkorn wheat (Triticum boeoticum) has two sets of chromosomes and small seeds. (b) Durum wheat (Triticum durum), which is used to make pasta, was bred to have four sets of chromosomes. It has medium-sized seeds. (c) Common wheat (Triticum aestivum), which is used for bread and other baked goods, was bred to have six sets of chromosomes. It has the largest seeds.
Concept Check
1. What is the assumption used when arranging species on a phylogenetic tree? 2. What is one requirement of allopatric speciation? 3. How does a polyploidy event immediately give rise to a new species?
Concepts
Death from tuberculosis. Tuberculosis patients being treated in a hospital in Tomsk, Russia. Tuberculosis, or TB, is a highly infectious disease caused by a bacterium (Mycobacterium tuberculosis). TB causes extensive tissue damage, weakness, night sweats, and bloody sputum. It is highly contagious; when an infected individual coughs or talks, bacteria are expelled and can survive in the air for several hours and infect other people. TB has killed people for thousands of years. In 2009, for example, researchers discovered that the preserved tissues of a woman who died 2,600 years ago and was mummified have genetic markers of the TB bacterium. Today, experts estimate that nearly one-third of the world’s human population is infected by the bacterium, although the bacterium remains inactive and does not cause any problems for most of these people. However, the bacterium does become active in about 9 million people every year. Around the world each year, 2 million people die of TB, which makes it the number one cause of death by infectious disease in the world. Fortunately, medical researchers have developed an inexpensive drug to
combat tuberculosis. While the drug has been highly effective in reducing the number of people infected with TB, the bacterium has started to evolve resistance to this drug. Drug-resistant tuberculosis is a rapidly growing problem around the world, particularly in Africa, Russia, and China. The reason is not a mystery. Bacteria can quickly grow to incredibly high numbers and, as you may recall from our discussion of evolution, very large populations are likely to have a few individuals that possess mutations. Occasionally, a mutation makes a bacterium more resistant and these mutations are heritable. Antibiotics represent a strong selective force that can quickly kill the vast majority of sensitive bacteria, thereby leaving resistant bacteria to flourish. One of the biggest contributors to the evolution of TB resistance is thought to be the behavior of TB patients. The typical drug treatment of TB requires taking pills daily for 1 year. Although many bacteria are killed early in the treatment, continued treatment helps eliminate every last pathogen. Sometimes patients stop taking their pills because they feel better after a few months or they simply lack the money to pay for an entire year of treatment. In either case, the most resistant bacteria remain in their bodies and they eventually become abundant, making the patient no longer responsive to the inexpensive TB drug. Drug-resistant TB is becoming a major problem. Researchers have developed new types of TB drugs to try to select different TB traits with the hope that evolving resistance to one drug will still make the pathogen susceptible to other drugs. However, there is now an increase in “Multiple Drug Resistant TB” or MDR-TB, a
strain of the bacterium that has evolved resistance to several different drugs. In Russia, for example, nearly 20 percent of all people infected with TB are carrying the MDR-TB strain. These strains are much harder for doctors to kill; the drugs required are 10 times more expensive than traditional ones, they may need to be taken for two years, and there are major side effects, including hearing loss. In 2016, researchers announced that a new regimen of seven drugs administered over nine months is turning out to be highly effective against MDR-TB. Even more sobering is the discovery of what is being called “Extensively Drug Resistant TB.” This version of TB has been detected in 45 countries, including Russia. Currently, drug regimens are only effective on 30 to 50 percent of patients, depending on the particular strain of bacteria and the state of the patient’s immune system. The evolution of TB resistance is an excellent example of why we need to understand the process of evolution. Knowing the sources of genetic variation and how selection operates on this variation helps us develop drug treatment programs that are better able to control pathogens without producing strains that have multiple drug resistance. SOURCES: Coghlan, A. 2016. Superfast therapy cracks multidrug-resistant tuberculosis. New Scientist, October 26. https://www.newscientist.com/article/2110555-superfast-therapy- cracks-multidrug-resistant-tuberculosis/. Altman, L. K. 2008. Drug-resistant TB rates soar in former Soviet regions. New York Times, February 27. http://www.nytimes.com/2008/02/27/health/27tb.html. Goozner, M. 2008. A report from the Russian front in the global fight
against drug-resistant tuberculosis. Scientific American, August 25. http://www.scientificamerican.com/article.cfm?id=siberia-drug- resistant-tuberculosis.
Summary of Chapter Concepts
7.1 The process of evolution depends on genetic variation. Among and within populations, genetic variation is caused by the presence of different alleles, which can be dominant, codominant, or recessive. Genetic variation can be generated through mutation or recombination. Key Terms: Deoxyribonucleic acid (DNA), Chromosomes, Alleles, Polygenic, Pleiotropy, Epistasis, Heterozygous, Homozygous, Codominant, Dominant, Recessive, Gene pool, Random assortment, Mutation, Recombination
7.2 Evolution can occur through random processes or through selection. The four random processes that cause evolution are mutation, genetic drift, bottleneck effects, and founder effects. Evolution can also occur by selection, which can be stabilizing, directional, or disruptive. Whether evolution occurs by random processes or by selection, scientists can use similarities in traits to arrange hypothesized patterns of relatedness among different groups on phylogenetic trees. Key Terms: Genetic drift, Bottleneck effect, Founder effect, Selection, Stabilizing selection, Directional selection, Disruptive selection, Strength of selection, Heritability
7.3 Microevolution operates at the population level. Populations can evolve due to artificial selection to produce breeds of domesticated animals and plants. Populations can also evolve due to natural selection, such as when predators selectively consume prey and when pesticides and antibiotics selectively kill the most sensitive individuals, allowing the most resistant individuals to survive and reproduce. Key Terms: Microevolution, Artificial selection, Industrial
7.4 Macroevolution operates at the species level and higher levels of taxonomic organization. The most common process causing macroevolution is allopatric speciation in which populations become geographically isolated and independently evolve to become distinct species over time. The less common process is sympatric speciation in which species become reproductively isolated without being geographically isolated, often by forming polyploids. Key Terms: Macroevolution, Speciation, Phylogenetic trees, Allopatric speciation, Sympatric speciation, Polyploid
Critical Thinking Questions
1. Given that mutations are rare in populations, how does a mutation spread through a population and become common? 2. Compare and contrast genetic variation caused by random assortment versus recombination. 3. The insecticide DDT has been widely used to control the mosquitoes that carry malaria. How would you explain the fact that many mosquito populations are now resistant to DDT? 4. How does the introduction of new individuals to a population help offset problems associated with genetic drift? 5. Compare and contrast evolution by artificial selection with evolution by natural selection. 6. Compare and contrast stabilizing, directional, and disruptive selection with regard to how each affects the mean phenotype of the population as well as the variance in the phenotype. 7. How does the breeding of domesticated animals provide evidence for the power of evolution on diverse phenotypes? 8. How is it that polyploidy can allow us to observe the evolution of a new species within one generation? 9. Distinguish between microevolution and macroevolution. 10. What is the difference between the processes involved in allopatric speciation and sympatric speciation?
#### Graphing the Data: Natural Selection of Finch Beaks
GRAPHING THE DATA Natural Selection of Finch Beaks The following table lists the frequency distributions of finch beak sizes, both before and after selection. Using a bar graph, plot the relationships between beak size and frequency. Then determine how much the mean beak size changed due to selection and decide which type of selection has occurred. Beak size (mm) Frequency before selection Frequency after selection R 10.00 0.00 0.00 10.20 0.00 0.00 10.40 0.02 0.00 10.60 0.04 0.00 10.80 0.08 0.00 11.00 0.16 0.00 11.20 0.20 0.00 11.40 0.20 0.00 11.60 0.16 0.02 11.80 0.08 0.04 12.00 0.04 0.08 12.20 0.02 0.16 12.40 0.00 0.20 12.60 0.00 0.20 12.80 0.00 0.16 13.00 0.00 0.08 13.20 0.00 0.04 13.40 0.00 0.02 13.60 0.00 0.00 13.80 0.00 0.00