8 Life Histories
8Life Histories Reproduction in frogs. While many frogs have a typical life of hatching into a tadpole that later metamorphoses into a frog, other species skip the entire tadpole stage by hatching into a tiny froglet. The Many Ways to Make a Frog When we think of a frog’s life, we often think of an egg hatching into a tadpole, the tadpole growing larger, and ultimately metamorphosing into a frog. Within this lifestyle, there is tremendous variation; some species of frogs spend up to 2 years as a tadpole, whereas other species spend as few as 10 days. In addition, the number of eggs laid can range from dozens to tens of thousands. However, this is only the beginning of the story of how different species of frogs live their lives. It turns out that the schedule of a frog’s life can take many different paths. While about half of all species lay eggs in the water and the eggs hatch into an aquatic tadpole, other species of frogs have evolved very different solutions. For example, some frogs hold their

eggs (and hatched tadpoles) on their back. The gastric-brooding frogs (Rheobatrachus spp.) hold their eggs in their stomach, which requires specialized adaptations, including the shutdown of acids in the stomach. Other species lay their eggs in foam nests on branches that hang over water; once the eggs hatch, the tadpoles drop into the water to begin an aquatic tadpole stage. Among all these species, tadpoles either feed themselves or they are given sufficient yolk from their mother to complete the tadpole stage without feeding. Still other species of frogs lay their eggs on land and completely skip the aquatic tadpole stage. In a process known as direct development, the embryo changes from an egg into a fully formed— albeit very tiny—frog (referred to as a froglet). For decades, scientists hypothesized that frogs with direct development likely had ancestors that began with the common aquatic tadpole stage, but over time they evolved to have terrestrial eggs, larvae that lived only on yolk, and then terrestrial eggs that could directly develop into froglets. “The schedule of a frog’s life can take many different paths.” To test this hypothesis, researchers in 2012 examined the reproductive patterns in 720 species of frogs to determine whether this hypothesis of gradual transitions was supported. Using genetic data, they could determine how the species were related to each other and the paths that frog evolution has taken over more than 200 million years. When they examined the patterns in the data, they found that many species of direct-developing frogs evolved by transitioning through the step of having terrestrial eggs. However, it was nearly as common to evolve from ancestors with fully aquatic eggs and tadpoles, thereby skipping the intermediate step of having terrestrial eggs. They also found that frog species with terrestrial eggs commonly produce fewer eggs, but the eggs are larger in diameter, which may provide space for more water and greater provisioning of
energy in the form of yolk. The diversity of frog reproduction highlights the notion that species on Earth have evolved a tremendous variety of reproductive strategies. As we shall see in this chapter, organisms have evolved a wide range of alternative strategies for growth, development, and reproduction, and these commonly reflect important fitness trade-offs. SOURCES: Gomez-Mestre, et al. 2012. Phylogenetic analyses reveal unexpected patterns in the evolution of reproductive modes in frogs. Evolution 66: 3687–3700.
Learning Objectives
After reading this chapter, you should be able to:
8.1 Describe life history traits as the schedule of an organism’s life.
8.2 Explain how life history traits are shaped by trade-offs.
8.3 Recognize that organisms differ in the number of times that they reproduce, but they all eventually become senescent.
8.4 Explain how life histories are sensitive to environmental conditions. As we have seen in previous chapters, organisms are generally well adapted to the conditions of their environments. Their form and function are influenced by physical and biological factors. Similarly, the strategies that organisms have evolved for sexual maturation, reproduction, and longevity are also shaped by natural selection. In this chapter, we will explore the wide array of strategies that species have evolved and the trade-offs among different traits.
#### 8.1 Life History Traits Represent the Schedule of an Organism’s Life
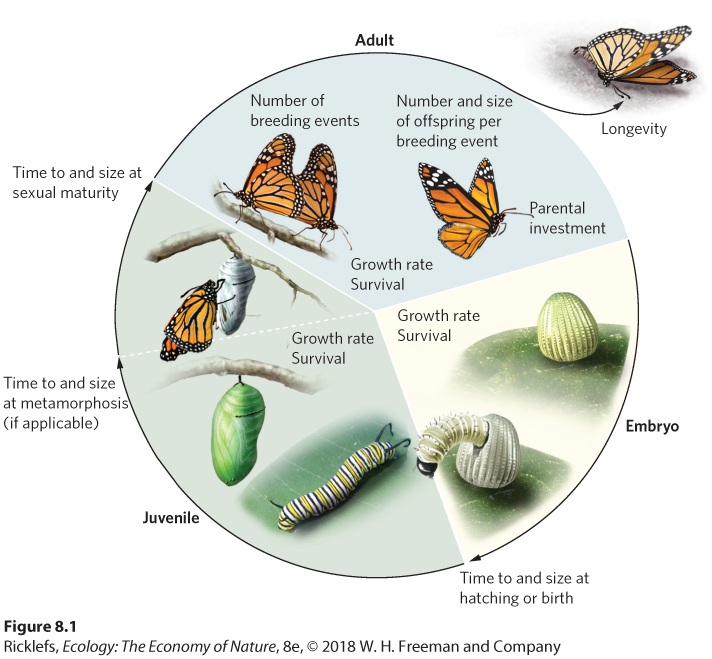
8.1 Life history traits represent the schedule of an organism’s life The schedule of an organism’s growth, development, reproduction, and survival makes up what ecologists call the life history of the organism. As you can see in Figure 8.1, the life history of an organism includes the traits connected to the birth or hatching of offspring. These include the time required to reach sexual maturity; fecundity, which is the number of offspring produced per reproductive episode; parity, which is the number of episodes of reproduction; parental investment, which is the amount of time and energy given to offspring; and longevity, or life expectancy, which is the life span of an organism. In essence, life history traits describe an organism’s strategy for obtaining evolutionary fitness throughout its lifetime. In addition, life history traits represent the combined effect of many morphological, behavioral, and physiological adaptations of organisms, all of which interact with environmental conditions to affect survival, growth, and reproduction. In this section, we will explore the wide range of life traits that exist in nature and how these traits are often organized into strategies that allow organisms to persist under different ecological conditions.
Figure 8.1 Life history traits. The schedule of an organism’s life begins as an embryo. It either hatches or is born at a particular size and time. During the subsequent juvenile stage, the organism grows and eventually becomes a sexually mature adult. To reach this stage, many species must first pass through metamorphosis. Adults can then reproduce in one or more breeding events with a particular level of fecundity, parental investment, and longevity. At all stages, species experience a characteristic amount of growth and some probability of surviving to the next stage. Life history The schedule of an organism’s growth, development, reproduction, and survival. Fecundity The number of offspring produced by an organism per reproductive episode. Parity The number of reproductive episodes an organism experiences. Parental investment The amount of time and energy given to an offspring by its parents. Longevity The life span of an organism. Also known as Life expectancy.
The Slow-To-Fast Life History Continuum
Life history traits vary widely among species and among populations within a species. An organism’s life history represents a solution to the problem of allocating limited time and resources to achieve maximum reproductive success. A remarkable fact about reproductive success is that the result is always nearly the same. On average, only one of the offspring that an individual produces lives to reproduce. In short, each individual replaces only itself. If this were not the case, populations would either dwindle to extinction because individuals fail to replace themselves or populations would continually expand. How organisms grow and produce offspring varies in all imaginable ways. A female sockeye salmon (Oncorhynchus nerka), after swimming up to 5,000 km from her Pacific Ocean feeding ground to the mouth of a coastal river in British Columbia, still faces an upriver journey of 1,000 km to her spawning ground. There, she lays thousands of eggs, and promptly dies, her body wasted from the exertion. A female African elephant gives birth to a single offspring at intervals of several years, lavishing intense care on her baby until it is old enough and large enough to fend for itself in the world of elephants. Thrushes, a group of birds that includes the American robin (Turdus migratorius), start to reproduce when they are 1 year old, and may produce several clutches—which are sets of eggs—each year, with each clutch containing three or four chicks. Adult thrushes rarely live beyond 3 or 4 years. In contrast, storm petrels, which are seabirds about the size of thrushes, do not begin to reproduce until they are 4 or 5 years old and then only rear a single chick each year. They may live for 30 or 40 years. This wide variation in life history traits among species has captured the interest of researchers who wish to understand ecological conditions that favor such disparate evolutionary outcomes. Two points can be made about this variation in life histories. First, life history traits often vary consistently with respect to life form, habitat, or conditions in the environment. Seed size, for example, is generally larger for trees than for grasses. Second, variation in one life history trait is often correlated with variation in other life history traits. For example, the number of offspring produced during a single breeding event is often negatively correlated with the size of the offspring. As a result, variations in many life history traits can be organized along a continuum of values. We can refer to one extreme as the “slow” end of the spectrum. At this
extreme, organisms such as elephants, albatrosses, giant tortoises, and oak trees require a long time to reach sexual maturity. They commonly have long life spans, low numbers of offspring, and a high parental investment in the energy given to their offspring, such as parental care, the amount of yolk in an egg, or the amount of energy stored in a seed. At the fast end of the spectrum are organisms such as fruit flies and herbaceous weedy plants, which exhibit short times to sexual maturity, high numbers of offspring, little parental investment, and short life spans.
Combinations of Life History Traits in Plants
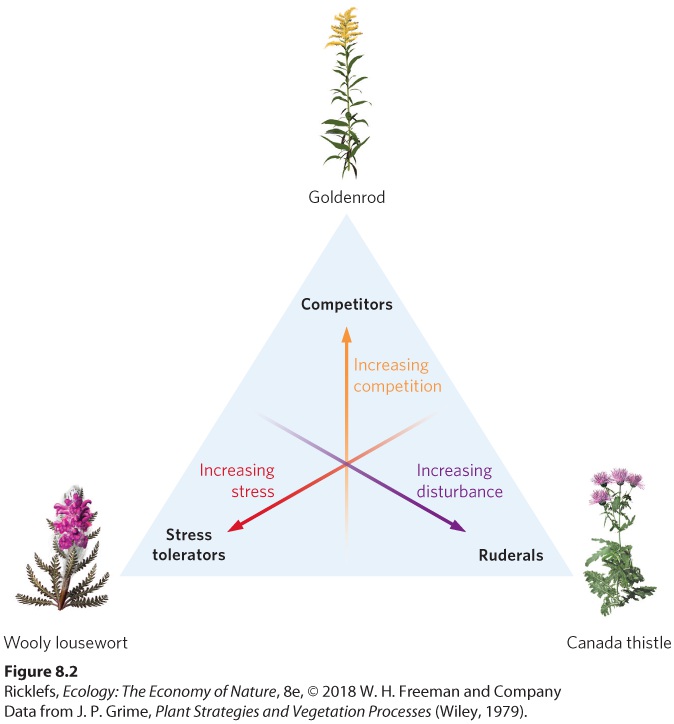
The English ecologist J. Philip Grime conceptualized the relationship between the life history traits and environmental conditions as a triangle, with each of the three points representing an extreme environmental condition: abiotic stress, competition, and the frequency of disturbances (see Figure 8.2). Grime proposed that plants functioning at the extremes of these three axes possessed combinations of traits that could be categorized as stress tolerators, competitors, or ruderals. Table 8.1 lists some of the major differences in these three plant strategies.
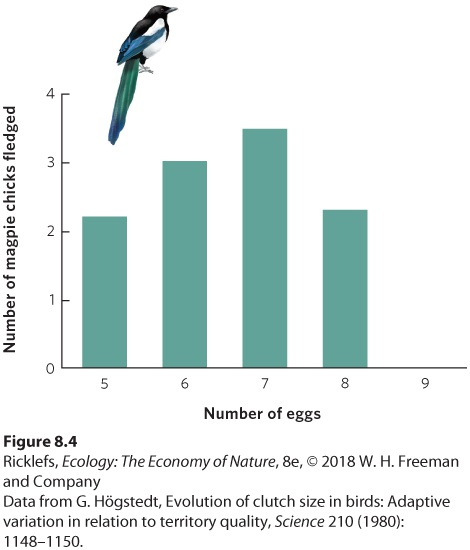
Figure 8.2 Combinations of life history traits in plants. Plants face the environmental challenges of competition, disturbance, and stress. Plant species living at each extreme environmental condition have evolved suites of life history traits that make the plants well suited to these environments.
TABLE 8.1 The Life History Traits of Plants That Are Positioned at the Environmental Extremes of Stress, Competition, and Disturbance Life history traits Stress tolerators Competitors Ruderals Potential growth rate Slow Fast Fast Age of sexual maturity Late Early Early Proportion of energy used to make seeds Low Low High Importance of vegetative reproduction Frequently important Often important Rarely important As their name implies, stress tolerators can survive and reproduce under extreme environmental conditions, such as very low water availability, very
cold temperatures, or high salt concentrations. For example, plants living in the tundra biome, such as the wooly lousewort (Pedicularis dasyantha), are typically small herbs that live for many years, grow very slowly, and achieve sexual maturity relatively late in life. Similarly, many plants that live in the desert, such as cactus, are stress tolerators because they can survive long periods of hot temperatures with no precipitation. Because growing from a seed is very difficult in such stressful environments, stress tolerators put little of their energy into seeds. Instead, they rely on vegetative reproduction, a form of asexual reproduction in which new plants develop from the roots and stems of existing plants. Where conditions for plant growth are less stressful, plants can evolve life history traits that fall along a continuum from competitors to ruderals. Without the abiotic stressors of extreme temperatures or an extreme lack of water and without frequent disturbances, plants can grow rapidly for long periods of time, and this creates more competition among plants for soil nutrients and light. Competitors can grow relatively quickly, achieve sexual maturity early in life, and still devote only a small proportion of their energy to seed production because they often spread by vegetative reproduction. Competitors also tend to grow to larger sizes and exhibit long life spans. In forests, most species of trees fit the competitor category. In abandoned farm fields, a strong competitor is the group of tall herbaceous plants known as goldenrods. At the other point of the triangle, with low stress and high frequency of disturbance, we find ruderals. These plants colonize disturbed patches of habitat, exhibit fast growth, early maturation, and use a high proportion of their energy to make seeds. Ruderals include many plants that we might call “weeds” in a garden, including dandelions (Taraxacum officinale), common ragweed (Ambrosia artemisiifolia), and Canada thistle (Cirsium arvense). Ruderals typically have seeds that are easily dispersed and that can persist in the environment for many years as they wait for favorable environmental conditions. This collection of traits allows ruderal plants to reproduce quickly and to disperse their seeds to other disturbed sites.
Concept Check
1. How would you differentiate fecundity and parity? 2. Using Grime’s categorization of plant life history traits, why might ruderals spread via easily dispersed seeds, whereas stress tolerators spread vegetatively? 3. Why do we expect that, on average, only one of the offspring that an individual produces survives to reproduce?
#### 8.2 Life History Traits Are Shaped by Trade-Offs
8.2 Life history traits are shaped by trade-offs If we consider the many types of life history traits, it would seem that an organism could have very high fitness if it could grow fast, achieve sexual maturity at an early age, reproduce at a high rate, and have a long life span. However, no organism has the best of all such life history traits, which highlights the fact that organisms face trade-offs. When one life history trait is favored, it often prevents the adoption of another advantageous life history trait. In some cases, there are physical constraints, such as the size of a mammal’s uterus, which places a limit on the total volume of offspring that can be produced at one time. Thus, a female can produce several small offspring, or a few large offspring, but not several large offspring. In other cases, the trade-off reflects the genetic makeup of the organism. Because some genes have multiple effects, selection that favors genes for one life history trait can cause other life history traits to change. For instance, in the plant known as the mouse-ear cress (Arabidopsis thaliana), artificial selection on genes that cause earlier flowering also results in reduced seed production because the genes have multiple effects. In still other cases, the trade-offs are the result of how an organism allocates a finite amount of time, energy, or nutrients. For example, the more time that a gazelle spends looking around for predators, the less time it has to spend looking for and eating food. In this section, we will discuss the principle of allocation and highlight some of the most common trade-offs that have been observed.
The Principle of Allocation
Organisms often have limited time, energy, and nutrients at their disposal. According to the principle of allocation, when these resources are devoted to one body structure, physiological function, or behavior, they cannot be allotted to another. As a result, natural selection will favor those individuals that allocate their resources in a way that achieves maximum fitness. Principle of allocation The observation that when resources are devoted to one body structure, physiological function, or behavior, they cannot be allotted to another. Selection on life history traits can be complex because when one trait is
altered, it often influences several components of survival and reproduction. As a result, the evolution of a particular life history trait can be understood only by considering the entire set of consequences. For example, an increase in the number of seeds an oak tree produces may contribute to higher fitness. However, if a higher number of seeds is achieved by making each seed smaller, and if smaller seeds experience lower survival, then producing more seeds could negatively affect the tree’s overall fitness. In this case, to achieve an outcome that maximizes overall fitness, evolution should favor a strategy that balances the trade-off between seed number and seed survival. From an evolutionary point of view, individuals exist to produce as many successful progeny as possible. Doing so, however, involves many allocation problems, including the timing of sexual maturity, the number of offspring to have at any one time, and the amount of parental care to bestow on the offspring. An optimized life history is one that resolves conflicts between the competing demands of survival and reproduction to the best advantage of the individual in terms of its fitness. Although it is widely believed that trade-offs constrain the specific combination of life history traits that a species can evolve, demonstrating this has proved difficult. In some cases, trade-offs can only be exposed by using experimental manipulations.
Offspring Number Versus Offspring Size
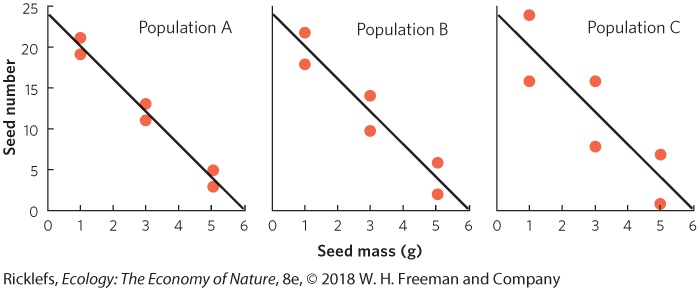
Most organisms face a trade-off between the number of offspring they can produce and the size of those offspring in a single reproductive event. As we noted for mammals, the number of offspring in any given pregnancy can only increase if the size of each individual offspring decreases. The trade-off between offspring number and offspring size for a given reproductive event can also be limited by energy and nutrients. An example of this can be seen in Figure 8.3, which illustrates the relationship between seed size and seed number in plants of the goldenrod genus (Solidago). Across populations and species, there is a negative correlation, demonstrating that goldenrod plants that produce more seeds also produce smaller seeds. While the trade-off between offspring number and offspring size can be seen in a number of species, the expected trade-off is often not observed. In some species, the number of offspring can be quite variable among individuals, but the size of offspring can be relatively constant. This suggests that selection often favors a uniform, perhaps even optimal, offspring size and that an individual able to
acquire additional energy can only use it to make greater numbers of offspring.
Figure 8.3 Offspring number versus offspring size. Among 14 populations and species of goldenrod plants, individuals that produce more seeds also produce smaller seeds, demonstrating that a trade-off exists between the two life history traits.
Offspring Number Versus Parental Care
The number of offspring produced in a single reproductive event can also cause a trade-off with the amount of parental care that can be provided. As the number of offspring increases, the efforts of the parents to provide food and protection will be increasingly spread thin. In a classic study of life history evolution, David Lack of Oxford University considered the number of offspring produced by songbirds. Lack observed that songbirds breeding in the tropics lay fewer eggs at a time—an average of 2 or 3 per nest—than songbirds breeding at higher latitudes,

##### Analyzing Ecology: Coefficients of Determination
which, depending on the species, typically lay 4 to 10 eggs. In 1947, he proposed that these different reproductive strategies evolved in response to differences between tropical and temperate environments. Lack recognized that birds could improve their overall reproductive success by increasing the number of eggs in each reproductive event, as long as a larger number of offspring did not cause a decrease in offspring survival. He hypothesized that the ability of parents to gather food for their young was limited and, if they could not provide enough, the offspring would be undernourished and therefore less likely to survive. We should therefore expect that parents produce the number of offspring that they can successfully feed. One difference between the low latitudes of the tropics and higher, temperate, latitudes is the number of daylight hours. Lack noted that parents at higher latitudes had more hours to gather food to feed their young. Therefore, assuming that the rate of food gathering is similar at low and high latitudes, he hypothesized that birds breeding at high latitudes could rear more offspring than birds breeding in the low latitudes of the tropics.
Analyzing Ecology
Coefficients of Determination When ecologists want to look for life history trade-offs, they commonly plot one life history variable against another life history variable and look for a negative relationship. We saw an example of this in the case of the goldenrod seeds; the researchers used a regression analysis to demonstrate a tradeoff between seed number and seed size. As we saw in Chapter 5, a regression analysis describes the mathematical relationship between two variables. For example, data points that tend to follow a straight line might be best described by using a linear regression described by the following equation: Y=mX+b where X and Y are the measured variables; m is the slope, which is negative in the case of the goldenrod example; and b is the intercept.
Although this equation reveals how one variable is associated with another variable, it does not tell us how strongly the two variables are related. For example, it would be valuable to know if the data points fit closely along the line or if the data points are highly variable around the line. We can answer this question using a statistic known as the coefficient of determination. The coefficient of determination, which is abbreviated as R2, is an index that tells us how well data fit to a line. Values can range from 0 to 1, with 0 indicating a very poor fit of the data and 1 indicating a perfect fit of the data. In terms of life history tradeoffs, higher R2 values indicate that a variation in one life history trait explains a large amount of the variation in the other life history trait. Coefficient of determination (R2) An index that tells us how well data fit to a line. Consider the following set of hypothetical data for plant seeds that all have the same linear relationship between seed mass and seed number, as given in Seed number = (-4 × Seed mass) + 24 Seed mass (g) Seed number for population A Seed number for population B Seed number for population C Ȳ = 12 Ȳ = 12 Ȳ = 12 For each set of data, we can graph the relationships and include the regression line, as shown in the graphs below. To calculate R2, we need to first calculate the mean Y value, which for all populations is a seed number of 12. We also need to determine the expected seed numbers if the data were to fall perfectly along the line.
Using the line equation and the above data for seed mass, the six expected seed numbers are 20, 20, 12, 12, 4, and 4. Next we need to calculate the total sums of squares, which is the sum of the squared differences between each observed seed number (yi) and the mean seed number (Ȳ): ∑ (yi-Y¯)2 Then we calculate the error sums of squares, which is the sum of the squared differences between each expected seed number (fi) and the observed seed number (yi): ∑ (yi-fi)2 Finally, we can calculate the value of R2 as follows: R2=1-(∑ (yi-fi)2∑ (yi-Y¯)2) For population A, we can calculate the value of R2 as R2=1-((21-20)2+(19-20)2+(13-12)2+(11-12)2+(5-4)2+(3-4)2(21-12)2+(19-12)2+(13-12)2+(11- 12)2+(5-2)2+(3-12)2)=1-(6262)=0.98 YOUR TURN Using the above formulas, calculate R2 for populations B and C. Based on your three R2 values, which set of data best fit the regression line? Which population gives you the strongest confidence that there is a negative relationship between seed size and seed number?
Lack made three important points. First, he stated that life history traits, such as the number of eggs laid in a nest, not only contribute to reproductive success but also influence evolutionary fitness. Second, he demonstrated that life histories vary consistently with respect to factors in the environment, such as the number of daylight hours available for gathering food for their young. This observation suggested that life history traits are molded by natural selection. Third, he hypothesized that the number of offspring parents can successfully rear is limited by food supply. To test this idea, one could add eggs to nests to create unnaturally high numbers of hatchlings. According to Lack’s hypothesis, parents should not be able to rear larger broods of chicks because they cannot gather the additional food that is required. Lack’s hypothesis has been supported by numerous experiments during the last several decades. For example, the European magpie (Pica pica) typically lays seven eggs in its nest. To determine whether this is the most fit strategy for the magpie, researchers manipulated the number of eggs in magpie nests by removing one or two eggs from several nests and adding these eggs to other nests. To control for their disturbance of the nests, they also switched eggs between nests without changing their number. The researchers then waited to see how many chicks could be raised to the fledgling stage, when the offspring can leave the nest. As shown in Figure 8.4, magpies that had fewer than seven eggs or more than seven eggs produced fewer fledgling birds. Nests that had eggs removed produced fewer fledglings because they started with fewer offspring. In contrast, nests with eggs added produced fewer total fledglings because the chicks had to share the food with more siblings. This competition with siblings caused the chicks to grow more slowly and experience higher mortality rates because the parents were unable to feed so many offspring. As Lack predicted, the number of eggs the magpie produces maximizes the number of offspring it can successfully raise.
Figure 8.4 Manipulating the number of eggs in a nest. In the European magpie, the adults typically lay seven eggs. When researchers removed one or two eggs or added one or two eggs, the number of chicks that survived to fledge declined. This suggests that the typical number of eggs laid may be the optimal number for the magpie. Despite the support for Lack’s hypothesis, research in 2015 revealed that the underlying premise that tropical birds lay fewer eggs than temperate birds because they are unable to provide as much food to their nestlings is not correct. By examining dozens of bird species across Arizona, Venezuela, and Malaysia, the study found that tropical birds had slower growth than temperate birds early in their nestling life, but they had faster growth later in their nestling life. Moreover, while temperate birds make more foraging trips to feed their total number of offspring, tropical birds make more foraging trips on a per-offspring basis, as shown in Figure 8.5. All this suggests that, contrary to Lack’s hypothesis, the tropical parents are quite capable of finding abundant food for their offspring. In addition, the tropical nestlings grow longer wings, and this makes them less susceptible to predators compared to temperate nestlings. In short, tropical birds produce a smaller number of higher-quality offspring, not because they are unable to feed more offspring but because they have a different overall strategy. The tropical birds’ strategy is to provide more food per nestling as a way to produce

offspring with higher survival rates after they leave the nest. In contrast, the temperate birds’ strategy is to produce more nestlings, at the cost of each nestling being of lower fitness.
Figure 8.5 Alternative life history strategies of temperate versus tropical birds. Researchers examined the relative rates of mortality and relative feeding effort of 72 bird species in temperate and tropical regions of the world. (a) Temperate birds, which typically lay one or two more eggs than closely related birds in the tropics, experience higher rates of adult mortality and conduct a greater number of foraging trips to feed their total clutch of offspring. (b) However, tropical birds make more foraging trips for their offspring on a per-nestling basis, resulting in the higher survival of those nestling while in the nest and throughout their later adult life. In all cases, relative rates are calculated after controlling for numbers other factors including differences in bird mass.
##### Fecundity and Parental Care versus Parental Survival
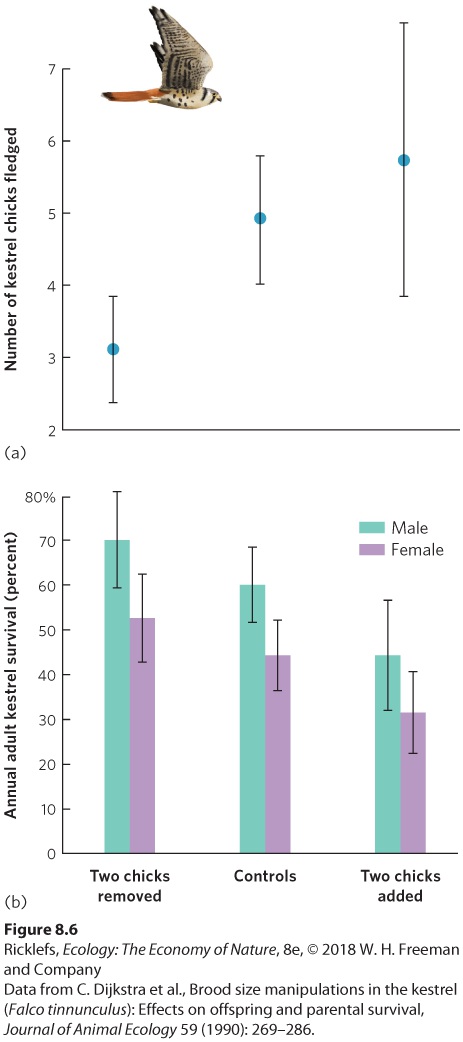
Survival
We saw how adding eggs to the nests of birds increases the effort for parents to supply food. Consequently, parents with small or intermediate size broods have the greatest fitness. Sometimes, however, having more mouths to feed stimulates parents to hunt harder for food for their chicks. In this case, an artificially enlarged brood might result in higher reproductive success in the short term. However, the additional parental effort can impose a cost on the parents that affects their subsequent fitness. European kestrels (Falco tinnunculus) provide an example of the trade-off between fecundity and parental survival. Kestrels are small falcons that feed on voles and shrews caught in open fields. While foraging requires a high rate of energy expenditure, the small mammals are so abundant that kestrel pairs normally can catch enough prey to feed their brood in a few hours each day. Kestrels lay an average of five eggs per brood. In one study, when chicks were about a week old, broods in a sample of nests were subjected to one of three manipulations: Investigators removed two chicks from a nest, switched chicks between nests without changing their number, or added two chicks to a nest. Investigators expected that parents of the artificially reduced and enlarged clutches would alter the amount of energy they expended looking for food for the chicks. While parents with fewer eggs ultimately had fewer chicks fledge than the control group, parents with additional eggs had more chicks fledge, as shown in Figure 8.6. However, in spite of the increased hunting efforts of their parents, chicks in the enlarged broods were somewhat undernourished and only 81 percent survived to fledge, compared with 98 percent survival in control nests and in nests with eggs removed. Consequently, the extra hunting effort to feed the additional two chicks only netted the parents an extra 0.8 chick, and this gain may have been diminished by the later deaths of some of the underweight fledglings. Moreover, the increased hunting efforts caused lower survival of these adults into the next breeding season. This means that increasing the number of offspring provides diminishing benefits to the parents in terms of the number of offspring that survive. Simultaneously, it causes greater adult mortality because parents must spend more energy securing food. At some point, the gains made by increasing current
reproduction, which requires a large increase in parental care, are offset by the increased adult mortality, which reduces the chance of future reproduction.
Figure 8.6 Parental care versus parental survival. (a) When researchers removed eggs from kestrel nests, fewer chicks fledged, but when researchers added eggs, more chicks fledged. Thus, having more eggs leads to more fledgling offspring. Error bars are standard
deviations. (b) However, adding more eggs causes a tradeoff. The parents work harder to feed the young, causing the adults to experience decreased survival. Error bars are standard errors.
Growth Versus Age of Sexual Maturity and Life Span
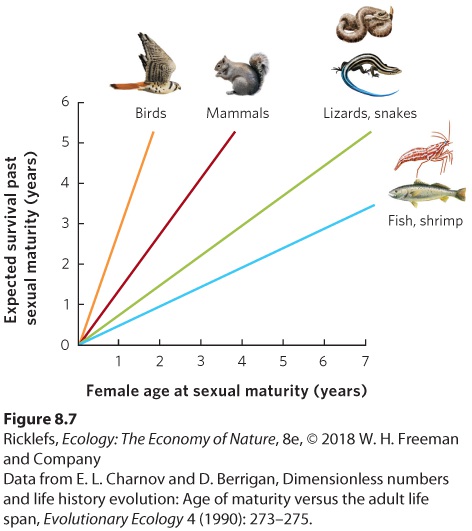
Organisms also commonly face a trade-off between putting their energy into growth or putting their energy into reproduction. In most species of birds and mammals, females grow to a particular size before they begin to reproduce. Once they initiate reproduction, females do not grow anymore—a phenomenon known as determinate growth. In contrast, many plants and invertebrates, as well as many fishes, reptiles, and amphibians, do not have a characteristic adult size. Rather, they continue to grow after initiating reproduction, a phenomenon known as indeterminate growth. Typically, indeterminate growth occurs at a decreasing rate over time. Determinate growth A growth pattern in which an individual does not grow any more once it initiates reproduction. Indeterminate growth A growth pattern in which an individual continues to grow after it initiates reproduction. Whether a species has determinate or indeterminate growth, the key feature shaping the trade-off between growth and reproduction is that larger females commonly produce more offspring. Because offspring production and growth draw on the same resources of assimilated energy and nutrients, increased fecundity during one year occurs at the cost of further growth that year. Moreover, for indeterminate growers, the reduced growth in one year can cause reduced fecundity in subsequent years. An organism with a long life expectancy should favor growth over fecundity during the early years of its life. In contrast, organisms with a short life expectancy should allocate their resources to producing eggs early in life rather than delaying reproduction and growing more. These predictions can be tested by examining the relationships among these life history traits across many different species and by conducting manipulative experiments on species in nature. Comparisons Across Species When we look across many different species, we see that long-lived
organisms typically begin to reproduce later in life than short-lived ones. Why is this the case? If an organism has a long life span and if delaying maturity allows an organism to grow larger and produce more offspring per year once reproduction begins, natural selection will favor delayed age of maturity in these organisms. An analysis of hundreds of populations and species of animals illustrates this relationship. As you can see in Figure 8.7, as the age of sexual maturity increases, there is an associated increase in the number of years that an animal will survive after reaching sexual maturity. Moreover, different taxonomic groups fall along different regression lines. For species that are expected to live 2 years after sexual maturity, birds and mammals have the shortest times to sexual maturity, whereas reptiles, fish, and shrimp have the longest times. This reflects the fact that endothermic animals can grow more rapidly than ectothermic animals. Among species with an age of maturity of 1 year, birds have the longest expected life spans after sexual maturity. This reflects the generally lower predation risk for birds due to their ability to fly.
Figure 8.7 Age of sexual maturity versus life span. Using hundreds of different populations and species, we can see that different groups of animals have different relationships between these two life history traits. Manipulative Experiments
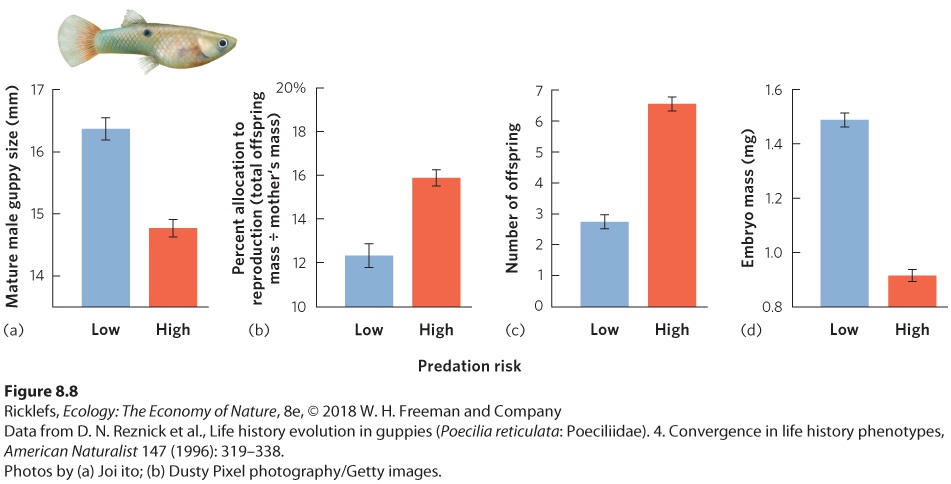
Another way to examine the trade-offs of growth, age of maturity, and life span is by conducting a manipulative experiment. The Trinidadian guppy (Poecilia reticulata), for instance, lives in the streams of Trinidad, a large island in the southern Caribbean Sea. In the lower reaches of these streams, the guppies live with a number of predatory fish species, including the pike cichlid (Crenocichla alta), which preys on adult guppies, and the smaller killifish (Rivulus hartii), which preys primarily on juvenile guppies. Because of this predation, these guppies have short life expectancies. However, at higher elevations, where guppies have been able to ascend numerous small waterfalls, they live in a relatively predator-free environment and have longer life expectancies. Figure 8.8 shows the life history traits of guppy populations. In populations that face high predation risk and short life spans, males mature at smaller sizes. Females allocate a larger percentage of their body mass to reproduction, they produce more offspring, and these offspring are smaller. As predicted, researchers found the opposite to be true for populations in the predator-free sections of the streams above the waterfalls: Males mature at larger sizes, and females allocate less of their body mass to reproduction and produce fewer, larger offspring.
Figure 8.8 Alternate life history strategies of guppies in Trinidad. Guppy populations in streams with a high risk of predation have shorter life spans, and those in streams with low predation risk have longer life spans. In response to this difference in longevity, guppies living in predator environments have evolved to (a) mature as smaller males, (b) allocate a greater fraction of energy to offspring, which is quantified as the percentage of the mother’s mass that is dedicated to the production of her offspring, (c) produce more offspring, and (d) produce smaller offspring. Error bars are standard deviations.
Researchers then conducted a manipulative experiment to test the hypothesis that increased mortality from predation altered life history strategies of the local guppy populations. They transplanted predators from the lower reaches of the streams to the areas above the waterfalls where predators did not historically exist. Within a few generations, the life histories of the populations above the waterfalls came to resemble those of the populations in the lower reaches of the stream. This finding not only confirmed the basic ideas about the optimization of life history patterns but also demonstrated that predation is a strong selective force in evolution.
Concept Check
1. Why are trade-offs among life history traits so commonly observed? 2. Why should organisms with low annual survival rates begin to reproduce at an early age? 3. Why do organisms face a fundamental trade-off between growth and fecundity?

#### 8.3 Organisms Differ in the Number of Times That They Reproduce, but They Eventually Become Senescent
8.3 Organisms differ in the number of times that they reproduce, but they eventually become senescent The number of times an individual reproduces during its lifetime varies a great deal among species. But in almost all species, individuals eventually experience a decline in body condition followed by death. For species that breed once, physiological decline and death follow rapidly. For species that breed many times, the decline in physiology comes more gradually. An interesting exception is in the case of bacteria, in which individuals do not always die. Instead, they undergo fission whereby a single-celled organism divides into two single-celled organisms. In this section, we will examine the life history strategies of breeding once versus breeding several times and investigate the causes of a gradual decline in physiology.
Semelparity and Iteroparity
Organisms can evolve to reproduce only once, a phenomenon known as semelparity, or multiple times throughout their life, a phenomenon known as iteroparity. Semelparity and iteroparity do not tell us whether an organism’s life history is annual, meaning a life span of one year, or perennial, referring to a life span of more than one year. For example, organisms that live for only one year can undertake more than one episode of reproduction, or even prolonged continuous reproduction, during that time. Iteroparity is a common life history that occurs in most species of birds, mammals, reptiles, and amphibians. Semelparity is relatively rare in vertebrate animals, but it occurs in insects and many species of plants. For example, semelparity is a common life history of crop plants, including wheat and corn. Semelparity When organisms reproduce only once during their life. Iteroparity When organisms reproduce multiple times during their life. Annual An organism that has a life span of one year. Perennial An organism that has a life span of more than one year.
Bamboos and Agaves Although most semelparous organisms are short-lived, the best-known cases of plants occur in the long-lived bamboos and agaves, two distinctly different groups. Most bamboos are plants of tropical or warm temperate climates, and they often form dense stands. Reproduction in bamboos does not appear to require substantial preparation or resources, as in the case of salmon. But bamboos probably have few opportunities for successful seed germination. Once a bamboo plant establishes itself in a disturbed habitat, it spreads for years by vegetative growth, continually sending up new stalks until the habitat in which it germinated is densely packed with bamboo shoots (Figure 8.9a).
Figure 8.9 Semelparous plants. (a) Bamboos and (b) agaves are two groups of plants that live for many years, reproduce one time, and then die. The flowering bamboos are from a location in Kyoto, Japan. The flowering agave is from Sodona, Arizona. In many species of bamboo, breeding is highly synchronous over large areas, such that every individual produces flowers and makes seeds in the same year. After reproduction, the future of the entire population rests with the crop of seeds. Synchronous breeding may facilitate fertilization in this wind-pollinated plant group and also may overwhelm seed predators, which cannot consume such a large crop of seeds. Some species of bamboo, such as Chinese bamboo (Phyllostachys bambusoides), have a 120-year cycle of germinating, growing, and then simultaneously flowering and producing
seeds. In contrast to bamboo plants, most species of agave plants live in arid climates with sparse and erratic rainfall. Ranging from the southwestern United States through Central America, agaves grow as a rosette of leaves over several years, with the duration of this growth varying among species. When the plant is ready to reproduce, it grows a gigantic flowering stalk that produces a large number of seeds (Figure 8.9b). The growth of the stalk is so rapid that it cannot be fully supported by photosynthesis in the stalk and uptake of water by the roots. Instead, the nutrients and water necessary for stalk growth are drawn from the leaves, which die soon after the seeds are produced. When we consider all the plants and animals that practice semelparity, we see that semelparity appears to arise when there is a massive amount of energy required for reproduction, such as the long migrations of salmon and the production of giant flowering stalks of agaves. These large energy requirements make it difficult for the individuals to survive after the reproductive event. Salmon Salmon are a group of species that vary a great deal in their parity. For example, Coho salmon (O. kisutch) lay eggs in rivers that empty into the North Pacific Ocean, from California to Alaska to eastern Russia. After growing in the river for a year, the fish swim out to the ocean, where they continue to eat and grow for 1 to 3 more years. When they are ready to reproduce, they migrate back to the same river where they hatched. The females make nests in the river bottom and deposit their eggs, which the males then fertilize with their sperm. Shortly after reproducing for the first time, both the male and female salmon rapidly lose strength and physiological abilities, and die. Several closely related species, including the chinook salmon (O. tshawytscha) and the sockeye salmon, also migrate to sea as juveniles, return for a single breeding event, and then die. In contrast, other salmon species, such as the rainbow trout, breed several times during their life. Some populations—known as resident rainbow trout —do not migrate out to the ocean but remain in freshwater rivers. Other populations of rainbow trout—commonly known as steelheads—migrate out to sea and return to the rivers to breed, much like the coho salmon. Unlike the
coho, however, steelheads migrate several times and breed each time they return to the freshwater rivers. Cicadas One of the most remarkable cases of semelparity in animals is the life cycle of periodical cicadas (Figure 8.10). Cicadas spend the first part of their life underground, where they obtain nutrients from the xylem tissue of plant roots and, after some time, emerge as adults. Their mating calls from the trees can be heard in the summer days in many parts of the Northern Hemisphere. Some cicada species have annual life cycles, while other species spend several years underground, with a fraction of them emerging each summer. Periodical cicadas, however, are different. They live as nymphs underground for 13 or 17 years and then emerge from the ground in synchrony to mate. The emergence of the periodical cicada is marked by nearly deafening noise as the males call to attract females during their brief mating period. The long life cycle gives the larvae time to grow to adulthood on a diet of low nutritional quality. The synchrony is probably a mechanism to overwhelm potential predators. Most of the occasional individuals that fail to emerge in synchrony come out a year earlier or later and are grabbed by predators attracted to the loud mating calls.
Figure 8.10 Periodical cicadas. The 13- and 17-year cicadas are semelparous insects. They spend many years underground, emerge as adults to breed, and die soon after. Pictured here is the pharaoh cicada (Magicicada septendecim), which is a species of 17-year cicada. Scientists long wondered how periodical cicadas know when to emerge from the ground. They speculated that the cicada might count the years by the warming and cooling of the soil, or by the physiological cycles of their hosts. Researchers performed a clever experiment by growing 17-year periodical cicadas on peach trees that had been artificially selected to drop their leaves and flower twice each year. The cicadas emerged after 17 fruiting seasons had passed, rather than 17 years, demonstrating that the cicadas are sensitive to the reproductive cycles of their hosts rather than annual physical changes in their environments. How they count to 17 remains a mystery.
Senescence
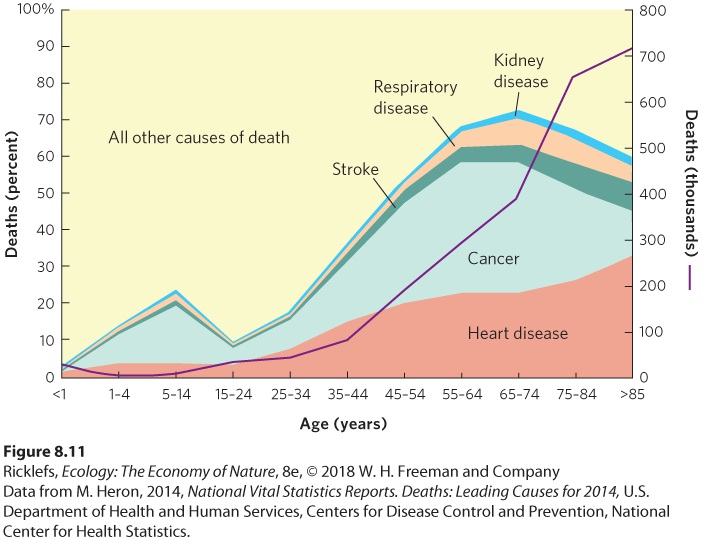
A few long-lived semelparous organisms die immediately after reproduction. However, iteroparous organisms experience a gradual deterioration in their physiological function over their lifetime. This leads to a gradual decrease in fecundity with an increase in the probability of mortality, a phenomenon known as senescence. Humans are an example of an organism that experiences senescence. Most physiological functions in humans decrease
between the ages of 30 and 85 years; for example, the rate of nerve conduction and basal metabolism decrease by 15 to 20 percent, the volume of blood circulated through the kidneys decreases by 55 to 60 percent, and maximum breathing capacity decreases by 60 to 65 percent. Over time the function of the immune system and other repair mechanisms also declines. Using data from the United States population in 2014, Figure 8.11 illustrates that the incidence of death from cancers and cardiovascular diseases rises sharply with age. Birth defects in offspring and infertility also occur with increasing prevalence in women after 30 years of age, and fertility decreases dramatically in men after 60 years.
Figure 8.11 Senescence. Throughout life, humans experience a gradual decrease in physiological function and an increase in cancer and cardiovascular diseases. These contribute to increased probability of mortality. Senescence A gradual decrease in fecundity and an increase in the probability of mortality. If maintaining high survival and reproduction would increase an individual’s fitness at any age, why does senescence exist? Studies of aging in a variety of animals demonstrate that senescence is an inevitable consequence of natural wear and tear. It is impossible to build a body that
will not wear out eventually, just as it is impossible to build an automobile that will not wear out. Senescence might simply reflect the accumulation of molecular defects that fail to be repaired. For example, ultraviolet radiation and highly reactive forms of oxygen break chemical bonds, macromolecules become inactivated, and DNA accumulates mutations. However, this wear and tear cannot be the entire explanation for patterns of aging because maximum longevity varies widely even among species of similar size and physiology. For instance, many small insectivorous bats live 10 to 20 years in captivity, whereas mice of similar size rarely live beyond 3 to 5 years. The rate of deterioration can be modified by a variety of physiological mechanisms that either prevent or repair damage. One major difference among groups that differ dramatically in life span is the cellular mechanisms for reducing the production of reactive forms of oxygen and for repairing damaged DNA and protein molecules. These appear to be better developed in long-lived animal species than in their short-lived relatives. Because these mechanisms are under genetic control, they can be modified by evolution. Mechanisms of prevention and repair require investments of time, energy, nutrients, and tissues. Therefore, the allocation of resources to these mechanisms depends on the expected life span of the individual. When a population has a low survival rate, selection should favor improvements in reproductive success at young ages and selection to delay senescence should be weak. In a population with a high survival rate, selection for delayed senescence should be strong. This prediction is consistent with observations of natural populations. For example, because bats and birds can fly to escape predators, they lead safer lives than do rodents of a similar size. As a result, the maximum potential life spans of birds and bats are much longer than the life spans of rodents. Storm petrels, a small seabird, have a body size and metabolic rate that is similar to those of many rodents. However, the storm petrel can live more than 40 years, whereas a rodent may only live 1 or 2 years. Because of their long life spans, birds and bats age more slowly than rodents of a similar size.
1. Compare and contrast semelparous and iteroparous life history
strategies. 2. What is a cause of senescence? 3. Across species, what is the expected relationship between longevity and senescence?
#### 8.4 Life Histories Are Sensitive to Environmental Conditions
8.4 Life histories are sensitive to environmental conditions As we saw in the discussion of phenotypic plasticity in Chapter 4, many traits show flexibility in response to different environmental conditions, and life history traits are no exception. As a result, researchers have continued to discover a fascinating array of life history traits that can be altered by changing environmental conditions.
Stimuli for Change
Many events in the life history of an organism are timed to match seasonal changes in the environment. The right timing is essential so behavior and physiology match changing environmental conditions. For instance, flowering plants must bloom when pollinators are present and most birds must breed when there is an abundance of food to feed their chicks. To get their timing right, organisms rely on various indirect cues in the environment. Virtually all organisms sense the amount of light that occurs each day, known as the photoperiod. Many can distinguish whether the photoperiod is getting shorter or longer. Within a single species, populations may be exposed to a variety of environmental conditions. Each population develops a particular response to the photoperiod in its environment. For example, consider the grass known as sideoats grama (Bouteloua curtipendula). Southern populations living at 30° N flower in autumn in response to a photoperiod of 13 hours per day. In contrast, northern populations living at 47° N flower in summer in response to a photoperiod that exceeds 16 hours each day. Because organisms experience longer days at higher latitudes, as we saw in Chapter 5, each population responds to a photoperiod that is indicative of summer at a particular latitude. Photoperiod The amount of light that occurs each day. Another example occurs in water fleas of the genus Daphnia. In Michigan, water fleas enter diapause in mid-September, when the photoperiod declines to fewer than 12 hours of sunlight, but related species in Alaska enter diapause in mid-August, when the photoperiod decreases to fewer than 20
hours of sunlight. Water fleas never see 20-hour days in Michigan, but Alaskan water fleas would perish from the cold if they waited for 12-hour days before entering diapause. From this we see that the critical stimulus for these organisms is the change in environmental conditions associated with a particular photoperiod that is relevant to a particular latitude. The sensitivity of individuals to these cues has been adjusted by natural selection so that the individual’s response to an environmental cue is well matched to the environmental condition.
The Effects of Resources
Many types of organisms undergo dramatic life history changes during the course of their development. One of the most striking changes is the process of metamorphosis in which a larva changes into a juvenile or adult organism. Metamorphosis can be seen in many species of insects and amphibians, as in the transformation from tadpole to frog. Organisms that metamorphose have a wide range of timing options. Environmental conditions that influence timing include the amount of resources available, the temperature, and the presence of enemies. Let’s consider the different options for timing of metamorphosis by looking at the two growth curves in Figure 8.12. These curves represent the change in the mass of the barking treefrog (Hyla gratiosa) raised under conditions of high or low food availability On any particular day early in life, the individuals raised under high food availability have a larger mass than those raised under low food availability. As time progresses, an individual with high food availability is able to metamorphose at a relatively large mass and young age. An individual with low food availability cannot achieve the same combination of mass and age, but it can follow several alternative strategies. It might wait to metamorphose when it achieves the same mass as individuals raised under high food availability, as shown by point D in the figure, though it will take longer to achieve that mass and the delay in reproduction could reduce its fitness. Alternatively, it might metamorphose at the same age as individuals raised under high food availability, as shown by point B in the figure, even though it will be significantly smaller. The drawback of this strategy is that a smaller size at metamorphosis makes the organism more vulnerable to predation. For most metamorphosing organisms, the optimum solution is usually a compromise between these two
strategies. So an organism with low food availability typically metamorphoses at an older age and smaller mass, as shown by the decision of the barking treefrog at point C in the figure.
Figure 8.12 Alternative growth curves of a metamorphosing organism. The amount of food available can affect an organism’s mass and age at metamorphosis. In the barking treefrog, an individual living under high food is able to metamorphose at a large mass and young age, represented by point A. An individual living under low food conditions could achieve the same age at metamorphosis if it emerges at a smaller mass, for example, at point B. It could achieve the same mass at metamorphosis if it took longer to metamorphose, for example, at point D. In reality, the tadpoles reach a compromise and metamorphose at a somewhat smaller mass and a somewhat later age, as indicated by point C. Error bars are standard deviations.
The Effects of Predation
The risk of predation is also an important factor in affecting the life history of organisms. As we saw in Chapter 4, predation can affect a wide range of life history traits, including time to and size at hatching, time to and size at metamorphosis, and the time to and size at sexual maturity. One of the more remarkable effects of predators is their impact on the embryos of many species of aquatic organisms. In a variety of groups, including fish and amphibians, the embryo that develops inside an egg can detect the presence
of an egg predator. Many embryos sense the chemical odors that predators emit, while other embryos can detect the vibrations produced by predators. When predators are detected, the embryos can speed up their time of hatching in an attempt to leave the egg before the predator eats it. For example, the red-eyed treefrog (Agalychis callidryas) lives in Central America and the adults lay their eggs on leaves that hang over water. When the embryos have developed sufficiently, they hatch and drop into the water. Should a cat-eyed snake (Leptodeira septentrionalis) appear, however, the frog embryos sense the vibrations of the approaching snake and begin to hatch earlier than usual and drop into the water to avoid the snake (Figure 8.13). However, this response comes at the cost of hatching at a smaller size that can make the hatchling tadpoles more susceptible to predators living in the water. Therefore, when egg predators are not present, the embryo stays in the egg longer and hatches at a larger and safer size.
Figure 8.13 Hatching early in response to predators. As the cat-eyed snake begins to attack the eggs of the red-eyed treefrog, the embryos are stimulated to hatch early. Note the
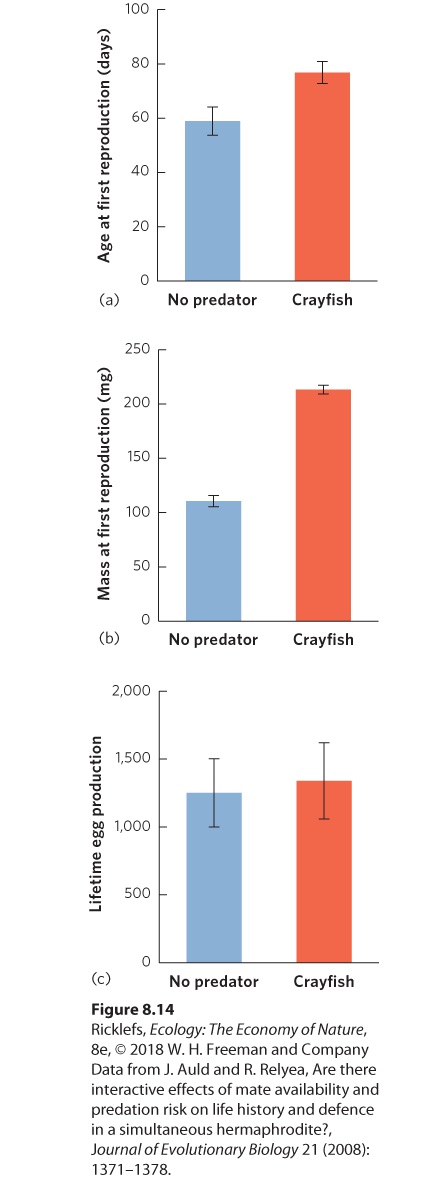
tadpole escaping the snake’s attempt to eat the egg in which the tadpole lived. This photo was taken at the Corcovado National Park in Costa Rica. Studies of metamorphosing animals also find that predators commonly play an important role in affecting the size and time at which metamorphosis takes place. For example, many high-elevation streams of western Colorado contain trout, an important predator of mayfly larvae, whereas other streams lack trout. Larval mayflies living with trout metamorphose and leave the streams earlier and at smaller sizes than mayflies in comparable streams that lack trout. Growth rates in the two types of streams are similar, so the difference in the time to and size at metamorphosis is entirely the result of predation risk. Predators can also affect when organisms achieve sexual maturity. Several species of freshwater snails, for example, face higher risks of predation when they are small. As a result, when predators are present, a snail is likely to do better if it delays reproduction and uses its energy to grow to a larger size before reproducing, as shown in Figure 8.14 a–b. Once it has grown to a safer size, it can reproduce. Although this strategy can improve the snail’s probability of survival in the presence of predators, the cost of delaying sexual maturity can be reduced fecundity. However, once the predatorinduced snails begin reproducing, they can produce more eggs in each clutch because they have larger bodies and they can live longer. In these cases, the snails can achieve the same lifetime fecundity as snails raised without predators, as shown in Figure 8.14c.
Figure 8.14 Snail life history strategies are altered in the presence of predators. When snails detect the smell of predatory crayfish in the water, (a) they delay their age at first
production in favor of (b) increasing their mass at first reproduction. Because snails living with the scent of predators experience an extended longevity, their (c) lifetime production of offspring is similar to that of snails not living with predators.
The Effects of Global Warming
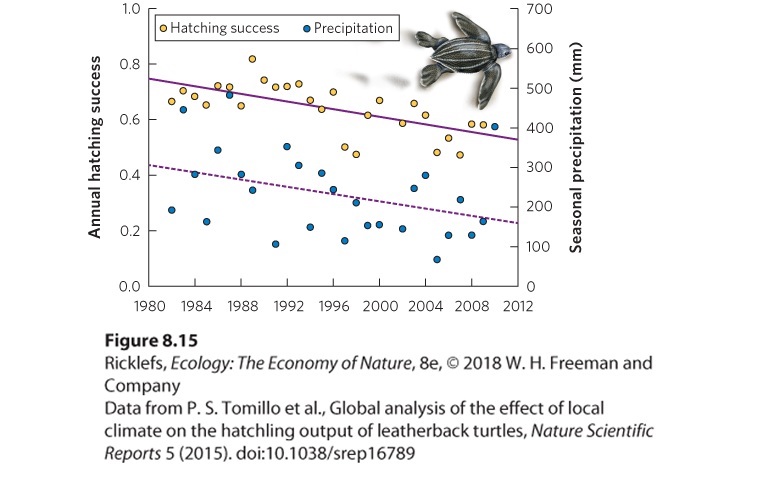
We have seen how the life history of organisms responds to different environmental conditions found in nature. Over the past 100 years, human activity has caused a warming trend on Earth. In many regions, the difference in temperature is relatively small—a rise of 1 °C or 2 °C. However, even small changes in temperature can have substantial impact on an organism’s physiological processes, as we saw in Chapter 2. During the past decade, researchers have begun to discover that the increase in global temperatures has caused changes in the breeding times of many animals and plants. Animal Breeding Researchers interested in the effect of global climate change on the life histories of animals have focused on the breeding times of numerous species. A change in breeding dates has been observed in several species of amphibians. In Britain, researchers monitored three species of frogs and three species of salamanders for 17 years. At the end of this period, they found that two of the three frog species monitored were breeding 2 to 3 weeks earlier and all three species of salamanders were breeding 5 to 7 weeks earlier. These changes in breeding times were correlated with the average maximum temperatures that occurred just prior to breeding, which generally increased over the 17 years. A similar study of North American amphibians, however, failed to find a relationship between changes in mean maximum air temperature over time and the initiation of breeding. At the present time, researchers do not know why amphibians in different regions of the world show different responses to global warming. While global climate change includes warming temperatures, it also includes changes in the pattern of precipitation. Researchers studying leatherback sea turtle (Dermochelys coriacea) breeding examined how the hatchling survive in light of changing precipitation patterns that have been observed at four sites around the world during the past several decades (1982‒2010). In some sites, no relationship existed between precipitation and
the proportion of turtle eggs that hatched. At other sites, however, there was a clear decline in precipitation over the decades as well as a decline in the proportion of turtle eggs that hatched (Figure 8.15). Such data suggest that the impacts of global climate change can vary among different geographic locations on Earth.
Figure 8.15 Hatching survival in leatherback sea turtles. At one of four sites where sea turtles breed, researchers monitored precipitation and the proportion of turtle eggs that hatched from 1982 to 2010. Plant Flowering Plants are also susceptible to climate change, which has the potential to alter the initiation of flower production. One of the longest studies began in the nineteenth century with the writer Henry David Thoreau, who is best known for having spent a year in a small cabin at Walden Pond in Concord, Massachusetts, and for his numerous essays about the natural world. Thoreau kept data on more than 500 species of flowering plants in Concord. From 1852 to 1858, he took notes on the dates when each plant species first began flowering in Concord. After Thoreau’s death, a local shopkeeper continued his work by observing the first flowering times of more than 700 plant species. More recently, two ecologists realized that the data could help them determine whether long-term changes in global temperatures might be associated with changes in the initial flowering times of plants. Because flowering time is sensitive to temperature
as well as to photoperiod, they predicted that warmer global temperatures would cause plants to flower earlier today than in Thoreau’s time. To test their hypothesis, they collected data for plant flowering times in Concord from 2003 to 2006. The researchers reported that over the 154-year period from 1852 to 2006, local temperatures in Concord had increased by 2.4 °C. You can see these data in Figure 8.16. They also found that for the 43 most common species of plants, flowering time today is an average of 7 days earlier than in Thoreau’s time. Interestingly, not all plants responded to the temperature change in the same way. In some species, initial flowering time remained unchanged, perhaps because these species use day length as their cue for flowering and day length has not changed. Other species, such as highbush blueberry (Vaccinium corymbosum) and yellow wood sorrel (Oxalis europaea), flower 3 to 4 weeks earlier now than they did in 1852. These unique data collected over a century and a half indicate that a seemingly small change in average annual temperature has been associated with dramatic changes in initial flowering time.
Figure 8.16 First flowering dates for plants in Concord, Massachusetts. (a) The mean flowering time today is 7 days earlier than in the 1850s. Error bars are standard errors. (b) The variation in first flowering time is associated with the mean temperature of the 1 or 2 months preceding each species’ flowering time. Consequences of Altered Breeding Events The changing breeding seasons of plants and animals in response to global warming do not by themselves cause any problems to the species that are responding. Problems can arise, however, when a species depends on the environment to provide the necessary
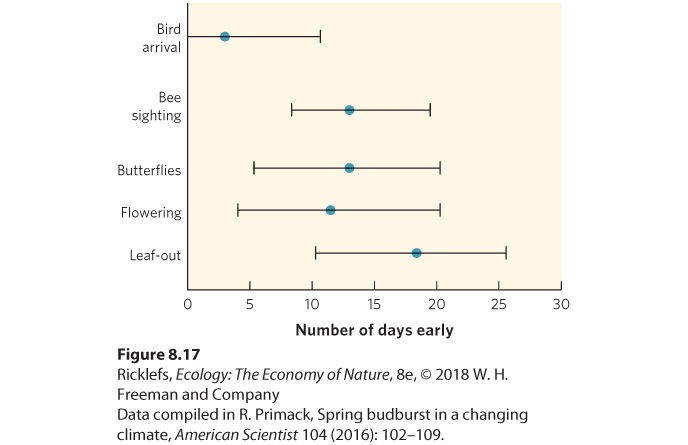
resources with an altered breeding season. The pied flycatcher (Ficedula hypoleuca), for example, is a bird that breeds in Europe each spring. In 1980, researchers in the Netherlands found that the date of egg hatching of the flycatcher began just a few days before the peak abundance of caterpillars, which are a major prey item for the flycatcher chicks. As spring temperatures warmed over the next 2 decades, however, tree leaves appeared 2 weeks earlier and the caterpillars reached their peak abundance 2 weeks earlier. The pied flycatcher, however, retained its normal date of egg hatching, which was 2 weeks later than the new time for peak caterpillar abundance. As a result, the chicks of the flycatcher no longer have a major source of food and the pied flycatcher population has declined by 90 percent. A similar phenomenon was observed by researchers who revisited the plants that Henry David Thoreau studied. From a series of studies conducted in the Boston area, these researchers noted a variety of organismal responses to warming temperatures. As you can see in Figure 8.17, the mean time for trees to leaf out is currently about 18 days earlier than Thoreau observed in the 1850s. The same team examined several other organisms’ responses to warming temperatures as well. Butterflies are being seen about 12 days earlier, but the arrival of migratory birds is only about 3 days earlier. As a result, changes in the historic timing of birds, insects, and plants beginning their spring activities have occurred, and this has the potential to dramatically alter how these species interact in regard to predation, herbivory, and pollination.
Figure 8.17 Effects of global warming on birds, plants, and insects. Since the 1850s in the Boston area, the dates of birds arriving, bees and butterflies being first observed, and plants flowering and leafing out from their winter buds are now all earlier than in the past. Error bars are 95% confidence intervals. From our discussion of life history traits, you can see that natural selection has favored a wide variety of strategies for the life history of different species. Different life histories evolve as a result of different selection pressures on traits such as mortality, fecundity, and longevity combined with a considerable number of potential trade-offs among life history traits. As is true for other traits, the genes that code for these traits interact with the environments that organism’s experience, ultimately to produce the life history traits of individuals.
Concept Check
1. Why is it important for organisms to adjust the timing of their life history to seasonal changes in the environment? 2. How do different resource levels affect the decision of when, and at what mass, an animal should metamorphose? 3. Why do prey species often delay their time to first reproduction in favor of increasing their mass at first reproduction?
###### Selecting on Life Histories with Commercial Fishing
Concepts
Selecting on Life Histories with Commercial Fishing Human selection on the life history of fish. For several decades, commercial fishing vessels, like this boat in Alaska, have harvested the largest individuals and thereby caused unintentional selection for smaller fish. Because of this selection, some species of fish now achieve sexual maturity earlier. Throughout this chapter, we have seen how natural selection has shaped the evolution of life histories by favoring individuals that are best suited to their environment. But what if a change in the selection process favors individuals with different suites of traits? This is precisely what happens in many commercial fisheries because they harvest only the largest individuals. For many years, this seemed like a wise way to manage the exploitation of wild populations because it protected the small individuals, allowing them a chance to grow. It is also
a common practice for state agencies to set a minimum size for fish, such as bass and salmon, that anglers are permitted to keep. Based on our discussion in this chapter, you should be able to predict what will happen to the life history traits of species that experience a great deal of fishing pressure, particularly from large commercial fishing boats that can collect thousands of fish. When the smallest fish are either not caught or are thrown back into the water, we impose a high mortality rate on large adults and leave the smaller, younger fish behind for breeding. As we have seen in this chapter, high adult mortality favors the evolution of smaller adult sizes, earlier times to maturity, higher fecundity, and shorter life span. During the past two decades, researchers investigated whether large-scale fishing can cause unintended evolution of fish life histories, and they have confirmed that commercial fishing imposes substantial selection on fished populations. In keeping with the requirements for evolution, there is sufficient heritability present in fish populations for the selection to cause a change in subsequent generations. For example, in the 1930s and 1940s, the northeast Atlantic Cod (Gadus morhua) had a median age at maturity that ranged between 9 and 11 years. By the 1960s and 1970s, age at maturity ranged between 7 and 9 years. The data collected by commercial fishing boats do not typically contain information on other life history traits such as fecundity and longevity. However, based on our knowledge of common life history trade-offs, it is reasonable to assume that an increase in adult mortality and a decline in age at maturity imposed by fishing practices coincide with increases in size-adjusted fecundity
and declines in longevity. One challenge in determining the effect of fishing pressure on life history is that it can cause other significant changes. As we noted earlier in this chapter, environmental changes—such as resource levels—can also affect life history traits. For example, fishing from commercial fishing boats could reduce competition among the remaining fish, which allows for a faster time to sexual maturity. In some cases, researchers cannot differentiate between the environmental induction of life history changes from reduced competition and the evolution of life history changes. In other cases, however, scientists have been able to document that a fish population held at low numbers for several decades continues to exhibit life history changes over time. In such cases, the changes in life history are more likely to be the result of evolution by artificial selection. To help clarify the role of selection on fish life histories without the complications of other environmental changes, including reduced competition, researchers in 2013 reported the results of an experiment in which they selected guppies by culling large individuals, culling small individuals, and culling the population randomly. After only three generations, the researchers found an evolutionary response to the selection on body size. Fish populations that experienced a culling of the largest individuals reached a smaller size at maturity and a younger age at maturity. In contrast, populations that experienced a culling of the smallest individuals reached a larger size at maturity and an older age at maturity.
Experimental selection on the life history of fish. Over six generations, populations selected for larger size evolved to become larger than the unselected control population, whereas populations selected for smaller size evolved to become smaller. Error bars are standard errors of the mean. The impact of human selection on natural populations is not limited to fish; similar impacts have been found in hunted mammals and some plants. In all these cases, identifying the factors that naturally cause the evolution of life histories and the ways in which various life history traits trade off with each other has helped fisheries managers understand how the human harvest of wild populations can have unintended consequences. SOURCES: van Wijk, S. J., et al. 2013. Experimental harvesting of fish populations drives genetically based shifts in body size and maturation. Frontiers in Ecology and the Environment 11: 181– 187. Law, R. 2000. Fishing, selection, and phenotypic evolution. CIES Journal of Marine Science 57: 659–668.
Darimont, C. T., et al. 2009. Human predators outpace other agents of trait change in the wild. Proceedings of the National Academy of Science 106: 952–954.
Summary of Chapter Concepts
8.1 Life history traits represent the schedule of an organism’s life. Species differ in a wide range of traits that help determine their fitness throughout their life. These traits are under the influence of natural selection and often evolve in particular combinations. Key Terms: Life history, Fecundity, Parity, Parental investment, Longevity
8.2 Life history traits are shaped by trade-offs. Trade-offs can occur because of physical constraints, time or energetic constraints that affect allocation, or genetic correlations that cause selection favoring one trait to come at the cost of another trait. Common tradeoffs include offspring number versus offspring size, and growth versus reproduction. Key Terms: Principle of allocation, Coefficient of determination (R2), Determinate growth, Indeterminate growth
8.3 Organisms differ in the number of times that they reproduce, but they all eventually become senescent. Semelparous organisms breed once, whereas iteroparous organisms breed more than once. All organisms ultimately experience a decay in physiological function followed by death. In semelparous organisms, this decay in function occurs rapidly after reproduction. In iteroparous organisms, the decay in function can be very gradual. Key Terms: Semelparity, Iteroparity, Annual, Perennial, Senescence
8.4 Life histories are sensitive to environmental conditions. Life history traits are the product of genes and environments interacting. Some of the most common environmental influences on life history traits include variation in resources and predators, both of which can
induce substantial changes in the life history of organisms. Current anthropogenic changes in the environment can also induce life history changes. Key Term: Photoperiod
Critical Thinking Questions
1. How should selection for greater longevity affect time to maturity and size at maturity? 2. Compare and contrast the concept of a regression versus a coefficient of determination. 3. What factors might favor the evolution of semelparity versus iteroparity in different species of salmon? 4. Compare and contrast the two leading arguments regarding why temperate bird species lay one or two more eggs than closely related tropical bird species. 5. Given your knowledge of how predators and resources can affect life history traits, hypothesize how the four combinations of high resources, low resources, predator presence, and predator absence would affect the size at maturity of a prey species. 6. Why might natural selection act more strongly on traits that improve reproductive success early in life rather than later in life? 7. Give two reasons why a mammal such as a gray wolf could face a trade-off between offspring number and offspring size. 8. What is the mechanism by which parental care can increase current fitness but decrease future fitness? 9. Why might an organism use cues such as photoperiod to predict the future state of its environment? 10. In terms of environmental cues, hypothesize why some, but not all, species of plants flower earlier in association with warmer spring temperatures.
#### Graphing the Data: Lizard Offspring Number Versus Offspring Mass 1
GRAPHING THE DATA Lizard Offspring Number Versus Offspring Mass The common lizard (Lacerta vivipara) can produce 2 to 15 offspring in a single breeding event. Using the following data, create a scatter graph to illustrate the relationship between the number of lizard offspring and the average mass of a lizard’s offspring. (You can review scatterplots in the graphing appendix.) After graphing the relationship, describe the relationship in words. Offspring number Offspring mass (g)