10 Social Behaviors
10Social Behaviors Leaf-cutter ants. Through an extensive division of labor, leaf-cutter ants work together to bring pieces of leaves back to their nest. In this photo, a large worker ant carries a cut piece of leaf, while a much smaller worker ant rides on top to discourage parasitoid flies from attacking the worker. The Life of a Fungus Farmer The leaf-cutter ant is an extraordinary farmer. Living in colonies of several million individuals, these ants leave the colony each day to harvest leaves from the surrounding forest. Using their sharp mandibles, they slice through leaves to cut off pieces that are many times larger than their bodies. They then carry the pieces back to the nest, which can rise several meters out of the ground, sink tens of meters below the ground, and expand more than 100 m wide underground. Back at the nest, the ants consume the sap in the leaves, but they do not eat the leaves. Instead, they use the leaves to grow a specialized species of fungus that they consume.

There are more than 40 species of leaf-cutter ants, which live primarily in Mexico, Central America, and South America. Like honeybees, the leaf-cutter ants form enormous societies of cooperating individuals. An ant colony normally has a single queen that can live for 10 to 20 years. Early in her life, the queen participates in a mating flight with males. The sperm she receives are then held inside her body and remain viable for the rest of her life. She uses them sparingly when fertilizing eggs to make daughters; in some species, the queen releases only one or two sperm per egg. Occasionally, she lays unfertilized eggs to make sons whose only function is to mate with other queens. The millions of individuals in the nest are daughters of the same queen and sisters to each other. They all forgo reproduction. Daughters in this ant society are the workers and the division of labor among the workers is amazingly complex. Scientists estimate that there are nearly 30 different jobs for the leaf-cutter ants and they have determined that different workers are suited to different jobs. The worker caste is composed of several subcastes, known as minims, minors, mediae, and majors. Ants in each subcaste differ dramatically in size and shape. The largest workers, the majors, can be 200 times more massive than the smallest workers, the minims. The differences in subcastes are thought to be a phenotypically plastic response to different diets the ants receive as larvae. In addition to size and shape differences, the jobs of a given worker can also change during its lifetime. For example, when workers are young, they spend most of their time inside the nest, where they build tunnels, cool the nest, and raise the larvae. When large leaf pieces arrive at the nest, another group of workers cut the leaves into smaller pieces and a third group of workers mash the leaf pieces into tiny bits. The smallest workers bring a strand of fungus to the mashed bits and tend the fungus gardens, a job that includes removing other species of pathogenic fungi, which can harm the ants. In 2016, researchers discovered that in one species of leaf-cutter ant (Mycocepurus smithii), the workers even adjust the nutrient mixture given to the fungus. Since the ants prefer to eat the fungal threads and not the reproductive mushrooms that the fungus produces, the workers adjust the amount of
carbohydrates and proteins that they give to the fungus to prevent the fungus from making mushrooms. “There are nearly 30 different jobs for the worker ants and different workers are suited to different jobs.” The oldest and largest ants act as soldiers and go out of the nest to collect leaves. The process of leaf collection is complex. Some ants climb into trees and cut large pieces of leaves that drop to the ground, while others relay leaf pieces back to the nest. When workers first begin leaf cutting, they have razor-sharp jaws, called mandibles, that are very effective at cutting tough leaves. Researchers have recently discovered that these mandibles become dull over time, causing older workers to take twice as long to cut a leaf. When an ant’s cutting performance declines, it shifts jobs from leaf cutting to leaf carrying. This change in job allows older individuals to continue contributing to the ant society. The division of labor also helps the group to maintain a high foraging efficiency. While the ants benefit from living in a very large group, the large colonies and conspicuous foraging behavior make them quite noticeable to their enemies. For example, a species of parasitoid fly that is specialized to hunt large leaf-cutter ants lays its eggs inside the necks of foraging ants. To reduce the risk of these attacks, leaf-cutter ants have evolved several tactics. Smaller individuals, which are less attractive to the fly, forage during the day. At night, when the flies do not hunt, the larger, more effective foragers go out to collect leaves. Small workers also guard larger ants by riding on leaves that the larger ants carry. When a fly approaches, these tiny guards prevent the fly from laying its eggs on the neck of the large ant that is carrying the leaf. When not repelling attacks, these hitchhiking individuals clean the leaf of undesirable microorganisms. The leaf-cutter ants illustrate an extreme case of social behavior
and living in groups. As we will see in this chapter, social groups are common in the animal world and there are both costs and benefits to social living that vary with the ecological conditions under which a species lives. SOURCES: Shik, J. Z., E. B. Gomez, P. W. Kooij, J. C. Santos, W. T. Wcislo, and J. J. Boomsma. 2016. Nutrition mediates the expression of cultivar–farmer conflict in a fungus-growing ant. Proceedings of the National Academy of Sciences 113:10121–10126. Hölldobler, B., and E. O. Wilson. 2011. The Leafcutter Ants. Norton. Schofield, R. M. S., et al. 2011. Leaf-cutter ants with worn mandibles cut half as fast, spend twice the energy, and tend to carry instead of cut. Behavioral Ecology and Sociobiology 65:969–982.
Learning Objectives
After reading this chapter, you should be able to:

10.1 Explain how living in groups has costs and benefits.
10.2 Illustrate the four types of social interactions.
10.3 Describe how eusocial species take social interactions to the extreme. In the course of a lifetime, an individual typically interacts with many members of its species. Interactions with mates, offspring, other relatives, and unrelated individuals in one’s species are known as social behaviors. Like most behaviors, social behaviors have a genetic basis and are therefore subject to natural selection. As a result, many types of social behaviors have evolved to favor the cohesiveness of family groups and populations. As we have mentioned in previous chapters, since social behaviors are traits, they are also affected by the environment in which the individuals live. Social behaviors Interactions with members of one’s own species, including mates, offspring, other relatives, and unrelated individuals.
Although the study of social behavior typically focuses on animals, many other organisms interact with conspecifics in ways that might be considered social. For example, bacteria and protists can sense the presence of individuals of the same species, often through chemical secretions, and react in “friendly” or “aggressive” ways. During parts of their life cycles, freeliving slime molds respond to others when they aggregate to form large fruiting bodies. Even plants communicate with one another. When one plant is damaged by herbivores, it emits volatile compounds. Other individuals detect the compounds and respond to them by producing chemical or structural defenses against future herbivore attacks. In this chapter, we will focus on the social behaviors of animals at the individual, population, community, and ecosystem levels. We will explore some of the implications of interacting within social groups, and describe various ways that individuals manage social relationships. We will also examine how different ecological conditions affect the evolution of social behaviors.
10.1 Living in groups has costs and benefits Animals are social for a variety of reasons. In some cases, offspring do not disperse but remain with their parents to form family groups. In other cases, individuals are attracted to each other for breeding. Individuals can also aggregate because they are independently attracted to the same habitat or resource. For instance, vultures gather around a carcass and dung flies congregate on cow pies. In this section, we will examine the costs and benefits of living in social groups and then discuss how animals use territories and dominance hierarchies in social interactions.
Benefits of Living in Groups
Animals generally form groups to increase their survival, rate of feeding, or success in finding mates. Survival While a single individual might not be able to fend off the attack of a predator, a group of individuals can be quite effective at doing so (Figure 10.1). Another survival mechanism available to social groups is a phenomenon known as the dilution effect. The dilution effect refers to the reduced, or diluted, probability of predation to a single animal when it is in a group. In an aggregation of prey, the predator has many prey choices, so the individual living in a group has a lower probability of being caught. The dilution effect is an important benefit of large groups such as herds of mammals, flocks of birds, and schools of fish.
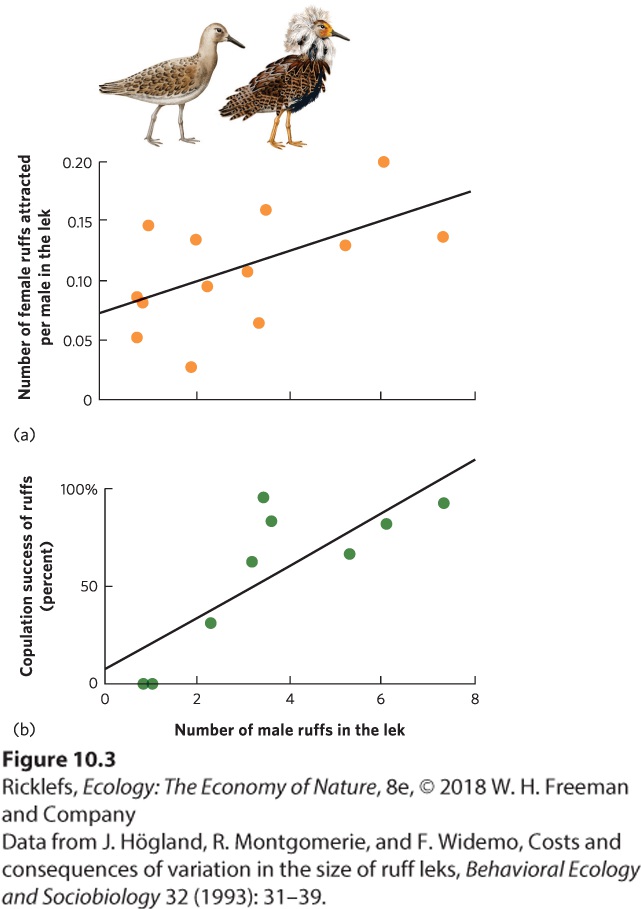
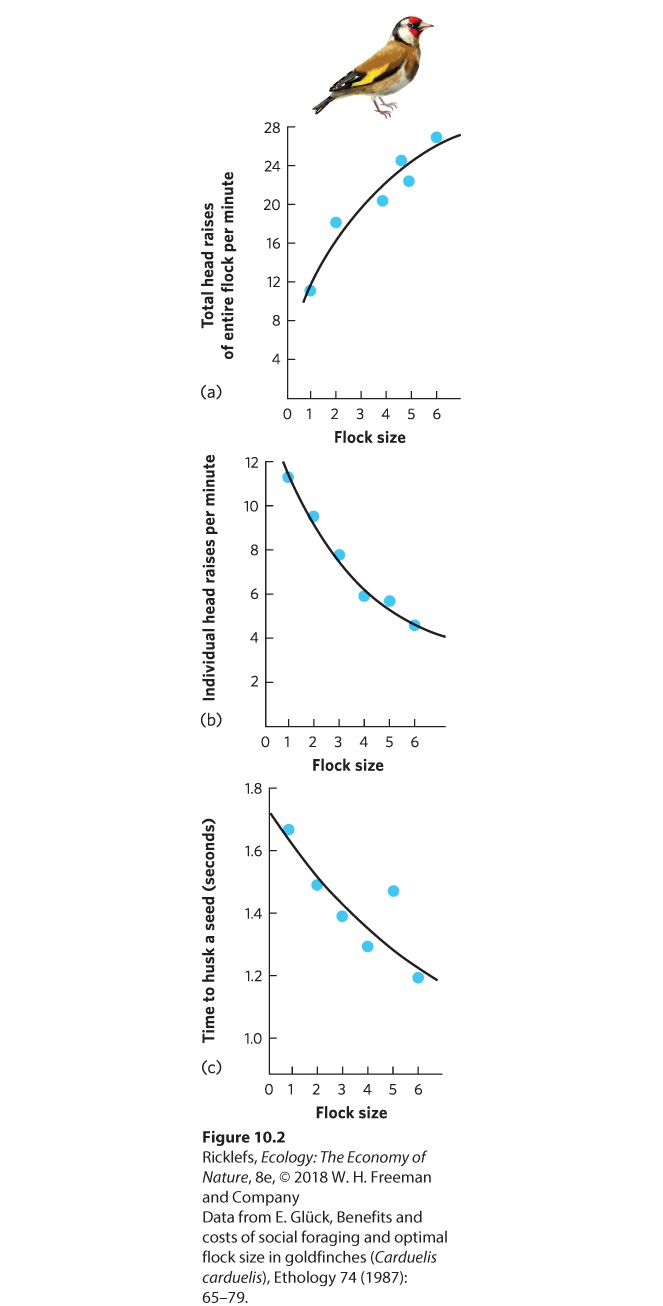
Figure 10.1 Group defense. Adult muskox (Ovibos moschatus), like these from Victoria Island, Canada, form an outward-facing circle and place the calves inside the circle, where they are safe from approaching predators. Dilution effect The reduced, or diluted, probability of predation to a single animal when it is in a group. A lower probability of predation in groups also allows individual prey animals to spend less time watching for predators. Consider the case of the European goldfinch (Carduelis carduelis), a small bird that feeds on the seed heads of plants in open fields and hedgerows. If you watch closely as the birds feed, you will notice that they raise their heads and look around for predators. The total number of head raises conducted by the group increases with flock size; as you can see in Figure 10.2a, the larger the group, the more eyes are on the lookout for predators. As flock size increases, however, each individual can raise its head less frequently, as shown in Figure 10.2b. Because each individual spends less time looking for predators, it can spend more time feeding. The data in Figure 10.2c show that when a goldfinch spends less time looking for predators, it can husk a seed much faster and therefore consume seeds more quickly.
Figure 10.2 Increased vigilance when living in a group. In the European goldfinch, an increase in flock size results in (a) an increase in the total number of head raises performed by the flock, (b) a decrease in the number of head raises performed by an individual, and (c) a decrease in the time required to husk a seed. Feeding Living in groups can also help animals locate and consume resources. Having many conspecifics all searching for food means that there are many more sets of eyes that might find food when it is rare. Furthermore, animals may find food easily but have difficulty in capturing and killing it when they are alone. In lions, for example, a lone female has a low probability of capturing and killing a zebra, but if she hunts with other lions, the probability of a capture goes up dramatically. Mating Socializing can also provide mating benefits since being social makes it easier to locate potential mates. An extreme example of socializing for mating benefits occurs when animals aggregate in large groups to attract members of the opposite sex by making calls or displaying in ways that capture the attention of potential mates. The location of the aggregation, known as a lek, is used only for displaying. The site has no other value either to the displaying sex or to the attracted sex. For example, males of the ruff (Philomachus pugnax)—a medium-sized wading bird that lives in northern Europe and Asia—come together at a lek and participate in mating displays to attract females. On the island of Gotland in Sweden, researchers observed ruff leks to determine whether lek size affected ruff mating. As you can see in
Figure 10.3a, males in larger leks were more successful at attracting females. In addition, as illustrated in Figure 10.3b, males in larger leks experienced a higher percentage of successful copulations with females, which confirms that forming social groups provides fitness benefits to the male birds.
Figure 10.3 Breeding benefits in a lek. Among displaying ruff males, those displaying to females in larger groups (a) are more likely to attract females and (b) have a higher probability of successfully copulating. Lek The location of an animal aggregation to put on a display to attract the opposite sex.
Costs of Living in Groups
The benefits of group living can certainly be substantial for many species, but group living can also come with costs that include predation and competition. Predation Groups of animals are much more conspicuous to predators than are individual animals. In a grassland, for example, it is easier for a predator to spot a herd of antelopes than an individual antelope. Given the propensity of antelopes to live in herds, this cost of being detected is outweighed by the benefits of the dilution effect and of more eyes to detect approaching predators.
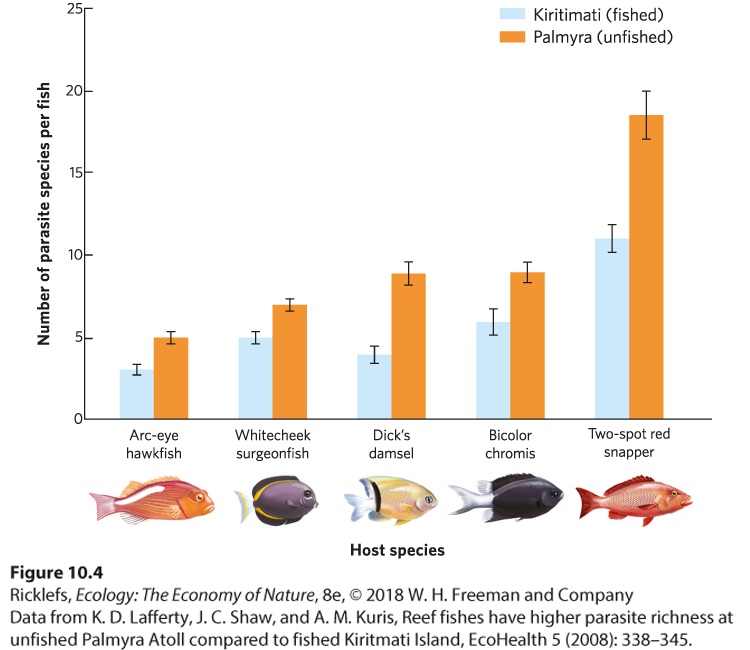
The risk of parasites and pathogens can also increase when living with conspecifics. Many species of parasites and pathogens spread from one host to another. High population density can increase the rate at which disease spreads and can lead to epidemics. For instance, coral reefs that experience fishing pressure generally have fewer fish compared to reefs that are protected from fishing. As a result, researchers examined the fish parasites from a protected and unprotected coral reef in the central Pacific Ocean. As shown in Figure 10.4, they found that fish on the protected reef were infested with a higher number of parasite species than the same species living in a coral reef that was fished. In addition, fish living in the protected reef frequently carried higher numbers of each species of parasite.
Figure 10.4 Parasite occurrence in coral reef fishes. Coral reefs protected from fishing have higher fish densities. A survey of five different species of fish found that fish living in a reef that is subject to fishing pressure contained fewer species of parasites than the same species living in a reef that is protected from fishing. Error bars are 95% confidence intervals. The parasite and disease costs of group living are also readily observed in modern aquaculture operations, which farm aquatic species for human consumption. These operations raise oysters, salmon, catfish, shrimp, and
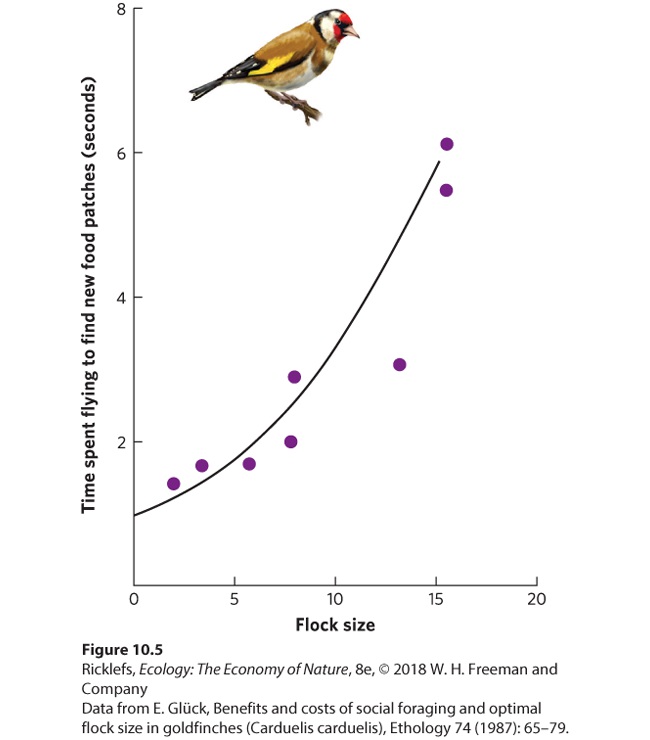
other edible species at very high densities. Under such conditions, a single infected individual can rapidly spread parasites and pathogens to the rest of the group. Increased transmission of parasites and diseases to groups that live in higher densities makes it undesirable for people to feed wild animals such as deer. When there is a steady, easily available source of food, deer form large aggregations around the food. This aggregating behavior makes it more likely that the animals will experience outbreaks of parasites compared to when they live in smaller family groups. Similar concerns exist for livestock operations in which animals are raised under very high densities. In this situation, diseases can jump to native wildlife populations and have dramatic effects with such viral diseases as rinderpest, avian influenza, and West Nile virus. Competition Another major cost of living in groups is competition for food. While larger groups are better at locating food, the food must then be shared among all the individuals in the group. Returning to the example of the European goldfinch, which experiences benefits from living in large groups, Figure 10.5 shows one consequence of sharing the food. Larger flocks consume the seeds in an area much faster than small flocks do, so larger flocks have to spend more time flying between patches of seeds. This causes each bird to spend more time and energy looking for food.
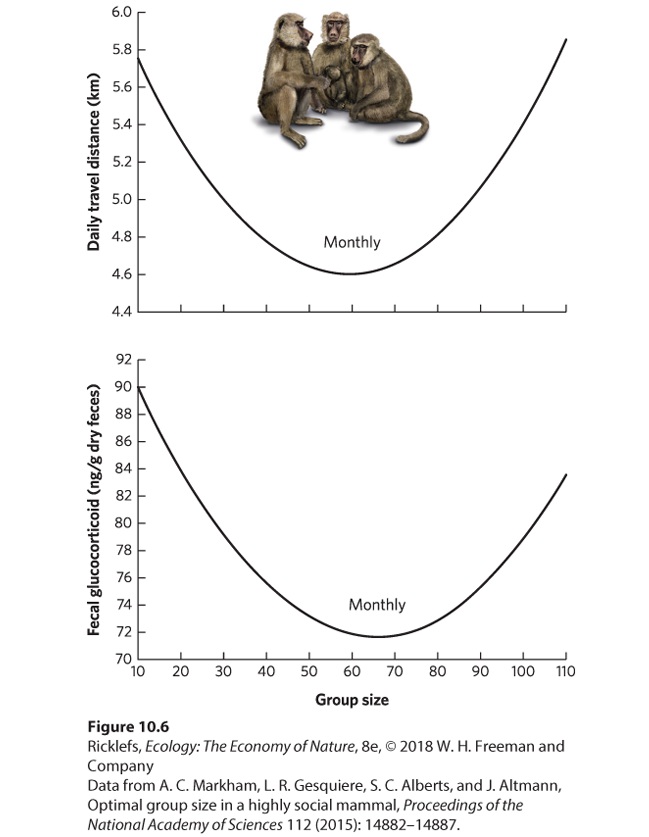
Figure 10.5 Large groups face increased competition for food. In the European goldfinch, larger flocks run out of food faster and must spend extra time and energy flying to find new patches of food. Each species that has evolved to live in groups faces different costs and benefits, which depend on the ecological conditions under which it lives. Assuming a genetic component for such social behavior, we expect natural selection to favor the evolution of group sizes that balance the costs and benefits for each species. A nice example of optimal group size can be found in yellow baboons (Papio cynocephalus). In 2015, researchers reported their observations of baboons for 11 years in East Africa (Figure 10.6). While baboon groups can range from 20 to 100 individuals, medium-size groups with 50‒75 individuals have the lowest levels of stress, which is assessed by hormone levels found in the baboon feces. Stress is known to harm an
individual’s health and survival. Medium-size groups have the lowest stress levels because they travel less than small and large groups. Large groups must travel farther to find food due to the large number of mouths to feed, whereas small groups must travel farther because they get pushed out of areas by large groups and by predators.
Figure 10.6 Optimal group size in yellow baboons. Groups of intermediate size have the lowest levels of stress and travel the shortest distances in search of food.
Territories
Many species of animals have evolved to live near their conspecifics by
establishing a territory or a dominance hierarchy. A territory is any area defended by one or more individuals against the intrusion of others. Territories can be either transient or relatively permanent, depending on the stability of the resources in the territory and how long an individual needs those resources. For example, many migratory species establish summer breeding territories and defend them for several months. Defending a highquality territory generally ensures greater resources, such as abundant food or nest sites. This typically improves the attractiveness of a territory holder as a mate, and therefore its fitness. When breeding is complete for the season, migratory species move on to their wintering grounds, where they establish new territories. Shorebirds that stop at several points along the way of their long migration defend feeding areas for a few hours or days and then continue their migratory trip. Hummingbirds and other nectar feeders defend individual flowering bushes and abandon them when those bushes cease producing flowers. As long as a resource can be defended and the benefits of defending the resource outweigh the costs, animals are likely to maintain territories. Territory Any area defended by one or more individuals against the intrusion of others.
Dominance Hierarchies
In some situations, defending a territory is impractical. This can occur when an individual is surrounded by so many conspecifics that it becomes impractical to defend a territory against them all, when resources are available for only short periods of time, or when the benefits of living in a group override the benefits of defending a territory. In such circumstances, individuals of many species form dominance hierarchies. A dominance hierarchy is a social ranking among individuals in a group, typically determined through a brief period of fighting or other contests of strength or skill (Figure 10.7). Once individuals order themselves into a dominance hierarchy, subsequent contests among them are resolved quickly by and in favor of higher-ranking individuals. In a linear dominance hierarchy, the firstranked member dominates all other members of the group, the second-ranked dominates all but the first-ranked, and so on down the line to the last-ranked individual, who dominates no one else in the group. In this way, all members

of the group can benefit from group living while accepting a dominance hierarchy within the group.
Figure 10.7 A dominance hierarchy. Bighorn sheep establish a dominance hierarchy. In this photo from Alberta Canada, the dominant male is followed by subordinate males. Dominance hierarchy A social ranking among individuals in a group, typically determined through fighting or other contests of strength or skill.
Concept Check
1. How is the dilution effect a benefit of group living? 2. Why are optimal group sizes often a compromise between the costs and benefits of group size? 3. How does group size affect disease transmission?
#### 10.2 There Are Many Types of Social Interactions
10.2 There are many types of social interactions Most social interactions can be considered as an action by one individual, the donor of the behavior, directed toward another individual, the recipient of the behavior. One individual delivers food, the other receives it; one individual attacks, the other is attacked. When the attacked individual responds—by standing its ground or by fleeing—it becomes the donor of this subsequent behavior. Every interaction between two individuals has the potential to affect the fitness of both individuals, either in a positive or negative way. To understand how an interaction affects both participants, it can be useful to categorize the interactions. In this section, we will explore the four types of social interactions between donors and recipients and then examine the conditions that favor a donor helping or harming a recipient. Donor The individual who directs a behavior toward another individual as part of a social interaction. Recipient The individual who receives the behavior of a donor in a social interaction.
The Types of Social Interactions
Social behaviors can be placed into one of four categories, as illustrated in
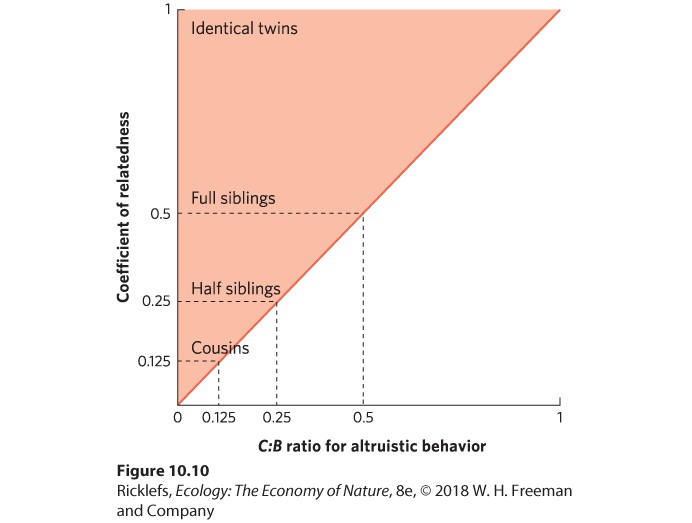
Figure 10.8: cooperation, selfishness, spitefulness, and altruism. When the donor and the recipient both experience increased fitness from the interaction, we call it cooperation. For instance, when one lion helps another kill a gazelle, and both feed from the kill, both individuals experience a fitness benefit. When the donor experiences increased fitness and the recipient experiences decreased fitness, we call it selfishness. Selfishness is a common interaction between two conspecifics that compete for a resource such as food, such as when a bald eagle attacks another to steal away a captured fish. The winner of the competition receives a fitness benefit, while the loser experiences a fitness loss. For both cooperation and selfishness, the interactions benefit the donor. We would therefore expect natural selection to favor any donors that engage in either cooperation or selfishness. Spitefulness occurs when a social interaction reduces the fitness of both donor and recipient. Spitefulness cannot be favored by natural selection under
any circumstance given that both participants experience lower fitness. Consistent with this prediction, spitefulness is not known to occur in wild animal populations. The fourth type of interaction, altruism, increases the fitness of the recipient but decreases the fitness of the donor. Explaining the evolution of altruism presents a unique challenge because it requires natural selection to favor individuals who improve the fitness of others while reducing their own fitness. We will explore this challenge in the next section.
Figure 10.8 The four types of social interactions. Cooperation occurs when the donor and recipient of a behavior both obtain a positive fitness effect. Selfishness occurs when the donor obtains a positive fitness effect while the recipient obtains a negative fitness effect. Altruism occurs when the donor obtains a negative fitness effect while the recipient obtains a positive fitness effect. Spitefulness occurs when the donor and recipient both obtain a negative fitness effect. Cooperation When the donor and the recipient of a social behavior both experience increased fitness from an interaction. Selfishness When the donor of a social behavior experiences increased fitness and the recipient experiences decreased fitness. Spitefulness When a social interaction reduces the fitness of both donor and recipient.
Altruism A social interaction that increases the fitness of the recipient and decreases the fitness of the donor.
Altruism and Kin Selection
Altruistic behavior is an interesting evolutionary behavior because it does not lead to an increase in direct fitness. Direct fitness is the fitness that an individual gains by passing on copies of its genes to its offspring. We would expect selfish individuals to prevail over altruistic individuals because selfishness directly increases the fitness of the donor, whereas altruism does not. Despite this expectation, altruism has evolved in many species. For example, some of the most extreme cases of altruism occur in colonial species, such as leaf-cutter ants and honeybees, in which workers forgo personal reproduction to rear the offspring of the dominant female. Direct fitness The fitness that an individual gains by passing on copies of its genes to its offspring. We can explain altruistic behavior by looking beyond direct fitness. When an individual has an altruistic interaction with a relative, it increases the fitness of the relative. Because you and your relative share some genes, due to sharing a common ancestor, when you help a relative improve its fitness, you are indirectly passing on more copies of your genes, which gives you indirect fitness. The key to understanding the evolution of altruism is to consider an individual’s inclusive fitness, which is the sum of its direct fitness and indirect fitness. When considering how selection operates, we say that direct fitness is favored by direct selection. Indirect fitness through relatives is favored by indirect selection, also known as kin selection. Indirect fitness The fitness that an individual gains by helping relatives pass on copies of their genes. Inclusive fitness The sum of direct fitness and indirect fitness. Direct selection Selection that favors direct fitness. Indirect selection Selection that favors indirect fitness. Also known as Kin selection.

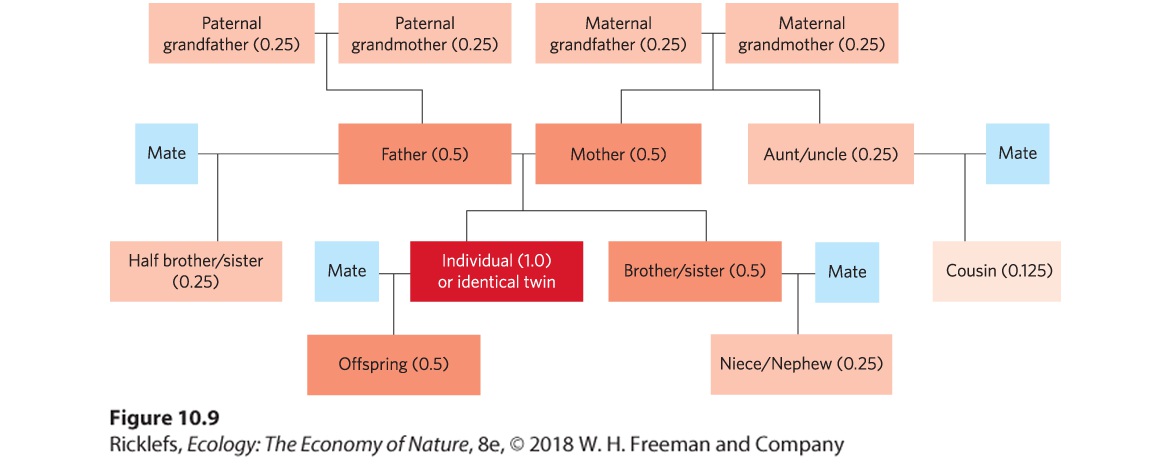
As we have noted, indirect or kin selection occurs because an individual and its relatives carry copies of some of the same genes inherited from a recent common ancestor. The probability that copies of a particular gene are shared by relatives is known as the coefficient of relatedness. As depicted in
Figure 10.9, its value for diploid organisms depends on the degree of relatedness between two individuals. If we focus on the individual in the red box of the family tree, we see that the coefficient between this individual and its offspring is 0.5 because a parent has two sets of genes but gives only one set to its offspring. As a result, the parent and offspring only have half of their genes in common. This also means that the coefficient of relatedness between our focal individual and its parent is also 0.5. If we next consider the focal individual and its siblings, we see that these two individuals have a 0.5 probability of receiving copies of the same gene from a parent. In the case of two cousins, the probability drops to 0.125 (one in eight) of inheriting copies of the same gene from one of their grandparents, which are their closest shared ancestors. Using these coefficients of relatedness, we can calculate the indirect fitness as the benefit given to a recipient relative (B) multiplied by the coefficient of relatedness between the donor and the recipient relative (r): Indirect fitness benefit=B×r
Figure 10.9 Coefficients of relatedness. The coefficient of relatedness is the probability that one individual possesses the same copy of a gene as another individual through a shared relative. In this family tree, we see that the individual in the red box has a 0.5 coefficient of relatedness with its parents, siblings, and offspring. More distant relatives have lower coefficients of relatedness, as indicated by the boxes with lighter shades of red. The coefficients of relatedness are based on the assumption that none of the mates are related to the highlighted individual. Coefficient of relatedness
The numerical probability of an individual and its relatives carrying copies of the same genes from a recent common ancestor. In the case of nonrelatives, there is a zero probability that an individual carries the same genes from a recent common ancestor. In examining these different coefficients of relatedness, we can see that an individual has a higher probability of leaving more copies of its genes in the next generation by promoting the fitness of its closest relatives and gains nothing by promoting the fitness of nonrelatives. Understanding the role of kin selection and coefficients of relatedness helps resolve the puzzle of how altruistic social interactions can evolve. Whereas selfish interactions provide direct fitness benefits to the donor, altruistic interactions provide indirect fitness benefits to the donor, weighted by the coefficient of relatedness between the donor and the recipient. If the inclusive fitness of altruistic behaviors exceeds the inclusive fitness of selfish behaviors, then altruism will be favored by natural selection. The evolution of altruistic behavior becomes clear when we examine the costs and benefits in an equation. Genes for altruistic behavior will be favored in a population when the benefit to the recipient (B) times the recipient’s coefficient of relatedness to the donor (r) is greater than the direct fitness cost to the donor (C): B×r>C If we rearrange this equation, we can show that for altruism to evolve, the cost-benefit ratio must be less than the coefficient of relatedness between donor and recipient: C/B<r

Figure 10.10 shows this relationship graphically. Based on this equation and the figure, we can see that altruism is favored when the cost to the donor is low, the benefit to the relative is high, and the donor and its relative are closely related.
Figure 10.10 Conditions that favor the evolution of altruistic behaviors. An altruistic behavior will evolve whenever the ratio of donor costs and recipient benefits (C/B) is less than the coefficient of relatedness between the donor and recipient. The region in red indicates the conditions that favor the evolution of altruistic behaviors. A study of wild turkeys (Meleagris gallopavo) in California has shown how altruistic behavior can be maintained through kin selection. Male turkeys display at leks by puffing up their feathers and strutting back and forth to attract females. Males may either display alone or with another male (Figure 10.11). When a pair of males displays together, only the dominant male copulates with the females that they attract. This raises the question of why the subordinate male in a pair spends its time and energy displaying when it will not be given a chance to produce any offspring with the female. Researchers obtained their first clue by using genetic data. Males in a coalition were more closely related than males drawn at random from the population. Indeed, the average coefficient of relatedness between groups of displaying males was 0.42, which suggests that paired males represent a mixture of full brothers (r = 0.5) and half brothers (r = 0.25).
Figure 10.11 Wild turkey coalition. When two or more male turkeys display together to attract a female, only the dominant male sires the offspring. These individuals live in Texas. The researchers then determined the average number of offspring sired by the different types of males. Dominant males in a coalition sired an average of 6.1 offspring; subordinate males in a coalition sired 0 offspring; and males that displayed alone sired an average of 0.9 offspring. With these data, we can evaluate the fitness of the subordinate male in the coalition. By being a part of the coalition, the subordinate male forgoes the ability to breed as a solo male, which would have allowed him to sire 0.9 offspring. Therefore, his cost of being altruistic is 0.9 offspring. By helping his brother or half brother to become highly successful at attracting females, he allows his brother to sire 6.1 offspring. As we have learned, the subordinate male’s average coefficient of relatedness to his brother is 0.42. Therefore, the indirect fitness benefit to the subordinate male can be calculated as Indirect fitness benefit=B×r=6.1×0.42=2.6 This means that a subordinate male obtains greater inclusive fitness by helping his brother than by going out on his own to attract females. The concept of kin selection has given ecologists a better understanding of the evolutionary reasons underlying a wide variety of altruistic and selfish behaviors in animals. In the next section, we will explore the evolution of an extreme form of altruistic behavior in which individuals completely forgo their reproduction to help others.
##### Analyzing Ecology: Calculating Inclusive Fitness
1. Why can altruism not be explained by direct fitness alone? 2. In the kin selection explanation for the evolution of altruism, why is the benefit to the recipient weighted by its coefficient of relatedness to the donor? 3. How do direct fitness and inclusive fitness differ?
Analyzing Ecology
Calculating Inclusive Fitness In the pied kingfisher (Ceryle rudis), a fisheating bird from Africa and Asia, adult males often forgo their own reproduction to help their parents raise offspring. Researchers have identified primary helpers and secondary helpers. Primary helpers are sons of the parents, and they work hard to protect the nest, and to bring food to the chicks. In some cases, one of the parents disappears and is replaced by an unrelated mate, so the son ends up helping a parent and a stepparent. Secondary helpers—which are unmated males from other families—are not related to the parents and do not work as hard to feed and protect the offspring. After helping for one year, both types of helpers set up their own nest the following year. A third group of males, known as delayers, do not help but simply delay reproduction until their second year. Researchers followed several nests of kingfishers and determined how much each helper improved the fitness of the parents in the first year (B1), the probability of surviving and finding a mate in the subsequent year (Psm), and the fitness of the helper when he bred independently in the second year (B2). They also quantified the coefficients of relatedness between the helpers and the parents being helped in the first year (r1) and between the helpers and their own offspring produced in the second year

(r2). The coefficient of relatedness for the primary helpers was 0.32 in year 1. This was the result of some nests retaining both parents of the helper (r = 0.5) and other nests having one parent of the helper plus a stepparent (r = 0.25). Based on these data, the researchers calculated the inclusive fitness of the primary helpers, secondary helpers, and delayers. As you can see, the primary helper had inclusive fitness that was a bit higher after 2 years. The primary helpers obtain about half of their inclusive fitness by helping their parents raise their siblings in year 1 and the other half by having their own offspring in year 2. In contrast, the secondary helpers did not gain any indirect fitness in year 1, but had a higher probability of surviving and finding a mate in year 2, which improved their direct fitness. The delayers obtained no indirect fitness in year 1 and had a poor ability to attract mates in year 2, leading to a low inclusive fitness. YOUR TURN Use the table below to calculate the change in inclusive fitness if the primary helpers only improved the fitness of their parents by 1.0 rather than 1.8. Under this scenario, which strategy of helping would be most favored by natural selection? Year 1 Year 2
Primary Helper
1.8 × 0.32 = 0.58 2.5 × 0.5 × 0.32 = 0.41 0.58 + 0.41 = 0.99
Secondary
1.3 × 0.00 = 0.00 2.5 × 0.5 × 0.67 = 0.84 0.00 + 0.84 = 0.84

0.0 × 0.00 = 0.00 2.5 × 0.5 × 0.23 = 0.29 0.00 + 0.29 = 0.29 Source: Data from H.-U. Reyer, Investment and relatedness: A cost/benefit approach of breeding and helping in the pied kingfisher, Animal Behaviour 32 (1984): 1163–1178.
#### 10.3 Eusocial Species Take Social Interactions to the Extreme
10.3 Eusocial species take social interactions to the extreme We have seen that many animals are social and interact with conspecifics in a variety of ways. Some animals are so extremely social that we call them eusocial. Eusocial (i.e., “truly” social) species are distinguished by their display of all four of the following characteristics: Eusocial A type of animal society in which individuals live in large groups with overlapping generations, cooperation in nest building and brood care, and reproductive dominance by one or a few individuals. 1. Several adults living together in a group 2. Overlapping generations of parents and offspring living together in the same group 3. Cooperation in nest building and brood care 4. Reproductive dominance by one or a few individuals, and the presence of sterile individuals Among the insects, eusocial species are limited to the order Hymenoptera, which includes bees, ants, and wasps, and the order Isoptera, which includes termites (Figure 10.12). Not only are these social insects of evolutionary interest, they are also major players in ecosystem processes. Eusocial insects pollinate plants, consume plant and animal material on a large scale, and recycle wood and organic detritus. Their dominance in the world is due in large part to the immense success of eusociality. Other than insects, the only animals known to be eusocial are two species of mammals that live in underground tunnels in Africa: the naked mole rat (Heterocephalus glaber) and the Damaraland mole rat (Fukomys damarensis).
Figure 10.12 Eusocial species. These Texas leaf-cutter ants (Atta texana) are one of many highly social species. Pictured here is a queen and her young daughters that are tending a fungus garden deep inside their underground nest. Eusocial species are fascinating to ecologists because most individuals in a eusocial group forgo sexual maturation and reproduction. Instead, they specialize at tasks that include defending the group, foraging for the group, or taking care of the subsequent offspring of their parents. Because nonreproductive individuals are specialized for these tasks, they are known as sterile castes. A caste consists of individuals within a social group who share a specialized form of behavior. For example, we saw in Chapter 9 that worker honeybees are a caste that works for the hive but typically does not reproduce. Similarly, the workers of the leaf-cutter ants represent a variety of castes that do different jobs but they do not reproduce. Caste Individuals within a social group sharing a specialized form of behavior. How can natural selection produce individuals that lack any reproductive output and therefore lack any direct fitness? To help answer this question, we will start by examining the unique breeding habits of the hymenopteran insects.
Eusociality in Ants, Bees, and Wasps
In Chapter 9, we talked about the unique reproductive behavior of the hymenopteran insects. You may recall that this group of insects produces
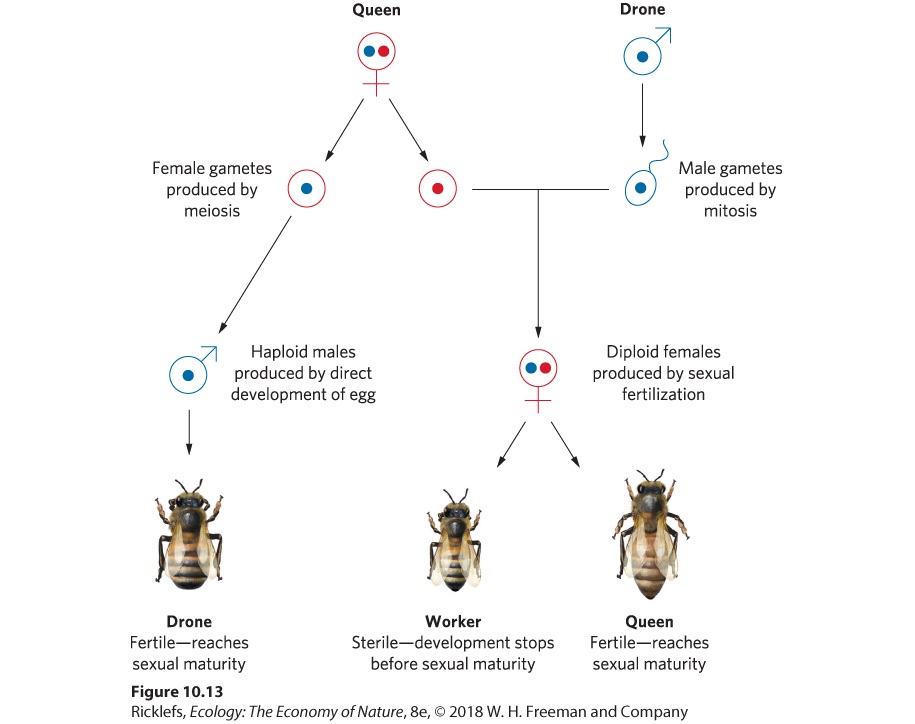
sons by laying unfertilized eggs and produces daughters by laying fertilized eggs. This method of reproduction is detailed in Figure 10.13. Because one sex is haploid and the other sex is diploid, the hymenopterans are said to have a haplodiploid sex-determination system. As we will see, this sexdetermination system helps to favor the evolution of eusociality.
Figure 10.13 The haplodiploid mating system of hymenopteran insects. In ants, bees, and wasps, sons are produced when a queen’s haploid gamete remains unfertilized and it develops into a haploid son. Daughters are produced when a queen’s haploid gamete is fertilized by a drone’s haploid gamete and it develops into a diploid daughter. Haplodiploid A sex-determination system in which one sex is haploid and the other sex is diploid. Social Organization Eusocial insect societies are dominated by one egg-laying female or a few egg-laying females, referred to as queens. The queens in colonies of ants, bees, and wasps mate only once during their lives and store enough sperm to produce all their offspring—up to a million or more offspring over a period of 10 to 15 years in some ant species. The nonreproductive progeny of a queen gather food and care for developing brothers and sisters, some of
which become sexually mature, leaving the colony to mate and to establish new colonies. Queen The dominant, egg-laying female in eusocial insect societies. As we discussed in Chapter 9, honeybee societies have a simple organization. The offspring of a queen are divided among a sterile worker caste that is female and a reproductive caste that consists of drones and future queens. Whether an individual becomes a sterile worker female or a fertile female is determined by the length of time the larva is fed royal jelly. As a result, the worker caste represents an arrested stage in the development of reproductive females; they stop developing before sexual maturity. Eusocial behavior appears in a large number of hymenopteran species. Coefficients of Relatedness Haplodiploidy is important to the evolution of eusocial animals because it creates strong asymmetries in coefficients of relatedness. The queen has the same genetic relationship to sons as she has to daughters (r = 0.50), so she can be relatively indifferent to the sex of her reproductive offspring—the drones and new queens that leave the nest—especially when the sex ratio among reproductive individuals in other local hives is nearly equal. However, the relatedness among siblings is unique in hymenopterans. If a queen has mated with a single drone, then all females have the same set of genes from their haploid father and they have a 50 percent probability of sharing the same genes from their mother. As a result, the coefficient of relatedness between a given female and her sisters is 0.75 in the haplodiploid mating system. This causes females to be more closely related to each other than is typical in a diploid mating system, where it is only 0.50. In contrast, her brothers are haploid, so the brothers receive no genes from the drone, but they have a 50 percent probability of sharing the same genes from their mother. As a result, the average coefficient of relatedness between a given female and her brother is 0.25. This skewed relatedness favors the evolution of eusocial groups. To understand how being haplodiploid favors eusociality, we can compare the different options for obtaining fitness in these organisms. For example, a
female worker that raises a fertile sister receives the fitness benefit of that individual multiplied by the coefficient of relatedness (r = 0.75). When a female raises its own offspring, it receives the same fitness benefit multiplied by the coefficient of relatedness between a mother and daughter (r = 0.50). This means that the indirect fitness obtained from caring for a sister exceeds the direct fitness that could be obtained from caring for a daughter. Thus, cooperation is likely to be greater among all-female castes than among male castes or, especially, among mixed castes. This may explain why workers in hymenopteran societies are all female. The fact that workers are three times more closely related to their sisters than to their brothers may also explain why broods of reproductive individuals usually favor females, by about three to one on a weight basis, in spite of the queen’s indifference to the sex ratio. When a female worker can help to rear more female than male reproductive individuals, her own inclusive fitness may actually be higher than it would be if she raised a brood of her own consisting of an equal number of males and females. Under these circumstances, it is not surprising that sterile castes have evolved.
Eusociality in Other Species
Although the hymenopterans are among the best known of the eusocial insects, there are a few other species that are also eusocial, including termites and a group of mammals known as naked mole rats. Unlike the hymenopterans, these latter two groups are both diploid, which present the challenge of understanding the conditions that favor eusociality in these species. Termites Termite colonies can be massive structures that are dominated by a mated pair called the king and queen (Figure 10.14a). Because the queen’s role is almost entirely limited to egg production, her abdomen is grossly distended to allow her to produce thousands of eggs (Figure 10.14b). The king and queen produce sons and daughters by sexual reproduction. With a few exceptions, these sons and daughters act as workers in the termite society but can become sexually mature if either the king or the queen dies. Many species of termites also have a second caste of nonreproductive individuals known as soldiers. As their name implies, soldiers help to defend the nest
against intruders such as ants. Soldiers typically have a very large head that can be used to block the openings of a termite nest to prevent ants from entering.
Figure 10.14 Termite colonies. (a) A massive termite colony located in the Northern Territory, Australia. (b) A queen termite (Macrotermes gilvus) living deep inside a colony surrounded by workers. Because her primary role is egg production, the queen’s abdomen is much larger than that of workers and soldiers. Mole Rats There are dozens of species of mole rats, but only two are known to be eusocial: the naked mole rat and the Damaraland mole rat. These rodents live in underground tunnels in African grasslands in colonies of up to 200 individuals. In naked mole rats, a single queen and several kings are responsible for all the reproduction in the colony, and all individuals are diploid (Figure 10.15). Although the workers in the group are capable of reproducing, they forgo reproduction in favor of caring for their younger siblings and taking care of the colony. Current research suggests that the subordinate males and females do not do this willingly. Instead, the dominant female harasses them, which causes them to become stressed. Stress reduces the levels of sex hormones in the subordinates and makes them less motivated to breed.
Figure 10.15 Naked mole rats. The naked mole rat is one of two species of mammals that are eusocial. In this photo, a queen is laying on top of the workers.
The Origins of Eusociality
From its distribution across a variety of distantly related species, ranging from hymenopterans to mammals, it is clear that eusociality has evolved independently many times. Even within the hymenopterans, eusociality appears to have evolved several times. It is tempting to conclude that eusociality is caused by a haplodiploid sex-determination system, but many haplodiploid species are not eusocial, and some eusocial species—such as termites and naked mole rats—are not haplodiploid. Based on these observations, we are left to conclude that while being haplodiploid appears to favor the evolution of eusocial behavior by providing large indirect fitness effects when workers forgo breeding to help their sisters, it is not required for the evolution of eusocial behavior. Throughout our discussion, we have focused on the importance of the coefficient of relatedness with the idea that the coefficient needs to be relatively high to favor eusociality. However, this equation could also be satisfied if the cost of forgoing personal reproduction is very low. If individuals that chose to leave a colony had a very low likelihood of surviving and setting up a new colony, then direct fitness would be very low. In naked mole rats, for example, some individuals leave the home colony to form new colonies, but most of these small new colonies do not persist
beyond one year. As a result, the cost of forgoing reproduction—in terms of lost direct fitness—would be very small. When this cost is small, a large coefficient of relatedness is no longer required to favor eusocial behavior. The origins of eusocial behavior are still actively debated. Many researchers have argued that it has been favored by the inclusive fitness of the altruistic sterile offspring. Others have argued that it has evolved to enhance the direct fitness of parents that force the offspring to forgo reproduction. Like many biological questions, continued research will likely provide new insights that will help us to understand eusocial societies. Behavioral relationships among the social insects represent one extreme along a continuum of social organization, from animals that live alone (except to breed) to those that aggregate in large groups organized by complex behavior. Regardless of their complexity, all social behaviors balance costs and benefits to the individual, and the magnitude of these costs and benefits is often determined by the ecological conditions in which these behaviors exist. Like morphology and physiology, behavior is strongly influenced by genetic factors and therefore is subject to evolutionary modification by natural selection. The evolution of behavior becomes complicated when individuals interact within a social setting, and the interests of individuals within a population may either coincide or conflict. Understanding the evolutionary resolution of social conflict in animal societies continues to be one of the most challenging and important concerns of biology.
Concept Check
1. What are the four characteristics of a eusocial species? 2. How does a haplodiploid sex-determination system favor the evolution of eusociality? 3. How does a low probability of successfully starting a new colony favor the evolution of eusocial behavior in naked mole rats?
Concepts
Hen-Pecked Chickens Domesticated chickens. Chicken survival and egg production depend on not only the traits of each chicken but also the way each chicken interacts with other chickens in its cage. The social behavior of animals in the wild is inherently interesting, but understanding social behavior also has practical applications for domesticated animals. For example, there are costs and benefits of raising domesticated animals in large social groups. Raising animals such as pigs and chickens in high densities is more cost effective because it requires less space. However, as we have seen, living at high densities has downsides that include increased risks of disease transmission, increased competition, and increased fighting. For farmers who raise domesticated animals, raising animals in high densities translates into fewer
healthy animals and smaller profits. However, during the past two decades, researchers with insights into group living have helped breed more productive domesticated animals. Some of the leading research has been conducted with chickens (Gallus gallus). Chickens are well known for fighting when raised under crowded conditions. For example, if one chicken receives a small injury that causes a spot of blood on its feathers, other chickens will begin to peck at the spot, causing greater injury and often death through cannibalism. In the process, some chickens will become dotted with blood spots and other chickens, in turn, will also peck them. This incessant behavior is the origin of the phrase “henpecked.” Fighting among chickens can reduce overall health and egg production, and can even cause death of the chickens. In fact, fighting can cause so much lost profit to poultry farms that there is a long-standing practice of cutting off the ends of chicken beaks to reduce injuries. Although beak trimming is effective at reducing fighting and cannibalism, it is a controversial practice and has been recently banned in several countries. This has motivated animal breeders to find alternative ways to reduce fighting among crowded domesticated animals. The traditional method of breeding animals has been to select for those individuals with the best survival, growth, or other traits such as increased egg production. After decades of selection on traits, genetic variation declines and becomes quite low, which makes it difficult to conduct further selection. In the mid-1990s, however, researchers discovered that they could cause large responses to selection if they based their selection on the bestperforming social groups of chickens living together rather
than just the best-performing individual chickens. Groups of hens were caged together and groups with the highest survival were used to generate offspring for the next generation. At the start of the experiment, annual hen mortality was 68 percent. After six generations of selection for the best surviving groups, annual hen mortality dropped to 9 percent, which was similar to the mortality rate of hens that were raised alone with no opportunities to fight. Moreover, lifetime egg production increased dramatically, from 91 eggs per hen in the first generation to 237 eggs per hen by the sixth generation. The increase in egg production was caused by the hens living longer and being in better physical condition. These data suggest that selection for the best social groups could reduce fighting to a level that would eliminate the need to trim chicken beaks. During the past 5 years, researchers have begun to understand why selection based on social groups of animals could cause such dramatic increases in animal production. Biologists have traditionally focused only on the heritability of traits for an isolated individual. However, the performance of an individual depends on not only the individual but also the way in which it interacts with other individuals. For example, if an individual is particularly aggressive and lives in a group of other aggressive individuals, the group will spend more time fighting and more deaths will occur. In contrast, a social group with individuals who fight less with each other will have lower stress, better health, and a reduced mortality rate. Because social animals interact with conspecifics, we need to consider the heritability of traits not just when an individual lives alone but when an individual lives in a social group. In chickens, for example, the heritability of survival for an
individual can be quite low, but the heritability of survival when social interactions are included can be two to three times higher because social interactions affect survival. Similar results have been observed in other farmed animals, including domesticated pigs (Sus scrofa) and mussels (Mytilus galloprovincialis). All this suggests that when we understand the social interactions of animals, we can dramatically improve agricultural production of domesticated animals. SOURCES: Muir, W. M. 1996. Group selection for adaptation to multiple-hen cages: Selection program and direct responses. Poultry Science 75: 447–458. Wade, M. J., et al. 2010. Group selection and social evolution in domesticated animals. Evolutionary Applications 3: 453–465.
Summary of Learning Objectives
10.1 Living in groups has costs and benefits. The benefits of social living include the dilution effect in which large groups of prey have a reduced likelihood of being killed by a predator, reduced need for personal vigilance, and increased ability to find food and mates. The costs include increased visibility to predators, increased risk of parasite and pathogen transmission, and increased competition for food. In response to social living, many species have evolved the ability to establish territories and dominance hierarchies to manage individual interactions. Key Terms: Social behaviors, Dilution effect, Lek, Territory, Dominance hierarchy
10.2 There are many types of social interactions. When we envision interactions in terms of donors and recipients, we can devise four types of social interactions: cooperation, selfishness, spitefulness, and altruism. Cooperation and selfishness of donors should be favored by natural selection, whereas spitefulness should not. Altruism can be favored when the recipient of an altruistic act is closely related to the donor, as measured by the coefficient of relatedness. As a result, altruism evolves because individuals experience an increase in inclusive fitness, which is the sum of direct and indirect fitness. Key Terms: Donor, Recipient, Cooperation, Selfishness, Spitefulness, Altruism, Direct fitness, Indirect fitness, Inclusive fitness, Direct selection, Indirect selection, Coefficient of relatedness
10.3 Eusocial species take social interactions to the extreme. Eusocial animals consist of many individuals living together with dominant individuals reproducing and subordinate individuals forgoing reproduction. Eusocial species are common among the haplodiploid species of bees, ants, and wasps, but also exist in diploid species of
termites and at least two species of mammals. A high coefficient of relatedness favors the evolution of eusocial behavior, but it is not required. Equally important may be the presence of a low cost of lost fitness in species that have a low probability of leaving the group and reproducing on their own. Key Terms: Eusocial, Caste, Haplodiploid, Queen
Critical Thinking Questions
1. If living in large groups has costs and benefits, under what conditions would natural selection favor group living? 2. Why might individuals give up defending territories if the density of their population increases? 3. Explain the costs and benefits that might influence the optimal flock size in birds. 4. Compare and contrast the conditions under which natural selection will favor cooperative versus altruistic behavior. 5. How could helping raise the offspring of another couple improve the helper’s fitness if the helper is not related? 6. Why are selfish behaviors less favored when the donor and recipient are related to each other? 7. Compare the coefficient of relatedness between brothers and sisters in diploid organisms with that of haplodiploid organisms. 8. Given that termites are diploid, what would you predict about the costs and benefits of the workers forgoing reproduction, which would favor the evolution of eusociality? 9. What evidence is there that a haplodiploid sex-determination system is not required for the evolution of eusociality? 10. Why might selection based on groups of domesticated goats result in greater meat production than selection on individual goats?
#### Graphing the Data: How Living in Groups Affects Predation Risk
GRAPHING THE DATA How Living In Groups Affects Predation Risk As we have discussed, living in groups has a number of potential costs and benefits. To determine if living in schools provided an antipredator benefit to minnows, researchers placed different numbers of minnows into aquaria and determined how often schools of different size were approached by a larger species of predatory fish. Using the data in the table and your knowledge about calculating sample standard deviations from Chapter 2, calculate the means and the standard deviations for the number of approaches by a predator per minute as a function of different school sizes. Then graph the means and sample standard deviations using a line graph. Minnow school size Trial 0.9 0.7 0.4 0.4 0.1 0.8 0.8 0.5 0.5 0.1 0.7 0.9 0.6 0.3 0.2 1.1 0.6 0.8 0.2 0.3 1.0 1.0 0.7 0.6 0.3 Mean Number of Approaches/Minute Standard Deviation Based on these data, what can you conclude about the effect of school size on the likelihood of predation by the larger species of fish?