9 Reproductive Strategies
9Reproductive Strategies A honeybee hive. In most populations of honeybees, a queen can reproduce by laying either haploid eggs that produce sons or fertilized diploid eggs that produce daughters. The Sex Life of Honeybees Honeybees (Apis mellifera) have a complicated sex life. They live in hives that may contain tens of thousands of bees, usually progeny of the same mother, known as the queen. Like many organisms, the queen bee produces sons and daughters, but she does so in a rather unique way. Early in her life, the queen bee flies out of the hive and mates in the air with a group of male bees. Male bees, known as drones, are smaller than the queen. A queen mates with several drones, but the larger drones contribute more sperm than the smaller drones. The queen stores sperm from this single mating event in a special organ in her body, known as the spermatheca, where it remains viable for several years. She uses these sperm to fertilize her eggs and make diploid daughters, known as workers. In contrast, she

creates drones by laying unfertilized haploid eggs. The drones a queen produces rarely mate with her, but instead mate with other queens outside the hive. After mating, the drones die. A key issue for a successful hive is the proper ratio of drones and workers. In a typical beehive, the queen bee may produce a few dozen drones but tens of thousands of workers. Since the workers do the vast majority of the work in the hive, producing many more workers than drones is in the best interest of the queen. Since workers only live for 4 to 7 weeks, the queen must constantly produce more of them. Genetically, the workers and the queen are quite similar. They are both female and they both arise from a fertilized, diploid egg. What makes them different is the food they are given as larvae. For the first few days of life, all larvae are fed royal jelly, a liquid produced by the worker bees, but after this, the larvae destined to be workers are switched to a diet of honey and pollen. These workers are not capable of mating with a drone, but they can lay unfertilized eggs. Larvae destined to be queens continue to be fed royal jelly, which allows the future queen to grow very large. The queen’s size allows her to eventually lay as many as 2,000 eggs per day. When a beehive experiences the decline of its queen, new larval queens are often already being produced as replacements. Sometimes, however, the death or departure of a queen happens unexpectedly and there are no replacements. In such cases, some of the workers lay eggs but, since the workers cannot mate, their eggs are haploid and destined to turn into drones. Without a queen to lay fertilized eggs, the colony will eventually die. “In a typical beehive, the queen bee may produce a few dozen drones but tens of thousands of workers.” Scientists have recently discovered an exception to this scenario in
a subspecies of the honeybee known as the Cape honeybee (Apis mellifera capensis) that is found in southern Africa. Cape honeybee workers can lay diploid eggs without ever mating by using an unusual form of meiosis, in which they produce haploid egg cells and then merge two haploid cells to form a diploid cell. Researchers recently discovered that multiple genes control this ability. As a result, depending on which alleles they carry, individual workers can lay either haploid eggs that give rise to drones or diploid eggs that give rise to workers. Although laying diploid is normally the queen’s job, evolving this behavior in workers can ensure the persistence of a beehive that has lost its queen. Even more interesting is that these workers can sneak into other beehives, eat the stored food of the hive, and lay their eggs. In doing so, a worker can have even greater fitness by having other hives rear her offspring. The complexity of honeybee reproduction is an excellent example of the variety of reproductive strategies that have evolved. These options include reproducing with or without a sexual partner, choosing the number of mating partners, selecting the best traits of a sexual partner, varying the way the sex of offspring is determined, and controlling the number of sons versus daughters. In this chapter, we will explore the wide range of reproductive strategies in a diversity of organisms. SOURCES: Wallberg, A., C. W. Pirk, M. H. Allsopp, and M. T. Webster. 2016. Identification of multiple loci associated with social parasitism in honeybees. PLoS Genetics 12: e1006097. Lattorff, H. M. G., R. F. A. Moritz, and S. Fuchs. 2005. A single locus determines thelytokous parthenogenesis of laying honeybee workers (Apis mellifera capensis). Heredity 94: 533– 537.
Learning Objectives
After reading this chapter, you should be able to:

9.1 Describe both sexual and asexual reproduction.
9.2 Explain how organisms can evolve as separate sexes or as hermaphrodites.
9.3 Describe why sex ratios of offspring are typically balanced, but can be modified by natural selection.
9.4 Explain how mating systems describe the pattern of mating between males and females.
9.5 Explain how sexual selection favors traits that facilitate reproduction. The evolution of reproductive strategies involves a large number of different factors, many of which are influenced by ecological conditions. For example, organisms can evolve to reproduce sexually or asexually; each strategy has unique costs and benefits, particularly when species interact with parasites and pathogens. Organisms can evolve to reproduce as separate sexes or as hermaphrodites that possess both female and male sexual organs. If they reproduce as hermaphrodites, they must also evolve solutions to the problems associated with self-fertilization. In many species, the sex ratio of offspring can be altered to respond to changing ecological conditions. Finally, we see many different mating strategies to improve fitness, including the number of mates and the preference for certain traits in the opposite sex. This chapter explores how ecological conditions affect the evolution of sex and the strategies that organisms have evolved to increase their fitness.
9.1 Reproduction can be sexual or asexual All organisms reproduce, but they have a variety of ways in which they accomplish this task. In plants, animals, fungi, and protists, reproduction may be accomplished through either sexual reproduction or asexual reproduction. In this section, we will look at the two processes and compare their costs and benefits.
Sexual Reproduction
As we discussed in Chapter 7, reproductive function in most animals and plants is divided between two sexes. When progeny inherit DNA from two parents, we say they are the result of sexual reproduction. Haploid germ cells, called gametes, are produced through meiosis within sex organs called gonads. Each gamete contains a single full set of chromosomes. In animals, these haploid cells can immediately act as gametes. In plants and many protists, the haploid cells develop into multicellular, haploid stages of the life cycle that eventually produce gametes. When two gametes combine to form an offspring, each parent contributes one set of chromosomes. During meiosis, the distribution of the chromosomes into the gametes is generally random and the subsequent mixing of chromosomes from the two parents results in new combinations of genes in the offspring. Ultimately, two gametes join together in an act of fertilization to produce a diploid zygote. Sexual reproduction A reproduction mechanism in which progeny inherit DNA from two parents.
Asexual Reproduction
In contrast to sexual reproduction, progeny produced by asexual reproduction inherit DNA from a single parent. This can be accomplished through either vegetative reproduction or parthenogenesis. Asexual reproduction A reproduction mechanism in which progeny inherit DNA from a single parent. Vegetative Reproduction Vegetative reproduction occurs when an individual is produced from the
nonsexual tissues of a parent. Many plants can reproduce by developing new shoots that sprout from leaves, roots, or rhizomes (i.e., underground shoots).
Figure 9.1 shows an example of this in a walking fern (Asplenium rhizophyllum), which produces offspring when the tips of its leaves touch the soil. If you have ever placed a plant clipping into a glass of water and watched it grow roots to become a new plant, you have witnessed this type of reproduction. Individuals that descend asexually from the same parent and bear the same genotype are known as clones. Many simple animals, such as hydras, corals, and their relatives, also reproduce in this way; they produce buds along their body that develop into new individuals. Bacteria and some species of protists reproduce by duplicating their genes and then dividing the cell into two identical cells, a process known as binary fission.
Figure 9.1 Vegetative reproduction. Organisms that use vegetative reproduction produce offspring from nonsexual tissues. The walking fern shown here produces offspring when the tips of its leaves touch the soil. Vegetative reproduction A form of asexual reproduction in which an individual is produced from the nonsexual tissues of a parent. Clones Individuals that descend asexually from the same parent and bear the same genotype. Binary fission Reproduction through duplication of genes followed by division of the cell into two identical cells.
Parthenogenesis In contrast to vegetative reproduction, some organisms reproduce asexually by producing an embryo without fertilization, a process known as parthenogenesis. In most cases, parthenogenetically produced offspring arise from diploid eggs, which do not require any genetic contribution from sperm. Parthenogenesis has evolved in plants and several groups of invertebrates, including water fleas, aphids, and the Cape honeybees mentioned at the beginning of this chapter. Animal species that reproduce only by parthenogenesis are typically composed entirely of females. Parthenogenesis A form of asexual reproduction in which an embryo is produced without fertilization. Parthenogenesis is relatively rare in vertebrates. It has never been observed as a natural occurrence in mammals but has been confirmed in a few species of lizards, amphibians, birds, and fishes. For a long time, it was thought that snakes and sharks were not capable of parthenogenesis. In 2007, however, researchers confirmed that a virgin female hammerhead shark (Sphyrna tiburo) gave birth to daughters that were genetically identical to the mother. In 2011, researchers discovered that an isolated female boa constrictor gave birth to two litters of daughters through parthenogenesis (Figure 9.2). The growing evidence suggests that parthenogenesis may be more common than we currently appreciate and that many species can reproduce through both sexual and asexual reproduction.
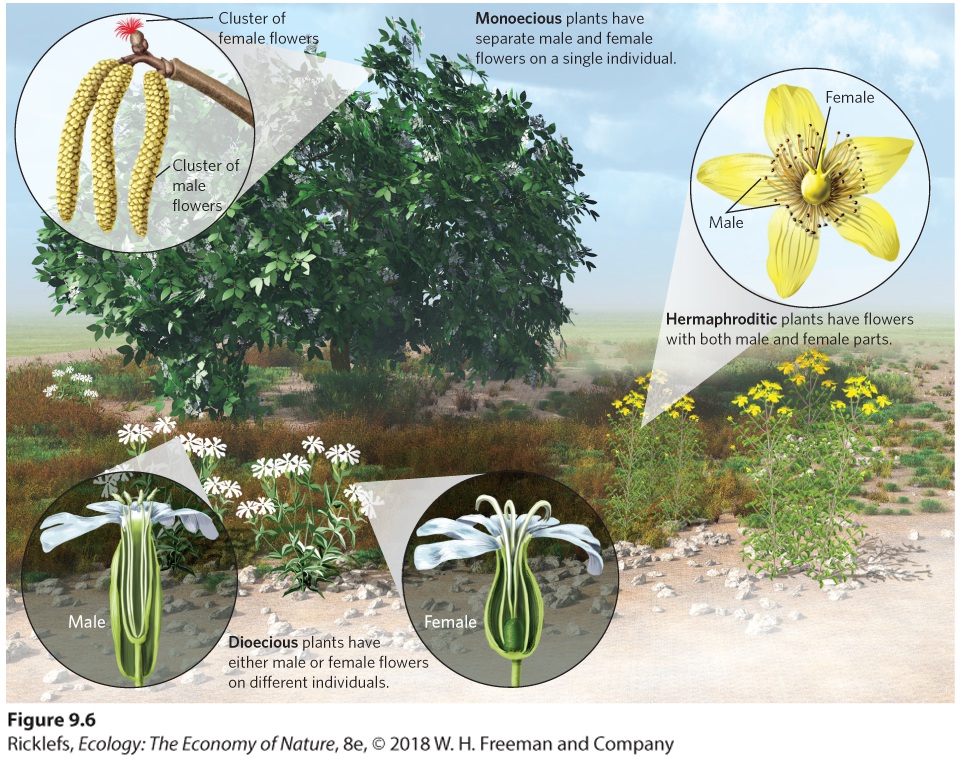
Figure 9.2 A fatherless boa constrictor. This female boa constrictor was the product of parthenogenesis. As a result, the recessive caramel coloration of the mother was passed on to all her clonal daughters. Parthenogenesis can produce offspring that are either clones of the parent or offspring that are genetically variable. Clones are produced when germ cells develop directly into egg cells without going through meiosis. In contrast, genetically variable offspring are produced when germ cells proceed partially or entirely through meiosis. In partial meiosis, germ cells pass through the first meiotic division, but suppression of the second meiotic division results in diploid egg cells. Although a sexual union is not involved, these eggs differ from one another genetically because of recombination between pairs of homologous chromosomes and the independent assortment of chromosomes during the first meiotic division. When the germ cells experience complete meiosis, the gamete-forming cells of the female are haploid and then fuse to form a diploid embryo. We saw an example of this in the Cape honeybee.
Costs of Sexual Reproduction
Sexual and asexual reproduction are both viable strategies, but sexual reproduction comes with a number of costs to the organism. For example, sexual organs require considerable energy and use resources that could be devoted to other purposes. In addition, mating itself can be a substantial task. Many plants must produce floral displays to attract pollinators and most
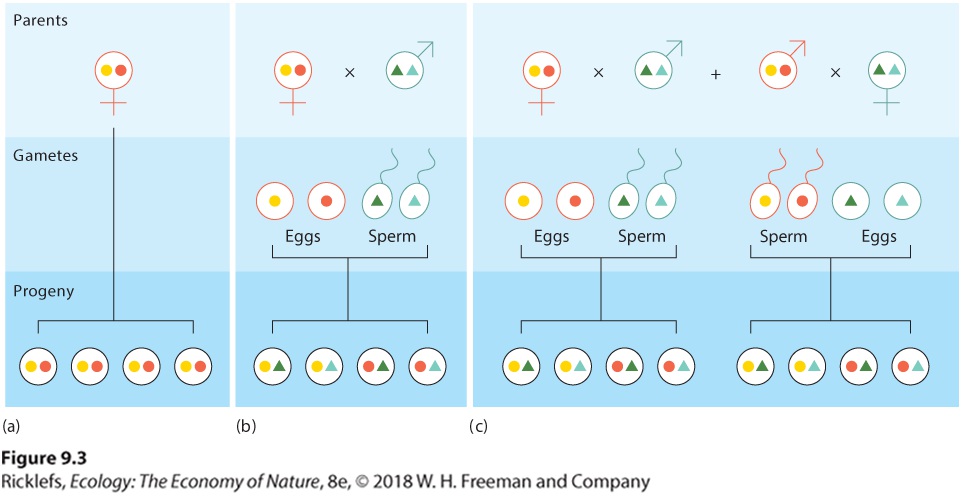
animals conduct elaborate courtship rituals to attract mates. These activities require time and resources. They can also elevate the risk of herbivory, predation, and parasitism. For organisms in which an individual is either male or female, sexual reproduction has an additional cost of reduced fitness. To understand this cost, we need to remember that every parent’s goal is to maximize their fitness by contributing as many copies of its genes as possible to the next generation. In the case of asexual reproduction, as shown in Figure 9.3a, a parent contributes two sets of chromosomes to each of its offspring. In the case of sexual reproduction, each parent contributes only one set of chromosomes to each offspring because the gametes produced by meiosis are haploid, as shown in Figure 9.3b. Females using either mode of reproduction produce the same number of offspring, but the female parent using sexual reproduction leaves behind half as many copies of its genes as a female using asexual reproduction. This 50 percent reduction due to sexual versus asexual reproduction is known as the cost of meiosis.
Figure 9.3 The cost of meiosis. If two hypothetical female organisms can only produce four eggs, (a) the female that uses asexual reproduction contributes eight copies of her genes. (b) In contrast, the female that uses sexual reproduction only contributes four copies of her genes. (c) A hermaphrodite can pass on eight copies of its genes by producing four eggs that are fertilized by another individual’s sperm and four copies of its genes when it uses its sperm to fertilize another individual’s eggs. Cost of meiosis The 50 percent reduction in the number of a parent’s genes passed on to the next generation via sexual reproduction versus asexual reproduction.
The cost of meiosis is counterbalanced in hermaphrodites, individuals that both possess both male and female function, a situation found in most plants and in many invertebrates. Such an individual can contribute one set of its genes to offspring produced through female function and another set to offspring produced through male function. As shown in Figure 9.3c, this allows a hermaphrodite to contribute twice as many copies of its genes to its offspring than is possible for an individual that can be only male or female. The cost of meiosis can also be offset when an individual is either male or female and the male helps the female take care of the offspring. For example, if a male’s parental assistance allows the female to raise twice as many offspring than she could care for alone, then the cost of meiosis is eliminated.
Benefits of Sexual Reproduction
If sexual reproduction is so costly, then it must persist because it provides substantial benefits. These benefits include purging harmful mutations and creating genetic variation that helps offspring deal with future environmental variation, including the existence of rapidly evolving parasites and pathogens. Purging Mutations Mutations occur in all organisms, and some are harmful. In asexually reproducing organisms, mutations are transmitted from one generation to the next, and so accumulate over generations, especially if the asexual parents produce clonal offspring. In contrast, during the random assortment of genes during meiosis in sexually reproducing organisms, deleterious mutations may not be transmitted to gametes. Further, of all the gametes that are produced, those that form zygotes may not contain the mutation. Alternatively, if both parents carry a copy of a gene with a harmful mutation, and some of the gametes produced do contain the mutation, then a union of two such gametes will be homozygous for the harmful mutation. An offspring that is homozygous for the harmful mutation is likely not viable and will fail to breed and pass the mutation to the next generation. Because species that reproduce only asexually do not have any means of purging mutations, deleterious mutations slowly accumulate over many generations. In time we would expect individuals of such a species to experience poor growth, survival, and reproduction, which would lead to eventual extinction. If this hypothesis is correct, then species that use asexual
reproduction would probably have gone extinct by now and species that use asexual reproduction would have adopted this mode of reproduction only recently. To test the hypothesis that asexual reproducing species do not persist in nature as long as sexual reproducing species, we can look at the patterns of asexual reproduction within a phylogeny. If the hypothesis is correct, we should observe that asexual reproduction has evolved relatively recently. For example, most vertebrate species that reproduce asexually belong to genera that have a sexual ancestor and contain mostly sexual species, with asexual species that are relatively recently evolved. We observe this pattern with salamanders in the genus Ambystoma, fishes in the genus Poeciliopsis, and lizards in the genus Cnemidophorus. These observations suggest that purely asexual species typically do not have long evolutionary histories. If they did, we would expect to see large groups of related species—such as most species within a genus—all using asexual reproduction. Such a pattern would suggest that their common ancestor used asexual reproduction. However, the longterm evolutionary persistence of asexual populations appears to be low, which supports the explanation that the accumulation of mutations and lack of genetic variation cause species that reproduce asexually to go extinct. However, not all asexual species fit this pattern. For example in the bdelloid rotifers, an ancient group of more than 300 species of freshwater and soil organisms, all species are asexual and entirely female. Similarly, some groups of protists have existed for hundreds of millions of years and appear to lack sexual reproduction. One way that such species could avoid extinction is by producing offspring more rapidly than new deleterious mutations arise. In this case, some individuals would always retain the nonmutated parental genotype and produce the next generation, a process known as clonal selection. However, groups such as these continue to challenge our efforts to understand the full range of costs and benefits that favor the evolution of sexual or asexual reproduction. Genetic Variation and Future Environmental Variation A second benefit of sexual reproduction is the production of offspring with greater genetic variation. If the environment were homogeneous across time and space, parents that are well adapted to the environment could use asexual reproduction to produce clonal offspring that are also well adapted. However,
as we have discussed in earlier chapters, environmental conditions typically change across time and space. As a result, offspring are likely to encounter different environmental conditions than their parents did. Because environmental conditions vary, offspring with genetic variation have an increased probability of possessing gene combinations that will help them adapt to different environmental conditions. However, most theoretical models conclude that a greater ability to adapt to temporal and spatial variation in the physical environment does not produce a large enough advantage to offset the cost of meiosis. However, temporal and spatial variation in the biotic environment—particularly variation in the occurrence of different pathogens—could provide a large advantage to the genetic variation created by sexual reproduction. Genetic Variation and Evolving Parasites and Pathogens To understand why sexual reproduction provides an evolutionary benefit when species experience variation in both types and abundance of pathogens, we first must realize that pathogens have much shorter generation times and much larger population sizes than most of the host species they infect. Because pathogens have the potential to evolve at a much faster rate than their hosts, they can evolve ways to counteract host defenses. Without rapid host evolution, pathogens could drive their hosts to low numbers or even to extinction. For example, in 1998 researchers described a newly discovered species of pathogen that was causing widespread amphibian deaths in Central America. The pathogen is a type of chytrid fungus (Batrachochytrium dendrobatidis) that can infect a wide variety of amphibian species. By 2012, the fungus had been detected on every continent inhabited by amphibians. In some parts of the world, including Central America, it appears that this deadly pathogen may have been recently introduced and that many species of frogs in the region have no adaptations to combat it. As a result, it appears that the pathogen has caused dozens of species to go extinct. The harmful effects of pathogens place a premium on hosts to evolve new defenses rapidly. As we have seen, sexual reproduction produces offspring with a greater range of genetic combinations, and some of these combinations might be better suited to combat the pathogen. In short, there is an evolutionary race between hosts, which are trying to evolve adaptations rapidly enough to combat the pathogen, and pathogens, which are trying to
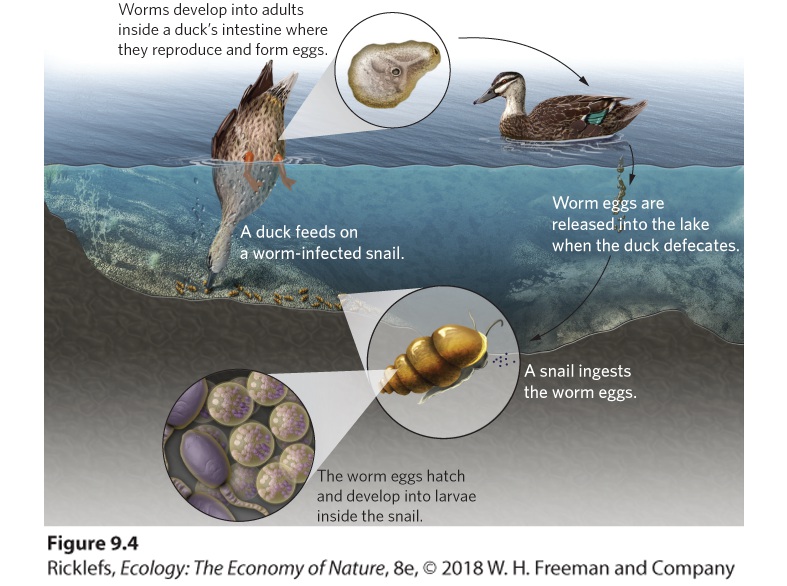
evolve adaptations rapidly enough to counter host defenses. The hypothesis that sexual selection allows hosts to evolve at a rate sufficient to counter the rapid evolution of parasites is called the Red Queen hypothesis, after the famous passage in Lewis Carroll’s Through the Looking-Glass, and What Alice Found There, in which the Red Queen tells Alice, “Now, here, you see, it takes all the running you can do, to keep in the same place.” Red Queen hypothesis The hypothesis that sexual selection allows hosts to evolve at a rate that can counter the rapid evolution of parasites. Testing the Red Queen Hypothesis One of the most compelling tests of the Red Queen hypothesis focuses on a species of freshwater snail (Potamopyrgus antipodarum) that is a common inhabitant of lakes and streams in New Zealand. The snails can become infected by parasites, including trematode worms in the genus Microphallus. The life cycle of the pathogen is shown in Figure 9.4. The worm’s life cycle begins when the snails ingest worm eggs. The eggs hatch into larvae that form cysts in the snail gonads, causing the snails to become sterile. Ducks then eat the infected snails and the pathogens mature sexually inside the intestines of the ducks, where they produce eggs asexually. These eggs exit the duck when it defecates in the water, thereby completing the cycle. Not surprisingly, Microphallus is most abundant in shallow waters of lakes where ducks feed.
Figure 9.4 A pathogen’s life cycle through snails and ducks. Eggs of the pathogenic worm Microphallus are inadvertently consumed by snails. The eggs hatch into larvae and form cysts in the snails, causing the snails to become sterile. When the snails are consumed by ducks in the shallow waters, the worm turns into an adult and reproduces in the ducks’ intestines. When the ducks defecate, the eggs are deposited back into the water. The snail’s mode of reproduction depends on the depth of water in which it lives. In the shallower regions of the lake, where the parasitic worm is more common, a higher proportion of the snails use sexual reproduction. Such populations contain about 13 percent males—enough to maintain some genetic diversity through sexual reproduction. In deeper regions, where the parasite is rare, a higher proportion of the snails use asexual reproduction. Although asexual populations reproduce faster than sexual populations, asexual clones cannot persist in the face of high rates of parasitism. As a result, the asexual snails do not survive well in the shallower regions of the lake, where they are more likely to encounter the parasitic worm. If the parasitic worms evolve to specialize on the snails with which they coexist, then parasites living in the shallow water should be good at infecting snail populations that live in shallow water. Similarly, other parasites that live in deep water should be good at infecting snail populations in that region. Researchers tested this hypothesis with parasites and snails from several different lakes in New Zealand. As shown in Figure 9.5, shallow-water snails

from several different lakes were infected most readily by shallow-water parasites, and deep-water snails were infected most readily by deep-water parasites. Averaged across all parasite sources, infection rates were relatively low in deep-water snails because few parasites live in this region and, therefore, have had less opportunity to evolve an ability to infect the deepwater snails. However, because deep-water habitats contain fewer parasites, asexual lineages of snails have a reproductive advantage over sexual lineages because of their more rapid reproduction.
Figure 9.5 Snail infection by a pathogen. Shallow water holds more parasites than deep water because ducks defecate and release pathogen eggs more frequently in shallow water. The large number of pathogens allows them to evolve rapidly in response to any changes in the snails. Snails in shallow water are challenged by a need to evolve defenses rapidly against the pathogens that respond quickly. As a result, a much higher percentage of shallowwater snails than deep-water snails become infected. The pathogens also show an increased ability to infect the populations of snails with which they coexist. Pathogens from shallow water are better able to infect shallow-water snails than deep-water snails. Similarly, pathogens from deep water are better able to infect deep-water snails than shallow-water snails. Recent studies have continued to support the Red Queen hypothesis. In the roundworm (Caenorhabditis elegans), for example, researchers raised individuals in the laboratory that were genetically destined to reproduce either sexually or asexually and then exposed populations containing two
different types of worms to a bacterial parasite. When researchers allowed the bacteria to evolve to infect the worms, the parasite quickly drove the asexual worms to extinction. In contrast, the sexual worms continually evolved resistance against the parasite and persisted. When the researchers prevented the bacteria from evolving, the asexual worms came to dominate the population.
Concept Check
1. What are the benefits of asexual reproduction? 2. What are the costs of sexual reproduction? 3. How does genetic variation help a population survive pathogens?
#### 9.2 Organisms Can Evolve as Separate Sexes or as Hermaphrodites
9.2 Organisms can evolve as separate sexes or as hermaphrodites Species in nature have evolved an incredible array of sexual strategies for male and female function, as you can see in Figure 9.6. Most vertebrates have separate sexes, whereas most plants are hermaphrodites. Hermaphroditic plants, such as the wildflower known as Saint-John’s-wort (Hypericum perforatum), possess male and female functions in the same flower. Flowers that have both male and female parts within each flower are known as perfect flowers. When the two functions are produced at the same time, we call it a simultaneous hermaphrodite. Examples of simultaneous hermaphrodites include many species of mollusks, worms, and plants. When an individual possesses one sexual function and then switches to the other, we call it a sequential hermaphrodite. Some species of plants are sequential hermaphrodites, as are some mollusks, echinoderms, and fishes.
Figure 9.6 Plant mating strategies. Hermaphroditic plants, such as the St.-John’s-wort wildflower, possess perfect flowers that contain male and female structures within a single flower. Monoecious plants, such as hazelnut shrubs, have separate male and female flowers, but a single plant possesses both types of flowers. Dioecious plants, such as the white campion wildflower, include some individuals that only contain male flowers and other
individuals that only contain female flowers. Perfect flowers Flowers that contain both male and female parts. Simultaneous hermaphrodites Individuals that possess male and female reproductive functions at the same time. Sequential hermaphrodites Individuals that possess male or female reproductive function and then switch to possess the other function. Some plants produce separate male and female flowers. Plants that have separate male and female flowers on the same individual plant are known as monoecious. For example, every hazelnut tree (Corylus americana) has separate male flowers and female flowers. When an individual plant contains only male flowers or only female flowers, the species is known as dioecious. For example, the white campion (Silene latifolia) is a wildflower that includes some individuals that produce only male flowers and other individuals that produce only female flowers. Although perfect-flowered hermaphrodites account for more than two-thirds of flowering plant species, nearly all-imaginable sexual patterns are known. Populations of some plant species can be composed of a complex mixture of hermaphrodites, males, females, and monoecious individuals. In other species, individual plants produce both perfect flowers and flowers that are only male or only female. Monoecious Plants that have separate male and female flowers on the same individual. Dioecious Plants that contain either only male flowers or only female flowers on a single individual.
Comparing Strategies
We would expect natural selection to favor the reproductive strategy with the highest fitness. For instance, in organisms such as flowering plants, a plant could evolve to make male flowers, female flowers, or hermaphrodite flowers. To determine whether evolution should favor separate sexes or hermaphrodites, we need to compare the amount of fitness an individual would gain by investing in only male or female reproduction versus the amount of fitness it would gain by investing in both male and female
reproduction. As depicted in Figure 9.7, if a male individual can invest in female function and gain a lot of female fitness while only giving up a small amount of male fitness, then selection will favor the evolution of hermaphrodites. A similar scenario can be considered for a female individual that adds male function. This occurs because the total fitness as a hermaphrodite through male plus female function exceeds the fitness of being only a male or only a female. In the case of flowers, the basic flower structure and the floral display necessary to attract pollinators—for those species that rely on them—are already in place in male flowers and female flowers. This should make the fitness costs of adding a sexual function relatively small while providing large fitness benefits. As we noted earlier, about two-thirds of all flowering plant species are hermaphrodites.
Figure 9.7 When hermaphrodites have a fitness advantage over separate sexes. When male or female individuals can add the other sexual function with little decline in original sexual function, they can achieve a higher total fitness by being a hermaphrodite than by being only male or female. In this example, the fitness of a hermaphrodite equals the fitness derived from male function (HM) plus the fitness derived from female function (HF). In some cases, the cost of investing in a second sexual function is too high to be offset by the benefits of being a hermaphrodite. Figure 9.8 illustrates how a reduction in female function can allow an investment in male function, but
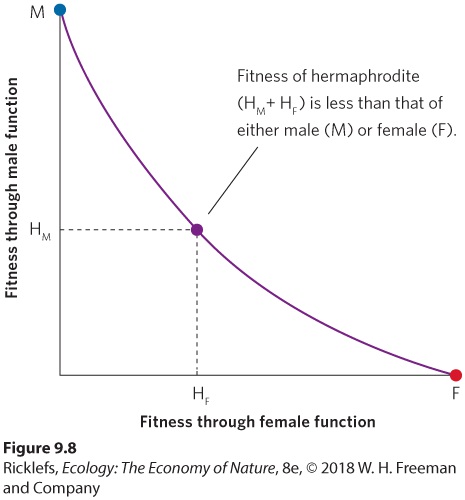
the total fitness is less than males and females would experience if they were to retain a single sexual function. For instance, sexual function in complex animals requires gonads, ducts, and other structures for transmitting gametes. Moreover, in many animals, being male requires large expenditures of time and energy for attracting mates and fighting with other males, while being female requires specializations for egg production or time and energy needed to care for offspring. Because these costs can be quite high, we would predict that hermaphroditism should occur only rarely among animal species that actively seek mates and engage in brood care. In contrast, we would predict that hermaphroditism should occur commonly among sedentary aquatic animals that mate by simply shedding their gametes into the water. Researchers have found evidence that supports both of these predictions.
Figure 9.8 When separate sexes have a fitness advantage over hermaphrodites. When male or female individuals add the other sexual function and experience a large decline in original sexual function, they can achieve a higher total fitness by staying as separate sexes rather than being a hermaphrodite. In this example, the fitness of a hermaphrodite equals the fitness derived from male function (HM) plus the fitness derived from female function (HF).
Selfing Versus Outcrossing of Hermaphrodites
One of the challenges for individuals that have both male and female function is the problem of self-fertilization. Self-fertilization, also known as selfing, occurs when an individual uses its male gametes to fertilize its own female
gametes. As we discussed in Chapter 4, self-fertilization poses a fitness cost due to inbreeding depression. Therefore, selection should favor individuals that do not use selfing when they have an opportunity to breed with other individuals, a strategy known as outcrossing. Some species avoid the problems of selfing by being sequential hermaphrodites. For example, the blue-headed wrasse (Thalassoma bifasciatum), a fish species common in coral reefs, can be functionally female when it is a small adult but then becomes functionally male later in life when it has grown larger. Similarly, if a plant releases the pollen in its anthers before making its stigma receptive to pollen, the flower will not be able to pollinate itself. Other species have selfincompatibility genes. Individuals with the same self-incompatibility genotype—including an individual mating with itself—cannot produce offspring.
Mixed Mating Strategies
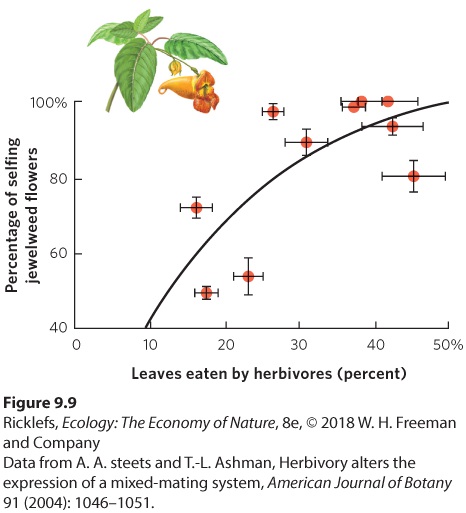
As we discussed in Chapter 4, some hermaphrodites use a mixture of mating strategies. When a mate can be found, the individual prefers to breed by outcrossing to avoid the costs of inbreeding. When a willing mate cannot be found, the individual self-fertilizes. Self-fertilizing will not provide as many viable offspring as outcrossing, but it might be better than not reproducing at all. In some cases, using a mixture of outcrossing and selfing is a response to a lack of resources. Attracting mates can be energetically expensive, as is the case with plants that must produce nectar to attract pollinators. For example, in orange jewelweed, the production of outcrossing flowers is more energetically expensive than the production of selfing flowers, which do not need to invest in nectar to attract pollinators. Plants that experience herbivory of their leaves have less energy to make outcrossing flowers. As a result, these plants produce a higher proportion of selfing flowers, as shown by the data for orange jewelweed in Figure 9.9.

Figure 9.9 Mixed mating strategies in orange jewelweed. In a survey of populations, those populations receiving higher amounts of herbivory also produced higher proportions of selfing flowers and, therefore, a lower proportion of outcrossing flowers. Error bars are standard errors.
1. What is the difference between simultaneous and sequential hermaphrodites? 2. When the fitness increment of increased male function results in a large cost in fitness of female function, would a population evolve separate sexes rather than hermaphrodites? 3. When the fitness increment of increased male function results in a small cost in fitness of female function, would a population evolve hermaphrodites?
#### 9.3 Sex Ratios of Offspring Are Typically Balanced, but They Can Be Modified by Natural Selection
9.3 Sex ratios of offspring are typically balanced, but they can be modified by natural selection In organisms with separate sexes, the sex ratio of male to female offspring is often one to one. The exceptions offer interesting insights into evolutionary forces that favor particular sex ratios in an individual’s offspring. In this section, we will examine the mechanisms that determine whether an offspring will be male or female. We will then examine the underlying reasons for the wide range of sex ratios observed in nature.
Mechanisms of Sex Determination
In previous chapters, we learned that the phenotypes of organisms are determined by both genetic and environmental factors. The sex of offspring is no different, although the influence of genetics and the environment on the sex of offspring differs among species. Genetic Sex Determination In mammals, birds, and many other organisms, sex is determined by inheritance of sex-specific chromosomes. In most mammals, females have two X chromosomes, whereas males have one X chromosome and one Y chromosome. Birds have the opposite pattern of genetic sex determination: males have two copies of the Z chromosome, whereas females have one Z chromosome and one W chromosome. In both cases, the sex that has two different chromosomes—male mammals and female birds—produces an approximately equal number of gametes containing each sex chromosome. On average, half the progeny in these populations will be female and half will be male. In insects, genetic sex determination is accomplished in a variety of ways. In grasshoppers and crickets, for instance, all individuals are diploid, but females have two sex chromosomes, whereas males have only one. In honeybees and other members of their order, including other bees, ants, and wasps, sex is determined by whether or not an egg is fertilized. Fertilized eggs, which receive two sets of chromosomes, become females, whereas unfertilized eggs become males.
Environmental Sex Determination In some species, sex is determined largely by the environment, in a process known as environmental sex determination. In reptiles, including several species of turtles, lizards, and alligators, the sex of an individual is determined by the temperature at which the egg develops. In turtles, embryos incubated at lower temperatures typically produce males, whereas those incubated at higher temperatures produce females. The reverse is generally true in alligators and lizards. This type of environmental sex determination is known as temperature-dependent sex determination. Because the genotype has the ability to produce multiple phenotypes, temperature-dependent sex determination is a type of phenotypic plasticity. Environmental sex determination A process in which sex is determined largely by the environment. For decades, biologists have wondered whether temperature-dependent sex determination is adaptive. For example, if the temperatures that cause eggs to become male produce the most fit males, and the temperatures that cause eggs to become female produce the most fit females, temperature-dependent sex determination would be adaptive. To test this hypothesis, one would need to produce male and female offspring from eggs incubated across a range of different temperatures. However, since temperature is the very factor that determines their sex, we cannot naturally produce males and females at each temperature and compare their performance. This problem was solved in a study of the Jacky dragon (Amphibolurus muricatus). In this lizard species from Australia, females are produced when incubated at low and high temperatures, whereas both males and females are produced when incubated at intermediate temperatures. Because of this pattern, researchers could easily produce females at all three temperatures. To produce males at the highest and lowest temperatures, the researchers injected a hormone inhibitor that prevented the embryos from becoming female, thereby overriding the normal response to temperature effects. This manipulation allowed the researchers to produce male and female lizards at all three temperatures. Once the lizards hatched, the animals were put outside in field enclosures for 3 years. At the end of 3 years, the researchers determined the number of offspring produced by adults that had been incubated as eggs at different temperatures. As shown
in Figure 9.10, males incubated at the intermediate temperature subsequently fathered more offspring than males incubated at the low and high temperatures. Females that incubated at the high temperature subsequently laid more eggs than females incubated at intermediate temperatures, although females incubated at low temperatures laid a similar number of eggs as females incubated at intermediate temperatures. This was one of the first studies in reptiles to demonstrate that temperature-dependent sex determination appeared to be adaptive.
Figure 9.10 Environmental sex determination in the Jacky dragon. Under natural conditions, the eggs of the Jacky dragon become female under the low and high temperatures. Eggs become both male and female under intermediate temperatures. Female fitness is highest for females raised at the high temperature. Male fitness is highest for males raised at the intermediate temperature. Error bars are standard errors. Not all cases of environmental sex determination are driven by temperature. In some species, sex determination is driven by the social environment in which an individual lives. As mentioned in our discussion of sequential hermaphrodites, the blue-headed wrasse is a coral reef fish that is able to change from female to male as it ages (Figure 9.11). Blue-headed
wrasses typically begin life as females and live in large schools along with one or a few dominant males. If the dominant male leaves or dies, the largest female will then change sex and become the new dominant male. Being large is important for males, because they must defend a territory from other males that attempt to breed with the females in the school.
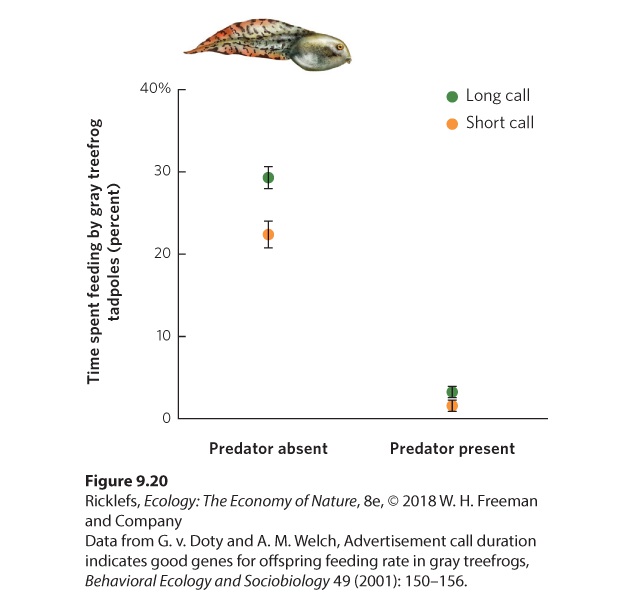
Figure 9.11 Blue-headed wrasse. Young fish are typically female and live in schools with a dominant male. If the male leaves or dies, the largest female changes into a new dominant male.
Offspring Sex Ratio
Now that we understand the mechanisms that help determine the sex of offspring, we can consider the factors that favor particular sex ratios of offspring. Biologists increasingly appreciate that a female can have a large influence on the sex ratios of her offspring. In species where the males have two different types of sex chromosomes, as in mammals, researchers have found that the females of some species can control whether X-chromosome or Y-chromosome sperm are allowed to fertilize their eggs. In species where the females contain two different types of sex chromosomes, as in birds, the females of some species can determine the sex ratio of their offspring by controlling the fraction of their eggs that receive the Z versus the W chromosome during meiosis. In the hymenopteran insects—bees, wasps, and ants—the female determines the sex of her offspring by whether or not she fertilizes the eggs.

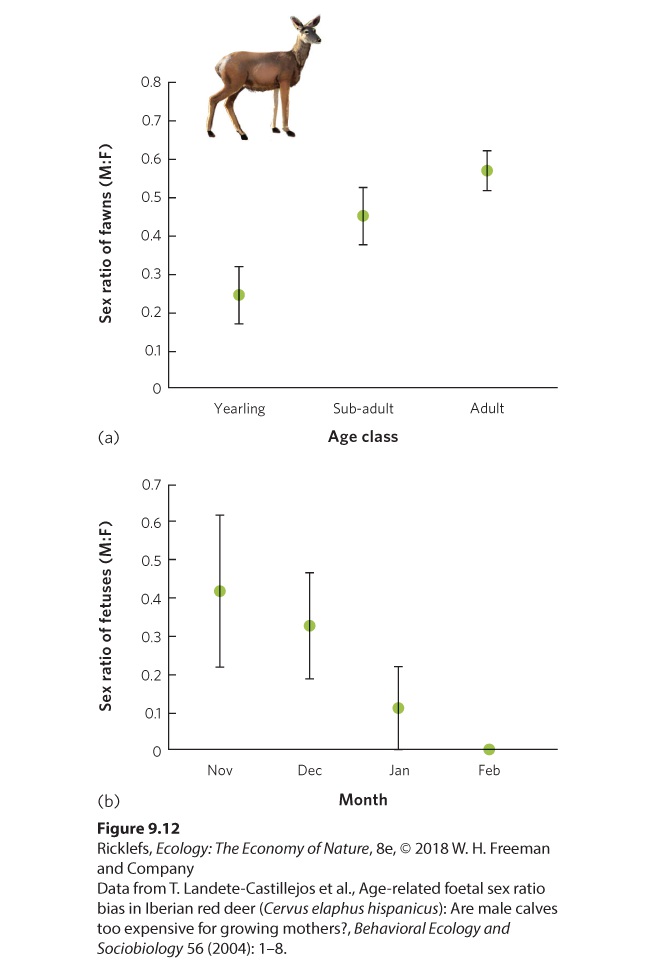
A different approach to controlling offspring sex ratio is through selective abortion. In red deer (Cervus elaphus), for example, adult females breed in early autumn and give birth the following spring. Sons are often larger at birth and require more of the mother’s milk than daughters. As a result, sons require a larger investment from the mother than daughters. Researchers in Spain examined the fetuses of 221 harvested red deer to determine whether the sex ratio of the fetuses was affected by the mother’s age—categorized as adult, subadult, or yearling—and whether the sex ratio changed through the gestation period. As shown in Figure 9.12a, adult mothers produced a nearly even offspring sex ratio. In contrast, yearling mothers, which are smaller and have less energy, were much more likely to produce the less energetically expensive daughters. Averaged across all months of the deer harvest, yearling mothers carried about 25 percent male fetuses and 75 percent female fetuses. To determine how the yearling mothers accomplish this uneven sex ratio, researchers examined yearling mothers that were harvested at different months throughout the winter, illustrated in Figure 9.12b. They found that the yearling mothers initially had a nearly even sex ratio of offspring. As winter progressed, however, the proportion of male fetuses declined sharply, suggesting that the yearling mothers were capable of selectively aborting the more expensive male fetuses as gestation progressed. This phenomenon is not restricted to red deer; it has also been observed in other mammals, including rats and mice.
Figure 9.12 Offspring sex ratios in red deer. (a) Yearling deer produce a low proportion of male fawns, but this proportion increases in older deer. (b) Among yearling deer, the proportion of male fetuses is initially high but then declines as gestation continues throughout the winter as female deer selectively abort male fetuses. Error bars are standard errors. Regardless of how the sex ratio is controlled, in most species the sex ratio of male to female offspring is nearly even. Is this a ratio produced by default, or are there adaptive reasons for it? To answer this question, we can compare
the conditions that favor a one-to-one sex ratio with the conditions that favor a deviation from that ratio. Frequency-Dependent Selection To understand how selection commonly favors a one-to-one sex ratio, we have to consider how natural selection might favor the offspring sex ratio that an individual produces when a population deviates from the one-to-one ratio. Let’s imagine, for instance, that a population has more females than males and each male breeds with only one female. In this situation, some females would remain unbred. A parent that produces all sons would have higher fitness than a parent that produces an even number of sons and daughters, because some of the daughters might not find mates. Similarly, if a population has a surplus of males and one male breeds with only one female, a parent that produces all daughters would have higher fitness than a parent that produces an even number of sons and daughters, because some of the sons might not find mates. As you can see in this example, individuals of the less abundant sex enjoy greater reproductive success because they compete with fewer individuals of the same sex for breeding. Thus, whenever the population has an abundance of one sex, natural selection will favor any parents that produce offspring of the less abundant sex. Over time, as the less abundant sex becomes more common and the common sex becomes less abundant, populations tend to be balanced at an approximate one-to-one ratio. Therefore, the best sex ratio strategy for parents depends on the frequencies of males and females in a population. The evolution of the sex ratio is said to be the product of frequency-dependent selection, which occurs when natural selection favors the rarer phenotype in a population. Frequency-dependent selection When the rarer phenotype in a population is favored by natural selection. Researchers are beginning to appreciate that human activities can cause unnatural changes in sex ratios. For example, in southwest Alaska, the sockeye salmon (Oncorhynchus nerka) males are typically larger than the females, but the magnitude of this size difference between the sexes varies naturally among the populations. Now consider that the regulations for salmon fishing require that anglers only
harvest the larger fish. This causes anglers to inadvertently harvest more males than females. In 2012, researchers reported that the sex ratios among the different populations range from 36 to 47 percent males due to the longterm effects of fishing, as illustrated in Figure 9.13.
Figure 9.13 Sex ratios in sockeye salmon populations. Among populations of sockeye salmon in southwest Alaska, those with much large males than females are experiencing declines in the ratios of males to females due to fishing regulations that require harvesting the largest fish. Highly Skewed Sex Ratios We have seen that natural selection favors a one-to-one sex ratio. In some cases, however, we observe highly skewed sex ratios, such as in Fig wasps (Pegoscapus assuetus). The female fig wasp arrives at a flower of a fig tree carrying pollen from another fig tree. She crawls in through a small hole in the flower and pollinates the flower. Once inside, the female lays her eggs in the developing fruit and then dies. Her eggs can be composed of up to 90 percent daughters! Similar to the bees that we discussed at the beginning of this chapter, wasps can easily adjust the sex ratio of their offspring because
##### Analyzing Ecology: Frequency-Dependent Selection
fertilized eggs become daughters, whereas unfertilized eggs become sons. Once the eggs hatch, the larvae feed on the fig fruit and seeds. The larvae metamorphose into adult wasps and breed with their siblings while still inside the flower. The young males breed with the young females, chew a hole in the side of the flower, and then die. The fertilized young females then escape through the hole and fly off to pollinate a new fig flower.
Analyzing Ecology
Frequency-Dependent Selection We can better understand frequency-dependent selection if we work though an example with real numbers. The American black vulture (Coragyps atratus) is a large bird that feeds on animal carcasses throughout much of North and South America. The female vulture typically lays two eggs in its nest. If a population is composed of five females and two males, and we assume that a male can breed with more than one female, how many copies of genes, on average, does each male and female contribute to the next generation? Total number of eggs = 10 Average female fitness = 10 female gene copies ÷ 5 females = 2 gene copies/female Average male fitness = 10 male gene copies ÷ 2 males = 5 gene copies/male If a population is composed of five females and eight males, how many copies of genes, on average, does each male and female contribute to the next generation? Total number of eggs = 10 Average female fitness = 10 female gene copies ÷ 5 females = 2 gene
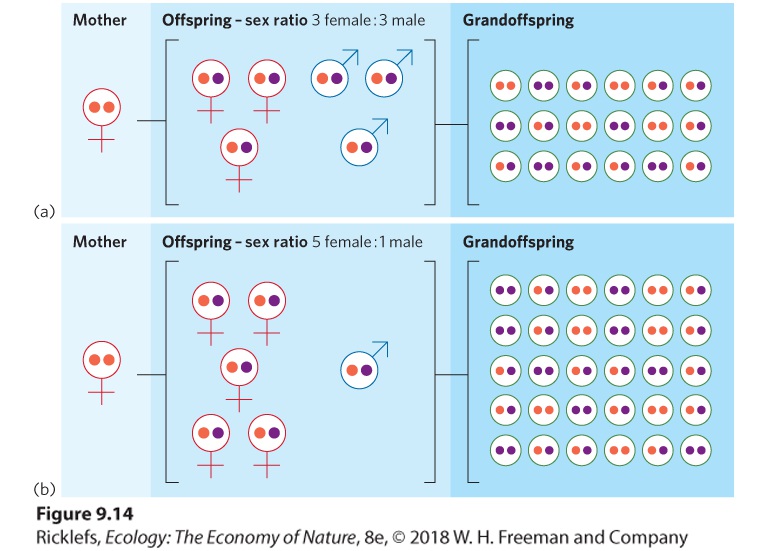
copies/female Average male fitness = 10 male gene copies ÷ 8 males = 1.25 gene copies/male In this example, you can see that when males are the less abundant sex, males have the higher fitness. When females are the less abundant sex, females have the higher fitness. YOUR TURN Using the same assumptions above, calculate the number of gene copies per male and per female under the following two scenarios: 1. Four males and five females 2. Six males and five females Given the four scenarios that we have explored, what sex ratio will be favored by natural selection over the long term? The high proportion of daughters among fig wasps occurs because male competition for females is among brothers. This phenomenon, known as local mate competition, occurs when competition for mates takes place in a limited area and only a few males are required to fertilize multiple females. When only one fertilized mother enters a fig flower, the only males available to fertilize her daughters are her sons. From the mother’s perspective, it does not matter which son passes along her genes to her grandoffspring. One son can fertilize multiple daughters, which means there is no fitness benefit to having multiple sons. The mother’s fitness is therefore a function of how many eggs can be produced by her daughters, so it is in the mother’s best interest to produce many daughters and just enough sons to fertilize those daughters. As you can see in Figure 9.14, when mating options are restricted such that the only mates available for daughters are their brothers, mothers that produce a higher proportion of daughters than sons will have more grandoffspring and therefore greater evolutionary fitness.
Figure 9.14 The effect of different sex ratios on a mother’s fitness. When one son can breed with multiple daughters in an isolated population, the sex ratio of the mother’s offspring will affect the number of grandoffspring that can be produced. In this example, we assume each female can produce six offspring. (a) If a mother produces three daughters and three sons, 18 grandoffspring can be produced. (b) If a mother produces five daughters and one son, 30 grandoffspring can be produced. Local mate competition When competition for mates occurs in a very limited area and only a few males are required to fertilize all the females. Sometimes, however, two or more females lay their eggs in the same flower. In this situation, a mother’s male offspring can mate either with their sisters or with the daughters of the other mothers. When this occurs, a mother will obtain greater fitness if she produces extra sons so she has enough sons to fertilize all the females in the flower. As expected, researchers observe that when a fig flower contains more than one mother laying eggs, the mothers lay a higher proportion of male eggs.
Concept Check
1. What is the difference between genetic and environmental sex determination?
2. When a population is composed of two sexes, why does the rarer sex have a fitness advantage? 3. How does local mate competition favor the production of femalebiased sex ratios in offspring?
#### 9.4 Mating Systems Describe the Pattern of Mating between Males and Females
9.4 Mating systems describe the pattern of mating between males and females Although many species of algae and fungi have gametes that are of similar size, animals and plants produce sperm that are a fraction of the size of eggs. This smaller sperm takes much less energy to produce than an egg. Because of this difference, a female’s reproductive success depends both on the number of eggs she can make and on the quality of the mates that she finds. Because most males can produce millions of sperm, a male’s reproductive success generally depends on how many females he can fertilize. In this section, we will discuss the mating system of species, which describes the number of mates each individual has and the permanence of the relationship among mates. Like sex ratios, the mating system of a population is subject to natural selection. Consequently, mating systems are often a product of the ecological conditions under which species live. Figure 9.15 illustrates the four mating systems: promiscuity, polyandry, polygyny, and monogamy.
Figure 9.15 Mating systems. Promiscuity occurs in species of outcrossing plants such as the prairie sunflower (Helianthus petiolaris). Polygyny exists when a male mates with several females, as in elk (Cervus canadensis). Polyandry occurs when a female mates with multiple males, as is the case for the western toad (Anaxyrus boreas). Monogamy for life was thought to be the rule in 90 percent of all birds, as in the sandhill crane (Grus canadensis). However, recent genetic analyses have confirmed that most bird species actually participate in extrapair copulations. Mating system The number of mates each individual has and the permanence of the relationship with those mates.
Promiscuity
Promiscuity is a mating system in which individuals mate with multiple partners and do not create a lasting social bond. Among animal taxa as a whole, promiscuity is by far the most common mating system. Promiscuity is universal among outcrossing plants because they send pollen out to fertilize the eggs of multiple individuals and receive pollen from multiple individuals. Promiscuity A mating system in which males mate with multiple females and females mate with multiple males and do not create a lasting social bond. When animals release eggs and sperm directly into the water or when pollen is released into the wind, much of the variation in mating success is random. Whether a particular sperm is the first to find an egg is largely a matter of chance. When males attract or compete for mates, however, reproductive success can be influenced by factors such as body size and the quality of courtship displays. Even when fertilization is random, males that produce the most sperm or pollen and males that produce the most competitive sperm or pollen are bound to father the most offspring.
Polygamy
Polygamy is a mating system in which a single individual of one sex forms long-term social bonds with more than one individual of the opposite sex. Most often, a male mates with more than one female, which is known as polygyny. In some species, polygyny evolves when males compete for females and the females all prefer only the few best males. In this case, the largest and healthiest males may mate with the vast majority of the females. Polygyny also can evolve when a male is able to defend a group of females from other males or when a male can defend a patchy resource that is attractive to multiple females. Consider the guanaco (shown in the opening to
Chapter 3), a relative of the camel that lives in the Patagonian region of South America. The land in Patagonia is generally dry, with patches of wet habitat that grow nutritious plants that guanacos prefer to eat. When a male guanaco defends a wet patch from other males, he is able to mate with many of the females who come to the wet patch to eat the plants.
Polygamy A mating system in which a single individual of one sex forms long-term social bonds with more than one individual of the opposite sex. Polygyny A mating system in which a male mates with more than one female. The opposite of polygyny is polyandry, a mating system in which a single female breeds with multiple males. The account of the queen honeybee at the beginning of this chapter is an example of polyandry because she mates with multiple drones. Polyandry commonly occurs when the female is in search of genetically superior sperm or has received material benefits from each male with whom she mates. For instance, in some species of insects, including some butterflies, the female receives a nutritional package of food—known as a spermatophore—from a male. The female uses the protein in the spermatophore to make her eggs. In polyandrous species, the more spermatophores that a female can collect from multiple males, the more eggs she can produce. Polyandry A mating system in which a female mates with more than one male.
Monogamy
Monogamy is a mating system in which a social bond between one male and one female persists through the period that is required for them to rear their offspring. In some cases, the bond may endure until one of the pair dies. Monogamy is favored when males can make important contributions to raising the offspring. Monogamy occurs in about 90 percent of bird species because male birds can offer much of the same care to offspring as females, such as incubating eggs, gathering food for the chicks, and protecting the chicks from predators. A male bird’s help enables his chicks to grow and survive much better than they would without his help. Therefore, his parenting improves his fitness. In mammals, however, fewer than 10 percent of species have a social bond with only one other individual. Because male mammals cannot provide the same offspring care as females, particularly because females lactate, the growth and survival of mammal offspring are less reliant on a male’s presence.
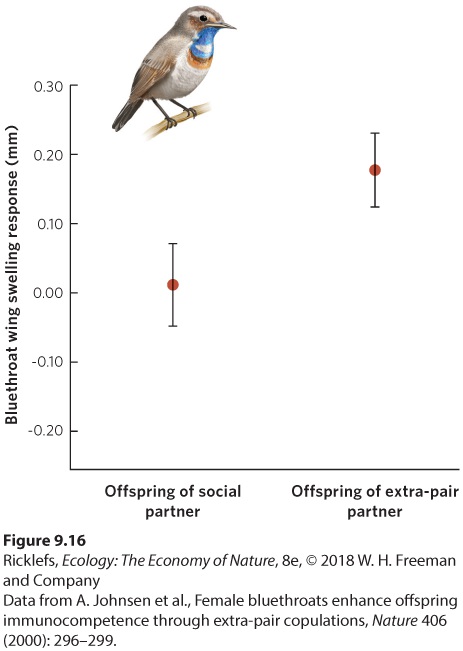
Monogamy A mating system in which a social bond between one male and one female persists through the period that is required for them to rear their offspring. Although most female birds form a single social bond with a male partner, DNA analysis of offspring in a nest has revealed that the offspring often have different fathers. This means that the mother copulated with other males. While these females had a social bond with only one male, they were actually breeding with other males, a behavior known as extra-pair copulation. In some monogamous species, a third or more of the clutches contain offspring sired by another male—usually, a male in a neighboring territory. Since neighboring males also have a social bond with their own partner, both males and females use extra-pair copulations. Thanks to DNA analysis, we now know that 90 percent of bird species thought to be monogamous engage in extra-pair copulations. Extra-pair copulations When an individual that has a social bond with a mate also breeds with other individuals. Extra-pair copulations surely increase the fitness of the neighboring males, but how does this behavior increase the fitness of the female? One way would be if her reproductive success is improved by creating greater genetic variation among her offspring so at least one might be better suited to future environmental conditions. She could also benefit if the neighboring males have better genotypes than her mate. In a bird known as the bluethroat (Luscinia svecica), for example, extra-pair copulations are common. Researchers in Norway examined the immune response of chicks whose father was the social partner of the female versus the extra-pair partner of the female. To do this, the researchers inject a small amount of foreign material —which is extracted from kidney beans—into a bird’s wing and then they measure the amount of swelling that occurs at the site of the injection; the greater the swelling, the stronger the bird’s immune response. As you can see in Figure 9.16, the amount of swelling in the wing was greater in offspring produced by extra-pair copulations, which demonstrates that they had a stronger immune response. This suggests that females seek extra-pair copulations as a way to obtain superior genotypes from another male and produce offspring that have superior immune systems.
Figure 9.16 Female benefits of extra-pair copulations. In bluethroats, the strength of the immune system can be measured in terms of a wing-swelling response when the wing is injected with a chemical extracted from kidney beans. Larger wing-swelling responses indicate stronger immune systems. Error bars are standard deviations. The male participating in a monogamous social bond does not benefit if his mate breeds with other males. The threat that she will participate in extra-pair copulations has selected for mate guarding, a behavior in which one partner prevents the other partner from participating in extra-pair copulations. A variety of mate guarding behaviors have evolved. In some species, an individual simply stays near its mate and keeps other potential suitors away, while in other cases, the mate makes future breeding physically impossible. For instance, researchers discovered that the golden orb-weaving spider (Argiope aurantia) inserts its sperm-transferring appendages into the two openings of the female’s reproductive system. Within a few minutes of transferring his sperm, the male’s heart stops and he dies with his appendages still stuck inside the female, making it impossible for the female to mate again. Such adaptations are effective at reducing the chance that an
individual’s partner will breed with another mate. Mate guarding A behavior in which one partner prevents the other partner from participating in extra-pair copulations. The various mating systems are the product of natural selection and are shaped by the ecological conditions under which each species lives. Because the mating systems often involve attracting a mate or defending against other members of the same sex, natural selection has also caused the evolution of many sex-specific traits, as we will see in the next section.
1. Under what conditions would natural selection favor the evolution of polygyny? 2. Under what conditions would natural selection favor the evolution of polyandry? 3. What behavior can a mate use to enforce monogamy?
#### 9.5 Sexual Selection Favors Traits That Facilitate Reproduction
9.5 Sexual selection favors traits that facilitate reproduction We have seen that a male’s reproductive success is typically determined by the quantity of females he can breed, whereas a female’s reproductive success is typically determined by the quality of males that can fertilize her limited number of eggs. This means that females should generally be the choosier sex. But what exactly should she be choosing? In broad terms, females should select those males that improve her fitness by the greatest amount. She might select the male with the best genotype or the male with the most resources for her and her offspring. Because females are choosy, males should compete strongly with other males for the opportunity to breed. This intense competition among males for mates has resulted in the evolution of male traits that are either used to attract females or used in contests and combat among males. Natural selection for sex-specific traits related to reproduction is referred to as sexual selection. In this section, we will explore how males and females have evolved different traits as a result of sexual selection, what traits females prefer in their mates, and how the fitness interests of males and females can cause conflict between the sexes. Sexual selection Natural selection for sex-specific traits related to reproduction.
Sexual Dimorphism
One result of sexual selection is sexual dimorphism, which is a difference in the phenotype between males and females of the same species. Sexual dimorphism is seen in honeybees and sockeye salmon, which we discussed earlier in this chapter. Sexual dimorphism includes differences in body size, ornaments, color, and courtship behavior. Traits related to fertilization—such as gonads—are referred to as primary sexual characteristics, while traits related to differences in body size, ornaments, color, and courtship are known as secondary sexual characteristics. Sexual dimorphism The difference in the phenotype between males and females of the same species.
Primary sexual characteristics Traits related to fertilization. Secondary sexual characteristics Traits related to differences between the sexes in terms of body size, ornaments, color, and courtship. Sexual dimorphism can evolve due to differences in life history between the sexes, contests between males, or mate choice by females. Body size differences are common between the sexes of many animals because there has been selection for an increased number of gametes produced or for an increase in parental care by one sex. In fish and spiders, for instance, egg production is directly related to body size. This selection for greater gamete production in females without concurrent selection for greater sperm production in males could be the underlying cause of the larger size of females than males in many species of spiders and fish (Figure 9.17).
Figure 9.17 Sexual size differences. In the golden silk spider (Nephila clavipes), females are much larger than males. Sexual dimorphism can also occur when males compete for mates. In this case, selection will favor the evolution of weapons for combat. Such weapons include the antlers of male elk, the horns of mountain sheep, and the leg spurs of chickens and turkeys. When fighting ability is also improved by having a large body, contests between males can also favor the evolution of larger
male bodies (Figure 9.18). Males that win such contests are more likely to gain access to females. Sexual dimorphism also commonly arises when one sex, usually the female, is particularly choosy when selecting a mating partner. In these cases, female selection of males with particular traits can cause the sexual selection for those traits, such as male deer growing antlers.
Figure 9.18 Male weapons. In some species, such as these elk in Alberta, Canada, males fight with each other for the right to breed females. Repeated contests cause selection for large weapons, including large antlers. Human activities can also affect sexual dimorphism. In species that are hunted, for example, hunters either prefer or are required to harvest only the largest males. This can be measured as the number of points on the antlers of deer or elk or the length of the curled horns in wild species of sheep. In doing so, we have created a situation of artificial selection that is opposite to what would occur naturally in sexual selection. In 2016, researchers reported the change in horn size in Stone’s sheep (Ovis salli stonei) based on differences in harvesting that occurred over a 37-year period across locations in British Columbia. Given that horn size is heritable in Stone’s sheep, greater harvesting represents stronger selection, which should then cause a strong evolutionary response, as we discussed in
Chapter 7. In the region of high hunting pressure, there were 2.7 times more hunters per unit area and 2.6 times more sheep harvested than the region of low-hunting pressure. Over the 37 years, the rate of horn growth declined by
12 percent in the region of high-hunting pressure, but showed no change in the region of low-hunting pressure. In short, increased hunting caused artificial selection for smaller horn size.
The Evolution of Female Choice
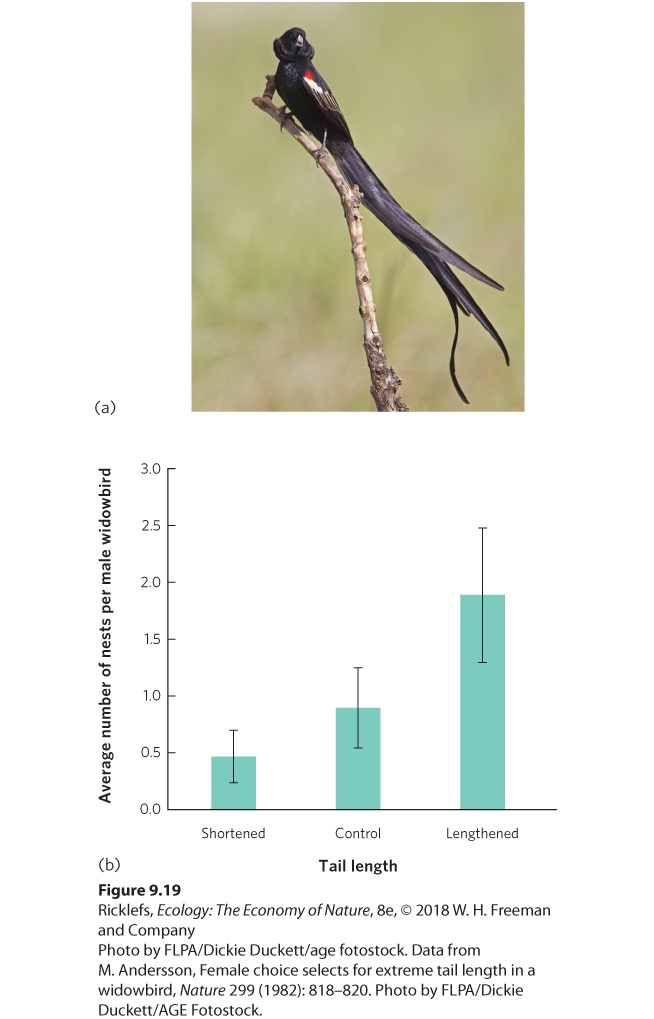
A female’s preference for particular male traits should be tied to those features that improve her fitness. In terms of broad categories, we can consider two types of female preferences: material benefits and nonmaterial benefits. Material benefits are those physical items that a male can provide a female, including a site for raising offspring, a high-quality territory, or abundant food. In these cases, the benefit to the female is straightforward—a site for raising offspring and resources for producing eggs and feeding the offspring should improve a female’s fitness. It has been more of a challenge for scientists to understand female choices when the female does not receive any material benefits from males. If the female choice is an adaptation, then there must be some benefit. One of the first demonstrations of female choice that provided no material benefit came from a study of tail length in male long-tailed widowbirds (Euplectes progne), a small, polygynous species that inhabits the open grasslands of central Africa. The females are mottled brown, short-tailed, and drab. In contrast, during the breeding season, males are jet black with a red shoulder patch, and they sport a half-meter-long tail that is conspicuously displayed in courtship flights (Figure 9.19a). The most successful males may attract up to a half dozen females to nest in their territories, but the males provide no care for their offspring. To determine what the females are choosing, researchers cut the tail feathers of some males to shorten them and glued the clipped feather ends onto the feathers of the tails of other males to lengthen them. As you can see in Figure 9.19b, males with experimentally elongated tails attracted significantly more females than those with shortened or unaltered tails.
Figure 9.19 Long-tailed widowbird. (a) During the breeding season, males have exceptionally long tails. (b) When researchers made male tails shorter, the same length, or longer than those of typical males, females preferred males with longer tails. Error bars are standard errors. Why would females choose a male based on his traits? There are two
possible hypotheses. According to the good genes hypothesis, an individual chooses a mate that possesses a superior genotype. In the gray treefrog, for example, females prefer males that can produce the longest mating calls. Long calls can only be produced by the largest and healthiest male frogs. If male size and health have a large genetic component, choosing this trait might benefit the female’s offspring. Indeed, when researchers forced females to breed with both long- and short-calling males, the offspring of long-calling males grew faster than the offspring of short-calling males. Subsequent research, illustrated in Figure 9.20, found that the offspring of long-calling males grew faster because they spend more time feeding compared to the offspring of short-calling males.
Figure 9.20 Good genes. In the gray treefrog, females who chose long-calling males produced offspring that spent more of their time eating than did the offspring of short-calling males, regardless of whether predators were present or absent. Error bars are standard errors. Good genes hypothesis
The hypothesis that an individual chooses a mate that possesses a superior genotype. According to a second hypothesis, known as the good health hypothesis, individuals choose the healthiest mates. Good health could be the outcome of either superior genetics or a superior upbringing with abundant resources. As a result, the good genes hypothesis and the good health hypothesis are not mutually exclusive. Females might prefer healthy males because such males may be both genetically superior and pose a lower risk of passing on a number of different parasites and diseases. Good health hypothesis The hypothesis that an individual chooses the healthiest mates.
Runaway Sexual Selection
Once female preference for a male trait has evolved, the trait may continue to evolve over time. For example, if females prefer longer tails in their mates, and there is genetic variation to select from, longer tails will continue to evolve in males. When selection for preference of a sexual trait and selection for that trait continue to reinforce each other, the result can be runaway sexual selection. Runaway sexual selection is thought to have favored the evolution of such extreme traits as the half-meter-long tails of the male widowbird, the giant tail fans of the peacock, and other large male ornaments such as horns, tusks, and antlers. Runaway selection continues until males either run out of genetic variation for the trait or until the fitness costs of possessing extreme traits begin to outweigh the reproductive benefits. Runaway sexual selection When selection for preference of a sexual trait and selection for that trait continue to reinforce each other.
The Handicap Principle
If sexually selected traits indicate intrinsic attributes of male quality—at least initially, before runaway sexual selection occurs—we are then faced with a paradox. Presumably, extreme traits burden males by requiring energy and resources to maintain them, and they make males more conspicuous to predators. If this is the case, it is hard to imagine how extreme traits indicate a higher-quality mate.
One intriguing possibility is that elaborate male secondary sexual characteristics act as handicaps. If a male can survive with sexual traits that require extra energy to build or that increase the risk of predation, these traits might signal a superior genotype. This idea, known as the handicap principle, argues that the greater the handicap an individual carries, the greater its ability to offset that handicap with other superior qualities. The handicap principle The principle that the greater the handicap an individual carries, the greater its ability to offset that handicap. One factor that might attract females to certain males is a genetically based high resistance to parasites and pathogens. As you know, parasites evolve rapidly and thereby continually apply selection for genetic resistance in the host. Because parasites and pathogens can impair the construction of secondary sex characteristics, a male bird that has an elaborate and showy plumage might communicate to females that because he has the energy to build elaborate feathers, he can resist parasites and pathogens. If this resistance can be inherited by the offspring of the males, then the secondary sex characteristics are honest signals of the genetic superiority of the males and indicate that only individuals with superior genes could resist parasites and maintain a bright and showy plumage. Numerous studies have found that parasites and pathogens affect male attractiveness. In rock doves (Columba livia)—also known as pigeons— hatchling birds can be infected by mites (Dermanyssus gallinae) that live in the nest. To determine the effect of the mites on the young doves, researchers fumigated one set of infected nests to kill the mites but did not fumigate a second set of infected nests. The nestlings living in the nests with mites experienced lower survival and slower growth as a nestling (Figure 9.21). Other researchers who have examined the affects of lice on rock doves have found that females prefer louse-free males to louse-infested males by a ratio of three to one. In ring-necked pheasants (Phasianus colchicus), females prefer males with longer spurs on their lower legs. Longer spurs are linked genetically to major histocompatibility complex (MHC) genes that influence susceptibility to disease. Males with longer spurs have MHC alleles that are linked to longer life spans. Therefore, females that choose to mate with long-
spurred males should produce offspring with a higher chance of surviving to reproduce as adults.
Figure 9.21 The effect of mites on pigeons. The pigeons on the left were raised in nests that were fumigated to reduce the mite population. The pigeons on the right were raised at the same time, but in unfumigated nests that had high mite populations. The mite infections cause lower survival, slower growth, and areas of missing feathers.
Sexual Conflict
Mating decisions were once thought to serve the mutual interest of both participants. More recently, scientists have come to appreciate that mating partners often behave according to their own self-interest. In lions, for example, when a new dominant male takes over the pride, he often kills newborn cubs that have been fathered by the previous dominant male so that the female loses her entire reproductive effort. The male obtains a fitness benefit because females without newborn cubs come into breeding condition more rapidly, which allows the new dominant male to father offspring sooner. Of course, this represents a fitness cost to the mother. One of the more dramatic examples of sexual conflict occurs in bedbugs (Cimex lectularius). The male bedbug has a sharp sperm-transferring appendage and fertilizes a female by stabbing her with the appendage and injecting his sperm into her circulatory system (Figure 9.22). These sperm make their way to the female’s ovaries. As a result, the males obtain more female mates, but females fertilized by several males live shorter lives and
lay fewer eggs. It is hypothesized that this aggressive behavior evolved because female bedbugs resisted copulation attempts by male bedbugs. Examples such as these demonstrate that sexual interactions can reflect different decisions that represent the self-interest of males and females.
Figure 9.22 Sexual conflict in bedbugs. (a) The male bedbug possesses a sharp spermtransferring appendage that it uses to pierce the underside of females in a specific location. (b) Using this appendage, located on the far end of its abdomen, males can mount females and pierce the females repeatedly. This causes the females to have a shorter life and to lay fewer eggs. We have seen the ecological conditions that favor sexual versus asexual reproduction as well as the factors that favor balanced versus biased sex ratios. We have also seen how the different investments into eggs versus sperm, as well as the need to care for offspring, have shaped the evolution of different mating systems and the evolution of secondary sex characteristics. The evolution of sex remains an active area of research and there is still much to be learned.
Concept Check
2. What traits might females prefer in a male when selecting a mate? 3. Why might exaggerated secondary sexual characteristics in males demonstrate a superior genotype to females?
Male-Hating Microbes
Infected with Wolbachia. The Wolbachia bacteria can infect a diversity of insect species, such as this mosquito (Plasmodium berghei). (a) The mosquito is seen under normal light. (b) When the mosquito and bacteria are treated to glow in different colors, the bacterial cells show up as red spots inside the mosquito’s body. Throughout this chapter, we have considered different mating strategies as the result of the individual’s genes and its environment. However, there is a growing realization that bacteria and other microorganisms living inside an organism can control its reproductive strategies. One of the most widespread groups of bacteria in insects belongs to the genus Wolbachia. Though first discovered
nearly a century ago living in mosquitoes, it was not until 1990 that scientists realized the bacteria could fundamentally change an individual’s reproductive strategies. Wolbachia infects a wide variety of invertebrates, including spiders, crustaceans, nematodes, and insects. In fact, it is currently estimated that 70 percent of all insect species experience infections by this bacteria, which can live throughout the tissues of its host. In 2007, researchers discovered that the parasite’s entire genome has even been incorporated into the genome of the tropical fruit fly (Drosophila ananassae). An important aspect of Wolbachia’s life history is that it is only passed down to offspring via the mother’s infected egg. From the microbe’s perspective, therefore, the host’s female offspring are important for improving the microbe’s fitness, whereas the host’s male offspring are useless. To improve its fitness, Wolbachia has evolved ways to exploit or eliminate these useless male hosts. In mosquitoes, Wolbachia alters sperm to prevent them from fertilizing the eggs of uninfected females, which ensures that subsequent generations will be dominated by offspring of infected females that carry the bacteria. In wasps, Wolbachia causes parthenogenesis and prevents some species from producing males. Researchers discovered this by treating some of the infected wasps with an antibiotic to kill Wolbachia; once the bacteria were gone, the wasps no longer produced offspring by parthenogenesis. Scientists now wonder if parthenogenesis in other species that we have discussed might also be the result of a bacterium rather than an adaptive strategy. In woodlice (Armadillidium vulgare), Wolbachia converts
males into females by suppressing male hormones. This ability for hormones to determine sex in embryos is similar to the effects of hormone manipulations by scientists investigating temperature-dependent sex determination in Jacky dragons that we saw earlier. In other species of invertebrates, the bacterium simply kills young males and prevents them from entering the adult population. In species with adults of both sexes, such as the Ugandan butterfly (Acraea encedon), the increase in females relative to males has caused a reversal in the typical sex roles; females now court males and compete with other females for breeding opportunities. Note that all these sexdetermining mechanisms, changes in offspring sex ratio, or mating systems are in the best interest of the bacteria, not the host. Wolbachia are not the only bacteria with such effects. Bacteria in the genus Rickettsia are well-known for causing diseases such as typhus and Rocky Mountain spotted fever, but other species in this genus commonly occur in invertebrates without causing disease. In 2011, researchers found a species of Rickettsia living in the sweet potato whitefly (Bemisia tabaci). In the southwestern United States, infection rates increased from 1 percent in 2000 to nearly 100 percent by 2009. This incredibly rapid spread of the bacterium occurred because infected female flies nearly doubled their reproductive output, the survival rate of their offspring increased, and the offspring sex ratio was altered from 50 percent daughters to nearly 85 percent daughters. This is interesting from the standpoint of host– parasite interactions and the evolution of reproductive strategies. It is also important for agriculture. The sweet potato white fly is a pest that sucks the sap out of the
leaves of many crops, including cotton, vegetables, and ornamental plants. Scientists worry that the massive increase in infection rates by Rickettsia will lead to a massive population increase of this pest. Infections of bacteria that are passed from mother to daughter are sometimes beneficial to humans. In 2016, researchers reported that they infected mosquitoes (Aedes aegypti) with Wolbachia pipientis, a species of Wolbachia that does not naturally infect this mosquito. After causing the infections, the researchers then tested whether the Wolbachia would affect the mosquito’s ability to transmit harmful human pathogens, including the yellow fever virus and the Zika virus. Amazingly, the mosquitoes infected by Wolbachia were less likely to be infected by and transmit the viruses. Based on this discovery, plans are being made to release large numbers of Wolbachia-infected mosquitoes in areas plagued by yellow fever and Zika. Collectively, these studies suggest that there is much more to the evolution of sex than we currently know, and that understanding the evolution of sex in these organisms can have important implications for humans, including the number of pests attacking our food supply. SOURCES: Aliota, M. T. Aliota, S. A. Peinado, I. D. Velez, and J. E. Osorio. 2016. The wMel strain of Wolbachia reduces transmission of Zika virus by Aedes aegypti. Scientific Reports 6: 28792. Himler, A. G., et al. 2011. Rapid spread of a bacterial symbiont in an invasive whitefly is driven by fitness benefits and female bias. Science 332: 254–256. Knight, J. 2011. Meet the Herod bug. Nature 412: 12–14.
Summary of Chapter Concepts
9.1 Reproduction can be sexual or asexual. Asexual reproduction occurs through vegetative reproduction or through parthenogenesis. Compared to asexual reproduction, sexual reproduction results in fewer copies of a parent’s genes in the next generation. This can be offset by adopting a hermaphrodite sexual strategy or by providing parental care that results in raising twice as many offspring. The benefits of sexual reproduction include purging harmful mutations and creating genetic variation to help offspring deal with future environmental variation. Key Terms: Sexual reproduction, Asexual reproduction, Vegetative reproduction, Clones, Binary fission, Parthenogenesis, Cost of meiosis, Red Queen hypothesis
9.2 Organisms can evolve as separate sexes or as hermaphrodites. If an individual possessing only male or female function can add a large amount of the other sexual function while giving up only a small amount of its current sexual function, selection will favor the evolution of hermaphrodites. If not, selection will favor the evolution of separate sexes. To avoid inbreeding depression, hermaphrodites have evolved adaptations to prevent selfing and mixed strategies for when selfing is the best option. Key Terms: Perfect flowers, Simultaneous hermaphrodites, Sequential hermaphrodites, Monoecious, Dioecious
9.3 Sex ratios of offspring are typically balanced, but they can be modified by natural selection. Depending on the species, sex may be determined largely by genetics or by the environment. In many species, females have the ability to manipulate the sex ratio by controlling which sperm are used to fertilize the eggs or which sex chromosomes end up in the eggs, or by selective abortion of the fertilized embryos. In most organisms, the sex ratio is approximately
one to one due to frequency-dependent selection. When offspring are isolated from the rest of the population and are subjected to local mate competition, highly skewed offspring sex ratios can be adaptive. Key Terms: Environmental sex determination, Frequencydependent selection, Local mate competition
9.4 Mating systems describe the pattern of mating between males and females. While many species are socially monogamous, recent studies have demonstrated that many individuals participate in extra-pair copulations. As a result of this infidelity, species have evolved a variety of mate guarding behaviors to prevent a reduction in their fitness. Key Terms: Mating system, Promiscuity, Polygamy, Polygyny, Polyandry, Monogamy, Extra-pair copulations, Mate guarding
9.5 Sexual selection favors traits that facilitate reproduction. The difference in the energetic costs of gametes and the costs of parental care typically causes female fitness to be a function of mate quality and male fitness to be a function of mate quantity. As a result, females are typically selective in choosing mates, whereas males compete strongly with each other to mate as often as possible. Male competition for female mates has favored the evolution of sexually dimorphic traits, including body size, ornaments, coloration, and courtship behaviors. Females choose particular males to obtain material benefits, such as nesting sites or food, or for nonmaterial benefits, such as good genes or good health. The best reproductive choices of males and females are often not reciprocal, which can cause conflicts between the sexes. Key Terms: Sexual selection, Sexual dimorphism, Primary sexual characteristics, Secondary sexual characteristics, Good genes hypothesis, Good health hypothesis, Runaway sexual selection, The handicap principle
Critical Thinking Questions
1. Compare and contrast the costs and benefits associated with sexual versus asexual reproduction. 2. How does the Red Queen hypothesis help us understand the fitness benefits of sexual reproduction? 3. Freshwater snails exhibit both sexual and asexual reproduction. Why might natural selection favor such a mixed breeding strategy? 4. Given that self-fertilization leads to inbreeding depression, under what conditions should a hermaphrodite use self-fertilization? 5. What strategies do monoecious plants use to avoid possible negative effects of inbreeding? 6. In addition to affecting sex ratios, what other traits might be affected by the artificial selection caused by intense fishing? 7. Turtle embryos incubated at lower temperatures typically produce males, whereas those incubated at higher temperatures produce females. In the face of global warming, how might turtles evolve to change their egg-laying behavior to maintain an even sex ratio? 8. Compare and contrast monogamy, polygyny, polyandry, and promiscuity. 9. Explain how extra-pair copulation has favored the evolution of mate guarding. 10. Why might exaggerated secondary sexual characteristics in males demonstrate a superior genotype to females?
#### Graphing the Data: Frequency-Dependent Selection
GRAPHING THE DATA Frequency-Dependent Selection In most species containing individuals that are either male or female, the proportion of males and females changes over time. Using a line graph, graph the proportion of male and female zebras in a population over a 10-year period. Based on these data, what happens whenever one sex becomes rare or common? Year Males Females 45% 55% 48% 52% 52% 48% 55% 45% 53% 47% 47% 53% 44% 56% 49% 51% 55% 45% 45% 55%