14 Predation and Herbivory
#### A Century-Long Mystery of the Lynx and the Hare

14Predation and Herbivory Canadian lynx and snowshoe hare. For nearly 100 years, ecologists have been examining the regular fluctuations in the populations of these species to determine the causes. A Century-Long Mystery of the Lynx and the Hare For centuries, naturalists, hunters, and trappers have noticed that populations of many species experience large fluctuations, and that some species fluctuate at regular intervals. In 1924, the ecologist Charles Elton drew attention to regular population fluctuations in many species of high-latitude animals in Canada, Scandinavia, and Siberia. In particular, he focused on snowshoe hares (Lepus americanus) and Canada lynx (Lynx canadensis). Elton examined data that had been compiled from the Hudson’s Bay Company, a Canadian firm that had been purchasing pelts from trappers for more than 70 years. He assumed that the number of purchased pelts over time reflected the abundance of the two species. Elton and his fellow ecologists were
fascinated by regular cycles of high and low density among lynx and hare populations that occurred approximately every 10 years. The 10year cycles in the abundance of lynx and hares were clear, but the mechanisms causing these cycles have been debated for nearly a century. There have been numerous hypotheses for these 10-year cycles. When Elton wrote his classic paper, some biologists hypothesized that animals possessed a “physiological rhythm” that caused both the lynx and the hare to reproduce abundantly in some years and sparingly in others. Elton rejected this hypothesis because it was very unlikely that such a rhythm would be synchronized for all individuals of different ages and for all individuals across large regions. Instead, he favored an explanation related to the 9- to 13-year cycle of sunspots, which are periods of increased solar activity. If the cycle of sunspots could substantially affect the climate and, therefore, the growing conditions of the plants that hares consumed, it could explain the cycles of the hares. The lynx cycle, which occurred about 2 years later than the hare cycle, was thought to reflect the fact that lynx primarily consume hares. When hares are abundant, lynx have more food and therefore reproduce more in subsequent years, but when hares are rare, the lynx reproduce poorly and many of them starve, which causes the lynx population to decline. Since Elton’s original work, ecologists have determined that although the sunspot cycle is similar in length to the hare cycle, it has never closely matched the timing of the hare cycle. Nor have they been able to find a climate-driven mechanism that connects the sunspot cycle to the hare cycle. With these hypotheses eliminated, researchers turned their attention to competition and predation. For many decades, a lively debate focused on the possibility that the hare cycles were caused by the hares exceeding their carrying capacity, which could explain the observation that hare reproduction declines as the hare population grows. Another hypothesis suggested that lynx predation caused the cycles. When lynx were rare, the hares survived better. As lynx became more numerous, they began consuming hares faster than the hares could reproduce, thereby causing the hare population to decline.
“The 10-year cycles in the abundance of lynx and hares have been clear, but the mechanisms causing these cycles have been debated for nearly a century.” It seemed impossible to determine the answer without conducting some experiments. From 1976 to 1985, a large experiment manipulated the presence or absence of supplemental food for the hares. Although the supplemental food increased the carrying capacity for hare populations, they still cycled in synchrony with populations that were not fed. This suggested that the hare population does not decline due to a lack of food. However, if the hares do not experience a lack of food, why does their rate of reproduction decline at higher densities? In a subsequent experiment, researchers built large fences so they could manipulate the presence or absence of supplemental food and the presence or absence of lynx predation. Both excluding lynx and adding food increased the peak population size of the hares, but the hare populations still cycled. However, the fence that excluded the lynx did not exclude other predators, including owls and hawks, which could fly into the fenced areas and continue to kill hares. Among the hares that died, more than 90 percent died from predation, whereas few died from starvation, which further confirmed that predation rather than food availability contributed to the cyclical decline of hares. A new insight came in 2009 when researchers discovered that the declining rate of hare reproduction under high hare densities is caused by high densities of predators, which induce high levels of stress in the hares. Eventually, the stress of the predation threat becomes so high that the hares experience reduced reproduction. Moreover, the faster that the hare population declines due to predators, the greater stress that the surviving hares experience. Once the hare population declines, there are many fewer predators so the hare stress level is
much reduced and hare reproduction returns to a high level. However, if the hares experienced especially high predation rates, the associated higher stress can affect them for multiple generations and cause the hare population to remain low in abundance for a longer amount of time. In short, while the abundance of food can affect the number of hares in the population, it appears that the lynx–hare population cycles can be attributed to a combination of direct predation and the indirect effects of predator stress that cause reduced hare reproduction. The century-long investigation of the lynx–hare cycles illustrates that consumers and the resources that they consume can interact in complex and interesting ways. In this chapter, we will examine how predators and herbivores can affect the populations of the species they consume, including how the abundance of predators and prey populations can cycle over time, how consumers catch their prey, and how prey defend themselves. SOURCES: Elton, C. S. 1924. Periodic fluctuations in the numbers of animals: Their causes and effects. British Journal of Experimental Biology 2: 119–163. Sheriff, M. J. 2009. The sensitive hare: Sublethal effects of predator stress on reproduction in snowshoe hares. Journal of Animal Ecology 78: 1249–1258. Krebs, C. J. 2011. Of lemmings and showshoe hares: The ecology of northern Canada. Proceedings of the Royal Society B. 278: 481–489. Sheriff, M. J., et al. 2015. Predator-induced maternal stress and population demography in showshoe hares: The more severe the risk, the longer the generational effect. Journal of Zoology 296: 305–310.
Learning Objectives
After reading this chapter, you should be able to:
14.1 Demonstrate how predators and herbivores can limit the abundance of populations.
14.2 Illustrate how populations of consumers and consumed populations fluctuate in regular cycles.
14.3 Explain how predation and herbivory favor the evolution of defenses. Most species consume resources and are also a resource for other species to consume. For example, plants and algae consume nutrients, water, and light; these resources allow plants and algae to photosynthesize and grow. While plants and algae are alive, they are consumed by herbivores, parasites, and pathogens. After they die, these producers are consumed by detritivores and decomposers. Similarly, animals consume a wide variety of other organisms at the same time that they are subject to consumption by carnivores, parasites, and pathogens. After they die, animals are consumed by scavengers, detritivores, and decomposers. As you can see, a tremendous number of interactions occur among species in nature. These interactions, critical to the composition of species in different communities, are the subjects of the next four chapters. In this chapter, we will focus on interactions between predators and their prey and between herbivores and producers. We will examine the conditions under which predators and herbivores can limit the population sizes of the species they consume. We will also look at models of predators and herbivores to help us understand how populations of these consumers fluctuate in relation to the populations of the species they consume. We will conclude the chapter by exploring how predators and herbivores have favored the evolution of defenses in prey and plants.
#### 14.1 Predators and Herbivores Can Limit the Abundance of Populations
14.1 Predators and herbivores can limit the abundance of populations Are populations limited primarily by what they eat or by what eats them? Studies of predation and herbivory attempt to answer this question by looking at whether or not predators and herbivores reduce the size of prey and producer populations below the carrying capacities set by resources. Understanding these relationships is of great practical concern to those interested in the management of crop pests, game populations, and endangered species. It also has far-reaching implications about the interactions among species that share resources—knowledge that helps us to understand the structure of ecological communities.
Predators
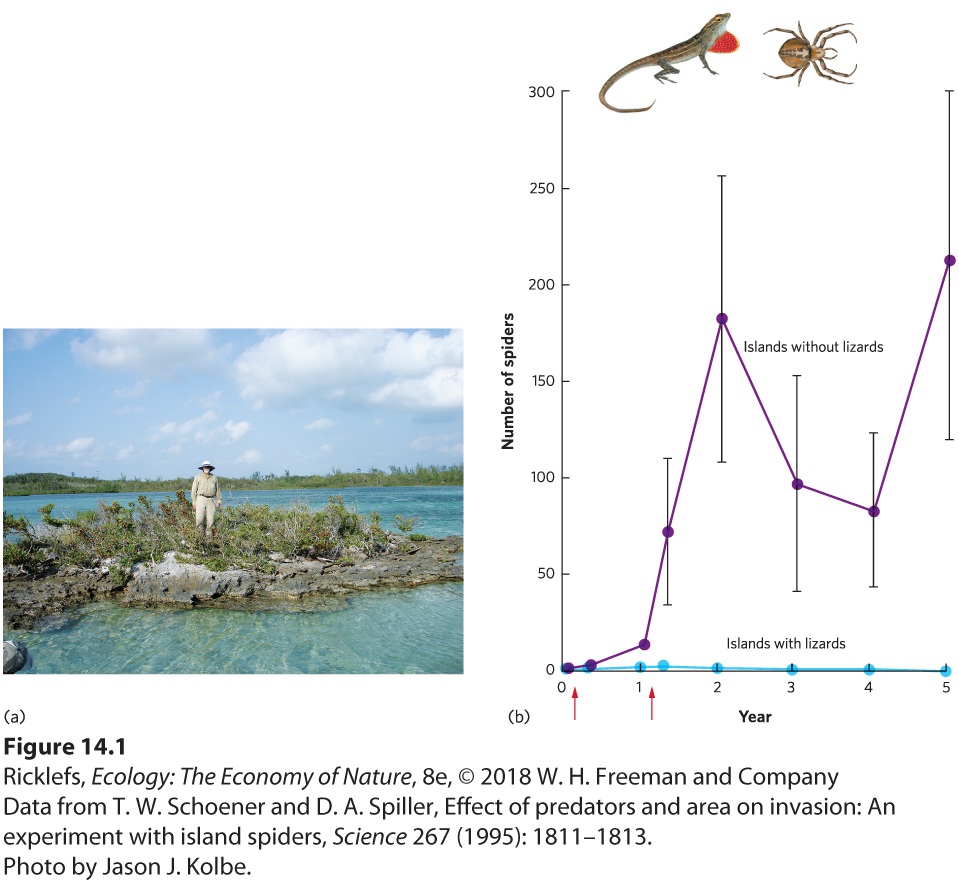
As we noted with lynx and hares at the beginning of this chapter, ecologists have long been fascinated by predator–prey interactions, in both nature and in manipulative experiments. For example, a survey of 93 Caribbean islands revealed that the smaller islands contained spiders, whereas the larger islands contained spiders and lizards, which are predators of the spiders. When the researchers compared the density of spiders on islands with and without the predatory lizards, they found that spiders were about 10 times more abundant on islands without the lizards. While these observations suggest that predator lizards play an important role in controlling spider populations, an experimental test would provide more definitive evidence that the predators were truly the cause. To that end, researchers conducted a manipulative experiment on islands in the Bahamas that ranged in size from about 200 to 4,000 m2. They selected five islands with brown anole lizards (Anolis sagrei) and five islands without lizards. On these 10 islands, they introduced a species of orb-weaving spider (Metepeira datona) as a prey that was not originally present on the islands. They introduced the spiders on two occasions, for a total of 20 spiders per island. Then they surveyed the spider populations 4 days after each introduction and detected lower spider densities on the islands containing lizards. This suggested that predation from lizards had already started to affect the spider populations. Over the next 5 years, the spider populations that lived on
islands without lizards became 10 times more abundant, as you can see in
Figure 14.1. In contrast, the spider populations living with lizards became either rare or extinct, which confirmed that predatory lizards reduce the densities of spiders.
Figure 14.1 Lizard predation on spiders. (a) In the Bahamas, researchers introduced a total of 20 spiders to each of five small islands with predatory lizards and to five small islands without lizards. (b) Spiders were introduced to each island on two dates, indicated by the red arrows. The spiders remained rare or absent on islands with lizards but increased 10-fold on islands without lizards. Error bars are standard errors. Sometimes the introduction of a predator is the result of an accident. When a species is introduced to a region of the world where it has not historically existed, we say that it is an introduced species, which is also known as an exotic species or non-native species. If the introduced species spreads rapidly and has negative effects on other species, human recreation, or human economies, we say that it is an invasive species. On the island of Guam, for example, many species of birds, bats, and lizards lived without predatory snakes for thousands of years.
Sometime after World War II, however, the brown tree snake (Boiga irregularis) was accidentally introduced to Guam by ships carrying supplies (Figure 14.2). While the snake was native to the South Pacific, the animals on Guam had no evolutionary history of living with snakes and many had no defenses against them. Over the next 20 years, as the population of the brown tree snake grew exponentially, it had a significant and devastating effect on the island fauna. Guam has experienced sharp declines or extinctions in nine species of forest-dwelling birds, all three species of bats, and several species of lizards.
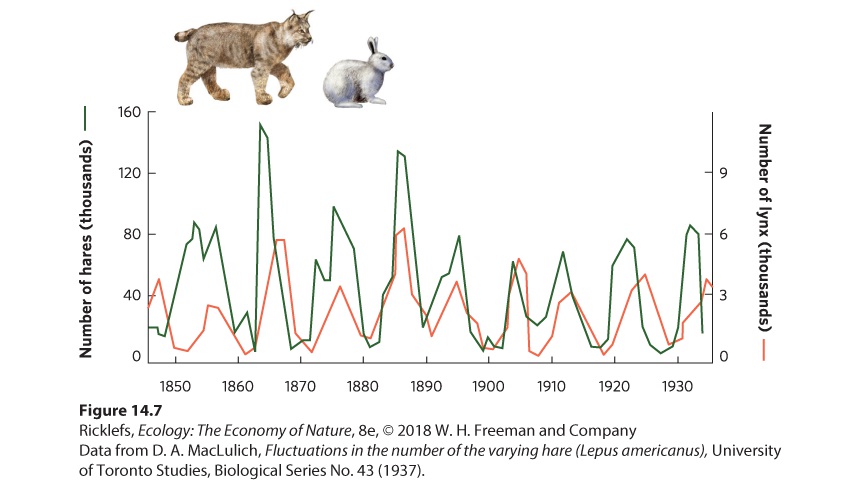
Figure 14.2 The introduced brown tree snake. (a) The introduction of this snake to the island of Guam led to the decline and local extinction of birds, bats, and lizards. (b) An example of the streamers that were attached to dead mice containing acetaminophen to poison the invasive snakes. Introduced species A species that is introduced to a region of the world where it has not historically existed. Also known as Exotic species or Non-native species. Invasive species An introduced species that spreads rapidly and has negative effects on other species, human recreation, or human economies. A variety of efforts have been made to reduce the snake population, including the use of snake traps and snake-detecting dogs. In a recent and very creative experiment, thousands of dead mice were injected with acetaminophen (the active ingredient in Tylenol, which is toxic to the snakes, but not other animals) and attached to streamers, which act as parachutes. These parachuted mice were then dropped from helicopters around the island. The hope is that the snakes will find the mice dangling from trees, consume
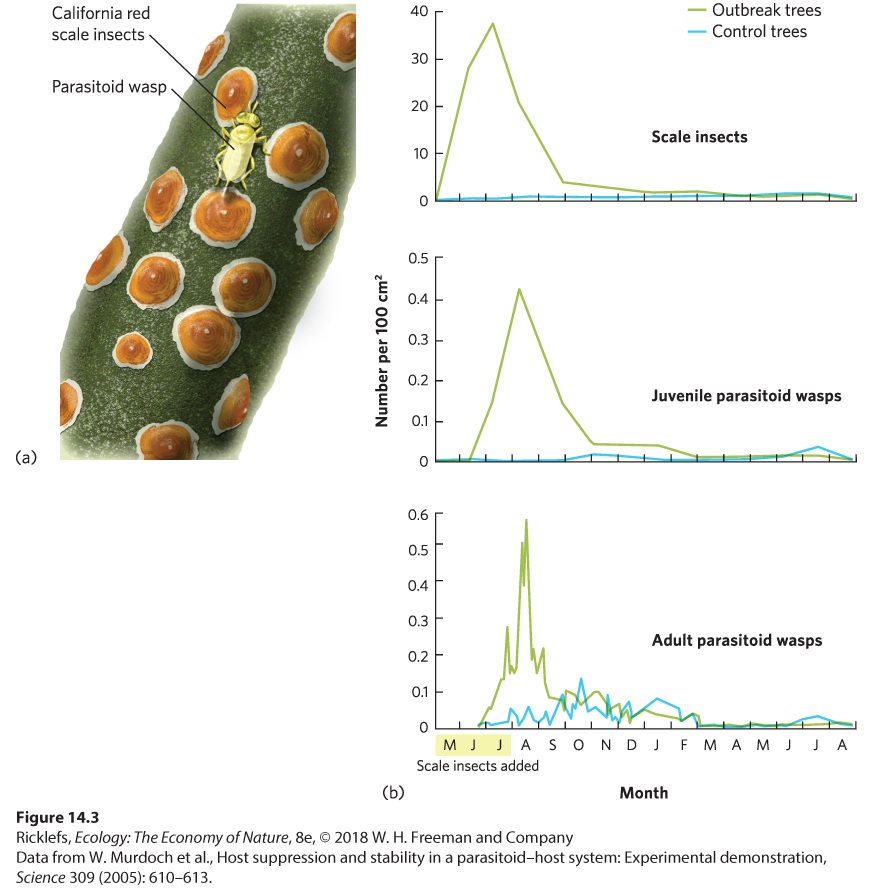
the mice, and then die from the drug. As we mentioned in Chapter 1, parasitoids are a unique type of predator that lives within and consumes the tissues of a living host, eventually killing it. Like many other predators, parasitoids also can limit the abundance of their prey. This can be seen in an example of wasps and scale insects. The California red scale insect (Aonidiella aurantii) is a worldwide pest in citrus orchards. It feeds on the leaves and fruits of the trees, which causes a great deal of damage and makes the fruit unmarketable. Fortunately, a small species of parasitoid wasp (Aphytis melinus) is able to control the abundance of the red scale insect by laying eggs inside the scale insects and ultimately killing them. To demonstrate the magnitude of this control, researchers in California mimicked an insect outbreak by adding large numbers of scale insects to four trees. Ten other trees with typical low numbers of scale insects were the controls. You can see the results of this experiment in Figure 14.3. Shortly after adding large numbers of scale insects, there was a significant increase in the number of juvenile and adult wasps. As the wasps became more abundant, the population of scale insects declined rapidly. Within a few months, the large population of scale insects was reduced to the same level found on trees that had never received an experimental addition of scale insects. Although the wasps were not able to completely eliminate the scale insects, they held the population of scale insects to a level that minimally harmed the citrus crops.
Figure 14.3 Effects of a parasitoid on populations of red scale insects. (a) The California red scale, a tiny insect with a hard shell, feeds on the sap of citrus trees and other plants. The parasitoid wasp lays its eggs under the hard shell of the scale insect. When the eggs hatch, the larvae slowly consume the scale insect. (b) After introducing large numbers of scale insects to trees—during the months indicated by yellow shading on the x axis—the scale insect population grew rapidly. Soon after the addition of the scale insects, the number of juvenile and adult parasitoid wasps began to rapidly increase. This increase in parasitoid wasps caused a subsequent rapid decline in scale insects.
Mesopredators
Two levels of predators often exist in ecological communities: mesopredators and top predators. Mesopredators such as coyotes, weasels, and feral cats are relatively small carnivores that consume herbivores. In contrast, top predators such as wolves, mountain lions, and sharks typically consume both herbivores and mesopredators. Throughout history, top predators have interfered with human activities such as ranching, farming, hunting, and fishing. To protect our livelihood, we have reduced or eliminated the world’s top predators, but this has caused
unintended consequences. It is estimated that the decline of the top predators in North America had allowed 60 percent of all mesopredators to expand their geographic ranges. Mesopredators Relatively small carnivores that consume herbivores. Top predators Predators that typically consume both herbivores and mesopredators. The expansion in the range and abundance of mesopredators has had dramatic effects on the prey they consume. For example, the reduction of shark populations in the Atlantic Ocean from overharvesting has led to an increase in the cownose ray (Rhinoptera bonasus), a major mesopredator that consumes bay scallops (Argopecten irridians). This has caused a large reduction in bay scallops. In some cases, researchers have found that the benefits to humans of removing a top predator are much smaller than the damage inflicted by a mesopredator that became more abundant after the top predator declined. In Australia, for example, there has long been a campaign to remove dingoes and feral dogs because they kill sheep. However, removing these top predators has caused an increase in red foxes (Vulpes vulpes), which also eat sheep. This increase in red foxes has led to more than triple the loss in sheep that occurred when dingoes and feral dogs helped to control the red fox population.
Herbivores
Like predators, herbivores can have substantial effects on the species they consume. One of the classic examples is the control of the prickly pear cactus in Australia. The prickly pear cactus is a group made up of several different species that are native to North and South America. In the nineteenth century, many of these species were brought to Australia for a variety of reasons, including use as ornamental plants and as “living fences” for pastures. The cacti rapidly spread across the continent to the point that they dominated thousands of hectares of pasture and rangeland. To combat the spread of the cactus, biologists in the 1920s collected cactus moths (Cactoblastis cactorum) in South America and introduced the moths to Australia. These moths are natural herbivores of the
cactus; the caterpillar stage of the moth consumes a portion of the cactus and the injuries it causes allow pathogens to infect the plants. As you can see in the before and after photographs of Figure 14.4, the moths quickly reduced the cactus to very low abundances. While the moth has not completely eradicated the cactus because the cactus is able to disperse to moth-free areas, today, the prickly pear cactus exists only in small pockets around Australia.
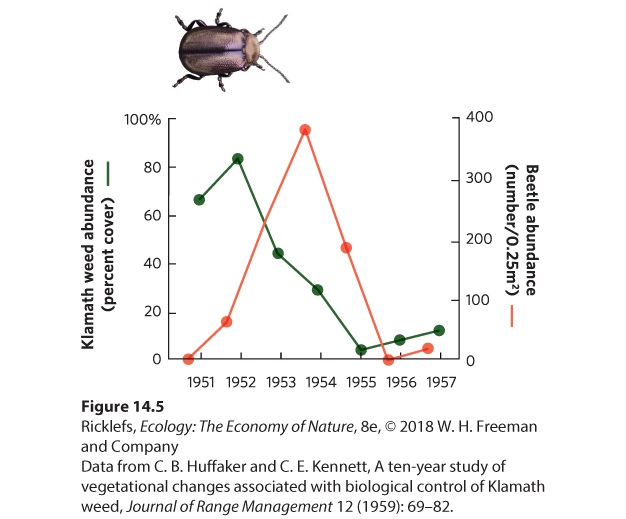
Figure 14.4 The prickly pear cactus in Australia. Following the import of the cactus from South America to Australia, its numbers increased dramatically. (a) To reduce the abundance of the cactus, the cactus moth was introduced to Australia from South America. (b) A site in Queensland, Australia, in 1926 prior to the introduction of the moth. (c) The same site 3 years after the introduction of the moth. Herbivorous insects have been used in similar ways in North America. In California, a plant known as Klamath weed (Hypericum perforatum), which is toxic to livestock, was accidentally introduced from Europe in the early 1900s. By 1944, the weed had spread over nearly a million hectares of rangeland in 30 counties. In the 1950s, biologists decided to introduce a leaf-feeding beetle (Chrysolina quadrigemina) that consumed Klamath weed in Europe. After introduction of the beetle, the weed rapidly declined in abundance, as you can see in Figure 14.5. Biologists estimate that the beetle has now eliminated 99 percent of the Klamath weed population in North America.
Figure 14.5 Beetle herbivory on Klamath weed. Following the introduction of leaf-feeding beetles, the beetle population initially increased and the plant began a rapid decline. Once the plant population declined, the beetle population also declined. The effects of herbivores are easily observed. For example, when domesticated animals such as cattle or sheep are raised at very high densities, little plant life remains. Similarly, deer and geese can consume large amounts of plants. Their impact can be demonstrated by fencing off areas to exclude animals to prevent grazing; the fenced areas have a greater total mass, or biomass, of plants and a higher composition of plants that are preferred by the herbivores (Figure 14.6). The plant species that remain in the areas where grazing occurs are those that the herbivores prefer not to eat.
Figure 14.6 Fencing out deer. In this deer enclosure in Gwaii Haanas National Park Reserve and Heritage Site, Haida Gwaii, BC, Canada, the long-term fencing of an area to prevent deer herbivory has allowed a much greater abundance of plants to grow. Herbivore effects also occur in aquatic habitats. In rocky shore communities, for example, sea urchins control populations of algae. When sea urchins are removed from an area, the biomass of algae increases rapidly, while the species composition of the algae also changes. In the presence of sea urchins, the remaining algae consist mostly of those species the sea urchins do not like and those that can withstand intense grazing. When sea urchins are removed, however, more palatable species such as the large brown algae become more abundant and can shade out the other species of algae. Such studies demonstrate that the influence of herbivores on the abundance of producers affects the species composition of the entire community.
Concept Check
1. What evidence is there that predators can control the abundance of prey? 2. How has the reduction of top predators had unintended
consequences on the abundance of prey? 3. What evidence is there that herbivores can control the abundance of plants?
#### 14.2 Populations of Predators and Prey Fluctuate in Regular Cycles
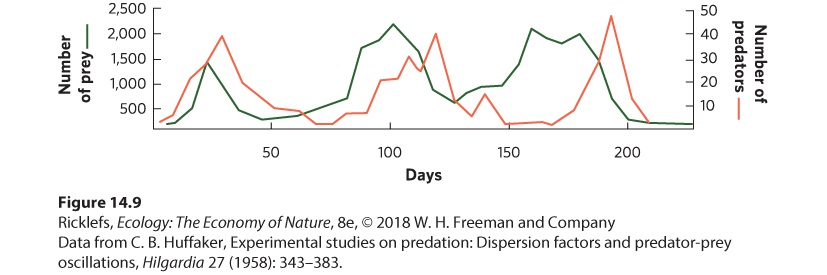
14.2 Populations of predators and prey fluctuate in regular cycles In the previous chapter, we discussed how populations can fluctuate over time and space, and we saw that some populations cycle. At the beginning of this chapter, we introduced a study of population cycles for lynx and snowshoe hares. You can see the data from this study in Figure 14.7. The hare and lynx populations both experienced cycles of 9 to 10 years, with the lynx cycles lagging about 2 years behind the hare cycles.
Figure 14.7 Cyclic fluctuations in the populations of snowshoe hares and lynx. Records from the Hudson’s Bay Company of pelts purchased from trappers demonstrate that the populations of hares and lynx went through cycles approximately every 10 years. It turns out that other large herbivores in the boreal and tundra regions of Canada—such as muskrat, ruffed grouse, and ptarmigan—also have population cycles of 9 to 10 years. Smaller herbivores, such as voles, mice, and lemmings, tend to have 4-year cycles. Studies of predators in these regions have revealed that some predators—including red foxes, marten, mink, goshawks, and horned owls—feed on larger herbivores and have long population cycles. In contrast, other predators—including Arctic foxes (Vulpes lagopus), rough-legged hawks (Buteo lagopus), and snowy owls (Bubo scandiacus)—feed on small herbivores and have short population cycles. The close synchrony of population cycles between predators and the prey they consume suggests that these oscillations are the result of
##### Creating Predator–Prey Cycles in the Laboratory
interactions between them. To understand the mechanisms underlying the predator–prey cycles, it is useful to examine them in the context of population models. CREATING PREDATOR–PREY CYCLES IN THE LABORATORY During the early twentieth century, biologists became interested in using predators and pathogens to control populations of crop and forest pests. One of the leading researchers in this effort was Carl Huffaker, a biologist at the University of California at Berkeley who pioneered the biological control of crop pests. Huffaker sought to understand the conditions that cause predator and prey populations to fluctuate. He chose two species of mites that lived on citrus trees: the six-spotted mite (Eotetranychus sexmaculatus) was the prey and the western predatory mite (Typhlodromus occidentalis) was the predator. In a series of experiments, he established populations on large trays that contained oranges—which were both habitat and food for the prey—with rubber balls interspersed among the oranges, as illustrated in Figure 14.8. On each tray, he varied the number and distribution of the oranges.
Figure 14.8 Huffaker’s predator–prey lab experiment. To determine the factors that cause predator–prey cycles, Huffaker manipulated the number and distribution of oranges in a tray filled with oranges and rubber balls. He also added posts that only the prey species were able to climb. The prey drifted from the posts to colonize new oranges. Because the predators had to walk from one orange to another and avoid Vaseline barriers, the prey stayed one step ahead of the predators. In most experiments, Huffaker introduced 20 female prey per tray and then
introduced two female predators 11 days later. Because both species reproduce parthenogenetically, no males were added. When the prey were introduced to trays without predators, the prey population leveled off at between 5,500 and 8,000 mites. When predators were added, the predator population increased rapidly and soon wiped out the prey population. Without prey to feed on, though, predators quickly became extinct. However, it took longer for predators and prey to go extinct if the oranges were spread far apart from each other because it took longer for the predators to locate the prey. Under these experimental conditions, predators and prey could not coexist over time. Huffaker reasoned that if predator dispersal could be further impeded, the two species might be able to coexist. To accomplish this, he introduced barriers to predator dispersal. The predatory mites disperse by walking, but the prey mites use a silk line that they spin to float on wind currents. Based on these differences, Huffaker modified his trays to give the prey a dispersal advantage by placing a mazelike pattern of Vaseline barriers among the oranges to slow the dispersal of the walking predators. He also placed vertical wooden pegs throughout the trays for the six-spotted mites to use as jumpingoff points. This arrangement produced a series of three population cycles during the 8-month experiment, as depicted in Figure 14.9. The distribution of predators and prey throughout the trays continually shifted as the prey, which were on their way to extermination on one orange, recolonized another orange and stayed one step ahead of their predators. In short, Huffaker created a metapopulation in the laboratory.
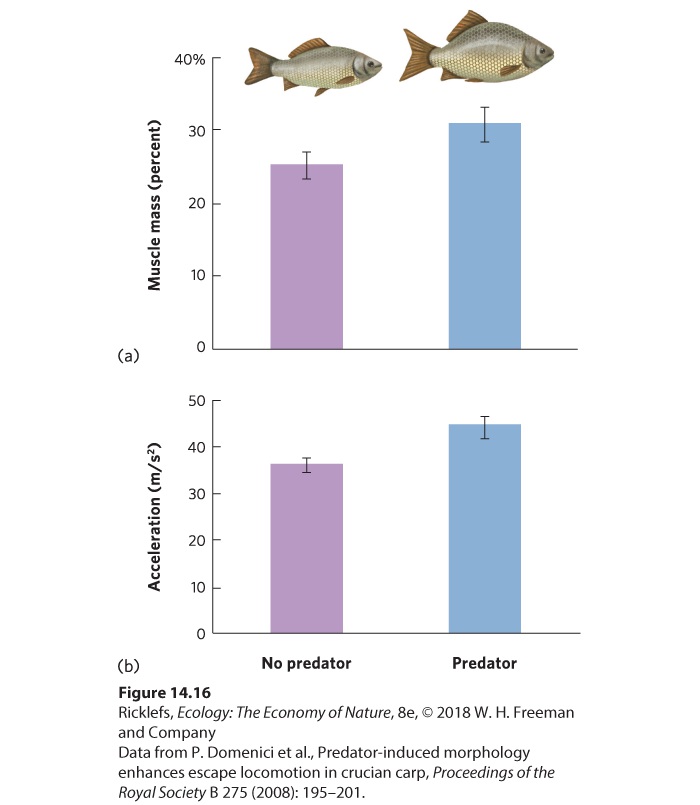
Figure 14.9 Predator–prey cycles in laboratory mites. When the prey species was given a dispersal advantage, it could colonize predator-free oranges and avoid going extinct. The cycling of the predator population lagged behind the prey population because it required more time to disperse to oranges containing prey and it required time to reproduce after finding prey.
Huffaker’s experiment demonstrates that predators and prey cannot coexist in the absence of suitable refuges for the prey. However, the predator and prey populations can coexist through time in a spatial mosaic of suitable habitats that provided a dispersal advantage to the prey. Two kinds of time delays caused the populations to cycle: one was the result of predators dispersing more slowly between food patches than their prey, and the other was the result of time needed for predator numbers to increase through reproduction. From this, we can conclude that stable population cycles can be achieved when the environment is complex enough that predators cannot easily find scarce prey. MATHEMATICAL MODELS OF PREDATOR–PREY CYCLES Even before Huffaker’s laboratory experiments on predator–prey cycles, mathematicians Alfred Lotka and Vito Volterra were developing models of predator–prey interactions. The Lotka–Volterra model incorporates oscillations in the abundances of predator and prey populations and shows predator numbers lagging behind those of their prey. It does this by calculating both the rate of change in the prey population and the rate of change in the predator population. Lotka–Volterra model A model of predator–prey interactions that incorporates oscillations in the abundances of predator and prey populations and shows predator numbers lagging behind those of their prey. Let’s begin with the prey population. Following the convention of our population models in Chapter 12, we will designate the number of prey as N and the number of predators as P. In words, the growth rate of the prey population depends on the rate of individuals being added to the prey population minus the rate of individuals being killed by predators: dNdt=rN-cNP The first term in this equation (rN) represents the exponential growth of a prey population based on the intrinsic growth rate (r), as we saw in Chapter 12. For simplicity, this term does not include density dependence. The second term (cNP) represents the loss of individuals due to predation. The model
assumes that the predation rate is determined by the probability of a random encounter between predators and prey (NP) and the probability of such an encounter leading to the prey’s capture (c) (think of c as “capture efficiency”). Next we turn to the predator population. The equation for the predator population is similar to that for the prey population in that it has two terms, one term that represents the birth rate of the predator population and a second term that represents the death rate of the predator population: dPdt=acNP-mP The first term in the equation (acNP) represents the birth rate of the predator population. It is determined by the number of prey consumed by the predator population (cNP), which we saw in the prey equation above, multiplied by the efficiency of converting consumed prey into predator offspring (a). The second term (mP) represents the death rate of the predator population and is determined by the per capita mortality rate of predators (m) multiplied by the number of predators (P). Changes in the Prey Population We can use the Lotka-Volterra equations to determine the conditions that allow the prey population to be stable. By definition, a population is stable when its rate of change is zero. We can write this as 0=rN−cNP We can rearrange this as rN=cNP This indicates that the prey population becomes stable when the addition of prey (rN) is equal to the consumption of prey (cNP). We can further simplify this equation as P=r÷c In other words, the prey population will be stable when the number of predators equals the ratio of the prey’s growth rate and the predator’s capture efficiency.
Now that we know the conditions that make the prey population stable, we can also explore the conditions that cause the prey population to increase or decrease. The prey population will increase when the addition of prey (rN) exceeds the consumption of prey (cNP), which we can write as rN>cNP We can rearrange this inequality as P<r÷c This inequality represents the number of predators that the population of prey can support and still increase. This number is higher when the growth potential of the prey population (r) is higher or when the predators are less efficient at capturing prey (c). Using the same logic, the prey population will decrease whenever P>r+c Changes in the Predator Population We can also use these equations to understand the conditions that make the predator population remain stable, increase, or decrease. Once again, the population is stable when the rate of change is zero: 0=acNP−mP We can rearrange this as acNP−mP This indicates that the predator population becomes stable when the production of new predators (acNP) is equal to the mortality of existing predators (mP). We can simplify this equation as N=m÷ac Given that this is the condition needed for a stable predator population, we can predict that the predator population will increase when the predator’s production of new predators exceeds the mortality of existing predators, which we can write as acNP>mP

We can rearrange this as N>m÷ac This inequality represents the number of prey required to support the growth of the predator population. This number is higher when the death rate of predators (m) is higher, and it is lower when predators are more efficient at capturing prey (c) and converting them into offspring (a). Using the same logic, the predator population will decrease whenever N<m÷ac Trajectories of Predator and Prey Populations Knowing the conditions under which predator and prey populations increase, decrease, or remain stable helps us understand why predator and prey populations sometimes cycle. Figure 14.10a plots the abundance of both populations. We can draw a horizontal line at the point where P = r ÷ c, which is the number of predators associated with a stable prey population. This line is called the equilibrium isocline, or zero growth isocline, for the prey because it indicates the points at which a population is stable. At any combination of predator and prey numbers in the region below the equilibrium isocline, there are relatively few predators and the prey population increases. In the region above this line, the prey population decreases because predators remove them faster than they can reproduce.
Figure 14.10 Equilibrium isoclines for prey and predator populations. (a) The prey population is stable when the number of predators is equal to r ÷ c. A higher number of predators causes the prey population to decrease, whereas a lower number of predators causes the prey population to increase. (b) The predator population is stable when the number of prey is equal to m ÷ ac. A higher number of prey causes the predator population to
increase, whereas a lower number of prey causes the predator population to decrease. In both figures, population decreases occur in the yellow-shaded areas, whereas population increases occur in the blue-shaded areas. Equilibrium isocline The population size of one species that causes the population of another species to be stable. Also known as Zero growth isocline. We can also plot the equilibrium isocline for the predator population, as shown in Figure 14.10b. In this graph, we draw a vertical line at the point where N = m ÷ ac, which is the number of prey that causes the predator population to be stable. Any combination of predator and prey numbers that lies in the region to the right of this line allows the predator population to increase because there is an increased abundance of prey to consume. In the region to the left, the predator population decreases because there is not enough prey available. We can now combine our understanding of how predator and prey populations change in abundance by considering the trajectory of both populations simultaneously, which is called a joint population trajectory. Joint equilibrium point The point at which the equilibrium isoclines for predator and prey populations cross.
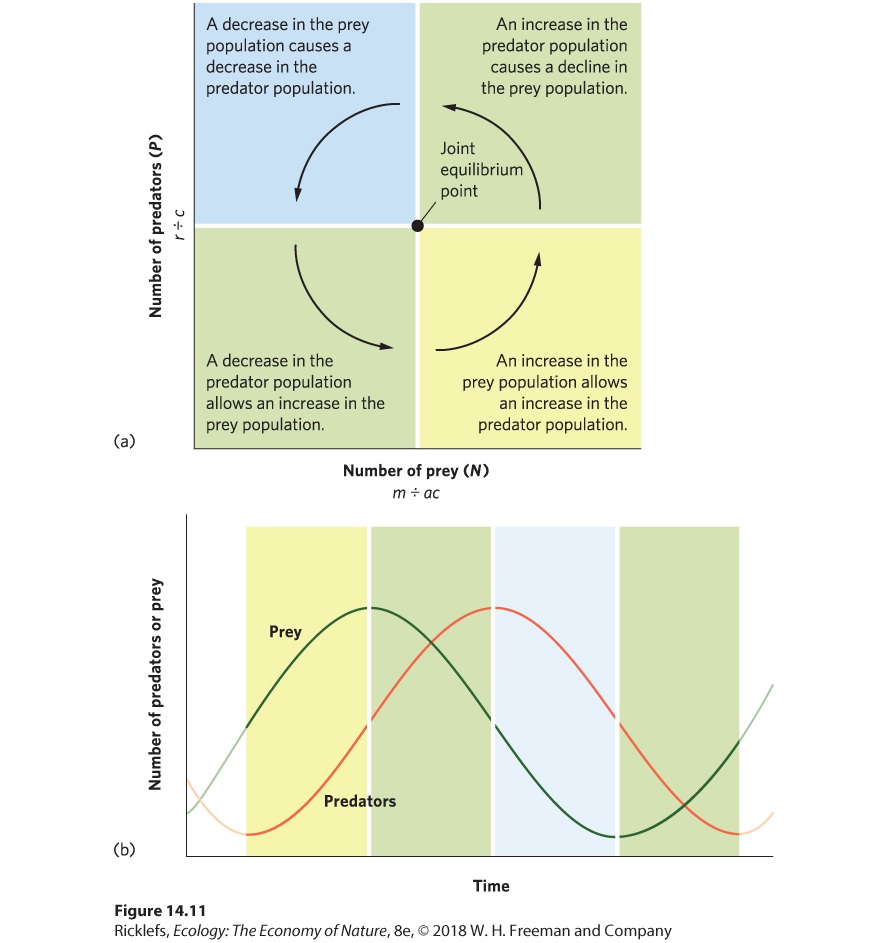
Figure 14.11a traces the path of the joint population trajectory. Beginning with the lower right region, predators and prey both increase, and their joint population trajectory moves up and to the right. In the upper right region, prey are still abundant enough that predators can increase, but the increasing number of predators depresses the prey population. Accordingly, the joint population trajectory moves up and to the left. In the upper left region, the continued decline in prey causes the predator population to decline, so the trajectory moves down and to the left. In the lower left region, the continued decline in predators allows the prey population to start increasing, which causes the trajectory to move down and to the right and completes the cycle. Together, the trajectories in the four regions define a counterclockwise cycling of predator and prey populations.
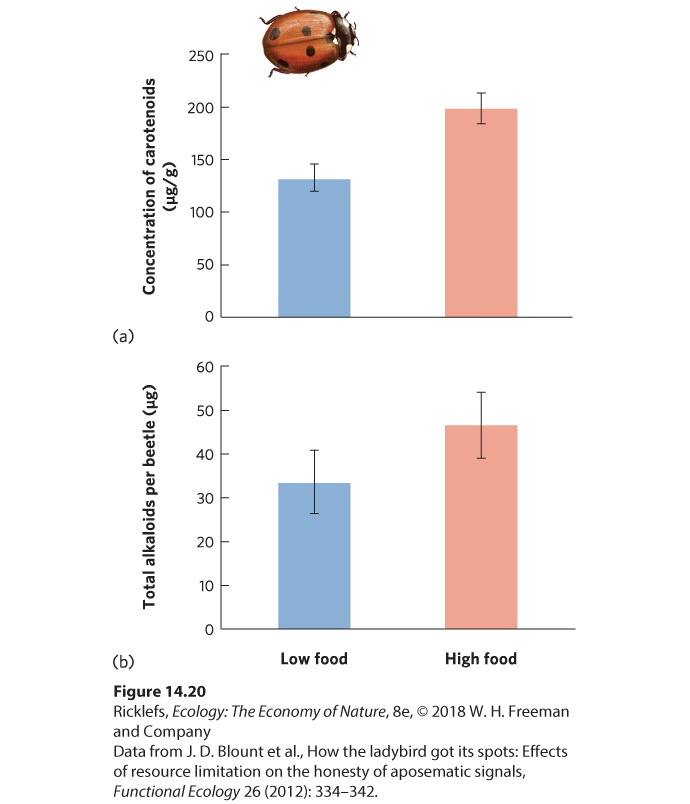
Figure 14.11 Predator–prey oscillations. The Lotka–Volterra model illustrates how the abundance of predator and prey populations cycle. (a) With a decline in predators, prey can increase in abundance. The increase in prey then provides more food for predators; this allows predators to increase their population size through reproduction. As predators grow more numerous, they start to kill prey at such a high rate that the prey population begins to decline. A decline in available prey reduces the survival and reproduction of the predator population, which starts to decline. (b) Over time, both populations cycle up and down in abundance, with predator abundance lagging behind prey abundance. At the center of Figure 14.11a, you can see the joint equilibrium point, which is the point at which the equilibrium isoclines for predator and prey populations cross. The joint equilibrium point represents the combination of predator and prey population sizes that falls exactly at this point and will not change over time. If either of the populations strays from the joint equilibrium point, the populations will oscillate around the joint equilibrium point rather than returning to it.
Joint population trajectory The simultaneous trajectory of predator and prey populations. Based on the joint population trajectory of the Lotka‒Volterra model, we can plot the changes in sizes of the two populations over time, as shown in
Figure 14.11b. This visualizes how the two populations cycle and reveals that the prey population remains one-fourth of a phase ahead of the predator population. Although some populations cycle due to time delays, as we discussed in Chapter 13, the predator–prey equations do not cycle due to time delays; the predator–prey equations are continuous equations in which population changes are immediate. The cycling in this case is the result of each population responding to changes in the size of the other population. This pattern is reminiscent of the lynx population, discussed at the beginning of this chapter, which follows the fluctuations in the hare population.
Functional and Numerical Responses
The Lotka-Volterra model provides an explanation for population cycles that relies on a very simplified version of nature. As we have already discussed, it does not include time delays or density dependence, and it does not incorporate the real foraging behavior of most predators. To get a more realistic picture of predator and prey relationships, we have to consider the functional response and the numerical response of predators. As we will see, both of these responses help to stabilize the cycling of predator and prey populations. The Functional Response The relationship between the density of the prey population and an individual predator’s rate of food consumption is known as the functional response of the predator. There are three potential categories of functional responses, as illustrated in Figure 14.12. For each of these categories, we can examine the number of prey consumed by each predator, shown in Figure 14.12a, or the proportion of prey consumed by each predator, shown in Figure 14.12b. A key point to remember is that whenever prey population density increases and a predator can consume a higher proportion of those prey, the predator has the ability to regulate the growth of the prey population.
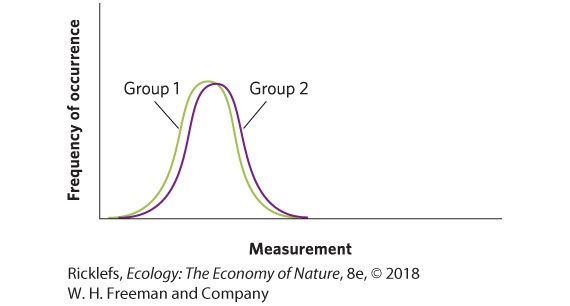
Figure 14.12 Functional responses of predators. (a) When we consider the number of prey consumed per predator, we see that a type I response exhibits a linear relationship between the density of the prey population and the number of prey a predator consumes until eventually the predator is satiated. In contrast, a type II response exhibits a slowing rate of prey consumption as prey density increases; the slowing rate is due to an increase in the time spent handling the additional prey. Eventually, the predator achieves satiation. A type III response also shows a slowing rate of consumption when prey density is high. In addition, it exhibits the effect of a predator learning a search image for prey as the prey density increases across the lower range. (b) When we consider the proportion of prey consumed, we see that the type I response results in a constant proportion of prey being consumed—prior to satiation —as prey density increases. A type II response results in a decreasing proportion of prey being consumed. Finally, a type III response initially causes an increased proportion of prey consumed, followed by a decrease. Functional response The relationship between the density of prey and an individual predator’s rate of food consumption. A type I functional response, indicated by the purple line, occurs when a predator’s rate of prey consumption increases linearly with an increase in prey density until the predator is satiated. As shown in Figure 14.12a, the increase in prey density results in an ever-increasing number of prey consumed by a predator until the predator becomes satiated and can consume no additional prey. Some species of predators, such as web-building spiders that catch an increasing number of prey as the density of the prey population increases, exhibit a type I functional response. As you can see in Figure 14.12b, this means that as prey population density increases, each predator continues to consume a constant proportion of the prey up until the point of predator satiation. Once the prey population becomes so dense that predator
satiation occurs, the predators consume a continuously decreasing proportion of the prey. This is the functional response used by the Lotka‒Volterra model that we discussed. Type I functional response A functional response in which a predator’s rate of prey consumption increases in a linear fashion with an increase in prey density until satiation occurs. The type II functional response, shown by the orange line, occurs when the number of prey consumed slows as prey population density increases and then plateaus when predator satiation occurs. The number of prey consumed slows because as predators consume more prey, they must spend more time handling the prey. For example, when a pelican catches a fish, it must take the time to manipulate the fish in its mouth and position the fish so that it will slide down the pelican’s throat. The more fish that the pelican catches, the more time it must spend handling fish, which leaves less time available to hunt for fish. Eventually, because the predator is spending so much of its time handling large numbers of fish and has little time remaining to catch additional fish, its predation rate levels off. In Figure 14.12b, we can see that in a type II functional response, the slowing rate of prey consumption causes a decline in the proportion of prey consumed by each predator. Type II functional response A functional response in which a predator’s rate of prey consumption begins to slow down as prey density increases and then plateaus when satiation occurs. In a type III functional response, illustrated by the green line in Figure 14.12a, prey consumption is low when prey population density is low, consumption is rapid when prey population density is moderate, and prey consumption slows when prey population density is high. Figure 14.12b shows how this type of functional response affects the proportion of prey consumed. As prey population density increases, there is an initial increase in the proportion of prey consumed. However, as the predators spend more time handling prey and become satiated, this proportion subsequently declines— just as we saw in the type II response. Type III functional response A functional response in which a predator exhibits low prey consumption under low prey
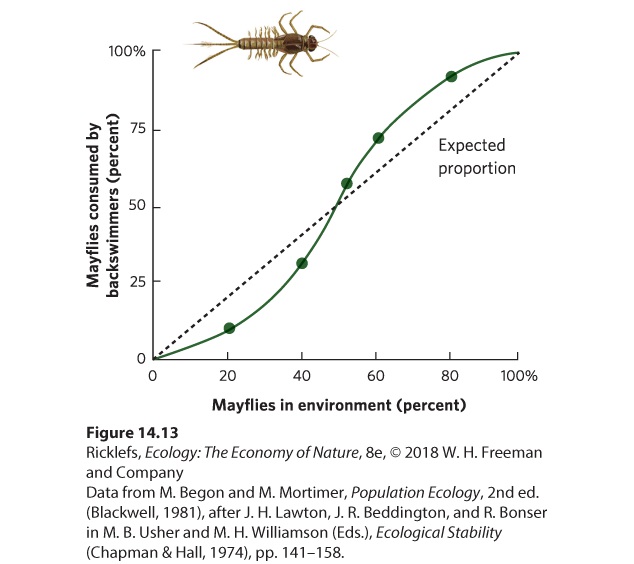
densities, rapid consumption under moderate prey densities, and slowing prey consumption under high prey densities. Low consumption at low prey population density can be the result of three factors. First, at very low prey density, the prey can hide in refuges where they are safe from predators. A predator can consume prey only after the prey become so numerous that some individuals are unable to find a refuge. Second, at low prey population density, predators have less practice locating and catching the prey and therefore are relatively poor at doing it. As prey population density increases, however, the predators learn to locate and identify a particular species of prey, a phenomenon known as a search image. A search image is a learned mental image that helps the predator locate and capture food, just as a person might locate a can of cola in a grocery store by searching for a small, red cylinder. Search image A learned mental image that helps the predator locate and capture food. A third factor that can cause relatively low consumption of prey at low population density is the phenomenon of prey switching. This occurs when one prey species is rare and a predator changes its preference to another prey species that is more abundant. If the population density of the first species then increases, the predator can switch back to it. Many laboratory and field studies have demonstrated type III functional responses. For example, researchers examined the feeding preferences of a predatory insect known as a backswimmer (Notonecta glauca). Figure 14.13 shows what happened when the predator was offered two types of prey— isopods (Asellus aquaticus) and larval mayflies (Cloeon dipterum)—and manipulated the proportions of each prey species. If the predator could not develop a search image, it would consume the two prey species in proportion to their availability. If the predator could develop a search image, it would eat fewer than expected of the rare prey and more than expected of abundant prey. As predicted, when the mayflies were rare, the backswimmer consumed fewer mayflies than expected, given their abundance. When mayflies were abundant, however, the backswimmers consumed more mayflies than expected, based on their abundance.
Figure 14.13 A type III functional response. Using predatory insects known as backswimmers, researchers manipulated the proportion of available prey: larval mayflies (shown above the graph) and isopods. The dashed black line indicates the expected proportion of mayflies consumed if the predator’s preference was not affected by the proportional abundance of mayflies. When mayflies were rare, the predator consumed fewer mayflies than expected at random. When mayflies were common, the predator consumed more mayflies than expected at random. The reason that the predator switched to the more abundant prey was because its success rate was higher with the prey that were abundant. For example, when larval mayflies were rare, predator attacks were successful less than 10 percent of the time. When the mayflies were common, predator attacks were successful nearly 30 percent of the time. Researchers attributed the improved capture rate to practice; the backswimmer became more proficient at catching the mayflies when it had more opportunities to catch them. The backswimmer showed no innate preference for either prey species, only a preference for the one that was more abundant. The Numerical Response As we have seen, the functional response considers changes in the number of

prey consumed by each predator. A predator’s functional response tells us how many prey can be consumed by predators and therefore the conditions under which predators can regulate prey populations. The numerical response is a change in the number of predators through population growth or population movement due to immigration or emigration. Populations of predators usually grow slowly relative to populations of their prey, although the movement of mobile predators from surrounding areas can occur quite rapidly when the population density of prey changes. Numerical response A change in the number of predators through population growth or population movement due to immigration or emigration. An example of numerical response occurs in local populations of the baybreasted warbler (Dendroica castanea), a small insectivorous bird of eastern North America. During outbreaks of the spruce budworm, populations of the warbler increase dramatically. In most years, the warbler populations live at densities of about 25 breeding pairs per square kilometer. However, during outbreak years, the warblers congregate in areas where the spruce budworm is abundant and warbler population densities can reach 300 breeding pairs per square kilometer. As a result of this rapid numerical response, the predator has the potential to reduce prey densities quickly and to regulate the prey’s abundance.
Concept Check
1. How does a prey population’s ability to disperse allow the prey to persist in the presence of predators? 2. Based on the predator–prey population equations, why is the prey population stable when rN = cNP? 3. What causes the difference between a predator exhibiting type II versus type III functional responses?

#### 14.3 Predation and Herbivory Favor the Evolution of Defenses
14.3 Predation and herbivory favor the evolution of defenses Given the large effects that predators can have on their prey and herbivores can have on producers, it is not surprising that many species have evolved strategies to defend themselves. In this section, we will review the types of defenses that prey have evolved and how some predators have evolved counterdefenses.
Defenses Against Predators
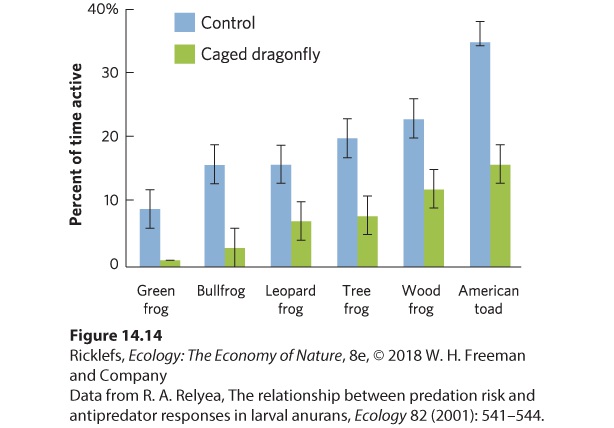
To understand the defenses prey use against predators, we first need to understand predator hunting strategies. Predator hunting strategies can be categorized as either active hunting or ambush hunting, also known as sitand-wait hunting. A predator that uses active hunting spends most of its time moving around looking for potential prey. For example, American robins actively hunt when they move around a lawn searching for earthworms. In contrast, a predator that uses ambush hunting lies in wait for a prey to pass by. Chameleons can sit very still as they wait for an insect to pass. When the insect is close enough, the chameleon shoots out its long tongue and the sticky, prehensile tip grabs the unsuspecting prey. Regardless of hunting mode, we can think of hunting by predators as a series of events: detecting the prey, pursuing the prey, catching the prey, handling the prey, and consuming the prey. As we will see, prey have evolved defenses to thwart the predator at different points in this series of events. Behavioral Defenses Some of the most common behavioral defenses against predators include alarm calling, spatial avoidance, and reduced activity. Alarm calling is used by many species of birds and mammals to warn their relatives that predators are approaching. Prey that use spatial avoidance move away from the predator. Prey that follow a strategy of reduced activity reduce their activity when predators are detected, making them less likely to come into contact with a predator. In a study of six species of tadpoles, the animals were placed in tubs of water with one of two treatments. The first treatment contained a
caged predator that could not kill the tadpoles but could emit chemical cues that tadpoles can detect. The second treatment was a control, which consisted of an empty cage. Once the experiment was set up, the tadpoles were watched to determine their activity level, which was defined as the percent of the time that the animals spent moving. As shown in Figure 14.14, each species exhibited a different level of activity when predators were absent and all species reduced their level of activity in the presence of the predator.
Figure 14.14 Behavioral defenses. Tadpoles commonly avoid predation by becoming less active, which is measured as the portion of the day that they spend moving. Being less active reduces the probability of being detected by a predator. Error bars are standard errors. Crypsis Another way to reduce the probability of being detected by a predator is through camouflage that either matches the environment or breaks up the outline of an individual to blend in better with the background environment, a phenomenon known as crypsis. Various animals resemble sticks, leaves, flower parts, or even bird droppings. These organisms are not so much concealed but mistaken for inedible objects and passed over by predators. Common species that use crypsis include stick insects, katydids, and horned lizards (Figure 14.15). Some species have a fixed color pattern that aids in crypsis, while other species, such as the octopus, are able to rapidly change color in ways that make themselves match their background.
Figure 14.15 Cryptic prey. Some prey avoid detection by blending into the environment. Crypsis is a strategy used by a number of animals including those shown here: (a) a katydid and (b) a horned lizard (Phrynosoma platyrhinos). Crypsis Camouflage that either allows an individual to match its environment or breaks up the outline of an individual to blend in better with the background environment. Structural Defenses Although some prey use behavior and crypsis to avoid being detected, other species employ mechanical defenses that reduce the predator’s ability to capture, attack, or handle the prey. One of the best-known examples of a mechanical defense is the barbed quills of the porcupine; more than 30,000 quills cover the porcupine’s body and can penetrate the flesh of an attacking predator. In other species, the structural defenses are phenotypically plastic and therefore induced only when the prey detects a predator in the environment. For example, water fleas—tiny freshwater crustaceans—that detect the chemical cues of predators early in their life can develop spines along different parts of their body to deter predators from consuming them. Other mechanical defenses involve changes in the overall shape of the body. For example, the crucian carp (Carassius carassius), a species of fish that lives in Europe and Asia, develops a deep, hump-shaped body over a period of many weeks when the carp smells a predatory fish in the water. As you can see in Figure 14.16, carp with a hump-shaped body have greater muscle mass and can accelerate more quickly away from a predator.
Figure 14.16 Structural defenses. In the crucian carp, individuals living with predatory fish develop a deep, hump-shaped body that allows the carp to escape predation. These predatorinduced fish have (a) greater muscle mass, which allows (b) greater acceleration while swimming away from predators. Error bars are standard errors. Chemical Defenses Prey can also use chemical defenses to deter a predator. Skunks, well known for using this strategy, spray potential threats with foul-smelling chemicals from posterior glands. Many insects also use chemical defenses. For example, when monarch butterfly caterpillars feed on milkweed, they store some of the milkweed toxins in their body, which make the butterfly very distasteful to predatory birds. Bombardier beetles take a different approach. Their abdomens contain two glands, each of which makes a distinct chemical. When agitated, the beetle mixes these two chemicals, causing a reaction that
makes the liquid approach 100 °C. They shoot the boiling hot liquid out of the abdomen, causing pain or death to small predators, as illustrated in Figure 14.17.
Figure 14.17 Chemical defenses. The bombardier beetle (Stenaptinus insignis) mixes two chemicals inside its abdomen that react to release a boiling hot fluid to deter predators. While chemical defenses are often effective in deterring predation, these defenses are even more effective if the prey can communicate their distastefulness to the predator before an attack occurs. In many species, distastefulness has evolved in association with very conspicuous colors and patterns, a strategy known as warning coloration, or aposematism. Predators quickly learn to avoid markings such as the black and orange stripes of monarch butterflies; the insect tastes so bitter that a single experience is well remembered. Conspicuous combinations of black, red, and yellow adorn such diverse animals as bombardier beetles, yellow-jacket wasps, and coral snakes. These color combinations consistently advertise distastefulness or a threat of harm; some predators have evolved innate aversions to such prey and, therefore, do not need to learn to avoid them. Warning coloration A strategy in which distastefulness evolves in association with very conspicuous colors and patterns. Also known as aposematism.
Mimicry of Chemical Defenses When predators avoid aposematic species, any individuals of palatable species that resemble the distasteful, aposematic species would also be favored by selection. Over generations, a palatable species can evolve to more closely resemble an aposematic species, a phenomenon called Batesian mimicry, named for Henry Bates, the nineteenth-century English naturalist who first described it. In his journeys to the Amazon region of South America, Bates found numerous cases of palatable insects that did not retain the cryptic patterns of their close relatives but instead had evolved to resemble brightly colored, unpalatable species (Figure 14.18).
Figure 14.18 Batesian mimicry. (a) The common wasp (Vespula vulgaris) possesses aposematic coloration as a warning to predators that an attack could result in being stung and injected with painful chemicals. Other harmless species that do not possess any stinging ability have evolved to resemble the color patterns of the wasp. These include (b) a species of hover fly (Helophilus pendulus) and (c) the hornet clearwing (Sesia apiformis), which is a species of moth. Their resemblance to the wasp reduces their risk of predation. Batesian mimicry When palatable species evolve warning coloration that resembles unpalatable species. Studies have demonstrated that mimicry does indeed confer an advantage to the mimics. For example, toads that consumed live bees and received a sting on the tongue subsequently avoided palatable drone flies, which mimic the appearance of bees. In contrast, when naive toads consumed dead bees with stingers removed, they consumed the bees and the drone flies. This result indicates that toads learned to associate the conspicuous and distinctive color patterns of live bees with an unpleasant experience. Another type of mimicry, called Müllerian mimicry, occurs when several unpalatable species evolve a similar pattern of warning coloration. Müllerian
mimicry is named after its discoverer, the nineteenth-century German zoologist Fritz Müller. When multiple species of prey have conspicuous color patterns and all are unpalatable, a predator that learns to avoid one prey species will later avoid all prey species with a similar appearance. For example, most of the bumblebees and wasps that co-occur in mountain meadows share a pattern of black and yellow stripes and they all have the ability to sting a predator. Similarly, in Peru, several species of poison dart frogs, all in the genus Ranitomeya, closely resemble each other. In four regions of that country, researchers have found three species that vary in coloration according to location, including one species (R. variabilis) that looks very different across two locations. A fourth species, R. imitator, is also unpalatable and has populations in each of these four locations that closely resemble the other species that is present (Figure 14.19).
Figure 14.19 Müllerian mimicry. Müllerian mimics are a collection of unpalatable species that share a pattern of warning coloration. At four locations in Peru, researchers have found pairs of species of poison dart frogs that are unpalatable and closely resemble each other. For each location, the frog on the left is a particular species of unpalatable frog. In all four locations, the frog on the right is the unpalatable R. imitator, so named because in each location it has evolved to imitate the coloration of the other frog. Müllerian mimicry When several unpalatable species evolve a similar pattern of warning coloration. Costs of Defenses Against Predators
Many types of defenses against predators can be costly, as we discussed in our coverage of predator- and herbivore-induced defenses in Chapter 4. For example, behavioral defenses such as spatial avoidance can result in reduced feeding or increased crowding as prey move to locations away from predators. In such cases, behavioral defenses often come at the cost of reduced growth and development. Similarly, most mechanical defenses, such as the 30,000 quills of a porcupine, are energetically expensive to produce. When the costs of defense are so high that they come at the cost of growth and reproduction, the presence of predators can cause smaller prey population sizes even when they do not consume the prey. Less is known about the costs of chemical defenses in prey, but studies suggest that these defenses are also energetically costly to produce. In ladybugs, which are technically known as ladybird beetles, many species are red with black spots. These warning colors communicate to predators that the ladybugs taste bad because of chemicals in their bodies known as alkaloids. However, there is a large amount of variation in the concentration of alkaloids that each ladybug can produce. In 2012, researchers reported that only beetles that consumed large amounts of food had sufficient energy to produce high concentrations of carotenoids, which give the ladybugs a more intense red color, as shown in Figure 14.20a. Moreover, as illustrated in Figure 14.20b, beetles that produced more carotenoids also produced higher concentrations of alkaloids. As a result, ladybugs with the highest energy diet can better advertise their level of toxicity to predators and thereby reduce their chances of being attacked.
Figure 14.20 Costly chemical defenses in ladybugs. In male ladybugs, individuals given a high amount of food produced (a) a higher concentration of carotenoids that make the ladybug’s body more intensely red, and (b) a greater amount of defensive alkaloid chemicals. Error bars are standard errors. Counter Adaptations of Predators If predation can select for prey to evolve a wide range of defenses, then prey defenses should favor the selection for counter-adaptation in predators. In this way, predators and prey experience an evolutionary arms race between prey defenses and predator offenses. When two or more species affect each other’s evolution, we call it coevolution. In the case of the porcupine, for example, the spines deter most predators. However, bobcats (Lynx rufus) and wolverines (Gulo gulo) have an effective solution. When these predators find a porcupine, they flip the porcupine on its back and attack the belly, which is
not defended by spines. Other common predator adaptations include highspeed locomotion to catch their prey and camouflage that allows them to ambush their prey. Coevolution When two or more species affect each other’s evolution. Some predators can also evolve to handle the toxic chemicals produced by prey. For example, the cane toad (Bufo marinus) is a species that was introduced into Australia in 1935. Like other species of toads, the cane toad contains toxins in its skin that can cause predators to become sick or die. As a result, predators in the native range of cane toads do not attack cane toads even though these same predators regularly consume other species of amphibians. When the toads were introduced to Australia, predators of amphibians, such as black snakes (Pseudechis porphyriacus), had no evolutionary experience with cane toads. When they ate the toads, most snakes had no resistance to the toxins and they died. However, some populations of the black snakes were consuming cane toads, which suggests that while most black snakes that consumed cane toads died, some snakes must have been resistant to the toad’s toxins and survived. Over time, the selection for snake resistance to the toad toxin must have resulted in the evolution of resistant populations. To test this hypothesis, the researchers fed samples of toad skin to snakes from different populations around Australia. They then examined how much the toad skin reduced the swimming speed of each snake, which indicates the susceptibility of each snake to the toxin. As you can see in Figure 14.21, the snake populations that coexisted with cane toads for the longest amount of time evolved the lowest susceptibility to the cane toad toxin. This evolution would have occurred in less than 70 years, which is a remarkably short time.
##### Analyzing Ecology: Understanding Statistical Significance
Figure 14.21 Counter-adaptations of predators to prey chemical defenses. Researchers measured the swimming speed of black snakes before and after being fed the skin of cane toads, which contains toxins. They found that snake populations that had coexisted for the longest times with the toad had the lowest susceptibility to the toad toxin.
Analyzing Ecology
Understanding Statistical Significance In examining adaptations of prey, counteradaptations of predators, or any other ecological measurements, we often consider experiments in which researchers find differences in the outcomes of experimental manipulations. Until now, we have not explored how ecologists assess when such differences are meaningful versus when the differences are due to chance. For any group of measurements, such as the concentration of toxins in ladybugs fed high and low amounts of food, there will be variation
among the individuals of each group. If we were to sample ladybugs at random from the high- and low-food treatment groups, we would find that the mean toxin concentration is higher in the high-food treatment group. However, the measurements taken on some of the individuals in the high-food treatment group might overlap with the measurements taken on some of the individuals in the low-food treatment group. When the means are similar and the distribution of the data from two groups is almost entirely overlapping, we would have to conclude that the two groups are nearly identical in whatever we are measuring, as shown in the figure below. In contrast, when the means are very far apart and the distribution of the data from two groups shows no overlap, as in the case of the next graph, we would feel confident that the two groups are completely different in whatever we are measuring.
Although it is rare for two groups to have completely overlapping or completely nonoverlapping distributions, we need to know if the degree of overlap between the two data sets is acceptable in order to conclude that the groups are different from each other, with regard to the variable being measured. Scientists agree that two distributions can be considered “significantly different” if we can sample the two distributions many times and find that the means of those distributions overlap less than 5 percent of the time. This somewhat arbitrary, but widely accepted, cutoff value is known as alpha (α). Thus, we say that our cutoff for statistical significance is α < 0.05. Determining that something has statistical significance is not the same as stating that a difference between two means is large, substantial, or important. In other words, the everyday use of “significant” is not synonymous with the scientific use of “significantly different.” YOUR TURN In Chapter 2, “Analyzing Ecology: Standard Deviation and Standard Error,” we mentioned that when data have a normal distribution, about 68 percent of the data fall within 1 standard deviation of the mean, 95 percent of the data fall within 2 standard deviations of the mean, and 99.7 percent of the data fall within 3 standard deviations of the mean. Based on this information, if you had two groups of data with identically shaped distributions of data, approximately how many standard deviations apart would they have to be in order for the two groups to be considered significantly different?
Defenses Against Herbivores
Just as selective pressure from predators has caused the evolution of prey defenses, selective pressure from herbivores has caused the evolution of defenses against herbivory. In some cases, these defenses are induced by an herbivore attack and are therefore phenotypically plastic, as we saw in
Chapter 4. In other cases, plant defenses are fixed and therefore expressed whether or not an herbivore has attacked the plant. In both cases, some herbivore species have evolved counter-adaptations. In fact, some species of herbivores are so specialized in countering the defenses of a particular plant species that they do not consume any other plants. Structural Defenses When it comes to structural defenses, plants have evolved a variety of traits to deter herbivores from consuming their leaves, stems, flowers, or fruit. Some plants, such as cacti, roses, and blackberries, have sharp spines and prickles that inflict pain on the mouths of herbivores. Other plants grow a wooly layer of hair over their leaf surfaces to make it difficult for herbivorous insects to penetrate them. Chemical Defenses A wide variety of chemical defenses have evolved in plants. Plant chemicals include sticky resins and latex compounds that are hard to consume. Some plants also produce alkaloids—including caffeine, nicotine, and morphine— that have a wide range of toxic effects on herbivores. Other chemicals in plants, such as tannins, are difficult for herbivores to digest. It is generally thought that the chemicals produced by plants are the by-products of a plant’s physiology. For many of these chemical defenses, one or more species of herbivore has evolved tolerance. For example, a shrub in Polynesia called the Tahitian noni (Morinda citrifolia) produces toxic chemicals with such a foul smell that the plant has the nickname “vomit fruit” (Figure 14.22). Most species of fruit flies avoid this plant because if they were to land on it, they would die. However, one species of fruit fly (Drosophila sechellia) has evolved the ability to tolerate the defensive chemicals. It lays its eggs on the vomit flower, and they have a distinct fitness advantage because they experience no competition
Figure 14.22 Chemical plant defenses and counter-adaptations of specialized herbivores. The Tahitian noni, also known as the vomit flower, is a shrub that produces such a foul smell that most species of fruit flies avoid it. However, one species of fruit fly has evolved that tolerates the chemicals and therefore lays its eggs on the plant. A similar situation exists for the monarch butterfly. As mentioned in our discussion of prey defenses, the monarch caterpillar specializes in feeding on milkweed plants, which produce toxic chemicals. The monarch caterpillar readily feeds on milkweed plants because it has evolved to resist the effects of a group of chemicals known as cardiac glycosides, which can stop the heart of many other herbivores. It also sequesters some of the chemicals to use as a defense against its own predators. In 2012, researchers made a striking discovery about the evolution of insects’ ability to consume the plants containing cardiac glycosides. They found that a diversity of insects from different orders—including flies, beetles, true bugs, and butterflies— independently evolved the same changes in a gene that offers resistance to the effects of the toxin. In short, the different groups of insects exhibited convergent evolution. Tolerance to Being Eaten
Some plants that have not evolved extensive defenses against herbivores take an alternative strategy of tolerating herbivory. Plants taking this strategy are able to grow new tissues rapidly to replace those that are consumed. For example, herbivores often consume the top meristem of a plant, which is the region of the plant where most growth occurs. When this meristem is removed, the meristems of lower stems begin to experience increased rates of growth, which still allows the plant to experience relatively high fitness despite being partially consumed by an herbivore. Costs of Herbivore Defenses For decades, researchers have investigated whether plant defenses come at the cost of reduced fitness. When defense traits are phenotypically plastic, researchers can compare the fitness of individuals with induced defenses against the fitness of noninduced individuals. For example, tobacco plants (Nicotiana sylvestris) respond to herbivory by producing chemical defenses including nicotine. Researchers damaged one group of tobacco plants to induce an increase in chemical defenses. As a control, they also damaged a second group of plants and then treated the damaged areas with a plant hormone that prevented the chemical defenses from responding. When they later counted the number of seeds produced by the two groups, they found that the group with increased chemical defenses produced fewer seeds, as shown in Figure 14.23.
Figure 14.23 Costs of defenses against herbivores in tobacco plants. Researchers damaged tobacco plants to simulate the act of herbivory. They then prevented a chemical response in some individuals by applying plant hormones to the damaged area of the leaves. (a) Damaged plants treated with the plant hormone produce a lower amount of chemical
defense than damaged plants not treated with the hormone. (b) Damaged plants treated with the plant hormone also experienced higher lifetime fitness, as measured by the mass of seeds produced. Error bars are standard errors. A second approach to quantifying the costs of defenses against herbivores is to make the genes responsible for defenses nonfunctional. For instance, researchers have examined the growth of different genotypes of mouse-ear cress, a tiny plant native to Europe and Asia. They reported that individuals with intact defense genes commonly grew more slowly than individuals with nonfunctional defense genes. This confirmed that plants pay a cost for defenses against herbivores.
Concept Check
1. What are four ways in which prey have evolved to reduce their risk of being killed by predators? 2. Why does natural selection favor counter-adaptations to prey defenses? 3. What are three ways in which plants have evolved to reduce their risk of being killed by herbivores?
Concepts
Introduced species on Macquarie Island. Introduced rabbits have caused devastating effects on the plants of the island, which do not have an evolutionary history of living with rabbits. Attempts to reduce the rabbit populations have caused feral cats to switch from eating rabbits to eating the island’s native birds. Islands throughout the world often contain a variety of endemic species that have evolved together for millions of years. As we have seen in this chapter, new species that are introduced to islands can have devastating effects on the native plants and animals. Because the native species do not share an evolutionary history with the introduced species, they have not evolved defenses against introduced predators and herbivores. These introductions happen commonly on islands. In response, we often attempt to remove the intruders to reverse the harmful effects, although the actual outcomes of these efforts can lead to unintended consequences. An example of this occurred on Macquarie Island, a small island located halfway between Australia and Antarctica. Island plants and animals existed together for eons in a tundra biome that contained a large number of
seabirds and land birds, as well as a diversity of plants that included tall grasses. Humans began to visit the island in the 1800s, and they brought several species of animals with them. In the early 1800s, the island was used by seal hunters as a place to rest and to resupply their ships. These visitors introduced house cats (Felis catus), which soon became feral. While there are no data on the impact of the feral cats, it is generally assumed that they were mesopredators and preyed on the abundant island birds. In 1878, the seal hunters introduced European rabbits (Oryctolagus cuniculus) to the island to serve as a food source whenever the sailors returned to the island. Despite the fact that the cats fed on the rabbits, over time the rabbit population grew to very large numbers. Data collected from the 1950s through the 1970s suggest that the rabbit population experienced large fluctuations approximately every 10 years, similar to those seen in hares and other animals that live in high northern latitudes. Because the plants on the island had no evolutionary history with the rabbits, periods of high rabbit numbers caused devastating effects on the abundance of palatable plant species; when the rabbit population experienced large declines, the vegetation rebounded.
Removal of introduced species on Macquarie Island. (a) By 2011, severe degradation of vegetation existed on Macquarie Island, resulting from introduced rabbit populations. (b) The problem was corrected with the removal of every last non-native mammal on the island. In 2014, this effort was declared a success with every rabbit and mouse removed, and a rapid improvement in the vegetation, native birds, and several native invertebrate animals that depend on the vegetation. Because periods of high rabbit populations caused such adverse effects on the island’s vegetation, scientists introduced the European rabbit flea (Spilopsyllus cuniculi) in 1968. The fleas can carry a Myxoma virus that causes the disease myxomatosis, which is fatal to rabbits. The scientists quickly learned that the virus did not persist well on the island, so beginning in 1978, they reintroduced it every year. The virus had its desired effect and caused the rabbit populations to plunge from a high of 130,000 individuals in 1978 to fewer than 20,000 by the 1980s. As the rabbit population declined, the vegetation of the island began to recover. However, the changes to the island community did not end there. With the rapid decline of the rabbits, the cats of the island exhibited prey-switching behavior—described in our discussion of predator functional responses—and added more birds to their diet. They consumed an estimated 60,000 seabirds per year and caused the extinction of two species of endemic land birds. To help save the island’s undefended birds, government officials decided to eradicate all the cats from the island. Starting in 1985, scientists removed 100 to 200 cats from the island each year. An analysis of the cat stomachs in 1997 indicated that while the cat population was killing
birds, the cat population was still consuming approximately 4,000 rabbits per year, so the cats—in combination with the Myxoma virus—were probably still regulating the rabbit population. When the last cat was removed in 2000, the island community responded rapidly. For example, five species of seabirds started breeding on the island, some of which had not bred there for more than 80 years. The rabbits also responded. From 1985 to 2000, the rabbit population increased from approximately 10,000 to more than 100,000 individuals. This increase was probably caused by a combination of fewer predatory cats, a rebounding abundance of plants, and a decline in the amount of virus distributed each year. By 2006, there were 130,000 rabbits, and these herbivores had consumed so much vegetation that parts of the island resembled a closely cropped lawn. These changes on the island underscored the importance of understanding all the interactions that introduced species can have on a community, including interactions among the introduced species themselves. In the case of Macquarie Island, it has become clear that removing the introduced cats was not enough to return the island community to its former condition. The introduced rabbits had to be removed as well. As a result, the Australian government agreed to spend $24 million to remove the rabbits from the island as well as several other introduced mammals, including mice and rats. In 2011, grain-containing poison was distributed around the island by helicopter, and this was very successful in killing the vast majority of the introduced mammal species. From 2012 to 2013, hunters with specially trained dogs were brought to the island to eliminate the few remaining rabbits,
mice, and rats. It is estimated that the hunters walked a total of 92,000 km in their search for every last non-native mammal on the island. In 2014, this effort was declared a success with every rabbit and mouse removed and a rapid improvement in the vegetation, native birds, and several native invertebrate animals that depend on the vegetation. SOURCES: Success on Macquarie Island. 2014. Macquarie Island Pest Eradication Project Newsletter, Issue 14, July. http://www.parks.tas.gov.au/file.aspx?id=36472. Bergstrom, D. M., et al. 2009. Indirect effects of invasive species removal devastate World Heritage Island. Journal of Applied Ecology 46: 83–81. Dowding, J. E., et al. 2009. Cats, rabbits, Mxyoma virus, and vegetation on Macquarie Island: A comment on Bergstrom et al. (2009). Journal of Applied Ecology 46: 1129–1132. Bergstrom, D. M., et al. 2009. Management implications of the Macquarie Island trophic cascade revisited: A reply to Dowding et al. (2009). Journal of Applied Ecology 46: 1133–1136. http://www.abc.net.au/news/2012-04-19/rabbit-hunters-head-to- macquarie-island/3961270.
Summary of Learning Objectives
14.1 Predators and herbivores can limit the abundance of populations. Using observations in nature and manipulative experiments, ecologists have found that predators commonly limit the abundance of prey and herbivores commonly limit the abundance of producers. Key Terms: Introduced species, Exotic species, Non-native species, Invasive species, Mesopredators, Top predators
14.2 Populations of consumers and consumed populations fluctuate in regular cycles. Cycling populations have been observed frequently in nature and recreated in laboratory experiments. Lags in the response times of predator movement and reproduction linked to changes in the abundance of prey cause these cycles. Mathematical models have been developed to mimic the cycling behavior of predator and prey populations. Key Terms: Lotka–Volterra model, Equilibrium isocline, Joint population trajectory, Joint equilibrium point, Functional response, Type I functional response, Type II functional response, Type III functional response, Search image, Numerical response
14.3 Predation and herbivory favor the evolution of defenses. Prey have evolved a wide variety of defenses, including behavioral defenses, mechanical defenses, chemical defenses, crypsis, and mimicry. Producers have evolved defenses against herbivores, including mechanical defenses, chemical defenses, and tolerance. Evolved defenses are commonly costly and can sometimes be countered by subsequent adaptations in predators. Key Terms: Crypsis, Warning coloration, Aposematism, Batesian mimicry, Müllerian mimicry, Coevolution
Critical Thinking Questions
1. How could you experimentally test whether herds of African antelope affect the abundance of plants on which they graze? 2. Explain why an herbivore that consumes many different species of plants might be less successful at regulating the abundance of a welldefended plant species compared to an herbivore that specializes on eating a single species of plant. 3. How do the results of the classic experiments of C. F. Huffaker, using mites and oranges in the lab, inform us about how predator and prey populations are able to persist in nature? 4. In evolutionary terms, explain why introduced species can often have harmful effects on native species but can also be controlled by an enemy that comes from the introduced species’ native region. 5. Using the lynx–hare interaction, explain in words the equations of the Lotka–Volterra model for the change in the population sizes of prey and predators. 6. According to the Lotka–Volterra model of predator–prey interactions, why do the populations of foxes and rodents cycle? 7. Compare and contrast a predator’s numerical response and functional response. 8. How might a type II functional response prevent a predator from controlling a large prey population? 9. What are the causes of an evolutionary arms race between consumers and the species that they consume? 10. For each of the five stages in a predation event, how might prey evolve a defense against predators?
#### Graphing the Data: the Functional Response of Wolves
GRAPHING THE DATA The Functional Response of Wolves In Gates of the Arctic National Park in Alaska, researchers monitored the densities of wolves and their major prey, including caribou (Rangifer tarandus). To understand whether the wolves could potentially regulate the growth of the caribou population, they wanted to know the shape of the wolves’ functional response. They could do this by determining the number of caribou killed by wolves in different areas and at different times of the year. Using the data from this study, plot the relationship between caribou density and the number of caribou killed per wolf. Then plot the relationship between caribou density and the proportion of caribou killed per wolf. Based on your graphs, what type of functional response do the wolves have?
Proportion of Caribou
KILLED PER WOLF (PER DAY) 0.1 0.50 1.80 0.2 0.70 0.90 0.3 0.90 0.50 0.4 0.95 0.30 0.5 0.98 0.22 1.0 1.00 0.15 1.5 1.01 0.10 2.0 1.02 0.07 2.5 1.03 0.05 3.0 1.03 0.04