15 Parasitism and Infectious Diseases
15Parasitism and Infectious Diseases A parasitized amber snail. The snail on the right has one normal eye stalk that is pale and slender and another that is infected by a parasitic flatworm, which causes the eye stalk to become enlarged and colorful. It also pulsates in a way that is attractive to predatory birds. The Life of Zombies Zombie films scare us because they depict zombies as the walking dead in search of victims they will turn into more zombies. But something similar to this happens commonly in nature; some species of parasites infect a host and take control of its life for their own benefit. Consider the case of the amber snail (Succinea putris) from Europe. This animal lives along the edges of streams and ponds. It normally spends its time in the shade of terrestrial vegetation, where it chews on leaves and remains hidden from the eyes of predatory birds. An

amber snail will occasionally consume bird feces, which sometimes contain the eggs of a parasitic flatworm (Leucochloridium paradoxum). These eggs hatch inside the snail and grow, but to reproduce, the parasites must spend the next stage of their lives inside a bird. To achieve this goal, the parasitic larvae slowly make their way into the snail’s eyestalks. These eyestalks are normally pale and slender, but the parasitic infection causes the eyestalks to become enlarged and banded with colors that pulsate in a way that resembles a moving caterpillar. The parasites also take control of the snail’s brain and force the snail to move up a plant’s stem, which is not something an amber snail normally does. Snails that move up plant stems are more easily noticed by predatory birds and, because the eyestalks look like caterpillars, birds consume the infected snails. The parasite completes the second stage of its life inside the bird and the cycle continues. The flatworm reproduces, its eggs leave the bird through feces, and snails consume them. Parasites can control the behavior of many different animals. In Thailand, for example, carpenter ants (Camponotus leonardi) normally spend their time living in nests in the hot, dry rainforest canopy. Occasionally, the carpenter ants travel from the canopy to the ground, where they can be exposed to the spores of the fungus Ophiocordyceps unilateralis, the so-called zombie-ant fungus, before heading back up to their nests. An ant that is infected subsequently moves down to the humid, understory vegetation. It stops at about 25 cm above the ground in a place that many previously infected ants have also gone, known to researchers as the “ant graveyard.” Here, it bites the underside of a leaf and maintains a death grip on a leaf vein as it dies. After the ant dies, the fungus grows a spore-producing structure out of the ant’s head and releases its spores into the environment.
An infected carpenter ant. In Thailand, carpenter ants that become infected by a fungus crawl down from the canopy and attach themselves to the underside of a leaf by biting the leaf vein and then dying. After death, a spore-producing stalk grows out of the ant’s head and releases its spores into the environment. “The ability of parasites to act as puppet masters over the behavior of their victims is just one way that parasites have evolved to improve their fitness.” This unusual ant behavior does not benefit the ant, but it greatly benefits the fungus. The higher humidity found lower in the rainforest is more suited to fungal growth than the drier conditions high in the canopy. Moreover, if the infected ant were to die in its nest in the canopy, its nest mates would remove the corpse before the fungus could grow the spore-producing structure that is critical to its reproduction. The ability of parasites to act as puppet masters over the behavior of their victims is just one way that parasites have evolved to improve their fitness. As we will discover in this chapter, parasites come in a wide variety of forms, and their effects on hosts can range from mild to lethal. Adaptations that enable parasites to infect hosts and adaptions
that help hosts to resist infections by parasites offer intriguing insights into the strategies of parasite–host interactions. SOURCES: Andersen, S. B., et al. 2009. The life of a dead ant: The expression of an adaptive extended phenotype. American Naturalist 174: 424–433. Hughes, D. P., et al. 2011. Behavioral mechanisms and morphological symptoms of zombie ants dying from fungal infection. BMC Ecology 11: 13. Lefevre, T., and F. Thomas. 2008. Behind the scene, something else is pulling the strings: Emphasizing parasitic manipulation in vector-borne diseases. Infection, Genetics and Evolution 8: 504–519. Wesolowska, W., and T. Wesolowski. 2014. Do Leucochloridium sporocysts manipulate the behavior of their snail hosts? Journal of Zoology 292: 151–155.
Learning Objectives
After reading this chapter, you should be able to:

15.1 Identify the many different types of parasites affecting the abundance of host species.
15.2 Describe how parasite and host dynamics are determined by the parasite’s ability to infect the host.
15.3 Illustrate how parasite and host populations commonly fluctuate in regular cycles.
15.4 Explain the process of parasites evolving offensive strategies, while hosts evolve defensive strategies. The struggle between parasites and hosts has produced many fascinating examples of ecological interactions and evolutionary adaptations. In Chapter 1, we defined a parasite as an organism that lives in or on another organism, called the host, and causes harmful effects as it consumes resources from the host. It is estimated that approximately half of all species on Earth are parasites. Some hosts have infection resistance, which is the ability of a host to prevent the parasite from causing an infection, while other hosts have

infection tolerance, which is the ability of a host to minimize the harm once an infection has occurred. The number of parasites of a given species that an individual host can harbor is known as the host’s parasite load. A parasite typically infects only one or a few species of hosts, whereas a given host species can contain dozens of species of parasites. Infection resistance The ability of a host to prevent an infection from occurring. Infection tolerance The ability of a host to minimize the harm once an infection has occurred. Parasite load The number of parasites of a given species that an individual host can harbor. Parasites that can cause an infectious disease are called pathogens. Infection by a pathogen does not always result in a disease. For example, humans can be infected with human immunodeficiency virus (HIV), but they may never experience the disease symptoms known as Acquired Immune Deficiency Syndrome (AIDS). In many cases, it is not known what causes a host to transition from being infected by a pathogen to having the disease. Infectious diseases take a large toll on people; the World Health Organization estimates that more than 25 percent of all human deaths are caused by infectious diseases. Note that only infectious diseases are caused by pathogens; there are many noninfectious diseases in which pathogens do not play a role, such as heart disease. In this chapter, we will focus on the interaction between parasites and their hosts. In later chapters, we will discuss the larger role that parasites can play in communities and ecosystems. We will begin by looking at the many different types of parasites that exist, including those that have large effects on crops, domesticated animals, and human health. We will then examine the factors that determine whether parasites can infect hosts, spread rapidly through a population, and cause widespread harmful effects. Because mathematical models can help us understand the population dynamics of interacting species, we will also discuss parasite–host models. Finally, we will consider how parasites have evolved to increase their chances of infecting hosts, and how hosts have evolved to combat the risk of infection.
#### 15.1 Many Different Types of Parasites Affect the Abundance of Host Species
15.1 Many different types of parasites affect the abundance of host species Parasites typically have specific habitat needs and, as a result, often live in particular places on a host organism. In humans, for instance, parasites find certain parts of the body to be highly suitable habitats. As illustrated in Figure 15.1, head lice insects live in the hair; liver flukes, which are flatworms, reside in the liver; the fungus that causes athlete’s foot resides in the feet; and so on.
Figure 15.1 Preferred habitats. Parasites have preferred habitats on their host. The human body, for example, offers a wide range of habitats for parasites.
We can categorize the wide variety of parasites common to both plants and animals as either ectoparasites or endoparasites. Ectoparasites live on the outside of organisms, whereas endoparasites live inside organisms. Each lifestyle has advantages and disadvantages, as summarized in Table 15.1. For example, because ectoparasites live on the outside of their hosts, they do not have to combat the immune system of a host, and they can easily move on and off a host. A disadvantage is that ectoparasites are exposed to the variable conditions of the external environment, including natural enemies, and they must find a way to pierce the flesh of their host to feed. In contrast, because endoparasites live inside their hosts, they must contend with the immune system of their host, and they may have a difficult time getting in and out of their host’s body. Endoparasites have the advantage of being protected from the external environment and are therefore not exposed to most of their enemies. In addition, living inside the host gives endoparasites easy access to the host’s body fluids on which they feed. In this section, we will discuss some of the most common types of ectoparasites and endoparasites and the effects they have on their hosts. Ectoparasite A parasite that lives on the outside of an organism. Endoparasite A parasite that lives inside an organism.
TABLE 15.1 Comparing the Consequences of Endoparasite and Ectoparasite Lifestyles Factor Ectoparasites Endoparasites Exposure to natural enemies High Low Exposure to external environment High Low Difficulty of movement to and from host, for parasite or its offspring Low High Exposure to host’s immune system Low High Ease of feeding on host Low High
Ectoparasites
A variety of organisms live as ectoparasites, as shown in Figure 15.2. Most ectoparasites that attack animals are arthropods, including two groups of arachnids—ticks and mites—and two groups of insects—lice and fleas. There

are many other groups of ectoparasites, including leeches and some species of lamprey fish. Each of these animals attaches to a host and consumes the host’s blood and other body fluids.
Figure 15.2 Ectoparasites. Ectoparasites are parasites that live attached to the outside of their host. Common ectoparasites include ticks, fleas, lice, mites, mistletoes, and nematodes. Plants also have ectoparasites, but they are most commonly either nematodes—also known as roundworms—or other species of plants. Nematodes that act as plant ectoparasites live in the soil and feed on plant roots. The 1-mm-long worms attach to a plant root, inject digestive enzymes that break down root cells, and then consume the resulting slurry. This parasitic behavior can reduce the growth, reproduction, and survival of the plant. Approximately 4,000 species of plants make their living as ectoparasites on other plants. For example, mistletoes embed rootlike organs into the branches of trees and shrubs. Because such plants have their roots inside the host and their shoots outside the host, they are sometimes categorized as hemiparasites. The mistletoe leaves conduct photosynthesis but obtain water and minerals from the host plants. Some species, such as dwarf mistletoes (in the genus Arceuthobium), cause death in conifer trees—particularly under drought conditions—by extracting too much of the host’s water and nutrients. In some species of mistletoe, the plant’s fruit is consumed by birds, which
inadvertently disperse the seeds from the fruit when they defecate onto the branches of other trees and shrubs. Other species of mistletoe are able to project seeds laterally up to 15 m.
Endoparasites
Endoparasites can be categorized as either intracellular or intercellular. As the names imply, intracellular parasites live inside the cells of a host, whereas intercellular parasites live in the spaces between cells that include the cavities of a host’s body. Intracellular parasites are very small; examples include viruses, small bits of proteins known as prions, and some types of bacteria and protists. Intercellular parasites are much larger; examples include some types of protozoa and bacteria, fungi, and a group of worms known as helminths, which include the nematodes. Each of these groups of endoparasites can have substantial effects on their host’s survival, growth, and reproduction. Because endoparasites often cause fatal diseases, they can alter the abundance of host species and change the composition of ecological communities. Viruses Late in the nineteenth century, researchers struggled to find the cause of a disease striking tobacco plants. They recognized that the pathogen was too small to be a bacterium. After many years of research, they were able to identify it as a virus, which they named tobacco mosaic virus. Plants that are infected exhibit a mosaic of colors in their leaves and may also have growths that resemble blisters (Figure 15.3). Although this virus was first discovered in tobacco, we now know that it can infect more than 150 species of plants. Viruses that infect plants can have a devastating effect on food production, which makes them a matter of great concern to farmers and consumers.

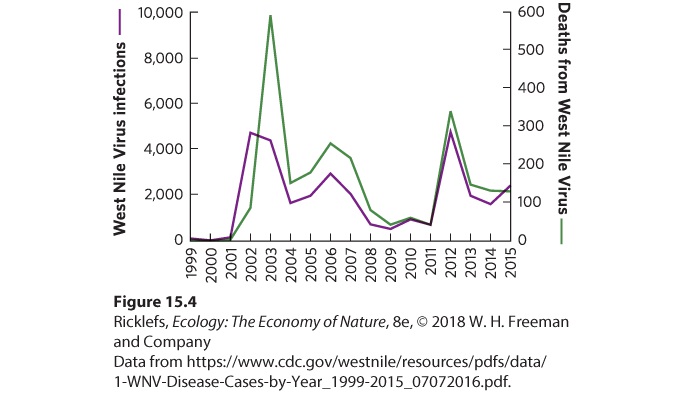
Figure 15.3 Tobacco mosaic virus. Plants infected with the tobacco mosaic virus, such as this tobacco plant, develop blisters and light areas on their leaves. These effects can cause substantial reductions in plant growth. Animals can also be infected by many different pathogenic viruses. For example, there are different genera of the poxvirus that can infect mammals: cowpox in cattle, monkeypox in primates and rodents, and smallpox in humans. Hoofed animals such as deer, cattle, and sheep are susceptible to bluetongue virus, which can cause high mortality rates. Similarly, the West Nile virus—named for the location of the first known human case in the West Nile region of Uganda in 1937—can be highly lethal to some species of birds. The mosquitoes that carry this virus usually transmit the disease to birds, but occasionally to horses and humans, both of which can die from the infection. West Nile virus arrived in the United States in 1999 and spread rapidly. Several species of North American birds experienced high rates of death, including blue jays (Cyanocitta cristata), American robins, and American crows (Corvus brachyrhynchos). At the same time, horses and people started to become infected. In the decade following the introduction of the virus into the United States, more than 1,000 people died. As a result, public health officials in the United States have made great efforts to reduce mosquito populations that spread the disease. These efforts, possibly combined with the evolution of the virus to
be less virulent, have caused the number of infections and deaths to decline dramatically from 2003 to 2011, as illustrated in Figure 15.4. However, in 2012, there was a sharp increase in the number of infections and deaths in the United States. The sharp increase is thought to be due to unusually warm and humid conditions combined with abundant rain, which created a lot of standing water where mosquitoes can breed and subsequently build up a large adult population. In subsequent years, the number of infections and deaths has declined.
Figure 15.4 Human infections and deaths from West Nile virus in the United States. Following the introduction of the virus to the United States in 1999, the number of human infections and deaths rose rapidly through 2003. Efforts to control mosquito populations, which carry the virus, caused a sharp decline in these effects on humans through 2011. In 2012, however, unusually warm temperatures with high precipitation in some regions of the country resulted in large mosquito populations and a new surge in West Nile infections and deaths. After 2012, infections and deaths declined substantially. Note that the two y axes have different scales. Both mammals and birds are susceptible to strains of influenza virus. For example, a strain of influenza known as the Spanish flu is caused by the H1N1 virus and normally infects only birds, but in 1918 it also infected humans. Researchers hypothesize that the virus experienced a mutation that allowed it to survive in humans and to infect other humans directly. They believe that humans initially contracted the disease by handling domesticated ducks and chickens. However, once the virus found its way into humans, it rapidly spread around the world and killed
up to 100 million people. More recently, the H1N1 virus has been found on pig farms, which is why the pathogen also has the name swine flu. In 2006, a similar strain of virus known as bird flu, or H5N1, infected and killed large numbers of birds and also jumped from domesticated birds to humans. Hundreds of thousands of domesticated birds were killed in an attempt to stop the spread of the virus (Figure 15.5). By 2017, the World Health Organization reported that 856 people had been infected with bird flu and more than half of them had died. The ability of pathogens to jump from traditional hosts to human hosts, as we will discuss in the next section, causes a great deal of concern for human health in the future.
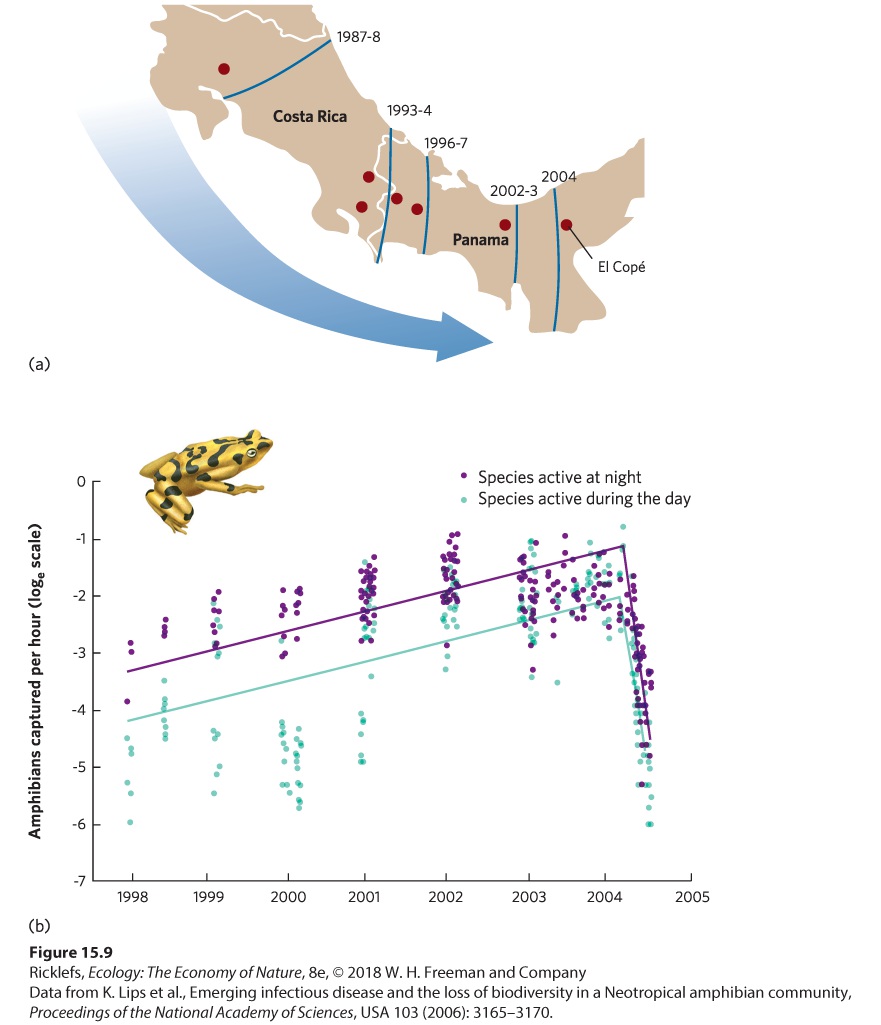
Figure 15.5 Controlling the spread of bird flu. In an attempt to stop the spread of bird flu, millions of domesticated birds in Asia were killed and their bodies burned. Prions Prions are a more recently discovered category of pathogenic parasite. All prions begin as a beneficial protein in the brain of an animal, but occasionally a protein folds into an incorrect shape and becomes pathogenic. Prions do not contain any RNA or DNA; instead, they replicate by coming into contact with normal proteins and causing the normal proteins to fold incorrectly, with the first prion serving as a template. As the number of prions increases in the body, they can kill cells and damage tissues.
One of the best-known diseases caused by prions is bovine spong encephalopathy, commonly known as mad cow disease. This disease, which is always fatal, gets its name from the way that infected cows (and sheep) lose control of their body, as if they have gone mad. While cows cannot transmit prions to each other, in the 1980s it was a common practice to feed cows the ground remains of other cows. Cows that consumed the mutated prions from the dead cattle became infected and ultimately passed on the infection to humans. Mad cow disease was most prevalent in the United Kingdom during the 1990s, where more than 180,000 cases of infected animals have occurred. Around the world, more than 220 people have died from mad cow disease. Today, new rules forbid feeding dead cattle and sheep to other cattle and sheep. As a result, the incidence of infected cattle and sheep is now rare, and mad cow disease in cows, sheep, and humans has declined. Chronic wasting disease is a more common prion disease. It infects members of the deer family, including white-tailed deer, mule deer (Odocoileus hemionus), elk, and moose. Individuals infected with chronic wasting disease excrete prions that are later consumed inadvertently by others. Infected individuals begin to lose weight and eventually die (Figure 15.6). Currently, chronic wasting disease is concentrated in Colorado and Wyoming, but small numbers of infected deer have been discovered in other regions of the United States and Canada.
Figure 15.6 Chronic wasting disease. Deer infected with prions that cause chronic wasting disease lose weight and eventually die. Protozoans Protozoans are a group of parasites that can cause a variety of diseases. For example, several protozoans cause diarrhea in humans and other animals. They are also the source of malaria in humans and avian malaria in birds. Malaria is transmitted by mosquitoes that acquire the protists when they feed on an infected individual and then transfer the protists to uninfected individuals they subsequently feed on. In Hawaii, as well as in some other parts of the world, there is no historic presence of avian malaria, so the birds of the Hawaiian Islands have not evolved defenses against the protists. Mosquitoes were accidentally introduced into Hawaii in the early 1800s, and by the early 1900s the protists that cause avian malaria had also arrived, which has contributed to the decline and extinction of many native bird species in these islands. Bacteria Bacterial parasites can cause a wide variety of plant and animal diseases. In plants, bacterial infections can cause spotted leaves, wilted stems, scabbed
surfaces, and large abnormal tissue growths known as galls (Figure 15.7). Because bacteria must enter a plant through a wound in the plant’s tissues, they commonly require the assistance of herbivores that pierce a plant’s tissues.
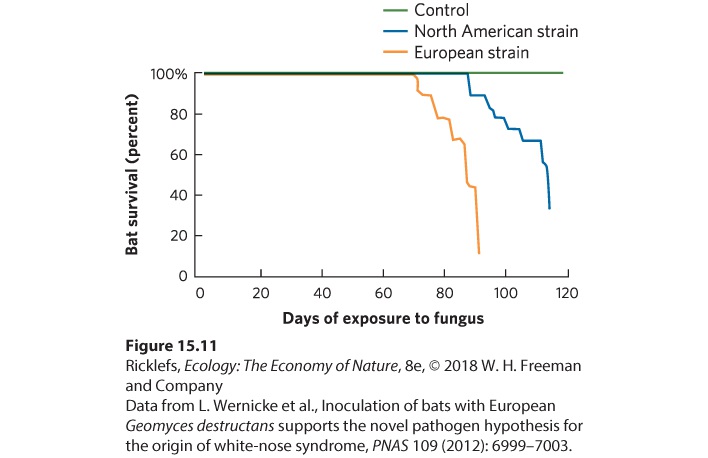
Figure 15.7 Bacterial disease of plants. Common plant diseases that are caused by bacteria include (a) shot-hole such as this Pseudomonas syringae mors-prunorum on a cherry tree and (b) Crown gall (Rhizobium radiobacter) shown on a birch tree in England. Bacterial infection in animals can also be quite harmful to the host. One of the earliest discovered species of deadly bacteria was anthrax (Bacillus anthracis). Anthrax, originally isolated from cows and sheep, is also highly lethal to humans. After an incubation period of about 1 week, the growing bacterial population begins to release toxic compounds that cause internal bleeding, which often brings rapid death. Other common bacterial pathogens include those that cause plague, pneumonia, salmonella, leprosy, and many sexually transmitted diseases, all of which occur in a wide variety of animals. Fungi Fungal parasites have large ecological impacts on a wide range of plants and animals. Fungal diseases have devastated many dominant plant species, including critical food sources for humans. A number of tree species in North America have suffered tremendous declines as a result of pathogenic fungi that have been introduced from Europe and Asia. The American chestnut tree was once one of the tallest tree species in the temperate forests of the eastern United States and composed up to 50 percent of all trees in the forest. It was also one of the most prized trees for lumber as well as for the crop of edible nuts that it
produced every autumn. Around 1900, however, species of Asian chestnut trees that were imported to New York inadvertently carried a fungus (Cryphonectria parasitica). While Asian chestnut trees had a long evolutionary history with the fungus and were resistant to its harmful effects, American chestnut trees had no such history. As a result, the American chestnut rapidly succumbed to the fungal disease, known as chestnut blight. The decline of the chestnut trees also likely had widespread effects on the many animal species that relied on the tree’s large crop of nuts for food. Today, the American chestnut tree is quite rare; young seedlings can emerge from seeds, but they ultimately become infected with the fungus and die early in life. In response, researchers are currently breeding more resistant varieties that they hope can someday be planted throughout forests in the eastern United States. Similar fungal disease problems have occurred with other trees, including the American elm tree (Ulmus americana). The elm was widely planted along North American streets because of its attractive arching branches that provided shade (Figure 15.8a). In the 1930s, a fungus from Asia was introduced into Europe and North America that caused a rapid decline in elm trees. Because the fungal disease was first described by researchers in Holland, it is known as Dutch elm disease. The American elm had no evolutionary history with the Asian fungus, and 95 percent of the elm trees across North America have succumbed (Figure 15.8b). The small number of remaining trees appears to be resistant to the fungal disease, and researchers are currently working to plant these resistant genotypes back into eastern forests.
Figure 15.8 Declining tree populations in North America. The introduction of a fungus that causes Dutch elm disease killed many American elms, which were once common along North
American streets. (a) A street in Detroit, Michigan, in 1974 before Dutch elm disease. (b) The same street in 1981 after Dutch Elm disease killed all the elm trees. Some fungal parasites also damage crop plants. One of the best-known examples is a group of fungi that causes a disease known as rust. Throughout human history, rust diseases have affected many of our most important crops, including wheat, corn, rice, coffee, and apples. The effect of rust can range from reduced food production to a complete loss of the crop, costing millions of dollars of damage. Widespread rust infections, such as wheat rust, have caused the starvation of millions of people and domesticated animals over the past two centuries. Animals can also be infected by fungi. One high-profile animal fungal disease is caused by a species of chytrid fungus (Batrachochytrium dendrobatidis) that infects amphibians, such as frogs and salamanders. During the 1990s, scientists working in Central America began to notice massive die-offs of amphibians and subsequently determined that the dead animals were infected with the fungus. The fungus, which lives in the outer layers of the amphibian’s skin, causes an imbalance of ions in the body that ultimately causes the animal’s heart to stop. For 2 decades, researchers surveyed amphibians from Costa Rica to Panama and found that the fungus was moving from the northwest to the southeast, as shown in Figure 15.9a. Between 2000 and July 2004, at a site in El Copé, Panama, more than 1,500 amphibians were sampled before the fungus arrived; not one individual tested positive. By October 2004, however, 21 of 27 sampled species were infected in at least 10 percent of their populations and by December 2004, 40 species tested positive for the fungus.

Figure 15.9 The deadly chytrid fungus in Central America. (a) Surveys of amphibians in Central America demonstrated that the fungus was spreading from the northwest to the southeast. (b) Surveys of amphibians at the El Copé site detected a sharp decline in amphibians, both in species that are active during the day and species that are active at night. The sizes of amphibian populations in El Copé were also estimated, using transect surveys of amphibians that were active either during the day or during the night. After the fungus arrived in 2004, the number of live amphibians declined sharply (Figure 15.9b). The dead amphibians included 38 different frog species. Moreover, 99 percent of the 318 dead individuals collected had moderate to severe chytrid infections. In 2010, researchers reported that of the 63 species of amphibians present at the site before the fungus arrived, 30 species were gone. As of 2016, many of these species have not been seen since chytird arrived. Relatively little is known about this
fungus, although the massive die-offs suggest that amphibians have few defenses against it, which indicates that the fungus may have been introduced to Central America. The fungus is now suspected to have caused the extinction of dozens of species of amphibians around the world. Helminths Helminths include several groups of roundworms and flatworms that can cause serious diseases. We have already discussed the impact of nematodes as ectoparasites on plant roots. Nematodes can also live as endoparasites within the tissues of plants, where they damage the plant’s growth, reproduction, and survival. In animals, disease-causing helminths include hookworms that feed on the blood of intestines, lungworms that live in the lungs, and echinostome worms that live in the kidneys. In livestock, infection by liver flukes has been a problem for centuries. Livestock inadvertently consume these parasites when they drink water containing fluke-infected snails or when they eat grass that contains a stage of the parasite that is excreted by snails. Liver flukes are a particular problem in sheep, where they may cause liver damage, hemorrhaging, and sudden death. Left untreated, liver flukes can kill up to 10 percent of a sheep population; fortunately, drugs can cure infected animals.
Emerging Infectious Diseases
Many infectious diseases have been infecting hosts for thousands of years, while others have only emerged recently. When a new disease is discovered, or a formerly common disease that declined in the past suddenly becomes common again, it is called an emerging infectious disease. New diseases typically emerge when a mutation allows a pathogen to jump to a new host species. We have already mentioned several emerging infectious diseases, including the chytrid fungus that has decimated amphibians around the world; the H5N1 bird flu that jumped from birds to humans; and mad cow disease that infected cattle, sheep, and humans. Since the 1970s, the world has experienced an average of one new emerging infectious disease each year. Emerging infectious disease A disease that is newly discovered or has been rare and then suddenly increases in occurrence.
In 2006, an emerging disease infecting bats in a cave near Albany, New York, was identified. These bats had a white-colored fungus (Geomyces destructans) growing on their noses, and they were dying in large numbers (Figure 15.10). Researchers named the disease white-nose syndrome. They hypothesized that the fungus causes bats hibernating in caves to come out of their normal torpor and to use up their fat reserves until they become emaciated and die. By 2016, the fungus had spread to 29 states and 5 Canadian provinces, and killed more than 5 million bats. In caves containing infected bats, the mortality rate can reach 100 percent. This is important because bats provide many important services, including the consumption of a large number of insects. Therefore, it is plausible that the death of so many bats will have substantial effects on many communities and ecosystems.
Figure 15.10 White-nose fungus in bats. This species, known as the little brown bat, is infected by the fungus Geomyces destructans. One symptom of the infection is that the bat’s nose turns white. When a bat colony becomes infected, a large proportion of the bats die. An interesting detail in the story of white-nose syndrome is that bats in Europe also carry the fungus but do not experience widespread death. This raises the possibility that the fungus is native to Europe and has only recently been introduced to North America. Alternatively, it could be that the fungus
has always been in North America but recently mutated to become much more lethal. To test these alternative hypotheses, researchers isolated strains of the fungus from European and North American bats. They then exposed the little brown bat (Myotis lucifugus), a species from North America, to fungal spores from the European strain, fungal spores from the North American strain, or a control treatment containing no fungal spores. After the exposures, they monitored bat survival for 120 days. In 2012, the results were reported, which you can see in Figure 15.11. In the control treatment, no bats died, but bats exposed to either fungal strain experienced high rates of death, which suggests that the fungus is a recent arrival from Europe and not a mutated North American native. Moreover, the bats died more slowly when exposed to the North American strain of fungus than the European strain, which may suggest that North American bats are already beginning to evolve some level of resistance to their local strain of fungus. Unfortunately, there is currently no treatment to prevent the continued death of bats from white-nose syndrome.
Figure 15.11 Testing North American versus European strains of white-nose fungus. Bats exposed to the control treatment did not die. In contrast, bats exposed to the North American fungus strain or the European fungus strain died in large numbers. However, bats died faster when exposed to the European strain than the American strain.
Concept Check
1. What are the advantages and disadvantages of being an ectoparasite? 2. What are the major groups of ectoparasites? 3. What are the major groups of endoparasites?
#### 15.2 Parasite and Host Dynamics Are Determined by the Parasite’s Ability to Infect the Host
15.2 Parasite and host dynamics are determined by the parasite’s ability to infect the host Parasites and their hosts have populations that fluctuate over time, much like the dynamics of predator and prey populations that we discussed in Chapter 14. Unlike predators, however, parasites generally have a higher reproductive rate than their hosts and often do not kill their hosts. If we know what causes fluctuations in parasite and host populations, we can predict when parasitism will be prevalent in a host population. This allows us to anticipate population changes in hosts and, in some cases, to intervene and reduce the harmful effects of a parasite on a species of concern. The probability that a host will become infected by a parasite depends on numerous factors that include the parasite’s mechanism of transmission, its mode of entry into a host’s body, its ability to jump between species, the existence of reservoir species, and the response of the host’s immune system.
Mechanisms of Parasite Transmission
The first factor that determines the risk of infection by a parasite is the mechanism of transmission. As illustrated in Figure 15.12, there are many different mechanisms. Parasites can move between hosts through horizontal transmission or vertical transmission. Horizontal transmission occurs when a parasite moves between individuals other than parents to their offspring. For example, horizontal transmission occurs when helminths are transmitted from snails to frogs or from frogs to birds. Horizontal transmission can also occur between conspecifics, such as the transmission of bird flu from one bird to another. Some parasites, such as the prions that cause mad cow disease, cannot be naturally transmitted from one individual to another. The risk of transmission is essentially zero in these cases. As we have seen with mad cow disease, dead infected cows were fed to other cows, which significantly raised the risk of horizontal transmission. At the other extreme of transmission are influenza viruses, which pass easily between individuals. You may have experienced the rapid transmission of the flu virus if you or someone in your family became ill with flu; most likely, other members of your family quickly became infected. For most hosts, the risk of becoming infected by a parasite generally increases with host population density
because higher densities mean that individuals are likely to come into contact more often with the parasites or with infected individuals.
Figure 15.12 Mechanisms of parasite transmission. Parasites can be transmitted vertically or horizontally. When transmission is horizontal, the parasite can be transmitted through a vector such as a mosquito, transmitted directly between two conspecifics, or transmitted to other species. When transmission is vertical, a parent host transmits the parasite to its offspring, such as when a mother bird transmits lice to its hatchlings at the nest. Horizontal transmission When a parasite moves between individuals other than parents and their offspring. Some parasites require another organism, known as a vector, to disperse from one host to another. For example, the West Nile virus is transmitted from one host to another by a mosquito that picks up the virus from one infected bird and transmits it to an uninfected bird or other animal that it bites. In this case, the mosquito is the vector. Vector An organism that a parasite uses to disperse from one host to another. Still other parasites require horizontal transmission to multiple host species to complete their life cycle, as we saw in the story of the amber snail. Some species of helminths, for example, spend the first stage of their life in a snail, the second stage of their life in an amphibian, and the final stage of their life in a bird. This requirement for multiple hosts poses a substantial challenge for the parasite, which must find all its required hosts during its lifetime.
Vertical transmission occurs when a parasite is transmitted from a parent to its offspring. In this case, the parasite must evolve in such a way that it does not cause the death of its host until after the host has reproduced and passed the parasite to its offspring. Many sexually transmitted diseases can be passed by vertical transmission. For example, chlamydia is a disease caused by several different species of bacteria within two genera, Chlamydia and Chlamydophila, that infect mammals, birds, and reptiles. In humans, chlamydia causes inflammation of the urethra and cervix but is typically not lethal. While the bacterium is often horizontally transmitted between individuals, it can also be transmitted vertically from an infected mother to her fetus. When this happens in humans, the newborn baby can suffer eye infections and pneumonia. Vertical transmission When a parasite is transmitted from a parent to its offspring. Researchers continue to learn about the wide range of pathways that parasites can take to infect a host. The existence of vertical transmission and various methods of horizontal transmission can make it very challenging to predict and control the spread of infectious diseases in humans, crops, domesticated animals, and wild organisms.
Modes of Entering the Host
The mode of entry into the host’s body also affects the ability of the parasite to infect the host. As we have discussed, some species of parasites, such as leeches, are able to pierce the tissues of the host. Other parasites, such as some viruses, bacteria, and protists, rely on another organism to penetrate the host’s tissues and use the damaged tissue as an entry point. The protist that causes avian malaria, for example, depends on mosquitoes to inject the protist into a bird’s body after the protist completes a critical life stage inside the mosquito’s body. Without the mosquito, the malaria life cycle could not be completed. Of course, another mode of entering a host occurs when the pathogen is inadvertently consumed, as we saw in the case of ducks eating infected snails in Chapter 9.
Jumping between Species
If a parasite specializes on only one host species and is able to cause a lethal disease in only that host species, then it might eventually run out of hosts and face extinction. One solution sometimes favored by natural selection is for the parasite to be nonlethal to the host; this allows a host population to persist. We will talk more about this strategy later in the chapter. Alternatively, the parasite might evolve the ability to infect other species. We saw an example of this with bird flu, which infected several species of birds before a mutation occurred that allowed the virus to infect humans. A similar scenario occurred with HIV. For many years, it was hypothesized that human HIV originated in chimpanzees, and in 2006 researchers identified a population of chimps in the West African nation of Cameroon that carried a genetically similar strain of the virus. The researchers suspect that the virus jumped from chimpanzees to humans when local hunters consumed the chimpanzees as food. Other examples of parasites jumping between species include the chytrid fungus, which can jump between amphibian species, and the canine parvovirus, which can jump from cats to dogs.
Reservoir Species
One way that parasite populations persist in nature is through the use of reservoir species. Reservoir species carry a parasite but do not succumb to the disease that the parasite causes in other species. Because reservoir species do not die from the infection, they serve as a continuous source of parasites as other susceptible host species become rare. For example, some species of birds can be infected with the protist that causes avian malaria, but they are resistant to developing the disease. However, mosquitoes that feed on these resistant species of birds can pick up the protist and transfer it to other species of susceptible birds that become infected and die. In some species, such as the Hawaiian honeycreepers (Figure 15.13), the individuals that are infected by avian malaria but survive through acquired immunity can become a reservoir for the pathogen. In this way, reservoir species and immune individuals favor the persistence of the parasite population over time.
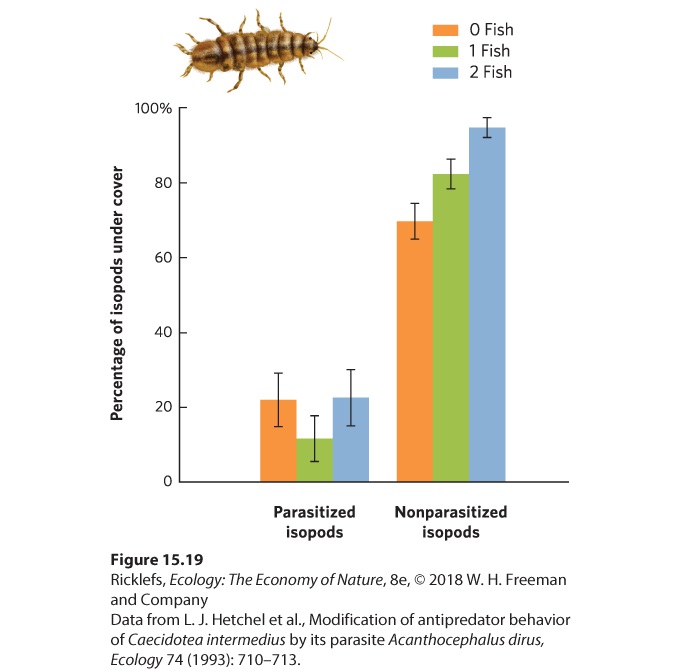
Figure 15.13 Reservoir species. Many species of birds such as this ‘Apapane (Himatione sanguinea, a species of Hawaiian honeycreeper) can survive infection and subsequently serve as a reservoir of avian malaria that can be spread to other birds through the bites of mosquitoes. Reservoir species Species that can carry a parasite but do not succumb to the disease that the parasite causes in other species. THE HOST’S IMMUNE SYSTEM A host’s immune system can play a role in combatting an infection from endoparasites. As a result, some parasites have evolved the ability to escape the immune system by making themselves undetectable. For example, when HIV enters a human cell, it can hide from the body’s immune system by living in the cytoplasm or by incorporating itself into the chromosomes of the cell. Because the body’s immune system searches for infections on the outside of cells, it cannot detect the virus. Other parasites have evolved additional strategies to escape the immune system. For instance, parasitic worms known as schistosomes produce a protective outer layer around their bodies that prevents them from being detected by the host’s immune system. Still others, such as the protists that cause African sleeping sickness, are able to continually change the compounds present on their outer surface so they become a moving target that stays one step ahead of the immune system as it tries to respond to the infection.
1. What are the different ways in which parasites can be transmitted between hosts? 2. What is the process that allows a parasite to jump to a new species? 3. How does a reservoir species help a parasite population persist over time?
#### 15.3 Parasite and Host Populations Commonly Fluctuate in Regular Cycles
15.3 Parasite and host populations commonly fluctuate in regular cycles In our discussion of predators and prey in Chapter 14, we saw that population fluctuations are common and sometimes occur in regular cycles. Because parasites and hosts represent consumers and resources, they show similar population dynamics. In this section, we will examine how parasites and hosts fluctuate over time. We will also look at a mathematical model that helps us describe and understand the behavior of infectious pathogens and hosts.
Population Fluctuations in Nature
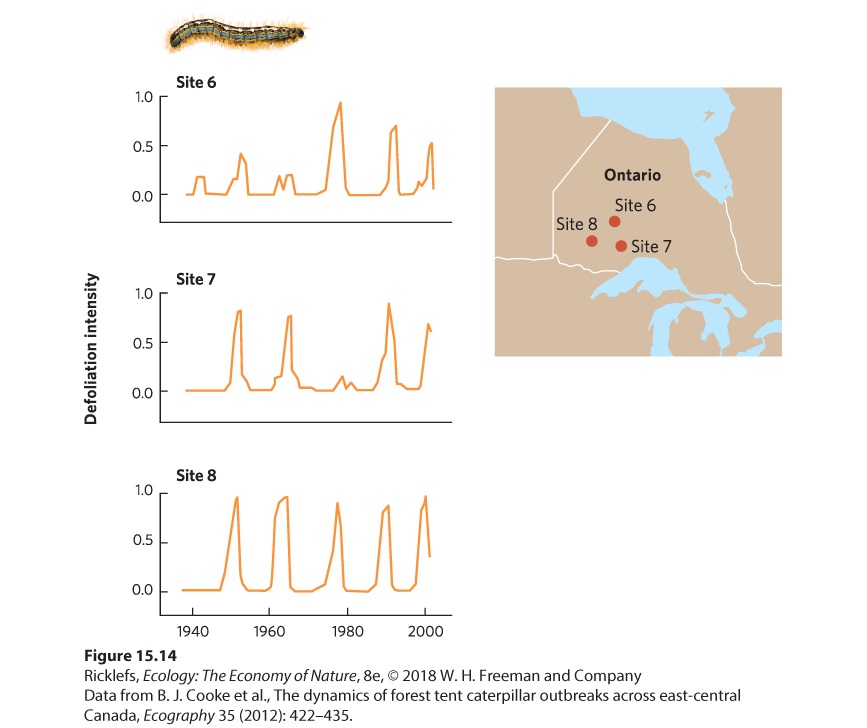
As we discussed in the previous chapter, the density of the host population can affect how easily parasites are transmitted from one host to another. An excellent example of this can be seen in the dynamic between the forest tent caterpillar (Malacosoma disstria)—a major herbivore of broadleaf trees in the United States and Canada—and a group of viruses that can infect and kill the caterpillar. The forest tent caterpillar can defoliate trees over thousands of square kilometers. In years of high caterpillar densities, they remove the majority of a tree’s leaves and tree growth is reduced up to 90 percent. Canadian researchers reported that caterpillar populations had cycles that last 10 to 15 years and that these fluctuations were fairly synchronous across large geographic areas. In three different locations in the province of Ontario, the fluctuations in caterpillar populations exhibit the same pattern of growth and decline over time, as shown in Figure 15.14. Although the caterpillar is susceptible to predators and parasitoids, viruses have the greatest ability to reduce its abundance. When the tent caterpillar population is high, the virus can more easily spread from one host to another, so each caterpillar is more likely to get the virus. Under these conditions, large numbers of caterpillars die. As host population density decreases, it becomes harder for the viruses to find a new host and the prevalence of the disease declines. Since fewer caterpillars become infected and die, the caterpillar population starts to increase again. These patterns are similar to the predator‒prey population cycles that we discussed in the previous chapter.

Figure 15.14 Cyclic population fluctuations of forest tent caterpillars. At three sites in the province of Ontario, researchers quantified the population sizes of tent caterpillars by measuring the intensity of leaf removal on the trees. Over a 60-year period, the caterpillars exhibited large population fluctuations every 10 to 15 years. The rapidly growing populations ultimately succumb to an outbreak of infection by a virus and the populations quickly die back. Fluctuations in parasite and host populations can also be caused by changes in the proportion of the host population that has achieved immunity. When a host species is able to become immune to a parasite after an initial infection, continued infection in the population causes an increasing proportion of the population to develop immunity. When a high proportion of the population is immune to the parasite, the spread of the parasite is slowed. Measles, for example, is a highly contagious viral disease that stimulates lifelong immunity in humans. In unvaccinated populations, measles typically produces epidemics at 2-year intervals. Once most of the population is infected and develops immunity, the number of new measles cases declines sharply. As humans continue to reproduce, however, there are enough new children born without immunity to initiate another measles outbreak after 2 years. The number of measles cases in London, England, from 1944 to 1968 —before vaccinations became available—shows a distinct pattern of 2-year
cycles, as you can see in Figure 15.15. Once vaccinations became available and a higher proportion of susceptible individuals were immunized, the number of new cases dropped sharply and the fluctuations declined and eventually disappeared.
Figure 15.15 The cyclical occurrence of measles in a human population. Before vaccinations became available in 1968, the human population in London, England, experienced cycles of measles every 2 years. Once vaccination became available and the number of people vaccinated increased, the number of measles cases declined and the fluctuations no longer occurred.
Modeling Parasite and Host Populations
For pathogenic parasites, we can understand the dynamics of the parasite and the host using models that are similar to the Lotka‒Volterra predator–prey model. However, the parasite–host model differs from the predator–prey model in two key ways. Parasites, unlike predators, do not always remove host individuals from a population, and hosts may develop immune responses that make some individuals resistant to the pathogen. The simplest model of infectious disease transmission that incorporates immunity is the Susceptible-Infected-Resistant (S-I-R) model. In this model, all individuals begin as susceptible to the pathogen (S). Of those, some number becomes infected (I). Of the infected individuals, some number develops resistance via immunity (R). We can use this model to examine the conditions that favor an epidemic versus the conditions that cause a disease to decline. The proportions of S, I, and R individuals in a population are determined by rates of transmission of the disease and acquisition of immunity, as well as the birth of new, susceptible individuals. Susceptible-Infected-Resistant (S-I-R) model The simplest model of infectious disease transmission that incorporates immunity. In a population of hosts, the first individual to be infected by a pathogen is

known as the primary case of the disease. Any individuals infected from this first individual are known as secondary cases. The rate at which the pathogen spreads through the population depends on two opposing factors. One factor is the rate of transmission between individuals (b), which includes both the rate of contact of susceptible individuals with an infectious individual and the probability of infection when there is contact. The other factor is the rate of recovery (g), which determines the period of time from when an individual is infected and can transmit the infection to when the individual’s immune system clears the infection and the individual becomes resistant to any future infections. The rate of infection and the rate of recovery can be used to determine whether an infectious disease will spread through a population. A disease will spread whenever the number of newly infected individuals is greater than the number of recovered individuals. To determine the number of newly infected individuals, we need to know the probability that an infected individual and a susceptible individual will come into contact with each other. If we assume that individuals in the population move around randomly, the probability of contact is the product of their proportions in the population: Probability of contact between susceptible and infected individuals=S×I Once infected and susceptible individuals come into contact, we also have to consider the rate of infection between them (b): Rate of infection between susceptible and infected individuals=S×I×b Next we have to determine how many individuals are recovering from the infection. We can do this by knowing the proportion of infected individuals (I) and the rate of recovery from an infection (g): Rate of recovery of infected individuals=I×g Now we can determine whether an infection will spread through a population. To do this, we need to calculate the reproductive ratio of the infection (R0), which is the number of secondary cases produced by a primary case during its period of infectiousness. The reproductive ratio of the infection is the ratio of new infections to recoveries:
R0=(S×I×b)÷(I×g)R0=S×(b÷g) If R0 > 1, the infection will continue to spread through the population and an epidemic will occur. This happens because each infected individual infects more than one other individual before it recovers from the disease and becomes resistant. When R0 < 1, the infection fails to take hold in the host population. This happens because each infected individual fails to infect another individual, on average, before it recovers and becomes resistant.
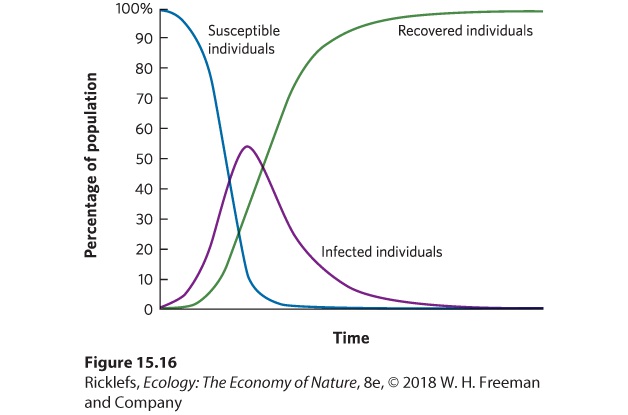
Figure 15.16 illustrates the dynamics of a typical disease. Imagine that we start with a population that is composed entirely of uninfected individuals. When an infectious disease arrives, there is initially a rapid increase in the number of infected individuals but, over time, infected individuals subsequently recover and become resistant (R). At this point, the number of susceptible individuals (S) decreases, so the value of R0 decreases. When R0 declines to <1, the growing epidemic can no longer sustain itself. As a result, the number of infected individuals achieves a peak in abundance and then declines.
Figure 15.16 The dynamics of an infection over time. In the basic S-I-R model, all individuals in a population are initially susceptible. When the infection is introduced at the beginning of the time period, there is an initial rapid growth in the number of infected individuals. As some infected individuals recover and become resistant, there are fewer susceptible individuals left to infect, so the number of infected individuals declines.
With this understanding of how R0 affects the spread of diseases, we can now look at the R0 values for different diseases that are caused by parasites. HIV is transmitted through rather limited mechanisms, including direct sexual contact, blood transfusion, or perinatal transmission from mother to offspring. HIV has a relatively low range of R0 values, from 2 to 5. Typical values for R0 in childhood diseases of humans—measles, chicken pox, and mumps, among others—range from 5 to 18 at the time a population is initially infected. At the extreme, malaria, which is transmitted by mosquitoes, has an R0 value greater than 100 in crowded human populations. This high value occurs because mosquitoes are excellent vectors for transmitting the parasite and infected people remain infectious for long periods of time. Assumptions of the S-I-R Model The basic S-I-R model has several important assumptions. For example, it assumes there are no births of new susceptible individuals and that individuals retain any resistance they develop. In such a model, the result is an epidemic that runs its course until all individuals in the population have become resistant or there are too few susceptible individuals remaining to sustain the spread of the disease. Some pathogens, such as influenza viruses, fit these assumptions. The basic model also explains why vaccinations slow or stop the spread of diseases such as influenza; by vaccinating individuals, we reduce the size of the susceptible population (S), which reduces the value of R0. This makes it harder for an epidemic to sustain itself. Other factors can be added to the model, including births of susceptible infants, lag times between when an individual is infected and becomes infectious to others, host mortality, host population dynamics, and transmission of disease from parent to offspring. These additional factors can have substantial effects on the predictions of the model. For example, the birth of new susceptible individuals can cause cyclic fluctuations in the model, as we observed in the case of the measles data from London (Figure 15.15). If a pathogen can kill the host, the pathogen should increase in abundance until the hosts begin to die. As the host population declines, the pathogen population will subsequently decline, and as the pathogen population
declines, the host population should subsequently recover. This is analogous to the predator–prey cycles we discussed in Chapter 14. However, some pathogens do not follow these dynamics because they do not attack a single host species. For instance, the chytrid fungus that we discussed earlier in this chapter infects dozens of species of amphibians. A host species can decline all the way to extinction, yet the fungus does not decline in abundance because it can infect other species of amphibians. Some of these newly infected species will become ill and die, while others never develop the disease and serve as reservoirs for the disease. When a pathogen is not restricted to a single host species, it has the ability to persist and spread even after it causes one of its hosts to go extinct.
Concept Check
1. How do lethal parasites cause host populations to cycle? 2. What is the effect of host immunity of parasite populations? 3. Why do we want to know the reproductive ratio of a parasite?
#### 15.4 Parasites Have Evolved Offensive Strategies, While Hosts Have Evolved Defensive Strategies
15.4 Parasites have evolved offensive strategies, while hosts have evolved defensive strategies Parasites gain fitness when they find a suitable host and reproduce. Hosts will experience higher fitness if they avoid being parasitized. Therefore, natural selection has favored the evolution of parasite offenses and host defenses. In this section, we will examine some of these strategies as well as look at evolutionary relationships between parasites and hosts.
Parasite Adaptations
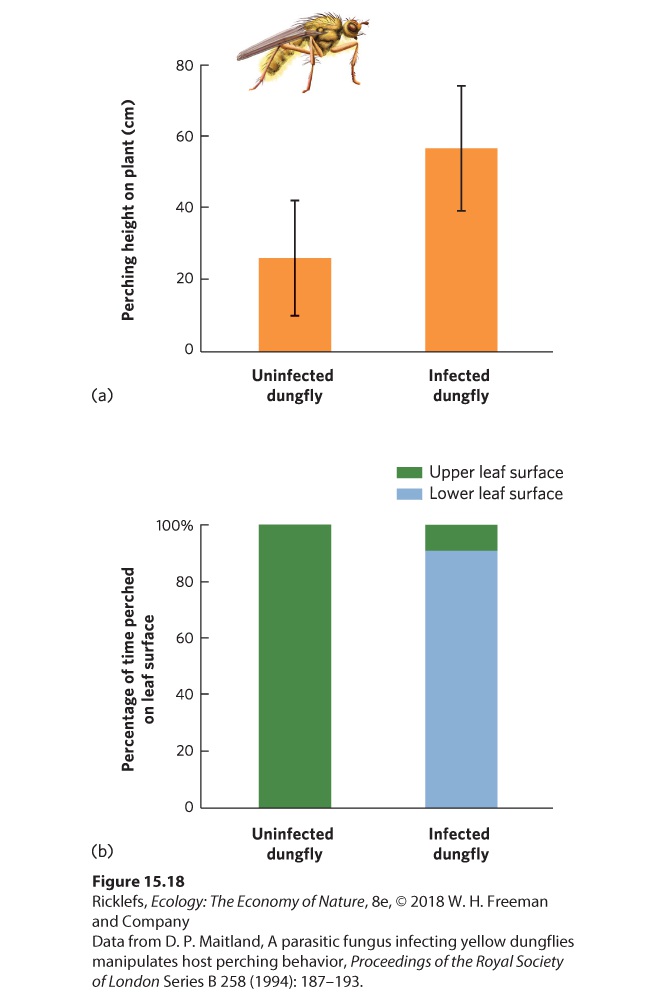
As we saw at the beginning of the chapter, parasites have evolved a wide array of strategies for finding and infecting hosts. Some of the most fascinating examples of parasite adaptation involve strategies that increase the probability of parasite transmission. For example, the pathogenic fungus Entomorpha muscae infects both houseflies (Musca domestica) and yellow dungflies (Scatophaga stercoraria). To improve its chance of transmitting its spores, it has two different strategies. When the fungus infects and kills female houseflies, the bodies become attractive to male houseflies looking for a mate. Although it is unclear what attractive cues the fungus produces, when a male housefly tries to mate with a dead female, spores of the fungus are transferred to the male’s body. When the same fungus infects the yellow dungfly, the fly climbs up nearby vegetation, moves to the upwind side of the plant, and hangs upside down on the underside of a leaf. The fly then moves its wings toward the leaf and moves its abdomen—which is swollen with fungal spores—away from the leaf. Once it reaches this position, the dungfly dies and the fungal spores erupt from its abdomen. The erupting spores can then potentially infect dungflies that pass below, as illustrated in Figure 15.17. When researchers compared the behavior of infected dungflies to uninfected dungflies, they found that uninfected flies perched lower on plants and never perched on the underside. You can view these data in Figure 15.18.
Figure 15.17 Transmission of fungal spores in the yellow dungfly. Infected flies climb up a plant, position themselves upside down on the underside of a leaf, and die as the spores erupt from their abdomen. In contrast, uninfected flies position themselves lower on plants and sit upright on the top sides of leaves.
Figure 15.18 Fungal control of the perching behavior in the yellow dungfly. (a) Compared to uninfected dungflies, infected dungflies perch higher on plants. Error bars are 1 standard deviation. (b) Uninfected dungflies never perch on the underside of a leaf, whereas infected flies do so 91 percent of the time. For parasites that require a series of different hosts to complete their life stages, it is challenging to find a way to get transmitted from one host to another. Some parasites—such as trematodes that first infect snails and
subsequently infect tadpoles—have evolved a simple strategy of leaving the body of the first host and then searching for the second host. Other parasites have evolved ways of manipulating the behavior of the first host to ensure that the second host consumes it. We saw an example of this with the amber snail discussed at the beginning of the chapter. There are many examples of parasites taking control of the host’s behavior. For example, mice can inadvertently consume a parasitic protist, Toxoplasma gondii, when feeding near cat feces. Once the mouse becomes infected, it no longer avoids an area marked by the smell of bobcat (Lynx rufus) urine but, instead, is mildly attracted to it. As a result, the mouse is more likely to be eaten by the bobcat, which is a valuable outcome for the protist since it can only reproduce in the gut of a cat. Similarly, small crustaceans known as isopods (Caecidotea intermedius) normally hide in refuges from predatory fish. However, if the isopods are infected by a parasitic worm (Acanthocephalus dirus), they spend less time in the refuge and more time out in the open where the fish are more likely to notice them, as you can see in Figure 15.19. This manipulation of the isopod’s behavior is beneficial to the parasite because fish serve as the parasite’s second host.
Figure 15.19 Parasite-induced changes in isopod behavior. When isopods are not infected by parasites, they generally stay under cover whether there are fish present or not. When they are infected, the isopods move out from under the cover, which makes them susceptible to predatory fish. Error bars are standard errors.
Host Adaptations
Host species have evolved a range of mechanical and biochemical defenses to combat parasite invaders. Some species of plants and animals can produce antibacterial and antifungal chemicals that kill bacterial and fungal parasites. For example, many species of amphibians naturally release antimicrobial peptides onto their skin, which inhibit the growth of the deadly chytrid fungus. In some cases, host species have evolved both mechanical and biochemical defenses to combat parasites. We see this with chimpanzees living in Tanzania that become infected with intestinal worms. Instead of eating their normal food diet, chimps that are ill with the intestinal worms pick a few leaves from plants in the genus Aspilia and swallow them whole (Figure
##### Analyzing Ecology: Comparing Two Groups with a t-Test
15.20). The leaves of these plants are covered in tiny hooks and, as the leaves pass through the chimp’s digestive system, the hooks pull nematode parasites (Oesophagostomum stephanostomum) out of the digestive tract, so they can be expelled from the chimp’s body with feces. Infected chimps also chew on bitter twigs from the Vernonia plant, something that healthy chimps do not do. The sick chimps become well within 24 hours because the twigs contain chemicals that kill a variety of different parasites. These plants are also consumed by people in the region when they experience symptoms of parasite infections. In short, both chimps and humans use plants to medicate themselves against parasitic infections.
Figure 15.20 Self-medicating chimpanzees. When chimpanzees become ill from intestinal parasites, they swallow whole leaves and chew bitter twigs of plants not normally in their diet. These leaves and twigs reduce the number of parasites in their body and help them recover.
Analyzing Ecology
Comparing Two Groups with a t-Test When the dungfly researchers examined the perching height of infected and uninfected flies,
they concluded that the infected flies perched significantly higher on vegetation. How did they come to that conclusion? They compared the mean heights for the infected and uninfected flies and used a statistical test to determine if the two means were different. In Chapter 14, we discussed how scientists typically consider two means to be different if we can sample their distributions many times and find that the means of those sampled distributions overlap less than 5 percent of the time. We commonly call this critical cutoff alpha— abbreviated as α. Therefore, we say that our critical cutoff is α = 0.05. To determine if the means of the two groups are significantly different, we need to know three things from each group: the mean, the variance, and the sample size. With these three parameters, we can calculate a t-test, which determines if the distributions of data from two groups are significantly different. An important assumption of the t-test is that the values from both groups are normally distributed. t-test A statistical test that determines if the distributions of data from two groups are significantly different. We begin by calculating the difference between the means of the two groups: X¯1-X¯2 where X¯1 is the mean of group 1 and X¯2 is the mean of group 2. Next we calculate the standard error of the difference in the two means: S12n1+S22n2 where S21 is the sample variance of group 1, S22 is the sample variance of group 2, n1 is the sample size of group 1, and n2 is the sample size of group 2 (you can review the concept of sample variances in Chapter 2).
The next step is to divide the first calculated value by the second calculated value: t=X¯1-X¯2S12n1+S22n2 As you can see from this equation, the value of t becomes larger as the difference between the means becomes larger, or the standard error of the difference in the means becomes smaller. For example, consider two groups of data with the following parameters: Group Mean Sample variance Sample size In this case, t=20-10105+105=102+2=5 The last thing we need to calculate is known as the degrees of freedom, which is defined as the sum of the two sample sizes minus 2. In this example, the degrees of freedom is 5+5−2=8 With this value of t calculated, we can then determine if our value exceeds a critical level of t and therefore whether the means are significantly different. We can find the critical value of t from a statistical table. If you look at the t-table in the “Statistical Tables” section at the back of this book, you can find the column that uses an alpha value of 0.05 and then find the row that contains 8 degrees of freedom. The critical t-value is the number found in this row and column. In this case, the critical t-value is 2.3. Because our calculated tvalue exceeds the critical t-value, we can conclude that the two means differ significantly.
YOUR TURN For the following data, calculate the t-value. Group Mean Sample variance Sample size Next calculate the degrees of freedom. Based on your calculated t-value and the critical t-value using α = 0.05, determine if the two groups are significantly different.
Coevolution
When we consider the evolutionary forces that affect adaptations of parasites and hosts, we often find that as one species in the interaction adapts, the other species responds by adapting as well. As we discussed in Chapter 14, when two or more species continue to evolve in response to each other’s evolution, we call it coevolution. When all species involved in an interaction coevolve together, no one species is likely to get an upper hand. For example, when a parasite evolves an advantage that makes it more effective at infecting hosts, the hosts will then be under stronger selection to evolve defenses against that parasite. We can see an example of coevolution between parasites and hosts in the dynamics of the rabbit population in Australia. In the 1800s, European rabbits were introduced to many regions of the world where they had not lived historically. In Australia, for instance, rabbits were first introduced from England in 1859. Within a few years, local ranchers were erecting fences to keep them out and organizing shooting parties to help control the size of the population. Eventually, hundreds of millions of rabbits ranged throughout most of the continent, destroying sheep pasturelands and thereby threatening wool production (Figure 15.21). The Australian government tried poisons, predators, and other control measures, all without success.
Figure 15.21 A growing rabbit population in Australia. After being introduced to Australia in 1859, the rabbit population exploded and consumed nearly all the vegetation that was needed by ranchers for raising sheep. In 1950, the government tried a new approach. A virus known as Myxoma had been discovered in South American rabbits. The virus—which was carried by mosquitoes—causes a disease known as myxomatosis. The disease had minor effects on South American rabbits because they had coevolved with the virus for a long time, but it turned out to be highly lethal to European rabbits; an infected rabbit dies within 48 hours. The Australian government introduced the virus and initially it had a devastating effect on the rabbits. Among infected rabbits, 99.8 percent died and the rabbit population declined to very low numbers. However, 0.2 percent of the infected rabbits proved to be resistant and passed on their resistant genes. Before the introduction of the Myxoma virus, a few rabbits possessed resistant genes, but they had never been favored. In a second outbreak of the disease, only 90 percent of infected rabbits died. By the third outbreak, only 40 to 60 percent of the rabbits died and the rabbit population of Australia started to increase. You can see the decline in rabbit mortality over time in Figure 15.22.
Figure 15.22 Coevolution of Australian rabbits and the Myxoma virus. In rabbit populations that had never been exposed to the virus, nearly all infected rabbits died. However, a small percentage of resistant rabbits survived and passed on their resistant genes to the next generation. At the same time, selection favored less lethal virus strains, which could reproduce and get transmitted by mosquitoes that only feed on live rabbits. Since rabbit populations evolved higher resistance and viruses evolved to be nonlethal, rabbit populations that experienced more epidemics of the disease experienced less mortality. The decline in rabbit mortality was due partly to the evolution of increased resistance in the rabbit population, but because the rabbit population was initially killed in large numbers, the rapidly dwindling host population also favored any virus strains that infected the rabbits without killing them. Dead rabbits represent a dead end for the virus, especially if the rabbit dies before it is bitten by a mosquito, which is the only way the virus can be transmitted. Over time, the interaction between the rabbits and the Myxoma virus resulted in a more resistant host population and a less lethal pathogen. This allowed the rabbit population to rebound and the virus to persist. This dynamic is probably what researchers observed in South America, where the rabbit population and Myxoma virus persisted together for a long time. To meet the Australian government’s goal of controlling the rabbit population in Australia, scientists continue to introduce new strains of the virus from South America that are highly lethal to Australian rabbits because the new strains have no history of coevolution with the Australian rabbits. In this way, they maintain the effectiveness of the Myxoma virus as a pest control agent.
Concept Check
1. What is an example of a parasite adaptation that helps promote its transmission from one host to another? 2. What is an example of a host adaptation that helps the host combat parasites? 3. What is the process of coevolution of parasite and host adaptations?
Concepts
Of Mice And Men . . . And Lyme Disease The black-legged tick. The tick, also known as a deer tick, is the
vector of the bacteria that cause Lyme disease in humans. Back in the 1970s, a number of children in a Connecticut community experienced a suite of mysterious symptoms. Many had a bull’s-eye– shaped rash on their skin followed by flulike symptoms that developed into other symptoms similar to arthritis and various neurological disorders. Physicians discovered that the children had been infected by a pathogenic bacterium (Borrelia burgdorferi). Because the community was in the town of Lyme, Connecticut, the illness came to be known as Lyme disease. The parasite that causes Lyme disease is carried by ticks that feed on wild animals. Occasionally, these ticks latch onto people and transmit the parasite. In North America, the primary vector is the black-legged tick (Ixodes scapularis), also known as the deer tick. The ticks and the parasite probably existed in North America for thousands of years before the disease was identified. According to the Centers for Disease Control and Prevention, currently 20,000 to 30,000 cases of Lyme disease occur every year in the United States. Most cases occur in the Northeast, although there is a growing number of cases in the Midwest. Over 4 decades, ecologists and medical researchers have determined that to understand Lyme disease requires a knowledge of the ecological community in which the parasite lives. First, they had to determine how ticks obtain the parasite. They found that 99 percent of newly hatched ticks do not carry the parasite, so vertical transmission is not likely. Instead, the tick obtains the parasite from infected hosts. But because the black-legged tick can live
on a wide variety of hosts, the next step was to determine which hosts affected the abundance of infected ticks. The cycle of Lyme disease. A diverse array of species affects the prevalence of the bacteria that causes Lyme disease. The cycle begins when ticks attach themselves to infected rodents and become infected with the bacteria. After they drop from the rodents, they molt into nymphs and spend the winter under leaves. In the spring, the nymphs attach themselves to hosts again and then drop and molt into adults. As adults, most ticks attach themselves to deer, search for mates, and then lay eggs. During years of high acorn production by oak trees, there is an abundance of food for rodents that produces large rodent populations. Large rodent populations lead to large tick populations that can infect a variety of animals, including deer and humans. By examining patterns of abundance in nature and conducting manipulative experiments, the researchers were able to work out the life cycle of the black-legged
ticks. The ticks are predominantly creatures of the forest. Larval ticks hatch from eggs each summer, but since they are unable to climb very high on vegetation, they can only infect small animals living close to the ground, such as birds and rodents. The most commonly infected rodents are chipmunks (Tamias striatus) and white-footed mice (Peromyscus leucopis). These two species can be infected without exhibiting any signs of disease, so they act as reservoir species that maintain the bacterial population. After feeding on the rodents for a few days, the ticks drop off their host and molt into nymphs. As nymphs, the ticks spend the winter living under the leaves on the forest floor. The following spring, they climb a short distance up in the vegetation and wait for another host to pass by. They most commonly attach to birds, small mammals, and any humans who might be taking a walk in the woods or in a garden. After attaching themselves to a host and becoming engorged with blood, the ticks drop off again and molt into adults by autumn. Adult ticks can climb higher up on vegetation, which allows them to jump onto larger mammals such as white-tailed deer. The deer’s body not only offers a blood meal but is also a place for adult male and female ticks to find each other and mate. From there, they drop off the deer and lay their eggs on the forest floor each autumn. All of this movement of ticks between multiple hosts results in many opportunities for the bacteria to be transmitted. In fact, researchers found that frequency of infection with the bacteria is 25 to 35 percent for nymphs and 50 to 70 percent for adults. Based on this research, it became clear that deer, mice, and chipmunks were keys to the tick’s life cycle. But what affects the abundances of these species? The researchers
discovered one more key player: oak trees. Oak trees produce a massive number of acorns every few years, and acorns are a major food item for deer, chipmunks, and mice. In high-acorn years, tick-carrying deer gather under the oak trees, which causes an aggregation of reproducing ticks that drop their eggs under the trees. Chipmunks and mice are also attracted to the acorns since the abundant food permits higher survival and increased reproduction. More rodents and an increased density of tick eggs cause an increase in the number of infected rodents the following summer. In low-acorn years, when both deer and rodents spend more time in maple forests, the prevalence of infected ticks shifts from the oak forests. To examine what steps can be taken to reduce the risk of Lyme disease to humans, the researchers also used mathematical models that included all the key species. These models suggested that, while reductions in deer densities would have little effect on the population of infected ticks unless the deer population was completely eradicated, reductions in the rodent population could cause a large reduction in the population of infected ticks. With the tremendous increase in cases of Lyme disease, researchers have looked back in time to determine how long the bacteria have been around. In 2012, the bacteria were found in the tissues of a frozen mummy that was discovered in the Alps between Austria and Italy. Given that the mummy lived 5,300 years ago, it suggests that Lyme disease has been infecting humans for thousands of years, despite the fact that physicians only recently identified the disease. The story of Lyme disease is an excellent example of how understanding the ecology of parasites allows us to
predict when and where they pose the greatest risk to wild organisms and to humans. SOURCES: Keller, A., et al. 2012. New insights into the Tyrolean Iceman’s origin and phenotype as inferred by whole-genome sequencing. Nature Communications 3: 698 Ostfeld, R. 1997. The ecology of Lyme-disease risk. American Scientist 85: 338–346. Ostfeld, R., et al. 2006. Climate, deer, rodents, and acorns as determinants of variation in Lyme-disease risk. PLOS Biology 4: 1058–1068.
Summary of Learning Objectives
15.1 Many different types of parasites affect the abundance of host species. Ectoparasites live on host organisms, whereas endoparasites live in host organisms. As a group, parasites include a wide range of species that include plants, fungi, protozoa, helminths, bacteria, viruses, and prions. Among parasites that cause diseases— known as pathogens—those that have recently become abundant are called emerging infectious diseases. Key Terms: Infection resistance, Infection tolerance, Parasite load, Ectoparasites, Endoparasites, Emerging infectious diseases
15.2 Parasite and host dynamics are determined by the parasite’s ability to infect the host. The transmission of parasites can be horizontal—either through direct transmission or transmission by a vector—or vertical from parent to offspring. The ability to infect a host also depends on the parasite’s mode of entering the host, its ability to infect reservoir species, its ability to jump to new host species, and its ability to avoid the host’s immune system. Key Terms: Horizontal transmission, Vector, Vertical transmission, Reservoir species
15.3 Parasite and host populations commonly fluctuate in regular cycles. These fluctuations occur because transmission increases with host density but decreases as an increased proportion of the host population develops immunity. These fluctuations can be modeled using the S-I-R model. Key Term: Susceptible-Infected-Resistant (S-I-R) model
15.4 Parasites have evolved offensive strategies, while hosts have evolved defensive strategies. Natural selection has favored
parasites that can improve their probability of transmission, including manipulations of host behavior. Hosts have evolved both specific and general immune responses to combat host infection. Hosts also can employ mechanical and biochemical defenses against parasites. Coevolution occurs when the parasite and host continually evolve in response to each other. Key Term: t-test
Critical Thinking Questions
1. Compare and contrast the advantages and disadvantages of life as an ectoparasite versus an endoparasite. 2. Why might parasites that are not very harmful to hosts in their native range be useful in controlling non-native hosts in an introduced range? 3. If a parasite has a reservoir host species, how effectively will the parasite population be controlled by immunizing a susceptible host species? 4. Given that there is currently no cure for mad cow disease, what is likely to be the most effective action to reduce its transmission? 5. Why might we continue to discover new emerging infectious diseases? 6. Compare and contrast horizontal versus vertical transmission of a parasite. 7. Using the basic S-I-R model of parasite and host dynamics, explain why the proportion of infected individuals in the population declines over time. 8. In the S-I-R model of parasite and host dynamics, how does the outcome change if we allow new susceptible individuals to be born into the population? 9. When using a t-test, what factors make it more likely that you will find a significan difference? 10. Explain why Dutch elm disease might become less lethal to its host over time.
Graphing the Data Time Series Data
As we saw in “Ecology Today: Applying the Concepts” at the end of this chapter, the number of black-legged ticks infected by the bacterium that causes Lyme disease is determined by the number of rodents, which in turn is affected by the production of acorns. In making these connections, researchers knew the abundance of the different species over time. Using the data provided in the table, plot the density of acorns, chipmunks, and ticks over time using a line graph. Based on your graph, what patterns do you see when changes in acorn density affect chipmunk density and when changes in chipmunk density affect tick density? Densities of acorns, chipmunks, and ticks over time Year Acorns/m2 Chipmunks/2.25-ha grid Ticks/100 m2