16 Competition
16Competition Garlic mustard is an invasive species that has spread throughout the eastern and midwestern United States. The invasive plant has a competitive advantage because it produces a toxin that harms native plants. Trying to Catch Up to Garlic Mustard Garlic mustard is a powerful forest foe. It was introduced to the United States from Europe 150 years ago and has since spread throughout eastern and midwestern forests. It not only spreads, it dominates as a competitor against native plant species. Where you find garlic mustard, you often find few other forest herbs. During the past decade, researchers have determined that the plant is a dominant competitor because it possesses a novel weapon. As mentioned in Chapter 1, many species of native plants depend on mutualistic relationships with soil fungi to obtain minerals. Without the fungi, these plants cannot grow well. The novel weapon of garlic

mustard is its production of sinigrin, a chemical that it releases into the soil from its roots and that is toxic to soil fungi. Because garlic mustard does not depend on soil fungi for its own growth, it has an immediate advantage over many native plant species. While sinigrin production helps garlic mustard survive and grow, researchers hypothesized that sinigrin production is costly. They hypothesized further that in forests where garlic mustard has been living for decades and has outcompeted most other plants, natural selection should favor reduced sinigrin production because at that point the cost comes with no benefit. To test this hypothesis, researchers collected seeds from locations where garlic mustard had recently invaded and locations where it has been present for up to 50 years. In total, they raised seeds from 44 different locations and discovered that populations with a longer history of being present at a given location produced less of the toxic chemical. Moreover, they saved the soil from each of these 44 populations and then raised three species of native trees in the saved soils. All three tree species grew better in the soils that had contained older populations of garlic mustard than soils with younger populations of garlic mustard. This confirmed that the garlic mustard was evolving to be less toxic to the soil fungi and, in turn, a less dominant competitor. The researchers also noticed that native plants were starting to become more abundant in forests with the oldest invasions of garlic mustard, which is consistent with the evolution of reduced sinigrin production. However, it is also possible that the native plants were simultaneously evolving a higher tolerance to competition from garlic mustard. To test this hypothesis, they collected a native forest herb from six forests that differed in their soil sinigrin concentrations. They raised the herb populations in the soil from which each had come, and then determined how well each herb population was able to compete against garlic mustard. They discovered that herb populations that derived from high-sinigrin soils were better able to compete against garlic mustard, which demonstrates that the native plants were evolving to tolerate the invader. After a decade of experiments, it has become clear that garlic
mustard is a highly effective competitor because it possesses a novel weapon. However, its competitive advantage continues to evolve as it experiences changes in the intensity of competition from native plants, while the native plants continue to coevolve to combat the invader. “Garlic mustard is a highly effective competitor because it possesses a novel weapon.” SOURCES: Evans, J. A., et al. 2016. Soil-mediated eco-evolutionary feedbacks in the invasive plant Alliaria petiolata. Functional Ecology 30: 1053–1061. Lankau, R. A. 2012. Coevolution between invasive and native plants driven by chemical competition and soil biota. PNAS 109: 11240–11245. Lankau, R. A., et al. 2009. Evolutionary limits ameliorate the negative impacts of an invasive plant. PNAS 106: 15362–15367.
Learning Objectives
After reading this chapter, you should be able to:
16.1 Illustrate that competition occurs when individuals experience limited resources.
16.2 Explain the theory of competition as an extension of logistic growth models.
16.3 Describe how the outcome of competition can be altered by abiotic conditions, disturbances, and interactions with other species.
16.4 Distinguish between exploitation competition, interference competition, and apparent competition. In Chapter 1, we defined competition as a negative interaction between two
species that depend on the same limiting resource to survive, grow, and reproduce. Competition can help determine where a species can live in nature and how abundant a population can become. It can occur among many groups of organisms, including predators, herbivores, and parasites. In this chapter, we will explore different types of competition and the resources for which species compete. We will then examine models of competition, which extend the Lotka‒Volterra models introduced in Chapter 14. Competition models allow us to predict the conditions that determine when a species will win a competitive interaction. Knowing how species compete for a resource is important, but we must also consider how other interactions might alter or even reverse the expected outcomes of competition, including abiotic effects, disturbances, and interactions with other species. At the end of the chapter, we will examine a variety of cases that appear to describe competition but actually reflect other processes, including predation and herbivory.
#### 16.1 Competition Occurs When Individuals Experience Limited Resources
16.1 Competition occurs when individuals experience limited resources When we study competition, we differentiate between intraspecific competition, which is competition among individuals of the same species, and interspecific competition, which is competition among individuals of different species. We considered intraspecific competition in our discussion of negative density dependence in Chapter 12, and saw that an increase in a population’s density causes a decline in the growth rate of the population. We have discussed interspecific competition much less. Interspecific competition can cause the population of either species to decline and eventually die out. Both intraspecific and interspecific competition play substantial roles in determining the distribution and abundance of species on Earth. Intraspecific competition Competition among individuals of the same species. Interspecific competition Competition among individuals of different species. In this section, we will examine how competition for a limited resource can cause one species to outcompete another. We will explore the wide variety of resources that are available to organisms, including resources that are fixed in abundance and those that are renewable. With our understanding of resources, we will then investigate the importance of the most limiting resource in a population and examine patterns of competition between closely and distantly related species.
The Role of Resources
A resource is anything an organism consumes or uses that causes an increase in the growth rate of a population when it becomes more available. For plants, resources generally include sunlight, water, and soil nutrients such as nitrogen and phosphorus. Each is used by most plants and plays a role in the growth of plant populations. Resources for animals generally include food, water, and space. For example, animals such as mussels and barnacles spend most of their lives attached to rocks in the intertidal zone along the ocean shores (Figure 16.1). Open space is a critical resource for them because as the

rocks become more crowded, there is less space to grow. With less space, the growth and fecundity of adult mussels and barnacles decline, and there are few places on the rocks for offspring to settle. Similarly, many birds may compete for a limited number of nest sites or cavities, and many prey species compete for a limited number of holes and crevices in which they can hide from predators. In each of these cases, when more space becomes available, the populations can increase. Resource Anything an organism consumes or uses that causes an increase in the growth rate of a population when it becomes more available.
Figure 16.1 Competition for space. Sessile organisms that live in the rocky intertidal biome compete for open space on the rocks as a point of attachment. This photo from Olympic National Park in Washington shows a number of sessile organisms at low tide, including mussels, giant green sea anemones (Anthopleura xanthogrammica), goose barnacles (Lepas anserifera), and ochre sea stars (Pisaster ochraceus). Ecological factors that cannot be consumed or used are not considered resources. Temperature, for instance, plays a major role in the growth and reproduction of organisms in nature, but temperature is not a resource because it is not consumed or used. The same is true for other abiotic factors, including pH and salinity. Renewable Versus Nonrenewable Resources We can categorize resources as either renewable or nonrenewable. Renewable resources are constantly regenerated. For example, rodents and
ants often compete for seeds, and every year new plants grow and renew the seed supply. Similarly, sunlight is continually generated by the Sun. In contrast, nonrenewable resources are not regenerated. For instance, space is a resource that typically has a fixed availability. If we think of the rocky intertidal habitat, there is a fixed number of rocks to which algae and animals can attach; space becomes available only when a competitor leaves or dies. Renewable resources Resources that are constantly regenerated. Nonrenewable resources Resources that are not regenerated. Renewable resources can originate from either inside or outside the ecosystem in which the competitors live. For example, dead leaves that fall into streams from the surrounding forest serve as food for stream insects. Because these resources come from outside the system, however, competition can reduce resource abundance but cannot affect the rate of the resource supply. Moreover, resources originating from outside the system do not respond to the rate of resource consumption. Sunlight continually strikes the surface of Earth regardless of the rate at which plants and algae consume it, and the amount of local precipitation is largely independent of the rate at which plants use water. Competitors can affect both the supply of and demand for resources that originate within the ecosystem. When competing herbivores or competing predators consume another species in the system, they reduce the supply of the species that they consumed. However, by reducing the abundance of a consumed species, the predators or herbivores also affect the future supply of that resource. For example, when rodents continue to consume seeds of abundant plants, the abundance of these plants declines over several years. In some cases, the supply rate of a renewable resource generated within an ecosystem is only indirectly affected by competitors. For example, plants take up nitrate from the soil and use it to grow. Competing herbivores that consume the plants return large amounts of nitrogen compounds back to the soil when they defecate and after death when their bodies decompose. These nitrogen compounds are further broken down by microorganisms, which release the nitrogen as nitrate, a form plants can use. The herbivores can
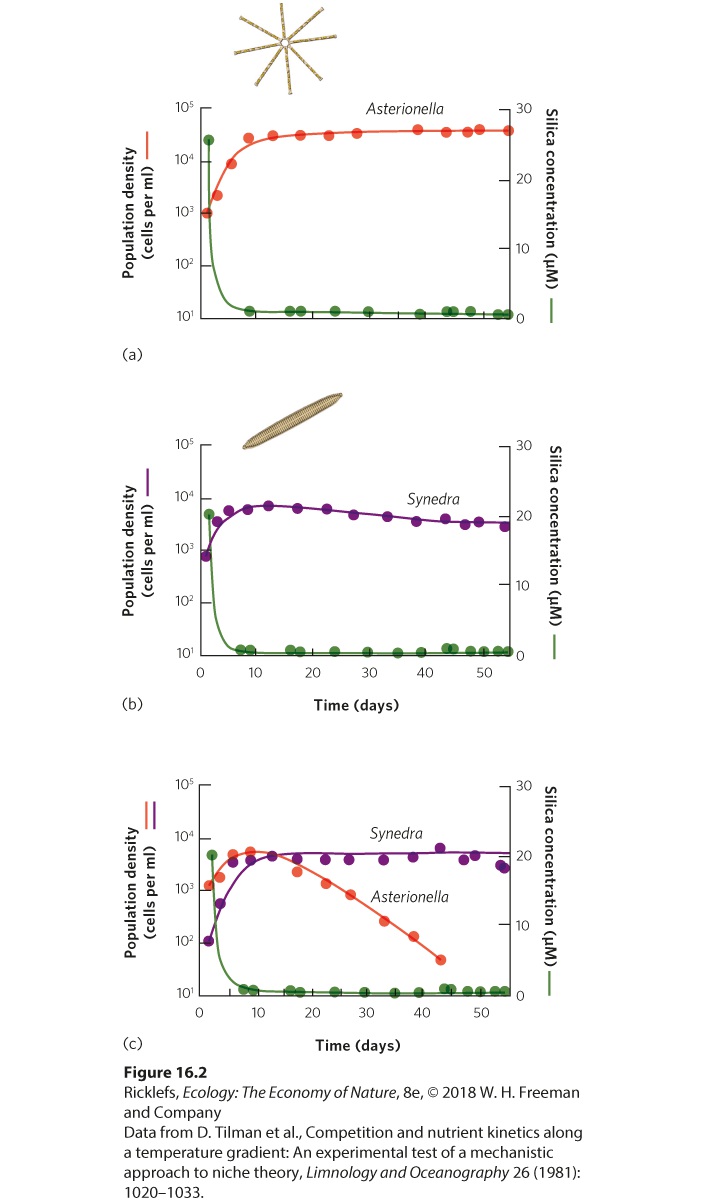
affect the supply of nitrate, but the chain of events has many links and takes such a long time that the herbivores do not have an immediate effect on the future plant population size through this indirect pathway. Leibig’s Law of the Minimum Although consumers can reduce the abundance of both renewable and nonrenewable resources, not all resources limit consumer populations. For example, all terrestrial animals require oxygen, but an increase in the population of an animal species does not depress the concentration of oxygen in the atmosphere to the point that population growth is limited. Well before the concentration of oxygen can become limiting, some other resource, such as food supply, will decline in abundance to the point that it limits the population’s growth. In 1840, Justus von Liebig, a German chemist, proposed that populations were limited by the single resource that was most scarce relative to demand. This idea is now known as Liebig’s law of the minimum, which states that a population increases until the supply of the most limiting resource prevents it from increasing further. Liebig’s law of the minimum Law stating that a population increases until the supply of the most limiting resource prevents it from increasing further. The amount of a resource that limits a population’s growth depends on the resource. For example, the microscopic, glass-shelled algae known as diatoms require both silicate (SiO2) and phosphate to grow and reproduce. When one diatom species, Cyclotella meneghiniana, is grown under different concentrations of each element, the population growth ceases whenever the concentration of silicate is reduced to 0.6 micromolar (μM) or whenever the concentration of phosphate is reduced below 0.2 μM. According to Liebig’s law of the minimum, whichever resource reaches its limiting value first will be the resource that regulates the growth of the diatom population. If we know the minimum amount of a resource that is required for populations to grow, we should be able to predict which species is the best competitor for the resource. An experiment with two species of diatoms, shown in Figure 16.2, demonstrates how competition for silicate reduces the
availability of silicate and affects the outcome of competition. When the two species are raised separately, Asterionella formosa and Synedra ulna both experience rapid population growth, followed by a plateau as they reach their carrying capacity, shown in Figures 16.2a and 16.2b. When Asterionella formosa reaches its carrying capacity, it drives down the abundance of silicate to 1 μM. However, when Synedra ulna reaches its carrying capacity, it drives down the abundance of silicate to 0.4 μM, which is not enough silicate to support the other diatom’s population. As a result, Synedra ulna should outcompete Asterionella formosa. When both species were put together, that is exactly what happened. As illustrated in Figure 16.2c, the two species drove the abundance of silicate down to a level that allowed Synedra ulna to persist but caused Asterionella formosa to decline to extinction. The outcome between these two diatoms is one that is commonly observed in nature: when two species compete for a single limiting resource, the species that persists is the one that can drive down the abundance to the lowest level.
Figure 16.2 Competition for the most limiting resource. When raised separately, (a) Asterionella and (b) Synedra diatoms grow rapidly and then persist with population sizes near their carrying capacity. At their carrying capacity, both species dramatically reduce the
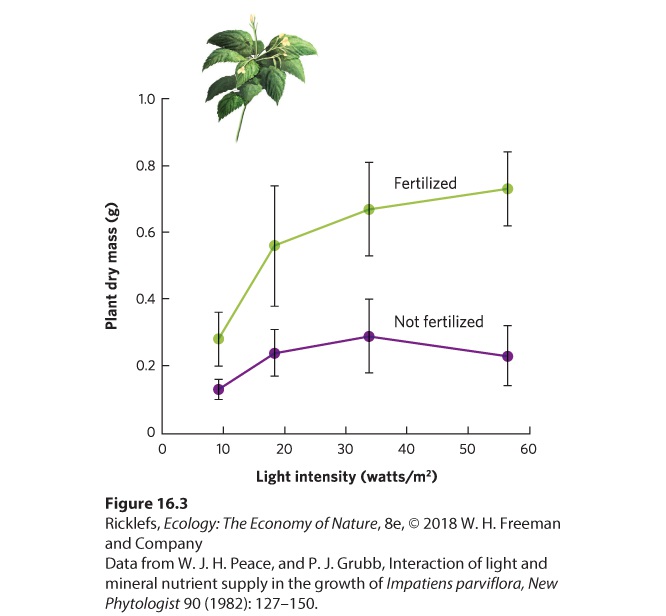
abundance of silicate, although Synedra causes the silicate to be driven down to a lower abundance than Asterionella. (c) When combined, the two species again drive down the abundance of silicate to the point that there is still enough to meet the minimum requirements of the Synedra population but not enough to meet the minimum requirements of the Asterionella population. As a result, the Asterionella population declines. Interactions Among Resources Leibig’s law of the minimum assumes that each resource has an independent effect on the growth of a population. In other words, it assumes that if a given resource limits the growth of individuals and populations, increasing the availability of other resources will not improve such growth. However, this is not always the case, as seen in research on small balsam (Impatiens parviflora), a plant that is common in English woodlands. Researchers wanted to know if increasing the abundance of one resource would cause the plant to become soon limited by a second resource. They sowed seeds of the plants in pots filled with soil. Control pots received only water, while fertilized pots received a solution containing both water and fertilizer. These two groups of pots were then grown under one of four different light intensities for 5 weeks. As you can see in Figure 16.3, plants grown in lownutrient soil experienced only a small increase in growth when the amount of light was increased. Similarly, plants grown at a low light intensity experienced a small growth increase when soil nutrients were increased. However, plants grown under high soil fertility and high light intensity experienced an increase in growth that was much larger than the sum of the separate effects of fertility and light intensity. This means that when the plants were given more nutrients, they soon became limited by light, whereas plants that were given more light soon became limited by nutrients.
Figure 16.3 Interactions among resources. The small balsam plant experienced little growth when light intensity was increased under low soil fertility. However, when light intensity and fertility were both increased, the large amount of growth that followed was much higher than the sum of the two separate effects. Error bars are 95 percent confidence intervals.
The Competitive Exclusion Principle
A classic study of competition between species was conducted by Russian biologist Georgyi Gause in the 1930s. As you may recall from Chapter 12, Gause conducted laboratory experiments that explored how populations of protists in the genus Paramecium grew when living alone or together with limited resources. He began by growing two species—P. aurelia and P. caudatum—separately in test tubes. The test tubes contained a fixed amount of food, which was a species of bacteria (Bacillus pyocyaneus). When grown separately, the population of each paramecium species initially experienced rapid growth and then began to plateau as it reached its carrying capacity. You can see these data in the first two graphs of Figure 16.4. However, when the two species were grown together in the test tube, Gause observed a different outcome. As you can see in Figure 16.4c, the population of P.
aurelia persisted in the test tube, but the population of P. caudatum declined to very low levels by the time the experiment ended. In short, P. aurelia was the superior competitor, presumably by driving down the abundance of the bacteria to such a low point that P. caudatum could not persist.
Figure 16.4 Competition between two species of protists. Gause raised different species of protists and measured the populations in terms of the total volume of cells in the container. When he raised each species separately on a fixed daily ration of bacteria, the population of (a) P. aurelia and (b) P. caudatum grew rapidly until they reached their carrying capacity. (c) When the two species were grown together, P. aurelia continued to grow well but P.
caudatum was reduced to a very low population density. Because P. caudatum was still present in low numbers, P. aurelia did not attain the same population size as when it was raised alone. Similar competition experiments have been conducted hundreds of times using a wide variety of organisms. These experiments frequently produce the same result: one species persists and the other dies out. This common pattern led to the development of the competitive exclusion principle, which states that two species cannot coexist indefinitely when they are both limited by the same resource. Researchers commonly find that when two species are limited by the same resource, one species either is better at obtaining the resource or is better able to survive when the resource is scarce. Competitive exclusion principle The principle that two species cannot coexist indefinitely when they are both limited by the same resource. Competition Among Closely Related Species Although competition can occur between closely and distantly related, organisms, Charles Darwin believed that competition is most intense between closely related species. He argued that closely related species possess very similar traits and likely consume similar resources. As a result, they have the potential to compete strongly. In consideration of Darwin’s prediction, naturalists observed that closely related species living in the same region often grow in different habitats within that region. They hypothesized that because these species would compete strongly for the same resources, natural selection would favor differences in habitat use. These differences would allow each species to have a competitive advantage within its preferred habitat and to have a competitive disadvantage in habitats preferred by closely related species. The first experimental test of this hypothesis was conducted in 1917 by the British botanist Arthur Tansley, who worked with two species of small perennial plants known as bedstraw. Heath bedstraw (Galium saxatile) typically lives on acidic soils, whereas white bedstraw (G. sylvestre) typically lives on alkaline soils. To determine whether each species competed best under the conditions where it naturally occurred, Tansley and his colleagues planted them separately and together in deep boxes containing either acidic or
alkaline soils, as illustrated in Figure 16.5. Because the boxes were located at a single site—the Botanic Garden in Cambridge, England—any differences in growth would be due to the types of soils in which the bedstraw was planted.
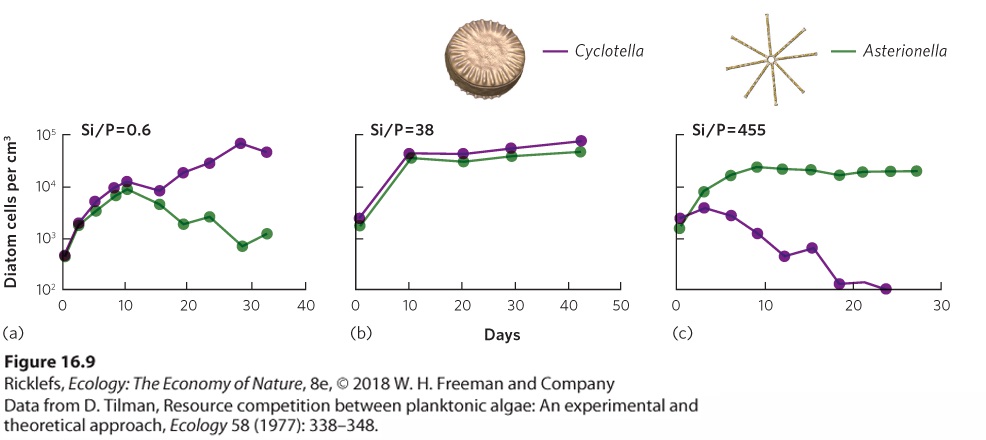
Figure 16.5 Tansley’s bedstraw experiment. White bedstraw naturally lives in alkaline soils and heath bedstraw naturally lives in acidic soils. Each species was planted either in its preferred soil or in the preferred soil of the other species; each species grew and survived best in its own preferred soil. This experiment showed that although closely related species can be intense competitors, the competition is reduced when competitors evolve to perform best in different habitats. When planted separately, each species germinated and grew in both types of soil. However, each species presented more vigorous germination and growth in the soil type characteristic of its natural habitat. When grown together on alkaline soils, the white bedstraw overgrew and shaded the heath
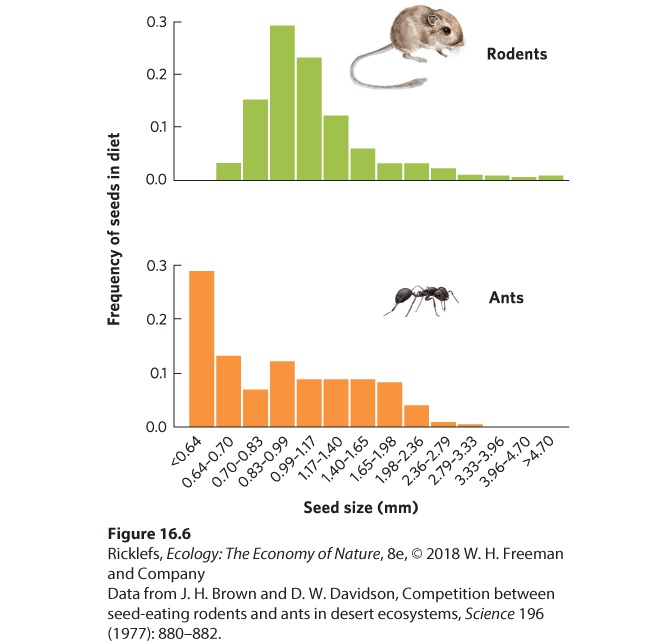
bedstraw. When grown together on acidic soils, heath bedstraw outcompeted the white bedstraw. Tansley concluded that although both species are capable of living in both soil types when they are grown separately, intense competition from a closely related species restricts their distribution in nature to the soil type that gives them a competitive advantage. Hundreds of studies have supported Tansley’s finding that many closely related species that live within the same region are intense competitors and are therefore distributed among different habitats in ways that reduce overlap, and thus competition, with each other. Competition Among Distantly Related Species While competition can be quite intense among closely related species, it can also be intense among distantly related species that consume a common resource. As we saw in Figure 16.1, open space on rocks in the intertidal biome is a resource used by numerous distantly related species, such as barnacles, mussels, algae, sponges, and others, all of which compete intensely for this limited space. As we will see later in this chapter, the outcome of this competition among so many competitors depends on their abilities to compete as well as their abilities to tolerate different abiotic conditions and predation. Another example of competition among different species occurs among animals that eat krill (Euphausia superba). Krill are shrimp-like crustaceans that live in the oceans surrounding Antarctica and are consumed by virtually every type of large marine animal, including fish, squid, penguins, seals, and whales. Commercial exploitation of whales in the Southern Hemisphere has caused a decline in whale populations, while penguin and seal populations have increased. This suggests that reducing the number of whales has eased competition for krill and allowed penguin and seal populations to grow. Intense competition can also occur among distantly related species in terrestrial ecosystems. On the forest floor, spiders, ground beetles, salamanders, and birds all consume the invertebrates that live in the leaf litter. In desert ecosystems, birds and lizards eat many of the same insect species. Moreover, desert ants and rodents both compete for seeds from a variety of plants. When we examine the patterns of seed sizes consumed by ants and rodents, as shown in Figure 16.6, we can see that the two groups both consume a wide range of seed sizes, but the ants tend to consume more of the small
seeds, whereas the rodents tend to consume more of the large seeds.
Figure 16.6 Seed size competition in rodents and ants. The two groups of herbivores in the deserts of Arizona consume seeds of similar size, although rodents prefer somewhat larger seeds and ants prefer somewhat smaller seeds. This overlap in seed consumption means that these two distantly related species can compete with each other.
Concept Check
1. What factors affect population growth but are not consumable resources? 2. Why might we expect closely related species to experience stronger competition with each other than would distantly related species? 3. What does the competitive exclusion principle predict about the outcome of two species competing for the same resource or resources?
#### 16.2 the Theory of Competition Is an Extension of the Logistic Growth Model
16.2 The theory of competition is an extension of the logistic growth model You may recall from Chapter 12 that population growth and regulation can be modeled using the logistic growth equation. Extensions of this equation can help us understand the population dynamics of two species that are competing with each other. We will begin by examining Lotka‒Volterra competition models when there is a single limiting resource and then expand our perspective and consider models that include multiple limiting resources. Although these models represent conditions that are a simplified version of what occurs in nature, they provide a useful foundation for exploring competitive interactions and the conditions under which species can coexist.
Competition for A Single Resource
To understand how the populations of two species compete for a single resource, let’s begin with the logistic growth equation introduced in Chapter 12: dNdt=rN(1-NK) As you may recall, the part of the equation in parentheses represents intraspecific competition for resources. As the size of the population, N, approaches the carrying capacity of the environment, K, the term in parentheses approaches zero. At this point, the population’s growth rate is zero, which means that the population has achieved a stable equilibrium. If we wish to use this equation to model competition between two species that compete for a single resource, we need to consider the carrying capacity of the environment relative to the number of individuals from both species that are being supported. For example, imagine an environment that has a carrying capacity of 100 rabbits. Let’s also assume that the food needed to support 100 rabbits could instead support 200 squirrels. The environment could also support many different combinations of rabbits and squirrels, such as 90 rabbits and 20 squirrels or 80 rabbits and 40 squirrels. To include a second species in our equation, we need to add two pieces of information: the number of individuals of the second species and how much

each individual of the second species affects the carrying capacity of the first species. For example, if we wanted to know the growth rate of a rabbit population, we would need to know the number of rabbits and squirrels in the area, and—in terms of resource consumption—how many squirrels equal one rabbit. Given that we want to model the changes in population for two species, we will use one equation for each. In these equations, we denote species 1 and 2 using the subscripts 1 and 2, respectively: dN1dt=r1N1(1-N1+αN2K1)dN2dt=r2N2(1-N2+βN1K2) The first equation tells us that the rate of change in the population of species 1 depends on its intrinsic rate of growth (r), the number of individuals of species 1 that are present (N1), and—in parentheses—the combined effects of species 1 and species 2 in consuming the resource relative to the carrying capacity. The second equation tells us the same thing for species 2. In these equations, we use two variables, the terms α and β, which are called competition coefficients. Competition coefficients are variables that convert between the number of individuals of one species and the number of individuals of the other species. In the preceding equations, α converts individuals of species 2 into the equivalent number of individuals of species 1. Similarly, β converts individuals of species 1 into the equivalent number of individuals of species 2. In our example of rabbits and squirrels competing for a common food resource, we can define rabbits as species 1 and squirrels as species 2. In that case, the value of α would be 0.5 because the resource consumption of one squirrel is equivalent to 0.5 rabbits. Similarly, the value of β would be 2.0 because the resource consumption of one rabbit is equivalent to two squirrels. Competition coefficients Variables that convert between the number of individuals of one species and the number of individuals of the other species. Using these two equations, we can create graphs that help us understand when each population will reach the point at which it no longer increases or decreases. This point is the population’s equilibrium point. We can start by
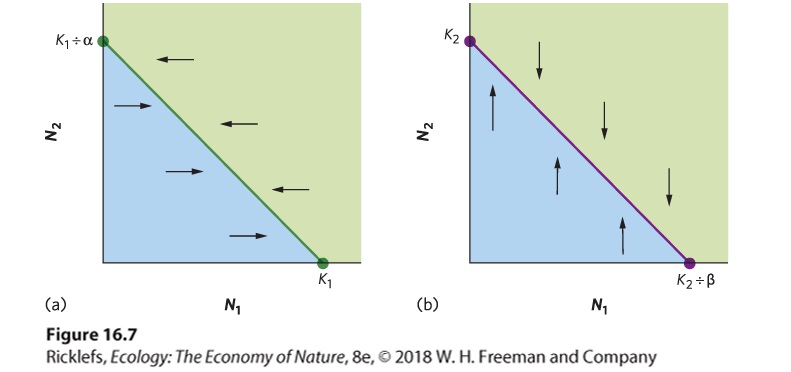
determining the conditions under which species 1 will have a population at equilibrium. This will happen whenever the change in population size per unit time is zero: dN1dt=r1N1(1-N1+αN2K1)=0r1N1(1-N1+αN2K1)=0 The equation indicates that there are two conditions that would put a population’s growth at equilibrium. One condition occurs when N1 = 0. Not surprisingly, if we have no individuals of species 1, there will be no growth of species 1. For the second condition, when N1 is not zero, we can find equilibrium by rearranging the equation: r1N1(1-N1+αN2K1)=0(1-N1+αN2K1)=1N1+αN2K1=1N1+αN2=K1N1=K1- αN2 In words, this equation tells us that the number of N1 individuals that can be present at equilibrium depends on the total carrying capacity for species 1 minus the amount of resources consumed by some number of N2 individuals. Based on this equation, we can draw a line that represents all combinations of N1 and N2 that would exist when the N1 population is at equilibrium. We can plot this line on a graph that has N1 on the x axis and N2 on the y axis, as shown in Figure 16.7a. To draw this line, we need to identify the x and y intercepts on the graph.
Figure 16.7 Equilibrium population sizes of two species. (a) The equilibrium isocline for species 1, shown as a green line, represents all combinations of species 1’s population sizes (N1) and species 2’s population sizes (N2) that cause species 1 to experience zero growth. When species 2 has an abundance of zero, species 1 will have an abundance of K1, which is the carrying capacity of species 1. (b) The equilibrium isocline for species 2, shown as a purple line, represents all combinations of species 1’s population sizes (N1) and species 2’s population sizes (N2) that cause species 2 to experience zero growth. When species 1 has an abundance of zero, species 2 will have an abundance of K2, which is the carrying capacity of species 2. The arrows indicate how each population either increases or decreases as it moves away from the equilibrium line. The x intercept can be found by setting N2 equal to zero. If we do this, the equation simplifies to N1=K1−α(0)N1=K1 As you can see from this equation, when there are no N2 individuals, the N1 population achieves equilibrium when it reaches its carrying capacity, K1. The y intercept can be found by setting N1 equal to zero. If we do this, the equation simplifies to 0=K1−αN2αN2=K1N2=K1÷α As you can see from this equation, when there are no N1 individuals, equilibrium occurs when the N2 population reaches K1 ÷ α individuals. In our example of squirrels and rabbits, this would mean that in the absence of any rabbits, the habitat could support 200 squirrels. Now that we have identified the x and y intercepts, we can construct the
line on the graph that represents all combinations of N1 and N2 that cause the population of N1 to be at equilibrium, as illustrated by the green line in Figure 16.7a. As you can see in this figure, as we increase N2, the equilibrium population size of N1 becomes smaller because species 2 is now consuming some of the resource that species 1 needs. Because this equation of a straight line represents all population sizes at which a population experiences zero growth, it is called a zero population growth isocline. Zero population growth isocline Population sizes at which a population experiences zero growth. The zero population growth isocline for species 1 represents the values of N1 that are in equilibrium across a range of N2 values. As a result, when the N1 population is to the left of the isocline, as indicated by the blue-shaded region, it will experience population growth and move to the right until it reaches the isocline. In contrast, when the N1 population is to the right of the isocline, as indicated by the green-shaded region, it will experience a population decline, thereby moving to the left until it reaches the isocline. Now that we have graphically represented the zero population growth isocline for species 1, we need to do the same for species 2. To do this, we set the second equation equal to zero and then rearrange the equation: dN2dt=r2N2(1-N2+βN1K2)=0r2N2(1N2+βN1K2)=01N2+βN1K2=0N2+βN1K2=0N2+βN1=K2N2=K2-βN1 You might notice that this equation looks very similar to the equation we derived for N1. To determine the x and y intercepts for the equilibrium line of species 2, we can go through the same mathematical steps that we used for species 1. For example, when we set N1 = 0, equilibrium will occur for

species 2 when it reaches its carrying capacity: N2=K2−β(0)N2=K2 If we then set N2 = 0, equilibrium will occur when 0=K1−βN1βN1=K2N1=K2÷β Using these two intercepts, we can plot a zero population growth isocline for species 2, as illustrated by the purple line in Figure 16.7b. As you can see in this figure, as N1 increases, the equilibrium population size of N2 decreases because species 1 is consuming the resource that species 2 needs. Because this is a zero population growth isocline for species 2, when the N2 population is above the isocline, as indicated by the green-shaded region, it will experience a population decline until it reaches the isocline. In contrast, when the population is below the isocline, as indicated by the blue-shaded region, it will experience population growth until it hits the isocline. In drawing both of these population isoclines, you might notice that the growth rates of the two populations (r1 and r2) have no effect on the position of the isocline. While the growth rates determine how quickly a population can reach equilibrium, they do not affect the location of the equilibrium. Predicting the Outcome of Competition Now that we know the conditions under which each of two species reaches an equilibrium population size, we can determine if one of the two species will win the competitive interaction or if the two species will coexist. We do this by overlapping the two zero population growth isoclines for species 1 and species 2. As illustrated in Figure 16.8, overlaying the two lines can produce four possible outcomes. In all four cases, when both species start with small population sizes, as indicated by the yellow-shaded regions in all four graphs, both species will experience population growth. In contrast, when both species start with large population sizes, as indicated by the green-shaded regions in all four graphs, both species will experience a population decline. It is the region that lies between these two extremes where we can determine the outcome of competition.
Figure 16.8 Predicting the outcome of competition for a single resource. Based on the Lotka–Volterra competition equations, there are four possible outcomes of competition. (a) If the zero population growth isocline for species 1 is farther out than species 2, species 1 will increase to its carrying capacity and species 2 will go extinct. (b) If the isocline for species 2 is farther out than species 1, species 2 will increase to its carrying capacity and species 1 will go extinct. (c) If the isoclines cross and the two carrying capacities are the outermost points on the two axes, the winner depends on the initial number of each species. (d) If the isoclines cross and the two carrying capacities are the innermost points on the two axes, then the two species will coexist. As in Figure 16.7, the green line represents the population isocline for species 1 and the purple line represents the population isocline for species 2. The thin black arrows represent increases and decreases in population sizes for each species, whereas the thick black arrows represent the net effects of population changes in both species. Open circles represent stable equilibria, which is the outcome of competition. In Figure 16.8a, the isocline for species 1 lies farther out than the isocline for species 2. As a result, any combination of the two species numbers that falls within the blue-shaded region means that species 1 is below its isocline and its population will grow, whereas species 2 is above its isocline and its population will decline. The net effect of species 1 moving to the right and species 2 moving down—both indicated by thin arrows—is that the
combination of the two species moves down and to the right, as indicated by the middle, thick arrow. This combination will continue to move until it reaches equilibrium, which is indicated by the open circle. At this point, N1 reaches its carrying capacity and N2 goes extinct. In Figure 16.8b, we see the opposite situation; the isocline for species 2 lies farther out than the isocline for species 1. In this case, any combination of the two species that falls within the blue-shaded region means that species 1 is above its isocline and its population will decline, whereas species 2 is below its isocline and its population will grow. With species 1 moving to the left and species 2 moving up, the net effect is that the combination of the two species moves up and to the left. As this process continues, the combination will reach equilibrium, which is again indicated by the open circle; at this point, N2 reaches its carrying capacity and N1 goes extinct. In Figure 16.8c, the two isoclines cross, with K1 and K2 as the two most extreme points on the axes. In this scenario, there are two possible equilibria, as indicated by the two open circles. If the combination of the two species falls within the blue-shaded region, the outcome of competition depends on the combination of N1 and N2 that are present. If the combination of N1 and N2 falls within the upper left blue region, the net movement is toward K2 and species 2 wins the competitive interaction. If the combination of N1 and N2 falls within the lower right blue region, the net movement is toward K1 and species 1 wins. In Figure 16.8d, the two isoclines also cross, but this time with K1 and K2 as the two innermost points on the axes. In this scenario, the movement of each population relative to its respective isocline causes a net movement toward a single equilibrium point where the two lines cross. This means that the two competing species will coexist; one species will not outcompete the other. In general, these equations tell us that coexistence of two competing species is most likely when interspecific competition is weaker than intraspecific competition—that is, when the competition coefficients α and β are less than 1. In other words, coexistence of two competing species occurs when individuals of each species compete more strongly with themselves (conspecifics) than with individuals of other species (heterospecifics).
Competition for Multiple Resources
Although it is simpler to think about two species competing for one resource, in reality, species in nature compete for multiple resources. For example, grassland plants might simultaneously compete for water, nitrogen, and sunlight. Let’s consider two species competing for two different resources. If species 1 is better able to sustain itself at low levels of both resources than species 2, then species 1 will win the competitive interaction. However, the outcome changes when species 1 is better at persisting at a low level of one resource but species 2 is better at persisting at a low level of another resource. In this case, because neither species can drive the other to extinction, the two species should coexist. This principle was first demonstrated by David Tilman in a series of experiments conducted with two species of diatoms: Asterionella formosa and Cyclotella meneghiniana. We encountered these species when we discussed Liebig’s law of the minimum. Both species require silicate (SiO2) to produce their glassy outer shells and both species require phosphorus (P) for their metabolic activities. However, each species differs in how much of each resource it needs. Cyclotella uses silicate more efficiently, so its population can persist when there is a low abundance of silicate. In contrast, Asterionella uses phosphorus more efficiently, so its population can persist when there is a low abundance of phosphorus. Based on these differences, Tilman predicted that if the diatoms were raised under a low ratio of SiO2/P, Cyclotella would dominate the competitive interaction because silicate would be low in abundance. If they were raised under a high ratio of SiO2/P, Asterionella would dominate the competitive interaction because phosphorus would be low in abundance. He also predicted that at intermediate ratios of SiO2/P, the two species would coexist because each is limited by a different resource. To test this hypothesis, Tilman raised the two species separately and together under a range of different SiO2/P ratios in laboratory containers, as shown in Figure 16.9. When raised separately, both species were able to survive under all the SiO2/P ratios. However, Cyclotella dominated the containers when they were raised together at a low SiO2/P ratio of 0.6, as shown in Figure 16.9a. In contrast, Asterionella dominated the containers
when they were raised at a high SiO2/P ratio of 455, as shown in Figure 16.9c. When they were raised at an intermediate SiO2/P ratio of 38, the two species coexisted and neither became dominant, as you can see in Figure 16.9b. These were the first results to demonstrate that competing species can coexist when there are multiple resources and each species is limited by a different resource.
Figure 16.9 Species competing for multiple resources. In the experiment, two species of diatoms compete for silicon and phosphorus. Cyclotella is more limited by a low abundance of phosphorus (P), whereas Asterionella is more limited by a low abundance of silicate. (a) When raised under a low ratio of SiO2/P, Cyclotella dominates the competitive interaction. (b) When raised under an intermediate ratio of SiO2/P, neither species dominates. Instead, the two species coexist. (c) When raised under a high ratio of SiO2/P, Asterionella dominates the competitive interaction.
1. What mathematical terms need to be added to the logistical growth equation to create the competition equation of population growth? 2. In the competition equation of population growth, why do we need to include competition coefficients? 3. Under what conditions can we expect the stable coexistence of two species competing for two resources?
#### 16.3 the Outcome of Competition Can Be Altered by Abiotic Conditions, Disturbances, and Interactions with Other Species

16.3 The outcome of competition can be altered by abiotic conditions, disturbances, and interactions with other species We have considered what happens when two species compete for shared resources. However, in nature, competition occurs in the context of many different environmental conditions and with other types of species interactions, including predation and herbivory. In this section, we will examine how competitive outcomes can be altered by abiotic conditions, disturbances, and interactions with other species.
Abiotic Conditions
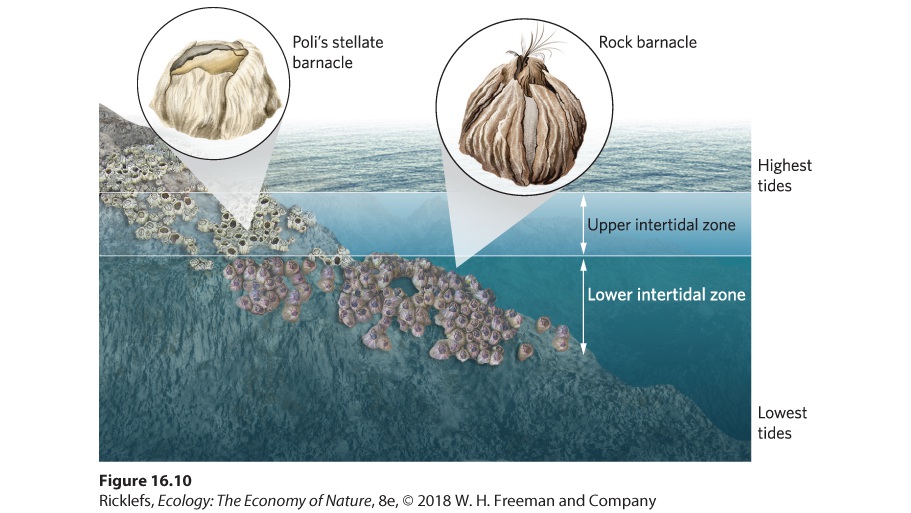
The ability to persist when resources are scarce is important to winning in a competitive situation, but it can be overwhelmed by the ability to persist under harsh abiotic conditions. We can see an example of this in a classic study of barnacles, conducted by Joseph Connell in Great Britain. Barnacles begin their lives as larvae that float on ocean currents and then settle on open, rocky spaces of coastal intertidal zones. They spend the rest of their lives as sessile adults, commonly living in dense groups. They feed on plankton that they filter from the water that washes over them. Because the barnacles cannot substantially reduce the vast amount of plankton that exists in coastal waters, plankton is not a limiting resource. Instead, barnacles are often limited by the amount of open space on rocks in the intertidal habitat. Joseph Connell observed that the larvae of two barnacle species on the coast of Scotland are distributed broadly along the upper and lower intertidal zones. However, adults are distributed in two separate zones; Poli’s stellate barnacles (Chthamalus stellatus) live in the upper intertidal zone, whereas rock barnacles (Semibalanus balanoides, formerly Balanus balanoides) live in the lower intertidal zone. The two species have only a small overlap in their distributions, as shown in Figure 16.10.
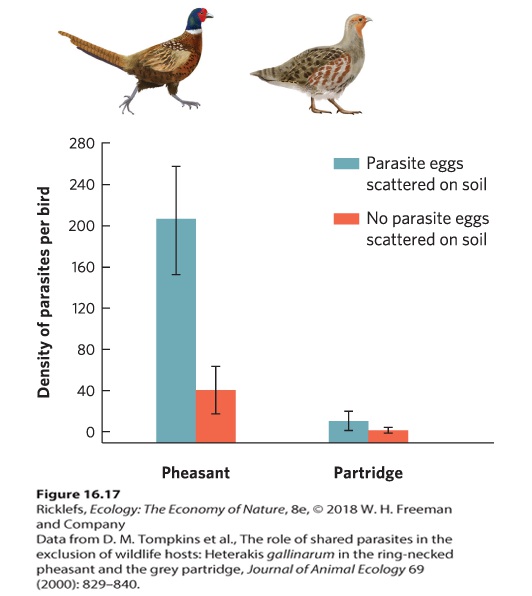
Figure 16.10 Competition between barnacles. In Great Britain, Poli’s stellate barnacle lives in the upper intertidal zone because it is more resistant to desiccation and it cannot compete successfully in the lower intertidal zone. In contrast, the rock barnacle lives in the lower intertidal zone because it is the superior competitor but is not resistant to the desiccating environment of the upper intertidal zone. To understand the reason for this distribution, Connell conducted a series of experiments in which he removed one or the other species. When he manually removed the rock barnacles from the lower part of the intertidal zone, the stellate barnacles quickly moved down and thrived. However, when he removed the stellate barnacle from rocks in the upper intertidal zone, the rock barnacles did not move up because they could not survive the desiccation that occurs in the upper intertidal zone. The stellate barnacles thrive in the upper intertidal zone because they are resistant to desiccation during periods of low tide. Though they could live in the lower intertidal zone, competition from the rock barnacle prevented it. The rock barnacle has a rapid growth rate and heavier shell than the stellate barnacle, which allows the rock barnacle to expand in size, pry the stellate barnacle off the rock, and take over the newly opened space. In short, the distribution of the two species is the result of a trade-off that many species experience—the trade-off between competitive ability and the ability to tolerate another challenging abiotic factor.
Disturbances
Competitive interactions among species can also be altered by disturbances. For example, in the southeastern United States, more than 36 million ha of
forest were once dominated by longleaf pine trees (Pinus palustris) with a very open, grassy understory. Many of the plants that live in this forest depend on frequent, low-intensity fires to persist (Figure 16.11). In fact, some species such as wiregrass (Aristida beyrichiana) can only reproduce after a fire has swept through the forest. Throughout the twentieth century, the natural fires were suppressed in much of the longleaf pine forest, which caused other species of woody plants to increase and outcompete the plants adapted to fire. Fire suppression altered the outcome of plant competition in the forest because it allowed the superior competitors previously removed by fire to persist.
Figure 16.11 Fire in a longleaf pine forest. These low-intensity fires, such as this one in Georgia, remove many species of woody plants and favor the growth of fire-adapted trees and grasses.
Predation and Herbivory
The outcome of competition can also be altered by the presence of predators and herbivores. We frequently find a trade-off between competitive ability and resistance to predators or herbivores; the most competitive plants are often the most susceptible to herbivores and the most competitive animals are typically the most susceptible to predators. For example, while many good competitors have high feeding rates because they move a lot in search of

food, this activity makes them more noticeable to predators and thus more likely to be killed by predators. Studies of aquatic communities have shown that predators can reverse the outcome of competition. For example, in one experiment, large outdoor tanks were used to simulate ponds. Each pond was supplied with 200 tadpoles of the spadefoot toad (Scaphiopus holbrookii), 300 tadpoles of the spring peeper (Pseudacris crucifer), and 300 tadpoles of the southern toad (Anaxyrus terrestris). After setting up a series of identical tanks, each one was randomly assigned a treatment of 0, 2, 4, or 8 predatory newts (Notophthalmus viridescens). In the absence of newts, the spadefoot toad’s superior competitive ability was clear. As depicted in Figure 16.12, survival was high for spadefoot toad tadpoles, moderate for southern toad tadpoles, and low for spring peeper tadpoles. Spadefoot toad tadpoles had high survival because they are very active and rapidly consume whatever algae are available. In contrast, southern toad tadpoles are somewhat less active and spring peeper tadpoles are very inactive and therefore grow slowly. When the predatory newts were added, the outcome changed dramatically. As the number of newts increased, the survival of the spadefoot toads dropped precipitously and the survival of the southern toads dropped modestly, which is consistent with the southern toad’s higher activity levels. Compared to the control treatment, the addition of eight newts reduced the survival of spadefoot toads by 75 percent and caused the survival of southern toad tadpoles to decrease by 32 percent. However, the survival of spring peepers increased by 26 percent. The reason for the improved survival of the spring peepers is that they are a very inactive tadpole species. This low activity helps them avoid being noticed by predators, but it comes at the cost of poor competitive ability. When newts are abundant and consume the superior competitors, the poorly competing spring peepers do much better and the outcome of competition among the three species of tadpoles is reversed.
Figure 16.12 Altering competition with predators. In the absence of predatory newts, the spadefoot toads and southern toads survive well because they are superior competitors. With the addition of predatory newts, the survival of the two toad species decreases, while the survival of the spring peepers increases. Error bars are standard errors. Herbivores can have a similar effect on competition. For example, several species of goldenrod dominate fallow fields throughout the northeastern United States. These plants can grow more than a meter high and cast shade upon their shorter competitors. A species of beetle (Microrhopala vittata) specializes in consuming the goldenrods. Every 5 to 15 years, the beetle population achieves very high densities, known as an insect outbreak, during which the beetles consume large amounts of the goldenrod plants. The impact of the beetles on the plant community was determined in a decade-long experiment in which some plots of plants were sprayed with an insecticide to kill the goldenrod-consuming beetles and prevent the insect outbreak, while other plots of plants were left unsprayed. The results were dramatic (Figure 16.13). In the sprayed plots that contained few herbivorous beetles, shown on the right side of the photo, the goldenrods grew tall, cast shade on the other species of plants, and dominated the community with their
superior competitive ability. In the unsprayed plots, shown on the left side of the photo, the beetles experienced a very large population increase midway through the experiment, and they consumed most of the goldenrod plants. The goldenrods that remained were substantially shorter, so they cast less shade on the inferior competitors. With fewer large goldenrods present, the inferior competitors became much more abundant. Based on these results, the researchers confirmed that the process of the herbivorous beetle eating the superior competitor caused a reversal in the outcome of competition among plants.
Figure 16.13 Altering competition with herbivores. Over a 10-year period, the plot on the right was sprayed with an insecticide to prevent beetles from consuming the goldenrods—the tall plants with yellow flowers—while the plot on the left was left unsprayed. The unsprayed plot experienced an outbreak of beetles, and this greatly reduced the density and height of the goldenrod plants. As a result, many species of plants that are inferior competitors became more numerous.
Concept Check
1. How can abiotic conditions alter the outcome of competition?
2. How do disturbances alter the outcome of competition? 3. What is the underlying trade-off that allows predators and herbivores to reverse the outcome of competition?
#### 16.4 Competition Can Occur Through Exploitation, Direct Interference, or Apparent Competition
##### Interference Competition: Aggressive Interactions
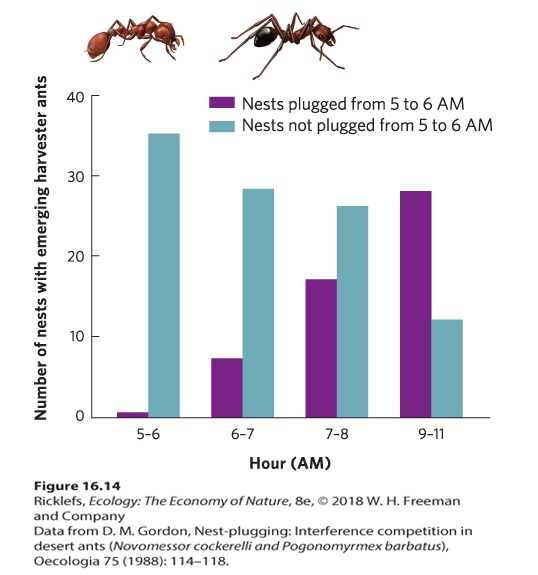
16.4 Competition can occur through exploitation, direct interference, or apparent competition Competition can be categorized in several ways. Thus far, we have seen numerous examples of exploitative competition, in which individuals consume and drive down the abundance of a resource to the point that other individuals cannot persist. For example, the different species of diatoms compete for silicate and phosphorus by consuming these resources and driving down their abundance. Sometimes competitors do not immediately consume resources but defend them, a type of competition known as interference competition. Whereas interference competition involves a direct interaction between two individuals, exploitative competition is considered an indirect interaction because it operates through a shared resource. Finally, two species can appear to compete for a shared resource but in fact cause a negative effect on each other due to mechanisms that are not competition, so we say that they experience apparent competition. Given our previous emphasis on examples of exploitative competition, in this section, we will explore the concepts of interference competition and apparent competition. Exploitative competition Competition in which individuals consume and drive down the abundance of a resource to the point that other individuals cannot persist. Interference competition When competitors do not immediately consume resources but defend them. INTERFERENCE COMPETITION: AGGRESSIVE INTERACTIONS Aggressive interactions are an effective form of interference competition that occurs between species of animals. For example, in the deserts of New Mexico, two species of ants—the long-legged ants (Novomessor cockerelli) and the red harvester ants (Pogonomyrmex barbatus)—compete for seeds and for any insects they can catch and subdue. However, the long-legged ants have a unique method of competing through interference. Early in the morning, they emerge from their nests, find the nests of the red harvester ants, and plug the nest entrances with stones and soil. As you can see in Figure 16.14, when researchers examined the effects of this aggression, they found
that harvester ants with plugged entrances required several hours to unplug their nests before they could begin foraging for seeds. As a result, the longlegged ants can forage for several hours in the morning with minimal competition from the harvester ants.
Figure 16.14 Interference competition in ants. Red harvester ants and long-legged ants compete for seeds and insects. When the harvester ant nest is not plugged by the longlegged ant, a large number of colonies emerge by 6 am. However, if the nest entrances are plugged by the long-legged ant, it can take several hours for the harvester ants to clear the openings and start foraging. INTERFERENCE COMPETITION: ALLELOPATHY Another type of interference, known as allelopathy, occurs when organisms use chemicals to interfere with their competitors. We saw an example of this in the earlier story of garlic mustard. As a second example, black walnut trees (Juglans nigra) produce juglone, an aromatic organic compound that inhibits
certain enzymes in other plants. The compound—found in the leaves, bark, roots, and seed husks of the black walnut tree—leaches out of the plant and into the soil, making it difficult for most other plant species to germinate and grow under a walnut tree. Allelopathy A type of interference that occurs when organisms use chemicals to harm their competitors. Allelopathy can also be an effective strategy for invasive plants, which invade and dominate a community. The wetland plant known as the common reed (Phragmites australis) is found throughout the world (Figure 16.15). In North America, some genetic strains are native and other strains have been introduced from Europe. In the Great Lakes region of North America, the introduced strain is spreading rapidly and has displaced many other species of native wetland plants. For many years, researchers hypothesized that the common reed was successful at spreading due to allelopathy, but they lacked evidence. Recently, it was discovered that the root of the common reed produced a chemical known as gallic acid, which is highly toxic to the roots of many other plants. By harming the roots of other species, the common reed impairs the growth of its interspecific competitors. The researchers also discovered that the root chemicals produced by the introduced strains of the reed were much more lethal to some plants than the root chemicals produced by the native strain, as shown in Figure 16.16. This result supports the observation that the introduced strains of the reed are able to invade a wetland and spread much more rapidly than the native strains.
Figure 16.15 The common reed. The common reed, a wetland plant, can be found in many locations around the world, including this site in Cape Cod, Massachusetts. In North America, some genetic strains are native, whereas others have been introduced from Europe.
Figure 16.16 Allelopathy in the common reed. Root chemicals from native or invasive strains of the common reed were extracted and then added to pots of mouse-ear cress or tobacco plants. The addition of the root chemicals caused both plant species to grow poorly compared to the control. In the case of the tobacco plant, chemicals from the invasive strain of the reed inhibited growth more than the native strain. Error bars are standard deviations. Allelopathy does not always take the form of toxic effects on interspecific competitors. In Australia, for example, flammable oils in the leaves of some eucalyptus could be an adaptation that promotes frequent fires in the leaf litter of the forest floor, which kill seedlings of competitors. Regardless of whether organisms use direct aggressive interactions or allelopathic chemicals, it is clear that interference competition can be an important
mechanism in determining the abundance and distribution of species.
Apparent Competition
Throughout this chapter, we have defined competition as a negative interaction between two individuals for a limited resource. Sometimes, however, two species can share a resource and have negative effects on each other without competing for a resource. When two species have a negative effect on each other through an enemy—including a predator, parasite, or herbivore—we call it apparent competition. As its name suggests, apparent competition causes an outcome that looks like competition, but the underlying mechanism is different. Apparent competition When two species have a negative effect on each other through an enemy—including a predator, parasite, or herbivore. Apparent competition can be observed in a variety of communities. For example, the ring-necked pheasant and the gray partridge (Perdix perdix) live in many of the same habitats in the United Kingdom. For 50 years, a large decline in the number of gray partridges had been attributed to increased agriculture, although researchers suspected that a parasitic nematode (Heterakis gallinarum) might also play a role. Infected pheasants experience few harmful effects from the parasite, but infected partridges can suffer weight loss, reduced fecundity, and death. To determine the susceptibility of each species to infection by parasites in the environment, researchers allowed both bird species to feed either in pens containing parasite eggs scattered across the soil or in pens without the eggs. After 50 days of living in the pens without added parasite eggs, they found that both species contained low parasite infections, as shown in Figure 16.17. However, in the pens that contained added parasite eggs, the pheasants became infected at rates that were more than 20 times higher than those for the partridges. Moreover, when both birds were fed the same number of nematode eggs over a 100-day period, the pheasants excreted over 80 times more nematode eggs than the partridges. In short, pheasants can carry a large number of parasites without being harmed, and they excrete a large number of parasites that subsequently infect and harm partridges. The presence of pheasants is associated with partridges that gain less weight and have a low
rate of survival. The relationship between the birds has the appearance of competitors for a shared resource, but the real cause of the declining partridges is a shared parasite.
Figure 16.17 Apparent competition in pheasants and partridges. Pheasants (left) and partridges (right) were held in pens with no parasite eggs, or with parasitic eggs of nematodes scattered across the soil. In the absence of parasite eggs, some birds exhibited a low level of background infection. In the presence of parasite eggs, pheasants had a much higher number of parasites in their bodies after 50 days. Pheasants are not harmed by the parasites, but they serve as a major reservoir of new parasite eggs, which they excrete onto the soil. Partridges, which experience harmful effects from the parasite, had a low number of parasites and therefore excreted few parasite eggs into the environment. The presence of pheasants causes apparent competition because they release large numbers of parasite eggs, which leads to a decline in the partridges. Error bars are standard errors. There are many examples of apparent competition involving pathogens.
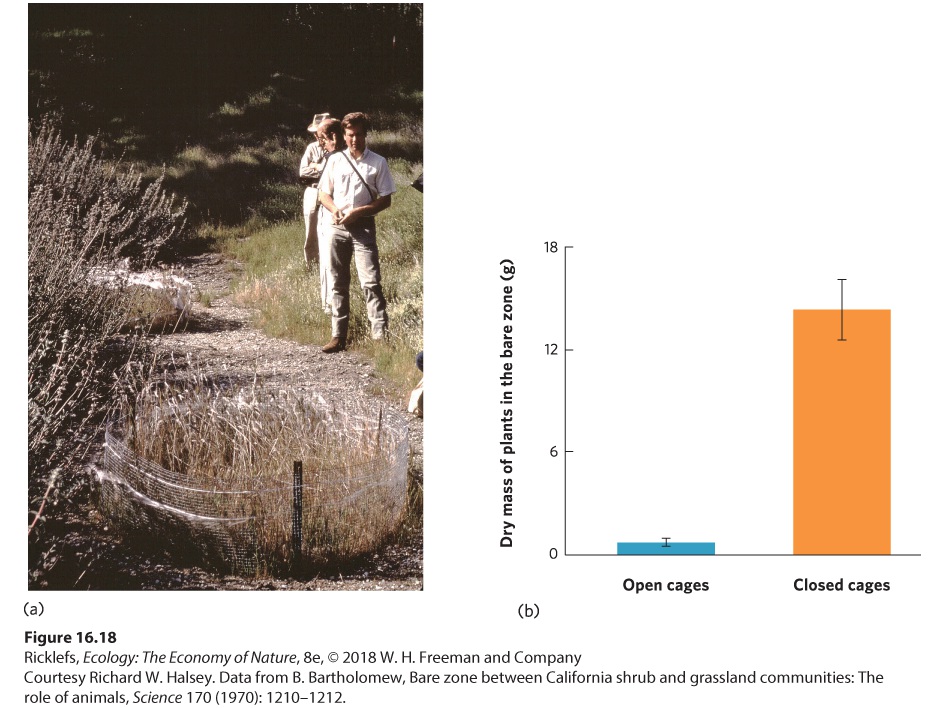
For instance, when Europeans colonized other continents, particularly North and South America, they brought with them diseases such as smallpox. The Europeans could tolerate many of these diseases because they had a long evolutionary history of living with them, but Native Americans lacked this evolutionary history and the introduced diseases devastated their populations. While the Europeans appeared to cause competition, the reduction of the Native American population occurred primarily because the Europeans brought new pathogens to the Western Hemisphere. In some cases, apparent competition occurs not because of a shared enemy, but because one species facilitates the enemy of another species. For example, on the coast of California, several species of shrubs commonly have a zone of bare soil immediately around them followed by a more distant zone made up of grasses (Figure 16.18a). Researchers initially hypothesized that the bare soil zone was caused by allelopathic chemicals produced by the shrubs, which made sense because some of the shrub species were known to produce chemicals that inhibited the growth of other plants when the chemicals were at high concentrations in the soil. However, since not all the shrub species produced these chemicals, the allelopathy hypothesis was not well supported.
Figure 16.18 Apparent competition between sage and grasses in California. (a) Researchers originally hypothesized that the bare zone surrounding the purple sage shrubs (Salvia leucophylla) occurred because of allelopathic chemicals the shrubs produced.
However, subsequent research discovered that herbivores used the shrubs as a refuge and only fed on grasses that were close to the safety of the shrub. To demonstrate that the effect was due to mice eating the plants surrounding the sage shrubs, researchers placed opensided and closed cages in the bare zone, as shown here in a photo of the researchers in the Santa Ynez Valley, California, in 1968. (b) One year later they found nearly 20 times more vegetation growing in the closed cages, which confirmed that herbivores were causing the bare zone of vegetation around the shrubs. Error bars are standard errors. Further research of the shrub growth showed that what appeared to be interference competition was not really competition at all. In one experiment, when seeds were placed on the bare zone and in the grass, 86 percent of the seeds were consumed in the bare zone but only 12 percent were consumed in the grass. When small mammal traps were set out in both areas, 28 mice were captured in the bare zone but only 1 mouse in the grass. Finally, two types of cages were placed in the bare zone: open-sided cages that allowed mice to move in and out of the cages and closed cages that excluded rodents. One year later, there was still little vegetation in the open cages but nearly 20 times more vegetation in the closed cages, as shown in Figure 16.18b. Collectively, these results demonstrate that the bare zone, which appeared to be the outcome of competition by allelopathy, actually occurred because the shrubs were a safe haven for mice from predators. The mice forage for seeds only a short distance from the shrubs and then run back to the shrub whenever predators appear. As a result, there are very few seeds surviving in close proximity to the shrubs, causing a bare zone that is created by herbivory rather than allelopathy.
Analyzing Ecology
Chi-Square Tests When the researchers in California tried to determine the causes of the bare zone around coastal shrubs, they captured 28 mice in the bare zone near the shrubs and 1 mouse in the grass. Although this is a substantial difference, how do we know if this difference is statistically significant? In Chapter 14, we discussed the importance of testing hypotheses using a criterion of α < 0.05 for statistical significance. In Chapter 15, we
discussed testing hypotheses using t-tests when we have estimates of mean values and standard deviations. Sometimes, however, our data do not consist of means and standard deviations, but rather of individuals that are counted, such as the number of mice in the bare zone versus those in the grass. When our data consist of counts, we can test whether the data are significantly different from an expected distribution using a chi-square test. A chi-square test is a statistical test that determines whether the number of observed events in different categories differs from an expected number of events, which is based on a particular hypothesis. In the case of the mice, the researchers sampled both habitats equally and they captured a total of 29 mice from both areas. If the 29 mice exhibited no habitat preference, we would have expected to find 14.5 mice in the bare zone and 14.5 mice in the grass. As a result, we want to statistically compare the observed distribution of mice—28 mice in the bare zone and 1 mouse in the grass—to an even distribution between the two habitats. Now that we know the observed distribution and the expected distribution for mice that exhibited no preference, we can create a small table and conduct the chi-square test. Chi-square test A statistical test that determines whether the number of observed events in different categories differs from an expected number of events, which is based on a particular hypothesis. Observed Expected Bare Zone 14.5 Grass 14.5 To calculate the chi-square value, we use an equation that includes our observed (O) and expected (E) values to calculate the value of chi-square (χ2): χ2=∑i=1n(Oi−Ei)2(Ei)χ2=(28−14.5)2+(1−14.5)214.5χ2=25.1
As was the case when we did a t-test in Chapter 15, we have to compare our calculated value to a table of values to determine if our observed distribution is significantly different from the expected distribution. The first step in that process is determining the degrees of freedom: Degrees of freedom=(number of observed categories−1)=(2−1)=1 Now we can examine the chi-square table provided in the statistical tables in the appendix. Using these tables, we can compare our calculated value (25.1) to the critical chi-square value when we have 1 degree of freedom and we set α = 0.05. In this case, the critical chi-square value is 3.841. Since our calculated value is greater than the critical value, we can conclude that the observed distribution of mice differed significantly from an equal distribution of mice between the two habitats. YOUR TURN Using a chi-square test, determine whether an observed distribution of 12 mice in the bare zone and 8 mice in the grass differs significantly from an expected even distribution of mice. Throughout this chapter, we have seen that competition is a major force affecting the distribution and abundance of organisms. This competition can occur by exploitation or interference and it can occur among species both closely and distantly related. As we will see in “Ecology Today: Finding the Forest in the Ferns,” understanding the role of competition and how it interacts with disturbance, herbivory, and apparent competition is important for understanding how the natural world works, including biomes that provide humans with valuable natural resources.
Concept Check
1. How is interference a form of competition? 2. Why is allelopathy considered a form of interference competition? 3. For what types of data do we use chi-square tests?
Concepts
Finding the Forest in the Ferns A mature temperate forest carpeted with hay-scented fern. The dense fern layer is associated with a low number of tree seedlings, as evidenced by the lack of small trees growing in the understory of this forest in Shenandoah National Park, Virginia. In forests around the world, adult trees produce offspring that will become the next generation of trees. For a long time, it was assumed that the composition of trees in the next generation would be determined by the composition of
seedlings produced by adult trees, the amount of shade, and the ability of the seedlings of each species to tolerate shade. For example, because seedlings of quaking aspen trees (Populus tremuloides) cannot tolerate shade from adult trees, they regenerate only when there are large openings in the forest that allow high amounts of sunlight to reach the ground. In contrast, seedlings of American beech trees are highly tolerant of shade and can grow in forests even under a dense canopy of leaves. While light gaps in the forest and variation in the shade tolerance of different tree species are important determinants of the composition of the next generation of forest, recent research has found that the effects of light gaps and shade tolerance can be overwhelmed by herbivory. An excellent example of this phenomenon has occurred in the eastern deciduous forests of the United States. Historically, these forests had a variety of mature tree species in the upper canopy and younger trees in the understory. The forests also had low numbers of large white-tailed deer and occasional natural fires. Over more than a century, the forests were logged, fires were suppressed, and top predators such as mountain lions and wolves were removed. The removal of top predators caused a dramatic increase in the whitetailed deer population, which consumed large numbers of tree seedlings. Although the logging increased the amount of sunlight reaching the ground, and this should promote the germination and growth of new tree seedlings, the deer became so abundant that they greatly reduced the survival of many seedling species. However, we now know that the impact of the deer eating tree seedlings is only part of the story.
When deer eat plants, they prefer the most palatable species and leave the others unharmed. In the forests of Pennsylvania, for example, deer do not eat the unpalatable hay-scented fern (Dennstaedtia punctilobula). Although the fern historically comprised less than 3 percent of the understory vegetation, today it is the dominant understory plant in more than one-third of Pennsylvania forests. This is an important change in the forest because few tree seedlings can emerge out of the dense fern layer. As a result, there are now many large trees in the forest canopy but few young trees to become the next generation. For many years, foresters were certain that few seedlings were emerging from the layer of ferns because the ferns were casting high amounts of shade over the seedlings. Competitive exclusion between these distantly related species made a great deal of sense. Recently, however, an alternative explanation has been proposed; perhaps the fern layer is not competing but is instead providing a refuge for rodents from predatory hawks and owls and that this refuge allows rodents to consume more tree seeds and tree seedlings.
Apparent competition from ferns in a forest. To determine whether the effect of ferns was due to competition or apparent competition, the researchers used fencing to exclude rodents and manipulated the presence of ferns. (a). Few seeds of beech trees were removed from fenced plots, but many seeds were removed from unfenced plots, especially when ferns were present. (b) Similarly, sugar maple seedlings experienced low rates of mortality in fenced plots but high rates of mortality in unfenced plots in which ferns remained in place. Error bars are standard errors.
Researchers reported the results of an experiment to determine which of these two hypotheses was correct: whether the negative effect of the ferns on tree seedlings was caused through competition for resources or through apparent competition by helping the rodents. They began by selecting plots of ferns within a forest, each with an area of 4 m2. For each plot, they either fenced out rodents or let rodents roam throughout the plot, and they either removed the ferns or left them in place. After replicating the four different manipulations several times, they examined how many rodents were captured in each plot. The most common rodent species were deer mice, chipmunks, and red-backed voles (Clethrionomys gapperi). Rodents rarely got inside the fenced plots, regardless of fern coverage. However, rodents were frequently captured in the unfenced plots, with more than twice as many rodents captured when the ferns remained in place. When beech seeds were added to the plots, only a small percentage of seeds were removed in the fenced plots— regardless of fern coverage—as shown in part (a) of the graph. However, a large percentage of seeds were removed in the unfenced plots, and even more seeds were removed when the ferns remained in place. Similar results were seen for many other species of seeds, including black cherry (Prunus serotina) and sugar maple (Acer saccharum). When the researchers set out seedlings of sugar maple, they observed low mortality in the fenced plots—regardless of whether ferns were removed—as shown in part (b) of the graph. In the unfenced plots, however, seedling mortality was higher, but only when ferns were present.
These results confirmed that the primary effect of the ferns on these tree seedlings was not as a competitor for light, but rather that they served as a refuge for rodents that eat the seeds and seedlings of beech, black cherry, and sugar maple. The results confirmed that the fern effects on American beech, black cherry, and sugar maple were due to apparent competition mediated through the fern’s protection of rodents. These experiments make clear that the regeneration of forests is complex, with multiple processes happening simultaneously. The success of new tree seedlings depends on competition for light, but also directly by large mammal herbivory and indirectly by the altered species composition of the understory that is caused by the rodent herbivory. In turn, these altered understories can appear to determine competitive outcomes through competition, but in fact they cause apparent competition by providing refuges for rodents. SOURCES: Nuttle, T., et al. 2013. Historic disturbance regimes promote tree diversity only under low browsing regimes in the eastern deciduous forest. Ecological Monographs 83: 3–17. Royo, A., and W. C. Carson. 2006. On the formation of dense understory layers in forests worldwide: Consequences and implications for forest dynamics, biodiversity, and succession. Canadian Journal of Forest Research 36: 1345–1362. Royo, A., and W. C. Carson. 2008. Direct and indirect effects of a dense understory on tree seedling recruitment in temperate forests: Habitat-mediated predation versus competition. Canadian Journal of Forest Research 38: 1634–1645.
Summary of Learning Objectives
16.1 Competition occurs when individuals experience limited resources. Competition can either be intraspecific or interspecific and occurs when there is a limited resource. Resources can either be renewable or nonrenewable, and they can be generated either from within an ecosystem or from outside it. Leibig’s law of the minimum states that a population will increase until the most limiting resource prevents further growth, although we now appreciate that different resources can have interactive effects on population growth. The competitive exclusion principle states that two species cannot coexist indefinitely when they are both limited by the same resource. Key Terms: Intraspecific competition, Interspecific competition, Resource, Renewable resources, Nonrenewable resources, Liebig’s law of the minimum, Competitive exclusion principle
16.2 The theory of competition is an extension of logistic growth models. The simplest models consider competition for a single resource and consider the zero population growth isoclines of two competing species. Using these models, we can make predictions regarding the conditions under which two species can win a competitive outcome or coexist. Under the more realistic situation of multiple limiting resources, we can have coexistence of multiple species of competitors when each species is limited by a different resource. Key Terms: Competitive coefficients, Zero population growth isocline
16.3 The outcome of competition can be altered by abiotic conditions, disturbances, and interactions with other species. If a species is competitively superior but not tolerant of extreme abiotic conditions, it will not be able to dominate areas that experience such conditions. Similarly, competitively superior species that cannot persist
with frequent disturbances, such as fire, cannot come to dominate inferior competitors. In the same way, superior competitors that are more vulnerable to herbivores or predators cannot outcompete inferior competitors because they are preferentially harmed or killed.
16.4 Competition can occur through exploitation or direct interference, or it may be apparent competition. Exploitative competition occurs when one species consumes enough of a resource that another species can no longer persist. In contrast, interference competition occurs when a species defends a resource and prevents other individuals from consuming it. Interference competition includes aggressive interactions among species and allelopathy. Sometimes species appear to be competing because the presence of one species has a negative effect on the population of the other. In cases of apparent competition, the underlying mechanism is not competition but another type of interaction, such as predation, herbivory, or parasitism. Key Terms: Exploitative competition, Interference competition, Allelopathy, Apparent competition, Chi-square test
Critical Thinking Questions
1. Compare and contrast renewable and nonrenewable resources. 2. How does Liebig’s law of the minimum explain how silicate controls the growth of diatom populations? 3. If two species require the same limiting resource, what situation would favor their coexistence? 4. Why are population growth rates at zero when two species are at equilibrium? 5. Under what conditions would distantly related species compete? 6. If natural fires and herbivores can both reduce the abundance of competitively superior plants, how should this affect the number of other plant species that can persist in the community? 7. Why it is critical to include competition coefficients in the Lotka– Volterra competition equations? 8. Why is allelopathy not a form of exploitative competition? 9. Under what conditions can two species competing for two resources not coexist? 10. Explain the difference between competition and apparent competition.
#### Graphing the Data: Competition for a Shared Resource
GRAPHING THE DATA Competition for a shared resource As we have seen in this chapter, the outcome of competition between two species for a common resource can be predicted if we know the carrying capacity and competition coefficients of each species. Using the following data, graph the zero population growth isoclines for each species. Based on your graph, what is the predicted outcome of competition between these two species? species 1: K1=100, α=0.4species 2: K2=50, β=0.3