19 Community Succession
19Community Succession Glacier Bay, Alaska. In 1794, a glacier that was more than 1,200 m thick covered all but the inlet of the bay. Since then, the glaciers have melted and retreated, leaving behind a large body of water. Retreating Glaciers in Alaska In the panhandle of Alaska near Juneau, a stretch of land has undergone incredible changes during the past 200 years. In 1794, Captain George Vancouver found an inlet that headed toward modern-day Juneau. To the north of the inlet was a massive glacier that measured more than 1 km thick and 32 km wide. When the naturalist John Muir visited the site 85 years later, he was shocked to find that the glacier was not located where Captain Vancouver had indicated on his map. Instead, it had receded nearly 80 km and left behind a large bay. Moreover, while most bays in Alaska were heavily forested, the shores of this new bay were relatively barren. As Muir explored, he found several locations with large stumps that were the
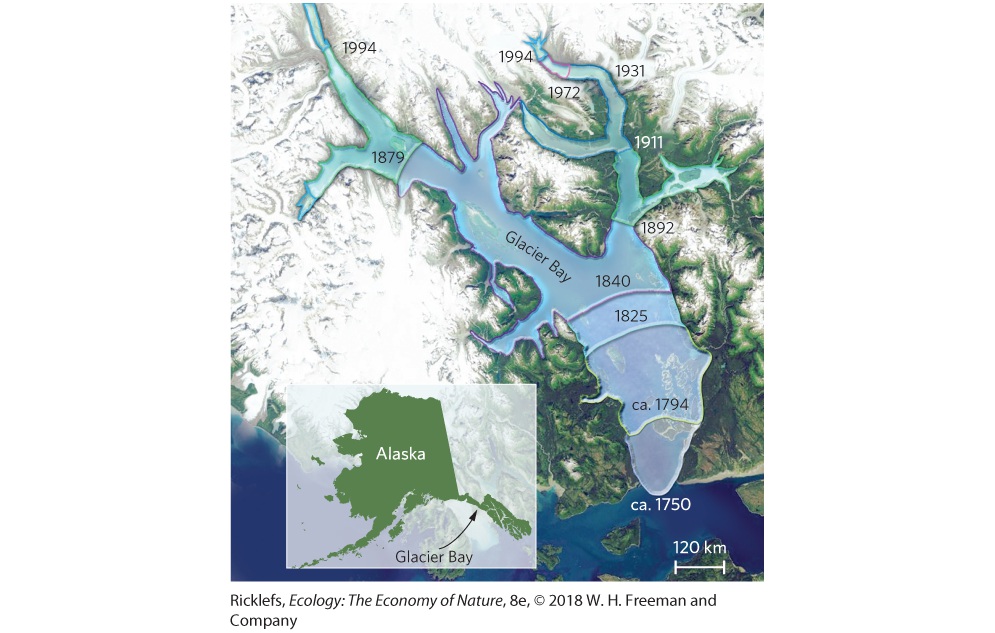
remnants of hemlock, black cottonwood (Populus trichocarpa), and Sitka spruce trees that had been sheared off by the glacier as it advanced centuries ago. Muir had no way of knowing that he was looking at one of the fastest-melting glaciers in the world. John Muir’s writings about the natural world were widely read. In 1916, William Cooper, an ecologist who was particularly intrigued by Muir’s stories of Alaska, made the first of several expeditions to the site that came to be known as Glacier Bay. On these expeditions, he surveyed the plants growing at various sites along the shore of the bay, including sites where the glacier existed during Vancouver’s visit, sites where the glacier existed during Muir’s visit, and sites where the glacier had receded only recently. He also set up quadrats around the bay to record the changes in vegetation at various locations that he planned to check when he returned on subsequent expeditions. “When the naturalist John Muir visited the site 85 years later, he was shocked to find that the glacier was not located where Captain Vancouver had indicated on his map.” Cooper reasoned that sites exposed for the longest duration would have the most time to grow back into the kind of forest that had existed before the glaciers had advanced centuries earlier. In contrast, he expected recently exposed sites to be bare rock and gravel, representing the first stages of a developing forest. On these recently exposed sites, Cooper found mosses, lichens, herbs, and low shrubs. Sites that had been exposed for 35 to 45 years had tall species of willow and alder shrubs and black cottonwood trees. Sites older than 100 years contained Sitka spruce trees, and sites older than 160 years had hemlock in the understory. By examining changes in the plant community at sites that had been exposed for different lengths of time, he was able to hypothesize how the forests of Alaska responded to the massive disturbance of an advancing and retreating glacier.
The observations of Vancouver, Muir, and Cooper paved the way for more than a century of subsequent ecological research at Glacier Bay. In fact, the original terrestrial quadrats set up by Cooper a century ago are still monitored today, as are local streams that have been created by the melting glacier. Researchers are also examining the soils and finding that as the plant communities shift from mosses to shrubs to trees, the soil becomes more nutrient rich. As a result, they found that forbs and grasses grow better in soils containing more nutrients. As we will see in this chapter, long-term changes in ecological communities follow predictable patterns, which are important for us to understand the ways in which terrestrial and aquatic communities change over time and the processes that underlie these changes. The rate of glacier retreat. Historic observations by Captain George Vancouver, John Muir, William Cooper, and current researchers indicate that the glacier at Glacier Bay has retreated rapidly during the past 200 years.
Castle, C., et al. 2016. Soil biotic and abiotic controls on plant performance during primary succession in a glacial landscape. Journal of Ecology 104: 1555–1565. Cooper, W. S. 1923. The recent ecological history of Glacier Bay, Alaska II: The present vegetation cycle. Ecology 4: 223–246. Fastie, C. L. 1995. Causes and ecosystem consequences of multiple pathways of primary succession at Glacier Bay, Alaska. Ecology 76: 1899–1916.
Learning Objectives
After reading this chapter, you should be able to:
19.1 Discuss how succession occurs in a community.
19.2 Describe the multiple mechanisms by which succession occurs.
19.3 Explain the ways in which succession does not always produce a single climax community. Because most ecological communities typically experience little change over weeks or months, it is tempting to think of a community as a static collection of species. However, communities commonly experience changes in species composition and relative abundance over longer periods. Such changes in communities over time are especially evident when a community experiences a major disturbance, like the receding glacier at Glacier Bay. Other examples include a field that has been plowed, a forest that has experienced an intense fire, or a pond that has dried during a drought and then refills. In these cases, the community slowly rebuilds and, given sufficient time, it often resembles the original community that existed prior to the disturbance. In this chapter, we will explore how the composition of species in communities changes over time. We will also discuss the evidence that scientists have used to quantify these changes in both terrestrial and aquatic communities and describe how modern studies have altered original hypotheses. Understanding such changes is critical for scientists trying to predict how communities might respond to future environmental disturbances, such as hurricanes and fires, or the results of anthropogenic activity such as logging and mining. Such an understanding is also critical for scientists who try to predict the impacts of larger disturbances such as global climate change.
#### 19.1 Succession Occurs in a Community When Species Replace Each Other over Time

19.1 Succession occurs in a community when species replace each other over time The process of succession in a community is the change in species composition over time. For example, when a field is plowed but not planted, grasses and wildflowers soon colonize it. In climates with sufficient precipitation, the grasses and wildflowers will eventually be replaced by shrubs and then by large trees. Each stage of community change during the process of succession is known as a seral stage. The earliest species to arrive at a site are known as pioneer species. These species typically have the ability to disperse long distances and arrive quickly at a disturbed site. The final seral stage in this process of succession is known as the climax community. A climax community is generally composed of the group of organisms that dominate in a given biome. As we will see, a climax community can be achieved through a variety of different sequences over time. Also, the climax community may continue to experience change. Succession The process by which the species composition of a community changes over time. Seral stage Each stage of community change during the process of succession. Pioneer species The earliest species to arrive at a site. Climax community The final seral stage in the process of succession. We begin this section by considering that succession can be observed either directly or indirectly. We will then look at how we examine successional patterns in a variety of terrestrial and aquatic environments. While most studies of succession focus on the changes in plant communities, there are also associated changes in the species of animals that depend on plants for food and habitat. Much less studied is the succession that occurs on dead organic material, such as rotting logs and animal carcasses.
Observing Succession
Observing succession in a community can take different amounts of time,

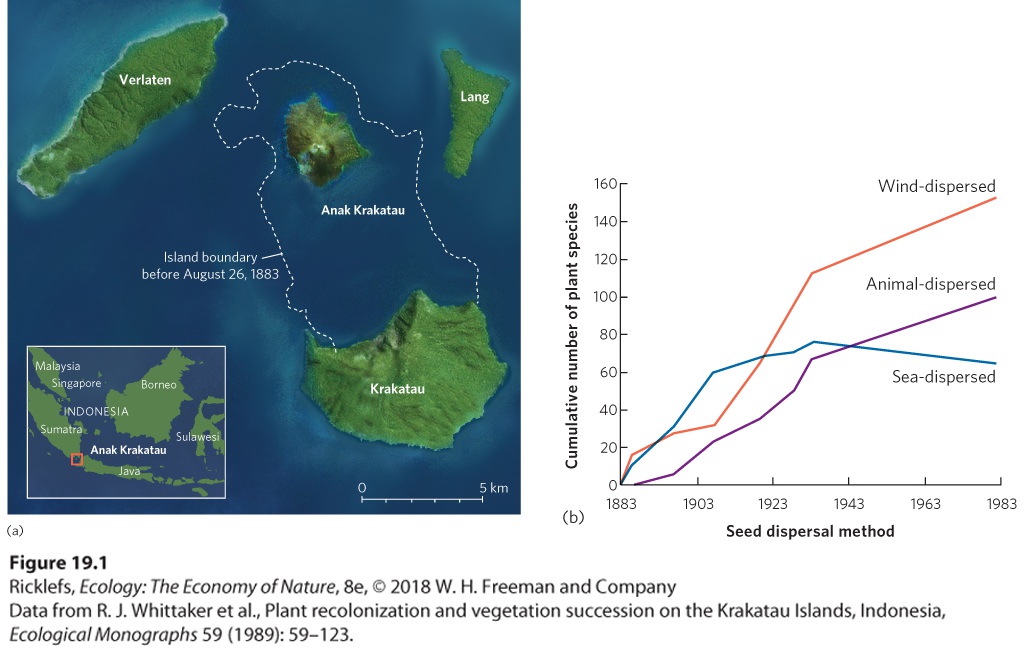
depending on the life histories of the species involved. For example, the succession of decomposers on a dead animal can happen in a matter of weeks or months. In contrast, the succession of a forest can take hundreds of years. When succession occurs over long time horizons, modern scientists can sometimes return to sites that have been studied by others decades or even centuries earlier and observe how the species composition of an area has changed over time. In other cases, when there are no historical data for an area, more indirect methods are used to estimate the pattern of succession. As we will see, taking both direct and indirect approaches to examine succession provides the most complete picture of how communities change over time. Direct Observations The clearest way to document succession in a community is by direct observation of the changes over time. We have already described the research at Glacier Bay, a well-known example of directly observed succession. Another example comes from the small island of Krakatau, Indonesia. In 1883, a massive volcanic eruption on Krakatau blew away nearly threequarters of the island, as depicted in Figure 19.1a. The remaining part of the island was covered with a layer of volcanic ash that obliterated all life. Researchers began to visit the island after the eruption to document how and when various species would return and whether the developing community would eventually be similar to the community that had existed on the island before the eruption. By 1886, 24 species of plants had colonized Krakatau, as you can see in Figure 19.1b. Ten of these species were sea-dispersed plants that are common on tropical shores throughout the region. Many of the other plants were wind-dispersed grasses and ferns whose seeds and spores had been blown to the island from surrounding islands. With more time, the seeds of trees arrived. By 1920, most of the island had developed into a forest community. The presence of forest habitats made the island more hospitable to many species of birds and bats. Bird and bat species that were fruit eaters brought a variety of seeds with them in their digestive systems. Many of the seeds that were excreted while the birds and bats were on the island subsequently germinated. Today, direct observations continue to monitor how succession on the island is affected by additional disturbances, such as volcanic eruptions, erosion of ash deposits by the ocean waves, and strong storms that pass through the region.
Figure 19.1 Direct observations of succession. (a) The island of Krakatau in Indonesia experienced a massive volcano in 1883 that blew away much of the original island and obliterated all life. (b) Since then researchers have documented the colonization of new plant species on the island over time. Initially, most plants colonizing the island as seeds were seaand wind-dispersed. As the forests developed, birds and bats that came to the island carried the seeds of many other species of plants. Succession is much easier to observe when it takes place over short timescales, such as the succession of species that occurs on decomposing organisms. For example, when a mammal dies, the dead tissues are rapidly colonized by bacteria and fungi. Within a few minutes, insects also begin to locate the carcass, often attracted by odors that are emitted by the dead animal. One of the first groups of insects to show up are the blowflies; they lay their eggs on the carcass and these hatch into larvae known as maggots (Figure 19.2). Given that insects are ectotherms, the rate at which the maggots grow, develop, and consume the carcass depends on the environmental temperature. Over time, more species of flies arrive as well as several species of beetles and predators. Once the tissues are consumed and only bones and hair remain, the number of insects drops sharply and the rate of decomposition slows. Scientists have directly observed such successions for centuries, and today this knowledge is also used to estimate the time of death for human carcasses based on the insects found on a body and the environmental temperature. These observations have formed the basis for the field known as forensic entomology.
Figure 19.2. Blowfly. The green bottle fly (Lucilia sericata) is a species of blowfly that is among the first groups of insects to arrive when an animal dies. Indirect Observations Because it is a challenge to directly observe succession in many types of communities, researchers have sought ways to determine the patterns of succession indirectly. The two most common methods attempt to look back in time from the present day. One approach examines regional communities that began succession at different times. For example, classic research by Henry Cowles in the late 1800s examined the succession of sand dunes in Indiana along the southern shore of Lake Michigan. Cowles knew that the water level in Lake Michigan had fallen since the last glaciation and that new sand dunes had formed along the edges of the receding shoreline. At the time Cowles visited Lake Michigan, the lake was surrounded by multiple ridges of sand dunes. The oldest dunes were far away from the current shoreline, while the youngest dunes were close to the shoreline. On these younger dunes, he found scattered plants such as beachgrass (Ammophila breviligulata) and bluestem grasses that represented the earliest stages of succession, much like the wind-dispersed grasses that appeared on Krakatau during the earliest years of succession. Farther away from the water, older dunes contained larger and more abundant plants that included herbs and several species of shrubs. Beyond these plants, still older dunes had pine trees, while the oldest dunes had beech, oak, maple, and hemlock trees (Figure 19.3). These observations led Cowles to propose a technique called chronosequencing, which is used to create a model of the sequence of communities that exist over time at a given location. The model, called a chronosequence, helps ecologists understand how succession has progressed over time in an area.
Many other ecologists subsequently used chronosequencing, including William Cooper, a graduate student of Cowles who examined chronosequences in Glacier Bay.

Figure 19.3 Chronosequences. The dunes of Lake Superior increase in age the farther one moves away from the water. Based on this relationship, researchers have examined the plant life on dunes of different ages as a way of estimating how succession proceeds over time. Chronosequence A sequence of communities that exist over time at a given location. It is also possible to look back in time by examining pollen and other plant parts preserved in distinct layers of lake and pond sediments. Flowering plants produce pollen grains with distinct sizes and shapes. When these pollen grains travel through the air and land on the surface of a lake, they sink and, over time, become preserved in layers of sediment at the bottom of the lake. Researchers can determine the age of each of these layers, by taking
a sample that penetrates through many layers of mud on the lake bottom and then using a technique known as carbon dating that identifies the age of the pollen in each layer. Dating the pollen helps determine changes in the species of plants around the lake over hundreds or even thousands of years.
Succession in Terrestrial Environments
Researchers investigating terrestrial environments have focused primarily on the succession of plant communities. In terrestrial environments, we can categorize succession into two types that are based on their starting conditions: primary succession and secondary succession. In both cases, we will begin by discussing a simplified version of terrestrial succession that is represented by an ordered progression over time. Later in this section, we will see that succession in terrestrial communities can be much more complex. Primary Succession Primary succession is the development of communities in habitats that are initially devoid of plants and organic soil, such as sand dunes, lava flows, and bare rock. These inhospitable environments are colonized by species, such as lichens and mosses, that require no soil, and that can live on the surfaces of rocks, and drought-tolerant grasses that are able to colonize dry sand dunes (Figure 19.4). The species that first colonize these places produce bits of organic matter that combine with the processes of rock weathering and microbial activity to create soils that make the site more hospitable for other species.
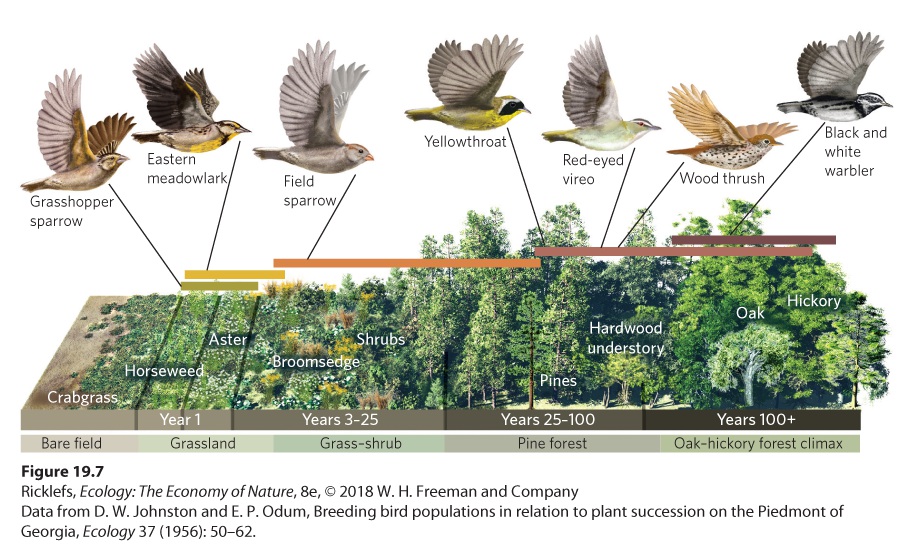
Figure 19.4 Primary succession. Primary succession occurs when organisms colonize sites that are initially devoid of life. (a) Bare rocks, such as these rocks in Wisconsin, are initially colonized by lichens and mosses. (b) Sand dunes, like these dunes along the shores of Lake Michigan, are initially colonized by grasses able to tolerate dry, sandy soils with little organic matter. Primary succession The development of communities in habitats that are initially devoid of plants and organic soil, such as sand dunes, lava flows, and bare rock. Secondary Succession Secondary succession is the development of communities in habitats that contain no plants but do have organic soil. For example, secondary succession occurs in fields that have been plowed or forests that have been
uprooted by a hurricane. Such habitats typically contain well-developed soils. These soils may also include plant roots and seeds, both of which contribute to rapid growth of new plants after the disturbance. Secondary succession The development of communities in habitats that have been disturbed and include no plants but still contain an organic soil. Secondary succession can be observed in a chronosequence of abandoned agricultural fields. An excellent example of this is Duke Forest, an outdoor laboratory in the Piedmont region of North Carolina composed of 1,900 ha of abandoned agricultural fields and forests that Duke University purchased in 1931. To understand the process of succession in this area, ecologist Henry Oosting visited multiple farm sites, each abandoned for different lengths of time. He found a clear pattern of succession that starts with annual plants and ends with large deciduous trees, as illustrated in Figure 19.5. Even before abandonment, crabgrass (Digitaria sanguinalis) is common in the fields. In the first summer after abandonment, crabgrass and horseweed dominate the fields. By the second summer, ragweed (Ambrosia artemisiifolia) and heath asters (Symphyotrichum ericoides) dominate the fields, and by the third summer, broomsedge (Andropogon virginicus) dominates. Next come shrubs, followed by pines. Within about 25 years, pines eventually crowd out earlier successional species. As the decades pass, deciduous tree species arrive and start displacing the pines. After about 100 years, deciduous trees dominate and constitute the last seral stage in the successional sequence.
Figure 19.5 Secondary succession. Secondary succession begins with a well-developed soil. In a temperate seasonal forest, such as this one from the Piedmont region of North Carolina, the climax community is a forest dominated by deciduous trees.
We distinguish between primary and secondary succession by the starting point of the community. For example, in the midwestern United States, an area containing bare rock will pass through seral stages of lichens and mosses, annual weeds, perennial weeds, shrubs, several species of pioneer tree species, and then a beech-maple forest. In contrast, an area containing bare soil will start with annual weeds and then pass through many of the same seral stages on its way to form a beech-maple forest. As we will see later in the chapter, this beech-maple forest can vary somewhat in the composition of dominant tree species. Sometimes the distinction between primary and secondary succession is unclear because the intensity of a disturbance can vary. For instance, because a tornado that levels a large area of forest usually does not harm soil nutrients and there are often seeds and living roots in the soil, succession follows quickly. In contrast, a severe fire that burns through organic layers of the forest soil requires the community to start over from scratch in a way that resembles primary succession; there is soil, but it contains few seeds or roots that can immediately sprout after the fire. The Complexity of Terrestrial Succession A given biome has a characteristic climax community, but the sequence of seral stages through which a single site passes on its way to this climax community can differ, depending on the initial conditions. For example, consider the succession of a field, a sand dune, and a wetland near Lake Michigan in Indiana. The abandoned field is typically colonized by asters that include horseweed and goldenrod. The sand dune is typically colonized by beachgrass and bluestem grasses, which are perennial grasses that can stabilize the sand dunes and add organic matter to the soil. The wetland supports plants such as cattails, which produce organic matter; over many years, this organic matter can fill in the wetland and create conditions that allow terrestrial plants to colonize the site. In all three cases, the progression of seral stages begins with a different community, but ends with the same climax community of a forest dominated by beech and maple trees. Although it is tempting to think that terrestrial succession is a simple linear process, this is often not the case. The sequence of seral stages can be quite variable. For example, the concept of chronosequences relies on the assumption that older sites pass through the same stages as the younger sites.
It also assumes that sites of different ages do not differ in other aspects, such as historic abiotic conditions, soil fertility, and human or natural disturbances. When chronosequences go back hundreds of years, it can be difficult or impossible to confirm that the sites did not differ in ways that affect succession. For example, sites of similar age in a given area sometimes contain important differences in species composition because of local disturbances, such as a tornado. Modern researchers will observe the changes in species composition, but they may not know that a tornado had passed through the area. The research at Glacier Bay provides a good example of the complexity of succession. For decades, ecologists presented a simplified scenario of how the communities in Glacier Bay have changed over time. Based on Cooper’s original observations, they identified primary succession as a linear process that begins with lichens, mosses, and herbs. Next is a seral stage containing low shrubs, followed by a stage containing tall shrubs that include willows, cottonwoods, and alders. The next stage is dominated by spruce trees, and the final seral stage is dominated by hemlock trees. More recent research has used tree rings to date the colonization of successional sequences and has reached a different conclusion regarding the path of succession. The number of growth rings tells us the age of the tree. From the tree’s age, we can determine when the tree first appeared at a site relative to when the retreating glacier opened the site to succession. For example, we now know that the older sites probably never had a seral stage that included cottonwood trees, whereas the younger sites currently pass through a stage of abundant cottonwoods. Furthermore, spruce and hemlock trees rapidly colonized sites that were exposed in the early 1800s, but not sites that were exposed in the late 1800s and early 1900s. You can view the data for Sitka spruce trees in Figure 19.6. This suggests that the soils in the older and younger sites may not have experienced the same changes as they underwent succession.
Figure 19.6 Complex succession. Using tree rings to date the Sitka spruce trees from different sites in Glacier Bay, we can see that older sites experienced rapid colonization by spruce trees after the glaciers retreated, whereas the younger sites experienced slow colonization by spruce trees after the glaciers retreated. Vertical red lines indicate the approximate year of glacial retreat at each site. Because the assumptions underlying chronosequences may not always be supported, the best approach is to use a combination of data and research methods, including chronosequences, pollen records, and long-term studies of single locations undergoing succession. Collectively, these studies provide the most accurate description of terrestrial succession. Animal Succession Ecologists have traditionally focused on changes in plant species when describing succession in terrestrial environments. However, the changes in the plant community cause substantial changes in the habitats that are available to animals, which in turn cause changes in the animal community. For example, a classic study by David Johnston and Eugene Odum examined the distribution of bird species in the Piedmont region of Georgia, along the same successional seral stages that Oosting surveyed in the Piedmont region of North Carolina. As illustrated in Figure 19.7, grasshopper sparrows (Ammodramus savannarum) and eastern meadowlarks (Sturnella magna) dominate the early-succession stages that contain annual plants. With the colonization of shrubs into the abandoned fields comes the arrival of many different bird species, including field sparrows (Spizella pusilla) and yellowthroats (Geothlypis trichas). As we move from the pine forest to the oak-hickory climax forest, other species appear that include the red-eyed vireo (Vireo olivaceus) and the wood thrush (Hylocichla mustelina). Although some species of birds specialize on a narrow range of the plant successional stages, many species inhabit multiple seral stages.
Figure 19.7 Animal succession. As the plant community experiences secondary succession, the habitat available to birds changes. In response to changes in the habitat, the species of birds that live in the community change.
Succession in Aquatic Environments
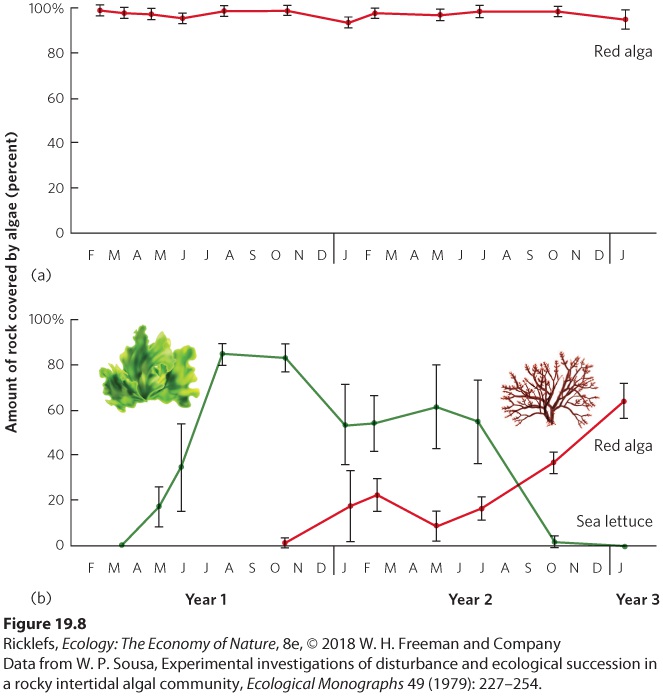
Succession also occurs in aquatic environments. In this section, we will examine how succession proceeds in three types of aquatic environments: intertidal zones, streams, and ponds. Intertidal Communities In contrast to most terrestrial communities, succession in intertidal communities can occur much more quickly, in part because the generation time of the dominant species is much shorter. In intertidal communities, powerful waves that occur during storms commonly remove organisms that are attached to boulders. For example, ecologist Wayne Sousa examined the succession of different species of algae on boulders of the intertidal zone in a classic experiment in Southern California. He examined some boulders that were not overturned by storms and others that had been overturned and had areas of bare rock exposed that could be colonized by algae. As shown in
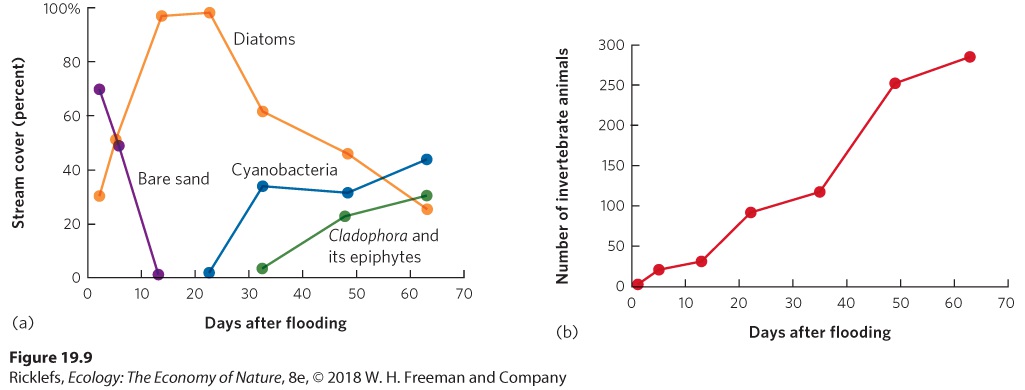
Figure 19.8, Sousa observed that the first species to arrive was a green alga known as sea lettuce (Ulva lactuca). Over the next year, sea lettuce came to dominate the rocky habitat and largely prevented a competing species of red alga, Gigartina canaliculata, from colonizing. However, as the sea lettuce became more dominant, it attracted crabs that eat it, which cleared areas on the boulders for the less edible red algae to colonize. Over time, red algae
Figure 19.8 Intertidal succession. (a) On undisturbed boulders in the intertidal zone, communities are dominated by a species of red alga, shown in the drawing on the left. (b) When strong waves overturn boulders, bare areas of rock are exposed and quickly colonized by sea lettuce, shown in the drawing on the right. Sea lettuce initially dominates the bare rock surface, as measured by the percent of the rock covered by the algae. As time passes, crabs begin to consume the sea lettuce, which creates space that allows the red algae to colonize the rock and ultimately dominate the boulder’s surface. Error bars are standard errors. Stream Succession Like intertidal habitats, streams also experience rapid succession largely because organisms can move downstream from sites that are less disturbed. Streams can experience major disturbances during heavy rainfall that increases both the volume of water the streams carry and the speed at which the water moves. With the greater speed of the rushing water, sand and rocks can tumble downstream and wipe out most plants, animals, and algae. In one
study, researchers looked at the effects of a flood event in Sycamore Creek in Arizona. The floodwater scoured the creek and eliminated nearly all of the algae and 98 percent of the invertebrates, leaving behind rocks and bare sand. The researchers then monitored how the community changed over the subsequent 2 months; you can view their data in Figure 19.9. Within just a few days after the flood, the stream became colonized with several species of algae known as diatoms. Within 5 days, the diatoms covered nearly 50 percent of the creek bottom, and after 13 days, the diatoms covered nearly 100 percent. After 3 weeks, cyanobacteria started to colonize the stream, followed by a species of filamentous green alga (Cladophora glomerata) along with associated diatoms that live as epiphytes on this green alga. As the three types of algae rebounded, adult insects from the surrounding terrestrial environment began to lay eggs in the stream. This returned a diverse group of larval insect species to the stream, which remained there until they metamorphosed into terrestrial adults.
Figure 19.9 Stream succession. After a major flood event, Sycamore Creek in Arizona experienced a nearly complete elimination of algae, leaving behind only rocks and bare sand. (a) Within just a few days, several species of diatoms came to dominate the bare sand and rocks in the stream. Later in the summer, other groups of algae colonized the stream and became more abundant, including cyanobacteria and a species of green filamentous alga (Cladophora) and its epiphytes. (b) As the different types of algae returned, adult flying insects from the terrestrial environment began to lay their eggs in the stream. As a result, the stream experienced a rapid increase in the number of invertebrate animals. Lake Succession For decades, ecologists have explained pond and lake succession using a paradigm of slow transformation, as depicted in Figure 19.10a. If we imagine a pond or shallow lake created by a receding glacier thousands of years ago or a beaver dam converting a stream into a pond, we start with a basin full of
water. Over time, the erosion of soil and the growth and death of organisms in the lake form sediments that gradually fill the basin. In addition, plants living along the shoreline slowly extend themselves out into the water to form a floating mat of vegetation. Underneath this live mat of vegetation is an accumulating layer of partially decomposed vegetation known as peat. Eventually, the entire basin has a floating mat of vegetation that continually contributes detritus to the peat layer in the basin. Microbial decomposition of the dead vegetation is slow because the water underneath the floating mat has little oxygen. As a result, detritus accumulates on the bottom of the basin, and over hundreds or thousands of years, the pond or lake becomes a bog. In North American bogs, sphagnum moss, sedges, and shrubs—such as leatherleaf and cranberry—become established along the edges and add to the development of a soil with progressively more terrestrial qualities. At the edges of the bog, shrubs may be followed by black spruce (Picea mariana) and tamarack trees (Larix laricina), which eventually give way to birch, maple, and fir trees, depending on the locality. In short, the classic explanation for lake succession was that it happened very slowly over long periods of time.
Figure 19.10 Succession in shallow lakes and ponds. (a) The classical explanation for succession in these habitats describes a gradual and steady accumulation of organic matter that eventually fills in the basin and converts it to a terrestrial habitat. (b) More recent studies have demonstrated that the process can occur in occasional large bursts when multi-year droughts allow vegetation to extend along the dry part of the basin. When water becomes
abundant again, the extended vegetation floats on the water’s surface and grows in thickness. Multiple drought events allow the vegetation to expand; eventually, it covers the water’s surface and fills in the basin. Recently, researchers proposed a new model of pond and lake succession. This model is based on carbon dating of core samples of a bog to determine when each plant species lived in the area. Contrary to the classic model of slow and continuous succession, the study illustrated that ponds and lakes can experience long periods of several hundred years in which little succession occurs, followed by brief episodes of rapid change. During times of prolonged drought that last for a decade or more, water levels drop and vegetation grows down onto the newly exposed shoreline. As you can see in
Figure 19.10b, when the drought ends and the water level rises again, the mat of vegetation releases its hold from the lake bottom and floats up to the water’s surface. With continued growth of the vegetation, this floating mat becomes thicker and deposits dead organic matter into the water below it. For example, after studying a 16-ha bog in Pennsylvania, it was estimated that 50 percent of the former lake became covered by bog vegetation in just a few decades during a severe drought in the late sixteenth century. In short, lake succession does not always have to be slow and steady; it can happen in brief bursts during rare, but prolonged, periods of drought.
Change in Species Diversity
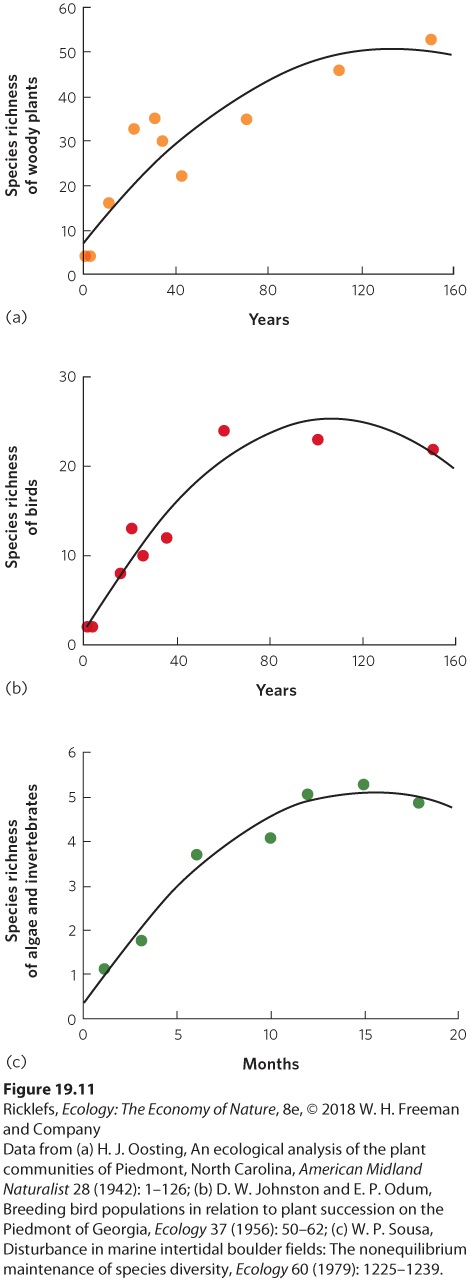
Across both terrestrial and aquatic habitats, the process of succession exhibits consistent effects on species richness. In most cases of succession, we begin with few or no species and then species richness increases rapidly at first. This is followed by a plateau and a small decline, as shown in Figure 19.11. Oosting’s survey of the Duke Forest, for example, found a rapid increase in the species richness of woody plants during the first 25 years and then a gradual decline in the rate of increase over the next 125 years. Similar patterns of richness over time can be seen in the number of bird species observed by Johnston and Odum in a forest community, and the number of algae and invertebrates observed by Sousa in an intertidal community.
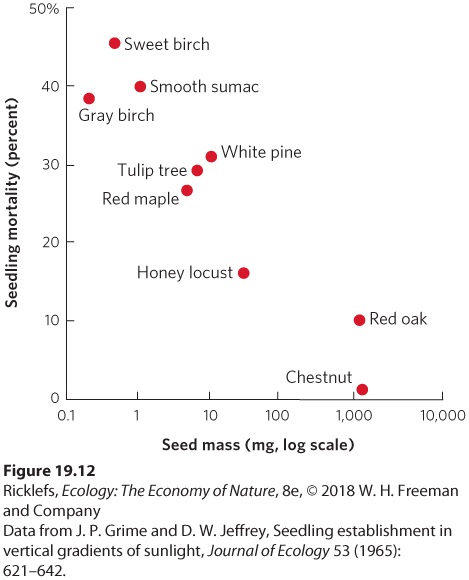
Figure 19.11 Succession effects on species richness. Across a variety of communities, succession is associated with a rapid increase in species richness that slows over time and eventually plateaus. Examples shown here include (a) woody plants in old fields in North Carolina, (b) birds in old fields in Georgia, and (c) algae and invertebrates from the intertidal boulders of southern California.
Concept Check
1. What is the fundamental difference in primary and secondary succession? 2. Why is it difficult to directly observe succession in most ecological communities? 3. How are chronosequences used to understand succession?
#### 19.2 Succession Can Occur Through Different Mechanisms
19.2 Succession can occur through different mechanisms Now that we have an idea of what succession looks like across a variety of communities, we can explore how succession actually occurs. In this section, we consider the traits of early- and late-succession species, compare the different mechanisms of succession, and then examine succession studies to determine which mechanisms are common in ecological communities.
Traits of Early- Versus Late-Succession Species
Early-succession and late-succession species possess different traits important to their performance. For example, pioneer species of terrestrial plants are typically better at dispersing seeds to newly created or disturbed sites. They produce many small seeds that are easily dispersed by wind or that stick to passing animals. These seeds can also persist in the soil for years and then germinate when a disturbance occurs. When they do germinate, these early-succession plants invest more resources into their shoots than their roots and grow rapidly and reproduce quickly. They are also typically quite tolerant of the harsh abiotic conditions that can exist in newly disturbed sites, including full sun and widely fluctuating temperatures and water availability. However, they are not tolerant of the high-shade conditions of late-succession plant communities. In contrast, climax species produce a relatively small number of large seeds that disperse poorly; some simply drop to the ground, whereas others are consumed by animals. The seeds have a relatively short viability, and once they germinate, they grow slowly, but their shade tolerance as seedlings and their large size as mature plants give them a competitive edge over earlysuccession species. As succession progresses, we see a shift in the balance between adaptations that promote dispersal, rapid growth, and early reproduction, and adaptations that enhance competitive ability. Table 19.1 summarizes the traits of early-succession and late-succession plants.
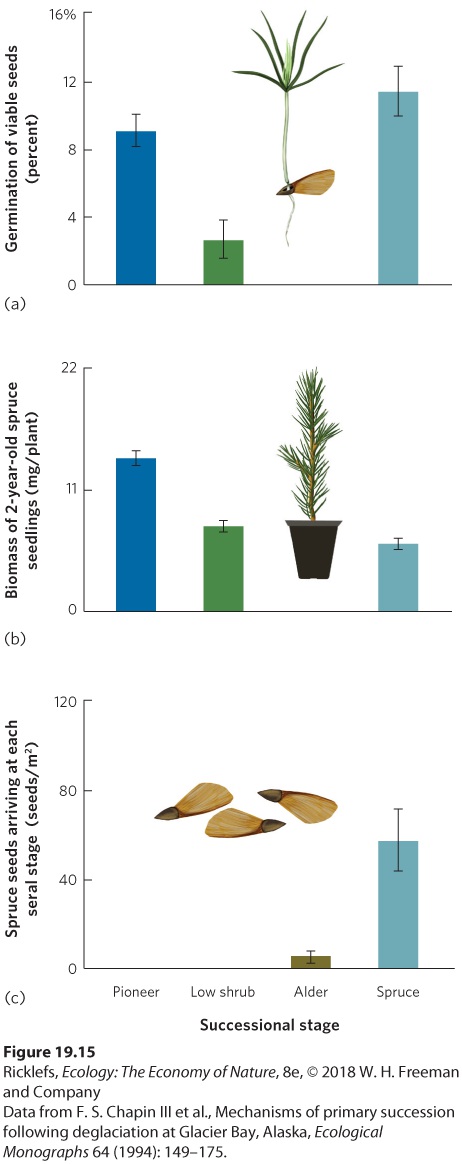
TABLE 19.1 General Characteristics of Early-Succession and Late-Succession Plants Trait Early Succession/Pioneer Species Late Succession/Climax Species
##### Analyzing Ecology: Quantifying Community Similarity
Number of seeds Many Few Seed size Small Large Mode of dispersal Wind or stuck to animals Gravity or eaten by animals Seed viability Long Short Root:shoot ratio Low High Growth rate Fast Slow Size at maturity Small Large Shade tolerance Low High
Analyzing Ecology
Quantifying Community Similarity When ecologists examine the species living in different communities, such as when examining chronosequences, they often quantify species abundance and richness. Although such data tell us about the species living in each community, they do not provide a measure of comparison between communities. To address this need, we use several indices of community similarity that can range from zero to one; a value of zero indicates that two communities have no species in common, whereas a value of one indicates that two communities have an identical composition of species. One of the most common ways to quantify similarity is Jaccard’s index of similarity, developed by the Swiss botanist Paul Jaccard in 1901. Jaccard’s index is calculated using the following equation: J=XA+B+X where A represents the number of species that are only present in Community A, B represents the number of species that are only present in Community B, and X represents the number of species present in both communities. For example, consider the table below that lists the species of fish found in each of three stream communities that are at different
stages of succession. We can now use Jaccard’s index to calculate the similarity between Community A and Community B: J=XA+B+XJ=31+4+3J=0.33 This value indicates that there is relatively low similarity in the species composition of Communities A and B. YOUR TURN Use Jaccard’s index to calculate the similarity between Communities A and C and between Communities B and C. Based on these calculations, which communities are the most similar to each other? Fish Species Stream Community A Stream Community B Stream Community C Rainbow Trout X X Brook Trout X X Brown Trout X X Mudminnow X Common Shiner X X Creek Chub X White Sucker X X Johnny Darter X X Smallmouth Bass X Mottled Sculpin X X The traits of early-succession and late-succession plants are different because they face inherent trade-offs, similar to the life history trade-offs that we discussed in Chapter 8. To test for trade-offs between early-succession and late-succession species, researchers examined the mass of a single seed from each of nine species of trees. They raised the seeds under low-light conditions to simulate the high amount of shade in mature forests. After 12
weeks, they examined the mortality of the germinated seedlings, which you can view in Figure 19.12. The large-seeded species, which are common in old forests, had low rates of mortality under low-light conditions since large seeds provide their seedlings with ample nutrients to get started in the lightlimited environment of the forest floor. In contrast, small-seeded species, which are common pioneer species that disperse well in large numbers, had high rates of mortality under low-light conditions. As a result, pioneer species cannot establish in mature forests.
Figure 19.12 Trade-offs between seed mass and seedling mortality under high-shade conditions. Species with large seeds, which are common in late-succession forests, produce seedlings that survive well in high shade. Species with small seeds, which are common in early-succession forests, produce seedlings that have a poor survival rate in high shade.
Facilitation, Inhibition, and Tolerance
The ability to disperse and the ability to persist under existing abiotic and biotic conditions determine which species will appear in different seral stages during succession. Organisms that disperse well and grow rapidly have an initial advantage and therefore dominate the early stages of succession. Species that disperse slowly or grow slowly once they colonize an area

typically become established later in succession. Early-succession species can also modify the environment in ways that affect whether late-succession species can become established. We must therefore consider whether a species has a positive, negative, or neutral effect on the probability of a second species becoming established. The mechanisms can be categorized as facilitation, inhibition, and tolerance. Facilitation is a mechanism of succession in which the presence of one species increases the probability that a second species can become established. Early-succession species do this by altering the environmental conditions of the site in a manner that makes it more suitable for other species to establish and less suitable for themselves. For example, alder shrubs, which are legumes, live in a mutualistic relationship with nitrogenfixing bacteria in their roots, as we discussed in Chapter 17. This relationship produces additional nitrogen in the soil, which facilitates the establishment of nitrogen-limited plants such as spruce trees. Over time, the spruce trees eventually grow tall and cast a deep shade that is not a favorable environment for the alder shrubs. Facilitation A mechanism of succession in which the presence of one species increases the probability that a second species can become established. Inhibition is a mechanism of succession in which one species decreases the probability that a second species will become established. Common causes of inhibition include competition, predation, and parasitism. That is, individuals of one species can inhibit those of other species by outcompeting them for resources, eating them, or attacking them with noxious chemicals or antagonistic behavior. Early in succession, inhibition can prevent movement toward a climax community, while late in succession, inhibition can prevent the pioneer species from colonizing and surviving. For example, in a mature forest in the northeastern United States, adult maple and beech trees cast a deep shade that prevents pioneer tree species from surviving. Inhibition A mechanism of succession in which one species decreases the probability that a second species will become established.
When inhibition occurs in a seral stage, the outcome of an interaction between two species depends on which species becomes established first. The arrival of one species at a site that affects the subsequent colonization of other species is known as a priority effect. We can see an example of the priority effect in the subtidal habitats of South Australia with bryozoans, a group of tiny invertebrate animals that live in colonies attached to rocks and that feed by filtering the water (Figure 19.13). If bryozoans become established first, they can prevent the establishment of tunicates and sponges—two other groups of filter-feeding animals that attach themselves to rocks. Sometimes the priority effect occurs because the first species to arrive has grown to a competitively superior adult stage, whereas the second species to arrive is in the competitively inferior immature stage. For example, if a beech tree gets established in a forest and grows to an adult stage, it shades the ground below, preventing the seedlings of most other tree species from conducting sufficient photosynthesis to survive. In short, the mechanism of inhibition can make the path of succession dependent on which species arrives at the site first.
Figure 19.13 Bryozoans. Bryozoans, such as these colorful individuals from Australia, are tiny invertebrate animals that live attached to rocks in the ocean. If they colonize the rocks first, they can prevent colonization by tunicates and sponges, which compete for space on the rocks. Priority effect When the arrival of one species at a site affects the subsequent colonization of other
species. Tolerance is a mechanism of succession in which the probability that a species can become established depends on its dispersal ability and its ability to persist under the physical conditions of the environment. For example, species that can tolerate the stressful environmental conditions of early succession—such as low moisture or more extreme temperature fluctuations —can become established quickly and dominate early stages of succession. Similarly, plants that can tolerate high-shade environments can become established in forests that cast deep shade. These species do not alter the environment in ways that either help or inhibit other species, but once a stress-tolerant species becomes established, it can be affected by interactions with other species. For example, superior competitors that arrive later will eventually replace the stress-tolerant species. Tolerance A mechanism of succession in which the probability that a species can become established depends on its dispersal ability and its ability to persist under the physical conditions of the environment.
Tests for the Mechanisms of Succession
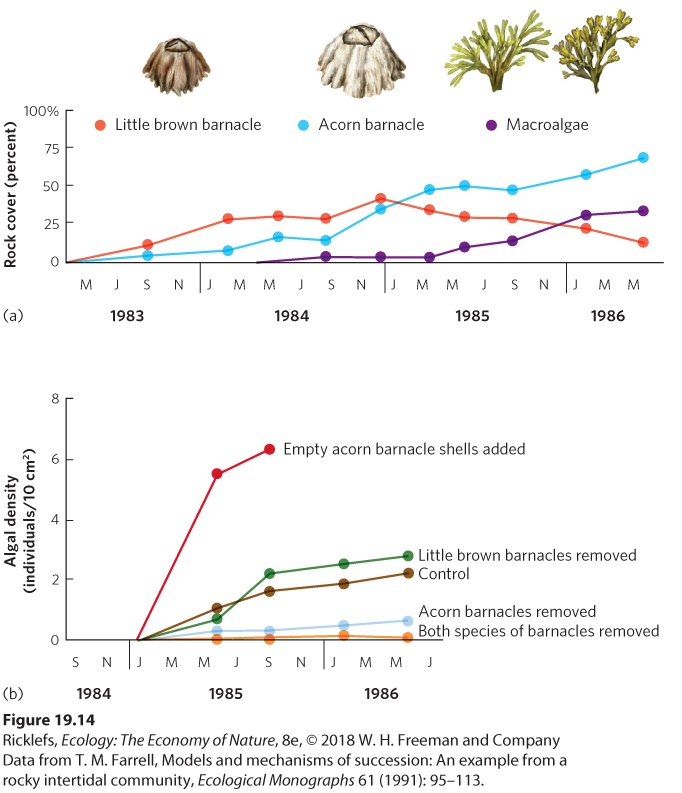
For many years, ecologists debated which of the three succession mechanisms were the most important determinants of the pattern of species replacement over time. Knowing the dominant mechanisms would allow scientists to predict how changes in communities proceed over time, especially since succession is often not a simple linear progression toward a climax community. To address this question, a great deal of research had to be done across a variety of biomes. Here, we discuss two such studies: one conducted in an intertidal community and one conducted in a forest community. Succession in Intertidal Communities Sousa’s research in Southern California showed that green algae prevented the colonization of red algae (see Figure 19.8). This suggested that the succession of intertidal communities is determined by inhibition. However, the intertidal communities off the Oregon coast consist not only of large species of algae—known as macroalgae—but also of several common species

of invertebrates that include the little brown barnacle (Chthamalus dalli), the common acorn barnacle (Balanus glandula), and several species of limpets, which are gastropods that consume algae. Observations of these communities showed that they were dominated by the common acorn barnacle and a brown macroalgae, Pelvetiopsis limitata. To determine how these communities came to be dominated by the two species, researchers scraped areas of rock clean and watched the succession of species over the subsequent 2.5 years. As illustrated in Figure 19.14a, the bare rocks were first colonized by the little brown barnacle. However, as more time passed, the larger acorn barnacle became established on the sites. As it became established, it slowly crushed the smaller little brown barnacle. Over time, the little brown barnacle became rare. Thus, the colonization of the acorn barnacle fit the tolerance model, but the decline of the brown barnacle fit the inhibition model. As the acorn barnacle became abundant, numerous species of macroalgae colonized the site and also became abundant. In fact, after 3 years, communities in the cleared sites closely resembled the control sites that had not been cleared.
Figure 19.14 Tests of succession mechanisms in an intertidal community. (a) When rocks were scraped bare of all organisms, the first species to dominate was the little brown barnacle. Over time, the acorn barnacle became more abundant, followed by an increase in several species of macroalgae. (b) To determine which mechanisms were responsible for these successional changes, the presence of each barnacle species was manipulated. The acorn barnacle was critical to facilitate the macroalgae, whereas the little brown barnacle was not. Which mechanisms of succession allowed the increase of macroalgae? To answer this question, researchers removed different species from the community. They knew that although the little brown barnacle would be the first to arrive due to its superior dispersal ability, the acorn barnacle would eventually arrive and be a superior competitor for space. They hypothesized that the presence of the acorn barnacles facilitated the colonization and survival of the macroalgae. The researchers began the experiment by once
again scraping multiple sites clear of all organisms. Each site then received one of five manipulations: (1) no subsequent removal of any organisms, (2) removal of any little brown barnacles that colonized, (3) removal of any acorn barnacles that colonized, (4) removal of both species of barnacles that colonized, or (5) attachment of empty acorn barnacle shells. The final manipulation was used to test whether the mere physical presence of the acorn barnacle facilitated the colonization and growth of macroalgae by providing the macroalgae with protected crevices to which they could attach. The density of the macroalgae was then monitored for 2 years. The results of this experiment are shown in Figure 19.14b. Starting near the bottom of the figure, you can see that the removal of the acorn barnacle, or both species of barnacles, caused a lower abundance of macroalgae than the control. In other words, the acorn barnacle helped the macroalgae establish itself. In contrast, the removal of the little brown barnacle had little effect on the density of the macroalgae compared to the control. This confirmed that the little brown barnacle neither helps nor inhibits the macroalgae. However, when empty acorn barnacle shells were added to an intact community that already included live acorn barnacles, there was a large increase in macroalgae. Additional experiments revealed that the acorn barnacle helps the macroalgae by providing crevices where young macroalgae can attach to the rocks without being consumed by herbivorous limpets. Collectively, these experiments in the intertidal zone indicate that the little brown barnacle persists because it is a good disperser that can rapidly colonize the frequently disturbed intertidal rocks and tolerate the conditions present on a bare rock. This is an example of tolerance. However, once the acorn barnacle arrives, it outcompetes the little brown barnacle, which is an example of inhibition. Finally, the succession of the macroalgae depends on the acorn barnacle, which is an example of facilitation. Thus, a given community can include all three mechanisms of succession. Succession in Forest Communities Like intertidal communities, forest communities can also exhibit a mixture of successional mechanisms. At Glacier Bay, for example, it was long hypothesized that each seral stage facilitates the establishment of species in the subsequent stage. This made sense because the soils of later seral stages
contained more organic matter, nitrogen, and moisture. If facilitation were the most important mechanism of succession, we would expect that a given species would grow and survive well in a seral stage where it dominates, but have a difficult time growing and surviving in earlier stages. To test this hypothesis, a group of researchers planted spruce seeds and seedlings in four seral stages of Glacier Bay: an early-succession pioneer stage containing lichens, mosses, and herbs; a low-shrub stage dominated by Drummond’s avens (Dryas drummondii); an alder stage dominated by dense thickets of tall alder shrubs (Alnus sinuata); and a spruce stage dominated by spruce trees. You can see the results in Figure 19.15a. When spruce seeds were planted in bare plots within each seral stage, germination of these seeds was high in the pioneer stage, low in the low-shrub and alder stages, and high in the spruce stage. When spruce seedlings were planted in plots within each seral stage, seedling growth was high in the pioneer and low-shrub stages, none survived in the alder stage, and growth was low in the spruce stage, as shown in Figure 19.15b. In short, the spruce seeds and seedlings germinated and grew quite well in the earliest seral stages at Glacier Bay, yet spruce was a rare plant in these early stages. Moreover, whereas spruce seeds and seedlings grew well in the low-shrub and alder stages, seedling growth was inhibited in the spruce stage.
Figure 19.15 Testing the mechanism of spruce succession. Across four successional seral stages in Glacier Bay, Alaska, researchers examined the (a) germination success of planted spruce seeds and (b) biomass of 2-year-old spruce seedlings. They also measured (c) the number of spruce seeds arriving naturally at each seral stage. These data demonstrate
that while spruce trees can germinate and grow well in earlier seral stages, they are absent from the early seral stages because very few seeds are dispersed to these sites. Error bars are standard errors. These observations refuted the hypothesis that each seral stage facilitates the next stage, but it raised the question of why spruce trees were not common in the earlier stages. Researchers decided to measure how many spruce seeds were dispersed by wind to each site. They discovered that very few spruce seeds arrived to colonize the pioneer and low-shrub stages. There were many more spruce seeds dispersed into the alder stage and even more dispersed into the spruce stage, as shown in Figure 19.15c. The researchers concluded that the dominance of spruce in later seral stages had little to do with facilitation or inhibition, but was due to differences in how many dispersing spruce seeds arrived in each location.
Concept Check
1. Why are early-succession species typically better at dispersing than late-succession species? 2. Why are priority effects considered part of the inhibition mechanism of ecological succession? 3. How did the spruce experiments determine that spruce trees did not depend on the facilitation by early-succession species?
#### 19.3 Succession Does Not Always Produce a Permanent Single Climax Community
19.3 Succession does not always produce a permanent single climax community The traditional view of succession was that a series of stages end with a climax community, which remains constant over space and time unless a major disturbance occurs. As we saw in the case of sand dune succession around Lake Michigan, a site can follow different pathways of succession and end up at the same climax community. However, the species composition of a climax community can still exhibit variation over space and time within a given biome. Moreover, a climax community can be short-lived if a disturbance wipes it out. In this section, we will examine how climax communities can change over time and how their composition varies along environmental gradients
Changes in Climax Communities over Time
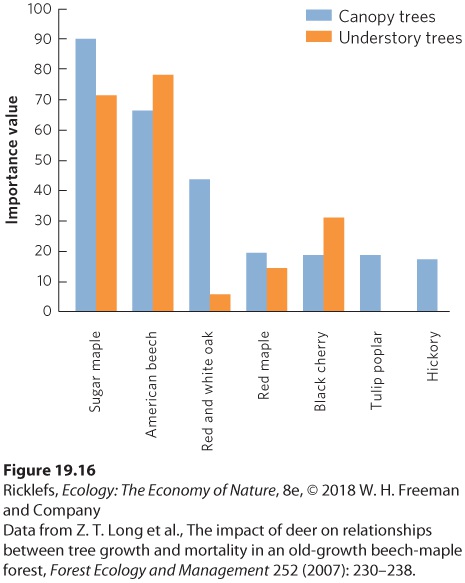
When succession occurs in a community, we typically observe changing environmental conditions and a progression from small life forms to large life forms. For example, primary succession on land begins with lichens and mosses and progresses to grasses and herbs. When sufficient moisture is available, as is the case in eastern North America, succession can continue to a stage that includes large trees. As succession occurs, the abiotic conditions are rapidly altered; areas with trees have less light at ground level, lower ground temperatures during hot summer days, and higher soil moisture. However, once a point is reached where the community contains the largest plants it can support, changes in environmental conditions occur more slowly. As a result, the changes in the community become less dramatic once the climax community develops. When environmental conditions become relatively stable, the composition of plant species that dominate the community also becomes relatively stable. However, the species found in a climax community can continue to change. For example, northern deciduous forests have a climax community dominated by large trees, but the composition of large tree species can slowly change over time. Initially, these large trees are mostly oak, hickory, and tulip poplar (Liriodendron tulipifera). However, over time the dominant species can change to sugar maple and beech. In an old-growth forest in
Pennsylvania, researchers surveyed the adult trees that lived in the canopy and the sapling trees that lived in the understory. They then calculated the importance value of each tree, which incorporates both the abundance and total area of the trunks for each species. As you can see in Figure 19.16, the canopy contained a variety of species with high importance values that included sugar maple, American beech, tulip poplar, several species of oaks, and hickories.
Figure 19.16 Changing species composition in a climax forest. There are many species of trees in the canopy of a forest in Pennsylvania, as indicated by the importance value of each species. However, in the understory, there are few oak trees and no tulip poplar or hickory trees because when they are young, these species are not tolerant of deep shade and they are susceptible to herbivory by deer. This suggests that the future forest canopy will experience a substantial change in the composition of dominant trees. In contrast to this adult distribution, there were few oaks and no tulip poplars or hickories in the understory. The oaks, tulip poplars, or hickories were not in the understory because these species are not very tolerant of the deep shade that occurs in a mature forest and they are also susceptible to being eaten by deer. In comparison, maple and beech trees are very tolerant of shade and less susceptible to being eaten by deer. As a result, maple and
beech trees can survive and grow in the understory of large trees, while other species cannot. Over time, the climax community of large trees will experience a gradual shift in the composition of the dominant species; as the current canopy species gradually die, there will be no younger oaks, tulip poplars, or hickories to replace them. In short, the composition of a climax community of large trees in a northern deciduous forest can continue to change over time.
Changes in Climax Communities over Space
We have seen that the composition of a climax community can vary over time. Climax communities can also vary in composition as one moves along environmental gradients. For instance, in the 1930s, plant ecologists described the climax vegetation of much of Wisconsin as a sugar maple– basswood (Tilia americana) forest. However, they later determined that the climax forest exhibited differences in various locations around the state. In the southern part of the state, beech trees were more common, while in the north, birch, spruce, and hemlock trees were more common. In the drier regions bordering prairies to the west, oaks became prominent. On drier upland sites, quaking aspen, black oak, and shagbark hickory—long recognized as successional species on moist, well-drained soils—came to be accepted as climax species.
Transient Climaxes
Sometimes a climax community is not persistent, a phenomenon known as a transient climax community. A transient climax occurs when a site is frequently disturbed so that the climax community cannot continue to perpetuate itself. A common example of a transient climax occurs in small wetlands, sometimes known as vernal pools, which fill with water each spring and then either dry up in summer or freeze solid in winter (Figure 19.17). Although drying and freezing events eliminate most species that comprise a pond community, some species have resting stages and persist in the soil until the wetland refills again in the spring. For example, many species of zooplankton produce resting eggs that can persist in the dry bottom of a wetland and then hatch when the wetland fills with water again. Similarly, some species of snails can aestivate, as discussed in Chapter 4, which allows them to live under the soil surface of the dry pond with their
metabolic processes largely shut down. When the water returns, plants, animals, and microbes come back to life from their resting stages. Many other species that live on land as adults, such as frogs, salamanders, aquatic beetles, and dragonflies, lay their eggs in the wetland. In this way, the community once again starts the process of succession, only to be destroyed by summer drying or winter freezing.
Figure 19.17 Transient climaxes. Vernal pool communities undergo rapid succession of the aquatic community, but (a) the climax community that is achieved throughout the spring and summer is frequently destroyed by (b) drying in the late summer or freezing in the winter. This vernal pool is in the French Creek Wildlife Area of Wisconsin. Transient climax community A climax community that is not persistent.
Creating Gaps in A Climax Community
Sometimes climax communities contain species that are not considered climax species. These species fill relatively large gaps created by small-scale disturbances in an area. In mature forests, for example, adult trees eventually die and leave a gap in the canopy that lets in sunlight (Figure 19.18). If the gap is not large, the surrounding branches of neighboring trees will likely grow in and close the gap. However, if the gap is large, the area of intense sunlight provides local conditions favoring species from earlier seral stages that have widely dispersed seeds and an ability to grow rapidly under high sunlight conditions. As a result, a mature forest that contains mostly climax species of trees can also contain a few early-succession trees. Gaps can occur in a variety of terrestrial and aquatic biomes. For example, in Chapter 18, we saw how gaps of increasing size in the intertidal zone favored the formation of alternative stable states (see Figure 18.23).
##### Climax Communities under Extreme Environmental Conditions
Figure 19.18 Gaps in a climax community. When a large tree dies and falls to the ground, it opens a gap in the forest that allows large amounts of light to reach the forest floor and favors the growth of species from earlier seral stages. This photo is from the Corcovado National Park in Costa Rica.
Conditions
As we have seen throughout this chapter, the composition of a climax community is determined by the environmental conditions that develop over time, including temperature, light, nutrients, and moisture. In some areas, however, additional environmental conditions also play a role, such as in communities affected by fire or grazing. Biomes in which fires occur at regular intervals favor the persistence of fire-tolerant species. For example, in Chapter 16, we saw that the pine forests in the southeastern United States experience periodic fires that kill oak trees and other species of broadleaf trees, but not pine trees. In fact, some species of pines do not even release seeds from their cones unless triggered by the heat of a fire. After a fire, pine seedlings grow rapidly because there is little or no competition from other understory species. As a result, forest succession reaches a climax that is dominated by pine trees. When a successional stage persists as the final seral stage due to periodic fires, we call it a fire-maintained climax community. Fire-maintained climax community
A successional stage that persists as the final seral stage due to periodic fires. The chaparral vegetation found in California is another community that has a fire-maintained climax. As we discussed in Chapter 6, the California chaparral is an example of the woodland/shrubland biome, which has cool, wet conditions in the winter and hot, dry conditions in the summer. As a result, plants can produce thick layers of detritus that become very susceptible to fires during the dry summers. When a successional stage persists as the final seral stage due to intense grazing, it is called a grazer-maintained climax community. Grazers can create a different climax community because they preferentially consume the most palatable plants and do not consume the less palatable or betterdefended plants. In the dry grasslands of Arizona, for example, intense cattle grazing can kill or severely damage many species of grass and leave behind less palatable plants, such as mesquite and cactus (Figure 19.19).
Figure 19.19 Grazer-maintained climax. In areas of dry grassland, such as this site in Catalina, Arizona, intense grazing eliminates many species of grasses and favors the growth of cactus and mesquite trees. Grazer-maintained climax community When a successional stage persists as the final seral stage due to intense grazing. In western North America, cattle grazing allows invasion by cheatgrass (Bromus tectorum), a grass native to Europe, Asia, and North Africa that was introduced to North America in the late 1800s. Cheatgrass is able to colonize
when grazing removes much of the competing grasses. Once it establishes, the detritus it produces is quite susceptible to fire. In fact, areas dominated by cheatgrass burn every 3–10 years, rather than the natural fire cycle of every 30–70 years. These more frequent fires promote the long-term persistence of cheatgrass as well as making the area more susceptible to other invasive plant species. In this case, the change in climax community is maintained by a combination of grazing and fire. Throughout this chapter, we have discussed how communities change over time after a major disturbance. We have emphasized that communities are always changing, both when we move from early to late seral stages and even when a community appears to have attained a climax state. By observing both what happens after a disturbance and the underlying mechanisms that cause the changes, we obtain new insights into the processes that regulate the structure of communities. It is also important to remember that the climate that influences succession is also changing, which means that succession toward a climax stage is actually heading toward a perpetually moving target.
Concept Check
1. What evidence is there that climax communities can continue to change over time? 2. How do grazers cause changes in the composition of the climax community? 3. Why is a vernal pool an example of a transient climax community?
Concepts
Strip mine succession. When strip mines are no longer used for the extraction of coal, grasses are commonly planted to rapidly hold the soil and prevent erosion, such as at this site in Muskingum County, Ohio. Over time, other plants will slowly colonize the area and succession will occur. Coal has long been a major source of energy in North America and Europe. For several centuries, humans have mined coal near the surface of the ground by removing the upper layers of soil and rock and then digging out the coal underneath. This process, known as strip mining, surface mining, or mountaintop-removal mining, is an efficient way of mining coal, but these mines eventually run out of coal and leave an immense barren landscape. Many areas of the world require the coal mining industry to return the barren land to a more natural condition. Since the ground may have little or no organic soil remaining after strip mining, it is a challenge to manipulate the primary succession of communities to reclaim the stripped land. Preventing soil erosion is critical and can be accomplished by rapidly colonizing the area with plants that will hold the soil and ultimately undergo succession in a way that promotes species richness and improves the functioning of the ecosystem. To promote succession on former mined areas, we need to understand how it works by observing the succession changes in plant communities.
In Spain, researchers examined a chronosequence of 26 strip mines in the northern region of the country that had been abandoned for 1 to 32 years. The mines all had similar soils and climate. When the researchers examined changes in species richness, they found that mines abandoned for 1 year had 8 plant species and that species richness peaked at 28 species after 10 years of being abandoned. This peak was caused by the persistence of some of the pioneer species combined with the colonization by later succession species. After 10 years, species richness exhibited a gradual decline; mines abandoned for 32 years had only 7 to 8 species. As with other communities discussed in this chapter, most of the species in the young abandoned sites were annual winddispersed plants. These species were able to tolerate the harsh conditions of the newly abandoned mine. After 10 years, there was an increase in the number of perennial herbs, and woody plants started to colonize the area. These included nitrogen-fixing plants that contributed large amounts of organic matter and nitrogen to the soil, which facilitated subsequent woody plants, including trees that began to dominate after 20 years. These changes correlated with increased nitrogen in the soil, which suggested that the plants that formed the earliest communities facilitated the colonization and growth of later species. Similar results were found in a 2016 study of succession in coal mines of the Czech Republic. Over several decades, subsoils with little or no organic matter were placed into piles next to the mine to get to the underlying coal. Because the mines were active for decades, these soil piles had experienced succession for 12, 20, 30, or 50
years. Researchers examined how well bushgrass (Calamagrostis epigejos) grew on these soils in the presence and absence of arbuscular mycorrhizal (AM) fungi, which we discussed in Chapter 17. The youngest soils contained low concentrations of phosphorus, and adding AM fungi caused greater plant growth. In contrast, the older soils contained higher concentrations of phosphorus, and adding AM fungi caused no additional growth improvement. Thus, AM fungi play a key role in promoting primary succession on the newly mined soils that lack abundant phosphorus. Understanding how succession proceeds helps scientists develop recommendations for speeding up succession. For example, researchers in Germany examined how succession would be affected under three different manipulations: let the site naturally undergo succession; sow the area with a mixture of herb and grass seeds; or cover the area with newly cut hay, which helps to reduce erosion and provides the seeds of dozens of herbs and grasses contained in the soil. After 4 years, the plots experiencing natural succession had only 35 percent plant cover, while the plots treated with either sown seeds or hay had more than 80 percent plant cover. These substantial differences in plant cover affected soil erosion; plots with sown seeds or green hay had channels that had eroded less than 5 cm deep, while plots undergoing natural succession had channels that had eroded up to 1.5 m deep. The plots experiencing natural succession also had lower species richness during the first year and a lower index of similarity, although the richness of the three plots converged by the end of the 9-year experiment. Collectively, these data confirm that while primary
succession will naturally occur on abandoned strip mines, we can use our knowledge of succession to speed up the process and rapidly move a barren landscape toward a much more natural community. However, as we have learned throughout this chapter, terrestrial communities may require centuries of succession after a disturbance such as strip mining before they can approximate the communities that were originally present.
Alday, J. G., et al. 2011. Functional groups and dispersal strategies as guides for predicting vegetation dynamics on reclaimed mines. Plant Ecology 212: 1759–1775. Baasch, A., et al. 2012. Nine years of vegetation development in a postmining site: Effects of spontaneous and assisted site recovery. Journal of Applied Ecology 49: 251–260. Rydlová, J., et al. 2016. Nutrient limitation drives response of Calamagrostis epigejos to arbuscular mycorrhiza in primary succession. Mycorrhiza 26: 757–767.
Summary of Learning Objectives
19.1 Succession occurs in a community when species replace each other over time. The process of succession can be observed through either direct observations over time or indirect observations that use chronosequences or parts of organisms, such as pollen, that have been naturally preserved over time. Succession occurs on land, where we can distinguish between primary and secondary succession, and in the water. Succession often does not follow a simple linear path of species replacements, but there is a common pattern of rapidly increasing species richness over time that plateaus and can subsequently exhibit a small decline. Key Terms: Succession, Seral stage, Pioneer species, Climax community, Chronosequence, Primary succession, Secondary succession
19.2 Succession can occur through different mechanisms. The mechanisms of succession can be categorized as facilitation, inhibition, and tolerance. More than one mechanism can operate in a community experiencing succession, and the traits of species help determine the mechanisms that develop and where each species occurs along the successional stages. Key Terms: Facilitation, Inhibition, Priority effect, Tolerance
19.3 Succession does not always produce a permanent single climax community. As succession proceeds, the environment continues to change until conditions reach a point of relative stability and the dominant species appear to be persistent. However, the climax community can continue to experience slow changes over time. The climax community can also differ within a region along environmental gradients, such as temperature and moisture. Some climax communities are transient because they experience regular disturbances that reset succession, such as vernal pools that dry each
summer. Extreme conditions, including fires and intense grazing, can also alter climax communities to produce a different composition of dominant organisms. Key Terms: Transient-climax community, Fire-maintained climax community, Grazer-maintained climax community
Critical Thinking Questions
1. In a pond that experiences succession, what relationship would you expect regarding changes in species richness over time? 2. If you were to use a chronosequence to document the pathway of succession in a tropical forest, what are the limitations of this method? 3. How would a trade-off between dispersal ability and competitive ability affect which types of species could colonize small versus large gaps in a community? 4. Why should we not expect a single climax community on recovering coal mines? 5. Compare and contrast the concepts of facilitation, inhibition, and tolerance in the context of ecological succession. 6. If two plant species have similar dispersal and competitive abilities, what factor might help determine which species occupies an early seral stage? 7. Compare and contrast the classic and modern explanations for the succession of ponds and lakes. 8. If two locations in the northeastern United States follow different paths of succession but end up at the same climax community, how will Jaccard’s coefficient of similarity change over time? 9. Why do early- and late-succession species tend to possess different adaptations? 10. Why might the community of insects on a decomposing animal be considered a transient climax?
#### Graphing the Data: Species Richness at Glacier Bay
GRAPHING THE DATA Species Richness at Glacier Bay We typically think of succession as a series of different species dominating a site over time. Ecologist William Reiners and his colleagues visited Glacier Bay sites that had been exposed by the retreating glacier at different times. They quantified the species richness for each of five types of vegetation. Using their data, given in the table, create a stacked bar graph that shows the species richness for each of the five types of vegetation at a given seral stage. Based on this graph, what happens to total species richness of the Glacier Bay sites as succession proceeds?
Data from Reiners, W., et al. 1971. Plant diversity in a chronosequence at Glacier Bay, Alaska. Ecology 52: 55–69. Successional Stage Mosses, Liverworts, and Lichens Low Shrubs and Herbs Tall Shrubs Trees Pioneer Low Shrub Tall Shrub Spruce Forest Hemlock Forest