21 Movement of Elements in Ecosystems
21Movement of Elements in Ecosystems The effects of a dead zone on fish populations. Because algae that bloom will eventually die, the decomposition uses up nearly all the oxygen in the water. These fish died from a dead zone that occurred in Lake Trafford, Florida. Living in a Dead Zone Each summer, as the Mississippi River flows into the Gulf of Mexico, an area develops where animals can’t survive. While fish, crawfish, and crabs remain abundant in other parts of the Gulf, the summer algal bloom in this area makes it uninhabitable. In many cases, the rapidly increasing populations of algae contain green pigments that turn the water green. When the algae contain red pigments, the algal bloom is called a red tide. Algal blooms can have both direct and indirect effects on aquatic organisms. A direct effect occurs when the species of algae or
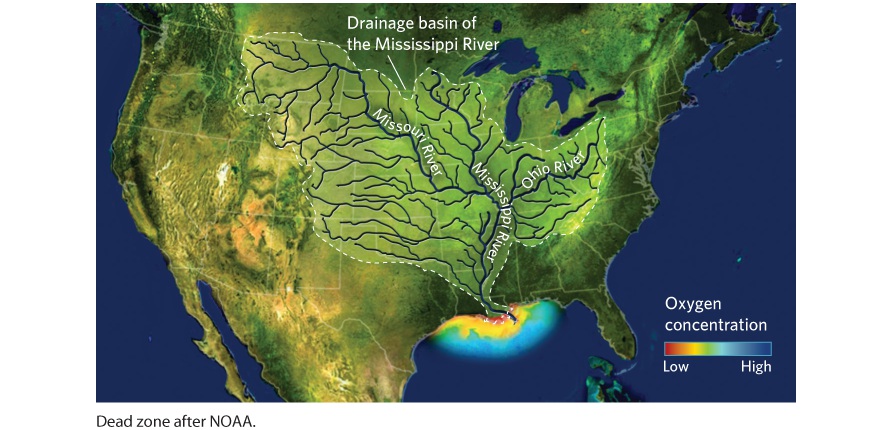
cyanobacteria that bloom produce toxins. At high algal densities, these toxins can accumulate to concentrations that impair the survival, growth, and reproduction of other species living in the area. One prominent example happened in Lake Erie in 2014, when tremendous amounts of algae grew in the bay next to Toledo, Ohio. The water was so green that is resembled pea soup, and the algae were producing such high concentrations of toxins that the city of Toledo had to shut off its water intake valves from the lake for several days to avoid harming its population of nearly 500,000 people. In response, the Ohio National Guard had to truck drinkable water into the city and many businesses and restaurants had to close. An indirect effect of algal blooms occurs after the algae, also known as phytoplankton, bloom and die. While live algae produce oxygen during photosynthesis, bacteria that consume the enormous biomass of dead algae use large amounts of oxygen. This leads to a dramatic reduction in oxygen in the water and causes many of the animals in the water to die from oxygen deprivation. Aquatic ecosystems that experience algal blooms and large animal die-offs are called dead zones. What causes the large algal blooms? Researchers have discovered that many rivers carry large amounts of nutrients, such as nitrogen and phosphorus, that come from fertilizers that run off lawns and agricultural fields when it rains. This water runoff enters streams and rivers that join before emptying into the ocean. Other sources of nutrients include wastewater with a range of components, including detergents that contain phosphorus and sewage, that is released from wastewater treatment systems when they are overwhelmed by large rain events. The nutrients that rivers dump into the ocean allow rapid algal growth, particularly during the warm summer months and this causes algal blooms. Because warm temperatures play a key role, global warming will likely favor an increase in the frequency and intensity of algal blooms today and in the future. “The resulting dead zone in the Gulf of Mexico can
cover more than 22,000 km2 during the summer— an area the size of New Jersey.” The Mississippi River drains 41 percent of the contiguous United States, so it carries nutrients gathered from a very large area. The resulting dead zone in the Gulf of Mexico can cover more than 22,000 km2 during the summer—an area the size of New Jersey. As autumn weather comes, fewer nutrients enter the Mississippi River and the temperatures in the Gulf of Mexico become cooler, conditions that make it more difficult for algal populations to increase. As a result, the dead zone disappears each winter. Human activities cause most dead zones, although some do have natural causes. The abundance of dead zones around the world is growing rapidly. In the 1910s, only four dead zones were known to exist. This number increased to 49 dead zones in the 1960s and 87 in the 1980s. The number of dead zones rose to 305 in 1995 and increased further to 405 by 2008, the most recent date for which there is an estimate. For example, a large dead zone occurs in the Chesapeake Bay on the east coast of North America, where up to 40 percent of the bay can become hypoxic, meaning the water is low in oxygen. Similarly, the bottom of Lake Erie becomes hypoxic each summer. Around the world, dead zones cover a total area of more than 205,000 km2. A dead zone at the mouth of the Mississippi River. The Mississippi River drains 41 percent
of the contiguous United States and carries nutrients from fertilizers leached from people’s yards and agricultural fields and from wastewater from communities located throughout the region. These nutrients facilitate the rapid growth of algae that ultimately cause a dead zone, which you can see at the mouth of the river. The existence of dead zones illustrates why we need to understand how nutrients—including water, nitrogen, and phosphorus—move within and between ecosystems and the important role that decomposition plays in recycling these nutrients. In this chapter, we will examine the movement of nutrients and how nutrients are regenerated in terrestrial and aquatic ecosystems. SOURCES: Diaz, R. J., and R. Rosenberg. 2008. Spreading dead zones and consequences for marine ecosystems. Science 321: 926–929. Neuhaus, L. 2016. A menace afloat. New York Times, July 19. Rabalais, N. N., et al. 2002. Gulf of Mexico hypoxia, a.k.a. The dead zone. Annual Review of Ecology and Systematics 33: 235–263.
Learning Objectives
After reading this chapter, you should be able to:
21.1 Describe how the hydrologic cycle moves many elements through ecosystems.
21.2 Explain why the carbon cycle is closely tied to the movement of energy.
21.3 Illustrate the ways in which nitrogen cycles through ecosystems in many different forms.
21.4 Describe how the phosphorus cycle moves between land and water.

21.5 Explain why most nutrients regenerate in the soil in terrestrial ecosystems.
21.6 Illustrate why most nutrients regenerate in the sediments in
aquatic ecosystems. Unlike energy—which moves through ecosystems—elements such as hydrogen, oxygen, carbon, nitrogen, and phosphorus cycle among the biotic and abiotic components of ecosystems. The movement of these elements through ecosystems is affected by chemical, physical, and biological processes. To understand the movement of these elements, which exist in a variety of chemical forms, it is helpful to think of different pools in which a given element resides, as well as the different processes that are responsible for moving an element from one pool to another. For example, two important pools for carbon are the CO2 that exists in the atmosphere and the biomass of producers that use carbon to build their tissues. In this example, the process that causes carbon to move from the atmosphere to producers is photosynthesis. Organisms contain large amounts of hydrogen, oxygen, and carbon. However, as we noted in Chapter 2, organisms also need seven major nutrients: nitrogen, phosphorus, sulfur, potassium, calcium, magnesium, and iron. Some elements are required in much smaller amounts; these include silicon, manganese, and zinc. In this chapter, we will examine the biogeochemical cycles of some of the major elements on Earth by looking at interactions among biological, geological, and chemical processes. We will also explore how human activities are currently altering these cycles in ways that have far-reaching effects on ecosystems.
#### 21.1 the Hydrologic Cycle Moves Many Elements Through Ecosystems
21.1 The hydrologic cycle moves many elements through ecosystems Throughout the previous chapter, we saw that water plays a critical role at every level of ecological study. In Chapter 2, we discussed how water is a key compound involved in the many chemical transformations that happen in the living and nonliving components of ecosystems. Chapter 5 examined the processes that determine global climates, including patterns of precipitation, and in Chapter 6, we discussed the various types of aquatic biomes. We begin this chapter by tracking the movement of water through ecosystems. Once we understand how water moves through ecosystems, we will look at the role water plays as it moves elements through ecosystems.
The Hydrologic Cycle
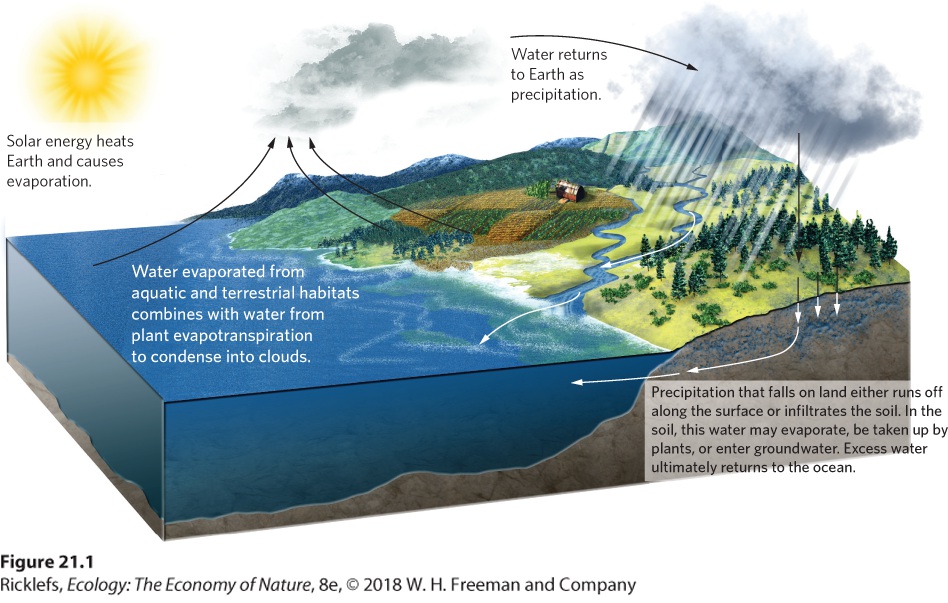
The movement of water through ecosystems and atmosphere, known as the hydrologic cycle, is driven largely by evaporation, transpiration, and precipitation. The largest pool of water, approximately 97 percent of all water on Earth, is found in the oceans. The remaining water exists in lakes, streams, rivers, wetlands, underground aquifers, and soil. Hydrologic cycle The movement of water through ecosystems and atmosphere. The hydrologic cycle is illustrated in Figure 21.1. Evaporation of water occurs from bodies of water, soil, and plants that experience evapotranspiration, which we discussed in Chapter 5. Solar energy provides the energy for the process of evaporation and evapotranspiration, which changes water from a liquid to a gas in the form of water vapor. There is a limit to the amount of water vapor that the atmosphere can contain. As additional water continues to evaporate, the water vapor in the atmosphere condenses into clouds that ultimately create precipitation in the form of rain, hail, sleet, or snow.

Figure 21.1 The hydrologic cycle. The movement of water is driven by the energy of the Sun, which causes evaporation from soil and water bodies, and evapotranspiration from plants. Evaporated water condenses into clouds that eventually return the water to Earth as precipitation. Water from precipitation either runs off on the surface or infiltrates the soil. Runoff flows along the surface of the ground until it enters streams and rivers. Water in the soil is taken up by plants or enters the groundwater. Ultimately, excess water returns to the ocean. When precipitation drops from the atmosphere, it can take several paths. Some precipitation falls directly onto the surface of aquatic ecosystems and the rest falls onto terrestrial ecosystems. Water that falls onto terrestrial ecosystems can travel along the surface of the ground or it can infiltrate the ground, where it is either absorbed by plants or moves deeper into the ground and becomes part of the underlying groundwater. The surface runoff and some of the groundwater will eventually find their way back into water bodies, thereby completing the cycle. Precipitation that falls on land either runs off along the surface or infiltrates the soil. In the soil, this water may evaporate, be taken up by plants, or enter groundwater. Excess water ultimately returns to the ocean. The rate of evaporation must balance the rate of precipitation or water would continually accumulate in one part of the cycle. When we consider the hydrologic cycle on a global scale, we find that precipitation exceeds evaporation in terrestrial ecosystems, whereas evaporation exceeds precipitation in aquatic ecosystems. To help maintain an overall balance, the excess water that is evaporated from aquatic ecosystems is transported by the
atmosphere and falls onto terrestrial ecosystems. At the same time, the excess water that falls on terrestrial ecosystems is transported in the form of runoff and groundwater into aquatic ecosystems.
Human Impacts on the Hydrologic Cycle
Water cycles through Earth’s biosphere with no net loss or gain over the long term. Therefore, any change in one part of the water cycle influences the other parts. For example, in large developed areas, construction materials such as roofing and paved parking lots are impervious to water infiltration. The amount of water that can percolate into the soil is significantly reduced, and we see an increase in surface runoff. Less water is able to infiltrate the soil for plants to use or to replenish the groundwater that many people rely on for drinking water. An increase in surface runoff also increases soil erosion. A similar effect occurs when we reduce the amount of plant biomass in a terrestrial ecosystem, as occurs during logging. Where there are fewer trees and other plants, much less precipitation is taken up by plant roots and subsequently released to the atmosphere through evapotranspiration. Consequently, the amount of surface runoff increases, which often causes severe soil erosion and flooding (Figure 21.2). Finally, when we pump out groundwater for irrigation or household use, we sometimes reduce the amount of groundwater at a rate that exceeds its replenishment. For example, in the Great Plains of the United States, a large supply of groundwater, known as the Ogallala aquifer, extends from South Dakota to Texas (Figure 21.3). This groundwater supplies about 30 percent of all water used for irrigation in the United States and provides drinking water for 82 percent of the people who live in the region. However, the extraction of this water has exceeded its rate of replenishment, and scientists are concerned that this critical supply of water for industry, households, and irrigation could run out sometime during this century.
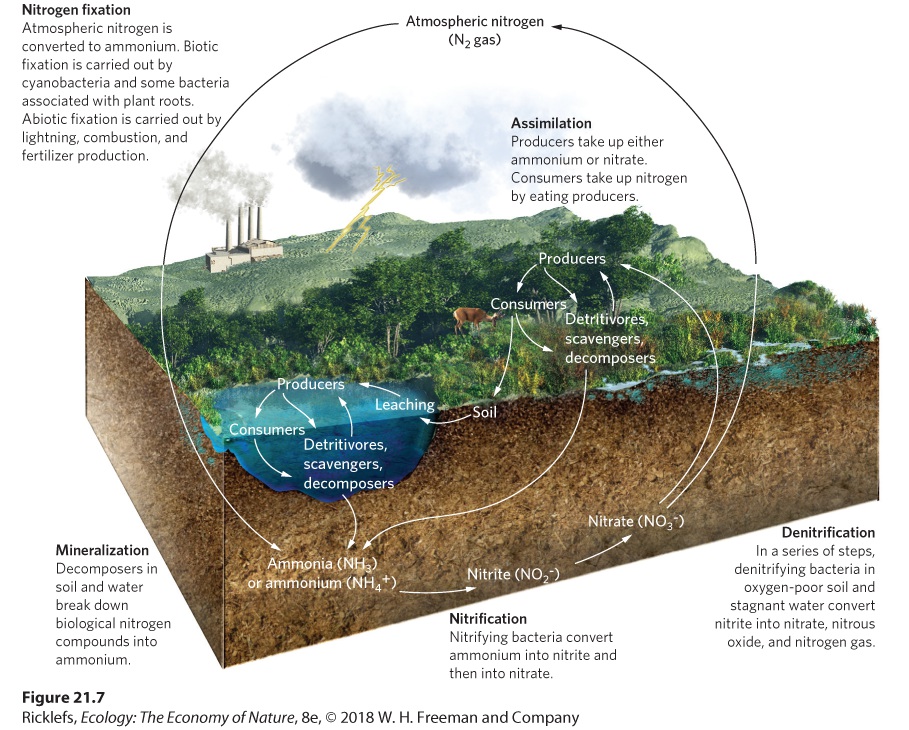
Figure 21.2 Altering the hydrologic cycle. When forests are logged, as in this site in Haiti, fewer plant roots are available to hold soil and the soil absorbs less rainwater. These changes cause increased surface runoff, more severe floods, and large amounts of soil erosion.
Figure 21.3 The Ogallala aquifer. The Ogallala aquifer is the largest source of ground water in the United States. Humans also alter the hydrologic cycle through activities that contribute to global warming. Scientists expect that as air and water temperatures rise, there will be an increase in the rate of water evaporation. An increase in the evaporation rate will cause water to move through the hydrologic cycle more quickly, potentially leading to an increased intensity of rain and snowstorms in various parts of the world.
Concept Check
1. What are the pathways that water can take when precipitation falls on terrestrial ecosystems? 2. How does an increase in the construction of buildings and water-impervious parking lots affect the hydrological cycle? 3. How does tree removal affect the hydrological cycle?
#### 21.2 the Carbon Cycle Is Closely Tied to the Movement of Energy
21.2 The carbon cycle is closely tied to the movement of energy Because all organisms are composed of carbon, the movement of carbon in ecosystems largely follows the same paths as the movement of energy. In this section, we consider the many pools and processes that are involved in the carbon cycle. We will then examine how human activities have altered the carbon cycle.
The Carbon Cycle
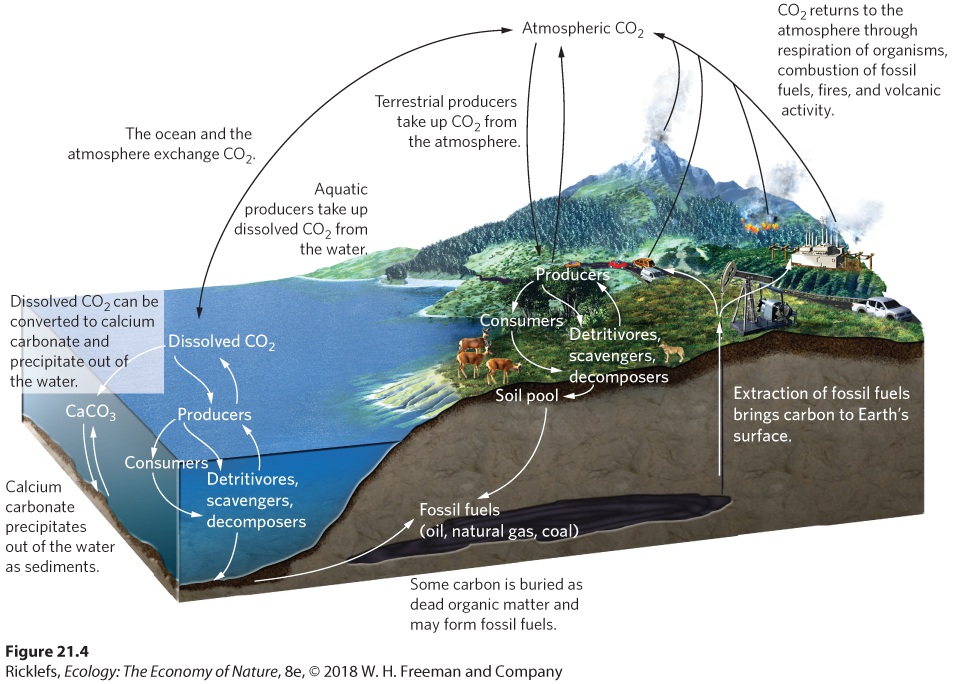
To understand how the carbon cycle works, we need to consider six types of transformations: photosynthesis, respiration, sedimentation and burial, exchange, extraction, and combustion. The carbon cycle is illustrated in
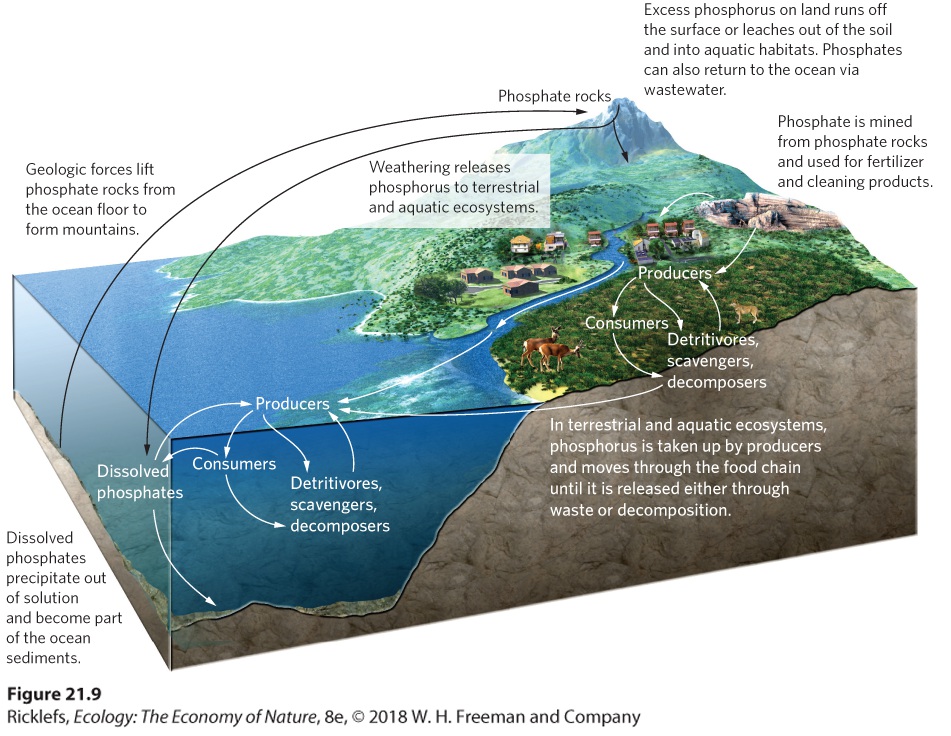
Figure 21.4 The carbon cycle. In the carbon cycle, producers take up CO2 from the atmosphere and the water. They transfer assimilated carbon to consumers, detritivores, scavengers, and decomposers. These organisms return CO2 to the atmosphere and oceans through respiration. Throughout an ecosystem, CO2 is exchanged between the atmosphere and the ocean and between the ocean and sediments. Carbon that has been stored underground for long periods turns into fossil fuels, which can be extracted. CO2 is returned to the atmosphere through the combustion of fossil fuels, burning in terrestrial ecosystems,
and volcanic activity. We begin our examination with the processes of photosynthesis and respiration. As we have discussed in previous chapters, producers use photosynthesis in terrestrial and aquatic ecosystems to take CO2 from the air and water and to convert it to carbohydrates. These carbohydrates are used to make other compounds, including proteins and fats. The carbon that is locked up in producers can then be transferred to consumers, scavengers, detritivores, and decomposers. All these trophic groups experience respiration, which releases CO2 back into the air or water. In some habitats, such as the waterlogged sediments of swamps or marshes, oxygen is not available for respiration. Under such anaerobic conditions, some species of archaea use carbon compounds for respiration. For example, some archaea use methanol (CH3OH) during respiration to produce CO2 in the following reaction: 4 CH3OH→CO2+2 H2O+3 CH4 As you can see from this reaction, the products are carbon dioxide, water, and methane. The methane that is released from swamps during anaerobic respiration is known as swamp gas. The production of methane through the process of anaerobic respiration is a growing concern because methane is a greenhouse gas and, on a per-molecule basis, it is 72 times more effective at absorbing and radiating infrared radiation back to Earth than CO2. Carbon dioxide is also exchanged between aquatic ecosystems and the atmosphere, as shown in Figure 21.4. The exchange occurs in both directions at a similar magnitude, which means there is little net transfer over time. As we discussed in Chapter 2, when CO2 diffuses from the atmosphere into the ocean, some of it is used by plants and algae for photosynthesis and some is converted to carbonate (CO3 2–) and bicarbonate ions (HCO3 –). The carbonate ions can then combine with calcium in the water to form calcium carbonate (CaCO3). Calcium carbonate has a low solubility in water, so it precipitates out of the water and becomes part of the sediments at the bottom of the ocean. Over millions of years, the calcium carbonate sediments that accumulate in the ocean bottoms, combined with the calcium carbonate
skeletons from tiny marine organisms, can develop into massive sources of carbon in the form of rocks known as dolomite and limestone. Humans mine dolomite and limestone for use in making concrete and fertilizer as well as for numerous industrial processes. Carbon can also be buried as organic matter before it fully decomposes. Over millions of years, some of this organic matter is converted to fossil fuels such as oil, gas, and coal. The rate of carbon burial is slow, and it is offset by the rate of carbon released into the atmosphere by the weathering of limestone rock and during volcanic eruptions. Because the process of sedimentation and burial can lock up carbon for millions of years, carbon moves through these pools very slowly.
Human Impacts on the Carbon Cycle
Now that we understand the pools and processes in the carbon cycle, we can examine how human activities have altered it. One major way in which humans have altered the carbon cycle is through the extraction and combustion of fossil fuels. During the past 2 centuries, extraction and combustion have been happening at an increasing rate to meet growing energy demands. Other forms of combustion by humans include burning land to prepare it for agriculture. In Chapter 4, we discussed the recent rise in atmospheric CO2 seen in measurements made atop Mauna Loa on the island of Hawaii. These measurements document an increase in CO2 from 316 ppm in 1958 to more than 405 ppm in 2017—a 28 percent increase in just 59 years. Although these measurements at Mauna Loa began only in 1958, human activities have affected CO2 concentrations in the atmosphere for much longer. To measure the concentrations of CO2 that were in the atmosphere hundreds of thousands of years ago, researchers have traveled to some of the coldest places on Earth. In locations such as Greenland and Antarctica, snowfall slowly compresses into ice that has tiny bubbles of air trapped inside. Because these bubbles contain tiny samples of the air from thousands of years ago, they can tell us about the climate conditions in the distant past. Each year, ice is formed by adding a new layer; the surface layers contain the youngest ice and the deepest layers contain the oldest ice. To sample the air that is trapped in the ice, researchers drill far down into the ice and extract long cylinders of ice
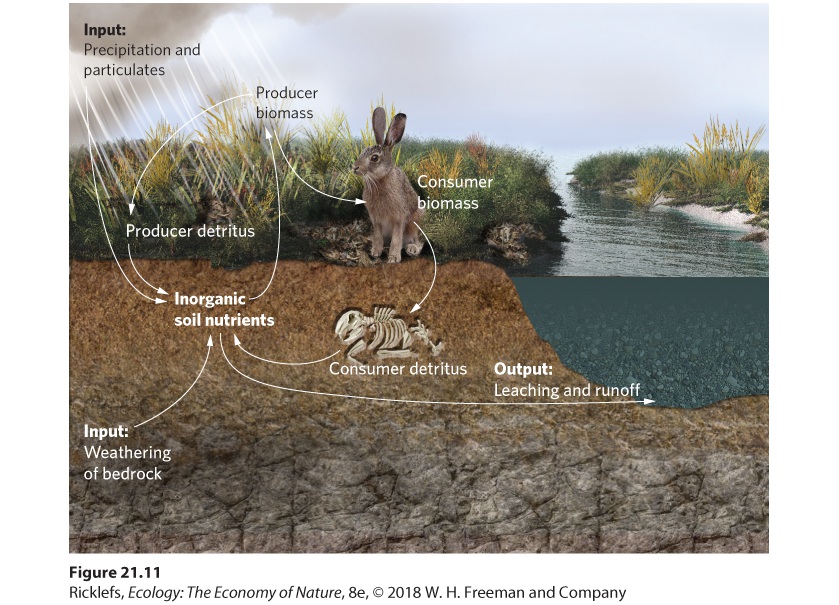
Figure 21.5 Ice cores. Researchers from Britain drill far down into ancient ice to collect ice cores that have been created in layers over the past 500,000 years. Ice cores contain ice that has been formed as far back as 500,000 years ago. After determining the age of different ice core layers, each layer is melted, which allows the release of trapped air bubbles of which the concentration of CO2 can be measured. Figure 21.6 shows some of the data from these ice cores. As you can see, CO2 concentrations in the atmosphere during the past 400,000 years have varied a great deal, from about 180 to 300 ppm. However, since 1800, as humans increasingly burned fossil fuels, CO2 concentrations have increased exponentially to the current value of 405 ppm. This means that the current concentration of CO2 in our atmosphere is 35 percent higher than the highest concentrations that existed during the past 400,000 years.
Figure 21.6 Atmospheric concentrations of CO2 over time. Using measurements of air bubbles from ice cores and, in modern times, direct measurements, researchers have documented that for more than 400,000 years CO2 concentrations never exceeded 300 ppm. During the past 200 years, the concentration of CO2 in the atmosphere has increased rapidly and is now more than 400 ppm. The rise in atmospheric CO2 is of great importance to humans because CO2 is a greenhouse gas that absorbs infrared radiation and radiates some of it back to Earth. Having CO2 in the atmosphere helps keep our planet warm, but an excessive amount of CO2 and other greenhouse gases in the atmosphere will cause our planet to become much warmer than it has been in a very long time. We know that the mean temperature on Earth is now 0.8 °C warmer than it was when the first temperature measurements were taken in the 1880s. While a mean increase of 0.8 °C may not seem like much, we can find some dramatic changes in specific locations. Some regions, such as parts of Antarctica, have experienced cooler temperatures, while other regions, such as the high latitudes of Alaska, Canada, and Russia, are now 4 °C warmer than they were a century ago. These high-latitude regions contain large deposits of frozen peat, which is a mixture of dead sphagnum moss and other plants. Peat thaws and decomposes more easily with warmer temperatures. Because peat decomposes under anaerobic conditions, the decomposition produces methane, which is a greenhouse gas. This means that the rise in temperatures due to increased atmospheric CO2 causes the release of additional greenhouse gases from the decomposing peat that exacerbate the problem. Temperature increases can have numerous effects around the world, such as reducing the size of the polar ice sheets (see Chapter 5), altering the length of plant
growing seasons, and changing the timing of plant and animal life histories. We will have much more to say about global warming in our discussion of global biodiversity conservation in Chapter 23.
1. What form of carbon is produced during anaerobic respiration? 2. What form of carbon precipitates out of water? 3. How has increased combustion of carbon sources over the past two centuries contribute to global warming?
#### 21.3 Nitrogen Cycles Through Ecosystems in Many Different Forms
21.3 Nitrogen cycles through ecosystems in many different forms Nitrogen is an important component of amino acids, the building blocks of proteins, and nucleic acids, which are the building blocks of DNA. Nitrogen exists in many different forms and is part of a complex set of pathways. In this section, we will examine the cycling of nitrogen and explore the ways that human activities have altered the nitrogen cycle.
The Nitrogen Cycle
A large pool of nitrogen gas (N2) exists in the atmosphere, where it comprises 78 percent of all atmospheric gases. Nitrogen moves through five major transformations, shown in Figure 21.7: nitrogen fixation, nitrification, assimilation, mineralization, and denitrification.
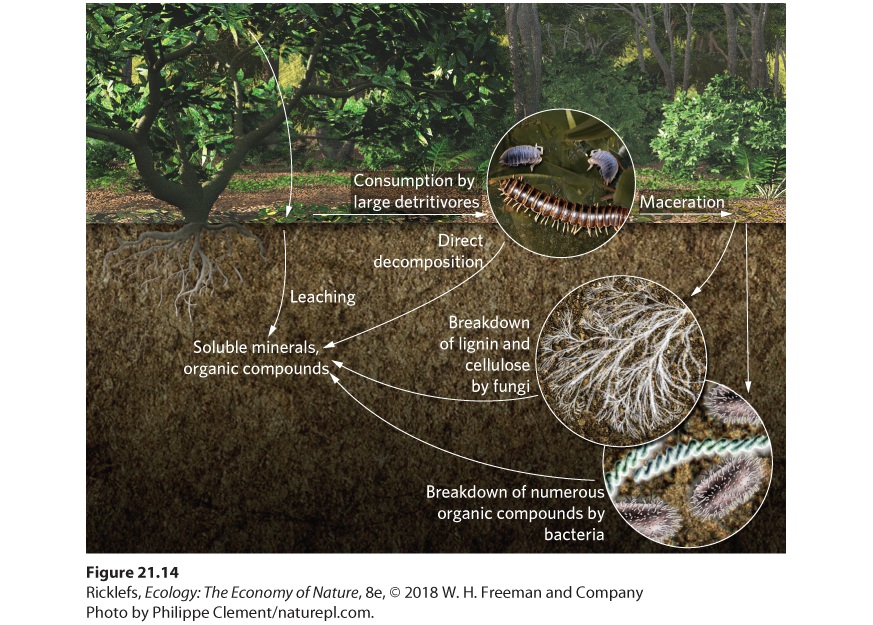
Figure 21.7 The nitrogen cycle. The nitrogen cycle begins with nitrogen gas in the atmosphere. The process of nitrogen fixation converts it into a form that producers can use. The fixed nitrogen can then be assimilated into producers and consumers; it ultimately decomposes into ammonium through the process of mineralization. The ammonium can be converted into nitrite and then nitrate through the process of nitrification. Under anaerobic conditions, the nitrate can be converted into nitrogen gas through the process of
denitrification. Nitrogen Fixation The process of converting atmospheric nitrogen into forms producers can use is known as nitrogen fixation. Nitrogen fixation converts nitrogen gas into either ammonia (NH3), which is rapidly converted to ammonium (NH4 +), or into nitrate (NO3 −). The compound that is formed depends on whether nitrogen fixation occurs by organisms, lightning, or the industrial production of fertilizers. Nitrogen fixation The process of converting atmospheric nitrogen into forms producers can use. As we discussed in previous chapters, some organisms are able to convert nitrogen gas into ammonia. Nitrogen fixation occurs in some species of cyanobacteria, in some free-living species of bacteria such as Azotobacter, and in mutualistic bacteria such as Rhizobium that live in the root nodules of some legumes and other plants (see Figure 17.4). Nitrogen fixation is an important source of required nitrogen, especially for early-succession plants colonizing habitats that have little available nitrogen. The process of nitrogen fixation requires a relatively high amount of energy, which nitrogen-fixing organisms obtain either by metabolizing organic matter from the environment or by acquiring carbohydrates from a mutualistic partner. As you can see in Figure 21.7, nitrogen fixation can also occur through abiotic processes. For example, lightning provides a high amount of energy that can convert nitrogen gas into nitrate in the atmosphere. Similarly, combustion that occurs during wildfires or when fossil fuels are burned also produces nitrates. In both cases, nitrates, which are suspended in the air after combustion, fall to the ground with precipitation. The industrial production of fertilizers that improve crop productivity converts nitrogen gas to either ammonia or nitrates. Like all nitrogen fixation, this process requires a great deal of energy and is powered mostly by the combustion of fossil fuels. The manufacture of nitrogen fertilizer has developed into such a large commercial endeavor that fixation conducted by humans now exceeds the nitrogen fixation that occurs through all natural processes.
Nitrification Another process in the nitrogen cycle is nitrification, which converts ammonium to nitrite (NO2 –) and then converts nitrite to nitrate (NO3 –): NH4+→NO2−→NO3− Nitrification The final process in the nitrogen cycle, which converts ammonium (NH4+) or ammonia (NH3) to nitrite (NO2−) and then to nitrate (NO3−). These conversions release much of the potential energy that is contained in ammonium. Each step is carried out by specialized bacteria and archaea in the presence of oxygen. In terrestrial and aquatic ecosystems, the conversion of ammonium to nitrites is carried out by Nitrosomonas and Nitrosococcus bacteria, whereas the conversion of nitrites to nitrates is carried out by Nitrobacter and Nitrococcus bacteria. Although nitrites are not an important nutrient for producers, plants can take them up and use them. Assimilation and Mineralization Producers can take up nitrogen from the soil or water as either ammonium or nitrates. Once producers take up nitrogen, they incorporate it into their tissues, a process known as assimilation, which was described in Chapter 20. When primary consumers ingest producers, they can either assimilate nitrogen from the producers or excrete it as waste. The same process occurs again with secondary consumers. Animal waste, as well as the biomass of dead producers and consumers, is broken down by scavengers, detritivores, and decomposers. The fungal and bacterial decomposers break down biological nitrogen compounds into ammonia. The process of breaking down organic compounds into inorganic compounds is known as mineralization. Mineralization The process of breaking down organic compounds into inorganic compounds. Denitrification Because nitrates produced by nitrification are quite soluble in water, they readily leach out of soils and into waterways, where they settle in the
sediments of wetlands, rivers, lakes, and oceans. These sediments are typically anaerobic. Under anaerobic conditions, nitrates can be transformed back into nitrites, which are then transformed into nitric oxide (NO): NO3−→NO2−→NO This reaction is accomplished by bacteria such as Pseudomonas denitrificans. Additional chemical reactions under anaerobic conditions in soils and water subsequently convert nitric oxide to nitrous oxide (N2O) and then to nitrogen gas, thereby completing the nitrogen cycle: NO→N2O→N2 This process of converting nitrates into nitrogen gas is known as denitrification. Denitrification The process of converting nitrates into nitrogen gas. Denitrification is necessary for breaking down organic matter in oxygendepleted soils and sediments. However, as you can see from the above reaction, it produces nitrogen gas (N2). Since N2 cannot be taken up by producers, the denitrification process causes nitrogen to leave the waterlogged soils and aquatic ecosystems in the form of a gas.
Human Impacts on the Nitrogen Cycle
Before human activities began dramatically altering the environment, the production of usable forms of nitrogen through the process of fixation was approximately offset, on a global scale, by the loss of usable nitrogen through denitrification. However, during the last 3 centuries, and especially during the past 50 years, human activities have nearly doubled the amount of nitrogen put into terrestrial ecosystems. These activities include combustion of fossil fuels that add nitric oxide to the air, production of nitrogen fertilizers, and planting nitrogenfixing crops. As nitric oxide enters the atmosphere from combustion, it reacts with water in the air to form nitrates, which then fall to the ground during precipitation events. Because nitrogen is often a limiting nutrient, we would expect the

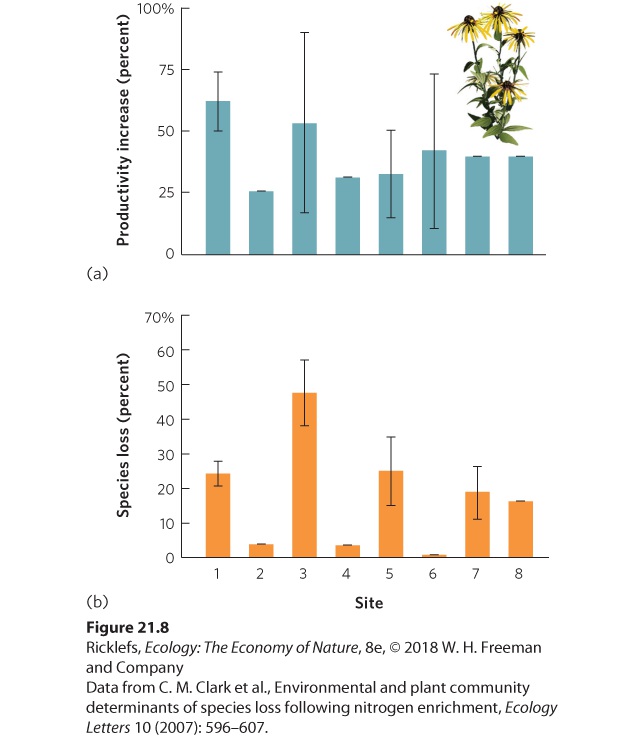
addition of nitrates to affect a variety of ecosystems. Over the years, there have been numerous investigations into whether adding nitrogen to terrestrial ecosystems in North America affects productivity and species richness. These study sites, which range north to south from Alaska to Arizona and west to east from California to Michigan, were recently compiled in an effort to determine whether their results showed a general pattern. When nitrogen was added in the form of nitrates and ammonium, all sites experienced an increase in primary productivity, shown in Figure 21.8a. This confirmed that nitrogen was a limiting resource at all sites. However, the sites differed in the proportion of species that were eliminated over time, as you can see in Figure 21.8b. The researchers explored a wide range of potential causes for this variation in species loss among sites. In addition to differences in temperature and soil among the sites, they found that the sites with the largest increases in productivity experienced the largest reduction in species richness. Adding nitrogen to these communities commonly caused a few plant species to grow very large and to dominate the community. These large plants shaded the lesscompetitive smaller plants, which caused the smaller species to decline. These results demonstrate that the increases in nitrogen in the environment due to human activities can reduce the species diversity of ecosystems.
Figure 21.8 Effects of nitrogen addition on plant productivity and species richness. Twenty-three experiments at eight locations around the United States added nitrogen to plant communities. (a) All sites experienced increases in primary productivity. (b) All sites experienced a loss in species richness, but the magnitude of the loss varied greatly among sites due to differences in existing nitrogen concentrations and differences in the cation exchange capacity of the soil. Error bars are standard errors.
Concept Check
1. What are three processes that cause nitrogen fixation? 2. Under what abiotic conditions does the process of denitrification occur? 3. Why does human production of nitrogen fertilizers alter plant species richness?

#### 21.4 the Phosphorus Cycle Moves between Land and Water
21.4 The phosphorus cycle moves between land and water Phosphorus is a critical element for organisms because it is used in bones and scales, teeth, DNA, RNA, and ATP, a molecule involved in metabolism. As we discussed in Chapter 20, phosphorus is also a common limiting nutrient in aquatic and terrestrial ecosystems. For this reason, phosphorus is a component of most fertilizers manufactured to boost the growth of crops. In this section, we will examine the phosphorus cycle and explore how human activities have altered it in ways that affect ecosystems.
The Phosphorus Cycle
The phosphorus cycle is considerably less complicated than the nitrogen cycle. As you can see in Figure 21.9, the atmosphere is not an important component of this cycle because phosphorus does not have a gas phase; phosphorus can only enter the atmosphere in the form of dust. Unlike nitrogen, phosphorus rarely changes its chemical form and typically moves as a phosphate ion (PO4 3–). Plants take up phosphate ions from soil or water and incorporate them directly into various organic compounds. Animals eliminate excess phosphorus in their diets by excreting urine containing either phosphate ions or phosphorus compounds that are converted into phosphate ions by phosphatizing bacteria.
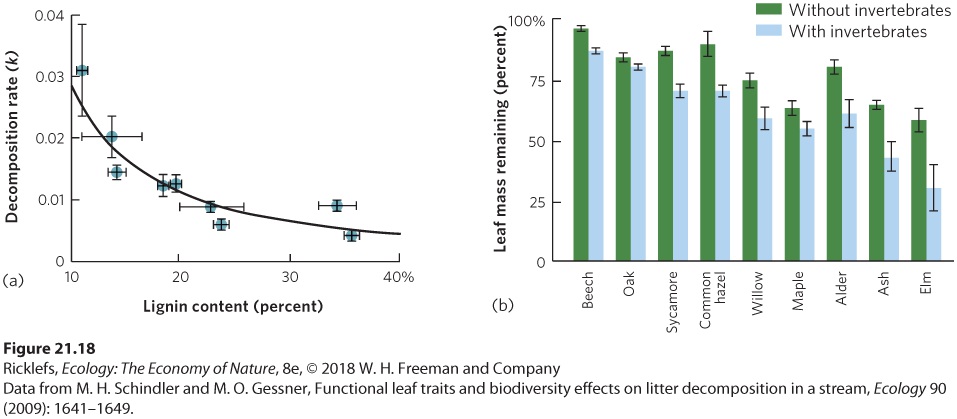
Figure 21.9 The phosphorus cycle. Phosphate rocks that are uplifted by geologic forces naturally weather over time to release phosphorus. These rocks are the source of phosphorus used in fertilizer and detergents. Phosphorus is taken up by producers and moves through the food chain until it is released either through waste or decomposition. Excess phosphorus on land runs off the surface or leaches out of the soil and into aquatic habitats. In the ocean, phosphorus combines with calcium or iron and precipitates out of the water, ultimately to form phosphate rocks again. We begin our exploration of the phosphorus cycle by examining phosphate rocks, which are the major source of phosphate. As you can see in Figure 21.9, over time calcium phosphate (Ca(H2PO4)2) precipitates out of ocean water and slowly forms sedimentary rock. Later, some of this rock is uplifted by geologic forces. Exposed rocks experience weathering, which causes them to slowly release phosphate ions. Phosphate rocks are also mined for phosphate that is used in fertilizers and in a variety of detergents. When phosphate ions enter terrestrial ecosystems, they can be either bound strongly to the soil or taken up by plants and passed through the terrestrial food web. Animal excretions and the decomposition of all terrestrial organisms release phosphorus back to the soil. Excess phosphorus that is not bound to the soil or taken up by plants either moves across the surface of the land during a rainstorm as runoff or leaches from the soil. When soil erosion occurs, the phosphorus that is bound to the soil is carried away with the
eroding soil particles. In either case, the phosphorus can be carried to a variety of aquatic ecosystems. When phosphate ions enter aquatic ecosystems, they are taken up by producers and enter the food web in a manner that is similar to the terrestrial food web. In well-oxygenated waters, phosphorus binds readily with calcium and iron ions and precipitates out of water to become part of the sediments. Thus, marine and freshwater sediments act as phosphorus sinks by continually removing phosphorus from the water. Under low-oxygen conditions, iron tends to combine with sulfur rather than phosphorus, so phosphorus remains more available in the water. Over time, the phosphate that precipitates down to ocean sediments is converted into calcium phosphate rocks, and the phosphorus cycle begins again.
Human Impacts on the Phosphorus Cycle
In Chapter 20, we discussed how phosphorus is commonly a limiting nutrient in terrestrial and aquatic ecosystems. Yet, adding phosphorus to these ecosystems can have harmful effects. As we saw at the beginning of this chapter, phosphorus, sometimes in combination with excess nitrates, contributes to algal blooms that cause dead zones where rivers empty into oceans. This phenomenon happens in locations around the world, as shown in Figure 21.10. An increase in the productivity of aquatic ecosystems is called eutrophication. An increase in the productivity of aquatic ecosystems caused by human activities is called cultural eutrophication.

Figure 21.10 Dead zones. In 2008, more than 400 dead zones were identified around the
world. This represents an increase of 33 percent from 1995. Eutrophication An increase in the productivity of aquatic ecosystems. Cultural eutrophication An increase in the productivity of aquatic ecosystems caused by human activities. From the 1940s to the 1990s, household detergents contained phosphates to improve their cleaning effectiveness. These detergents became part of the wastewater that traveled through public sewage systems, ultimately emptying into rivers, lakes, and oceans. People began to realize that these detergents significantly increased phosphorus in waterways, which contributed to eutrophication and dead zones. In 1994, the United States banned phosphates in laundry detergents after several states had already done so. In 2010, 16 states banned phosphates in dishwashing detergents. In 2011, the European Union agreed to similar restrictions on phosphates in laundry and dishwasher detergents to help reduce the problems of cultural eutrophication and dead zones.
1. What are the two paths that dissolved phosphates can take in aquatic ecosystems? 2. In what ways is mined phosphate used by people? 3. How has laundry detergent led to cultural eutrophication?
#### 21.5 in Terrestrial Ecosystems, Most Nutrients Regenerate in the Soil
21.5 In terrestrial ecosystems, most nutrients regenerate in the soil We have seen that elements cycle through the ecosystem and how they are both used and regenerated. In this section, we will examine how nutrients are regenerated in terrestrial ecosystems by the weathering of bedrock and by the breakdown of organic matter.
The Importance of Weathering
Terrestrial ecosystems constantly lose nutrients because many are leached out of the soil and transported away in streams and rivers. To maintain a stable level of productivity, the loss of nutrients from an ecosystem must be balanced by an input of nutrients. For some nutrients, such as nitrogen, inputs come from the atmosphere. For most other nutrients, such as phosphorus, the inputs come from the weathering of bedrock beneath the soil. As we described in Chapter 5, weathering is the physical and chemical alteration of rock material near Earth’s surface. Substances such as carbonic acid in rainwater and organic acids produced by the decomposition of plant litter react with minerals in the bedrock and release various elements that are essential to plant growth. Determining the rate of weathering can be difficult because bedrock often exists far below the surface of the soil. One solution has been to measure the nutrients that enter a terrestrial ecosystem from precipitation and the nutrients that leave an ecosystem by leaching out of the soil and into a stream. You can see a diagram of nutrient inputs and outputs in Figure 21.11. If the system is in equilibrium—if nutrient inputs equal nutrient outputs—the difference between the nutrients entering the system by precipitation and particulate matter and the amount of nutrients leaving the system by leaching and runoff should equal the amount of nutrients made available by weathering.
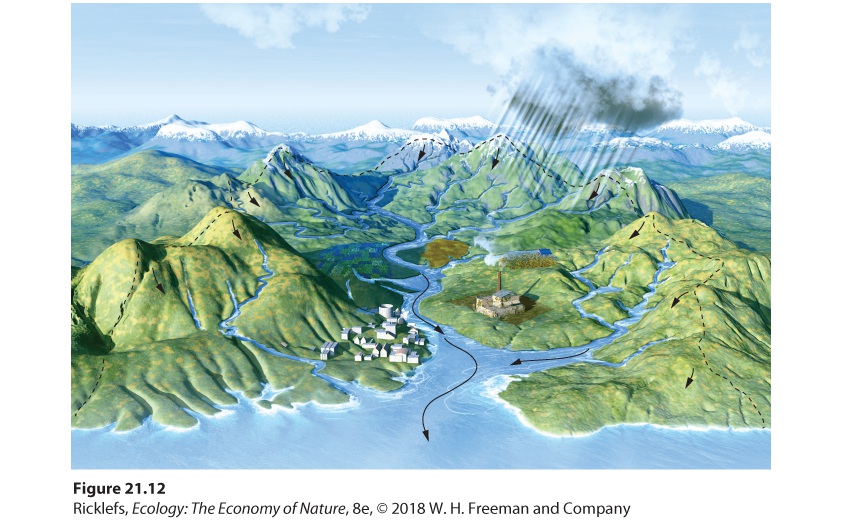
Figure 21.11 Quantifying nutrient weathering. Soil nutrients cycle through producers, consumers, and detritus. This cycle also has inputs from precipitation and weathering, and outputs of groundwater and surface water runoff. Ecologists commonly determine the rate of nutrient regeneration through weathering by quantifying nutrient inputs and outputs from a watershed. A watershed is an area of land that drains into a single stream or river, as illustrated in Figure 21.12. In a watershed, the rate of weathering is estimated by measuring the net movement of several highly soluble nutrients, such as calcium (Ca2+), potassium (K+), sodium (Na+), and magnesium (Mg2+). These nutrients easily leach out of the soil and move into streams, where their concentrations can be measured as the stream leaves the watershed.
Figure 21.12 A watershed. A watershed is an area of land that drains down into a single stream or river. The dashed black line indicates the boundaries of the watershed and the black arrows indicate the directions of water movement down the mountain. Watershed An area of land that drains into a single stream or river. An example of this approach was recently reported for 21 small watersheds in the Canadian province of Quebec. Each watershed contained a small lake with a stream flowing out of the lake that drained the watershed. The watersheds were all forested and had little human activity, so the researchers assumed that nutrient movement was at equilibrium. Measurements were made of the amount of calcium, potassium, sodium, and magnesium entering each watershed through precipitation, the amount of each element present in the soil and bedrock, and the amount of each element coming out of the watershed in the stream that drained it. By knowing the inputs and outputs of a watershed, scientists were able to determine rates of bedrock weathering. When the data were placed on a map of the study area, as shown in Figure 21.13, they found that weathering rates varied geographically. The rate of weathering was highest in the southwest region of the province, probably because of regional differences in temperatures, precipitation, and soil conditions.
Figure 21.13 Weathering rates in 21 Canadian watersheds in Quebec. When researchers quantified the combined inputs and outputs of calcium, magnesium, potassium, and sodium, they found that the rates of weathering differed a great deal across the landscape.
The Breakdown of Organic Matter
Although weathering of bedrock provides nutrients to terrestrial ecosystems, it is a very slow process. Therefore, primary production largely depends on rapid regeneration of nutrients from decomposition, a process that breaks down organic matter into smaller and simpler chemical compounds and is conducted primarily by bacteria and fungi. As we discussed in Chapter 1, scavengers consume dead animals and detritivores break up organic matter into smaller particles. As illustrated in Figure 21.14, the breakdown of plant matter in a forest occurs in four ways: soluble minerals and small organic compounds leach out of organic matter, large detritivores consume organic matter, fungi break down the woody components and other carbohydrates in leaves, and bacteria decompose almost everything. Leaching removes 10 to 30 percent of soluble substances from organic matter, which includes most salts, sugars, and amino acids. What remains behind are complex carbohydrates, such as cellulose, and other large organic compounds, such as proteins and lignin. Lignin determines the toughness of leaves and many of the structural qualities of
wood. The lignin content of plants is a particularly important determinant of decomposition rate because it resists decomposition more than cellulose. However, some fungi and bacteria can break down cellulose and lignin. They secrete enzymes that break down the plant matter into simple sugars and amino acids that they then absorb. Some portion of the lignins, as well as other plant compounds that resist decomposition, may never break down in the surface soils but can form fossil fuels when buried for millions of years.
Figure 21.14 Decomposition of organic matter. Organic matter decomposes through the leaching action of water, consumption by invertebrates, mineralization by fungi, and mineralization by bacteria. Large detritivores, including millipedes, earthworms, and wood lice, also play an important role in decomposition. These animals can consume 30 to 45 percent of the energy available in leaf litter, but they consume a much lower fraction of the energy available in wood. The importance of large detritivores is twofold; they decompose organic matter directly and they macerate organic matter into smaller pieces of detritus, which have a greater surface-area-to-volume ratio. This gives bacteria and fungi more surfaces on which to act and increases the rate of decomposition. Bacteria and fungi play an important role in decomposition because they help convert organic matter into inorganic nutrients. Fungi play a special role
because the hyphae of fungi can penetrate the tissues of leaves and wood that large detritivores and bacteria cannot penetrate on their own. If you have ever walked through a forest, you may have seen the fruiting bodies of many different fungi, including the impressive shelf fungi that emerge from the sides of dead logs (Figure 21.15).
Figure 21.15 Fungi decomposing a dead log. Fungi play a key role in the decomposition of organic matter in terrestrial ecosystems by penetrating the dead tissues of plants. The fruiting bodies of this oyster bracket fungus (Pleurotus ostreatus), commonly referred to as mushrooms, are emerging from a dead log in Belgium. In terrestrial ecosystems, 90 percent of all plant matter produced in a given year is not consumed directly by herbivores but is ultimately decomposed. Many plants resorb some of the nutrients from their leaves before the leaves are dropped. The aboveground dead plant biomass, combined with the organic matter of dead animals and animal waste, drops onto the soil surface where nutrients are leached. Here, decomposition is primarily aerobic, and plant roots and their associated mycorrhizal fungi have ready access to the nutrients that are released by the decomposers. Because plant growth and decomposition are biochemical processes, nutrient cycling between producers and decomposers in terrestrial ecosystems is influenced by temperature, pH, and moisture. The rate of decomposition is also affected by the ratio of carbon and nitrogen in the organic matter. As we discussed in Chapter 20, differences in the stoichiometry of an organism’s
food can affect the consumption of the food and the number of consumers that can be supported by the food supply. In the case of decomposition, if the decomposers require high amounts of nitrogen, then low nitrogen availability in the organic matter can cause slower rates of decomposition. A study of leaf decomposition in Costa Rica provides insights into how several factors affect the rate of decomposition. In this study, leaves recently shed from 11 species of tropical trees were placed into coarse-mesh bags, as shown in Figure 21.16. The bags were positioned on the ground in the forest where invertebrates had access to the leaves, while the leaves remained in the bag. These invertebrates shredded the leaves into smaller pieces with an increased surface area, which led to more rapid decomposition. The leaf bags were weighed at different times over a 230-day period to determine the rate of leaf decomposition. You can view the data for several of these species in
Figure 21.17a. Leaf species with curves that decline faster over time, such as the Brazilian firetree (Schizolobium parahyba), have the highest rate of decomposition.
Figure 21.16 Researchers collected recently fallen leaves and placed them into mesh bags to determine the rate of daily mass loss over time in the forest.
Figure 21.17 Decomposition rates of leaves in a tropical forest. (a) A sample of 4 of the 11 species examined reveals a range of different decomposition rates. Error bars are standard errors. (b) During the first 50 days of decomposition, the decomposition rates of the 11 leaf species were positively correlated with the solubility of compounds that could be leached from each species. (c) After 50 days, the decomposition rates of the 11 leaf species were considerably lower and negatively correlated to the amount of lignin in each leaf species relative to the amount of nitrogen. Such species have relatively low amounts of lignin and cellulose and they have high leaf solubility, which allows more nutrients to be leached out of the leaf. The daily rate of mass loss, denoted as k, can be calculated for each curve. Species that decompose faster have a higher value of k. In “Analyzing Ecology: Calculating Decomposition Rates of Leaves,” we discuss these calculations in more detail. To examine how leaf traits affected decomposition rate, measured as the
##### Analyzing Ecology: Calculating Decomposition Rates of Leaves
daily rate of mass loss (k), the researchers focused on distinct time periods: from day 0 to 50 when most decomposition would occur due to leaching, and from day 51 to day 230 when most decomposition would be due to invertebrates, fungi, and bacteria. During the first 50 days, there was a positive relationship between the decomposition rate and the fraction of soluble compounds in the different leaf species that can be leached, as shown in Figure 21.17b. During the latter portion of the experiment, there was a negative relationship between the decomposition rate and the ratio of lignin to nitrogen, as you can see in Figure 21.17c. This means that leaf species with a relatively high amount of lignin experience a slower rate of decomposition. From this study, we see that the decomposition of organic matter depends on both the environmental conditions and chemical traits of the organic matter. Once researchers calculated values of k for each species, they could examine how both precipitation and the traits of the leaves affect the leaf decomposition rate. For example, they manipulated the amount of precipitation that some of the mesh bags received. When precipitation was reduced to 50 or 25 percent of normal levels, it caused a 10 to 20 percent decline in the rate of decomposition.
Analyzing Ecology
Calculating Decomposition Rates of Leaves As we discussed in the study of leaf decomposition in Costa Rica, researchers often want to examine the rate of leaf litter decomposition to determine how fast nutrients can be regenerated so they are available to producers, such as algae that form the base of an aquatic food web, plants that form the basis of a terrestrial food web, or last year’s crops that decompose and provide nutrients to the current year’s crop. The rate of decomposition commonly follows a negative exponential curve; there is initially a rapid loss in mass that slows over time. This negative exponential curve can be described by the following equation: mt=moe−kt
where mt is the mass of leaf litter that remains at a particular time, mo is the original mass of leaf litter, e is the base of the natural log, k is the daily rate of mass loss, and t is time, which is measured in days. The decay constant k is the key parameter in this equation because it determines the shape of the curve; leaves that decompose at a faster rate have a larger value of k. The value of k can be estimated using statistical software that determines the line that best fits a set of data for decomposition over time. Once the value of k is known, we can estimate the amount of leaf litter at any point in time, provided that the decomposition occurs under similar environmental conditions. For example, if we start with 100 g of leaves and k = 0.01, we can estimate the mass of leaves that have not decomposed after 10, 50, and 100 days: After 10 days: mt=mo e−kt=100 e−(0.01)(10)=90 gAfter 50 days: mt=mo e−kt=100 e−(0.01) (50)=61 gAfter 100 days: mt=mo e−kt=100 e−(0.01)(100)=37 g YOUR TURN Estimate the remaining mass of leaves that has not decomposed after 10, 50, and 100 days for two other leaf species: one with a daily decomposition rate of k = 0.05, and the other with a daily decomposition rate of k = 0.10.
Decomposition Rates Among Terrestrial Ecosystems
Because environmental conditions are a key determinant for rates of decomposition, terrestrial ecosystems differ a great deal in their decomposition rates. Comparative studies of temperate and tropical forests show that detritus in the tropics decomposes more rapidly because of warmer temperatures and higher amounts of precipitation. For example, we can compare the amount of dead plant matter on the forest floor—including leaves, branches, and logs—versus the total biomass of vegetation and detritus in a forest. The proportion of dead plant matter is about 20 percent in temperate coniferous forests, 5 percent in temperate hardwood forests, and only 1 to 2 percent in tropical rainforests. Of the total organic carbon in terrestrial ecosystems, more than 50 percent occurs in soil and litter in
northern forests, but less than 25 percent occurs in tropical rain forests, where the majority of the organic matter exists in the living biomass. These differences in litter decomposition rates mean that tropical forests have a much larger proportion of the total organic matter in living vegetation than in detritus. This has important implications for tropical agriculture and conservation. For example, when tropical forests are cleared and burned, a large fraction of the nutrients are mineralized by burning and by subsequent high rates of decomposition. Together, these processes create an abundance of nutrients during the first 2 to 3 years of crop growth, but any surplus nutrients not taken up by the crops quickly leach away. Traditionally, tropical areas burned for agricultural fields would be farmed for 2 to 3 years and then left to undergo natural succession for 50 to 100 years to rebuild the fertility of the soil. However, many regions have human populations that are too dense to allow rotation of agriculture into different areas over several decades. Without rotation, the soils cannot replenish their nutrients and the fertility of the land rapidly degrades.
Concept Check
1. What causes bedrock weathering? 2. How does taking a whole watershed approach aid researchers in quantifying rates of bedrock weathering? 3. What factors influence the rate at which organic matter is broken down?
#### 21.6 in Aquatic Ecosystems, Most Nutrients Regenerate in the Sediments
21.6 In aquatic ecosystems, most nutrients regenerate in the sediments Because most cycling of elements takes place in an aqueous medium, the chemical and biochemical processes involved are similar in terrestrial and aquatic ecosystems. However, the location of decomposition differs between terrestrial and aquatic ecosystems. In terrestrial ecosystems, nutrients regenerate close to the location, where they are taken up by producers. In aquatic ecosystems, however, most nutrients regenerate in sediments, which are often far below the surface waters that contain dominant producers, such as phytoplankton. In addition, in terrestrial ecosystems, aerobic decomposition is most common, whereas decomposition in the sediments and deep waters of aquatic ecosystems is typically anaerobic, which is considerably slower. In this section, we will investigate how decomposition operates in streams and wetlands, which receive much of their energy from leaves that blow in from the surrounding terrestrial environment. We will then examine the important role of sedimentation in rivers, lakes, and oceans and explore how stratification of water affects the movement of regenerated nutrients in lakes and oceans.
Allochthonous Inputs to Streams and Wetlands
As we have discussed in Chapter 6, streams and small, forested wetlands receive a major portion of their energy from the surrounding terrestrial environment from allochthonous inputs in the form of dead leaves that fall into the water. The decomposition process of leaves in a stream is similar to the process on land. As the leaves settle onto the bottom of the stream, the first stage is the leaching of soluble compounds, followed by the shredding of the leaves into smaller pieces by invertebrates such as amphipods, isopods, and larval caddisflies. At the same time that leaves are being shredded, fungi and bacteria are working to decompose the leaves much as they do on land. Given the similar processes in both the terrestrial and stream ecosystems, it is perhaps not surprising that the rate of leaf decomposition depends on the temperature of the water and the chemical composition of the leaves. To determine the rate at which leaves decompose in streams, aquatic ecologists follow a protocol that is similar to the protocol used by terrestrial ecologists.
Newly fallen leaves are collected, weighed, and placed into mesh bags that are submerged in the stream. The mesh bag allows aquatic invertebrates to enter the bag without losing any of the leaves. Over time, the bags are removed from the stream, dried, and reweighed to determine the amount of leaf mass that remains. To determine how leaf composition affects leaf decomposition rates, researchers placed coarse-mesh bags of nine leaf species into a stream located in the Black Forest of Germany and removed the bags over time. The nine leaf species differed a great deal in their nitrogen, phosphorus, and lignin content. The findings were that leaf decomposition rate is not related to the amount of nitrogen or phosphorus in the leaves but it is strongly associated with the lignin content of the leaves, as shown in Figure 21.18a.
Figure 21.18 Leaf decomposition in a stream. (a) When nine species of leaves in mesh bags were added to a stream in Germany, the decomposition rate was slower in leaf species that contained more lignin in their tissues. (b) The researchers also assessed the contribution of aquatic invertebrates to leaf decomposition by using large-mesh and fine-mesh leaf bags. After 55 days, the leaves in the fine-mesh bags, which excluded invertebrates, experienced 20 percent less decomposition compared to the large-mesh bags, which allowed invertebrates to enter. Error bars are standard errors. The researchers also were interested in knowing how important the invertebrates were in the decomposition process. To answer this question, a second set of leaves was placed into fine-mesh leaf bags that prevented invertebrates from entering. As you can see in Figure 21.18b, leaf decomposition was about 20 percent higher in bags where invertebrates had been allowed to enter. This demonstrates that the invertebrates play an important role in breaking down the organic matter. The small wetlands that exist in forests also receive a large proportion of
##### Decomposition and Sedimentation in Rivers, Lakes, and Oceans
their energy from leaves. Similar to leaves in streams, the decomposition of leaves in forested wetlands is largely tied to the lignin content. Moreover, the breakdown rate of the leaves has widespread effects on the entire food web and functioning of the ecosystem.
And Oceans
In most rivers, lakes, and oceans, organic matter sinks to the bottom and accumulates in deep layers of sediments. While some nutrients are recycled in the surface waters when animals excrete waste or when microbes in the surface water decompose organic matter, most organic matter sinks to the sediments. As a result, most nutrients must come from the sediments, although they will return slowly to the productive surface waters. The process of nutrient regeneration in sediments of aquatic ecosystems helps us understand many patterns of ecosystem productivity. For example, you might recall from Chapter 20 that the least productive aquatic ecosystems are the deep oceans where the benthos is far from the surface waters (see
Figure 20.6). Shallow oceans are more productive, in part, because the sediments are much closer to the surface waters and can therefore regenerate nutrients to the surface water more quickly through decomposition. The upwelling of water from the deep sediments to the surface brings nutrients from the site of regeneration to the site of algal productivity. In addition, some of the most productive regions of the ocean occur at the upwelling of water along the coasts of continents, where currents draw deep, nutrient-rich water up to the surface (see Figure 5.11).
Stratification of Lakes and Oceans
The stratification of water, a phenomenon that we discussed in Chapter 6, further affects the availability of nutrients in aquatic ecosystems. The vertical mixing of water from the sediments to the surface can be hindered whenever surface waters have a different temperature and therefore a different density than that found in deep waters. The vertical mixing can affect primary production in two opposing ways: it can bring the deep, nutrient-rich water to the surface where phytoplankton can use it; but it can also carry phytoplankton down to the deep water, where they will die because of the low light conditions. When phytoplankton die, primary production may shut
down in the deep water and little primary production will occur in the nutrient-rich waters at the surface. Stratification happens in temperate and tropical lakes when the surface waters are warmed by the summer sunlight, while the deeper waters stay cold and dense. Such stratification does not happen in polar lakes because their surfaces never become warm enough. In estuaries and oceans, stratification of the water happens when an input of less dense freshwater from rivers or melting glaciers is positioned above a layer of the denser ocean saltwater. Occasionally, stratified aquatic ecosystems experience periods of vertical mixing. For example, temperate lakes in the spring and fall experience changes in the temperature of surface waters that eventually match the temperature of the deeper waters. When the temperatures become equal, spring and fall winds that blow along the surface of the lakes cause the entire lake to mix. Mixing can also happen in oceans. In areas where the surface water is less salty than the deep water, sunlight can slowly cause the surface waters to evaporate and leave the salt behind. At some point, the surface water becomes saltier than the deeper water and the surface water sinks, thereby causing the ocean water to circulate. In this chapter, we have learned that elements cycle within and between ecosystems and that this cycling determines the availability of elements to organisms. We have also seen that human activities commonly affect element cycles in ways that are harmful to ecosystem function. Finally, we have seen how elements regenerate through weathering and the decomposition of organic matter. In “Ecology Today: Applying the Concepts,” we will apply these concepts to understand how logging and global change alter the functioning of an entire forest.
1. Why is ocean upwelling an important process in regenerating nutrients in deep ocean waters? 2. What processes contribute to the breakdown of allochthonous inputs in streams? 3. Why is most decomposition typically anaerobic in the deep waters of lakes and oceans?
Concepts
Nutrient Cycling in New Hampshire Monitoring nutrient flows from a watershed. At the Hubbard Brook Experimental Forest in New Hampshire, the soils lie on bedrock that prevents water from percolating down, so all nutrients leached from the soils find their way into the streams that drain the watershed. At the bottom of the watershed, researchers constructed a catchment device to monitor water volumes and nutrient flow from the watershed. As we have seen throughout this chapter, producers assimilate nutrients from the environment and hold the nutrients in their tissues to be passed on to consumers and decomposers. However, assessing the degree to which plants affect
nutrient cycling in an ecosystem is a daunting task, in part because ecosystems are large and complex. Fifty years ago, researchers at the Hubbard Brook Experimental Forest in New Hampshire took on this challenge by selecting a number of forested watersheds and logging some of them. Then they monitored the changes in how nutrients moved in each watershed. The Hubbard Brook Experimental Forest was an ideal location for this grand experiment. A layer of impenetrable rock under the soil prevents water from percolating into the deep groundwater. Because of this, all the precipitation falling on the watershed is either taken up by plants or passes over and through the soil, ultimately ending up in the stream that leaves the watershed. This allows the researchers to monitor the stream to measure the amount of water and nutrients leaving the ecosystem. In 1962, researchers removed all the trees from an entire watershed and sprayed it with herbicides for several years to suppress plant growth. As a control, adjacent watersheds were not logged. Over the past 50 years, ecologists have tracked how the ecosystem has responded to this disturbance. Without plants to take up water and nutrients, the movement of elements in the ecosystem changed dramatically. For example, the amount of water leaving the watershed in the stream increased several-fold. In addition, because the nitrates available in the soil were no longer being used by plants, there was a large increase in the amount of nitrates that leached out of the soil and into the streams. In forests that were not logged, there was a net gain of soil nitrogen over time at an annual rate of 1 to 3 kg per ha, due to precipitation and nitrogen fixation. In the logged forest, however, there was a net loss of nitrogen
at an annual rate of 54 kg per ha. Because nitrates do not bind well to soil, they leached out of the soil and into the stream after logging. Logging a watershed. To determine the role of plants in the cycling of nutrients, researchers at the Hubbard Brook Experimental Forest logged an entire watershed and then applied herbicides for several years to prevent plants from growing. Other watersheds were logged and allowed to regrow, while still others were left unmanipulated as controls. Many other nutrients were also affected by the logging. For example, the researchers tracked the movement of calcium ions that come from precipitation and weathering of the bedrock. They found the vast majority of calcium ions came from the decomposition of detritus, whereas less than 10 percent of the calcium derived from weathering. When the region experienced years of acid precipitation, the inputs of calcium from precipitation and weathering could not keep pace with the calcium that leached from the soil. As a result, the forest experienced a net loss of calcium.
Today, the Hubbard Brook Experimental Forest continues to provide insights into nutrient cycling. For example, researchers recently reported that over a 5decade period, the amount of nitrates leaving forested watersheds through the streams each year declined by more than 90 percent. Changes in the composition of tree species had only a small impact on the decline in nitrates leaving the watershed. This small impact was due to maple trees being naturally replaced by beech trees due to ecological succession; the leaves of beech trees decompose and release nitrates more slowly than the leaves of maple trees. Climate change was responsible for about 40 percent of the reduction in nitrates in the stream; warmer temperatures in the late fall and early spring in recent years give plants a longer time to take up the nitrates, so fewer nitrates leach out of the soil and into the stream. The remaining 50 to 60 percent of the nitrate reduction over the 5 decades was due to a recovery from historic disturbances that removed more than 20 percent of the trees, including logging that took place in 1906 and 1917, a hurricane in 1938, and an ice storm in 1998. Just as we saw with the clear-cutting of a watershed, tree removal causes a short-term flush of nitrates out of the soils and into the stream. As the forests begin to come back, however, the soil recovers and holds on to more of the available nitrates. As the soil accumulates nitrates, the amount of nitrates leached into the stream declines. Another surprising result came in 2016 when researchers reported on the effects of adding calcium to part of the forest in an attempt to reverse the long-term harmful effects of acid rain. The hypothesis was that the
added calcium would improve tree growth and that the trees would, in turn, take up more nitrates from the soil. Surprisingly, they found that calcium caused a 30-fold greater release of nitrates from the soil and into the streams. The revised hypothesis is that the addition of calcium created a more favorable environment for the microbes, allowing the microbes to increase their breakdown of organic matter, including an increase in the release of nitrates. The long-term experiment at Hubbard Brook shows us how natural events and human activities can dramatically alter the movement of elements within and between ecosystems in ways that can have major consequences for terrestrial and aquatic ecosystems. SOURCES: Bernal, S., et al. 2012. Complex response of the forest nitrogen cycle to climate change. PNAS 109: 3406–3411. Likens, G. E. 2004. Some perspectives on long-term biogeochemical research from the Hubbard Brook Ecosystem Study. Ecology 85: 2355–2362. Rosi-Marshall, E. J., et al. 2016. Acid rain mitigation shifts a forested watershed from a net sink to a net source of nitrogen. PNAS 113: 7580–7583.
Summary of Learning Objectives
21.1 The hydrologic cycle moves many elements through ecosystems. Water evaporates from water bodies, soil, and plants and moves as water vapor into the atmosphere. This water vapor condenses into clouds and eventually falls back to Earth as precipitation. This precipitation can be taken up by plants, run across the surface of the ground, or infiltrate the groundwater. This water moves to streams and lakes and ultimately makes its way back to the oceans. Humans can alter this cycle by reducing infiltration due to the construction of impermeable surfaces, by logging trees and thereby increasing runoff, and by causing global warming, which increases the rate of evaporation. Key Term: Hydrologic cycle
21.2 The carbon cycle is closely tied to the movement of energy. Carbon exists in the atmosphere in the form of CO2. This CO2 can be used by terrestrial producers and, after dissolving into the water, by aquatic producers. The producers, consumers, scavengers, detritivores, and decomposers in terrestrial and aquatic ecosystems can produce CO2 when they respire. Carbon can also leave the water by sedimentation and be buried both in the water and on land. Buried carbon can be extracted in the form of fossil fuels. The combustion of fossil fuels and the combustion of organic matter during fires release CO2 into the atmosphere. Humans can alter this cycle primarily by affecting the extraction and combustion of carbon.
21.3 Nitrogen cycles through ecosystems in many different forms. Nitrogen gas in the atmosphere can be converted into ammonia and nitrates by lightning, nitrogen-fixing bacteria, and the manufacture of fertilizers. Producers can take up these forms of nitrogen and assimilate them. This nitrogen is then transferred through terrestrial and aquatic food webs. During decomposition, nitrogen in
organisms and their wastes can be converted to ammonia by the process of mineralization. Ammonia can be converted into nitrites and nitrates through the process of nitrification, and nitrates can be converted to nitrous oxide and nitrogen gas by the process of denitrification. Humans alter this cycle primarily by manufacturing and applying large amounts of fertilizer and combusting fossil fuels, which produces nitric oxide in the air that later mixes with precipitation and falls to the ground as nitrates. These activities alter the fertility of terrestrial and aquatic environments. Key Terms: Nitrogen fixation, Nitrification, Mineralization, Denitrification
21.4 The phosphorus cycle moves between land and water. Most phosphorus is released by weathering of rocks. This phosphorus is taken up by terrestrial and aquatic producers, which pass it to consumers, scavengers, detritivores, and decomposers. The phosphorus from excretions and decomposed organisms dissolves in the water of soil or in the water of streams, rivers, lakes, and oceans. In the ocean, phosphorus precipitates into sediments that are slowly converted into rocks. Humans affect the phosphorus cycle primarily by mining rocks for fertilizer. This phosphorus-rich fertilizer can alter the fertility of terrestrial and aquatic habitats and lead to algal blooms and then eutrophication. Key Terms: Eutrophication, Cultural eutrophication
21.5 In terrestrial ecosystems, most nutrients regenerate in the soil. The nutrients in terrestrial ecosystems are primarily regenerated in soils. Some nutrients such as phosphorus are regenerated by weathering of rocks, and all nutrients are regenerated by the decomposition of dead organic matter. Because decomposition rates are faster under warm temperatures and high precipitation, tropical ecosystems have high decomposition rates and low amounts of dead organic matter. Boreal and other cold ecosystems have low rates of decomposition and large amounts of dead organic matter.
21.6 In aquatic ecosystems, most nutrients regenerate in the sediments. In many streams and some wetlands, allochthonous inputs of leaves from the surrounding terrestrial environment are the major source of nutrients. The rate of leaf decomposition is determined, primarily, by the water temperature and the lignin content of the leaves. In rivers, lakes, and streams, much of the organic matter settles out of the water and onto the bottom sediments, where it decomposes. When lakes and oceans stratify, the movement of nutrients that have been released by decomposition is hindered from moving to the more productive surface waters. Although nitrogen and phosphorus are the most common limiting nutrients in aquatic ecosystems, some regions of the ocean are limited by the availability of other nutrients, including silicon and iron.
Critical Thinking Questions
1. How does energy from the Sun drive the movement of water from the oceans to the continents and back to the oceans again? 2. How might the ocean reduce the effects of fossil fuel combustion on CO2 concentrations in the atmosphere? 3. Why is methane gas commonly produced in swamps? 4. Given that the bottom of the ocean is anaerobic, what process in the nitrogen cycle is likely to be occurring in this location? 5. How might nitrogen-fixing bacteria living in symbiosis with a plant affect the types of environments in which the plant could live? 6. Why is the weathering of bedrock in a New Hampshire forest responsible for such a small fraction of the nutrients available to plants? 7. Why do tropical and temperate soils have different rates of nutrient regeneration? 8. Why do agricultural soils in boreal Canada retain their nutrients for many more years than agricultural soils in tropical South America? 9. How might global warming cause the release of CO2 from boreal forest soils? 10. What is the likely chain of events in which the dumping of raw sewage into the Mississippi River leads to fish kills in the dead zone in the Gulf of Mexico?
#### Graphing the Data: the Decomposition of Organic Matter
GRAPHING THE DATA The Decomposition of Organic Matter As we have seen in this chapter, dead organic matter commonly decomposes in a pattern that follows a negative exponential curve. To compare the decomposition rates of leaves from cherry trees and maple trees in a stream, scientists placed leaf bags into a stream and retrieved them over time. They conducted three replicates of this experiment, and obtained the data shown in the table. For each species of leaf litter, calculate the mean amount of litter remaining at each time point and then graph the mean amount of litter remaining over time using a scatterplot. LEAF LITTER MASS (g) 0 DAYS 10 DAYS 30 DAYS 60 DAYS 100 DAYS