2 Microbial Cell Structure and Function
## Chapter 2 Microbial Cell Structure and Function
Exploring the Microbial Cell
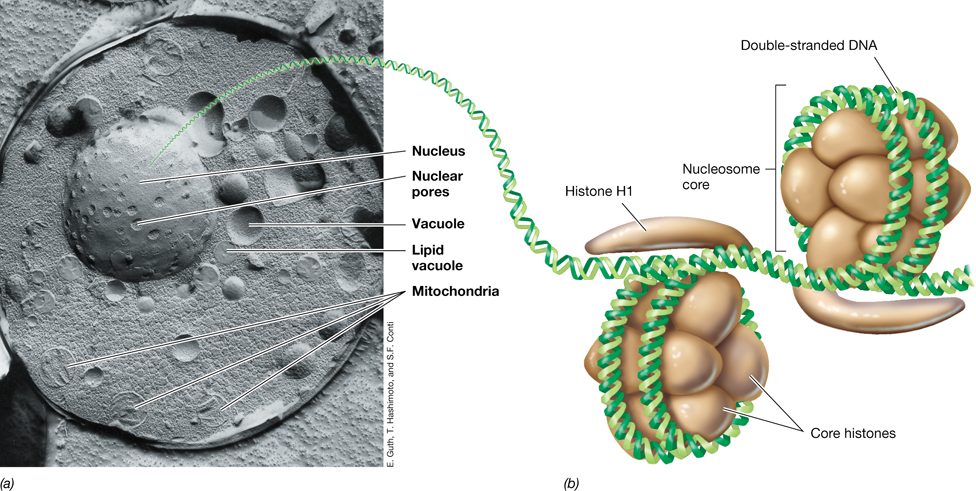
New microscopic techniques are changing how we view microbial cells. Many cellular processes are performed by macromolecules and structures whose forms dictate their function. These structures are incredibly small and impossible to resolve with standard light microscopy. Cryogenic electron tomography (cryoET) images cells embedded within vitreous ice at ultra-low temperatures so they can be viewed in their native state without dehydration. During cryoET, samples are tilted in an electron microscope and images made at many different angles, much in the same way that a CT scan images a human body. These images are integrated using computer algorithms to generate complex, high-resolution, three-dimensional images that provide unprecedented structural detail. CryoET images have led to the discovery of new features within microbial cells and stunning new insights on well-known structures.
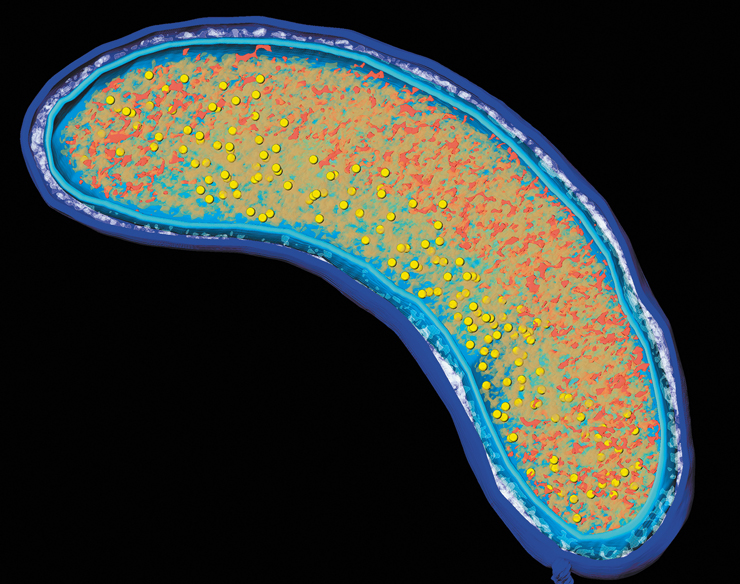
“Candidatus Pelagibacter ubique,” one of the most abundant organisms on Earth, lives in microbial communities suspended in the upper waters of oceans worldwide. Cells of Pelagibacter are quite small (about 0.8 μm long), but their structures are readily visualized by cryoET (see image). Colors in the cryoET reconstruction indicate the outer membrane (blue), cytoplasmic membrane (cyan), peptidoglycan (white), cytoplasm (orange), nucleoid (red), and ribosomes (yellow). These new images reveal that the periplasm (the space between inner and outer membrane) in these cells is larger than expected and can occupy a remarkable 50–70% of cell volume in nongrowing cells. These images also reveal that the cytoplasm is divided into distinct regions that contain either nucleoid or ribosomes. Finally, mysterious structures such as membrane vesicles have been observed on the extracellular surface of the outer membrane and tethered to the cytoplasmic membrane (not shown). The function of these vesicles is unknown. New imaging techniques such as cryoET move microbiology forward and are revealing surprising complexity and diversity in the structures of prokaryotic cells.
Source: Zhao, X., et al. 2017. Three-dimensional structure of the ultraoligotrophic marine bacterium “Candidatus Pelagibacter ubique.” Appl. Environ. Microbiol. 83: e02807-16.
In this chapter, we consider the structure and function of microbial cells, both prokaryotic and eukaryotic. We will explore the components of microbial cells that control nutrient transport and cellular integrity but also several internal structures, which can be found in certain cells. We will also examine structures that give microbial cells the ability to move within their environment. We begin with two critical cell structures—the cytoplasmic membrane and cell wall—that collectively compose the cell envelope.
I The Cell Envelope
The cytoplasm of the prokaryotic cell is surrounded by several layers that include a membrane, a cell wall, and other structures specific to the kind of cell. Collectively, these layers form the cell envelope and are essential for preserving the integrity and functioning of the cell.
The cell envelope consists of a series of layered structures that surround the cytoplasm and govern cellular interactions with the external environment. The cell envelope has many important functions: It governs transport of nutrients into the cell and wastes out of the cell, it is the site of energy conservation, it governs cell shape, it protects the cell from mechanical stress, and it can help the cell attach to surfaces and even protect the cell from attack.
The diversity of microorganisms is in part reflected in the composition of the cell envelope; but we will also learn that certain envelope structures are highly conserved and that knowledge of cell envelope structure can help us to identify and classify microorganisms. In the first few sections, we will learn about major components of cell envelope structure including the cytoplasmic membrane, cell wall, outer membrane, and S-layers. We start our tour by considering the cytoplasmic membrane, a structure found in all cells.
2.1 The Cytoplasmic Membrane
The cytoplasmic membrane surrounds the cytoplasm—the mixture of macromolecules and small molecules inside the cell—and separates it from the environment. The cytoplasmic membrane is physically rather weak but is an ideal structure for its major cellular function: selective permeability. In order for a cell to grow, nutrients must be transported inwards and waste products outwards. Both of these events occur across the cytoplasmic membrane. A variety of proteins located in the cytoplasmic membrane facilitate these reactions, and many other membrane proteins play important roles in energy metabolism.
Bacterial Cytoplasmic Membranes
The cytoplasmic membrane of all bacterial and eukaryal cells is a phospholipid bilayer containing embedded proteins. The cytoplasmic membrane is only 8–10 nanometers wide but can be resolved easily by transmission electron microscopy (**Figure 2.1*a***). Phospholipids are composed of both hydrophobic (water-repelling) and hydrophilic (water-attracting) components (Figure 2.1b). In Bacteria and Eukarya, the hydrophobic component consists of fatty acid “tails” and the hydrophilic component consists of a glycerophosphate (a glycerol molecule bound to a phosphate) and one of several other functional groups (such as sugars, ethanolamine, or choline) also bonded to the phosphate (Figure 2.1c). The membrane is comprised of two phospholipid layers in which the fatty acid tails associate together to form a hydrophobic region, leaving the hydrophilic “head groups” exposed to either the environment or the cytoplasm (Figure 2.1b). That is, the outer surface of the cytoplasmic membrane faces the environment while the inner surface faces the cytoplasm and interacts with the cytoplasmic milieu. This type of membrane structure is called a lipid bilayer, or a unit membrane.
Figure 2.1 Phospholipid bilayer membrane.

(a) Transmission electron micrograph of a cell with membrane region shown in detail. (b) General architecture of a bilayer membrane; phospholipids are composed of a hydrophilic head group (blue spheres) with fatty acid tails (yellow lines). The phospholipid head groups of the bilayer are visible in a as parallel dark lines, between which can be found a lighter region comprising the hydrophobic region of the membrane. (c) Structure of the phospholipid phosphatidylethanolamine. Each fatty acid side chain is connected to the head group by an ester bond (boxed with a red dashed line); ester linkages are a characteristic feature of lipids from Bacteria and Eukarya but not those of Archaea.
Mastering Microbiology
Art Activity: Figure 2.5 Structure of the cytoplasmic membrane
A variety of proteins are attached to or integrated into the cytoplasmic membrane; membrane proteins typically have hydrophobic domains that span the membrane and hydrophilic domains that contact the environment or the cytoplasm (Figure 2.2). Proteins significantly embedded in the membrane are called integral membrane proteins. Many, though not all, integral membrane proteins extend completely across the membrane, and these are called transmembrane proteins. By contrast, peripheral membrane proteins are more loosely attached. Some peripheral membrane proteins are lipoproteins, proteins that contain a hydrophobic lipid tail that anchors the protein into the membrane. Other peripheral membrane proteins have residues that associate with the hydrophilic head groups of phospholipids, or they associate indirectly with membranes by binding to other proteins anchored in the membrane. Peripheral membrane proteins typically interact with integral membrane proteins in important cellular processes such as energy metabolism and transport.
Figure 2.2 Structure of the cytoplasmic membrane.

The inner surface (In) faces the cytoplasm and the outer surface (Out) faces the environment. Phospholipids compose the matrix of the cytoplasmic membrane with proteins embedded (integral) or surface associated (peripheral). The general design of the cytoplasmic membrane is similar in both prokaryotic and eukaryotic cells, although there can be differences in the chemistry between different species. Note that this membrane is shown in a relaxed shape to better illustrate its inner and outer surfaces; in a living cell, cytoplasmic turgor pressure would cause the membrane to have convex curvature.
Archaeal Cytoplasmic Membranes
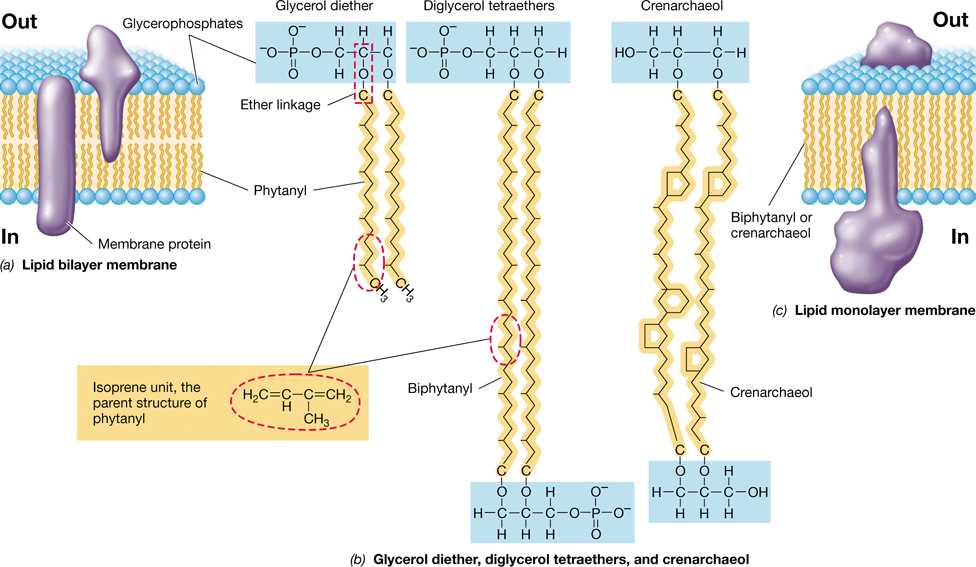
The cytoplasmic membrane of Archaea is structurally similar to those of Bacteria and Eukarya, but the chemistry is somewhat different. In the lipids of Bacteria and Eukarya the hydrophobic fatty acid tails are bound to glycerol by ester linkages (Figure 2.1); in contrast, the lipids of Archaea have hydrophobic isoprenoid (rather than fatty acid) tails, which are bound to glycerol by ether bonds (Figure 2.3). The hydrophobic region of archaeal membranes is formed from repeating units of the five-carbon hydrocarbon isoprene, rather than from fatty acids (compare Figures 2.1 and 2.3).
Figure 2.3 Major lipids of ***Archaea*** and the architecture of archaeal membranes.

(a, b) Archaea can have lipid bilayers composed of phosphoglycerol diether lipids. The hydrophobic portions of archaeal lipids are comprised of isoprenoid chains synthesized from repeated units of isoprene (in dashed red ovals); this contrasts with the lipids of Bacteria and Eukarya, which have fatty acid tails (Figure 2.1). Note that these isoprenoids are bonded to glycerol by an ether linkage (in dashed red box). (b, c) Some Archaea can also have lipid monolayers composed of diphosphoglycerol tetraether lipids or other isoprenoid lipids such as crenarchaeol. The isoprenoid lipids in b are phytanyl (C20), biphytanyl (C40), and crenarchaeol. Isoprene lipids can often contain 5- and 6-carbon rings such as those present in crenarchaeol. The membrane structure in Archaea may form a lipid bilayer or a lipid monolayer (or a mix of both).
The cytoplasmic membrane of Archaea is constructed from either phosphoglycerol diethers, which can have C20 side chains (called a phytanyl group), or diphosphoglycerol tetraethers (C40 side chains, called a biphytanyl group) (Figure 2.3). In the tetraether lipid structure, the ends of the inwardly pointing isoprenoid chains are covalently linked at their termini to form a lipid monolayer (Figure 2.3c) instead of a lipid bilayer (Figure 2.3a) membrane.
Archaeal lipids can have many different isoprenoid chains including some that contain ring structures. For example, crenarchaeol, a common membrane lipid in cells of Thaumarchaeota (a major phylum of Archaea, Section 17.5) contains four C5 rings and one C6 ring (Figure 2.3b). These rings affect the chemical properties of the lipids and thus influence membrane function. As in other organisms, the polar head groups in archaeal lipids can be sugars, ethanolamine, or a variety of other molecules.
Despite differences in chemistry between the cytoplasmic membranes of Archaea and organisms in the other phylogenetic domains, the fundamental construction of the archaeal cytoplasmic membrane—inner and outer hydrophilic surfaces and a hydrophobic interior—is the same as that of membranes in all cells. Obviously, evolution has selected this fundamental design as the best solution to the major functions of the cytoplasmic membrane, an issue we turn to now.
Cytoplasmic Membrane Function
The cytoplasmic membrane has at least three major functions (Figure 2.4). First, it is the cell’s permeability barrier, preventing the passive leakage of solutes into or out of the cell. Second, the cytoplasmic membrane anchors several proteins that catalyze a suite of key cell functions. And third, the cytoplasmic membrane of Bacteria and Archaea plays a major role in energy conservation and consumption.
Figure 2.4 The major functions of the cytoplasmic membrane.

Although physically weak, the cytoplasmic membrane controls at least three critically important cellular functions: maintaining selective permeability, anchoring proteins, and conserving energy.
The cytoplasmic membrane is a barrier to the diffusion of most substances, especially polar or charged molecules. Because the cytoplasmic membrane is so impermeable, most substances that enter or leave the cell must be carried in or out by transport proteins. These are not simply ferrying proteins but instead function to accumulate solutes against the concentration gradient, a process that diffusion alone cannot do (Figure 2.5). Transport, which requires energy, ensures that the cytoplasm has sufficient concentrations of the nutrients it needs to perform biochemical reactions efficiently.
Figure 2.5 The importance of transport in membrane function.


In both types of transport, the uptake rate shows saturation at relatively low external solute concentrations. Both high-affinity and low-affinity transport systems are depicted.
Transport proteins typically display high sensitivity and high specificity. If the concentration of a solute is high enough to saturate the transporter, which often occurs at the very low concentrations of nutrients found in nature, the rate of uptake can be near maximal (Figure 2.5). Some nutrients are transported by a low-affinity transporter when the nutrient is present at high external concentration and by a separate, typically higher-affinity, transporter when the nutrient is present at low concentration (Figure 2.5). Moreover, many transport proteins transport only a single kind of molecule while others carry a related class of molecules, such as different sugars or different amino acids. This economizing reduces the need for separate transport proteins for each different sugar or amino acid.
In addition to its permeability and transport functions, the cytoplasmic membrane of Bacteria and Archaea is a major site of both energy conservation and energy consumption. We discuss in Chapter 3 how the cytoplasmic membrane can be energized when protons (H+) are separated from hydroxyl ions (OH−) across the membrane surface (Figure 2.4c). This charge separation creates an energized state of the membrane called the proton motive force, analogous to the potential energy present in a charged battery. Dissipation of the proton motive force can be coupled to several energy-requiring reactions, such as transport, cell locomotion, and the biosynthesis of ATP. In eukaryotic microbial cells, although transport across the cytoplasmic membrane is just as necessary as it is in prokaryotic cells, energy conservation takes place in the membrane systems of the cell’s key organelles, the mitochondrion (respiration) and chloroplast (photosynthesis), as we will see later in this chapter.
Check Your Understanding
Draw the basic structure of a lipid bilayer and label the hydrophilic and hydrophobic regions. Why is the cytoplasmic membrane a good permeability barrier?
How are the membrane lipids of Bacteria and Archaea similar, and how do they differ?
Describe the major functions of the cytoplasmic membrane.
2.2 Transporting Nutrients into the Cell
In Section 2.1 we learned how the structure of the cytoplasmic membrane is an effective barrier to leakage; solutes leak neither into nor out of a living cell. However, selective transport is a major function of the cytoplasmic membrane. In order for cells to survive and grow, they must transport nutrients across the membrane and into the cell and they must export wastes out of the cell. Substances are transported across the cytoplasmic membrane through membrane-spanning integral membrane proteins (Figure 2.2). Some transporters also require the action of peripheral membrane proteins. We consider the most common of these transport systems here, with a focus on the well-studied transporters widespread in Bacteria and Archaea.
Active Transport and Transporters
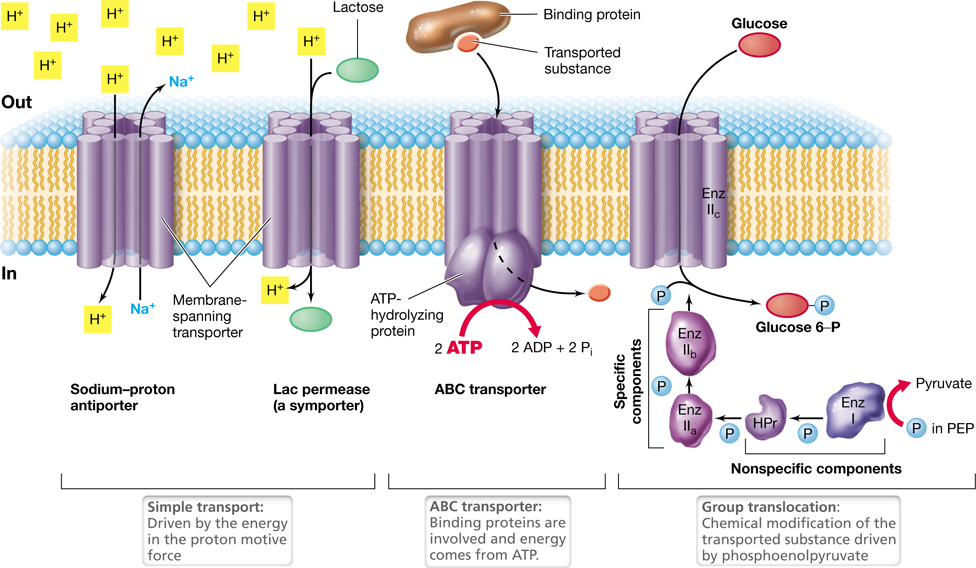
Active transport is the process by which cells accumulate solutes against the concentration gradient. Three basic mechanisms of active transport are found in prokaryotic cells. A simple transport system consists only of a transmembrane transport protein, group translocation employs a series of proteins in the transport event, and ABC transport systems consist of three components: a binding protein, a transmembrane transporter, and an ATP-hydrolyzing protein (Figure 2.6). Each of these transport systems is energy-driven, be it from the proton motive force, ATP, or some other energy-rich compound.
Figure 2.6 The three classes of transport systems.

Transmembrane transporters are generally composed of a polypeptide that has 12 α-helices (each shown as a cylinder) that aggregate to form a channel through which solutes can cross the membrane. In simple transport the movement of a solute is coupled with the dissipation of an electrochemical gradient such as the proton motive force. ABC transporters have three components: a binding protein that has high affinity for a substrate, a transmembrane protein channel, and a cytoplasmic ATP-hydrolyzing protein, which supplies the energy required to drive substrate transport. In group translocation, the substance transported is chemically modified upon entering the cell. For example, the glucose group translocation system has five proteins: Enzyme (Enz) I, Enzymes IIa, IIb, and IIc, and HPr. A phosphate cascade occurs from phosphoenolpyruvate (PEP) to Enz IIc, and the latter protein actually transports and simultaneously phosphorylates the sugar. Proteins HPr and Enz I are nonspecific and participate in the transport of any sugar, while the three components of Enz II are specific for a particular sugar. Note how simple transporters and the ABC system transport substances without chemically modifying them, whereas group translocation results in chemical modification (in this case phosphorylation) of the transported substance.
The transmembrane component of virtually all transport systems is composed of a polypeptide containing 12 regions (called domains) that weave back and forth through the membrane to form a channel, and it is through this channel that the solute is transported into the cell. Transport is linked to a conformational change in this transmembrane protein complex that occurs when it binds its specific solute. Like a gate swinging open, this conformational change sweeps the solute into the cell.
Simple Transporters and Group Translocation
Simple transport reactions are driven by the energy inherent in the proton motive force (Figure 2.4c). The two major transport events catalyzed are either symport reactions (where a solute and a proton are cotransported in the same direction) or antiport reactions (where a solute and a proton are transported in opposite directions) (Figure 2.6). A classic example of a simple transporter is the uptake of the sugar lactose by way of the lac permease, a well-studied symporter in Escherichia coli. As each lactose molecule enters the cell, the potential energy in the proton motive force is diminished slightly by the cotransport of a proton (Figures 2.4c and 2.6). The net result is the energy-driven accumulation of lactose in the cytoplasm against the concentration gradient. Many other solutes enter by the activity of their own simple symporters, including phosphate, sulfate, and several different organic compounds.
Group translocation differs from simple transport in two important ways: (1) the transported substance is chemically modified during the transport process, and (2) an energy-rich organic compound (rather than the proton motive force) drives the transport event. The best-studied group translocation systems transport the sugars glucose, mannose, and fructose in E. coli. During uptake, these compounds are phosphorylated by the phosphotransferase system (Figure 2.6). The phosphotransferase system consists of a family of five proteins that work in concert to transport any given sugar. Before the sugar is transported, the proteins in the phosphotransferase system are themselves alternately phosphorylated and dephosphorylated in a cascading fashion until Enzyme IIc phosphorylates the sugar as it enters the cytoplasm (Figure 2.6). A protein called HPr, the enzyme that phosphorylates HPr (Enzyme I), and Enzyme IIa are all cytoplasmic proteins. By contrast, Enzyme IIb is a peripheral membrane protein and Enzyme IIc is the transmembrane component.
In the phosphotransferase system, HPr and Enzyme I are nonspecific components and participate in the uptake of several different sugars. By contrast, distinct Enzyme II proteins exist, one set for each different sugar transported. Energy to drive the phosphotransferase system comes from phosphoenolpyruvate, an energy-rich intermediate in glycolysis (Sections 3.4 and 3.6).
ABC Transporter Systems
ABC transporters are modular systems that have three components: a binding protein, a transmembrane protein channel, and an ATP-hydrolyzing protein (Figure 2.6). The ABC stands for ATP-binding cassette, a structural feature of proteins that bind ATP. More than 200 different ABC transport systems are known, and these catalyze the uptake of a wide variety of organic and inorganic compounds.
Substrate-binding proteins are present outside of the cell, where they bind to a specific substrate and enable its transport into the cell. A characteristic property of binding proteins is their extremely high substrate affinity. These proteins can bind their specific substrate even when it is present at extremely low concentration; for example, less than 1 micromolar (10−6 M) Once its specific substrate is bound, the binding protein interacts with its respective transmembrane component to transport the substrate into the cell driven by the energy in ATP (Figure 2.6).
We move on now from our coverage of the cytoplasmic membrane to consider components of the cell envelope that confer structural strength on the cell, something the membrane cannot do.
Check Your Understanding
Compare and contrast simple transporters, the phosphotransferase system, and ABC transporters in terms of (1) energy source, (2) chemical alterations of the solute during transport, and (3) number of proteins required.
Which major characteristic of ABC transport systems makes them ideal for organisms living in nutrient-poor environments?
2.3 The Cell Wall
The cytoplasm of prokaryotic cells maintains a high concentration of dissolved solutes that creates significant osmotic pressure—about 2 atm (203 kPa); this is about the same as the pressure in an automobile tire. This osmotic pressure is sufficient to cause the cell membrane to burst and the cell to die—a process called cell lysis. To withstand this turgor pressure, the cell envelopes of most Bacteria and Archaea have a layer outside the cytoplasmic membrane called the cell wall. Besides protecting against osmotic lysis, cell walls also maintain cell shape and rigidity.
The cell envelopes of most Bacteria can be classified as being either gram-positive or gram-negative based on their organization and cell wall structures. The structures of gram-positive and gram-negative cell envelopes differ markedly as viewed in the electron microscope (Figure 2.7). The cell envelope of a gram-positive cell typically contains a cytoplasmic membrane and a thick cell wall, whereas a gram-negative cell has a cytoplasmic membrane, a thin cell wall, an outer membrane (Figure 2.7), and a periplasm, which is a compartment between the cytoplasmic and outer membranes. We will consider the outer membrane and periplasm in the next section. The names we use to describe the typical gram-positive and gram-negative cell envelopes are based on their Gram stain reactions (Section 1.8). The Gram stain reaction is determined primarily by the thickness of the cell wall rather than the number of layers in the cell envelope, and so Gram stain reaction does not always correlate with cell envelope structure. However, the Gram stain reaction is sufficiently predictive of cell envelope structure in Bacteria that the names of the two most common bacterial cell envelopes—gram-positive and gram-negative—are based on their typical reactions to the Gram stain.
Figure 2.7 Cell envelopes of ***Bacteria.***

(a, b) Schematic diagrams of gram-positive and gram-negative cell envelopes; the Gram stain procedure was described in Section 1.8 and shown in Figure 1.23. The photo of Gram-stained bacteria in the center shows cells of Staphylococcus aureus (purple, gram-positive) and Escherichia coli (pink, gram-negative). (c, d) Transmission electron micrographs showing the cell wall of a gram-positive bacterium and a gram-negative bacterium, respectively. (e, f) Scanning electron micrographs of gram-positive and gram-negative bacteria, respectively. Note differences in surface texture. Each cell is about 1 μm wide.
Knowledge of cell wall envelope structure and function is important not only for understanding the biology of microbial cells but also for medical reasons. Certain antibiotics, for example, the penicillins and cephalosporins, target bacterial cell wall synthesis, leaving the cell susceptible to osmotic lysis. Since human and other animal cells lack cell walls and are therefore not a target of such antibiotics, these drugs are of obvious benefit for treating bacterial infections. The major component of the bacterial cell wall, and a target of many antibiotics, is a molecule called peptidoglycan, and we consider this molecule in detail now.
Bacterial Cell Walls
The cell walls found in Bacteria contain a rigid polysaccharide called peptidoglycan that confers structural strength on the cell. Peptidoglycan is found in all Bacteria that contain a cell wall, but it is unique to Bacteria and is not found in Archaea or Eukarya. The sugar backbone of peptidoglycan is composed of alternating repeats of two modified glucose residues called N-acetylglucosamine and N-acetylmuramic acid joined by a β-1,4 linkage (Figure 2.8). Attached to the latter residue is a short peptide side chain. The amino acid composition of this peptide side chain can vary considerably between bacterial species. In Escherichia coli this peptide contains the amino acids L-alanine, D-alanine, D-glutamic acid, and diaminopimelic acid (DAP), though in other bacteria, L-lysine can be substituted for DAP. The presence of d stereoisomer amino acids, D-alanine and D-glutamic acid, is an unusual feature of peptidoglycan since proteins are always constructed of L-amino acids. These constituents are connected in an ordered way to form the glycan tetrapeptide (Figure 2.8), and long chains of this basic unit form peptidoglycan.
Figure 2.8 Structure of the repeating unit in peptidoglycan, the glycan tetrapeptide.

The structure given is that for the peptidoglycan of Escherichia coli and most other gram-negative Bacteria. Cross-links can be formed between adjacent peptide side chains at residues having free amino and carboxyl groups (circled in blue). For example, cross-links in E. coli most commonly occur between the amino group of diaminopimelic acid on one peptide and the terminal carboxyl group of D-alanine on a different peptide.
Strands of peptidoglycan run parallel to each other around the circumference of the cell (Figure 2.9). The peptide side chains of adjacent peptidoglycan strands are cross-linked together by covalent peptide bonds (Figure 2.9a), and in this way, the peptidoglycan forms one single enormous molecule. In gram-negative bacteria, the cross-links form primarily between the amino group of DAP on one glycan strand and the carboxyl group of the terminal D-alanine on the adjacent glycan strand (Figure 2.9a). The cell wall in the gram-negative cell envelope is 2–7 nm thick consisting primarily of a single layer of peptidoglycan, though it can be up to three layers thick in some places (Figure 2.9c).The peptidoglycan mesh so formed is flexible and porous, but strong enough to resist turgor pressure and prevent rupture of the cytoplasmic membrane and cell lysis. Additional strength against osmotic lysis in gram-negative bacteria is provided by the outer membrane, as discussed in the next section.
Figure 2.9 Peptidoglycan structure in the cell wall.

(a) Gram-negative cells that have thin cell walls, such as the cell wall of E. coli, mostly have direct cross-links between peptide side chains. (b) Gram-positive cells that have thick cell walls, such as S. aureus, can also have peptide interbridges that extend between cross-linked peptide side chains. (c) Conformation of peptidoglycan in the gram-negative cell wall. G, N-acetylglucosamine; M, N-acetylmuramic acid. Note how glycosidic bonds confer strength on peptidoglycan around the circumference of the cell whereas peptide bonds confer strength along the axis of the cell.
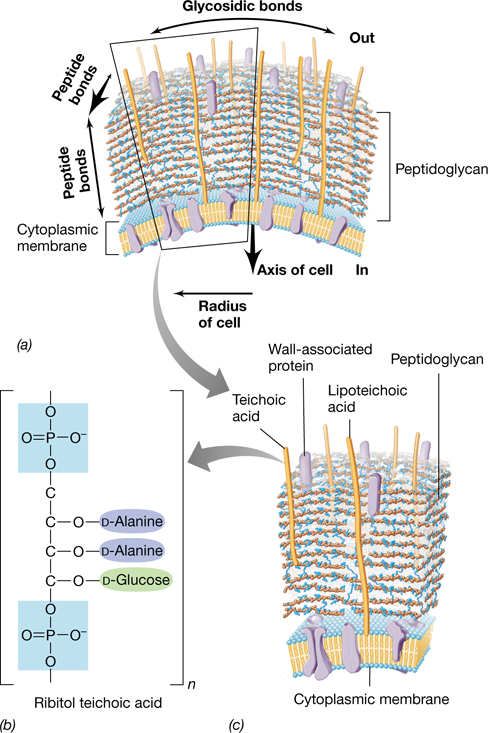
The typical bacterial gram-positive cell envelope contains a thick peptidoglycan cell wall, which can measure 20 to 35 nm in thickness and is usually much thicker than the wall of gram-negative organisms. As much as 90% of the gram-positive cell envelope can consist of peptidoglycan. Whereas the gram-negative cell wall typically contains only a single layer of peptidoglycan, the gram-positive cell wall can be 15 or more layers thick (**Figure 2.10*a***). The peptidoglycan of the gram-positive cell wall is stabilized three-dimensionally by peptide cross-links, which form between adjacent peptidoglycan strands both horizontally and vertically. In gram-positive bacteria, peptide cross-links often contain a short peptide “interbridge,” the kinds and numbers of amino acids in the interbridge varying between species. In the gram-positive bacterium Staphylococcus aureus, for example, the interbridge often consists of five glycines (Figure 2.9b).
Figure 2.10 Structure of the gram-positive bacterial cell wall.

(a) Schematic of a gram-positive cell wall showing the internal architecture of the peptidoglycan and its relationship to teichoic acids. Peptide cross-links form between peptidoglycan strands that are adjacent both horizontally and vertically, and peptidoglycan can also form covalent bonds to teichoic acids. (b) Structure of a ribitol teichoic acid. The teichoic acid is a polymer of the repeating ribitol unit shown here. (c) Summary diagram of the gram-positive bacterial cell wall. Lipoteichoic acids tether the cell wall to the cell membrane.
In addition to peptidoglycan, many gram-positive bacteria produce acidic molecules called teichoic acids embedded in their cell wall (Figure 2.10). Teichoic acids are composed of glycerol phosphate or ribitol phosphate with attached molecules of glucose or D-alanine (or both). Individual alcohol molecules are then connected through their phosphate groups to form long strands, and these are then covalently linked to peptidoglycan (Figure 2.10b). Some teichoic acids are covalently bonded to membrane lipids rather than to peptidoglycan, and these are called lipoteichoic acids.
Mastering Microbiology
Art Activity: Figure 2.12c Parts of the Gram-positive cell wall
Peptidoglycan can be destroyed by lysozyme, an enzyme that cleaves the glycosidic bond between N-acetylglucosamine and N-acetylmuramic acid (Figure 2.8). This weakens the peptidoglycan and can cause cell lysis. Lysozyme is present in human secretions including tears, saliva, and other bodily fluids, and functions as a major line of defense against bacterial infection. Many antibiotics, including penicillin, also target peptidoglycan. Whereas lysozyme destroys preexisting peptidoglycan, penicillin blocks the formation of peptide cross-links, which compromises the strength of the peptidoglycan, leading to cell lysis.
Archaeal Cell Walls
The cell envelopes of Archaea differ in fundamental ways from those of Bacteria. We have already learned that the cytoplasmic membranes of Archaea, while functionally analogous to those of Bacteria, differ in chemical structure (Section 2.1). Another major difference is that Archaea lack peptidoglycan. In addition, Archaea typically lack an outer membrane (see Section 2.4). One consequence of these differences is that the Gram stain reaction is not very useful for predicting the structures of archaeal cell envelopes and so we typically do not use the terms gram-positive and gram-negative to describe cells of Archaea. Most Archaea lack a polysaccharide-containing cell wall and instead have an S-layer (see Section 2.5), which is a rigid protein shell that functions to prevent osmotic lysis just as does the bacterial cell wall.
While some Archaea do have cell walls, these walls have unique chemical structures not found in Bacteria. For example, the cell walls of certain methane-producing Archaea (methanogens) contain a polysaccharide called pseudomurein (Figure 2.11), which is structurally remarkably similar to peptidoglycan (the term murein is from the Latin word for “wall” and was an old term for peptidoglycan). The backbone of pseudomurein is formed from alternating repeats of N-acetylglucosamine (also present in peptidoglycan) and N-acetyltalosaminuronic acid; the latter replaces the N-acetylmuramic acid of peptidoglycan. Pseudomurein also differs from peptidoglycan in that the glycosidic bonds between the sugar derivatives are β-1,3 instead of β-1,4, and the amino acids are all of the l stereoisomer (compare Figures 2.9 and 2.11).
Figure 2.11 Pseudomurein.
Structure of pseudomurein, the cell wall polymer of Methanobacterium species. Note the similarities and differences between pseudomurein and peptidoglycan (Figures 2.8 and 2.9).
Because in many respects they are so similar, it is likely that peptidoglycan and pseudomurein are variants of a cell wall polysaccharide originally present in the common ancestor of Bacteria and Archaea. However, although they are structurally and functionally very similar, they differ sufficiently that pseudomurein is immune from destruction by both lysozyme and penicillin, molecules that destroy peptidoglycan.
Check Your Understanding
Describe the major differences between the cell walls of gram-negative and gram-positive bacteria.
Explain whether you expect the enzyme lysozyme to be equally effective against Bacteria and Archaea.
2.4 LPS: The Outer Membrane
Most of the gram-negative cell envelope is composed of the outer membrane. The outer membrane is a second lipid bilayer found external to the cell wall (Figure 2.12), but its structure and function differs from that of the cytoplasmic membrane. The outer membrane and cytoplasmic membrane are similar in that they both contain phospholipid and protein, but a major difference is that the outer membrane also contains polysaccharide molecules covalently bound to lipids (Figure 2.12). Hence, the outer membrane is often called the lipopolysaccharide layer, or simply LPS for short.
Mastering Microbiology
Art Activity: **Figure 2.14*a*** Gram-negative cell wall structure
Figure 2.12 The gram-negative bacterial cell envelope.

(a) Arrangement of lipopolysaccharide, lipid A, phospholipid, porins, and Braun lipoprotein in the outer membrane. See Figure 2.13 for details of the structure of LPS. (b) Transmission electron micrograph of a cell of Escherichia coli showing the cytoplasmic membrane and wall. (c) Molecular model of porin proteins showing their hollow pores that allow solute transport across the outer membrane. The view of the porin is perpendicular to the plane of the membrane. The black dashed circle highlights some of the hydrophilic amino acids that line the inside of the pore.
LPS molecules have several unique functions: They can facilitate surface recognition, they are important virulence factors for some bacterial pathogens, and they contribute to the mechanical strength of the cell. We will see that another major difference between the cytoplasmic and outer membranes is that the outer membrane contains porins, which are transmembrane proteins that allow for the nonspecific transport of solutes. Hence, we will see that the outer membrane is far more permeable than is the cytoplasmic membrane.
Structure and Activity of LPS
While the precise chemistry of LPS can vary among different species of bacteria, these molecules have several common features. As seen in Figure 2.13, LPS contains a polysaccharide that consists of two components, the core polysaccharide and the O-specific polysaccharide (see also Figure 2.12a). In Salmonella species, where LPS has been well studied, the core polysaccharide consists of ketodeoxyoctonate (KDO), various seven-carbon sugars (heptoses), the hexose sugars glucose and galactose, and N-acetylglucosamine. Connected to the core is the O-specific polysaccharide, which typically contains galactose, glucose, the hexoses rhamnose and mannose, and one or more dideoxyhexoses, such as abequose, colitose, paratose, or tyvelose. These sugars are connected in four- or five-membered sequences, which often are branched. When the sequences repeat, the long O-specific polysaccharide is formed. Within the outer membrane these negatively charged polysaccharides can be linked together tightly when adjacent LPS molecules mutually form ionic bonds to divalent cations (such as Ca2+ and Mg2+). The presence of these ionic bonds confers considerable strength to the outer membrane, which rivals the gram-negative cell wall in its mechanical strength.
Figure 2.13 Structure of bacterial lipopolysaccharide.

The chemical structures of lipid A and polysaccharides can vary among gram-negative Bacteria, but the major components (lipid A–KDO–core–O-specific) are typically invariant. The O-specific polysaccharide is highly variable among species. KDO, ketodeoxyoctonate; Hep, heptose; Glu, glucose; Gal, galactose; GluNac, N-acetylglucosamine; GlcN, glucosamine; P, phosphate. Glucosamine and the lipid A fatty acids are linked through the amine groups of GlcN. The lipid A portion of LPS can be toxic to animals and comprises the endotoxin complex. Compare the art here with that of Figure 2.12 and follow the LPS components by their color-coding.
The lipid portion of the LPS, called lipid A, is not a typical glycerol lipid (see Figure 2.1c); instead the fatty acids are bonded through the amine groups from a disaccharide composed of glucosamine phosphate. The disaccharide is attached to the core polysaccharide through KDO (Figure 2.13). Fatty acids typically found in lipid A include caproic (C6), lauric (C12), myristic (C14), palmitic (C16), and stearic (C18) acids. LPS replaces much of the phospholipid in the outer half of the outer membrane, and although the outer membrane is technically a lipid bilayer, its many unique components distinguish it from the cytoplasmic membrane. The outer membrane is anchored to the peptidoglycan layer by the Braun lipoprotein, a molecule that spans the gap between the LPS layer and the peptidoglycan layer (in the periplasm, discussed in the next subsection) (Figure 2.12a).
An important biological activity of LPS is its toxicity to animals. Common gram-negative pathogens for humans include species of Salmonella, Shigella, and Escherichia, among many others, and some of the gastrointestinal symptoms these pathogens elicit are due to their toxic outer membrane components. Toxicity is specifically linked to the LPS layer, in particular, to lipid A. The term endotoxin refers to this toxic component of LPS. Some endotoxins cause violent symptoms in humans, including gas, diarrhea, and vomiting, and the endotoxins produced by Salmonella and enteropathogenic strains of Escherichia coli transmitted in contaminated foods are classic examples of this. We discuss major gram-negative enteric pathogens in Chapter 33 and endotoxin in Section 25.8.
The Periplasm and Porins
The outer membrane is impermeable to proteins and other very large molecules. In fact, a major function of the outer membrane is to prevent cellular proteins whose activities must occur outside the cytoplasm from diffusing away from the cell. These extracellular proteins reside in the periplasm, a space of about 15 nm located between the outer surface of the cytoplasmic membrane and the inner surface of the outer membrane (Figure 2.12a, b).
The periplasm may contain several different classes of proteins. These include hydrolytic enzymes, which function in the initial degradation of polymeric substances; binding proteins, which begin the process of transporting substrates (Section 2.2 and Figure 2.6); chemoreceptors, which are proteins that govern the chemotaxis response (Section 2.11); and proteins that construct extracellular structures (such as peptidoglycan and the outer membrane) from precursor molecules secreted through the cytoplasmic membrane. Most periplasmic proteins reach the periplasm by way of a protein-exporting system present in the cytoplasmic membrane (Sections 6.12 and 6.13).
The outer membrane is relatively permeable to small molecules because of proteins called porins that function as channels for the entrance and exit of solutes (Figure 2.12a, c). Porins are unique to the outer membrane of Bacteria and should not be confused with aquaporins, which are a different class of proteins (aquaporins facilitate water transport across the cytoplasmic membrane). Several porins are known, including both specific and nonspecific classes. Nonspecific porins form water-filled channels through which most very small hydrophilic substances can pass. By contrast, specific porins contain a binding site for one or a group of structurally related substances. Porins are transmembrane proteins composed of three identical polypeptides; the proteins are arranged to form channels through which solutes can diffuse (Figure 2.12c).
While the cell envelopes of many Bacteria conform to either the gram-positive or the gram-negative model, we have already learned that Archaea have cell envelopes that diverge from both of these model structures. In the next section we will see that there is considerable diversity in cell envelope structure across the microbial world.
Check Your Understanding
Describe and contrast the cell envelope structure of gram-negative and gram-positive bacteria.
What is the function of porins, and where are they located in the gram-negative cell envelope?
What component of the gram-negative cell envelope has endotoxin properties?
2.5 Diversity of Cell Envelope Structure
While many Bacteria have a gram-positive or gram-negative cell envelope organization (Figure 2.7), a variety of cell envelope structures are known. One way in which cell envelopes can vary between cells is in the presence of an S-layer.
S-Layers
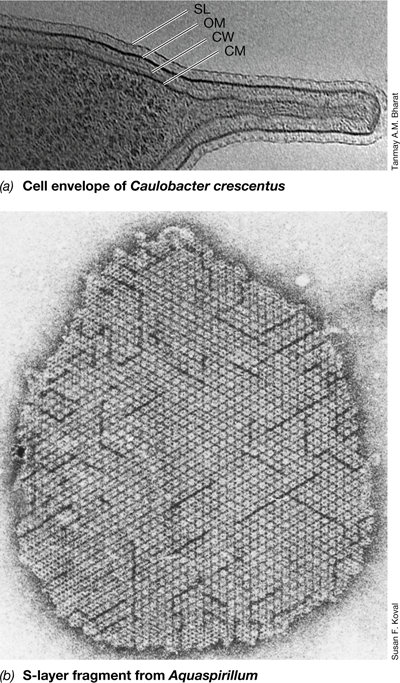
S-layers are found in many Bacteria and in nearly all Archaea. An S-layer consists of a paracrystalline monolayer of interlocking molecules of protein or glycoprotein (Figure 2.14). When an S-layer is present, it is always the outermost layer of the cell envelope (Figure 2.14a). The S-layer is usually composed of only one or a few subunits self-organized into repeating structures, which can have hexagonal, tetragonal, or trimeric symmetry. These repeating units form a rigid yet permeable paracrystalline lattice (Figure 2.14b), which can be as much as 5–20 nm thick in Bacteria and up to 70 nm thick in some Archaea.
Figure 2.14 S-layers.

(a) The S-layer (SL), outer membrane (OM), peptidoglycan cell wall (CW), and cytoplasmic membrane (CM) can be clearly seen in this electron tomographic slice through a cell of Caulobacter crescentus, a stalked bacterium that has a gram-negative cell envelope plus an outer S-layer. (b) Transmission electron micrograph of a portion of an S-layer removed from the bacterium Aquaspirillum and flattened to show the paracrystalline nature and hexagonal symmetry common to S-layers. The stalk in a has an outer diameter of about 150 nm and both images are at the same scale.
S-layers have many important functions. In many Archaea, thick S-layers can take on the role of the cell wall and are responsible for providing structural strength, protecting the cell from osmotic lysis, and conferring cell shape. The S-layer can also create a periplasmic-like space in Archaea. S-layers function as molecular sieves and have pore sizes in the range of 2–10 nm in diameter. These pores are large enough to allow low-molecular-weight compounds to pass but small enough to trap large molecules—such as most proteins—in the space between the S-layer and the cytoplasmic membrane. The compartment formed between the cytoplasmic membrane and the S-layer thus functions much as the periplasm does in gram-negative bacteria, including forming a site near the cytoplasmic membrane where reactions important to cellular metabolism can occur. As the outermost layer of the cell, S-layers can also facilitate cell surface interactions, such as attachment. S-layers can also increase the ability of some bacterial pathogens to cause disease by either promoting adhesion or protecting the cell from host defenses.
Alternative Configurations of the Cell Envelope
We have now learned about the most common components found in the cell envelope: the cytoplasmic membrane (CM), the cell wall (CW), the outer membrane (OM), and S-layers (SL). While certain configurations of these structures are common, a range of different configurations are possible (Figure 2.15). A common variation on cell envelope structure is to find an outer S-layer surrounding an otherwise gram-positive or gram-negative bacterium (compare Figure 2.15a with Figure 2.15b). In addition, many Archaea have only an S-layer outside of their cytoplasmic membrane, and these layers, while always constructed of some type of paracrystalline protein or glycoprotein, can vary considerably in their molecular structures (compare Figure 2.15b with Figure 2.15c). Some methanogenic Archaea also have cell walls made of pseudomurein, and such cell walls may or may not have an outer S-layer as well. Finally, though unusual, Archaea such as the heat-loving Ignicoccus actually have an outer membrane. This structure in Ignicoccus is unlike that of gram-negative bacteria in that it is composed largely of archaeal isoprenoid lipids and lacks LPS.
Figure 2.15 Alternative cell envelope structures.

Cell envelope structures including cytoplasmic membranes (CM), cell walls (CW), outer membranes (OM), and S-layers (SL) can be found in both bacterial and archaeal species. (a) Vibrio cholerae has a classic gram-negative type bacterial cell envelope. (b) Caulobacter crescentus is a bacterium with a gram-negative envelope and an S-layer (see also Figure 2.14a). (c) Nitrosopumilus maritimus has a typical archaeal cell envelope containing a CM and an SL. (d) Mycoplasma pneumoniae is a pathogenic bacterium whose cell envelope consists of only a CM. S-layers, while typically composed of a paracrystalline protein or glycoprotein layer (see also Figure 2.14b) can vary considerably in molecular structure (compare the S-layers in b and c). All images are transmission electron tomographs, a technique in which a special transmission electron microscope passes electrons through a specimen at different angles and then consolidates the views to form a final three-dimensional image.
Although this is uncommon, a few Bacteria and Archaea lack cell walls altogether. These include in particular the mycoplasmas (Figure 2.15d) and other pathogenic Bacteria that grow within a host cell, and Archaea such as Thermoplasma and its relatives. Lacking a cell wall, these cells would be expected to contain unusually tough cytoplasmic membranes, and chemical analyses show that they do. For example, most mycoplasmas contain sterols in their cytoplasmic membranes; these molecules function to add strength and rigidity to the membrane as they do in the cytoplasmic membranes of eukaryotic cells. Mycoplasmas may also have little need for a cell wall because they experience little osmotic pressure when living within the cytoplasm of another cell. In addition, the loss of peptidoglycan may help mycoplasmas evade the host immune system because host defenses recognize bacterial cell wall components as one of many signals of bacterial invasion (Chapter 26).
Check Your Understanding
What is the structure of an S-layer, and what are its functions?
What are some alternate arrangements of cell envelope structure?
II Cell Surface Structures and Inclusions
Many prokaryotic cells contain a cell surface layer that can have a variety of functions. Cytoplasmic inclusions may also be present and function as food reserves or bestow upon the cell a unique capacity of ecological value.
The cell envelope governs many aspects of how microbes interact with their environments and with other cells. Likewise, many other structures visible to microscopy can have profound impacts on cellular function. In particular, structures found on the cell surface and inclusions—structures present within the cell’s cytoplasm—have many important functions that govern microbial interactions with the world around them.
2.6 Cell Surface Structures
Many Bacteria and Archaea secrete sticky or slimy materials on their cell surface that consist of either polysaccharide or protein. However, these are distinct from and external to the cell envelope. The terms “capsule” and “slime layer” are used to describe these layers. These outer layers can mediate attachment, they can protect the cell from attack and from environmental stresses, and they can alter the diffusive environment of the cell.
Capsules and Slime Layers
The terms capsule and slime layer are used to describe a sticky coat of polysaccharide formed outside of the cell envelope. If the polysaccharide layer is organized in a tight matrix that excludes small particles and is tightly attached to the cell, it is called a capsule. Capsules are readily visible by light microscopy if cells are treated with India ink, which contains particulates that stain the background but cannot penetrate the capsule; capsules can also be seen in the electron microscope (Figure 2.16b–d). By contrast, if the surface layer is easily deformed and loosely attached, it will not exclude particles and is more difficult to see microscopically. Such a loosely attached polysaccharide coat is called a slime layer, and it is easily detected in colonies of slime-forming species such as the lactic acid bacterium Leuconostoc (**Figure 2.16*a***).
Figure 2.16 Bacterial capsules and slime formation.

(a) A semisolid colony of the bacterium Leuconostoc mesenteroides (lifted up by an inoculating loop) contains a thick dextran (glucose polymer) slime layer formed by the cells. (b) Capsules of Acinetobacter species observed by phase-contrast microscopy after negative staining with India ink. India ink does not penetrate the capsule and so the capsule appears as a light area surrounding the cell, which appears black. (c) Transmission electron micrograph of a thin section of cells of Rhodobacter capsulatus with capsules (arrows) clearly evident; cells are about 0.9 μm wide. (d) Transmission electron micrograph of Rhizobium leguminosarum biovar trifolii stained with ruthenium red to reveal the capsule. The cell is about 0.7 μm wide.
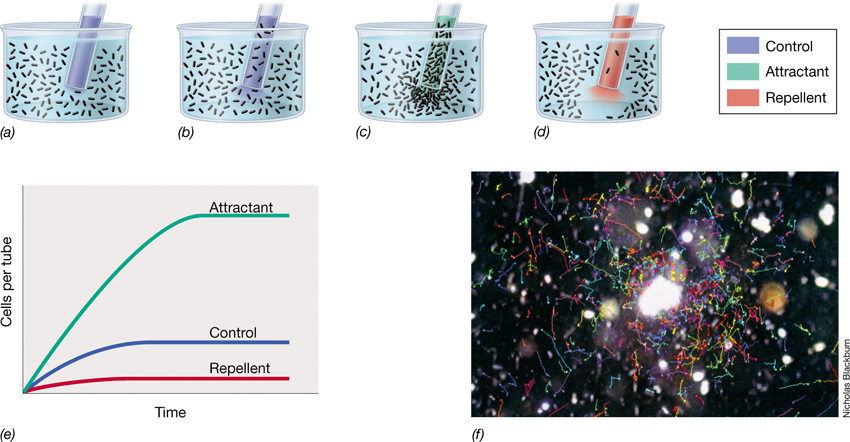
Outer surface layers have several functions. Surface polysaccharides assist in the attachment of microorganisms to solid surfaces. As we will see later, pathogenic microorganisms that enter the body by specific routes usually do so by first binding to specific surface components of host tissues; this binding is often facilitated by bacterial cell surface polysaccharides. When the opportunity arises, many bacteria will bind to solid surfaces, often forming a thick layer of cells called a biofilm (Section 4.9). Extracellular polysaccharides play a key role in the development and maintenance of biofilms as well.
Besides attachment, outer surface layers have other functions. These include contributing to the infectivity of a bacterial pathogen and preventing dehydration. For example, the causative agents of the diseases anthrax and bacterial pneumonia—Bacillus anthracis and Streptococcus pneumoniae, respectively—each contain a thick capsule of either protein (B. anthracis) or polysaccharide (S. pneumoniae). Encapsulated cells of these bacteria avoid destruction by the host’s immune system because the immune cells that would otherwise recognize these pathogens as foreign and destroy them are blocked from doing so by the bacterial capsule. In addition to this role in disease, bacterial outer surface layers bind water, and this helps protect the cell from desiccation in periods of dryness.
Fimbriae, Pili, and Hami
Pili are thin (2–10 nm in diameter) filamentous structures made of protein that extend from the surface of a cell and can have many functions. Short pili that mediate attachment are often called fimbriae (Figure 2.17). Pili enable bacterial cells to stick to surfaces, including animal tissues, or to form pellicles (thin sheets of cells on a liquid surface) or biofilms on solid surfaces. All gram-negative bacteria produce pili of one sort or another, and many gram-positive bacteria also contain these structures. Pili, by allowing bacteria to attach to other cells, often contribute to the virulence of pathogens.
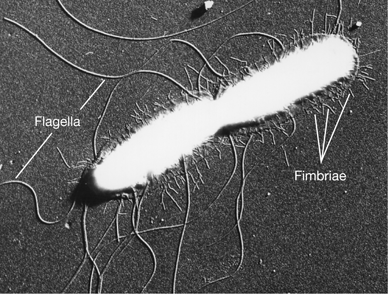
Figure 2.17 Fimbriae.

Electron micrograph of a dividing cell of Salmonella enterica (typhi), showing flagella and fimbriae. A single cell is about 0.9 μm wide.
Many classes of pili are known, and they can have diverse functions. As already mentioned, pili can enable bacteria to adhere to surfaces and this function can allow pathogens to target and invade specific host tissues. However, pili are diverse and they can have several other important functions as well. For example, conjugative pili facilitate genetic exchange by causing cell-to-cell attachment (Figure 2.18) during a process called conjugation (Section 9.8). In addition, electrically conductive pili (also known as nanowires, Section 23.3) can conduct electrons toward or away from the cell and in so doing play an important role in the energy metabolism of diverse microbes (Sections 14.13 and 15.13). Lastly, a type of pili called type IV pili not only facilitate adhesion but also support an unusual form of cell movement called twitching motility in certain bacterial species (see Section 2.10).
Figure 2.18 Pili.

The pilus on an Escherichia coli cell that is undergoing conjugation. The cells are about 0.8 μm wide. The visibility of the pilus in this electron micrograph has been improved because it is coated with viral particles that bind to the pilin protein.
Twitching motility allows cells to move along a solid surface. In twitching motility, pili are extended away from the cell, attach to a surface, and are subsequently retracted, dragging the cell forward. ATP supplies the energy necessary for extension and retraction of the pilus. On rod-shaped cells that move by twitching, type IV pili are present only at the cell poles. Type IV pili assist in infectivity by certain pathogens, including the gram-negative bacteria Vibrio cholerae (cholera) and Neisseria gonorrhoeae (gonorrhea) and the gram-positive bacterium Streptococcus pyogenes (strep throat and scarlet fever). The twitching motility of these organisms assists them in locating specific sites for attachment to initiate the disease process. Type IV pili are also widespread in the Archaea, functioning in surface adhesion and cell aggregation events that lead to biofilm formation.
An unusual group of Archaea, the SM1 group, forms a unique attachment structure called a hamus (plural, hami) that resembles a tiny grappling hook (**Figure 2.19a,*b***). The SM1 group inhabits anoxic groundwater in Earth’s deep subsurface, and hami function to affix cells to a surface to form a networked biofilm (Figure 2.19c). Hami structurally resemble type IV pili except for their barbed terminus, which functions to attach cells both to surfaces and to each other (Figure 2.19c). The biofilms formed by SM1 Archaea are likely an ecological strategy that allows these microbes to more efficiently trap the scarce nutrients present in their deep subsurface habitat. Although cells of the SM1 group are not as small as the groundwater ultramicrobacterial cells described in Chapter 1 (see Explore the Microbial World, “Tiny Cells”), they are less than 1 μm in diameter and live in a similar nutrient-limiting habitat. Thus, their hami likely play an important role in preventing cells from being washed away in groundwater flowage.
Figure 2.19 Unique attachment structures in the SM1 group of ***Archaea***: Hami.

(a) Cells of SM1 Archaea showing the pili-like surface structures called hami. (b) Transmission electron micrograph of isolated hami. A hamus “grappling hook” (labeled “Hook” in the micrograph) is about 60 nm in diameter. (c) A biofilm of SM1 cells showing the network of hami connecting individual cells.
In addition to the cell surface, the cytoplasm of a prokaryotic cell may contain enclosed and sometimes rigid structures that benefit the cell in one way or another, and we consider these now.
Check Your Understanding
Could a bacterial cell dispense with a cell wall if it had a capsule? Why or why not?
How do fimbriae differ from pili, both structurally and functionally?
2.7 Cell Inclusions
Prokaryotic cells often contain inclusions of one sort or another. Many inclusions store energy or nutrients (such as carbon or phosphorus), but some have other highly specialized functions that confer unique properties on the cells that contain them. Inclusions are often visible in cells with the light microscope and are enclosed by a single-layer (as opposed to a unit) membrane composed of proteins that partitions off the inclusion in the cytoplasm. Storing carbon or other substances in an insoluble form within the cytoplasm reduces osmotic stress and takes up less space compared with storing these substances in a soluble form.
Carbon Storage Polymers
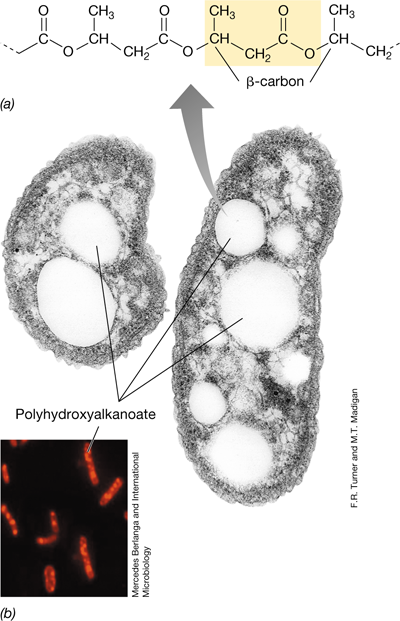
One of the most common inclusion bodies in prokaryotic organisms is poly-β-hydroxybutyric acid (PHB), a lipid that is formed from β-hydroxybutyric acid units. The monomers of PHB polymerize by ester linkage and then the polymer aggregates into granules; the granules can be seen by either light or electron microscopy (Figure 2.20). The monomer in the polymer is usually hydroxybutyrate (C4) but can vary in length from as short as C3 to as long as C18. Thus, the more generic term poly-β-hydroxyalkanoate (PHA) is often used to describe this class of carbon- and energy-storage polymers. PHAs are synthesized by cells when there is an excess of carbon and are broken down as carbon or energy sources when conditions warrant.
Figure 2.20 Poly-β-hydroxyalkanoates (PHAs).

(a) Chemistry of poly-β-hydroxybutyrate, a common PHA. A monomeric unit is highlighted in color. Other PHAs are made by substituting longer-chain hydrocarbons for the –CH3 group on the β-carbon. (b) Electron micrograph of a thin section of cells of a bacterium containing granules of PHB. Color photo: Nile red–stained cells of a PHA-containing bacterium.
Another carbon storage inclusion is glycogen, which is a polymer of glucose; like PHA, glycogen is a reservoir of both carbon and energy and is produced when carbon is in excess. Glycogen resembles starch, the major storage reserve of plants, but differs slightly from starch in the manner in which the glucose units are linked together.
Polyphosphate, Sulfur, and Carbonate Minerals
Many prokaryotic and eukaryotic microbes accumulate inorganic phosphate (PO43−) in the form of polyphosphate granules (**Figure 2.21*a***). These granules are formed when phosphate is in excess and can be drawn upon as a source of phosphate for nucleic acid and phospholipid biosynthesis when phosphate is limiting. In addition, in some organisms, polyphosphate can be broken down to synthesize the energy-rich compound ATP from ADP.
Figure 2.21 Polyphosphate and sulfur storage products.

(a) Phase-contrast photomicrograph of cells of Heliobacterium modesticaldum showing polyphosphate as dark granules; a cell is about 1 μm wide. (b) Bright-field photomicrograph of cells of the purple sulfur bacterium Isochromatium buderi. The periplasmic inclusions are sulfur globules formed from the oxidation of hydrogen sulfide (H2S). A cell is about 4 μm wide.
Many gram-negative Bacteria and several Archaea oxidize reduced sulfur compounds, such as hydrogen sulfide (H2S); these organisms are the “sulfur bacteria,” discovered by the great Russian microbiologist Sergei Winogradsky (Section 1.13). The oxidation of sulfide generates electrons for use in energy metabolism (chemolithotrophy) or CO2 fixation (autotrophy). In either case, elemental sulfur (S0) from the oxidation of sulfide may accumulate in the cell in microscopically visible granules (Figure 2.21b). This sulfur remains as long as the source of reduced sulfur from which it was derived is still present. However, as the reduced sulfur source becomes limiting, the S0 in the granules is oxidized to sulfate (SO42−), and the granules slowly disappear. Interestingly, although sulfur globules appear to reside in the cytoplasm (Figure 2.21b), they are actually present in the periplasm (Section 2.4). In these cells the periplasm expands outward to accommodate the growing globules as H2S is oxidized to S0 and then contracts inward as S0 is oxidized to SO42−.
Filamentous cyanobacteria have long been known to form carbonate minerals on the external surface of their cells. However, some cyanobacteria also form carbonate minerals inside the cell, as cell inclusions. For example, the unicellular cyanobacterium Gloeomargarita forms intracellular granules of benstonite, a carbonate mineral that contains barium, strontium, and magnesium (Figure 2.22). The microbiological process of forming minerals is called biomineralization. It is unclear why benstonite is formed by Gloeomargarita, although it might function as ballast to maintain cells of this cyanobacterium in their habitat, deep in an alkaline lake. Alternatively (or in addition), the mineral could be a way to sequester carbonate (a source of CO2) to support autotrophic growth.
Figure 2.22 Biomineralization by a cyanobacterium.

Electron micrograph of a cell of the cyanobacterium Gloeomargarita containing granules of the mineral benstonite [(Ba,Sr)6(Ca,Mn)6Mg(CO3)13]. A cell is about 2 μm wide.
Gas Vesicles
Some Bacteria and Archaea can float because they contain gas vesicles, structures that confer buoyancy and allow the cells to position themselves in regions of the water column that best suit their metabolisms. The most dramatic examples of gas-vesiculate microbes are those cyanobacteria that form massive accumulations called blooms in lakes or other bodies of water. These blooms are commonly on or near the lake surface (**Figure 2.23*a***) where sunlight is most intense and photosynthesis can occur at maximal rates.
Figure 2.23 Buoyant cyanobacteria and their gas vesicles.

(a) Flotation of a bloom of gas-vesiculate cyanobacteria in a freshwater lake. (b) Phase-contrast photomicrograph of Anabaena. Clusters of gas vesicles form phase-bright gas vacuoles (small arrows). (c) Transmission electron micrograph of a dividing cell of Microcystis. Gas vesicles are arranged in bundles, here seen in both longitudinal and cross section. A cell of Microcystis is about 5 μm wide.
Gas vesicles are conical-shaped structures composed of two different proteins; they are hollow yet rigid and of variable length and diameter (Figure 2.23b, c). Gas vesicles in different species vary from 300 to more than 1000 nm in length and from 45 to 120 nm in width. Gas vesicles may number from a few to hundreds per cell and are impermeable to water and solutes but permeable to gases. The composition and pressure of the gas inside a gas vesicle is that in which the organism is suspended. This could be air at 1 atm in cyanobacteria on a lake surface (Figure 2.23a), or a mixture of gases such as N2, CO2, and H2 at greater than 1 atm in gas vesiculate species that inhabit anoxic zones deeper in the lake. The presence of gas vesicles in cells can be detected either by light microscopy, where clusters of vesicles, called gas vacuoles, appear as irregular bright inclusions (Figure 2.23b), or by transmission electron microscopy of cell thin sections (Figure 2.23c).
Magnetosomes
Some bacteria can orient themselves within a magnetic field because they contain magnetosomes. These structures are biomineralized particles of the magnetic iron oxides magnetite [Fe(II)Fe(III)2O4] or greigite [Fe(II)Fe(III)2S4] (Figure 2.24). Magnetosomes impart a magnetic dipole on a cell, allowing it to orient itself in a magnetic field. This allows the cell to undergo magnetotaxis, the process of migrating along Earth’s magnetic field lines (see Section 2.12).
Figure 2.24 Magnetotactic bacteria and magnetosomes.

(a) Differential interference contrast micrograph of coccoid magnetotactic bacteria; note chains of magnetosomes (arrows). A cell is 2.2 μm wide. (b) Magnetosomes isolated from the magnetotactic bacterium Magnetospirillum magnetotacticum; each particle is about 50 nm wide. (c) Transmission electron micrograph of magnetosomes from an unnamed magnetic coccus. The arrow points to the membrane that surrounds each magnetosome. A single magnetosome is about 90 nm wide.
Magnetosome synthesis begins with insertion of magnetosome-specific proteins into the cytoplasmic membrane followed by invagination of the membrane to form a vesicle. The vesicle is then filled with iron—primarily iron in the Fe(II) oxidation state—and biomineralization proceeds through the activities of the magnetosome proteins, which includes an iron oxidase enzyme that catalyzes formation of the Fe(III) needed to form the magnetic minerals. The morphology of magnetosomes varies and appears to be species-specific; several morphologies are possible, but square, rectangular, or spike-shaped magnetosomes are most common.
We now consider a special case of a cytoplasmic enclosed cell structure—the endospore.
Check Your Understanding
Under what nutritional conditions would you expect PHAs or glycogen to be produced?
Why would it be impossible for gram-positive bacteria to store sulfur as gram-negative sulfur-oxidizing chemolithotrophs can?
How are magnetosomes and the Gloeomargarita inclusions similar and how do they differ? What are gas vesicles made of and what is inside them?
2.8 Endospores
Many microbes produce spore structures that allow them to survive unfavorable conditions. However, certain species of Bacteria produce specialized spores called endospores (Figure 2.25). Endospores (the prefix endo- means “within”) are highly differentiated dormant cells that function as survival structures and can tolerate harsh environmental conditions, including extreme heat, radiation, chemical exposure, drying, and nutrient depletion. Endospores are not reproductive structures, such as the spores of fungi, but are rather the dormant stage of a bacterial life cycle: vegetative cell → endospore → vegetative cell (Figure 2.26).
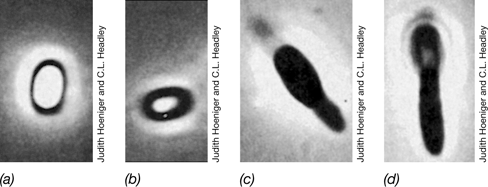
Figure 2.25 The bacterial endospore.
Phase-contrast photomicrographs showing different intracellular locations of endospores in different species of bacteria. Endospores appear bright by phase-contrast microscopy.
Figure 2.26 The life cycle of an endospore-forming bacterium.

The phase-contrast photomicrographs are of cells of Clostridium pascui. A vegetative cell is about 0.8 μm wide.
Many microbes can form spores or spore-like structures, including the myxospores of myxobacteria (Section 15.16), the arthrospores of actinomycetes (Section 16.12), and various forms of cysts, but endospores are exceptional for their extreme resistance to environmental challenges. Most notably, endospores are the only type of spore that can tolerate high heat, being able to survive for hours in boiling water. And, because of their unique structure, endospores can remain dormant for hundreds and perhaps even thousands of years, only to germinate and grow when conditions become favorable. Endospores are easily dispersed by wind, water, or through the animal gut, and hence endospore-forming bacteria are widely distributed in nature.
Endospores are only produced by two groups of bacteria, the Bacillales and Clostridiales, both of which are gram-positive bacteria of the phylum Firmicutes. These bacteria share an ancestor and so it is likely that the ability to form endospores evolved only once, though many species within these groups have lost the ability to form endospores over time. The best-studied endospore formers are in the genera Bacillus and Clostridium. These bacteria are found widely in soil and other environments, and some are well-known pathogens of humans and other animals. In particular, endospore-forming bacteria are a major cause of food spoilage and foodborne disease. Botulism, tetanus, and several foodborne bacterial infections are caused by species of endospore-forming bacteria (Chapters 32 and 33).
Endospore Formation and Germination
The process of cellular differentiation that results in endospore formation is called sporulation. During endospore formation, a vegetative cell is converted through a process of cellular differentiation into a nongrowing, heat-resistant, and light-refractive structure (Figure 2.26). Sporulation is typically triggered when some nutrient becomes limiting. When the cell senses this, the developmental process that converts a vegetative cell into an endospore begins and is controlled by a complex regulatory system we will consider later (Section 8.6). Our focus here is not on regulation but instead on the major steps in the process.
An endospore can remain dormant for years, but when conditions are favorable for growth, it can convert back to a vegetative cell rapidly through the process of germination (Figure 2.26). Germination is usually triggered by the availability of nutrients, such as certain amino acids or sugars. This process occurs in three steps: activation, germination, and outgrowth (Figure 2.27). The overall process occurs over a period of just a few minutes, and it is characterized by hydration of the spore, which results in loss of its heat and chemical resistance, the loss of specific spore structures and the regeneration of vegetative cell structures, and ultimately, the onset of vegetative growth by binary fission.
Figure 2.27 Endospore germination in ***Bacillus***.

Conversion of an endospore into a vegetative cell. This series of phase-contrast photomicrographs shows the sequence of events starting from (a) a highly refractile free endospore. (b) Activation: The spore becomes less refractile as the spore is hydrated. (c) Germination: the spore begins to develop into a vegetative cell. (d) Outgrowth: the vegetative cell emerges and begins to divide.
Endospore Structure and Features
Endospores differ in many ways from vegetative cells (Table 2.1). Endospores are visible by light microscopy as strongly refractile structures (Figures 2.25 and 2.26). Endospores are impermeable to most dyes, so occasionally they are seen as unstained regions within cells that have been stained with basic dyes such as methylene blue. To stain endospores, special stains and procedures must be used. In the classical endospore-staining protocol, the stain malachite green is used and is infused into the spore with steam.
Table 2.1 Differences between endospores and vegetative cells

The structure of the endospore as seen with the electron microscope differs distinctly from that of the vegetative cell (Figure 2.28). The endospore contains many layers absent from the vegetative cell. The innermost region of the endospore is called the core; this contains DNA and ribosomes and develops from the cytoplasm of the vegetative cell. Surrounding the core is the inner membrane, the cortex, and the outer membrane (Figure 2.28). The inner membrane develops from the cytoplasmic membrane of the vegetative cell, the cortex is composed of peptidoglycan, and the outer membrane is a special membrane formed during sporulation (and should not be confused with the LPS-containing outer membrane of gram-negative cells). Beyond the outer membrane is the endospore coat (Figure 2.28), composed of layers of spore-specific proteins, and some (but not all) endospores also have an outer proteinaceous layer called the exosporium (Figure 2.28).
Figure 2.28 Structure of the bacterial endospore.

(a) Transmission electron micrograph of a thin section through an endospore of Bacillus megaterium. Note that the composition of the endospore “outer membrane” is not the same as the outer membrane (LPS layer) of gram-negative bacteria shown in Figure 2.12. (b) Fluorescent photomicrograph of a cell of Bacillus subtilis undergoing sporulation. The green color is a fluorescent dye that specifically stains a sporulation protein in the endospore coat.
The secret to the endospore toughness, and the reasons that endospores are so highly refractile, lies in the dehydration of the core. The endospore core contains less than one-quarter of the water found in the vegetative cell (Table 2.1). Dehydration greatly increases heat and chemical resistance and causes enzymes in the core to become inactive (but not denatured). Dehydration of the core is facilitated by the accumulation of a substance called dipicolinic acid, a distinctive characteristic of endospores. Endospores also contain large amounts of calcium (Ca2+), most of which is complexed with dipicolinic acid. The calcium–dipicolinic acid complex forms about 10% of the dry weight of the endospore and functions to bind water, helping to dehydrate the developing endospore. In addition, the complex inserts between bases in DNA, which helps stabilize DNA against heat denaturation.
The endospore core also contains high levels of small acid-soluble spore proteins (SASPs). These proteins are only made during the sporulation process and have at least two functions. SASPs bind tightly to DNA in the core and protect it from potential damage from ultraviolet radiation, desiccation, and dry heat. Ultraviolet resistance is conferred when SASPs alter the physical structure of DNA, causing it to become more compact. This change in DNA structure causes it to be more resistant to mutations and other forms of potential damage caused by harsh chemicals or UV radiation (Section 12.4) and also increases its resistance to thermal denaturation. In addition, SASPs function as a carbon and energy source for the outgrowth of a new vegetative cell from the endospore during germination.
The Sporulation Cycle
Sporulation is a form of cellular differentiation (Figure 1.5), and many genetically directed events occur during the conversion from vegetative growth to sporulation. The structural changes in sporulating cells of Bacillus are shown in Figure 2.29. In Bacillus subtilis, which has been studied in detail, the conversion of a vegetative cell into an endospore takes about 8 hours and begins with asymmetric cell division and the formation of a forespore (Figure 2.29). Engulfment of the forespore by the mother cell results in the formation of the outer membrane that surrounds the developing endospore; the outer membrane forms from part of the mother cell’s cytoplasmic membrane. Key events in endospore formation such as asymmetric cell division, cortex formation, and SASP production take place in a defined sequence and at specific times in the sporulation cycle (Figure 2.29). Genetic studies of mutants of Bacillus subtilis, each blocked at one of the stages of sporulation, indicate that more than 200 sporulation-specific genes exist. These genes are turned on and off in a genetic program that governs cellular differentiation, a process we will consider in detail when we examine the genetic regulation of sporulation (Section 8.6).
Figure 2.29 Major events in endospore formation.

The steps depicted are those defined from genetic and microscopic analyses of sporulation in Bacillus subtilis, the model organism for studies of sporulation. SASPs, small acid-soluble proteins.
Prokaryotic cells are not static entities, and so we now turn our attention to how and why cell movements occur.
Check Your Understanding
What features differentiate an endospore from a vegetative cell?
Why might endospore-forming organisms be a problem for human health or the food industry?
How is the outer membrane of an endospore formed, and how does this structure differ from the outer membrane in the gram-negative cell envelope?
III Cell Locomotion
Prokaryotic cells move about their environments by powering motility devices that allow them to swim or crawl, and they can move in specific directions in response to chemical or physical signals.
We finish our survey of prokaryotic cell structure and function by examining cell locomotion. Many microbial cells can move under their own power. Motility allows cells to reach different parts of their environment, and in nature, a new location may offer a cell additional resources or protection from harmful substances and ultimately spell the difference between life and death.
We examine here the two major types of prokaryotic cell movement, swimming and gliding. We then consider how motile cells are able to move in a directed fashion toward or away from particular stimuli (a phenomenon called taxis) and present examples of these simple behavioral responses.
2.9 Flagella, Archaella, and Swimming Motility
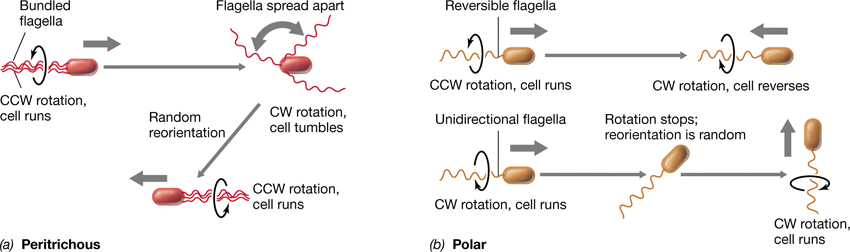
Many Bacteria are motile by swimming due to a structure called the flagellum (plural, flagella) (Figure 2.30); an analogous structure called the archaellum is present in many Archaea. Flagella and archaella are tiny rotating machines that function to push or pull the cell through a liquid.
Figure 2.30 Bacterial flagella.

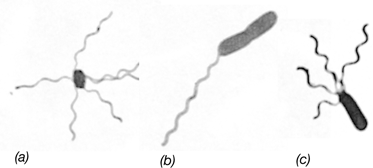
Classic light photomicrographs taken by Einar Leifson of bacteria containing different arrangements of flagella. Cells are stained with the Leifson flagella stain. (a) Peritrichous. (b) Polar. (c) Lophotrichous.
Flagella and Flagellation
Bacterial flagella are long, thin appendages (15–20 nm wide, depending on the species) free at one end and anchored into the cell at the other end. Flagella can be stained and observed by light microscopy (Figure 2.30) or electron microscopy (Figure 2.31).
Figure 2.31 Bacterial flagella as observed by negative staining in the transmission electron microscope.

(a) A single polar flagellum. (b) Peritrichous flagella. Both micrographs are of cells of the phototrophic bacterium Rhodocista centenaria, which are about 1.5 μm wide. Cells of R. centenaria are normally polarly flagellated but under certain growth conditions form peritrichous flagella. See Figure 2.41b for a photo of colonies of R. centenaria cells that move toward an increasing gradient of light (phototaxis).
Flagella can be anchored to a cell in different locations. In polar flagellation, the flagella are attached at one or both ends of a cell (Figure 2.30b). Occasionally, a group of many flagella (called a tuft) may arise at one end of the cell, a type of polar flagellation called lophotrichous (Figure 2.30c). Tufts of flagella can sometimes be seen in large unstained cells by dark-field or phase-contrast microscopy (Figure 2.32). When a tuft of flagella emerges from both poles of the cell, flagellation is called amphitrichous. In contrast to these more specific sites of flagellation, in peritrichous flagellation (Figures 2.30a and 2.31b), flagella are inserted around the cell surface.
Figure 2.32 Bacterial flagellar tufts observed in living cells.

(a) Dark-field photomicrograph of a group of large rod-shaped bacteria with flagellar tufts at each pole (a condition called amphitrichous flagellation). A single cell is about 2 μm wide. (b) Phase-contrast photomicrograph of cells of the large phototrophic purple bacterium Rhodospirillum photometricum that contain a tuft of lophotrichous flagella emerging from one of the poles. A cell measures about 4×25 μm.
Flagella do not rotate at a constant speed but increase or decrease their rotational speed in relation to the strength of the proton motive force. Flagella can rotate at up to 1000 revolutions per second to support a swimming speed of up to 60 cell-lengths/sec. The fastest known land animal, the cheetah, can move at about 25 body-lengths/sec. Thus, a bacterium swimming at 60 cell-lengths/sec is actually moving over twice as fast—relative to its size—as the fastest animal!
The swimming motions of polarly and lophotrichously flagellated organisms differ from those of peritrichously flagellated organisms, and these can be distinguished microscopically (Figure 2.33). Peritrichously flagellated organisms typically move slowly in a straight line, stop and then head off in a new direction. By contrast, polarly flagellated organisms often move more rapidly and continuously, and some are able to reverse their direction. The different behavior of flagella on polar and peritrichous organisms, including differences in reversibility of the flagellum, is illustrated in Figure 2.33.
Figure 2.33 Movement in peritrichously and polarly flagellated prokaryotic cells.

(a) Peritrichous: Forward motion is imparted by all flagella forming into a bundle and rotating counterclockwise (CCW). Clockwise (CW) rotation causes the bundle to break apart and the cell to tumble. A return to counterclockwise rotation leads the cell off in a new direction. (b) Polar: Cells change direction by reversing flagellar rotation (thus pulling instead of pushing the cell) or, with unidirectional flagella, by stopping periodically to reorient and then moving forward by clockwise rotation of its flagella. The gray arrow above each cell shows the direction the cell is traveling.
Flagella Structure and Activity
Bacterial flagella are rigid and helical (unlike eukaryal flagella, which are whiplike). The main part of the flagellum, called the filament, is composed of many copies of a protein called flagellin. The amino acid sequence of flagellin is highly conserved in Bacteria, suggesting that flagellar motility evolved early and has deep roots within this domain. In addition to the filament, a flagellum consists of several other components. A wider region at the base of the filament called the hook consists of a single type of protein and connects the filament to the flagellum motor in the basal body (Figure 2.34).
Figure 2.34 Structure and function of the flagellum in gram-negative ***Bacteria***.

The L ring is embedded in the LPS and the P ring in peptidoglycan. The MS ring is embedded in the cytoplasmic membrane and the C ring is found within the cytoplasm. The stator (Mot proteins) is embedded both within the cytoplasmic membrane and the peptidoglycan. Proton translocation through channels within the stator cause the MS ring to rotate, thereby driving rotation of the attached rod and flagellum. Flagellin molecules move from the export apparatus through a narrow channel in the rod and filament to reach the site of flagellar synthesis at the filament tip. The Mot proteins function as the flagellar motor. The flagellar motor rotates the filament to propel the cell through the medium. Inset photos: Top left, a cell of the purple sulfur bacterium Chromatium containing a tuft of polar flagella; Top right, transmission electron micrograph of a flagellar basal body from a cell of Salmonella enterica with the various rings labeled.
The flagellum motor is a reversible rotating machine composed of more than 25 proteins and anchored in the cytoplasmic membrane and cell wall. The motor consists of a central rod that passes through a series of rings. In gram-negative bacteria, an outer ring, called the L ring, is anchored in the outer membrane (Section 2.4). A second ring, called the P ring, is anchored in the peptidoglycan layer (Section 2.3). A third set of rings, called the MS and C rings, are located within the cytoplasmic membrane (Section 2.1) and the cytoplasm, respectively (Figure 2.34). In gram-positive bacteria, which lack an outer membrane, only the inner pair of rings is present. Surrounding the inner rings and anchored in the cytoplasmic membrane and the peptidoglycan is the stator, which is composed of Mot proteins. On the cytoplasmic side of the MS ring is the export apparatus, a type III secretion system (Section 6.13) that facilitates synthesis of the flagellum.
The flagellum motor contains two main components: the rotor and the stator. The rotor consists of the central rod and the L, P, C, and MS rings. The stator is comprised of Mot proteins, which surround the rotor and function to generate torque. Collectively, these structures make up the flagellar basal body (Figure 2.34). Rotation of the flagellum occurs at the expense of the proton motive force (Section 2.1), and it is thought that rotation is caused by a type of “proton turbine” process. In this model, proton translocation through channels within the stator complex cause the MS ring to rotate, thereby driving rotation of the attached rod and flagellum. The L and P rings act like bushings within which the rod rotates. Protons flowing through the Mot proteins exert electrostatic forces on helically arranged charged residues on rotor proteins and cause the MS ring to rotate. About 1200 protons are translocated by each rotation of the flagellum. The rotational speed of the flagellum is set by the proton flow rate through the Mot proteins, which is a function of the intensity of the proton motive force. The flagellar motors of different microbes are able to generate different amounts of torque, causing significant differences in swimming speed; such adaptations are driven by adding or subtracting subunits from the stator and C ring, which presumably changes the “gear ratio” of the motor.
Flagellar Synthesis
Several genes encode the motility apparatus of Bacteria. In Escherichia and Salmonella species, in which motility studies have been extensive, over 50 genes are linked to motility in one way or another. These genes encode the structural proteins of the flagellum and motor apparatus, of course, but also encode proteins that export the structural proteins through the cytoplasmic membrane to the outside of the cell and proteins that regulate the synthesis of new flagella.
A flagellar filament grows not from its base, as does an animal hair, but from its tip. The MS ring is synthesized first and inserted into the cytoplasmic membrane. Then other anchoring proteins are synthesized along with the hook and the cap before the filament forms (Figure 2.35). Flagellin molecules synthesized in the cytoplasm are exported through the export apparatus (Figure 2.34) present on the cytoplasmic side of the basal body. The export apparatus shuttles flagellin molecules into a 3-nm channel that runs through the center of the basal body and the hollow flagellar filament. The flagellin subunits are ultimately passed to the end of the filament where they attach to the terminus of the growing flagellum. Cap proteins assist flagellin molecules to assemble in the proper fashion at the flagellum tip (Figure 2.35). Approximately 20,000 flagellin protein molecules are needed to make one filament. The flagellum grows more or less continuously until it reaches its final length. Broken flagella still rotate and can be repaired with new flagellin units passed through the filament channel to replace the lost ones.
Figure 2.35 Flagella biosynthesis.

Synthesis begins with assembly of MS and C rings in the cytoplasmic membrane, followed by the other rings, the hook, and the cap. Flagellin protein flows through the hook to form the filament and is guided into position by cap proteins.
Archaella
As in Bacteria, swimming motility is widespread among species of Archaea and is driven by the rotation of their flagellum analog, the archaellum. However, despite their similar function, archaella proteins are unrelated to those of flagella, and in evolutionary terms are more closely related to type IV pili (Section 2.6). Archaella are somewhat smaller than flagella, measuring about 10–13 nm in width (Figure 2.36), which is only slightly wider than type IV pili. Archaella are not hollow and are assembled from their bases much like type IV pili, whereas flagella are hollow and are assembled at their tips (Figure 2.35). Archaellar rotation, like the extension and retraction of type IV pili, is driven by ATP hydrolysis, whereas the energy for flagellar rotation is derived from the proton motive force. In addition, the archaellar motor is structurally simpler than the flagellar motor (compare Figure 2.36 with Figure 2.34), and only 7–12 genes are required to encode the major proteins that make up the archaellar motor, in contrast to the more than 25 genes that encode components of the flagellar motor.
Figure 2.36 Archaella.

(a) Depiction of an archaellum embedded in an S-layer and cytoplasmic membrane; compare with the bacterial flagellum in Figure 2.34. The archaellum and its motor differ from flagella in that the archaellum has a smaller diameter, it is not hollow, its motor has fewer proteins, and the energy for rotation comes from ATP hydrolysis and not from consumption of the proton motive force. (b) Negatively stained transmission electron micrograph of purified archaella. (c) Scanning electron micrograph of cells of Methanocaldococcus containing multiple archaella.
Studies of swimming Archaea, such as Halobacterium, show that they swim at speeds only about one-tenth that of Escherichia coli, and in general, Archaea swim much more slowly than Bacteria. This could be due to the smaller diameter of the archaellum compared to the flagellum, which should reduce the torque the structure can generate. It could also be due to differences in the construction of the motor and how it is powered. However, some Archaea have found a way to overcome these limitations. For example, cells of Methanocaldococcus (Figure 2.36c) swim nearly 50 times faster than cells of Halobacterium and 10 times faster than cells of Escherichia coli. In fact, Methanocaldococcus swims at nearly 500 cell-lengths per second, which in a relative sense makes it one of the fastest organisms on Earth!
We now compare swimming motility with mechanistically quite different forms of cell movement but ones that still allow the cell to move about and explore its environment.
Check Your Understanding
Cells of Salmonella are peritrichously flagellated, those of Pseudomonas polarly flagellated, and those of Spirillum lophotrichously flagellated. Using a sketch, show how each organism would appear in a flagella stain.
Compare flagella and archaella in terms of their structure, function, and energy source.
How would you expect the flagellar basal body to differ between gram-negative and gram-positive bacteria?
2.10 Surface Motility
Many motile microbes are unable to swim and instead crawl over surfaces in various ways. These forms of surface motility require attachment to a surface and are independent of flagella or archaella. Surface motility results in distinctive colony morphology because cells can move out and away from the center of the colony (Figure 2.37). All forms of surface motility are considerably slower than swimming motility, usually less than 10 μm/sec. Finally, whereas nearly all swimming motility in Bacteria and Archaea is driven by flagella and archaella (Section 2.9), surface motility can be produced by a diversity of different systems, the most well characterized of which are twitching motility and gliding motility.
Figure 2.37 Gliding bacteria.
(a, b) The large filamentous cyanobacterium Oscillatoria has cells about 35 μm wide. (b) Oscillatoria filaments gliding on an agar surface. (c) Masses of the bacterium Flavobacterium johnsoniae gliding away from the center of the colony (the colony is about 2.7 mm wide). (d) Nongliding mutant strain of F. johnsoniae showing typical colony morphology of nongliding bacteria (the colonies are 0.7–1 mm in diameter). See also Figure 2.41.
Twitching Motility
Twitching motility requires type IV pili (Section 2.6), which extend from one pole of the cell, attach to a surface, and then retract to pull the cell forward (**Figure 2.38*a***). The energy required to catalyze the activity of type IV pili comes from ATP hydrolysis. Twitching motility has been described in many Bacteria and some Archaea. For example, species of the gram-negative bacterium Pseudomonas are often capable of twitching motility and this trait is important to their ability to form biofilms (Section 4.9). Twitching motility has also been well described in myxobacteria, which are social predatory bacteria whose cells can work together to consume other bacteria. Myxobacteria exhibit two forms of motility: social motility, which is caused by twitching, and adventurous motility, which is caused by gliding (see below). Twitching motility allows cells to move together in groups, and this trait is facilitated by the production of both type IV pili and the secretion of extracellular polysaccharides that aid in cellular cohesion.
Figure 2.38 Surface motility.

(a) Twitching motility employs type IV pili. These structures require ATP hydrolysis to extend (up to several micrometers) and then retract, causing the cell to move forward; movement occurs in discrete increments. (b) Gliding motility requires a helical intracellular protein track that interacts with gliding motors and extracellular adhesion proteins. The proton motive force drives rotation of gliding motors that translate this force to the helical track, causing adhesion proteins to move in a helical pattern; this results in continuous forward motion and clockwise rotation of the cell.
Gliding Motility
Gliding motility (Figure 2.38b) is defined as a smooth motion along the long axis of a cell without the aid of external propulsive structures (such as pili or attachment organelles). Gliding motility is a continuous form of movement, very unlike swimming motility, in which cells frequently stop and then swim off in a random new direction. Gliding bacteria are typically filamentous or rod-shaped in morphology, and while gliding has been observed among diverse species of Bacteria, no gliding Archaea are known. The adventurous motility of myxobacteria is a form of gliding that has been well studied, and many species of the Bacteroidetes such as Flavobacterium also exhibit gliding motility.
Gliding has been best studied in Myxococcus and Flavobacterium (Figure 2.37c), and the mechanism of gliding in these two organisms is likely to be quite similar. These organisms both possess a helical intracellular track made of proteins that run in a continuous loop around the cell (Figure 2.38b). Associating with this track are “gliding motors,” rotary motors that are driven by the proton motive force, and which are evolutionarily and operationally similar to the flagellar motor (see Figure 2.34). These organisms also have adhesion proteins that grab onto surfaces on the outside of the cell. While the exact mechanism of gliding remains unknown, it is thought that the gliding motor associates with the helical track, and rotation of the motor causes displacement of the track relative to the motor (or of the motor relative to the track). This movement is in some way transduced to the surface adhesion proteins, causing them to move in a helical direction around the surface of the cell. The net result is that the cell is propelled forward over the surface and caused to rotate around its axis as it moves forward (Figure 2.38b).
We now consider how these various forms of prokaryotic cell motility can move cells in specific directions, which allows them to both exploit useful nutrients and avoid harmful substances.
Check Your Understanding
How does gliding motility differ from swimming motility in both mechanism and requirements?
Contrast the mechanism of twitching motility with that of gliding motility.
2.11 Chemotaxis
Cells of Bacteria and Archaea have evolved numerous systems that allow them to sense and respond to chemical and physical stimuli in their environments. For example, microbes are able to sense and move toward a food source. Such a directed movement is called taxis (plural, taxes). Chemotaxis, a response to chemicals, and phototaxis, a response to light, are two well-studied forms of taxis. The ability of a cell to move toward or away from various stimuli has ecological significance in that the directed movement may enhance a cell’s access to resources or allow it to avoid harmful substances that could damage or kill it.
Chemotaxis has been well studied in Bacteria that use swimming motility; these organisms sense environmental stimuli and transmit signals to the flagellum causing it to alter its rotation. In addition, most Archaea that exhibit swimming motility are also chemotactic, and several of the proteins that control chemotaxis in Bacteria are similar to those found in Archaea. Gliding bacteria are also often chemotactic and can alter their direction in response to environmental stimuli. Phototaxis is often observed in filamentous cyanobacteria (Figure 2.37a, b) which move toward or away from a light source. In Section 7.6 we examine in detail the molecular mechanism of chemotaxis and its genetic regulation in the model bacterium Escherichia coli. For now, we will focus on the general principles by which chemotaxis operates.
Chemotaxis in Peritrichously Flagellated *Bacteria*
Much research on chemotaxis has been done with the peritrichously flagellated bacterium E. coli. In the absence of an attractant, cells move in a random fashion that includes runs, in which the cell is swimming forward in a smooth fashion, and tumbles, when the cell stops and jiggles about randomly (Figure 2.39). During a run, the flagellar motor rotates counterclockwise, causing the flagella to coil together into a bundle that propels the cell forward. By contrast, when flagella rotate clockwise, the bundle of flagella pushes apart, forward motion ceases, and the cells tumble randomly (**Figure 2.39*a***). Following a tumble, the cell will begin a new run in a random direction. Thus, in the absence of an attractant, the cell moves about its environment in random fashion through a series of runs and tumbles.
Figure 2.39 Chemotaxis in a peritrichously flagellated bacterium.

(a) In the absence of a chemical attractant, the cell swims randomly in runs, changing direction during tumbles. (b) In the presence of an attractant, runs become biased, and the cell moves up the gradient of the attractant. The attractant gradient is depicted in green, with the highest concentration where the color is most intense.
To understand chemotaxis, consider what happens when E. coli encounters a chemical attractant, such as the sugar glucose (**Figure 2.39*b***). Because of diffusion, the concentration of the attractant will increase when the cell is moving toward the attractant. During chemotaxis cells will tend to move toward the attractant over time, but cells do not move continuously nor directly toward the attractant; instead, they exhibit a behavior known as a biased random walk. The key to this mechanism is that cells sample the concentration of attractant not over space, but rather over time. As the cell is swimming in a random direction, it continues to sense the concentration of attractant over time. If the concentration goes up, runs become longer and tumbles less frequent, but if the concentration goes down, runs become shorter and tumbles more frequent. The biased random walk causes net movement toward the attractant (**Figure 2.39*b***), and this mechanism is remarkably effective for bringing cells to the source of an attractant. For a repellent, the same mechanism applies, but the response is reversed, with longer runs triggered by a decrease in concentration of the repellent. Attractants and repellents are sensed by a series of membrane proteins called chemoreceptors. Chemoreceptors sense the concentration of particular chemicals and transduce this information to flagella, causing them to alter their rotation (Section 7.6).
Chemotaxis in Polarly Flagellated *Bacteria*
Chemotaxis in polarly flagellated cells is similar but not identical to chemotaxis in peritrichously flagellated cells such as E. coli. Many polarly flagellated bacteria, such as Pseudomonas species, which have a single polar flagellum rather than a tuft, do not tumble when they reverse their flagellum. Instead, they swim backwards when their flagellum reverses its direction of rotation (**Figure 2.33*b***). In contrast, the phototrophic bacterium Rhodobacter has a single polar flagellum that is not reversible; it can only start and stop its rotation. Both of these polarly flagellated bacteria rely on Brownian motion when stopped to randomly reorient the cell (**Figure 2.33*b***). Then as the flagellum begins to rotate again, the cell moves off in a new direction. Despite this seemingly random activity, cells of Rhodobacter are strongly chemotactic to various organic compounds and to oxygen and light. Chemotactic organisms, by employing a biased random walk, can navigate effectively through their environments toward conditions that favor growth and away from those that could inhibit growth or otherwise cause harm.
Measuring Chemotaxis
Bacterial chemotaxis can be measured in a chemotaxis assay in which a small glass capillary tube containing an attractant is immersed into a suspension of motile bacteria. A chemical gradient extends from the tip of the capillary into the surrounding medium, with the chemical concentration decreasing with distance from the tip (Figure 2.40). When an attractant is present, chemotactic bacteria will move toward it, forming a swarm around the open tip (**Figure 2.40*c***) with many of the bacteria swimming into the capillary tube. Of course, because of random movements some chemotactic bacteria will swim into the capillary even if it does not contain an attractant (control solution, Figure 2.40b). However, when an attractant is present, the number of cells within the capillary will increase significantly (Figure 2.40e). If the capillary contains a repellent, the number of bacteria within the capillary will be fewer than in the control (**Figure 2.40*d***). This capillary method can be used to screen chemicals to see if they are attractants or repellents for a given bacterium.
Figure 2.40 Measuring chemotaxis using a capillary tube assay.

(a) Insertion of the capillary into a bacterial suspension. As the capillary is inserted, a gradient of the chemical begins to form. (b) A control capillary contains a salt solution that is neither an attractant nor a repellent. Cell concentration inside the capillary becomes the same as that outside. (c) Accumulation of bacteria in a capillary containing an attractant. (d) Repulsion of bacteria by a repellent. (e) Time course showing cell numbers in capillaries containing various chemicals. (f) Tracks of motile bacteria in seawater swarming around an algal cell (large white spot, center) photographed with a tracking video camera system attached to a microscope. The bacterial cells are showing positive aerotaxis by moving toward the oxygen-producing algal cell. The alga is about 60 μm in diameter.
Chemotaxis can also be observed microscopically. It is possible to quantify chemotactic behavior using a video camera that captures the positions of bacterial cells with time and records the motility tracks of each cell (Figure 2.40f). This method has been used to study chemotaxis of microbes in natural environments. In nature, nutrients are often excreted from larger microbial cells whether they are living or dead. Algae, for example, produce both organic compounds and oxygen (O2, from photosynthesis), which can trigger chemotactic movements of bacteria toward the algal cell (**Figure 2.40*f***). Chemotaxis can result in complex cellular interactions in nature including competition for resources and cooperation by recycling metabolic products.
Check Your Understanding
Describe how bacteria use a biased random walk to move toward an attractant.
Contrast the motility of a peritrichously flagellated bacterium such as Escherichia coli with that of a polarly flagellated bacterium such as a species of Pseudomonas.
In a capillary tube assay, why is it necessary to use a control that lacks attractant?
2.12 Other Forms of Taxis
Chemotaxis is a directed movement with respect to a chemical gradient, but many other forms of sensory response can govern microbial motility. For example, osmotaxis is directed movement with respect to a gradient of ionic strength, hydrotaxis is directed movement with respect to a gradient of available water, aerotaxis is directed movement with respect to gradients of O2, and phototaxis is directed movement with respect to a gradient in light intensity. Here we will consider phototaxis and aerotaxis, two taxes that have been particularly well studied.
Phototaxis
Many phototrophic microorganisms can move toward light, a process called phototaxis. Phototaxis allows a phototrophic organism to position itself most efficiently to receive light for photosynthesis. For example, if motile phototrophic purple bacteria are placed on a microscope slide that is illuminated with a spectrum of light they will move preferentially toward certain wavelengths (**Figure 2.41*a***). The wavelengths of light at which the bacteria accumulate correspond to the wavelengths of light that are absorbed by their photosynthetic pigments. These pigments include, in particular, bacteriochlorophylls and carotenoids (Chapter 14).
Figure 2.41 Phototaxis of phototrophic bacteria.

(a) Scotophobic accumulation of the phototrophic purple bacterium Thiospirillum jenense at wavelengths of light at which its pigments absorb. A light spectrum was displayed on a microscope slide containing a dense suspension of the bacteria; after a period of time, the bacteria had accumulated selectively and the photomicrograph was taken. The wavelengths at which the bacteria accumulated are those at which the photosynthetic pigment bacteriochlorophyll a absorbs. (b) Phototaxis of an entire colony of the purple phototrophic bacterium Rhodocista centenaria. These strongly phototactic cells move in unison toward the light source at the top. See Figure 2.31 for electron micrographs of flagellated R. centenaria cells.
Two different light-mediated taxes are observed in phototrophic bacteria. One, called scotophobotaxis, is easily observed in the microscope. Scotophobotaxis occurs when a phototrophic bacterium happens to swim into darkness outside the illuminated field of view of the microscope. Entering darkness negatively affects photosynthesis, and thus cellular energy levels, and this signals the cell to tumble with greater frequency until it reenters the light by a biased random walk. Scotophobotaxis is a mechanism to prevent phototrophic cells from swimming away from a lighted zone into darkness, and this likely improves their competitive success in nature.
Phototaxis differs from scotophobotaxis in that cells move with respect to a gradient of light intensity, whereas scotophobotaxis is a response only to the absence of light. Phototaxis is analogous to chemotaxis except that the attractant is light instead of a chemical. In some phototactic organisms, such as the highly motile phototrophic purple bacterium Rhodocista centenaria (Figure 2.31), entire colonies of cells show phototaxis and move in unison toward the light (Figure 2.41b).
Several components of the regulatory system that control chemotaxis also control phototaxis. A photoreceptor, a protein that functions similarly to a chemoreceptor but which senses a gradient of light instead of chemicals, is the initial sensor in the phototaxis response. The photoreceptor then interacts with the same cytoplasmic proteins that control flagellar rotation in chemotaxis, maintaining the cell in a run if it is swimming toward an increasing intensity of light. Section 7.6 describes the activities of these proteins in more detail.
Aerotaxis and Magnetotaxis
Aerotaxis is directed motility with respect to O2 (see Figure 2.40f). Aerobic organisms require O2 and may exhibit a positive aerotactic response, swimming toward increasing concentrations of O2. We will see in the next chapter, however, that O2 is toxic to many microbes. Microaerophiles are aerobic organisms that require O2 but are killed when the concentration of O2 is too high. Microaerophiles often use aerotaxis to position themselves at an optimum concentration of O2 (usually between 1% and 5% O2). Microaerophily can be a challenging lifestyle because either too much or too little O2 can mean death, and O2 gradients are highly dynamic because many organisms either produce or consume O2. Hence, some microaerophiles have evolved aerotactic mechanisms that help them better navigate dynamic gradients of O2. Indeed, a movement of only a few tens of micrometers may be necessary to place aerotactic cells in a more suitable gaseous environment.
Magnetosomes (Section 2.7) are found in specialized microaerophiles, referred to as magnetotactic bacteria, which are found in sediments, lakes, and ponds where concentrations of O2 are naturally low. Because Earth is a sphere, its magnetic field lines have a significant vertical component where they intersect the surface. Magnetosomes allow magnetotactic bacteria to align themselves with these magnetic field lines. This causes the bacterial cells to point up or down so that they can swim either toward or away from O2 at the surface. These so-called magnetotactic bacteria do not actually exhibit directed motility toward magnetic fields but instead are exhibiting aerotaxis; their magnetosomes allow them to reduce a three-dimensional biased random walk to a two-dimensional biased random walk and this vastly improves their ability to position themselves in their environment with respect to O2.
We now segue from our consideration of the structure and function of major components of prokaryotic cells to examine structure–function issues in eukaryotic cells, many of which are microbial and coexist with prokaryotic cells in the microbial world.
Check Your Understanding
Explain how magnetosomes contribute to the motility of magnetotactic bacteria.
IV Eukaryotic Microbial Cells
The cytoplasm of eukaryotic cells contains membrane-bound organelles including the nucleus, mitochondria, and chloroplasts, and several other distinct structures. During cell division, the nucleus divides by mitosis and reproductive structures are formed by the process of meiosis.
Compared with prokaryotic cells, microbial eukaryotes typically have structurally more complex and much larger cells. Microbial eukaryotes include the fungi and a vast diversity of protists as we will see in Chapter 18. Many aspects of eukaryotic cell structure set Eukarya apart from Archaea and Bacteria, but the defining feature of the eukaryotic cell is its nucleus.
2.13 The Nucleus and Cell Division
A double membrane–enclosed nucleus is a universal feature of the eukaryotic cell. Mitochondria are also nearly universal among eukaryotic cells, although a few unusual protists lack mitochondria. Chloroplasts are found only in phototrophic eukaryotic cells, including plants and diverse algae. Other characteristic eukaryotic structures include the Golgi complex, lysosomes, endoplasmic reticula, and microtubules and microfilaments (Figure 2.42). Some microbial eukaryotes have flagella or cilia—structures that confer motility—and a cell wall is present in many eukaryotic cells, such as the fungi and algae.
Figure 2.42 Cutaway schematic of a cell of a microbial eukaryote.

Although all eukaryotic cells contain a nucleus, not all organelles and other structures shown are present in all microbial eukaryotes. Not shown is the cell wall, found in fungi, algae, plants, and a few protists.
Eukaryotic cell membranes contain sterols. These rather rigid organic molecules, which are a subclass of steroids and absent from all but a few prokaryotic cells, lend structural strength to the eukaryotic cell, something especially important to those eukaryotes that lack a cell wall, such as many protists and animal cells.
The Nucleus
The nucleus contains the chromosomes of the eukaryotic cell. DNA within the nucleus is wound around basic (positively charged) proteins called histones, which tightly pack the negatively charged DNA to form nucleosomes (Figure 2.43); the latter are then organized into chromosomes. Archaea also contain histones and nucleosomes, and these are evolutionarily related to those found in Eukarya. Moreover, a few Bacteria also contain histone-like proteins that function to help organize their DNA.
Figure 2.43 The nucleus and DNA packaging in eukaryotes.

(a) Electron micrograph of a yeast cell prepared in such a way as to reveal a surface view of the nucleus. The cell is about 8 μm wide. (b) Packaging of DNA around histone proteins to form a nucleosome. Nucleosomes are arranged along the DNA strand like beads on a string and aggregate to form chromosomes during the process of mitosis (see Figure 2.44).
Only cells of Eukarya contain a nucleus. The nucleus is enclosed by a pair of membranes, each with its own function, separated by a space. The innermost membrane is a simple sac while the outermost membrane is in many places continuous with the endoplasmic reticulum. The inner and outer nuclear membranes specialize in interactions with the nucleoplasm and the cytoplasm, respectively. The nuclear membranes contain pores (Figures 2.42 and 2.43a), formed from holes where the inner and outer membranes are joined. The pores allow transport proteins to import and export other proteins and nucleic acids into and out of the nucleus, a process called nuclear transport.
Within the nucleus is found the nucleolus (Figure 2.42), the site of ribosomal RNA (rRNA) synthesis. The nucleolus is rich in RNA, and ribosomal proteins synthesized in the cytoplasm are transported into the nucleolus and combine with rRNA to form the small and large subunits of eukaryotic ribosomes. These are then exported to the cytoplasm, where they associate to form the intact ribosome and function in protein synthesis.
Cell Division
Eukaryotic cell division requires a special process, called mitosis, in which the chromosomes are replicated, the nucleus is disassembled, the chromosomes are segregated into two sets, and a nucleus is reassembled in each daughter cell (Figure 2.44). Whereas many (though not all) prokaryotic cells are genetically haploid, microbial eukaryotes often alternate between haploid and diploid states. Diploid cells have two copies of each chromosome whereas haploid cells have only one. For example, the brewer’s yeast Saccharomyces cerevisiae can exist in the haploid state (in which cells contain 16 chromosomes) as well as in the diploid state (in which cells contain 32 chromosomes). However, regardless of its genetic state, during cell division the chromosome number is first doubled and later halved to give each daughter cell its correct complement of chromosomes. During mitosis, the chromosomes condense, divide, and are separated into two sets, one for each daughter cell. Each of these distinct stages of nuclear division has a common name used widely in biology: prophase, metaphase, anaphase, and telophase (Figure 2.44a–d, respectively).
Figure 2.44 Light micrograph of eukaryotic cells undergoing mitosis.

(a) Interphase, distinct chromosomes are not apparent. (b) Metaphase. Homologous chromosomes are lining up along the cell center; compare with Figure 2.47b. (c) Anaphase. Homologous chromosomes are pulling apart. (d) Telophase. Chromosomes have separated into the newly forming daughter cells.
In contrast to mitosis, meiosis converts a diploid cell into several haploid cells. Meiosis consists of two successive cell divisions. In the first meiotic division, pairs of chromosomes segregate into separate cells, changing the genetic state from diploid to haploid. The second meiotic division is essentially the same as mitosis, as the two haploid cells divide to form a total of four haploid cells called gametes. This form of cell division is typically used in organisms that reproduce sexually. In animals, these gametes are the eggs and sperm; in eukaryotic microorganisms, they can be reproductive spores or other reproductive structures.
We now consider the classic organelles of the eukaryotic cell: the mitochondrion and the chloroplast.
Check Your Understanding
How is DNA arranged in the chromosomes of eukaryotes?
What are the major differences between mitosis and meiosis?
2.14 Mitochondria and Chloroplasts
Organelles that specialize in energy metabolism in eukaryotes include the mitochondrion and the chloroplast. These organelles have evolutionary roots within the Bacteria and provide ATP to the eukaryotic cell either from the oxidation of organic compounds or from light.
Mitochondria
In aerobic eukaryotic cells, respiration occurs in the mitochondrion. Mitochondria are the size of a typical bacterium, they have their own DNA and ribosomes, they can take on many shapes (Figure 2.45), and they reproduce independently of the cell. The number of mitochondria per cell varies depending on the cell type, its size, and its physiological state. A yeast cell may have only a few mitochondria per cell, whereas an animal cell may have over a thousand. The mitochondrion is enclosed by a double membrane system. Like the nuclear membrane, the outermost mitochondrial membrane is somewhat permeable and contains pores that allow the passage of small molecules. The innermost membrane is much less permeable, and its structure more closely resembles that of the cytoplasmic membrane of Bacteria.
Figure 2.45 Structure of the mitochondrion.

(a) Diagram showing the overall structure of the mitochondrion; note the inner and outer membranes. (b, c) Transmission electron micrographs of mitochondria from rat tissue showing the variability in morphology; note the cristae. Both mitochondria are about 0.7 μm wide.
Mitochondria also contain folded internal membranes called cristae. These membranes, formed by invagination of the inner membrane, contain the enzymes needed for respiration and ATP production. Cristae also contain transport proteins that regulate the passage of key molecules such as ATP into and out of the matrix, the innermost compartment of the mitochondrion (Figure 2.45a). The matrix contains enzymes for the oxidation of organic compounds, in particular, enzymes of the citric acid cycle, the major pathway for the combustion of organic compounds to CO2 (Section 3.6).
Chloroplasts
Chloroplasts are the chlorophyll-containing organelles found in plants and algae and are the site of photosynthesis. Chloroplasts are about the size of unicellular cyanobacteria and are readily visible with the light microscope (Figure 2.46). Like mitochondria, the number of chloroplasts per cell varies among species, and chloroplasts contain their own DNA and ribosomes.
Figure 2.46 Chloroplasts of a diatom and a green alga cell.
(a) Fluorescence photomicrograph of a diatom shows chlorophyll fluorescence; arrows, chloroplasts. The cell is about 40 μm wide. (b) Phase-contrast photomicrograph of the filamentous green alga Spirogyra showing the characteristic spiral-shaped chloroplasts (arrows) of this phototroph. A cell is about 20 μm wide. (c) Transmission electron micrograph showing a chloroplast of a diatom; note the thylakoids.
Also like mitochondria, chloroplasts are enclosed by a double membrane composed of a permeable outer membrane and a less-permeable inner membrane. The innermost membrane surrounds the stroma, analogous to the matrix of the mitochondrion (**Figure 2.46*c***). The stroma contains large levels of the enzyme ribulose bisphosphate carboxylase (RuBisCO), the key enzyme of the Calvin cycle, which is the series of biosynthetic reactions by which phototrophs convert CO2 to organic compounds (Section 3.12). The permeability of the outermost chloroplast membrane allows glucose and ATP produced during photosynthesis to diffuse into the cell cytoplasm where they are consumed in biosynthesis.
Chlorophyll and all other components needed for ATP synthesis in chloroplasts are located in a series of flattened membrane discs called thylakoids (Figure 2.46c). Like the cytoplasmic membrane, the thylakoid membrane is highly impermeable, and its major function is to form a proton motive force (Figure 2.4c) that results in ATP synthesis.
The Endosymbiotic Origin of Organelles
On the basis of their relative autonomy, size, and morphological resemblance to bacteria, it was hypothesized over 100 years ago that mitochondria and chloroplasts were descendants of respiratory and phototrophic bacterial cells, respectively. According to this hypothesis, when these symbiotic bacteria associated with nonphototrophic hosts, the hosts gained new forms of energy metabolism while the bacterial partners received a stable and supportive growth environment inside the host cell. Then, over time, these originally free-living symbionts became an intimate part of the eukaryotic cell. The idea that symbiotic bacteria are ancestors to the mitochondrion and chloroplast is called the endosymbiotic theory and is well accepted in biology today (Sections 13.4 and 18.1). Several lines of evidence support the endosymbiotic theory. These include, most notably, the fact that mitochondria and chloroplasts contain their own genomes and ribosomes whose structures are similar to the genomes and ribosomes of Bacteria but not to those of Eukarya. These organellar genomes are also circular, typical of bacterial chromosomes (Section 10.4).
Eukarya are a distinct domain of life, a domain that evolved after Bacteria and Archaea. Nevertheless, eukaryotic cells share many genes with both Bacteria and Archaea. In this respect, Eukarya are genetic chimeras that contain genes from two domains of life. The domain Eukarya is hypothesized to have originated from a symbiotic fusion between an archaeal host cell and a mitochondrial endosymbiont (derived from Bacteria). Sometime later, a second major symbiotic event occurred, in which a eukaryotic host cell acquired a chloroplast endosymbiont (derived from cyanobacteria), becoming the ancestor of all phototrophic eukaryotes (including plants and algae).We consider these important concepts in detail in Chapter 13.
Check Your Understanding
What key reactions occur in the mitochondrion and in the chloroplast, and what key products are made in each?
What is the endosymbiotic theory, and what evidence is there to support it?
2.15 Other Eukaryotic Cell Structures
Several other complex structures are present within the cytoplasm of microbial eukaryotes in addition to mitochondria and chloroplasts (Figure 2.42). These include the endoplasmic reticulum, the Golgi complex, lysosomes, and a dynamic cytoskeleton. Unlike the nucleus, mitochondria, and chloroplasts, these structures lack both DNA and a double membrane and are not of endosymbiotic origin. In addition, these structures control the transport of proteins and nutrients within the cell and govern cell shape and movement. Cell walls are also present in certain microbial eukaryotes (such as fungi and algae), though there is great diversity in cell wall structure among eukaryotic cells.
Cytoskeleton
The eukaryotic cytoplasm is crisscrossed by a series of dynamic protein filaments, which are used to transport substances, to position cell structures, and to control cell movement. This internal support network consists of microtubules, microfilaments, and intermediate filaments; together, these structures form the cell cytoskeleton (Figure 2.42). Some cytoskeleton-like elements are also important for determining the shape and molecular organization of prokaryotic cells, but the eukaryotic cytoskeleton is notable for its complexity and dynamism.
Microtubules are hollow tubes about 25 nm in diameter and are composed of the proteins α-tubulin and β-tubulin. Microtubules (**Figure 2.47*a***) have many functions including maintaining cell shape and facilitating cell motility, moving chromosomes during mitosis (Figures 2.44 and 2.47b), and in the movement of organelles within the cell. Microfilaments (Figure 2.47c) are smaller than microtubules, about 7 nm in diameter, and are polymers of two intertwined strands of the protein actin. Microfilaments function in maintaining or changing cell shape, in cell motility by cells that move by amoeboid movement, and during cell division. Intermediate filaments are fibrous keratin proteins that are arranged into fibers 8–12 nm in diameter and function in maintaining cell shape and positioning organelles in the cell.
Figure 2.47 Tubulin and microfilaments.
(a) Fluorescence photomicrograph of a cell of the ciliated protozoan Tetrahymena thermophila with red- and green-labeled antitubulin antibodies (the two dyes combine to give yellow) and with DAPI, which stains DNA (blue, nucleus). A cell is about 10 μm wide. (b) A mouse cell showing the role of tubulin (green) in separating chromosomes (blue) during metaphase of mitosis (cytoplasmic proteins stain red). Compare with Figure 2.44c. (c) Electron microscopic image of the cellular slime mold Dictyostelium discoideum showing the network of actin microfilaments that along with microtubules functions as the cell cytoskeleton. Microfilaments are about 7 nm in diameter.
Endoplasmic Reticulum, the Golgi Complex, and Lysosomes
The endoplasmic reticulum (ER) is a network of membranes continuous with the nuclear membrane. Two types of endoplasmic reticulum exist: rough ER, which contains attached ribosomes, and smooth ER, which does not (Figure 2.42). Smooth ER participates in the synthesis of lipids and in some aspects of carbohydrate metabolism. Rough ER, through the activity of its ribosomes, is a major producer of glycoproteins and also produces new membrane material that is transported throughout the cell to enlarge the various membrane systems before cell division.
The Golgi complex is a stack of membrane-bound sacs. In the Golgi complex, products of the ER are chemically modified and sorted into those destined for secretion versus those that will function in other membranous structures in the cell. Many of the modifications made in the Golgi complex are glycosylations (addition of sugar residues) that convert the proteins into glycoproteins that can then be targeted to specific locations in the cell.
Lysosomes (Figure 2.42) are membrane-enclosed compartments that contain digestive enzymes that hydrolyze proteins, fats, and polysaccharides. The lysosome fuses with food that enters the cell in vacuoles and then releases digestive enzymes that break down the foods for biosynthesis and energy generation. Lysosomes also function in degrading damaged cellular components and recycling these materials for new biosyntheses. The lysosome thus allows the cell’s lytic activities to be partitioned away from the cytoplasm proper. Following the degradation of macromolecules in the lysosome, the resulting nutrients pass from the lysosome into the cytoplasm for use by cytoplasmic enzymes.
Flagella and Cilia
Flagella and cilia are present on the surface of many eukaryotic microbes and function as motility structures, allowing cells to move by swimming. As we learned when we considered motility in prokaryotic cells (Sections 2.9, 2.10, 2.11 and 2.12), motility has survival value, as the ability to move allows motile organisms to move about their habitat and exploit new resources. Cilia are essentially short flagella that beat in synchrony to propel the cell—usually quite rapidly—through the medium. Flagella, by contrast, are long appendages present singly or in groups that propel the cell along—typically more slowly than by cilia—through a whiplike motion (**Figure 2.48*a***). In cross section, eukaryotic cilia and flagella appear similar. Each contains a bundle of nine pairs of microtubules surrounding a central pair of microtubules (Figure 2.48b). A protein called dynein is attached to the microtubules and uses ATP to drive motility. Movement of flagella and cilia is similar. In both cases, movement is the result of the coordinated sliding of microtubules against one another in a direction toward or away from the base of the cell. This movement confers the whiplike motion on the flagellum or cilium that results in cell propulsion.
Figure 2.48 Motility organelles in eukaryotic cells: Flagella and cilia.

(a) Flagella can be present as single or multiple filaments. Cilia are structurally similar to flagella but much shorter. Eukaryotic flagella move in a whiplike motion. (b) Cross section through a flagellum of the fungus Blastocladiella showing the outer sheath, the outer nine pairs of microtubules, and the central pair of microtubules.
Eukaryotic flagella differ structurally and functionally from the flagella of Bacteria and the archaella of Archaea, and these three structures should not be confused (contrast Figure 2.48 with Figure 2.34). Eukaryotic flagella are much larger than flagella or archaella, are enclosed by the cytoplasmic membrane and contain a cytoskeleton, and do not rotate but rather whip back and forth through the activity of their cytoskeleton proteins.
Armed with the major principles of microbial cell structure and function, we move on to Chapter 3 where the metabolic events that provide the energy necessary to build cellular components will unfold as a prelude to our consideration of the building process itself.
Check Your Understanding
Why are the activities in the lysosome partitioned from the rest of the cytoplasm?
Describe the structural elements that comprise the cytoskeleton.
In terms of structure and function, how do the flagella of eukaryotic cells differ from those of prokaryotic cells?
Chapter Review
Go to Mastering Microbiology for videos, animations, practice tests, and more.
I The Cell Envelope
2.1 The cytoplasmic membrane is a highly selective permeability barrier constructed of lipids and proteins that form a bilayer, with a hydrophobic interior and hydrophilic exterior. In contrast to Bacteria and Eukarya, where fatty acids are ester-linked to glycerol, Archaea contain ether-linked lipids, and some form monolayer instead of bilayer membranes. The major functions of the cytoplasmic membrane are permeability, transport, and energy conservation, and nutrient accumulation requires energy.
**Q Describe in a single sentence the structure of the cytoplasmic membrane. Describe the major structural differences between membranes of Bacteria and Archaea. How do solutes enter a cell through the cytoplasmic membrane?**
2.2 The active transport of nutrients into the cell is an energy-requiring process driven by ATP (or some other energy-rich compound) or by the proton motive force. Three transporters are employed by prokaryotic cells: simple, group translocation, and ABC systems. Each mechanism accumulates solutes against the concentration gradient.
**Q Cells of Escherichia coli transport lactose via lac permease, glucose via the phosphotransferase system, and maltose via an ABC-type transporter. For each of these sugars describe: (1) the components of the transport system and (2) the source of energy that drives the transport event.**
2.3 Peptidoglycan is a polysaccharide found only in Bacteria that consists of an alternating repeat of N-acetylglucosamine and N-acetylmuramic acid, the latter cross-linked by tetrapeptides in adjacent strands. The enzyme lysozyme and the antibiotic penicillin both destroy peptidoglycan, leading to cell lysis.
Q Why is the rigid layer of the bacterial cell wall called peptidoglycan? What are the structural reasons for the rigidity that is conferred on the cell wall by the peptidoglycan structure?
2.4 Gram-negative Bacteria have an outer membrane consisting of LPS, protein, and lipoprotein. Porins allow for permeability across the outer membrane. The gap between the outer and cytoplasmic membranes is called the periplasm and contains proteins that function in transport, sensing chemicals, and other important cell functions.
**Q List several functions of the outer membrane in gram-negative Bacteria. What is lipid A composed of?**
2.5 Cell envelopes can exhibit a range of different structures. One common variation is the presence of an outer S-layer composed of protein or glycoprotein. S-layers function as the cell wall for many Archaea and they are also found in many Bacteria.
Q What functions can S-layers provide to those cells that make them?
II Cell Surface Structures and Inclusions
2.6 Many prokaryotic cells have capsules, slime layers, or pili. These structures have several functions, including attachment, genetic exchange, and twitching motility. Hami, present on the surface of certain Archaea, function as miniature grappling hooks to attach cells to a surface or to one another.
Q What function(s) do polysaccharide layers outside the cell wall have in prokaryotic cells?
2.7 Prokaryotic cells can contain inclusions of sulfur, polyphosphate, carbon polymers, various minerals formed by biomineralization, or atmospheric gases. These substances function as nutrient storage materials, control magnetotaxis (in the case of magnetosomes), or confer buoyancy (in the case of gas vesicles).
Q How does an inclusion of poly-β-hydroxybutyric acid differ from a magnetosome in terms of structure and function?
2.8 The endospore is a highly resistant and differentiated structure produced by certain gram-positive Bacteria. Endospores are highly dehydrated and contain calcium dipicolinate and small acid-soluble spore proteins, both of which are absent from vegetative cells. Endospores can remain dormant indefinitely but can germinate quickly when conditions warrant.
Q In a few sentences, indicate how the bacterial endospore differs from the vegetative cell in structure, chemical composition, and ability to resist extreme environmental conditions. In addition, how do endospores differ from other types of spores?
III Cell Locomotion
2.9 Swimming motility in prokaryotic cells is due to flagella (Bacteria) or archaella (Archaea). Both structures are composed of several proteins, are anchored in the cell wall and cytoplasmic membrane, and cause cells to swim by their rotation. However, flagella and archaella differ in both structure and mechanism of operation.
**Q Compare and contrast the differences in structure and mechanism between the flagella of Bacteria and the archaella of Archaea.**
2.10 Some microbes are capable of surface motility in which they move along solid surfaces by one of several mechanisms including twitching or gliding.
**Q Contrast the mechanisms of gliding motility in Flavobacterium with twitching motility in Pseudomonas and swimming motility in Escherichia coli.**
2.11 Swimming bacteria respond to chemical and physical gradients in their environment by controlling the lengths of runs and frequency of tumbles, resulting in a biased random walk. Tumbles are controlled by the direction of rotation of the flagellum, which in turn is controlled by a network of sensory and response proteins.
Q In a few sentences, explain how a swimming bacterium is able to sense the direction of an attractant and move toward it.
2.12 Phototaxis describes directed motility in response to light intensity, while scotophobotaxis describes directed motility away from the dark. Aerotaxis describes directed motility in response to O2 gradients.
Q Magnetotactic bacteria do not exhibit directed motility in response to magnetic fields and hence their name is a misnomer. Describe the role of magnetosomes and aerotaxis in the directed movements of magnetotactic bacteria.
IV Eukaryotic Microbial Cells
2.13 Microbial eukaryotes contain several organelles including the nucleus, mitochondria, and chloroplasts. The nucleus contains the cell’s DNA wrapped around histone proteins. Cells of microbial eukaryotes divide following the process of mitosis and may also undergo meiosis if a haploid/diploid life cycle occurs.
Q List at least three features of eukaryotic cells that clearly differentiate them from prokaryotic cells.
2.14 Mitochondria and chloroplasts are energy-generating organelles. According to the endosymbiotic theory, these organelles were once free-living Bacteria that later established symbiotic relationships within ancestral cells of Eukarya.
**Q Describe the evidence that supports the idea that the major organelles of eukaryotes were once Bacteria.**
2.15 Endoplasmic reticula are membranous structures in eukaryotes that either contain attached ribosomes (rough ER) or do not (smooth ER). Flagella and cilia are means of motility and in eukaryotic cells move by a whiplike mechanism instead of by rotation. Lysosomes specialize in degrading proteins and other macromolecules no longer needed by the cell. Microtubules, microfilaments, and intermediate filaments function as internal cell structures that combine to form the cell cytoskeleton.
Q Compare and contrast the structure and function of eukaryotic flagella with those of prokaryotic cells.
Application Questions
Assume you are given two cultures, one of a species of gram-negative Bacteria and one of a species of Archaea. Discuss at least four different ways you could analyze these two cultures and tell which culture was which.
Gram-negative and gram-positive bacteria have different susceptibility to antibiotics because of differences in cell envelope structure. Assume that you are given an antibiotic that is effective against gram-positive cells but ineffective against gram-negative cells because it cannot cross the outer membrane. How might you chemically modify this antibiotic to increase its efficacy against gram-negative organisms?
Chapter Glossary
a membrane transport system consisting of three proteins, one of which hydrolyzes ATP; the system transports specific nutrients into the cell Archaellum
a long, thin cellular appendage present in some Archaea that rotates and is responsible for swimming motility Basal body
the “motor” portion of the bacterial flagellum, embedded in the cytoplasmic membrane and cell wall Capsule
a polysaccharide or protein outermost layer, usually rather slimy, present on some bacteria Cell envelope
the system of layered structures that surround the cytoplasm and define the outer boundary of the cell Chemotaxis
directed movement of an organism toward (positive chemotaxis) or away from (negative chemotaxis) a chemical gradient Chloroplast
the photosynthetic organelle of phototrophic eukaryotes Cristae
the internal membranes of a mitochondrion Cytoplasmic membrane
a semipermeable barrier that separates the cell interior (cytoplasm) from the environment Cytoskeleton
the cellular scaffolding typical of eukaryotic cells, made of microtubules, microfilaments, and intermediate filaments Dipicolinic acid
a substance unique to endospores which, when complexed with Ca2+, confers heat resistance on these structures Endospore
a highly heat-resistant, thick-walled, differentiated structure produced by certain gram-positive Bacteria Endosymbiotic theory
the idea that mitochondria and chloroplasts originated from Bacteria Flagellum
a long, thin cellular appendage that rotates in Bacteria or has a whiplike motion in Eukarya and is responsible for swimming motility Gas vesicles
gas-filled cytoplasmic structures bounded by a single layered protein membrane that confer buoyancy on cells Gliding motility
a form of surface motility characterized by smooth continuous movement Group translocation
an energy-dependent transport system in which the substance transported is chemically modified during the process of being transported by a series of proteins Histones
highly basic proteins that compact and wind DNA in the nucleus of eukaryotic cells Intermediate filament
a filamentous polymer of fibrous keratin proteins, supercoiled into thicker fibers, that functions in maintaining cell shape and the positioning of certain organelles in the eukaryotic cell Lipopolysaccharide (LPS)
a combination of lipid with polysaccharide and protein that forms the major portion of the outer membrane in gram-negative Bacteria Lysosome
an organelle containing digestive enzymes for hydrolysis of proteins, fats, and polysaccharides Magnetosome
a particle of magnetite (Fe3O4) or greigite (Fe3S4) enclosed by a single membrane in the cytoplasm of magnetotactic Bacteria Meiosis
the nuclear division that halves the diploid number of chromosomes to the haploid Microfilament
a filamentous polymer of the protein actin that helps maintain the shape of a eukaryotic cell Microtubule
a filamentous polymer of the proteins α-tubulin and β-tubulin that functions in eukaryotic cell shape and motility Mitochondrion
the respiratory organelle of eukaryotic organisms Mitosis
nuclear division in eukaryotic cells in which chromosomes are replicated and partitioned into two daughter cells during cell division Nucleus
a membrane-enclosed structure in eukaryotic cells that contains the cell’s DNA genome Outer membrane
a phospholipid- and polysaccharide-containing unit membrane that lies external to the peptidoglycan layer in cells of gram-negative Bacteria and is a component of the cell envelope in these organisms Peptidoglycan
a polysaccharide composed of alternating repeats of N-acetylglucosamine and N-acetylmuramic acid arranged in adjacent layers cross-linked by short peptides; a component of the cell envelope of virtually all Bacteria Periplasm
a region between the outer surface of the cytoplasmic membrane and the inner surface of the lipopolysaccharide layer of gram-negative Bacteria Peritrichous flagellation
having flagella located in many places around the surface of the cell Phototaxis
thin, filamentous structures that extend from the surface of a cell and, depending on type, facilitate cell attachment, genetic exchange, or twitching motility Polar flagellation
having flagella emanating from one or both poles of the cell poly-β-hydroxybutyric acid (PHB)
a common storage material of prokaryotic cells consisting of a polymer of β-hydroxybutyrate or another β-alkanoic acid or mixtures of β-alkanoic acids S-layer
an outermost cell surface layer composed of protein or glycoprotein present on some Bacteria and many Archaea Simple transport system
a transporter that consists of only a membrane-spanning protein and is typically driven by energy from the proton motive force Stroma
the lumen of the chloroplast, surrounded by the inner membrane Teichoic acid
a phosphorylated polyalcohol found in the cell wall of some gram-positive Bacteria Thylakoids
membrane stacks containing the photosynthetic pigments in cyanobacteria or in the chloroplast of eukaryotic phototrophs Twitching motility
a form of surface motility caused by extension and retraction of type IV pili