4 Microbial Growth and Its Control
I Culturing Microbes and Measuring Their Growth
III Environmental Effects on Growth: Temperature
IV Environmental Effects on Growth: pH, Osmolarity, and Oxygen
Growing Their Own Way
When thinking of microbial growth we typically think of binary fission, the splitting of one cell into two, and indeed this form of growth is common and widespread in the microbial world. But microbial growth takes many different forms. Streptomyces, for example, have a complex life cycle, as antibiotic producers often do. Streptomyces are common soil bacteria that produce secondary metabolites such as those called geosmin, which give soil its characteristic odor. Streptomyces also produce a wide range of other secondary metabolites, including many clinically useful antibiotics. Indeed, the antibiotic streptomycin provided the first cure for tuberculosis, and its discovery earned the soil microbiologist Selman Waksman a Nobel Prize in 1952. Progress in understanding the Streptomyces life cycle is now being made using techniques to grow and image Streptomyces in microfluidic chambers.
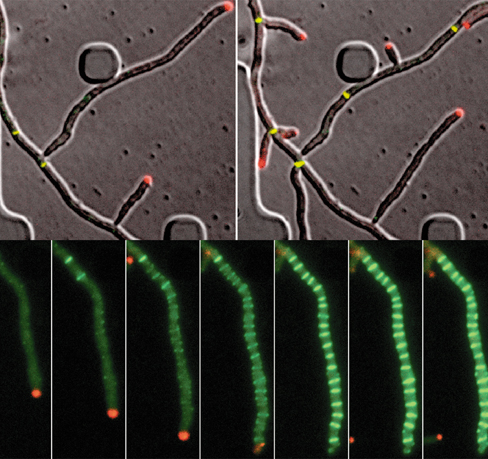
The life cycle of Streptomyces begins with vegetative growth, during which filamentous cells grow by elongating from the tip without cell division. This elongation without division can be visualized (upper image set) in stains where a red fluorescent protein has been fused to a protein that localizes to the growth tip. Cell division is visualized by a yellow fluorescent protein fused to FtsZ, a protein that mediates septa formation.
Nutrient limitation triggers the formation of aerial hyphae and sporulation. Streptomyces sporulation proceeds through multiple fission (lower image set). That is, one filamentous cell divides to form many spores. As seen in the image, many septa are formed simultaneously, resulting in up to 20 spores. Genetic analyses, facilitated by live imaging of growing cells, has revealed that dynamin-like proteins are required during this process. Dynamin is present in eukaryotic cells where it mediates membrane remodeling. This discovery in Streptomyces suggests that dynamin-like proteins may have similar functions in bacteria and shows that we still have a lot to learn about microbial growth.
Source: Schlimpert, S., et al. 2017. Two dynamin-like proteins stabilize FtsZ rings during Streptomyces sporulation. Proc. Natl. Acad. Sci. USA E6176.
In previous chapters we discussed fundamental aspects of microbiology including cell structure and function (Chapter 2) and the principles of microbial metabolism (Chapter 3). In this chapter we explore the areas of microbial growth and cultivation. We will also learn about environmental factors that influence microbial growth and consider some of the major techniques for the control and prevention of microbial growth.
In the first few sections we consider how microbes are grown in laboratory culture and how microbial growth is measured. Recall that when we use the term microbial growth we are referring to an increase in population size as the result of cell division (Section 1.2). Culturing microbes and assessing their growth are major events in the daily routine of many microbiologists and microbiology laboratories.
I Culturing Microbes and Measuring Their Growth
Microorganisms require a suite of nutrients as foodstuffs to produce new microbial cells. Several methods exist for quantifying cell numbers in either natural environments or laboratory cultures.
4.1 Feeding the Microbe: Cell Nutrition
Because the metabolic capacities of microbes differ, their nutrient requirements also differ. However, all microbes require a core set of nutrients. Some nutrients, called macronutrients, are required in large amounts, while others, called micronutrients, are required in minute amounts. We begin by dissecting the cell to reveal its chemical composition and then consider the nutrients that all cells require.
Chemical Makeup of a Cell
A single cell of Escherichia coli weighs, on average, just about 10−12 g, and over 75% of that mass is water. However, when considering the composition of cells, it is typical to refer to cell dry weight, which for E. coli is about 184×10−15 g (184 fg) per cell. Just a handful of the chemical elements predominate in living systems. The elements carbon (C), oxygen (O), nitrogen (N), hydrogen (H), phosphorus (P), and sulfur (S) account for about 96% of the dry weight of an average bacterial cell (Figure 4.1), and these six elements are universally required by all life forms. The next 3.7% of a cell’s mass is composed of potassium (K), sodium (Na), calcium (Ca), magnesium (Mg), chlorine (Cl), and iron (Fe) (Figure 4.1c), and these elements are required by most (but not all) microorganisms. In addition, at least another 20 elements can be found in cells of E. coli (Figure 4.1a) and in cells across the three domains of life. In total, microbes can metabolize 62 different elements in one way or another (Figure 4.1a).
Figure 4.1 Elemental and macromolecular composition of a bacterial cell.

(a) A microbial periodic table of the elements. With the exception of those shown in row 7, other elements in row 7 and elements in rows beyond row 7 are not known to be metabolized. (b) Relative abundance of macromolecules in a bacterial cell. Data from Escherichia coli and Salmonella typhimurium: Cellular and Molecular Biology. ASM, Washington, DC (1996). (c) Distribution of the elements in an average E. coli cell (dry weight). “Other” includes in order of abundance: Se, B, Cu, Mn, Zn, F, Si, As, Cs, Sr, Ba, V, Co, W, and Ni.
Cells are comprised mostly of macromolecules, including proteins, lipids, polysaccharides, lipopolysaccharides, and nucleic acids, which together comprise more than 96% of the dry weight of an E. coli cell (Figure 4.1b). The vast majority of these are proteins and RNAs (Figure 4.1b). Interestingly, as important as DNA is to a cell (the cell genome), it contributes a very small percentage of a cell’s dry weight (Figure 4.1b).
Carbon, Nitrogen, and Other Macronutrients
Carbon and nitrogen are present in large amounts in all cells. Heterotrophs (organisms that require organic carbon) obtain carbon from the breakdown of organic polymers or from the direct uptake of their monomeric constituents: the amino acids, fatty acids, organic acids, sugars, nitrogen bases, and other organic compounds. Some microbes are autotrophs and can synthesize organic compounds from carbon dioxide (CO2). The bulk of nitrogen in nature exists in proteins, ammonia (NH3), nitrate (NO3−), or nitrogen gas (N2). Virtually all microorganisms can use NH3 as their nitrogen source and many can also use nitrate (NO3−). Some microbes can use organic nitrogen sources, such as amino acids, and a few can use N2 (the nitrogen-fixing bacteria).
In addition to C and N (and O and H from H2O), many other macronutrients are needed by cells but typically in smaller amounts (Figure 4.1c). Phosphorus is required for nucleic acids and phospholipids and is usually assimilated by microbes as inorganic phosphate (PO43−). Sulfur is present in the amino acids cysteine and methionine and also in several vitamins, including thiamine, biotin, and lipoic acid. Most microbes can assimilate inorganic forms of S such as sulfate (SO42−) or sulfide (H2S) while others assimilate S from organic sulfur compounds. Potassium (K) is required for the activity of several enzymes, whereas magnesium (Mg) stabilizes ribosomes, membranes, and nucleic acids and is also required for the activity of many enzymes. Calcium (Ca) and sodium (Na) are essential nutrients for only a few organisms. For example, most marine organisms have a strict requirement for NaCl.
Micronutrients: Trace Metals and Growth Factors
Many enzymes require a metal ion or small organic molecule as a cofactor in order to catalyze their specific reaction (Section 3.5). For this reason, the growth of microorganisms requires various metals, and chief among these is iron (Fe). Iron is present in cytochromes and several other enzymes that play major roles in cellular respiration or related oxidation–reduction reactions. Besides iron, many other metals may be required or otherwise metabolized by microorganisms (Figure 4.1a). Collectively these metals are called trace metals because they are required in such small amounts. Table 4.1 lists the major trace metals and other trace elements of life and examples of enzymes or other molecules in which each is found. Enzymes with a trace metal requirement may be synthesized by the cell but will not function properly if the cell is starved for these metals.
Table 4.1 Micronutrients needed by microorganismsa


aNot all trace elements or growth factors are needed by all organisms, and many growth factors are biosynthesized and not required from the environment.
bIron is typically needed in larger amounts than the other trace metals shown.
cThe production of methane (CH4) by methanogens (Archaea).
Growth factors differ from trace metals in that they are organic micronutrients rather than metals (Table 4.1). Vitamins, which typically function as coenzymes (Section 3.5), are the most frequently required growth factors, and a few common ones are shown in Table 4.1. While some microbes are able to biosynthesize all of their own growth factors, many organisms (including animals) need to assimilate diverse growth factors from the environments (or from their diet). Besides vitamins, other common growth factors can include amino acids, purines, pyrimidines, and several other organic molecules.
Growth factor requirements vary widely among microorganisms. For example, cyanobacteria are autotrophic microbes that inhabit aquatic environments such as lakes and oceans; they can synthesize all of their own growth factors and need little if any supplementation. By contrast, the lactic acid bacteria Streptococcus, Lactobacillus, and Leuconostoc inhabit environments rich in nutrients, such as certain foods and in the guts of animals (Sections 16.6 and 24.2), and as a result have evolved to rely on their organic-rich environments for these required nutrients.
If a cell is to grow and divide, it must take up macronutrients and micronutrients from the environment and transform them into new cell material. Hence, to grow microbes in the laboratory, we need to satisfy their growth requirements. If the nutrients, trace metals, and growth factors that a microbe needs are not made available to it, the organism will not grow. In the next section we consider how these important factors are taken into account to support the robust growth of microbes in laboratory culture media.
Check Your Understanding
Which four chemical elements make up the bulk of a cell’s dry weight?

Which two classes of macromolecules contain most of a cell’s nitrogen?
Differentiate between trace metals and growth factors. How are these used by the cell?
4.2 Growth Media and Laboratory Culture
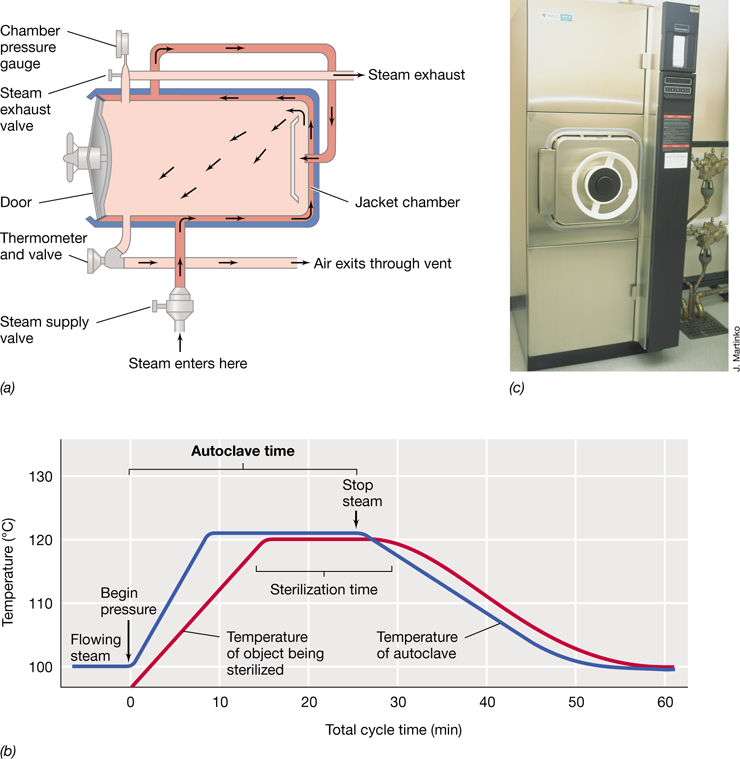
Laboratory cultures of microorganisms are grown in culture media (singular, medium), nutrient solutions tailored to the particular organism to be grown (Section 1.13). Because laboratory culture of a microbe is required for its detailed study, careful attention must be paid to the selection and preparation of media for laboratory culture to be successful. Culture media must be sterilized before use, and sterilization is typically achieved by heating the medium under pressure in an autoclave. We discuss the operation and principles of the autoclave in Section 4.17, along with other methods for sterilizing culture media and laboratory utensils and other devices.
Classes of Culture Media
Although thousands of recipes for microbial culture media exist, two broad classes of culture media are widely used in microbiology: defined media and complex media. Defined media are prepared by adding precise amounts of pure inorganic or organic chemicals to distilled water. Therefore, the exact composition of a defined medium (in both a qualitative and quantitative sense) is known. Of major importance in any culture medium is the carbon source because all cells need large amounts of carbon to make new cell material (Figure 4.1). The particular carbon source and its concentration depend on the organism to be cultured. Table 4.2 lists recipes for four different culture media. Some defined media, such as the one listed for Escherichia coli, are considered “simple” because they contain only a single carbon source. In such a medium, E. coli biosynthesizes all of its cellular materials from glucose.
Table 4.2 Examples of culture media for microorganisms with simple and demanding nutritional requirementsa

aThe photos are tubes of (a) the defined medium for Escherichia coli, and (b) the complex medium described. Note how the complex medium is colored from the various organic extracts and digests that it contains. Photo credits: Cheryl L. Broadie and John Vercillo, Southern Illinois University at Carbondale.
For culturing many microorganisms, knowledge of the exact composition of a medium is not essential. In these instances, complex media may suffice and may even be advantageous. Complex media are made from digests of microbial, animal, or plant products, such as milk protein (casein), beef (beef extract), soybeans (tryptic soy broth), yeast cells (yeast extract), or any of a number of other highly nutritious substances (Table 4.2). For example, the media originally used by Pasteur in his classic early studies in microbiology were complex media containing yeast extract (Section 1.11). Such digests are commercially available in dehydrated form and need only be hydrated with distilled water to form a culture medium. However, the disadvantage of a complex medium is that its exact nutritional composition is unknown.
Culture media are sometimes prepared that are selective or differential (or both), especially media used in medical microbiology. A selective medium contains compounds that inhibit the growth of some microorganisms but not others. Selective media are commercially available for the isolation of certain common gut pathogens, such as the enteric bacteria Salmonella or those strains of E. coli that cause foodborne illnesses. For example, bile salts are often added to culture media for the selective isolation of these bacteria because bile salts kill many bacteria that are unable to grow in the gut. A differential medium is one to which an indicator (typically a dye) is added, which reveals by a color change whether a particular metabolic reaction has occurred during growth. For example, the detection of lactic acid bacteria, which secrete lactic acid and cause the pH to drop, can be facilitated by incorporating a pH-sensitive dye into the culture medium that changes color when conditions become acidic. Differential media are useful for distinguishing bacteria and are widely used in clinical diagnostic microbiology and in microbial taxonomy. Differential and selective media are discussed in a clinical microbiology context in Chapter 29.
Nutritional Requirements and Biosynthetic Capacity
Of the four culture medium recipes in Table 4.2, three are defined and one is complex. The complex medium, which is rich in nutrients, is easiest to prepare and it supports growth of both Escherichia coli (a common enteric bacterium) and Leuconostoc mesenteroides (a lactic acid bacterium often associated with fermented foods), the examples used in Table 4.2. By contrast, only E. coli grows in the simple defined medium because L. mesenteroides has more nutritional requirements (it requires more growth factors) than E. coli. The nutritional needs of L. mesenteroides can be satisfied by preparing either a highly supplemented defined medium, a laborious undertaking because of all the individual nutrients that need to be added (Table 4.2), or by preparing a complex medium, a much easier and quicker operation.
The fourth medium listed in Table 4.2 supports growth of the bacterium Thiobacillus thioparus, an aerobic sulfur-oxidizing chemolithotroph (Section 3.11). T. thioparus derives all of its carbon from CO2 (that is, it is autotrophic) and conserves energy from the oxidation of sulfur compounds such as thiosulfate (Na2S2O3). T. thioparus is common in low-nutrient sulfur-rich environments where it only has access to inorganic nutrients. Thus, T. thioparus has evolved to biosynthesize all of its needed growth factors and does not require organic nutrients of any type.
The take-home lesson from Table 4.2 is that different microorganisms may have vastly different nutritional requirements. For successful cultivation, it is necessary to understand an organism’s physiology and nutritional requirements and then supply it with the nutrients it needs in both the proper form and amount.
Laboratory Culture
Laboratory media can be either liquid (Table 4.2) or solid (Figure 4.2). Culture media are solidified with agar, an algal polysaccharide first used in the classical studies of Robert Koch (Section 1.12). Solid media immobilize cells so that as they grow, they accumulate in a pile to form visible, isolated cell masses called colonies (Figure 4.2). Microbial colonies vary in shape, color, texture, and size depending on the organism, the culture conditions, the nutrient supply, and other physiological parameters. Colony appearance is consistent for a given organism on a particular type of medium. Hence, colony morphology—the visible characteristics of a colony—can sometimes be used to identify microorganisms but is used routinely to determine if a culture is pure (only one microbe is present), contaminated (undesired organisms co-occur with a desired organism), or mixed (many microbes are present). Plates inoculated from a mixed culture (such as a natural sample, Figure 4.2e) or from a contaminated pure culture will typically contain more than one colony type.
Figure 4.2 Bacterial colonies.
Colonies are visible masses of cells formed from the division of one or a few cells and can contain over a billion (109) individual cells. (a) Serratia marcescens, grown on MacConkey agar. (b) Close-up of colonies outlined in part a. (c) Pseudomonas aeruginosa, grown on trypticase soy agar. (d) Shigella flexneri, grown on MacConkey agar. (e) An agar plate containing many different bacterial colonies that developed from plating a dilution of seawater.
Propagation of microbial cultures requires aseptic technique (Figure 4.3), a series of steps by which microbes are transferred between growth media without contamination. Contamination can be introduced from microbes in the air, in liquid droplets, or on surfaces. With liquid medium, the goal is to transfer a microbial culture while protecting the culture vessel from air currents or contact with nonsterile surfaces (Figure 4.3a). With agar plates, the plan is basically the same but with a greater emphasis on keeping the surface of the agar protected from aerosols or particulate matter in air (Figure 4.3b). A mastery of aseptic technique is required for maintaining pure cultures, as airborne contaminants are virtually everywhere, even in what may appear to be a very clean microbiology laboratory.
Figure 4.3 Aseptic technique.

(a) Liquid media: After the tube is recapped at the end, the loop is resterilized. (b) Solid media: making a streak plate to obtain pure cultures. The plate cover is opened just enough to permit streaking manipulations. In streaking a plate, the microbial cells are separated by the streaking process to yield widely separated single cells that then grow and divide to form colonies.
Mastering Microbiology
Art Activity: Figure 4.3b Streaking a Petri plate
The primary method for obtaining pure cultures that contain a single microbe, and of verifying culture purity, is to perform the streak plate technique (Figure 4.3b). In this method, an inoculating loop is used to spread a sample across a solidified medium on an agar plate. Any microbes that are able to grow will form colonies, and colony morphology can be examined to determine the number and type of microbes present.
Many techniques for obtaining pure cultures have been developed, and some of the more common ones will be discussed in Chapter 19. But for now, we turn our attention away from general methods of culturing microbes to explore methods for quantifying their growth.
Check Your Understanding
In which medium shown in Table 4.2, defined or complex, do you think Escherichia coli would grow the fastest?
What is meant by the word sterile? Why is aseptic technique necessary for successful maintenance of pure cultures in the laboratory?
How many E. coli cells would be present in a single bacterial colony whose dry weight was 0.2 mg?
4.3 Microscopic Counts of Microbial Cell Numbers
Assessing cell numbers gives quantitative information on the state of a laboratory microbial culture or a microbial community in nature. Several methods for enumerating a microbial population have been developed, each having their own strengths and caveats. We begin with the classic “total count” carried out by microscopic examination of a culture or natural sample.
Total Cell Count
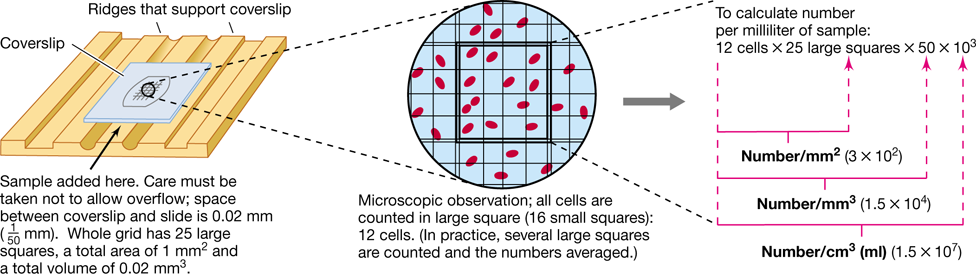
Total counts of microbial numbers in a culture or natural sample can be done by simply observing and enumerating the cells present by a microscopic cell count. Microscopic counts can be performed either on samples dried on slides or on liquid samples. Dried samples can be stained to increase contrast between cells and their background (Section 1.8) when observed by bright-field microscopy, but a phase-contrast microscope is essential for observing unstained preparations. With liquid samples, counting chambers consisting of a grid with squares of known area etched on the surface of a glass slide are used (Figure 4.4). When the coverslip is placed on the chamber, each square on the grid has a precise volume. The number of cells per unit area of grid can be counted under the microscope, giving a measure of the number of cells per small chamber volume. The number of cells per milliliter of suspension is calculated by employing a conversion factor based on the volume of the chamber sample (Figure 4.4).
Figure 4.4 Direct microscopic counting procedure using the Petroff–Hausser counting chamber.

A phase-contrast microscope is typically used to count the cells to avoid the necessity for staining.
Microscopic counting is a quick and easy way of estimating microbial cell numbers. However, it has several limitations that restrict its usefulness. For example, without special staining techniques, dead cells cannot be distinguished from live cells, and precision is difficult to achieve, even when replicate counts are made. Moreover, small cells are often difficult to see under the microscope, which can lead to erroneous counts, and cell suspensions of low density (less than about 106 cells/milliliter) will have few if any cells in a microscope field unless the sample is first concentrated and resuspended in a small volume. Finally, motile cells must be killed (usually with formaldehyde) or otherwise immobilized before counting, and debris in the sample may easily be mistaken for microbial cells.
Microscopic Cell Counts in Microbial Ecology
Despite the many potential caveats, microbial ecologists often use microscopic cell counts on natural samples. But they do so by using stains to visualize the cells, often stains that yield phylogenetic or other key information about the cells, such as their metabolic properties (Section 19.5).
Many fluorescing stains can be used in a general way. For example, the stain DAPI (Section 1.8 and Figure 1.25e) stains all cells because it binds tightly to DNA. Other general fluorescent stains can differentiate live from dead cells by detecting whether a cell’s cytoplasmic membrane is intact or not. By contrast, fluorescent stains that are highly specific for certain organisms or groups of related organisms can be prepared by attaching fluorescent dyes to specific nucleic acid probes. For example, phylogenetic stains that stain only species of Bacteria or only species of Archaea can be used in combination with nonspecific stains to determine the proportion of each domain present in a sample (Section 19.5). Other fluorescent probes have been developed that target genes encoding enzymes that catalyze specific metabolic processes; if a cell is stained by one of these probes, a metabolism can be inferred that may reveal the cell’s ecological role in the microbial community. In all of these cases, if cells in the sample are present in only low numbers, for example in a sample of ocean water, this limitation can be overcome by first concentrating the cells on a filter and then counting them after staining.
Because they are easy to do and often yield useful baseline information, microscopic cell counts are common in ecological studies of natural microbial environments. We pursue this theme in more detail in Chapter 19. In the next section we consider quantifying microbes using growth-based methods.
Check Your Understanding
What are some of the problems that can arise when unstained preparations are enumerated in microscopic counts?
Using microscopic techniques, how might you determine the number of Archaea present in an alpine lake?
4.4 Viable Counting of Microbial Cell Numbers
A viable cell is one that is alive and able to grow. Some viable microbes are able to grow on laboratory culture media; thus, while microscopic counts often fail to distinguish viable microbes from dead microbes, we can use culture media to perform viable counts. A viable count is performed by spreading microbes on solid media and counting colonies, and hence is often called a plate count because agar plates are required. The assumption made in a viable count is that each viable cell will grow and divide to yield a single colony, and hence, colony numbers are taken as an measure of cell numbers.
Methods for Viable Counts
There are at least two ways of performing a plate count: the spread-plate method and the pour-plate method (Figure 4.5). In the spread-plate method, a volume (usually 0.1 ml or less) of an appropriately diluted culture is spread over the surface of an agar plate using a sterile glass spreader. In the pour-plate method, a known volume (usually 0.1–1.0 ml) of culture is pipetted into an empty sterile Petri plate. Molten agar medium, tempered to just above gelling temperature (50 °C), is then added and gently mixed before allowing the agar to solidify.
Figure 4.5 Two methods for the viable count.

Only surface colonies form in the spread plate technique. By contrast, in the pour-plate method, colonies form within the agar as well as on the agar surface. On the far right are photos of colonies of Escherichia coli formed from cells plated by the spread-plate method (top) or the pour-plate method (bottom).
With both the spread-plate and pour-plate methods, it is important that the number of colonies developing on or in the medium not be too many or too few. On crowded plates some cells may not form colonies, and some colonies may fuse, leading to erroneous measurements. If the number of colonies is too small, the statistical significance of the calculated count will be low. The usual practice, which is most valid statistically, is to count colonies only on plates that contain between 30 and 300 colonies.
A major assumption of the viable plate count technique is that one cell forms one colony. However, we have already learned that some bacteria are filamentous and many form clusters or sticky biofilms (Chapter 1). For example, a bacterial filament might contain 10 or more cells, but since these cells are firmly attached together, such a filament would form only one colony. Hence, data from viable counts are typically expressed as the number of colony-forming units (CFU) obtained rather than the actual number of viable cells, to account for clumps containing more than one viable cell.
Diluting a Sample
Samples often contain thousands, millions, or even billions of viable bacteria, and so samples must typically be diluted to obtain a suspension that will yield a countable number of colonies. A series of 10-fold dilutions are commonly used prior to performing viable plate counts (Figure 4.6). To make a 10-fold dilution, 1 ml of sample can be added to 9 ml of diluent (usually water, a dilute salts solution, or growth medium). In this way, one milliliter of this 10-fold dilution contains 0.1 ml (10−1 ml) of the original sample and 0.9 ml of diluent.
Figure 4.6 Procedure for viable counting using serial dilutions of the sample and the pour-plate method.

The sterile liquid used for making dilutions can simply be water, but a solution of mineral salts or actual growth medium may yield a higher recovery. The dilution factor is the reciprocal of the dilution.
With dense cultures, several serial dilutions are needed to yield a countable number of colonies (Figure 4.6). For example, if a sample is suspected to contain more than 105 cells then the sample can be diluted 1000-fold by performing three successive 10-fold dilutions (1/10×1/10×1/10=1/1000). In this way, one milliliter of a 1000-fold dilution contains 0.001 ml (10−3 ml) of sample and 0.999 ml of diluent. To determine the number of cells in the sample, the number of colonies observed is divided by the amount of sample plated. Hence, if 1 ml of a 1000-fold dilution is plated, this is equivalent to plating 0.001 ml of the original sample. If 159 colonies form from this sample, the original sample must have contained 159×103 (which is equivalent to 1.59×105) CFU (Figure 4.6). Because the number of bacteria in a sample is often not known ahead of time, it is usually necessary to make multiple dilutions to ensure a countable number of colonies (Figure 4.6).
Applications of the Plate Count
Despite the caveats associated with viable counting, the procedure is quick and easy to do and so is widely used in microbiology. For example, in food, dairy, medical, and aquatic microbiology, viable counts are employed routinely. The method has the virtue of high sensitivity, because as few as one viable cell per sample plated can be detected. This feature allows for the sensitive detection of microbial contamination of foods or other materials.
The use of highly selective culture media and growth conditions allows the plate count to be used to target particular species in a sample containing many organisms. For example, a complex medium containing 10% NaCl is very useful in isolating species of potentially pathogenic Staphylococcus from skin, because the salt inhibits growth of most other skin bacteria. In practical applications such as in the food industry, viable counting performed using both a complex medium and a selective medium on the same sample allows for simultaneous quantitative and qualitative assessments of food quality and safety. That is, with a single food sample, the complex medium yields a total cell count—a relative indicator of freshness and shelf life—while the selective medium indicates the presence or absence of a particular pathogen that may be transmitted in the food.
Targeted counting is also common in wastewater and other water analyses. For instance, enteric bacteria such as Escherichia coli originate from feces and are easy to recover from natural samples using selective media; if enteric bacteria are detected in a water sample from a swimming site, for example, their presence is a signal that the water contains fecal matter and is therefore unsafe for human contact.
Caveats to Plate-Counting Methods
Direct microscopic counts of natural samples typically reveal far more microbes than are recoverable on any single culture medium. Thus, although a very sensitive technique, plate counts can be highly unreliable when used to assess total cell numbers of natural samples, such as soil and water. In microbiology, this has been referred to as the great plate count anomaly.
Why do plate counts reveal lower numbers of cells than direct microscopic counts? The reason is that different organisms, even those present in a very small natural sample, have vastly different requirements for nutrients and growth conditions in laboratory culture (Sections 4.1 and 4.2 and Table 4.2). Thus, one medium and set of growth conditions can only be expected to support the growth of one subset of the total microbial community. For example, if a sample has 109 viable cells but only 106 them are capable of growing on a given culture medium, then the viable plate count will miss 99.9% of the viable cell population, a vast underestimation of the actual number of organisms present in the sample.
Plate count results thus carry a large caveat. Plate counts targeted to specific organisms using highly selective media (Section 4.2), as in, for example, the microbial analysis of sewage or food, can often yield quite reliable data, since the physiology of the targeted organisms is known and so the recovery of viable cells is near 100%. By contrast, “total” cell counts of the same samples using a single medium and set of growth conditions may be, and usually are, underestimates of actual cell numbers by one to several orders of magnitude. Because of this problem, a wide variety of molecular methods have been developed to detect and quantify specific organisms in natural samples without the need for cultivating them in the laboratory. We explore these methods and the important information they can reveal in Chapter 19.
Check Your Understanding
Why is a viable count more sensitive than a microscopic count? What major assumption is made in relating plate count results to cell number?
Describe how you would dilute a bacterial culture by 10−7.
4.5 Turbidimetric Measures of Microbial Cell Numbers
4.5 Turbidimetric Measures of Microbial Cell Numbers
4.5 Turbidimetric Measures of Microbial Cell Numbers
Though small, microbial cells are large enough to scatter light, and the amount of light scattered in a cell suspension is proportional to the number of cells present. A cell suspension looks cloudy (turbid) to the eye because cells scatter light that passes through the suspension. The more cells that are present, the more light is scattered, and hence the more turbid the suspension. Hence, turbidity measurements offer a rapid and widely used approach to estimating cell numbers in a solution.
Optical Density and Its Relationship to Cell Numbers
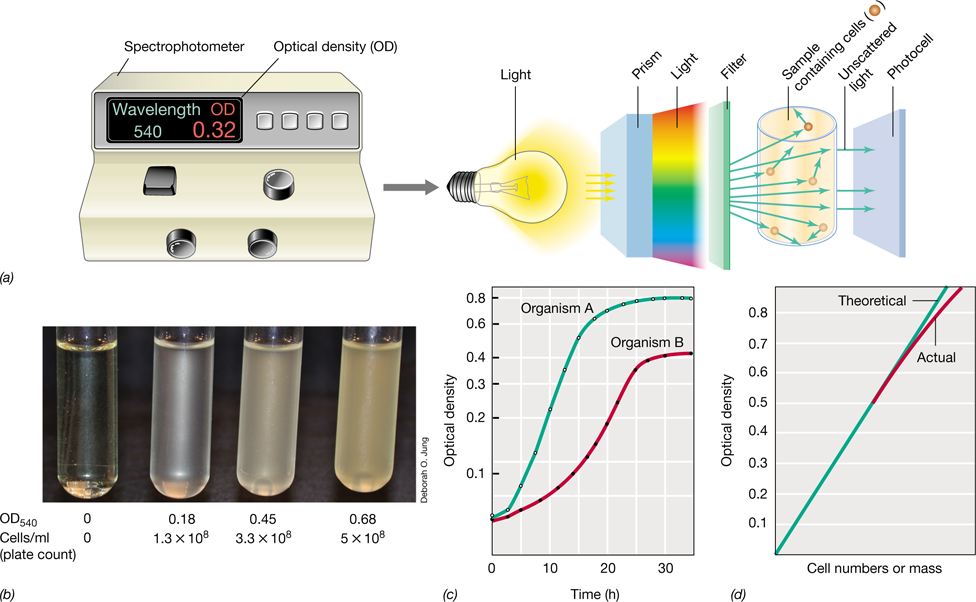
Turbidity is measured with a spectrophotometer, an instrument that measures the unscattered light that passes through a sample (Figure 4.7). A spectrophotometer employs a prism or diffraction grating to generate incident light of a specific wavelength (Figure 4.7a). Commonly used wavelengths for microbial turbidity measurements include 480 nm (blue), 540 nm (green), and 660 nm (red). Sensitivity is best at shorter wavelengths, but measurements of dense cell suspensions are more accurate at longer wavelengths.
Figure 4.7 Turbidity measurements of microbial growth.

(a) Measurements of turbidity are made in a spectrophotometer. The photocell measures incident light unscattered by cells in suspension and gives readings in optical density units. (b) Liquid cultures of Escherichia coli; the tube on the far left is sterile medium. The increasing (left to right) optical density (OD540) of each culture is shown below the tube, as is the actual cell number measured in a viable count. (c) Typical growth curve data for two organisms growing at different growth rates. For practice, calculate the generation time of the two cultures using the formula n=3.3 (log Nt−log N0) where Nt and N0 are two different OD readings with a time interval t between the two (see Section 4.7). Which organism is growing faster, A or B? (d) Relationship between cell number or dry weight and turbidity readings. Note that the direct relationship between these values breaks down at high turbidities.
The unit of turbidity is optical density (OD) at the wavelength specified, for example, OD540 for measurements at 540 nm (Figure 4.7). For unicellular organisms, optical density is proportional, within certain limits, to cell number. Turbidity readings can therefore be used as a substitute for total or viable counting methods. However, before this can be done, a standard curve must be prepared that relates cell number (microscopic or viable count) to turbidity. As can be seen in such a plot, proportionality only holds within limits (Figure 4.7d). At high cell densities, light scattered away from the spectrophotometer’s photocell by one cell can be scattered back toward the photocell by another, and as a result, the one-to-one correspondence between cell number and turbidity deviates from linearity. Nevertheless, when a standard has been prepared, turbidimetric estimates of bacterial numbers are very useful.
Other Issues with Turbidimetric Growth Estimates
On the one hand, turbidity measurements are quick and easy to perform and can be made without destroying or significantly disturbing the sample. For these reasons, turbidity measurements are widely employed to monitor growth of pure cultures of Bacteria and Archaea and can also be used with many microbial eukaryotes. With turbidimetric assays, the same sample can be checked repeatedly over time and the measurements plotted on a semilogarithmic plot versus time (Figure 4.7c) to measure growth of a microorganism over time.
On the other hand, turbidity measurements can occasionally be problematic. Although many microorganisms grow evenly distributed in suspensions in liquid medium, many do not. Some bacteria routinely form small to large clumps, and in such instances, OD measurements may be quite inaccurate as a measure of total microbial mass. In addition, many bacteria form biofilms on the sides of tubes or other growth vessels. Hence for OD measurements to be an accurate reflection of cell mass (and thus cell numbers) in a liquid culture, clumping and biofilms have to be minimized. This can be accomplished by stirring, shaking, or in some way keeping the cells well mixed during the growth process to prevent the formation of cell aggregates and the sticking of swimming cells to surfaces. Some bacteria are just naturally planktonic and stay well suspended in liquid medium for long periods. But if a solid surface is available, most bacteria will eventually develop a static biofilm, and accurately quantifying cell numbers by turbidity in such a case can be difficult or even impossible.
Check Your Understanding
List two advantages of using turbidity as a measure of cell growth.
Describe how you could use a turbidity measurement to tell how many colonies you would expect from plating a culture of a given OD.
II Dynamics of Microbial Growth
Microorganisms that divide by binary fission grow at exponential rates into cell populations whose numbers over time are predictable from simple mathematical models. A few microbes grow by budding, and some can form sticky cell masses called biofilms.
In Part II of this chapter we consider the actual process of microbial growth. Here we consider the different ways in which microbes grow, explore the basic quantitative relationships displayed by growing microbial cultures, and introduce an important laboratory tool—the chemostat—for manipulating exponentially growing microbial cultures to better answer specific questions.
4.6 Binary Fission and the Microbial Growth Cycle
Growth is the result of cell division and is the ultimate process in the life of a microbial cell. In microbiology, growth is defined as an increase in the number of cells. As macromolecules accumulate in the cytoplasm of a cell, they assemble into major cell structures, such as the cell envelope, cytoplasmic membrane, ribosomes, and so on, eventually leading to cell division. In a growing culture of a rod-shaped bacterium such as Escherichia coli, cells elongate to approximately twice their original length and then form a partition that constricts the cell into two identical daughter cells (Figure 4.8). This process is called binary fission (“binary” to indicate that two cells have arisen from one).
Figure 4.8 Binary fission in a rod-shaped bacterium.

Cell numbers (and all components of the cells) double every generation.
The partition that forms between dividing cells is called a septum and results from the inward growth of the cell envelope from opposing directions; septum formation continues until the two daughter cells are pinched off. There are some variations in this general pattern of binary fission. In some bacteria, such as Bacillus subtilis, a septum forms without cell wall constriction (Figure 4.9), while in the budding bacterium Caulobacter (Figure 2.14a, and see Figure 4.18) constriction occurs but no septum is formed.
Figure 4.9 Septa.

The septum that separates dividing cells of the bacterium Bacillus subtilis is clearly visible in this series of fluorescent micrographs. (a) DAPI stains the entire cell. (b) The green fluorescent protein lights up entire cells. (c) A dye that stains only the cytoplasmic membrane shows that septa contain membrane (and cell wall) material.
When one cell eventually separates to form two cells (Figure 4.8), we say that one generation has occurred, and the time required for this process is called the generation time (or doubling time). Each daughter cell is essentially identical, having received a copy of the chromosome(s) and sufficient copies of all cellular materials required to begin life as an independent entity. Microbes differ in their doubling times, and the doubling time of any given microbe varies depending on growth conditions. Under optimal growth conditions, the generation time of E. coli is about 20 min. The fastest-growing microbes known can double in less than 10 minutes while the slowest can have generation times of several months or even longer.
The Microbial Growth Cycle
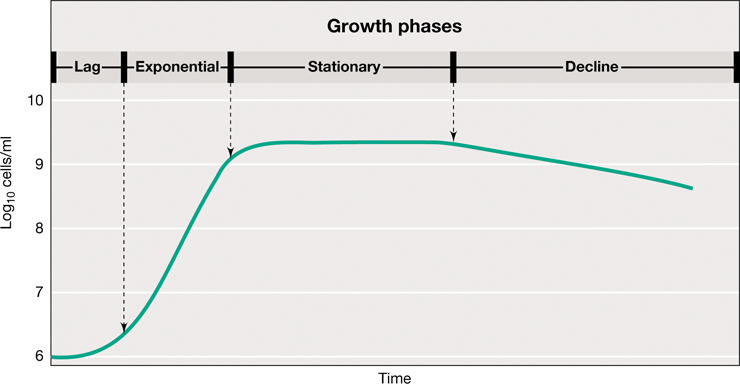
Under optimal conditions, growth by binary fission can cause a rapid increase in the number of cells present, but conditions are rarely optimal, even in laboratory culture. Typically, microbes are grown in batch culture, which describes the growth of microbes in a fixed volume of liquid enclosed within a container such as a test tube or a flask. The nutrients in such a culture flask are finite and cannot support growth indefinitely. Hence, in batch culture microbes typically exhibit a growth cycle called the microbial growth curve (Figure 4.10). The growth curve is composed of four phases: lag phase, exponential phase, stationary phase, and decline phase.
Figure 4.10 Typical growth curve for a bacterial population.

A viable count measures the cells in the culture that are capable of reproducing. Optical density (turbidity), a quantitative measure of light scattering by a liquid culture (see Figure 4.7), increases with the increase in cell number.
Lag and Exponential Phases
When a microbial culture is inoculated into fresh growth media (Section 4.2), there is typically an initial pause during which the cells do not grow. This period between inoculation and the onset of growth is called the lag phase (Figure 4.10). Microbes exhibit a lag phase because transfer to fresh medium represents new environmental conditions for the cells, which need to alter their metabolic state to respond to these new conditions.
The lag phase may be brief or extended depending on the previous history of the cells (old culture, young culture, type of growth medium, and so on), the choice of growth medium, and the growth conditions. For example, if an actively growing culture is transferred into a new flask containing the same type of medium under the same growth conditions, there will be very little lag. This is because the cell needs to make very few metabolic adjustments to this new condition. However, if the inoculum is from an old culture, there is usually a longer lag because cell viability may be low, or the cells may need to synthesize many enzymes and macromolecules before they can begin growing. A long lag is also observed when a microbial culture is transferred from a nutrient-rich to a nutrient-poor culture medium. In order to grow, cells must have a complete set of enzymes to biosynthesize the essential metabolites absent from the nutrient-poor medium. The more enzymes and molecules that a cell must manufacture in order to grow in a new environment, the longer the lag.
The exponential phase of growth is the period when the growing cell population doubles at regular intervals (Figure 4.10). This phase is also called balanced growth because cells are as close to being metabolically identical throughout this period as they can be. Hence, in order to minimize experimental variation, most experiments are performed with microbes taken from cultures in the exponential phase of growth. Rates of exponential growth vary greatly and are influenced by choice of media, growth conditions, and the organism itself. Exponential growth continues until conditions in the batch culture can no longer sustain growth.
Stationary and Death Phases
In a batch culture, exponential growth cannot continue forever. Consider the fact that a single cell weighing one-trillionth (10−12) of a gram that doubled unabated every 20 min for 48 hours would produce a population of cells that weighed 4000 times more than planet Earth. Obviously, this is impossible because the carrying capacity of an environment will always limit growth of the organisms within it. In the case of batch cultures, growth is limited because of either nutrient depletion or the accumulation of microbial waste products. When exponential growth ceases for one (or both) of these reasons, the population enters stationary phase (Figure 4.10).
In stationary phase, there is no net increase or decrease in cell number and thus the growth rate of the population is zero. During stationary phase, cellular metabolism shifts away from growth as the cell prepares for maintenance and survival. Despite growth arrest, energy metabolism and biosynthetic processes in stationary phase cells may continue, but typically at a greatly reduced rate. Eventually, the population will enter the decline phase of the growth cycle as the total number of cells decreases due to cell death. Cell division may still occur for some cells in the population during the stationary and decline phases, but this meager increase in number is balanced by the death of other cells. Some microbial cultures exhibit cryptic growth during stationary and decline phases as subpopulations of cells adapt to cannibalize and reuse resources released from dying cells. Also, if a culture is prevented from drying, some number of cells may remain for months or even years.
We now consider how exponential growth can be put on a quantitative footing using some basic mathematical relationships and how these relationships can be exploited in laboratory studies of growing microbial cultures.
Check Your Understanding
Under what conditions would a lag phase not occur?
Define the term generation. What is meant by the term generation time?
4.7 Quantitative Aspects of Microbial Growth
During cell division, one cell becomes two. During that doubling time (the generation time), both the total cell number and cell mass double (Figure 4.8). As we will see, cell numbers in a bacterial culture can quickly become very large, and so it is necessary to deal with the topic of microbial growth using quantitative methods.
Plotting Growth Data
Exponential growth of a population occurs during a period when cell number doubles at regular intervals (defined as the doubling or generation time, Section 4.6). For example, consider a single cell that initiates exponential growth with a generation time of 30 minutes (Figure 4.11). When we plot the change in cell number over time on arithmetic (linear) coordinates, we obtain an upwardly sweeping curve with a continuously increasing slope (Figure 4.11b). By contrast, when the cell number is plotted on a logarithmic (log10) scale as a function of time (a semilogarithmic graph) the points fall on a straight line (Figure 4.11b). This straight-line function reflects the fact that the cells are growing exponentially and that the population is doubling in a constant time interval.
Figure 4.11 The rate of growth of a microbial culture.

(a) Data for a cell population that doubles every 30 min. (b) Data plotted on arithmetic (left ordinate) and logarithmic (right ordinate) scales.
Semilogarithmic graphs are also convenient for estimating the generation time of a culture from actual growth data, since generation times can be inferred directly from the graph as shown in Figure 4.12. For example, the generation time can be determined by finding the time it takes for the number of cells to double (Figure 4.12). In a more formal sense, we define generation time (g) as the time (t) per generation (n), which is g=t/n. Hence, a population that takes six hours (t=6 h) to double once (n=1) has a generation time of 6 h (Figure 4.12a). Several of the terms and expressions useful in quantifying microbial growth are defined in Figure 4.12c.
Figure 4.12 Calculating microbial growth parameters.

Method of estimating the generation times (g) of exponentially growing populations with g of (a) 6 h and (b) 2 h from data plotted on semilogarithmic graphs. The slope of each line is equal to 0.301/g, and n is the number of generations in the time t. All numbers are expressed in scientific notation; that is, 10,000,000 is 1×107, 60,000,000 is 6×107, and so on. (c) Useful expressions for analyzing microbial growth.
The Mathematics of Bacterial Growth
Starting with a single cell and considering the fact that the cell number of an exponentially growing culture doubles every generation, the number of cells present at generation n can be expressed as 2n. For example, after 3 generations there will be 23=8 cells, after 10 generations there will be 210=1024 cells, and so on (Figure 4.11a). If exponential growth began with two cells instead of one, the number of cells at any generation would be doubled (for example, 2×210=2048 cells), and if we started with three cells, the number of cells at any generation would be tripled (3×210=3072 cells). Hence, it is possible to calculate the number of cells (N) in an exponentially growing culture at any point in time from the following expression: Nt=N02n(1)
where Nt is cell number at time t, N0 is the initial cell number (the number at time 0), and n is the number of generations during the period of exponential growth.
The equation Nt=N02n can be solved for n by taking the logarithms of both sides and using algebra to yield n=[(log Nt−logN0)/log 2](2)
This equation makes it possible to calculate generation times in terms of measurable quantities, Nt and N0. Values for Nt and N0 can be determined from the viable or microscopic counting methods described earlier; alternatively, OD readings from a spectrophotometer can be used for these calculations (Sections 4.3, 4.4 and 4.5 describe methods for quantifying cell numbers). As an example, consider actual growth data from the plot in Figure 4.12b, in which Nt=108, N0=5×107, and t=2 h: n=[(log Nt−log N0)/log 2]n=[log(108)−log(5×107)]/log 2n=(8−7.69)/log 2=(0.301)/0.301=1
Thus, in this example n=1 and so we can use the equation g=t/n to determine that g=2 h. If exponential growth continued for another 2 h, the cell number would be 2×108. Two hours later, the cell number would be 4×108, and so on.
The value of g can also be calculated by evaluating the slope of the line on a semilogarithmic plot of exponential growth. On such a plot (Figure 4.12b) the slope is defined as [(log Nt−log N0)/t]. Combining this with expression (2) and the understanding that g=t/n, the slope of the curve reduces to [log 2/g], or simply, 0.301/g. In the example shown in Figure 4.12b, the slope is 0.15 h−1, and since g=0.301/slope, the generation time is [0.301/0.15 h−1]=2 h.
Specific Growth Rate
Other expressions are often useful in describing exponential growth, and chief among these is the specific growth rate (k). The specific growth rate expresses the rate at which the population is growing at any instant (by contrast, g is the mean time required for the cell population to double); k is expressed in units of reciprocal hours (h−1).
The specific growth rate is a function of the change in cell number (dN/dt) over time and is expressed as dN/dt=kN, where the specific growth rate (k) indicates the rate at which the cell number (N) is increasing at any point in time. Integration of this equation using natural logarithms (loge), gives the expression Nt=N0ekt. To estimate k, the log10 of both sides of this equation is taken to obtain the expression log Nt=kt/2.303+log N0 (log10 is used such that N can be plotted against t in a semilog plot, Figure 4.12). From this equation, [(log Nt−log N0)/t] reduces to (k/2.303); that is, the slope of the line on a semilogarithmic plot of growth can be calculated as k/2.303. Also, since 0.301/g=k/2.303, k can be expressed directly as 0.693/g.
Armed with knowledge of n and t, one can calculate g and k for different microorganisms growing under various conditions (Figure 4.12). This is often useful for optimizing culture conditions for a newly isolated organism and for testing the positive or negative effect of some treatment on a bacterial culture. For example, comparison with an unamended control allows factors that stimulate or inhibit growth to be identified by measuring their effect on the various growth parameters presented here.
Consequences of Exponential Growth
During exponential growth, the increase in cell number is initially rather slow but increases at an ever-faster rate. In the later stages of exponential growth, this results in an explosive increase in cell numbers. For example, in the experiment shown in Figure 4.11, the rate of cell production in the first 30 min of growth is 1 cell per 30 min. However, between 4 and 4.5 h of growth, the rate of cell production is 256 cells per 30 min, and between 5.5 and 6 h of growth it is 2048 cells per 30 min. Because of this, cell numbers in laboratory cultures of bacteria can quickly become very large, and population sizes of >109 cells/ml are common.
Besides being a theoretical consideration, exponential growth can have implications in everyday life. Consider something we have all experienced, the spoilage of milk. The lactic acid bacteria responsible for the soured flavor of spoiled milk contaminate the milk during its collection and exist in fresh, pasteurized milk in low numbers; these organisms grow slowly at refrigerator temperature (4 °C) but much faster at room temperature. If a bottle of fresh milk stands at room temperature overnight, some lactic acid is made, but not enough to affect milk quality. However, if week-old milk, which now contains a week’s worth of slow bacterial growth (and thus much higher cell numbers), is left standing under the same conditions, a huge amount of lactic acid is made, and spoilage results.
Check Your Understanding
What is a semilogarithmic plot and what information can we derive from it?
For an exponentially growing culture that increases from 5×106 cells/ml to 5×108 cells/ml in 8 h, calculate g, n, and k.
For testing a bacterium’s response to an inhibitory substance, why would g be useful information?
4.8 Continuous Culture
Up to this point our consideration of microbial population growth has been confined to batch cultures. The environment in a batch culture is constantly changing because of nutrient consumption and waste production. These limitations can be circumvented in a continuous culture device. The most common type of continuous culture device is the chemostat (Figure 4.13). Unlike a batch culture, which is a closed system, a continuous culture is an open system. In a chemostat, a known volume of sterile medium is added at a constant rate while an equal volume of spent culture medium (which also contains cells) is removed at the same rate (Figure 4.13). Once in equilibrium, the culture volume, cell number, and nutrient/waste product status remain constant, and the culture attains steady state.
Figure 4.13 Continuous culture device (chemostat).
The population density is controlled by the concentration of a limiting nutrient in the reservoir, and the growth rate is controlled by the dilution rate; both parameters are set by the experimenter. (a) Chemostat components. (b) Photo of a chemostat setup.
The Chemostat and the Concept of Steady State
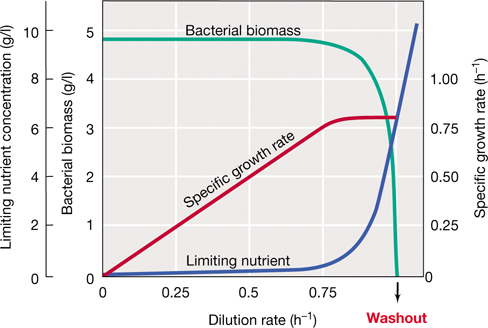
A chemostat enables control over both the specific growth rate (k, Figure 4.14) and growth yield (biomass per ml, Figure 4.15) of a microbial culture. In a chemostat, fresh sterile medium is added to a culture vessel and spent media washed out at equal rates, resulting in a culture that maintains a fixed volume (Figures 4.13 and 4.14). The supply of medium is defined by the dilution rate (D) which is expressed as F/V, where F is the flow rate (volume per unit time), and V is the culture volume. In the chemostat, the specific growth rate is controlled by the dilution rate (D), and the growth yield is controlled by the concentration of a limiting nutrient in the fresh medium added to the vessel.
Figure 4.14 Steady-state relationships in the chemostat.

The dilution rate is determined from the flow rate and the volume of the culture vessel. Note that at a high dilution rate, growth cannot balance dilution, and the population washes out. Note also that although the population density (bacterial biomass) remains constant and the concentration of the growth-limiting nutrient remains near zero during steady state, the specific growth rate can vary over a wide range.
Figure 4.15 The effect of nutrients on growth in a chemostat.

Relationship between nutrient concentration, growth rate, and growth yield (bacterial biomass) in a chemostat culture. Only at low nutrient concentrations are both growth rate and growth yield affected.
In a chemostat at steady state, cells grow at the same rate that they are removed by outflow from the system; that is, k=D (Figure 4.14). At steady state, a large number of cells are all competing for a limiting nutrient. The nutrient added in the fresh medium is consumed rapidly by the cells within the chemostat, thereby limiting their growth rate. As the flow of nutrients increases, the cells can grow faster and they continue to keep pace with the dilution rate until it is so fast that it exceeds the maximal growth rate of the organism. At this point, the cells can no longer keep up and they are washed out of the culture vessel (Figure 4.14).
While the specific growth rate is controlled by D, the growth yield is controlled by the concentration of the limiting nutrient in the medium (Figure 4.15). As shown in Figure 4.14, a change in D does not change growth yield in terms of biomass present until washout occurs. However, more nutrients in the medium mean that more cells can be made (Figure 4.15) but has no impact on the growth rate (except at very low nutrient concentrations when cells are starving). Thus, by varying D or the concentration of the growth-limiting nutrient, one can establish cultures that are growing exponentially at a defined growth rate and a defined cell density.
Experimental Uses of the Chemostat
A practical advantage of the chemostat is that a cell population can be maintained in the exponential growth phase for long periods—weeks or even months. Exponential phase cells are usually most desirable for physiological experiments. Such cells are available at any time in the chemostat, and the vessel can be sampled repeatedly. Chemostat cultures have been used extensively to study bacterial physiology and have also been used in studies of microbial ecology and evolution. For example, because the chemostat can mimic the low substrate concentrations often found in nature, it is possible to ask which organisms in mixed cultures of known composition compete best at various specific growth rates or when particular nutrients are limiting. Chemostats have also been used for the direct enrichment and isolation of bacteria from nature. From a natural sample, one can select a stable population under the chosen conditions of nutrient concentration and D and then slowly increase D until a single organism remains. In this way, microbiologists studying the growth rates of various soil bacteria isolated a bacterium with a 6-min doubling time—the fastest-growing bacterium known!
Check Your Understanding
How do microbial populations in a chemostat differ from microorganisms in a batch culture?
What happens in a chemostat if the dilution rate exceeds the maximal specific growth rate of the organism?
4.9 Biofilm Growth
Thus far we have considered microbial growth in liquid suspensions only. Such growth of free-floating or free-swimming cells is called planktonic growth. Planktonic growth is a natural state for many microbes, and most laboratory cultures are maintained in this state. However, sessile growth, meaning growth while attached to a surface, is quite common in the microbial world. Microbes that are attached to surfaces often develop into biofilms. Biofilm growth has tremendous importance in medical and industrial applications, and it allows microbes to exhibit different properties from those of planktonic cells.
Biofilm Formation
A biofilm is a population of cells enmeshed in a polysaccharide matrix that is attached to a surface (Figure 4.16). Biofilms form in stages, beginning with the attachment of planktonic cells to a surface. Attachment is often mediated by flagella, fimbriae, or pili (Chapter 2). Colonization of the surface starts when microbes begin to grow and produce sticky extracellular polysaccharides (EPS). Microbial colonization and growth on the surface causes changes in the biofilm that lead to development. During development, cells in the biofilm begin to change their metabolism. These changes can cause biofilms to develop complex systems of mushroomlike columns and channels that trigger metabolic differentiation of microbes at the surface of the biofilm from those at its base. Finally, dispersal of cells from a mature biofilm allows microbes to colonize new sites (Figure 4.16). Dispersal is usually prompted by changes in the environment, such as nutrient limitation or other forms of stress.
Figure 4.16 ***Pseudomonas aeruginosa*** biofilm development.

Confocal scanning laser micrographs of a developing Pseudomonas aeruginosa biofilm in a flow cell continuously irrigated with nutrient-rich medium. P. aeruginosa cells first attach to the glass surface (day 0), then rapidly grow and move on the surface to cover the entire surface (day 1); by day 4 mushroom-shaped microcolonies over 0.1 mm high have developed.
Biofilms can be studied in a flow chamber, in which liquid media flows continuously between two layers of glass. Flow chambers are designed so that biofilm growth can be monitored microscopically over time (Figure 4.16). Pseudomonas aeruginosa is a model organism that has been used to study biofilms. P. aeruginosa is a relevant model because it forms biofilms in the lungs of humans with the genetic disease cystic fibrosis. Biofilm formation allows cells of P. aeruginosa to persist and resist removal. Biofilms also tend to make bacteria more resistant to drugs because they provide a penetration barrier and promote metabolic differentiation. Hence, biofilms allow complex microbial interactions not observed in planktonic microbes.
Biofilms and Humans
Biofilms are a common growth form for bacteria in nature because the intensely interwoven nature of the structure prevents harmful chemicals from reaching cells deep within the biofilm structure. Biofilms also provide a physical barrier that protects cells from grazing by protists and allows them to remain in a safe and favorable habitat. Some biofilms form multilayered sheets with different organisms present in the individual layers, and these biofilms are called microbial mats. Mats composed of various phototrophic and chemotrophic bacteria are common in the outflows of hot springs (**Figure 4.17*a***) and marine intertidal regions and can form crusty matlike structures quickly in puddles of water that stay moist for as little as a few days.
Figure 4.17 Examples of biofilms.

(a) A microbial mat of the purple phototrophic bacterium Thermochromatium tepidum that developed in a small sulfidic hot spring in Yellowstone National Park (USA). (b) Scanning electron micrograph of a biofilm of cells of Staphylococcus aureus that formed on an indwelling catheter.
Biofilms affect many aspects of our lives, including human health. For example, biofilms have been implicated in difficult-to-treat infections of human joints, implanted medical devices such as artificial heart valves and joints, and indwelling devices, such as catheters (Figure 4.17b). Biofilms are also responsible for the formation of dental caries (cavities) and are a cause of gum disease. In industrial and municipal settings, biofilms can cause fouling, plugging, and corrosion in pipes used to transport liquids. Biofilms can even form in fuel storage tanks, where they contaminate fuel by producing corrosive chemicals such as H2S. Finally, the formation of biofilms on the hulls of ships is a major problem because they decrease speed and energy efficiency and thereby increase shipping times and shipping costs.
Biofilms are of tremendous significance and so we will revisit different aspects of the biology of biofilms in Chapters 7 and 20.
Check Your Understanding
What are the stages and events that occur during biofilm formation?
Why is biofilm formation such a major problem in human medicine?
4.10 Alternatives to Binary Fission
Planktonic cells that grow by binary fission undergo balanced growth during exponential phase. All cells in such a culture are nearly identical, genetically and metabolically, and the uniformity of such cultures makes them desirable as experimental systems. We have already learned that cells within biofilms do not exhibit balanced growth because their metabolic characteristics and growth rates vary widely depending on their position within the biofilm. Here we consider microbes that do not display binary fission and whose growth dynamics are not well explained by standard growth equations.
Budding Cell Division
Although binary fission, which produces identical daughter cells, is common among Bacteria and Archaea, many bacteria grow by other means. Budding bacteria, for example, exhibit unequal cell growth and produce daughter cells that have different characteristics (Figure 4.18). In budding division a new cell emerges and buds off from a mother cell, the latter of which retains its original identity (Figure 4.18).
Figure 4.18 Cell division in different morphological forms of bacteria.

The contrast is shown between cell division in conventional bacteria (cells that divide by binary fission) and in various budding and stalked bacteria.
Some budding bacteria form cytoplasmic extensions such as stalks or hyphae, and classic examples are the genera Caulobacter (Figure 2.14) and Hyphomicrobium (Figure 4.18). Hyphomicrobium forms a long stalk from which a new cell emerges and buds. In the case of Caulobacter, the mother cell is attached to a surface by its stalk. The daughter cell that buds off is motile; it lacks a stalk and has a flagellum that allows it to disperse by swimming motility. Other budding bacteria such as the aquatic bacterium Ancalomicrobium produce multiple appendages that resemble arms extending away from the cell (Figure 15.53b). The appendages increase the surface-to-volume ratio of the cell (Section 1.3), which increases its ability to extract nutrients from oligotrophic (very dilute) habitats. Many budding bacteria also have distinctive life cycles, and we consider these and the group as a whole in Section 15.18.
Hyphal Growth and Multiple Fission
Actinomycetes are gram-positive filamentous bacteria that are common in soils and grow as long thin filaments called hyphae; species of the genus Streptomyces are typical actinomycetes (Figure 4.19). Hyphal growth occurs only at the tip of an elongating filament and is unlike binary fission because growth of the cell is not linked directly to cell division. That is, cells elongate and replicate DNA as they grow but do not produce septa. Instead, hyphae form cross-walls away from the point of cell growth. These cross-walls do not define independent cells but instead allow transport to occur between adjacent compartments in the hyphal filament. Hyphae often weave together to form mycelia, which form from complex hyphal filaments, and mature mycelia often form arthrospores. Arthrospores are survival structures but differ from endospores in their mechanism of development and their lack of resistance to heat and harsh chemicals.
Figure 4.19 Colorized scanning electron micrograph of cells of ***Streptomyces***, a filamentous bacterium that produces arthrospores.

Various species of Streptomyces are major producers of antibiotics.
Arthrospores develop by multiple fission, in which a single hyphal filament forms many septa simultaneously along its length. This causes many cells to form all at once along the filament, each of which ultimately differentiates into a mature arthrospore. Multiple fission is also seen in certain cyanobacteria (a defining feature of the Pleurocapsales; see Figure 15.2b). Cells of the cyanobacterium Stanieria, for example, begin about 1 μm in diameter and grow to as large as 30 μm in diameter before undergoing multiple fission, a process in which the large cell subdivides into tens (or even hundreds) of smaller cells, each of which begins the replication cycle anew. Finally, some bacteria even form intracellular offspring. Bacteria such as the giant-celled Epulopiscium (Figure 1.6a) grow by forming multiple daughter cells within the cytoplasm of the mother cell. Once the daughter cells are mature, they burst out of the mother cell and begin a new round of daughter cell production.
Now we turn our attention from microbial growth itself to environmental characteristics that control microbial growth. We will see that the microbial world is ripe with exceptional organisms able to grow under punishing conditions that can only be labeled extreme.
Check Your Understanding
Why would it be incorrect to apply the growth equations in Figure 4.12 to an organism like Stanieria?
In a solid environment such as soil, what advantage might hyphal growth provide to a bacterium relative to those that grow by binary fission?
Compare and contrast the life cycle of Caulobacter to the development of a biofilm.
III Environmental Effects on Growth: Temperature
Having the right set of chemical nutrients in a culture medium is only half the secret of growing microbial cells; the other half is to ensure that the environment is suitable for growth. Temperature is a key factor, but what’s warm for one organism may be cold for another.
Even when a microbe is provided with an optimal array of its required nutrients, growth is not a sure thing unless the chemical and physical state of its environment is also suitable. Four environmental factors control microbial growth in a major way: temperature, pH, water availability, and oxygen. If any one of these factors is beyond the limits that an organism can tolerate, growth will not occur, even in an otherwise ideal culture medium. In Parts III and IV of this chapter we examine these important environmental factors, beginning with temperature, the key factor affecting the growth and survival of microorganisms.
4.11 Temperature Classes of Microorganisms
At either too cold or too hot a temperature, microorganisms will not be able to grow and may even die. The minimum and maximum temperatures supporting growth vary greatly among different organisms and usually reflect the temperature range and average temperature of the environments the organisms inhabit.
Cardinal Temperatures
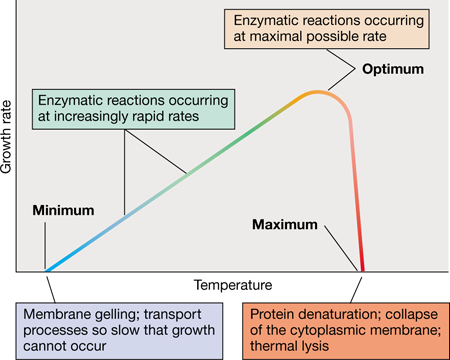
Temperature affects microorganisms in two opposing ways. As temperatures rise, the rate of enzymatic reactions increases and growth becomes faster. However, above a certain temperature, proteins or other critical cell components may be denatured or otherwise irreversibly damaged. For every microorganism there is a minimum temperature below which growth is not possible, an optimum temperature at which growth is most rapid, and a maximum temperature above which growth is not possible. These three temperatures, called the cardinal temperatures (Figure 4.20), are characteristic of any given microorganism and can differ dramatically between different species. For example, some organisms have growth temperature optima near 0 °C, whereas the optima for others can be higher than 100 °C. The temperature range throughout which microbial growth is possible is even wider than this, from as low as −15 °C to at least 122 °C. However, no single organism can grow over this whole temperature range, as the range for any given organism is typically less than 40 °C.
Mastering Microbiology
Art Activity: Figure 4.20 The cardinal temperatures: minimum, optimum, and maximum
Figure 4.20 The cardinal temperatures: minimum, optimum, and maximum.

The actual values may vary greatly for different organisms (see Figure 4.21).
The maximum growth temperature of an organism reflects the temperature above which denaturation of one or more essential cell components, such as a key enzyme, occurs. The factors controlling an organism’s minimum growth temperature are not as clear. However, the cytoplasmic membrane must remain in a semifluid state for nutrient transport and bioenergetic functions to take place. That is, if an organism’s cytoplasmic membrane stiffens to the point that it no longer functions properly in transport or can no longer develop or consume a proton motive force, the organism cannot grow. In contrast to the minimum and maximum, the growth temperature optimum reflects a state in which all or most cellular components are functioning at their maximum rate and typically lies closer to the maximum than to the minimum (Figures 4.20 and 4.21).
Temperature Classes of Organisms
Although there is a continuum of organisms, from those with very low temperature optima to those with high temperature optima, it is possible to distinguish four broad classes of microorganisms in relation to their growth temperature optima: psychrophiles, with low temperature optima; mesophiles, with midrange temperature optima; thermophiles, with high temperature optima; and hyperthermophiles, with very high temperature optima (Figure 4.21).
Figure 4.21 Temperature and growth response in different temperature classes of microorganisms.

The temperature optimum of each example organism is shown on the graph.
Mesophiles are widespread in nature and are the most commonly studied microorganisms. Mesophiles are found in the intestines of endothermic (warm-blooded) animals and in terrestrial and aquatic environments in temperate and tropical latitudes. Escherichia coli is a typical mesophile, and its cardinal temperatures have been precisely defined. The optimum temperature for most strains of E. coli is 39 °C, the maximum is 48 °C, and the minimum is 8 °C. Thus, the temperature range for E. coli is about 40 degrees (Figure 4.21).
Psychrophiles and thermophiles are found in unusually cold and unusually hot environments, respectively. Hyperthermophiles are found in extremely hot habitats such as hot springs, where temperatures can be as hot as 100 °C, and deep-sea hydrothermal vents, where temperatures can exceed 100 °C. We now consider these fascinating microbes and examine some of the physiological problems they face and some of the biochemical solutions they have evolved to thrive under these extreme conditions.
Check Your Understanding
How does a hyperthermophile differ from a psychrophile?
What are the cardinal temperatures for Escherichia coli ? To what temperature class does it belong?
E. coli can grow at a higher temperature in a complex medium than in a defined medium. Why?
4.12 Microbial Life in the Cold
Because humans live and work in places where temperatures are moderate, it is natural to consider very hot and very cold environments as “extreme.” However, many microbial habitats are indeed very hot or very cold, and organisms that inhabit these environments—called extremophiles (Section 1.5 and Table 1.2)—actually thrive in these punishing environments. We consider the biology of these fascinating organisms here and in the next section.
Cold Environments
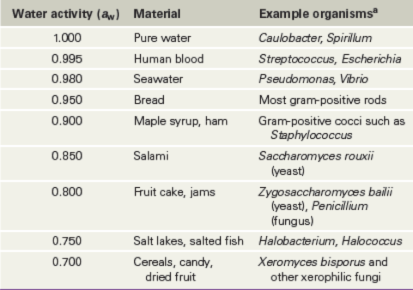
Much of Earth’s surface is cold. The oceans, which make up over half of Earth’s surface, have an average temperature of 5 °C, and the depths of the open oceans have constant temperatures of 1–3 °C. Vast areas of the Arctic and Antarctic are permanently frozen or are unfrozen for only a few weeks in summer (Figure 4.22). These cold environments support diverse microbial life, as do glaciers (**Figure 4.22*e***), where the networks of liquid water channels that run through and under the glacier are teeming with microorganisms. Even in solidly frozen materials there remain small pockets of liquid water where solutes have concentrated and microorganisms can metabolize and grow, albeit very slowly.
Mastering Microbiology
Art Activity: Figure 4.21 Temperature and growth responses in different temperature classes of microorganisms
Figure 4.22 Antarctic microbial habitats and microorganisms.

(a) A core of frozen seawater from McMurdo Sound, Antarctica. The core is about 8 cm wide. Note the dense coloration due to pigmented microorganisms. (b) Phase-contrast micrograph of phototrophic microorganisms from the core shown in part a. Most organisms are either diatoms or green algae (both eukaryotic phototrophs). (c) Transmission electron micrograph of Polaromonas, a gas vesiculate bacterium that lives in sea ice and grows optimally at 4 °C. Cells are about 0.8 μm in diameter. (d) Lake Bonney, McMurdo Dry Valleys, Antarctica. Although the lake is permanently ice-covered, the water column under the ice contains a diverse array of Bacteria, Archaea, and microbial eukaryotes. (e) Garwood Glacier, McMurdo Dry Valleys, Antarctica. The edge of the glacier (arrow) is about 20 m high. Glaciers and subglacial lakes are teeming with microbial life.
In considering cold environments as microbial habitats, it is important to distinguish between environments that are constantly cold and those that are only seasonally cold. The latter, characteristic of temperate climates, may have summer temperatures as high as 40 °C. A temperate lake, for example, may have ice cover in the winter, but the water may remain at 0 °C for only a relatively brief time. By contrast, Antarctic lakes contain a permanent ice cover several meters thick (Figure 4.22d), and the water column below the ice in these lakes remains at 0 °C or colder year-round. Marine sediments and glaciers are also constantly cold, as are subglacial lakes—lakes deep beneath the glacier surface—and all of these are teeming with microbial life. It is thus not surprising that the best examples of microbes well adapted to cold temperatures have emerged from these environments.
Psychrophilic and Psychrotolerant Microorganisms
A psychrophile is a microbe with an optimal growth temperature of 15 °C or lower, a maximum growth temperature below 20 °C, and a minimum growth temperature of 0 °C or lower. By contrast, microbes that grow at 0 °C but have optima of 20–40 °C are called psychrotolerant. Psychrophiles are found in environments that are constantly cold and may even be killed by warming to moderate temperatures. For this reason, the laboratory study of psychrophiles requires that great care be taken to ensure that they never warm up during sampling, transport to the laboratory, isolation, or other manipulations.
Psychrophilic algae and bacteria often grow in dense masses within and under sea ice (frozen seawater that forms seasonally) in polar regions (Figure 4.22a–c). They can also be found on the surfaces of permanent snowfields and glaciers where they impart a distinctive coloration to the surface (Figure 4.23). The snow alga Chlamydomonas nivalis is an example of this, the carotenoid pigment astaxanthin in its spores (Figure 4.23 inset) being responsible for the brilliant red color of the snow surface. This alga grows within the snow as a green-pigmented vegetative cell and then sporulates. As the snow dissipates by melting, erosion, and ablation (evaporation and sublimation), the spores become concentrated on the surface. Related species of snow algae contain different carotenoid pigments, and thus fields of snow algae can also be green, orange, brown, or purple.
Figure 4.23 Snow algae.

Snow bank in the Sierra Nevada, California, with red coloration caused by the presence of snow algae. Inset: photomicrograph of red-pigmented spores of the snow alga Chlamydomonas nivalis. Spores are about 18 μm in diameter. The spores germinate to yield motile green algal cells.
Several psychrophilic Bacteria and a few psychrophilic Archaea have been isolated, and some of these show very low growth temperature optima. The permafrost bacterium Planococcus halocryophilus grows slowly at −15 °C, the lowest growth temperature documented for any bacterium. However, theoretical considerations of bacterial metabolism suggest that the lower temperature limit for bacterial metabolism is likely to be considerably colder than this. For example, microbial respiration (as measured by CO2 production) has been measured in tundra soils at nearly −40 °C. At a temperature of −20 °C, pockets of liquid water can exist in “frozen” materials, and studies have shown that enzymes from cold-active bacteria still function under such conditions. Growth at such temperatures, if possible, would be extremely slow. However, if an organism can grow, even if only at a very low rate, it can remain competitive and maintain a population in its habitat.
Psychrotolerant microorganisms are more widely distributed in nature than are psychrophiles and can be isolated from soils and water in temperate climates as well as from meat, dairy products, cider, vegetables, and fruit stored at standard refrigeration temperatures (4 °C). Although psychrotolerant microorganisms grow at 0 °C, most do not grow well, and one must often wait weeks before visible growth is seen in laboratory cultures. By contrast, the same organism cultured at 30 °C may grow at rates similar to that of many mesophiles. Various Bacteria, Archaea, and microbial eukaryotes are psychrotolerant.
Molecular Adaptations to Life in the Cold
Psychrophiles produce enzymes that function—often optimally—in the cold and that may be denatured or otherwise inactivated at even very moderate temperatures. The molecular basis for this is not entirely understood, but clearly it is linked to protein structure. Several cold-active enzymes whose structure is known show a greater content of α-helix and lesser content of β-sheet secondary structure (Section 6.7) than do enzymes that show little or no activity in the cold. Because β-sheet secondary structures tend to be more rigid than α-helices, the greater α-helix content of cold-active enzymes allows these proteins greater flexibility for catalyzing their reactions at cold temperatures. Cold-active enzymes also tend to have greater polar and lesser hydrophobic amino acid content (Figure 6.27 for structures of amino acids) and lower numbers of weak bonds, such as hydrogen and ionic bonds, compared with the corresponding enzyme from mesophiles. Collectively, these molecular features are likely to keep cold-active enzymes flexible and functional under cold conditions.
Another characteristic feature of psychrophiles is that their cytoplasmic membranes remain functional at low temperatures. Cytoplasmic membranes from psychrophiles tend to have a higher content of unsaturated and shorter-chain fatty acids, and this helps the membrane remain in a semifluid state at low temperatures to carry out important transport and bioenergetic functions. Some psychrophilic bacteria even contain polyunsaturated fatty acids; unlike monounsaturated or fully saturated fatty acids that tend to stiffen at low temperatures, polyunsaturated fatty acids remain flexible even at very cold temperatures.
Other molecular adaptations to cold temperatures include “cold shock” proteins and cryoprotectants, and these are not limited to psychrophiles. Cold shock proteins are a type of molecular chaperone (Section 6.11) and have several functions that include maintaining cold-sensitive proteins in an active form or binding specific mRNAs and facilitating their translation under cold conditions. Cryoprotectants include dedicated antifreeze proteins or specific solutes—such as glycerol or certain sugars—that are produced in large amounts at cold temperatures; these agents help prevent the formation of ice crystals that can puncture the cytoplasmic membrane. Highly psychrophilic bacteria often produce abundant levels of exopolysaccharide cell surface slime, and these slime layers confer cryoprotection as well.
Although “freezing” temperatures may prevent microbial growth, they do not necessarily cause death. Indeed, just the opposite may occur, and this has been exploited for the preservation of bacterial cells in microbial culture collections. Cells suspended in growth medium containing 10% dimethyl sulfoxide (DMSO) or glycerol as a cryoprotectant and frozen at −80 °C (ultracold freezer) or −196 °C (liquid nitrogen) remain viable in the frozen state for years.
Check Your Understanding
How do psychrotolerant organisms differ from psychrophilic organisms?
What molecular adaptations to cold temperatures are seen in the cytoplasmic membrane of psychrophiles? Why are they necessary?
4.13 Microbial Life at High Temperatures
Microbial life flourishes in high-temperature environments, from sun-heated soils and pools of water to boiling hot springs, and the organisms that live in these environments are typically highly adapted to their environmental temperature. We examine these organisms now and pick up on them again in several later chapters.
Thermal Environments
Organisms whose growth temperature optimum exceeds 45 °C are called thermophiles and those whose optimum exceeds 80 °C are called hyperthermophiles (Figure 4.21). The surface of soils subject to full sunlight can be heated to above 50 °C at midday, and some surface soils may warm to as high as 70 °C. Fermenting materials such as compost piles and silage can also reach temperatures of 70 °C. Thermophiles abound in such environments. The most extreme high-temperature environments in nature, however, are hot springs, and these are home to a huge diversity of thermophiles and hyperthermophiles.
Many terrestrial hot springs have temperatures at or near boiling, while those at the bottom of the ocean, called hydrothermal vents, can have temperatures of 350 °C or greater. Hot springs are found throughout the world, but they are especially abundant in the western United States, New Zealand, Iceland, Japan, Italy, Indonesia, Central America, and central Africa. The largest concentration of hot springs in the world is in Yellowstone National Park, Wyoming (USA). Although some hot springs vary widely in temperature, many have nearly constant high temperatures, varying less than a degree or two over many years. In addition, different springs have different chemical compositions and pH values. In habitats hotter than 65 °C, only prokaryotic cells can thrive (Table 4.3), but the diversity of Bacteria and Archaea in such environments is often extensive.
Table 4.3 Presently known upper temperature limits for growth of living organisms

Hyperthermophiles and Thermophiles
A variety of hyperthermophiles inhabit boiling hot springs (Figure 4.24), including both chemoorganotrophic and chemolithotrophic species. Growth rates of hyperthermophiles can be studied in the field by immersing a microscope slide into a spring and then examining it microscopically over time. The slide is an excellent surface for microbial attachment and subsequent growth, and so small microbial colonies form (Figure 4.24b) and growth rates can be calculated from cell number data. Ecological studies such as this have shown that growth rates in boiling springs are often quite high, with generation times (g) as short as 1 h not uncommon.
Figure 4.24 Growth of hyperthermophiles in boiling water.

(a) Boulder Spring, a small boiling spring in Yellowstone National Park. This spring is superheated, having a temperature 1−2 °C above the boiling point. The mineral deposits around the spring consist mainly of silica and sulfur. (b) Photomicrograph of a microcolony of Archaea that developed on a microscope slide immersed in such a boiling spring.
Cultures of diverse hyperthermophiles have been obtained, and a variety of morphological and physiological types of both Bacteria and Archaea are known. Some hyperthermophilic Archaea have growth-temperature optima above 100 °C, while no species of Bacteria have yet been discovered that grow above 95 °C. Growing laboratory cultures of organisms with optima above the boiling point requires pressurized vessels that permit temperatures in the growth medium to rise above 100 °C without boiling. The most heat-tolerant organisms known inhabit hydrothermal vents, with the most thermophilic example thus far being Methanopyrus, a methane-producing genus of Archaea capable of growth at up to 122 °C (Section 17.12).
In contrast to hyperthermophiles, thermophiles (optima 45–80 °C) inhabit moderately hot or intermittently hot environments. As boiling water leaves a hot spring, it gradually cools, setting up a thermal gradient. Along this gradient, microorganisms become established, with different species growing in the different temperature ranges (**Figure 4.25*a***). By studying the species distribution along such natural thermal gradients, it has been possible to determine the upper temperature limits for various classes of microbes (Table 4.3). Thermophilic Bacteria and Archaea have also been found in artificial thermal environments, such as hot water heaters. Hot water discharges from power plants and other artificial thermal sources also provide sites where thermophiles can flourish.
Figure 4.25 Hot spring microbes and their heat-stable enzymes.

(a) Characteristic V-shaped pattern (shown by the dashed white lines) formed by cyanobacteria at the upper temperature for phototrophic life, 70–73 °C, in the thermal gradient formed from a boiling hot spring in Yellowstone National Park. The pattern develops because the water cools more rapidly at the edges than in the center of the channel. (b) In the spring source, hyperthermophiles thrive, and some have been used as sources of heat-stable enzymes, such as pullulanase from Pyrococcus (Archaea). Ca2+ stabilizes the enzyme above the boiling point.
Protein and Membrane Stability at High Temperatures
How do thermophiles and hyperthermophiles survive high temperatures? First, their enzymes and other proteins are much more heat-stable than are those of mesophiles and actually function optimally at high temperatures. The heat stability of an enzyme from a hyperthermophile is often due to subtle changes in amino acid sequence from the corresponding enzyme of a mesophile, and these changes affect protein structure and function to resist heat denaturation. Heat-stable proteins also typically show increased ionic bonding between basic and acidic amino acids and have highly hydrophobic interiors, factors that also prevent unfolding. Finally, solutes such as di-inositol phosphate, diglycerol phosphate, and mannosylglycerate are produced at high levels in certain hyperthermophiles, and these are thought to help stabilize their proteins against thermal denaturation.
Enzymes from thermophiles and hyperthermophiles have significant commercial uses. Heat-stable enzymes catalyze biochemical reactions at high temperatures and are in general more stable than enzymes from mesophiles, thus prolonging the shelf life of commercial enzyme preparations (Figure 4.25b). A classic example of this is the DNA polymerase isolated from Thermus aquaticus—Taq polymerase—used to automate the repetitive steps in the polymerase chain reaction (PCR), a technique for amplifying DNA and a major tool of modern biology (Section 12.1). Several other heat-stable enzymes are commercially available for specific industrial applications (Figure 4.25b).
Besides enzymes and other macromolecules in the cell, the cytoplasmic membranes of thermophiles and hyperthermophiles must be heat-stable. Heat naturally works to peel apart the lipid bilayer that makes up the cytoplasmic membrane (Section 2.1). In thermophiles and most hyperthermophilic Bacteria, the cytoplasmic membrane has a higher content of long-chain and saturated fatty acids and a lower content of unsaturated fatty acids than are found in the cytoplasmic membranes of mesophiles. Saturated fatty acids form a stronger hydrophobic environment than do unsaturated fatty acids, and longer-chain fatty acids have a higher melting point than shorter-chain fatty acids; collectively, these properties increase membrane stability.
Hyperthermophiles, most of which are Archaea, do not contain fatty acids in their membranes but instead have C40 hydrocarbons composed of repeating units of isoprene bonded by ether linkage to glycerol phosphate (Figure 2.3). In addition, however, the architecture of the cytoplasmic membranes of many hyperthermophiles takes a unique twist: The membrane forms a lipid monolayer rather than a lipid bilayer (Figure 2.3c). The monolayer structure covalently links both halves of the membrane and prevents it from melting at the high growth temperatures of hyperthermophiles.We consider other aspects of heat stability in hyperthermophiles, including that of DNA stability, in Chapter 17.
Check Your Understanding
Which phylogenetic domain includes species with optima of >100 °C? What special techniques are required to culture them?
How does the membrane structure of hyperthermophilic Archaea differ from that of Escherichia coli and why is this structure helpful for growth at high temperature?
IV: Environmental Effects on Growth: pH, Osmolarity, and Oxygen
IV: Environmental Effects on Growth: pH, Osmolarity, and Oxygen
IV Environmental Effects on Growth: pH, Osmolarity, and Oxygen
Although temperature has an overarching effect on microbial growth, pH, salinity, and atmospheric conditions also control growth of laboratory cultures and natural populations. The best conditions for one organism may differ considerably from those for another.
As we have seen, temperature has a major effect on the growth of microorganisms. But many other environmental factors can affect microbial growth as well, including pH, osmolarity, and oxygen.
4.14 Effects of pH on Microbial Growth
Acidity or alkalinity of a solution is expressed by its pH on a logarithmic scale in which neutrality is pH 7 (Figure 4.26). pH values less than 7 are acidic and those greater than 7 are alkaline. In analogy to a temperature range (Figure 4.21), every microorganism has a pH range, typically about 2–3 pH units, within which growth is possible. Also, each organism shows a well-defined pH optimum, where growth occurs best. Most natural environments have a pH between 3 and 9, and organisms with pH growth optima in this range are most common. Terms used to describe organisms that grow best in particular pH ranges are shown in Table 4.4.
Table 4.4 Relationships of microorganisms to pH

aPicrophilus and Natronobacterium are Archaea; all others are Bacteria.
Figure 4.26 The pH scale.

Although some microorganisms can live at very low or very high pH, the cell’s internal pH remains near neutrality.
Acidophiles
Organisms that grow optimally at a pH value in the range termed circumneutral (pH 5.5 to 7.9) are called neutrophiles. For example, the bacterium Escherichia coli is a neutrophile (Table 4.4). By contrast, organisms that grow best below pH 5.5 are called acidophiles. There are different classes of acidophiles, some growing best at moderately acidic pH and others at very low pH. Many fungi and bacteria grow best at pH 5 or even below, while a more restricted number grow best below pH 3. An even more restricted group grow best below pH 2 and those with pH optima below 1 are extremely rare. Most acidophiles cannot grow at pH 7 and many cannot grow at pH values more than two units above their optimum.
A critical factor governing acidophily is the stability of the cytoplasmic membrane. When the pH is raised to neutrality, the cytoplasmic membranes of strongly acidophilic bacteria are destroyed and the cells lyse. This indicates that these organisms are not just acid-tolerant but that high concentrations of protons are actually required for cytoplasmic membrane stability. For example, the most acidophilic microbe known is Picrophilus oshimae, a species of Archaea that grows optimally at pH 0.7 and 60 °C. Above pH 4, cells of P. oshimae spontaneously lyse. As one would predict, P. oshimae inhabits extremely acidic thermal soils associated with volcanic activity.
Alkaliphiles
A few extremophiles have very high pH optima for growth, sometimes as high as pH 10, and some of these can still grow, albeit poorly, at even higher pH. Microorganisms showing pH optima of 8 or higher are called alkaliphiles. Alkaliphilic microorganisms are typically found in highly alkaline habitats, such as soda lakes and high-carbonate soils. The best-studied alkaliphilic bacteria are certain Bacillus species, such as Bacillus firmus. This organism is alkaliphilic but has an unusually broad range for growth, from pH 7.5 to 11. Some extremely alkaliphilic microbes are also halophilic (salt-loving), and most of these are Archaea (Section 17.1). Some phototrophic purple bacteria (Sections 15.4 and 15.5) are also strongly alkaliphilic. Certain alkaliphiles have commercial uses because they excrete hydrolytic enzymes such as proteases and lipases that maintain their activities at alkaline pH. These enzymes are added to laundry detergents to remove protein and fat stains, respectively, from clothing.
Managing membrane bioenergetics is an obvious problem for alkaliphiles. B. firmus uses sodium (Na+) rather than H+ to drive transport reactions and rotate its flagellum; that is, it forms a sodium motive force instead of a proton motive force. Remarkably, however, B. firmus uses a proton motive force to drive ATP synthesis even though the external membrane surface is highly alkaline. Exactly how this happens is unclear, although it is thought that hydrogen ions are in some way kept very near the outer surface of the cytoplasmic membrane such that they cannot spontaneously combine with the abundant hydroxyl ions to form water.
Cytoplasmic pH and Buffers
The optimal pH for growth of an organism refers to the extracellular environment only; the intracellular pH must be maintained at a value consistent with the stability of macromolecules, a range of about 4 pH units from pH 5 to 9. Thus, despite conditions in their habitats, extreme acidophiles and alkaliphiles maintain cytoplasmic pH values nearer to neutrality.
To prevent major shifts in pH during microbial growth in batch cultures, buffers are commonly added to culture media along with the nutrients required for growth. However, any given buffer works over only a relatively narrow pH range. For neutrophilic species, potassium phosphate (KH2PO4) or sodium bicarbonate (NaHCO3) is often employed. Various organic buffers are available for the growth of acidophiles and alkaliphiles and are widely used for assaying enzymes extracted from cells. The buffer keeps the enzyme solution at optimal pH during the assay, thus ensuring that the enzyme remains catalytically active and unaffected by any protons or hydroxyl ions generated in the enzymatic reaction.
Check Your Understanding
How does the concentration of H+ change when a culture medium at pH 5 is adjusted to pH 9?
What terms are used to describe organisms whose growth pH optimum is very high? Very low?
In terms of pH, what class of organism is the bacterium Escherichia coli?
4.15 Osmolarity and Microbial Growth
Water is the solvent of life, and water availability is an important factor affecting the growth of microorganisms. Water availability depends not only on how moist or dry an environment is but also on the concentration of solutes (salts, sugars, or other substances) dissolved in the water that is present. Solutes bind water, making it less available to organisms. Hence, for organisms to thrive in high-solute environments, physiological adjustments are necessary. Water availability is expressed in terms of water activity (aw), the ratio of the vapor pressure of air in equilibrium with a substance or solution to the vapor pressure of pure water. Values of aw vary between 0 (no free water) and 1 (pure water); some aw values are listed in Table 4.5.
Table 4.5 Water activity of several substances

aSelected examples of Bacteria and Archaea or fungi capable of growth in culture media adjusted to the stated water activity.
Water diffuses from regions of higher water concentration (lower solute concentration) to regions of lower water concentration (higher solute concentration) in the process of osmosis. The cytoplasm of a cell typically has a higher solute concentration than the environment, so the tendency for water is to diffuse into the cell. Under such conditions, the cell is said to be in positive water balance, which is the normal state of the cell. However, when a cell is placed in an environment where the solute concentration exceeds that of the cytoplasm, water will flow out of the cell. If a cell has no strategy to counteract this, it will become dehydrated and unable to grow.
Table 4.6 Compatible solutes of microorganisms

a See Figure 6.27 for the structures of amino acids.
bStructures not shown. Like sucrose, trehalose is a C12 disaccharide; glucosylglycerol is a C9 alcohol; mannitol is a C6 alcohol.
cTo achieve an osmotic aw lower than about 0.77, solutes other than just NaCl are necessary; for example, other salts (MgCl2, MgSO4, or CaCl2) or nonsalts, such as glycerol or sucrose. For most organisms listed (other than for the xerophiles), the lower aw for growth can be extended downward somewhat by additional solutes.
dGrowth of Zygosaccharomyces tested in high-sucrose medium. Germination of Xeromyces spores tested using matric water potential.
Although halophiles require at least some NaCl for growth, the NaCl optimum varies with the organism and is habitat dependent (Figure 4.27). For example, marine microorganisms typically grow best with 1–4% NaCl, organisms from hypersaline environments (environments that are more salty than seawater) grow best at 3–12% NaCl, and organisms from extremely hypersaline environments require even higher levels of NaCl. Organisms isolated from brackish waters (a mixture of freshwater and seawater) may or may not be halophilic.
Figure 4.27 Effect of NaCl concentration on growth of microorganisms of different salt tolerances or requirements.

The optimum NaCl concentration for marine microorganisms such as Aliivibrio fischeri is about 5%; for extreme halophiles, it is between 15 and 30%, depending on the organism.
Mastering Microbiology
Art Activity: Figure 4.27 Effect of NaCl on growth of microorganisms of different salt tolerances or requirements
In contrast to halophiles, halotolerant organisms can tolerate some level of dissolved solutes but grow best in the absence of the added solute (Figure 4.27). Halophiles capable of growth in very salty environments are called extreme halophiles (Figure 4.27). These organisms require very high levels of NaCl, typically 15–30%, for optimum growth and are often unable to grow at all at NaCl concentrations below this. Organisms able to live in environments high in sugar are called osmophiles, and those able to grow in very dry environments (made dry by lack of water rather than by dissolved solutes) are called xerophiles. Examples of these various classes of organisms are given in Table 4.6.
From growth data obtained from extremely halophilic representatives of all three domains of life, there appears to be a common lower water activity limit for living organisms, and this limit is 0.61. This lower limit is likely set by the physiochemical constraints on obtaining water in osmotic environments of aw less than 0.6 that cannot be overcome through biochemical adaptations by the cell. Matric water activity, a measure of water bound to a surface, is measured in the same way as osmotic water activity but can drop to significantly lower than 0.6 and still contain viable microbial communities. For example, hyper-arid hot desert soils can have matric aw values as low as 0.1 during daylight hours. But these environments absorb moisture at night and during rain events, and these increase the water activity to above 0.6, making conditions suitable for microbial metabolism and growth.
Compatible Solutes
When an organism is transferred from a medium of high aw to one of low aw, it maintains positive water balance by increasing its internal solute concentration. This is possible either by pumping solutes into the cell from the environment or by synthesizing a cytoplasmic solute (Table 4.6). In either case, the solute must not inhibit biochemical processes in the cell and is thus called a compatible solute.
Compatible solutes are highly water-soluble organic molecules and include sugars, alcohols, and amino acid derivatives (Table 4.6). Glycine betaine, an analog of the amino acid glycine, is widely distributed among halophilic bacteria. Other common compatible solutes include sugars such as sucrose and trehalose, dimethylsulfoniopropionate (produced by marine algae), and glycerol, a common solute in xerophilic fungi, organisms that grow at the lowest water activities known (Table 4.6). In contrast to these organic solutes, KCl is the compatible solute of extremely halophilic Archaea, such as Halobacterium (Section 17.1), and of a few extremely halophilic Bacteria.
The concentration of compatible solute in a cell is a function of the levels of solute in its environment, and adjustments are made in response to the challenge from external solutes. However, in any given organism, the maximal level of compatible solute tolerated is a genetically encoded characteristic. As a result, different organisms have evolved to thrive in habitats of different salinities (Tables 4.5 and 4.6). In fact, organisms designated as nonhalotolerant, halotolerant, halophilic, or extremely halophilic (Figure 4.27) are to some extent a reflection of their genetic capacity to produce or accumulate compatible solutes.
Check Your Understanding
What is the aw of pure water? What is the lower limit of aw for life?
What are compatible solutes, and when and why are they needed by the cell? What is the compatible solute of Halobacterium?
4.16 Oxygen and Microbial Growth
Oxygen (O2) is an essential nutrient for many microbes; they are unable to metabolize or grow without it. Other microbes, by contrast, cannot grow in the presence of O2 and may even be killed by it. We therefore see, just as we did for other environmental factors considered in this chapter, classes of microorganisms based on their needs or tolerance of O2.
Oxygen Classes of Microorganisms
Microorganisms can be grouped according to their relationship with O2 as outlined in Table 4.7. Aerobes can grow at full oxygen tensions (air is 21% O2) and respire O2 in their metabolism. Microaerophiles, by contrast, are aerobes that can use O2 only when it is present at levels reduced from that in air (microoxic conditions). This is because of the limited capacity of these organisms to respire or because they contain some O2-sensitive molecule such as an O2-labile enzyme. Many aerobes are facultative, meaning that under the appropriate nutrient and culture conditions they can grow in the absence of O2.
Table 4.7 Oxygen relationships of microorganisms

aLetters in parentheses indicate phylogenetic status (B, Bacteria; A, Archaea). Representatives of either domain of prokaryotic cells are known in each category. Most eukaryotes are obligate aerobes, but facultative aerobes (for example, yeast) and obligate anaerobes (for example, certain protozoa and fungi) are known.
bListed are typical habitats of the example organism; many others could be listed.
Some organisms cannot respire oxygen and are called anaerobes. There are two kinds of anaerobes: aerotolerant anaerobes, which can tolerate O2 and grow in its presence even though they cannot respire, and obligate anaerobes, which are inhibited or even killed by O2 (Table 4.7). Anoxic (O2-free) microbial habitats are common in nature and include muds and other sediments, bogs, marshes, water-logged soils, intestinal tracts of animals, sewage sludge, the deep subsurface of Earth, and many other environments. Because there are many habitats for anaerobes, they are very common in nature and highly diverse. As far as is known, obligate anaerobiosis is characteristic of only three groups of microorganisms: a wide variety of Bacteria and Archaea, a few fungi, and a few protozoa.
Some of the best-known prokaryotic anaerobes are Clostridium, a genus of gram-positive endospore-forming Bacteria, and the methanogens, a group of methane-producing Archaea. Among obligate anaerobes, the sensitivity to O2 varies greatly. Many clostridia, for example, although requiring anoxic conditions for growth, can tolerate traces of O2 or even full exposure to air. Others, such as the methanogens, are killed rapidly by O2 exposure.
Culture Techniques for Aerobes and Anaerobes
For the growth of aerobes, it is necessary to provide extensive aeration. This is because the O2 that is consumed by the organisms during growth is not replaced fast enough by diffusion from the air. Therefore, forced aeration of liquid cultures is needed and can be achieved by either vigorously shaking the flask or tube on a shaker or by bubbling sterilized air into the medium through a fine glass tube or porous glass disk.
For the culture of anaerobes, the problem is not to provide O2 but to exclude it. Bottles or tubes filled completely to the top with culture medium and fitted with leakproof closures provide suitably anoxic conditions for organisms that are not overly sensitive to small amounts of O2. A chemical called a reducing agent may be added to such vessels to remove traces of O2 by reducing it to water (H2O). An example is thioglycolate, which is present in thioglycolate broth, a medium commonly used to test an organism’s requirements for O2 (Figure 4.28).
Figure 4.28 Growth versus O2 concentration.

From left to right, aerobic, anaerobic, facultative, microaerophilic, and aerotolerant anaerobe growth, as revealed by the position of microbial colonies (depicted in this drawing as black dots) within tubes of thioglycolate broth culture medium. A small amount of agar has been added to keep the liquid from becoming disturbed. The redox dye resazurin, which is pink when oxidized and colorless when reduced, has been added as a redox indicator. (a) O2 penetrates only a short distance into the tube, so obligate aerobes grow only close to the surface. (b) Anaerobes, being sensitive to O2, grow only away from the surface. (c) Facultative aerobes are able to grow in either the presence or the absence of O2 and thus grow throughout the tube. However, growth is better near the surface because these organisms can respire O2. (d) Microaerophiles grow away from the most oxic zone. (e) Aerotolerant anaerobes grow throughout the tube. Growth is not better near the surface because these organisms can only ferment. In nature, many different habitats exist for each of these oxygen classes. In addition, a single habitat, such as a soil particle, may support growth of both aerobes and anaerobes.
Thioglycolate broth is a complex medium containing a small amount of agar, making the medium viscous but still fluid. After thioglycolate reacts with O2 throughout the tube, O2 can penetrate only near the top of the tube where the medium contacts air. Obligate aerobes grow only at the top of such tubes. Facultative organisms grow throughout the tube but grow best near the top. Microaerophiles grow near the top but not right at the top. Anaerobes grow only near the bottom of the tube, where O2 cannot penetrate, whereas aerotolerant anaerobes grow throughout the tube. The redox indicator dye resazurin is present in thioglycolate broth to signal oxic regions; the dye is pink when oxidized and colorless when reduced and so gives a visual assessment of the degree of penetration of O2 into the medium (Figure 4.28).
To remove all traces of O2 for the culture of strict anaerobes, one can incubate tubes or plates in a glass jar flushed with an O2-free gas or fitted with an O2 consumption system (**Figure 4.29*a***). For manipulating cultures in an anoxic atmosphere, special enclosures called anoxic glove bags permit work with open cultures in completely anoxic atmospheres (Figure 4.29b).
Figure 4.29 Incubation under anoxic conditions.

(a) Anoxic jar. A chemical reaction in the envelope in the jar generates H2+CO2. The H2 reacts with O2 in the jar on the surface of a palladium catalyst to yield H2O; the final atmosphere contains N2, H2, and CO2. (b) Anoxic glove bag for manipulating and incubating cultures under anoxic conditions. The airlock on the right, which can be evacuated and filled with O2-free gas, serves as a port for adding and removing materials to and from the glove bag.
Why Is Oxygen Toxic?
Why are anaerobic microorganisms inhibited in their growth or even killed by oxygen? Molecular oxygen (O2) per se is not toxic, but O2 can be converted to toxic oxygen by-products, and it is these that can harm or kill cells not able to deal with them. These include superoxide anion (O2−), hydrogen peroxide (H2O2), and hydroxyl radical (OH⋅). All of these are by-products of the reduction of O2 to H2O in respiration (Figure 4.30). Flavoproteins, quinones, and iron–sulfur proteins, electron carriers found in virtually all cells (Section 3.8), also catalyze some of these reductions. Thus, regardless of whether it can respire O2, an organism exposed to O2 will experience toxic forms of oxygen, and if not destroyed, these molecules can wreak havoc in cells. For example, superoxide anion and OH⋅ are strong oxidizing agents that can oxidize macromolecules and any other organic compounds in the cell. Peroxides such as H2O2 can also damage cell components but are not as toxic as O2− or OH⋅.
Figure 4.30 Four-electron reduction of O2 to H2O by stepwise addition of electrons.

All the intermediates formed are reactive and toxic to cells; water is not.
Superoxide Dismutase and Other Enzymes That Destroy Toxic Oxygen
A major requirement for inhabiting an oxic world is to keep toxic oxygen molecules under control. Microbes accomplish this in much the same way as plants and animals do. Superoxide anion and H2O2 are the most abundant toxic oxygen species, and all aerobic and aerotolerant cells have enzymes that destroy these compounds (Figure 4.31). The enzymes catalase and peroxidase attack H2O2, forming O2 and H2O, respectively (Figure 4.31 and Figure 4.32). Superoxide anion is destroyed by the enzyme superoxide dismutase, an enzyme that generates H2O2 and O2 from two molecules of O2− (Figure 4.31c). Superoxide dismutase and catalase (or peroxidase) thus work in series to convert O2− to harmless products (Figure 4.31d).
Figure 4.31 Enzymes that destroy toxic oxygen species.

(a) Catalases and (b) peroxidases are porphyrin-containing proteins, although some flavoproteins may consume toxic oxygen species as well. (c) Superoxide dismutases are metal-containing proteins, the metals being copper and zinc, manganese, or iron. (d) Combined reaction of superoxide dismutase and catalase. (e) Superoxide reductase catalyzes the one-electron reduction of O2− to H2O2.
Figure 4.32 Method for testing a microbial culture for the presence of catalase.

A heavy loopful of cells from an agar culture was mixed on a slide (right) with a drop of 30% hydrogen peroxide. The immediate appearance of bubbles is indicative of the presence of catalase. The bubbles are O2 produced by the reaction H2O2+H2O2→2 H2O+O2.
Aerobes and facultative aerobes typically contain both superoxide dismutase and catalase. Superoxide dismutase is an essential enzyme for aerobes. Some aerotolerant anaerobes lack superoxide dismutase and use protein-free manganese complexes instead to carry out the dismutation of O2− to H2O2 and O2. Such a system is not as efficient as superoxide dismutase, but it is sufficient to protect the cells from O2− damage. In some strictly anaerobic Archaea and Bacteria, superoxide dismutase is absent and instead the enzyme superoxide reductase functions to remove O2−. Unlike superoxide dismutase, superoxide reductase reduces O2− to H2O2 without the production of O2 (Figure 4.31e), thus avoiding exposure of the organism to O2.
Check Your Understanding
How does an obligate aerobe differ from a facultative aerobe?
How does a reducing agent work? Give an example of a reducing agent.
How does superoxide dismutase or superoxide reductase protect a cell?
V Controlling Microbial Growth
Chemical and physical agents used to control the growth of microorganisms include heat, radiation, and toxic chemicals. Microbial growth control—whether simple disinfection or actual sterilization—is part of our everyday lives and helps curb the transmission of infectious microbes.
Thus far in this chapter we have discussed microbial growth from the perspective of conditions that promote their growth. We close by considering the opposite side of the coin, microbial growth control.
Many aspects of microbial growth control have significant practical applications. For example, we routinely wash fresh fruits and vegetables to remove attached microorganisms and we inhibit microbial growth on body surfaces by washing. However, neither of these processes kills or removes all microorganisms. Only sterilization—the killing or removal of all microorganisms (including viruses)—ensures that this is the case. In many circumstances, sterility is not required. In others, however, sterilization is absolutely essential.
4.17 General Principles and Microbial Growth Control by Heat
4.17 General Principles and Microbial Growth Control by Heat
4.17 General Principles and Microbial Growth Control by Heat
The effects of microorganisms can often be controlled by simply limiting or inhibiting growth. Methods for inhibiting microbial growth include decontamination, the treatment of an object or surface to make it safe to handle, and disinfection, a process that directly targets pathogens although it may not eliminate all microorganisms. Decontamination can be as simple as wiping off food utensils to remove food fragments (and their attached organisms) before using them, while disinfection requires agents called disinfectants that actually kill microorganisms or severely inhibit their growth. Physical methods of microbial growth control are used extensively in industry, medicine, and the home, and we consider three classes of physical controls in this section and the next: heat, radiation, and filtration. Of these three, heat is the most widely used method of physically treating an object or substance to render it sterile.
Heat Sterilization
The effectiveness of heat as a sterilant is quantified by the time required at a given temperature for a 10-fold reduction in the viability of a microbial population. This is called the decimal reduction time (D). The relationship between D and temperature is exponential, as the logarithm of D plotted against temperature yields a straight line (Figure 4.33). Moreover, heat killing proceeds more rapidly as the temperature rises. The type of heat is also important: Moist heat has better penetrating power than dry heat and, at a given temperature, inhibits growth or kills cells more quickly than does dry heat.
Figure 4.33 The effect of temperature on the heat killing of microorganisms.

(a) The decimal reduction time (D) is the time at which only 10% of the original population of a given organism (in this case, a mesophile) remains viable at a given temperature. For 70 °C, D=3 min; for 60 °C, D=12 min; for 50 °C, D=42 min. (b) D values for model organisms of different temperature classes: A, mesophile; B, thermophile; C, hyperthermophile.
Another way to characterize the heat sensitivity of an organism is to measure its thermal death time, the time it takes to kill all cells at a given temperature. To determine the thermal death time, samples of a cell suspension are heated for different times, mixed with culture medium, and incubated. If all the cells have been killed, no growth is observed in the incubated samples. However, unlike a decimal reduction time measurement that is independent of the original cell number, the thermal death time is greatly affected by population size; a longer time is required to kill all cells in a large population than in a small one.
The presence of endospore-forming bacteria in a heat-treated sample can influence both the decimal reduction and thermal death times. Recall that the mature endospore is very dehydrated and contains calcium dipicolinate and small acid-soluble spore proteins (SASPs) that help confer heat stability on the structure (Section 2.8). The medium in which heating takes place also influences the rate of killing, and this is especially relevant in food canning procedures. Microbial death is more rapid at acidic pH, and acidic foods such as tomatoes, fruits, and pickles are easier to sterilize than neutral-pH foods such as corn and beans. Moreover, high concentrations of sugars, proteins, and fats decrease heat penetration and usually increase the resistance of organisms to heat, whereas high salt concentrations may either increase or decrease heat resistance, depending on the organism.
The Autoclave and Pasteurization
The autoclave is a sealed heating device that uses steam under pressure to kill microorganisms (Figure 4.34). Killing of heat-resistant endospores requires heating at temperatures above the boiling point of water at 1 atm. The autoclave places steam under a pressure of 1.1 kg/cm2 (15 lb/in2), which yields a temperature of 121 °C. At 121 °C, the time to achieve sterilization of small amounts of endospore-containing material is about 15 min (Figure 4.34b). If the object to be autoclaved is bulky or large volumes of liquids are to be sterilized, heat transfer to the interior is retarded, and thus the total heating time must be extended. Note that it is not the pressure inside the autoclave that kills the microorganisms but the high temperatures that are achieved when steam is placed under pressure.
Figure 4.34 The autoclave and moist heat sterilization.

(a) The flow of steam through an autoclave. (b) A typical autoclave cycle. The temporal heating profile of a fairly bulky object is shown. The temperature of the object rises and falls more slowly than the temperature of the autoclave. The temperature of the object must reach the target temperature and be held for 10–15 min to ensure sterility, regardless of the temperature and time recorded in the autoclave. (c) A modern research autoclave containing a pressure-lock door and automatic cycle controls.
Pasteurization, a process named for the famous early microbiologist Louis Pasteur (Section 1.11), is not the same as sterilization. Pasteurization uses heat to significantly reduce rather than totally eliminate the microorganisms found in liquids, such as milk. At the temperatures and times standardized for the pasteurization of food products, all known pathogenic bacteria are killed. In addition, however, by decreasing the overall microbial load, pasteurization increases the shelf life of perishable liquids.
To pasteurize milk, the liquid is passed through a tubular heat exchanger. Careful control of flow rate and the size and temperature of the heat source raises the temperature of the milk to 71 °C for 15 sec (or even higher temperatures for shorter time periods; see time–temperature relationships in Figure 4.33), after which it is rapidly cooled. This process is called flash pasteurization. Ultrahigh-temperature (UHT) pasteurization of milk requires heat treatment at 135 °C for 1–2 sec and actually sterilizes the milk such that it can be stored at room temperature for long periods without spoilage.
Check Your Understanding
What steps are necessary to ensure the sterility of material contaminated with bacterial endospores?
Distinguish between the sterilization of microbiological media and the pasteurization of dairy products.
4.18 Other Physical Control Methods: Radiation and Filtration
4.18 Other Physical Control Methods: Radiation and Filtration
4.18 Other Physical Control Methods: Radiation and Filtration
In addition to heat, radiation, in particular ultraviolet (UV) light, X-rays, and gamma rays, are also effective microbial killing agents. However, each type of energy has a different mode of action and killing efficacy and thus their applications vary widely.
Ultraviolet and Ionizing Radiation
Ultraviolet radiation between 220 and 300 nm is absorbed by DNA and can cause mutations or have other serious effects on DNA that lead to death of the exposed organism (Section 9.4). UV radiation is useful for disinfecting surfaces and air and is widely used to decontaminate and disinfect the work surface of laboratory laminar flow hoods equipped with a “germicidal” UV light (Figure 4.35) and air circulating in hospital and food preparation rooms. However, UV radiation has very poor penetrating power, limiting its use to the disinfection of exposed surfaces or air rather than bulk objects such as canned foods or surgical clothing.
Figure 4.35 A laminar flow hood.

An ultraviolet light source prevents contamination of the hood when it is not in use. When the hood is in use, air is drawn into the cabinet through a HEPA filter. The filtered air inside the cabinet is exhausted out of the cabinet, preventing contamination of the inside of the hood. The cabinet provides a contaminant-free workspace for microbial and tissue culture manipulations.
Ionizing radiation is electromagnetic radiation of sufficient energy to produce ions and other reactive molecular species from molecules with which the radiation particles collide. The unit of ionizing radiation is the roentgen, and the standard for sterilization is the absorbed radiation dose, measured in rads (100 erg/g) or grays (1 Gy=100 rad). Ionizing radiation is typically generated from X-ray sources or the radioactive nuclides 60Co and 137Cs. These nuclides produce X-rays or gamma rays, both of which have sufficient energy and penetrating power to kill microorganisms in bulk items such as food products and medical supplies.
Table 4.8 shows the dose necessary for a 10-fold reduction (D10) in number of selected microorganisms. The D10 value is analogous to the decimal reduction time (D) for heat sterilization, and the killing curve of ionizing radiation yields a similar plot (Figure 4.36; compare with Figure 4.33). As is also true of heat treatments, killing endospores with ionizing radiation is more difficult than killing vegetative cells, and viruses are more difficult to kill than bacteria (Table 4.8). In addition, microorganisms in general are much more resistant to ionizing radiation than are multicellular organisms. For example, the lethal radiation dose for humans can be as low as 10 Gy if delivered over a short time period.
Table 4.8 Radiation sensitivity of some representative microorganisms

aD10 is the amount of radiation necessary to reduce the initial population or activity level 10-fold (1 logarithm, see Figure 4.36). Gy, grays. 1 Gy is equivalent to 100 rads. The lethal dose for humans is 10 Gy.
Figure 4.36 Relationship between the survival fraction and the radiation dose of a microorganism.

The D10, which is the decimal reduction dose, can be interpolated from the data as shown.
In the United States, radiation is used for the sterilization of such diverse items as surgical supplies, plastic labware, drugs, and even tissue grafts. Certain foods and food products such as fresh produce, poultry, meat products, and spices are also routinely irradiated to ensure that they are sterile or at least free of pathogens and insects.
Filter Sterilization
Heat is an effective way to decontaminate most liquids, but not all substances are stable at high temperature and so heat-sensitive liquids that are not sterilized by radiation are typically sterilized by filtration. For sterilization, a filter with pores of average size 0.2 μm is a minimum requirement; however, even such tiny holes will not trap most viruses. Commonly used filter pore sizes for the filter sterilization of small volumes, such as laboratory solutions, are 0.45 μm and 0.2 μm.
Several types of filters are in routine use in microbiology, including depth filters, membrane filters, and nucleopore filters. A depth filter is a fibrous sheet made from an array of overlapping paper or glass fibers that traps particles in the network of fibers (Figure 4.37a). Depth filters are important in biosafety applications such as in a biological safety cabinet where air, both into and out of the cabinet, flows through a depth filter called a HEPA filter, or high-efficiency particulate air filter (**Figure 4.37*a***). HEPA filters typically remove 0.3- μm or larger particles from an airstream with an efficiency of greater than 99.9%; however, this does not ensure sterilization.
Figure 4.37 Microbiological filters.

Scanning electron micrograph showing the structure of (a) a depth filter, (b) a conventional membrane filter, and (c) a nucleopore filter. (d) Scanning electron micrograph of various aquatic microbes trapped on a nucleopore 0.2- μm-pore-size membrane filter.
Membrane filters are the most common filters used for liquid sterilization in the microbiology laboratory (Figure 4.37b and Figure 4.38). Membrane filters are composed of high-tensile-strength polymers manufactured in such a way as to contain a large number of tiny pores. Filtration is accomplished by using a syringe or a pump to force the liquid through the filtration apparatus into a sterile collection vessel (Figure 4.38). Another type of membrane filter is the nucleopore filter. Nucleopore filters are made from a thin polycarbonate film that is treated with radiation and then etched with a chemical, yielding very uniform-sized holes (Figure 4.37c). Nucleopore filters are commonly used to isolate specimens for scanning electron microscopy. Microorganisms removed by filtration from a liquid or a natural sample, such as lake water, can then be observed directly on the filter (Figure 4.37d).
Figure 4.38 Membrane filters.

Disposable, presterilized, and assembled membrane filter units. Left: a filter system designed for small volumes forced through the filter by a syringe. Right: a system employing a peristaltic pump for filtering larger volumes.
Check Your Understanding
Why is ionizing radiation more effective than UV radiation for sterilization of food products?
Distinguish between the major types of sterilization filters used in the microbiology laboratory.
4.19 Chemical Control of Microbial Growth
Chemicals are routinely used to control microbial growth, and an antimicrobial agent is a natural or synthetic chemical that kills or inhibits the growth of microorganisms. Agents that actually kill are called -cidal agents, with a prefix indicating the type of microorganism killed. Thus, bactericidal agent, fungicidal, and viricidal agents kill bacteria, fungi, and viruses, respectively. Agents that do not kill but only inhibit growth are called -static agents, and include bacteriostatic agent, fungistatic, and viristatic compounds. We focus here on chemicals commonly used as disinfectants and reserve discussion of the activities of a very important class of chemicals—the antibiotics—for later (Sections 8.11 and 28.5).
Effect of Antimicrobial Agents on Growth
Antibacterial agents are classified as -static, -cidal, or -lytic (cell lysing) by observing their effects on cultures using viable and turbidimetric growth assays (Figure 4.39). Bacteriostatic agents are typically inhibitors of some important biochemical process and bind relatively weakly; if the agent is removed, the cells can resume growing. Some antibiotics, such as the sulfonamides, work in this way. The clinical value of bacteriostatic antibiotics is their ability to keep a pathogenic bacterium from multiplying until the immune system can rid the body of the pathogen. By contrast, bactericidal agents, for example formaldehyde, bind tightly to their cellular targets and by definition kill the cell. However, the dead cells are not lysed, and total cell numbers, reflected in the turbidity of the culture, remain constant (Figure 4.39b). Bacteriolytic agents kill cells by lysing them, and this affects both viable and total cell numbers (Figure 4.39c). Examples of bacteriolytic agents would be a detergent that ruptures the cytoplasmic membrane or the antibiotic penicillin, which inhibits bacterial cell wall synthesis, resulting in cell lysis.
Figure 4.39 Different types of antimicrobial agents.

(a) Bacteriostatic agents inhibit but do not kill. (b) Bactericidal agents kill. (c) Bacteriolytic agents lyse cells. In each graph, the arrow indicates the time at which a growth-inhibitory concentration of an antimicrobial agent was added to an exponentially growing culture. The turbidity and viable counts shown are characteristic of each type of agent.
Mastering Microbiology
Art Activity: Figure 4.39 Different types of antimicrobial agents
Assaying Antimicrobial Activity
Antimicrobial activity can be measured by determining the smallest amount of the agent needed to inhibit the growth of a test organism, a value called the minimum inhibitory concentration (MIC). One way to determine the MIC of a given agent is to inoculate a series of tubes of liquid growth medium (Figure 4.40) containing a test organism and dilutions of the agent. Following incubation, the tubes are scored for growth (turbidimetrically), and the MIC is revealed as the lowest concentration of antimicrobial agent that completely inhibits growth.
Figure 4.40 Antimicrobial agent susceptibility assay using dilution methods.

The assay defines the minimum inhibitory concentration (MIC). A series of increasing concentrations (from left to right) of antimicrobial agent is prepared in the culture medium. Each tube is inoculated with a specific concentration of a test organism, followed by a defined incubation period. Growth, measured as turbidity, occurs in those tubes with antimicrobial agent concentrations below the MIC.
Antimicrobial activity can also be assessed using solid media (Figure 4.41). Known amounts of an antimicrobial agent are added to filter-paper disks and the disks are arranged on the surface of a uniformly inoculated agar plate. During incubation, the agent diffuses from the disk into the agar, establishing a gradient; the farther a chemical diffuses away from a disk, the lower its concentration. Following an incubation period, a zone of growth inhibition forms around disks that released effective chemicals. The zone is a function of several factors, including the amount of antimicrobial agent added to the disk, its solubility and diffusion coefficient, and, most importantly, its overall effectiveness. The disk diffusion assay is routinely used to test clinically isolated pathogenic bacteria for their antibiotic susceptibility (Section 28.5).
Figure 4.41 Antimicrobial agent susceptibility assay using diffusion methods.

The antimicrobial agent diffuses from paper disks into the surrounding agar, inhibiting growth of susceptible microorganisms.
Chemical Antimicrobial Agents
Several antimicrobial agents are used to prevent the growth of human pathogens on inanimate surfaces and on external body surfaces. These include sterilants, disinfectants, sanitizers, and antiseptics (Table 4.9).
Table 4.9 Antiseptics, sterilants, disinfectants, and sanitizersa

aAlcohols, hydrogen peroxide, and iodophors can be antiseptics, disinfectants, sanitizers, or sterilants depending on concentration, length of exposure, and form of delivery.
Sterilants destroy all microorganisms, including endospores. Chemical sterilants are used for decontamination or sterilization in situations where it is impractical or impossible to use heat or radiation. Hospitals, clinics, and laboratories, for example, must routinely decontaminate and sterilize heat-sensitive materials such as thermometers, lensed instruments, polyethylene tubing, catheters, and reusable medical and dental equipment. This process of cold sterilization, as it is called, employs gases such as ethylene oxide or aldehydes such as formaldehyde or glutaraldehyde to sterilize the devices.
Disinfectants are chemicals that kill microorganisms but not necessarily endospores and are primarily used on surfaces. For example, phenol and cationic detergents are used to disinfect floors, tables, bench tops, walls, and so on (Table 4.9) and are important agents of infection control in hospitals and other medical settings. Sanitizers, by contrast, are less harsh than disinfectants and reduce microbial numbers but do not sterilize. Sanitizers are widely used in the food industry to treat surfaces such as mixing and cooking equipment, dishes, and utensils, and are also used for dry hand washing when water is unavailable. Antiseptics, often called germicides, are chemicals that kill or inhibit the growth of microorganisms but are sufficiently nontoxic to animals to be applied to living tissues. Most germicides are used for hand washing or for treating surface wounds (Table 4.9). Certain antiseptics are also effective disinfectants. Ethanol, for example, can be both an antiseptic and a disinfectant, depending on the concentration and exposure time employed.
Several factors affect the efficacy of any chemical antimicrobial agent. For example, many antimicrobial agents are bound and inactivated by organic matter; thus, disinfecting a kitchen countertop littered with spilled foods is more difficult than disinfecting a clean countertop. Furthermore, bacteria can form biofilms (Section 4.9), covering surfaces of tissue or soiled medical devices with microbial cells embedded in polysaccharides. Biofilms may slow or even completely prevent penetration of antimicrobial agents, reducing or negating their effectiveness. Thus, the ultimate efficacy of any antimicrobial agent must be determined empirically and under the actual conditions of use. Only by actually testing the chemical and assaying for microbial growth both before and after treatment can one be confident that the agent is working as it should.
Check Your Understanding
Distinguish between the antimicrobial effects of -static, -cidal, and -lytic agents.
Describe how the minimum inhibitory concentration of an antibacterial agent is determined.
Distinguish between a sterilant, a disinfectant, and an antiseptic. What is cold sterilization?
Chapter Review
Go to Mastering Microbiology for videos, animations, practice tests, and more.
I Culturing Microbes and Measuring Their Growth
4.1 Cells are primarily composed of the elements H, O, C, N, P, and S. Nutrients required by a cell in large amounts are called macronutrients while those required in very small amounts, such as trace elements or growth factors, are micronutrients. Proteins are the most abundant class of macromolecules in the cell.
Q Why are carbon and nitrogen macronutrients but cobalt is a micronutrient?
4.2 Culture media supply the nutritional needs of microorganisms and are either defined or complex. Other media, such as selective and differential media, are used for specific purposes. Many microorganisms can be grown in liquid or solid culture media, and pure cultures can be maintained if aseptic technique is practiced.
Q Why would the following medium not be considered a chemically defined medium: glucose, 5 grams (g); NH4Cl, 1 g; KH2PO4, 1 g; MgSO4, 0.3 g; yeast extract, 5 g; distilled water, 1 liter? What is aseptic technique and why is it necessary?
4.3 Total cell counts can be done under the microscope using counting chambers and are useful for assessing total cell numbers in a microbial habitat or laboratory culture. Certain stains can be used to target specific cell populations in a sample.
Q Are total cell counts useful if one does not know the viability of the culture or sample?
4.4 Viable cell counts measure only the living population present in the sample with the assumption that each colony originates from the growth and division of a single cell. Depending on the growth medium and conditions employed, and the nature of the sample, viable counts can be fairly accurate assessments or highly unreliable.
Q How does a viable count differ from a total count?
4.5 Turbidity measurements are a rapid and useful method of measuring microbial growth and are based on the fact that cells in suspension scatter light. In order to relate a turbidity value to a cell number, a standard curve plotting these two parameters against one another must first be established.
Q How can turbidity be used as a measure of cell numbers?
II Dynamics of Microbial Growth
4.6 Microbial growth is defined as an increase in cell numbers and is the final result of the doubling of all cell components prior to actual division that yields two daughter cells. Microorganisms show a characteristic growth pattern when inoculated into a fresh culture medium. There is usually a lag phase and then growth commences in an exponential fashion. As essential nutrients are depleted and/or toxic products accumulate, growth ceases and the population enters the stationary phase. Further incubation can lead to cell death.
Q Describe the growth cycle of a population of bacterial cells from the time this population is first inoculated into fresh medium.
4.7 Microbial cells undergo exponential growth, and a semilogarithmic plot of cell numbers with time can reveal the doubling time of the population. Various growth expressions can be calculated from cell number data obtained from an exponentially growing culture. Key expressions here are n, the number of generations; t, time; g, generation time; and k, the instantaneous growth rate constant.
**Q How is the generation time (g) of an exponentially growing culture determined?**
4.8 The chemostat is an open system used to maintain cell populations in exponential growth for extended periods. In a chemostat, the rate at which a culture is diluted with fresh growth medium controls the growth rate of the population, while the cell density (cells/ml) is controlled by the concentration of a growth-limiting nutrient present in the fresh medium.
Q How does a chemostat regulate growth rate and cell density independently?
4.9 Biofilms are an alternative growth mode to a suspended (planktonic) lifestyle and consist of a microbial community enmeshed in a sticky polysaccharide matrix attached to a surface. Biofilms promote intercellular communication and offer protection from predation, harmful substances, and being washed away.
Q What happens during each of the steps that occur in the formation of a microbial biofilm?
4.10 Most microorganisms grow by binary fission but some grow by budding. Budding bacteria show unequal cell growth and produce daughter cells that have different characteristics. Some bacteria such as the actinomycetes form long filaments called hyphae that grow from their tips.
Q How does the process of binary fission differ from that of budding division?
III Environmental Effects on Growth: Temperature
4.11 Temperature is a major environmental factor controlling microbial growth. An organism’s cardinal temperatures are the minimum, optimum, and maximum temperatures at which it grows. Microorganisms can be grouped by their cardinal temperature from cold-loving to heat-loving as psychrophiles, mesophiles, thermophiles, and hyperthermophiles.
Q Examine the graph in Figure 4.21. Why is the optimum temperature for an organism usually closer to its maximum than its minimum?
4.12 Organisms with temperature optima below 15 °C are called psychrophiles, and the most extreme representatives inhabit constantly cold environments. Psychrophiles synthesize macromolecules that remain flexible and functional at cold temperatures but that can be unusually sensitive to warm temperatures.
Q Why are psychrophiles more likely to be found in constantly cold rather than intermittently cold environments?
4.13 Organisms with growth temperature optima between 45 and 80 °C are called thermophiles while those with optima greater than 80 °C are hyperthermophiles. These organisms inhabit hot environments that can have temperatures even above 100 °C. Thermophiles and hyperthermophiles produce heat-stable macromolecules.
Q How do cells of hyperthermophiles prevent heat from destroying them?
IV Environmental Effects on Growth: pH, Osmolarity, and Oxygen
4.14 The acidity or alkalinity of an environment can greatly affect microbial growth. Some organisms grow best at low or high pH (acidophiles and alkaliphiles, respectively), but most organisms grow best between pH 5.5 and 8. The internal pH of a cell must stay relatively close to neutral to prevent the spontaneous destruction of macromolecules at pH extremes.
Q Concerning the pH of the environment and of the cell cytoplasm, in what ways are acidophiles and alkaliphiles different? In what ways are they similar?
4.15 The water activity of an aqueous environment is a function of its solute concentration. To survive in high-solute environments, organisms produce or accumulate compatible solutes to maintain positive water balance. Some microorganisms grow best at reduced water potential and some even require high levels of salts for growth.
Q How does a halophile maintain positive water balance while growing in a solution high in NaCl?
4.16 Aerobes require O2 while anaerobes do not and may even be killed by O2. Facultative organisms can live with or without O2. Special techniques are needed to grow aerobic and anaerobic microorganisms. Several toxic forms of oxygen can form in the cell, but enzymes are present that neutralize most of them. Superoxide is a major toxic form of oxygen.
Q Contrast an aerotolerant and an obligate anaerobe in terms of sensitivity to O2 and ability to grow in the presence of O2. Compare and contrast the enzymes catalase, superoxide dismutase, and superoxide reductase in regard to their substrates and products.
V Controlling Microbial Growth
4.17 Sterilization is the killing of all microbes including viruses, and heat is the most widely used method of sterilization. An autoclave employs moist heat under pressure to achieve temperatures above the boiling point of water. Pasteurization does not sterilize liquids, but it reduces the microbial load and kills pathogens.
Q Contrast the terms thermal death time and decimal reduction time. How would the presence of bacterial endospores affect either value? What time and temperature is necessary to ensure sterility in the autoclave?
4.18 Radiation can effectively inhibit or kill microorganisms. Ultraviolet radiation is used for decontaminating surfaces and air, whereas ionizing radiation is used for sterilization where penetration is required. Filters remove microorganisms from air or liquids. Membrane and nucleopore filters are used for sterilization of heat-sensitive liquids and to examine the contents of filtration by microscopy.
Q Describe the effects of a lethal dose of ionizing radiation at the molecular level. What type of filter would be used to filter sterilize a heat-sensitive liquid?
4.19 Chemicals that kill organisms are called -cidal agents while those that arrest growth but do not kill are called -static agents. Antimicrobial agents are tested for efficacy by determining their ability to inhibit growth. Sterilants, disinfectants, and sanitizers are used to decontaminate nonliving material, while antiseptics (germicides) are used to reduce the microbial load on living tissues.
**Q Describe the procedure for obtaining the minimum inhibitory concentration (MIC) for a chemical that is bactericidal for Escherichia coli. Contrast the action of disinfectants and antiseptics.**
Application Questions
A medium was inoculated with 5×106 cells/ml of Escherichia coli cells. Following a 1-h lag, the population grew exponentially for 5 h, after which the population was 5.4×109 cells/ml. Calculate g and k for this growth experiment.
Escherichia coli but not Pyrolobus fumarii will grow at 40 °C, while P. fumarii but not E. coli will grow at 110 °C. What is happening (or not happening) to prevent growth of each organism at the nonpermissive temperature?
In which direction (into or out of the cell) will water flow in cells of Escherichia coli (an organism found in your large intestine) suddenly suspended in a solution of 20% NaCl? What if the cells were suspended in distilled water? If growth nutrients were added to each cell suspension, which (if either) would support growth, and why?
Chapter Glossary
an organism that grows best at low pH, typically below pH 5.5 Aerobe
an organism that can use O2 in respiration; some require O2 Aerotolerant anaerobe
a microorganism unable to respire O2 but whose growth is unaffected by it Alkaliphile
an organism that has a growth pH optimum of 8 or higher Anaerobe
an organism that cannot use O2 in respiration and whose growth is typically inhibited by O2 Antimicrobial agent
a chemical compound that kills or inhibits the growth of microorganisms Antiseptic (germicide)
a chemical agent that kills or inhibits growth of microorganisms and is sufficiently nontoxic to be applied to living tissues Aseptic technique
a series of steps taken to prevent contamination of laboratory cultures and media Autoclave
a sealed heating device that destroys microorganisms with temperature and steam under pressure Bactericidal agent
an agent that inhibits bacterial growth Batch culture
a closed-system microbial culture of fixed volume Binary fission
cell division following enlargement of a cell to twice its minimum size Biofilm
colonies of microbial cells encased in a porous organic matrix and attached to a surface Budding division
a cell division process whereby new cell material is produced from a single point instead of along the entire cell Cardinal temperatures
the minimum, maximum, and optimum growth temperatures for a given organism Chemostat
a device that allows for the continuous culture of microorganisms with independent control of both growth rate and cell number Colony morphology
the visible appearance of a colony grown on a solid medium Compatible solute
a molecule that is accumulated in the cytoplasm of a cell for adjustment of water activity but that does not inhibit biochemical processes Complex medium
a culture medium of highly nutritious substances for which the exact composition is unknown Culture medium
a nutrient solution required for growing a laboratory culture of a specific microorganism Decline phase
the component of a microbial growth curve during which the rate of cell death exceeds the rate of cell growth Defined medium
a culture medium for which the exact composition is known Disinfectant
an antimicrobial agent used only on inanimate objects Disinfection
rendering a surface or object free of all pathogenic microorganisms Exponential phase (exponential growth)
a condition during which the cell numbers in a microbial population double at regular intervals Extreme halophile
a microorganism that requires very large amounts of NaCl, usually greater than 10% and in some cases near saturation, for growth Facultative
with respect to O2, an organism that can grow in either its presence or absence Generation time
the time required for a population of microbial cells to double Germicide (antiseptic)
a chemical agent that kills or inhibits growth of microorganisms and is sufficiently nontoxic to be applied to living tissues Growth curve
the dynamics of microbial growth observed in batch culture, consisting of four phases: lag, exponential, stationary, and decline Halophile
a microorganism that requires NaCl for growth Halotolerant
a microorganism that does not require NaCl for growth but can grow in the presence of NaCl, in some cases, substantial levels of NaCl Hyperthermophile
a species of Bacteria or Archaea whose growth temperature optimum is 80 °C or greater Hyphal growth
a form of growth in which cells elongate only at the tip of a growing filament Lag phase
the component of a microbial growth curve prior to the onset of growth Mesophile
an organism that grows best at temperatures between 20 and 40 °C Microaerophile
an aerobic organism that can grow only when O2 tensions are reduced from that present in air Minimum inhibitory concentration (MIC)
The smallest amount of an antimicrobial substance that prevents growth of the target microorganism Neutrophile
an organism that grows best at neutral pH, between pH 5.5 and 8 Obligate anaerobe
an organism that cannot grow in the presence of O2 Osmophile
an organism that grows best in the presence of high levels of solute, typically a sugar Pasteurization
the heat treatment of milk or another liquid to reduce its total number of microorganisms pH
the negative logarithm of the hydrogen ion (H+) concentration of a solution Plate count
a method for counting viable cells; the number of colonies on a plate is used as a measure of cell numbers Psychrophile
an organism with a growth temperature optimum of 15 °C or lower and a maximum growth temperature below 20 °C Psychrotolerant
capable of growing at low temperatures but having an optimum above 20 °C Sanitizer
an agent that reduces microorganisms to a safe level, but may not eliminate them Stationary phase
the component of a microbial growth curve during which the rate of growth and death are approximately equal Sterilant
a chemical agent that destroys all forms of microbial life Sterilization
the killing or removal of all living organisms and viruses Thermophile
an organism whose growth temperature optimum lies between 45 and 80 °C Viable
a measurement of the number of live cells in a population Water activity
the ratio of the vapor pressure of air in equilibrium with a solution to the vapor pressure of pure water Xerophile
an organism that is able to live, or that lives best, in very dry environments