6 Molecular Information Flow and Protein Processing
6 Molecular Information Flow and Protein Processing
## Chapter 6 Molecular Information Flow and Protein Processing
II Copying the Genetic Blueprint: DNA Replication
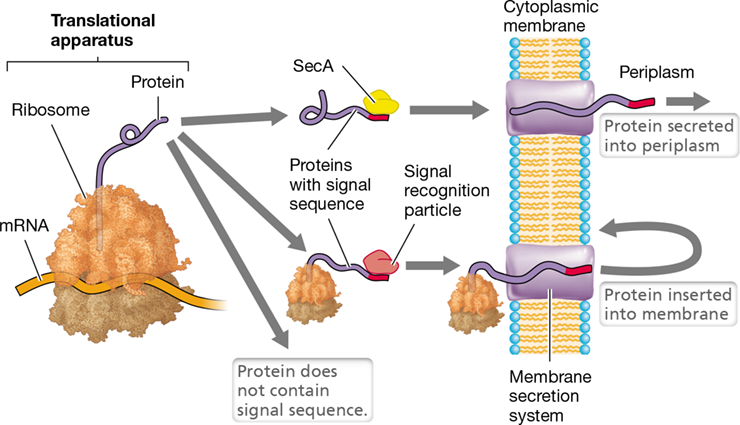
V Protein Processing, Secretion, and Targeting
Injectisomes: *Salmonella*’s Mode of Attack
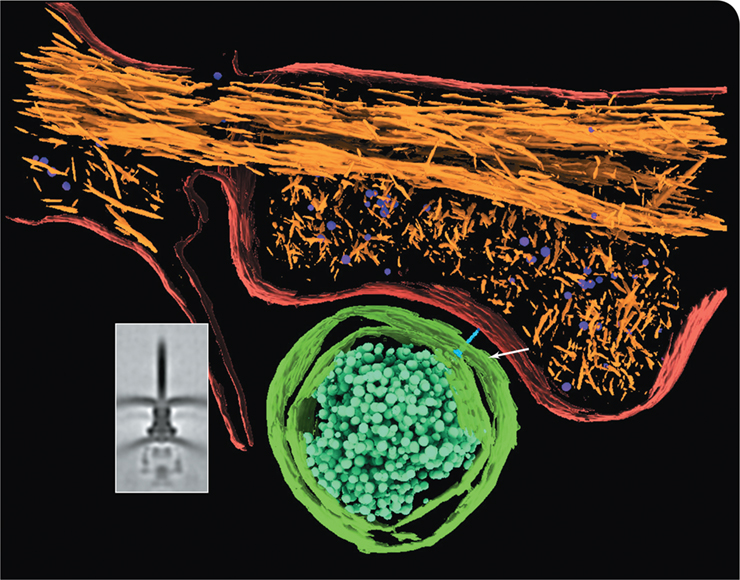
Salmonella is a genus of gram-negative Bacteria that causes salmonellosis in humans. Once Salmonella colonizes the intestines, the pathogen produces proteins called effectors to manipulate host epithelial cells and trigger disease symptoms. In fact, this pathogen delivers over 40 different effectors, many of which are toxins, directly into host epithelial cells during infection. How does Salmonella accomplish such an aggressive feat?
Salmonella possesses a secretion system known as type III that moves effectors all the way through its membranes and cell wall into a host cell. The inset in the photo here shows a cryo-electron tomograph of a type III secretion system. The machinery can be divided into three parts: bottom—a platform that resides inside the bacterial cytoplasm; middle—a basal body within the periplasm and an outer membrane–anchored base; and top—a protruding needle filament. Based on this syringe-like morphology, type III secretion systems have been termed “injectisomes.”
The actual injection of effector proteins into host cells requires an orchestrated series of events that is triggered when the injectisome tip makes contact with the host cell, and the colored tomograph illustrates this cell-to-cell contact. Bacterial cell membranes and ribosomes are green, and a blue injectisome (white arrow) is shown making contact with the host cell membrane (red). Host cell actin filaments and ribosomes are depicted in orange and purple, respectively.
Injectisome contact does not actually stab a hole into the host cell as one might expect. Instead, the contact leads to the formation of a protein channel (not shown) that allows for the passage of the toxin proteins from one domain of life to the other. Injectisomes are deployed by several other pathogens including Yersinia (plague), Shigella (gastrointestinal distress), Pseudomonas (various infections), and Chlamydia (sexually transmitted infections), and thus the success of this structure in facilitating disease symptoms is readily apparent.
Source: Park, D., et al. 2018. Visualization of the type III secretion mediated Salmonella–host cell interface using cryo-electron tomography. eLife 7: e39514.
In this chapter we take a brief detour from our coverage of microbiology per se to consider some basic principles of molecular biology, principles that govern the activities of all cells regardless of domain. The goal is to review how an organism orchestrates the series of macromolecular events that precede cell division and place these events in the context of modern molecular microbiology.
I Molecular Biology and Genetic Elements
Genetic information flow is the hallmark of all cells, as the central dogma of cell biology is that DNA begets RNA begets protein. These processes are invariant in cells of all domains of life and can be initiated from many different DNA genetic elements, the chromosome being chief among them.
In Part I we examine the structure of DNA and the basic processes in genetic information flow and then move on to consider the various types of genetic elements. Part I lays the groundwork for a detailed consideration of DNA replication in prokaryotic cells (Part II) and the mechanisms behind RNA and protein synthesis (Parts III and IV).
6.1 DNA and Genetic Information Flow
An overview of the major macromolecules and processes of molecular biology is shown in **Figure 6.1*a***. In addition, the functional unit of genetic information is the gene, and genes make up parts of chromosomes or other large molecules known collectively as genetic elements; the total complement of genetic elements is the genome. Genetic information is embedded in the sequence of nucleotides in the nucleic acids DNA (deoxyribonucleic acid) and RNA (ribonucleic acid). DNA carries the cell’s genetic blueprint while RNA, produced in transcription, carries a copy of this blueprint. One form of RNA called messenger RNA is converted by translation into defined amino acid sequences in proteins. Collectively, nucleic acids and proteins are called informational macromolecules (Figure 6.1a).
Figure 6.1 Genetic information flow and the components of the nucleic acids.

(a) An overview of the types of informational macromolecules. (b) Part of a DNA chain. The numbers on the sugar of the nucleotide contain a prime (′) to differentiate them from the numbering on the rings of the nitrogen bases. In DNA, a hydrogen is present on the 2′-carbon of the pentose sugar. In RNA, an OH group occupies this position (indicated by the red dashed circle). The nucleotides are linked by a phosphodiester bond. (c) The nitrogen bases of DNA and RNA and the specific pairing between cytosine (C) and guanine (G) and between thymine (T) and adenine (A) via hydrogen bonds. Uracil (U) instead of thymine is present in RNA as indicated by the red dashed circle. Note the numbering system of the rings in that a pyrimidine base bonds through N-1 to the sugar–phosphate backbone and that a purine base bonds through N-9. Atoms that are found in the major groove of the double helix (see Figure 6.3) and that interact with proteins are highlighted in pink.
The monomers of nucleic acids are called nucleotides and so DNA and RNA are polynucleotides. A nucleotide has three components: a pentose sugar (either ribose in RNA or deoxyribose in DNA), a nitrogenous base, and a molecule of phosphate, PO43− (Figure 6.1b). A nucleoside has a pentose sugar and a nitrogenous base but does not include a phosphate group. The nitrogenous bases in nucleic acids are either pyrimidines or purines. The purines guanine and adenine and the pyrimidine cytosine are present in both DNA and RNA, whereas the pyrimidines thymine and uracil are only present (with minor exceptions) in DNA and RNA, respectively (Figure 6.1c).
Properties of the Double Helix
The nucleic acid backbone is a polymer of alternating sugar and phosphate molecules, and nucleotides are linked by phosphate between the 3′-carbon of one sugar and the 5′-carbon of the next sugar, the phosphodiester bond (Figure 6.1b, Figure 6.2). The sequence of nucleotides in a DNA or RNA molecule is its primary structure and encodes the genetic information. In cells, DNA is double-stranded, the strands being held together by hydrogen bonds between the bases in the two strands (Figure 6.1c). Specific base pairing, A with T and G with C, ensures that the two strands of DNA are complementary in base sequence, and this complementarity is essential for the faithful replication of the molecule. The two strands of the DNA molecule are also arranged in an antiparallel fashion; one strand runs 5′ to 3′ (top to bottom), whereas its complement runs 5′ to 3′ (bottom to top) (Figure 6.2).
Figure 6.2 DNA structure.

Complementary and antiparallel nature of DNA. Note that one chain ends in a 5′-phosphate group, whereas the other ends in a 3′-hydroxyl. The purple bases represent the pyrimidines cytosine (C) and thymine (T), and the yellow bases represent the purines adenine (A) and guanine (G).
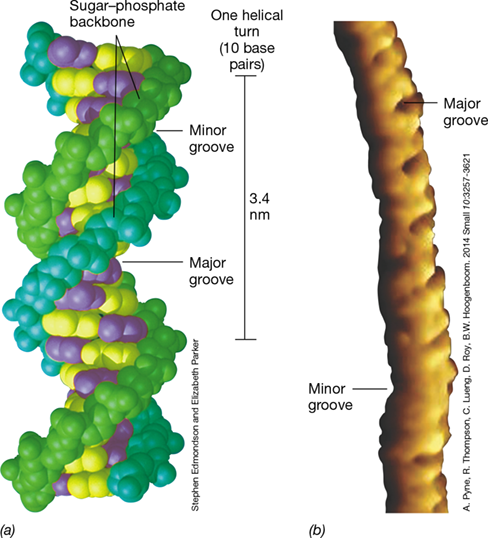
The complementary and antiparallel strands of DNA are wrapped around each other to form a double helix (Figure 6.3). The helix naturally forms two distinct grooves, the major groove and the minor groove. Most proteins that interact specifically with DNA bind in the major groove, where space is abundant. Because the double helix is a regular structure, some atoms of each base are always exposed in the major groove (and some in the minor groove). These key regions of the DNA that are important in interactions with proteins can be seen in Figure 6.3, and the atoms of the major groove that interact with proteins are highlighted in pink in Figure 6.1c.
Figure 6.3 Arrangement of the DNA double helix.

(a) A computer model of a short segment of DNA showing one of the sugar–phosphate backbones in blue-green and the other in light green. The pyrimidine bases are shown in purple and the purines in yellow. Note the locations of the major and minor grooves. One helical turn contains 10 base pairs. (b) Atomic force microscopy showing the biomolecular structure of a small piece of DNA. Note the locations of the major and minor grooves.
Size, Shape, and Supercoiling of DNA
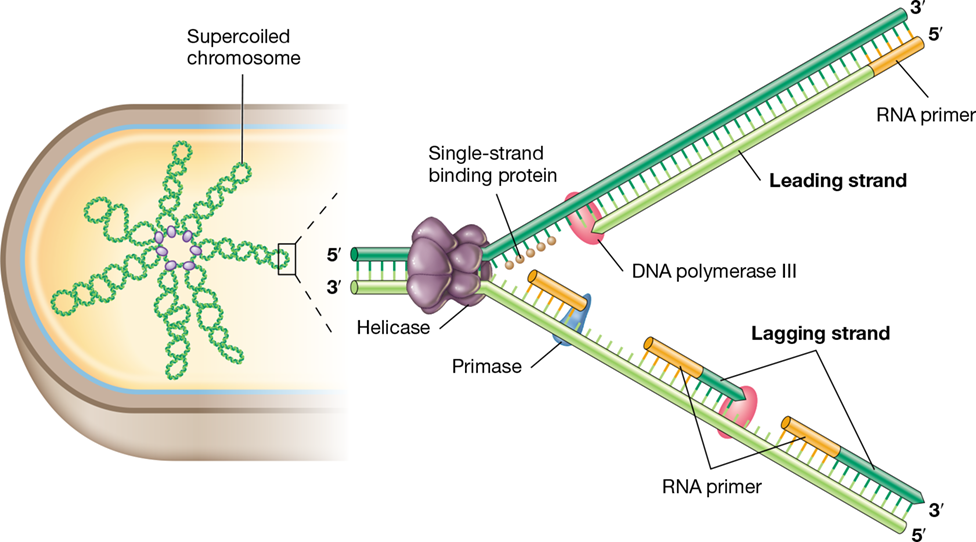
The size of a DNA molecule is expressed as its total number of nucleotide base pairs. Thus, a double-stranded DNA molecule consisting of 1000 bases is one kilobase pair (kbp) of DNA. The bacterium Escherichia coli has about 4640 kbp (4.64 megabase pairs, Mbp) of DNA in its genome. If this molecule were extended linearly it would be several hundred times longer than the cell itself. To accommodate their genome, cells of Bacteria and Archaea must compact the DNA, and this is done by the process of supercoiling (Figure 6.4).
Figure 6.4 Supercoiled DNA and DNA gyrase.

(a) Schematic showing the introduction of negative supercoiling into circular DNA by the activity of DNA gyrase (topoisomerase II), which makes double-strand breaks. (b) The double-stranded DNA in the bacterial chromosome is arranged not in one supercoil but in several supercoiled domains, as shown here. (c) Atomic force microscopy of the Escherichia coli nucleoid. (d) Simultaneous phase-contrast and fluorescence image of E. coli illustrating the location of the nucleoid within growing cells. Cells were treated with a fluorescent dye specific for DNA and the color was inverted to show the nucleoids as black.
Supercoils are inserted or removed in DNA by enzymes called topoisomerases. The activity of supercoiling puts the DNA molecule under torsion (Figure 6.4), and DNA can be supercoiled in either a positive or a negative manner. Negative supercoiling results when the DNA is twisted about its axis in the opposite sense from the right-handed double helix and is the form found in most cells. In the E. coli chromosome, more than 100 supercoiled domains exist, each stabilized by specific proteins bound to the DNA. Inserting supercoils into DNA requires energy from ATP, whereas releasing supercoils does not. In Bacteria and most Archaea, the topoisomerase DNA gyrase inserts negative supercoils into DNA by making double-strand breaks (Figure 6.4a). As we see in Chapters 4 and 17, some Archaea live at very high temperatures—above the boiling point in some cases. These species have chromosomes that are positively (instead of negatively) supercoiled, and this genomic feature helps to maintain DNA structure (that is, it prevents the two strands from melting apart) at such high temperatures (Section 17.13). Supercoiling is not a feature of eukaryotes since their genomic DNA is linear rather than circular. However, eukaryotic DNA must still be compacted, and this occurs when the DNA is highly wound around histone proteins.
Genes and the Steps in Biological Information Flow
Genetic information flow is a fundamental process in all cells and is the central dogma of molecular biology (Figure 6.1a and Figure 6.5). When genes are expressed, the genetic information encoded in DNA is transferred to ribonucleic acid (RNA). While several classes of RNA exist in cells, three main classes of RNA participate in protein synthesis. Messenger RNAs (mRNAs) are single-stranded molecules that carry the genetic information from DNA to the ribosome. Transfer RNAs (tRNAs) help convert the genetic information in the nucleotide sequences of RNA into a defined sequence of amino acids in proteins. Ribosomal RNAs (rRNAs) are important catalytic and structural components of the ribosome. The molecular processes of genetic information flow can be divided into three stages (Figure 6.5):
Figure 6.5 Synthesis of the three types of informational macromolecules in the processes of replication (DNA→DNA), transcription (DNA→RNA), and translation (RNA→protein).

Note that for any particular gene only one of the two strands of the DNA double helix is transcribed.
Replication. During replication, the DNA double helix is duplicated. Replication is catalyzed by the enzyme DNA polymerase.
Transcription. The transfer of genetic information from DNA to RNA is called transcription. Transcription is catalyzed by the enzyme RNA polymerase.
Translation. The formation of a polypeptide using the genetic information transferred to mRNA by DNA is a process that occurs on the ribosome.
Many different RNA molecules can be transcribed from a relatively short region of the long DNA molecule. In eukaryotes, each gene is transcribed to yield a single mRNA, whereas a single mRNA molecule may encode several different proteins in Bacteria and Archaea. However, a linear correspondence exists between the base sequence of a gene and the amino acid sequence of a polypeptide, and as we will see, each group of three bases on an mRNA molecule encodes a single amino acid (Section 6.9).
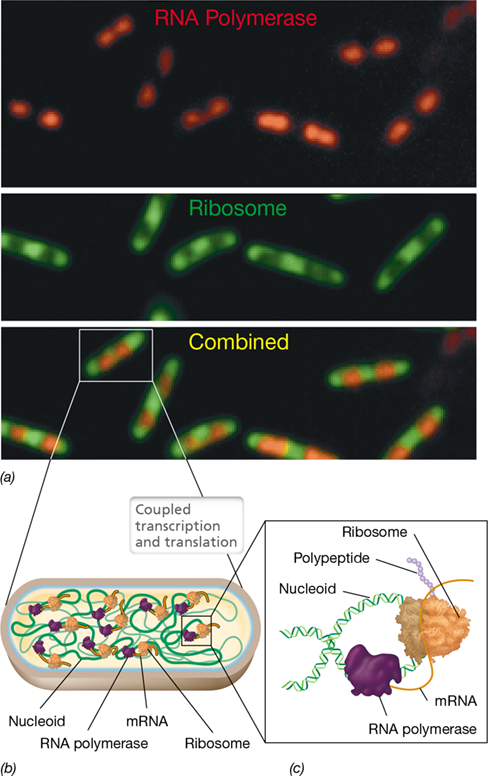
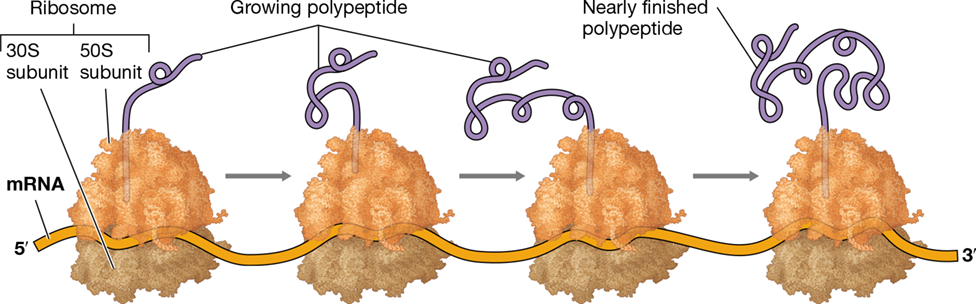
Eukaryotes differ from Bacteria and Archaea in that the first two steps of the central dogma, replication and transcription (Figures 6.1 and 6.5), occur in the nucleus. Because ribosomes are not present in the nucleus, mRNAs as well as other RNAs must be transported outside of the nucleus for translation. By contrast, in prokaryotic cells, mRNAs do not have to be exported from an organelle to be translated. Because of this fundamental difference, transcription and translation in Bacteria and Archaea can occur simultaneously in a process known as coupled transcription and translation (Figure 6.6). During this process, a ribosome initiates translation of an mRNA before RNA polymerase has finished synthesizing it. This allows rapidly growing cells to produce proteins at a maximal rate and also allows the cell to rapidly adapt to changes in growth conditions by quickly expressing the new protein sets required.
Figure 6.6 Coupled transcription and translation in prokaryotic cells.

(a) Fluorescence microscopy and protein tagging of actively growing Escherichia coli cells illustrating the position of RNA polymerases and ribosomes performing transcription and translation, respectively. The combined photo (bottom image) shows that transcription and translation are occurring concurrently in the cell. (b) Location of the nucleoid, RNA polymerases, mRNA, and ribosomes in the cell during coupled transcription and translation. (c) Schematic illustration in a single cell of a ribosome actively translating an mRNA as it is being synthesized by RNA polymerase.
While the central dogma of molecular biology (Figures 6.1a and 6.5) is invariant in cells, we will see later that some viruses (which are not cells, Section 1.4) violate the dogma in many interesting ways (Chapter 11). But for now we move on to consider the different genetic elements present in prokaryotic cells.
Check Your Understanding
What is a genome and what is it composed of? What is the central dogma of molecular biology?
Define the terms complementary and antiparallel as they pertain to DNA.
Why is supercoiling essential to a bacterial cell? What enzyme facilitates this process?
6.2 Genetic Elements: Chromosomes and Plasmids
Structures containing genetic material (DNA in cells but RNA in some viruses) are called genetic elements, and the main genetic element in prokaryotic cells is the chromosome. However, other genetic elements play important roles in microbes and these include virus genomes(discussed in Chapters 5 and 11), plasmids, organellar genomes, and transposable elements (Table 6.1). Most Bacteria and Archaea contain a single circular chromosome containing all (or most) of the organism’s genes. Although a single chromosome is the rule in prokaryotic cells, there are exceptions, as a few contain two or even three chromosomes. Eukaryotic genomes, by contrast, are composed of two or more chromosomes containing linear DNA. The genomes of viruses consist of either DNA or RNA and can be single- or double-stranded and either linear or circular.
Table 6.1 Kinds of genetic elements
Plasmids are circular or linear double-stranded DNA molecules that replicate separately from the chromosome and are typically much smaller than chromosomes. Transposable elements are sequences of DNA that are inserted into other DNA molecules but can move from one site on the DNA molecule to another, either within the same molecule or on a different DNA molecule. Chromosomes, plasmids, virus genomes, and any other type of DNA molecule may host a transposable element. Transposable elements are found in both prokaryotic and eukaryotic cells and play important roles in genetic variation (Section 9.11).
Chromosomal Gene Arrangements
Thousands of genomes from species of Bacteria and Archaea have been completely sequenced, thus revealing the number and location (the genetic map) of the genes they possess. The genetic map of the 4,639,675-bp (4.639 Mb) chromosome of a widely studied strain of Escherichia coli is presented in Figure 6.7, with only a few of the organism’s several thousand genes depicted. Analysis of the E. coli genome has revealed 4288 possible protein-encoding genes that account for 88% of the E. coli genome. Approximately 1% of the genome encodes tRNAs and rRNAs, and the remaining genes include regulatory sequences that may or may not be transcribed (but are not translated) and sequences that have other functions. The compact genomes of Bacteria and Archaea stand in contrast to the genomes of eukaryotes, which typically contain much more DNA than is needed to encode all the proteins required for cell function. This “extra” DNA in eukaryotes is present as intervening DNA between coding sequences (the intervening sequences are removed before translation) or as repetitive sequences, some of which are repeated hundreds or thousands of times (Chapter 10).
Figure 6.7 The chromosome of *Escherichia coli* strain K-12.

Map distances are given in 100 kilobases of DNA. The chromosome contains 4,639,675 base pairs and 4288 open reading frames (genes). Depending on the DNA strand, the locations of a few genes and operons are indicated. Replication (Figure 6.5 and see Figures 6.15 and 6.16) proceeds in both directions from the origin of DNA replication, oriC, indicated in red.
Genetic mapping of the genes encoding the enzymes that function in steps of the same biochemical pathway in E. coli has shown that these genes are sometimes clustered. On the genetic map in Figure 6.7, a few such clusters are shown (for example, the gal, trp, and his clusters); each of these groups is called an operon, which we will discuss in Section 6.5. In contrast to these, the genes for many other biochemical pathways in E. coli are not clustered. For example, genes for maltose degradation (mal genes, Figure 6.7) are scattered throughout the chromosome. In fact, analysis of the E. coli chromosome has shown that over 70% of the predicted or known transcriptional units are of only a single gene and that only 6% of operons have four or more genes. Thus operons, as efficient an arrangement of genes as they may be, appear to be the exception rather than the rule.
Plasmids
Many Bacteria and Archaea contain plasmids in addition to their chromosome(s). Most plasmids are nonessential since with rare exception they do not contain genes required for growth under all conditions. Thousands of different plasmids are known, and over 300 different plasmids have been isolated from strains of E. coli alone. Virtually all plasmids consist of double-stranded DNA and exist in the cytoplasm as free DNA. Most plasmids are circular, but many are linear and vary in size from approximately 1 kbp to more than 1 Mbp.
Typical plasmids are less than 5% of the size of the chromosome (Figure 6.8), and some bacteria contain several different plasmids. Moreover, different plasmids may be present in different copy number. For example, some plasmids may be present in only one or a few copies per cell, whereas others may be present in over 100 copies. Enzymes that replicate chromosomal DNA also replicate plasmids. Some of the genes encoded on a plasmid function to direct the initiation of plasmid replication and to partition replicated plasmids between daughter cells.
Figure 6.8 The bacterial chromosome and bacterial plasmids, as seen in the electron microscope.


The plasmids (arrows) are the circular structures and are much smaller than the main chromosomal DNA. The cell (large, tan structure) was broken gently so the DNA would remain intact.
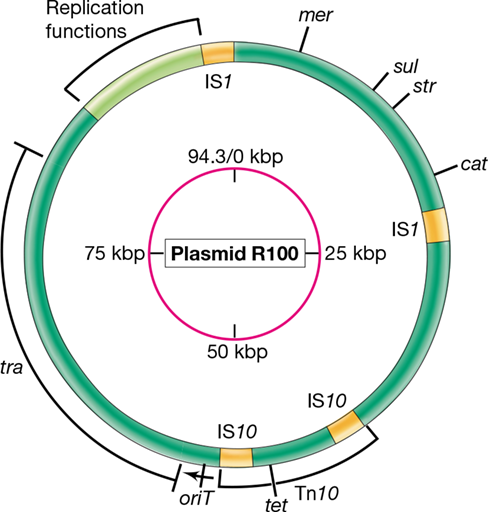
Although by definition plasmids do not encode functions essential to the host, plasmids may carry genes that profoundly influence host cell physiology; for example, plasmid genes may encode enzymes for some special metabolism that ensures survival under certain conditions. Among the most widespread and well-studied groups of plasmids are the resistance plasmids, called R plasmids, which confer resistance to antibiotics or other growth inhibitors. The resistance genes encode proteins that either inactivate the antibiotic or protect the cell in some other way (Section 8.11), and several antibiotic resistance genes can be encoded on a single R plasmid. Plasmid R100 (Figure 6.9), for example, encodes resistance to sulfonamides, streptomycin, spectinomycin, fusidic acid, chloramphenicol, and tetracycline, as well as the toxic heavy metal mercury. Pathogenic bacteria resistant to antibiotics are of considerable medical significance, and their increasing incidence is correlated with the increasing use of antibiotics for treating infectious diseases in humans and animals (Chapter 28).
Figure 6.9 Genetic map of the resistance plasmid R100.

The inner circle shows the size in kilobase pairs. The outer circle shows the location of major antibiotic resistance genes and other key functions: mer, mercuric ion resistance; sul, sulfonamide resistance; str, streptomycin resistance; cat, chloramphenicol resistance; tet, tetracycline resistance; oriT, origin of conjugative transfer; tra, transfer functions. The locations of insertion sequences (IS) and the transposon Tn10 are also shown. Genes for plasmid replication are found in the region from 88 to 92 kbp.
Pathogenic bacteria express a variety of plasmid-encoded virulence factors that assist them in establishing infections. For example, the ability of a pathogen to attach to and colonize specific host tissues and to produce toxins, enzymes, and other invasive molecules that damage the host are sometimes plasmid encoded. Some bacteria also produce proteins called bacteriocins that inhibit or kill closely related species of bacteria (or even different strains of the same species of bacteria), and the genes encoding these bacteriocins and other proteins that protect the producing organism are typically found on plasmids.
In a few cases plasmids encode properties that are fundamental to the ecology of the bacterium. For example, the ability of the soil bacterium Rhizobium to form nitrogen-fixing nodules on the roots of plants (Section 23.4) requires certain functions encoded by plasmids. Other plasmids confer special metabolic properties. For example, the ability to degrade hydrocarbons or toxic pollutants, such as polychlorinated biphenyls (PCBs) and herbicides or other pesticides, is often plasmid encoded. In addition, plasmids play a crucial role in the horizontal gene transfer process called conjugation that we consider in detail later (Chapter 9).
Check Your Understanding
Approximately how large is the Escherichia coli genome in base pairs? How many genes does it contain?
What properties does an R plasmid confer on its host cell?
II: Copying the Genetic Blueprint: DNA Replication
II: Copying the Genetic Blueprint: DNA Replication
II Copying the Genetic Blueprint: DNA Replication
DNA replication must occur in a cell before it can divide. This process has a unique dimension in prokaryotic cells because their circular chromosome allows bidirectional DNA synthesis.
DNA replication is necessary for cells to divide, whether to reproduce new organisms, as in unicellular microorganisms, or to produce new cells as part of a multicellular organism. To successfully transmit genetic information from a mother cell to an identical daughter cell, DNA replication must be extremely accurate. We review the basic principles of DNA replication here as a prelude to focusing on the process as it o ccurs in prokaryotic cells.
6.3 Templates, Enzymes, and the Replication Fork
DNA exists in cells as a double helix of two complementary strands (Figures 6.2 and 6.5), and if the helix is opened up, a new strand can be synthesized as the complement of each parental strand. As shown in **Figure 6.10*a***, replication is thus a semiconservative process; in semiconservative replication, the two resulting double helices each consist of one new strand and one parental strand. The DNA strand that is used to make a complementary daughter strand is called the template strand, and in DNA replication, each parental strand is a template for one newly synthesized strand (Figure 6.10a). The precursor of each new nucleotide in the DNA strand is a deoxynucleoside 5′-triphosphate. During insertion of this molecule, the two terminal phosphates are removed and the remaining phosphate is bonded to a deoxyribose of the growing chain (Figure 6.10b). This addition of the incoming nucleotide requires the presence of a free hydroxyl group, which is available only at the 3′ end of the molecule. This leads to the important principle that DNA replication always proceeds from the 5′ end to the 3′ end, the 5′-phosphate of the incoming nucleotide being attached to the 3′-hydroxyl of the previously added nucleotide (Figure 6.10b).
Figure 6.10 Overview of DNA replication.

(a) In cells, DNA replication is a semiconservative process. Note that the new double helices each contain one new daughter strand (shown in red) and one parental strand. (b) Extension of a DNA chain by adding a deoxyribonucleoside triphosphate at the 3′ end. Growth proceeds from the 5′-phosphate to the 3′-hydroxyl end. DNA polymerase catalyzes the reaction. The four precursors are deoxythymidine triphosphate (dTTP), deoxyadenosine triphosphate (dATP), deoxyguanosine triphosphate (dGTP), and deoxycytidine triphosphate (dCTP). Upon nucleotide insertion, the two terminal phosphates of the triphosphate are split off as pyrophosphate (PPi). Thus, two energy-rich phosphate bonds are consumed when adding each nucleotide.
Replication Enzymes
Enzymes that catalyze the polymerization of deoxynucleotides are called DNA polymerases (abbreviated DNA Pol), and there are five different enzymes in Escherichia coli, DNA Pol I–V. DNA Pol III is the primary enzyme for replicating chromosomal DNA, although DNA Pol I also plays a lesser role. The other DNA polymerases function to repair damaged DNA (Section 9.4). DNA Pol enzymes are just some of the many enzymes that are required for DNA replication (Table 6.2).
Table 6.2 Major enzymes that participate in DNA replication in ***Bacteria***

All DNA polymerases synthesize DNA in the 5′→3′ direction, but none of them can initiate a new chain de novo; they can only add a nucleotide onto a preexisting 3′-OH group. Thus, in order to start a new DNA chain, a primer, a nucleic acid molecule to which DNA polymerase can attach the first nucleotide, is required, and this primer is a short stretch of RNA rather than DNA (Figure 6.11). When the DNA helix is first opened, the enzyme primase makes this RNA primer, synthesizing a short stretch (11–12 nucleotides) of RNA complementary in base pairing to the template-strand DNA. At the growing end of this RNA primer is a 3′-OH group to which DNA polymerase adds the first deoxyribonucleotide. Continued extension of the molecule thus occurs as DNA rather than RNA (Figure 6.11), and the primer is eventually removed and replaced with DNA (as described shortly).
Figure 6.11 The RNA primer.

Structure of the RNA–DNA hybrid formed during initiation of DNA synthesis. Orange depicts the RNA primer.
Initiation of DNA Synthesis
Before replication can begin, the double helix must be unwound to expose the template strands at the so-called replication fork. The enzyme DNA helicase unwinds the double helix (using energy from ATP) and exposes a short single-stranded region (Figure 6.12). Helicase moves along the DNA and separates the strands just in advance of the replication fork. The single-stranded region is immediately covered with copies of single-strand binding protein to stabilize the single-stranded DNA and prevent the double helix from re-forming. DNA synthesis begins at a single site on the chromosome, the origin of replication (oriC), where the protein DnaA (Table 6.2) binds and opens up the double helix. Next to assemble is helicase (DnaB), which is helped onto the DNA by a loader protein (DnaC) (Figure 6.12b). Finally, primase and DNA polymerase enzymes are loaded onto the DNA behind the helicase and initiation of DNA replication begins. As replication proceeds, the replication fork appears to move along the DNA (Figure 6.12a).
Mastering Microbiology
Microbiology Animation: DNA Replication: Replication Proteins
Figure 6.12 DNA helicase unwinding a double helix.

(a) In this schematic, the helicase is seen pulling the two antiparallel strands of DNA apart beginning from the right and moving to the left. (b) A three-dimensional model of the helicase (DnaB) along with its loader protein (DnaC, yellow and orange) based on cryo-electron microscopy.
Ernesto Arias-Palomo and James Berger, Johns Hopkins School of Medicine
Leading and Lagging Strands and the Replication Process
Figure 6.13 depicts DNA replication at the replication fork. Recall that replication always proceeds from 5′ to 3′ (5′→3′), always adding a new nucleotide to the 3′-OH of the growing chain. On the strand growing from the 5′-PO42− to the 3′-OH, called the leading strand, DNA synthesis occurs continuously because there is always a free 3′-OH at the replication fork to which a new nucleotide can be added; the leading strand must therefore be primed only once. By contrast, on the opposite strand, called the lagging strand, DNA synthesis occurs discontinuously because there is no 3′-OH at the replication fork to which a new nucleotide can attach; on this strand, primase must synthesize multiple RNA primers in order to provide free 3′-OH groups for DNA Pol III (Figure 6.13). As a result, the lagging strand forms from several short DNA fragments that are combined later to yield a continuous strand of DNA.
Figure 6.13 Events at the DNA replication fork on the nucleoid.

Note the polarity and antiparallel nature of the DNA strands. Helicase unwinds the DNA while primase adds the RNA primer. For the steps in introducing and removing supercoils from DNA, see Figure 6.4. Further events in DNA synthesis including sealing replicated fragments are shown in Figure 6.14.
After synthesizing the RNA primer, primase is replaced by DNA Pol III. This enzyme complex (Table 6.2) is held on the DNA by a “sliding clamp,” which encircles and slides along the single template strands of DNA. Consequently, the replication fork contains two polymerase core enzymes and two sliding clamps, one set for each strand. After assembly on the lagging strand, the elongation activity of DNA Pol III adds deoxyribonucleotides sequentially until it reaches previously synthesized DNA (Figure 6.14); at this point, activity of DNA Pol III stops.
Figure 6.14 Sealing two fragments on the lagging strand.

Unlike the leading strand, where synthesis occurs in a continuous fashion, on the lagging strand, DNA fragments need to be sealed to form the intact DNA strand. (a) DNA polymerase III is synthesizing DNA in the 5′→3′ direction toward the RNA primer of a previously synthesized fragment on the lagging strand. (b) On reaching the fragment, DNA polymerase III leaves and is replaced by DNA polymerase I. (c) DNA polymerase I continues synthesizing DNA while removing the RNA primer from the previous fragment, and DNA ligase replaces DNA polymerase I after the primer has been removed. (d) DNA ligase seals the two fragments together. (e) The final product, complementary and antiparallel double-stranded DNA.
To complete DNA synthesis, DNA Pol I catalyzes two different reactions. Besides synthesizing DNA, Pol I has a 5′→3′ exonuclease activity that removes the RNA primer (Figure 6.14). When the primer has been excised and replaced with DNA, Pol I is released. The very last phosphodiester bond in replicating DNA is made by DNA ligase. This enzyme seals nicks in DNAs that have an adjacent 5′-PO42− and 3′-OH (something that DNA Pol I and Pol III are unable to do), and along with DNA Pol I, it also participates in DNA repair. DNA ligase is also important for sealing genetically manipulated DNA during molecular cloning ( Section 12.2).
We now put DNA synthesis in the context of Archaea and Bacteria to see how replication events occur around the covalently closed and circular chromosomes typical of these organisms.
Check Your Understanding
What is the difference between a template strand and a daughter strand of DNA?
To which end (5′ or 3′) of a newly synthesized strand of DNA does DNA polymerase add a nucleotide?
In DNA replication, what is the primer composed of and why are there leading and lagging strands?
What are the functions of DNA Pol I and III, DNA helicase, and DNA ligase?
6.4 Bidirectional Replication, the Replisome, and Proofreading
6.4 Bidirectional Replication, the Replisome, and Proofreading
6.4 Bidirectional Replication, the Replisome, and Proofreading
The circular nature of the bacterial and archaeal chromosome accelerates the genomic replication process. In Escherichia coli—and probably in all cells with circular chromosomes—DNA replication occurs bidirectionally from the origin of replication. There are thus two replication forks on each chromosome, each moving in opposite directions. In circular DNA, bidirectional replication leads to the formation in the replicating molecules of characteristic shapes (so-called “theta structures”) as synthesis proceeds in both a leading and a lagging fashion on each template strand (Figure 6.15). In an actively growing cell of E. coli, DNA Pol III adds nucleotides at the rate of about 1000 per second; hence, replication of the entire chromosome takes about 40 min.
Figure 6.15 Replication of circular DNA: the theta structure.

In circular DNA, bidirectional replication from an origin forms an intermediate structure resembling the Greek letter theta (θ). The blowup shows dual replication forks in the circular chromosome. In Escherichia coli, the origin of replication is recognized by the DnaA protein and the terminus of replication is recognized by the Tus protein. Note that DNA synthesis is occurring in both a leading and a lagging manner on each of the new daughter strands until the replication forks hit the terminus. Compare this figure with the illustration of the replisome in Figure 6.16.
The Replisome
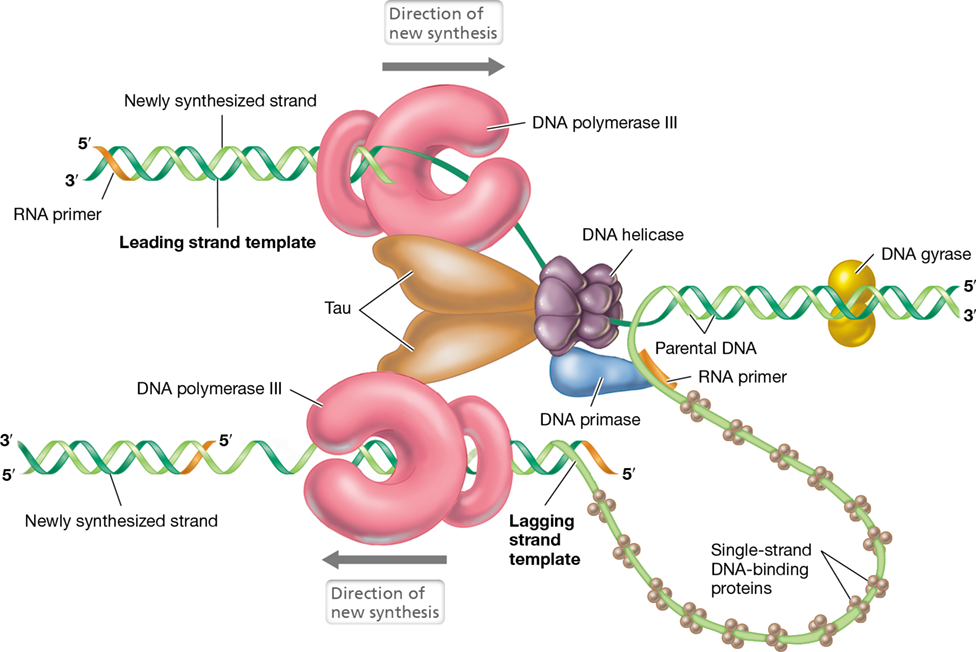
Figure 6.13 shows the enzymes that participate in replication, and from such a schematic it may appear that the enzymes are working independently. However, this is not the case. Instead, replication proteins aggregate to form a large replication complex called the replisome (Figure 6.16). The lagging strand of DNA actually loops out to allow the replisome to move smoothly along both strands, the complex literally pulling the DNA template through it as replication proceeds. In addition to the replisome, helicase and primase form their own subcomplex within the replisome called the primosome. This close association facilitates the sequential activities of these two enzymes during the replication process (Figure 6.16). Table 6.2 summarizes the functions of proteins essential for DNA replication in Bacteria.
Figure 6.16 The replisome.
The replisome consists of two copies of DNA polymerase III and DNA gyrase, plus helicase and primase (together forming the primosome), and many copies of single-strand DNA-binding protein. The Tau subunits hold the two DNA polymerase assemblies and helicase together. Just upstream of the rest of the replisome, DNA gyrase removes supercoils in the DNA to be replicated. Note that the two polymerases are replicating the two individual strands of DNA in opposite directions. Consequently, the lagging-strand template loops around so that the whole replisome moves in the same direction along the chromosome.
Eventually the activity of the replisome is finished, and this is signaled when the replication forks collide at the terminus of replication, a site located on the opposite side of the chromosome from the origin. In the terminus region are several DNA sequences called Ter sites that are recognized by a protein called Tus, whose function is to block progress of the replication forks. When replication of the circular chromosome is complete, the two circular molecules are linked together, much like the links of a chain. After replication, the DNA is partitioned so that each daughter cell receives a copy of the chromosome; DNA partitioning is facilitated by FtsZ, a protein that orchestrates several key events in the cell division process (Chapter 8).
Fidelity of DNA Replication: Proofreading
DNA replication occurs with a remarkably low error rate. Nevertheless, when errors do occur, a mechanism exists to detect and correct them. Errors in DNA replication introduce mutations, changes in DNA sequence. Mutation rates in cells are extremely low, between 10−8 and 10−11 errors per base pair inserted. This accuracy is achieved because DNA polymerases get two chances to incorporate the correct base at any given site. The first chance comes when DNA Pol III inserts bases according to the base-pairing rules (Figure 6.1c). The second chance comes when a process called proofreading takes place (Figure 6.17).
Figure 6.17 Proofreading by the 3′→5′ exonuclease activity of DNA polymerase III.

(a) A mismatch in base pairing at the terminal base pair leads to a distortion in DNA topology that causes the polymerase to pause briefly. This signals the proofreading activity to cut out the mismatched nucleotide, after which the correct base is inserted by the polymerase activity. (b) Cryo-electron micrograph of Escherichia coli DNA polymerase III bound to DNA (green strands) in proofreading mode. Image credit: Fernandez-Leiro, R., Conrad, J., Scheres, S.H.W., and Lamers, M.H., EMDataBank: EMD-4141.
During replication, if an incorrect base has been inserted, a mismatch in base pairing (Figure 6.1c) occurs. Both DNA Pol I and Pol III possess a 3′→5′ exonuclease activity that can remove such mismatched nucleotides. The polymerase detects the error because incorrect base pairing causes a slight distortion in the topology of the double helix. After the removal of a mismatched nucleotide, the polymerase then gets a second chance to insert the correct nucleotide (Figure 6.17; part b shows the polymerase working in “proofread mode”). With the extremely low error rate of DNA polymerases, the chance of inserting the wrong base at the same site twice is vanishingly small. Exonuclease proofreading occurs in Bacteria and Archaea, eukaryotes, and viral DNA replication systems.
We now move on from replicating genes to consider gene expression as a prelude to examining synthesis of the proteins encoded by the transcribed genes.
Check Your Understanding
What is the replisome and what are its components?
How are errors in DNA replication kept extremely low?
III RNA Synthesis: Transcription
Transcription transforms the genetic information inscribed in DNA into RNA, a form of nucleic acid that the cell’s protein-synthesizing machinery can recognize and process into the many different types of proteins that a given cell needs.
Transcription—RNA synthesis off of a DNA template—yields three main forms of RNA: messenger (mRNA), transfer (tRNA), and ribosomal (rRNA) (Section 6.1). Several other minor classes of RNA exist, but most of these function in regulation (Chapter 7). RNA is both a genetic and a functional molecule. At the genetic level, mRNA encodes genetic information from the genome for the synthesis of proteins and carries this information to the ribosome. In contrast, rRNAs play both a structural and a functional role in ribosomes, while tRNAs function as the carriers of amino acids to the ribosome for protein synthesis.
There are two key differences in the chemistry of RNA and DNA: (1) RNA contains ribose instead of deoxyribose; and (2) RNA contains uracil instead of thymine. The change from deoxyribose to ribose dramatically affects the chemistry of a nucleic acid, and enzymes that act on DNA typically have no effect on RNA, and vice versa. However, the change from thymine to uracil does not affect base pairing, as these two bases pair with adenine equally well.
With the exception of a few viruses that contain double-stranded RNA genomes (Chapter 11), RNA is a single-stranded molecule. However, the primary structure (sequence of nucleotides) of some RNAs allows them to fold and exploit complementary base pairing. The term secondary structure refers to this folding, and the functional role an RNA plays in the cell may depend critically on its secondary structure. For example, messenger RNAs, which are typically unfolded, exist in Bacteria (and Archaea) for only a few minutes before enzymes called ribonucleases degrade them. By contrast, rRNAs and tRNAs (referred to as stable RNAs) are long-lived because their secondary structures prevent ribonuclease attack. The rapid turnover of mRNAs in Bacteria and Archaea allows them to quickly adapt to changing environmental conditions and halt the translation of mRNAs whose products are no longer needed.
We begin with transcription in Bacteria and contrast this in the following section with transcriptional events in Archaea and Eukarya.
6.5 Transcription in *Bacteria*
Transcription is catalyzed by the enzyme RNA polymerase. Like DNA polymerase, RNA polymerase forms phosphodiester bonds but between the ribonucleotides rather than deoxyribonucleotides (Figure 6.1b). Polymerization is driven by energy released from the hydrolysis of two energy-rich phosphate bonds of the incoming ribonucleoside triphosphates. The mechanism of RNA synthesis is thus quite similar to that of DNA synthesis (Figure 6.10b): During elongation of an RNA chain, ribonucleoside triphosphates are added to the 3′-OH of the ribose of the preceding nucleotide. Thus chain growth is 5′→3′ just as in DNA synthesis, and the newly synthesized strand of RNA runs antiparallel to the DNA template strand it was transcribed from. A summary of bacterial transcription is illustrated in Figure 6.18.
Figure 6.18 Transcription.

(a) Steps in RNA synthesis. The initiation site (promoter) and termination site are specific nucleotide sequences on the DNA. Sigma binds to the promoter region and then RNA core polymerase binds to begin transcription. RNA polymerase moves down the DNA chain, temporarily opening the double helix (creating a transcription bubble) and transcribing one of the DNA strands. (b) Electron micrograph illustrates transcription by multiple polymerases along a gene on the Escherichia coli chromosome. Transcription is proceeding from left to right, with the shorter transcripts on the left becoming longer as transcription proceeds.
Mastering Microbiology
RNA polymerase uses DNA as a template, but for any given gene, only one of the two strands is transcribed. Unlike DNA polymerase, RNA polymerase can initiate new RNA on its own; no priming is necessary as it is for DNA synthesis (Figure 6.11). Transcription continues until specific sequences called transcription terminators are reached, but unlike DNA replication, which copies the entire genome, transcription occurs on much smaller units of DNA, often as little as a single gene. This system allows the cell to transcribe different genes at different frequencies, depending on the needs of the cell for different proteins. Said a different way, gene expression is a highly regulated process. Transcriptional regulation can occur in bacterial and archaeal cells in many different ways, but the different mechanisms have a common outcome: Cell resources are conserved and cell fitness enhanced (Chapter 7).
RNA Polymerases and the Promoter Sequence
The RNA polymerase from Bacteria, which has the simplest structure and about which most is known, consists of five different subunits, designated β, β′, α, ω (omega), and σ (sigma), with α present in two copies (Figure 6.19). The subunits form an enzyme complex called the RNA polymerase holoenzyme. Sigma is not as tightly bound as the other subunits and easily dissociates to yield the RNA polymerase core enzyme, α2ββ′ω. The core enzyme alone synthesizes RNA, and sigma functions only to recognize the appropriate site on the DNA to initiate transcription (sigma dissociates from the holoenzyme once a short sequence of RNA has been formed; Figure 6.18).
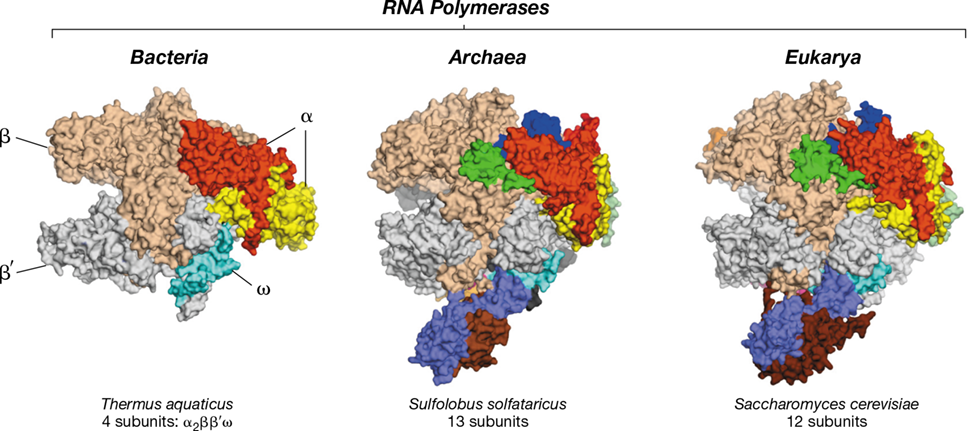
Figure 6.19 RNA polymerase from the three domains.

Surface representation of multi-subunit cellular RNA polymerase structures from Bacteria (left, Thermus aquaticus core enzyme), Archaea (center, Sulfolobus solfataricus), and Eukarya (right, Saccharomyces cerevisiae RNA Pol II). Orthologous subunits are depicted by the same color. A unique subunit in the S. solfataricus RNA polymerase is not shown in this view.
To begin transcription, RNA polymerase must first recognize initiation sites on the DNA; these sites are called promoters. In Bacteria, promoters are recognized by sigma (Figure 6.20). Once RNA polymerase has bound to a promoter, transcription can proceed (Figure 6.18). In this process, the DNA helix at the promoter site is opened up by RNA polymerase, and as the polymerase moves, it unwinds the DNA in short segments to expose template DNA, creating a transcription bubble (Figure 6.18). Thus, an enzyme with the activity of DNA helicase is not needed. Because some genes reside on one strand of DNA while other genes reside on the other strand of DNA, promoters are present on both strands; as a result, transcription occurs in opposite directions on the two different strands of DNA.
Figure 6.20 The interaction of RNA polymerase with a bacterial promoter.

(a) Cryo-electron micrograph of Escherichia coli RNA polymerase with sigma factor (together constituting the holoenzyme) bound to closed promoter region. Image credit: Glyde, R., Ye, F.Z., Darbari, V.C., Zhang, N., Buck, M., and Zhang, X.D. EMDataBank: EMD-3695. (b) Six different promoter sequences identified in the E. coli genome. The contacts of the RNA polymerase and sigma factor with the −35 region and the Pribnow box (−10 sequence) are shown. Transcription begins at a unique base just downstream from the Pribnow box. Below the actual sequences at the −35 and Pribnow box regions are consensus sequences derived from comparing many promoters. Although the consensus sequences are not identical, they constitute a sufficiently similar pattern recognized through contacts between the bases in the DNA and specific domains of the sigma factor. Note that although sigma recognizes the promoter sequences on the 5′→3′ (dark green) strand of DNA, the RNA polymerase core enzyme will actually transcribe the light green strand (that runs 3′→5′) because core enzyme synthesizes only in a 5′→3′ direction (Figure 6.13).
Sigma Factors, Consensus Sequences, and Transcriptional Termination
Promoters are specific DNA sequences; Figure 6.20b shows the sequence of several promoters from Escherichia coli. All of these sequences are recognized by the same E. coli sigma factor called σ70 (the superscript 70 indicates the size of this protein in kilodaltons, a measure of atomic mass). Although these sequences are not identical, sigma recognizes two highly conserved regions within the promoter. These conserved sequences are upstream of (prior to) the transcription start site. One is 10 bases upstream, the −10 region, or Pribnow box. Although promoter sequences differ slightly, comparison of many −10 regions gives a consensus sequence of TATAAT. The second conserved region is about 35 bases upstream of the start site and its consensus sequence is TTGACA (Figure 6.20). In E. coli, promoters that conform most closely to the consensus sequence are more effective in binding RNA polymerase. Such promoters are called strong promoters and are very useful in genetic engineering, where it is desirable to have only one or a restricted set of genes heavily transcribed in order to produce more of a particular protein product (Chapter 12).
While most genes in E. coli require σ70 for transcription, several alternative sigma factors exist that recognize different consensus sequences (Table 6.3). Each alternative sigma factor is specific for a group of genes required under special circumstances, and thus the presence or absence of a specific sigma factor is a mechanism for regulating gene expression (Chapter 7). That is, by changing the rate of either synthesis or degradation of a particular sigma factor, the cell can control the transcription of entire gene families.
Table 6.3 Sigma factors in *Escherichia coli*
aSuperscript number indicates size of protein in kilodaltons. Many factors also have other names, for example, σ70 is also called σD. See Figure 6.20 for a description of how the recognition (consensus) sequence is recognized by the sigma factor.
Units of Transcription and Polycistronic mRNA
Genetic information is organized into transcriptional units, segments of DNA that are transcribed into a single RNA molecule bounded by their initiation and termination sites. Some transcriptional units contain RNA transcribed from a single gene, whereas others are formed from two or more genes (cotranscribed genes). Most genes encode proteins, but others encode nontranslated RNAs, such as ribosomal or transfer RNAs. For example, prokaryotic cells produce three size classes of rRNA: 16S, 23S, and 5S (the S refers to Svedberg units, a sedimentation coefficient that provides a measure of particle size based on sedimentation rate), and their genes are cotranscribed to form a single transcriptional unit that also includes a tRNA (Figure 6.21). This transcriptional unit is subsequently “processed” by proteins that cut them to form the individual rRNAs or tRNAs.
Figure 6.21 A ribosomal rRNA transcription unit from ***Bacteria*** and its subsequent processing.

In Bacteria, all rRNA transcription units have the genes in the order 16S rRNA, 23S rRNA, and 5S rRNA (shown approximately to scale). In this particular transcriptional unit, the spacer between the 16S and 23S rRNA genes contains a tRNA gene. In other transcription units this region may contain more than one tRNA gene. Often one or more tRNA genes also follow the 5S rRNA gene and are cotranscribed. Escherichia coli contains seven rRNA transcription units.
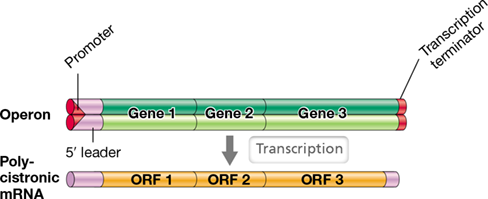
As we have previously considered, genes that encode several enzymes of a particular metabolic pathway in prokaryotic cells, for example the biosynthesis of a particular amino acid, are often clustered together in an operon (Section 6.2). An operon is transcribed to form a single mRNA that encodes several different proteins and is regulated as a unit. Assembling genes for the same biochemical pathway or genes needed under the same conditions into an operon allows for their coordinate expression. During transcription, RNA polymerase proceeds through the operon and transcribes the entire set of genes into a single mRNA called a polycistronic mRNA (Figure 6.22). Polycistronic mRNAs contain multiple open reading frames, portions of the mRNA that actually encode amino acids (Section 6.9). When this mRNA is translated, several polypeptides are synthesized sequentially by the same ribosome. Bacterial and archaeal genomes can also possess regulons, which are multiple operons that encode genes whose products are needed under the same conditions. We discuss the regulation of operons and the coordinated expression of regulons in Chapter 7.
Figure 6.22 Operon and polycistronic mRNA structure.

Note that a single promoter controls the three genes within the operon and that the polycistronic mRNA molecule contains an open reading frame (ORF) corresponding to each gene.
Termination of Transcription
In a growing bacterial cell, only those genes that need to be expressed are usually transcribed; therefore, it is critical that transcription end at the correct position. Termination of transcription is governed by specific base sequences on the DNA. In Bacteria a common termination signal is a GC-rich sequence containing an inverted repeat with a central nonrepeating segment. When such a DNA sequence is transcribed, the RNA forms a stem–loop structure by intra-strand base pairing (Figure 6.23). Coding sequences for stem–loops followed by a run of adenines in the DNA template (which yields a run of uridines in the RNA) are strong transcription terminators because a stretch of U–A base pairs are formed that hold the RNA and DNA together. However, this structure is very weak since U–A base pairs have only two hydrogen bonds rather than the three that form in G–C pairs (Figure 6.1c). Thus, RNA polymerase pauses at the stem–loop, and the DNA and RNA dissociate at the run of uridines, terminating transcription.
Figure 6.23 Inverted repeats and transcription termination.

(a) Inverted repeats in transcribed DNA form a stem–loop structure in the RNA that terminates transcription when followed by a run of uracils. (b) Schematic indicating the formation of the terminator stem–loop in the RNA within the RNA polymerase.
A second mechanism for terminating transcription is catalyzed by the terminator protein Rho. Rho does not bind to RNA polymerase or to the DNA but binds tightly to RNA and moves down the chain toward the RNA polymerase–DNA complex. Once RNA polymerase has paused at a Rho-dependent termination site (a specific nucleotide sequence on the DNA template), Rho causes both the RNA and RNA polymerase to be released from the DNA, thus terminating transcription.
Now that we have grasped the essentials of transcription in Bacteria, we turn our attention to this crucial cell process in Archaea and Eukarya, where the phylogenetic connection between these two domains (Section 1.15) will be apparent in their mechanisms of transcription.
Check Your Understanding
What enzyme catalyzes transcription? What is a promoter, and what protein recognizes promoters in Bacteria?
What is the role of messenger RNA (mRNA)? What are the other two classes of RNA?
How does polycistronic mRNA allow for gene families to be controlled as a group?
What type of structures lead to transcription termination?
6.6 Transcription in *Archaea* and *Eukarya*
Here we discuss key elements of transcription in Archaea and Eukarya that differ from those of Bacteria. Although in both Archaea and Eukarya the overall flow of genetic information is the same as in Bacteria, some details differ, and in eukaryotic cells the presence of the nucleus complicates the routing of genetic information. Many of the details of transcription (and translation) in Archaea resemble those in Eukarya more closely than Bacteria. However, Archaea also share some transcriptional similarities with Bacteria, such as operons. We begin our discussion at center stage with a consideration of RNA polymerase.
Archaeal and Eukaryotic RNA Polymerases, Promoters, and Terminators
Archaeal and eukaryotic RNA polymerases are similar and more complex than those of Bacteria. Archaea contain only a single RNA polymerase while eukaryotes have three. The archaeal RNA polymerase most closely resembles eukaryotic RNA polymerase II and is composed of 11–13 subunits, depending on the species (eukaryotic RNA polymerase II has 12 or more subunits). These contrast with the comparatively simple four-subunit RNA polymerase core enzyme of Bacteria (Figure 6.19).
We learned the importance of the promoter and its recognition sequences to the overall process of transcription in Section 6.5. The most important recognition sequence in archaeal and eukaryotic promoters is the 6- to 8-base-pair “TATA” box, located 18–27 nucleotides upstream of the transcriptional start site (Figure 6.24). The TATA box is recognized by the TATA-binding protein (TBP), one of the many transcription factors present in Archaea and eukaryotes. Upstream of the TATA box is the B recognition element (BRE) sequence that is recognized by archaeal transcription factor B (TFB). In addition, a specific initiator element sequence is located at the start of transcription. Once TBP has bound to the TATA box and TFB has bound to the BRE, then archaeal RNA polymerase can bind and initiate transcription. This process is similar in eukaryotes except that several additional transcription factors are required.
Figure 6.24 Promoter architecture and transcription in ****Archaea****.

(a) Three promoter elements are critical for promoter recognition in Archaea: the initiator element (INIT), the TATA box, and the B recognition element (BRE). The TATA-binding protein (TBP) binds the TATA box; transcription factor B (TFB) binds to both BRE and INIT. Once both TBP and TFB are in place, RNA polymerase binds. Note that the colors of the different transcription factors match those in the molecular model shown to the right. (b) Surface representation of the archaeal pre-initiation complex (with TBP and TFB) and including transcription factor E (TFE). TFE is an optional transcription factor frequently associated with the archaeal pre-initiation complex.
Less is known about transcription termination in Archaea than in Bacteria, although some archaeal genes have inverted repeats followed by an AT-rich sequence similar to those present in many bacterial transcription terminators (Section 6.5). One other type of suspected transcription terminator lacks inverted repeats but contains repeated runs of thymines. While no Rho-like proteins (Section 6.5) have been found in either Archaea or Eukarya, a separate transcriptional termination protein has been identified in Euryarchaeota (a major phylum of Archaea) called Eta. Eta binds to DNA upstream of the transcription bubble, and when it collides with the archaeal RNA polymerase, it bumps the polymerase off of the DNA. In eukaryotes, the termination process differs depending on the RNA polymerase and often requires a specific termination factor protein.
RNA Processing in Eukaryotes and Intervening Sequences in *Archaea*
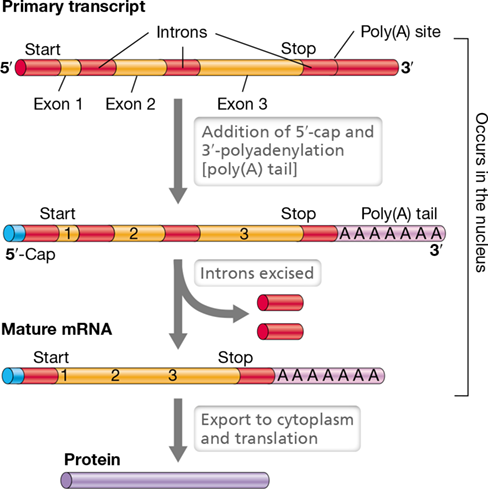
In contrast to Bacteria, Eukarya contain many genes that are split into two or more coding regions separated by noncoding regions. The coding sequences are called exons while the intervening noncoding regions are called introns. Thus, the transcripts from exons typically require alterations—known as RNA processing—to form mature RNAs suitable for translation. The term primary transcript refers to the RNA molecule that is originally transcribed before the introns are removed to form the mature mRNA containing only exons. The process by which introns are removed and exons are joined is called splicing (Figure 6.25).
Figure 6.25 Activity of the spliceosome.

Removal of an intron from the primary transcript of a protein-coding gene in a eukaryote. (a) A primary transcript containing a single intron. The sequence GU is conserved at the 5′ splice site, and AG is conserved at the 3′ splice site. There is also an interior A that serves as a branch point. (b) Several small ribonucleoprotein particles (shown in purple) assemble on the RNA to form a spliceosome. Each of these particles contains distinct small RNA molecules that take part in the splicing mechanism. (c) The 5′ splice site has been cut with the simultaneous formation of a branch point. (d) The 3′ splice site has been cut and the two exons have been joined. Note that overall, two phosphodiester bonds were broken, but two others were formed. (e) The final products are the joined exons (the mRNA) and the released intron.
Mastering Microbiology
Art Activity: Figure 6.26 Processing of the primary transcript into mature mRNA in eukaryotes
RNA splicing occurs in the nucleus by the activity of a macromolecular complex containing both RNA and protein called the spliceosome. The proteins of the spliceosome excise the intron(s) from the primary transcript and link the flanking exons to form a contiguous protein-encoding mature mRNA (Figure 6.25). While intervening sequences in genes encoding proteins are extremely rare in Archaea, several archaeal tRNA- and rRNA-encoding genes contain introns that must be removed after transcription to generate the mature tRNA or rRNA. In analogy to the introns of eukaryotes, these intervening sequences are called “archaeal introns”; however, their processing is catalyzed by a special ribonuclease rather than by a spliceosome-type complex.
Two other steps in the processing of mRNA in Eukarya are unique to this domain, and both steps take place in the nucleus prior to splicing (Figure 6.26). The first step, called capping, occurs before transcription is complete. Capping is the addition of a methylated guanine nucleotide at the 5′-phosphate end of the mRNA (Figure 6.26). The cap is added in reverse orientation relative to the rest of the mRNA molecule and is needed to initiate translation. The second step, polyadenylation, consists of trimming the 3′ end of the primary transcript and adding 100–200 adenine residues, called the poly(A) tail (Figure 6.26). The poly(A) tail stabilizes mRNA against nuclease attack, and following translation, it must be removed before the mRNA can be degraded.
Figure 6.26 Processing of the primary transcript into mature mRNA in eukaryotes.

The processing steps include adding a cap at the 5′ end, removing the introns, and clipping the 3′ end of the transcript while adding a poly(A) tail. All these steps are carried out in the nucleus. The location of the start and stop codons to be used during translation are indicated.
We now consider the culmination of genetic information flow: protein synthesis. In Part IV we will see several events common to all cells with a few exceptions that once again link Archaea with Eukarya.
Check Your Understanding
What three major components make up an archaeal promoter?
What specific eukaryotic enzyme does the archaeal RNA polymerase resemble?
What steps take place in the processing of eukaryotic RNA?
IV Protein Synthesis: Translation
The culmination of replication and transcription is translation, the actual production of proteins by the cell’s remarkable biomolecular machine, the ribosome.
Once transcription has occurred, the mRNAs are translated into protein. Translation requires many proteins and RNAs (in addition to mRNA) that combine to form a key cellular structure called the ribosome. How an mRNA and a ribosome interact to produce a cell’s array of proteins is what we consider now, and we begin with a refresher section on the basic properties of proteins.
6.7 Amino Acids, Polypeptides, and Proteins
Proteins play major roles in cell function. Three major classes of cellular proteins are catalytic proteins, structural proteins, and regulatory proteins. Enzymes are the catalysts for chemical reactions that occur in cells (Section 3.5). Structural proteins are parts of the major structures of the cell: membranes, the cell envelope, ribosomes, and so on (Chapter 2). Regulatory proteins control most cell processes by a variety of mechanisms, including binding to DNA and affecting transcription (Chapter 7). However, regardless of function, all proteins have certain basic features in common.
Composition
Proteins are polymers of amino acids, organic compounds that contain both an amino group (−NH2) and a carboxylic acid group (−COOH) attached to the α-carbon (**Figure 6.27a). Bonds between the carboxyl carbon of one amino acid and the amino nitrogen of a second (formed through the elimination of water) are known as peptide bonds (Figure 6.28**). Two amino acids bonded by peptide linkage constitute a dipeptide; three amino acids, a tripeptide; and so on. When many amino acids are linked, they form a polypeptide. A protein consists of one or more polypeptides. The number of amino acids differs greatly from one protein to another, from as few as 15 to as many as 10,000 (see Figure 6.41).
Figure 6.27 Structure of the 22 genetically encoded amino acids.

(a) General structure. (b) R group structure. The three-letter codes for the amino acids are to the left of the names, and the one-letter codes are in parentheses to the right of the names. Pyrrolysine has thus far been found only in certain methanogenic Archaea.
Figure 6.28 Peptide bond formation.
R1 and R2 refer to the side chains of the amino acids. Note that following peptide bond formation, a free OH group is present at the C-terminus for formation of the next peptide bond.
Each amino acid has a unique side chain (abbreviated R) bonded to its α-carbon; it is this side chain that governs the chemical properties of the amino acid. Side chains vary considerably, from as simple as a hydrogen atom in the amino acid glycine to aromatic rings in phenylalanine, tyrosine, and tryptophan (Figure 6.27b). Amino acids with chemically related side chains often show similar chemical properties and are thus grouped into related amino acid “families” (Figure 6.27b). For example, the side chain may contain a carboxylic acid group, as in aspartic acid or glutamic acid, rendering the amino acid acidic. Others contain additional amino groups, making them positively charged and basic. Several amino acids contain hydrophobic side chains and are grouped together as nonpolar amino acids. Cysteine contains a sulfhydryl group (−SH). Using their sulfhydryl groups, two cysteines can form a disulfide linkage (R–S–S–R) that connects two polypeptide chains.
Protein Diversity and Structures
The diversity of chemically distinct amino acids makes possible an enormous number of structurally unique proteins that can have widely different biochemical properties. If one assumes that an average polypeptide contains 300 amino acids, then 22300 different polypeptide sequences are theoretically possible. No cell has anywhere near this many different proteins. A cell of Escherichia coli contains around 2000 different kinds of proteins; the exact number of different kinds produced is highly dependent on the resources (nutrients) and growth conditions employed.
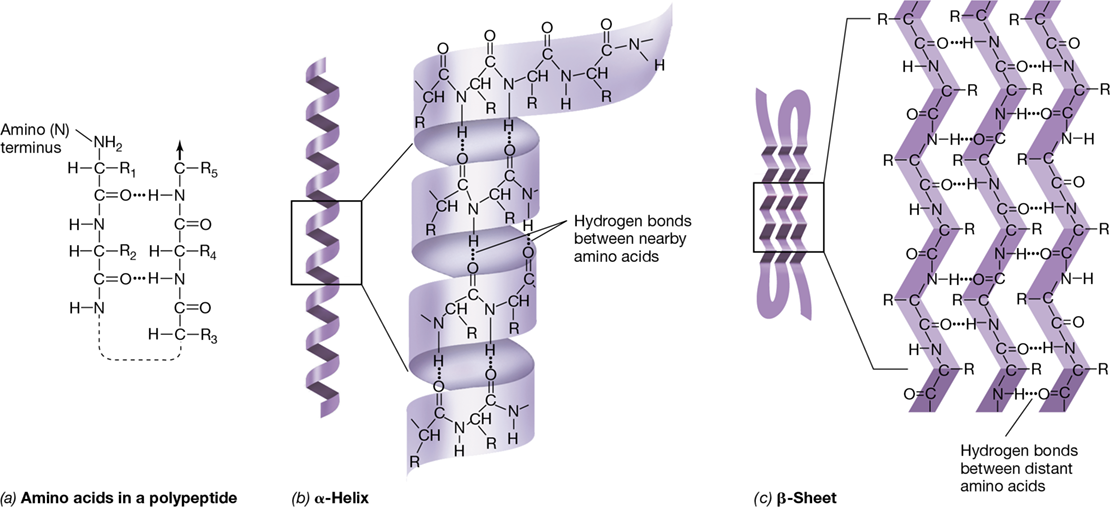
The linear sequence of amino acids in a polypeptide is its primary structure. This ultimately determines the folding pattern of the polypeptide, which in turn determines its biological activity. Even as little as a single amino acid change in the primary structure of a protein can affect its folding and thus its activity. Once formed, a polypeptide proceeds to fold to form a more stable structure. Hydrogen bonding between the oxygen and nitrogen atoms of two peptide bonds generates the secondary structure, either as an α-helix (imagine a polypeptide wound around a cylinder) or as a β-sheet (a repeated “back and forth” type of folding) (Figure 6.29). A single polypeptide can contain regions, called domains, of α-helix and regions of β-sheet secondary structure. The type of folding and its location in the molecule are determined by the primary structure and the available opportunities for hydrogen bonding.
Figure 6.29 Secondary structure of polypeptides.

(a) Hydrogen bonding in protein secondary structure. R represents the side chain of the amino acid. (b) α-Helix secondary structure. (c) β-Sheet secondary structure. Note that the hydrogen bonding is between atoms in the peptide bonds and does not involve the R groups.
Interactions between the R groups of the amino acids in a polypeptide generate higher-order structures. A protein’s tertiary structure depends largely on hydrophobic interactions, with lesser contributions from hydrogen bonds, ionic bonds, and disulfide bonds, and generates the overall three-dimensional form of the polypeptide (Figure 6.30). Many proteins consist of two or more polypeptides and thus show quaternary structure. In such proteins, the quaternary structure describes the number and secondary structure of polypeptides (referred to as subunits) in the molecule. Quaternary structures may be stabilized by various interactions and also by disulfide bonds; if cysteines located in two different polypeptides are joined, the disulfide bond links the two subunits.
Figure 6.30 Tertiary structure of polypeptides.

(a) Insulin, a protein containing two polypeptide chains; note how the B chain contains both α-helix and β-sheet secondary structure and how disulfide linkages (S—S) help in dictating folding patterns (tertiary structure). (b) Ribonuclease, a large protein with several regions of α-helix and β-sheet secondary structure.
When proteins are exposed to extremes of heat or pH or to certain chemicals that affect their folding, they may undergo denaturation. This results in the loss of a protein’s secondary, tertiary, and quaternary structure along with its biological properties. However, because peptide bonds are usually not broken, the denatured polypeptide retains its primary structure. Depending on the severity of the denaturing conditions, the polypeptide may properly refold after the denaturant is removed. However, if refolding is not correct, the protein is permanently inactive and is degraded by proteases to release its amino acids for new protein synthesis.
Check Your Understanding
Draw the structure of a dipeptide containing the amino acids alanine and tyrosine and outline the peptide bond.
Differentiate between the different classes of protein structure.
What is denaturation and why is the process harmful to a cell?
6.8 Transfer RNA
With a primer on proteins behind us, we now consider protein synthesis. But to do so, we must first understand the role of transfer RNA (tRNA). Transfer RNAs function to carry amino acids to the translation machinery. To ensure that they carry the correct amino acid, each tRNA contains a specific three-nucleotide sequence called the anticodon, the group of three bases that recognizes a codon (a three-base code for an amino acid) on the mRNA (Section 6.10). The correct amino acid (called the cognate amino acid) is linked to a specific tRNA by an enzyme called an aminoacyl-tRNA synthetase. For each amino acid, a separate aminoacyl-tRNA synthetase exists that specifically binds both the cognate amino acid and the tRNA that contains the corresponding anticodon, thus ensuring that each tRNA receives its correct amino acid. If an amino acid is encoded by more than one codon, a single aminoacyl-tRNA synthetase will recognize the anticodon on any of the tRNAs for that amino acid.
General Structure of Transfer RNA
There are about 60 different tRNAs in prokaryotic cells and 100–110 in human cells. Transfer RNAs are short (73–93 nucleotides), single-stranded molecules that contain extensive secondary structure. Certain base sequences and secondary structures are invariant for tRNAs, whereas other parts are variable. Transfer RNAs also contain some purine and pyrimidine bases that are modified from the bases found in other classes of RNA, and these modifications occur after transcription. These unusual bases include pseudouridine (Ψ), inosine, dihydrouridine (D), ribothymidine, methyl guanosine, dimethyl guanosine, and methyl inosine. The mature and active tRNA also contains extensive double-stranded regions formed by internal base pairing when the single-stranded molecule folds back on itself (Figure 6.31).
Figure 6.31 Structure of a transfer RNA.

(a) The conventional cloverleaf structural drawing of yeast phenylalanine tRNA. The amino acid is attached to the ribose of the terminal A at the acceptor end. A, adenine; C, cytosine; U, uracil; G, guanine; T, thymine; Ψ, pseudouracil; D, dihydrouracil; m, methyl; Y, a modified purine. (b) The tRNA molecule folds so that the D loop and TΨC loops are close together and associate by hydrophobic interactions.
A tRNA can be depicted in the shape of a cloverleaf (Figure 6.31a). Some regions of tRNA secondary structure are named after the modified bases found there (for example, the TΨC and D loops) or after their functions (for example, the anticodon loop and acceptor stem). The three-dimensional model of a tRNA shown in Figure 6.31b is a more realistic view of the molecule and shows how bases that appear widely separated in the cloverleaf model are actually much closer together when viewed in 3D. This close proximity allows some of the bases in one loop to pair with bases in another loop (Figure 6.31b).
At the 3′ end (the acceptor end) of all tRNAs are three unpaired nucleotides. The sequence of these three nucleotides is always cytosine-cytosine-adenine (CCA), and they are absolutely essential for function. However, in most organisms the 3′ CCA is not encoded in the tRNA gene on the chromosome; instead, each nucleotide is added sequentially by a protein called CCA-adding enzyme, using CTP and ATP as substrates. The cognate amino acid is then covalently attached to the terminal adenosine of the CCA end of its corresponding tRNA. From this location, the amino acid is incorporated into the growing polypeptide chain on the ribosome by a mechanism to be described in Section 6.10.
Recognition, Activation, and Charging of tRNAs
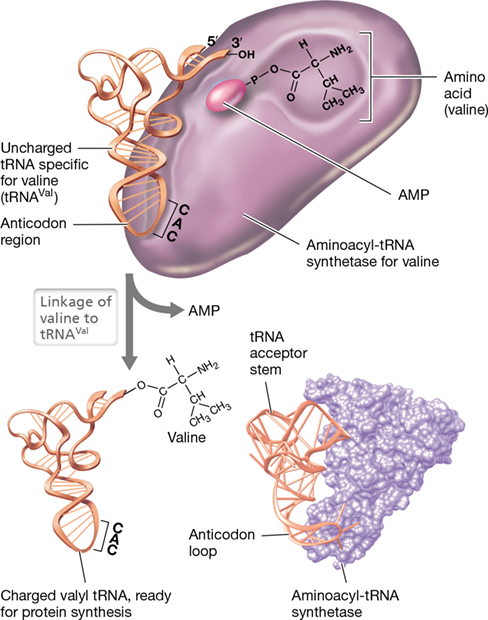
Recognition of the correct tRNA by an aminoacyl-tRNA synthetase is obviously crucial to the fidelity of translation and requires that specific contacts be made between regions of the tRNA and the synthetase (Figure 6.32). As might be expected because of its unique sequence, the anticodon of the tRNA is important in recognition by the synthetase. However, other contact sites between the tRNA and the synthetase are also important, including parts of the acceptor stem and D loop of the tRNA (Figure 6.31a).
Figure 6.32 Aminoacyl-tRNA synthetase.

Mode of activity of an aminoacyl-tRNA synthetase. Recognition of the correct tRNA by a particular synthetase involves contacts between specific nucleic acid sequences in the D loop and acceptor stem of the tRNA and specific amino acids of the synthetase. In this diagram, valyl-tRNA synthetase (specific for the amino acid valine) is shown catalyzing the final step of the reaction, where the valine in valyl-AMP is transferred to tRNA. The computer model on the bottom right shows the interaction of the prolyl-tRNA synthetase from Thermus thermophilus with its tRNA.
Modified from G. Caetano-Anollés and A. Nasir. 2012.
The specific reaction between amino acid and tRNA catalyzed by the aminoacyl-tRNA synthetase begins with activation of the amino acid by reaction with ATP: Amino acid+ATP↔aminoacyl−AMP+P−P
The aminoacyl-AMP intermediate formed remains bound to the tRNA synthetase until collision with the appropriate tRNA molecule. Then, as shown in Figure 6.32, the activated amino acid is bonded to the CCA stem of its tRNA to form a charged tRNA: Aminoacyl−AMP+tRNA↔aminoacyl−tRNA+AMP
The pyrophosphate (PPi) formed in the first reaction is split into two molecules of inorganic phosphate. Because ATP is used and AMP is formed in these reactions, a total of two energy-rich phosphate bonds are expended to charge a tRNA with its cognate amino acid. After activation and charging, the aminoacyl-tRNA leaves the synthetase. In the next step, it will be bound by a ribosome where actual polypeptide synthesis occurs.
Check Your Understanding
What is the function of the acceptor stem of a tRNA?
6.9 Translation and the Genetic Code
The heart of genetic information transfer is the correspondence between the nucleic acid template and the amino acid sequence of a polypeptide. This correspondence is rooted in the genetic code. An mRNA triplet of three bases, called a codon, encodes each specific amino acid (the codons themselves are encoded by the organism’s genome). The 64 possible codons (four bases taken three at a time=43) are shown in Table 6.4. Note that in addition to the codons for amino acids, there are also codons for starting and stopping translation. Here we focus on translation in Bacteria, with Escherichia coli as the model.
Table 6.4 The genetic code as expressed by triplet base sequences of mRNA

aAUG encodes N-formylmethionine at the beginning of polypeptide chains of Bacteria.
Properties of the Genetic Code
There are 22 naturally occurring amino acids and because there are 64 codons, several amino acids can be encoded by more than one codon. A code such as this that lacks one-to-one correspondence between “word” (that is, the amino acid) and code (codon) is called a degenerate code. A codon is recognized by specific base pairing with a complementary sequence on the anticodon, located on a tRNA (Section 6.8 and Figures 6.31 and 6.32). If this base pairing were always the standard pairing of A with U and G with C, then at least one specific tRNA would be needed to recognize each codon. In some cases, this is true. For instance, there are six different tRNAs in Escherichia coli for the amino acid leucine, one for each codon (Table 6.4). By contrast, some tRNAs can recognize more than one codon. For example, although there are two lysine codons in E. coli, there is only one lysyl tRNA, whose anticodon can base-pair with either AAA or AAG. In these cases, the anticodon forms standard base pairs at only the first two positions of the codon and tolerates irregular base pairing at the third position. This phenomenon is called wobble and is illustrated in Figure 6.33.
Figure 6.33 The wobble concept.

Base pairing is more flexible for the third base of the codon than for the first two. Only a portion of the tRNA is shown here.
Mastering Microbiology
If an amino acid is encoded by multiple codons, the codons are typically closely related in base sequence, usually differing at only their third position (Table 6.4) to allow for wobble (Figure 6.33). Interestingly, not all multiple codons for a given amino acid are used at the same frequency, leading to a codon bias that varies from organism to organism. Codon bias is correlated with a corresponding bias in the concentration of different tRNA molecules. That is, a tRNA whose anticodon corresponds to a rarely used codon is typically produced at low levels.
Start and Stop Codons and Reading Frames
Messenger RNA is translated in Bacteria beginning with its start codon (AUG, Table 6.4), which encodes a chemically modified methionine called N-formylmethionine (although AUG at the beginning of a coding region encodes N-formylmethionine, AUG within the coding region encodes methionine). By contrast, the translational machinery in Archaea and Eukarya insert an unmodified methionine as the first amino acid in a polypeptide.
With a triplet code it is critical for translation to begin at the correct nucleotide. If it does not, the whole reading frame of the mRNA will be shifted and thus an entirely different (and likely inactive) protein will be made. Alternatively, if the shift introduces a stop codon (Table 6.4) into the reading frame, the polypeptide will terminate prematurely. The reading frame that when translated yields the polypeptide encoded by the gene is called the 0 (zero) frame; the other possible reading frames (−1 and +1) do not encode the same amino acid sequence (Figure 6.34). Reading frame fidelity is governed by interactions between mRNA and rRNA within the ribosome. In Bacteria, ribosomal RNA recognizes a specific AUG on the mRNA as a start codon with the aid of an upstream sequence in the mRNA called the ribosome-binding site (RBS) (also called the Shine–Dalgarno sequence after its discoverers). This upstream alignment requirement explains why some mRNAs from Bacteria can use other start codons, such as GUG. However, because of the RBS, even these unusual start codons direct the incorporation of N-formylmethionine as the initiator amino acid (see Figure 6.36).
Figure 6.34 Possible reading frames in an mRNA.

An interior sequence of an mRNA is shown. (a) The amino acids that would be encoded if the ribosome is in the correct reading frame (designated the “0” frame). (b) The amino acids that would be encoded by this region of the mRNA if the ribosome were in the −1 reading frame. (c) The amino acids that would be encoded if the ribosome were in the +1 reading frame.
The codons UAA, UAG, and UGA (Table 6.4) are stop codons, and they signal the termination of translation of a protein-coding sequence on the mRNA. Stop codons are also called nonsense codons, because they interrupt the “sense” of the growing polypeptide when they terminate translation. In a few rare cases in Bacteria and Archaea, nonsense codons encode the unusual amino acids selenocysteine and pyrrolysine (Figure 6.27). When this occurs, specific tRNAs are employed whose anticodons read these stop codons. What controls this unusual occurrence is a recognition sequence just downstream of the now-coding stop codon that signals the incorporation of tRNAs containing selenocysteine or pyrrolysine rather than stopping translation. A few other microbes use conventional stop codons to encode amino acids, but in these cases the organisms have simply dispensed with using these particular stop codons as translational stop sites.
If an mRNA can be translated, it is because it contains an open reading frame (ORF): a start codon followed by a number of codons and then a stop codon in the same reading frame as the start codon. Using computational methods, a DNA base sequence can be scanned to search for open reading frames. In addition to looking for start and stop codons, computer analyses may include a search for promoters and ribosome-binding sequences as well in order to confirm the ORF as protein encoding.
The search for ORFs is central to the field of genomics (Chapter 10), for if an unknown piece of DNA has been sequenced, the presence of an ORF implies that it can encode protein. In this way, computational analyses of sequenced microbial genomes can reveal much about the biology of an organism.
Check Your Understanding
What are start codons and stop codons? Why is it important for the ribosome to read “in frame”?
If you were given a nucleotide sequence, how would you find ORFs?
6.10 The Mechanism of Protein Synthesis
Protein synthesis is a dynamic process that can be broken down into three major steps: initiation, elongation, and termination. In addition to mRNA, tRNA, and ribosomes, translation requires a number of initiation, elongation, and termination proteins and the energy-rich compound guanosine triphosphate (GTP) to provide the energy to drive the process.
Ribosomes and the Initiation of Translation
Ribosomes are large complexes of proteins and RNAs where the cell’s proteins are biosynthesized (Figure 6.35). A cell may have many thousands of ribosomes, the number increasing at higher growth rates. Each ribosome consists of two subunits. Bacteria and Archaea have 30S and 50S ribosomal subunits that yield intact 70S ribosomes. Each ribosomal subunit contains specific ribosomal RNAs (rRNAs) and ribosomal proteins. The 30S subunit contains 16S rRNA and 21 proteins, and the 50S subunit contains 5S and 23S rRNA and 31 proteins. Thus, in Escherichia coli, there are 52 distinct ribosomal proteins, most present at one copy per ribosome. The ribosome is a highly dynamic structure, and its subunits alternately associate and dissociate during the translational process (see Figures 6.36 and 6.37) while interacting with many other proteins. In addition, several cytoplasmic proteins called translation factors are essential for translation and interact with the ribosome at various stages of the translational process.
Figure 6.35 Ribosome structure.

Crystal structure of the Thermus thermophilus 70S ribosome. 50S and 30S subunits are indicated. Each subunit is composed of several distinct proteins; the 50S subunit contains 23S and 5S rRNAs, and the small subunit contains 16S rRNA.
In Bacteria, initiation of protein synthesis begins with a free 30S ribosomal subunit (Figure 6.36). From this, an initiation complex forms consisting of the 30S subunit, mRNA, formylmethionine tRNA (the initiator tRNA in Bacteria; after polypeptide completion, the formyl group is removed), and the initiation factors IF1, IF2, and IF3. GTP is also required for this step. Next, a 50S ribosomal subunit is added to the initiation complex to form the active 70S ribosome (Figures 6.35 and 6.36). Just preceding the start codon on the mRNA is a sequence of three to nine nucleotides that compose the ribosome-binding site (Figure 6.36). This site is toward the 5′ end of the mRNA and is complementary to base sequences in the 3′ end of the 16S rRNA, which is part of the ribosome. Base pairing between these two RNAs holds the ribosome–mRNA complex securely together in the correct reading frame. Polycistronic mRNA (Section 6.5) contains several RBS sequences, one upstream of each coding sequence. This allows bacterial ribosomes to translate several genes on the same mRNA because the ribosome can locate each initiation site within a message by binding to its RBS.
Figure 6.36 The ribosome and initiation of protein synthesis.

The mRNA and initiator tRNA, carrying N-formylmethionine (“Met”), bind first to the small subunit of the ribosome. Initiation factors (not shown) use energy from GTP to promote the addition of the large ribosomal subunit. The initiator tRNA starts out in the P site (labeled in the structure on the right-hand side of the arrow).
Elongation, Translocation, and Termination
During translation, the mRNA threads through the ribosome as its nucleotides bind to the 30S subunit. The ribosome contains other sites where the tRNAs interact. Two of these sites are located primarily on the 50S subunit and are termed the A (acceptor) site and the P (peptide) site (Figure 6.37). The A site is where the incoming charged tRNA first attaches, and the loading of a tRNA into the A site is assisted by the elongation factor EF-Tu. The P site is where the growing polypeptide chain is attached to the prior tRNA. During peptide bond formation, the growing polypeptide chain moves to the tRNA at the A site as a new peptide bond is formed. In addition to EF-Tu, elongation factor EF-Ts, as well as more GTP, is required (Figure 6.37).
Figure 6.37 Elongation cycle of translation.

1. Elongation factors (not shown) use GTP to install the incoming tRNA into the A site. 2. Peptide bond formation is then catalyzed by the 23S rRNA. 3. Translocation of the ribosome along the mRNA from one codon to the next requires hydrolysis of another GTP and results in movement of the tRNA with the growing peptide to the P site. The outgoing tRNA is released from the E site. 4. The next charged tRNA binds to the A site and the cycle repeats. The genetic code, expressed as codons of mRNA, is shown in Table 6.4. Translation of a given polypeptide ends when the ribosome reaches any one of three stop codons (Table 6.4) because no tRNA has an anticodon sequence (Figure 6.33) complementary to the three-base sequence in a stop codon.
Following elongation, the tRNA holding the polypeptide is translocated from the A site to the P site, thus opening the A site for a new charged tRNA; this requires elongation factor EF-G and one molecule of GTP for each translocation event (Figure 6.37). In each translocation, the ribosome advances three nucleotides (one codon) along the mRNA, exposing a new codon at the A site. Translocation pushes the now amino acid–free tRNA to a third site, called the E (exit) site, and it is from here that the tRNA is released from the ribosome. The precision of the translocation step is critical to the accuracy of protein synthesis. That is, the ribosome must move exactly one codon at each step or the reading frame will change and the fidelity of translation will be compromised (Section 6.9).
Several ribosomes can translate a single mRNA molecule simultaneously, forming a complex called a polysome (Figure 6.38). Polysomes increase both the speed and efficiency of translation because each ribosome in the polysome makes a complete polypeptide. Note in Figure 6.38 how ribosomes in the polysome that are closest to the 5′ end (the beginning) of the mRNA molecule have short polypeptides attached to them because only a few codons have been read, while ribosomes closest to the 3′ end of the mRNA have nearly finished polypeptides.
Figure 6.38 Polysomes.

Translation by several ribosomes on a single messenger RNA forms the polysome. Note how the ribosomes nearest the 5′ end of the message are at an earlier stage in the translation process than ribosomes nearer the 3′ end, and thus only a relatively short portion of the final polypeptide has been made.
Translation terminates when the ribosome reaches a stop codon (Table 6.4) because no tRNA binds to a stop codon. Instead, proteins called release factors (RFs) recognize the stop codon and cleave the attached polypeptide from the final tRNA, releasing the finished product. Following this, the ribosomal subunits dissociate, and the 30S and 50S subunits are then free to form new initiation complexes (Figure 6.36) and repeat the process.
Role of Ribosomal RNA in Protein Synthesis
Ribosomal RNA plays major roles in all stages of translation, from initiation to termination. By contrast, the primary role of the proteins in the ribosome is to form a scaffold to position key sequences in the ribosomal RNAs (Figure 6.35). In Bacteria, 16S rRNA facilitates initiation by base pairing with the ribosome-binding site on the mRNA, and, along with ribosomal proteins, it holds the mRNA in position on either side of the A and P sites. Ribosomal RNA also plays a role in ribosome subunit association, as well as in positioning tRNAs on the ribosome (Figures 6.36 and 6.37). Although charged tRNAs recognize the correct codon by codon–anticodon base pairing (Figure 6.33), they are also bound to the ribosome by interactions between the anticodon stem–loop of the tRNA and specific sequences in the 16S rRNA. Moreover, the acceptor end of the tRNA (Figures 6.36 and 6.37) base-pairs with sequences in 23S rRNA.
In addition to roles in mRNA alignment and translocation along the transcript, ribosomal RNA also catalyzes the actual formation of peptide bonds. The peptidyl transferase reaction (formation of peptide bonds, Figure 6.28) occurs on the 50S subunit of the ribosome and is catalyzed solely by 23S rRNA. This rRNA also plays a role in translocation and interacts with the elongation factors. Thus, in addition to its role as the structural backbone of the ribosome, ribosomal RNA plays a major catalytic role in the translation process.
Freeing Trapped Ribosomes and *Trans*-Translation
A defective mRNA that lacks a stop codon cannot be properly translated. Such a defect may arise, for example, from a mutation that removed the stop codon, from defective synthesis of the mRNA, or when partial degradation of an mRNA occurs before it is translated. If a ribosome reaches the end of an mRNA molecule and there is no stop codon, release factor cannot bind and the ribosome cannot be released from the mRNA; the ribosome is effectively “trapped.”
To deal with this problem, bacterial cells initiate a process called trans-translation. This process produces a small RNA molecule called tmRNA that frees stalled ribosomes (Figure 6.39). The “tm” in its name refers to the fact that tmRNA mimics both tRNA, in that it carries an amino acid (alanine), and mRNA, in that it contains a short stretch of RNA that can be translated. When tmRNA collides with a stalled ribosome, it binds alongside the defective mRNA. Protein synthesis can then proceed, first by adding the alanine on the tmRNA and then by translating the short tmRNA message. The tmRNA contains a stop codon that allows release factor to bind and disassemble the ribosome. The protein made as a result of this rescue operation is defective and is subsequently degraded. This is accomplished by a short sequence of amino acids encoded by tmRNA and added to the end of the defective protein; the sequence is a signal for a specific protease to degrade the protein. Thus, through the activity of trans-translation, stalled ribosomes are not inactivated but instead are freed up to participate in protein synthesis once again.
Figure 6.39 Freeing of a stalled ribosome by tmRNA.

A defective mRNA lacking a stop codon stalls a ribosome that has a partly synthesized polypeptide attached to a tRNA (blue) in the P site. Binding of tmRNA (yellow) in the A site allows translation to continue up to the stop codon provided by the tmRNA.
We complete this chapter by considering what happens to proteins after they are made but before they become functional. For some proteins the answer is “nothing,” but for others, functionality depends on additional activities, and we survey these now.
Check Your Understanding
What are the components of a ribosome? What functional roles does rRNA play in protein synthesis?
How is a completed polypeptide chain released from the ribosome?
V Protein Processing, Secretion, and Targeting
Not all proteins that exit the ribosome are “ready to go,” as some need additional folding or other processing and others need to be forwarded to specific cellular locations—such as the cytoplasmic membrane—if they are to carry out their functions properly.
While translation is the final step in genetic information flow (Figures 6.1a and 6.5), some proteins require subsequent processing or targeting before they become functional. These activities often require accessory proteins to assist with folding and transport as well as intrinsic signals within the translated protein itself.
6.11 Assisted Protein Folding and Chaperones
In Section 6.7 we saw how proteins show several levels of structure including primary, secondary, tertiary, and, in multi-subunit proteins, quaternary structure. Many proteins fold spontaneously into their active form, even while they are being synthesized (Figure 6.38). However, some do not, and these require assistance to achieve an active functional state.
Major Chaperones of *Bacteria Bacteria* produce a series of proteins called chaperones that catalyze a variety of macromolecular folding events. These events include folding proteins that do not fold spontaneously, refolding partially denatured proteins, assembling multiprotein complexes, preventing the improper aggregation of proteins, untangling RNAs, and incorporating cofactors into enzymes. Chaperones are found in all domains of life, and their amino acid sequences are highly conserved among all organisms.
Four key chaperones in Escherichia coli are the proteins DnaK, DnaJ, GroEL, and GroES. DnaK and DnaJ are ATP-dependent enzymes that bind to newly formed polypeptides and prevent them from folding too quickly, which would increase the risk of improper folding (**Figure 6.40*a***). If the DnaKJ complex is unable to fold the protein properly, it may transfer the partially folded protein to the two multi-subunit proteins GroEL and GroES. The protein first enters GroEL, a large, barrel-shaped protein that uses the energy of ATP hydrolysis to fold the protein properly. GroES assists in this (Figure 6.40a). It is estimated that about 100 of the several thousand different proteins in a cell of E. coli need help from the GroEL–GroES complex to become properly folded, and of these, a dozen or so are proteins essential for the survival of the bacterium.
Figure 6.40 The activity of chaperone proteins.

(a) An improperly folded protein can be refolded by either the DnaKJ complex or by the GroEL–GroES complex. In both cases, energy for refolding comes from ATP. Inset: Crystal structure of GroELS complex. (b) Formation of an active nitrate reductase enzyme (Section 3.10 and Figure 3.23b) by chaperone-facilitated incorporation of a cofactor. The chaperone protein NarJ carries the molybdenum cofactor (Moco) to the inactive cytoplasmic NarGH complex. After incorporation of Moco, NarJ dissociates and the complex binds to NarI in the membrane, becoming an active enzyme.
Other Functions of Chaperones
Chaperones can also refold proteins that have partially denatured in the cell. A protein may denature for many reasons, but often it is because the organism has temporarily experienced high temperatures. Chaperones are thus one type of heat shock protein, and their synthesis is greatly accelerated when a cell is stressed by excessive heat (Section 7.11). The heat shock response is an attempt by the cell to refold its partially denatured proteins for reuse before proteases recognize them as improperly folded and destroy them. In contrast to heat shock proteins, cold shock proteins are produced when the cell experiences a sudden shift to very cold temperatures. Cold (but nonfreezing) conditions do not affect most proteins but can affect RNAs. CspA, a major cold shock protein of E. coli, is an RNA chaperone instead of a protein chaperone and prevents the spontaneous formation of secondary structures in mRNA that could compromise their ability to be translated. Many other cold shock proteins are known in E. coli including ones that refold cold-sensitive proteins rather than acting on mRNAs.
Some chaperones help assemble cofactor-containing enzymes such as those that catalyze redox or electron transport reactions (Section 3.8). For example, several multi-subunit molybdoenyzmes such as the E. coli membrane-bound nitrate reductase (Section 3.10 and Figure 3.23b) require that a molybdenum cofactor (Moco) be inserted during folding, and cofactor insertion is assisted by enzyme-specific chaperone proteins (Figure 6.40b). Nitrate reductase is a key enzyme in anaerobic respiration (Section 3.10 and Section 14.11) and is composed of three subunits: NarG, NarH, and NarI. For the enzyme to be functional, the cytoplasmic chaperone protein NarJ must first insert the Moco group into NarG. Once this cofactor has been inserted, the NarG and NarH subunits associate with the membrane-bound NarI subunit to form the active nitrate reductase enzyme complex (Figure 6.40b).
Check Your Understanding
What are molecular chaperones and why are they necessary?
What macromolecules are protected by heat shock proteins?
How do chaperones assist the Escherichia coli cell to make nitrate reductase?
6.12 Protein Secretion: The Sec and Tat Systems
While many proteins exist in the cell’s cytoplasm, some must be transported outside the cytoplasmic membrane into the periplasm (of gram-negative cells) or inserted into the cytoplasmic or outer membranes to facilitate nutrient transport or bioenergetic events. Other proteins such as toxins and extracellular enzymes (exoenzymes) must be secreted from the cell entirely to be active in the environment or to invade another cell. These secreted proteins must get from their site of synthesis on ribosomes into or through the cytoplasmic membrane in order to be functional, and the cell has evolved several systems that require either ATP (or GTP) or the proton motive force (Chapter 3) to get this job done.
One example of an unusually large protein that must be secreted to perform its function is halomucin. Halomucin is produced by the halophilic archaeon Haloquadratum walsbyi and shares a structural similarity with animal mucin, which protects eyes and bronchial epithelium from dehydration. Thus halomucin is believed to protect H. walsbyi from desiccation when growing in its highly saline brine pool habitat (Section 4.15). At 9159 amino acids, halomucin is one of the largest proteins produced by any life form (the average cellular protein is <500 amino acids). For this large protein to confer protection to H. walsbyi, the cell must specifically recognize halomucin in the cytoplasm and orchestrate its secretion across the cytoplasmic membrane (Figure 6.41). While little is known about the mechanisms H. walsbyi uses to secrete proteins, however this is accomplished, a significant energy input would be needed to secrete a protein the extreme size of halomucin.
Figure 6.41 Secretion of halomucin.

Fluorescent micrograph illustrating secreted halomucin (labeled with green fluorescent protein, Section 12.5) by the archaeon Haloquadratum walsbyi (red). Note that cells of H. walsbyi are square and are visualized here by using a red stain for storage polymers (poly-β-hydroxyalkanoates; Section 2.7).
Zenke, R. et al. 2015. Frontiers in Microbiology 6: 249
The Signal Sequence
Proteins called translocases transport specific proteins into or through the membranes of Bacteria and Archaea. For example, the Sec translocase system both exports unfolded proteins and inserts integral membrane proteins into the cytoplasmic membrane. By contrast, the Tat translocase system transports previously folded proteins through the cytoplasmic membrane. Both the general secretion (Sec) and Tat systems are universal in Bacteria and Archaea.
Most proteins that must be transported into or through bacterial or archaeal cytoplasmic membranes are synthesized with an amino acid sequence of 15–20 residues—called the signal sequence—at the beginning (N-terminus, Figure 6.28) of the protein molecule. Signal sequences are variable, but they typically contain a few positively charged amino acids at the beginning, a central region of hydrophobic residues, and then a more polar region at their end. The signal sequence is so named because it “signals” to a translocase system that this particular protein is to be exported and also helps prevent the protein from completely folding, a process that could interfere with its secretion. Because the signal sequence is the first part of the protein to be synthesized, the early steps in export may actually begin before the protein is completely synthesized (Figure 6.42).
Figure 6.42 Export of proteins via the SecA secretory system.

The signal sequence is recognized either by SecA or by the signal recognition particle, which carries the protein to the membrane secretion system. The signal recognition particle binds proteins that are inserted into the membrane, whereas SecA binds proteins that are secreted across the cytoplasmic membrane.
Sec and Tat Translocases
In the Sec system, unfolded proteins to be exported from the cytoplasm are recognized by either the SecA protein or the signal recognition particle (SRP) (Figure 6.42). Typically, SecA binds polypeptides that are to be exported into the periplasm and transports the unfolded polypeptide to a membrane-bound channel. The polypeptide is then translocated through the channel using ATP as the energy source. Conversely, the SRP binds polypeptides that are to be inserted into the membrane (but not released on the other side) through a process called cotranslation. The bacterial SRP, which consists of a single protein and a small, noncoding RNA molecule (4.5S RNA), coordinates to both recognize polypeptides that are destined for the membrane and to move the ribosome actively translating these proteins to the membrane surface. Thus during cotranslation, the polypeptide is actively translated into the membrane, driven by the hydrolysis of GTP. After the protein has been inserted into the membrane (SRP-mediated) or has crossed the membrane (SecA-mediated), the signal sequence is removed by a protease and the proteins fold to their active form.
Some proteins must be folded before they are translocated because they contain cofactors that must be inserted as the protein folds; for example, iron–sulfur proteins, cytochromes, and other respiratory enzymes (Section 3.8). Such proteins are processed by the Tat translocase system (for example, nitrate reductase shown in Figure 6.40b). Tat stands for “twin-arginine translocase” because the transported proteins contain a short signal sequence that has a pair of arginine residues. This signal sequence is recognized by TatBC, which carries the folded protein to TatA—the membrane transporter. Following transport, which is fueled by the proton-motive force, the signal sequence is removed by a protease.
Check Your Understanding
What is the signal sequence and what does it signal?
What is the signal recognition particle composed of?
How do the translocases Sec and Tat differ in the molecules they secrete?
6.13 Protein Secretion: Gram-Negative Systems
In order to insert proteins or other small molecules known as effectors into the outer membrane of gram-negative Bacteria, or to secrete them outside of the cell, special secretion systems known as types I through VI are used. Proteins secreted by these systems are in their final folded form once outside of the cell. Gram-positive bacteria and Archaea possess secretion systems similar to some of these, but in these organisms, the machinery only has to transport the proteins or effector molecules across the cytoplasmic membrane.
Types I–VI secretion systems facilitate several cellular activities including symbiotic interactions, biofilm formation, extracellular enzyme secretion, transfer of DNA, release of antibiotics, and delivery of proteins into host cells. Thus, molecules secreted by these systems allow bacteria to interact with the environment and other organisms. Each of these systems specifically recognizes its substrate based on amino acid residues and is composed of a large complex of proteins that forms a translocase channel spanning one or more membranes through which the secreted molecule travels (Figure 6.43).
Figure 6.43 Secretion systems in gram-negative bacteria.

(a) Types I, II, and V secrete proteins outside of the bacterial cell. Type I systems secrete proteins in a single step. Types II and V first require the Sec or Tat system to transport the protein to be secreted across the inner membrane. Note that during type V secretion, the Sec system first transports the unfolded transporter domain linked to the unfolded secretion protein through the inner membrane. The translocase then folds in the outer membrane and delivers the folded secretion protein through the outer membrane. (b) Types III, IV, and VI secrete molecules outside of the bacterial cell and into a host cell. Type III systems have been termed the injectisome, while type IV systems are similar to a pilus (Section 2.6) and also secrete DNA into a host cell. Type VI systems contain a sheath or needle in the cytoplasm that contracts to deliver a protein into a host cell.
Types II and V Secretion Systems
Secretion systems can be grouped into one-step and two-step classes. Type II and type V systems are two-step translocases because they depend on either the Sec or Tat system (Section 6.12) to transport the secreted protein or a portion of the translocase channel from the cytoplasm through the inner membrane (Figure 6.43a).
Type II systems are found in a wide variety of gram-negative pathogenic and nonpathogenic bacteria, and they transport proteins from the periplasm to the extracellular environment. The translocase machinery of type II systems includes a secretion pore in the outer membrane that is anchored to the cytoplasmic membrane by proteins that span the periplasm (Figure 6.43a). While the type II system does possess cytoplasmic membrane and periplasmic components, proteins to be secreted do not translocate through these and are instead delivered to the secretion pore in the outer membrane by either the Sec or Tat system. A key to the specificity of the type II system is that the secretion pore only opens to proteins specific to the type II system. Examples of proteins secreted by a type II system include the toxin produced by Vibrio cholerae (the causative agent of cholera) and a glucanase exoenzyme produced by Klebsiella pneumoniae to degrade large extracellular starch molecules.
Type V systems are the structurally simplest of the secretion systems (Figure 6.43a) and are also called autotransporters in that the protein to be secreted is fused to a transmembrane protein domain essential to the protein’s transport across the outer membrane. After this unfolded multidomain protein is itself transported through the cytoplasmic membrane by the Sec system, a transmembrane domain called the transporter forms a secretion pore in the outer membrane that allows the remainder of the protein (the passenger domain) to pass through and ultimately be secreted outside of the cell (Figure 6.43a). Thus proteins to be moved outside of the cell by a type V system are initially transported in the unfolded state, and both the transporter and passenger domains require chaperone proteins (Section 6.11) for proper folding. The folding of the passenger domain (instead of ATP hydrolysis) drives type V secretion. Examples of proteins secreted by the type V system are adhesion proteins used by Escherichia coli and Haemophilus influenzae (a causative agent of pneumonia) to attach to host cells.
Types I, III, IV, and VI Secretion Systems
The second class of translocases moves proteins through the outer membrane in a single step. Types I, III, IV, and VI are one-step systems because they form channels through both the cytoplasmic and outer membranes and do not require Sec or Tat. Type I systems are ABC transporters (Section 2.2) that are characterized by three protein components: (1) a cytoplasmic membrane transporter coupled to (2) an outer membrane pore by (3) a membrane fusion protein. The cytoplasmic membrane transporter binds specifically to the protein to be secreted and requires ATP to initiate transport to the outside of the cell (Figure 6.43a). While the cytoplasmic membrane transporter is specific to its substrate, a wide range of polypeptide sizes can be secreted. Type I systems secrete small molecules such as a toxic protein (bacteriocin) produced by E. coli to kill competing bacterial cells and can also secrete large proteins such as those necessary for biofilm formation on plant surfaces by the soil bacterium Pseudomonas fluorescens.
Type III systems are commonly used by pathogenic bacteria not only to secrete toxic proteins outside of the cell but to inject these molecules directly into eukaryotic host cells (Figure 6.43b). The entire type III structure is highly complex and composed of over 100 proteins that facilitate substrate recognition, coordinated assembly of the machinery for translocations, and the transport process itself. ATP hydrolysis provides the energy for the secretion and injection. The entire structure, 30–70 nm in length, has been termed the “injectisome” for its similarity to a syringe in both structure and function (**Figure 6.44*a***). Proteins injected into host cells by the type III system often aid in pathogen infection and host invasion. Type III–injected proteins include certain effector molecules of Chlamydia (the cause of trachoma and a sexually transmitted disease) and Salmonella (a foodborne and waterborne pathogen). However, type III secretion systems are not limited to pathogens. Nitrogen-fixing bacteria deliver molecules critical for establishing a symbiotic relationship with plant roots (root nodules) through type III secretion machinery (Section 23.4).
Figure 6.44 Electron micrographs of secretion machinery.

(a) Purified type III injectisomes from Salmonella enterica (typhimurium). (b) Purified type VI contractile sheaths from Vibrio cholerae.
Type IV systems (Figure 6.43b) are the most ubiquitous and are present in many Bacteria and Archaea. This system is also used to deliver secreted proteins or other molecules into other cells. While this system can be used to transport toxins into host cells, its primary role is to transfer DNA to other cells though the process of conjugation (Section 9.8), one mechanism that prokaryotic cells have to exchange genes. The type IV translocase is similar to a pilus that extends through the outer membrane into another cell during conjugation. Once the tip of the pilus makes contact with a receptor on the host cell, the DNA is recognized by an inner membrane protein of the donor and then transferred using ATP-facilitated transport (Section 9.8). Not only can DNA be transferred from one bacterial cell to another using a type IV system, DNA from the plant pathogenic bacterium Agrobacterium can be transferred to host plant cells through the system encoded on the Ti plasmid (resulting in crown gall disease, Section 23.6).
Type VI systems are widely distributed in gram-negative bacteria, and like type IV systems they are capable of delivering a diversity of proteins directly into the cytoplasm of other cells using a one-step, ATP-requiring process similar to type III and IV systems. However, unlike the injectisome or pilus-like structure of the type III and IV translocase systems, the type VI translocase is cytoplasmic and forms a needle-like protein with a pore-forming protein that contracts all the way through the donor cell’s two membranes and directly into a host cell once a substrate molecule is recognized (Figures 6.43b and 6.44b). The overall delivery process is similar to the mechanism that the tailed bacteriophage T4 uses to deliver DNA into an E. coli cell through tail contraction (Section 5.5). Type VI systems are used by bacteria as weapons to compete with other bacterial cells or to attack eukaryotic cells. In Pseudomonas aeruginosa, a type VI system is used to inject a toxin into competing bacterial cells. Similarly, Vibrio cholerae uses its type VI system (Figure 6.44b) to inject enzymes into competing bacteria; the enzymes degrade the cell wall and membrane. V. cholerae also uses the system to deliver toxins to human intestinal cells during a cholera infection.
It should be obvious by now that protein secretion is a crucial aspect of the biology of Bacteria and Archaea and that these cells have evolved diverse structures and unique mechanisms to deal with protein secretion. In fact, three other systems (types VII–IX) have also been discovered, but their mechanisms have not been fully described. All of these elaborate systems are of interest for reasons beyond basic science. Since many of these systems are essential virulence factors (Section 25.3) for the pathogenic bacteria that produce them, they are currently targets for vaccine development. The rationale here is quite simple: If the activity of these systems could be blocked by the immune system, the pathogen would be unable to establish an infection and cause disease.
We now move on from our focus on the central dogma of biology, protein processing, and protein secretion to tackle the concepts of Chapter 7, where we will examine how gene expression and protein activity are regulated. Without regulatory systems, cells would not be able to quickly respond to ever-changing conditions in their environments, and thus their very survival would be at stake.
Check Your Understanding
Compared with gram-positive bacteria, why is it important for gram-negative bacteria to have additional secretion pathways?
How do the types I–VI secretion processes differ from Sec and Tat secretion?
Chapter Review
Go to Mastering Microbiology for videos, animations, practice tests, and more.
I Molecular Biology and Genetic Elements
6.1 The informational content of a nucleic acid is determined by the sequence of nitrogenous bases along the polynucleotide chain. Both RNA and DNA are informational macromolecules, as are the proteins they encode. DNA forms a double-stranded helix whose strands are complementary and antiparallel. DNA is supercoiled before it is packaged into cells. The three key processes of macromolecular synthesis are: (1) DNA replication; (2) transcription (RNA synthesis); and (3) translation (protein synthesis).
Q Describe the central dogma of molecular biology. With regards to DNA, what is supercoiling and what is meant by the terms antiparallel and complementary?
6.2 In addition to the chromosome, other genetic elements can exist in cells. Plasmids are DNA molecules that exist separately from the chromosome and may confer a selective growth advantage under certain conditions. Viruses contain an RNA or DNA genome, and transposable elements exist as a part of other genetic elements.
Q How are chromosomes and plasmids similar, and how do they differ? What are R plasmids and why are they of medical concern?
II Copying the Genetic Blueprint: DNA Replication
6.3 Both strands of the DNA helix are templates for the synthesis of new strands (semiconservative replication). The new strands are elongated by addition of deoxyribonucleotides to the 3′ end. DNA polymerases require a primer made of RNA by the enzyme primase, and synthesis begins at an origin of replication. The double helix is unwound by helicase and is stabilized by single-strand binding proteins. Extension of the DNA occurs continuously on the leading strand but discontinuously on the lagging strand, resulting in fragments that must be joined together.
Q What is meant by the term semiconservative replication? What are the functions of DNA Pol I and III, helicase, and primase? How does a leading strand differ from a lagging strand?
6.4 Starting from a single origin on a circular chromosome, two replication forks synthesize DNA simultaneously in both directions until the forks meet at the terminus region. The proteins at the replication fork form a large complex known as the replisome. Most errors in base pairing that occur during replication are corrected by the proofreading functions of DNA polymerases.
Q What is the replisome and what does it contain? Why can replication occur faster on circular DNA than on linear DNA? What is proofreading and why is it important?
III RNA Synthesis: Transcription
6.5 In Bacteria, promoters are recognized by the sigma subunit of RNA polymerase. Alternative sigma factors allow joint regulation of large families of genes in response to growth conditions. Transcription by RNA polymerase continues until specific sites called transcription terminators are reached. These terminators function at the level of RNA. In Bacteria and Archaea a single mRNA molecule may encode more than one polypeptide. A cluster of genes that are transcribed together from a single promoter is called an operon.
Q How does RNA polymerase know where to begin transcription? How does it know where to end?
6.6 The transcription apparatus and the promoter architecture of Archaea and Eukarya have many features in common, although the components are usually relatively more simple in Archaea. In contrast, the processing of eukaryotic primary transcripts is unique and has three distinct steps: splicing, capping, and adding a poly(A) tail.
**Q How does the archaeal RNA polymerase differ from that in Bacteria? How does the initiation of transcription in the two domains differ? Why do eukaryotic mRNAs have to be “processed” whereas most prokaryotic RNAs do not?**
IV Protein Synthesis: Translation
6.7 Polypeptide chains contain many amino acids linked via peptide bonds. Twenty-two different amino acids are genetically encoded. The primary structure of a protein is its amino acid sequence, but the higher-order structure (folding) of the polypeptide determines its cellular function.
Q Describe the two types of secondary structure a polypeptide can attain. Which proteins can achieve quaternary structure? Which protein structure(s) are altered by denaturation?
6.8 One or more tRNAs exist for each amino acid incorporated into polypeptides by the ribosome. Enzymes called aminoacyl-tRNA synthetases bond amino acids to their cognate tRNAs.
Q Why are tRNAs important in translation? Do genes for tRNAs have promoters, and are tRNAs translated? What are aminoacyl-tRNA synthetases, what are their substrates, and what do they do?
6.9 The genetic code is expressed as RNA, and a single amino acid may be encoded by several different but related codons. In addition to the stop (nonsense) codons, there is also a specific start codon that signals the initiation of translation.
Q Why is the genetic code a degenerate code? What is wobble and how does it accommodate fidelity in the genetic code?
6.10 Translation occurs on the ribosome and requires mRNA and aminoacyl-tRNAs. The ribosome has three sites: acceptor, peptide, and exit (A, P, and E). During each step of translation, the ribosome advances one codon along the mRNA, and the tRNA in the acceptor site moves to the peptide site. Protein synthesis terminates when a stop codon, which does not have a corresponding tRNA, is reached.
Q Where on the ribosome do tRNAs bind, and what is the energy source that supports translocation?
V Protein Processing, Secretion, and Targeting
6.11 Polypeptides do not remain linear in structure following translation but require proper folding and additional processing for functional activity. Proteins called chaperones help fold some proteins that are unable to fold spontaneously and also assist in the incorporation of cofactors and refolding partially denatured proteins.
Q What proteins are involved in refolding misfolded proteins? What other functions do they have?
6.12 Many proteins need to be transported into or through the cytoplasmic membrane. These proteins contain a signal sequence that is recognized by the cellular translocase systems of Sec and Tat.
Q What are the functions of the Sec and Tat systems and why are they necessary? How do these systems know which proteins to act upon?
6.13 A large diversity of secretion systems are employed by gram-negative bacteria to secrete proteins into or through their outer membrane and even into other cells, and types I–VI secretion systems function in this regard. Types II and V are two-step translocases and rely on the activity of Sec or Tat, whereas types I, III, IV, and VI are one-step translocases. In most cases, energy is required to drive secretion of proteins outside the cell.
Q What are the major differences between one-step and two-step translocases? Which are used to secrete proteins into other cells?
Application Questions
The genome of the bacterium Neisseria gonorrhoeae consists of one double-stranded DNA molecule that contains 2220 kilobase pairs. If 85% of this DNA molecule is made up of the open reading frames of genes encoding proteins, and the average protein is 300 amino acids long, how many protein-encoding genes does Neisseria have? What kind of genetic information is present in the other 15% of the DNA?
Compare and contrast the activity of DNA and RNA polymerases. What is the function of each? What are the substrates of each? What is the main difference in the behavior of the two polymerases?
What would be the result (in terms of protein synthesis) if RNA polymerase initiated transcription one base upstream of its normal starting point? Why? By inspecting Table 6.4, discuss how the genetic code has evolved to help minimize the impact of mutations.
Chapter Glossary
one of the 22 different monomers that are attached to one another by peptide bonds to form proteins; chemically, a two-carbon carboxylic acid containing an amino group and a characteristic substituent on the alpha carbon Aminoacyl-tRNA synthetase
an enzyme that catalyzes attachment of an amino acid to its cognate tRNA Anticodon
a sequence of three bases in a tRNA molecule that base-pairs with a codon during protein synthesis Antiparallel
in reference to double-stranded DNA, the two strands run in opposite directions (one runs 5′→3′ and the complementary strand 3′→5′) Bacteriocin
a toxic protein secreted by bacteria that inhibits or kills other, related bacteria Chaperone
a protein that helps other proteins fold or refold from a partly denatured state Chromosome
a genetic element, usually circular in prokaryotic cells, carrying genes essential to cellular function Codon
a sequence of three bases in mRNA that encodes an amino acid Codon bias
the nonrandom usage of multiple codons encoding the same amino acid; the relative proportions of different codons encoding the same amino acid vary in different organisms. Same as codon usage Complementary
nucleic acid sequences that can base-pair with each other Denaturation
loss of the correct folding of a protein, leading (usually) to protein aggregation and loss of biological activity DNA (deoxyribonucleic acid)
a polymer of deoxyribonucleotides linked by phosphodiester bonds; the deoxyribonucleotide sequence encodes genetic information DNA gyrase
an enzyme found in Bacteria and most Archaea that introduces negative supercoils in DNA DNA helicase
an enzyme that uses ATP to unwind the double helix of DNA DNA ligase
an enzyme that seals nicks in the backbone of DNA DNA polymerase
an enzyme that synthesizes a new strand of DNA in the 5′→3′ direction using an antiparallel DNA strand as a template Exons
the coding DNA sequences in a split gene (contrast with intron) Gene
a segment of DNA specifying a protein (via mRNA), a tRNA, an rRNA, or any other noncoding RNA Genetic code
the correspondence between nucleic acid sequence and amino acid sequence of proteins Genetic element
a structure that carries genetic information, such as a chromosome, a plasmid, or a virus genome Genome
the total complement of genes contained in a cell or virus Informational macromolecule
any large polymeric molecule that carries genetic information, including DNA, RNA, and protein Introns
the intervening noncoding DNA sequences in a split gene (contrast with exons) Lagging strand
the new strand of DNA that is synthesized in short pieces and then joined together later Leading strand
the new strand of DNA that is synthesized continuously during DNA replication Messenger RNA (mRNA)
an RNA molecule that contains the genetic information to encode one or more polypeptides Nonsense codon
a nitrogenous base (adenine, guanine, cytosine, thymine, or uracil) plus a sugar (either ribose or deoxyribose) but lacking phosphate Nucleotide
a monomer of a nucleic acid containing a nitrogenous base (adenine, guanine, cytosine, thymine, or uracil), one or more molecules of phosphate, and a sugar, either ribose (in RNA) or deoxyribose (in DNA) Open reading frame (ORF)
a sequence of DNA or RNA that could be translated to give a polypeptide Operon
two or more genes transcribed into a single RNA and under the control of a single regulatory site Peptide bond
a type of covalent bond linking amino acids in a polypeptide Phosphodiester bond
a type of covalent bond linking nucleotides together in a polynucleotide Plasmid
an extrachromosomal genetic element that is usually not essential to the cell Polynucleotide
a polymer of nucleotides bonded to one another by covalent bonds called phosphodiester bonds Polypeptide
a polymer of amino acids bonded to one another by peptide bonds Primary structure
the precise sequence of monomers in a macromolecule such as a polypeptide or a nucleic acid Primary transcript
an unprocessed RNA molecule that is the direct product of transcription Primase
the enzyme that synthesizes the RNA primer used in DNA replication Primer
an oligonucleotide to which DNA polymerase attaches the first deoxyribonucleotide during DNA synthesis Promoter
a site on DNA to which RNA polymerase binds to commence transcription Protein
a polypeptide or group of polypeptides forming a molecule of specific biological function Purine
one of the nitrogenous bases of nucleic acids that contain two fused rings; adenine and guanine Pyrimidine
one of the nitrogenous bases of nucleic acids that contain a single ring; cytosine, thymine, and uracil Quaternary structure
in proteins, the number and types of individual polypeptides in the final protein molecule Replication
in cells, synthesis of DNA using DNA as a template Replication fork
the site on the chromosome where DNA replication occurs and where the enzymes replicating the DNA are bound to untwisted, single-stranded DNA Replisome
a DNA replication complex that consists of two copies of DNA polymerase III, DNA gyrase, helicase, primase, and copies of single-strand binding protein Ribosomal RNA (rRNA)
the types of RNA found in the ribosome; some participate actively in protein synthesis Ribosome
a cytoplasmic particle composed of ribosomal RNA and protein, whose function is to synthesize proteins RNA (ribonucleic acid)
a polymer of ribonucleotides linked by phosphodiester bonds that plays many roles in cells, in particular, during protein synthesis RNA polymerase
an enzyme that synthesizes RNA in the 5′→3′ direction using a complementary and antiparallel DNA strand as a template RNA processing
the conversion of a primary transcript RNA to its mature form Secondary structure
the initial pattern of folding of a polypeptide or a polynucleotide, usually dictated by opportunities for hydrogen bonding Semiconservative replication
DNA synthesis yielding two new double helices, each consisting of one parental and one progeny strand Signal sequence
a special N-terminal sequence of approximately 20 amino acids that signals that a protein should be incorporated into or exported across the cytoplasmic membrane Spliceosome
a complex of ribonucleoproteins that catalyze the removal of introns from primary RNA transcripts Start codon
a special codon, usually AUG, that signals the start of a protein Stop codon
a codon that signals the end of a protein Termination
stopping the elongation of an RNA molecule at a specific site Tertiary structure
the final folded structure of a polypeptide that has previously attained secondary structure Transcription
the synthesis of an RNA molecule complementary to one of the two strands of a double-stranded DNA molecule Transfer RNA (tRNA)
a small RNA molecule used in translation that possesses an anticodon at one end and has the corresponding amino acid attached to the other end Translation
the synthesis of protein by a ribosome using the genetic information in a messenger RNA as a template Transposable element
a genetic element able to move (transpose) from one site to another on host DNA molecules Wobble
a less rigid form of base pairing allowed only in codon–anticodon pairing