17 Diversity of Archaea
**II Thaumarchaeota and Cryptic Archaeal Phyla**
Methanogens and Global Climate Change
A diversity of methanogens exists within the domain Archaea. Methanogens are among the oldest microbes on Earth and have a tremendous impact on our biosphere. Methanogens in nature promote the anaerobic decomposition of organic matter by consuming the products of fermentative and syntrophic metabolisms. Methanogens release methane as a waste product, and this methanogenesis is a natural part of Earth’s carbon cycle. However, atmospheric methane has been increasing dramatically since the dawn of the industrial age. Methane, like CO2, traps heat in the atmosphere, contributing to the greenhouse effect. However, molecule for molecule, methane traps far more heat than does CO2. Moreover, whereas the increase in atmospheric CO2 comes mostly from burning fossil fuels, the increase in atmospheric methane comes mostly from human activities that promote methanogenesis.

Human activities have created many new opportunities for methanogens. In particular, domesticated ruminants such as cows, sheep, and goats harbor methanogens within their rumens. In addition, during rice production, farmers flood the soil, generating artificial wetlands that release huge amounts of methane. These two agricultural activities alone account for a major fraction of global methane emissions. Remarkably, however, human activities have even created new habitats for methanogens under glaciers. The meltwater from this Greenland glacier (photo) is supersaturated with methane and contains DNA from cells of the Methanomicrobiales, Methanosarcinales, and Methanobacteriales, highly diverse groups of methanogens. As climate change accelerates glacier melting, the meltwater makes available ancient organic carbon buried beneath the ice. This meltwater creates large subglacial wetlands that are rapidly colonized by cold-active methanogens.
Massive amounts of organic carbon have been locked away for eons in frozen Arctic soils. As temperatures on Earth increase, this carbon thaws, creating new habitats for methanogens. Thus, accurately predicting the consequences of global climate change will require us to understand methanogen diversity and how humans affect it.
Source: Lamarche-Gagnon, G., et al. 2019. Greenland melt drives continuous export of methane from the ice-sheet bed. Nature 565: 73.
The domain Archaea is named for the Archaean eon, the period of geological history when life first spread across Earth (Figure 13.1). In the Archaean, high temperatures and an atmosphere devoid of O2 and thick in toxic gases enveloped Earth. Archaea were once thought to be remnants of this forgotten age since many Archaea live in extreme environments such as volcanic systems or salt ponds. We now know, however, that Archaea occupy a wide range of habitats and they perform important biogeochemical reactions in soil, the oceans, wetlands, and even in the guts of animals. While Archaea and Bacteria are both single-celled organisms with prokaryotic cell structure (Chapter 2), these domains are highly differentiated both genetically and physiologically. In many ways, Archaea share more features with Eukarya than with Bacteria. Indeed, it is likely that archaeal cells contributed fundamentally to the origin of the domain Eukarya (Section 13.4 and Figure 13.10). In this chapter, we will learn about the enormous phylogenetic and physiological diversity found within the domain Archaea.
The Archaea are nearly as diverse as the Bacteria, but the vast majority of Archaea have thus far proven difficult to grow in culture. Hence, it is likely that we still have much to learn about the breadth of characteristics in organisms in this domain. To date, most well-characterized species of Archaea come from only two phyla: the Euryarchaeota and the Crenarchaeota (Figure 17.1). In addition, several species have been isolated from the phylum Thaumarchaeota. By contrast, the phyla Korarchaeota and Nanoarchaeota are represented only by strains grown in coculture or studied using enrichment techniques (Section 19.1). The use of metagenomics (Section 10.7) and single-cell genome sequencing (Section 10.11) has led to an explosion in our understanding of the phylogenetic and physiological diversity of the archaeal domain. As a result, many new phyla of Archaea such as representatives of the DPANN, TACK, and Asgard groups (Figure 17.1) have been discovered relatively recently.
Figure 17.1 Schematic representation of the phylogeny of the major taxonomic orders within the domain *Archaea*.

Five major archaeal phyla (studied in pure or highly enriched cultures) and their most representative orders are indicated in color, while other archaeal taxa are indicated in black. While archaeal phyla are nearly as numerous as Bacteria, far fewer Archaea have been cultivated. Archaeal phyla can be organized into four major branches, or superphlya, consisting of the Euryarchaeota, DPANN (represented here by Nanoarchaeota, see Section 17.6), TACK (composed of Thaumarchaeota, Aigarchaeota, Crenarchaeota, Korarchaeota, and related lineages, see Section 17.8), and Asgard Archaea (see Section 17.8). The root of the archaeal tree lies between these four branches though the exact position remains uncertain.
While all Archaea share certain features, this domain encompasses considerable physiological diversity. Common traits of all Archaea include ether-linked lipids, a lack of peptidoglycan in cell walls (Chapter 2), and structurally complex RNA polymerases that resemble those of Eukarya (Section 6.6). These features were likely present in the last common ancestor of the Archaea. Genomic analyses suggest that the earliest Archaea lived at high temperature, oxidized H2 as an energy source, and fixed CO2 using the reductive acetyl-CoA pathway (Section 14.14 and Figure 14.34). Despite their shared phylogenetic origin, contemporary species of Archaea are metabolically diverse. Archaea can be chemoorganotrophs or chemolithotrophs, they can be respiratory or fermentative, and they can be aerobic or anaerobic using a wide diversity of electron donors and acceptors (Chapter 14).
In addition, certain physiological capabilities are uniquely found within the domain Archaea. Methane production, for example, is a unique characteristic of Archaea called methanogens (Section 14.15, Figures 14.38 and 14.39). Methanogenesis evolved very early within the archaeal domain, and all well-characterized methanogens belong to the phylum Euryarchaeota. Methanogenesis is a globally important process that has produced virtually all of the natural gas on Earth and has a significant effect on climate because methane is a strong “greenhouse gas” (Sections 21.1, 21.2; and see page 556). Archaea are also well known for containing many examples of extremophiles (Section 1.5 and Table 1.2) including hyperthermophiles (organisms with growth temperature optima above 80 °C), as well as halophiles, acidophiles, and psychrophiles (Chapter 4).
With this brief background and the phylogeny of Archaea firmly in mind (Figure 17.1), we now consider the organismal diversity of this fascinating domain of life.
I *Euryarchaeota*
**The phylum Euryarchaeota contains most cultured species of Archaea. The group is metabolically diverse and includes methanogens, halophiles, thermophiles, and hyperthermophiles.**
Euryarchaeota comprise a large and physiologically diverse group of Archaea. This phylum includes methanogens as well as many genera of extremely halophilic (salt-loving) Archaea. As a study in physiological contrasts, these two groups are remarkable: Methanogens are the strictest of anaerobes while extreme halophiles are primarily obligate aerobes. Other groups of Euryarchaeota include the hyperthermophiles Thermococcus and Pyrococcus, the hyperthermophilic methanogen Methanopyrus, and the cell wall–less Thermoplasma, an organism phenotypically similar to the mycoplasmas (Section 16.9). We begin our review of Euryarchaeota by reviewing the extremely halophilic Archaea.
Key Genera: *Halobacterium, Haloferax, Natronobacterium*
Extremely halophilic Archaea, often given the nickname “haloarchaea,” are a diverse group that inhabits environments high in salt. These include naturally salty environments, such as solar salt evaporation ponds and salt lakes, and artificial saline habitats such as the surfaces of heavily salted foods, for example, certain fish and meats. Such salty habitats are called hypersaline (Figure 17.2). The term extreme halophile is used to indicate that these organisms not only are halophilic but require a very high level of salt, in some cases at levels near saturation (Section 4.15 and Figure 4.27).
Figure 17.2 Hypersaline habitats for halophilic *Archaea*.

Hypersaline habitats are home to the halophilic Archaea. These organisms not only tolerate salt but require salt, and typically in large amounts. (a) The north arm of Great Salt Lake, Utah, a hypersaline lake in which the ratio of ions is similar to that of seawater, but in which absolute concentrations of ions are several times that of seawater. The green color is primarily from cells of cyanobacteria and green algae. (b) Aerial view near San Francisco Bay, California, of a series of seawater evaporating ponds where solar salt is prepared. The red-purple color is predominantly due to bacterioruberins and bacteriorhodopsin in cells of haloarchaea. (c) Lake Hamara, Wadi El Natroun, Egypt. A bloom of pigmented haloalkaliphiles is growing in this pH 10 soda lake. Note the deposits of trona (NaHCO3⋅Na2CO3⋅2 H2O) around the edge of the lake. (d) Scanning electron micrograph of halophilic bacteria including square Archaea present in a saltern in Spain.
An organism is considered an extreme halophile if it requires 1.5 M (about 9%) or more sodium chloride (NaCl) for growth. Most species of extreme halophiles require 2–4 M NaCl (12–23%) for optimal growth and can grow at salinities as high as 5.5 M NaCl (32%, the limit of saturation for NaCl), although some species grow very slowly at this salinity. Some phylogenetic relatives of extremely halophilic Archaea, for example species of Haloferax and Natronobacterium, are able to grow at much lower salinities, such as at or near that of seawater (about 2.5% NaCl).
Hypersaline Environments: Chemistry and Productivity
Hypersaline habitats are common throughout the world, but extremely hypersaline habitats are rare. Most such environments are in hot, dry areas of the world. Salt lakes can vary considerably in ionic composition. The predominant ions in a hypersaline lake depend on the surrounding topography, geology, and general climatic conditions. Great Salt Lake in Utah (USA) (Figure 17.2a), for example, is essentially concentrated seawater. In this hypersaline lake the relative proportions of the various ions [e.g., sodium (Na+), chloride (Cl−), and sulfate (SO4 2−)] are those of seawater, although the overall concentration of ions is much higher. In addition, the pH of this hypersaline lake is slightly alkaline.
Soda lakes, in contrast, are highly alkaline hypersaline environments. The water chemistry of soda lakes resembles that of hypersaline lakes such as Great Salt Lake, but because high levels of carbonate minerals are also present in the surrounding strata, the pH of soda lakes is quite high. Waters of pH 10–12 are not uncommon in these environments (Figure 17.2c). In addition, the divalent cations Ca2+ and Mg2+ are present in only trace amounts in soda lakes because they precipitate out as carbonate salts at high pH and carbonate concentrations.
The diverse chemistries of hypersaline habitats have selected for a large diversity of halophilic microorganisms. Some organisms are unique to one environment while others are widespread. Moreover, despite their extreme conditions, salt lakes can be highly productive ecosystems (the word productive here means high levels of autotrophic CO2 fixation). Archaea are not the only microorganisms present. The eukaryotic alga Dunaliella (Figure 18.34a) is the major, if not the sole, oxygenic phototroph in most salt lakes. In highly alkaline soda lakes where Dunaliella is absent, anoxygenic phototrophic purple bacteria of the genera Ectothiorhodospira and Halorhodospira (Section 15.4) predominate. Organic matter originating from primary production by oxygenic or anoxygenic phototrophs sets the stage for growth of haloarchaea, which are chemoorganotrophic organisms. In addition, a few extremely halophilic chemoorganotrophic Bacteria, such as Halanaerobium, Halobacteroides, and Salinibacter, thrive in such environments.
Marine salterns are also habitats for extreme halophiles. Marine salterns are enclosed basins filled with seawater left to evaporate, eventually yielding solar sea salt (Figure 17.2b, d). As salterns approach the minimum salinity limits for haloarchaea, the waters turn a reddish-purple color due to the massive growth—called a bloom—of cells (the red coloration apparent in Figure 17.2b and c is due to carotenoids and other pigments to be discussed later). Morphologically unusual Archaea are often present in salterns, including species with a square or cup-shaped morphology (Figure 17.2d). Extreme halophiles are also present in highly salted foods, such as certain types of sausages, marine fish, and salted pork.
Taxonomy and Physiology of Extremely Halophilic *Archaea*
The extremely halophilic Archaea are found within the taxonomic orders Halobacteriales, Natrialbales, and Haloferacales, which share a common ancestor within the Euryarchaeota (Figure 17.1). These three orders constitute the haloarchaea, but they are also sometimes called “halobacteria” because the genus Halobacterium (Figure 17.3), the best-studied representative of the extreme halophiles, was discovered and characterized long before the Archaea were recognized as an independent domain. Many genera of Natrialbales, including Natronobacterium, Natronomonas, and their relatives, differ from other extreme halophiles in being extremely alkaliphilic as well as halophilic. As befits the geochemistry of their soda lake habitat (Figure 17.2c), these natronobacteria grow optimally at low Mg2+ concentrations and alkaline pH (9–11).
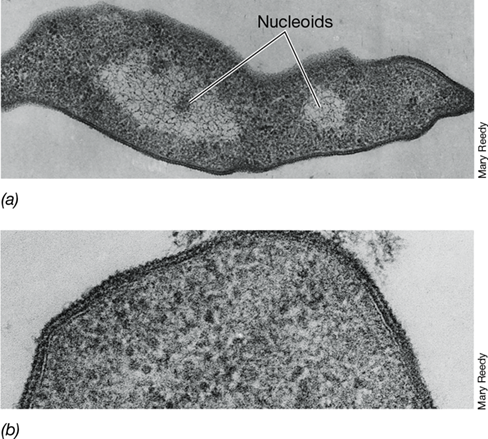
Figure 17.3 Electron micrographs of thin sections of the extreme halophile *Halobacterium salinarum*.


A cell is about 0.8 μm in diameter. (a) Longitudinal section of a dividing cell showing the nucleoids. (b) High-magnification electron micrograph showing the glycoprotein subunit structure of the cell wall. Archaeal cell walls are discussed in Section 2.5.
Haloarchaea stain gram-negatively, reproduce by binary fission, and do not form spores. Cells of the various cultured genera are rod-shaped, cocci, or cup-shaped, but even cells that form squares are known (Figure 17.2d). Cells of Haloquadratum are square and only about 0.1 μm thick. Haloquadratum also forms gas vesicles (Section 2.7) that allow cells of this organism to float in its hypersaline habitat, probably as a means to be in contact with air since most extreme halophiles are obligate aerobes. Many other extremely halophilic Archaea also produce gas vesicles. A few strains of extreme halophiles are weakly motile by archaella, the archaeal analog of bacterial flagella, that rotate to propel the cell forward (Section 2.9), but most halophiles lack archaella. The genomes of Halobacterium and Halococcus are unusual in that large plasmids (Section 6.2) containing up to 30% of the total cellular DNA are present and the G+C base ratio of these plasmids (near 60% G+C) differs significantly from that of chromosomal DNA (66–68% G+C). Plasmids from extreme halophiles are among the largest naturally occurring plasmids known.
Most haloarchaea use amino acids or organic acids as electron donors aerobically and require a number of growth factors such as vitamins for optimal growth. A few haloarchaea oxidize carbohydrates aerobically, but this capacity is rare; sugar fermentation does not occur. Electron transport chains containing cytochromes of the a, b, and c types are present in Halobacterium, and energy is conserved via a proton motive force arising from electron transport (Sections 3.8 and 3.9). Some haloarchaea can grow anaerobically, as growth by anaerobic respiration linked to the reduction of nitrate or fumarate (Sections 3.10, 14.11, and 14.13) has been demonstrated in certain species.
Water Balance in Extreme Halophiles
Extremely halophilic Archaea require large amounts of NaCl for growth. Detailed salinity studies of Halobacterium have shown that the requirement for Na+ cannot be satisfied by any other ion, even the chemically related ion potassium (K+). However, cells of Halobacterium need both Na+ and K+ for growth because each plays an important role in maintaining osmotic balance.
As we learned in Section 4.15, microbes must withstand the osmotic forces they face in their habitats. To do so in a high-solute environment such as the salt-rich habitats of Halobacterium, organisms must either accumulate or synthesize solutes intracellularly. These solutes are called compatible solutes (Table 4.6). These compounds counteract the tendency of the cell to become dehydrated under conditions of high osmotic strength by placing the cell in positive water balance with its surroundings. Cells of Halobacterium, however, do not synthesize or accumulate organic compounds but instead pump large amounts of K+ from the environment into the cytoplasm. This ensures that the concentration of K+ inside the cell is even greater than the concentration of Na+ outside the cell (Table 17.1). This ionic condition maintains positive water balance.
Table 17.1 Concentration of ions in cells of *Halobacterium salinarum*a


aData from Christian, J.H.B, and Waltho, J.A. 1962. Biochim. Biophys. Acta 65: 506.
The Halobacterium cell wall (Figure 17.3b) is composed of glycoprotein and is stabilized by Na+. Sodium ions bind to the outer surface of the Halobacterium wall and are absolutely essential for maintaining cellular integrity. When insufficient Na+ is present, the cell wall breaks apart and the cell lyses. This is a consequence of the exceptionally high content of the acidic (negatively charged) amino acids aspartate and glutamate in the glycoprotein of the Halobacterium cell wall. The negative charge on the carboxyl group of these amino acids is bound to Na+; when Na+ is diluted away, the negatively charged parts of the proteins tend to repel each other, leading to cell lysis.
Mastering Microbiology
Art Activity: Figure 17.4 Model for the mechanism of bacteriorhodopsin
Halophilic Cytoplasmic Components
Like cell wall proteins, cytoplasmic enzymes of Halobacterium are highly acidic, but it is K+, not Na+, that is required for activity. This makes sense because K+ is the predominant cation in the cytoplasm of cells of Halobacterium (Table 17.1). Besides having a highly acidic amino acid composition, halobacterial cytoplasmic proteins typically contain lower levels of hydrophobic amino acids and lysine, a positively charged (basic) amino acid, than do proteins of nonhalophiles. This is also to be expected because in a highly ionic cytoplasm, more polar proteins would tend to remain in solution whereas less polar proteins would tend to cluster and perhaps lose activity. The ribosomes of Halobacterium also require high K+ levels for stability, whereas ribosomes of nonhalophiles have no K+ requirement.
Extremely halophilic Archaea are thus well adapted to life in a highly ionic environment. Cellular components exposed to the external environment require high Na+ levels for stability, whereas cytoplasmic components require high K+ levels. With the exception of a few extremely halophilic species of Bacteria that also use K+ as a compatible solute, in no other group of prokaryotic cells do we find this unique requirement for such high amounts of specific cations.
Bacteriorhodopsin and Light-Mediated ATP Synthesis in Haloarchaea
Certain species of haloarchaea can catalyze a light-driven synthesis of ATP. This form of phototrophy is not linked to CO2 fixation, and does not require chlorophyll pigments, and so it is not photosynthesis in the traditional sense. However, other light-sensitive pigments are present, including red and orange carotenoids—primarily C50 pigments called bacterioruberins—and inducible pigments involved in energy conservation; we discuss these pigments here.
Under conditions of low aeration, Halobacterium salinarum and some other haloarchaea synthesize a protein called bacteriorhodopsin and insert it into their cytoplasmic membranes. Bacteriorhodopsin is so named because of its structural and functional similarity to rhodopsin, the visual pigment of the eye. Conjugated to bacteriorhodopsin is a molecule of retinal, a carotenoid-like molecule that can absorb light energy and pump a proton across the cytoplasmic membrane. The retinal gives bacteriorhodopsin a purple hue. Thus cells of Halobacterium that are switched from growth under high-aeration conditions to O2-limiting growth conditions (a trigger of bacteriorhodopsin synthesis) gradually change color from orange-red to purple-red (Figure 17.2) as they synthesize bacteriorhodopsin and insert it into their cytoplasmic membranes.
Bacteriorhodopsin absorbs green light around 570 nm. Following absorption, the retinal of bacteriorhodopsin, which normally exists in a trans configuration (RetT), becomes excited and converts to the cis (RetC) form (Figure 17.4). This transformation is coupled to the translocation of a proton across the cytoplasmic membrane. The retinal molecule then decays to the trans isomer along with the uptake of a proton from the cytoplasm, and this completes the cycle and resets the proton pump to repeat the process (Figure 17.4). As protons accumulate on the outer surface of the membrane, a proton motive force is generated that is coupled to ATP synthesis through the activity of a proton-translocating ATPase (Figure 17.4; Section 3.9).
Figure 17.4 Model for the mechanism of bacteriorhodopsin.

Light of 570 nm (hv570nm) converts the protonated retinal of bacteriorhodopsin from the trans form (RetT) to the cis form (RetC), along with translocation of a proton to the outer surface of the cytoplasmic membrane, thus establishing a proton motive force. ATPase activity is driven by the proton motive force.
Bacteriorhodopsin-mediated ATP production in H. salinarum supports slow growth of this organism under anoxic conditions. The light-stimulated proton pump of H. salinarum also functions to pump Na+ out of the cell by activity of a Na+−H+ antiport system and to drive the uptake of nutrients, including the K+ needed for osmotic balance. Amino acid uptake by H. salinarum is indirectly driven by light as well, because amino acids are cotransported into the cell with Na+ by an amino acid−Na+ symporter (Section 2.2); removal of Na+ from the cell occurs by way of the light-driven Na+−H+ antiporter.
Other Rhodopsins
Besides bacteriorhodopsin, at least three other rhodopsins are present in the cytoplasmic membrane of cells of H. salinarum. Halorhodopsin is a light-driven chloride (Cl−) pump that brings Cl− into the cell as the anion for K+. The retinal of halorhodopsin binds Cl− and transports it into the cell. Two other light sensors, called sensory rhodopsins, are present in H. salinarum. These light sensors control phototaxis (movement toward light, Section 2.12) by the organism. Through the interaction of a cascade of proteins similar to those in chemotaxis (Sections 2.11 and 7.6), sensory rhodopsins affect archaellar rotation, moving cells of H. salinarum toward light where bacteriorhodopsin can function to make ATP (Figure 17.4).
We will learn when we consider marine microbiology (Sections 20.11 and 20.12) that diverse species of chemoorganotrophic Bacteria that inhabit the upper layers of the ocean contain bacteriorhodopsin-like proteins called proteorhodopsins. As far as is known, proteorhodopsin functions like bacteriorhodopsin except that different spectral forms exist, each form being tuned to the absorption of its own specific wavelength of light. Although the energy generated from proteorhodopsin alone is insufficient to sustain growth, these marine bacteria use proteorhodopsin as a supplement to the ATP they generate from respiration. Proteorhodopsin as a mechanism for energy conservation in marine bacteria makes good ecological sense because levels of dissolved organic matter in the open oceans are typically very low, and thus a strictly chemoorganotrophic lifestyle would be difficult.
Check Your Understanding
Why do cells of Halobacterium require high levels of Na+ for growth?

What benefit does bacteriorhodopsin confer on Halobacterium salinarum?
Key Genera: *Methanobacterium, Methanocaldococcus, Methanosarcina, Methanopyrus*
Many Euryarchaeota are methanogens, microorganisms that produce methane (CH4) as an integral part of their energy metabolism (methane production is called methanogenesis). In Section 14.15 we considered the unique biochemistry of methanogenesis. Later, we will learn how methanogenesis is a major component of the global carbon cycle, serving as the terminal step in the biodegradation of organic matter in many anoxic habitats (Sections 21.1, 21.2, and 21.9). Methanogens are important in a wide range of anoxic habitats including freshwater sediments, wetlands, rice paddies, wastewater treatment plants, geothermal systems, the subsurface of the Earth’s crust, and within the guts of many animals.
Diversity and Physiology of Methanogens
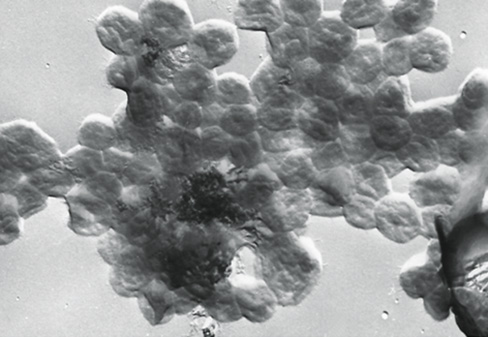
Methanogens, often called by their nickname “methanoarchaea,” occur in at least eight taxonomic orders, including Methanobacteriales, Methanococcales, Methanopyrales, Methanomassiliicoccales, Methanomicrobiales, Methanocellales, Methanosarcinales, and Methanonatronarchaeales (Figure 17.1). Methanogens exhibit considerable morphological and physiological diversity (Figure 17.5 and see Table 17.2). As can be seen in Figure 17.1, methanogens are widespread within the Euryarchaeota and do not represent a single coherent phylogenetic group. We have already been introduced to the factors that cause inconsistency between phylogenetic diversity and functional diversity (Section 15.1). In the case of methanogenesis it is most likely that the ability to reduce CO2 to CH4 evolved only once within the Euryarchaeota, and that gene loss caused lineages such as the haloarchaea and Thermoplasmatales to lose the capacity to produce methane. We will also see that Archaeoglobus (Section 17.4) still retains some of the genes encoding methanogenesis and can actually produce methane under certain growth conditions.
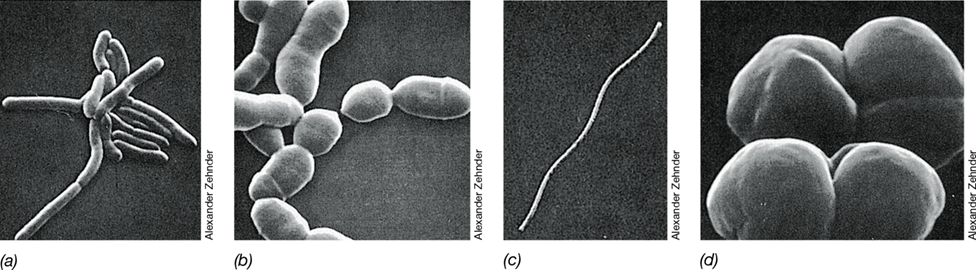
Figure 17.5 Scanning electron micrographs of cells of diverse species of methanogenic *Archaea*.

(a) Methanobrevibacter ruminantium. A cell is about 0.7 μm in diameter. (b) Methanobrevibacter arboriphilus. A cell is about 1 μm in diameter. (c) Methanospirillum hungatei. A cell is about 0.4 μm in diameter. (d) Methanosarcina barkeri. A cell is about 1.7 μm wide.
Methanogens have diverse physiological characteristics, but they are united by their ability to produce methane and by their intolerance to O2. Several cofactors required by methanogenic pathways are inhibited by O2, and hence all methanogens are obligate anaerobes. As a result, strict anoxic techniques are necessary for their cultivation in isolation. While most methanogens are mesophilic and do not live in extreme environments, a diversity of species have been described including some that grow optimally at very high (see Figure 17.7) or very low temperatures, at very high salt concentrations, or at extremes of pH.
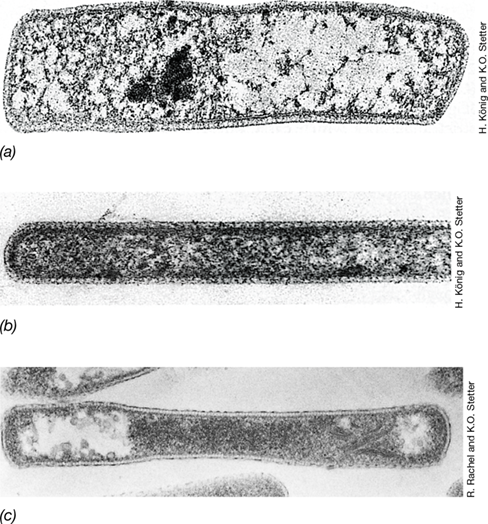
Methanogens possess a diversity of cell envelope and cell wall configurations. Methanobacterium species and relatives (**Figure 17.6*a***) have cell walls composed of pseudomurein (Section 2.3). Methanosarcina and relatives (Figure 17.6b) have cell walls composed of methanochondroitin (so named because of its structural resemblance to chondroitin, the connective tissue polymer of vertebrate animals). In addition, Methanocaldococcus (**Figure 17.7*a***) and Methanoplanus species have cell walls composed of protein or glycoprotein, respectively. Finally, species such as Methanospirillum (Figure 17.5c) use an S-layer (Section 2.5) as their cell wall.
Figure 17.6 Transmission electron micrographs of thin sections of methanogenic *Archaea*.

(a) Methanobrevibacter ruminantium. A cell is 0.7 μm in diameter. (b) Methanosarcina barkeri, showing the thick cell wall and the manner of cell segmentation and cross-wall formation. A cell is 1.7 μm in diameter.
Figure 17.7 Hyperthermophilic and thermophilic methanogens.

(a) Methanocaldococcus jannaschii (temperature optimum, 85 °C), shadowed preparation electron micrograph. A cell is about 1 μm in diameter. (b) Methanotorris igneus (temperature optimum, 88 °C), thin section. A cell is about 1 μm in diameter. (c) Methanothermus fervidus (temperature optimum, 88 °C), thin-sectioned electron micrograph. A cell is about 0.4 μm in diameter. (d) Methanosaeta thermophila (temperature optimum, 60 °C), phase-contrast micrograph. A cell is about 1 μm in diameter. The refractile bodies inside the cells are gas vesicles.
Methanogenic Pathways
Methanogenesis occurs through three different pathways (Table 17.2): CO2 reduction (Figure 14.38), methylotrophic methanogenesis (Figure 14.39a), and acetoclastic methanogenesis (Figure 14.39b). Each of these pathways relies on coenzyme M; methane is ultimately produced by reduction of methyl-CoM to methane (Section 14.15). Through these pathways, methanogens convert a limited number of substrates into methane (Table 17.2). Interestingly, these substrates do not include such common compounds as glucose, organic acids (other than pyruvate), or fatty acids (other than acetate). Methanogens often form syntrophic associations with fermentative anaerobes (Section 14.22, Section 21.2). In these cooperative systems, the fermentative organisms degrade a wide range of organic molecules into H2, CO2, and acetate, which are ultimately used as substrates for methanogenesis.
Table 17.2 The three methanogenic pathways and their substrates

aIn a dismutation, one part of the molecule is reduced by oxidizing another part of the same molecule.
The three methanogenic pathways are distributed across different phylogenetic groups of methanogens (Table 17.3). Methanogenesis by CO2 reduction occurs widely across the known diversity of methanogens, but not all methanogens can reduce CO2. Notably, many species of Methanosarcinales, and all species in the Methanomassiliicoccales, Methanonatronarchaeales, and the genus Methanosphaera have lost the ability to produce methane by CO2 reduction (Table 17.3). The CO2 reduction pathway also allows some methanogens to produce methane from formate or carbon monoxide. In addition, while H2 is the typical electron donor for methanogenesis by CO2 reduction, this pathway also allows some methanogens to reduce CO2 by using electrons from pyruvate or certain alcohols (Tables 17.2 and 17.3).
Table 17.3 Characteristics of some methanogenic *Archaea*a

aTaxonomic orders are listed in bold. An order is a taxonomic rank that consists of several families; families consist of several genera (Table 13.1).
bMethanogenic pathways I–III are summarized in Table 17.2.
cMethylamines can include the substrates methylamine (CH3NH3+), dimethylamine ((CH3)2NH2+), and trimethylamine ((CH3)3NH+); methyl sulfides can include dimethyl sulfide ((CH3)2S) and methyl mercaptan (CH3SH).
Acetoclastic methanogenesis is performed by few methanogens and is found exclusively within the Methanosarcinales (Table 17.3). Methanogenesis from acetate is a dominant source of methane production in a wide range of environments (Section 21.2). In acetoclastic methanogenesis, electron flow is branched, producing both CO2 and CH4 as products. Acetoclastic methanogens perform a dismutation of acetate in which the carboxyl group is oxidized to CO2 while the methyl group is reduced to CH4 (Table 17.2, Figure 14.39b). The reductive acetyl-CoA pathway is then used to assimilate carbon into cell biomass.
Methylotrophic methanogenesis can take many forms, but all methylotrophic methanogens use one of two pathways that differ in important ways. Some methylotrophic methanogens use a branched pathway that produces both CH4 and CO2 as products and allows for carbon assimilation from methylated compounds. Other methylotrophic methanogens use a methyl-reducing pathway that produces only CH4 as a product and does not allow for carbon assimilation from methylated compounds. The branched pathway is found only within the family Methanosarcinales. Branched pathways for methylotrophic methanogenesis are similar conceptually to the acetoclastic pathway with a major difference being that methylotrophs do not perform a dismutation, instead oxidizing one molecule to CO2 so that they can reduce other molecules to CH4 (Table 17.2). For example, many species of Methanosarcinales oxidize one molecule of CH3OH to CO2 to generate the six electrons needed to reduce three molecules of CH3OH to 3 CH4 and 3 H2O (Figure 14.39a). In addition, methylotrophic methanogens that use a branched pathway funnel carbon from their oxidative branch into the reductive acetyl-CoA pathway in order to assimilate carbon from methylated compounds.
The methyl-reducing pathway for methylotrophic methanogenesis, in contrast, does not result in the production of CO2, and methanogens using the methyl-reducing pathway lack the ability to assimilate carbon from methylated compounds. Methylotrophic methanogens such as Methanosphaera stadtmanae are unable to oxidize methyl groups to CO2 and therefore require an external electron donor—either H2 or formate—to reduce the methyl group to methane (Tables 17.2 and 17.3). Hence, M. stadtmanae produces CH4 but not CO2 from methylated substrates. In addition, although M. stadtmanae and metabolically similar methanogens can make methane from methylated compounds, they are unable to incorporate them as a carbon source. Instead, they typically require an organic carbon source such as acetate, though a few species can also fix CO2. Besides M. stadtmanae, this pattern of methylotrophic methanogenesis is also characteristic of Methanomicrococcus blatticola and species in the orders Methanomassiliicoccales and Methanonatronarchaeales (Table 17.3).
*Methanocaldococcus jannaschii* as a Model Methanogen
The genomes of the hyperthermophilic methanogen Methanocaldococcus jannaschii (Figure 17.7a) and many other methanogens have been sequenced. The 1.66-megabase-pair (Mbp) circular genome of M. jannaschii, an organism that has been used as a model for the molecular study of methanogenesis and archaeal motility, contains about 1700 genes, and genes encoding enzymes of methanogenesis and several other key cell functions have been identified. Interestingly, the majority of M. jannaschii genes encoding functions such as central metabolic pathways and cell division are similar to those in Bacteria. By contrast, most of the M. jannaschii genes encoding core molecular processes such as transcription and translation more closely resemble those of eukaryotes. These findings reflect the various traits shared by organisms in the three cellular domains and are consistent with our understanding of how the three domains of cells evolved, as discussed in Chapter 13. However, analyses of the M. jannaschii genome also show that fully 40% of its genes have no counterparts in genes from either of the other domains. Some of these are genes that encode the enzymes needed for methanogenesis, of course, but many others likely encode novel cellular functions absent from cells in the other domains or may encode redundant functions carried out by classes of enzymes distinct from those found in Bacteria and Eukarya.
*Methanopyrus*, a Hyperthermophilic Methanogen
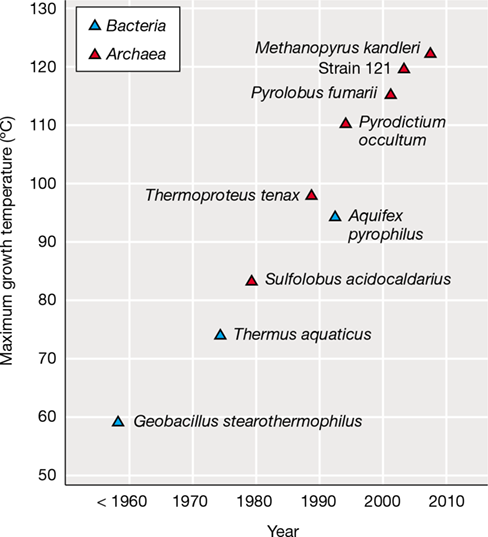
Methanopyrus (Figure 17.8), the only genus in the order Methanopyrales, is a rod-shaped hyperthermophilic methanogen that shares phenotypic properties with both the hyperthermophiles (see Section 17.12) and the methanogens. Methanopyrus was isolated from hot sediments near submarine hydrothermal vents and from the walls of “black smoker” hydrothermal vent chimneys (Section 17.11; Section 20.16 and Figure 20.46). Methanopyrus produces CH4 only from H2+CO2 and grows rapidly for an autotrophic organism (generation time, 1 h at 100 °C). In special pressurized vessels, growth of one strain of Methanopyrus has been recorded at 122 °C, the highest temperature yet shown to support microbial growth (Sections 17.12 and 17.13).
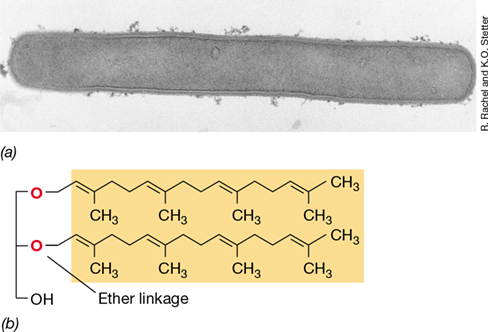
Figure 17.8 *Methanopyrus*.

Methanopyrus grows optimally at 100 °C and can make CH4 only from CO2+H2. (a) Electron micrograph of a cell of Methanopyrus kandleri, the most thermophilic of all known organisms (upper temperature limit, 122 °C). This cell measures 0.5×8 μm. (b) Structure of the novel lipid of M. kandleri. This is the normal glycerol diether of Archaea (Figure 2.3b) except that the side chains are an unsaturated form of phytanyl (called geranylgeraniol) rather than phytanyl.
In addition to its remarkable tolerance to high temperature, Methanopyrus is also unusual because it contains membrane lipids found in no other known organism. Recall that in the lipids of Archaea, the glycerol side chains contain phytanyl rather than fatty acids bonded in ether linkage to the glycerol (Section 2.1 and Figure 2.3a). In Methanopyrus, this ether-linked lipid is an unsaturated form of the otherwise saturated biphytanyl tetraethers found in all other hyperthermophilic Archaea (Figure 17.8b). These unusual lipids are thought to help stabilize the cytoplasmic membrane of Methanopyrus at its unusually high growth temperatures.
Some Euryarchaeota lack cell walls and we consider them next.
Check Your Understanding
What are the three major pathways of methanogenesis and in what phylogenetic groups are they found?
What physiological and structural features distinguish the Methanosarcinales from other methanogens?
Key Genera: *Thermoplasma, Picrophilus, Ferroplasma*
A phylogenetically distinct line of Archaea contains thermophilic and extremely acidophilic genera: Thermoplasma, Ferroplasma, and Picrophilus. These organisms are among the most acidophilic of all known microbes, with Picrophilus being capable of growth even below pH 0. Most are thermophilic as well. These genera also form their own taxonomic order within the Euryarchaeota, the Thermoplasmatales (Figure 17.1). We begin with a description of the mycoplasma-like organisms Thermoplasma and Ferroplasma.
*Archaea* Lacking Cell Walls
Thermoplasma and Ferroplasma lack cell walls, and in this respect they resemble the mycoplasmas (Section 16.9). Thermoplasma (Figure 17.9) is a chemoorganotroph that grows optimally at 55 °C and pH 2 in complex media. Two species of Thermoplasma have been described, Thermoplasma acidophilum and Thermoplasma volcanium. Species of Thermoplasma are facultative aerobes, growing either aerobically or anaerobically by sulfur respiration (Section 14.12). Most strains of T. acidophilum have been obtained from self-heating coal refuse piles. Coal refuse contains coal fragments, pyrite (FeS2), and other organic materials extracted from coal. When dumped into piles in surface mining operations, coal refuse heats as a result of microbial metabolism bringing it to combustion temperature (Figure 17.10). This sets the stage for growth of Thermoplasma, which likely metabolizes organic compounds leached from the hot coal refuse. The second species, T. volcanium, has been isolated in hot acidic soils throughout the world and is highly motile by multiple archaella (Figure 17.9b).
Figure 17.9 *Thermoplasma* species.

(a) Thermoplasma acidophilum, an acidophilic and thermophilic mycoplasma-like archaeon; electron micrograph of a thin section. The diameter of cells varies from 0.2 to 5 μm. The cell shown is about 1 μm in diameter. (b) Shadowed preparation of cells of Thermoplasma volcanium isolated from hot springs. Cells are 1–2 μm in diameter. Notice the abundant archaella (the rotating structures that confer swimming motility on Archaea, Section 2.9) and irregular cell morphology.
Figure 17.10 A typical self-heating coal refuse pile, habitat of *Thermoplasma*.

The pile, containing coal debris, pyrite, and other microbial substrates, self-heats as a result of microbial metabolism.
To survive the osmotic stresses of life without a cell wall and to withstand the dual environmental extremes of low pH and high temperature, Thermoplasma has evolved a unique cytoplasmic membrane structure. The membrane contains a lipopolysaccharide-like material called lipoglycan. This substance consists of glycolipids containing sugars such as mannose and glucose (Figure 17.11), and these glycolipids form a tetraether lipid monolayer membrane. The hydrophobic core of this glycolipid consists of biphytanyl (Section 2.1 and Figure 2.3b). In Thermoplasma and similar organisms such as Sulfolobus, this basic biphytanyl structure can be modified to include one to four cyclopentane rings, with the number of rings tending to increase in proportion to the temperature of the environment. These glycolipids constitute a major fraction of the total lipids of Thermoplasma. The membrane also contains glycoproteins and glycophospholipids, but not sterols. These molecules render the Thermoplasma membrane stable to hot, acidic conditions.
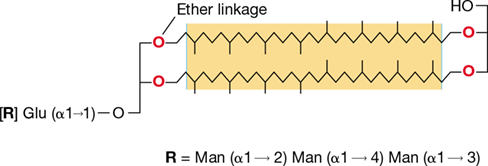
Figure 17.11 Structure of a tetraether glycolipid of *Thermoplasma acidophilum*.

The dominant glycolipids of T. acidophilum have two polar head groups that are connected by ether linkages to a hydrophobic core. This structure causes them to form a thermostable lipid monolayer (Figure 2.3b). One or both of the polar head groups typically contains a mono- or oligosaccharide that contains glucose (Glu), mannose (Man), and other sugars. The hydrophobic core consists of the straight-chain caldarchaeol (shown), which can be modified to include one to four cyclopentane rings (not shown).
Like mycoplasmas (Section 16.9), Thermoplasma contains a relatively small genome (1.5 Mbp). In addition, Thermoplasma DNA is complexed with a highly basic DNA-binding protein that organizes the DNA into globular particles resembling the nucleosomes of eukaryotic cells. This protein is homologous to the histone-like DNA-binding protein HU of Bacteria, which plays an important role in organization of the DNA in the cell. In contrast, several other Euryarchaeota contain basic proteins homologous to the DNA-binding histone proteins of eukaryotic cells.
Ferroplasma
Ferroplasma is a chemolithotrophic relative of Thermoplasma. Ferroplasma is a strong acidophile; however, it is not a thermophile, as it grows optimally at 35 °C. Ferroplasma oxidizes ferrous iron (Fe2+) to ferric iron (Fe3+) to obtain energy and uses CO2 as its carbon source. The oxidation of Fe2+ results in acidification of the environment. Ferroplasma grows in mine tailings containing pyrite, which is its energy source. The extraordinary acidophily of Ferroplasma allows it to drive the pH of its habitat down to extremely acidic values. After moderate acidity is generated from Fe2+ oxidation by acidophilic organisms such as Acidithiobacillus ferrooxidans and Leptospirillum ferrooxidans (Sections 21.6, 22.1, and 22.2), Ferroplasma becomes active and subsequently generates the very low pH values typical of acid mine drainage. Acidic waters at pH 0 can be generated by the activities of Ferroplasma.
Picrophilus
A phylogenetic relative of Thermoplasma and Ferroplasma is Picrophilus. Although Thermoplasma and Ferroplasma are extreme acidophiles, Picrophilus is even more so, growing optimally at pH 0.7 and capable of growth at pH values lower than 0. Picrophilus also has a cell wall (an S-layer; Section 2.5, Figures 2.14 and 2.36a) and a much lower DNA G+C base ratio than does Thermoplasma or Ferroplasma. Although phylogenetically related, Thermoplasma, Ferroplasma, and Picrophilus have quite distinct genomes. Two species of Picrophilus have been isolated from acidic Japanese solfataras, and like Thermoplasma, both grow heterotrophically on complex media.
The physiology of Picrophilus is of interest as a model for extreme acid tolerance. Studies of its cytoplasmic membrane point to an unusual arrangement of lipids that forms a highly acid-impermeable membrane at very low pH. By contrast, at moderate acidities such as pH 4, the membranes of cells of Picrophilus become leaky and disintegrate. Obviously, this organism has evolved to survive only in highly acidic habitats and shows the most extraordinary acidophily of any known microbe.
Check Your Understanding
In what ways are Thermoplasma and Picrophilus similar? In what ways do they differ?
How does Thermoplasma strengthen its cytoplasmic membrane to survive without a cell wall?
Key Genera: *Thermococcus, Pyrococcus, Archaeoglobus, Ferroglobus*
A few euryarchaeotes thrive in thermal environments and some are hyperthermophiles. We consider here the Thermococcales and Archaeoglobales, which are two orders of Euryarchaeota that contain hyperthermophilic species (Figure 17.1).
Figure 17.12 Spherical hyperthermophilic *Euryarchaeota* from submarine volcanic areas.

(a) Thermococcus celer; electron micrograph of shadowed cells (note tuft of archaella). (b) Dividing cell of Pyrococcus furiosus; electron micrograph of thin section. Cells of both organisms are about 0.8 μm in diameter.
Pyrococcus (Figure 17.12b) is morphologically similar to Thermococcus. Pyrococcus differs from Thermococcus primarily by its higher temperature requirements; Pyrococcus grows between 70 and 106 °C with an optimum of 100 °C. Thermococcus and Pyrococcus are also metabolically quite similar. Proteins, starch, or maltose are oxidized as electron donors, and S0 is the terminal electron acceptor and is reduced to hydrogen sulfide (H2S). Both Thermococcus and Pyrococcus form H2S when S0 is present, but form H2 when S0 is absent (see Table 17.4).
Figure 17.13 *Archaeoglobales*.

(a) Transmission electron micrograph of the sulfate-reducing hyperthermophile Archaeoglobus fulgidus. The cell measures 0.7 μm in diameter. (b) Freeze-etched electron micrograph of Ferroglobus placidus, a ferrous iron–oxidizing, nitrate-reducing hyperthermophile. The cell measures about 0.8 μm in diameter.
Archaeoglobus and methanogens share some characteristics. We learned in Section 14.15 about the unique biochemistry of methanogenesis. Briefly, this process requires a series of novel coenzymes, and with rare exceptions, these coenzymes have only been found in methanogens. Archaeoglobus, however, also contains many of these coenzymes, and cultures of this organism can actually produce small amounts of CH4. In addition, the genome of Archaeoglobus, which contains about 2400 genes, shares many genes in common with methanogens (Section 17.2). It seems likely that the ancestor of Archaeoglobus was a methanogen that lost many of the genes required for methanogenesis. Furthermore, genome analysis suggests that the ancestor of Archaeoglobus acquired genes for sulfate reduction as a result of horizontal gene transfer from sulfate-reducing bacteria within the Deltaproteobacteria (Section 15.11).
Ferroglobus (Figure 17.13b) is related to Archaeoglobus but is not a sulfate reducer. Instead, Ferroglobus is an iron-oxidizing chemolithotroph, conserving energy from the oxidation of Fe2+ to Fe3+ coupled to the reduction of nitrate (NO3 −) to nitrite (NO2 −) (see Table 17.4). Ferroglobus grows autotrophically and can also use H2 or H2S as electron donor in its energy metabolism. Ferroglobus was isolated from a shallow marine hydrothermal vent and grows optimally at 85 °C.
Ferroglobus is interesting for several reasons, but especially for its ability to oxidize Fe2+ to Fe3+ under anoxic conditions. This process might help explain the origin of some Fe3+ found in ancient rocks dated to before the predicted appearance of cyanobacteria on Earth (Section 13.2). With organisms like Ferroglobus, it would have been possible for Fe2+ oxidation to proceed without the need for O2 as an electron acceptor. The metabolism of Ferroglobus thus has implications for dating the origin of cyanobacteria and the subsequent oxygenation of Earth. Certain anoxygenic phototrophic bacteria can also oxidize Fe2+ under anoxic conditions (Section 14.8), and so several anaerobic routes to ancient Fe3+ are possible. This makes it difficult to estimate when cyanobacteria first appeared on Earth and to what degree nonphototrophic organisms helped trigger the Great Oxidation Event (Figure 13.1).
We move on now to Part II where we survey the most recently recognized groups of Archaea, some of which are thermophiles but many of which inhabit common environments such as soil, lakes, and the oceans.
Check Your Understanding
Compare the energy-yielding metabolisms of Archaeoglobus and Ferroglobus.
II *Thaumarchaeota* and Cryptic Archaeal Phyla
**Many Archaea remain uncultured and are only known from metagenomic analyses of various microbial habitats. Thaumarchaeota, a group of nitrifying Archaea, were first detected in this way, but have now been cultured and found to be the most abundant Archaea in nature.**
Our understanding of Archaea has been revolutionized by the development of molecular phylogeny (Sections 1.15 and 13.11) and methods for studying microorganisms without needing to cultivate them in the laboratory (Sections 19.4, 19.5, 19.6, 19.7 and 19.7 and 19.8). The discovery and characterization of Thaumarchaeota, Nanoarchaeota, and Korarchaeota were all enabled by the use of molecular techniques that target 16S ribosomal RNA genes. From these initial efforts, species representing each of these phyla were subsequently isolated or at least grown in enrichment cultures. In addition, metagenomic and single-cell genome sequencing techniques (Sections 10.7 and 10.11) have made it possible to obtain genome sequence data from organisms without the need to obtain cultures, and this advance has expanded greatly our knowledge of the archaeal domain. We begin our consideration of these newly discovered phyla with the Thaumarchaeota.
Key Genera: *Nitrosopumilus, Nitrososphaera*
Early surveys of 16S ribosomal RNA genes from open-ocean microbial communities resulted in the shocking conclusion that Archaea were abundant and widespread in the oceans. At the time, the archaeal domain was considered to contain only extremophiles and obligate anaerobes, and their presence in O2-rich temperate and even polar oceanic environments was a major surprise. Even more remarkable, these novel Archaea were also discovered to be widespread in soils worldwide.
Phylogenetic analyses of 16S ribosomal RNA gene sequences initially suggested that this novel group of Archaea was a deeply divergent lineage of the Crenarchaeota, a major group of hyperthermophilic Archaea we will consider in Section 17.9. It was only after genome sequence analysis of the marine nitrifier Nitrosopumilus maritimus that it became clear that the Thaumarchaeota constitute a unique phylum of Archaea (Figure 17.1).
Physiological Characteristics of *Thaumarchaeota*
The physiology of Thaumarchaeota remained a mystery until the isolation of Nitrosopumilus maritimus (Figure 17.14). N. maritimus grows chemolithotrophically by oxidizing ammonia (NH3) to nitrite (NO2 −) aerobically, the first step in nitrification (Sections 14.9, 15.10, and Section 21.3). This organism uses CO2 as its sole carbon source, as do most nitrifying Bacteria (Section 15.10). Prior to the discovery of Thaumarchaeota—for over 100 years in fact—microbiologists thought only certain species of Bacteria catalyzed ammonia oxidation. However, unlike ammonia-oxidizing Bacteria such as Nitrosomonas, N. maritimus is adapted to life under extreme nutrient limitation, as would befit an organism indigenous to open ocean waters. N. maritimus can grow at NH3 concentrations that are a hundred times lower than those required by bacterial nitrifiers, and many archaeal nitrifiers are actually inhibited at the higher NH3 concentrations required to support growth of nitrifying species of Bacteria.
Figure 17.14 *Nitrosopumilus maritimus*, a nitrifying species of marine *Thaumarchaeota*.

This organism can oxidize NH3 present at the very low amounts typical of marine environments. (a) Phase-contrast photomicrograph. (b) Scanning electron micrograph. A single cell of N. maritimus is about 0.2 μm in diameter.
Several species of Thaumarchaeota have been isolated and characterized, revealing a number of properties common to this group. Species have been isolated from habitats including the oceans, marine sediments, estuaries, soils, and hot springs. All existing isolates are chemolithotrophic ammonia-oxidizers (Section 14.9), and most species, like N. maritimus, are able to grow at very low concentrations of NH3. The membranes of all Thaumarchaeota also contain a unique lipid called *crenarchaeo**l(Figure 2.3b,c), a compound limited to species of this phylum. In addition, autotrophy inThaumarchaeotais supported by the 3-hydroxypropionate/4-hydroxybutyrate cycle, a finding that further distinguishes archaeal nitrifiers from nitrifyingBacteriathat employ the Calvin cycle for CO2 fixation (Sections 3.12 and 14.2). The 3-hydroxypropionate/4-hydroxybutyrate cycle also allows for the assimilation of organic carbon, and some archaeal nitrifiers have been shown to assimilate pyruvate during mixotrophic growth. Growth temperatures ofThaumarchaeota* vary widely, as some species thrive in polar seas while others inhabit hot spring environments up to about 75 °C.
Nitrososphaera viennensis was the first species of Thaumarchaeota isolated from soil (Figure 17.15). Cells of N. viennensis form irregular coccoids, and the organism is both mesophilic and neutrophilic. N. viennensis is a mixotroph (an organism that can fix CO2 but grows best with an organic carbon source) and it grows as an aerobic chemolithotroph using either ammonia or urea as electron donor. When growing on urea, N. viennensis uses the enzyme urease to produce ammonia, which is the ultimate electron donor. Like marine species of Thaumarchaeota, N. viennensis can grow at low concentrations of NH3, but unlike archaeal marine nitrifiers, N. viennensis can also tolerate high levels (up to 10 mM) of ammonium at neutral pH. Hence, N. viennensis and related archaeal nitrifiers may be active in soils containing high levels of ammonia, and in these environments, they likely compete directly with bacterial nitrifiers. N. viennensis was isolated from a backyard garden soil (Figure 17.15a) by using cultivation-independent molecular analyses to monitor its growth in different culture media and tailor enrichment media to the growth needs of the organism. This discovery reminds us that when we open our eyes to the microbial world, major discoveries are often “just underfoot.”
Figure 17.15 *Nitrososphaera viennensis*, a nitrifying species of soilborne *Thaumarchaeota*.

This organism can oxidize NH3 and urea across a wide range of concentrations. (a) A Viennese garden soil used to enrich N. viennensis. (b) Cells of N. viennensis can take on many shapes (a condition called pleomorphism), as shown in this transmission electron micrograph of sectioned cells. A single cell of N. viennensis is about 0.5 μm in diameter.
Figure 17.16 Cold-dwelling *Thaumarchaeota*.

(a) Photo of the Antarctic Peninsula taken from shipboard. The frigid waters that lie under the surface ice are habitats for cold-dwelling Thaumarchaeota. (b) Fluorescence photomicrograph of seawater treated with a fluorescent FISH probe (Section 19.5) specific for species of Thaumarchaeota (green cells). Blue cells are stained with DAPI, a fluorescent DNA stain that stains all cells.
Thaumarchaeota are also common in soils, sometimes outnumbering nitrifying Bacteria by 1000-fold, and are found across a wide range of soil pH, from about 3 to 9. While widely present in soils, Thaumarchaeota may be particularly important in acid soils (pH<5.5), which make up more than 30% of all soils. Nitrifiers oxidize NH3, but at low pH, NH4 + predominates and is thus unavailable for nitrification. While nitrification occurs in acid soils, and often at high rates, bacterial nitrifiers have not been observed to grow below pH 6.3. In contrast, the thaumarchaeotal species Nitrosotalea devanaterra, isolated from acidic agricultural soil, grows optimally at pH 4–5. The ability of Thaumarchaeota to grow at low NH3 concentrations explains how they can be successful in acidic soils where free NH3 is present in very low concentration.
Check Your Understanding
How does the organism Nitrosopumilus maritimus conserve energy and obtain carbon?
In what environments might you expect to find species of Thaumarchaeota?
Key Genus: *Nanoarchaeum*
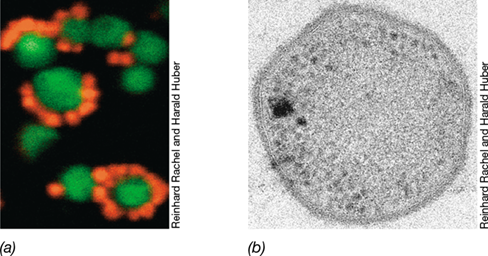
The Nanoarchaeota are represented by a single species, the highly unusual Nanoarchaeum equitans (Figure 17.17). N. equitans is one of the smallest cellular organisms known and has the smallest genome among species of Archaea (0.49 Mb). The coccoid cells of N. equitans are very small, about 0.4 μm in diameter, and have only about 1% of the volume of an Escherichia coli cell. They cannot grow in pure culture and replicate only when attached to the surface of their host organism, Ignicoccus hospitalis (Section 17.11), a hyperthermophilic species of Crenarchaeota whose name means “the hospitable fireball.” N. equitans grows to 10 or more cells per Ignicoccus cell and lives an apparently parasitic lifestyle, making it the only known archaeal symbiont. Indeed, in agreement with its lifestyle, the species epithet equitans means “riding,” as in “riding the fireball.”
Figure 17.17 *Nanoarchaeum equitans*.

(a) Fluorescence micrograph of cells of N. equitans (red) attached to cells of Ignicoccus (green). Cells were stained by FISH (Section 19.5) using specific nucleic acid probes targeted to each organism. (b) Transmission electron micrograph of a thin section of a cell of N. equitans. Note the distinct cell wall. Cells of N. equitans are about 0.4 μm in diameter.
*Nanoarchaeum* and Its Host
N. equitans and its host Ignicoccus were first isolated from a submarine hydrothermal vent (Section 20.16) off the coast of Iceland. However, environmental sampling of 16S ribosomal RNA genes (Section 19.6) indicates that organisms phylogenetically similar to N. equitans exist in other submarine hydrothermal vents and in terrestrial hot springs, so Archaea of this kind are probably distributed worldwide in suitable hot habitats. Like its host Ignicoccus, N. equitans grows at temperatures from 70 to 98 °C and optimally at 90 °C.
The metabolism of Nanoarchaeum is not fully understood, but it likely depends on its host for many metabolic functions. Ignicoccus is an autotroph that uses H2 as an electron donor and S0 as an electron acceptor and probably supplies N. equitans with organic carbon. From analyses of its genome (see next subsection), N. equitans is incapable of metabolizing H2 and S0 for energy. Thus, whether it generates ATP from substances obtained from Ignicoccus or obtains its ATP directly from its host is unknown. The appearance of N. equitans cells is typical of Archaea, with a cell wall consisting of an S-layer (Section 2.5) that overlays what appears to be a periplasmic space (Figure 17.17b).
The *Nanoarchaeum equitans* Genome
The sequence of the N. equitans genome provides insight into this organism’s obligately parasitic lifestyle. Its single, circular genome is only 490,885 nucleotides long, one of the smallest cellular genomes yet sequenced (Table 10.1). Genes for several important metabolic functions are missing from the N. equitans genome, including those for the biosynthesis of amino acids, nucleotides, coenzymes, and lipids. Also missing are genes encoding proteins for widely distributed catabolic pathways, such as glycolysis. Presumably, all of these functions are carried out for N. equitans by its Ignicoccus host, with transfer of needed substances from Ignicoccus to the attached N. equitans cells. N. equitans also lacks some of the genes necessary to encode ATPase, and this indicates that it may not synthesize a functional ATPase. If true, this would be the only cellular organism that lacks ATPase. If no ATPase is present and substrate-level phosphorylation does not occur (because a glycolytic pathway is lacking), then N. equitans would be dependent on Ignicoccus for energy as well as carbon.
With so many genes missing, which genes remain in the N. equitans genome? N. equitans contains genes encoding the key enzymes for DNA replication, transcription, and translation as well as genes for DNA repair enzymes. In addition to its small size, the genome of N. equitans is also among the most gene dense of any organism known, as over 95% of the N. equitans chromosome encodes proteins—a value higher than that of most all other prokaryotic cells (Section 10.3).
The DPANN Superphylum
Metagenomic analyses of numerous habitats have identified diverse archaeal phyla based on their genome sequences (see Section 17.8). Many of these phyla affiliate phylogenetically with Nanoarchaeota in the DPANN superphylum (Figure 17.1). This large cluster of Archaea is named for the phyla Diapherotrites, Parvarchaeota, Aenigmarchaeota, Nanohaloarchaeota, and Nanoarchaeota, although recently additional phyla have been discovered to reside in this group. These Archaea escaped detection for so long because their 16S ribosomal RNA genes are so highly divergent from those of other Archaea that they were not recovered using standard metagenomic techniques for surveys of 16S rRNA genes in the environment (Section 19.8 and Figure 19.21).
Members of the DPANN superphylum, like N. equitans, appear to be extremely small cells that have highly reduced genomes. Most have very limited metabolic capabilities lacking many core biosynthetic pathways such as those for the synthesis of certain nucleotides, amino acids, and lipids. The DPANN Archaea are predicted to have obligately dependent lifestyles relying on other Archaea or Bacteria to provide them with the metabolites they need to survive. Like N. equitans, many of the DPANN Archaea are likely to be symbiotic or parasitic, living on or within other cells. Sequence analyses of complete or nearly complete DPANN genomes recovered from natural samples suggest that these organisms are obligate anaerobes that rely on fermentative forms of metabolism. DPANN Archaea have been observed in a wide range of environments including extreme environments (geothermal, acidic, and hypersaline), temperate environments (lake and marine sediments), and the deep subsurface (Section 20.8 and Figure 20.16).
Check Your Understanding
Which aspects of the biology of Nanoarchaeum equitans make it especially interesting from an evolutionary point of view?
Why can it be said that N. equitans is both a carbon and an energy parasite?
Key Genus: *Korarchaeum*
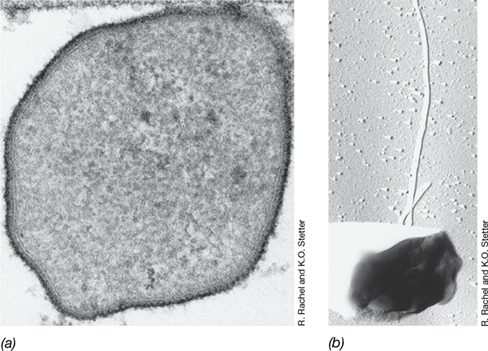
Ribosomal RNA sequences of Korarchaeota (Figure 17.1) have been retrieved from a range of geothermal habitats, both submarine and terrestrial. However, Korarchaeum cryptofilum, whose name means “the secret filament of youth,” is the only characterized species in the phylum Korarchaeota.
First observed as a 16S ribosomal RNA gene phylotype (Figures 19.16 and 19.17) recovered from Obsidian Pool, a hot spring in Yellowstone National Park, USA, K. cryptofilum has yet to be grown in pure culture. However, its genome sequence has been determined from metagenomic analyses of a highly enriched culture. K. cryptofilum is an obligately anaerobic chemoorganotroph and a hyperthermophile, growing at 85 °C. Cells are long, thin (<0.2-μm diameter) filaments of variable length (**Figure 17.18a–*c***), with most filaments being around 15 μm long but some reaching as much as 100 μm. Filaments of K. cryptofilum have a tough paracrystalline S-layer (Figure 17.18d), which maintains cell integrity in its extremely hot habitat.
Figure 17.18 *Korarchaeum cryptofilum*.

(a) Fluorescence in situ hybridization (FISH; Section 19.5) first identified the morphology of Korarchaeota growing in an enrichment culture at 85 °C. (b) Phase-contrast image of filaments of K. cryptofilum. (c) Scanning electron micrograph of a K. cryptofilum filament. (d) Transmission electron micrograph of the surface of a K. cryptofilum filament showing the paracrystalline S-layer (Section 2.5). Filaments of K. cryptofilum are about 0.17 μm wide and 15 μm long.
Though K. cryptofilum cannot be grown in isolation, its genome sequence provides clues about its lifestyle. K. cryptofilum lacks the ability to perform anaerobic respiration and lives a fermentative lifestyle. Similar to other archaeal hyperthermophiles, K. cryptofilum grows by fermentation of peptides or amino acids (see Table 17.4). K. cryptofilum lacks many core genes in biosynthesis including the ability to synthesize purines, coenzyme A, and several essential cofactors. Presumably K. cryptofilum obtains these essential components from its environment. The inability of K. cryptofilum to synthesize molecules essential for its own growth may be explained by the evolution of mutual dependence in microbial communities (Section 13.8 and Figure 13.18). This dependence on other members of the hot spring microbial community for key necessities may also explain why K. cryptofilum has not yet been obtained in pure culture.
Phylogenetic analyses indicate affinity between Thaumarchaeota, Crenarchaeota, and Korarchaeota (Figure 17.1). These lineages, along with a group called the Aigarchaeota (see next section), form a cluster of phyla known as the TACK superphylum (Figure 17.1). Like the DPANN superphylum (Section 17.6), many of the species in the TACK superphylum are only known from metagenomic analyses, but such information is often important for piecing together the nutritional secrets of uncultured microbes and obtaining their successful laboratory culture.
Check Your Understanding
What is the most likely reason that Korarchaeum cryptofilum has been difficult to isolate in pure culture?
17.8 Other Cryptic Archaeal Phyla
Advances in DNA sequencing technologies have allowed metagenomic analyses to become almost routine. As powerful DNA sequencing approaches are applied to a wider range of habitats, more and more genomes of unusual Archaea are being discovered such as those just discussed in the DPANN and TACK superphyla (Sections 17.6, 17.7). These discoveries have greatly expanded our knowledge of archaeal diversity. Thus far, our only insight into these newly discovered archaeal phyla comes from the analysis of genome data assembled from metagenomes. For most of these groups, no cultures are available and there is little direct evidence pertaining to their physiology or ecology. However, metagenomic data allows us to form many hypotheses.
Asgard *Archaea*
The Asgard Archaea is a large group of related phyla discovered and characterized through metagenomic analyses (Section 20.8 and Figure 20.16). The first of the Asgard Archaea to be discovered were those in the phylum Lokiarchaeota. Lokiarchaeota were discovered through metagenomic analyses of microbial communities that inhabit deep marine sediments near a highly sulfidic hydrothermal vent system known as Loki’s Castle (**Figure 17.19*a***), located along the Mid-Atlantic Ridge between Greenland and Norway. Remarkably, the genomes of Lokiarchaeota contain a number of eukaryotic signature genes; in particular, genes associated with membrane remodeling and the development of a cellular cytoskeleton. These results suggest that features uniquely associated with the eukaryotic cell may also be found within the domain Archaea. The presence of these eukaryotic-like genes in Lokiarchaeota also support well-established findings that the processes of transcription and translation in Archaea bear greater resemblance to those in Eukarya than to those in Bacteria (Section 6.6).
Figure 17.19 *Lokiarchaeota* and *Aigarchaeota.*

(a) Loki’s Castle, an ocean floor hydrothermal vent system at the Mid-Atlantic Ridge (depth, 2350 m). These vent chimneys are rich in iron–sulfide minerals, and when they were active, emitted water at nearly 300 °C; the white areas on the surface of the mounds contain elemental sulfur and sulfur-oxidizing bacteria (see also Figure 13.11). The first genomes of Lokiarchaeota (Asgard Archaea, Figure 17.1) were assembled from metagenomic analyses of the microbial community living near this hydrothermal vent. (b) The “pink streamer” microbial community that develops at pH 8 and 82 °C in the outflow channel of Octopus Spring, Yellowstone National Park, contains the bacterium Thermocrinis (Section 16.19) and several Archaea. The mass of pink streamers is about 10 cm in diameter. (c) In a sample of the Octopus Spring streamers subjected to fluorescence in situ hybridization (FISH) analysis (Section 19.5), a cell of Aigarchaeota (about 15 μm long) stains pink while cells of Thermocrinis stain green. Photos in (b) and (c) taken by J. Beam and W. Inskeep under research permits YELL-2012-SCI-5068 (for part b) and YELL-2013-SCI-5068 (for part c).
Since the discovery of the Lokiarchaeota, related Archaea have been found in many marine and freshwater sediments as well as in geothermal systems. These organisms have been given the phylum names Thorarchaeota, Heimdallarchaeota, and Odinarchaeota after the legendary names of Norse gods. Together, these Archaea comprise the Asgard Archaea. Like the Lokiarchaeota, the genomes of other Asgard Archaea show many features that were once thought to be uniquely eukaryal. And, since Asgard Archaea diverge deeply within the domain Archaea (Figure 17.1), it has been hypothesized that they are close relatives of Eukarya. Thus far we know relatively little about these mysterious organisms, although their genomes point to an anaerobic and fermentative lifestyle similar to some cultured Archaea and many Archaea known only from their genomes.
*Aigarchaeota* and *Bathyarchaeota*
As with the Asgard Archaea, all of our knowledge of Aigarchaeota and Bathyarchaeota comes from genomes assembled from metagenomic data. The Aigarchaeota (Figure 17.1) belong to the TACK superphylum. The first characterized genome from this group came from “Candidatus Caldiarchaeum subterraneum,” identified in a metagenome obtained from geothermally heated water found in a Japanese gold mine deep underground. Additional genomes have been identified from other geothermal systems including hot springs. For example, in the hot water runoff channel of a Yellowstone hot spring where Thermocrinis, a hyperthermophilic species of Bacteria (Section 16.19), forms pink-colored “streamers” (Figure 17.19b; see also Figure 16.52), a species of Aigarchaeota has been found that grows enmeshed in the Thermocrinis biomass. Although not yet cultured, this organism, given the name “Candidatus Calditenuis aerorheumensis” has been identified directly in natural samples using FISH probes (Section 19.5). The probes revealed an organism that is a short filament (Figure 17.19c), and metagenomic analyses suggest that it is an aerobic chemoorganotroph capable of using a variety of organic compounds as energy sources including acetate, sugars, and fatty and amino acids and that it requires a number of vitamins and cofactors.
The Bathyarchaeota (Figure 17.1) are also members of the TACK superphylum. The first genomes from this phylum were discovered in metagenomes retrieved from deep-sea sediments. Additional genomes have been obtained from a range of freshwater and shallow marine sediments as well as subsurface habitats. Several genomes of Bathyarchaeota contain genes encoding enzymes of the reductive acetyl-CoA pathway. This pathway is used by a wide variety of microbes for CO2 fixation, acetogenesis, and methanogenesis (Section 14.14 and Figure 14.34). It has been hypothesized that some Bathyarchaeota are indeed methanogens, although this remains to be proven. The genomes of Bathyarchaeota that have been characterized suggest this phylum likely contains a diversity of metabolic types, including some species capable of degrading complex polymers as well as others that might metabolize C1 compounds (methylotrophy, Section 14.16).
Check Your Understanding
What unique feature of the Asgard Archaea could provide insight on the origin of the domain Eukarya?
What technique has led to the recent discovery and characterization of many new phyla of Archaea?
III Crenarchaeota
**The phylum Crenarchaeota contains many cultured isolates, most of which are thermophiles or hyperthermophiles that use sulfur compounds as either electron donors or electron acceptors to thrive in their punishing environments.**
Among Archaea in laboratory culture, the Crenarchaeota are mostly hyperthermophiles and include species growing optimally above the boiling point of water. Many hyperthermophiles are chemolithotrophic autotrophs, and because no phototrophs can survive such temperatures, these organisms are the sole primary producers in these habitats.
17.9 Habitats and Energy Metabolism of Crenarchaeota
17.9 Habitats and Energy Metabolism of Crenarchaeota
17.9 Habitats and Energy Metabolism of *Crenarchaeota*
Most hyperthermophilic Archaea have been isolated from geothermally heated soils or waters containing S0 and H2S, and most species metabolize sulfur in one way or another. In terrestrial environments, sulfur-rich springs, boiling mud, and soils may have temperatures up to 100 °C and are mildly to extremely acidic owing to the production of sulfuric acid (H2SO4) from the biological oxidation of H2S and S0(Section 14.7 and Section 21.4). Such hot, sulfur-rich environments, called solfataras, are found throughout the world (Figure 17.20), including Italy, Iceland, New Zealand, and Yellowstone National Park in Wyoming. Depending on the surrounding geology, solfataras can be mildly acidic to slightly alkaline (pH 5–8) or extremely acidic, with pH values below 1. Hyperthermophilic crenarchaeotes have been obtained from all of these environments, but most inhabit neutral or weakly acidic thermal habitats.
Figure 17.20 Terrestrial habitats of hyperthermophilic *Archaea:* Yellowstone National Park.

(a) A typical solfatara; steam rich in H2S rises to the surface. (b) Sulfur-rich hot spring, a habitat containing dense populations of Sulfolobus. The acidity in solfataras and sulfur springs comes from the oxidation of H2S and S0 to H2SO4 (sulfuric acid) by Sulfolobus and related sulfur-oxidizing microbes. (c) A typical neutral pH boiling spring, Imperial Geyser. Many different species of hyperthermophilic Archaea may reside in such a habitat. (d) An acidic iron-rich geothermal spring, another Sulfolobus habitat; here the oxidation of Fe2+ to Fe3+ generates acidity (Section 22.2 and Figure 22.4c).
Hyperthermophilic Crenarchaeota also inhabit undersea hot springs called hydrothermal vents. We discuss the geology and microbiology of these fascinating microbial habitats in Section 20.16. Here we only note that submarine waters can be much hotter than surface waters because the water is under hydrostatic pressure. Indeed, all hyperthermophiles with growth temperature optima above 100 °C originate from submarine sources. These sources include shallow (2–10 m depth) vents such as those off the coast of Vulcano, Italy, as well as deep (2000–4000 m depth) vents near ocean-spreading centers (see Figure 17.26). Deep hydrothermal vents are the hottest habitats so far known to yield viable life forms.
Most hyperthermophilic Crenarchaeota are obligate anaerobes. Their energy-yielding metabolism is either chemoorganotrophic or chemolithotrophic (or both, for example, in Sulfolobus) and is dependent on diverse electron donors and acceptors. Fermentation is rare and most bioenergetic strategies employ anaerobic respiration (Table 17.4). Energy is conserved during these respiratory processes by the same general mechanism widespread in Bacteria: electron transfer within the cytoplasmic membrane leading to the formation of a proton motive force from which ATP is made by way of proton-translocating ATPases (Sections 3.8, 3.9 and 3.10).
Table 17.4 Energy-yielding reactions of hyperthermophilic *Archaea* by nutritional class

aAnR, anaerobic respiration; AeR, aerobic respiration; F, fermentation.
A variety of metabolisms are practiced by crenarchaeotes, and all of these were discussed in Chapter 14. For example, many hyperthermophilic crenarchaeotes grow chemolithotrophically under anoxic conditions with H2 as the electron donor and S0 or NO3 − as the electron acceptor; a few can also oxidize H2 aerobically (Table 17.4). H2 respiration with ferric iron (Fe3+) as electron acceptor also occurs in several hyperthermophiles. Other chemolithotrophic lifestyles include the oxidation of S0 and Fe2+ aerobically or Fe2+ anaerobically with NO3 − as the acceptor, and a variety of organic compounds are oxidized by one or another species of crenarchaeote (Table 17.4). Only one sulfate-reducing hyperthermophile is known (the euryarchaeote Archaeoglobus, Section 17.4). The only bioenergetic option apparently absent from hyperthermophilic crenarchaeotes is photosynthesis, a means of energy conservation that is apparently limited to temperatures below 74 °C (see Figure 17.30). In the next two sections we will meet some of these fascinating microbes, microbes that have contributed significantly to our understanding of the physiochemical limits to life.
Check Your Understanding
What form of energy metabolism is widespread among hyperthermophiles?
How might the temperature and pH tolerance of a hyperthermophile living in a solfatara differ from that of a hyperthermophile living in a hydrothermal vent?
17.10 Crenarchaeota from Terrestrial Volcanic Habitats
17.10 Crenarchaeota from Terrestrial Volcanic Habitats
Key Genera: *Sulfolobus, Acidianus, Thermoproteus, Pyrobaculum*
Terrestrial volcanic habitats can have temperatures as high as 100 °C and are thus suitable for hyperthermophilic Archaea. Two phylogenetically related organisms isolated from these environments are Sulfolobus and Acidianus. These genera form the heart of an archaeal order called the Sulfolobales (Table 17.5). In addition, Sulfolobus has been a model organism for molecular biology studies of Archaea.
Table 17.5 Properties of some well-characterized hyperthermophilic *Crenarchaeota*

bAlso known by the unofficial taxonomic name of “Geogemma barossii.”
Sulfolobales
Sulfolobus grows in sulfur-rich acidic thermal areas (Figure 17.20) at temperatures up to 90 °C and at pH values of 1–5. Species of Sulfolobus are typically aerobic chemoorganotrophs, though some can also grow as chemolithotrophs by oxidizing H2S or S0 to H2SO4 and fixing CO2. Cells of Sulfolobus are more or less spherical but contain distinct lobes (**Figure 17.21*a***). Cells adhere tightly to sulfur crystals, where they can be seen with a microscope after staining with fluorescent dyes (Figure 14.19b). Besides the aerobic respiration of sulfur or organic compounds, some Sulfolobus can also oxidize Fe2+ to Fe3+, and this ability has been harnessed for the high-temperature leaching of iron and copper ores (Sections 21.6 and 22.1 and 22.2).
Figure 17.21 Acidophilic hyperthermophilic *Archaea*, the *Sulfolobales*.

(a) Sulfolobus acidocaldarius; electron micrograph of a thin section. (b) Acidianus infernus; electron micrograph of a thin section. Cells of both organisms vary from 0.8 to 2 μm in diameter. Sulfolobales typically show temperature optima below 90 °C (Table 17.5).
A facultative aerobe resembling Sulfolobus also lives in acidic solfataric springs. This organism, Acidianus (Figure 17.21b), differs from Sulfolobus most clearly by its ability to grow using S0 both anaerobically as well as aerobically. Under aerobic conditions the organism uses S0 as an electron donor, oxidizing S0 to H2SO4, with O2 as an electron acceptor. Under anaerobic conditions, by contrast, Acidianus uses S0 as an electron acceptor with H2 as the electron donor, forming H2S as the reduced product. Thus, the metabolic fate of S0 in cultures of Acidianus depends on the presence or absence of O2. Like Sulfolobus, Acidianus is roughly spherical in shape but is not as lobed (Figure 17.21b). It grows at temperatures from 65 °C up to a maximum of 95 °C. As a group, then, the Sulfolobales contain the most thermophilic of all highly acidophilic Archaea.
Thermoproteales
Key genera within the Thermoproteales are Thermoproteus, Thermofilum, and Pyrobaculum. The genera Thermoproteus and Thermofilum consist of rod-shaped cells that inhabit neutral or slightly acidic hot springs. Cells of Thermoproteus are rigid rods about 0.5 μm in diameter and highly variable in length, ranging from short cells of 1–2 μm (**Figure 17.22*a***) up to filaments 70–80 μm long. Filaments of Thermofilum are thinner, some as little as 0.17–0.35 μm wide, with filament lengths ranging up to 100 μm (Figure 17.22b).
Figure 17.22 Rod-shaped hyperthermophilic *Archaea*, the *Thermoproteales*.

(a) Thermoproteus neutrophilus; electron micrograph of a thin section. A cell is about 0.5 μm in diameter. (b) Thermofilum librum. A cell is about 0.25 μm in diameter. (c) Pyrobaculum aerophilum. Transmission electron micrograph of a thin section; a cell measures 0.5×3.5 μm. Although the temperature optimum of P. aerophilum is 100 °C, optima for other Thermoproteales are all below 90 °C (Table 17.5).
Both Thermoproteus and Thermofilum are strict anaerobes that carry out an S0-based anaerobic respiration (Table 17.4). Most Thermoproteus isolates can grow chemolithotrophically on H2 or chemoorganotrophically on complex carbon substrates such as yeast extract, small peptides, starch, glucose, ethanol, malate, fumarate, or formate (Table 17.4). Pyrobaculum (Figure 17.22c) is a rod-shaped hyperthermophile but is physiologically distinct from other Thermoproteales in that some species of Pyrobaculum can respire aerobically. However, Pyrobaculum can also grow by anaerobic respiration with NO3 −, Fe3+, or S0 as electron acceptors and H2 as an electron donor (that is, they can grow chemolithotrophically and autotrophically). Other species of Pyrobaculum can grow anaerobically on organic electron donors, reducing S0 to H2S. The growth temperature optimum of Pyrobaculum is 100 °C, and species of this organism have been isolated from both terrestrial hot springs and hydrothermal vents.
Check Your Understanding
What are the similarities and differences between Sulfolobus and Pyrobaculum?
Among Thermoproteales, what is unusual about the metabolism of Pyrobaculum?
17.11 Crenarchaeota from Submarine Volcanic Habitats
17.11 Crenarchaeota from Submarine Volcanic Habitats
Key Genera: *Pyrodictium, Pyrolobus, Ignicoccus, Staphylothermus*
We now consider the microbiology of submarine volcanic habitats, homes to the most thermophilic of all known Archaea. These habitats include both shallow-water thermal springs and deep-sea hydrothermal vents. We discuss the geology of these fascinating microbial habitats in Section 20.16 and the interesting animal communities that develop there in Section 23.11. The organisms to be described here constitute an order of Archaea called the Desulfurococcales (Table 17.5).
Figure 17.23 *Desulfurococcales* with growth temperature optima >100°C.

(a) Pyrodictium occultum (growth temperature optimum, 105 °C), dark-field micrograph. (b) Thin-section electron micrograph of P. occultum. Cells are highly variable in diameter from 0.3 to 2.5 μm. (c) Thin section of a cell of Pyrolobus fumarii, one of the most thermophilic organisms ever described (growth temperature optimum, 106 °C); a cell is about 1.4 μm in diameter. (d) Negative stain of a cell of “Strain 121,” capable of growth at 121 °C; a cell is about 1 μm wide. Although the Desulfurococcales contain the greatest number of hyperthermophiles capable of growth above 100 °C, the most thermophilic of all known Archaea is actually a euryarchaeote, Methanopyrus (Section 17.2).
Pyrolobus fumarii (Figure 17.23c) is one of the most thermophilic of the hyperthermophiles. Its growth temperature maximum is 113 °C (Table 17.5). P. fumarii lives in the walls of “black smoker” hydrothermal vent chimneys (Section 20.16 and Figures 20.43, 20.44, and 20.46) where its autotrophic abilities contribute organic carbon to this otherwise inorganic environment. P. fumarii cells are coccoid-shaped (Figure 17.23c), and the cell wall is composed of protein. The organism is an obligate H2 chemolithotroph, growing by the oxidation of H2 coupled to the reduction of NO3 − to ammonium (NH4 +), thiosulfate (S2O3 2−) to H2S, or very low concentrations of O2 to H2O. Besides its extremely thermophilic nature, P. fumarii can withstand temperatures substantially above its growth temperature maximum. For example, cultures of P. fumarii survive autoclaving (121 °C, Section 4.17) for 1 h, a condition that even bacterial endospores (Section 2.8) cannot withstand.
Another organism in this group shares with Pyrolobus a growth temperature optimum of 106 °C. However, “Strain 121,” as this organism has been called, actually shows weak growth at 121 °C, and cells remain viable for 2 h at 130 °C. Only Methanopyrus, a hyperthermophilic methanogen, can grow at a higher temperature (122 °C, Section 17.2). Strain 121 consists of coccoid cells that are motile by rotating archaella (Figure 17.23d); the organism is also a strict anaerobe and grows chemolithotrophically and autotrophically with Fe3+ as electron acceptor and formate or H2 as electron donors. Thus, it is clear that the Pyrodictium/Pyrolobus group collectively contains many of the most hyperthermophilic examples of all known microbes.
*Desulfurococcus* and *Ignicoccus*
Other notable members of the Desulfurococcales include Desulfurococcus, the genus for which the order is named (**Figure 17.24*a***), and Ignicoccus. Desulfurococcus is a strictly anaerobic S0-reducing organism like Pyrodictium, but differs from this organism in its phylogeny and the fact that it is much less thermophilic, growing optimally at about 85 °C.
Figure 17.24 *Desulfurococcales* with growth temperature optima < 100°C.

(a) Thin section of a cell of Desulfurococcus saccharovorans; a cell is 0.7 μm in diameter. (b) Thin section of a cell of Ignicoccus islandicus; the cell proper is surrounded by an extremely large periplasmic compartment. The cell itself measures about 1 μm in diameter and the cell plus periplasm measures 1.4 μm. Ignicoccus is a host for the tiny parasitic microbe Nanoarchaeum (see Figure 17.17).
Ignicoccus species grow optimally at about 90 °C, and their energy metabolism is based on H2 as an electron donor and S0 as an electron acceptor, as is that of so many hyperthermophilic Archaea (Table 17.4). Some Ignicoccus species are hosts to the small parasitic archaeon Nanoarchaeum equitans (Section 17.6). Ignicoccus (Figure 17.24b) has a novel cell structure that lacks an S-layer and possesses a unique outer cellular membrane. This outer cellular membrane is distinct in several ways from the outer membrane of gram-negative Bacteria (Section 2.4). Most notably, the outer cellular membrane of Ignicoccus contains ATPase and is the site of energy conservation. Ignicoccus also contains an inner cellular membrane that contains the cytoplasm and the enzymes responsible for biosynthesis and information processing. In this way neither the outer nor inner cellular membrane satisfies the typical definition of a cytoplasmic membrane (Section 2.1).
Between the inner and outer cellular membranes of Ignicococcus is a large intermediate compartment that is analogous to the periplasm of gram-negative Bacteria but is much larger in volume, representing some two to three times the volume of the cytoplasm (Figure 17.24b). The periplasm of Ignicoccus also contains membrane-bound vesicles (Figure 17.24b) that may function in exporting substances outside the cell. In this way, the cell structure of Ignicoccus resembles that of Eukarya. Because of this, Ignicoccus has been proposed to be a modern descendant of the ancestral cell type that gave rise to the origin of eukaryotic cells, a topic we covered earlier (Section 13.4 and Figure 13.10).
Staphylothermus
A morphologically unusual member of the order Desulfurococcales is the genus Staphylothermus (Figure 17.25). Cells of Staphylothermus are spherical, about 1 μm in diameter, and form aggregates of up to 100 cells, much like its morphological but mesophilic counterpart among the Bacteria, Staphylococcus (Figure 16.20 and Figure 31.28). Unlike many hyperthermophiles, Staphylothermus is not a chemolithotroph but instead a chemoorganotroph, with species that grow optimally at 92 °C. Organic carbon sources are used as electron donors in sulfur reduction, producing H2S, acetate, and isovalerate as products (Table 17.4).
Figure 17.25 The hyperthermophile *Staphylothermus marinus*.

Electron micrograph of shadowed cells. A single cell is about 1 μm in diameter.
Isolates of Staphylothermus have been obtained from both shallow marine hydrothermal vents and very hot black smokers (see Figure 17.26; Section 20.16). This organism is apparently widely distributed in submarine thermal areas, where it is likely to play a significant role in consuming proteins released from dead organisms.
Check Your Understanding
What archaeal genera contain species having growth optima at temperatures greater than 100 °C?
What unusual structural features are present in Ignicoccus and Staphylothermus?
IV Evolution and Life at High Temperature
**The ancestor of all Archaea was likely a hyperthermophile, and virtually all such organisms are Archaea. Hyperthermophilic Archaea have several biochemical adaptations that make life possible at high temperatures, and the widespread use of H2 in their metabolisms may reflect early Earth conditions.**
Most of the hyperthermophiles discovered so far are species of Archaea and some grow near to what may be the upper temperature limit for life. Here we consider the major factors that likely define the upper temperature limit for life and the biological adaptations of hyperthermophiles that permit them to exist at the exceptionally high temperatures of 100 °C and higher. We end with a discussion of the importance of hydrogen (H2) metabolism to the biology of hyperthermophiles.
17.12 An Upper Temperature Limit for Microbial Life
17.12 An Upper Temperature Limit for Microbial Life
17.12 An Upper Temperature Limit for Microbial Life
Habitats that contain liquid water—a prerequisite for cellular life—and that have temperatures higher than 100 °C are only found where geothermally heated water flows out of vents or rifts in the ocean floor (Figure 13.3 and Figures 20.43 and 20.44). The hydrostatic pressure that overlies the water keeps it from boiling, allowing it to reach temperatures of up to about 400 °C in vents at several thousand meters’ depth. In contrast, terrestrial hot springs can boil and therefore only attain temperatures near 100 °C. It is not surprising, then, that hydrothermal vents have been rich sources of hyperthermophilic Archaea with growth temperature optima above 100 °C (Table 17.5).
Black smokers emit hydrothermal vent fluid at 250–350 °C or higher. Metallic mounds or more upright structures called chimneys form from the metal sulfides that precipitate out of the hot fluid as it mixes with the surrounding, much cooler seawater (Figure 17.19a and Figure 17.26). As far as is known, the superheated vent water itself is sterile. However, hyperthermophiles thrive in mounds or smoker chimney walls where temperatures are compatible with their survival and growth (Figures 20.45 and 20.46). By studying structures such as these, we can address the question, “What is the upper temperature for microbial (and presumably all forms of) life?”
Figure 17.26 Hydrothermal vents.

Hydrothermal mound from the Rainbow vent field, Mid-Atlantic Ridge hydrothermal system. The hydrothermal fluid emitting from the two short chimneys is >300 °C and rich in sulfidic minerals.
Anna-Louise Reysenbach and Woods Hole Oceanographic Institution
What Is the Upper Temperature Limit for Life?
How high a temperature can hyperthermophiles withstand? Over the past several decades, the known upper temperature limit for life has been pushed higher and higher with the isolation and characterization of new species of thermophiles and hyperthermophiles (Figure 17.27). For some time the record holder was Pyrolobus fumarii (Figure 17.23c), with its upper temperature limit for growth of 113 °C. The current record holder, Methanopyrus (Section 17.2 and Figure 17.8), however, has pushed the limit somewhat higher, with the ability to grow at 122 °C and to survive substantial periods at even higher temperatures. Given the trend over the past several years (Figure 17.27), one can predict that Archaea even more hyperthermophilic than Methanopyrus may inhabit hydrothermal environments but have yet to be isolated. Indeed, many experts predict that the upper temperature limit for Archaea is likely to exceed 140 °C, perhaps even 150 °C, and that the maximum temperature allowing survival but not growth is hotter yet.
Figure 17.27 Thermophilic and hyperthermophilic *Bacteria* and *Archaea*.

The graph gives the species that were, in turn, the record holders for growing at the highest temperature, from before 1960 to the present.
Mastering Microbiology
Art Activity: Figure 17.27 Thermophilic and hyperthermophilic Bacteria and Archaea
Biochemical Problems at Supercritical Temperatures
Whatever the upper temperature limit is for life, it is likely to be defined by one or more biochemical challenges that evolution has been unable to solve. There is obviously an upper limit, but we do not yet know what it is. Water samples taken directly from superheated (>250 °C) hydrothermal vent discharges are devoid of measurable biochemical markers (DNA, RNA, and protein) that would signal life as we know it, while vents emitting water at temperatures below about 150 °C yield evidence of macromolecules. These results are consistent with laboratory experiments on the stability of key biomolecules. For example, ATP is degraded almost instantly at 150 °C. Thus, above 150 °C, any life forms would have to deal with the heat lability of a molecule that is, as far as is known, a universal energy currency in all cells.
This conclusion has a caveat, however, in that it is possible that the stability of small molecules such as ATP may be significantly greater under cytoplasmic conditions of high levels of dissolved solutes than in pure solutions tested in the laboratory. Nevertheless, if life forms exist at temperatures above 150 °C, they must be unique in at least one of two ways—they must either produce novel biomolecules absent from cells as we know them or deploy protective systems that maintain known cellular molecules in a sufficiently stable state to support biochemistry. Only time and more microbiological research and exploration for new organisms growing at ever higher temperatures will reveal which of these possibilities (or perhaps some of both) is correct.
Check Your Understanding
Where would you expect to find hyperthermophiles that grow at temperatures above 100 °C? Why would it be impossible for organisms to grow at 200 or 300 °C?
17.13 Molecular Adaptations to Life at High Temperature
17.13 Molecular Adaptations to Life at High Temperature
17.13 Molecular Adaptations to Life at High Temperature
Because all cellular structures and activities are affected by heat, hyperthermophiles are likely to exhibit multiple adaptations to the exceptionally high temperatures of their habitats. Here we briefly examine some adaptations employed by hyperthermophiles to protect their proteins and nucleic acids at high temperatures.
Protein Folding and Thermostability
Because most proteins denature at high temperatures, much research has been done to identify the properties of thermostable proteins. Protein thermostability derives from the folding of the molecule itself (Figure 6.29), not because of the presence of any special amino acids. Perhaps surprisingly, however, the amino acid composition of thermostable proteins is not particularly unusual except perhaps in their slight bias for increased levels of amino acids that promote alpha-helical secondary structures. In fact, many enzymes from hyperthermophiles contain the same major structural features in both primary and higher-order structure (Section 6.7) as their heat-labile counterparts from organisms that grow best at much lower temperatures.
Thermostable proteins typically do display some structural features that likely improve their thermostability. These include having highly hydrophobic cores, which decrease the tendency of the protein to unfold in an ionic environment, and more ionic interactions on the protein surfaces, which also help hold the protein together and work against unfolding. Ultimately, it is the folding of the protein that most affects its heat stability, and noncovalent ionic bonds called salt bridges on a protein’s surface likely play a major role in maintaining the biologically active structure. But, as previously stated, many of these changes are possible with only minimal changes in primary structure (amino acid sequence), as seen when thermostable and heat-labile forms of the same protein are compared.
Chaperones: Assisting Proteins to Remain in Their Native State
Earlier we discussed a class of proteins called chaperones (heat shock proteins; Section 6.11) that function to refold partially denatured proteins. Hyperthermophilic Archaea have special classes of chaperones that function only at the highest growth temperatures. In cells of Pyrodictium abyssi (Figure 17.28), for example, a major chaperone is the protein complex called the thermosome. This complex keeps other proteins properly folded and functional at high temperature, helping cells survive even at temperatures above their maximal growth temperature. Cells of P. abyssi grown near its maximum temperature (110 °C) contain high levels of the thermosome. Possibly because of this, the cells can remain viable following a heat shock, such as a 1-h treatment in an autoclave (121 °C). In cells experiencing such a treatment and then returned to the optimum temperature, the thermosome, which is itself quite heat resistant, is thought to refold sufficient copies of key denatured proteins that P. abyssi can once again begin to grow and divide. Thus, as a result of chaperone activity, the upper temperature limit at which many hyperthermophiles can survive is higher than the upper temperature at which they can grow. The “safety net” of chaperone activity probably ensures that cells in nature that briefly experience temperatures above their growth temperature maximum are not killed by the exposure.
Figure 17.28 *Pyrodictium abyssi*, scanning electron micrograph.

Pyrodictium has been studied as a model of macromolecular stability at high temperatures. Cells are enmeshed in a sticky glycoprotein matrix that binds them together.
DNA Stability: Solutes, Reverse Gyrase, and DNA-Binding Proteins
What keeps DNA from melting at high temperatures? Various mechanisms are known to contribute. One such mechanism increases cellular solute levels, in particular potassium (K+) or compatible organic compounds. For example, the cytoplasm of the hyperthermophilic methanogen Methanopyrus (Section 17.2) contains greater than 1 M potassium cyclic 2,3-diphosphoglycerate. This solute prevents chemical damage to DNA, such as depurination or depyrimidization (loss of a nucleotide base through hydrolysis of the glycosidic bond) from high temperatures, events that can lead to mutation (Section 9.2). This compound and other compatible solutes such as potassium di-myo-inositol phosphate, which protects against osmotic stress, and the polyamines putrescine and spermidine, which stabilize both ribosomes and nucleic acids at high temperature, help maintain key cellular macromolecules in hyperthermophiles in their active forms.
A unique protein found only in hyperthermophiles is responsible for DNA stability in these organisms. All hyperthermophiles produce a special DNA topoisomerase called reverse DNA gyrase. This enzyme introduces positive supercoils into the DNA of hyperthermophiles (in contrast to the negative supercoils introduced by DNA gyrase present in Bacteria and most Archaea; Section 6.1). Positive supercoiling stabilizes DNA to heat and thereby prevents the DNA helix from spontaneously unwinding. The noticeable absence of reverse DNA gyrase in organisms whose growth temperature optima lie below 80 °C strongly suggests a specific role for this enzyme in DNA stability at high temperatures.
Most Archaea also contain highly basic (positively charged) DNA-binding proteins that are remarkably similar in amino acid sequence and folding properties to the core histones of the Eukarya (Figure 2.43b). Archaeal histones from the hyperthermophilic methanogen Methanothermus fervidus (Figure 17.7c) have been particularly well studied. These proteins wind and compact DNA into nucleosome-like structures (Figure 17.29) and maintain the DNA in a double-stranded form at very high temperatures. Archaeal histones are found in most Euryarchaeota, including extremely halophilic Archaea, such as Halobacterium. However, because the extreme halophiles are not thermophiles, archaeal histones may have other functions besides DNA stability, in particular in assisting in gene expression by opening the helix to allow transcriptional proteins to bind.
Figure 17.29 Archaeal histones.

Electron micrograph of linearized plasmid DNA wrapped around copies of archaeal histone Hmf (from the hyperthermophilic methanogen Methanothermus fervidus) to form the roughly spherical, darkly stained nucleosome structures (arrows).Compare this micrograph with an artist’s depiction of the histones and nucleosomes of Eukarya shown in Figure 2.43b.
Lipid and Ribosomal RNA Stability
How have the lipids and the protein-synthesizing machinery of hyperthermophiles adjusted to high temperatures? Virtually all hyperthermophilic Archaea synthesize lipids of the biphytanyl tetraether type (Section 2.1). These lipids are naturally heat resistant because the phytanyl units forming each half of the membrane structure are covalently bonded to one another; this yields a lipid monolayer membrane instead of the normal lipid bilayer (Figure 2.3). This structure resists the tendency of heat to pull apart a lipid bilayer constructed of fatty acid or phytanyl side chains that are not covalently bonded.
A final point on molecular adaptations to life at high temperatures is the base composition of ribosomal RNAs. Ribosomal RNAs are key structural and functional components of the ribosome, the cell’s protein-synthesizing apparatus (Section 6.10). Hyperthermophilic species of both Bacteria and Archaea show as much as a 15% greater proportion of G+C base pairs in their small ribosomal subunit RNAs compared with organisms that grow at lower temperatures. G+C base pairs form three hydrogen bonds compared to the two of AU base pairs (Figure 6.1c), and thus the higher G+C content of the ribosomal RNAs should confer greater thermal stability on the ribosomes of these organisms and this should assist protein synthesis at high temperatures. By contrast to ribosomal RNAs, the G+C content of genomic DNA of hyperthermophiles is often rather low, which suggests that the thermal stability of ribosomal RNA might be an especially significant factor for life under hyperthermophilic conditions.
Check Your Understanding
How do hyperthermophiles keep proteins and DNA from being destroyed by high heat?
How are the lipids and ribosomes of hyperthermophiles protected from heat denaturation?
17.14 Hyperthermophilic Archaea, H2, and Microbial Evolution
17.14 Hyperthermophilic *Archaea*, H2, and Microbial Evolution
When cellular life first arose on Earth nearly 4 billion years ago (Figures 1.10a and 13.1), temperatures almost certainly were far hotter than they are today. Thus, for hundreds of millions of years, Earth may have been suitable only for hyperthermophiles. Given the discussion above on the temperature limits to life, it has been hypothesized that biological molecules, biochemical processes, and the first cells arose on Earth around hydrothermal springs and vents on the seafloor as they cooled to temperatures compatible with biological molecules (Section 13.1 and Figures 13.3 and 13.4). The phylogeny of modern hyperthermophiles (Figure 17.1), as well as the similarities in their habitats and metabolism to those of early cells on Earth, suggests that hyperthermophiles may be the closest remaining descendants of ancient cells and are a living window into the biology of ancient microbial life.
Hyperthermophilic Habitats and H2 as an Energy Source
The oxidation of H2 linked to the reduction of Fe3+, S0, NO3 −, or, rarely, O2 is a widespread mechanism of energy conservation in hyperthermophiles (Table 17.4 and Figure 17.30). This, coupled with the likelihood that these hyperthermophiles best characterize early Earth phenotypes, points to the important role H2 has played in the evolution of microbial life. Hydrogen metabolism may have evolved in primitive organisms because of the ready availability of H2 and suitable inorganic electron acceptors in their primordial environments, but also because known examples of H2-based energy conservation require relatively few proteins (Figure 13.5). As chemolithotrophs, these organisms may have obtained all of their carbon from CO2 or might have assimilated available organic compounds directly for biosynthetic needs. Either way, it is likely that the oxidation of H2 was the energetic driving force for maintaining life processes.
Figure 17.30 Upper temperature limits for energy metabolism.

The record holder for phototrophy is Synechococcus lividus (Bacteria, cyanobacteria); for chemoorganotrophy, Pyrodictium occultum (Archaea); for chemolithotrophy with S0 as electron donor, Acidianus infernus (Archaea); for chemolithotrophy with Fe2+ as electron donor, Ferroglobus placidus (Archaea); and for chemolithotrophy with H2 as electron donor, Methanopyrus kandleri (Archaea, 122 °C).
Mastering Microbiology
Art Activity: Figure 17.30 Upper temperature limits for energy metabolism
If one compares microbial energy conservation mechanisms as a function of temperature from data of cultured Bacteria and Archaea, only chemolithotrophic organisms are known at the hottest temperatures (Figure 17.30). Chemoorganotrophy occurs up to at least 110 °C, as this is the upper temperature limit for growth of Pyrodictium occultum, an organism that can conserve energy and grow by fermentation and by chemolithotrophic growth on H2 with S0 as electron acceptor (Table 17.4). Photosynthesis is apparently the least heat-tolerant of all bioenergetic processes, with no hyperthermophilic representatives known and an upper temperature limit of 73 °C. This is consistent with the conclusion that anoxygenic photosynthesis first appeared on Earth some hundreds of millions of years after the first life forms are thought to have appeared (Figure 13.1).
Comparisons of bioenergetic options as a function of temperature (Figure 17.30) point to the H2-oxidizing hyperthermophilic Archaea and Bacteria as the most likely extant examples of Earth’s earliest cellular life forms. More so than any other microbes, these organisms retain the metabolic and physiological traits one would predict to be necessary for existence on a hot early Earth.
Check Your Understanding
What phylogenetic and physiological evidence suggests that today’s hyperthermophiles are the closest living links to Earth’s earliest cells?
Which mechanism of energy conservation is least heat tolerant?
Which chemolithotrophic lifestyle seems best suited to life at the highest temperatures?
Chapter Review
Go to Mastering Microbiology for videos, animations, practice tests, and more.
I *Euryarchaeota*
17.1 Extremely halophilic Archaea require large amounts of NaCl for growth and accumulate large levels of K+ in their cytoplasm as a compatible solute. These salts affect cell wall stability and enzyme activity. The light-mediated proton pump bacteriorhodopsin helps extreme halophiles make ATP.
**Q Contrast the roles of bacteriorhodopsin, halorhodopsin, and sensory rhodopsin in *Halobacterium salinarum.***
17.2 Methanogenic Archaea are strict anaerobes whose metabolism is linked to the production of CH4. Methane can be produced by CO2 reduction by H2, from methyl substrates such as CH3OH, or from acetate.
**Q What are two unique features of the methanogen Methanopyrus?**
17.3 Thermoplasma, Ferroplasma, and Picrophilus are extremely acidophilic thermophiles that form their own phylogenetic family of Archaea. Cells of Thermoplasma and Ferroplasma lack cell walls, resembling the mycoplasmas in this regard.
**Q What two major physiological features unify species of Thermoplasmatales? Why does this allow some of them to successfully colonize coal refuse piles?**
17.4 Archaeoglobus and Ferroglobus are related anaerobic Archaea that carry out different anaerobic respirations. Archaeoglobus is a sulfate reducer and Ferroglobus is a nitrate reducer that oxidizes ferrous iron.
**Q What metabolic property is shared by the archaean Archaeoglobus and the bacterium Desulfovibrio?**
II *Thaumarchaeota* and Cryptic Archaeal Phyla
17.5 Thaumarchaeota are widespread and abundant in soils and marine environments. All cultivated species of thaumarchaea are autotrophic ammonia-oxidizers and these organisms are important in the global nitrogen cycle.
**Q What is physiologically unusual about the thaumarchaeotal species Nitrosopumilus maritimus?**
17.6 Nanoarchaeum equitans is a hyperthermophile that forms its own phylum, the Nanoarchaeota, and is a parasite of the crenarchaeote Ignicoccus. N. equitans has a tiny, highly compact genome and depends on Ignicoccus for most of its cellular needs, including both carbon and energy.
**Q What features of Nanoarchaeota and other members of the DPANN superphylum distinguish them from other Archaea?**
17.7 Korarchaeum cryptofilum forms its own phylum, the Korarchaeota, and is a hyperthermophile that lacks important biosynthetic pathways, obtaining key building blocks from its environment.
**Q What is a likely explanation for the fact that organisms like Korarchaeum cryptofilum have proven so difficult to cultivate in pure culture?**
17.8 Lokiarchaeota and other Asgard Archaea have eukaryotic signature genes that were once thought to be unique to the domain Eukarya. Bathyarchaeota and Aigarchaeota are members of the TACK superphylum known only from their genome sequences obtained from metagenomic studies.
**Q What pathway, observed in the genomes of certain Bathyarchaeota, has led to the hypothesis that these organisms are capable of acetogenesis or possibly methanogenesis?**
III *Crenarchaeota*
17.9 A wide variety of chemoorganotrophic and chemolithotrophic energy metabolisms have been found in hyperthermophilic Crenarchaeota, including fermentation and anaerobic respirations. Strictly autotrophic lifestyles are common, but photosynthesis is absent.
**Q What forms of energy metabolism are present in Crenarchaeota? What form is not present?**
17.10 Hyperthermophilic Crenarchaeota thrive in terrestrial hot springs of various chemistries. These include in particular organisms such as Sulfolobus, Acidianus, Thermoproteus, and Pyrobaculum.
**Q What is unusual about the metabolism of S0 by Acidianus?**
17.11 In deep-sea hydrothermal systems, Crenarchaeota such as Pyrolobus, Pyrodictium, Ignicoccus, and Staphylothermus thrive. With the exception of the methanogen Methanopyrus (Euryarchaeota), species of these genera grow at the highest temperatures of all Archaea, in many cases well above the boiling point of water.
**Q Why is it likely that organisms such as Pyrolobus fumarii are limited to hydrothermal habitats rather than terrestrial hot springs?**
IV Evolution and Life at High Temperature
17.12 Life as we know it is probably limited to temperatures below 150 °C. Key small molecules, such as ATP, are quickly destroyed above this temperature.
Q What organism is the current record holder for the upper temperature limit for growth? What is predicted to be the absolute upper temperature limit for growth, and why?
17.13 Macromolecules in hyperthermophiles are protected from heat denaturation by their heat-stable folding patterns (proteins), solutes and binding proteins (DNA), unique monolayer membrane architecture (lipids), and the high GC content of their ribosomal RNAs.
Q What is reverse DNA gyrase and why is it important to hyperthermophiles?
17.14 Hydrogen metabolism is likely to have been the driving force behind the energetics of the earliest cells on Earth. Chemolithotrophic metabolisms based on H2 as an elec‑tron donor are found in the most heat-tolerant of all known Bacteria and Archaea.
Q Why might H2 metabolism have evolved as a mechanism for energy conservation in the earliest organisms on Earth?
Application Questions
Using the phylogenetic tree in Figure 17.1 as a guide, discuss what indicates that bacteriorhodopsin may have been a late evolutionary invention and that anaerobic respiration with S0 as electron acceptor might have been an early evolutionary invention.
Use Figure 17.1 to make a hypothesis about the distribution of methanogenesis in the domain Archaea. When do you think this trait evolved? What evolutionary processes have resulted in the distribution of this trait in the archaeal family tree?
Chapter Glossary
a large collection of related archaeal phyla including the Lokiarchaeota, members of which are notable for containing many eukaryotic signature genes Bacteriorhodopsin
a protein containing retinal that is found in the membranes of certain extremely halophilic Archaea and that is involved in light-mediated ATP synthesis Compatible solute
a molecule that is accumulated in the cytoplasm of a cell for adjustment of water activity but that does not inhibit biochemical processes Crenarchaeota
a large phylum of Archaea that contains hyperthermophilic organisms DPANN superphylum
a large collection of related archaeal phyla characterized by extremely small cells and reduced genomes, Nanoarchaeum equitans is the best-characterized species Euryarchaeota
a large phylum of Archaea that contains primarily methanogens, extreme halophiles, Thermoplasma, and some marine hyperthermophiles Extreme halophile
a microorganism that requires very large amounts of NaCl for growth, usually greater than 10% and in some cases near saturation, for growth Extremophiles
microorganisms that inhabit environments characterized by extremes of temperature, pH, pressure, or salinity Halorhodopsin
a light-driven chloride pump that accumulates Cl− within the cytoplasm Hydrothermal vent
a deep-sea hot spring emitting warm (∼20 °C) to superheated (>300 °C) water Hyperthermophile
a species of Bacteria or Archaea whose growth temperature optimum is 80 °C or greater Korarchaeota
a phylum of Archaea that contains the hyperthermophile Korarchaeum cryptophilum Methanogen
a methane-producing species of the Archaea Nanoarchaeota
a phylum of Archaea that contains the hyperthermophilic parasite Nanoarchaeum equitans Phytanyl
a branched-chain hydrocarbon containing 20 carbon atoms and commonly found in the lipids of Archaea Reverse DNA gyrase
a protein universally present in hyperthermophiles that introduces positive supercoils into circular DNA Solfatara
a hot, sulfur-rich, generally acidic environment commonly inhabited by hyperthermophilic Archaea TACK superphylum
a large collection of related archaeal phyla including the eponymous Thaumarchaeota, Aigarchaeota, Crenarchaeota, and Korarchaeota Thaumarchaeota
a phylum of Archaea that contains widespread species capable of aerobic ammonia oxidization Thermosome
a heat shock (chaperone) protein complex that functions to refold partially heat-denatured proteins in hyperthermophiles