The Chemical Senses
BOX 8.1 OF SPECIAL INTEREST: Strange Tastes: Fat, Starch, Carbonation, Calcium, Water?
BOX 8.2 OF SPECIAL INTEREST: Memories of a Very Bad Meal
BOX 8.4 PATH OF DISCOVERY: Channels of Vision and Smell, by Geoffrey Gold
Spatial and Temporal Representations of Olfactory Information
Life evolved in a sea of chemicals. From the beginning, organisms have floated or swum in water containing chemical substances that signal food, poison, or sex. In this respect, things have not changed much in three billion years. Animals, including humans, depend on the chemical senses to help identify nourishment (the sweetness of honey, the aroma of pizza), noxious substances (the bitterness of plant poisons), or the suitability of a potential mate. Of all the sensory systems, chemical sensation is the oldest and most pervasive across species. Even brainless bacteria can detect, and tumble toward, a favorable food source.
Multicellular organisms must detect chemicals in both their internal and their external environments. The variety of chemical detection systems has expanded considerably over the course of evolution. Humans live in a sea of air, full of volatile chemicals; we put chemicals into our mouth for a variety of reasons; and we carry a complex sea within us in the form of blood and the other fluids that bathe our cells. We have specialized detection systems for the chemicals in each environment. The mechanisms of chemical sensation that originally evolved to detect environmental substances now serve as the basis for chemical communication between cells and organs, using hormones and neurotransmitters. Every cell in every organism is responsive to many chemicals.
This chapter considers the most familiar of our chemical senses: taste, or gustation, and smell, or olfaction. Although taste and smell reach our awareness most often, they are not our only important chemical senses. Many types of chemically sensitive cells, called chemoreceptors, are distributed throughout the body. For example, some nerve endings in skin and mucous membranes warn us of irritating chemicals. A wide range of chemoreceptors report subconsciously and consciously about our internal state: Nerve endings in the digestive organs detect many types of ingested substances, receptors in arteries of the neck measure carbon dioxide and oxygen levels in our blood, and sensory endings in muscles respond to acidity, giving us the burning feeling that comes with exertion and oxygen debt.
Gustation and olfaction have a similar task: the detection of environmental chemicals. In fact, only by using both senses together can the nervous system perceive flavor. Gustation and olfaction have unusually strong and direct connections with our most basic internal needs, including thirst, hunger, emotion, sex, and certain forms of memory. However, the systems of gustation and olfaction are separate and different, from the structures and mechanisms of their chemoreceptors, to the gross organization of their central connections, to their effects on behavior. The neural information from each system is processed in parallel and is merged at rather high levels in the cerebral cortex.
Humans evolved as omnivores (from the Latin omnis, “all,” and vorare, “to eat”), opportunistically eating the plants and animals they could gather, scavenge, or kill. A sensitive and versatile system of taste was necessary to distinguish between new sources of food and potential toxins. Some of our taste preferences are inborn. We have an innate fondness for sweetness, satisfied by mother’s milk. Bitter substances are instinctively rejected, and indeed, many kinds of poisons are bitter. However, experience can strongly modify our instincts, and we can learn to tolerate and even enjoy the bitterness of such substances as coffee and quinine. The body also has the capacity to recognize a deficiency of certain key nutrients and develop an appetite for them. For example, when deprived of essential salt, we may crave salty foods.
Although the number of different chemicals is practically endless and the variety of flavors seems immeasurable, it is likely that we can recognize only a few basic tastes. Most neuroscientists put the number at five. The four obvious taste qualities are saltiness, sourness, sweetness, and bitterness. A fifth taste quality is umami, meaning “delicious” in Japanese; it is defined by the savory taste of the amino acid glutamate; monosodium glutamate, or MSG, is the familiar culinary form. The five major categories of taste qualities seem to be common across human cultures, but there may be additional types of taste qualities (Box 8.1).
Strange Tastes: Fat, Starch, Carbonation, Calcium, Water?
Are there specific taste receptors beyond the classic five: salt, sour, bitter, sweet, umami? The answer is yes, probably. New types of taste receptors have been difficult to identify, but evidence is slowly accumulating.
People love fatty foods and for a good reason. Fat is a concentrated source of calories and essential nutrients. Keen observers as far back as Aristotle have suggested that a taste for fat is basic. But fat stimulates other sensory systems, and this complicates the question of its essential taste. Triglycerides, the fundamental fat molecules, impart a distinct texture to food in the mouth: they feel oily, slippery, creamy. These properties are detected by the somatic sensory system, not taste receptors. Fat also includes many volatile chemicals we can detect with our olfactory system. These odors may be pleasant or foul. Free fatty acids, which are breakdown products of triglycerides, sometimes smell putrid; think of rancid fats. They can also be irritants, sensed again by receptors of the somatic sensory system. But do we also taste fats? Yes, probably. Mice prefer water when it is spiked with some types of fatty acids. Mice also have a type of taste cell that is sensitive to fatty acids and expresses a presumed fatty acid receptor protein. A similar receptor is found in some human taste cells, which may be dedicated fat detectors.
People also love starchy foods, such as pasta, bread, and potatoes. Starch is a complex carbohydrate, specifically a polymer of glucose, the essential sugar in our bodies. Perhaps we like starch because we taste the glucose in it? Experiments on rodents suggest this is not the case; a rat’s preferences for sugars and glucose polymers seem quite distinct. In a recent study, mice were tested for their ability to detect sugar and starchy molecules after the T1R3 protein—a key subunit of sweet and umami receptors (see Figure 8.6)—was genetically knocked out. The knockout mice seemed to be indifferent to sugar, as expected, but they continued to seek out starchy foods. Perhaps mice, at least, have dedicated starch detectors.

FIGURE 8.6 Taste receptor proteins. (a) There are about 25 types of bitter receptors, comprising a family of T2R proteins. Bitter receptors are probably dimers consisting of two different T2R proteins. (b) There is only one type of sweet receptor, formed from the combination of a T1R2 and a T1R3 protein. (c) There is only one type of umami receptor, formed from the combination of a T1R1 and a T1R3 protein. Description
Many people also love carbonated drinks such as soft drinks, club soda, or beer. Water becomes carbonated when substantial amounts of the gas CO2 are dissolved in it. As with fats, we can often feel carbonation as the fizziness vibrates the skin of the mouth and tongue. Mice, and to a lesser extent people, can also smell CO2. We may even hear the bubbles bursting. The CO2 level of blood is a critical measure of respiration, and cells in special arterial detectors sense it. But can we also taste carbonation? Yes, probably. Mice have taste cells with an enzyme called carbonic anhydrase that catalyzes the combination of CO2 and H2O to form protons (H+) and bicarbonate (HCO3–). High levels of protons (i.e., a low pH) taste sour, implying that sour taste cells can detect carbonation. That is at least part of the answer, but how do we discriminate between simple sourness and carbonation? The answer still is not clear. Carbonation sensation may require the proper combination of sour taste and tingling somatic sensation.
People may not love calcium, but they certainly need it for healthy bones, brains, and all other organs. Many animals seem to find calcium salts very tasty when they have been deprived of calcium but reject them when they are sated with calcium. One hypothesis is that Ca2+ is sensed as a combination of bitter and sour tastes. Recent experiments suggest a more interesting possibility. Oddly, an aversion to the taste of Ca2+ in mice requires the T1R3 protein, and the taste of Ca2+ in humans is attenuated by a substance that binds to T1R3. It is possible, although far from proven, that T1R3 is part of a dedicated calcium taste receptor.
Finally, water. Water is crucial for life, and its consumption is regulated by thirst. Wetness, like fattiness and carbonation, can be felt with the somatic sensory system. But can we taste water? Distilled water given to humans has been variously described as sweet, salty, or bitter, depending on the test conditions. A specific taste receptor for water would seem to be a useful adaptation, and there is strong evidence for such receptors in insects. But water receptors in mammalian taste cells have not been identified so far.
The correspondence between chemistry and taste is obvious in some cases. Most acids taste sour, and most salts taste salty. But the chemistry of substances can vary considerably while their basic taste remains the same. Many substances are sweet, from familiar sugars (like fructose, present in fruits and honey, and sucrose, which is white table sugar) to certain proteins (monellin, from the African serendipity berry) to artificial sweeteners (saccharin and aspartame, the second of which is made from two amino acids). Surprisingly, sugars are the least sweet of all of these; gram for gram, the artificial sweeteners and proteins are 10,000–100,000 times sweeter than sucrose. Bitter substances range from simple ions like K+ (KCl actually evokes both bitter and salty tastes) and Mg2+ to complex organic molecules such as quinine and caffeine. Many bitter organic compounds can be tasted even at very low concentrations, down to the nanomolar range. There is an obvious advantage to this, as poisonous substances are often bitter.
With only a handful of basic taste types, how do we perceive the countless flavors of food, such as chocolate, strawberries, and barbecue sauce? First, each food activates a different combination of the basic tastes, helping make it unique. Second, most foods have a distinctive flavor as a result of their combined taste and smell occurring simultaneously. For example, without the sense of smell (and sight), a bite of onion can be easily mistaken for the bite of an apple. Third, other sensory modalities contribute to a unique food-tasting experience. Texture and temperature are important, and pain sensations are essential to the hot, spicy flavor of foods laced with capsaicin, the key ingredient in hot peppers. Therefore, to distinguish the unique flavor of a food, our brain actually combines sensory information about its taste, its smell, and its feel.
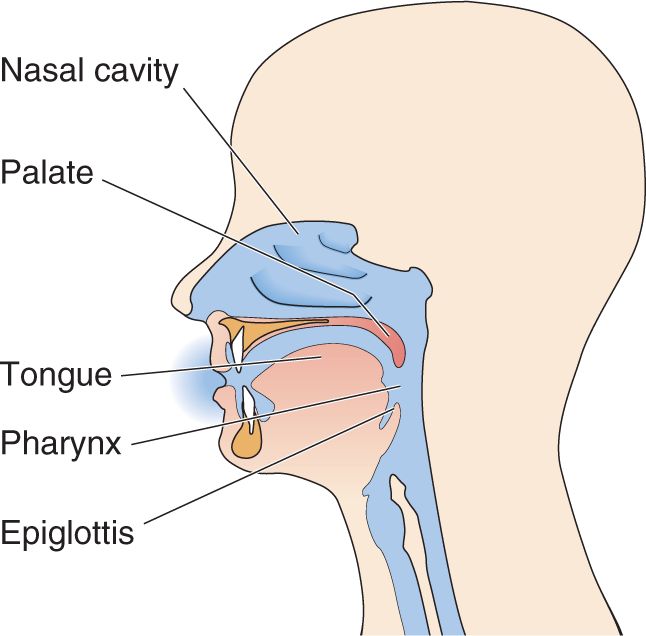
Experience tells us that we taste with our tongue; but other areas of the mouth, such as the palate, pharynx, and epiglottis, are also involved (Figure 8.1). Odors from the food we are eating can also pass, via the pharynx, into the nasal cavity, where they can be detected by olfactory receptors. The tip of the tongue is most sensitive to sweetness, the back to bitterness, and the sides to saltiness and sourness. This does not mean, however, that we taste sweetness only with the tip of our tongue. Most of the tongue is sensitive to all basic tastes.

FIGURE 8.1 Anatomy of the mouth, throat, and nasal passages. Taste is primarily a function of the tongue, but regions of the pharynx, palate, and epiglottis also have some sensitivity. Notice how the nasal passages are located so that odors from ingested food can enter through the nose or the pharynx, thereby easily contributing to perceptions of flavor through olfaction.
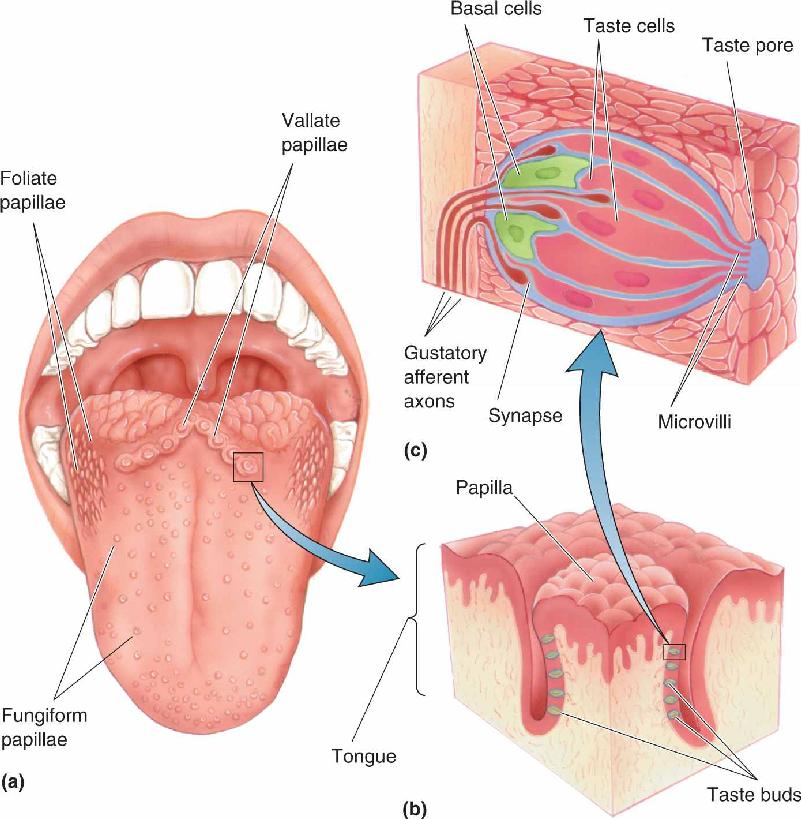
Scattered about the surface of the tongue are small projections called papillae (Latin for “bumps”). Papillae are shaped like ridges (foliate papillae), pimples (vallate papillae), or mushrooms (fungiform papillae) (Figure 8.2a). Facing a mirror, stick your tongue out and shine a flashlight on it, and you will see your papillae easily—small, rounded ones at the front and sides, and large ones in the back. Each papilla has from one to several hundred taste buds, visible only with a microscope (Figure 8.2b). Each taste bud has 50–150 taste receptor cells, or taste cells, arranged within the bud like the sections of an orange. Taste cells comprise only about 1% of the tongue epithelium. Taste buds also have basal cells that surround the taste cells, plus a set of gustatory afferent axons (Figure 8.2c). A person typically has 2000–5000 taste buds, although in exceptional cases, there are as few as 500 or as many as 20,000.

FIGURE 8.2 The tongue, its papillae, and its taste buds. (a) Papillae are the taste-sensitive structures. The largest and most posterior are the vallate papillae. Foliate papillae are elongated. Fungiform papillae are relatively large toward the back of the tongue and much smaller along the sides and tip. (b) A cross-sectional view of a vallate papilla, showing the locations of taste buds. (c) A taste bud is a cluster of taste cells (the receptor cells), gustatory afferent axons and their synapses with taste cells and basal cells. Microvilli at the apical end of the taste cells extend into the taste pore, the site where chemicals dissolved in saliva can interact directly with taste cells. Description
Using tiny droplets, it is possible to expose a single papilla on a person’s tongue to low concentrations of various basic taste stimuli (something almost purely sour, like vinegar, or almost purely sweet, like a sucrose solution). Concentrations too low will not be tasted, but at some critical concentration, the stimulus will evoke a perception of taste; this is the threshold concentration. At concentrations just above threshold, most papillae tend to be sensitive to only one basic taste; there are sour-sensitive papillae and sweet-sensitive papillae, for example. When the concentrations of the taste stimuli are increased, however, most papillae become less selective. Whereas a papilla might have responded only to sweet when all stimuli were weak, it may also respond to sour and salt if they are made stronger. We now know that each papilla has multiple types of taste receptor cells and that each receptor type is specialized for a different category of taste.
The chemically sensitive part of a taste receptor cell is its small membrane region, called the apical end, near the surface of the tongue. The apical ends have thin extensions called microvilli that project into the taste pore, a small opening on the surface of the tongue where the taste cell is exposed to the contents of the mouth (see Figure 8.2c). Taste receptor cells are not neurons according to standard histological criteria. However, they do form synapses with the endings of the gustatory afferent axons near the bottom of the taste bud. Taste receptor cells also make both electrical and chemical synapses onto some of the basal cells; some basal cells synapse onto the sensory axons, and these may form a simple information-processing circuit within each taste bud. Cells of the taste bud undergo a constant cycle of growth, death, and regeneration; the lifespan of one taste cell is about 2 weeks. This process depends on an influence of the sensory nerve, because if the nerve is cut, the taste buds will degenerate.
When an appropriate chemical activates a taste receptor cell, its membrane potential changes, usually by depolarizing. This voltage shift is called the receptor potential (Figure 8.3a). If the receptor potential is depolarizing and large enough, some taste receptor cells, like neurons, may fire action potentials. In any case, depolarization of the receptor membrane causes voltage-gated calcium channels to open; Ca2+ enters the cytoplasm, triggering the release of transmitter molecules. This is basic synaptic transmission, from taste cell to sensory axon. The transmitter released depends on the taste receptor cell type. Sour and salty taste cells release serotonin onto gustatory axons, whereas sweet, bitter, and umami taste cells release adenosine triphosphate (ATP) as their primary transmitter. In both cases, the taste receptor’s transmitter excites the postsynaptic sensory axon and causes it to fire action potentials (Figure 8.3b), which communicate the taste signal into the brain stem. Taste cells may also use other transmitters, including acetylcholine, GABA, and glutamate, but their functions are unknown.

FIGURE 8.3 Taste responsiveness of taste cells and gustatory axons. (a) Three different cells were sequentially exposed to salt (NaCl), bitter (quinine), sour (HCl), and sweet (sucrose) stimuli, and their membrane potential was recorded with electrodes. Notice the different sensitivities of the three cells. (b) In this case, the action potential discharge of the sensory axons was recorded. This is an example of extracellular recording of action potentials. Each vertical deflection in the record is a single action potential. Description
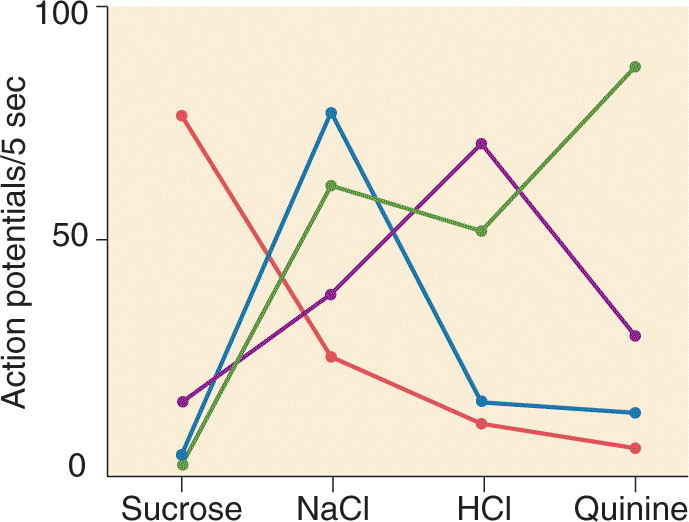
Evidence from recent studies of mice suggests that most taste receptor cells respond primarily or even exclusively to just one of the five basic tastes. Examples are cells 1 and 3 in Figure 8.3a, which give strong depolarizing responses to salt (NaCl) and sweet (sucrose) stimuli, respectively. Some taste cells and many gustatory axons show multiple response preferences, however. Each of the gustatory axons in Figure 8.3b is influenced by several of the basic tastes, but each has a clearly different bias.
Figure 8.4 shows the results of similar recordings from four gustatory axons in a rat. One responds strongly only to salt, one only to sweet, and two to all but sweet. Why will one cell respond only to a single chemical type while another responds to three or four categories of chemicals? The answer is that the responses depend on the particular transduction mechanisms present in each cell.

FIGURE 8.4 Action potential firing rates of four different primary gustatory nerve axons in a rat. The taste stimuli were sweet (sucrose), salt (NaCl), sour (HCl), and bitter (quinine). Each colored line represents measurements from a single axon. Notice the differences in selectivity between axons. (Source: Adapted from Sato, 1980, p. 23.) Description
The process by which an environmental stimulus causes an electrical response in a sensory receptor cell is called transduction (from the Latin transducere, “to lead across”). The nervous system has myriad transduction mechanisms that make it sensitive to chemicals, pressures, sounds, and light. The nature of the transduction mechanism determines the specific sensitivity of a sensory system. We see because our eyes have photoreceptors. If our tongue had photoreceptors, we might see with our mouth.
Some sensory systems have a single basic type of receptor cell that uses one transduction mechanism (e.g., the auditory system). However, taste transduction involves several different processes, and each basic taste uses one or more of these mechanisms. Taste stimuli, or tastants, may (1) directly pass through ion channels (salt and sour), (2) bind to and block ion channels (sour), or (3) bind to G-protein-coupled receptors in the membrane that activate second messenger systems that, in turn, open ion channels (bitter, sweet, and umami). These are familiar processes, very similar to the basic signaling mechanisms present in all neurons and synapses, which were described in Chapters 4, 5, and 6.
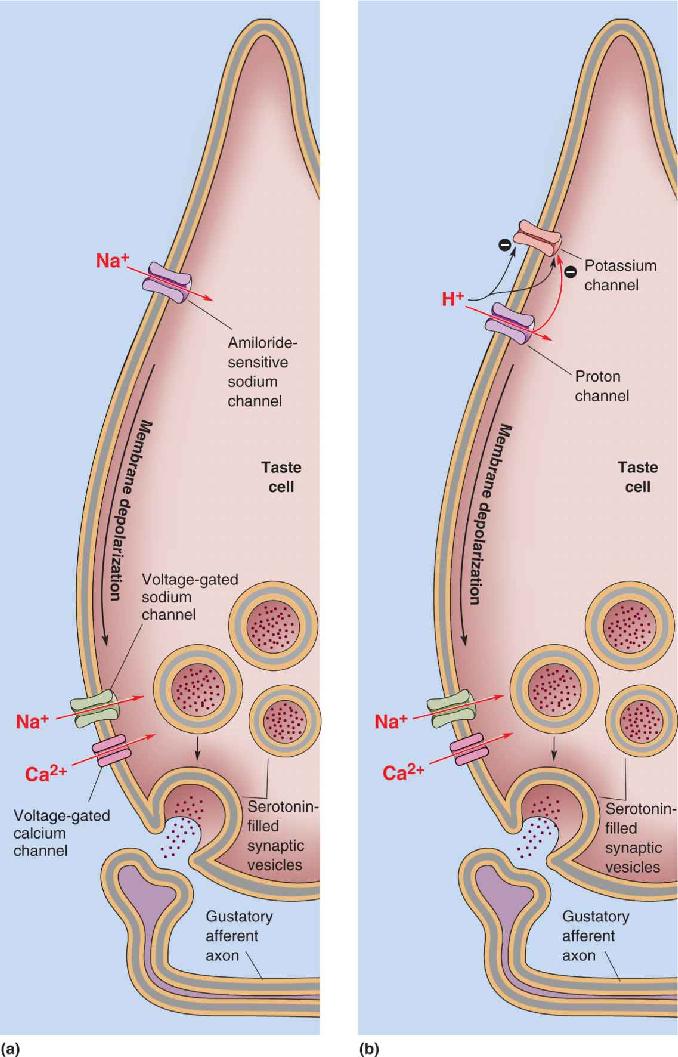
Saltiness. The prototypical salty chemical is table salt (NaCl), which, apart from water, is the major component of blood, the ocean, and chicken soup. Salt is unusual in that relatively low concentrations (10–150 mM) taste good, whereas higher concentrations tend to be disagreeable and repellent. The taste of salt is mostly the taste of the cation Na+, but taste receptors use very different mechanisms to detect low and high concentrations of it. To detect low concentrations, salt-sensitive taste cells use a special Na+-selective channel that is common in other epithelial cells and which is blocked by the drug amiloride (Figure 8.5a). Amiloride is a diuretic (a drug that promotes urine production) used to treat some types of hypertension and heart disease. The amiloride-sensitive sodium channel is quite different from the voltage-gated sodium channel that generates action potentials; the taste channel is insensitive to voltage and generally stays open. When you sip chicken soup, the Na+ concentration rises outside the receptor cell, and the gradient for Na+ across the membrane is made steeper. Na+ then diffuses down its concentration gradient, which means it flows into the cell, and the resulting inward current causes the membrane to depolarize. This depolarization—the receptor potential—in turn causes voltage-gated sodium and calcium channels to open near the synaptic vesicles, triggering the release of neurotransmitter molecules onto the gustatory afferent axon.

FIGURE 8.5 Transduction mechanisms of (a) salt and (b) sour tastants. Tastants can interact directly with ion channels either by passing through them (Na+ and H+) or by blocking them (H+ blocking the potassium channel). The membrane voltage then influences calcium channels on the basal membrane, which in turn influence the intracellular [Ca2+] and transmitter release. Description
Animals avoid very high concentrations of NaCl and other salts, and humans usually report that they taste bad. It appears that high salt levels activate bitter and sour taste cells, which normally trigger avoidance behaviors. How very salty substances stimulate bitter and sour taste cells is still a mystery.
The anions of salts affect the taste of the cations. For example, NaCl tastes saltier than Na acetate, apparently because the larger anion, acetate, inhibits the salty taste of the cation. The mechanisms of anion inhibition are poorly understood. Another complication is that as the anions become larger, they tend to take on tastes of their own. Sodium saccharin tastes sweet because the Na+ concentrations are far too low for us to taste the saltiness, and the saccharin potently activates sweetness receptors.
Sourness. Foods taste sour because of their high acidity (otherwise known as low pH). Acids, such as HCl, dissolve in water and generate hydrogen ions (protons or H+). Thus, protons are the causative agents of acidity and sourness. Protons may affect sensitive taste receptors in several ways, from either inside or outside the taste cell membrane, although these processes are poorly understood (Figure 8.5b). It is likely that H+ can bind to and block special K+-selective channels. When the K+ permeability of a membrane is decreased, it depolarizes. H+ may also activate or permeate a special type of ion channel from the superfamily of transient receptor potential (TRP) channels, which are common in many kinds of sensory receptor cells. The cation current through the TRP channels can also depolarize sour receptor cells. pH can affect virtually all cellular processes, and there may be other mechanisms of sour taste transduction. It is possible that a constellation of effects evokes the sour taste.
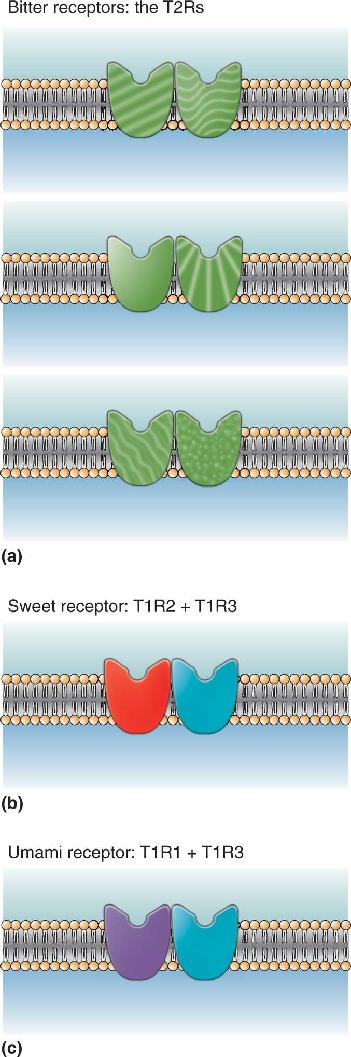
Bitterness. The transduction processes underlying bitter, sweet, and umami tastes rely on two families of related taste receptor proteins called T1R and T2R. The various subtypes of T1R and T2R are all G-protein-coupled taste receptors that are very similar to the G-protein-coupled receptors that detect neurotransmitters. There is good evidence that the protein receptors for bitter, sweet, and umami tastes are dimers; dimers are two proteins affixed to one another (Figure 8.6). Tightly associated proteins are common in cells (see Figure 3.6); most ion channels (see Figure 3.7) and transmitter-gated channels (see Figure 5.14) consist of several different bound proteins, for example.
Bitter substances are detected by the 25 or so different types of T2R receptors in humans. Bitter receptors are poison detectors, and because we have so many, we can detect a vast array of different poisonous substances. Animals are not very good at telling different bitter tastants apart, however, probably because each bitter taste cell expresses many, and perhaps most, of the 25 bitter receptor proteins. Since each taste cell can send only one type of signal to its afferent nerve, a chemical that can bind to one of its 25 bitter receptors will trigger essentially the same response as a different chemical that binds to another of its bitter receptors. The important message the brain receives from its taste receptors is simply that a bitter chemical is “Bad! Not to be trusted!” And the nervous system apparently does not distinguish one type of bitter substance from another.
Bitter receptors use a second messenger pathway to carry their signal to the gustatory afferent axon. In fact, the bitter, sweet, and umami receptors all seem to use the same second messenger pathway to carry their signals to the afferent axons. The general pathway is illustrated in Figure 8.7. When a tastant binds to a bitter (or sweet or umami) receptor, it activates its G-proteins, which stimulate the enzyme phospholipase C, thereby increasing the production of the intracellular messenger inositol triphosphate (IP3). IP3 pathways are ubiquitous signaling systems in cells throughout the body (see Chapter 6). In taste cells, IP3 activates a special type of ion channel that is unique to taste cells, causing it to open and allow Na+ to enter, thus depolarizing the taste cell. IP3 also triggers the release of Ca2+ from intracellular storage sites. This Ca2+ in turn triggers neurotransmitter release in an unusual way. The taste cells for bitter, sweet, and umami lack conventional transmitter-filled presynaptic vesicles. Instead, increases of intracellular Ca2+ activate a special membrane channel that allows ATP to flow out of the cell. The ATP serves as a synaptic transmitter and activates purinergic receptors on postsynaptic gustatory axons.

FIGURE 8.7 Transduction mechanisms for bitter, sweet, and umami tastants. Tastants bind directly to G-protein-coupled membrane receptors, which activate phospholipase C, which increases the synthesis of IP3. IP3 then triggers the release of Ca2+ from internal storage sites, and Ca2+ opens a taste-specific ion channel, leading to depolarization and transmitter release. The main transmitter is ATP, which is released from the taste cell by diffusing through ATP-permeable channels. Description
Sweetness. There are many different sweet tastants, some natural and some artificial. Surprisingly, all of them seem to be detected by the same taste receptor protein. Sweet receptors resemble bitter receptors, in that they are all dimers of G-protein-coupled receptors. A functioning sweet receptor requires two very particular members of the T1R receptor family: T1R2 and T1R3 (see Figure 8.6). If either one of them is missing or mutated, an animal may not perceive sweetness at all. In fact, all species of cats and some other carnivores lack the genes encoding T1R2 and are indifferent to the taste of most molecules that we consider sweet.
Chemicals binding to the T1R2 + T1R3 receptor (i.e., the sweet receptor) activate exactly the same second messenger system that the bitter receptors activate (Figure 8.7). So, why don’t we confuse bitter chemicals with sweet ones? The reason is that bitter receptor proteins and sweet receptor proteins are expressed in different taste cells. Bitter taste cells and sweet taste cells, in turn, connect to different gustatory axons. The activity of different gustatory axons reflects the chemical sensitivities of the taste cells that drive them, so the messages about sweetness and bitterness are delivered to the central nervous system along different transmission lines.
Umami (Amino Acids). “Amino acids” may not be the answer at the tip of your tongue when asked to list your favorite tastes, but recall that proteins are made from amino acids and that they are also excellent energy sources. In short, amino acids are the foods your mother would want you to eat. Most amino acids also taste good, although some taste bitter.
The transduction process for umami is identical to that for sweetness, with one exception. The umami receptor, like the sweet receptor, is composed of two members of the T1R protein family, but in this case, it is T1R1 + T1R3 (see Figure 8.6). The sweet and umami receptors share the T1R3 protein, so it is the other T1R that determines whether the receptor is sensitive to amino acids or sweet tastants. When the gene encoding for the T1R1 protein is removed from mice, they are unable to taste glutamate and other amino acids, although they still demonstrate a sense for sweet chemicals and other tastants.
Similar to other types of taste receptors, the genetics of different mammalian species lead to interesting taste preferences and deficits. Most bats, for example, do not have a functioning T1R1 receptor and presumably cannot taste amino acids. Vampire bats lack functional genes for both umami and sweet taste. The ancestors of bats presumably did have umami and sweet receptors; why they were lost is unknown.
Considering how similar the umami receptor is to the sweet and bitter receptors, it will not surprise you that all three use exactly the same second messenger pathways (Figure 8.7). Then why don’t we confuse the taste of amino acids with sweet or bitter chemicals? Once again, the taste cells selectively express only one class of taste receptor protein. There are umami-specific taste cells, just as there are sweet-specific taste cells and bitter-specific taste cells. The gustatory axons they stimulate are, in turn, delivering messages of umami, sweetness, or bitterness to the brain.
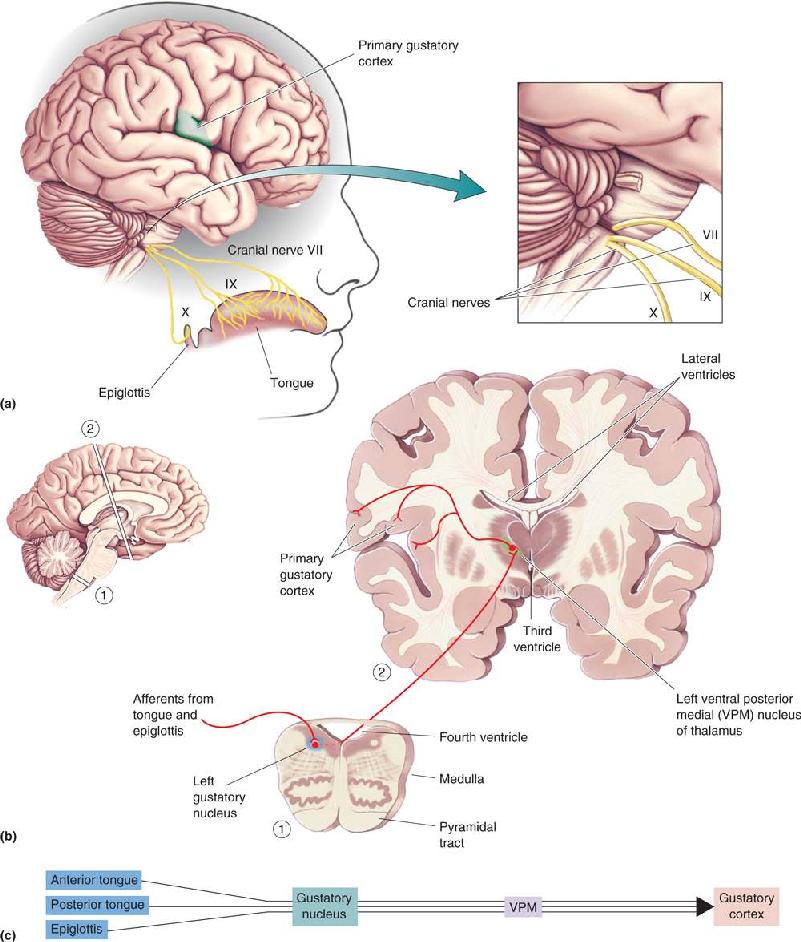
The main flow of taste information is from the taste buds to the primary gustatory axons, into the brain stem, up to the thalamus, and to the cerebral cortex (Figure 8.8). Three cranial nerves carry primary gustatory axons and bring taste information to the brain. The anterior two-thirds of the tongue and the palate send axons into a branch of cranial nerve VII, the facial nerve. The posterior third of the tongue is innervated by a branch of cranial nerve IX, the glossopharyngeal nerve. The regions around the throat, including the glottis, epiglottis, and pharynx, send taste axons to a branch of cranial nerve X, the vagus nerve. These nerves are involved in a variety of other sensory and motor functions, but their taste axons all enter the brain stem, bundle together, and synapse within the slender gustatory nucleus, a part of the solitary nucleus in the medulla.

FIGURE 8.8 Central taste pathways. (a) Taste information from the tongue and mouth cavity is carried by three cranial nerves (VII, IX, and X) to the medulla. (b) Gustatory axons enter the gustatory nucleus within the medulla. Gustatory nucleus axons synapse on neurons of the thalamus, which project to primary gustatory cortex in regions of the postcentral gyrus and insular cortex. The enlargements show planes of section through ① the medulla and ② the forebrain. (c) The central taste pathways summarized. Description
From the gustatory nucleus, taste pathways diverge. The conscious experience of taste is presumably mediated by the cerebral cortex. The path to the neocortex via the thalamus is a common one for sensory information. Neurons of the gustatory nucleus synapse on a subset of small neurons in the ventral posterior medial (VPM) nucleus, a portion of the thalamus that deals with sensory information from the head. The VPM taste neurons then send axons to the primary gustatory cortex (located in Brodmann’s area 36 and the insula-operculum regions of cortex). The taste pathways to the thalamus and cortex are primarily ipsilateral to the cranial nerves that supply them. Lesions within the VPM thalamus or the gustatory cortex—as a result of a stroke, for example—can cause ageusia, the loss of taste perception.
Gustation is important to basic behaviors such as the control of feeding and digestion, both of which involve additional taste pathways. Gustatory nucleus cells project to a variety of brain stem regions, largely in the medulla, that are involved in swallowing, salivation, gagging, vomiting, and basic physiological functions such as digestion and respiration. In addition, gustatory information is distributed to the hypothalamus and related parts of the basal telencephalon (structures of the limbic system; see Chapter 18). These structures seem to be involved in the palatability of foods and the forces that motivate us to eat (Box 8.2). Localized lesions of the hypothalamus or amygdala, a nucleus of the basal telencephalon, can cause an animal to either chronically overeat or ignore food, or alter its preferences for food types.
When one of us was 14 years old, he ended an entertaining day at an amusement park by snacking on one of his favorite New England foods, fried clams. Within an hour he became nauseated, vomited, and had a most unpleasant bus ride home. Presumably, the clams had been spoiled. Sadly, for years afterward, he could not even imagine eating another fried clam, and the smell of them alone was repulsive. The fried clam aversion was quite specific. It did not affect his enjoyment of other foods, and he felt no prejudice against amusement park rides, buses, or the friends he had been with the day he got sick.
By the time the author reached his thirties, he could happily dine on fried clams again. He also read about research that John Garcia, working at Harvard Medical School, had done just about the same time as the original bad-clam experience. Garcia fed rats a sweet liquid, and, in some cases, he then gave them a drug that made them briefly feel ill. After even one such trial, rats that had received the drug avoided the sweet stimulus forever. The rats’ aversion was specific for the taste stimulus; they did not avoid sound or light stimuli under the same conditions.
Extensive research has shown that flavor aversion learning results in a particularly robust form of associative memory. It is most effective for food stimuli (taste and smell both contribute); it requires remarkably little experience (as little as one trial); and it can last a very long time—more than 50 years in some people! And learning occurs even when there is a very long delay between the food (the conditioned stimulus) and the nausea (the unconditioned stimulus). This is obviously a useful form of learning in the wild. An animal can’t afford to be a slow learner when new foods might be poisonous. For modern humans, this memory mechanism can backfire; many perfectly good fried clams have remained uneaten. Food aversion can be a more serious problem for patients undergoing radiation or chemotherapy for cancer, when the nausea induced by their treatments makes many foods unpalatable. On the other hand, taste aversion learning has also been used to prevent coyotes from stealing domestic sheep and to help people reduce their dependence on alcohol and cigarettes.
If you were going to design a system for coding tastes, you might begin with many specific taste receptors for many basic tastes (sweet, sour, salty, bitter, chocolate, banana, mango, beef, Swiss cheese, etc.). Then you might connect each receptor type, by separate sets of axons, to neurons in the brain that also respond to only one specific taste. All the way up to the cortex, you would expect to find specific neurons responding to “sweet” and “chocolate,” and the flavor of chocolate ice cream would involve the rapid firing of these cells and very few of the “salty,” “sour,” and “banana” cells.
This concept is the labeled line hypothesis, and at first it seems simple and rational. At the start of the gustatory system—the taste receptor cells—something like labeled lines are used. As we have seen, individual taste receptor cells are often selectively sensitive to particular classes of stimuli: sweet, bitter, or umami. Some of them, however, are more broadly tuned to stimuli; that is, they are less specific in their responses. They may be excited to some extent by both salt and sour, for example (see Figure 8.3). Primary taste axons are even less specific than receptor cells, and most central taste neurons continue to be broadly responsive all the way into the cortex. In other words, the response of a single taste cell is often ambiguous about the food being tasted; the labels on the taste lines are uncertain rather than distinct.
Cells in the taste system are broadly tuned for several reasons. If one taste receptor cell has two different transduction mechanisms, it will respond to two types of tastants (although it may still respond most strongly to one of them). In addition, there is convergence of receptor cell input onto afferent axons. Each receptor cell synapses onto a primary taste axon that also receives input from several other receptor cells in that papilla as well as its neighbors. This means that one axon may combine the taste information from numerous taste cells. If one of those cells is mostly sensitive to sour stimuli and another to salt stimuli, then the axon will respond to salt and sour. This pattern continues into the brain: Neurons of the gustatory nucleus receive synapses from many axons of different taste specificities, and they may become less selective for tastes than the primary taste axons.
All of this mixing of taste information might seem like an inefficient way to design a coding system. Why not use many taste cells that are highly specific? In part, the answer might be that we would need an enormous variety of receptor types, and even then we could not respond to new tastes. So, when you taste chocolate ice cream, how does the brain sort through its apparently ambiguous information about the flavor to make clear distinctions between chocolate and thousands of other possibilities? The likely answer is a scheme that includes features of roughly labeled lines and population coding, in which the responses of a large number of broadly tuned neurons, rather than a small number of precisely tuned neurons, are used to specify the properties of a particular stimulus, such as a taste.
Population coding schemes seem to be used throughout the sensory and motor systems of the brain, as we shall see in later chapters. In the case of taste, receptor cells are sensitive to a small number of taste types, often only one; gustatory axons and the neurons they activate in the brain tend to respond more broadly—for example, strongly to bitter, moderately to sour and salt, and not at all to sweet (see Figure 8.4). Only with a large population of taste cells, with different response patterns, can the brain distinguish between specific alternative tastes. One food activates a certain subset of neurons, some of them firing very strongly, some moderately, some not at all, others perhaps even inhibited below their spontaneous firing rates (i.e., their nonstimulated rates); a second food excites some of the cells activated by the first food, but also others; and the overall patterns of discharge rates are distinctly different. The relevant population may even include neurons activated by the olfactory, temperature, and textural features of a food; certainly, the coldness and creaminess of chocolate ice cream contribute to our ability to distinguish it from chocolate cake.
Olfaction brings both good news and bad news. It combines with taste to help us identify foods, and it increases our enjoyment of many of them. But it can also warn of potentially harmful substances (spoiled meat) or places (smoke-filled rooms). In olfaction, the warnings from bad smells may outweigh the benefits of good smells; by some estimates, we can smell several hundred thousand substances, but only about 20% of them smell pleasant. Practice helps in olfaction, and professional perfumers and whiskey blenders can actually distinguish among thousands of different odors.
Smell is also a mode of communication. Chemicals released by the body, called pheromones, are important signals for reproductive behaviors, and they may also be used to mark territories, identify individuals, and indicate aggression or submission. (The term is from the Greek pherein, “to carry,” and horman, “to urge or excite.”) Although systems of pheromones are well developed in many other animals, their importance to humans is not clear (Box 8.3).
Smells are surer than sounds and sights to make your heart strings crack.
Odors can certainly sway emotions and arouse memories, but just how important are they to human behavior? Each of us has a distinctive set of odors that marks our identity as surely as our fingerprints or genes do. In fact, variations in body odor are probably genetically determined. Bloodhounds have great difficulty distinguishing between the smells of identical twins, but not between those of fraternal siblings. For some animals, odor identity is essential: When her lamb is born, the ewe establishes a long-term memory of its specific smell and develops an enduring bond based largely on olfactory cues. In a newly inseminated female mouse, the smell of a strange male (but not the smell of her recent mate, which she remembers) will trigger an abortion of the pregnancy.
Humans have the ability to recognize the scents of other humans. Infants as young as 6 days old show a clear preference for the smell of their own mother’s breast over that of other nursing mothers. The mothers, in turn, can usually identify the odor of their own infant from among several choices.
About 30 years ago, researcher Martha McClintock reported that women who spend a lot of time together (e.g., college roommates) often find that their menstrual cycles synchronize. This effect is probably mediated by pheromones. In 1998, McClintock and Kathleen Stern, working at the University of Chicago, found that odorless compounds from one group of women (the “donors”) could influence the timing of the menstrual cycles of other women (the “recipients”). Body chemicals were collected by placing cotton pads under the arms of the donors for at least 8 hours. The pads were then wiped under the noses of the recipients, who agreed not to wash their faces for 6 hours. The recipients were not told the source of the chemicals on the pads and did not consciously perceive any odor from them except the alcohol used as a carrier. Nevertheless, depending on the donor’s time in her menstrual cycle, the recipient’s cycle was either shortened or lengthened. These dramatic results are the best evidence yet that humans can communicate with pheromones.
Many animals use the accessory olfactory system to detect pheromones and mediate a variety of social behaviors involving mother, mating, territory, and food. The accessory system runs parallel to the primary olfactory system. It consists of separate chemically sensitive regions in the nasal cavity, in particular the vomeronasal organ, which projects to the accessory olfactory bulb and from there provides input to the hypothalamus. Alas, it seems likely that the vomeronasal organ is absent or vestigial in most mature people, and even when it is identifiable, it does not appear to have functional receptor proteins or direct connections to the brain. This by itself does not mean humans lack pheromonal signals, since these could pass through the main olfactory organs.
Napoleon Bonaparte once wrote to his love Josephine, asking her not to bathe for the 2 weeks until they would next meet, so he could enjoy her natural aromas. The scent of a woman may indeed be a source of arousal for sexually experienced males, presumably because of learned associations. But there is still no hard evidence for human pheromones that might mediate sexual attraction (for members of either sex) via innate mechanisms. Considering the commercial implications of such a substance, we can be sure the search will continue.
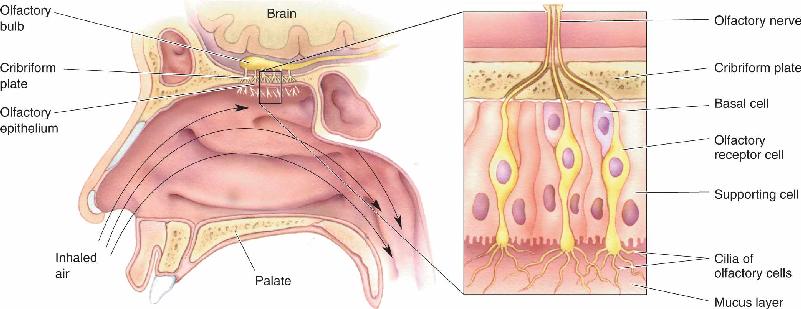
We do not smell with our nose. Rather, we smell with a small, thin sheet of cells high up in the nasal cavity called the olfactory epithelium (Figure 8.9). The olfactory epithelium has three main cell types. Olfactory receptor cells are the sites of transduction. Unlike taste receptor cells, olfactory receptors are genuine neurons, with axons of their own that penetrate into the central nervous system. Supporting cells are similar to glia; among other things, they help produce mucus. Basal cells are the source of new receptor cells. Olfactory receptors (similar to taste receptors) continually grow, die, and regenerate, in a cycle that lasts about 4–8 weeks. In fact, olfactory receptor cells are one of the very few types of neurons in the nervous system that are regularly replaced throughout life.

FIGURE 8.9 The location and structure of the olfactory epithelium. The olfactory epithelium consists of a layer of olfactory receptor cells, supporting cells, and basal cells. Odorants dissolve in the mucus layer and contact the cilia of the olfactory cells. Axons of the olfactory cells penetrate the bony cribriform plate on their way to the CNS. Description
Sniffing brings air through the convoluted nasal passages, but only a small percentage of that air passes over the olfactory epithelium. The epithelium exudes a thin coating of mucus, which flows constantly and is replaced about every 10 minutes. Chemical stimuli in the air, called odorants, dissolve in the mucus layer before they reach receptor cells. Mucus consists of a water base with dissolved mucopolysaccharides (long chains of sugars); a variety of proteins, including antibodies, enzymes, and odorant binding proteins; and salts. The antibodies are critical because olfactory cells can be a direct route by which some viruses (such as the rabies virus) and bacteria enter the brain. Also important are odorant binding proteins, which are small and soluble and which may help concentrate odorants in the mucus.
The size of the olfactory epithelium is one indicator of an animal’s olfactory acuity. Humans are relatively weak smellers (although even we can detect some odorants at concentrations as low as a few parts per trillion). The surface area of the human olfactory epithelium is only about 10 cm2. The olfactory epithelium of certain dogs may be over 170 cm2, and dogs have over 100 times more receptors in each square centimeter than humans. By sniffing the aromatic air above the ground, dogs can detect the few molecules left by someone walking there hours before. Humans may be able to smell the dog, however, only when it licks our face.
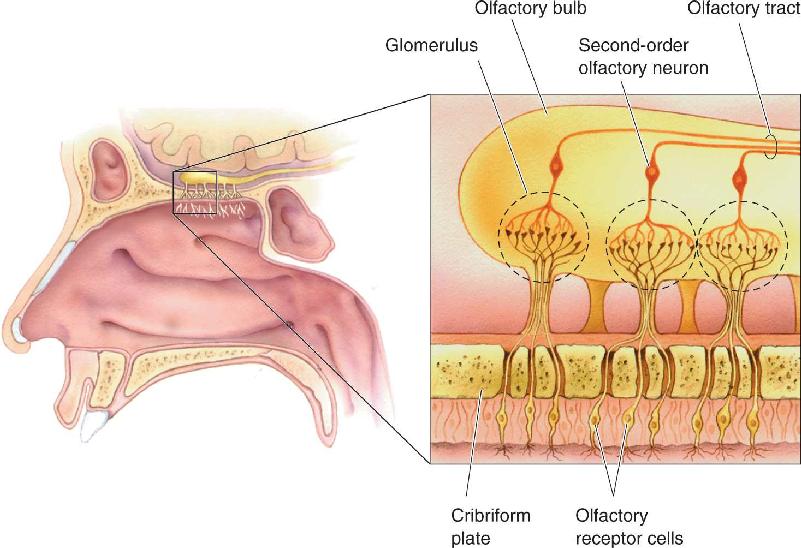
Olfactory receptor neurons have a single, thin dendrite that ends with a small knob at the surface of the epithelium (see Figure 8.9). Waving from the knob, within the mucus layer, are several long, thin cilia. Odorants dissolved in the mucus bind to the surface of the cilia and activate the transduction process. On the opposite side of the olfactory receptor cell is a very thin, unmyelinated axon. Collectively, the olfactory axons constitute the olfactory nerve (cranial nerve I). The olfactory axons do not all come together as a single nerve bundle, as in other cranial nerves. Instead, after leaving the epithelium, small clusters of the axons penetrate a thin sheet of bone called the cribriform plate, then course into the olfactory bulb (see Figure 8.9). The olfactory axons are fragile, and in a traumatic injury, such as a blow to the head, the forces between the cribriform plate and surrounding tissue can sever the olfactory axons. After this type of injury, the axons cannot regrow, resulting in anosmia, the inability to smell.
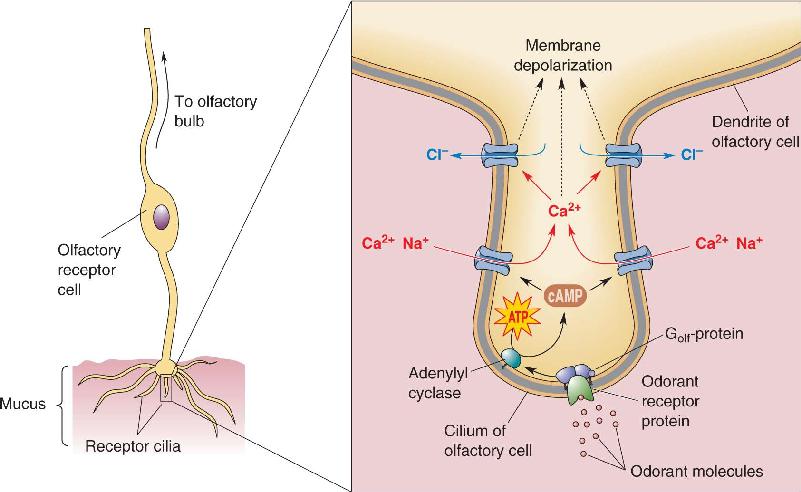
Olfactory Transduction. Although taste receptor cells use several different molecular signaling systems for transduction, olfactory receptors probably use only one (Figure 8.10). All of the transduction molecules are located in the thin cilia. The olfactory pathway can be summarized as follows:

FIGURE 8.10 Transduction mechanisms of vertebrate olfactory receptor cells. The drawing at right shows a single cilium of an olfactory receptor cell and the signaling molecules of olfactory transduction that it contains. Golf is a special form of G-protein found only in olfactory receptor cells. Description
Bind cAMP to a cyclic nucleotide-gated cation channel →
Open cation channels and allow influx of Na+ and Ca2+ →
Cause current flow and membrane depolarization (receptor potential).
Once the cation-selective cAMP-gated channels open, current flows inward, and the membrane of the olfactory neuron depolarizes (Figures 8.10 and 8.11). In addition to Na+, the cAMP-gated channel allows substantial amounts of Ca2+ to enter the cilia. In turn, the intracellular Ca2+ triggers a Ca2+-activated Cl– current that may amplify the olfactory receptor potential. (This is a switch from the usual effect of Cl– currents, which inhibit neurons; in olfactory cells, the internal Cl– concentration must be unusually high so that a Cl– current tends to depolarize rather than hyperpolarize the membrane.) If the resulting receptor potential is large enough, it will exceed the threshold for action potentials in the cell body, and spikes will propagate out along the axon into the central nervous system (CNS) (Figure 8.11).

FIGURE 8.11 Voltage recordings from an olfactory receptor cell during stimulation. Odorants generate a slow receptor potential in the cilia; the receptor potential propagates along the dendrite and triggers a series of action potentials within the soma of the olfactory receptor cell. Finally, the action potentials (but not the receptor potential) propagate continuously along the olfactory nerve axon. Description
The olfactory response may terminate for several reasons. Odorants diffuse away, scavenger enzymes in the mucus layer often break them down, and cAMP in the receptor cell may activate other signaling pathways that end the transduction process. Even in the continuing presence of an odorant, the strength of a smell usually fades because the response of the receptor cell itself adapts to an odorant within about a minute. Decreased response despite the continuing presence of a stimulus is called adaptation, and we will see that it is a common feature of receptors in all the senses.
This signaling pathway has two unusual features: the receptor binding proteins at the beginning and the cAMP-gated channels near the end.
Olfactory Receptor Proteins. Receptor proteins have odorant binding sites on their extracellular surface. Because of your ability to discriminate among thousands of different odorants, you might guess that there are many different types of odorant receptor proteins. You would be right, and the number is very large indeed. Researchers Linda Buck and Richard Axel, working at Columbia University in 1991, found that there are more than 1000 different odorant receptor genes in rodents, making this by far the largest family of mammalian genes yet discovered. This important and surprising discovery earned Buck and Axel the Nobel Prize in 2004.
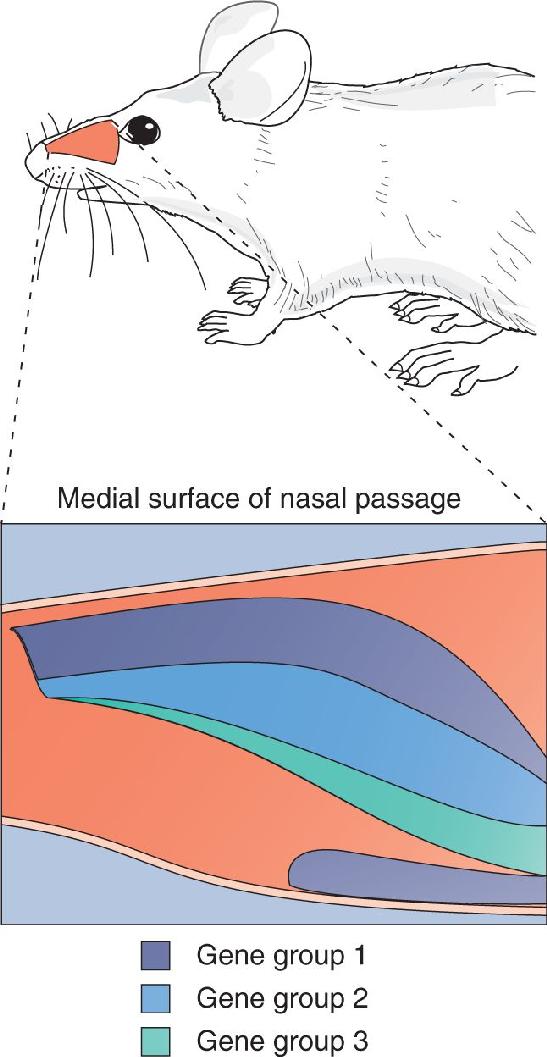
Humans have fewer odorant receptor genes than rodents—about 350 that code for functional receptor proteins—but this is still an enormous number. Odorant receptor genes comprise about 3–5% of the entire mammalian genome. The receptor genes are scattered about on the genome, and nearly every chromosome has at least a few of them. Each receptor gene has a unique structure, allowing the receptor proteins encoded by these genes to bind different odorants. It is also surprising that each olfactory receptor cell seems to express very few of the many types of receptor genes, in most cases just one. Thus, in mice, there are more than 1000 different types of receptor cells, each identified by the particular receptor gene it expresses. The olfactory epithelium is organized into a few large zones, and each zone contains receptor cells that express a different subset of receptor genes (Figure 8.12). Within each zone, individual receptor types are scattered randomly (Figure 8.13a).

FIGURE 8.12 Maps of the expression of different olfactory receptor proteins on the olfactory epithelium of a mouse. Three different groups of genes were mapped in this case, and each had a different, nonoverlapping zone of distribution. (Source: Adapted from Ressler et al., 1993, p. 602.)

FIGURE 8.13 Broad tuning of single olfactory receptor cells. (a) Each receptor cell expresses a single olfactory receptor protein (here coded by cell color), and different cells are randomly scattered within a region of the epithelium. (b) Microelectrode recordings from three different cells show that each one responds to many different odors, but with differing preferences. By measuring responses from all three cells, each of the four odors can be clearly distinguished. Description
The receptor neurons in the vomeronasal organ of mice, dogs, cats, and many other mammals express their own sets of receptor proteins. The structures of odorant receptor proteins and vomeronasal receptor proteins are surprisingly different. There are many fewer functional vomeronasal receptor proteins (about 180 in mice and perhaps none in humans) than odorant receptor proteins. The types of chemicals vomeronasal receptors detect are only partially known, but it is likely that some of them are pheromones (see Box 8.3).
Olfactory receptor proteins belong to the large family of proteins called G-protein-coupled receptors, all of which have seven transmembrane alpha helices. The G-protein-coupled receptors also include a variety of neurotransmitter receptors, which were described in Chapter 6, and the bitter, sweet, and umami receptors described earlier in this chapter. All of these receptors are coupled to G-proteins, which in turn relay a signal to other second messenger systems within the cell (olfactory receptor cells use a particular type of G-protein, called Golf). Increasing evidence indicates that the only second messenger mediating olfactory transduction in vertebrates is cAMP. Some of the most compelling studies have used genetic engineering to produce mice in which critical proteins of the olfactory cAMP pathway have been knocked out (e.g., Golf); these mice are inevitably anosmic for a wide variety of odors.
cAMP-Gated Channels. In neurons, cAMP is a common second messenger, but the way it acts in olfactory transduction is quite unusual. Tadashi Nakamura and Geoffrey Gold, working at Yale University in 1987, showed that a population of channels in the cilia of olfactory cells responds directly to cAMP; that is, the channels are cAMP-gated. In Chapter 9, we will see that a similar version of cyclic nucleotide-gated channel is used for visual transduction. This is another demonstration that biology is conservative and evolution recycles its good ideas: Smelling and seeing use some very similar molecular mechanisms (Box 8.4).
The discovery of cyclic nucleotide-gated ion channels in olfactory receptor cells provides illuminating examples of how scientific orthodoxy can inhibit progress. Ironically, the story begins with work on vision. Studies of visual transduction began in earnest following the discovery, in 1971, that light caused the breakdown of cyclic guanosine monophosphate (cGMP) in photoreceptors. Yet, it was not until 1985 that the patch-damp method was used to demonstrate a direct effect of cGMP on ion channels from photoreceptors. This long delay was not due to lack of interest because there were at least a dozen labs working on the visual transduction mechanism. Rather, I think the widespread acceptance of protein phosphorylation as the mechanism of action of cyclic nucleotides in most cells effectively suppressed curiosity about other (direct) effects of cyclic nucleotides on ion channels. The cGMP-gated channel of photoreceptors was discovered by a group in the former Soviet Union, perhaps because these scientists were less influenced by the reigning dogma in Western countries.
The discovery of the olfactory cyclic nucleotide-gated channel by Tadashi Nakamura and me also emphasizes the importance of listening to the beat of your own drum. After the odorant-stimulated adenylyl cyclase was discovered in 1985, only a few months after the discovery of the photoreceptor cGMP-gated channel, we (and probably others) thought that olfactory cilia might contain a cyclic nucleotide-gated channel. This was because the biochemical similarities between visual and olfactory transduction suggested an evolutionary relationship between photoreceptors and olfactory receptor cells. Thus, we hypothesized that if the biochemical reactions of sensory transduction were conserved throughout evolution, ion channels might also be conserved. However, we knew that the transduction process was located in the cilia, and structures as small as cilia, which are about 0.2 μm in diameter, had never before been studied with the patch-damp technique. Indeed, most people I talked to thought it would be impossible to excise patches from the cilia. Nevertheless, we reasoned that it should be possible if only we could make patch pipettes with tip openings smaller than the ciliary diameter. This proved easy to accomplish; it only required fire polishing (melting) the tips of the patch pipettes slightly longer than was customary. Once we obtained high-resistance seals on cilia, patch excision and current recording were done in the conventional way.
Perhaps the most ironic thing about this story is that the photoreceptor channel was discovered by a group led by E. E. Fesenko, whose previous (and subsequent) work was on olfactory receptor proteins, whereas our work prior to the discovery of the olfactory channel was on phototransduction. That just goes to show how useful it can be for people to move into new areas. I like to point out that ours was a project that never would have been funded by the conventional grant review process because it was so unlikely to work.
How do the 1000 types of receptor cells used by mice discriminate among tens of thousands of odors? Like taste, olfaction involves a population-coding scheme. Each receptor protein binds different odorants more or less readily, so its receptor cell is more or less sensitive to those odorants (Figure 8.13b). Some cells are more sensitive to the chemical structure of the odorants they respond to than other cells are, but in general each receptor is quite broadly tuned. A corollary is that each odorant activates many of the 1000 types of receptors. The concentration of odorant is also important, and more odorant tends to generate stronger responses, until response strength saturates. Thus, each olfactory cell yields very ambiguous information about odorant type and strength. It is the job of the central olfactory pathways to respond to the full package of information arriving from the olfactory epithelium—the population code—and use it to classify odors further.
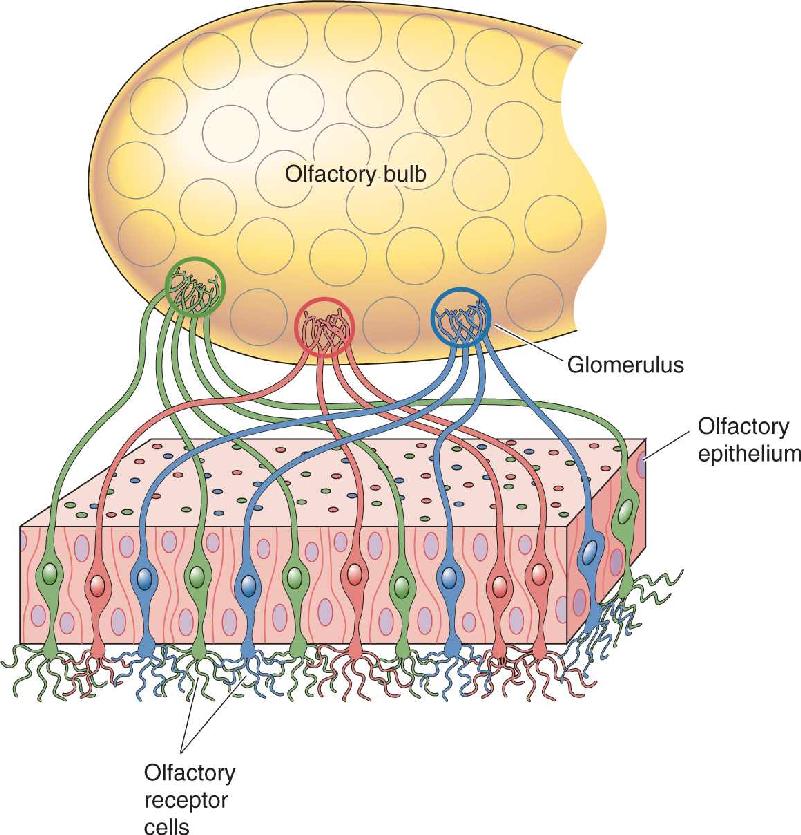
Olfactory receptor neurons send axons into the two olfactory bulbs (Figure 8.14). The bulbs are a neuroscientist’s wonderland, full of neural circuits with numerous types of neurons, fascinating dendritic arrangements, unusual reciprocal synapses, and high levels of many different neurotransmitters. The input layer of each bulb in mice contains about 2000 spherical structures called glomeruli, each about 50–200 μm in diameter. Within each glomerulus, the endings of about 25,000 primary olfactory axons (axons from the receptor cells) converge and terminate on the dendrites of about 100 second-order olfactory neurons.

FIGURE 8.14 The location and structure of an olfactory bulb. Axons of olfactory receptor cells penetrate the cribriform plate and enter the olfactory bulb. After multiple branching, each olfactory axon synapses upon second-order neurons within a spherical glomerulus. The second-order neurons send axons through the olfactory tract further into the brain.
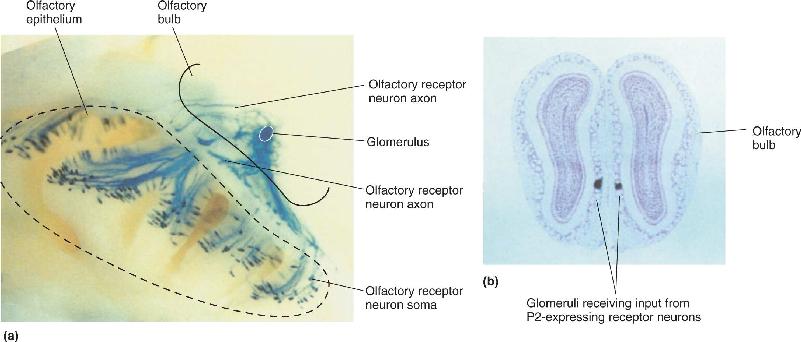
Recent studies revealed that the mapping of receptor cells onto glomeruli is astonishingly precise. Each glomerulus receives receptor axons from a large region of the olfactory epithelium. When molecular labeling methods are used to tag each receptor neuron expressing one particular receptor gene of the mouse—in this case, a gene called P2—we can see that the P2-labeled axons all converge onto only two glomeruli in each bulb, one of which is shown in Figure 8.15a. No axons seem to be out of place, but our knowledge of axonal pathfinding during development cannot yet explain the targeting accuracy of olfactory axons (see Chapter 23).

FIGURE 8.15 The convergence of olfactory neuron axons onto the olfactory bulb. Olfactory receptor neurons expressing a particular receptor gene all send their axons to the same glomeruli. (a) In a mouse, receptor neurons expressing the P2 receptor gene were labeled blue, and every neuron sent its axon to the same glomeruli in the olfactory bulbs. In this image, only a single glomerulus with P2 axons is visible. (b) When the two bulbs were cut in cross section, it was possible to see that the P2-containing receptor axons project to symmetrically placed glomeruli in each bulb. (Source: Adapted from Mombaerts et al., 1996, p. 680.) Description
This precision mapping is also consistent across the two olfactory bulbs; each bulb has only two P2-targeted glomeruli in symmetrical positions (Figure 8.15b). The positions of the P2 glomeruli within each bulb are consistent from one mouse to another. Finally, it seems that each glomerulus receives input from only receptor cells of one particular type. This means that the array of glomeruli within a bulb is a very orderly map of the receptor genes expressed in the olfactory epithelium (Figure 8.16), and, by implication, a map of odor information.

FIGURE 8.16 Specific mapping of olfactory receptor neurons onto glomeruli. Each glomerulus receives input only from receptor cells expressing a particular receptor protein gene. Receptor cells expressing a particular gene are color-coded.
Olfactory information is modified by inhibitory and excitatory interactions within and among the glomeruli and between the two bulbs. Neurons in the bulbs are also subject to modulation from systems of axons that descend from higher areas of the brain. While it is obvious that the elegant circuitry of the olfactory bulbs has important functions, it is not entirely clear what those functions are. It is likely that they begin to segregate odorant signals into broad categories, independent of their strength and possible interference from other odorants. The precise identification of an odor probably requires further processing in the next stages of the olfactory system.
Many brain structures receive olfactory connections. The output axons of the olfactory bulbs course through the olfactory tracts and project directly to several targets, some of which are illustrated in Figure 8.17. Among the most important targets are the primitive region of cerebral cortex called the olfactory cortex and some of its neighboring structures in the temporal lobes. This anatomy makes olfaction unique. All other sensory systems first pass information through the thalamus before projecting it to the cerebral cortex. The olfactory arrangement produces an unusually direct and widespread influence on the parts of the forebrain that have roles in odor discrimination, emotion, motivation, and certain kinds of memory (see Chapters 16, 18, 24 and 25). Conscious perceptions of smell may be mediated by a path from the olfactory tubercle, to the medial dorsal nucleus of the thalamus, and to the orbitofrontal cortex (situated right behind the eyes).

FIGURE 8.17 Central olfactory pathways. Axons of the olfactory tract branch and enter many regions of the forebrain, including the olfactory cortex. The neocortex is reached only by a pathway that synapses in the medial dorsal nucleus of the thalamus. Description
Spatial and Temporal Representations of Olfactory Information
In olfaction, there is an apparent paradox similar to the one in gustation. Individual receptors are broadly tuned to their stimuli; that is, each cell is sensitive to a wide variety of chemicals. However, when we smell those same chemicals, we can easily tell them apart. How is the whole brain doing what single olfactory cells cannot? We will discuss three important ideas: (1) Each odor is represented by the activity of a large population of neurons; (2) the neurons responsive to particular odors may be organized into spatial maps; and (3) the timing of action potentials may be an essential code for particular odors.
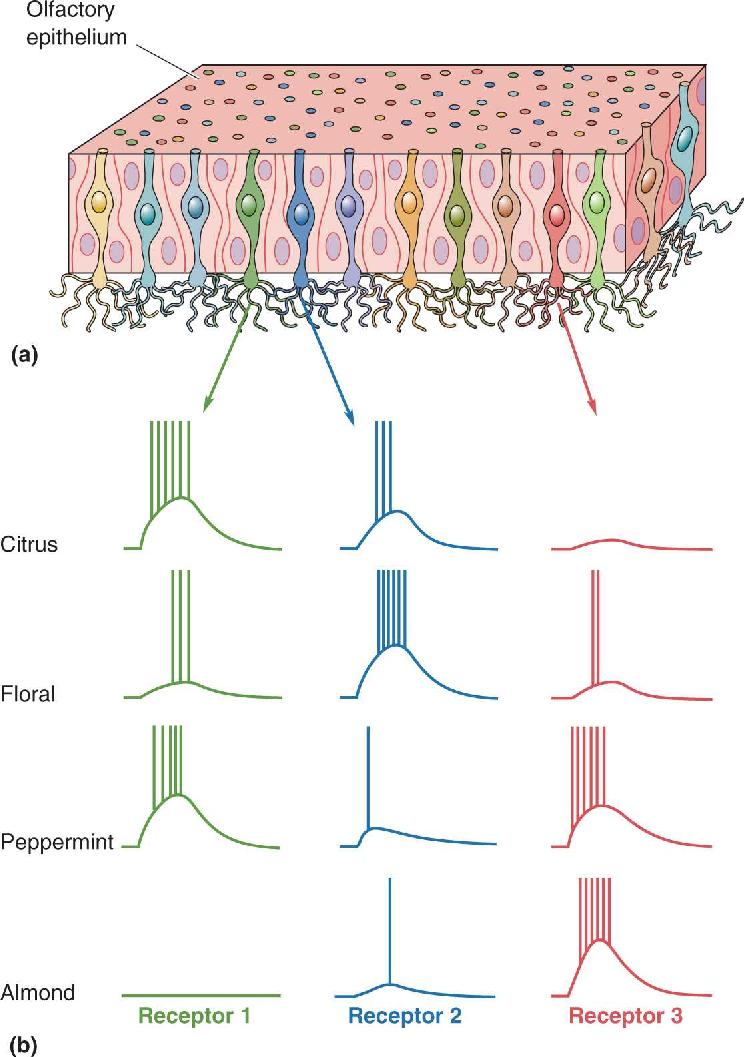
Olfactory Population Coding. As in gustation, the olfactory system uses the responses of a large population of receptors to encode a specific stimulus. A simplistic example was shown in Figure 8.13b. When presented with a citrus smell, none of the three different receptor cells can individually distinguish it clearly from the other odors. But by looking at the combination of responses from all three cells, the brain could distinguish the citrus smell unambiguously from floral, peppermint, and almond. By using such population coding, you can imagine how an olfactory system with 1000 different receptors might be able to recognize many different odors. In fact, by one recent estimate humans can discriminate at least one trillion different combinations of odor stimuli.
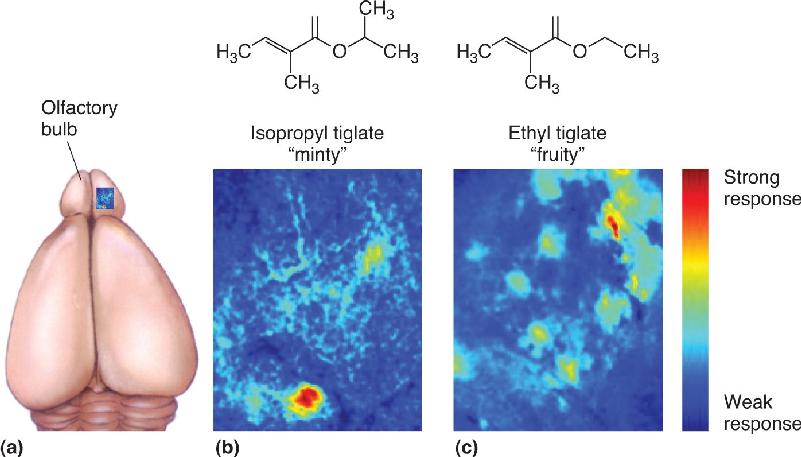
Olfactory Maps. A sensory map is an orderly arrangement of neurons that correlates with certain features of the environment. Microelectrode recordings show that many receptor neurons respond to the presentation of a single odorant and that these cells are distributed across a broad area of the olfactory epithelium (see Figure 8.13). This is consistent with the widespread distribution of each receptor gene. However, we have seen that the axons of each receptor cell type synapse upon particular glomeruli in the olfactory bulbs. Such an arrangement yields a sensory map in which neurons in a specific place in the bulb respond to particular odors. The maps of regions activated by one chemical stimulus can be visualized with special recording methods. Experiments reveal that while a particular odor activates many bulb neurons, the neurons’ positions form complex but reproducible spatial patterns. This is evident from the experiment shown in Figure 8.18, in which a minty-smelling chemical activates one pattern of glomeruli and a fruity scent activates quite a different pattern. Thus, the smell of a particular chemical is converted into a specific map defined by the positions of active neurons within the “neural space” of the bulbs, and the form of the map depends on the nature and concentration of the odorant.

FIGURE 8.18 Maps of neural activation of the olfactory bulb. The activity of neurons in the glomeruli of a mouse olfactory bulb was recorded with a specialized optical method. The cells expressed a fluorescent protein sensitive to intracellular Ca2+ levels, and neural activity was then signaled by changes in the amount of light emitted by the protein. The colors on the maps represent differing levels of neural activity; hotter colors (red and orange) imply more activity. Activated glomeruli show up as spots of color. (a) The blue box shows the area of one olfactory bulb that was mapped. Different olfactants evoked different spatial patterns of neural activation in the bulb: (b) isopropyl tiglate, which smells minty to humans, and (c) ethyl tiglate, which smells fruity, activate completely different patterns of glomeruli. (Source: Adapted from Blauvelt et al., 2013, Figure 4.) Description
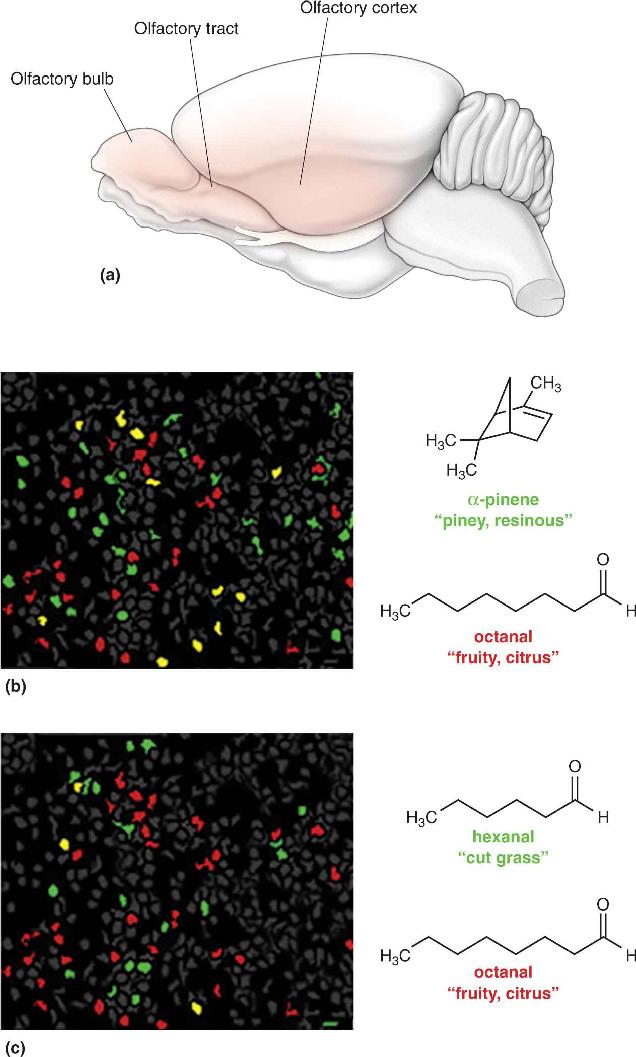
You will see in subsequent chapters that every sensory system uses spatial maps, perhaps for many different purposes. In most cases, the maps correspond obviously to features of the sensory world. For example, in the visual system, there are maps of visual space; in the auditory system, there are maps of sound frequency; and in the somatic sensory system, there are maps of the body surface. The maps of the chemical senses are unusual in that the stimuli themselves have no meaningful spatial properties. Although seeing a skunk walking in front of you may tell you what and where it is, smell by itself can reveal only the what. (By moving your head about, you can localize smells only crudely.) The most critical feature of each odorant is its chemical structure, not its position in space. Because the olfactory system does not have to map the spatial pattern of an odor in the same way that the visual system has to map the spatial patterns of light, neural odor maps may have other functions, such as discrimination among a huge number of different chemicals. Recent studies of the olfactory cortex show that each distinct odor triggers activity in a different subset of neurons. In the experiment shown in Figure 8.19, the orange-smelling octanal excites a group of neurons that are mostly different from the neurons excited by either the piney-scented α-pinene or the grassy-smelling hexanal (Figure 8.19).

FIGURE 8.19 Maps of neural activation of the olfactory cortex. The activity of many neurons in the mouse olfactory cortex was recorded with a specialized optical method. The cells were loaded with a Ca2+-sensitive fluorescent dye, and neural activity was then signaled by changes in the amount of light emitted. (a) The olfactory regions are shaded orange. (b) Neurons responsive to the piney scent of α-pinene are color-coded green, neurons responsive to the fruity scent of octanal are red, and cells responsive to both are yellow. (c) Neurons responsive to the grassy scent of hexanal are coded green, those responsive to the fruity scent of octanal are red, and cells responsive to both are yellow. Each of the three odorants activated a distinctly different pattern of cortical neurons. (Source: Adapted from Stettler and Axel, 2009, p. 858.) Description
But are neural odor maps actually used by the brain to distinguish among chemicals? We don’t know the answer. For a map to be useful, there must be something that reads and understands it. With practice and very specialized goggles, we might be able to read the “alphabet” of odors mapped on the surface of the olfactory bulb with our eyes. This may roughly approximate what higher regions of the olfactory system do, but so far there is no evidence that the olfactory cortex has this capability. An alternative idea is that spatial maps do not encode odors at all but are simply the most efficient way for the nervous system to form appropriate connections between related sets of neurons (e.g., receptor cells and glomerular cells). With orderly mapping, the lengths of axons and dendrites can be minimized. Neurons with similar functions can interconnect more easily if they are neighbors. The spatial map that results may be simply a side effect of these developmental requirements, rather than a fundamental mechanism of sensory coding itself.
Temporal Coding in the Olfactory System. There is growing evidence that the temporal patterns of spiking in olfactory neurons are essential features of olfactory coding. Compared to many sounds and sights, odors are inherently slow stimuli, so the rapid timing of action potentials is not necessary for encoding the timing of odors. Temporal coding, which depends on the timing of spikes, might instead encode the quality of odors. Hints about the possible importance of timing are easy to find. Researchers have known for many decades that the olfactory bulb and cortex generate oscillations of activity when odors are presented to the receptors, but the relevance of these rhythms is still unknown. Temporal patterns are also evident in the spatial odor maps, as they sometimes change shape during the presentation of a single odor.
Recent studies of insects and rodents have provided some of the most convincing evidence for temporal odor codes (Figure 8.20). Recordings from the olfactory systems of mice and insects demonstrate that odor information is encoded by the detailed timing of spikes within cells and between groups of cells as well as by the number, temporal pattern, rhythmicity, and cell-to-cell synchrony of spikes.

FIGURE 8.20 Spiking patterns may include changes of number, rate, and timing. As a mouse inspired (black trace at top) first air and then air with odorants, the activity of two neurons in the olfactory bulb was recorded. Spikes from the two cells are represented by red and blue lines. Both cells tended to spike twice during inspiration of air alone. Odor 1 did not change the numbers of spikes, but it did cause the red cell to spike much later in the respiratory cycle. Odor 2 increased the numbers of spikes without changing their timing much. Odor 3 increased spike numbers and delayed their onset. (Source: Dhawale et al., 2010, p. 1411.) Description
As with spatial maps, however, demonstrating that information is carried by spike timing is only a first step; proving that the brain actually uses that information is much more difficult. In a fascinating experiment with honeybees, Gilles Laurent and his colleagues at the California Institute of Technology were able to disrupt the rhythmic synchrony of odor responses without otherwise affecting their spiking responses. This loss of synchronous spiking was associated with a loss of the bees’ ability to discriminate between similar odors, although not between broad categories of odors. The implication is that the bee analyzes an odor not only by keeping track of which olfactory neurons fire but also by when they fire. It will be very interesting to learn whether similar processes occur in a mammalian olfactory system.
The chemical senses are a good place to begin learning about sensory systems because smell and taste are the most basic of sensations. Gustation and olfaction use a variety of transduction mechanisms to recognize the enormous number of chemicals we encounter in the environment. Yet, the molecular mechanisms of transduction are very similar to the signaling systems used in every cell of the body, for functions as diverse as neurotransmission and fertilization. We will see that the transduction mechanisms in other sensory systems are also highly specialized and derive from common cellular processes. Remarkable parallels have been discovered, such as the molecular similarity between the sensory cells of smelling and seeing.
Common sensory principles also extend to the level of neural systems. Most sensory cells are broadly tuned for their stimuli. This means that the nervous system must use population codes to represent and analyze sensory information, resulting in our remarkably precise and detailed perceptions. Populations of neurons are often arranged in sensory maps within the brain. And the timing of action potentials may function to represent sensory information in ways that are not yet understood. In the chapters that follow, we will see similar trends in the anatomy and physiology of systems dealing with light, sound, and pressure.
1. Most tastes involve some combination of the five basic tastes. What other sensory factors can help define the specific perceptions associated with a particular food?
2. The transduction of saltiness is accomplished, in part, by a Na+-permeable channel. Why would a sugar-permeable membrane channel be a poor mechanism for the transduction of sweetness?
3. Chemicals that have sweet, bitter, and umami tastes all activate precisely the same intracellular signaling molecules. Given this fact, can you explain how the nervous system can distinguish the tastes of sugars, alkaloids, and amino acids?
4. Why would the size of an animal’s olfactory epithelium (and consequently the number of receptor cells) be related to its olfactory acuity?
5. Receptor cells of the gustatory and olfactory systems undergo a constant cycle of growth, death, and maturation. Therefore, the connections they make with the brain must be continually renewed as well. Can you propose a set of mechanisms that would allow the connections to be remade in a specific way, again and again, over the course of an entire lifetime?
6. If the olfactory system does use some kind of spatial mapping to encode specific odors, how might the rest of the brain read the map?
Kinnamon SC. 2013. Neurosensory transmission without a synapse: new perspectives on taste signaling. BMC Biology 11:42.
Liberles SD. 2014. Mammalian pheromones. Annual Review of Physiology 76:151–175.
Liman ER, Zhang YV, Montell C. 2014. Peripheral coding of taste. Neuron 81: 984–1000.
Murthy VN. 2011. Olfactory maps in the brain. Annual Review of Neuroscience 34:233–258.
Stettler DD, Axel R. 2009. Representations of odor in the piriform cortex. Neuron 63: 854–864.
Zhang X, Firestein S. 2002. The olfactory receptor gene superfamily of the mouse. Nature Neuroscience 5:124–133.
Additional figures