The Eye
BOX 9.1 OF SPECIAL INTEREST: Demonstrating the Blind Regions of Your Eye
BOX 9.4 PATH OF DISCOVERY: Seeing Through the Photoreceptor Mosaic, by David Williams
Regional Differences in Retinal Structure and Their Visual Consequences
BOX 9.5 OF SPECIAL INTEREST: The Genetics of Color Vision
Vision is remarkable; it lets us detect things as tiny and close as a mosquito on the tip of our nose, or as immense and far away as a galaxy hundreds of thousands of light-years away. Sensitivity to light enables animals, including humans, to detect prey, predators, and mates. Based on the light bounced into our eyes from objects around us, we somehow make sense of a complex world. While this process seems effortless, it is in reality extremely complicated. Indeed, it has proven quite difficult to make computer visual systems with even a small fraction of the capabilities of the human visual system.
Light is electromagnetic energy that is emitted in the form of waves. We live in a turbulent sea of electromagnetic radiation. Like any ocean, this sea has large waves and small waves, short wavelets, and long rollers. The waves crash into objects and are absorbed, scattered, reflected, and bent. Because of the nature of electromagnetic waves and their interactions with the environment, the visual system can extract information about the world. This is a big job, and it requires a lot of neural machinery. However, the mastery of vision over the course of vertebrate evolution has had surprising rewards. It has provided new ways to communicate, given rise to brain mechanisms for predicting the trajectory of objects and events in time and space, allowed for new forms of mental imagery and abstraction, and led to the creation of a world of art. The significance of vision is perhaps best demonstrated by the fact that more than a third of the human cerebral cortex is involved with analyzing the visual world.
The mammalian visual system begins with the eye. At the back of the eye is the retina, which contains photoreceptors specialized to convert light energy into neural activity. The rest of the eye acts like a camera and forms crisp, clear images of the world on the retina. Like a camera, the eye automatically adjusts to differences in illumination and focuses on objects of interest. The eye can also track moving objects (by eye movement) and can keep its transparent surfaces clean (by tears and blinking).
While much of the eye functions like a camera, the retina does much more than passively register light levels across space. In fact, as mentioned in Chapter 7, the retina is actually part of the brain. (Think about that the next time you look deeply into someone’s eyes.) In a sense, each eye has two overlapping retinas: one specialized for low light levels that we encounter from dusk to dawn, and another specialized for higher light levels and for the detection of color from sunrise to sunset. Regardless of the time of day, however, the output of the retina is not a faithful reproduction of the intensity of the light falling on it. Rather, the retina is specialized to detect differences in the intensity of light falling on different parts of it. Image processing is well under way in the retina before any visual information reaches the rest of the brain.
Axons of retinal neurons are bundled into optic nerves, which distribute visual information (in the form of action potentials) to several brain structures that perform different functions. Some targets of the optic nerves are involved in regulating biological rhythms, which are synchronized with the light–dark daily cycle; others are involved in the control of eye position and optics. However, the first synaptic relay in the pathway that serves visual perception occurs in a cell group of the dorsal thalamus called the lateral geniculate nucleus, or LGN. From the LGN, visual information ascends to the cerebral cortex, where it is interpreted and remembered.
This chapter explores the eye and the retina. We’ll see how light carries information to our visual system, how the eye forms images on the retina, and how the retina converts light energy into neural signals that can be used to extract information about luminance and color differences. In Chapter 10, we will pick up the visual pathway at the back of the eye and take it through the thalamus to the cerebral cortex.
The visual system uses light to form images of the world around us. Let’s briefly review the physical properties of light and its interactions with the environment.
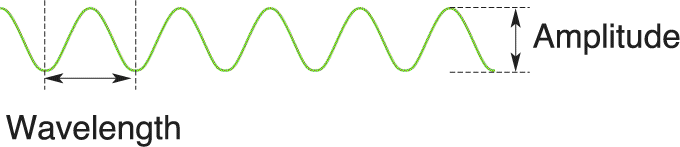
Electromagnetic radiation is all around us. It comes from innumerable sources, including radio antennas, mobile phones, X-ray machines, and the sun. Light is the electromagnetic radiation that is visible to our eyes. Electromagnetic radiation can be described as a wave of energy. Like any wave, electromagnetic radiation has a wavelength, the distance between successive peaks or troughs; a frequency, the number of waves per second; and an amplitude, the difference between wave trough and peak (Figure 9.1).

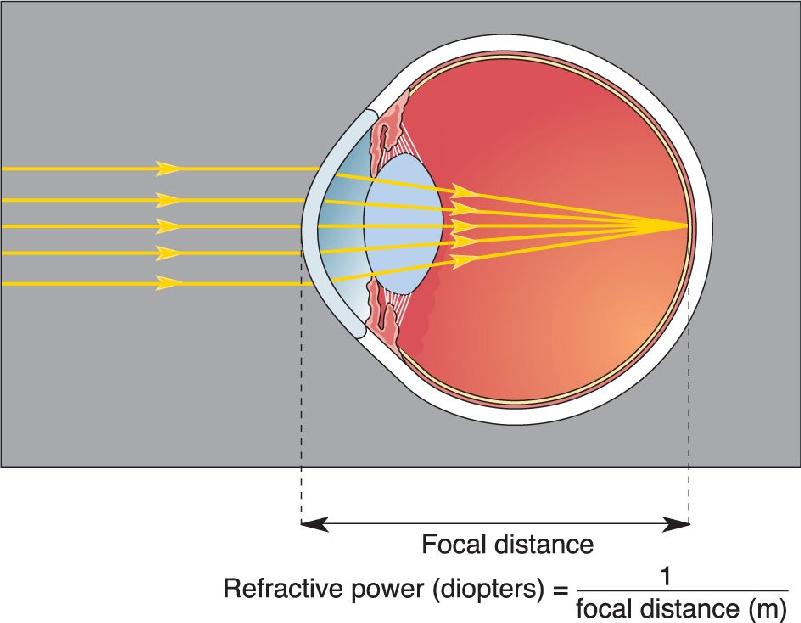
FIGURE 9.1 Characteristics of electromagnetic radiation. Description
The energy content of electromagnetic radiation is proportional to its frequency. Radiation emitted at a high frequency (short wavelengths) has the highest energy content; examples are gamma radiation emitted by some radioactive materials and X-rays used for medical imaging, with wavelengths less than 10–9 m (<1 nm). Conversely, radiation emitted at lower frequencies (longer wavelengths) has less energy; examples are radar and radio waves, with wavelengths greater than 1 mm. Only a small part of the electromagnetic spectrum is detectable by our visual system; visible light consists of wavelengths of 400–700 nm (Figure 9.2). As first shown by Isaac Newton early in the eighteenth century, the mix of wavelengths in this range emitted by the sun appears to humans as white, whereas light of a single wavelength appears as one of the colors of the rainbow. It is interesting to note that a “hot” color like red or orange consists of light with a longer wavelength, and hence has less energy, than a “cool” color like blue or violet. Clearly, colors are themselves “colored” by the brain, based on our subjective experiences.

FIGURE 9.2 The electromagnetic spectrum. Only electromagnetic radiation with wavelengths of 400–700 nm is visible to the human eye. Within this visible spectrum, different wavelengths appear as different colors. Description
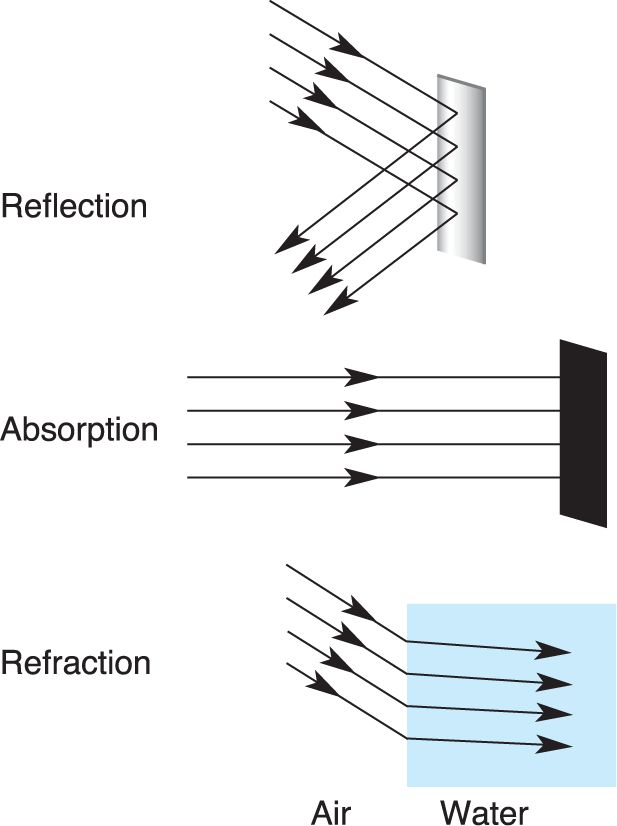
In a vacuum, a wave of electromagnetic radiation travels in a straight line and thus can be described as a ray. Light rays in our environment also travel in straight lines until they interact with the atoms and molecules of the atmosphere and objects on the ground. These interactions include reflection, absorption, and refraction (Figure 9.3). The study of light rays and their interactions is called optics.

FIGURE 9.3 Interactions between light and the environment. When light strikes an object in our environment, the light may be reflected, absorbed, or some combination of both. Visual perception is based on light coming directly into the eye from a luminous object such as a neon sign or being reflected off objects. Images are formed in the eye by refraction. In this example of light passing through air and then water, the light rays bend toward a line perpendicular to the air–water interface. Description
Reflection is the bouncing of light rays off a surface. The manner in which a ray of light is reflected depends on the angle at which it strikes the surface. A ray striking a mirror perpendicularly is reflected 180° back upon itself, a ray striking the mirror at a 45° angle is reflected 90°, and so on. Most of what we see is light that has been reflected off objects in our environment.
Absorption is the transfer of light energy to a particle or surface. You can feel this energy transfer on your skin on a sunny day, as visible light is absorbed and warms you up. Surfaces that appear black absorb the energy of all visible wavelengths. Some compounds absorb light energy only in a limited range of wavelengths and reflect the remaining wavelengths. This property is the basis for the colored pigments of paints. For example, a blue pigment absorbs long wavelengths but reflects a range of short wavelengths centered on 430 nm that are perceived as blue. As we will see in a moment, light-sensitive photoreceptor cells in the retina also contain pigments and use the energy absorbed from light to generate changes in membrane potential.
Images are formed in the eye by refraction, the bending of light rays that can occur when they travel from one transparent medium to another. When you dangle your leg into a swimming pool, for example, the odd way the leg appears to bend at the surface is a result of refraction. Consider a ray of light passing from the air into a pool of water. If the ray strikes the water surface perpendicularly, it will pass through in a straight line. However, if light strikes the surface at an angle, it will bend toward a line that is perpendicular to the surface. This bending of light occurs because the speed of light differs in the two media; light passes through air more rapidly than through water. The greater the difference between the speed of light in the two media, the greater the angle of refraction. The transparent media in the eye bend light rays to form images on the retina.
The eye is an organ specialized for the detection, localization, and analysis of light. Here we introduce the structure of this remarkable organ in terms of its gross anatomy, ophthalmoscopic appearance, and cross-sectional anatomy.
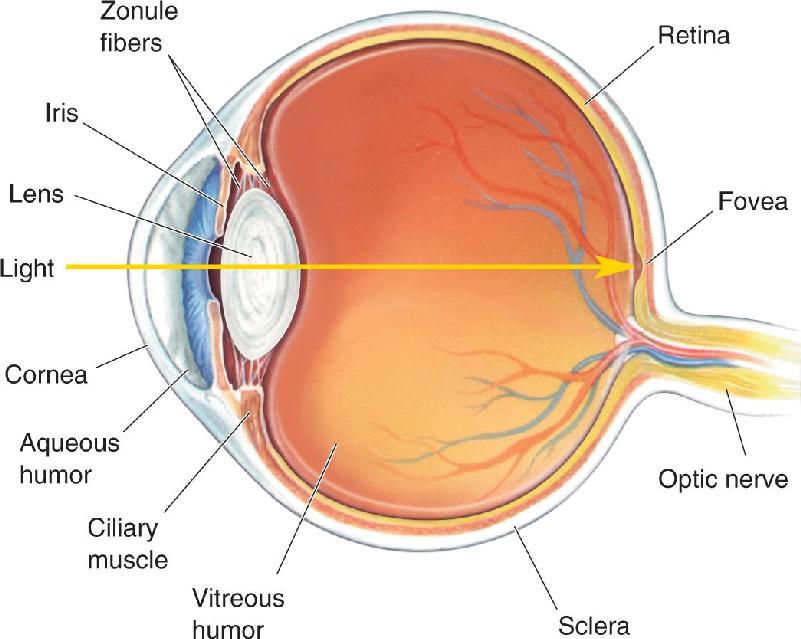
When you look into someone’s eyes, what are you really looking at? The main structures are shown in Figure 9.4. The pupil is the opening that allows light to enter the eye and reach the retina; it appears dark because of the light-absorbing pigments in the retina. The pupil is surrounded by the iris, whose pigmentation provides what we call the eye’s color. The iris contains two muscles that can vary the size of the pupil; one makes it smaller when it contracts, the other makes it larger. The pupil and iris are covered by the glassy transparent external surface of the eye, the cornea. The cornea is continuous with the sclera, the “white of the eye,” which forms the tough wall of the eyeball. The eyeball sits in a bony eye socket in the skull, also called the eye’s orbit. Inserted into the sclera are three pairs of extraocular muscles, which move the eyeball in the orbit. These muscles are normally not visible because they lie behind the conjunctiva, a membrane that folds back from the inside of the eyelids and attaches to the sclera. The optic nerve, carrying axons from the retina, exits the back of the eye, passes through the orbit, and reaches the base of the brain near the pituitary gland.

FIGURE 9.4 Gross anatomy of the human eye. Description
Another view of the eye is afforded by the ophthalmoscope, a device that enables one to peer into the eye through the pupil to the retina (Figure 9.5). The most obvious feature of the retina viewed through an ophthalmoscope is the blood vessels on its surface. These retinal vessels originate from a pale circular region called the optic disk, which is also where the optic nerve fibers exit the retina.

FIGURE 9.5 The retina, viewed through an ophthalmoscope. The dotted line through the fovea represents the demarcation between the side of the eye nearer the nose (nasal retina) and the side of the eye nearer the ear (temporal retina). The imaginary line crosses through the macula, which is in the center of the retina (it appears slightly to one side here because the photograph was taken to include the optic disk off to the nasal side of the retina).
It is interesting to note that the sensation of light cannot occur at the optic disk because there are no photoreceptors here, nor can it occur where the large blood vessels exist because the vessels cast shadows on the retina. And yet, our perception of the visual world appears seamless. We are not aware of any holes in our field of vision because the brain fills in our perception of these areas. However, there are tricks by which we can demonstrate the “blind” retinal regions (Box 9.1).
A look through an ophthalmoscope reveals that there is a sizable hole in the retina. The region where the optic nerve axons exit the eye and the retinal blood vessels enter the eye, the optic disk, is completely devoid of photoreceptors. Moreover, the blood vessels coursing across the retina are opaque and block light from falling on photoreceptors beneath them. Although we normally don’t notice them, these blind regions can be demonstrated. Look at Figure A. Hold the book about 1.5 feet away, close your right eye, and fixate on the cross with your left eye. Move the book (or your head) around slightly, and eventually you will find a position where the black circle disappears. At this position, the spot is imaged on the optic disk of the left eye. This region of visual space is called the blind spot for the left eye.

The blood vessels are a little tricky to demonstrate, but give this a try. Get a standard household flashlight. In a dark or dimly lit room, close your left eye (it helps to hold the eye closed with your finger so you can open your right eye fully). Look straight ahead with the open right eye, and shine the flashlight at an angle into the outer side of the right eye. Jiggle the light back and forth, up and down. If you’re lucky, you’ll see an image of your own retinal blood vessels. This is possible because the illumination of the eye at this oblique angle causes the retinal blood vessels to cast long shadows on the adjacent regions of retina. For the shadows to be visible, they must be swept back and forth on the retina, hence the jiggling of the light.
If we have all these light-insensitive regions in the retina, why does the visual world appear uninterrupted and seamless? The answer is that mechanisms in the visual cortex appear to “fill in” the missing regions. Perceptual filling-in can be demonstrated with the stimulus shown in Figure B. Fixate on the cross with your left eye and move the book closer and farther from your eye. You’ll find a distance at which you will see a continuous uninterrupted line. At this point, the space in the line is imaged on the blind spot, and your brain fills in the gap.
At the middle of each retina is a darker-colored region with a yellowish hue. This is the macula (from the Latin word for “spot”), the part of the retina for central vision (as opposed to peripheral vision). Besides its color, the macula is distinguished by the relative absence of large blood vessels. Notice in Figure 9.5 that the vessels arc from the optic disk to the macula; this is also the trajectory of the optic nerve fibers from the macula en route to the optic disk. The relative absence of large blood vessels in this region of the retina is one of the specializations that improves the quality of central vision. Another specialization of the central retina can sometimes be discerned with the ophthalmoscope: the fovea, a dark spot about 2 mm in diameter. The term is from the Latin for “pit,” and the retina is thinner in the fovea than elsewhere. Because it marks the center of the retina, the fovea is a convenient anatomical reference point. Thus, the part of the retina that lies closer to the nose than the fovea is referred to as nasal, the part that lies near the temple is called temporal, the part of the retina above the fovea is called superior, and the part below is inferior.
A cross-sectional view of the eye shows the path taken by light as it passes through the cornea toward the retina (Figure 9.6). The cornea lacks blood vessels and is nourished by the fluid behind it, the aqueous humor. This view reveals the transparent lens located behind the iris. The lens is suspended by ligaments (called zonule fibers) attached to the ciliary muscle, which forms a ring inside the eye. If you imagine using toothpicks to center a strawberry in the hole of a bagel, you have the right mental picture: The strawberry is the lens, the toothpicks represent the zonule fibers, and the bagel is the ciliary muscle that attaches to the sclera. As we shall see, changes in the shape of the lens enable our eyes to adjust their focus to different viewing distances.

FIGURE 9.6 The eye in cross section. Structures at the front of the eye regulate the amount of light allowed in and refract light onto the retina, which wraps around the inside of the eye.
The lens also divides the interior of the eye into two compartments containing slightly different fluids. The aqueous humor is the watery fluid that lies between the cornea and the lens. The more viscous, jellylike vitreous humor lies between the lens and the retina; it serves to keep the eyeball spherical.
Although the eyes do a remarkable job of delivering precise visual information to the rest of the brain, a variety of disorders can compromise this ability (Box 9.2).
Once you know the basic structure of the eye, you can understand how a partial or complete loss of vision results from abnormalities in various components. For example, if there is an imbalance in the extraocular muscles of the two eyes, the eyes will point in different directions. Such a misalignment or lack of coordination between the two eyes is called strabismus, and there are two varieties. In esotropia, the directions of gaze of the two eyes cross, and the person is said to be cross-eyed. In exotropia, the directions of gaze diverge, and the person is said to be wall-eyed (Figure A). In most cases, both types of strabismus are congenital; it can and should be corrected during early childhood. Treatment usually involves the use of prismatic glasses or surgery to the extraocular muscles to realign the eyes. Without treatment, conflicting images are sent to the brain from the two eyes, degrading depth perception and, more importantly, causing the person to suppress input from one eye. The dominant eye will be normal but the suppressed eye will become amblyopic, meaning that it has poor visual acuity. If medical intervention is delayed until adulthood, the condition cannot be corrected.

Figure A Exotropia. (Source: Newell, 1965, p. 330.)
A common eye disorder among older adults is cataract, a clouding of the lens (Figure B). Many people over 65 years of age have some degree of cataract; if it significantly impairs vision, surgery is usually required. In a cataract operation, the lens is removed and replaced with an artificial plastic lens. Although the artificial lens cannot adjust its focus as the normal lens does, it provides a clear image, and glasses can be used for near and far vision (see Box 9.3).

Figure B Cataract. (Source: Schwab, 1987, p. 22.)
Glaucoma, a progressive loss of vision associated with elevated intraocular pressure, is a leading cause of blindness. Pressure in the aqueous humor plays a crucial role in maintaining the shape of the eye. As this pressure increases, the entire eye is stressed, ultimately damaging the relatively weak point where the optic nerve leaves the eye. The optic nerve axons are compressed, and vision is gradually lost from the periphery inward. Unfortunately, by the time a person notices a loss of more central vision, the damage is advanced and a significant portion of the eye is permanently blind. For this reason, early detection and treatment with medication or surgery to reduce intraocular pressure are essential.
The light-sensitive retina at the back of the eye is the site of numerous disorders that pose a significant risk of blindness. You may have heard of a professional boxer having a detached retina. As the name implies, the retina pulls away from the underlying wall of the eye from a blow to the head or by shrinkage of the vitreous humor. Once the retina has started to detach, fluid from the vitreous space flows through small tears in the retina resulting from the trauma, thereby causing more of the retina to separate. Symptoms of retinal detachment include abnormal perception of shadows and flashes of light. Treatment often involves laser surgery to scar the edge of the retinal tear, thereby reattaching the retina to the back of the eye.
Retinitis pigmentosa is characterized by a progressive degeneration of the photoreceptors. The first sign is usually a loss of peripheral vision and night vision. Subsequently, total blindness may result. The cause of this disease is unknown. Some forms clearly have a strong genetic component, and more than 100 genes have been identified that can contain mutations leading to retinitis pigmentosa. There is currently no cure, but taking vitamin A may slow its progression.
In contrast to the tunnel vision typically experienced by patients with retinitis pigmentosa, people with macular degeneration lose only central vision. The condition is quite common, affecting more than 25% of all Americans over 65 years of age. While peripheral vision usually remains normal, the ability to read, watch television, and recognize faces is lost as central photoreceptors gradually deteriorate. Laser surgery can sometimes minimize further vision loss, but the disease currently has no known cure.
The eye collects the light rays emitted by or reflected off objects in the environment, and focuses them onto the retina to form images. Bringing objects into focus involves the combined refractive powers of the cornea and lens. You may be surprised to learn that the cornea, rather than the lens, is the site of most of the refractive power of the eyes. This occurs because light reaches the eye from air and the cornea is mostly made of water. A good deal of refraction occurs because light travels significantly slower in water than air. In comparison there is less refraction by the lens because the aqueous humor, lens, and vitreous humor are all composed largely of water.
Consider the light emitted from a distant source, perhaps a bright star at night. Light rays are emitted in all directions from the star, but because of the great distance, the rays that reach our eye on earth are virtually parallel. These parallel rays hit our cornea from one side to the other. Nonetheless, we see the star as a point of light, rather than a bright splotch filling our field of view, because the eye uses refraction to focus all the star’s light reaching the cornea into a small point on the retina.
Recall that as light passes into a medium where its speed is slowed, it will bend toward a line that is perpendicular to the border, or interface, between the media (see Figure 9.3). This is precisely the situation as light strikes the cornea and passes from the air into the aqueous humor. As shown in Figure 9.7, light that enters the eye perpendicular to the corneal surface passes straight to the retina, but light rays that strike the curved surface of the cornea at angles other than perpendicular are bent such that they converge on the back of the retina. The distance from the refractive surface to the point where parallel light rays converge is called the focal distance. Focal distance depends on the curvature of the cornea—the tighter the curve, the shorter the focal distance. The equation in Figure 9.7 shows that the reciprocal of the focal distance in meters is a unit of measurement called the diopter. The cornea has a refractive power of about 42 diopters, which means that parallel light rays striking the corneal surface will be focused 0.024 m (2.4 cm) behind it, about the distance from cornea to retina. To get a sense of the large amount of refraction produced by the cornea, note that many prescription eyeglasses have a power of only a few diopters.

FIGURE 9.7 Refraction by the cornea. The cornea must have sufficient refractive power, measured in diopters, to focus light on the retina at the back of the eye. Description
Remember that refractive power depends on the slowing of light at the air–cornea interface. If we replace air with a medium that passes light at about the same speed as the internal structures of the eye, the refractive power of the cornea would be eliminated. This is why things look blurry when you open your eyes underwater; the water–cornea interface has very little focusing power. Wearing swimmer’s goggles or a scuba mask restores the air–cornea interface and, consequently, the refractive power of the eye.
Although the cornea performs most of the eye’s refraction, the lens also contributes another dozen or so diopters to the formation of a sharp image of a distant point. However, the lens is involved more importantly in forming crisp images of objects located closer than about 9 m from the eye. With near objects, the light rays originating at a point are no longer parallel. Rather, these rays diverge from a light source or a point on an object, and greater refractive power is required to bring them into focus on the retina. This additional focusing power is provided by changing the shape of the lens, a process known as accommodation (Figure 9.8).

FIGURE 9.8 Accommodation by the lens. (a) To focus the eye on a distant point, relatively little refraction is required. The ciliary muscle relaxes, stretching the zonule fibers and flattening the lens. (b) Near objects require greater refraction provided by a more spherical lens. This is achieved by contracting the ciliary muscle so there is less tension in the zonule fibers.
Recall that the ciliary muscle forms a ring around the lens. During accommodation, the ciliary muscle contracts and swells in size, thereby making the area inside the ring smaller (i.e., a smaller hole inside the bagel in our analogy) and decreasing the tension in the suspensory ligaments. Consequently, the lens becomes rounder and thicker because of its natural elasticity. This rounding increases the curvature of the lens surfaces, thereby increasing their refractive power. Conversely, relaxation of the ciliary muscle increases the tension in the suspensory ligaments, and the lens is stretched into a flatter shape.
The ability to accommodate changes with age. An infant’s eyes can focus on objects just beyond his or her nose, whereas many middle-aged adults cannot clearly see objects closer than about arm’s length. Fortunately, artificial lenses can compensate for this and other defects of the eye’s optics (Box 9.3).
When the ciliary muscles are relaxed and the lens is flat, the eye is said to be emmetropic if parallel light rays from a distant point source are focused sharply on the back of the retina. (The word is from the Greek emmetros, meaning “in proper measure,” and ope, “sight.”) Stated another way, the emmetropic eye focuses parallel light rays on the retina without the need for accommodation (Figure A), and accommodation is sufficient to focus the images of objects over a wide range of distances.

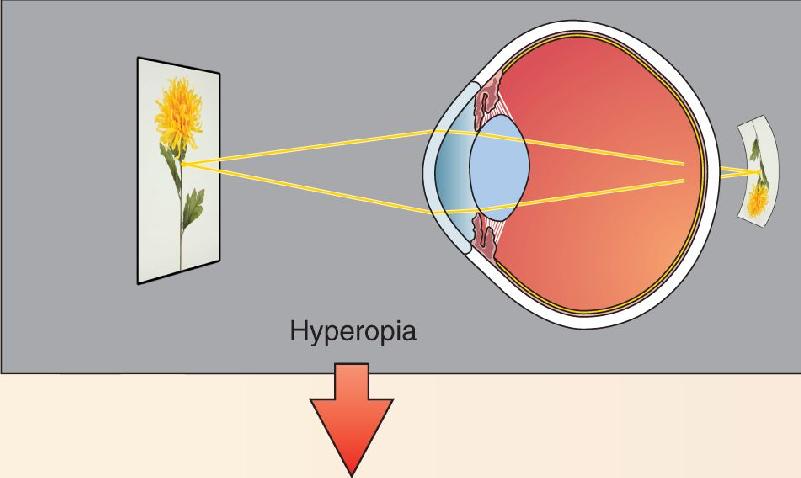
Now consider what happens when the eyeball is too short from front to back. Without accommodation, parallel light rays are focused at a point behind the eye. This condition is known as hyperopia, or farsightedness. The eye has enough accommodation to achieve good focus for distant objects, but even with the maximum amount of accommodation, near objects are focused at a point behind the retina (Figure B). Farsightedness can be corrected by placing a convex glass or plastic lens in front of the eye (Figure C). The curved front edge of the lens, like the cornea, bends light toward the center of the retina. Also, as the light passes from glass into air as it exits the lens, the back of the lens also increases the refraction (light going from glass to air speeds up and is bent away from the perpendicular).
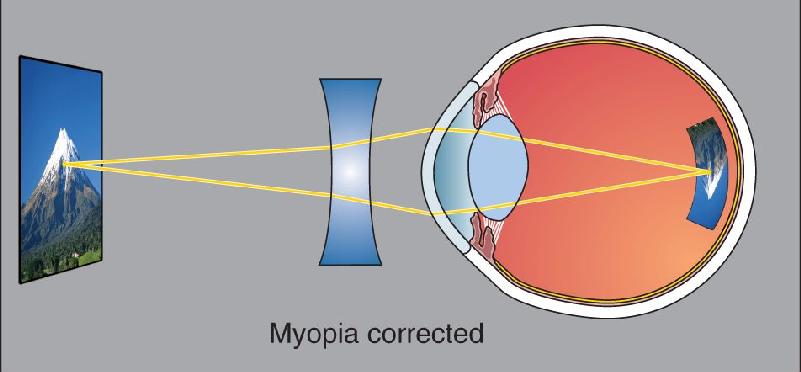
If the eyeball is too long rather than too short, parallel rays will converge before the retina, cross, and again be imaged on the retina as a blurry circle. This condition is known as myopia, or nearsightedness. There is more than enough refraction to image close objects, but even with the least amount of accommodation, distant objects are focused in front of the retina (Figure D). For the nearsighted eye to see distant objects clearly, artificial concave lenses must be used to move the image back onto the retina (Figure E).
Some eyes have irregularities such that the curvature and refraction in the horizontal and vertical planes is different. This condition is called astigmatism, and it can be corrected with an artificial lens that is curved more along one axis than others.
Even if you are fortunate enough to have perfectly shaped eyeballs and a symmetrical refractive system, you probably will not escape presbyopia (from the Greek meaning “old eye”). This condition is a hardening of the lens that accompanies the aging process and is thought to be explained by the fact that while new lens cells are generated throughout life, none are lost. The hardened lens is less elastic, leaving it unable to change shape and accommodate sufficiently to focus on both near and far objects. The correction for presbyopia, first introduced by Benjamin Franklin, is a bifocal lens. These lenses are concave on top to assist far vision and convex on the bottom to assist near vision.
In hyperopia and myopia, the amount of refraction provided by the cornea is either too little or too great for the length of the eyeball. But modern techniques can now change the amount of refraction the cornea provides. In radial keratotomy, a procedure to correct myopia, tiny incisions through the peripheral portion of the cornea relax and flatten the central cornea, thus reducing the amount of refraction and minimizing the myopia. The most recent techniques use lasers to reshape the cornea. In photorefractive keratectomy (PRK), a laser is used to reshape the outer surface of the cornea by vaporizing thin layers. Laser in situ keratomileusis (LASIK) has become very common; you’ve probably seen clinics offering this procedure in shopping areas or malls. In LASIK, a microkeratome or laser is used to make a thin flap in the outer cornea. With the cornea flap temporarily lifted, the laser can reshape the cornea from the inside (Figure F). Nonsurgical methods are also being used to reshape the cornea. A person can be fitted with special retainer contact lenses or plastic corneal rings that alter the shape of the cornea and correct refractive errors.
In addition to the cornea and the lens, the pupil contributes to the optical functioning of the eye by continuously adjusting for different ambient light levels. To check this for yourself, stand in front of a bathroom mirror with the lights out for a few seconds, and then watch your pupils shrink when you turn the lights on. This pupillary light reflex involves connections between the retina and neurons in the brain stem that control the muscles that constrict the pupils. An interesting property of this reflex is that it is consensual; shining a light into only one eye causes the constriction of the pupils of both eyes. It is unusual, indeed, when the pupils are not the same size; the lack of a consensual pupillary light reflex is often taken as a sign of a serious neurological disorder involving the brain stem.
A beneficial effect of the pupil constriction that results from an increase in light level is an increase in the depth of focus (the range of distances from the eye that appear in focus), just like decreasing the aperture size (increasing the f-stop) of a camera lens. To understand why this is true, consider two points in space, one close and the other far away. When the eye accommodates to the closer point, the image of the farther point on the retina no longer forms a point but rather a blurred circle. Decreasing the aperture—constricting the pupil—reduces the size of this blurred circle so that its image more closely approximates a point. In this way, distant objects appear to be less out of focus.
The structure of the eyes, and where they are positioned in the head, limits how much of the world we can see at any one time. Let’s investigate the extent of the space seen by one eye. Holding a pencil vertically in your right hand, close your left eye and look at a point straight ahead. Keeping your eye fixated on this point, slowly move the pencil to the right (toward your right ear) across your field of view until the pencil disappears. Repeat this exercise, moving the pencil to the left where it will disappear behind your nose, and then up and down. The points where you can no longer see the pencil mark the limits of the visual field for your right eye. Now look at the middle of the pencil as you hold it horizontally in front of you. Figure 9.9 shows how the light reflected off this pencil falls on your retina. Notice that the image is inverted; the left visual field is imaged on the right side of the retina, and the right visual field is imaged on the left side of the retina. Similarly, the upper visual field is imaged on the bottom of the retina and the lower visual field is imaged on the upper retina.

FIGURE 9.9 The visual field for one eye. The visual field is the total amount of space that can be viewed by the retina when the eye is fixated straight ahead. Notice how the image of an object in the visual field (pencil) is left–right reversed on the retina. The visual field extends nearly 100° to the temporal side but only about 60° to the nasal side of the retina, where vision is blocked by the nose.
The ability of the eye to distinguish two points near each other is called visual acuity. Acuity depends on several factors but especially on the spacing of photoreceptors in the retina and the precision of the eye’s refraction.
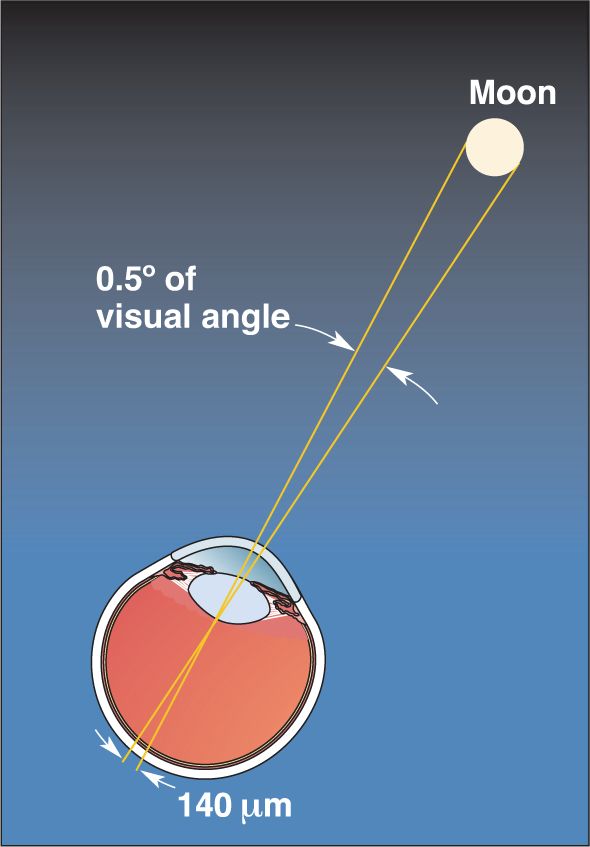
Distance across the retina can be described in terms of degrees of visual angle. A right angle subtends (spans) 90°, and the moon, for example, subtends an angle of about 0.5° (Figure 9.10). At arm’s length, your thumb is about 1.5° across and your fist about 10°. We can speak of the eye’s ability to resolve points that are separated by a certain number of degrees of visual angle. The Snellen eye chart, which we have all read at the doctor’s office, tests our ability to discriminate letters and numbers at a viewing distance of 20 feet. Your vision is 20/20 when you can recognize a letter that subtends an angle of 0.083° (equivalent to 5 minutes of arc, where 1 minute is 1/60 of a degree).

FIGURE 9.10 Visual angle. Distances across the retina can be expressed as degrees of visual angle.
Now that we have an image formed on the retina, we can get to the neuroscience of vision: the conversion of light energy into neural activity. To begin our discussion of image processing in the retina, we must introduce the cellular architecture of this bit of brain.
The basic system of retinal information processing is shown in Figure 9.11. The most direct pathway for visual information to exit the eye is from photoreceptors to bipolar cells to ganglion cells. The photoreceptors respond to light, and they influence the membrane potential of the bipolar cells connected to them. The ganglion cells fire action potentials in response to light, and these impulses propagate along the optic nerve to the rest of the brain. Besides the cells in this direct path from photoreceptor to brain, retinal processing is influenced by two additional cell types. Horizontal cells receive input from the photoreceptors and project neurites laterally to influence surrounding bipolar cells and photoreceptors. A wide variety of types of amacrine cells generally receive input from bipolar cells and project laterally to influence surrounding ganglion cells, bipolar cells, and other amacrine cells.

FIGURE 9.11 The basic system of retinal information processing. Information about light flows from the photoreceptors to bipolar cells to ganglion cells, which project axons out of the eye in the optic nerve. Horizontal cells and amacrine cells modify the responses of bipolar cells and ganglion cells via lateral connections. Description
There are three important points to remember here:
- With one exception, the only light-sensitive cells in the retina are the rod and cone photoreceptors. All other cells are influenced by light only via direct and indirect synaptic interactions with the photoreceptors. (We will see in a moment that recently discovered types of ganglion cells are also light sensitive, but these uncommon cells do not appear to play a major role in visual perception.)
- The ganglion cells are the only source of output from the retina. No other retinal cell type projects an axon through the optic nerve.
- With the exception of certain amacrine cells, ganglion cells are the only retinal neurons that fire action potentials; this is essential for transmitting information outside the eye. All other retinal cells depolarize or hyperpolarize, with a rate of neurotransmitter release that is proportional to the membrane potential, but they do not fire action potentials.
Now let’s take a look at how the different cell types are arranged in the retina.
Figure 9.12 shows that the retina has a laminar organization: Cells are organized in layers. Notice that the layers are seemingly inside-out; light must pass from the vitreous humor through the ganglion cells and bipolar cells before it reaches the photoreceptors. Because the retinal cells above the photoreceptors are relatively transparent, image distortion is minimal as light passes through them. One reason the inside-out arrangement is advantageous is that the pigmented epithelium that lies below the photoreceptors plays a critical role in the maintenance of the photoreceptors and photopigments. The pigmented epithelium also absorbs any light that passes entirely through the retina, thus minimizing the scattering of light within the eye that would blur the image. Many nocturnal animals, such as cats and raccoons, have a reflective layer beneath the photoreceptors, called the tapetum lucidum, which bounces light back at the photoreceptors if it passes through the retina. The animal is thus more sensitive to low light levels at the expense of reduced acuity. An interesting side effect of the reflective tapetum can be seen when you shine a light at or take a flash photograph of nocturnal animals: There is striking “eyeshine” in which the pupils seem to glow (Figure 9.13).

FIGURE 9.12 The laminar organization of the retina. Notice how light must pass through several cell layers before it reaches the photoreceptors at the back of the retina. Description

FIGURE 9.13 Eyeshine results from the reflective tapetum in cats.
The cell layers of the retina are named in reference to the middle of the eyeball. Don’t get confused by thinking about the head instead of the eye: The photoreceptors are the outermost part of the retina even though they are the farthest from the front of the eye and the deepest inside the head. The innermost retinal layer is the ganglion cell layer, which contains the cell bodies of the ganglion cells. Moving outward, there are two other layers that contain the cell bodies of neurons: the inner nuclear layer, which contains the cell bodies of the bipolar cells, horizontal cells, and amacrine cells, and the outer nuclear layer, which contains the cell bodies of the photoreceptors.
Between the ganglion cell layer and the inner nuclear layer is the inner plexiform layer (“plexiform” means a network of connections), which contains the synaptic contacts between bipolar cells, amacrine cells, and ganglion cells. Between the outer and inner nuclear layers is the outer plexiform layer, where the photoreceptors make synaptic contact with the bipolar and horizontal cells. Finally, the layer of photoreceptor outer segments contains the light-sensitive elements of the retina. The outer segments are embedded in the pigmented epithelium.
The conversion of electromagnetic radiation into neural signals occurs in the photoreceptors at the back of the retina. Every photoreceptor has four regions: an outer segment, an inner segment, a cell body, and a synaptic terminal. The outer segment contains a stack of membranous disks. Light-sensitive photopigments in the disk membranes absorb light, thereby triggering changes in the photoreceptor membrane potential (discussed later). Figure 9.14 shows the two types of photoreceptors in the retina, easily distinguished by the appearance of their outer segments. Rod photoreceptors have a long, cylindrical outer segment, containing many disks. Cone photoreceptors have a shorter, tapering outer segment with fewer membranous disks. The greater number of disks and higher photopigment concentration in rods makes them over 1000 times more sensitive to light than cones. It is estimated that there are about 5 million cones and 92 million rods in each human retina.

FIGURE 9.14 Rods and cones. (a) Rods contain more disks and make vision possible in low light; cones enable us to see in daylight. (b) Scanning electron micrograph of rods and cones. (Source: Courtesy of J. Franks and W. Halfter.) Description
The large differences in the structure and sensitivity of rods and cones led investigators to say humans have a duplex retina: essentially two complementary systems in one eye. Some animals have only rods or only cones and thus do not have a duplex retina. The structural differences between rods and cones correlate with important functional differences. For example, in nighttime lighting, or scotopic conditions, only rods contribute to vision. Conversely, in daytime lighting, or photopic conditions, cones do the bulk of the work. At intermediate light levels (indoor lighting or outdoor traffic lighting at night), or mesopic conditions, both rods and cones are responsible for vision.
Rods and cones differ in other respects as well. All rods contain the same photopigment, but there are three types of cones, each containing a different pigment. The variations among pigments make the different cones sensitive to different wavelengths of light. As we shall see in a moment, only the cones, not the rods, are responsible for our ability to see color. David Williams at the University of Rochester has used clever imaging techniques to reveal the distribution of human cones in exquisite detail. Surprisingly, rather than an arrangement like the neat arrangement of pixels in a computer display, human retinas show striking diversity in the arrangement and distribution of cone photoreceptors (Box 9.4).
When I started graduate school in 1975, we knew almost nothing about the topography of the three classes of cones that form the basis of trichromatic color vision in the human eye. Although Thomas Young had deduced 175 years earlier that color vision depended on three fundamental channels, we did not know the relative numbers of the three kinds of cones or how they were arranged in the retina. With my graduate advisor, Don MacLeod at the University of California, San Diego, I used psychophysical methods to map retinal sensitivity to violet light. We found that the S cones, which are stimulated by this light, are sparsely sprinkled across a mosaic of L and M cones. We also discovered that a person can reliably detect a flash of light that stimulates just one of the 5 million or so cones in the retina.
Later, I kept returning to the topography of the trichromatic mosaic. After several failed attempts over many years, I hit upon a solution from a completely unexpected direction. I’d had a long-standing interest in the limits of visual acuity. While exploring various technologies to prevent the normal optical blurring of the retinal image, I came upon adaptive optics, in which astronomers use a deformable mirror to correct for the blur caused by the turbulent atmosphere when imaging stars with telescopes on earth.
A major obstacle to using adaptive optics in vision science was that such telescope mirrors cost about $1 million. Fortunately, we found an engineer who made us an affordable deformable mirror. We were also fortunate that the military’s work with adaptive optics had been recently declassified, so my post-doc, Junzhong Liang, and I were allowed to visit the Starfire Optical Range (SOR), a $16 million satellite-tracking telescope equipped with adaptive optics. I was discouraged by the legions of engineers and expensive optical system needed to run the facility—but then something remarkable happened. Bob Fugate, director of the SOR, was trying to measure atmospheric aberrations by reflecting light from a high-power laser off a mirror left on the moon’s surface in the Apollo program. I heard Bob say, “Move that beam over to the right, you missed the whole damn moon!” Suddenly I realized that they were learning from fumbling just the same as in my lab—so there was hope after all.
Liang and I hurried back to the University of Rochester and with another post-doc, Don Miller, built the first adaptive optic system that could correct all of the eye’s monochromatic aberrations. This started a minor revolution in optometry and ophthalmology because now many more of the eye’s optical defects could be corrected than had been possible before. A person’s visual acuity using an adaptive optics system can be better than with the most carefully prescribed spectacles. This led to a better way to correct vision with laser refractive surgery as well as improved designs for contact lenses and intraocular lenses.
We also equipped a camera with adaptive optics to take the sharpest pictures ever of the living human retina, so sharp that the individual cones in the photoreceptor mosaic can be seen. Could we also use adaptive optics to identify which of the three photopigments resides in each cone imaged in the living eye, solving the problem I’d started with in graduate school? Using adaptive optics and another technique called retinal densitometry, two post-docs in my lab, Austin Roorda and later Heidi Hofer, finally answered the question definitively. It turns out the three classes of cones are remarkably disorganized (Figure A), unlike the highly regular mosaics of many insect eyes. Moreover, the relative numbers of the M and L cones vary enormously from one person to another, despite the similarity of color vision in people (Figure B). Joe Carroll, another former postdoc, has gone on to reveal the mosaic organization in colorblind eyes and those with many different genetic mutations.
Figure A The arrangement of the three cone classes in the human eye. (Source: Roorda and Williams, 1999.)

Figure B Variation in the relative numbers of cones in eyes with normal color vision. (Source: Hofer et al., 2005; Roorda and Williams, 1999.)
Adaptive optics is also being used to image many other cells in the retina, including ganglion cells, and is a valuable tool in the diagnosis and treatment of retinal disease. I certainly could never have foreseen that advances in astronomic technology would provide these tools for vision research—or that my graduate school interest in the trichromacy of the cone mosaic would 20 years later spawn these advances in vision correction and single cell imaging.
Regional Differences in Retinal Structure and Their Visual Consequences
Retinal structure varies from the fovea to the retinal periphery. Most of the 5 million cones are in the fovea, and the proportion diminishes substantially in the retinal periphery. There are no rods in the central fovea, but there are many more rods than cones in the peripheral retina. Rod and cone distributions in the retina are summarized in Figure 9.15.

FIGURE 9.15 Regional differences in retinal structure. (a) Cones are found primarily in the central retina, within 10° of the fovea. Rods are absent from the central fovea and are found mainly in the peripheral retina. (b) In the central retina, relatively few photoreceptors feed information to a ganglion cell; in the peripheral retina, many photoreceptors provide input. This arrangement makes the peripheral retina better at detecting dim light but the central retina better for high-resolution vision. (c) This magnified cross section of the human central retina shows the dense packing of cone inner segments. (d) At a more peripheral location on the retina, the cone inner segments are larger and appear as islands in a sea of smaller rod inner segments. A 10-micron scale bar for the images in c and d is shown to the right in d. (Source for parts c and d: Curcio et al., 1990, p. 500.) Description
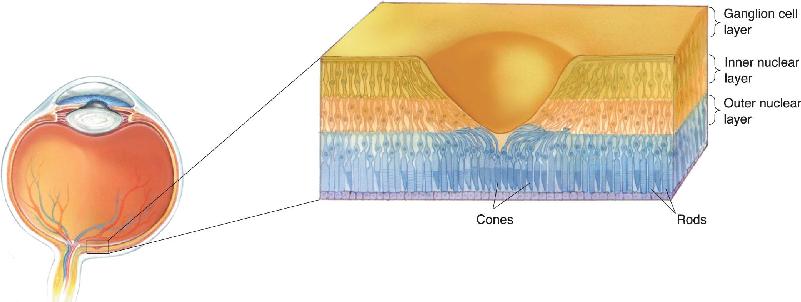
The differences in rod and cone numbers and distribution across the retina have important visual consequences. At photopic light levels (daylight), the most obvious is that we have much greater spatial sensitivity on our central retina. Visual acuity is measured when we look directly at symbols on a test chart, placing the critical features in our cone-rich fovea. Recall that the fovea is a thinning of the retina at the center of the macula. In cross section, the fovea appears as a pit in the retina. This appearance is due to the lateral displacement of the cells above the photoreceptors, allowing light to strike the photoreceptors without passing through the other retinal cell layers (Figure 9.16). This structural specialization maximizes visual acuity at the fovea by pushing aside other cells that might scatter light and blur the image. If you were to take an eye test while looking slightly away from the test chart, or if you try to read the titles of books on a shelf using your peripheral vision, you would need the letters to be much larger for you to read them. Less obvious than the fovea’s high spatial acuity is that we are poorer at discriminating colors on our peripheral retina because of the smaller number of cones. You might be able to demonstrate this by looking straight ahead and moving a small colored object slowly to the side.

FIGURE 9.16 The fovea in cross section. The ganglion cell layer and the inner nuclear layer are displaced laterally to allow light to strike the foveal photoreceptors directly.
The consequences of rod and cone distribution differences are very different with the light at dim scotopic levels, when we see only with rods. For example, we are more sensitive to low levels of light on our peripheral retina. Put another way, our central vision is blind at scotopic light levels. This is because rods respond more strongly to low light levels than cones, there are more rods in the peripheral retina (and none in the central fovea), and more rods project to single bipolar and ganglion cells in the peripheral retina (thereby aiding the detection of dim light). You can demonstrate the greater sensitivity of your peripheral retina to yourself on a starry night. (It’s fun; try it with a friend.) First, spend about 20 minutes in the dark getting oriented, and then gaze at a bright star. Fixating on this star, search your peripheral vision for a dim star. Then move your eyes to look at this dim star. You will find that the faint star disappears when it is imaged on the central retina (when you look straight at it) but reappears when it is imaged on the peripheral retina (when you look slightly to the side of it).
Because cones alone make the perception of color possible, we are unable to perceive color differences at night when rods are active but cones are not. A green tree, a blue car, and a red house all appear to have vaguely the same color (or lack of color). The peak sensitivity of rods is to a wavelength of about 500 nm, and thus at scotopic light levels objects tend to look dark blue-green. The loss of color as the sun goes down is a huge perceptual effect, but one that we hardly notice because of its familiarity.
Nighttime vision for modern humans is not due exclusively to rods, however. In densely populated areas, we actually can perceive some color at night because streetlights and neon signs emit sufficient light to activate the cones. This fact is the basis for differing points of view about the design of automobile dashboard indicator lights. One view is that the lights should be dim blue-green to take advantage of the spectral sensitivity of the rods. An alternate view is that the lights should be bright red because this wavelength affects mainly cones, leaving the rods unsaturated and thus resulting in better night vision.
The photoreceptors convert, or transduce, light energy into changes in membrane potential. We begin our discussion of phototransduction with rods, which outnumber cones in the human retina by about 20 to 1. Most of what has been learned about phototransduction by rods has proven to be applicable to cones as well.
As we discussed in Part I of this book, one way information is represented in the nervous system is as changes in the membrane potential of neurons. Thus, we look for a mechanism by which the absorption of light energy can be transduced into a change in the photoreceptor membrane potential. In many respects, this process is analogous to the transduction of chemical signals into electrical signals that occurs during synaptic transmission. At a G-protein-coupled neurotransmitter receptor, the binding of transmitter to the receptor activates G-proteins in the membrane, which in turn stimulate various effector enzymes (Figure 9.17a). These enzymes alter the intracellular concentration of cytoplasmic second messenger molecules, which (directly or indirectly) change the conductance of membrane ion channels, thereby altering membrane potential. Similarly, in the photoreceptor, light stimulation of the photopigment activates G-proteins, which in turn activate an effector enzyme that changes the cytoplasmic concentration of a second messenger molecule. This change causes a membrane ion channel to close, and the membrane potential is thereby altered (Figure 9.17b).

FIGURE 9.17 Light transduction and G-proteins. G-protein-coupled receptors and photoreceptors use similar mechanisms. (a) At a G-protein-coupled receptor, the binding of neurotransmitter activates G-proteins and effector enzymes. (b) In a photoreceptor, light begins a similar process using the G-protein transducin. Description
Recall from Chapter 3 that a typical neuron at rest has a membrane potential of about –65 mV, close to the equilibrium potential for K+. In contrast, in complete darkness, the membrane potential of the rod outer segment is about –30 mV. This depolarization is caused by the steady influx of Na+ through special channels in the outer segment membrane (Figure 9.18a). The movement of positive charge across the membrane, which occurs in the dark, is called the dark current. Sodium channels are stimulated to open—are gated—by an intracellular second messenger called cyclic guanosine monophosphate, or cGMP. cGMP is produced in the photoreceptor by the enzyme guanylyl cyclase, keeping the Na+ channels open. Light reduces cGMP, causing the Na+ channels to close, and the membrane potential becomes more negative (Figure 9.18b). Thus, photoreceptors hyperpolarize in response to light (Figure 9.18c).

FIGURE 9.18 The hyperpolarization of photoreceptors in response to light. Photoreceptors are continuously depolarized in the dark because of an inward sodium current, the dark current. (a) Sodium enters the photoreceptor through a cGMP-gated channel. (b) Light leads to the activation of an enzyme that destroys cGMP, thereby shutting off the Na+ current and hyperpolarizing the cell. (c) In a dark auditorium, the membrane potential of our photoreceptors is –30 mV (left). At intermission, we move to a bright lobby and the cells hyperpolarize (middle). The slower depolarization that follows is adaptation. Returning to the dark room sends membrane potential back to –30 mV (right). Description
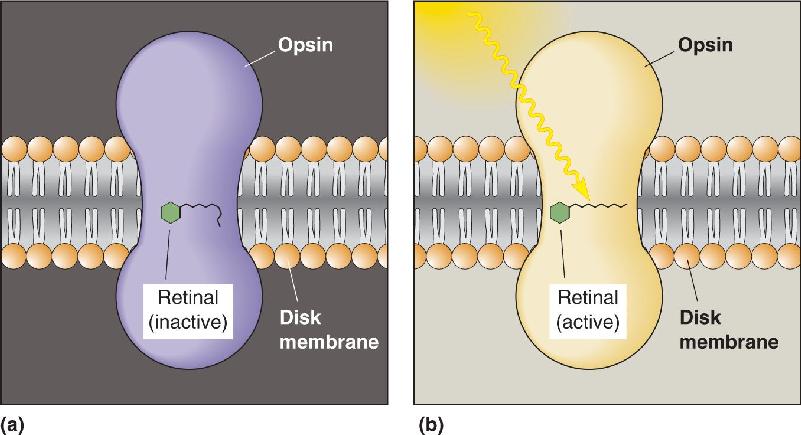
The hyperpolarizing response to light is initiated by the absorption of electromagnetic radiation by the photopigment in the membrane of the stacked disks in the rod outer segments. In the rods, this pigment is called rhodopsin. Rhodopsin can be thought of as a receptor protein with a prebound chemical agonist. The receptor protein is called opsin, and it has the seven transmembrane alpha helices typical of G-protein-coupled receptors throughout the body. The prebound agonist is called retinal, a derivative of vitamin A. The absorption of light causes a change in the conformation of retinal so that it activates the opsin (Figure 9.19). This process is known as bleaching because it changes the wavelengths absorbed by the rhodopsin (the photopigment literally changes color from purple to yellow). The bleaching of rhodopsin stimulates a G-protein called transducin in the disk membrane, which in turn activates the effector enzyme phosphodiesterase (PDE), which breaks down the cGMP that is normally present in the cytoplasm of the rod (in the dark). The reduction in cGMP causes the Na+ channels to close and the membrane to hyperpolarize.

FIGURE 9.19 The activation of rhodopsin by light. Rhodopsin consists of opsin, a protein with seven transmembrane alpha helices, and retinal, a small molecule derived from vitamin A. (a) In the dark, retinal is inactive. (b) Retinal undergoes a change in conformation when it absorbs light, thereby activating (bleaching) the opsin. Description
One of the interesting functional consequences of using a biochemical cascade for transduction is signal amplification. Many G-proteins are activated by each photopigment molecule, and each PDE enzyme breaks down more than one cGMP molecule. This amplification gives our visual system incredible sensitivity to small amounts of light. Rods are more sensitive to light than cones because they contain more disks in their outer segments and thus more photopigment, and also because they amplify the response to light more than cones do. The combined result is, incredibly, that rods give a measureable response to the capture of a single photon of light, the elementary unit of light energy.
To summarize, here are the steps in the transduction of light by rods:
- Phosphodiesterase (PDE), the effector enzyme, is activated.
- Na+ channels close, and the cell membrane hyperpolarizes.
The complete sequence of events of phototransduction in rods is illustrated in Figure 9.20.

FIGURE 9.20 The light-activated biochemical cascade in a photoreceptor. (a) In the dark, cGMP gates a sodium channel, causing an inward Na+ current and depolarization of the cell. (b) The activation of rhodopsin by light energy causes the G-protein (transducin) to exchange guanosine diphosphate for guanosine triphosphate (see Chapter 6), which in turn activates the enzyme phosphodiesterase (PDE). PDE breaks down cGMP and shuts off the dark current. Description
In bright light, cGMP levels in rods fall to the point where the response to light becomes saturated; increasing the light level causes no additional hyperpolarization. Thus, vision during the day depends entirely on the cones, whose photopigments require more energy to become bleached.
The process of phototransduction in cones is virtually the same as in rods; the only major difference is in the type of opsins in the membranous disks of the cone outer segments. The cones in our retinas contain one of three opsins that give the photopigments different spectral sensitivities. Thus, we can speak of short-wavelength or “blue” cones that are maximally activated by light with a wavelength of about 430 nm, medium-wavelength or “green” cones that are maximally activated by light with a wavelength of about 530 nm, and long-wavelength or “red” cones that are maximally activated by light with a wavelength of about 560 nm (Figure 9.21). Note that each type of cone is activated by a broad range of different wavelengths of light, and there is overlap in the wavelengths that affect the three cone types. Commonly the cones are referred to as blue, green, and red, but this can be confusing because different colors are perceived when different wavelengths of light, within the broad sensitive range of a particular cone type, are presented. The short, medium, long terminology is safer.

FIGURE 9.21 The spectral sensitivity of the three types of cone pigments. Each photopigment absorbs a wide range of wavelengths from the color spectrum (see Figure 9.2). Description
Color Perception. The color that we perceive is largely determined by the relative contributions of short-, medium-, and long-wavelength cones to the retinal signal. The fact that our visual system perceives colors in this way was actually predicted over 200 years ago by British physicist Thomas Young. Young showed in 1802 that all the colors of the rainbow, including white, could be created by mixing the proper ratio of red, green, and blue light (Figure 9.22). Young proposed, quite correctly, that the retina contains three receptor types, each type being maximally sensitive to a different spectrum of wavelengths. Young’s ideas were later championed by Hermann von Helmholtz, an influential nineteenth-century German physiologist. (Among his accomplishments is the invention of the ophthalmoscope in 1851.) This theory of color vision came to be known as the Young–Helmholtz trichromacy theory. According to the theory, the brain assigns colors based on a comparative readout of the three cone types. When all types of cones are equally active, as in broad-spectrum light, we perceive “white.” Novel colors arise from other mixtures. For example, orange is a mixture of red and yellow, and it looks somewhat red and somewhat yellow (red, orange, and yellow are neighbors in the color spectrum). But note that some color mixtures are perceptually different: No color looks simultaneously red and green or blue and yellow (and these “opponent” colors are not neighbors in the color spectrum). As we will see later, this may be a reflection of further “color opponent” processing by ganglion cells.

FIGURE 9.22 Mixing colored lights. The mixing of red, green, and blue light causes equal activation of the three types of cones, and the perception of “white” results.
The color vision nomenclature can be puzzling, so be careful to not confuse the color of light with the “color name” of a cone photoreceptor. It is wrong to think that lights perceived as red consist of a single wavelength of light or that this wavelength is absorbed only by long-wavelength cones. The reality is that colored lights generally contain a broad and complex spectrum of wavelengths that may partially activate all three types of cones. The ratios of activations determine color. Various forms of color blindness result when one or more of the cone photopigment types is missing (Box 9.5). And, as discussed earlier, if we had no cones, we would be unable to perceive color differences at all.
The color we perceive is largely determined by the relative amounts of light absorbed by the red, green, and blue visual pigments in our cones. This means it’s possible to perceive any color of the rainbow by mixing different amounts of red, green, and blue light. For example, the perception of yellow light comes from an appropriate mixture of red and green light. Because we use a “three-color” system, humans are referred to as trichromats. However, not all normal trichromats perceive colors exactly the same. For example, if a group of people are asked to choose the wavelength of light that most appears green without being yellowish or bluish, there will be small variations in their choices. However, significant abnormalities of color vision extend well beyond this range of normal trichromatic vision.
Most abnormalities in color vision result from small genetic errors that lead to the loss of one visual pigment or a shift in the spectral sensitivity of one type of pigment. The most common abnormalities involve red–green color vision and are much more common in men than in women. The reason for this pattern is that the genes encoding the red and green pigments are on the X chromosome, whereas the gene that encodes the blue pigment is on chromosome 7. Men have abnormal red–green vision if there is a defect on the single X chromosome they inherit from their mother. Women have abnormal red–green vision only if both parents contribute abnormal X chromosomes.
About 6% of men have a red or green pigment that absorbs somewhat different wavelengths of light than do the pigments of the rest of the population. These men are often called “colorblind,” but they actually see quite a colorful world. They are more properly referred to as anomalous trichromats because they require somewhat different mixtures of red, green, and blue to see intermediate colors (and white) than other people. Most anomalous trichromats have normal genes to encode the blue pigment and either the red or the green pigment, but they also have a hybrid gene that encodes a protein with an abnormal absorption spectrum between that of normal red and green pigments. For example, a person with an anomalous green pigment can match a yellow light with a red–green mixture containing less red than a normal trichromat. Anomalous trichromats perceive the full spectrum of colors that normal trichromats perceive, but in rare instances they disagree about the precise color of an object (e.g., blue versus greenish blue).
About 2% of men actually lack either the red or the green pigment, making them red–green colorblind. Because this leaves them with a “two-color” system, they are referred to as dichromats. People lacking the green pigment are less sensitive to green, and they confuse certain red and green colors that appear different to trichromats. A “green dichromat” can match a yellow light with either red or green light; no mixture is needed. In contrast to the roughly 8% of men who either are missing one pigment or have an anomalous pigment, only about 1% of women have such color abnormalities.
People without one color pigment are considered colorblind, but they actually do see colors. Estimates of the number of people lacking all color vision vary, but less than about 0.001% of the population is thought to have this condition. In one type, both red and green cone pigments are missing, in many cases because mutations of the red and green genes make them nonfunctional. These people are called blue cone monochromats and live in a world that varies only in lightness, like a trichromat’s perception of a black-and-white movie.
Although achromatopsia (lack of color vision) is rare in humans, on the tiny Micronesian island of Pingelap more than 5–10% of the population is colorblind and many more are unaffected carriers. It is known that the underlying basis for the disorder is a genetic mutation associated with incomplete development of cones that leaves them nonfunctional. But why is achromatopsia so common on Pingelap? According to islanders, in the late eighteenth century a typhoon killed all but about 20 of the inhabitants. Those afflicted with achromatopsia appear all to be descendants of one man who was a carrier; in subsequent generations the incidence of achromatopsia grew with inbreeding in the small surviving population.
Recent research has shown that, precisely speaking, there may not be such a thing as normal color vision. In a group of males classified as normal trichromats, it was found that some require slightly more red than others to perceive yellow in a red–green mixture. This difference, which is tiny compared to the deficits discussed above, results from a single alteration of the red pigment gene. The 60% of males who have the amino acid serine at site 180 in the red pigment gene are more sensitive to long-wavelength light than the 40% who have the amino acid alanine at this site. Imagine what would happen if a woman had different red gene varieties on her two X chromosomes. Both red genes should be expressed, leading to different red pigments in two populations of cones. In principle, such women should have a super-normal ability to discriminate colors because of their tetrachromatic color vision, a rarity among all animals.
The transition from all-cone daytime vision to all-rod nighttime vision is not instantaneous; depending on how high the initial light level is, it can take minutes to nearly an hour to reach the greatest light sensitivity in the dark (hence the time needed to get oriented in the stargazing exercise described earlier). This phenomenon is called dark adaptation, or getting used to the dark. Sensitivity to light actually increases a millionfold or more during this period.
Dark adaptation is explained by a number of factors. Perhaps the most obvious is dilation of the pupils, which allows more light to enter the eye. However, the diameter of the human pupil only ranges from about 2–8 mm; changes in its size increase sensitivity to light by a factor of only about 10. The larger component of dark adaptation involves the regeneration of unbleached rhodopsin and an adjustment of the functional circuitry of the retina so that information from more rods is available to each ganglion cell. Because of this tremendous increase in sensitivity, when the dark-adapted eye goes back into bright light, it is temporarily saturated. This explains what happens when you first go outside on a bright day. Over the next 5–10 minutes, the eyes undergo light adaptation, reversing the changes in the retina that accompanied dark adaptation. This light-dark adaptation in the duplex retina gives our visual system the ability to operate in light intensities ranging from moonless midnight to bright high noon.
Calcium’s Role in Light Adaptation. In addition to the factors discussed earlier, the ability of the eye to adapt to changes in light level relies on changes in calcium concentration within the cones. When you step out into bright light from a dark theater, initially the cones are hyperpolarized as much as possible (i.e., to EK, the equilibrium potential for K+). If the cones stayed in this state, we would be unable to see further changes in light level. As discussed earlier, the constriction of the pupil helps a bit in reducing the light entering the eye. However, the most important change is a gradual depolarization of the membrane back to about –35 mV (see Figure 9.18c).
The reason this happens stems from the fact that the cGMP-gated sodium channels discussed previously also admit calcium (Figure 9.23). In the dark, Ca2+ enters the cones and has an inhibitory effect on the enzyme (guanylyl cyclase) that synthesizes cGMP. When the cGMP-gated channels close, the flow of Ca2+ into the photoreceptor is curtailed along with the flow of Na+; as a result, more cGMP is synthesized (because the synthetic enzyme is less inhibited), thereby allowing the cGMP-gated channels to open again. Stated more simply, when the channels close, a process is initiated that gradually reopens them even if the light level does not change. Calcium also appears to affect photopigments and phosphodiesterase in ways that decrease their response to light. These calcium-based mechanisms ensure that the photoreceptors are always able to register relative changes in light level, though information about the absolute level is lost.

FIGURE 9.23 The role of calcium in light adaptation. Ca2+ enters a cone through the same cGMP-gated channels as Na+; it inhibits the synthesis of cGMP. Description
Local Adaptation of Dark, Light, and Color. The effect that pupil size has on light and dark adaptation is the same for all photoreceptors. However, photoreceptor bleaching and other adaptational mechanisms, such as calcium’s influence on cGMP, can occur on a cone-by-cone basis. You can demonstrate this to yourself with Figure 9.24. First, fixate on the black cross at the center of the gray square in part a for about a minute. Cones imaging the dark spots will become dark-adapted and cones imaging the white spots will be relatively light-adapted. Now look at the cross at the middle of the large light square in part b. Because of the local adaptation of the cones, you should now see white spots where you adapted to black and black spots where you adapted to white. The same idea applies to color adaptation. First look at the yellow or green square in part c or d of the figure, and selectively adapt your cones. Then move your fixation to the light square in part b. You should see blue if you adapted to yellow and red if you adapted to green (the exact colors depend on the inks used in printing). These demonstrations use abnormally long fixations to reveal the critical adaptational processes that are always at work to keep photoreceptors providing useful information.

FIGURE 9.24 Light and color adaptation. Fixate on the cross in (a) for 1 minute and then look at (b) to see the effects of local retinal adaptation to the light and dark spots. After adaptation to the color in (c) or (d), the gray in (b) will appear to have an opponent color. Description
Now that we have seen how light is first converted into neural activity, we turn to the process by which information about light is passed beyond the eye to the rest of the brain. As the sole source of output from the retina is the action potentials arising from ganglion cells, our goal is to understand what information ganglion cells convey. It is interesting to note that researchers were able to explain some of the ways the retina processes visual images well before the discovery of how photoreceptors work. Since about 1950, neuroscientists have studied the action potential discharges of retinal ganglion cells as the retina is stimulated with light. The pioneers of this approach were neurophysiologists Keffer Hartline and Stephen Kuffler working in the United States and Horace Barlow working in England. Their research uncovered which aspects of a visual image were encoded as ganglion cell output. Early studies of horseshoe crabs and frogs gave way to investigations of cats and monkeys. Researchers learned that similar principles are involved in retinal processing across a wide range of species.
Progress in understanding how ganglion cell properties are generated by synaptic interactions in the retina has been slower. This is so because only ganglion cells fire action potentials; all other cells in the retina (except some amacrine cells) respond to stimulation with graded changes in membrane potential. The detection of such graded changes requires technically challenging intracellular recording methods, whereas action potentials can be detected using simple extracellular recording methods (see Box 4.1). It was not until the early 1970s that John Dowling and Frank Werblin at Harvard University were able to show how ganglion cell responses are built from the interactions of horizontal and bipolar cells.
The most direct path for information flow in the retina is from a cone photoreceptor to bipolar cell to ganglion cell. At each synaptic relay, the responses are modified by the lateral connections of horizontal cells and amacrine cells. Photoreceptors, like other neurons, release neurotransmitter when depolarized. The transmitter released by photoreceptors is the amino acid glutamate. As we have seen, photoreceptors are depolarized in the dark and are hyperpolarized by light. We thus have the counterintuitive situation in which photoreceptors actually release fewer transmitter molecules in the light than in the dark. However, we can reconcile this apparent paradox if we take the point of view that dark rather than light is the preferred stimulus for a photoreceptor. Thus, when a shadow passes across a photoreceptor, it responds by depolarizing and releasing more neurotransmitter.
In the outer plexiform layer, each photoreceptor is in synaptic contact with two types of retinal neurons: bipolar cells and horizontal cells. Recall that bipolar cells create the direct pathway from photoreceptors to ganglion cells; horizontal cells feed information laterally in the outer plexiform layer to influence the activity of neighboring bipolar cells and photoreceptors (see Figures 9.11 and 9.12). We now turn to the response properties of bipolar and then ganglion cells by analyzing their receptive fields.
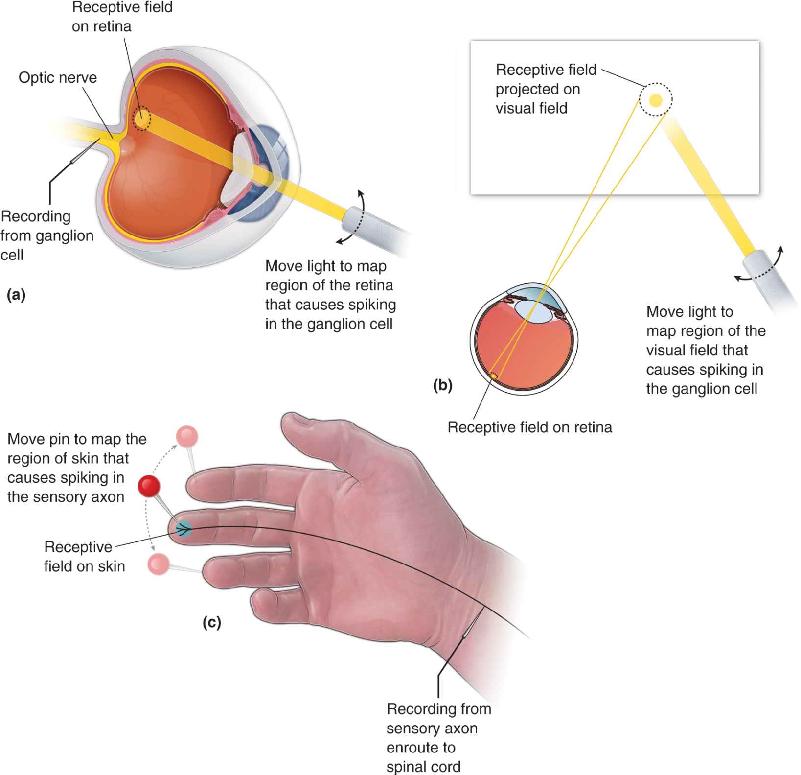
Suppose you have a flashlight that can project a very small spot of light onto the retina while you monitor the activity of a visual neuron, such as the output of a retinal ganglion cell. You would find that light applied to only a small portion of the retina would change the firing rate of the neuron (Figure 9.25a). This area of the retina is called the neuron’s receptive field. Light anywhere else on the retina, outside this receptive field, would have no effect on firing rate. This same procedure can be applied to any neuron in the eye or elsewhere that is involved in vision; its receptive field is specified by the pattern of light on the retina that elicits a neural response. In the visual system, the optics of the eye establish a correspondence between locations on the retina and the visual field. Therefore, it is customary to also describe visual receptive fields as areas of visual space interchangeably with areas of the retina (Figure 9.25b). “Receptive field” is actually a general term useful in describing the stimulus specificity of neurons across sensory systems. For example, we will see in Chapter 12 that receptive fields of neurons in the somatosensory system are small areas of the skin that, when touched, produce a response in a neuron (Figure 9.25c).

FIGURE 9.25 The receptive field. (a) The receptive field of a ganglion cell is located by recording from the neuron’s axon in the optic nerve. A small spot of light is projected onto various parts of the retina; the receptive field consists of the locations that increase or decrease the ganglion cell’s firing rate. By moving the recording electrode, the same procedure can be used to locate receptive fields for neurons at other locations in the brain and other retinal neurons (seen as changes in membrane potential if they do not fire action potentials). (b) A receptive field on the retina corresponds to light coming from a particular location in the visual field. (c) The receptive field is a useful concept in other sensory systems; for example, a small patch of skin is a receptive field for touch. Description
As we move farther into the visual pathway, we find that receptive fields change in shape and correspondingly in the sort of stimulus that makes the neurons most active. In the retina, spots of light give optimal responses from ganglion cells, but in different areas of the visual cortex, neurons respond best to bright lines and even complex shapes of biological significance such as hands and faces. These changes may reflect important differences in the kind of information represented at each stage. (We will have a lot more to say about this in Chapter 10.) Receptive fields receive a great deal of attention because of the functional interpretation given to them. An instructive example comes from the early work of Horace Barlow, who made recordings from the frog retina. He found that a frog will jump and snap at a small black spot waved in front of it, and the same stimulus elicits a strong response from retinal ganglion cells. Might this be a “bug detector” used by the frog to hunt? We will see that inferences about function based on receptive field properties are also made for monkeys and humans, and they are as exciting as they are speculative.
Bipolar cells and receptive fields can be categorized into two classes, ON and OFF, based on the response to the glutamate released by photoreceptors. The circuitry that gives rise to bipolar receptive fields consists of direct input from photoreceptors and indirect photoreceptor input relayed by horizontal cells (Figure 9.26a). Let’s start by considering only the interactions between cones and bipolar cells in the direct path (no horizontal cells involved), shown in Figure 9.26b. Light shined onto a cone will hyperpolarize some bipolar cells. These are called OFF bipolar cells, because light effectively turns them off. Conversely, light shined onto a cone will depolarize other bipolar cells. These cells “turned on” by light are called ON bipolar cells. Evidently the cone-to-bipolar synapse inverts the signal from the cone: The cone hyperpolarizes to light, but the ON bipolar cell depolarizes.

FIGURE 9.26 Direct and indirect pathways from photoreceptors to bipolar cells. (a) Bipolar cells receive direct synaptic input from a cluster of photoreceptors, constituting the receptive field center. In addition, they receive indirect input from surrounding photoreceptors via horizontal cells. (b) An ON-center bipolar cell is depolarized by light in the receptive field center via the direct pathway. (c) Light in the receptive field surround hyperpolarizes the ON-center bipolar cell via the indirect pathway. Because of the intervening horizontal cell, the effect of light on the surround photoreceptors is always opposite the effect of light on the center photoreceptors. Description
How can different bipolar cells give opposite responses to direct cone input? The answer is that there are two kinds of receptors that receive glutamate released by the photoreceptors. OFF bipolar cells have ionotropic glutamate receptors, and these glutamate-gated channels mediate a classical depolarizing excitatory postsynaptic potential from the influx of Na+. Hyperpolarization of the cone causes less neurotransmitter to be released, resulting in a more hyperpolarized bipolar cell. On the other hand, ON bipolar cells have G-protein-coupled (metabotropic) receptors and respond to glutamate by hyperpolarizing. Each bipolar cell receives direct synaptic input from a cluster of photoreceptors. The number of photoreceptors in this cluster ranges from one at the center of the fovea to thousands in the peripheral retina.
In addition to direct connections with photoreceptors, bipolar cells also are connected via horizontal cells to a circumscribed ring of photoreceptors that surrounds the central cluster (see Figure 9.26a). The synaptic interactions of photoreceptors, horizontal cells, and bipolar cells are complex, and research is ongoing. For our purposes here, there are two key points. First, when a photoreceptor hyperpolarizes in response to light, output is sent to horizontal cells that also hyperpolarize. Second, the effect of horizontal cell hyperpolarization is to counteract the effect of light on neighboring photoreceptors. In Figure 9.26c, light is shined onto two photoreceptors connected through horizontal cells to a central photoreceptor and bipolar cell. The effect of this indirect path input is to depolarize the central photoreceptor, counteracting the hyperpolarizing effect of light shined directly on it.
Let’s synthesize this discussion: The receptive field of a bipolar cell consists of two parts: a circular area of retina providing direct photoreceptor input, the receptive field center, and a surrounding area of retina providing input via horizontal cells, the receptive field surround. The response of a bipolar cell’s membrane potential to light in the receptive field center is opposite to that of light in the surround. Thus, these cells are said to have antagonistic center-surround receptive fields. Receptive field dimensions can be measured in millimeters across the retina or, more commonly, in degrees of visual angle. One millimeter on the retina corresponds to a visual angle of about 3.5°. Bipolar cell receptive field diameters range from a fraction of a degree in the central retina to several degrees in the peripheral retina.
The center-surround receptive field organization is passed on from bipolar cells to ganglion cells via synapses in the inner plexiform layer. The lateral connections of the amacrine cells in the inner plexiform layer also contribute to the elaboration of ganglion cell receptive fields and the integration of rod and cone input to ganglion cells. Numerous types of amacrine cells have been identified, which make diverse contributions to ganglion cell responses.
Most retinal ganglion cells have essentially the same concentric center-surround receptive field organization as bipolar cells. ON-center and OFF-center ganglion cells receive input from the corresponding type of bipolar cell. An important difference is that, unlike bipolar cells, ganglion cells fire action potentials. Ganglion cells actually fire action potentials whether or not they are exposed to light, and light in the receptive field center or surround increases or decreases the firing rate. Thus, an ON-center ganglion cell will be depolarized and respond with a barrage of action potentials when a small spot of light is projected onto the center of its receptive field. An OFF-center cell will fire fewer action potentials when a small spot of light is projected to the center of its receptive field; it will fire more action potentials if a small dark spot covers the receptive field center. In both ON and OFF types of cell, the response to stimulation of the center is canceled by the response to stimulation of the surround (Figure 9.27). The surprising implication is that most retinal ganglion cells are not particularly responsive to changes in illumination that include both the receptive field center and the receptive field surround. Rather, it appears that the ganglion cells are mainly responsive to differences in illumination that occur within their receptive fields.

FIGURE 9.27 A center-surround ganglion cell receptive field. (a, b) An OFF-center ganglion cell responds with a barrage of action potentials when a dark spot is imaged on its receptive field center. (c) If the dark spot is enlarged to include the receptive field surround, the response is greatly reduced. Description
To illustrate this point, consider the response generated by an OFF-center cell as a light–dark edge crosses its receptive field (Figure 9.28). Remember that in such a cell, dark in the center of the receptive field causes the cell to depolarize, whereas dark in the surround causes the cell to hyperpolarize. In uniform illumination, the center and surround cancel to yield some low level of response (Figure 9.28a). When the edge enters the surround region of the receptive field without encroaching on the center, the dark area has the effect of hyperpolarizing the neuron, leading to a decrease in the cell’s firing rate (Figure 9.28b). As the dark area begins to include the center, however, the partial inhibition by the surround is overcome, and the cell response increases (Figure 9.28c). To understand why the response in Figure 28c is increased, consider that there is 100% excitation of the cell because darkness covers the entire receptive field center, but there is only partial inhibition from the portion of the surround in darkness. But when the dark area finally fills the entire surround, the center response is canceled (Figure 9.28d). Notice that the cell response in this example is only slightly different in uniform light and in uniform dark; the response is modulated mainly by the presence of the light–dark edge in its receptive field.

FIGURE 9.28 Responses to a light–dark edge crossing an OFF-center ganglion cell receptive field. The response of the neuron is determined by the fraction of the center and surround that are filled by light and dark. (See text for details.) Description
Now let’s consider the output of all the OFF-center ganglion cells at different positions on the retina that are stimulated by a stationary light–dark edge. The responses will fall into the same four categories illustrated in Figure 9.28. Thus, the cells that will register the presence of the edge are those with receptive field centers and surrounds that are differentially affected by the light and dark areas. The population of cells with receptive field centers “viewing” the light side of the edge will be inhibited (see Figure 9.28b). The population of cells with centers “viewing” the dark side of the edge will be excited (see Figure 9.28c). In this way, the difference in illumination at a light–dark edge is not faithfully represented by the difference in the output of ganglion cells on either side of the edge. Instead, the center-surround organization of the receptive fields leads to a neural response that emphasizes the contrast at light–dark edges.
There are many visual illusions involving the perception of light level. The organization of ganglion cell receptive fields suggests an explanation for the illusion shown in Figure 9.29. Even though the two central squares are the same shade of gray, the square on the lighter background appears darker. Consider the two ON-center receptive fields shown on the gray squares. In both cases, the same gray light hits the receptive field center. However, the receptive field on the left has more light in its surround than the receptive field on the right. This will lead to more inhibition and a lower response and may be related to the darker appearance of the left central gray square.

FIGURE 9.29 The influence of contrast on the perception of light and dark. The central boxes are identical shades of gray, but because the surrounding area is lighter on the left, the left central box appears darker. ON-center receptive fields are shown on the left and right of the figure. Which would respond more? Description
Structure-Function Relationships. Most ganglion cells in the mammalian retina have a center-surround receptive field with either an ON or an OFF center. They can be further categorized based on their appearance, connectivity, and electrophysiological properties. In the macaque monkey retina and human retina, two major types of ganglion cells are distinguished: large M-type ganglion cells and smaller P-type ganglion cells. (M stands for magno, from the Latin for “large”; P stands for parvo, from the Latin for “small.”) Figure 9.30 shows the relative sizes of M and P ganglion cells at the same location on the retina. P cells constitute about 90% of the ganglion cell population, M cells constitute about 5%, and the remaining 5% are made up of a variety of nonM–nonP ganglion cell types that are less well characterized.

FIGURE 9.30 M-type and P-type ganglion cells in the macaque monkey retina. (a) A small P cell from the peripheral retina. (b) An M cell from a similar retinal location is significantly larger. (Source: Watanabe and Rodieck, 1989, pp. 437, 439.)
The visual response properties of M cells differ from those of P cells in several ways. They have larger receptive fields, they conduct action potentials more rapidly in the optic nerve, and they are more sensitive to low-contrast stimuli. In addition, M cells respond to stimulation of their receptive field centers with a transient burst of action potentials, while P cells respond with a sustained discharge as long as the stimulus is on (Figure 9.31). We will see in Chapter 10 that the different types of ganglion cells appear to play different roles in visual perception.

FIGURE 9.31 Different responses to light of M-type and P-type ganglion cells. Description
Color-Opponent Ganglion Cells. Another important distinction between ganglion cell types is that some P cells and nonM–nonP cells are sensitive to differences in the wavelength of light. The majority of these color-sensitive neurons are called color-opponent cells, reflecting the fact that the response to one color in the receptive field center is canceled by showing another color in the receptive field surround. The two types of opponency are red versus green and blue versus yellow. Consider, for example, a cell with a red ON center and a green OFF surround (Figure 9.32). The center of the receptive field is fed by red cones, and the surround is fed by green cones via an inhibitory circuit (i.e., the indirect horizontal cell path). To understand how the neuron with this receptive field responds to light, recall Figure 9.21, which shows that red and green cones absorb different, but overlapping, wavelengths of light.

FIGURE 9.32 Color opponency in ganglion cells. (a) A color-opponent center-surround receptive field of a P-type ganglion cell. (b) A strong response is elicited by red light to the receptive field center, which receives input from long-wavelength-sensitive (red) cones. (c) Extending the red light into the surround inhibits the response because the green cones providing input to the surround are also somewhat sensitive to long wavelengths of light. (d) Even stronger inhibition is produced by green light in the receptive field surround, which optimally drives the green cones. Description
If red light bathes the receptive field center, the neuron responds with a strong burst of action potentials (Fig. 9.32b). If the red light is extended to cover both the center and the surround of the receptive field, the neuron is still excited but much less so (Fig. 9.32c). The reason that red light has an effect in the green OFF surround is that, as shown in Fig. 9.21, red wavelengths of light are partially absorbed by green cones, and their activation inhibits the response of the neuron. To fully activate the inhibitory surround of the receptive, green light is needed. In this case, the red ON center response is canceled by the green OFF surround response (Fig. 9.32d). The shorthand notation for such a cell is R+G–, meaning simply that it is optimally excited by red in the receptive field center and inhibited by green in the surround.
What would be the response to white light on the entire receptive field? Because white light contains all visible wavelengths (including both red and green), both center and surround would be equally activated, canceling each other, and the cell would not respond to the light.
Blue–yellow color opponency works the same way. Consider a cell with a blue ON center and a yellow OFF surround (B+Y–). The receptive field center receives input from blue cones, and the surround gets input from both red and green cones (hence yellow) via an inhibitory circuit. Blue light hitting the receptive field center would give a strong excitatory response that could be canceled by yellow light in the surround. Diffuse blue light covering both the center and the surround of the receptive field would also be an effective stimulus for this cell. Why is diffuse blue light a strong stimulus here, whereas diffuse red light was a much weaker stimulus in R+G– cells? The answer lies in the absorption curves (see Figure 9.21); in the R+G– cell, red light is absorbed by the “green” photoreceptors, but in the B+Y– neuron, very little blue light is absorbed by the “red” and “green” photoreceptors that make up the receptive field surround. Diffuse white light would not be an effective stimulus because it contains red, blue, and green wavelengths, and the center and surround responses would cancel.
Finally, note that M ganglion cells lack color opponency. This doesn’t mean they won’t respond to colored light; rather, their responses are not color-specific. So, for example, red light in the receptive field center or surround would have the same effect as green light. The lack of color opponency in M- cells is accounted for by the fact that both the center and the surround of the receptive field receive input from more than one type of cone. M-cell receptive fields are therefore denoted as simply either ON center/OFF surround or OFF center/ON surround. The color and light sensitivity of M and P ganglion cells suggest that the overall ganglion cell population sends information to the brain about three different spatial comparisons: light versus dark, red versus green, and blue versus yellow.
The retinal circuit we have described, in which rods and cones project to bipolar and then ganglion cells, implies that rods and cones are responsible for all phototransduction and that ganglion cells play a different role relaying visual information to the rest of the brain. But even back in the 1980s and 1990s, there were odd findings that were difficult to reconcile with this view. For example, mutant mice lacking rods and cones appeared to synchronize their sleeping and waking with the rise and setting of the sun even though they otherwise behaved as if they were blind. A subset of humans who are totally blind also appear to unconsciously synchronize their behavior to daily changes in sunlight.
The resolution of these mysteries came from the discovery in the 1990s that a small percentage of retinal ganglion cells actually transduce light. These few thousand neurons, known as intrinsically photosensitive retinal ganglion cells (ipRGCs), use melanopsin as a photopigment, an opsin previously studied in frog skin! The ipRGCs function as normal ganglion cells that receive input from rods and cones and send axons out the optic nerve; in addition they are photoreceptors. The photosensitivity of ipRGCs differs from that of rods and cones in important ways, however. Unlike the hyperpolarization that light causes in rods and cones, ipRGCs depolarize to light. The ipRGCs also have very large dendritic fields; because the dendrites are photosensitive, this means the cells sum light input over much larger areas of the retina than rods and cones (Figure 9.33). The small number of ipRGCs and their large receptive fields are not ideal for, and the ipRGCs are not used in, fine pattern vision. As we will see in Chapter 19, an important function of the ipRGCs is providing input to subcortical visual areas that synchronize behavior to daily changes in light level (circadian rhythms). Since their initial discovery, a spectrum of different ipRGCs has been found that vary in their morphology, physiology, and connections to other retinal neurons. Research continues to investigate the multiple roles of these unusual cells in unconscious and conscious vision.

FIGURE 9.33 Intrinsically photosensitive retinal ganglion cells. (a) Rods (blue) and cones (green) project to bipolar cells and then conventional ganglion cells (black) that send axons to the thalamus. In addition to receiving rod and cone inputs, ipRGCs (red) transduce light themselves. Unlike rods and cones, these photosensitive neurons send axons out of the eye without additional neurons and synaptic connections. The dendrites of the ipRGCs spread over a much broader area than conventional ganglion cell dendrites, making these neurons sensitive to light over a larger area. (b) A micrograph of an ipRGC shows the long winding dendrites and the axon (arrow). Note how much larger the dendritic field is compared to those of the regular ganglion cells in Figure 9.30. (Source: Berson, 2003, Fig. 1.) Description
One of the important concepts that emerge from our discussion of the retina is the idea of parallel processing in the visual system. Parallel processing means that different visual attributes are processed simultaneously using distinct pathways. For example, we view the world with not one but two eyes that provide two parallel streams of information. In the central visual system, these streams are compared to give information about depth, the distance of an object from the observer. A second example of parallel processing is the independent streams of information about light and dark that arise from the ON-center and OFF-center ganglion cells in each retina. Finally, ganglion cells of both ON and OFF varieties have different types of receptive fields and response properties. M cells can detect subtle contrasts over their large receptive fields and are likely to contribute to low-resolution vision. P cells have small receptive fields that are well suited for the discrimination of fine detail. P cells and nonM–nonP cells are specialized for the separate processing of red–green and blue–yellow information.
In this chapter, we have seen how light emitted by or reflected off objects can be imaged by the eye onto the retina. Light energy is first converted into membrane potential changes in the mosaic of photoreceptors. Interestingly, the transduction mechanism in photoreceptors is very similar to that in olfactory receptor cells, both of which involve cyclic nucleotide-gated ion channels. Photoreceptor membrane potential is converted into a chemical signal (the neurotransmitter glutamate), which is again converted into membrane potential changes in the postsynaptic bipolar and horizontal cells. This process of electrical-to-chemical-to-electrical signaling repeats again and again, until the presence of light or dark or color is finally converted to a change in the action potential firing frequency of the ganglion cells.
Information from 97 million photoreceptors is funneled into 1 million ganglion cells. In the central retina, particularly the fovea, relatively few photoreceptors feed each ganglion cell, whereas in the peripheral retina, thousands of receptors do. Thus, the mapping of visual space onto the array of optic nerve fibers is not uniform. Rather, in “neural space,” there is an overrepresentation of the central few degrees of visual space, and signals from individual cones are more important. This specialization ensures high acuity in central vision but also requires that the eye move to bring the images of objects of interest onto the fovea.
As we shall see in the next chapter, there is good reason to believe that the different types of information that arise from different types of ganglion cells are, at least in the early stages, processed independently. Parallel streams of information—for example, from the right and left eyes—remain segregated at the first synaptic relay in the lateral geniculate nucleus of the thalamus. The same can be said for the M-cell and P-cell synaptic relays in the LGN. In the visual cortex, it appears that parallel paths may process different visual attributes. For example, the distinction in the retina between neurons that do or do not convey information about color is preserved in the visual cortex. In general, each of the more than two dozen visual cortical areas may be specialized for the analysis of different types of retinal output.
intrinsically photosensitive retinal ganglion cell (ipRGC) (p. 327)
1. What physical property of light is most closely related to the perception of color?
2. Name eight structures in the eye that light passes through before it strikes the photoreceptors.
3. Why is a scuba mask necessary for clear vision underwater?
5. Give three reasons explaining why visual acuity is best when images fall on the fovea.
6. How does the membrane potential change in response to a spot of light in the receptive field center of a photoreceptor? Of an ON bipolar cell? Of an OFF-center ganglion cell? Why?
7. What happens in the retina when you “get used to the dark”? Why can’t you see color in the dark?
8. In what way is retinal output not a faithful reproduction of the visual image falling on the retina?
9. In retinitis pigmentosa, early symptoms include the loss of peripheral vision and night vision. The loss of what type of cells could lead to such symptoms?
Arshavsky VY, Lamb TD, Pugh EN. 2002. G proteins and phototransduction. Annual Review of Physiology 64:153–187.
Berson DM. 2003. Strange vision: ganglion cells as circadian photoreceptors. Trends in Neurosciences 26:314–320.
Field GD, Chichilinsky EJ. 2007. Information processing in the primate retina: circuitry and coding. Annual Review of Neuroscience 30:1–30.
Nassi JJ, Callaway EM. 2009. Parallel processing strategies of the primate visual system. Nature Reviews Neuroscience 10:360–372.
Solomon SG, Lennie P. 2007. The machinery of colour vision. Nature Reviews Neuroscience 8: 276–286.
Wade NJ. 2007. Image, eye, and retina. Journal of the Optical Society of America 24:1229–1249.
Wassle H. 2004. Parallel processing in the mammalian retina. Nature Reviews Neuroscience 5: 747–757.
Additional figures