Brain Control of Movement
BOX 14.1 OF SPECIAL INTEREST: Paresis, Paralysis, Spasticity, and Babinski

The Contributions of Posterior Parietal and Prefrontal Cortex
BOX 14.2 OF SPECIAL INTEREST: Behavioral Neurophysiology
Direct and Indirect Pathways through the Basal Ganglia
BOX 14.3 OF SPECIAL INTEREST: Do Neurons in Diseased Basal Ganglia Commit Suicide?
BOX 14.4 OF SPECIAL INTEREST: Destruction and Stimulation: Useful Therapies for Brain Disorders
THE INITIATION OF MOVEMENT BY PRIMARY MOTOR CORTEX
BOX 14.5 PATH OF DISCOVERY: Distributed Coding in the Superior Colliculus, by James T. McIlwain
BOX 14.6 OF SPECIAL INTEREST: Involuntary Movements—Normal and Abnormal
In Chapter 13, we discussed the organization of the peripheral somatic motor system: the joints, skeletal muscles, and their sensory and motor innervation. We saw that the final common pathway for behavior is the alpha motor neuron, that the activity of this cell is under the control of sensory feedback and spinal interneurons, and that reflex movements reveal the complexity of this spinal control system. In this chapter, we’ll explore how the brain influences the activity of the spinal cord to command voluntary movements.
The central motor system is arranged as a hierarchy of control levels, with the forebrain at the top and the spinal cord at the bottom. It is useful to think of this motor control hierarchy as having three levels (Table 14.1). The highest level, represented by the association areas of neocortex and basal ganglia of the forebrain, is concerned with strategy: the goal of the movement and the movement strategy that best achieves the goal. The middle level, represented by the motor cortex and cerebellum, is concerned with tactics: the sequences of muscle contractions, arranged in space and time, required to smoothly and accurately achieve the strategic goal. The lowest level, represented by the brain stem and spinal cord, is concerned with execution: activation of the motor neuron and interneuron pools that generate the goal-directed movement and make any necessary adjustments of posture.
To appreciate the different contributions of the three hierarchical levels to movement, consider the actions of a baseball pitcher preparing to pitch to a batter (Figure 14.1). The cerebral neocortex has information—based on vision, audition, somatic sensation, and proprioception—about precisely where the body is in space. Strategies must be devised to move the body from the current state to one in which a pitch is delivered and the desired outcome is attained (a swing and a miss by the batter). Several throwing options are available to the pitcher—a curve ball, a fast ball, a slider, and so on—and these alternatives are filtered through the basal ganglia and back to the cortex until a final decision is made, based in large part on past experience (e.g., “This batter hit a home run last time I threw a fast ball”). The motor areas of cortex and the cerebellum then make the tactical decision (to throw a curve ball) and issue instructions to the brain stem and spinal cord. Activation of neurons in the brain stem and spinal cord then causes the movement to be executed. The properly timed activation of motor neurons in the cervical spinal cord generates a coordinated movement of the shoulder, elbow, wrist, and fingers. Simultaneously, brain stem inputs to the thoracic and lumbar spinal cord command the appropriate leg movements along with postural adjustments that keep the pitcher from falling over during the throw. In addition, brain stem motor neurons are activated to keep the pitcher’s eyes fixed on the catcher, his target, as his head and body move about.

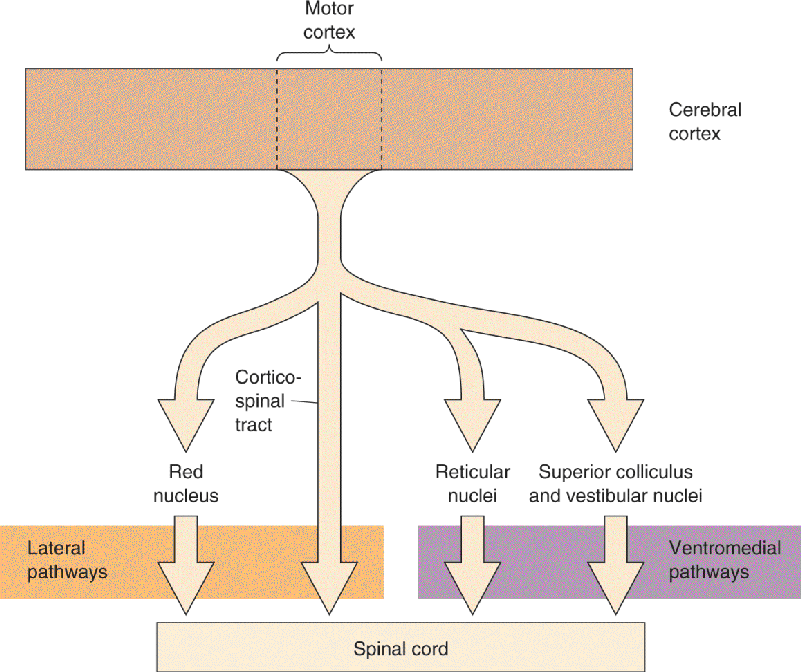
FIGURE 14.1 The contributions of the motor control hierarchy. As a baseball pitcher plans to pitch a ball to a batter, chooses which pitch to throw, and then throws the ball, he engages the three hierarchical levels of motor control.
According to the laws of physics, the movement of a thrown baseball through space is ballistic, referring to a trajectory that cannot be altered. The movement of the pitcher’s arm that throws the ball is also described as ballistic because it cannot be altered once initiated. This type of rapid voluntary movement is not under the same type of sensory feedback control that regulates antigravity postural reflexes (see Chapter 13). The reason is simple: The movement is too fast to be adjusted by sensory feedback. But the movement does not occur in the absence of sensory information. Sensory information before the movement was initiated was crucial in order to decide when to initiate the pitch, to determine the starting positions of the limbs and body, and to anticipate any changes in resistance during the throw. And sensory information during the movement is also important, not necessarily for the movement at hand, but for improving subsequent similar movements.
The proper functioning of each level of the motor control hierarchy relies so heavily on sensory information that the motor system of the brain might properly be considered a sensorimotor system. At the highest level, sensory information generates a mental image of the body and its relationship to the environment. At the middle level, tactical decisions are based on the memory of sensory information from past movements. At the lowest level, sensory feedback is used to maintain posture, muscle length, and tension before and after each voluntary movement.
In this chapter, we investigate this hierarchy of motor control and how each level contributes to the control of the peripheral somatic motor system. We start by exploring the pathways that bring information to the spinal motor neurons. From there we will ascend to the highest levels of the motor hierarchy, and then we’ll fill in the pieces of the puzzle that bring the different levels together. Along the way, we’ll describe how pathology in specific parts of the motor system leads to particular movement disorders.
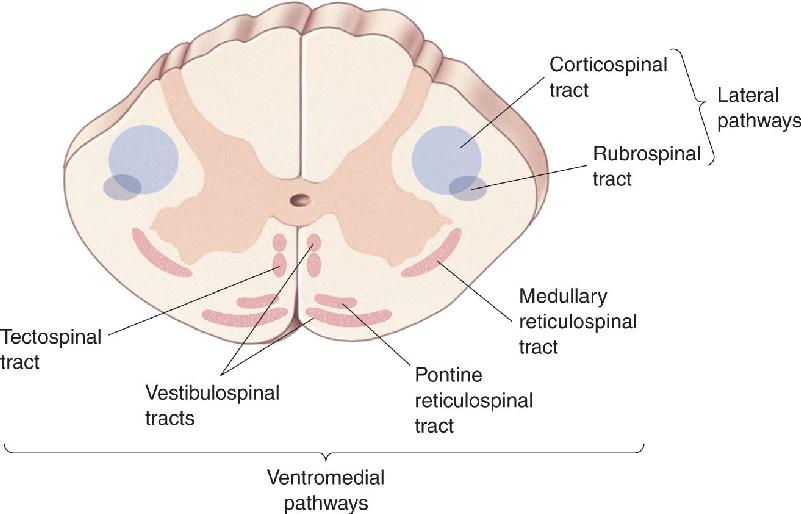
How does the brain communicate with the motor neurons of the spinal cord? Axons from the brain descend through the spinal cord along two major groups of pathways, shown in Figure 14.2. One is in the lateral column of the spinal cord, and the other is in the ventromedial column. Remember this rule of thumb: The lateral pathways are involved in voluntary movement of the distal musculature and are under direct cortical control, and the ventromedial pathways are involved in the control of posture and locomotion and are under brain stem control.

FIGURE 14.2 The descending tracts of the spinal cord. The lateral pathways, consisting of the corticospinal and rubrospinal tracts, control voluntary movements of the distal musculature. The ventromedial pathways, consisting of the medullary reticulospinal, pontine reticulospinal, vestibulospinal, and tectospinal tracts, control postural muscles. Description
The most important component of the lateral pathways is the corticospinal tract (Figure 14.3a). Originating in the neocortex, it is the longest and one of the largest central nervous system (CNS) tracts (106 axons in humans). Two-thirds of the axons in the tract originate in areas 4 and 6 of the frontal lobe, collectively called motor cortex. Most of the remaining axons in the corticospinal tract derive from the somatosensory areas of the parietal lobe and serve to regulate the flow of somatosensory information to the brain (see Chapter 12). Axons from the cortex pass through the internal capsule bridging the telencephalon and thalamus, course through the base of the cerebral peduncle, a large collection of axons in the midbrain, then pass through the pons, and collect to form a tract at the base of the medulla. The tract forms a bulge, called the medullary pyramid, running down the ventral surface of the medulla. When cut, the tract’s cross section is roughly triangular, explaining why it is also called the pyramidal tract.

FIGURE 14.3 The lateral pathways. Origins and terminations of (a) the corticospinal tract and (b) the rubrospinal tract. These tracts control fine movements of the arms and fingers. Description
At the junction of the medulla and spinal cord, the pyramidal tract crosses, or decussates, at the pyramidal decussation. This means that the right motor cortex directly commands the movement of the left side of the body, and the left motor cortex controls the muscles on the right side. As the axons cross, they collect in the lateral column of the spinal cord and form the lateral corticospinal tract. The corticospinal tract axons terminate in the dorsolateral region of the ventral horns and intermediate gray matter, the location of the motor neurons and interneurons that control the distal muscles, particularly the flexors (see Chapter 13).
A much smaller component of the lateral pathways is the rubrospinal tract, which originates in the red nucleus of the midbrain, named for its distinctive pinkish hue in a freshly dissected brain (rubro is from the Latin for “red”). Axons from the red nucleus decussate in the pons, almost immediately, and parallel those in the corticospinal tract in the lateral column of the spinal cord (Figure 14.3b). A major source of input to the red nucleus is the very region of frontal cortex that also contributes to the corticospinal tract. Indeed, it appears that this indirect corticorubrospinal pathway has largely been replaced by the direct corticospinal path over the course of primate evolution Thus, while the rubrospinal tract contributes importantly to motor control in many mammalian species, in humans it appears to be reduced, most of its functions subsumed by the corticospinal tract.
The Effects of Lateral Pathway Lesions. Donald Lawrence and Hans Kuypers laid the foundation for the modern view of the functions of the lateral pathways in the late 1960s. Experimental lesions in both corticospinal and rubrospinal tracts in monkeys rendered them unable to make fractionated movements of the arms and hands; that is, they could not move their shoulders, elbows, wrists, and fingers independently. For example, they could grasp small objects with their hands but only by using all the fingers at once. Voluntary movements were also slower and less accurate. Despite this, the animals could sit upright and stand with normal posture. By analogy, a human with a lateral pathway lesion would be able to stand on the pitcher’s mound but would be unable to grip the ball properly and throw it accurately.
Lesions in the monkeys’ corticospinal tracts alone caused a movement deficit as severe as that observed after lesions in the lateral columns. Interestingly, however, many functions gradually reappeared over the months following surgery. In fact, the only permanent deficit was some weakness of the distal flexors and an inability to move the fingers independently. A subsequent lesion in the rubrospinal tract completely reversed this recovery, however. These results suggest that the corticorubrospinal pathway was able, over time, to partially compensate for the loss of the corticospinal tract input.
Strokes that damage the motor cortex or the corticospinal tract are common in humans. Their immediate consequence can be paralysis on the contralateral side, but considerable recovery of voluntary movements may occur over time (Box 14.1). As in Lawrence and Kuypers’ lesioned monkeys, it is the fine, fractionated movements of the fingers that are least likely to recover.
The neural components of the motor system extend from the highest reaches of the cerebral cortex to the farthest terminals of the motor axons in muscles. Its sheer size makes the motor system uncommonly vulnerable to disease and trauma. The site of motor system damage has a big effect on the types of deficits patients experience.
Damage to the lower parts of the motor system—alpha motor neurons or their motor axons—leads to easily predicted consequences. Partial damage may cause paresis (weakness). Complete severing of a motor nerve leads to paralysis, a loss of movement of the affected muscles, and areflexia, an absence of their spinal reflexes. The muscles also have no tone or resting tension; they are flaccid and soft. Damaged motor neurons can no longer exert their trophic influence on muscle fibers (see Chapter 13). The muscles profoundly atrophy (decrease in size) with time, losing up to 70–80% of their mass.
Damage to the upper parts of the motor system—the motor cortex or the various motor tracts that descend into the spinal cord—can cause a distinctly different set of motor problems. These are common after a stroke, which damages regions of the cortex or brain stem by depriving them of their blood supply, or traumatic injury, such as a knife or gunshot wound, or even a demyelinating disease that damages axons (see Box 4.5).
Immediately following severe upper motor system damage, there is a period of spinal shock: reduced muscle tone (hypotonia), areflexia, and paralysis. Paralysis is known as hemiplegia if it occurs on one side of the body, paraplegia if it involves only the legs, and quadriplegia if it involves all four limbs. With the loss of descending brain influences, the functions of the spinal cord appear to shut down. Over the next several days, some of its reflexive functions mysteriously reappear. This is not necessarily a good thing. A condition called spasticity sets in, often permanently. Spasticity is characterized by a dramatic and sometimes painful increase of muscle tone (hypertonia) and spinal reflexes (hyperreflexia), compared to normal. Overactive stretch reflexes often cause clonus, rhythmic cycles of contraction and relaxation when limb muscles are stretched.
Another indication of motor tract damage is the Babinski sign, described by the French neurologist Joseph Babinski in 1896. Sharply scratching the sole of the foot from the heel toward the toes causes reflexive upward flexion of the big toe and an outward fanning of the other toes. The normal response to this stimulus, for anyone older than about 2 years, is to curl the toes downward. Normal infants also exhibit the Babinski sign, presumably because their descending motor tracts have not yet matured.
By systematically testing a patient’s reflexes, muscle tone, and motor ability across his body, a skilled neurologist can often deduce the site and severity of motor system damage with impressive precision.
The ventromedial pathways contain four descending tracts that originate in the brain stem and terminate among the spinal interneurons controlling proximal and axial muscles. These tracts are the vestibulospinal tract, the tectospinal tract, the pontine reticulospinal tract, and the medullary reticulospinal tract. The ventromedial pathways use sensory information about balance, body position, and the visual environment to reflexively maintain balance and body posture.
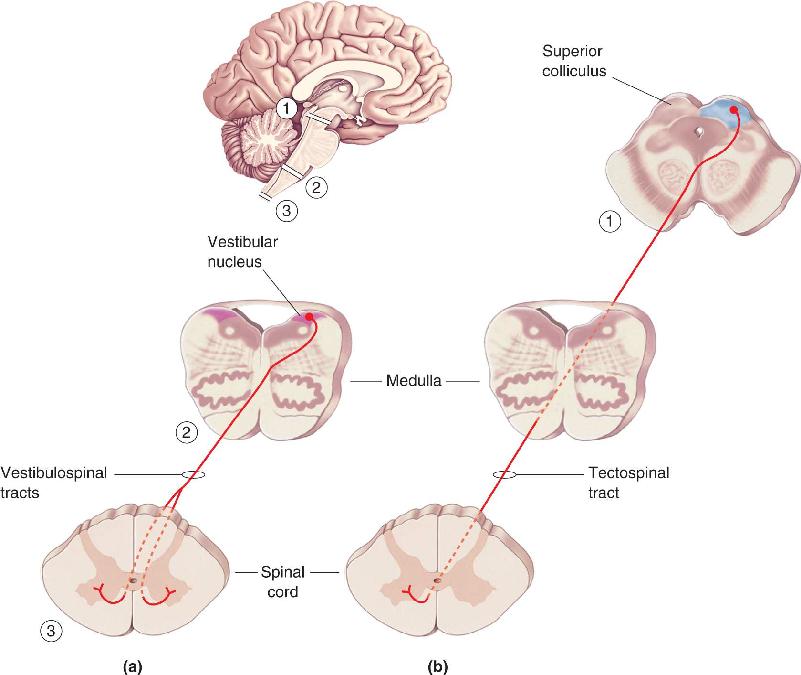
The Vestibulospinal Tracts. The vestibulospinal and tectospinal tracts function to keep the head balanced on the shoulders as the body moves through space and to turn the head in response to new sensory stimuli. The vestibulospinal tracts originate in the vestibular nuclei of the medulla, which relay sensory information from the vestibular labyrinth in the inner ear (Figure 14.4a). The vestibular labyrinth consists of fluid-filled canals and cavities in the temporal bone that are closely associated with the cochlea (see Chapter 11). The motion of the fluid in this labyrinth, which accompanies movement of the head, activates hair cells that signal the vestibular nuclei via cranial nerve VIII.

FIGURE 14.4 The ventromedial pathways. Origins and terminations of (a) the vestibulospinal tracts and (b) the tectospinal tract. These tracts control the posture of the head and neck. Description
One component of the vestibulospinal tracts projects bilaterally down the spinal cord and activates the cervical spinal circuits that control neck and back muscles and thus guide head movement. Stability of the head is important because the head contains our eyes, and keeping the eyes stable, even as our body moves, ensures that our image of the world remains stable. Another component of the vestibulospinal tracts projects ipsilaterally as far down as the lumbar spinal cord. It helps us maintain an upright and balanced posture by facilitating extensor motor neurons of the legs.
The Tectospinal Tract. The tectospinal tract originates in the superior colliculus of the midbrain, which receives direct input from the retina (Figure 14.4b). (Recall from Chapter 10 that optic tectum is another name for the superior colliculus.) Besides its retinal input, the superior colliculus receives projections from visual cortex, as well as afferent axons carrying somatosensory and auditory information. From this input, the superior colliculus constructs a map of the world around us; stimulation at one site in this map leads to an orienting response that directs the head and eyes to move so that the appropriate point of space is imaged on the fovea. Activation of the colliculus by an image of a runner sprinting toward second base, for example, would cause the pitcher to orient his head and eyes toward this important new stimulus.
After leaving the colliculus, axons of the tectospinal tract quickly decussate and project close to the midline into cervical regions of the spinal cord, where they help to control muscles of the neck, upper trunk, and shoulders.
The Pontine and Medullary Reticulospinal Tracts. The reticulospinal tracts arise mainly from the reticular formation of the brain stem, which runs the length of the brain stem at its core, just under the cerebral aqueduct and fourth ventricle. A complex meshwork of neurons and fibers, the reticular formation receives input from many sources and participates in many different functions. For the purposes of our discussion of motor control, the reticular formation may be divided into two parts that give rise to two different descending tracts: the pontine (medial) reticulospinal tract and the medullary (lateral) reticulospinal tract (Figure 14.5).

FIGURE 14.5 The pontine (medial) and medullary (lateral) reticulospinal tracts. These components of the ventromedial pathway control posture of the trunk and the antigravity muscles of the limbs. Description
The pontine reticulospinal tract enhances the antigravity reflexes of the spinal cord. Activity in this pathway, by facilitating the extensors of the lower limbs, helps maintain a standing posture by resisting the effects of gravity. This type of regulation is an important component of motor control: Keep in mind that most of the time, the activity of ventral horn neurons maintains, rather than changes, muscle length and tension. The medullary reticulospinal tract, however, has the opposite effect; it liberates the antigravity muscles from reflex control. Activity in both reticulospinal tracts is controlled by descending signals from the cortex. A fine balance between them is required as the pitcher goes from standing on the mound to winding up and throwing the ball.
Figure 14.6 provides a simple summary of the major descending spinal tracts. The ventromedial pathways originate from several regions of the brain stem and participate mainly in the maintenance of posture and certain reflex movements. Initiation of a voluntary, ballistic movement, such as throwing a baseball, requires instructions that descend from the motor cortex along the lateral pathways. The motor cortex directly activates spinal motor neurons and also liberates them from reflex control by communicating with the nuclei of the ventromedial pathways. It is clear that the cortex is critical for voluntary movement and behavior, so we will now focus our attention there.

FIGURE 14.6 A summary of the major descending spinal tracts and their origins. Description
Although cortical areas 4 and 6 are called motor cortex, it is important to recognize that the control of voluntary movement engages almost all of the neocortex. Goal-directed movement depends on knowledge of where the body is in space, where it intends to go, and on the selection of a plan to get it there. Once a plan has been selected, it must be held in memory until the appropriate time. Finally, instructions must be issued to implement the plan. To some extent, these different aspects of motor control are localized to different regions of the cerebral cortex. In this section, we explore some of the cortical areas implicated in motor planning. Later we’ll look at how a plan is converted into action.
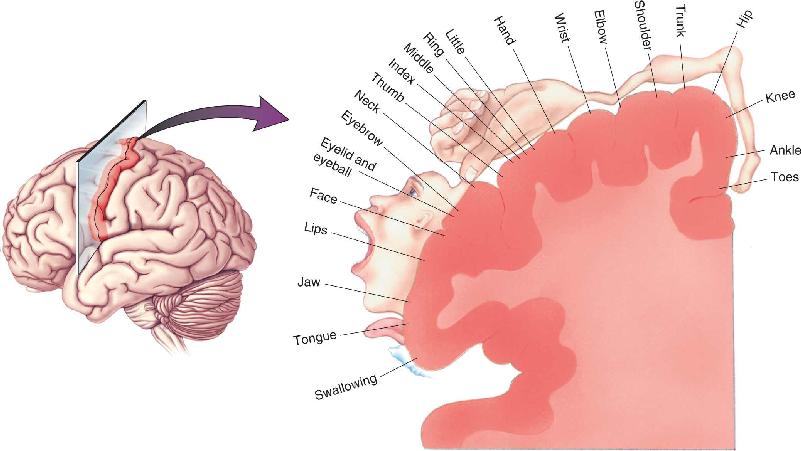
The motor cortex is a circumscribed region of the frontal lobe. Area 4 lies just anterior to the central sulcus on the precentral gyrus, and area 6 lies just anterior to area 4 (Figure 14.7). The definitive demonstration that these areas constitute motor cortex in humans came from the work of neurosurgeon Wilder Penfield. Recall from Chapter 12 that Penfield electrically stimulated the cortex in patients who were undergoing surgery to remove bits of brain thought to be inducing epileptic seizures. The stimulation was used in an attempt to identify which regions of cortex were so critical that they should be spared from the knife. In the course of these operations, Penfield discovered that weak electrical stimulation of area 4 in the precentral gyrus would elicit a twitch of the muscles in a particular region of the body on the contralateral side. Systematic probing of this region established that there is a somatotopic organization in the human precentral gyrus much like that seen in the somatosensory areas of the postcentral gyrus (Figure 14.8). Area 4 is now often referred to as primary motor cortex or M1.

FIGURE 14.7 Planning and directing voluntary movements. These areas of the neocortex are involved in the control of voluntary movement. Areas 4 and 6 constitute the motor cortex. Description

FIGURE 14.8 A somatotopic motor map of the human precentral gyrus. Area 4 of the precentral gyrus is also known as primary motor cortex (M1). Description
The foundation for Penfield’s discovery had been laid nearly a century before by Gustav Fritsch and Eduard Hitzig, who in 1870 had shown that stimulation of the frontal cortex of anesthetized dogs would elicit movement of the contralateral side of the body (see Chapter 1). Then, around the turn of the century, David Ferrier and Charles Sherrington discovered that the motor area in primates was located in the precentral gyrus. By comparing the histology of this region in Sherrington’s apes with that of the human brain, Australian neuroanatomist Alfred Walter Campbell concluded that cortical area 4 is motor cortex.
Campbell speculated that cortical area 6, just rostral to area 4, might be an area specialized for skilled voluntary movement. Penfield’s studies 50 years later supported the conjecture that this was a “higher” motor area in humans by showing that electrical stimulation of area 6 could evoke complex movements of either side of the body. Penfield found two somatotopically organized motor maps in area 6: one in a lateral region he called the premotor area (PMA) and one in a medial region called the supplementary motor area (SMA) (see Figure 14.7). These two areas appear to perform similar functions but on different groups of muscles. While the SMA sends axons that innervate distal motor units directly, the PMA connects primarily with reticulospinal neurons that innervate proximal motor units.
The Contributions of Posterior Parietal and Prefrontal Cortex
Recall the baseball player standing on the mound, preparing to pitch. It should be apparent that before the detailed sequence of muscle contractions for the desired pitch can be calculated, the pitcher must have information about the current position of his body in space and how it relates to the positions of the batter and the catcher. This mental body image seems to be generated by somatosensory, proprioceptive, and visual inputs to the posterior parietal cortex.
Two areas are of particular interest in the posterior parietal cortex: area 5, which is a target of inputs from the primary somatosensory cortical areas 3, 1, and 2 (see Chapter 12); and area 7, which is a target of higher order visual cortical areas such as MT (see Chapter 10). Recall that human patients with lesions in these areas of the parietal lobes, as can occur after a stroke, show bizarre abnormalities of body image and the perception of spatial relations. In its most extreme manifestation, the patient will simply neglect the side of the body, and even the rest of the world, opposite the parietal lesion.
The parietal lobes are extensively interconnected with regions in the anterior frontal lobes that in humans are thought to be important for abstract thought, decision making, and anticipating the consequences of action. These “prefrontal” areas, along with the posterior parietal cortex, represent the highest levels of the motor control hierarchy, where decisions are made about what actions to take and their likely outcome (a curve ball followed by a strike). The prefrontal cortex and parietal cortex both send axons that converge on cortical area 6. Recall that areas 6 and 4 together contribute most of the axons to the descending corticospinal tract. Thus, area 6 lies at the junction where signals encoding what actions are converted into signals that specify how the actions will be carried out.
This general view of higher order motor planning received dramatic support in a series of studies on humans carried out by Danish neurologist Per Roland and his colleagues. They used positron emission tomography (PET) to monitor changes in the patterns of cortical activation that accompany voluntary movements (see Box 7.3). When the subjects were asked to perform a series of finger movements from memory, the following regions of cortex showed increased blood flow: the somatosensory and posterior parietal areas, parts of the prefrontal cortex (area 8), area 6, and area 4. These are the very regions of the cerebral cortex that, as discussed earlier, are thought to play a role in generating the intention to move and converting that intention into a plan of action. Interestingly, when the subjects were asked only to mentally rehearse the movement without actually moving the finger, area 6 remained active but area 4 did not.
Experimental work on monkeys further supports the idea that area 6 (SMA and PMA) plays an important role in the planning of movement, particularly complex movement sequences of the distal musculature. Using a method developed in the late 1960s by Edward Evarts at the National Institutes of Health, researchers have recorded the activity of neurons in the motor areas of awake, behaving animals (Box 14.2). Cells in the SMA typically increase their discharge rates about a second before the execution of a hand or wrist movement, consistent with their proposed role in planning movement (recall Roland’s findings in humans). An important feature of this activity is that it occurs in advance of the movements of either hand, suggesting that the supplementary areas of the two hemispheres are closely linked via the corpus callosum. Indeed, movement deficits observed following an SMA lesion on one side, in both monkeys and humans, are particularly pronounced for tasks requiring the coordinated actions of the two hands, such as buttoning a shirt. In humans, a selective inability to perform complex (but not simple) motor acts is called apraxia.
Showing that a brain lesion impairs movement and that brain stimulation elicits movement does not tell us how the brain controls movement. To address this problem, we need to know how the activity of neurons relates to different types of voluntary movement in the intact organism. PET scans and fMRI are extremely valuable for plotting out the distribution of activity in the brain as behaviors are performed, but they lack the resolution to track the millisecond-by-millisecond changes in the activity of individual neurons. The best method for this purpose is extracellular recording with metal microelectrodes (see Box 4.1). But how is this done in awake, behaving animals?
This problem was solved by Edward Evarts and his colleagues at the National Institutes of Health. Monkeys were trained to perform simple tasks; when the tasks were performed successfully, the monkeys were rewarded with a sip of fruit juice. For example, to study the brain’s guidance of hand and arm movements, the monkey might be trained to move its hand toward the brightest of several spots on a computer screen. Pointing to the correct spot earned it a juice reward. After training, the animals were anesthetized. In a simple surgical procedure, each monkey was fitted with a small headpiece so that a microelectrode could be introduced into the brain through a small opening in the skull. When the animals recovered from surgery, they showed no signs of discomfort from either the headpiece or the insertion of a microelectrode into the brain (recall from Chapter 12 that there are no nociceptors in the brain). Evarts and his colleagues then recorded the discharges of individual cells in the motor cortex as the animals made voluntary movements. In the example above, one could then see how the neuron’s response changes when the animal points to different spots on the screen.
This is an example of what is now called behavioral neurophysiology, the recording of cellular activity in the brain of awake, behaving animals. By altering the task that the animal performs, the same method can be applied to the investigation of a wide range of neuroscientific topics, including attention, perception, learning, and movement. Some types of human neurosurgery are also done with the patient awake, at least during part of the procedure. By applying the techniques of behavioral neurophysiology to informed, consenting adults, we have also learned some fascinating information about uniquely human skills.
In recent years, technical developments have made it possible to insert large numbers of microelectrodes into the same or different parts of an animal’s brain and to record from dozens or even hundreds of neurons simultaneously. This approach yields a massive amount of information about brain activity and its relationship to behavior. Understanding this relationship is one of the greatest challenges in neuroscience.
You’ve heard the expression “ready, set, go.” The preceding discussion suggests that readiness (“ready”) depends on activity in the parietal and frontal lobes, along with important contributions from the brain centers that control levels of attention and alertness. “Set” may reside in the supplementary and premotor areas, where movement strategies are devised and held until they are executed. A good example is shown in Figure 14.9, based on the work of Michael Weinrich and Steven Wise at the National Institutes of Health. They monitored the discharge of a neuron in the PMA as a monkey performed a task requiring a specific arm movement to a target. The monkey was first given an instruction stimulus informing him what the target would be (“Get set, monkey!”), followed after a variable delay by a trigger stimulus informing the monkey that it was OK to move (“Go, monkey!”). Successful performance of the task (i.e., waiting for the “go” signal and then making the movement to the appropriate target) was rewarded with a sip of juice. The neuron in the PMA began firing if the instruction was to move the arm to the left, and it continued to discharge until the trigger stimulus came on and the movement was initiated. If the instruction was to move to the right, this neuron did not fire (presumably another population of PMA cells became active under this condition). Thus, the activity of this PMA neuron reported the direction of the upcoming movement and continued to do so until the movement was made. Although we do not yet understand the details of the coding taking place in the SMA and PMA, the fact that neurons in these areas are selectively active well before movements are initiated is consistent with a role in planning the movement.

FIGURE 14.9 The discharge of a neuron in the premotor area before a movement. (a) Ready: A monkey sits before a panel of lights. The task is to wait for an instruction stimulus that will inform him of the movement required to receive a juice reward, then perform the movement when a trigger stimulus goes on. The activity of a neuron in PMA is recorded during the task. (b) Set: The instruction stimulus (one of the square red lights) occurs at the time indicated by the upward arrow, resulting in the discharge of the neuron in PMA. (c) Go: A trigger stimulus (a blue light in one of the buttons) tells the monkey when and where to move. Shortly after the movement is initiated, the PMA cell ceases firing. (Source: Adapted from Weinrich and Wise, 1982.) Description
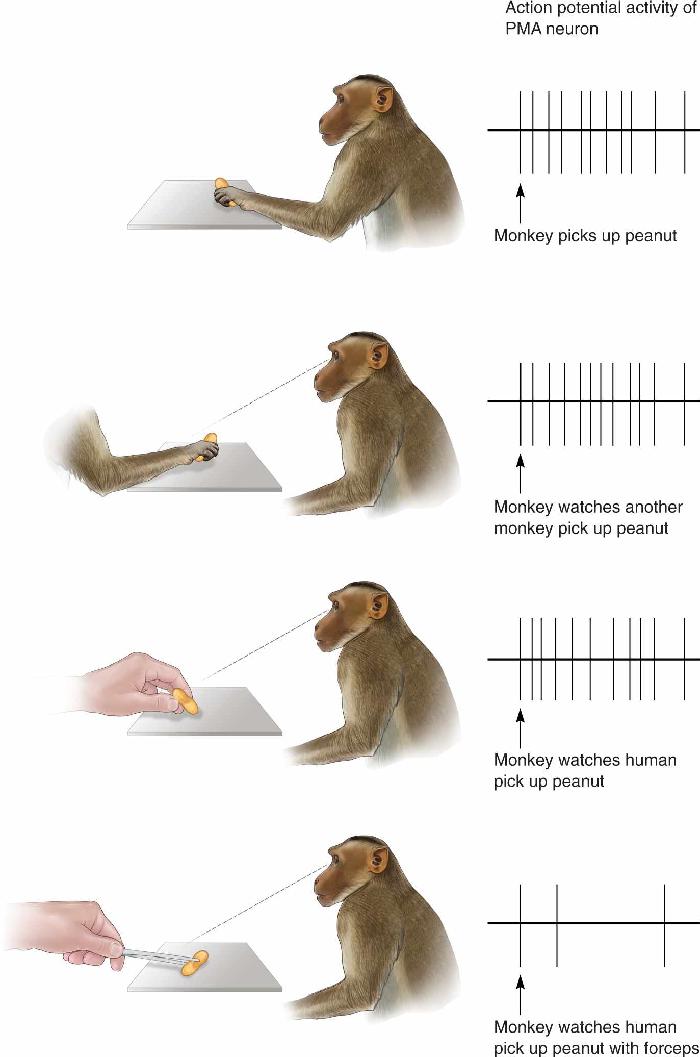
We mentioned previously that some neurons in cortical area 6 respond not only when movements are executed but also when the same movement is only imagined—mentally rehearsed. Remarkably, some neurons in motor areas of cortex fire not only when a monkey makes a specific movement himself but also when the monkey simply observes another monkey, or even a human, making the same type of movement (Figure 14.10). These cells were called mirror neurons by Giacomo Rizzolatti and his colleagues when they discovered them in the PMA of monkeys at the University of Parma in the early 1990s. Mirror neurons seem to represent particular motor acts, such as reaching, grasping, holding, or moving objects, regardless of whether a monkey actually performs the act or merely observes others doing it. Each cell has very specific movement preferences; a mirror neuron that responds when its monkey grasps a food tidbit will also respond to the sight of another monkey making a similar grasp of a tidbit but not when either monkey waves its hand. Many mirror neurons even respond to the unique sounds another monkey produces during a specific movement (e.g., cracking open a peanut), as well as to the sight of that movement. In general, mirror neurons seem to encode the specific goals of motor acts rather than particular sensory stimuli.

FIGURE 14.10 The discharge of a mirror neuron. (a) A PMA mirror neuron fires action potentials when a monkey reaches for a peanut. (b) The same mirror neuron fires when the monkey sees another monkey reach for a peanut. (c) The neuron also fires when the monkey sees a human reach for a peanut. (d) When the human reaches for a peanut using a forceps, the mirror neuron is not activated. (Source: Adapted from Rizzolatti et al., 1996.) Description
It is very likely that humans also have mirror neurons in PMA and other cortical areas, although the evidence for this, mainly from studies using functional magnetic resonance imaging (fMRI) (see Boxes 7.2 and 7.3), is still indirect.
Mirror neurons may be part of an extensive brain system for understanding the actions and even the intentions of others. This is an exciting and attractive hypothesis. It implies that we use the same motor circuits both for planning our own movements and for understanding the actions and goals of others. When one pitcher watches another pitcher throw a ball, the first pitcher may activate the same motor planning neurons that allow him to throw his own ball. In a sense, he may be experiencing the action of the other pitcher by running his own neural program for the same type of action. More expansive versions of this hypothesis suggest that mirror neurons are also responsible for our ability to read the emotions and sensations of others and to empathize. Some investigators have even suggested that dysfunctional mirror neurons are responsible for certain features of autism, such as the impaired ability to understand the thoughts, intentions, feelings, and ideas of others (see Box 23.4). As intriguing as these hypotheses about mirror neuron functions are, there is still scant evidence for any of them. As methods for recording directly from human neurons improve, it will be fascinating to test these ideas directly.
Now, let’s again consider our baseball pitcher standing on the mound. He has made the decision to throw a curve ball, but the batter abruptly walks away from the plate to adjust his helmet. The pitcher stands motionless on the mound, muscles tensed. He knows the batter will return, so he waits. The pitcher is “set”; a select population of neurons in the premotor and supplementary motor cortex (the cells that are planning the curve ball movement sequence) are firing away in anticipation of the throw. Then the batter steps up to the plate, and an internally generated “go” command is given. This command appears to be implemented with the participation of a major subcortical input to area 6, which is the subject of the next section. After that, we’ll examine the origin of the “go” command, the primary motor cortex.
The major subcortical input to area 6 arises in a nucleus of the dorsal thalamus, called the ventral lateral (VL) nucleus. The input to this part of VL, called VLo, arises from the basal ganglia buried deep within the telencephalon. The basal ganglia, in turn, are targets of the cerebral cortex, particularly the frontal, prefrontal, and parietal cortex. Thus, we have a loop where information cycles from the cortex through the basal ganglia and thalamus and then back to the cortex, particularly the supplementary motor area (Figure 14.11). One of the functions of this loop appears to be the selection and initiation of willed movements.

FIGURE 14.11 A summary of the motor loop from the cortex to the basal ganglia to the thalamus and back to area 6. Description
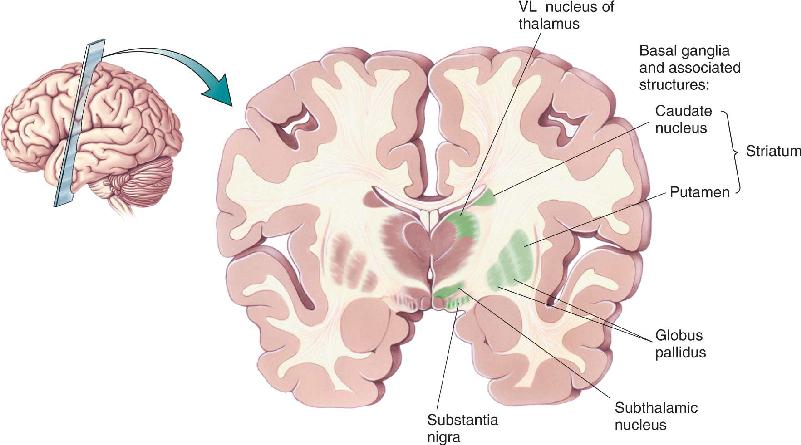
The basal ganglia consist of the caudate nucleus, the putamen, the globus pallidus (consisting of an internal segment, GPi, and an external segment, GPe), and the subthalamic nucleus. In addition, we can add the substantia nigra, a midbrain structure that is reciprocally connected with the basal ganglia of the forebrain (Figure 14.12). The caudate and putamen together are called the striatum, which is the target of the cortical input to the basal ganglia. The globus pallidus is the source of the output to the thalamus. The other structures participate in various side loops that modulate the direct path:

FIGURE 14.12 The basal ganglia and associated structures. Description
Through the microscope, the neurons of the striatum appear randomly scattered, with no apparent order such as that seen in the layers of the cortex. But this bland appearance hides a degree of complexity in the organization of the basal ganglia that we only partially understand. It appears that the basal ganglia participate in a large number of parallel circuits, only a few of which are strictly motor. Other circuits are involved in certain aspects of memory and cognitive function. We will try to give a concise account of the motor function of the basal ganglia, simplifying this very complex and poorly understood part of the brain.
Direct and Indirect Pathways through the Basal Ganglia
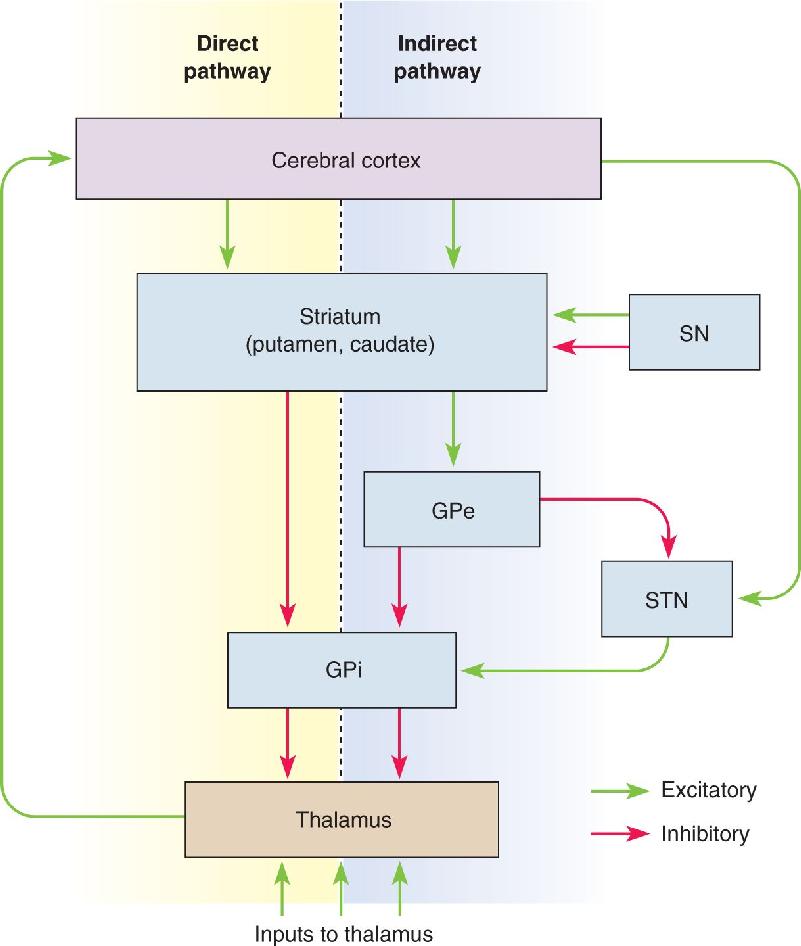
The motor loop through the basal ganglia originates with excitatory connections from the cortex. In the direct pathway through the basal ganglia, synapses from cortical cells excite cells in the putamen, which make inhibitory synapses on neurons in the globus pallidus, which in turn make inhibitory connections with the cells in VLo. The thalamocortical connection (from VLo to SMA) is excitatory and facilitates the discharge of movement-related cells in the SMA. This direct motor loop is summarized in Figure 14.13.

FIGURE 14.13 A wiring diagram of the basal ganglia motor loop. Synapses marked with a plus (+) are excitatory; those with a minus (–) are inhibitory. Description
In general, the direct pathway allows the basal ganglia to enhance the initiation of desired movements. Cortical activation of the putamen leads to excitation of the SMA by VL. Let’s work out how this happens. A critical point is that neurons in the internal segment of the globus pallidus are spontaneously active at rest, and therefore they tonically inhibit VL. Cortical activation (1) excites putamen neurons, which (2) inhibit GPi neurons, which (3) release the cells in VLo from inhibition, allowing them to become active. The activity in VLo boosts the activity of the SMA. Thus, this part of the circuit acts as a positive-feedback loop that may serve to focus, or funnel, the activation of widespread cortical areas onto the supplementary motor area of cortex. We can speculate that the “go” signal for an internally generated movement occurs when activation of the SMA is boosted beyond some threshold amount by the activity reaching it through this basal ganglia “funnel.”
There is also a complex indirect pathway through the basal ganglia that tends to antagonize the motor functions of the direct pathway. Information from the cortex flows through the direct and indirect pathways in parallel, and the outputs of both pathways ultimately regulate the motor thalamus (Figure 14.14). The most unique features of the indirect pathway are the GPe and the subthalamic nucleus. Striatal neurons inhibit cells of the GPe, which then inhibit cells of both the GPi and subthalamic nucleus. The subthalamic nucleus is also excited by axons from the cortex, and its projections excite the neurons of the GPi, which of course inhibit thalamic neurons.

FIGURE 14.14 The direct and indirect pathways through the basal ganglia. Dopaminergic neurons of the substantia nigra (SN) modulate the putamen and caudate nucleus. The GPe and the subthalamic nucleus (STN) are part of the indirect pathway.” Description
Whereas activation of the direct pathway by the cortex tends to facilitate the thalamus and information passing through it, activation of the indirect pathway by the cortex tends to inhibit the thalamus. In general, the direct pathway may help to select certain motor actions while the indirect pathway simultaneously suppresses competing, and inappropriate, motor programs.
Basal Ganglia Disorders. Studies of several human diseases have supported the view that the direct motor loop through the basal ganglia functions to facilitate the initiation of willed movements. According to one model, increased inhibition of the thalamus by the basal ganglia underlies hypokinesia, a paucity of movement, whereas decreased basal ganglia output leads to hyperkinesia, an excess of movement.
Parkinson’s disease exemplifies the first condition. This disorder, which affects about 1% of all people over age 60, is characterized by hypokinesia. Its symptoms include slowness of movement (bradykinesia), difficulty in initiating willed movements (akinesia), increased muscle tone (rigidity), and tremors of the hands and jaw, which are most prominent at rest when the patient is not attempting to move. Many patients also suffer deficits of cognition as the disease progresses. The organic basis of Parkinson’s disease is a degeneration of certain substantia nigra neurons and their inputs to the striatum (Box 14.3). These inputs use the neurotransmitter dopamine (DA). The actions of DA are complex because it binds to multiple types of striatal DA receptors that mediate quite different effects (see Figure 14.14). Dopaminergic synapses terminate on striatal neurons closely adjacent to the synaptic inputs from the cortex, and DA can enhance the cortical inputs to the direct pathway. DA facilitates the direct motor loop by activating cells in the putamen (which releases VLo from GPi-induced inhibition). In essence, the depletion of dopamine in Parkinson’s disease closes the funnel that feeds activity to the SMA via the basal ganglia and VLo. At the same time, DA inhibits the neurons in the striatum that send inhibitory outputs, via the indirect pathway, to the GPe.
Do Neurons in Diseased Basal Ganglia Commit Suicide?
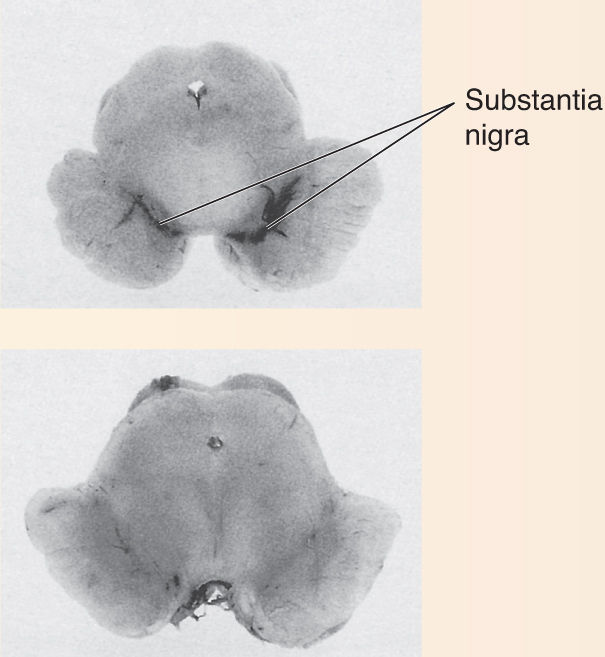
Several devastating neurological diseases involve the slow, progressive death of neurons. Patients with Parkinson’s disease have usually lost more than 80% of the dopamine-utilizing neurons in their substantia nigra (Figure A). Neurons of the striatum and other regions slowly degenerate in sufferers of Huntington’s disease (Figure B). Why do these neurons die? Ironically, it may be that natural forms of cell death are involved. A process called programmed cell death is essential for normal brain development; certain neurons commit suicide as part of the “program” by which the nervous system forms (see Chapter 23). All cells have several “death genes” that trigger a set of enzymes that destroy cellular proteins and DNA. Some forms of cancer occur when normal programmed cell death is prevented and cells proliferate wildly. Some neurological diseases may result when programmed cell death is unnaturally activated.

Figure A Normal (top); Parkinson’s disease (bottom). (Source: Strange, 1992, Fig. 10.3.)
Figure B Normal (left); Huntington’s disease (right). (Source: Strange, 1992, Fig. 11.2.)
Huntington’s disease is caused by a dominant gene that codes for a large brain protein called huntingtin. The normal molecule has a chain of 10–34 glutamines at one end, but people with a repeat of more than 40 glutamines develop Huntington’s disease. The abnormally long huntingtins aggregate; globs of them accumulate and trigger neuronal degeneration. The function of normal huntingtin is unknown, but it may counterbalance the triggers for programmed cell death. Thus, Huntington’s disease may arise from normal processes of neuronal degeneration gone awry.
Parkinson’s disease is usually a disease of aging, and the vast majority of cases occur after age 60. However, in 1976 and again in 1982, several relatively young drug abusers in Maryland and California developed severe Parkinsonian symptoms within a few days. This was extraordinary because usually, symptoms accumulate over many years. Medical detective work unraveled the cause of the addicts’ affliction. Each had taken street versions of a synthetic narcotic that contained the chemical MPTP. The incompetent basement chemists who had synthesized the illegal drug tried to shortcut the procedure, thereby creating a chemical by-product that kills dopaminergic neurons. MPTP has since helped us understand Parkinson’s disease better. We now know that MPTP is converted in the brain to MPP+; dopaminergic cells are selectively vulnerable to it because their membrane dopamine transporters mistake MPP+ for dopamine, and they selectively accumulate this chemical Trojan horse. Once inside the cell, MPP+ disrupts energy production in mitochondria, and the neurons apparently die because their ATP is depleted.
The effect of MPTP supports the idea that common forms of Parkinson’s disease might be caused by chronic exposure to a slowly acting toxic chemical in the environment. Unfortunately, no one has identified such a toxin. Research has shown that MPTP can induce a form of programmed neuronal death in the substantia nigra. Dopaminergic neurons of Parkinson’s patients may degenerate for a similar reason. About 5% of Parkinson’s cases are inherited, and mutations in several different genes are now known to cause these rarer types of the syndrome. One hypothesis is that Parkinsonian genes encode mutant proteins that are misfolded, aggregate, accumulate in neurons, and trigger or facilitate the death of dopaminergic neurons.
By understanding how and why neurons self-destruct, we may eventually be able to devise strategies of cellular suicide prevention that halt or avert a variety of terrible neurological diseases.
A central goal of most therapies for Parkinson’s disease is to enhance the levels of dopamine delivered to the caudate nucleus and putamen. This is most easily done by administering the compound L-dopa (L-dihydroxyphenylalanine, introduced in Chapter 6), which is a precursor to dopamine. L-dopa crosses the blood–brain barrier and boosts DA synthesis in the cells that remain alive in the substantia nigra, thus alleviating some of the symptoms. DA agonists are also useful drugs in the treatment of Parkinson’s disease. However, treatments with L-dopa or DA agonists do not alter the progressive course of the disease, nor do they alter the rate at which substantia nigra neurons degenerate. They also have significant side effects. (We will return to the topic of dopamine neurons in Chapter 15.) The symptoms of some Parkinson’s disease patients can also be improved with brain surgery and stimulation (Box 14.4). There are also a variety of experimental treatment strategies. One of them is to graft DA-producing cells into the basal ganglia. A promising possibility is to use human stem cells that have been manipulated developmentally or genetically to produce DA. These may one day provide an effective treatment, perhaps even a cure, for Parkinson’s disease, but we are not there yet.
Destruction and Stimulation: Useful Therapies for Brain Disorders
Brain disorders can be very difficult to treat, and useful therapies are often counterintuitive. Advanced Parkinson’s disease, for example, is sometimes treated with small surgical lesions of the brain or by implanting electrodes for deep brain stimulation (DBS). Destruction and stimulation are alternative strategies with the same therapeutic goal—to relieve patients of their severely abnormal movements.
The most common treatment for the early phase of Parkinson’s disease, L-dopa, can be tremendously helpful. Unfortunately, with time, the effects of the drug usually diminish, and new types of abnormal and debilitating movements, dyskinesias, may appear. Numerous other drugs can be useful at this stage, but their effectiveness varies and they have side effects of their own.
Surgery for movement disorders began in the 1880s with Victor Horsley, a pioneering British neurosurgeon, who treated a patient’s uncontrollable spontaneous movements by removing part of his motor cortex. The abnormal movements ceased, but the patient’s limb was paralyzed. Between the 1940s and 1970s, surgeons found that making small lesions in the globus pallidus, thalamus, or subthalamic nucleus could often improve the tremor, rigidity, and akinesia of Parkinson’s disease without inducing paralysis. With the introduction of L-dopa in 1968 and a backlash against unjustified types of neurosurgery (see Box 18.4), surgical treatments for Parkinson’s disease fell out of favor for a while. Currently, targeted surgical lesions of basal ganglia and thalamus are still used in some Parkinson’s patients, but DBS has become an increasingly popular form of treatment.
The ancient Greeks and Egyptians were early advocates of the therapeutic power of electrical shocks. Their medical devices were electric eels and rays, and it was said that direct application of such a stimulating fish could help alleviate pain and headache, hemorrhoids, gout, depression, and even epilepsy. The modern use of DBS for movement disorders began in the 1980s. Taking a cue from their experience with lesions, and noting the promising effects of stimulation in the operating room, surgeons began systematically testing whether high-frequency stimulation (DBS) could reduce abnormal movements over the long term. Several clinical trials have shown that it can. The U.S. Food and Drug Administration approved the use of DBS for treatment of Parkinson’s disease in 2002.
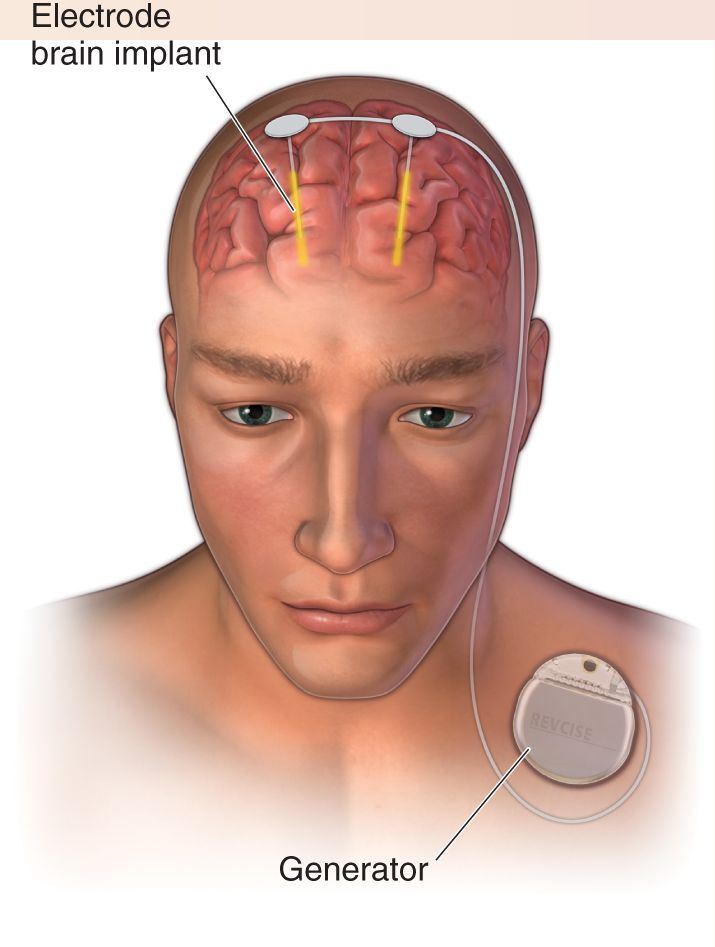
The current approach to DBS is to surgically implant bilateral electrodes with their tips in the subthalamic nuclei or, less often, in the GPi nuclei (Figure A). Advanced brain imaging methods, neuronal recordings, and trial stimulation are used in the operating room to ensure that the electrodes are placed precisely. Power and control of the electrodes come from small batteries and computers implanted under the skin below the collarbone. Postoperatively, therapists work with the patients to tune the properties of the stimulation for optimal effectiveness and minimal side effects.

Considering the complexities of the brain’s functions and dysfunctions, DBS is a very crude tool. The most effective stimulation pattern tends to be a continuous stream of brief shocks at very high frequency (130–180 Hz). Since this does not resemble any natural neural pattern in the brain, how does DBS work? Research on this question has been intense, but the answer remains unknown. High-frequency stimulation can block abnormal firing in some cases. Stimulation may also “jam” or suppress abnormal patterns of firing. DBS may activate inhibitory neurons that suppress dysfunctional brain activity. It may trigger release of neurotransmitters that modulate cells and synapses. The mechanism of DBS may also vary with the brain structure being stimulated. It would not be surprising if all of these effects, and more, were important for the efficacy of DBS.
DBS can be quite effective in controlling both hyperkinetic and hypokinetic symptoms and improving patients’ quality of life overall. It is not a panacea, however. DBS is not a useful treatment for most of the nonmotor features of the disease, including disturbances of cognition, mood, gait, and speech. There are also side effects and the usual risks of surgery. Batteries need to be surgically replaced every few years, although some DBS systems are now rechargeable.
DBS has therapeutic promise far beyond Parkinson’s disease. It can reduce the symptoms of several other movement disorders. It may be helpful for a surprising range of other psychiatric and neurological conditions, including major depression, obsessive-compulsive disorder, Tourette syndrome, schizophrenia, epilepsy, tinnitus, chronic pain, and Alzheimer’s disease. The best brain stimulation sites vary for each disorder. DBS is still an experimental treatment for nearly all these conditions, and only further research will determine whether its benefits outweigh its risks and costs.
If Parkinson’s disease lies at one end of the spectrum of basal ganglia disorders, Huntington’s disease lies at the other. Huntington’s disease is a hereditary, progressive, inevitably fatal syndrome characterized by hyperkinesia and dyskinesias (abnormal movements), dementia (impaired cognitive abilities), and a disorder of personality. Luckily, it is quite rare, afflicting 5–10 people per 100,000 worldwide. The disease is particularly insidious because its symptoms usually do not appear until well into adulthood. In the past, patients often unwittingly passed the gene on to their children before they knew they had the disease. It is now possible to perform a genetic test that reveals whether a person carries the Huntington gene. People with Huntington’s disease exhibit changes in mood, personality, and memory. The most characteristic sign of the disease is chorea—spontaneous, uncontrollable, and purposeless movements with rapid, irregular flow and flicking motions of various parts of the body. The most obvious pathology of their brains is a profound loss of neurons in the caudate nucleus, putamen, and globus pallidus, with additional cell loss in the cerebral cortex and elsewhere (see Box 14.3). The damage to structures in the basal ganglia and consequent loss of its inhibitory output to the thalamus seem to account for the disorders of movement in Huntington’s patients. Cortical degeneration is primarily responsible for their dementia and personality changes.
Hyperkinesia can also result from other types of lesions that affect the basal ganglia. One example is ballism, which is characterized by violent, flinging movements of the extremities (somewhat like our baseball pitcher unintentionally throwing the ball while sitting in the dugout). The symptoms usually occur on just one side of the body, and the condition is then called hemiballismus. As with Parkinson’s disease, the cellular mechanisms associated with ballism are known; it is caused by damage to the subthalamic nucleus (usually resulting from an interruption of its blood supply caused by a stroke). The subthalamic nucleus, part of another side loop within the basal ganglia, excites neurons in the globus pallidus that project to VLo (see Figure 14.14). Remember that excitation of the globus pallidus inhibits VLo (see Figure 14.13). Thus, a loss of excitatory drive to the globus pallidus facilitates VLo, in effect opening the funnel of activity to the SMA.
In summary, the basal ganglia may facilitate movement by focusing activity from widespread regions of cortex onto the SMA. Importantly, however, they also serve as a filter that keeps inappropriate movements from being expressed. We saw in Roland’s PET studies that activity in the SMA does not automatically trigger movement. The initiation of voluntary movement also requires activation of area 4, the subject of the next section.
THE INITIATION OF MOVEMENT BY PRIMARY MOTOR CORTEX
SMA is heavily interconnected with M1, cortical area 4 on the precentral gyrus (see Figure 14.7). The designation of area 4 as primary motor cortex is somewhat arbitrary because this is not the only cortical area that contributes to the corticospinal tract or to movement. Nonetheless, since the time of Sherrington, neuroscientists have recognized that this area has the lowest threshold for the elicitation of movement by electrical stimulation. In other words, stimulation intensities that are unable to evoke movement in other cortical areas are still effective in evoking movement when applied to area 4, meaning that area 4 has dense, strong synaptic connections with the motor neurons and the spinal interneurons that drive them. Focal electrical stimulation of area 4 evokes the contraction of small groups of muscles, and, as we discussed earlier, the somatic musculature is mapped systematically in this area. This ribbon of cortex that stretches the full length of the precentral gyrus is sometimes also called the motor strip.
The pathway by which motor cortex activates lower motor neurons originates in cortical layer V. Layer V has a population of pyramidal neurons, some of which can be quite large (soma diameters approaching 0.1 mm). The largest cells were first described as a separate class by Russian anatomist Vladimir Betz in 1874 and are therefore called Betz cells. In humans, many of the large corticospinal cells of layer V project to pools of lower motor neurons and excite them monosynaptically. The same corticospinal axons can also branch and excite local inhibitory interneurons. By controlling selected groups of motor neurons and interneurons, a single corticospinal neuron may generate coordinated effects on antagonist muscles. For example, the motor cortex neurons in Figure 14.15 excite pools of extensor motor neurons and simultaneously inhibit pools of flexor motor neurons. This is similar to the reciprocal inhibition that we saw in the spinal reflex circuitry in Chapter 13 (see Figure 13.25).

FIGURE 14.15 Corticospinal tract axons control pools of motor neurons. Large pyramidal neurons in layer V of the motor cortex project axons, via the corticospinal tract, to the ventral horns of the spinal cord. In this case, the axons directly excite pools of extensor motor neurons and indirectly (via interneurons) inhibit pools of flexor motor neurons that serve as antagonists of the extensors. (Source: Adapted from Cheney et al., 1985.) Description
The layer V pyramidal cells in M1 receive their inputs primarily from two sources: other cortical areas and the thalamus. The major cortical inputs originate in the areas adjacent to area 4: area 6 immediately anterior; and areas 3, 1, and 2 immediately posterior (see Figure 14.7). The thalamic input to M1 arises mainly from another part of the ventral lateral nucleus, called VLc, which relays information from the cerebellum. Besides projecting directly to the spinal cord, layer V pyramidal cells also send axon collaterals to many subcortical sites involved in sensorimotor processing, especially the brain stem.
Researchers previously thought the motor cortex consisted of a detailed mapping of the individual muscles, such that the activity of a single pyramidal cell would lead to activity in a single motor neuron pool. However, the view that has emerged from more recent work is that individual pyramidal cells can drive numerous motor neuron pools from a group of different muscles involved in moving a limb toward a desired goal. Recordings from M1 neurons in behaving animals have revealed that a burst of activity occurs immediately before and during a voluntary movement, and that this activity appears to encode two aspects of the movement: force and direction.
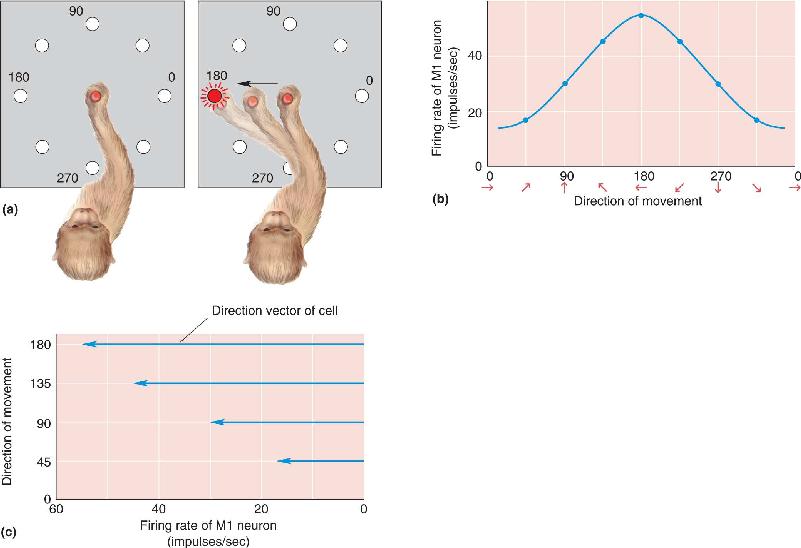
Because cortical microstimulation studies had suggested the existence of a fine-grained movement map in M1, the discovery that the movement direction tuning of individual M1 neurons is rather broad came as a surprise. This breadth of tuning is shown clearly in a type of experiment devised by Apostolos Georgopoulos and his colleagues, then working at Johns Hopkins University. Monkeys were trained to move a joystick toward a small light whose position varied randomly around a circle. Some M1 cells fired most vigorously during movement in one direction (180° in the example in Figure 14.16a) but also discharged during movement angles that varied considerably from the preferred direction. The coarseness in the directional tuning of the corticospinal neurons was certainly at odds with the high accuracy of the monkey’s movements, suggesting that the direction of movement could not be encoded by the activity of individual cells that command movement in a single direction. Georgopoulos hypothesized that movement direction was encoded instead by the collective activity of a population of neurons. Recall the role of neuronal population coding in the sensory systems, where the responses of many broadly tuned neurons are used to specify the properties of a particular stimulus (for example, see Chapter 8). Population coding in the motor system implies that groups of neurons are broadly tuned for the properties of movements.

FIGURE 14.16 Responses of an M1 neuron during arm movements in different directions. (a) As the monkey moves a handle toward a small light, the responses of an M1 neuron are monitored. When the monkey moves in directions around the clock, the relationship between the cell’s discharge rate and movement direction can be determined. (b) A tuning curve for an M1 neuron. This cell fires most during movements to the left. (c) Because the cell in part b responds best to leftward movement, it is represented by a direction vector pointing in that direction. The length of the vector is proportional to the firing rate of the cell. Notice that as the movement direction changes, the length of the direction vector changes. (Source: Adapted from Georgopoulos et al., 1982.) Description
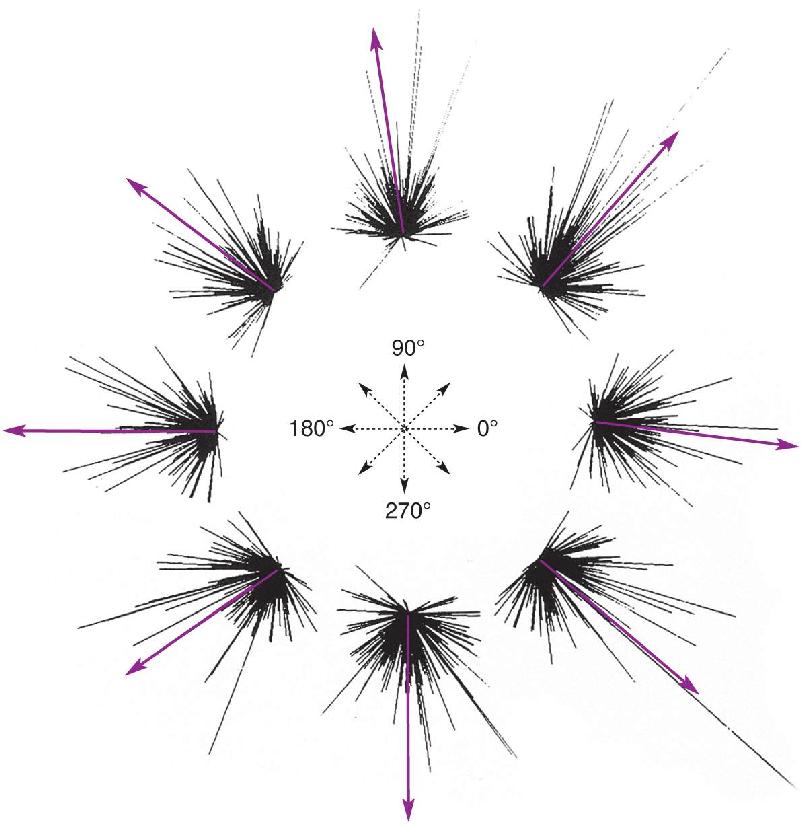
To test the feasibility of the idea of population coding for movement direction, Georgopoulos and his colleagues recorded from over 200 different neurons in M1; for each cell, they constructed a directional tuning curve such as that shown in Figure 14.16b. From these data, the researchers knew how vigorously each of the cells in the population responded during movement in each direction. The activity of each cell was represented as a direction vector pointing in the direction that was best for that cell; the length of the vector represented how active that cell had been during a particular movement (Figure 14.16c). The vectors representing each cell’s activity could be plotted together for each direction of movement, then averaged to yield what the researchers called a population vector (Figure 14.17). They found a strong correlation between this average vector, representing the activity of the entire population of M1 cells, and the actual direction of movement (Figure 14.18).

FIGURE 14.17 Direction vectors and population vectors. (a) The tuning curves for two cells in the motor cortex (see Figure 14.16). Both cells fire during movement in a range of directions, but cell 1 fires best when movement is upward, while cell 2 responds best when movement is from left to right. (b) The response of each cell is represented as a direction vector, which points in the preferred direction for the neuron, but its length depends on the number of action potentials the cell fires during movement over a range of directions. For any direction, the direction vectors of individual cells are averaged to yield a population vector, reflecting the strength of the response of both cells during this movement. Description

FIGURE 14.18 Predicting the direction of movement by population vectors. Each cluster of lines reflects the direction vectors of many cells in M1. Line length reflects the discharge rate of each cell during a movement in one of eight different directions. Arrows represent the average population vectors, which predict the movement direction of the monkey’s arm. (Source: Georgopoulos et al., 1983.) Description
These studies suggest three important conclusions about how M1 commands voluntary movement: (1) much of the motor cortex is active for every movement, (2) the activity of each cell represents a single “vote” for a particular direction of movement, and (3) the direction of movement is determined by a tally (and averaging) of the votes registered by each cell in the population. Although this population-coding scheme remains hypothetical in M1, experiments on the superior colliculus by James McIlwain at Brown University and David Sparks at the University of Alabama showed conclusively that a population code is used by this structure to command precisely directed eye movements (Box 14.5).
During the 1960s and 1970s, it became relatively easy to record the electrical activity of single neurons in the brain. The power and promise of this method led to the idea of the “feature-detector neuron,” whose discharge was thought to announce the presence of the stimulus feature to which it was most sensitive. This view rarely tempted students of olfaction and gustation because they had found that the discharge of single chemosensory neurons is highly ambiguous with respect to the identity of the stimulus. Not so in vision research, where the cells with the smallest spatial receptive fields and the most refined preferences for specific stimuli seemed most interesting. Any neuron responsive to a range of stimuli was regarded as crudely selective and unsuited to processes requiring high resolution.
Neurophysiologists who examined visual areas of the brain stem soon encountered a paradox. The receptive fields of cells in the superior colliculus turned out to be very large, yet this structure was clearly important for the execution of highly accurate saccadic eye movements, which change the direction of gaze to a stimulus of interest. The superior colliculus receives orderly input from both the retina and the visual cortex, and damage to it impairs an animal’s ability to direct its gaze at novel stimuli. Focal electrical stimulation of the colliculus evokes saccades whose directions and amplitudes are correlated with the visual receptive fields of cells at the stimulus site. A small change in the position of the stimulating electrode results in small changes in the saccade’s direction and amplitude. Certain collicular neurons discharge in association with saccadic eye movements, as if they are part of the control mechanism that specifies the dimensions of the movement. This activity occurs in association with saccades that terminate across a restricted zone of visual space called the movement field of the cell, by analogy to the sensory receptive field. How could such cells specify the target of a saccade with any accuracy if their movement fields and visual receptive fields are very large?
The answer began to emerge from experiments on the cat’s superior colliculus in my laboratory at Brown University and from studies of the primate’s superior colliculus by David Sparks, then at the University of Alabama. We asked the inverse of the traditional receptive field question or movement field question. From the sensory side, instead of asking where a point of light must be located to activate a collicular neuron, we asked where in the superior colliculus are the cells that have the point in their receptive fields—that “see” the point. Similarly, the key consideration on the motor side is the location of the cells that discharge before a saccade to a given target, rather than the size of their individual movement fields. The analyses from both laboratories revealed that these regions of activity are widespread, occupying considerable fractions of the collicular tissue. As the stimulus or target location moves around in visual space, the corresponding patch of neural activity moves around in the superior colliculus.
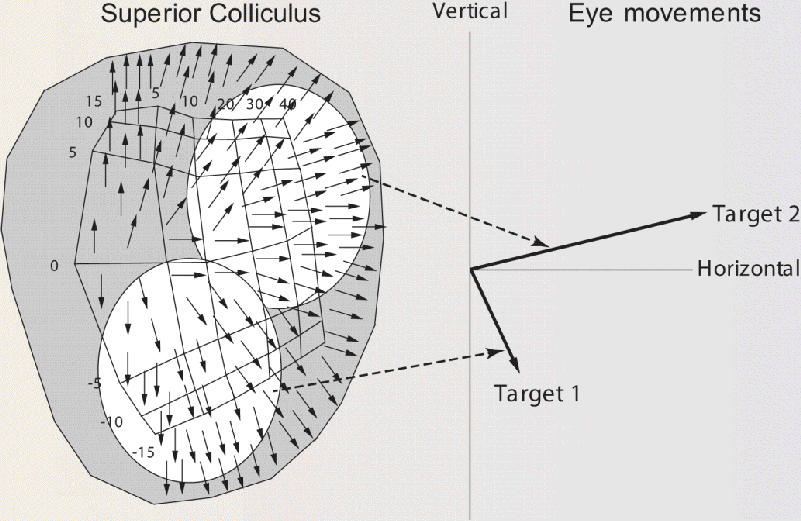
A general idea of how a system of such neurons may encode a saccade is shown in Figure A. On the left, each arrow on the retinotopic map of the superior colliculus symbolizes the contribution of its location to the code for the direction of a saccade. The more closely packed the arrows, the stronger the signal from that region to brain stem circuits that shape the motor commands for the saccade. The distribution of the arrows is consistent with the effects of focal electrical stimulation. Thus, for example, stimulation at successively lower points in the lower half of the map (representing the lower visual field) leads to downwardly directed saccades of increasing amplitude. If the appearance of target 1 in the right side of the figure excited cells in the lower unshaded oval of the map, the collective activity of the cells would specify the downward and horizontal components needed for a saccade to the target. The appearance of target 2 would activate the upper unshaded area, assembling the components of the correct, upwardly directed saccade. In this model, changes in the position of the target modify the composition of the output signal to yield a saccade matching any position of the target.

The large size of the receptive fields and movement fields of collicular neurons means that information about the location of a visual point or a saccade target is distributed among many neurons. The model of Figure A shows how the active population of collicular neurons may indicate the target’s position as a motor code. The simple and incomplete model shown here represents only one of several ideas about how the superior colliculus accomplishes its task. It seems certain, though, that target location and saccade measurements are encoded in the distribution of activity across a population of neurons.
The Malleable Motor Map. This scheme for motor control leads to an interesting prediction: the larger the population of neurons representing a type of movement, the finer the possible control. From the motor map shown in Figure 14.8, we would predict that finer control should be possible for the hands and the muscles of facial expression, and indeed this is normally the case. Of course, fine movements of other muscles can be learned with experience; consider the finger, wrist, elbow, and shoulder movements of an accomplished cellist. Does this mean that cortical cells in M1 can switch allegiance from participation in one type of movement to another as skills are learned? The answer appears to be yes. John Donoghue, Jerome Sanes, and their students at Brown University collected evidence indicating that such plasticity of the adult motor cortex is possible. For example, in one series of experiments, they used cortical microstimulation in rats and mapped the regions of M1 that normally elicit movements of the forelimb, facial whiskers, or muscles around the eye (Figure 14.19a). Then they cut the motor nerve that supplies the muscles of the snout and its whiskers and found that regions of M1 that had evoked whisker movements now would elicit either forelimb or eye movements (Figure 14.19b). The motor map had been reorganized. These neuroscientists speculated that similar types of cortical reorganization might provide a basis for learning fine motor skills.

FIGURE 14.19 Malleable motor maps. (a) This map represents motor cortex from a normal rat. (b) This map represents a rat that had the motor nerve cut that supplies the whiskers (vibrissae). Notice the cortical regions that previously evoked movement of the vibrissae now cause muscle movement in the forelimb or around the eyes (periocular). (Source: Adapted from Sanes and Donoghue, 1997.) Description
From the preceding discussion, we can imagine that when the time has come for our baseball pitcher to wind up, his motor cortex generates a torrent of activity in the pyramidal tract. What could appear to be a discordant voice to a neurophysiologist recording from a single M1 neuron is part of a clear chorus of activity to the spinal motor neurons that generate the precise movements necessary to propel the baseball accurately.
It is not enough to simply command the muscles to contract. Throwing a ball requires a detailed sequence of muscle contractions, each one generating exactly the right amount of force at precisely the right time. These critical motor control functions belong to the cerebellum (introduced in Chapter 7). The importance of the cerebellum in this aspect of motor control is plainly revealed by cerebellar lesions; movements become uncoordinated and inaccurate, a condition known as ataxia.
Take this simple test. Lay your arms in your lap for a moment, then touch your nose with one finger. Try it again with your eyes closed. No problem, right? Patients with cerebellar damage are often incapable of performing this simple task. Instead of smoothly and simultaneously moving the shoulder, elbow, and wrist to bring the finger to rest on the nose, they move each joint sequentially—first the shoulder, then the elbow, and finally the wrist. This is called dyssynergia, decomposition of synergistic multijoint movement. Another characteristic deficit shown by these patients is that their finger movement will be dysmetric; they will either come up short of the nose or shoot past it, poking themselves in the face. You may recognize these symptoms as similar to those that accompany ethanol intoxication. Indeed, much of the clumsiness that accompanies alcohol abuse is a direct consequence of the depression of cerebellar circuits (Box 14.6).
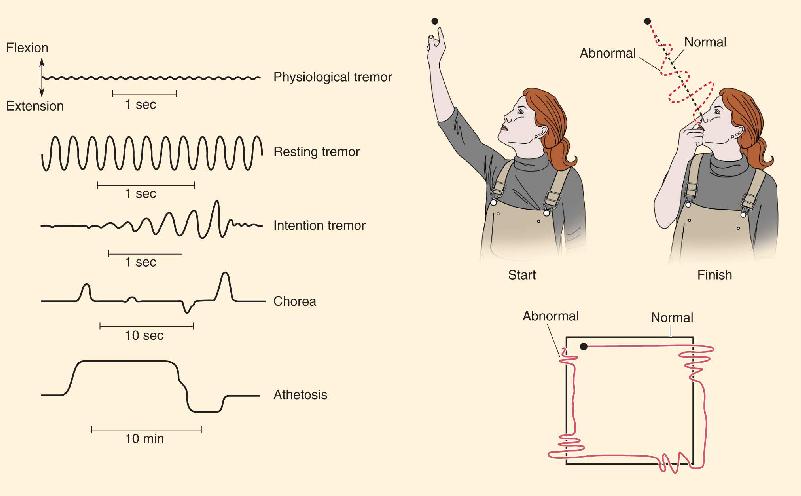
Raise your hand in front of your face, and try to hold it as still as possible. You will see a very slight trembling of your fingers. This is called physiological tremor, a small, rhythmic oscillation of about 8–12 Hz. It is perfectly normal, and there is nothing you can do to stop it short of resting your hand on the table. A variety of everyday circumstances—stress, anxiety, hunger, fatigue, fever, too much caffeine—can enhance the tremor.
As we have discussed in this chapter, some neurological diseases lead to more dramatic involuntary movements, with distinctive characteristics. Parkinson’s disease is often associated with a large resting tremor of about 3–5 Hz. Movement is at its worst when the patient is not attempting to move. Strangely enough, the tremor immediately disappears during a voluntary movement. On the other hand, people with cerebellar damage have no abnormal tremor at rest but often show dramatic intention tremor when they try to move. Cerebellar tremor is an expression of ataxia, the uncoordinated contractions of the muscles used in the movement. For example, as the patient tries to move her finger from one point in space to another, or track a pathway with her finger, she makes large errors; in trying to correct an error, she makes more errors, and so on, as the finger wobbles inaccurately toward its destination.
Huntington’s disease causes chorea (from the Greek for “dance”)—quick, irregular, involuntary but relatively coordinated movements of the limbs, trunk, head, and face. Other types of basal ganglia disease can lead to athetosis—much slower, almost writhing movements of the neck and trunk. The unique properties of each abnormal movement can help in the diagnosis of neurological diseases, and they have taught us volumes about the normal functions of the damaged parts of the brain.

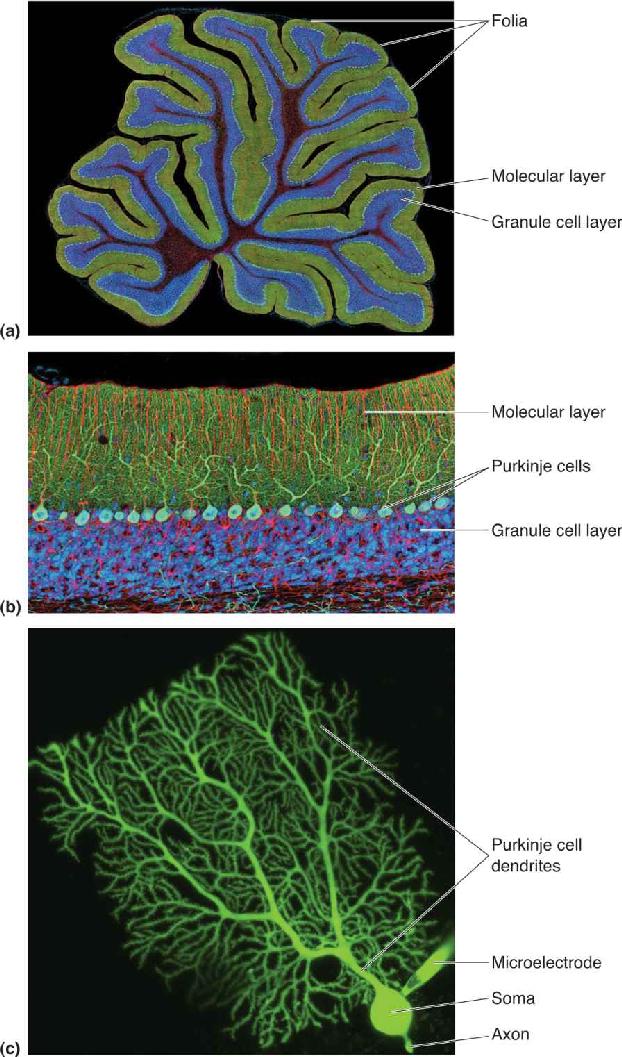
Cerebellar anatomy is shown in Figure 14.20. The cerebellum sits on stout stalks of axons called peduncles that rise from the pons; the whole structure resembles a piece of cauliflower. The visible part of the cerebellum is actually a thin sheet of cortex, which is repeatedly folded. The dorsal surface is characterized by a series of shallow ridges called folia (singular: folium), which run transversely (from side to side). In addition, there are deeper transverse fissures, revealed by making a sagittal slice through the cerebellum; these divide the cerebellum into 10 lobules. Together, folia and lobules serve to greatly increase the surface area of the cerebellar cortex, as the gyri of the cerebrum do for the cerebral cortex. Neurons are also embedded deep within the white matter of the cerebellum, forming the deep cerebellar nuclei, which relay most of the cerebellar cortical output to various brain stem structures. The cerebellum constitutes only about 10% of the total volume of the brain, but its cortex has an astonishingly high density of neurons. The vast majority of these are tiny excitatory neurons called granule cells, whose somata lie in the granule cell layer (Figure 14.21a,b). The number of granule neurons in the cerebellum is about equal to the number of other neurons in the entire CNS. The largest neuron in the cerebellar cortex is the inhibitory Purkinje cell, which receives excitatory input from granule cells in the molecular layer and sends its inhibitory axons to the deep cerebellar nuclei (Figure 14.21c).

FIGURE 14.20 The cerebellum. (a) A dorsal view of the human cerebellum, showing the vermis and hemispheres. (b) A midsagittal view of the brain, showing the lobules of the cerebellum. (c) A cross section of the cerebellum, showing the cortex and deep nuclei. Description

FIGURE 14.21 Neurons of the cerebellar cortex. (a) A histological section through the folia of the cerebellar cortex. Fluorescent stains color the molecular layer green and the granular cell layer blue. (b) A close-up of the layers of the cerebellar cortex. (c) A Purkinje cell after it has been injected with a fluorescent dye through the tip of a microelectrode. (Source: Parts a and b adapted from Tom Deerinck and Mark Ellisman, National Center for Microscopy and Imaging Research; part c adapted from Tetsuya Tatsukawa, RIKEN Brain Science Institute, Wako, Japan) Description
Unlike the cerebrum, the cerebellum is not obviously split down the middle. At the midline, the folia appear to run uninterrupted from one side to the other. The only distinguishing feature of the midline is a bump that runs like a backbone down the length of the cerebellum. This midline region is called the vermis (from the Latin for “worm”), and it separates the two lateral cerebellar hemispheres from each other. The vermis and the hemispheres represent important functional divisions. The vermis sends output to the brain stem structures that contribute to the ventromedial descending spinal pathways, which, as already discussed, control the axial musculature. The hemispheres are related to other brain structures that contribute to the lateral pathways, particularly the cerebral cortex. For the purpose of illustration, we’ll focus on the lateral cerebellum, which is particularly important for limb movements.
The simplest circuit involving the lateral cerebellum constitutes yet another loop, shown schematically in Figure 14.22. Axons arising from layer V pyramidal cells in the sensorimotor cortex—frontal areas 4 and 6, somatosensory areas on the postcentral gyrus, and the posterior parietal areas—form a massive projection to clusters of cells in the pons, the pontine nuclei, which in turn feed the cerebellum. To appreciate the size of this pathway, consider that the corticopontocerebellar projection contains about 20 million axons; that’s 20 times more than in the pyramidal tract! The lateral cerebellum then projects back to the motor cortex via a relay in the ventral lateral nucleus of the thalamus (VLc).

FIGURE 14.22 A summary of the motor loop through the cerebellum. Description
From the effects of lesions in this pathway, we can deduce that it is critical for the proper execution of planned, voluntary, multijoint movements. Indeed, once the cerebellum has received the signal for movement intent, its activity appears to instruct the primary motor cortex with respect to movement direction, timing, and force. For ballistic movements, these instructions are based entirely on predictions about their outcome (because such movements are too fast for sensory feedback to be of much immediate use). Such predictions are based on past experience; that is, they are learned. Therefore, the cerebellum is another important site for motor learning; it is a place where what is intended is compared with what has happened. When this comparison fails to meet expectations, compensatory modifications are made in certain cerebellar circuits.
Programming the Cerebellum. We will return to cerebellar circuitry and how it is modified by experience in Chapter 25. But for now, think about the process of learning a new motor skill (e.g., skiing, piano playing, juggling, knitting, throwing a curve ball). Early on, you must concentrate on new movements, and you tend to perform them in a disjointed and uncoordinated fashion. Practice makes perfect, however. As you master the skill, the movements will become smooth, and eventually you will be able to perform them almost unconsciously. This process represents the creation of a new motor program that generates the appropriate movement sequences on demand without the need for conscious control.
Recall that the word “cerebellum” derives from the Latin for “little brain.” The cerebellum acts as a small brain within the brain, unconsciously determining that the programs for skilled movement are executed properly and are adjusted whenever their execution fails to meet expectations.
Let’s return to the example of the baseball pitcher one last time to put the different pieces of the motor control puzzle together. Imagine the pitcher walking to the mound. The spinal circuits of the crossed extensor reflex are engaged and coordinated by descending commands on the ventromedial pathways. Extensors contract, flexors relax; flexors contract, extensors relax.
Once on the mound, the pitcher is joined by the umpire. Into his outstretched hand the umpire drops a new baseball. The added weight stretches the flexors of the arm. Group Ia axons become more active and cause monosynaptic excitation of the motor neurons innervating the flexors. The muscles contract to hold the ball up against gravity.
He is now ready to pitch. His neocortex is fully engaged and active as he looks at the catcher for the hand signal that tells him the type of pitch to throw. At the same time, the ventromedial pathways are working to maintain his standing posture. Although his body is still, the neurons of the ventral horns of the spinal cord are firing steadily under the influence of the ventromedial pathways, keeping the extensors of the lower leg activated.
The catcher flashes the sign for the curve ball. The sensory information is communicated from the occipital cortex to the parietal and prefrontal cortex. These regions of cortex along with area 6 begin planning the movement strategy.
The batter steps up to the plate and is ready. Activity cycling through the basal ganglia increases, triggering the initiation of the pitch. In response to this input to the cerebral cortex, SMA activity increases, followed immediately by the activation of M1. Now instructions are sweeping down the axons of the lateral pathways. The cerebellum, activated by the corticopontocerebellar inputs, uses these instructions to coordinate the timing and force of the descending activity so the proper sequence of muscle contractions can occur. Cortical input to the reticular formation leads to the release of the antigravity muscles from reflex control. Finally, lateral pathway signals engage the motor neurons and interneurons of the spinal cord, which cause muscles of the arms and legs to contract.
The pitcher winds up and throws. The batter swings. The ball sails over the left-field fence. The crowd jeers; the manager curses; the team owner frowns. Even as the pitcher’s cerebellum goes to work making adjustments for the next pitch, his body reacts. His face flushes; he sweats; he’s angry and anxious. But these latter reactions are not the stuff of the somatic motor system. These are topics of Part III, The Brain and Behavior.
The Initiation of Movement by the Primary Motor Cortex
1. List the components of the lateral and ventromedial descending spinal pathways. Which type of movement does each path control?
2. You are a neurologist presented with a patient who has the following symptom: an inability to independently wiggle the toes on the left foot, but with all other movements (walking, independent finger movement) apparently intact. You suspect a lesion in the spinal cord. Where?
3. PET scans can be used to measure blood flow in the cerebral cortex. What parts of the cortex show increased blood flow when a subject is asked to think about moving her right finger?
4. Why is L-dopa used to treat Parkinson’s disease? How does it act to alleviate the symptoms?
5. Individual Betz cells fire during a fairly broad range of movement directions. How might they work together to command a precise movement?
6. Sketch the motor loop through the cerebellum. What movement disorders result from damage to the cerebellum?
Alstermark B, Isa T. 2012. Circuits for skilled reaching and grasping. Annual Review of Neuroscience 35:559–578.
Blumenfeld H. 2011. Neuroanatomy Through Clinical Cases, 2nd ed. Sunderland, MA: Sinauer.
Donoghue J, Sanes J. 1994. Motor areas of the cerebral cortex. Journal of Clinical Neurophysiology 11:382–396.
Foltynie T, Kahan J. 2013. Parkinson’s disease: an update on pathogenesis and treatment. Journal of Neurology 260:1433–1440.
Glickstein M, Doron K. 2008. Cerebellum: connections and functions. Cerebellum 7:589–594.
Graziano M. 2006. The organization of behavioral repertoire in motor cortex. Annual Review of Neuroscience 29:105–134.
Lemon RN. 2008. Descending pathways in motor control. Annual Review of Neuroscience 31:195–218.
Rizzolatti G, Sinigaglia C. 2008. Mirrors in the Brain: How Our Minds Share Actions and Emotions. New York: Oxford University Press.
Additional figures