Sex and the Brain
The Control of Sex Hormones by the Pituitary and Hypothalamus

BOX 17.1 PATH OF DISCOVERY: Bonding with Voles, by Thomas Insel
BOX 17.2 OF SPECIAL INTEREST: Bird Songs and Bird Brains
Direct Genetic Effects on Behavior and Sexual Differentiation of the Brain
BOX 17.3 OF SPECIAL INTEREST: David Reimer and the Basis of Gender Identity
Brain Changes Associated with Maternal and Paternal Behavior
Estrogen Effects on Neuron Function, Memory, and Disease
Without sex, there is no human reproduction. And without offspring, no species can survive. Those are the simple facts of life, and over millions of years, the human nervous system evolved for the survival of the species. The drive to reproduce can be compared to the powerful motivation to eat or drink, which we discussed in Chapter 16. For the sake of survival, life-maintaining functions, such as reproduction and eating, are not left entirely to the whims of conscious thought. They are regulated by subcortical structures, and thoughtful conscious control is provided by the cerebral cortex.
In this chapter, we will explore what is known about sex and the brain. Our goal is not a discussion of the birds and the bees; we will assume you have picked up the basics about human sexual behavior from your parents, teachers, friends, or the Internet. Instead, we will look at the neural machinery that makes reproduction possible. For the most part, the neural control of sexual organs uses the same somatosensory and motor pathways we have examined in earlier chapters. Sexual and reproductive behaviors are clearly different in men and women, but just how different are the brains of the two sexes? We will explore this question and see whether brain differences pertain to only reproductive behaviors or more generally to behavior and cognition.
Ultimately, the origin of most distinctions between males and females is the genes inherited from the parents. Under the guidance of certain genes, the human body produces a small number of sex hormones that have powerful effects on the sexual differentiation of the body, as well as adult sexual physiology and behavior. The reproductive organs (the ovaries and testes), which secrete sex hormones, are outside the nervous system, but they are activated by the brain. Recall from Chapter 15 that the hypothalamus controls the release of diverse hormones from the anterior pituitary. In the case of reproductive function, the hormones released by the anterior pituitary regulate secretions from the ovaries and testes. Sex hormones have obvious effects on the human body, but they also influence the brain. There appear to be effects of hormones on gross brain structure and also at the level of neurites on individual neurons. Sex hormones may even influence resistance to certain neurological diseases.
Another point to consider is what it means to be male or female. Is gender determined by genetics, anatomy, or behavior? The answer is not simple; there are cases of gender identities that do not correlate with biological and behavioral factors. And what about sexual orientation? Is an attraction to members of the opposite or same sex determined by experiences in childhood or the structure of the brain? These are challenging questions that address how we perceive ourselves and others. We will examine the extent to which we can answer such questions by looking at the anatomy and physiology of the nervous system.
The words sex and gender both concern distinctions between male and female and are often used synonymously. However, there is disagreement about the meanings of the two terms and the distinction between them. For the sake of clarity, our starting point is the definitions accepted by the World Health Organization. Thus, sex is the biological state of being male or female, and it is determined by chromosomes, hormones, and body anatomy (Figure 17.1). Gender is the set of behaviors and attributes a culture associates with men and women (i.e., masculine and feminine). Of course, it is not always easy or possible to determine whether an expectation or behavior of men and women is a consequence of biology (nature), society (nurture), or both. And, as we will discuss, there are situations in which sex and gender assessments conflict.

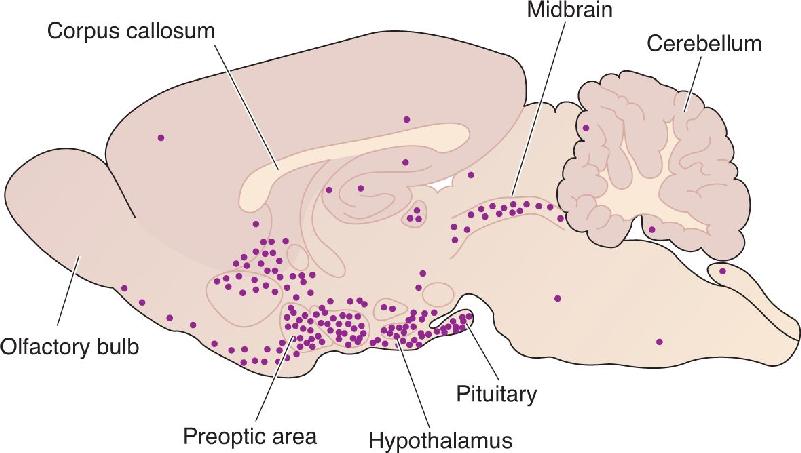
FIGURE 17.1 Biological and behavioral gender differences. Pheasants are just one of countless animal examples of highly divergent sex traits. The male is dramatically colored and large with a long tail and wattle; it plays little role in raising offspring. The female is small and brownish and an involved mother. (Source: ChrisO at the English Wikipedia.)
The behavioral and cultural implications of a person’s sex start at birth. With a newborn we ask the parents, “Is the baby a boy or a girl?” The answer to this question often leads to innumerable assumptions about the life experiences the child will have. We don’t typically inquire about an adult’s sex because it’s usually obvious from appearance. However, identifying someone as female or male still involves many assumptions, as our ideas about sex and gender are associated with numerous biological and behavioral traits. Gender-specific behaviors result from complex interactions among introspection, upbringing, life experiences, societal expectations, genetics, and hormones. These behaviors are related to gender identity—our perception of our own gender. In this section, we will discuss some of the genetic and developmental origins of sex.
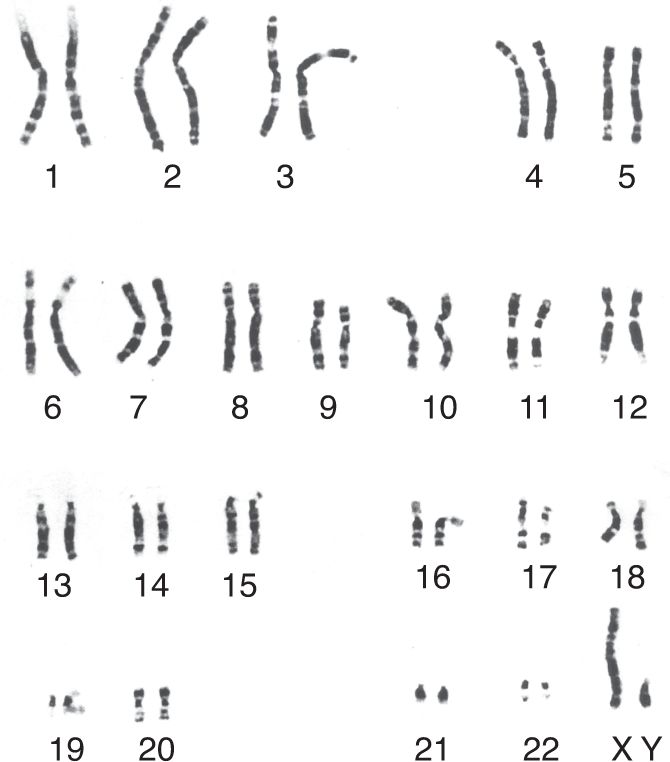
Within the nucleus of every human cell, DNA provides a person’s genetic blueprint, all the information needed to build an individual. The DNA is organized into 46 chromosomes: 23 from the father and 23 from the mother. Each of us has two versions of the chromosomes 1 through 22, conventionally numbered in order of decreasing size (Figure 17.2). The only exceptions to this pair system are the sex chromosomes, X and Y. Thus, it is usually stated that there are 44 autosomes (22 pairs of matching chromosomes) and two sex chromosomes. Females have two X chromosomes, one from each parent. Males have an X chromosome from the mother and a Y chromosome from the father. Therefore, the female genotype is denoted XX and the male genotype XY. These genotypes specify a person’s genetic sex. Because the mother contributes an X chromosome to every child regardless of sex, the child’s genetic sex is determined by the X or Y contribution from the father. In some nonhuman animals, such as birds, it is instead the mother’s contribution that determines the genetic sex of the offspring.

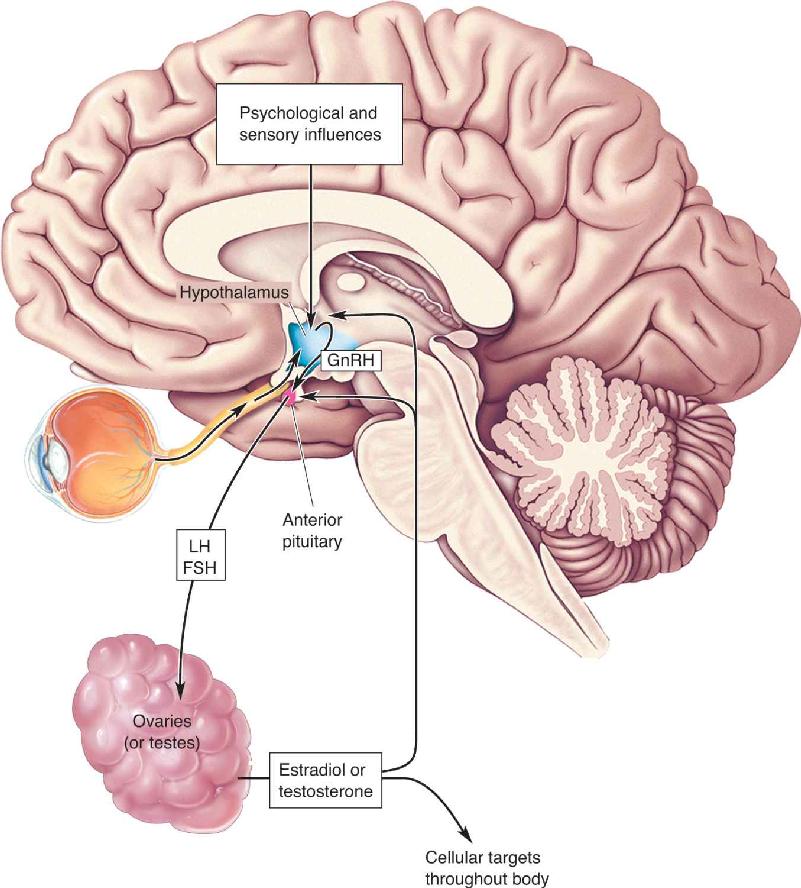
FIGURE 17.2 Human chromosomes. These 23 pairs of chromosomes are from a man. Notice how much smaller the Y chromosome is than the X chromosome. (Source: Yunis and Chandler, 1977.)
The DNA molecules that make up chromosomes are some of the largest molecules known, and they contain genes, the basic units of hereditary information. The piece of DNA comprising a single gene provides the unique information needed to construct a particular protein. There are about 25,000 genes in the human genome, although the number varies depending on the technique used to make the estimate (see Box 2.2).
As you can see in Figure 17.2, the X chromosome is significantly larger than the Y chromosome. Corresponding to this size difference, scientists estimate that the X chromosome contains about 800 genes, whereas the Y chromosome probably contains about 50. You might joke that men are genetically shortchanged, and in a sense, that’s right: The XY genotype has serious medical consequences. If a female has a defective gene on an X chromosome, she may experience no negative consequence if her gene on the other X chromosome is normal. However, any defect in the single X chromosome of a male can lead to a developmental defect. Such a defect is called an X-linked disease, and there are many. For example, red–green color blindness is relatively common in males (see Box 9.5). Other X-linked diseases that occur more often in men than women are hemophilia and Duchenne muscular dystrophy.
Compared to the X chromosome, the smaller Y chromosome has fewer genes and less diverse functions. Most importantly for sex determination, it contains a gene called the sex-determining region of the Y chromosome (SRY), which codes for a protein called testis-determining factor (TDF). A human with a Y chromosome and the SRY gene develops as a male, and without it, the individual develops as a female. The SRY gene was found to be located on the short arm of the Y chromosome in 1990 by Peter Goodfellow, Robin Lovell-Badge, and their colleagues at the Medical Research Council in London (Figure 17.3). If this bit of the Y chromosome is artificially incorporated into the DNA of a fetal XX mouse, the mouse will develop as a male instead of a female. However, this doesn’t mean that SRY is the only gene involved in sex determination, as SRY is known to regulate genes on other chromosomes. Also, male-specific physiology, such as sperm production, relies on other genes on the Y chromosome. Nonetheless, we will see shortly that expression of the SRY gene causes the development of the testes, and the hormones from the testes are largely responsible for making a male fetus develop differently from a female fetus.

FIGURE 17.3 The location of the SRY gene on the Y chromosome. In 1959, researchers found that TDF depended on the Y chromosome, and in 1966, the important location was further restricted to the short (p) arm. Research in the 1980s established that TDF is coded by the SRY gene, a small segment near the tip of the short arm of the Y chromosome. (Source: Adapted from McLaren, 1990, p. 216.) Description
Sex Chromosome Abnormalities. In rare cases, a person has too few or too many sex chromosomes, with health consequences that range from minimal to lethal. Turner syndrome is a partial or complete absence of one X chromosome in a female (XO genotype), affecting about one in 2500 female births. Miscarriage is thought to occur with most XO fetuses. The girls who survive have a variety of characteristics, including short stature, a receding jaw, a webbed neck, and visuospatial and memory difficulties. Their ovaries are abnormal, and estrogen replacement therapy is generally needed for breast development and menstruation. Presumably because the loss of the X chromosome in a male is lethal, there are no known individuals with a YO genotype.
In some cases, people are born with additional sex chromosomes. When this occurs, the sex is always determined by the presence or absence of the Y chromosome. In about one in 1000 male births, there is an extra X chromosome; this defect is known as Klinefelter syndrome. These XXY individuals are male because of the presence of the SRY gene on the Y chromosome. In some cases, there are no obvious indications of the XXY genotype, but possible symptoms include a less muscular body, less body hair, and increased breast tissue because of lower testosterone production. XYY and XXYY genotypes also occur, and these individuals are male, whereas an XXX person is female.
Differences between males and females are numerous, from average body size and muscle development to endocrine function. We know it is ultimately the genes of the child that normally determine its sex. But during development, when and how does the fetus differentiate into one sex or the other? How does the genotype of the child lead to the male or female development of the gonads?
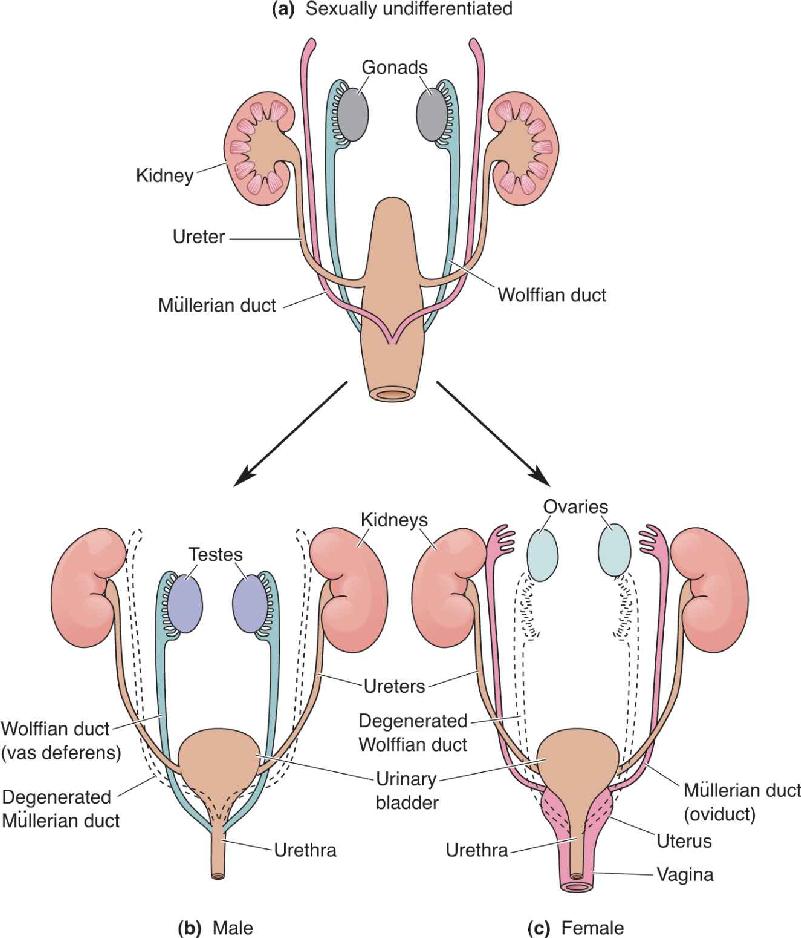
The answer involves the unique characteristics of the gonads during development. Unlike organs such as the lung and liver, the rudimentary cells that develop into the gonads are not committed to a single developmental pathway. During the first 6 weeks of pregnancy, the gonads are in an indifferent stage that can develop into either ovaries or testes. The uncommitted gonads possess two key structures, the Müllerian duct and the Wolffian duct (Figure 17.4). If the fetus has a Y chromosome with an SRY gene, testosterone is produced, and the Wolffian duct develops into the male internal reproductive system. At the same time, the Müllerian duct is prevented from developing by a hormone called Müllerian-inhibiting factor. Conversely, if there is no Y chromosome and no upsurge of testosterone, the Müllerian duct develops into the female internal reproductive system, and the Wolffian duct degenerates.

FIGURE 17.4 Development of the reproductive organs. (a) The undifferentiated urogenital system has both Müllerian and Wolffian ducts. (b) If an SRY gene is present, the Wolffian duct develops into the male reproductive organs. (c) If there is no SRY gene, the Müllerian duct develops into the female reproductive organs. (Source: Adapted from Gilbert, 1994, p. 759.) Description
The external genitals of both males and females develop from the same undifferentiated urogenital structures. This is why it is possible for a person to be born with genitals intermediate in form between those of typical males and females, a condition known as hermaphroditism.
Hormones are chemicals, released into the bloodstream, that regulate physiological processes. The endocrine glands we are primarily interested in are the ovaries and testes because they release sex hormones and the pituitary because it regulates this release. The sex hormones are crucial to the development and function of the reproductive system and sexual behavior. The sex hormones are steroids (mentioned in Chapter 15), and some of them are familiar, such as testosterone and estrogen. Steroids are molecules synthesized from cholesterol that have four carbon rings. Small alterations in the basic cholesterol structure have profound consequences for the effects of hormones. For example, testosterone is the most crucial hormone for male development, but it differs from the important female steroid estradiol in only a few places on the molecule.
Steroid sex hormones are often referred to as “male” or “female,” but men also have “female” hormones and women also have “male” hormones. The designation reflects the fact that men have higher concentrations of androgens, or male hormones, and women have more estrogens, or female hormones. For example, testosterone is an androgen and estradiol is an estrogen. In the series of chemical reactions that lead from cholesterol to sex hormones, one of the principal female hormones, estradiol, is actually synthesized from the male hormone testosterone (Figure 17.5). This reaction takes place with the aid of an enzyme called aromatase.

FIGURE 17.5 Cholesterol and the synthesis of the principal steroid sex hormones. Broken arrows indicate where one or more intermediate reactions occur. The enzyme aromatase converts testosterone into estradiol. Description
Steroids act differently from other hormones because of their structure. Some hormones are proteins and therefore cannot cross the lipid bilayer of a cell membrane. These hormones act at receptors with extracellular binding sites. In contrast, steroids are fatty and can easily pass through cell membranes and bind to receptors within the cytoplasm, giving them direct access to the nucleus and gene expression. Differences in the concentration of various receptors result in steroid effects localized to different areas of the brain (Figure 17.6).

FIGURE 17.6 The distribution of estradiol receptors in a sagittal section of the rat brain. High concentrations of these receptors are found in the pituitary and hypothalamus, including the preoptic area of the anterior hypothalamus. These brain areas are all involved in sexual and reproductive behaviors. Description
The testes are primarily responsible for the release of androgens, although small amounts are secreted in the adrenal glands and elsewhere. Testosterone is by far the most abundant androgen and is responsible for most masculinizing hormonal effects. Prenatally, elevated testosterone levels are essential for the development of the male reproductive system. Increases in testosterone much later, at puberty, regulate the development of secondary sex characteristics, ranging from increased muscular development and facial hair in human males to the mane of a lion. Oddly, for those with a genetic predisposition, testosterone also causes baldness in men. Female concentrations of testosterone are roughly 10% of those found in males. Male testosterone levels vary during the course of the day because of numerous factors, including stress, exertion, and aggression. It is not clear whether an increase in testosterone is a cause or an effect, but it is correlated with social challenges, anger, and conflict.
The principal female hormones are estradiol and progesterone, which are secreted by the ovaries. As already mentioned, estradiol is an estrogen; progesterone is a member of another class of female steroid hormones called progestins. Quite low during childhood, estrogen levels increase dramatically at puberty and control the maturation of the female reproductive system and the development of breasts. As in the male, blood concentrations of sex hormones are quite variable in the female. However, whereas in men fluctuations occur rapidly each day, in women, hormonal levels follow a regular cycle of approximately 28 days.
The Control of Sex Hormones by the Pituitary and Hypothalamus
The anterior pituitary gland secretes two hormones that are particularly important for normal sexual development and function in both women and men: luteinizing hormone (LH) and follicle-stimulating hormone (FSH). These hormones are also called gonadotropins. LH and FSH are secreted by specialized cells scattered throughout the anterior pituitary, comprising about 10% of the total cell population. Recall from Chapter 15 that the secretion of hormones from the anterior pituitary is under the control of hypophysiotropic hormones released by the hypothalamus. Gonadotropin-releasing hormone (GnRH) from the hypothalamus does what the name suggests, causing the release of LH and FSH from the pituitary. GnRH is also referred to as LHRH, for luteinizing hormone-releasing hormone because it causes a much greater increase in LH than FSH. Neuronal activity in the hypothalamus is influenced by numerous psychological and environmental factors that indirectly affect the secretion of gonadotropins from the anterior pituitary.
The chain of events from hypothalamic input to the secretion of gonadal hormones is illustrated in Figure 17.7. Neural input from the retina to the hypothalamus causes changes in the release of GnRH based on daily variations in light level. In some nonhuman species, strong seasonal variations in reproductive behavior and gonadotropin secretion occur. Light inhibits the production of the hormone melatonin in the pineal gland, increasing gonadotropin secretion because of the inhibitory effect of melatonin on gonadotropin release. By means of this circuit, reproductive activity can be influenced by the length of daylight during the course of the year, and offspring are born seasonally when they have the best chance of survival. In humans, there is also an inverse relationship between gonadotropin release and melatonin levels, but whether melatonin actually modulates reproductive behavior is not known.

FIGURE 17.7 Bidirectional interactions between the brain and the gonads. The hypothalamus is influenced by both psychological factors and sensory information, such as light hitting the retina. GnRH from the hypothalamus regulates gonadotropin (LH and FSH) release from the anterior pituitary. The testes secrete testosterone and the ovaries secrete estradiol, as directed by the gonadotropins. The sex hormones have diverse effects on the body and also send feedback to the pituitary and hypothalamus. Description
In males, LH stimulates the testes to produce testosterone. FSH is involved in the maturation of sperm cells within the testes. Sperm maturation also requires testosterone, meaning that both LH and FSH play key roles in male fertility. Because there is cortical input to the hypothalamus, it is possible for psychological factors to decrease male fertility by inhibiting gonadotropin secretion and sperm production.
In females, LH and FSH cause the secretion of estrogens from the ovaries. In the absence of gonadotropins, the ovaries are inactive, which is the situation throughout childhood. Cyclic variations in LH and FSH levels in adult females cause periodic changes in the ovaries, and the timing and duration of LH and FSH secretion determine the nature of the reproductive cycle, or menstrual cycle. In the follicular phase of the cycle, these hormones (particularly FSH) increase the growth of a small number of follicles, the cavities in the ovaries that enclose and maintain the ova (egg cells). In the luteal phase after egg expulsion, the small cells that surround the egg undergo chemical changes in a process called luteinization, which depends on LH release from the pituitary. The duration of the follicular and luteal phases of the reproductive cycle vary significantly for different mammals. The phases are roughly equal in length in the primate menstrual cycle.
In the estrous cycle of non-primate mammals, such as rats and mice, the luteal phase is much shorter. In other estrous animals, such as dogs, cats, and farm animals, the phases are more nearly equal in duration. Many estrous animals have only one cycle per year, usually in the spring. Presumably, this timing is for the production of offspring when the weather and food supply are optimal. At the other extreme are animals such as rats, which are said to be polyestrous because they have short periods of estrus, or “heat,” throughout the year.
Sexual behavior is a vast, complex, and provocative topic, ranging from the mechanical and biological facts of copulation to the myriad cultural practices of human societies. Here, we will touch on only certain aspects of this subject. We begin with the autonomic and spinal neurons that control the genitals, then discuss various strategies of mating, and conclude with some research about brain mechanisms that are important for monogamy and parenting.
Despite the obvious structural differences between female and male reproductive organs, their neural regulation (to the extent it is understood) is surprisingly similar. Sexual arousal of adult men and women can result from erotic psychological and sensory stimuli (including visual, olfactory, and somatosensory), as well as from tactile stimulation of the external sex organs. A full sexual response cycle consists of arousal followed by plateau, orgasm, and resolution phases. Although the duration of each phase can vary widely, the physiological changes associated with each are relatively consistent. Neural control of the sexual response comes in part from the cerebral cortex—where erotic thoughts occur—but the spinal cord coordinates this brain activity with sensory information from the genitals and generates the critical outputs that mediate the sexual responses of the genital structures.
The major external and internal sex organs are shown in Figure 17.8. Research on the physiology of the human sexual response has tended to focus unduly on men, but we will try to summarize some of what is known about both sexes. Sexual arousal causes certain parts of the external genitals of both women and men to become engorged with blood, and thus to swell. In women, these structures include the labia and the clitoris; in men, it is primarily the penis. The external genitals are densely innervated by mechanoreceptors, particularly within the clitoris and the glans of the penis. Stimulation of these sensory endings can, by itself, be enough to cause engorgement and erection. The best evidence that engorgement can be generated by a simple spinal reflex is that most men who have suffered a complete transection of the spinal cord at the thoracic or lumbar level can nevertheless experience an erection when their penis is mechanically stimulated. The mechanosensory pathways from the genitals are components of the somatosensory system (see Chapter 12), and their anatomy follows the usual pattern: Axons from mechanoreceptors in the penis and clitoris collect in the dorsal roots of the sacral spinal cord. They then send branches into the dorsal horns of the cord, and into the dorsal columns, through which they project toward the brain.

FIGURE 17.8 The neural control of human sex organs. Sensory information from the sex organs follows the dorsal column–medial lemniscal pathway to the brain. Description
Engorgement and erection are controlled primarily by axons of the parasympathetic division of the ANS (see Figure 15.9). Within the sacral spinal cord, the parasympathetic neurons can be excited by either mechanosensory activity from the genitals (which can directly trigger reflexive erection) or by axons descending from the brain (which account for responses mediated by more cerebral stimuli) (see Figure 17.8). Engorgement of the clitoris and penis depends on dramatic changes in blood flow. Parasympathetic nerve endings are thought to release a potent combination of acetylcholine, vasoactive intestinal polypeptide (VIP), and nitric oxide (NO) directly into the erectile tissues. These neurotransmitters cause the relaxation of smooth muscle cells in the arteries and the spongy substance of the clitoris and penis. The usually flaccid arteries then become filled with blood, thereby distending the organs. (Sildenafil, better known by its trade name Viagra, is a treatment for erectile dysfunction that works by enhancing the effects of NO.) As the penis becomes longer and thicker, the spongy internal tissues swell against two thick, elastic outer coverings of connective tissue that give the erect penis its stiffness. In order to keep the organs sliding easily during copulation throughout the plateau phase, parasympathetic activity also stimulates the secretion of lubricating fluids from the woman’s vaginal wall and from the man’s bulbourethral gland.
Completing the sexual response cycle requires activity from the sympathetic division of the ANS. As sensory axons, particularly from the penis or clitoris, become highly active, they, together with activity descending from the brain, excite sympathetic neurons in the thoracic and lumbar segments of the spinal cord (see Figure 17.8). In men, the sympathetic efferent axons then trigger the process of emission: Muscular contractions move sperm from storage sites near the testes through two tubes called the vas deferens, combine the sperm with fluids produced by various glands, and propel the resulting mixture (called semen) into the urethra. During ejaculation, a series of coordinated muscular contractions expel the semen from the urethra, usually accompanied by the intense sensations of orgasm. In women, stimulation adequate to trigger orgasm probably also activates the sympathetic system. Sympathetic outflow causes the outer vaginal wall to thicken and, during orgasm itself, triggers a series of strong muscular contractions.
Studies of the neural basis of orgasm are challenging and relatively new. One can only imagine the “technical” challenges that would be encountered fitting two people inside a magnetic resonance imaging (MRI) machine, but a more scientific problem is the investigation of feelings themselves (more on this in Chapters 18 and 21). For example, research has shown that the feelings of orgasm are accompanied by neural activity in widespread cortical and subcortical structures, but we don’t know which areas are actually responsible for the feelings and, more generally, it is a complete mystery how patterns of neural activity evoke feelings—why is one activity pattern pleasurable and another painful? Studies in people who experience epileptic seizures give us clues to brain areas particularly relevant for orgasm. In rare cases, the aura that precedes seizures may be sexually arousing, and the loci of such seizures are most commonly in the temporal lobe. In the surgical treatment of epilepsy, electrical stimulation of the medial temporal lobe or the basal forebrain has been reported to cause sexual arousal in some patients. Electrical stimulation of the medial temporal lobe has also been found to produce feelings of orgasm in at least a few patients. Studies in additional patients and in non-epileptic brains are needed to confirm the association between orgasm and temporal lobe activation.
Following an orgasm, some time must pass before another orgasm can be triggered in men. The orgasmic experience of women tends to be considerably more variable in frequency and intensity. The resolution phase, which ends the sexual response cycle, includes a draining of blood from the external genitals through veins, and a loss of erection and other signs and sensations of sexual excitement.
Mammals practice a dazzling range of mating behaviors. Each one is a strategy that ultimately meets the single evolutionary objective: to maximize the survival of offspring and parental genes. Species variations in preferred mating systems seem to depend on the investment that males and females make in raising their offspring, although there are exceptions. Very common among mammals is polygyny (from the Greek for “many women”), in which the male mates with many females but the female mates with only one male for one or multiple mating seasons. Polygynous mating (practiced by giraffes, orangutans, and most other mammals) usually has a “one-night stand” quality to it, and the male never looks back to check on the outcome of his many liaisons or the mate. Sometimes, polygyny takes the form of a harem; one male forms a lasting and exclusive association with a group of females, as practiced by gorillas, elephant seals, and a very small number of traditional human cultures.
Polyandry (“many men”), in which one female mates with many males but the males mate with only that female, is rare among mammals and vertebrates in general. One exception is the phalarope, a shorebird that breeds in the cold tundra. Some species practice simultaneous polyandry in which the female bird mates with and lays eggs in the nests of multiple males who raise the young within her territory. Other species practice sequential polyandry in which the female has nothing to do with a mate or offspring once the eggs are laid. Some marmosets and tamarins also appear to be polyandrous. Although historical examples of human polyandry have had a wide geographic distribution, these cases are rare and polyandry appears in only a tiny fraction of societies today. Polygyny and polyandry are both examples of polygamy—having more than one mate.
In monogamy (“one spouse”), a male and a female form a tightly bound relationship that includes exclusive (or nearly exclusive) mating with each other. Only about 3% of mammalian species are monogamous, although monogamy is practiced by roughly 12% of primate species (and 90% of bird species). The exclusive relationship may last a lifetime or until a new partner is chosen (serial monogamy).
Nearly every type of mating strategy occurs among humans in various cultures and eras. On balance, humans have a strong tendency toward (at least temporary) monogamy, although some cultures condone polygyny. Interestingly, even where polygyny is socially acceptable, most marriages are monogamous. Polyandry for reproductive purposes is rare, and most cultures have penalized women accused of it. Although there has been much speculation about evolutionary explanations for human mating patterns, determining the precise influences of genetics and culture on mating behaviors is virtually impossible.
Regardless of an animal’s choice of reproductive strategy—staying faithful to a mate and devoted to one’s children, or wandering promiscuously and abandoning offspring—complex social behaviors are involved. It would be remarkable if the tendency to be monogamous or polygamous were controlled by a few simple brain chemicals. Yet recent work on mouselike rodents called voles suggests that certain well-known pituitary hormones do precisely that (at least in voles).
Voles are a wonderful natural experimental model because closely related species of voles have very different reproductive behaviors. The prairie vole (Microtus ochrogaster) lives in the American grasslands, practicing solid “family values” (Figure 17.9). It is highly social and as reliably monogamous as any mammal known. After an intense period of initial matings, the male and female form a tight lifelong pair-bond and live together in one nest. The male will fiercely defend his mate, and both parents cooperate in the long-term care of their young. In contrast, the montane vole (Microtus montanus) is asocial and promiscuous. Each one lives in an isolated nest, the males take no part in parenting, and the females care for their offspring only briefly before letting them fend for themselves in the world.
FIGURE 17.9 Studying reproductive behavior. The prairie vole is a valuable experimental model, representing monogamy and the care of offspring by both parents. (Source: Copyright 2005, Wendy Shattil/Bob Rozinski.)
Vole pair-bonding has been studied in the laboratory by testing the preference of an animal for spending time with a partner or an unfamiliar animal (Figure 17.10). After mating, a female prairie vole spends more time with its partner than by itself or with a stranger. Female montane voles, on the other hand, spend most of their time in a neutral area alone rather than with their recent mating partner or a stranger.

FIGURE 17.10 Pair-bonding in prairie and montane voles. (a) To experimentally measure partner preference, the vole is placed in a neutral chamber and allowed to choose whether to stay alone or visit neighboring chambers, where a partner or a stranger is held. (b) After mating, montane voles spend most of their time alone and away from their partner (blue), whereas prairie voles choose to spend most of their time with the partner (purple). (Source: Adapted from Insel and Young, 2001.) Description
Because these two vole species are physically and genetically quite similar, relatively few biological factors might account for their different reproductive behaviors. Thomas Insel and his coworkers at Emory University and the National Institute of Mental Health have investigated the subtle brain differences that appear to account for the very different mating strategies in the two species of voles (Box 17.1). Based on clues from previous studies of maternal and territorial behavior, research has focused on the roles of oxytocin and vasopressin in voles. Recall that these peptide hormones are synthesized in the hypothalamus and can be released into the bloodstream by neurosecretory terminals in the posterior pituitary gland (see Figure 15.4). Circulating vasopressin (also known as ADH, antidiuretic hormone) helps regulate water and salt levels in the body, mainly by affecting the kidneys; oxytocin stimulates smooth muscle, causing uterine contractions during childbirth and milk letdown during lactation. However, oxytocin and vasopressin are also released onto CNS neurons, and, like most signaling molecules, they bind to specific receptors scattered about the brain. Because oxytocin and vasopressin are protein hormones, they bind with extracellular receptors.
I had never heard of a vole, let alone met one in vivo. I trained as a physician and then a psychiatrist. After clinical training, I really had no idea about science. Almost completely by serendipity, I got a job at the National Institutes of Health (NIH) in Bethesda, Maryland. In the early 1980s, NIH seemed to have a Nobel laureate on almost every floor, and the intellectual environment, especially for the fast-emerging field of neuroscience, was infectious. Neuropeptides were the rage, with a new neuropeptide or neuropeptide receptor being discovered nearly every month. And tools for studying slow and fast neurotransmitters were evolving rapidly, so that any young investigator with a new technique could quickly begin to run experiments.
But neuroscience at the NIH in the 1980s also felt a bit crowded. There were lots of talented and technique-savvy scientists working on the neural basis of stress, sadness, and pain. My instinct had always been to go to less crowded places where I could focus on problems without rushing to keep up or ahead of others. And since I had no formal scientific training, I needed time to learn the hard lessons of science. I moved to the Laboratory of Brain Evolution and Behavior in the NIMH, founded by Paul MacLean on a farm in Poolesville, Maryland.
I chose to focus on stress research but decided to study development, focusing on the recently discovered ultrasonic calls rat pups emit when they are separated from their mothers. My behavioral neuroscience career seemed to be progressing until my first post-doctoral fellow arrived, returning with some ambivalence from maternity leave. The idea of listening to rat pups cry after being separated from their mothers was not an ideal project for her. To her great credit and my lasting gratitude, Marianne Wamboldt pointed out that we might study the experience of the mothers and not just the separation distress of the pups.
At that time, very few people were interested in the neurobiology of positive behaviors like parental care, affiliation, or attachment. A robust community of scientists was studying reproductive behavior in rodents and working out the role of gonadal steroids and neuropeptides, but most of this research focused on the motor or sensory aspects of reproduction, not on the emotional or affective experience. With the discovery that neuropeptides, like oxytocin, could modify parental behavior, and with a new post-doc who cared a lot about maternal affect, we were off into a new frontier. Using tools for mapping oxytocin receptors in the brain, we were able to demonstrate the pathways critical for rats becoming maternal, a profound behavioral transition that takes place right at the time of parturition.
These studies helped us to understand the neural mechanisms of maternal care, but what about attachment between adults? Laboratory rats and mice are not ideal for studying attachment. They are highly social but not selective. We needed a species that was monogamous, forming selective and enduring pair bonds. Again, through serendipity, I met a brilliant behavioral endocrinologist, Sue Carter, who was then at the University of Maryland. Sue schooled me in behavioral biology and introduced me to her favorite animal, the prairie vole.
If nature had set out to evolve a species perfect for social neuroscience, prairie voles could well be the result: They are highly affiliative, easy to breed in the lab, and profoundly monogamous. Sue Carter had studied these critters in both the lab and the field, working out simple but rigorous behavioral measures of partner preference and attachment. Bringing the neuroscience expertise from our lab in Poolesville together with the behavioral expertise in Sue’s lab, we were able to show the profound effects of oxytocin and vasopressin on affiliative behaviors and attachment.
The story became even more interesting after the lab moved to Emory University in 1994. Joined by Larry Young and Zuoxin Wang, we were able to bring transgenic and viral vector tools to answer questions about the mechanisms by which oxytocin and vasopressin influence social cognition and social behavior. Two insights emerged. First, altering the regional expression of receptors in brain could change social organization, inducing or preventing mating-induced attachment. This was quite a shock because it meant that release of the same peptide had completely different effects in different species. And as we compared monogamous and nonmonogamous species, we noticed a surprising pattern. In monogamous rodents and primates, oxytocin receptors were found in brain areas associated with reward, as if this single receptor linked the social world to the circuitry for motivation. Today, oxytocin is also being studied in autism and schizophrenia.
Of course, the vole work has raised questions about monogamy in humans. I have always been reluctant to extrapolate from voles to mice, so extrapolating from voles to humans seems a fool’s errand. But that does not mean the prairie vole is irrelevant. “Nature’s gift to social neuroscience” reminds us that neuroanatomy, especially the distribution of receptors, is important for understanding function. Thanks to prairie voles, the neural basis of attachment is now an exciting area of neuroscience. And, whatever the role of oxytocin and vasopressin in human social behavior, we have gained some basic principles for understanding the relationship of form and function in the brain.
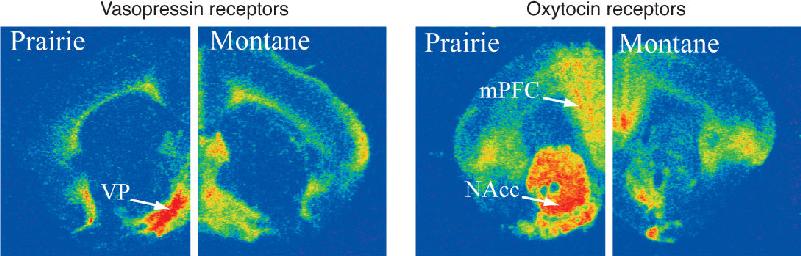
As shown in Figure 17.11, maps of these receptors are strikingly different in the brains of prairie voles and montane voles, whereas maps of other types of neurotransmitter and hormone receptors are very similar in the two species. The receptor differences correlate well with reproductive behavior even in other species of voles. Furthermore, the maps are plastic: When the female montane vole gives birth and assumes a maternal role (however briefly), her receptor maps temporarily change to resemble those of the prairie vole.

FIGURE 17.11 The role of oxytocin and vasopressin receptors in reproductive behavior. These coronal brain sections show the distribution of oxytocin and vasopressin in the brains of montane voles and prairie voles. The red areas have the highest receptor densities. Compared to montane voles, prairie voles have high vasopressin receptor density in the ventral pallidum (VP) and high oxytocin receptor density in the medial prefrontal cortex (mPFC) and the nucleus accumbens (NAcc). (Source: Young et al., 2011.) Description
The distinctive maps of oxytocin and vasopressin receptors tell us that each hormone activates a different network of neurons in the polygynous and monogamous vole brains. This alone does not prove that the hormones have anything to do with sex-related behaviors. But together with the effects of the hormones and drugs that antagonize them, this evidence makes a strong case for cause and effect. When a pair of prairie voles copulates, levels of vasopressin (in males) and oxytocin (in females) rise sharply. Vasopressin antagonists given to a male prairie vole before mating prevent him from forming a pair-bond relationship. This disruption of pair-bonding can be produced with the antagonist selectively infused into the ventral pallidum (the anterior portion of the globus pallidus). Oxytocin antagonists have no such effect. When a male is given vasopressin while he is exposed to a new female, he quickly forms a strong preference for her even without the intense mating that usually precedes pair-bonding. In females, oxytocin appears to be necessary to establish a preference for her mate, while vasopressin has little effect.
A study by Lim et al. provides more direct evidence that vole pair-bonding can be significantly altered by a small change in vasopressin receptors. A virus was used to deliver genes to the ventral pallidum of male montane voles, causing an overexpression of vasopressin receptors. Consequently, the male montane voles had numbers of vasopressin receptors in the ventral pallidum comparable to prairie voles. The manipulated montane voles also pair-bonded like prairie voles. If this cause-and-effect link is supported by further studies, it will dramatically show that a complex social behavior can be altered by the overexpression of a single protein at one location in the brain.
Oxytocin and vasopressin are also involved in vole parenting habits. Vasopressin increases the male prairie vole’s paternal proclivities, causing him to spend more time with his pups, and oxytocin similarly stimulates maternal behaviors in the female. The research on voles suggests a very interesting hypothesis about the evolution of complex social behaviors. If genetic mutations change the anatomical distribution of a particular hormone’s receptors, then that hormone may evoke an entirely new repertoire of behaviors. Consistent with this idea, administering vasopressin or oxytocin to the naturally promiscuous montane voles does not evoke the effects on pair-bonding and parenting seen in prairie voles, perhaps because they don’t have receptors in the necessary places.
The vole story is a fascinating example of how brain chemicals can regulate critical behaviors. But what does all this have to do with human relationships, faithfulness, and love? It is too early to be sure, but intriguing pieces of evidence suggest that voles may teach us something about the human brain and behavior. For example, there is evidence that human plasma oxytocin levels increase during breastfeeding in mothers and during sexual intercourse in men and women.
In a series of experiments, Andreas Bartels and Semir Zeki at University College London have used functional magnetic resonance imaging (fMRI) to explore human brain activity associated with maternal and romantic love and bonding. In an experiment investigating maternal love, while brain scans were being taken, mothers saw pictures of their child intermixed with pictures of other familiar children. In a second experiment studying romantic love, brain activity was compared when men and women viewed pictures of partners and pictures of friends. The differences in brain activity for one’s child versus other children and one’s partner versus friends are shown in Figure 17.12. Several brain areas, including the anterior cingulate cortex, the caudate nucleus, and the striatum, are more activated by one’s child and partner than by pictures of unrelated people. The heightened responses to child and partner overlap significantly; other brain areas respond differently with the two types of relationship. Many of the areas that are active with both maternal and romantic attachment are part of the brain’s reward circuitry (see Chapter 16). We can speculate that the brain activation demonstrates the strong reinforcing nature of partner and parental relationships. Also interesting, and relevant to the vole story, is the finding that many of the brain areas activated by pictures of people’s partners and children are rich in oxytocin and vasopressin receptors.

FIGURE 17.12 Imaging maternal and romantic love in the human brain. Brain activation is shown in (a) sagittal, (b) horizontal, and (c,d) two different coronal planes. Yellow areas were more active when mothers saw pictures of their own child than with pictures of other familiar children. Red areas were more activated by pictures of romantic partners than to pictures of friends. Some of the highlighted areas are labeled: PAG, periaqueductal gray; aC, anterior cingulate cortex; hi, hippocampus; I, insula; C, caudate nucleus; S, striatum. (Source: Bartels and Zeki, 2004.) Description
These fMRI studies suggest that oxytocin and vasopressin play roles in human bonding, perhaps similarly to what we see in voles. But surely, the tendency of humans to be monogamous or not isn’t as simple as in rodents. Although human behavior undoubtedly involves more complex factors than vole behavior, surprising evidence suggests that vasopressin influences human bonding as well. Hasse Walum and a team of Swedish and American scientists studied 552 pairs of same-sex Swedish twins who were married or had long-term partners. Of particular interest was the gene sequence that codes for vasopressin receptors and the tendency toward monogamy. The DNA sequences that code vasopressin receptors in montane and prairie voles are nearly identical, but the monogamous prairie voles have a DNA sequence adjacent to the gene that encodes the V1aR vasopressin subtype (called a gene variant). When this gene variant is transgenically introduced into nonmonogamous mice, their social behavior becomes more like the prairie voles. In the human twin study, the scientists investigated whether vasopressin gene variants might influence pair-bonding in humans too. In women, there was no connection between the vasopressin gene variants and the quality of their marriage as assessed by a variety of questionnaires. In the men, however, an intriguing correlation was found: Men with a particular gene variant scored significantly lower on measures of the quality of their marriage and were twice as likely to report that a marital crisis had occurred in the year before the survey. The wives of the men with the gene variant also reported lower marital quality than the wives of the men without the variant. The function of this gene variant is not known, but these results suggest that, even in humans, vasopressin receptors might play a role in pair-bonding.
Sexual reproduction depends on a variety of individual and social behaviors—finding, attracting, and keeping a mate; copulating; giving birth; and nursing and nurturing the offspring—and in each case, the behavior of males and females is usually quite different. Since all behaviors ultimately depend on the structure and function of the nervous system, we can make the strong prediction that male and female brains are also somehow different; that is, they have sexual dimorphisms (from the Greek dimorphos, “having two forms”). Another good reason to expect that male and female brains differ is simply that male and female bodies differ. The body parts that are unique to each sex require neural systems that have evolved specifically to control them. For example, male rats have a particular muscle at the base of the penis, and their spinal cord has a small cluster of motor neurons that control that muscle; females lack both the muscle and the related motor neurons. Body size and general shape also vary with gender, and thus somatosensory and motor maps must adjust to fit them.
Sexual dimorphisms vary widely across species. In the brain, dimorphisms are sometimes found, but they are significant in some species and nonexistent in others. An example of an animal with a large dimorphism is the Icelandic stickleback fish, in which the male brain is much larger than the female brain, perhaps because of the cognitive demands of nest construction, courtship, and childcare (Figure 17.13), which are carried out only by the male. In rodents, the trained eye can tell male from female brains with no ambiguity because of differences in the hypothalamus. The diversity of brain dimorphisms across species is sometimes associated with remarkable variations in sexual behaviors. For example, in some songbird species, only males sing, and, not surprisingly, only males have large singing-related brain nuclei. In human brains, dimorphisms have so far proven to be subtle, few, and of unknown function. Differences between human male and female brains tend to vary along a continuum, with lots of overlap. A particular hypothalamic nucleus might be larger in women than men on average, for example, but size variations of the nucleus may be so great that many men have a larger nucleus than many women.

FIGURE 17.13 Dimorphism in brain size. These brains are from adult female (left) and male (right) three-spined stickleback fish that were the same length and weight. The male brain is larger and 23% heavier than the female brain. The scale bar indicates 1 mm. (Source: Kotrschal et al., 2012.)
In the rest of this section, we will describe sexual dimorphisms in the nervous systems of humans and other species, focusing on examples that illuminate the relationship between the brain and behavior. We will also discuss some of the neurobiological mechanisms that generate these dimorphisms.
Few dimorphic neural structures are related to their sexual functions in an obvious way. One structure that is related is the collection of spinal motor neurons that innervates the bulbocavernosus (BC) muscles surrounding the base of the penis. These muscles have a role in penile erection and help to eject urine. Both women and men have a BC muscle. In women, it surrounds the opening of the vagina and serves to constrict it slightly. The motor neuron pool controlling the BC muscles in humans, called Onuf’s nucleus, is located in the sacral spinal cord. Onuf’s nucleus is moderately dimorphic (there are more motor neurons in men than women) because the male BC muscles are larger than those of females.
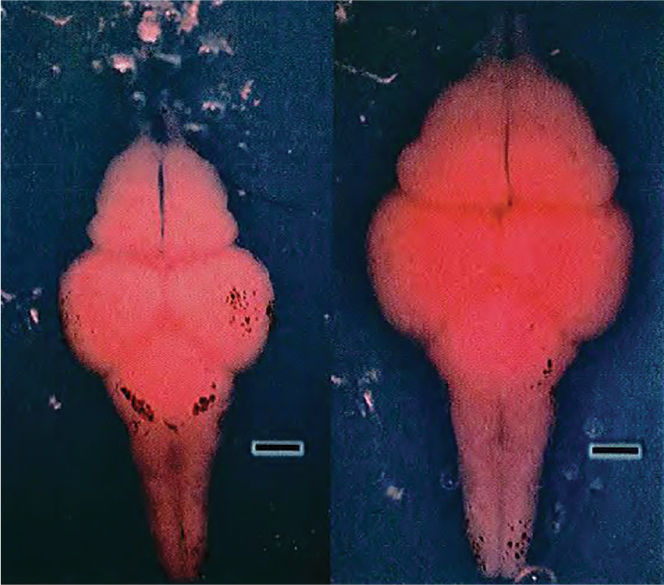
The most distinct sexual dimorphisms in the mammalian brain are clustered around the third ventricle, within the preoptic area of the anterior hypothalamus. This region seems to have a role in reproductive behaviors. In rats, lesions of the preoptic area disrupt the estrous cycle in females and reduce the frequency of copulation in males. Histological sections of male and female preoptic areas from rats show an obvious difference: The aptly named sexually dimorphic nucleus (SDN) is five to eight times larger in males than in females (Figure 17.14).

FIGURE 17.14 Sexual dimorphism in rats. The sexually dimorphic nucleus (SDN) in the hypothalamus of male rats (left) is much larger than the SDN in female rats (right). (Source: Adapted from Rosenzweig et al., 2005, Fig 12.21. Photos courtesy of Roger Gorski.) Description
The preoptic area of humans may also have dimorphisms, but the differences are small and controversial. There are four clusters of neurons called the interstitial nuclei of the anterior hypothalamus (INAH). In different studies INAH-1, INAH-2, and INAH-3 have all been reported to be larger in men than women. INAH-1 may be the human analogue of the rat SDN, but researchers disagree about whether INAH-1 is dimorphic. The clearest dimorphism appears to be in INAH-3, which was first reported to be twice as large in men as women by Laura Allen, Roger Gorski, and their colleagues at UCLA. Evidence of the involvement of the INAH in sexual behavior is thus far inconclusive. Various neurons of the medial preoptic area in male rhesus monkeys fire vigorously during specific phases of sexual behavior, including arousal and copulation. In addition, there may be subtle differences in the size of certain hypothalamic nuclei that correlate with sexual orientation in people.
Human brain dimorphisms outside the hypothalamus have been difficult to prove conclusively, although many have been reported. For example, some studies have found that the corpus callosum is larger in men, but this may be a consequence of men having slightly larger brains (and bodies). In other reports, the posterior end of the corpus callosum, called the splenium, is larger in women than men. But even if there is a dimorphism in the size or shape of the corpus callosum, what could it mean? We can only guess. The callosum has no obvious role in mediating specifically sex-related behaviors, but it is important for a variety of cognitive functions that involve coordinated activity between hemispheres. Observations of stroke patients in whom only one hemisphere has been damaged suggest that the functions of female brains may be less lateralized, that is, less dependent on one cerebral hemisphere more than the other. But this conclusion, too, has been challenged. As a rule, sexual dimorphisms of the brain are difficult to prove because male and female brains are very similar, and because within populations of male and female brains, there is considerable individual variation.
Perhaps, the most reliable conclusion we can draw about sexual dimorphisms in human brain structure is that there are so few of them. This probably should not come as a surprise, since the vast majority of women’s and men’s behaviors are very similar, if not indistinguishable. The gross anatomy of the brain provides only a crude view of the organization of the nervous system. To determine the reasons for sexually dimorphic behavior, we need to look deeper into the patterns of neural connections, the neurochemistry of the brain, and the influence of sex-related hormones on neural development and function.
Even if there are no major differences in the brain structures of men and women, there may still be differences in cognitive abilities. Reports of cognitive dimorphisms are sometimes accompanied by an evolutionary explanation: Men evolved as hunters and relied more on their abilities to navigate their environment. Women evolved the behavior of staying closer to home to care for children, so they were more social and verbal.
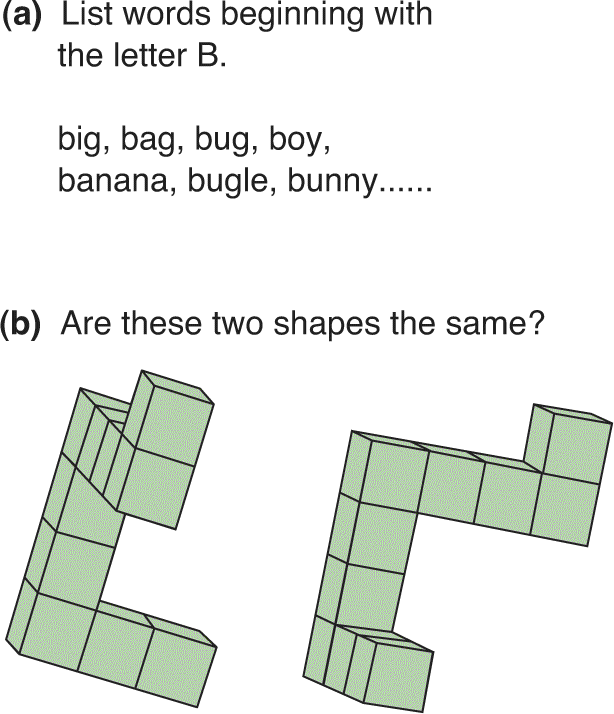
Numerous studies have reported that women are better at verbal tasks than men. Starting at around age 11 years, girls perform slightly better on tests of comprehension and writing, and this effect is sometimes said to extend through high school and beyond. Perhaps, it reflects a difference in the rates of brain development in the two sexes. Specific tasks at which women excel include naming objects of the same color, listing words beginning with the same letter, and verbal memory (Figure 17.15a).

FIGURE 17.15 Cognitive tasks that may favor women or men. (a) Women may outperform men in listing words beginning with the same letter. (b) Men appear to be somewhat better at spatial rotation tasks, such as deciding whether two three-dimensional objects are the same. (Source: Adapted from Kimura, 1992, p. 120.) Description
In other sorts of tasks, men appear to outperform women. Tasks that reportedly favor men include map reading, maze learning, and mathematical reasoning. Researchers speculate that these male advantages evolved from the days when men roamed large areas to hunt wild animals. One of the largest reported differences between the sexes is mental rotation of objects, a task that appears to favor men (Figure 17.15b).
While thinking about dimorphisms of cognition, we need to consider a few things. First, not all studies yield the same results. In some cases, one sex performs better, and in others, there is no difference. Second, across large groups of people of both sexes, there are huge differences in performance. But most of the variation is the result of differences among individuals, rather than being sex-specific. Third, it is not clear whether performance differences (or brain dimorphisms) are innate or the result of differences in experience. Typical males and females experience different things and they may, on average, develop slightly different skills. This may in turn influence neural circuitry.
A common interpretation of sex-based differences in performance is that the distinctive hormonal environments of male and female brains make them work somewhat differently. Perhaps, there is a benefit or penalty associated with estrogens or androgens for each task. Consistent with this conjecture are reports that spatial reasoning in women correlates with the menstrual cycle, better performance being observed when estrogen levels are lowest. Evidence also indicates that administering testosterone enhances spatial performance in older men with low testosterone levels. However, cognition cannot be so simply related to hormones, as there is no reliable correlation between performance on verbal or spatial tasks and hormone levels. This doesn’t mean that hormones do not affect cognitive function, but we must be cautious about overgeneralizing.
A variety of factors ranging from genetics to culture and life experiences may make a behavior more common in one sex than the other, but ultimately, all behaviors are controlled by the brain. Even if there are no gross anatomical dimorphisms, male and female brain circuitry must be somewhat different to account for sex-specific behaviors, whether singing by male birds or human sexual behavior. Recall that the kinds of sex hormones circulating in the blood are determined by the gonads, and the dimorphism of the gonads is ordinarily specified by our genes. As described earlier, people with a Y chromosome express a factor (testis-determining factor) that causes the undifferentiated gonads to become testes; people lacking a Y chromosome do not produce TDF, and their gonads differentiate into ovaries. The differentiation of testes or ovaries sets off a cascade of developmental events in the body. Most importantly for the sexual differentiation of the brain, the testes produce androgens, which trigger the masculinization of the nervous system by regulating the expression of a variety of sex-related genes. In the absence of androgens, there is a feminization of the brain through a different pattern of gene expression.
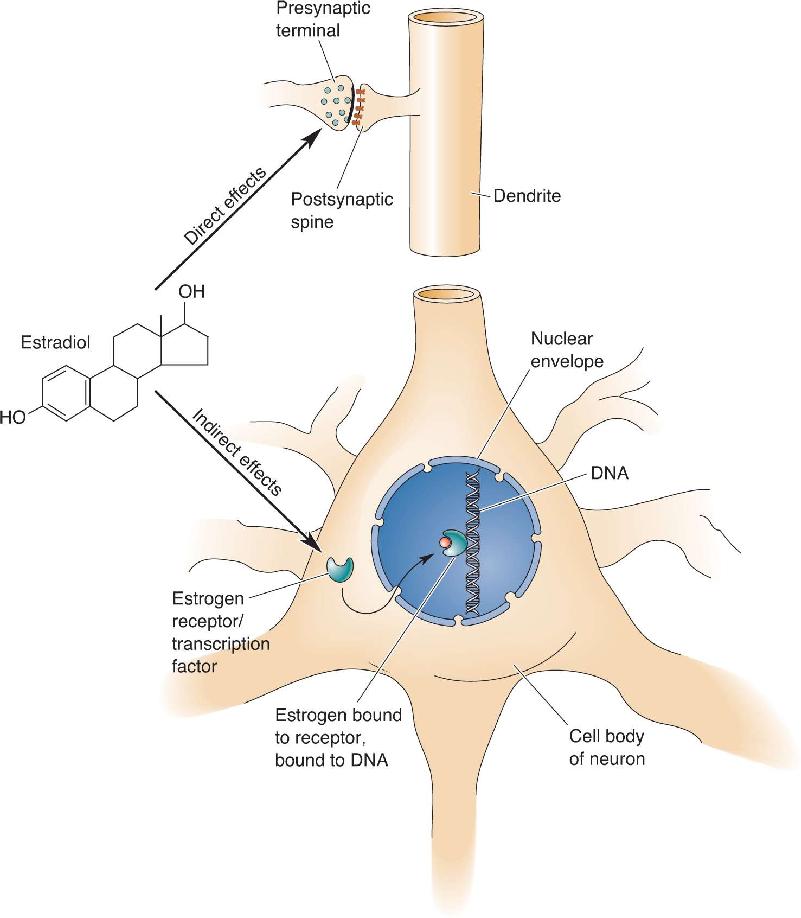
There is nothing fundamentally unique about the brain’s sensitivity to hormones. It is just one more body tissue waiting for a hormonal signal to decide its specific pattern of growth and development. Androgens provide a unitary signal for masculinization in the brain, just as in the various other tissues of the body that are sexually dimorphic. Steroids can influence neurons in two general ways (Figure 17.16). First, they can act quickly (within seconds or less) to alter membrane excitability, sensitivity to neurotransmitters, or neurotransmitter release. Steroids do this, in general, by directly binding to, and modulating the functions of, various enzymes, channels, and transmitter receptors. For example, certain metabolites (breakdown products) of progesterone bind to the inhibitory GABAA receptor and potentiate the amount of chloride current activated by GABA. The effects of these progesterone metabolites are quite similar to the sedative and anticonvulsant effects of the benzodiazepine class of drugs (see Figure 6.22). Second, steroids can diffuse across the outer membrane and bind to specific types of steroid receptors in the cytoplasm and nucleus. Receptors with bound steroid can either promote or inhibit the transcription of specific genes in the nucleus, a process that can take minutes to hours. Specific receptors exist for each type of sex hormone, and the distributions of each receptor type vary widely throughout the brain (see Figure 17.6).

FIGURE 17.16 The direct and indirect effects of steroids on neurons. Steroids can directly affect transmitter synthesis, transmitter release, or postsynaptic transmitter receptors. They can indirectly influence gene transcription. Description
Steroid hormones can exert effects on the brain and body throughout life, but their influence early in development is often fundamentally different from their effects after the animal is mature. For example, the ability of testosterone to alter very young genitals and brain circuitry, leading to distinctly male genitals and masculine behaviors later in life, can be thought of as the organizational effects of the hormone. The hormone organizes the perinatal tissues in irreversible ways that allow it to generate male functions after sexual maturity has been reached. For a mature animal to express sexual behaviors fully, however, it is often necessary for steroid hormones to circulate again during periods of sexual activity, providing activational effects on the nervous system. Thus, for example, testosterone levels might surge in the body of a male songbird in the spring, activating changes in certain parts of his brain that are essential for normal reproductive behavior (Box 17.2). Activational effects are usually temporary.
To our ears, the singing of birds may be simply a pleasant harbinger of spring, but for birds, it is part of the serious business of sex and reproduction. Singing is strictly a male function in many species, performed for the purpose of attracting and keeping a mate and for warning off potential rivals. Studies of two bird species with different habits of reproduction and singing have revealed some fascinating clues about the control and diversity of sexual dimorphisms in the brain.
Zebra finches, which are popular pets, live in the wild in the harsh Australian desert. To breed successfully, birds require dependable sources of food, but in the desert, food comes only with sporadic and unpredictable rains. Zebra finches must therefore be ready and willing to breed whenever food and a mate are available, in any season. Wild canaries, on the other hand, live in the more predictable environment of the Azores and (where else?) the Canary Islands. They breed seasonally during spring and summer and do not reproduce during fall and winter. The males of both species are passionate singers, but they differ greatly in the size of their repertoires. Zebra finches belt out one simple ditty all their lives and cannot learn new ones. Canaries learn many elaborate songs, and they add new ones each spring. The different behaviors of zebra finches and canaries require different mechanisms of neural control.
The birds’ sexually dimorphic behavior—singing—is generated by dramatically dimorphic neural structures. Birds sing by forcing air past a special muscularized organ called the syrinx, which encircles the air passage. The muscles of the syrinx are activated by motor neurons of the nucleus of cranial nerve XII, which are in turn controlled by a set of higher nuclei collectively called the vocal control regions, or VCRs (Figure A). In zebra finches and canaries, VCR size is five or more times larger in males than in females.

Figure A Blue circles represent the vocal control regions in male and female zebra finches. Description
The development of VCRs and singing behavior is under the control of steroid hormones. However, the very different seasonal requirements of zebra finches and canaries are paralleled by distinctly different modes of steroidal control. Zebra finches apparently require early doses of steroids to organize their VCRs, and later androgens to activate them. If a hatchling female zebra finch is exposed to testosterone or estradiol, her VCRs will be larger than those of normal females when she reaches adulthood. If the masculinized female is given more testosterone as an adult, her VCRs will grow larger still, and she will then sing like a male. Females that are not exposed to steroids when young are unresponsive to testosterone as adults.
In contrast, the song system in canaries seems to be independent of early steroid exposure, yet it bursts into full service each spring. If female canaries are given androgens for the first time as adults, they will begin singing within a few weeks. The androgens of males surge naturally each spring; their VCRs double in size as neurons grow larger dendrites and more synapses, and singing commences. Remarkably, neurogenesis, the birth of neurons, continues throughout adulthood in songbird brains, further contributing to the VCR circuitry during the mating season. By fall, male androgen levels drop, and the canary song system shrinks in size as his singing abates. In a sense, the male canary rebuilds much of his song control system anew each year as courtship begins. This may enable him to learn new songs more easily and, with his enlarged repertoire, gain some advantage in attracting a mate.
Masculinization of the Fetal Brain. Prenatally, elevated testosterone levels are essential for the development of the male reproductive system. Ironically, it is a “female” hormone, not testosterone, which causes the changes in gene expression responsible for masculinization of the male brain. Remember that testosterone is converted within neuronal cytoplasm into estradiol in a single chemical step, catalyzed by the enzyme aromatase (see Figure 17.5). The rise in testosterone that occurs prenatally actually leads to an increase in estrogen, which binds to estradiol receptors, thereby triggering masculinization of the developing nervous system. What is not clear is which genes are regulated in various parts of the brain by sex hormones to account for masculinization. Female gonads do not produce an early surge of testosterone or estrogen, so female brains are not affected in the same way as male brains.
An interesting and important complication arises regarding the response of the fetal brain to circulating hormones. In addition to the estrogens and androgens produced by the fetal gonads, hormones coming from the pregnant mother’s placenta reach the circulation of the fetus. A reasonable question is why estrogens from the mother do not alter sexual development of the fetal brain. We have said that estrogen, rather than testosterone, is actually responsible for masculinization, so why doesn’t the female fetus become masculinized in response to estrogens passed to the fetus from the mother? In rats and mice, the answer to this dilemma is that α-fetoprotein, a protein found in high concentrations in fetal blood, binds estrogens and protects the female fetus from masculinization. As odd as it sounds, the female fetus must be protected from “female hormones” to keep its brain from becoming more masculine. Knockout mice that lack α-fetoprotein are sterile and do not exhibit normal sexual behaviors. The role of α-fetoprotein in humans is less clear; there are conflicting reports about whether human α-fetoprotein binds estrogen as it does in rodents. Also noteworthy is the diagnostic measurement of α-fetoprotein levels in maternal blood or amniotic fluid; unusually high levels are a possible indicator of neural tube defects, and unusually low levels are found in Down syndrome.
A study by Amateau and McCarthy shows that one factor in brain masculinization “downstream” from testosterone and estradiol is the production of prostaglandins. Prostaglandins are compounds derived from arachidonic acid, a fatty acid present in the brain and other organs. One of the enzymes involved in the synthesis of prostaglandins is cyclooxygenase (COX). Prostaglandins have numerous roles; most notably, they are produced after tissue damage and are involved in inducing pain and fever. Amateau and McCarthy found that fetal and neonatal male rats exposed to COX inhibitors showed reduced copulatory behavior as adults. Conversely, female rats treated with COX inhibitors exhibited male-like copulatory behaviors; the female rat behavior, and brain, were partially masculinized despite the lack of the usual sex hormones playing this role, thus moving us one step downstream from estradiol in the chain reaction that causes masculinization. A fascinating side note to this study is that human pain is frequently treated with COX inhibitors such as aspirin. Only time will tell whether a human mother’s use of analgesics during pregnancy affects the future sexual behavior of her children.
Mismatches between Genetic Sex and Hormone Action. Under normal circumstances, the genetic sex of an animal or person determines hormonal function and, consequently, the sexual characteristics of the nervous system. However, in situations in which hormonal function is altered, it is possible for genetic males to have female brains and genetic females to have male brains. For example, in all mammalian species studied, treatment with testosterone early in development leads to decreases in at least some features of adult female sexual behavior. Activating fully masculine behavior usually requires extended testosterone treatment before and after birth. If genetically female (XX) rats are exposed to testosterone during the few days around birth, they will fail to elicit the typical female mating posture, called lordosis, when they reach maturity. Female guinea pigs treated in utero with enough testosterone to masculinize their external genitals will, as adults, energetically mount and attempt to mate with females in estrus. In the absence of human intervention, when a cow carries twin calves that include both a male and a female, the female calf is exposed in utero to some testosterone produced by her male twin. As an adult, the female, known as a freemartin, will invariably be infertile and behave more like a bull than a cow.
Some humans also experience mismatches between their chromosomes and sex hormones. For example, genetic males (XY) who carry a defective androgen receptor gene may have profound androgen insensitivity. The androgen receptor gene is on the X chromosome; males thus have only one copy of it, and males with the defective gene cannot produce functioning androgen receptors. These individuals develop normal testes but they remain undescended in the abdomen. The testes produce ample testosterone, but outwardly these individuals appear entirely female because their tissues cannot respond to androgen; they have a vagina, a clitoris, and labia, and at puberty, they develop breasts and a female body shape. The testes also produce normal levels of Müllerian-inhibiting factor so the Müllerian duct does not develop into the female reproductive system, the individuals do not menstruate, and they are infertile. Androgen-insensitive genetic males not only look like normal genetic females, but they also behave like them. Even when they understand the circumstances of their biology, they prefer to call themselves women; they dress like women, and they choose men as their sex partners.
Occasionally, genetic females have a condition called congenital adrenal hyperplasia (CAH), which literally means overgrown adrenal glands present at birth. Although they are genetically female, because their adrenal glands secrete unusually large amounts of androgens, CAH females are exposed to abnormally high levels of circulating androgens early in their development. At birth, they have normal ovaries and no testes, but their external genitals are intermediate in size between a normal clitoris and a penis. Surgery and medications are the usual treatments after birth. Nevertheless, CAH girls (and their parents) are more likely to describe their behavior as aggressive and tomboyish. As adults, most CAH women are heterosexual, but, compared to other women, a higher percentage of CAH women are homosexual. Presumably, by analogy to the animal studies, prenatal exposure to high levels of androgens causes a somewhat male-like organization of certain brain circuits in CAH women. We have to be particularly cautious about drawing conclusions about the causes of human behavior, however (Box 17.3). It is very hard to determine whether masculine behavior of a CAH female is due entirely to early androgen exposure and male-like brain dimorphisms, whether her behavior is the product of subtle differences in the way she is treated by others (particularly parents faced with a child who has ambiguous genitals), or both.
David Reimer was a normal healthy baby boy when he was born in 1965. But during a circumcision, an accident with an electrocautery device burned his entire penis. David’s parents were referred to Johns Hopkins University, where they met Dr. John Money. Because it was not possible to restore David’s male genitals, it was recommended that the boy be castrated and undergo cosmetic surgery, followed by estrogen treatment at puberty, to turn him into a girl. Dr. Money’s recommendation was based on his hypothesis that, at birth, babies are essentially gender-neutral; their identity as male or female is determined by their subsequent life experiences and identification with their anatomy. Faced with a terrible decision, David’s parents were eventually convinced that surgery combined with a female upbringing gave their child the best chance at a normal life.
Dr. Money’s accounts of David’s life after his transformation sound as if the child adapted well and became a happy normal girl. In publications, Dr. Money refers to “John” having been successfully changed into “Joan.” The case even made it into the popular press, as evidenced by a 1973 article in Time magazine: “This dramatic case . . . provides strong support . . . that conventional patterns of masculine and feminine behavior can be altered. It also casts doubt on the theory that major sex differences, psychological as well as anatomical, are immutably set by the genes at conception.”1 At that time, dramatic societal changes were taking place in the roles of men and women, and the success of David as a female appeared to confirm that society created gender identity as much as, or more than, biology.
Unfortunately, a follow-up report revealed that David’s gender transformation was a disaster from the outset. According to David and his twin brother, David’s behavior was always much more like other boys than girls. David rebelled at wearing girls’ clothing and playing with traditional girls’ toys. Despite cosmetic surgery and female indoctrination, as an adult, he said that he had suspected he was a boy as early as the second grade and imagined growing up to be a muscular man. As a child, David was incessantly teased and ostracized. He knew nothing about the failed circumcision and subsequent surgery, nor the fact that he was genetically a male. However, as he got older, he was more attracted to girls than boys, and he expressed the opinion that he felt like a boy trapped in a girl’s body. By the age of 14, after being on estrogen for 2 years, he looked increasingly like a girl, but he stopped living as one (Figure A). David’s father finally told him what had happened when he was young. David immediately requested sex-change hormonal therapy and surgery. For years, David dealt with the overwhelming emotional problems resulting from his past. He married, adopted his wife’s children, and happily worked a physically demanding job as a janitor in a slaughter house. In the 1990s, David collaborated on a book about his life. Tragically, after numerous traumatic events in his life, including the death of his twin brother and the breakup of his marriage, David committed suicide in 2004 at the age of 38.

Figure A David Reimer (aka John/Joan) and his twin brother Brian shortly before they were told the truth about David’s childhood. (Source: Courtesy of Jane Reimer.)
David Reimer’s experiences demonstrate that, rather than being gender-neutral, he had a “male brain” from the outset. Evidently, his genetically determined sex could not be suppressed even with sex-change surgery, hormonal therapy, and a female upbringing. Clearly, gender identity involves a complex interplay of genetics, hormones, and life experiences.
Direct Genetic Effects on Behavior and Sexual Differentiation of the Brain
The classic view of sexual differentiation, and the one discussed in this chapter, gives genetics only an indirect role in determining the gender of an individual: Genes direct the development of the gonads, and the hormonal secretions of the gonads control sexual differentiation. While there is no question that hormones are extremely important in sexual development, recent research suggests that genes may sometimes be more directly involved in sexual differentiation, at least in some species. The most compelling evidence comes from studies of birds. In one particularly dramatic study, Agate et al. examined the body and brain of a rare, naturally occurring zebra finch. This bird, technically a gynandromorph (meaning that it has both male and female tissues), was genetically female on the left side of its body and brain and genetically male on the right side (Figure 17.17). As both sides of the brain are exposed to the same circulating hormones, they should be equally masculine or feminine if hormones are entirely responsible for sexual differentiation. However, the brain areas associated with singing (see Box 17.2) were masculine on the right side and feminine on the left side, suggesting that different gene expression and not sex hormones in the two brain halves led to the brain dimorphisms. In related studies of gynandromorphic chickens, researchers found that most cells on the male side of the body carry male sex chromosomes and cells on the other side have female chromosomes, the result of a mutation early in development.

FIGURE 17.17 Brain analysis of a gynandromorphic zebra finch. (a) This bird had female plumage on its left side and male plumage on its right. (b) The HVC nucleus (hyperstriatum ventrale, pars caudalis) controls singing. It is larger on the genetically male right side (dark patch of labeled neurons). (c) An autoradiogram showing the expression of a gene normally expressed only in females labels only the left side of the brain. (d) An autoradiogram showing the expression of a gene normally more highly expressed in males labels the right side of the brain more than the left. (Source: Arnold, 2004, Fig. 4.) Description
Recently, attempts have been made to find direct genetic effects on sexual differentiation in mammals. In a species in which the male and female forms are not as strikingly different as the zebra finch, a gynandromorph might not be as easily recognized. Eric Vilain and his colleagues at UCLA have found 51 genes that are expressed at different levels in male and female mouse brains before the formation of the gonads. The function of these genes is not yet known.
In addition to influencing sexual differentiation, genes can play a role in surprisingly complex sex behaviors. Some of the best evidence comes from studies of the fruit fly Drosophila melanogaster. The male courts the female with an enticing set of behaviors, including orienting toward and following her, singing a courtship song, and tapping her with his forelegs before attempting to mate. The female chooses whether to accept or reject the male’s advances. Evidently, these behaviors are genetically coded, as males know how to court even if they have never seen courtship by other flies. Many genes may play some role in the courting behaviors, but there appear to be a very small number of critical regulatory genes (genes that regulate the expression of other genes). For example, the fru gene (short for fruitless) may be essential for male courtship behaviors. In males, the fru gene is expressed in a variety of cell types, and this leads to the development of a male central nervous system. The male CNS develops such that male courtship behaviors automatically occur. In females without fru expression, a complete CNS also develops, but its wiring is somewhat different and female behaviors are “built in.” If the fru gene is absent in a male fly, male courtship behaviors are significantly reduced or nonexistent. Conversely, females made to express fru exhibit male courtship behaviors and resist courtship by males.
Another gene involved in sexual differentiation is dsx (for double sex). The dsx gene plays an important role in the sexual differentiation of the body (the development of male or female genitals) and also interacts with fru in controlling sexual differentiation of the CNS and sex-specific behaviors. In the case of fru, the gene is either expressed (males) or not expressed (females). The dsx gene is different in that it is expressed in both males and females, but alternative splicing leads to the production of male-specific and female-specific proteins. How the fru- and dsx-influenced structure of the CNS assures sex-specific behaviors is a challenging mystery yet to be explained in detail.
Long after sex hormones have determined the structure of the reproductive organs, they can have activational effects on the brain. These effects range in scale from temporary modifications in brain organization to changes in the structure of neurites. In men, testosterone has a two-way interaction with sexual behavior. On the one hand, testosterone levels rise in anticipation of a sexual act or even while fantasizing about it. Conversely, reduced testosterone levels are associated with decreased sexual interest. It has been reported that women are more likely to initiate sex when estradiol levels are highest during their menstrual cycle. Through unknown mechanisms, hormone levels in both sexes influence the brain and an individual’s interest in sexual behavior.
Brain Changes Associated with Maternal and Paternal Behavior. Patterns of sexual behavior vary over time. In some species, reproduction occurs only during a particular season, and mating may occur only during a specific phase of that season. Obviously, females of all species nurse their offspring only after birth and only temporarily. In most animals, but not in humans, sexual attractiveness and copulation occur only during certain phases of the estrous cycle. Sexually dimorphic changes in the brain are sometimes transient or cyclical, coinciding with the sexual behavior to which they are related.
In Chapter 16, we saw that appetite is controlled in part by blood levels of the hormone leptin that is secreted by fat cells: Higher leptin levels modulate cells in the hypothalamus and suppress eating. During pregnancy, the mother needs more food to supply energy for her growing fetus, and indeed, food intake increases early in pregnancy. Consequently, body fat accumulates and leptin levels rise. Paradoxically, researchers have observed in rats that even though leptin levels rise during pregnancy, appetite and food intake increase rather than decrease. This occurs because hormonal changes associated with pregnancy lead to leptin resistance in the hypothalamus.
Another behavior unique to maternal rats occurs with lactation and nursing. In female rats, the somatosensory cortex contains a sensory representation of the ventral skin surrounding the nipples. Within a few days of the start of nursing, the tactile stimulation leads to a dramatic increase in the representation of the ventral skin (Figure 17.18) and a shrinkage of receptive fields to half their normal size. This interesting example of somatosensory map plasticity (see Chapter 12) appears to be temporary, as receptive fields are normal in size several months after weaning.

FIGURE 17.18 The effect of lactation on a sensory representation in the cortex. (a) The ventral skin of a nursing rat mother, showing the location of the nipples along the right side. (b) The rat’s brain and left primary somatosensory cortex, which contains a map of the right ventral skin (top). The boxed area, expanded below, illustrates how the cortical region that responds to the ventral skin around the nipples is enlarged in a postpartum lactating mother (left) compared to a postpartum nonlactating mother (right). Regions of somatosensory cortex subserving other regions of the body were not affected by the lactating state. (Source: Adapted from Xerri et al., 1994.) Description
Lactation also appears to lead to brain changes that reinforce this behavior, which is critical for the survival of mammalian offspring. Despite their very different pharmacological and behavioral effects, all addictive drugs appear to enhance the influence of dopamine released by neurons projecting from the ventral tegmental area (VTA) into the nucleus accumbens (NA) (see Box 16.5). It is becoming increasingly clear that a variety of reinforcing or addictive behaviors also modify this VTA-NA circuit. In one study, fMRI scans of lactating female rats nursing their pups were compared with those of virgin female rats after an injection of cocaine. Brain activation was surprisingly similar in both, with particular activation in the NA. The hypothesis is that in both instances, there is stimulation of the dopamine system associated with reward and addiction. The tactile stimulation of the suckling pups may make nursing a reinforcing behavior to promote mother–infant bonding and, ultimately, the survival of the pups.
Even though fathers do not experience the dramatic body changes associated with pregnancy and lactation, their interactions with children may alter their brains in fundamental ways. A hint of this comes from a study in Elizabeth Gould’s lab at Princeton University, where she examined the brains of marmosets. Marmoset fathers are unusually involved in child care; indeed, they carry the babies around for the first few months of life. It is known that across species, the prefrontal cortex is involved in complex, goal-directed behaviors. It is also established that environment can change neurons; for example, dendritic branching and spine density are enhanced when animals are housed in an enriched environment. To see whether fatherhood alters brain structure, Gould’s group compared the prefrontal cortex of marmoset fathers with that of non-fathers who were part of a mating pair. Two interesting differences were found: The density of dendritic spines on pyramidal cells was significantly higher in the fathers, and there appeared to be more vasopressin receptors on the spines. The functional consequences of these changes are not known, but they suggest that brains in other species that invest much time in childrearing, whether by the male or female, might be structurally changed by the experience.
Estrogen Effects on Neuron Function, Memory, and Disease. Estrogens have powerful activational effects on the structure and function of neurons. Within minutes of experimental application, estradiol alters the intrinsic excitability of neurons in a broad range of brain areas. By modulating the flow of potassium ions, estradiol depolarizes some neurons and causes them to fire more action potentials. A dramatic example of the effects estrogen can have on cell structure is shown in Figure 17.19. Dominique Toran-Allerand at Columbia University found that estradiol treatment of tissue taken from the hypothalamus of newborn mice causes a great degree of neurite outgrowth. Other studies have shown that estradiol increases cell viability and spine density. Taken together, these findings suggest that estrogens play an important role in forming neuronal circuitry during brain development.

FIGURE 17.19 The effect of estrogen on neurite growth in the hypothalamus. Below the bottom of each photograph is a piece of hypothalamic tissue from a newborn mouse. (a) Without the addition of estrogen, a relatively small number of neurites grow from the tissue. (b) With the addition of estrogen, there is an exuberant growth of neurites. (Source: Toran-Allerand, 1980.) Description
Working at Rockefeller University, Elizabeth Gould, Catherine Woolley, Bruce McEwen, and their colleagues reported a fascinating example of estradiol’s activational effects. They counted dendritic spines on neurons in the hippocampus of female rats and found that the number of spines fluctuated dramatically during the 5-day estrous cycle. Spine density and estradiol levels peaked together, and treatment with injected estradiol also increased the numbers of spines in animals whose natural estradiol levels were kept low (Figure 17.20). Since spines are the major site of excitatory synapses on dendrites (see Chapter 2), this provides a possible explanation for the fact that hippocampal excitability also seems to track the estrous cycle. The hippocampus of experimental animals more easily generates seizures, for example, when estrogen levels increase (Figure 17.21). Note that estradiol and progesterone levels peak during the proestrus phase (Figure 17.21a,b) and at this time seizure thresholds are lowest (Figure 17.21c). Woolley and McEwen showed that it is indeed estradiol itself that triggers the increase in spine numbers and that as hippocampal neurons grow more spines, they also grow more excitatory synapses.

FIGURE 17.20 An activational effect of steroid hormones. Estradiol treatment causes an increase in the number of dendritic spines on hippocampal neurons. (Source: Adapted from Woolley et al., 1997.) Description

FIGURE 17.21 Fluctuations of hormone levels during the estrous cycle and hippocampal seizure threshold. The circulating levels of (a) estradiol and (b) progesterone vary during the estrous cycle. Levels of both hormones peak during the proestrus phase. (c) The threshold (in terms of stimulating current) for triggering a seizure in the hippocampus of a female rat varies over the estrous cycle and is lowest during the proestrus phase. The phases of the estrous cycle are D, diestrus; P, proestrus; E, estrous. (Sources: Parts a and b, Smith et al., 1975; part c, Terasawa et al., 1968.) Description
How does estradiol increase the numbers of hippocampal spines and excitatory synapses? Although the details of this mechanism are not all clear, it appears that estradiol increases the capacity for synaptic plasticity in the hippocampus in multiple ways. In the presence of estradiol, postsynaptic responses to glutamate are larger than without estradiol. As we will see in Chapter 25, such enhanced responses at excitatory synapses cause the synapses to strengthen. Estradiol may also alter hippocampal function by decreasing synaptic inhibition. Estradiol causes some inhibitory cells to produce less GABA, their neurotransmitter, and therefore synaptic inhibition becomes less effective. Less inhibition increases neural activity, complementing the estradiol effect at excitatory synapses. When the pieces are put together, it seems that estradiol produces a hippocampus with less effective inhibitory synapses and stronger excitatory synapses, thereby triggering an increase in the number of spines on the pyramidal cells.
In rats, the hippocampus is particularly important for spatial memory and navigational skills, and several studies demonstrate that estradiol enhances such forms of memory formation. In these experiments, rats are trained to run a maze or remember objects or places. Estradiol administered shortly before or after training enhances performance on these tasks when they are performed again hours later as a test of memory. It is interesting to note that the memory benefit associated with the estradiol disappears if the estrogen is given 2 hours after training. Evidently, estrogens can facilitate memory, but they must be present near in time to the learning experience.
Woolley notes that the peak in hippocampal spine number coincides with the rat’s peak in fertility. During that period, the female actively seeks out mates, which may require the heightened spatial ability that might come with a more excitable, NMDA-receptor-filled hippocampus. Thus, the female rat’s brain may fine-tune itself on a 5-day cycle in order to meet changing reproductive needs.
Estradiol has also been shown to have a protective effect on neurons that may help combat disease. In neuronal cultures, cells are more likely to survive hypoxia, oxidative stress, and exposure to various neurotoxic agents if they are exposed to estradiol. Clinically, estrogen appears to minimize or delay neural damage in a variety of situations. For example, it may protect against stroke in humans, although the mechanism is unclear. This observation could be related to the finding that tamoxifen, an estrogen receptor antagonist often used in the treatment of breast cancer, is associated with an increased risk of stroke in women. Estrogen replacement therapy appears to aid in the treatment of certain neurological disorders. Researchers have observed that increased levels of sex hormones during pregnancy are associated with a reduced severity of multiple sclerosis, and there is some evidence that estrogen may benefit women with multiple sclerosis. It also appears that estrogen replacement therapy might delay the onset of Alzheimer’s disease and decrease tremors in Parkinson’s disease. The actions of estrogens in these diseases have been difficult to pin down, in part because a variety of cell types express estrogen receptors. Indeed, recent evidence suggests that the benefits of estrogen may come from effects on astrocytes as well as neurons.
Estimates are that about 3–10% of the American population is homosexual. In light of the behavioral differences between, for example, male homosexuals and male heterosexuals, is the anatomy or physiology of the brain different? Is there a biological basis for sexual orientation? In some sense, this must be true if we believe that all behavior is based on brain activity. However, there is no evidence that sexual orientation is related to activational effects of hormones in adults. For example, administering androgens or estrogens to adults, or removing the gonads, has no effect on sexual orientation. As an alternative, perhaps homosexual and heterosexual brains are structurally different due to organizational effects.
We saw earlier that in animals, there are sex differences in the anterior hypothalamus. In rats, the SDN (sexually dimorphic nucleus) in the preoptic area of the anterior hypothalamus is much larger in males than females. After a surgical lesion to this brain area, male rats spend more time with sexually active males than with sexually receptive females, a reversal of their preference before surgery. Another piece of suggestive evidence comes from studies of Rocky Mountain bighorn sheep, in which researchers estimate that about 8% of the male population prefers mounting other males rather than females. The SDN is found to be about half the size in these male-oriented rams compared to female-oriented rams. Thus, it appears that the size of hypothalamic nuclei in some animals may correlate with sexual preference. Unfortunately, the causal relationship between SDN size and sexual orientation is unclear.
In humans, the INAH-3 nucleus (one of the interstitial nuclei of the anterior hypothalamus) is about twice as large in men as in women, a difference that may be related to sexually dimorphic behavior. Some studies of the INAH suggest that there are differences between homosexual and heterosexual brains that might be related to sexual orientation. Simon LeVay, then working at the Salk Institute, found that INAH-3 in gay men is only half the size of the nucleus in straight men (Figure 17.22). In other words, INAH-3 in homosexual men is similar in size to that of women. While this finding may indicate a biological basis for homosexuality, it is difficult to interpret this in terms of complex human behavior. Moreover, subsequent studies have not always confirmed a correlation between INAH-3 size and sexual orientation.

FIGURE 17.22 The location and size of INAH-3. (a) The location of the four INAH nuclei in the hypothalamus. In the micrographs, arrows indicate INAH-3 in (b) a heterosexual man and (c) a homosexual man. In the homosexual, the nucleus is smaller and the cells more scattered. (Source: Micrographs from LeVay, 1991, p. 1035.) Description
Other research has found the anterior commissure and suprachiasmatic nucleus to be larger in male homosexuals than in male heterosexuals. One study reported that the bed nucleus of the stria terminalis is larger in men than women and that male-to-female transsexuals have a nucleus comparable in size to females. Collectively, these studies offer the intriguing prospect that complex aspects of human sexual behavior may ultimately be linked to distinct brain organization. However, the difficulties involved in comparing brains, as well as the history of brain dimorphisms, suggest that caution is advised until a research consensus is reached.
The subject of sex and the brain is complicated by the subtleties of the biological and cultural mechanisms that determine sexual behavior. Particularly in humans, anatomical differences between the female and male nervous systems are not readily apparent, and indeed, most human behavior is not distinctly masculine or feminine. Where small brain differences between the sexes occur, any adaptive purpose they may serve is not clear. And in no case is the neurobiological basis for sex differences in cognition known.
Nevertheless, the essential biological imperative—reproduction—demands sex-specific behaviors, at least for mating and giving birth. For the most concretely sexual structures (such as the muscles and motor neurons controlling the penis or the sensory afferents innervating the clitoris), identifying some of the peripheral and spinal neural systems involved is fairly easy. The powerful role of sex hormones in sexual development and behavior is also clear. But the more complex aspects of sexual behavior and the brain systems that generate them are still quite mysterious.
We have touched on only a few of the issues in the study of sex and the brain, and most of the basic questions remain unanswered. Scientific research about sex was long hampered by society’s reluctance to talk openly about the subject, and sexual politics still tend to muddy the scientific waters today. But sexual behavior is a defining feature of being human, and understanding its neural basis is a worthy challenge.
sex-determining region of the Y chromosome (SRY) (p. 582)
interstitial nuclei of the anterior hypothalamus (INAH) (p. 597)
1. Suppose you have just been captured by aliens who have landed on Earth to learn about humans. The aliens are all one sex, and they are curious about the two human sexes. To earn your freedom, all you must do is tell them how to reliably distinguish between males and females. What biological and/or behavioral tests do you tell them to conduct? Be sure to describe any exceptions that might violate your tests; you don’t want the aliens to get angry!
2. Figure 17.18 shows an interesting but unexplained observation: In the brain of a mother rat during periods of lactation, the size of the somatosensory cortex representing the skin around the nipples expands. Speculate about a likely mechanism for this phenomenon. Suggest a reason why such brain plasticity might be advantageous.
3. Estradiol is usually described as a female sex hormone, but it also plays a critical role in the early development of the male brain. Explain how this happens and why the female brain is not similarly affected by estradiol at the same stage of development.
4. Where and how can steroid hormones influence neurons in the brain at the cellular level?
5. What evidence supports the hypothesis that sexual differentiation of the body and brain is not entirely dependent on sex hormones?
6. Suppose that a research team has just claimed that a small, obscure nucleus in the brain stem, nucleus X, is sexually dimorphic and essential for certain “uniquely male” sexual behaviors. Discuss the kinds of evidence you would need to accept these claims about (a) the existence of a dimorphism, (b) the definitions of uniquely male behaviors, and (c) the involvement of nucleus X in these sexual behaviors.
Arnold AP. 2004. Sex chromosomes and brain gender. Nature Reviews Neuroscience 5:701–708.
Bartels A, Zeki S. 2004. The neural correlates of maternal and romantic love. Neuroimage 21:1155–1166.
Colapinto J. 2001. As Nature Made Him: The Boy Who Was Raised as a Girl. New York: Harper Collins.
De Boer A, van Buel EM, ter Horst GJ. 2012. Love is more than just a kiss: a neurobiological perspective on love and affection. Neuroscience 201:114–124.
Hines M. 2011. Gender development and the human brain. Annual Review of Neuroscience 34:69–88.
Pfaus JG. 2009. Pathways of sexual desire. Journal of Sexual Medicine 6:1506–1533.
Valente SM, LeVay S. 2003. Human Sexuality. Sunderland, MA: Sinauer.
Wooley CS. 2007. Acute effects of estrogen on neuronal physiology. Annual Review of Pharmacology and Toxicology 47:657–680.
Wu MV, Shah NM. 2011. Control of masculinization of the brain and behavior. Current Opinion in Neurobiology 21:116–123.
Young KA, Gobrogge KL, Liu Y, Wang Z. 2011. The neurobiology of pair bonding: insights from a socially monogamous rodent. Frontiers in Neuroendocrinology 32:53–69.
Additional figures