Brain Mechanisms of Emotion
BOX 18.1 OF SPECIAL INTEREST: Butterflies in the Stomach

Difficulties with the Concept of a Single System for Emotions
BOX 18.3 PATH OF DISCOVERY: Concepts and Names in Everyday Science, by Antonio Damasio
BOX 18.4 OF SPECIAL INTEREST: The Frontal Lobotomy
Neural Components of Anger and Aggression Beyond the Amygdala
To appreciate the significance of emotions, just imagine life without them. Instead of the daily highs and lows we all experience, life would likely feel like a great empty plain of existence with little significance. Emotional experiences are a large part of being human. In books and movies, aliens and robots may look like people, but they usually seem inhuman simply because they exhibit no emotions.
Affective neuroscience is the investigation of the neural basis of emotion and mood. In this chapter, we explore emotion. Affective disorders, also known as mood disorders, are discussed in Chapter 22. You might be wondering how we can study something as ephemeral as one’s feelings. When studying a sensory system, in contrast, you can present a stimulus and seek the neurons that respond to it. You can manipulate the stimulus to determine the stimulus attributes (light intensity, sound frequency, etc.) that are best for evoking a response. It is not as straightforward to study emotions in animals that cannot tell us their subjective feelings, however. What we observe are behaviors presumed to be expressions of internal emotions. We must be careful to distinguish between emotional experience (feelings) and emotional expression. What we know about the brain mechanisms of emotion has been derived from a synthesis of animal and human studies. In animals, brain activity and the effects of brain lesions on behavior have been noted and interpreted in the context of emotions, even though we cannot determine the animals’ feelings. Studies in humans have examined brain activity associated with emotional experience and the recognition of emotion in others.
We are not yet at a stage where we can map out an emotion system the same way that sensory systems have been outlined. Indeed, we will see that earlier ideas about a single emotion system or multiple systems involving brain areas dedicated to particular emotions have been replaced by theories in which emotions are based on distributed networks of brain activity.
Emotions—love, hate, happiness, sadness, fear, anxiety, and so on—are feelings we all experience at one time or another. But what precisely defines those feelings? Are they sensory signals from our body, diffuse patterns of activity in our cortex, or something else?
In the nineteenth century, several highly regarded scientists, including Darwin and Freud, considered the role of the brain in the expression of emotion (Figure 18.1). This early research was based on the careful study of emotional expression in animals and humans and emotional experience in humans. It may seem like common sense to many now, but Darwin made the important observations that people in different cultures experience the same emotions and that animals appear to express some of the same emotions as humans. Later in the nineteenth and twentieth centuries, scientists developed theories for the physiological basis of emotion and the relationship between emotional expression and experience.

FIGURE 18.1 Expressions of anger in animals and humans. These drawings are from Darwin’s book The Expression of the Emotions in Man and Animals; they were used to support his claim that there are basic universal emotions. Darwin conducted one of the first extensive studies of emotional expression. Reproduced with permission from John van Wyhe, ed. 2002. The Complete Work of Charles Darwin Online. (http://darwin-online.org.uk/)
One of the first well-articulated theories of emotion was proposed in 1884 by the renowned American psychologist and philosopher William James. Similar ideas were proposed by Danish psychologist Carl Lange. This theory, commonly known as the James–Lange theory of emotion, proposed that we experience emotion in response to physiological changes in our body. To understand why many contemporaries of James and Lange considered this idea counterintuitive, consider an example.
Suppose you wake one morning to find a malicious-looking spider hanging from a web above your bed. If you like many people have arachnophobia, you may experience a fight-or-flight response that involves changes in heart rate, muscle tone, and lung function (see Chapter 15). According to the James–Lange theory, your visual system sends an image of the spider to your brain and in response the brain issues commands to the somatic and autonomic nervous systems that alter muscle and organ function. These responses of the body follow directly from the sensory input without any emotional component. The emotion you experience consists of your feelings that result from the changes in the body. In other words, rather than jumping out of bed in response to being scared, you actually feel scared because you become aware of your racing heart and tensed muscles. This seems like a backward idea to many people today, as it did to many contemporaries of James and Lange. Until this theory was proposed, the common conception was that an emotion is evoked by a situation and the body changes in response to the emotion: You become scared when you see a spider and then your body reacts. The James–Lange theory is the exact opposite.
Consider one of the thought experiments suggested by James. Suppose you are boiling with anger about something that has just happened. Try to strip away all the physiological changes associated with the emotion. Your pounding heart is calmed, your tensed muscles are relaxed, and your flushed face is cooled. As James said, it is hard to imagine maintaining rage in the absence of any of these physiological responses.
Even if it is true that emotion follows from changes in the body’s physiological state, this doesn’t mean that emotion cannot be felt in the absence of obvious physiological signs (a point even James and Lange would concede). But for strong emotions that are typically associated with physical change, the James–Lange theory says the bodily changes cause the emotion rather than the other way around.
Although the James–Lange theory became popular in the early twentieth century, it soon came under attack. In 1927, American physiologist Walter Cannon published a paper that offered several compelling criticisms of the James–Lange theory and proposed a new theory. Cannon’s theory was modified by Philip Bard and became known as the Cannon–Bard theory of emotion. It proposed that emotional experience can occur independently of emotional expression.
One of Cannon’s arguments against the James–Lange theory was that emotions can be experienced even if physiological changes cannot be sensed. To support this claim, he offered the cases of animals he and others studied after transection of the spinal cord. Such surgery eliminated body sensations below the level of the cut, but it did not appear to abolish emotion. To the extent possible with muscular control of just the upper body or head, the animals still exhibited signs of experiencing emotions. Similarly, Cannon noted human cases in which a transected spinal cord did not diminish emotion. If emotional experience occurs when the brain senses physiological changes in the body, as the James–Lange theory proposed, then eliminating sensation should also eliminate emotions, and this did not appear to be the case.
A second observation of Cannon’s that seems inconsistent with the James–Lange theory is the lack of a reliable correlation between the experience of emotion and the physiological state of the body. For example, fear is accompanied by increased heart rate, inhibited digestion, and increased sweating. However, these same physiological changes accompany other emotions, such as anger, and even nonemotional conditions of illness, such as fever. How can fear be a consequence of the physiological changes when these same changes are associated with states other than fear?
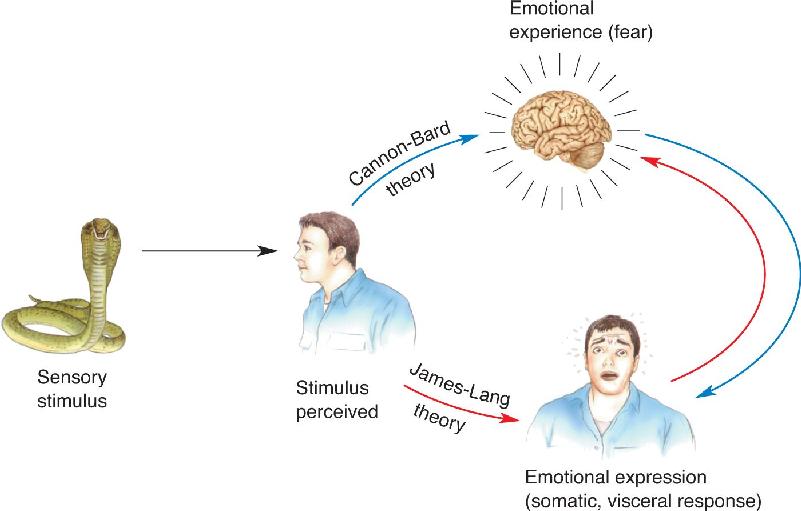
Cannon’s new theory focused on the idea that the thalamus plays a special role in emotional sensations. The theory proposes that sensory input is received by the cerebral cortex, which in turn activates certain changes in the body. But according to Cannon, this stimulus–response neural loop is devoid of emotion. Emotions are produced when signals reach the thalamus either directly from the sensory receptors or by descending cortical input. In other words, the character of the emotion is determined by the pattern of activation of the thalamus irrespective of the physiological response to the sensory input. An example may clarify the difference between this and the James–Lange theory. According to James and Lange, you feel sad when you sense that you are crying; if you could prevent the crying, the sadness should go, too. In Cannon’s theory, you don’t have to cry to feel sad; there simply has to be the appropriate activation of your thalamus in response to the situation. The James–Lange and Cannon–Bard theories of emotion are compared in Figure 18.2.

FIGURE 18.2 A comparison of the James–Lange and Cannon–Bard theories of emotion. In the James–Lange theory (red arrows), the man perceives the threatening animal and reacts. When he senses his body’s response to the situation, he becomes afraid. In the Cannon–Bard theory (blue arrows), the threatening stimulus first causes the feeling of fear, and the man’s reaction follows. Description
Many theories of emotion have been proposed since the days of the James–Lange and Cannon–Bard theories. Subsequent work has demonstrated that each of the older theories has merits as well as flaws. For instance, contrary to Cannon’s statements, fear and rage have been shown to be associated with distinguishable physiological responses, even though they both activate the sympathetic division of the autonomic nervous system (ANS). Although this does not prove that these emotions result from distinct physiological responses, the responses are at least different (Box 18.1). Research has also shown that to some extent we can be aware of our body’s autonomic function (called interoceptive awareness), a key component of the James–Lange theory. For example, people have been shown to be capable of judging the timing of their heartbeat, and increased activity is found in certain brain areas when this task is performed.
Human language has colorful ways of describing emotional experiences. If someone hesitates before bungee jumping off a high bridge, we describe their fear by saying they have “cold feet.” At the other end of the temperature spectrum, a person who readily becomes angry is described as a “hot head.” Nervous before going out with someone new? You may be experiencing “butterflies in the stomach.” These descriptive terms are fun, but do they have the slightest relationship to the physiological experience of emotion?
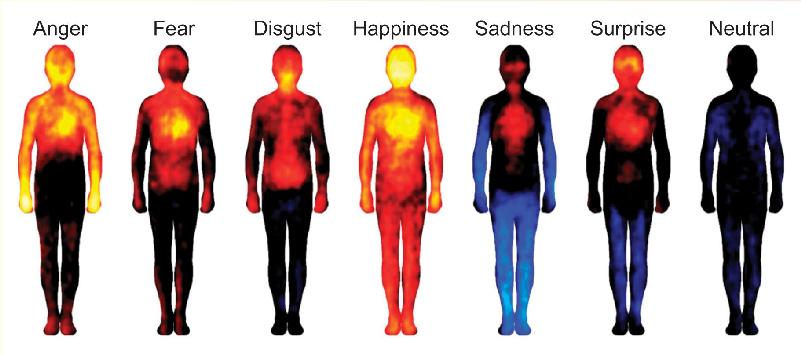
An intriguing study conducted by scientists at Aalto University in Finland suggests that basic emotions and some other emotions may indeed be associated with unique maps of sensory changes spread across the body. This conclusion was reached based on online testing of over 700 people in Finland, Sweden, and Taiwan. In order to itemize which parts of the body were felt to be affected by an emotion, the experimenters asked participants to color a map of the body using warm colors where they felt an emotion made the body more active and cool colors where the body was made less active. Emotion maps were made in response to a variety of stimuli including emotion words, pictures of emotional facial expressions, emotional experiences in short stories, and emotional scenes in movies. The hope was that by studying participants in different cultures and languages, universal emotional experiences could be mapped rather than cultural stereotypes.
The figure shows maps of presumptive body activity averaged across many observers. Red and yellow indicate elevated activity, and blue reduced activity, relative to a neutral (black) state. Some traits, such as elevated head and chest activity (elevated heart and respiration rates?), were common to multiple emotions. Other characteristics were more unique. Happiness was unusual in the extent to which the entire body showed increased activity, and sadness had a unique lowering of activity in the extremities. The body map for disgust had an odd elevation in activity around the digestive tract and throat (a gag reflex?). What do these colorful maps represent? We can only speculate, but perhaps they are related to patterns of sensation and activation in the autonomic nervous system. Obviously, one has to be cautious interpreting the maps, but it is intriguing that the different emotion maps are distinguishable, and this was even true to some extent for emotions not considered “basic.” Also interesting is the finding that the emotion maps are similar across cultures. Even though we can’t take a snapshot of butterflies in the stomach, these findings are consistent with Darwin’s idea that at least some emotions are unique experiences that are universal across cultures.

Figure A Color maps of six basic emotions. Estimates of body activation range from low (blue) to high (yellow). (Source: Adapted from Nummenmaa L, Glerean E, Hari R, Hietanen JK. 2014. Bodily maps of emotions. Proceedings of the National Academy of Science 111:646–651, Figure 1.)
Another interesting challenge to the Cannon–Bard theory, demonstrated by later studies, is that emotion is sometimes affected by damage to the spinal cord. In one study of adult men with spinal injuries, there was a correlation between the extent of sensory loss and reported decreases in emotional experiences, although other studies of people with spinal injuries have not always found a similar correlation. Shortly, we will examine more recent theories of emotion in the context of experimental results that suggest the brain structures involved in the experience and expression of emotion.
Although the findings may be counterintuitive, some studies suggest that sensory input can have emotional effects on the brain without our being aware of the stimuli. Several related experiments were conducted by Arne Öhman, Ray Dolan, and their colleagues in Sweden and England. They first showed that if an angry face is briefly flashed and quickly followed by a briefly flashed photo of an expressionless face, subjects report seeing only the expressionless face. The angry face is said to be perceptually “masked,” and the expressionless face is the masking stimulus.
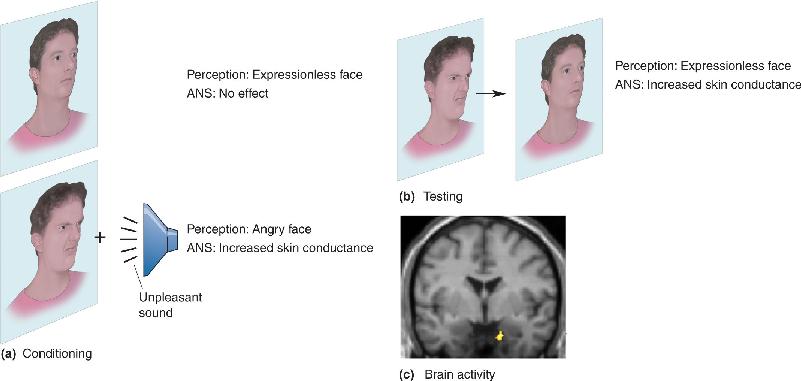
In one experiment, subjects were shown a variety of faces without a masking stimulus; and each time an angry face was shown, the subject’s finger got a mild electrical shock. After aversive conditioning like this, subjects exhibited altered autonomic activity, such as increased skin conductance (sweaty palms), when the angry faces were shown again. The researchers were interested in what happens when the angry faces are occasionally shown after training but the masking stimulus is reintroduced. Surprisingly, when angry faces were shown, the subjects had an autonomic response (increased skin conductance), even though they were unaware of the angry faces. These findings indicate that the subjects responded to the angry expressions on the aversive face stimuli even though they were not perceptually aware of seeing the faces at all. The concept of an unconscious emotion is based on this observation.
In a second experiment, subjects were shown angry faces with or without a loud unpleasant sound (Figure 18.3). As before, the subjects did not perceive the angry faces when a subsequent masking stimulus was presented. Nonetheless, skin conductance showed that the subjects responded to the angry faces that had been paired with the sound. In addition, positron emission tomography (PET) imaging was used to record brain activity while the photos were presented. The brain images revealed that the angry faces conditioned to be unpleasant evoked greater activity in the brain in a particular location, the amygdala. We will have more to say about the amygdala later in the chapter. For now, the important point to remember is that measures of both autonomic response and amygdala activity correlate with the presentation of angry faces that are conditioned to be unpleasant despite the fact that the faces are not perceived.

FIGURE 18.3 Unconscious emotional brain activity. (a) Human subjects were conditioned using photos of expressionless and angry faces. Subjects responded to the angry face, paired with a loud unpleasant sound, with increased ANS activity (skin conductance). (b) In the testing phase, an angry face was shown briefly, followed immediately by an expressionless face. Subjects reported seeing only the expressionless face, but increased skin conductance still occurred. (c) Despite the fact that the angry face was not perceived in the testing phase, amygdala activation (red and yellow) occurred only when an angry face preceded the masking stimulus. (Source: Morris, Öhman, and Dolan, 1998.) Description
If sensory signals can have emotional impact on the brain without our being aware of it, this seems to rule out theories of emotion in which emotional experience is a prerequisite for emotional expression. But even with this conclusion, there are many possible ways for the brain to process emotional information. We now turn to the pathways in the brain that link sensations (inputs) to the behavioral responses (outputs) that characterize emotional experience. In the remainder of this chapter, we will see that different emotions may depend on different neural circuits, but some parts of the brain are important for multiple emotions.
Previous chapters discussed how sensory information from peripheral receptors is processed along clearly defined, anatomically distinct pathways to the neocortex. The components of a pathway collectively constitute a system. For example, neurons located in the retina, lateral geniculate nucleus, and striate cortex work together to serve vision, so we say they are part of the visual system. Is there a system, in this sense, that processes the experiencing of emotions? Beginning around 1930, some scientists argued that there is, and it came to be known as the limbic system. Shortly, we will discuss the difficulties of trying to define a single system for emotion. But first, let’s examine the origin of the limbic system concept.
In a paper published in 1878, French neurologist Paul Broca noted that, on the medial surface of the cerebrum, all mammals have a group of cortical areas that are distinctly different from the surrounding cortex. Using the Latin word for “border” (limbus), Broca named this collection of cortical areas the limbic lobe because they form a ring or border around the brain stem (Figure 18.4). According to this definition, the limbic lobe consists of the cortex around the corpus callosum (mainly the cingulate gyrus), the cortex on the medial surface of the temporal lobe, and the hippocampus. Broca did not write about the importance of these structures for emotion, and for some time, they were thought to be primarily involved in olfaction. However, the word limbic, and the structures in Broca’s limbic lobe, were subsequently closely associated with emotion.

FIGURE 18.4 The limbic lobe. Broca defined the limbic lobe as the structures that form a ring around the brain stem and corpus callosum on the medial walls of the brain. The main structures in the limbic lobe labeled here are the cingulate gyrus, medial temporal cortex, and the hippocampus. The brain stem has been removed in the illustration to make the medial surface of the temporal lobe visible.
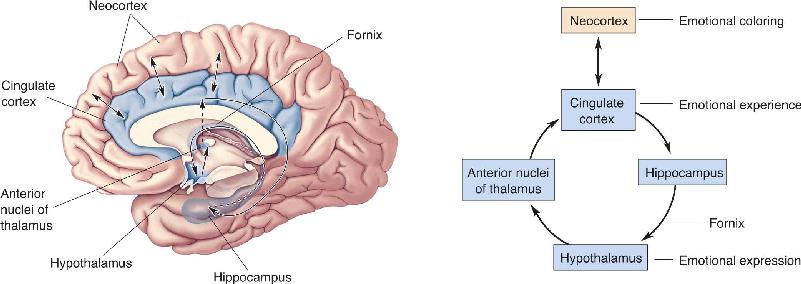
By the 1930s, evidence suggested that a number of limbic structures are involved in emotion. Reflecting on the earlier work of Cannon, Bard, and others, American neurologist James Papez proposed that there is an “emotion system,” lying on the medial wall of the brain that links the cortex with the hypothalamus. Figure 18.5 shows the group of structures that have come to be called the Papez circuit. Each is connected to another by a major fiber tract.

FIGURE 18.5 The Papez circuit. Papez believed that the experience of emotion was determined by activity in the cingulate cortex and, less directly, other cortical areas. Emotional expression was thought to be governed by the hypothalamus. The cingulate cortex projects to the hippocampus, and the hippocampus projects to the hypothalamus by way of the bundle of axons called the fornix. Hypothalamic effects reach the cortex via a relay in the anterior thalamic nuclei. Description
Papez believed, as do many scientists today, that the cortex is critically involved in the experience of emotion. Following damage to certain cortical areas, there are sometimes profound changes in emotional expression with little change in perception or intelligence (Box 18.2). Also, tumors located near the cingulate cortex are associated with certain emotional disturbances, including fear, irritability, and depression. Papez proposed that activity evoked in other neocortical areas by projections from the cingulate cortex adds “emotional coloring” to our experiences.
Brain damage can sometimes have a profound influence on a person’s personality. One of the most famous examples is the case of Phineas Gage. On September 13, 1848, while tamping explosive powder into a hole to prepare for blasting at a railroad construction site in Vermont, Phineas made the mistake of looking away from what he was doing. His tamping iron hit a rock and sparked an explosion. The consequences are described by Dr. John Harlow in an 1848 article entitled “Passage of an Iron Rod Through the Head.” When the charge went off, it sent Phineas’s meter-long, 6-kg iron tamping rod into his head just below his left eye. After passing through his left frontal lobe, the rod exited the top of Gage’s head.
Incredibly, after being carried to an ox cart, Gage sat upright on the ride to a nearby hotel and walked up a long flight of stairs to go inside. When Harlow first saw Gage at the hotel, he commented that “the picture presented was, to one unaccustomed to military surgery, truly terrific” (p. 390). As you might imagine, the projectile destroyed a considerable portion of the skull and left frontal lobe, and Gage lost a great deal of blood. The hole through his head was more than 9 cm in diameter. Harlow was able to stick the full length of his index finger into the hole from the top of Gage’s head, and also upward from the hole in his cheek. Harlow dressed the wound as best he could. Over the following weeks, considerable infection developed. No one would have been surprised if the man died. But about a month after the accident, he was out of bed and walking around town.
Harlow corresponded with Gage’s family for many years and in 1868 published a second article, “Recovery from the Passage of an Iron Bar Through the Head,” describing Gage’s life after the accident. After Gage recovered from his wounds, he was apparently normal except for one thing: His personality was drastically and permanently changed. When he tried to return to his old job as construction foreman, the company found he had changed so much for the worse that they wouldn’t rehire him. According to Harlow, before the accident Gage was considered “the most efficient and capable foreman. . . . He possessed a well-balanced mind, and was looked upon by those who knew him as a shrewd, smart business man, very persistent in executing all his plans of operation” (pp. 339–340). After the accident, Harlow described him as follows:
The equilibrium or balance, so to speak, between his intellectual faculties and animal propensities, seems to have been destroyed. He is fitful, irreverent, indulging at times in the grossest profanity (which was not previously his custom), manifesting but little deference for his fellows, impatient of restraint or advice when it conflicts with his desires, at times pertinaciously obstinate, yet capricious and vacillating, devising many plans of future operation, which are no sooner arranged than they are abandoned in turn for others appearing more feasible. . . . His mind was radically changed, so decidedly that his friends and acquaintances said he was “no longer Gage.” (pp. 339–340)
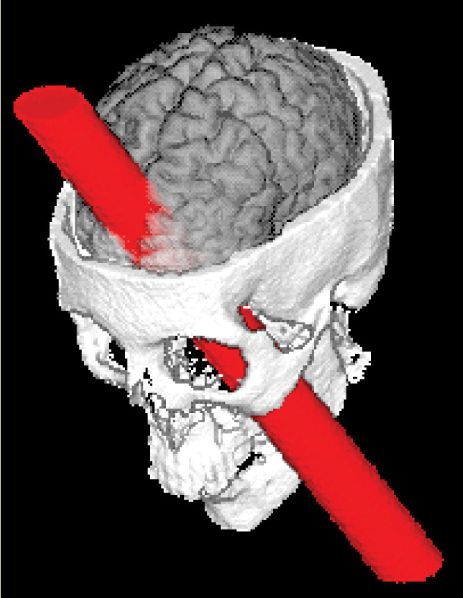
Phineas lived for another 12 years; when he died, no autopsy was performed. However, Gage’s skull and the tamping iron have been preserved in a museum at Harvard Medical School. In 1994, Hanna and Antonio Damasio and their colleagues at the University of Iowa made new measurements of the skull and used modern imaging techniques to assess the damage to Gage’s brain. Their reconstruction of the tamping iron’s path is shown in Figure A. The iron rod severely damaged the cerebral cortex in both hemispheres, particularly the frontal lobes. It was presumably this damage that led to Gage’s emotional outbursts and the drastic changes in his personality.

Figure A Phineas Gage and the rod that passed through his brain. (Source: Wikimedia.)

Figure B The path of the iron rod through Gage’s skull. (Source: Damasio et al., 1994, p. 1104.)
We saw in Chapter 15 that the hypothalamus integrates the actions of the ANS. In the Papez circuit, the hypothalamus governs the behavioral expression of emotion. The hypothalamus and neocortex are arranged so that each can influence the other, thus linking the expression and experience of emotion. In the circuit, the cingulate cortex affects the hypothalamus via the hippocampus and fornix (the large bundle of axons leaving the hippocampus), whereas the hypothalamus affects the cingulate cortex via the anterior thalamus. The fact that communication between the cortex and hypothalamus is bidirectional means that the Papez circuit is compatible with both the James–Lange and the Cannon–Bard theories of emotion.
While anatomical studies demonstrated that the components of the Papez circuit were interconnected as Papez had indicated, there was only suggestive evidence that each was involved in emotion. One reason Papez thought the hippocampus is involved in emotion is that it is affected by the virus responsible for rabies. An indication of rabies infection, and an aid in its diagnosis, is the presence of abnormal cytoplasmic bodies in neurons, especially in the hippocampus. Because rabies is characterized by hyperemotional responses such as exaggerated fear and aggressiveness, Papez reasoned that the hippocampus must be involved in normal emotional experience. Although little evidence related to the role of the anterior thalamus, other clinical reports at the time stated that lesions in this area led to apparent emotional disturbances, such as spontaneous laughing and crying.
You may have noticed the correlation between the elements composing the Papez circuit and Broca’s limbic lobe. Because of their similarity, the group of structures in the Papez circuit are often referred to as the limbic system, even though Broca’s anatomical notion of the limbic lobe originally had nothing to do with emotion. The term limbic system was popularized in 1952 by American physiologist Paul MacLean. According to MacLean, the evolution of a limbic system enabled animals to experience and express emotions and freed them from the stereotypical behavior dictated by their brain stem.
Difficulties with the Concept of a Single System for Emotions
We have defined a group of interconnected anatomical structures roughly encircling the brain stem as a limbic system. Experimental work supports the hypothesis that some of the structures in Broca’s limbic lobe and the Papez circuit play a role in emotion. On the other hand, some of the components of the Papez circuit are no longer thought to be important for the expression of emotion, such as the hippocampus.
The critical point seems to be conceptual, concerning the definition of an emotion system. Given the diversity of emotions, we experience and the different brain activity associated with each, there is no compelling reason to think that only one system, rather than several, is involved. Conversely, solid evidence indicates that some structures involved in emotion are also involved in other functions; there is not a one-to-one relationship between structure and function in this case. Although the term limbic system is still commonly used in discussions of brain mechanisms of emotion, it has become increasingly clear that there is not a single, discrete emotion system.
Early theories of emotion and subsequent descriptions of the limbic system were built on a combination of introspection and inference based primarily on instances of brain injury and brain disease: If damage to a brain structure alters the experience or expression of emotion, we infer that the structure is important for normal emotional function. Unfortunately, studies of disease and the consequences of lesions are not ideal for revealing normal function. Before getting into the nitty-gritty of experiments investigating neural mechanisms of emotion, it may be helpful to consider the representation of emotions from a broad perspective.
If the limbic system is not a monolithic system for the experience and expression of all emotions, as now seems the case, another possibility that has been explored is that some emotions are at least associated with distinct patterns of activity in the brain and unique physiological responses in the body (see Box 18.1). In basic theories of emotion, certain emotions are thought to be unique, indivisible experiences that are innate and universal across cultures, an idea that seems a logical extension of Darwin’s early observations on the universality of a small number of emotions. Commonly, these basic emotions are considered anger, disgust, fear, happiness, sadness, and surprise. From a neural perspective, one might hypothesize that basic emotions have distinct representations or circuits in the brain, perhaps analogous to distinct representations for sensory experiences. For example, it has been claimed that sadness correlates best with activity in the medial prefrontal cortex and fear with activity in the amygdala. In a moment, we will take an in-depth look at evidence suggesting a special role for the amygdala in fear. But first, let’s look into the general question of brain activity associated with emotion.
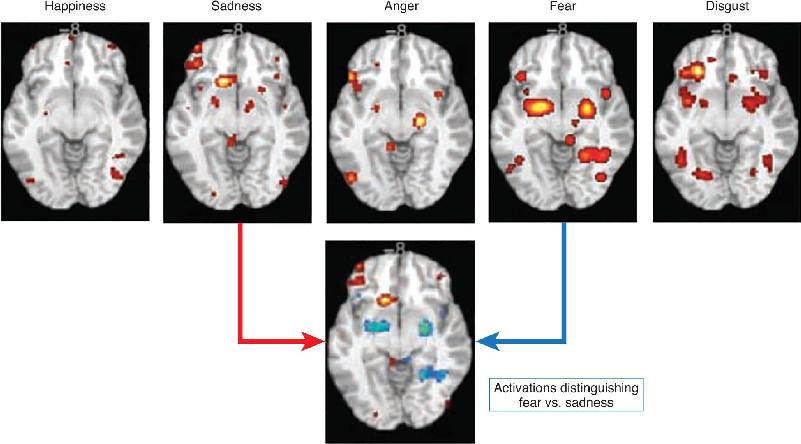
One way to get a broad perspective on the representation of emotion is to compare human functional magnetic resonance imaging (fMRI) or PET brain recordings while people experience different emotions. Numerous experiments of this sort have been conducted in which people are induced to experience emotions or shown pictures that evoke different emotions while they lie in a brain imaging machine. Figure 18.6 shows a summary of brain images collected in this way. Several observations can be made from these images. First, there are different “hotspots”—areas of particularly high brain activity associated with each emotion. Second, each emotion is associated with an array of smaller and larger patches of lesser brain activity. Finally, some activated regions are associated with more than one emotion. The bottom portion of the figure compares brain activations for sadness and fear, emotions that could be reliably discriminated from the activation patterns. Consistent with distinct circuits for different emotions, amygdala activity is more associated with fear than with sadness, and medial prefrontal activity is more associated with sadness.

FIGURE 18.6 Brain activation associated with five basic emotions. For each emotion, the strength of brain activation is indicated by color (yellow greater than red). The lower brain image compares activations associated with sadness (red and yellow = greater sadness activity) and fear (blue = greater fear activity). (Source: Hamann, 2012, p. 460.)
One interpretation of the data in Figure 18.6 is that the most highly activated region uniquely represents an emotion, such as the medial prefrontal cortex for sadness. This might be like the face-selective patches of visual cortex in the temporal lobes (see Chapter 10). Alternatively, the pattern of activation could be the basis of the emotion and each active brain region a piece of the puzzle. If either single areas or networks of areas uniquely represent emotions, we might, in principle, be able to scan people’s brains and know what they are feeling. This would be consistent with the concept of basic emotions thought to have unique and distinct representations. At this time we do not know which of these interpretations is correct. As we will now see, there are also alternative theories for the nature of the brain’s representation of emotion.
There is an intuitive appeal to the idea that each basic emotion we experience is based on brain activity in a specialized brain area or network of areas; how convenient that would be for us scientists! Unfortunately, we have learned enough about the brain to know it doesn’t always do what we find intuitive. An interesting analogy is the coding of body movement. The firing rate of a neuron in the motor cortex might code something rather straightforward such as the contraction properties of a single muscle (e.g., length, force). However, there is evidence that neural activity might represent something more complex, such as the input to a spectrum of muscles that makes up a portion of a complex behavior (e.g., swinging a golf club, dancing a pirouette).
An alternative to basic theories are dimensional theories of emotion. These theories are based on the idea that emotions, even basic emotions, can be broken down into smaller fundamental elements combined in different ways and differing amounts, just as all the elements of the periodic table are made of protons, neutrons, and electrons. Examples of proposed affective dimensions are valence (“pleasant–unpleasant”) and arousal (“weak emotion–strong emotion”). Imagine a two-dimensional graph with axes labeled in these ways; each emotional experience would be located in a different part of the graph (Figure 18.7). Of course, for any particular emotion, such as happiness, there would be some normal range along a dimension such as emotional strength (arousal). In different theories there are different numbers of dimensions, sometimes with different names. Look again at Figure 18.6, in which we first considered the patches of activity in each brain slice to be, as a group, a possible representation of one basic emotion. Might the patches instead be distinct subpatterns, one subpattern associated with the degree of pleasantness, another with emotional strength, and perhaps still others with additional dimensions? The answer to this question is, at present, unclear.

FIGURE 18.7 A dimensional representation of basic emotions. In a dimensional theory, emotions such as happiness and sadness consist of differing amounts of brain activation corresponding to affective dimensions such as valence and arousal. (Source: Hamann, 2012, p. 461.) Description
Psychological constructionist theories of emotion are a variation on dimensional theories. These theories are similar to dimensional theories in the sense that emotions are said to consist of smaller building blocks. A key difference is that in the constructionist models, the dimensions do not carry affective weight. Instead of dimensions such as pleasantness, an emotional state is constructed from physiological processes that, on their own, do not concern only emotion. Examples of nonemotional psychological components that construct emotion are things like language, attention, internal sensations from the body, and external sensations from the environment. The emotion is an emergent consequence of the combination of these components, just as a cake results from the combination of ingredients in a recipe.
From before the time of Darwin, there has been speculation about the nature of human emotions. Some researchers contend that a small set of basic emotions has evolved and these emotions are common to humans around the globe and animals as well. Other affective neuroscientists believe that emotions are constructed from building blocks that do or do not have emotional weight themselves. At present, there is a great diversity of perspectives on the nature of emotions, going beyond what we have discussed. One of the leaders in this field is Antonio Damasio at the University of Southern California, who has investigated the nature of emotions, the distinction between emotions and feelings, and the relationship between emotion and other brain functions such as decision making (Box 18.3). Aside from the nature of emotions, a related issue is the neural basis of emotions: Is each emotion represented by activity in a specialized area of the brain, a network of areas, or a more diffuse network of neurons? We do not yet have answers to these questions. Our hope for answering them relies on a convergence of approaches including behavioral observations, physiological recordings, and studies of the effects of lesions and disease, among others. In the next section, we focus on two emotions, fear and anger/aggression. We could have chosen other emotions just as well, but the research on fear and anger provides good examples that nicely bring together studies in humans and experimental animals.
It would appear that the clarity of a concept or of a scientific hypothesis is the trait that counts most in determining the acceptance and impact of an idea. But not so fast: The name given to the concept or hypothesis plays a role in how they succeed, or not. Three examples from my own work illustrate the point.
First: For the past 20 years I have been insisting on a principled distinction between the concepts of emotion and feeling.1 Emotions are programs of actions that rapidly modify the state of several components of our bodies in response, for instance, to a threat or an opportunity. Quite differently, feelings are the mental experiences of body states, including, of course, those that are caused by emotions. That the two sets of phenomena are distinct is quite clear, and yet the general public, not to mention several scientists, have persisted in lumping them together as if they were one and the same. Worse, when people do make the distinction, they often call the phenomenon by the wrong name (e.g., referring to feeling when they mean emotion or vice versa). Why so much confusion? Surely it cannot be mere carelessness. Well, it so happens that, given the long-standing historical conflation, no distinct words have evolved for the emotion or the feeling of a specific affective state. When I use the word “fear,” I could be referring to the actual emotion fear or to the feeling that results from deploying the said emotion. And even worse: One of my intellectual heroes, William James, who is responsible for first sketching a credible physiology of emotion and how it may lead to the feeling experience, is guilty of confusing the two within the very paragraph in which he so well articulated the distinction! One lesson: One should have different and unambiguous terms to designate different phenomena.
Second: Unambiguous naming is only part of what is needed for the success of new ideas. The more transparent one can be about what one means, the more likely it is that people will retain a clear message. About the same time, I began insisting on the emotion–feeling distinction, I also advanced a hypothesis regarding how affect—emotions and feelings, conscious or not—intervene, for better or for worse, in the decision-making process, and, importantly, how they need to be factored in the decision process alongside knowledge and cool reason. I called this the somatic marker hypothesis.2 Why the term “somatic”? Because emotions alter the state of the body, the soma, and feelings originate in that same body, the soma. And why the term “marker”? Because the affective state of the body, by virtue of its natural valence, marks a certain option as good, bad, or indifferent. Well, the designation caught. People do refer to it and usually get the gist of the idea from the name. It found a niche.
Third: I had no such luck when I used the terms “convergence” and “divergence” to describe, quite accurately, a connectional neural architecture with two distinct features: (a) neurons project, hierarchically, from a primary sensory cortex to smaller and smaller cortical association fields, thus converging into a narrower brain territory; and (b) other neurons reciprocate the favor in the opposite direction, thus diverging from “convergence–divergence zones” toward the originating points.3 The reality of this anatomical arrangement in the human brain is unquestionable and is quite evident in the cerebral cortex, for example. The importance of the arrangement to help explain how memory works, in terms of learning and recall, is also high. The correctness of the terms “convergence” and “divergence” is not in question either. And yet, these names did not catch and that did hurt the diffusion of my idea. About the same time, however, the terms “hub” and “spoke” began to be used to designate the same general architecture. Rather than focusing on the actual direction of the neural messages, or the functional role that each direction of the projections played, hub and spoke succinctly described the resulting flow diagram. Amusingly, after deregulation, U.S. airlines stopped flying everywhere each way and instead operated their flights to and from a few major urban hubs connected by spokes to smaller hubs, in smaller cities, by what else, spokes! Advertisements used “hub and spokes” to refer successfully to the airlines’ route system. Guess what: hub and spoke stuck for the neural architecture as well. The word “hub,” in particular, captured in three letters what I described as “convergence–divergence zones and regions.”
What’s in a name? A lot. A rose by another name is still a rose but it may not smell as well. My prize for the catchiest term to convey a scientific idea goes to mirror neurons. Ironically, mirror neurons depend on a convergence–divergence neuronal architecture and operate in a hub and spoke network!4
1. Damasio AR. 1994. Descartes’ Error. New York: Penguin Books.
2. Damasio A, Carvalho GB. 2013. The nature of feelings: evolutionary and neurobiological origins. Nature Reviews Neuroscience 14: 143–152.
3. Damasio AR. 1996. The somatic marker hypothesis and the possible functions of the prefrontal cortex. Transactions of the Royal Society (London) 351:1413–1420.
4. Damasio AR. 1989. Time-locked multiregional retroactivation: a systems level proposal for the neural substrates of recall and recognition. Cognition 33:25–62.
5. Meyer K, Damasio A. 2009. Convergence and divergence in a neural architecture for recognition and memory. Trends in Neurosciences 32(7):376–382.
As we’ve seen, there remains considerable uncertainty about the brain’s representation of emotions. Human brain imaging gives us pictures of brain activity associated with different emotions, but these pictures cannot tell us how or which brain areas actually contribute to an emotion’s experience or expression. That said, there is one brain structure that, more than any other, has a reputation as being critical for emotion: the amygdala. It has been claimed that the amygdala plays a special role in fear. While we explore the evidence connecting the amygdala to fear, keep in mind that other brain structures also appear to be involved in fear and that the amygdala is active also in other emotional states.
Shortly after Papez’s proposal of an emotion circuit in the brain, neuroscientists Heinrich Klüver and Paul Bucy, at the University of Chicago, found that bilateral removal of the temporal lobes, or temporal lobectomy, in rhesus monkeys has a dramatic effect on the animals’ aggressive tendencies and responses to fearful situations. The surgery produces numerous bizarre behavioral abnormalities collectively referred to as the Klüver–Bucy syndrome.
After temporal lobectomy, the monkeys appeared to have good visual perception but poor visual recognition. Placed in a new environment, the monkeys moved about exploring objects they saw. However, unlike normal animals they appeared to rely on placing objects into their mouth to identify them. If a hungry monkey was shown a group of objects it had seen before intermixed with food, the monkey would still go through the process of picking up each object for study before consuming the food bits. A normal hungry monkey in the same situation would make a beeline for the food. The monkeys also showed a markedly increased interest in sex.
The emotional changes in monkeys with Klüver–Bucy syndrome were most dramatically represented by decreases in fear and aggression. For example, a normal wild monkey will avoid humans and other animals. In the presence of an experimenter, it usually crouches in a corner and remains still; if approached, it will dash off to a safer corner or make an aggressive stand. These behaviors were not exhibited by the monkeys with bilateral temporal lobectomies. These otherwise wild monkeys would not only approach and touch the human but would even let the human stroke them and pick them up. They had the same placid demeanor in the presence of other animals that monkeys normally fear. Even after approaching and being attacked by a natural enemy such as a snake, the monkey would go back and try to examine it again. There was also a corresponding decrease in the vocalizations and facial expressions usually associated with fear. It appeared that both the normal experience and the normal expression of fear and aggression were severely decreased by the temporal lobectomy.
Virtually all the symptoms of the Klüver–Bucy syndrome reported in monkeys have also been seen in humans with temporal lobe lesions and, more specifically, amygdala lesions. In addition to visual recognition problems, oral tendencies, and hypersexuality, these people appear to have “flattened” emotions.
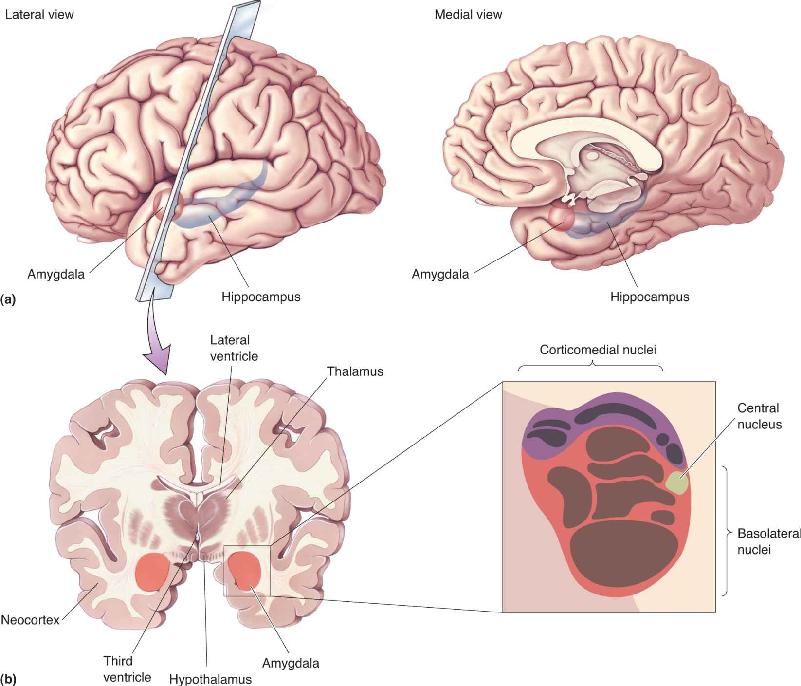
The amygdala is situated in the pole of the temporal lobe, just below the cortex on the medial side. Its name is derived from the Greek word for “almond” because of its shape.
The human amygdala is a complex of nuclei that are commonly divided into three groups: the basolateral nuclei, the corticomedial nuclei, and the central nucleus (Figure 18.8). Afferents to the amygdala come from a large variety of sources, including the neocortex in all lobes of the brain, as well as the hippocampal and cingulate gyri. Of particular interest here is the fact that information from all the sensory systems feeds into the amygdala, particularly the basolateral nuclei. Each sensory system has a different projection pattern to the amygdala nuclei, and interconnections within the amygdala allow the integration of information from different sensory modalities. Two major pathways connect the amygdala with the hypothalamus: the ventral amygdalofugal pathway and the stria terminalis.

FIGURE 18.8 A cross section of the amygdala. (a) Lateral and medial views of the temporal lobe, showing the location of the amygdala in relation to the hippocampus. (b) The brain is sectioned coronally to show the amygdala in cross section. The basolateral nuclei (surrounded by red) receive visual, auditory, gustatory, and tactile afferents. The corticomedial nuclei (surrounded by purple) receive olfactory afferents. Description
Researchers have demonstrated in several species that lesions of the amygdala have the effect of flattening emotion in a manner similar to the Klüver–Bucy syndrome. Bilateral amygdalectomy in animals can profoundly reduce fear and aggression. Reports claim that rats so treated will approach a sedated cat and nibble its ear and that a wild lynx will become as docile as a house cat.
Numerous studies in humans have examined the effect of lesions that include the amygdala on the ability to recognize emotional facial expressions. While there is a consensus that the lesions usually impair the recognition of emotional expression, researchers disagree about which emotions are affected. In different studies, deficits associated with fear, anger, sadness, and disgust have been reported. The variety of deficits probably reflects, in part, differences in the damage; two lesions are rarely alike, and they typically include damage to other brain structures in addition to the amygdala. Nonetheless, the most commonly reported symptom of lesions involving the amygdala is an inability to recognize fear in facial expressions.
Very few cases of humans with bilateral damage isolated to the amygdala have been documented. However, Ralph Adolphs, Antonio Damasio and their colleagues, then at the University of Iowa, studied a 30-year-old woman known as S.M., who had bilateral destruction of the amygdala resulting from Urbach–Wiethe disease, a rare disorder characterized by thickening of the skin, mucus membranes, and certain internal organs. S.M. was somewhat unusual in the extent to which she was indiscriminately friendly and trusting, perhaps indicating she experienced less fear than other people. She had normal intelligence and was perfectly able to identify people from photographs. When asked to categorize the emotion expressed in a person’s face, she correctly described happiness, sadness, and disgust. She was somewhat less likely to describe an angry expression as angry, however, and the most abnormal response was that she was much less likely to describe a fearful expression as afraid. Interestingly, S.M. could recognize fear from a person’s tone of voice. It appears that the amygdala lesion selectively decreased her ability to recognize fear in faces from visual input alone.
Ten years after the initial examination of S.M., a follow-up study probed her deficit in more detail by comparing her abilities to recognize happiness and fear. In the 10 intervening years, her ability to recognize fear in faces had not improved. The fascinating finding of the later study was that her inability to detect fear and some other emotions resulted from her not looking at the eyes of people in the test photographs. Evidently because she consistently looked at their mouths, she was able to recognize happiness. By comparison, control subjects routinely spent a high percentage of time looking at the eyes as they explored faces. S.M.’s exploratory eye movements were unusual in not fixating on the eyes of the people in the photographs. When she was explicitly instructed to look at a person’s eyes, she did so and then was able to correctly recognize fear. Surprisingly, after the more recent tests, she reverted to abnormal eye movements and poor fear recognition. To explain this curious set of results, the scientists hypothesized that fear is normally recognized by a two-way interaction between the amygdala and the visual cortex. Visual information is delivered to the amygdala, which then instructs the visual system to move the eyes and examine the visual input to determine the emotional expression in a face. Without the amygdala, this interaction does not take place, and S.M.’s abnormal eye movements did not allow her to recognize fear.
If removing the amygdala reduces the expression and recognition of fear, what happens if the intact amygdala is electrically stimulated? Depending on the site, amygdala stimulation can lead to different effects, including a state of increased vigilance or attention. Stimulation of the lateral portion of the amygdala in cats can elicit a combination of fear and violent aggression. Electrical stimulation of the amygdala in humans has been reported to lead to anxiety and fear. Not surprisingly, the amygdala figures prominently in current theories about anxiety disorders, as we will see in Chapter 22.
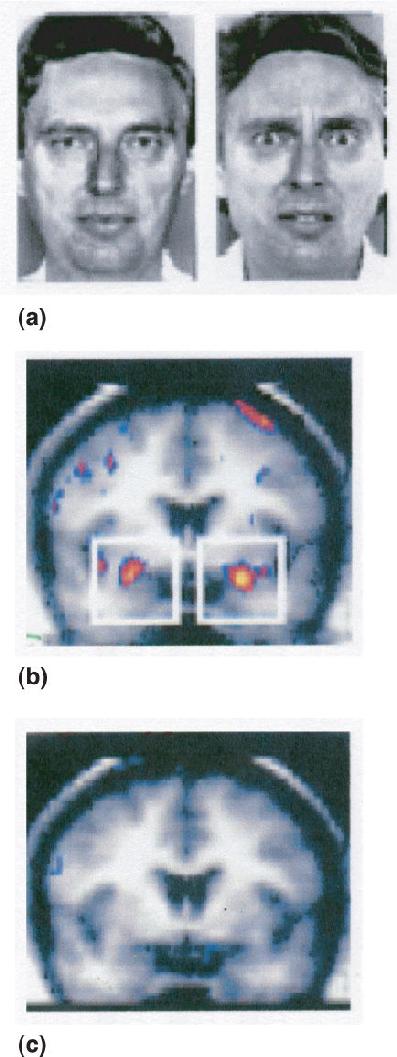
Functional brain imaging demonstrates that neural activity in the amygdala is consistent with its role in fear, as seen in Figure 18.6. For example, in an experiment performed by Breiter et al., subjects were positioned in an fMRI machine and brain activity was monitored as they viewed pictures of neutral, happy, and fearful faces (Figure 18.9a). Brain activity in response to fearful faces showed more amygdala activity than in response to faces with neutral expressions (Figure 18.9b). The amygdala activation was specific to fear, as no difference in activity occurred in response to happy and neutral facial expressions (Figure 18.9c). Other studies have reported amygdala activation in response to other facial expressions including happiness, sadness, and anger. The function the amygdala plays in these various emotions is not yet resolved, but all the evidence together suggests that the amygdala plays a key role in detecting fearful and threatening stimuli.

FIGURE 18.9 Human brain activity in response to emotional stimuli. (a) Neutral and fearful faces were used as visual stimuli. (b) Fearful faces produced greater activity in the amygdala (red and yellow areas within white squares) than neutral faces. (c) No difference in amygdala activity occurred in response to happy and neutral faces. (Source: Breiter et al., 1996.) Description
Experiments in animals and humans, as well as introspection, indicate that memories for emotional events are particularly vivid and long-lasting. This is undoubtedly true for learned fear. Through socialization or painful experience, we all learn to avoid certain behaviors for fear of being hurt. If you ever received a painful shock as a child by pushing a paper clip into an electrical outlet, you probably never did it again. Memories associated with fear can form quickly and be long-lasting. As we will see in Chapter 22, in post-traumatic stress disorder, intense fear resulting from a traumatic experience can interfere with normal life for many years. Although the amygdala is not thought to be a primary location for memory storage, synaptic changes in the amygdala appear to be involved in forming memories for emotional events.
A number of different experiments suggest that neurons in the amygdala can “learn” to respond to stimuli associated with pain, and after such learning, these stimuli evoke a fearful response. In an experiment performed by Bruce Kapp and his colleagues at the University of Vermont, rabbits were conditioned to associate the sound of a tone with mild pain. A normal sign of fear in rabbits is a change in heart rate. In the experiment an animal was placed in a cage, and at various times it would hear one of two tones. One tone was followed by a mild electrical shock to the feet through the metal floor of the cage; the other tone was benign. After conditioning, Kapp’s group found that the rabbit’s heart rate developed a fearful response to the tone associated with pain but not to the benign tone. Before conditioning, neurons in the central nucleus of the amygdala had not responded to the tones used in the experiment. After conditioning, however, neurons in the central nucleus of the amygdala responded to the shock-related tone (but not the benign one). Joseph LeDoux of New York University has shown that after this type of fear conditioning, amygdala lesions eliminate the learned visceral responses, such as the changes in heart rate and blood pressure. It appears that the conditioned response in the amygdala arises from synaptic changes in the basolateral nuclei.
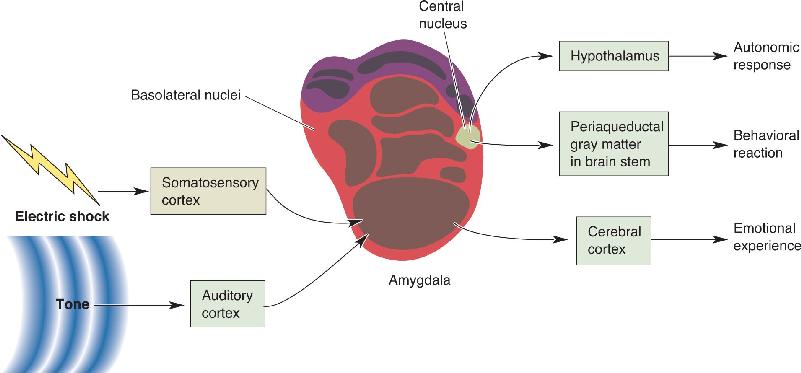
Figure 18.10 shows a proposed circuit to account for learned fear. Sensory information, for example, the tone the animal heard and the electrical shock it felt, is sent to the basolateral region of the amygdala, where cells in turn send axons to the central nucleus. The pairing of a benign tone with a painful stimulus leads to changes in synaptic strength that enhance the amygdala’s response to the tone after conditioning (Chapters 24 and 25 discuss the neural changes that occur with conditioning). Efferents from the central nucleus project to the hypothalamus, which can alter the state of the ANS, and to the periaqueductal gray matter in the brain stem, which can evoke behavioral reactions via the somatic motor system. The emotional experience is thought to be based on activity in the cerebral cortex.

FIGURE 18.10 A neural circuit for learned fear. Through conditioning, a sound tone becomes associated with the pain of an electrical shock. The fear response is mediated by the amygdala. The benign tone and the painful shock reach the basolateral nuclei of the amygdala by way of auditory and somatosensory cortex, and the signal is relayed to the central nucleus. The pairing of these stimuli leads to synaptic change in the amygdala and an enhanced response to the benign tone. Efferents from the amygdala project to the brain stem periaqueductal gray matter, causing the behavioral reaction to the conditioned tone, and to the hypothalamus, resulting in the autonomic response. The experience of an unpleasant emotion presumably involves projections to the cerebral cortex. Description
Recent research suggests that the role of the amygdala in learned fear, first studied in rabbits and rats, extends to humans. In one study, subjects were shown a number of visual stimuli and conditioned to expect a mild electrical shock when a particular visual stimulus was presented. An fMRI machine monitored brain activity. The fMRI images show that the “feared” visual stimulus activated the amygdala significantly more than visual stimuli not associated with a shock.
In another study using PET imaging of brain activity conducted by Hamann et al., subjects first viewed a series of pictures. Some of the pictures were pleasant (appealing animals, sexually arousing scenes, appetizing food); some were frightening or aversive (frightening animals, mutilated bodies, violence); and some were neutral (household scenes, plants). Compared to the neutral objects, both the pleasant and unpleasant stimuli affected physiological measures such as heart rate and skin conductance, and they evoked greater activity in the amygdala. These measurements confirm the role of the amygdala in emotional processing, as we have already discussed. In the second phase of the experiment, subjects were put back in the PET imaging machine and shown a variety of pictures. They were asked to use their memory and identify which pictures they had seen in the initial conditioning session. As expected, subjects recalled the emotional pictures better than the neutral ones. The enhanced memories for emotional pictures correlated with recorded amygdala activity (Figure 18.11). There was no such correlation for the neutral pictures.

FIGURE 18.11 Amygdala activity associated with enhanced emotional memory. Subjects first viewed pictures of emotional and neutral stimuli while PET imaging recorded brain activity. Later the original and new pictures were viewed. The recall of emotional stimuli was associated with an enhanced response in the amygdala, shown in yellow. (Source: Hamann et al., 1999.)
Anger is a basic emotion. Many things can make us angry: frustration, hurt feelings, stress, and so on. Aggression is not an emotion but is one possible behavioral result of anger; an angry drunk might punch someone in the nose. In studies of humans, aggression and the feeling we call anger can be readily distinguished, since people can state that they are angry even if they do not act on that feeling. As we have already seen, emotions are more difficult to study in animals because we cannot ask an animal how it feels but can only measure its physiological or behavioral manifestations. We can infer that an animal is angry only by the aggressive behaviors it exhibits, such as making a loud scary sound, a menacing facial expression, or a threatening posture. Because aggression and anger are often intertwined in animals, we will discuss them together here.
We can distinguish different forms of aggression in humans, ranging from self-defense to murder. Likewise, there are different types of aggression in animals. One animal may act aggressively toward another for many reasons: to kill for food, to defend offspring, to win a mate, or to scare off a potential adversary. There is some evidence that different types of aggression are regulated differently by the nervous system.
Aggression is a multifaceted behavior that is not a product of a single isolated system in the brain. One factor that influences aggression is the level of male sex hormones, or androgens (see Chapter 17). In animals, seasonal androgen levels and aggressive behavior correlate. Consistent with one of the roles of androgens, injections of testosterone can make an immature animal more aggressive, and castration can reduce aggressiveness. In humans, the relationship is less clear, although some have claimed that aggressive behavior in violent criminals is connected to testosterone levels. You may have heard of “roid rage,” an uncontrolled outburst of anger and aggression sometimes reported in athletes taking anabolic steroids, which have similar effects on the body as testosterone. In any case, there is strong evidence for a neurobiological component to aggression, which is our focus here.
A useful distinction can be made between predatory aggression and affective aggression. Predatory aggression involves attacks against a member of a different species for the purpose of obtaining food, such as a lion hunting a zebra. Attacks of this type are typically accompanied by relatively few vocalizations, and they are aimed at the head and neck of the prey. Predatory aggression is not associated with high levels of activity in the sympathetic division of the ANS. Affective aggression is for show rather than to kill for food, and it involves high levels of sympathetic activity. An animal exhibiting affective aggression typically makes vocalizations while adopting a threatening or defensive posture. A cat hissing and arching its back at the approach of a dog is a good example. The behavioral and physiological manifestations of both types of aggression must be mediated by the somatic motor system and the ANS, but the pathways must diverge at some point to account for the dramatic differences in the behavioral responses.
Several lines of evidence indicate that the amygdala is involved in aggressive behavior. American scientist Karl Pribram and his colleagues in 1954 showed that amygdala lesions had a major effect on social interactions in a colony of eight male rhesus monkeys. Having lived together for some time, the animals had established a social hierarchy. The investigators’ first intervention was to make bilateral amygdala lesions in the brain of the most dominant monkey. After this animal returned to the colony, it fell to the bottom of the hierarchy, and the monkey previously second in dominance now became dominant. Presumably, the second monkey in the hierarchy discovered that the “top banana” had become more placid and less difficult to challenge. After an amygdalectomy was performed on the new dominant monkey, it likewise fell to the bottom of the hierarchy. This pattern suggested that the amygdala is important for the aggression normally involved in maintaining a position in the social hierarchy. This is consistent with the finding that electrical stimulation of the amygdala can produce a state of agitation or affective aggression.
Surgery to Reduce Human Aggression. In the 1960s, amygdala surgery in violent humans was first performed in the hope that lesions would reduce aggression as they do in animals. It was thought by some that violent behavior frequently resulted from seizures in the temporal lobe. In a human amygdalectomy, electrodes are passed through the brain and down into the temporal lobe. By making neural recordings along the way and imaging the electrodes with X-rays, it is possible to position the tip of the electrode in the amygdala. Electrical current is then passed through the electrode, or a solution is injected, to destroy all or part of the amygdala. The lesions that are produced have a “taming effect” in some patients, reducing the incidence of aggressive outbursts. Brain surgery used as a method of treating psychiatric disorders is called psychosurgery. Early in the twentieth century, treating severe disorders involving anxiety, aggression, or neuroses with psychosurgical techniques, including the frontal lobotomy, was a common practice (Box 18.4). By today’s standards, psychosurgery is a drastic procedure to be considered only as a treatment of last resort. Although amygdalectomies are still occasionally performed to treat aggressive behavior, medication is the usual treatment.
Ever since the discoveries by Klüver, Bucy, and others that brain lesions can alter emotional behavior, clinicians have attempted surgery as a means of treating severe behavioral disorders in humans. Today, it is difficult for many people to imagine that destroying a large portion of the brain was once thought to be therapeutic. Indeed, in 1949 the Nobel Prize in Medicine was awarded to Dr. Egas Moniz for his development of the frontal lobotomy technique. Even stranger is the fact that Moniz was shot in the spine and partially paralyzed by a lobotomized patient. Although lobotomies are no longer being performed, tens of thousands were performed following World War II.
Little theory supported the development of the lobotomy. In the 1930s, John Fulton and Carlyle Jacobsen of Yale University reported that frontal lobe lesions had a calming effect in chimpanzees. It has been suggested that frontal lesions have this effect because of the destruction of limbic structures and, in particular, connections with frontal and cingulate cortex. Moniz proposed that ablations of the frontal cortex might be effective in treating psychiatric diseases.
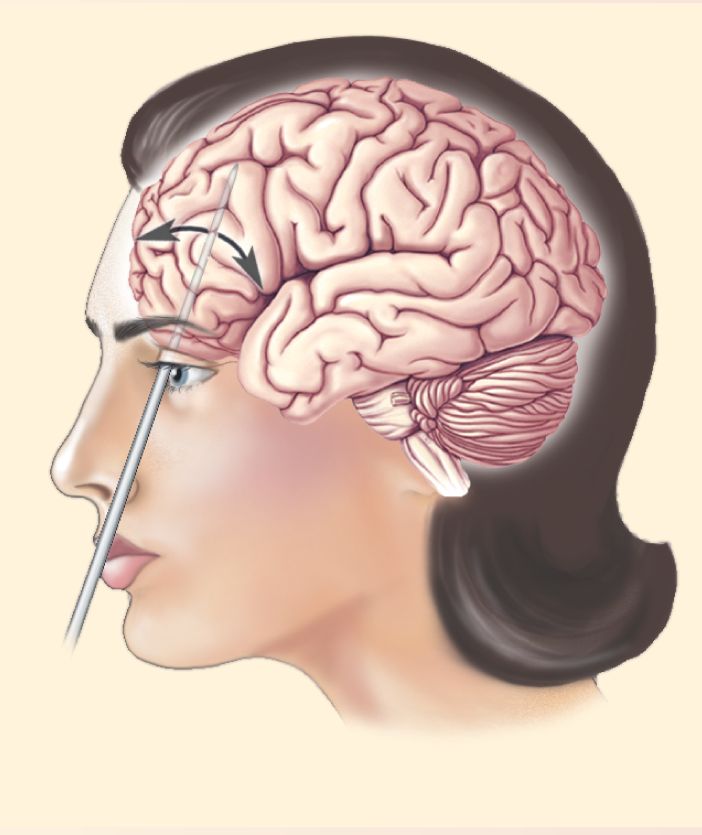
A frightening variety of techniques were used to produce lesions in the frontal lobes. The procedure became more commonplace with the development of a technique known as transorbital lobotomy (Figure A). In this procedure, a leucotome, a 12-cm steel rod that tapers to a point, was driven through the thin bone at the top of the eye’s orbit with a hammer. The handle was then swung medially and laterally to destroy cells and interconnecting pathways. Thousands of people were lobotomized with this technique, sometimes called “ice pick psychosurgery”; it was so simple it could be performed in the physician’s office. Note that although this surgery left no outward scars, the physician could not see what was being destroyed.

Frontal lobotomy reportedly had beneficial effects on people with a number of disorders, including psychosis, depression, and various neuroses. The effect of the surgery was described as a relief from anxiety and escape from thoughts that were unendurable. Only later did a pattern of less pleasant side effects emerge. While frontal lobotomy can be performed with little decrease in IQ or loss of memory, it did have other profound effects. The changes that appear to be related to the limbic system are a blunting of emotional responses and a loss of the emotional component of thoughts. In addition, lobotomized patients often developed “inappropriate behavior” or an apparent lowering of moral standards. Like Phineas Gage, patients had considerable difficulty planning and working toward goals. Lobotomized patients also had trouble concentrating and were easily distracted.
With our modest understanding of the neural circuitry underlying emotion and other brain functions, it is hard to justify destroying a large portion of the brain. Fortunately, treatment with lobotomy decreased fairly rapidly, and today, instead, drug therapy is primarily used for serious emotional disorders.
Neural Components of Anger and Aggression Beyond the Amygdala
In addition to the amygdala, a variety of brain structures have been reported to be involved in anger and aggression. For example, human brain imaging studies have found that there is greater activity in orbitofrontal cortex and anterior cingulate cortex when subjects recall past experiences that made them angry. Interpreting these patterns of brain activation involves the same challenges we have discussed for other emotions. Historically, studies of anger and aggression have been important for their implications for the involvement of subcortical structures in emotion. We now look at a few of these important research milestones.
Anger, Aggression, and the Hypothalamus. One of the earliest structures linked to anger and aggressive behavior is the hypothalamus. Experiments performed in the 1920s showed that a remarkable behavioral transformation took place in cats or dogs whose cerebral hemispheres had been removed. Animals that were not easy to provoke prior to the surgery would go into a state of violent rage with the least provocation after the surgery. For instance, a violent response might be produced by an act as mild as scratching a dog’s back. This state was called sham rage because the animal demonstrated all the behavioral manifestations of rage but in a situation that normally would not cause anger. It was also a sham in the sense that the animals would not actually attack as they normally might.
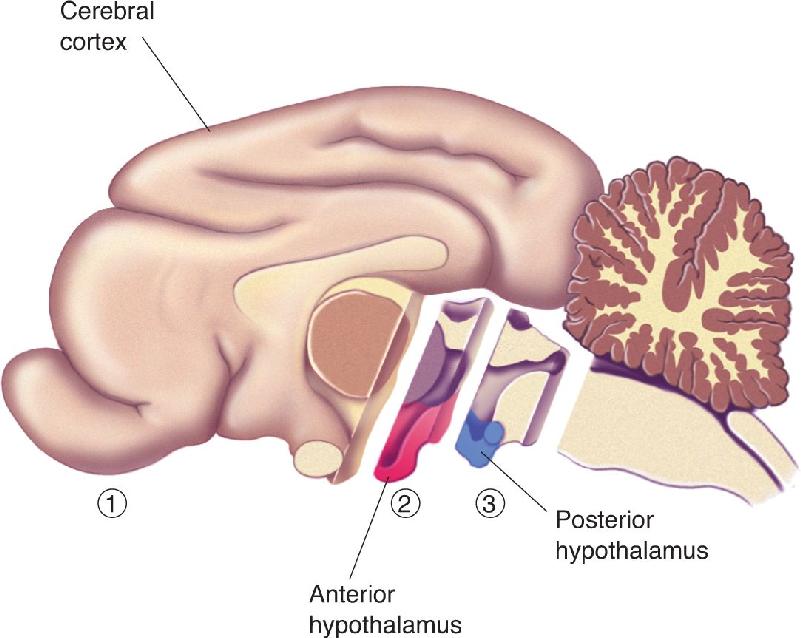
While the extreme behavioral condition called sham rage resulted from removing all of both cerebral hemispheres (the telencephalon), remarkably, the behavioral effect can be reversed by making the lesion just a little bit larger to include portions of the diencephalon, particularly the hypothalamus. Sham rage is observed if the anterior hypothalamus is destroyed along with the cortex but is not seen if the lesion is extended to include the posterior half of the hypothalamus (Figure 18.12). The implication is that the posterior hypothalamus may be particularly important for the expression of anger and aggression and that normally it is inhibited by the telencephalon. But we must bear in mind that these lesions were large, and something other than the posterior hypothalamus may have been destroyed with the larger lesion.

FIGURE 18.12 Brain transections and sham rage. ➀ If the cerebral hemispheres are removed and the hypothalamus is left intact, sham rage results. ➀ and ➁ A similar result is obtained if the anterior hypothalamus is removed in addition to the cerebral cortex. ➀, ➁, and ➂ If the posterior hypothalamus is removed in addition to the anterior hypothalamus, sham rage does not result. Description
In a series of pioneering studies begun in the 1920s, W.R. Hess at the University of Zurich investigated the behavioral effects of electrically stimulating the diencephalon. Hess made small holes in the skulls of anesthetized cats and implanted electrodes in the brain. After the animal awoke, a small electrical current was passed through the electrodes, and behavioral effects were noted. Various structures were stimulated, but here we will focus on the effects of stimulating different regions of the hypothalamus.
The variety of complex responses to stimulating slightly different portions of the hypothalamus is amazing, considering that it is such a small part of the brain. Depending on where the electrode is placed, stimulation may cause the animal to sniff, pant, eat, or express behaviors characteristic of fear or anger. These reactions illustrate the two primary functions of the hypothalamus discussed in Chapters 15 and 16: homeostasis and the organization of coordinated visceral and somatic motor responses. Responses related to emotional expression can include changes in heart rate, pupillary dilation, and gastrointestinal motility, to name a few. Because stimulation of some parts of the hypothalamus also elicits behavior characteristic of fear and anger, we hypothesize that the hypothalamus is an important component of the system normally involved in expressing these emotions.
The expression of rage Hess evoked by hypothalamic stimulation was similar to the sham rage seen in animals whose cerebral hemispheres had been removed. With a small application of electrical current, a cat would spit, growl, and fold its ears back, and its hair would stand on end. This complex and highly coordinated set of behaviors would normally occur when the cat feels threatened by an enemy. Sometimes the cat would suddenly run as if fleeing an imaginary attacker. If the intensity of the stimulation was increased, the animal might make an actual attack, swatting with a paw or leaping onto an imaginary adversary. When the stimulation was stopped, the rage disappeared as quickly as it started, and the cat might even curl up and go to sleep.
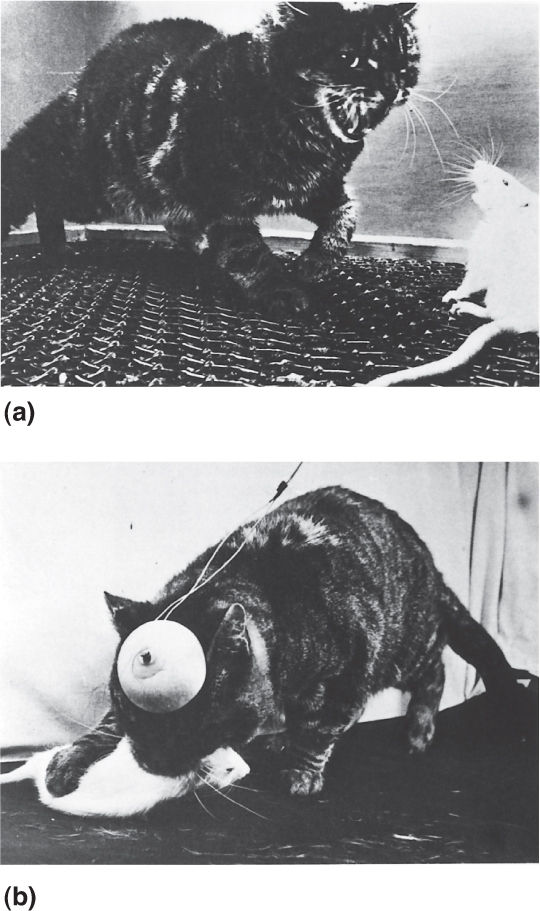
In experiments conducted at the Yale University Medical School in the 1960s, John Flynn found that affective aggression and predatory aggression could be elicited by stimulating different areas of a cat’s hypothalamus (Figure 18.13). Affective aggression, also known as a threat attack, was observed after stimulating specific sites in the medial hypothalamus. Similar to the rage response reported by Hess, the cat would arch its back, hiss, and spit but would usually not actually attack a victim, such as a nearby rat. Predatory aggression, which Flynn called a silent-biting attack, was evoked by stimulating parts of the lateral hypothalamus. While the back might be somewhat arched and the hair slightly on end, predatory aggression was not accompanied by the dramatic threatening gestures of affective aggression. Nonetheless, in this “quiet attack,” the cat would move swiftly toward a rat and viciously bite its neck. Despite the crudeness of such experimentation by today’s standards, the early research involving lesions and electrical stimulation of the hypothalamus are consistent in suggesting that this structure is important for the expression of anger and aggression in animals.

FIGURE 18.13 Rage reactions in cats with hypothalamic stimulation. (a) Stimulation of the medial hypothalamus produces affective aggression (threat attack). (b) Stimulation of the lateral hypothalamus evokes predatory aggression (silent-biting attack). (Source: Flynn, 1967, p. 45.) Description
The Midbrain and Aggression. There are two major pathways by which the hypothalamus sends signals involving autonomic function to the brain stem: the medial forebrain bundle and the dorsal longitudinal fasciculus. Axons from the lateral hypothalamus make up part of the medial forebrain bundle, and these project to the ventral tegmental area in the midbrain. Stimulation of sites within the ventral tegmental area can elicit behaviors characteristic of predatory aggression, just as stimulation of the lateral hypothalamus does. Conversely, lesions in the ventral tegmental area can disrupt offensive aggressive behaviors. One finding suggesting that the hypothalamus influences aggressive behavior via its effect on the ventral tegmental area is that hypothalamic stimulation will not evoke aggression if the medial forebrain bundle is cut. Interestingly, aggressive behavior is not entirely eliminated by this surgery, suggesting that this route is important when the hypothalamus is involved, but that the hypothalamus need not always be involved.
The medial hypothalamus sends axons to the periaqueductal gray matter (PAG) of the midbrain by way of the dorsal longitudinal fasciculus. Electrical stimulation of the PAG can produce affective aggression, and lesions there can disrupt this behavior. Interestingly, the hypothalamus and the midbrain periaqueductal gray matter appear to influence behavior partially based on input from the amygdala. Figure 18.14 shows a simplified circuit for anger and aggression involving the structures we have discussed.

FIGURE 18.14 A neural circuit for anger and aggression. (a) The hypothalamus can influence aggressive behavior through projections to the ventral tegmental area and the periaqueductal gray matter. (b) In this simplified scheme, the expression of anger and aggression is controlled by a neural pathway from the amygdala through the hypothalamus, periaqueductal gray matter (PAG), and ventral tegmental area. Description
A variety of studies suggest that the neurotransmitter serotonin plays an important role regulating anger and aggression. Serotonin-containing neurons are located in the raphe nuclei of the brain stem, and they ascend in the medial forebrain bundle and project to the hypothalamus and various limbic structures involved in emotion (see Figure 15.13). For the most part, experimental evidence supports the serotonin deficiency hypothesis, which states that aggression is inversely related to serotonergic activity.
One link between serotonin and aggression comes from studies of induced aggression in rodents. If male mice are isolated in a small cage for several weeks, about half of them will become hyperactive and unusually aggressive when they subsequently encounter other mice. Although the isolation has no effect on the level of serotonin in the brain, there is a decrease in the turnover rate (the rate of synthesis, release, and resynthesis) of this neurotransmitter. Moreover, this decrease is found only in the mice that later become unusually aggressive and not in those relatively unaffected by the isolation. Also, female mice typically do not become aggressive following isolation, and they show no decrease in serotonin turnover. Evidence indicates that drugs that block the synthesis or release of serotonin increase aggressive behavior. For instance, in one study, when the drug parachlorophenylalanine (PCPA), which blocks serotonin synthesis, was administered, the injected animals increased their attacks on others in their cage.
There are at least 14 serotonin receptor subtypes, and it appears that the 5-HT1A and 5-HT1B subtypes are involved in modulating anger and aggression. For example, a number of experiments have shown in mice that agonists of 5-HT1B receptors decrease aggressiveness while antagonists of these receptors increase aggressiveness. Based on these pharmacological results, one would predict that mice lacking the 5-HT1B receptor would be more aggressive than normal animals. Consistent with this prediction, in some studies, 5-HT1B receptor knockout mice are reported to show more aggressive behavior. Other experiments paint a somewhat different picture, however, suggesting that rather than simply being more aggressive, the knockout mice are more impulsive.
The relationship between serotonin and aggression is similar in primates that have been studied. For example, researchers found that the dominance hierarchy in a colony of vervet monkeys could be manipulated by injecting animals with drugs that either increased or decreased serotonergic activity. The behavior of these animals was consistent: More aggression was associated with less serotonergic activity. However, there was one interesting sociological twist; aggression did not correlate with dominance in the group. If the dominant male was removed, the top position was taken by an animal with artificially enhanced serotonergic activity (i.e., one injected with a serotonin precursor or reuptake inhibitor who was less aggressive). Conversely, the injection of drugs that reduced serotonin function (serotonin antagonists) was correlated with animals becoming subordinate. The subordinate animals were actually significantly more likely to initiate aggression. Interestingly, the less aggressive dominant male garnered his status by his skills in recruiting females to support his position.
In humans, there are a large number of reports of a negative correlation between serotonin activity and aggression. For example, in a study of men in the military who had been diagnosed with personality disorders, aggression was found to be inversely related to cerebrospinal fluid levels of the serotonin metabolite 5-hydroxyindoleacetic acid (5-HIAA). Questions have been raised, however, about the generality of the correlation between serotonin and aggression when people of different ages and people without personality disorders are examined. As with the animal studies, a correlation is often reported, but the reality is probably more complex.
Many scientists would agree that serotonin is involved in the modulation of anger and aggressive behavior. The evidence presented here suggests a straightforward negative correlation between aggression and activity in the brain’s serotonergic system. However, some scientists in the field consider this relationship overly simplistic. Animals exhibit aggressive behaviors for a variety of reasons, and serotonin is not involved equally in all forms. From a mechanistic standpoint, the system is complex. Serotonergic neurons project broadly across the brain. 5-HT1A and 5-HT1B receptors are distributed widely, and they and other serotonin receptors provide interactions with other neurotransmitter systems. As well, there is negative feedback in the system because many of the 5-HT1A and 5-HT1B receptors are autoreceptors (see Chapter 5). Some autoreceptors are presynaptic on the raphe neurons that send serotonin widely to the brain. Activation of them globally inhibits serotonin release. With this negative feedback, serotonin release affects the raphe neurons in such a way as to decrease further release. Because of the diversity of receptor locations and functions, interpretation of pharmacological and knockout experiments is challenging; new approaches are needed to tease out the details of the relationships among serotonin, anger, and aggressive behavior.
We all know what emotions are—those feelings we have that we call happiness, sadness, and so on. But what exactly are those feelings? As evidenced by the diverse theories we have outlined, there is a great deal of uncertainty. More than a hundred years after the James–Lange theory was proposed, there remains controversy about the extent to which emotions cause changes in the body or bodily changes cause emotions. We do know from brain imaging studies that emotions are associated with widespread brain activation. Some of the structures involved are part of the limbic system, and other structures are not. But even with images of brain activity in various emotional states, understanding the neural basis of emotional experience is challenging. We don’t know which of the active areas are responsible for the feelings. Is it the most active area, all of the areas, or something else? What should we make of the observation that some brain structures are activated in multiple emotional states while others are more specific to particular emotions? For that matter, is it even correct to think of brain activity as reflecting feelings, or might feelings be emergent sensations based on combinations of active neurons, none of which independently signals an emotion?
In this chapter, we have focused on a handful of brain structures for which there is particularly strong evidence for involvement in emotion. A way to look at our current state of understanding is that the combined lesion, stimulation, and brain imaging studies have done a good job identifying structures that are candidates for emotional processing. It will take a good deal more work to figure out what various cortical and subcortical areas contribute.
Emotional experiences are the result of complex interactions among sensory stimuli, brain circuitry, past experiences, and the activity of neurotransmitter systems. In light of this complexity, we probably should not be surprised that humans exhibit a broad spectrum of emotional and mood disorders, as we will see in Chapter 22.
When thinking about the neural basis of emotion, keep in mind that the structures apparently involved in emotion also have other functions. For a considerable time after Broca defined the limbic lobe, it was thought to be primarily an olfactory system. And even though our perspective has changed much since Broca’s time, parts of the brain involved in olfaction have been included in the definition of the limbic system. We will see in Chapter 24 that some of the limbic structures are also important for learning and memory. Emotions are nebulous experiences that influence our brains and behavior in many ways, so it seems logical that emotional processing should be intertwined with other brain functions.
psychological constructionist theories of emotion (p. 628)
1. According to the James–Lange and Cannon–Bard theories of emotion, what is the relationship between the anxiety you would feel after oversleeping through an exam and your physical responses to the situation? Would you experience anxiety before or after the increase in your heart rate?
2. How have the definition of the limbic system and thoughts about its function changed since the time of Broca?
3. What procedures produce an abnormal rage reaction in an experimental animal? Can we know that the animal feels angry?
4. What changes in emotion were observed following temporal lobectomy in monkeys by Klüver and Bucy? Of the numerous anatomical structures they removed, which is thought to be closely related to changes in temperament?
5. Why might performing bilateral amygdalectomy on a dominant monkey in a colony result in that monkey becoming a subordinate?
6. What assumptions about limbic structures underlie the surgical treatment of emotional disorders?
7. The drug fluoxetine (Prozac) is a serotonin-selective reuptake inhibitor. How might this drug affect a person’s level of anxiety and aggression?
8. What distinguishes basic emotion, dimensional, and psychological constructionist theories of emotion?
9. How do patterns of brain activation differ for sadness and fear?
Barrett LF, Satpute AB. 2013. Large-scale networks in affective and social neuroscience: towards an integrative functional architecture of the brain. Current Opinion in Neurobiology 23:361–372.
Dagleish T. 2004. The emotional brain. Nature Reviews 5:582–589.
Dolan RJ. 2002. Emotion, cognition, and behavior. Science 298:1191–1194.
Duke AA, Bell R, Begue L, Eisenlohr-Moul T. 2013. Revisiting the serotonin-aggression relation in humans: a meta-analysis. Psychological Bulletin 139:1148–1172.
Gendron M, Barrett LF. 2009. Reconstructing the past: a century of ideas about emotion in psychology. Emotion Review 1:316–339.
Gross CT, Canteras NS. 2012. The many paths to fear. Nature Reviews Neuroscience 13:651–658.
Hamann S. 2012. Mapping discrete and dimensional emotions onto the brain: controversies and consensus. Trends in Cognitive Sciences 16:458–466.
LeDoux J. 2012. Rethinking the emotional brain. Neuron 73:653–676.
Lindquist KA, Wager TD, Kober H, Bliss-Moreau E, Barrett LF. 2012. The brain basis of emotion: a meta-analytic review. Behavioral and Brain Sciences 35:121–143.
McGaugh JL. 2004 The amygdala modulates the consolidation of memories of emotionally arousing experiences. Annual Review of Neuroscience 27:1–28.
Additional figures