Language
BOX 20.1 OF SPECIAL INTEREST: Thinking in Different Languages
Genetic Factors in Specific Language Impairment and Dyslexia
THE DISCOVERY OF SPECIALIZED LANGUAGE AREAS IN THE BRAIN
BOX 20.2 OF SPECIAL INTEREST: Assessing Hemispheric Language Dominance
BOX 20.3 PATH OF DISCOVERY: Uncovering Language Areas of the Brain, by Nina Dronkers

The Wernicke–Geschwind Model of Language and Aphasia
ASYMMETRICAL LANGUAGE PROCESSING IN THE TWO CEREBRAL HEMISPHERES
LANGUAGE STUDIES USING BRAIN STIMULATION AND HUMAN BRAIN IMAGING
BOX 20.4 OF SPECIAL INTEREST: Hearing Sight and Seeing Touch
Language is a remarkable system for communication that has an enormous impact on our lives. You can walk into a café and order a large skinny mocha cappuccino with a dash of vanilla and be reasonably certain you will not be handed a bucket of mud. You can speak on the telephone with someone thousands of miles away and explain to them both the complexities of quantum physics and the emotional toll the physics course is having on your social life. There are endless debates about whether animals also have language, but there is no question that the complex and flexible system of language we use is unique to humans. Without language, we could not learn most of what we study in school, and this would greatly limit what we could accomplish.
More than just sounds, language is a system by which sounds, symbols, and gestures are used for communication. Language comes into our brains through the visual and auditory systems, and we produce speech and writing with our motor system. But the brain processing between the sensory and motor systems is the essence of language. Because animals are of limited use in studying human language, for many years, language was studied primarily by linguists and psychologists rather than neuroscientists. Much of what we know about the brain mechanisms responsible for language is derived from studies of language deficits resulting from brain damage. Numerous different aspects of language can be selectively disrupted, including speech, comprehension, and naming, suggesting that language is processed in multiple, anatomically distinct, stages. More recently, the imaging of activity in the human brain with fMRI and PET has given us intriguing insights into the complex circuits underlying language.
Language is universal in human society, perhaps because of specialized brain organization. It is estimated that there are over 5000 languages and dialects throughout the world, and languages differ in many ways, such as the order in which nouns and verbs are arranged. But despite differences in syntax, from Patagonia to Katmandu, all languages convey the subtleties of human experience and emotion. Consider the fact that no mute tribe of people has ever been found, not even in the remotest corner of the world. Many scientists believe the universality of language is a consequence of the fact that the human brain has evolved special language-processing systems.
Language is a system for representing and communicating information that uses words combined according to grammatical rules. Language can be expressed in a variety of ways including gestures, writing, and speech. Speech is an audible form of communication built on the sounds humans produce. Speech comes naturally to humans: Even with no formal training, children raised in a normal language environment will invariably learn to understand spoken language and to speak. Reading and writing, on the other hand, require years of formal training, and more than 10% of the world’s population is illiterate.
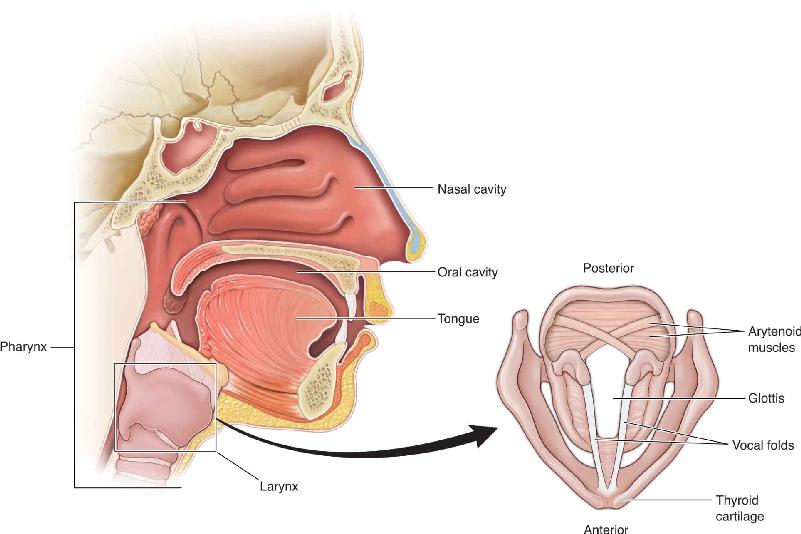
Across the animal kingdom, a variety of systems are used to produce sounds, but we’ll focus on the basics of sound production in humans (Figure 20.1). Human speech involves a remarkable coordination of over 100 muscles ranging from those controlling the lungs to those of the larynx and the mouth. Ultimately all of these muscles are controlled by motor cortex, something we will have a good deal more to say about. Human sound begins when a person exhales air out of the lungs. The air passes through the larynx, also known as the voice box. What we call the Adam’s apple in the neck is the larynx’s anterior wall of cartilage. Within the larynx are the vocal folds, two bands of muscle also called vocal chords, which form a V. The space between the vocal folds is the glottis. Sounds are produced by vibrations in tightened vocal folds, somewhat as you can make a buzzing sound by blowing on a tightly held blade of grass. If the vocal fold muscles are relaxed, no sound will be produced, just as blowing on a loose blade of grass will make no sound; this is what happens when we exhale without speaking. The pitch of the sound results from the frequency of the vocal fold vibrations: Greater tension in the vocal folds produces higher-frequency vibrations and higher pitch sound. The sound is modified at further stages of the vocal tract, including the pharynx (especially the throat between larynx and mouth), mouth, and nose. Finally, rapid changes in the positions of the tongue, lips, and soft palate modulate sound for speech. The fundamental sounds that a language uses to communicate are called phonemes. Different spoken languages have different phonemes that build words unique to the language. Interestingly, research suggests that the words used in a language may have subtle effects on how people using that language think (Box 20.1).

FIGURE 20.1 The human vocal tract. Air exhaled from the lungs sets the vocal folds, located in the larynx, into vibration. The sound produced by the vibration is modified by later structures including the pharynx, mouth, and nose. Description
There are hundreds of different human cultures around the globe, each with its own customs, beliefs, and ways of life. We are used to the idea that people’s attitudes about what is beautiful, delicious, and appropriate vary a good deal from place to place. Thinking may be affected by culture, but it shouldn’t be influenced by language, which is simply the way people express themselves, right? Is it possible that the language a person uses alters how the person thinks? In the 1950s, Benjamin Lee Whorf proposed that the language people speak constrains their thoughts, perceptions, and actions, an extreme stance that has been largely abandoned. However, language does appear to impact thought in a few subtle and intriguing situations. Consider the use of gender in language. Most European languages, unlike English, assign a gender to inanimate objects. English speakers may occasionally assign gender to an object—“I like that car over there, she’s a beauty!”—but this is rare. Contrast that with Italian, in which objects are masculine or feminine: teeth, flowers, and the sea—all masculine, who knows why. Or consider French, in which the same three things are feminine. The German dinner table is a funny arrangement with a masculine spoon, a feminine fork, and a neutral knife. It’s enough to make a college language student’s head spin.
Research suggests that the seemingly random gender assignments used in many languages may influence the way people think about the objects. In one study, French and Spanish speakers were asked to help make a movie in which objects would come to life. The subjects were shown pictures of the objects and asked to assign a man’s or woman’s voice. The objects were a mixture of ones that have the same gender in French and Spanish (e.g., “ballerina”) and others that have the opposite gender (e.g., “broom”). The idea of using pictures was to avoid the bias that might come from saying the masculine or feminine words of each language. The experimental results showed that both French and Spanish speakers were significantly more likely to say an object should use a female voice when the object has a female gender in their language. This was true when the languages agreed about gender, but when these languages had different genders for the same object, the French and Spanish speakers disagreed about the appropriate gender for the voice.
In another experiment, Spanish and German speakers were asked to memorize object–name pairs in which each inanimate object that had male or female gender was paired with a human male or female name. Even though the experimental subjects were not asked any questions about the qualities of the inanimate objects, speakers of both languages found it more difficult to remember object–name pairs when the paired human name conflicted with the gender of the object in their language. The researchers speculated that the gender associated with the objects affected the subjects’ memory of them as well as their thinking about them. Perception of masculine and feminine properties of inanimate objects is just one example of the subtle effects of language. Other effects may include influences on people’s descriptions of color, time, and spatial location.
Deutscher G. 2010. Through the Looking Glass: Why the World Looks Different in Other Languages. New York: Picador.
Animals communicate with each other in a variety of ways, from the dance of a honey bee to the watery bellow of a whale. Animals and humans also communicate with each other in many ways, such as the verbal command we give our dogs, if we’re lucky, to jump down from the mailman. But do animals use language as we humans do? Human language is a remarkably complex, flexible, and powerful system for communication that involves the creative use of words according to the rules of a systematic grammar. Do other animals have anything similar? Actually, there are two questions we want to examine: Do animals naturally use language? Can animals be taught human language? These questions are difficult to investigate, but the answers have major implications for the evolution of human language.
Let’s first consider the possible use of vocalizations for language in nonhuman primates. In the wild, chimpanzees have been reported to make tens of different vocalizations. There are alarm calls that express fear and warn others of a predator and pant-hoots that announce a chimp’s presence and excitement. However, compared to humans, nonhuman primates appear to have a very limited range of vocalizations, and there is little evidence that they are based on rules (phonological rules) as in humans. Most of the vocalizations chimpanzees make may be stereotyped responses to behavioral situations. In comparison, human language is highly creative; limited only by the rules of grammar, it is effectively infinite. New word combinations and sentences are constantly being made by humans, and the combinations have clear meaning according to the meaning of the individual words plus the rules used in arranging them.
But maybe we haven’t been fair in our comparison; perhaps animal speech requires that they have a language tutor, just as human children must be exposed to language. A variety of animals, including bottlenose dolphins and chimpanzees, have undergone training in an attempt to get them to speak human language. The harbor seal known as Hoover raised by fishermen in Maine learned to utter phrases that sounded something like a drunken man with a New England accent saying “hey hey hello there” or “get outta there.” In the 1940s, several psychologists tried raising baby chimpanzees just like human children, including teaching them to speak. Despite extensive training, the chimps and other animals never learned to utter anything like the range of sounds and words used by humans. In the 1960s, the physician and inventor John Lily, famous for his later development of the sensory deprivation tank and studies with psychedelic drugs, flooded a small house with a few feet of water so that a dolphin could live around the clock with humans. As the trainer moved between a wet bed and a floating desk, she tried to teach the dolphin to speak, such as counting with numbers. Despite positive reports, later experiments did not corroborate the results.
The lack of a rich chimpanzee spoken language and their inability to speak human language are not surprising, given that their vocal tracts are not structured to make the sounds humans make. For example, in chimpanzees and other nonhuman primates, the larynx is much higher, closer to the mouth, making it impossible to make the range of sounds used in human speech. An alternative or adjunct to vocalizations for communication in chimpanzees is the use of gestures and facial expressions. There is evidence indicating that chimpanzees make gestures with the intention of influencing the behavior of other animals. In a recent study, Catherine Hobaiter and Richard Byrne from the University of St. Andrews reported the results of analyzing thousands of gestures categorized into 66 types. Behavioral reactions in other chimpanzees viewing the gestures were used to infer the meaning and intended outcome of each gesture. Examples are “groom me,” “follow me,” and “stop that.” Some gestures appeared to have a particular intention, and others were used more flexibly. This is clearly a much more elaborate system of communication than the dance of a honey bee.
To test and quantify the abilities of animals, a range of studies have attempted to teach them nonverbal communication using words represented by American Sign Language gestures, plastic objects with various patterns and shapes, or keys with different colors and patterns on a keyboard. Well-known examples include the chimpanzee named Washoe, trained by Allen and Beatrix Gardner; the gorilla named Koko, trained by Francine Patterson; and the bonobo named Kanzi, raised by Sue Savage-Rumbaugh. Without question, these animals learned the meanings of gestures or symbols. They showed an ability to understand the meaning of phrases in human language, and they were able to use the improvised communication systems to request objects and actions from the scientists.
Whether we can conclude from such studies that animals use or can use language is controversial. Animals certainly communicate, and to some scientists, their systems of communication are sophisticated enough to be considered rudimentary language. These simpler systems may hint at the origins of human language. Other scientists think the distance between human and animal language is too great, that animals do not use language, defined as communication that is flexible, able to describe new things, and systematic according to rules of grammar. Regardless of your conclusion, it is important to distinguish between language, thought, and intelligence. Language is not a requirement for intelligence or thought. Nonhuman primates, dolphins, and humans raised without any language can do many things requiring abstract reasoning. Many creative people say they do some of their best thinking without words. Albert Einstein claimed that many of his ideas about relativity came from visually thinking of himself riding on a beam of light while looking at clocks and other objects. In any event, Fido does think even if he doesn’t use language as we do (Figure 20.2).

Language processing in the adult human brain relies on carefully orchestrated interactions among a number of cortical areas and subcortical structures that we will look at in a moment. But how does the brain learn to use language? Learning a language, language acquisition, is a remarkable and fascinating process that proceeds in a similar manner in all cultures. The gurgles of newborns turn into babbling around 6 months of age. By 18 months of age, children understand about 150 words and can speak about 50. Interestingly, even at this early age, infants start to lose the ability to distinguish sounds they could discriminate earlier; an example is a Japanese child’s difficulty discriminating the English “r” and “l” sounds because these sounds are not used in Japanese. By 1–2 years of age, children’s speech has the tones, rhythm, and accent of the language they are exposed to. A 3-year-old can produce full sentences and knows roughly 1000 words. By adulthood, a person knows tens of thousands of words. On the other hand, after puberty, learning a second language becomes more difficult. A critical period for language acquisition is suggested by the difficulty older children have learning a second language compared to the first and the difficulty acquiring a first language if they were not exposed to any spoken language before puberty.
The speed at which infants learn language belies the challenges involved. When we first hear a foreign language, it sounds rapidly spoken, and it is difficult to determine where one word stops and the next begins. This is one of the problems faced by infants learning their native tongue. By 1 year of age, however, infants can already recognize the sounds of their language and words even though they don’t understand the words. Spoken language does not reliably indicate the divisions between words; it’s rather like reading text without the spaces (Figure 20.3). Yet, infants must learn to understand thousands of words that are all constructed from the same small pool of language-specific sounds. Jenny Saffran and her colleagues at the University of Wisconsin found that this is achieved by statistical learning in the infant. In other words, the child learns that some combinations of sounds are far more likely than others. When a low probability combination occurs, it suggests the possible location of a word boundary. For example, in the phrase “pretty baby,” the probability that “ty” follows “pret” in a single word is higher than “ba” following “ty” in a single word. Another cue that infants learn to use is the syllable emphasis most common in the language. For example in English, the stress is usually on the first syllable, and this helps determine where words start and stop. Adults of both genders when talking to infants often use “motherese,” in which the speech is slower and exaggerated and vowel sounds are more clearly articulated. Motherese may assist the child in learning speech sounds.

FIGURE 20.3 Word boundaries in spoken and written English. (a) Acoustic analysis of a spoken sentence demonstrates that word boundaries cannot be determined simply from sounds. (b) An analogous situation might be reading text without spaces between words. Indeed, some patterns of letters could form more than one sentence. (Source: Kuhl, 2004.) Description
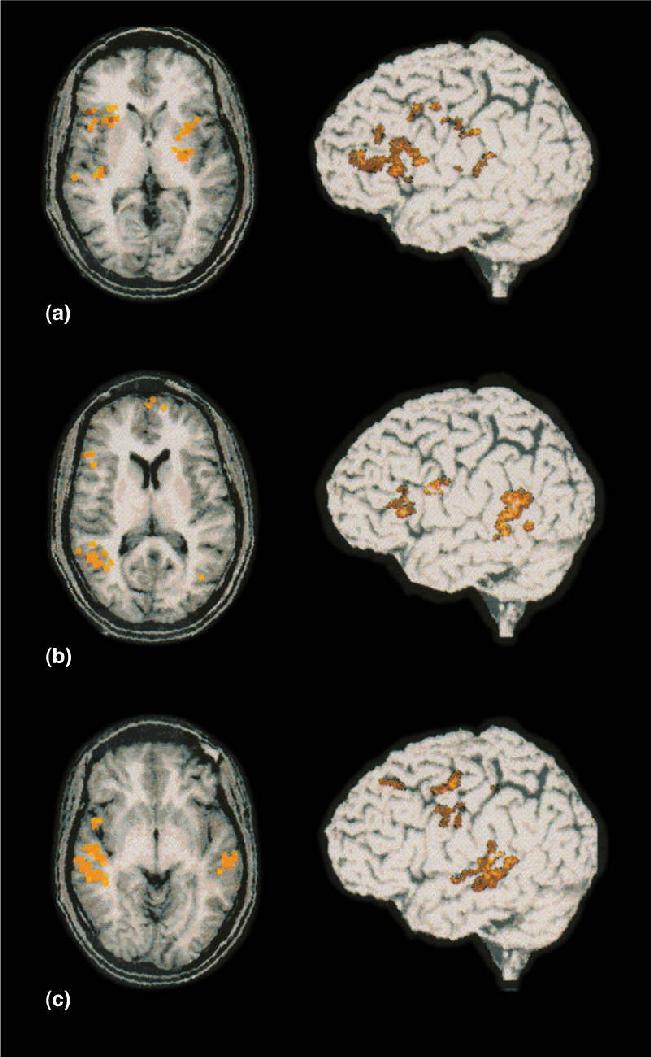
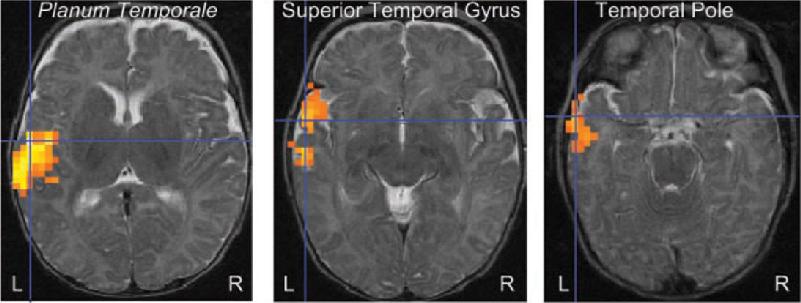
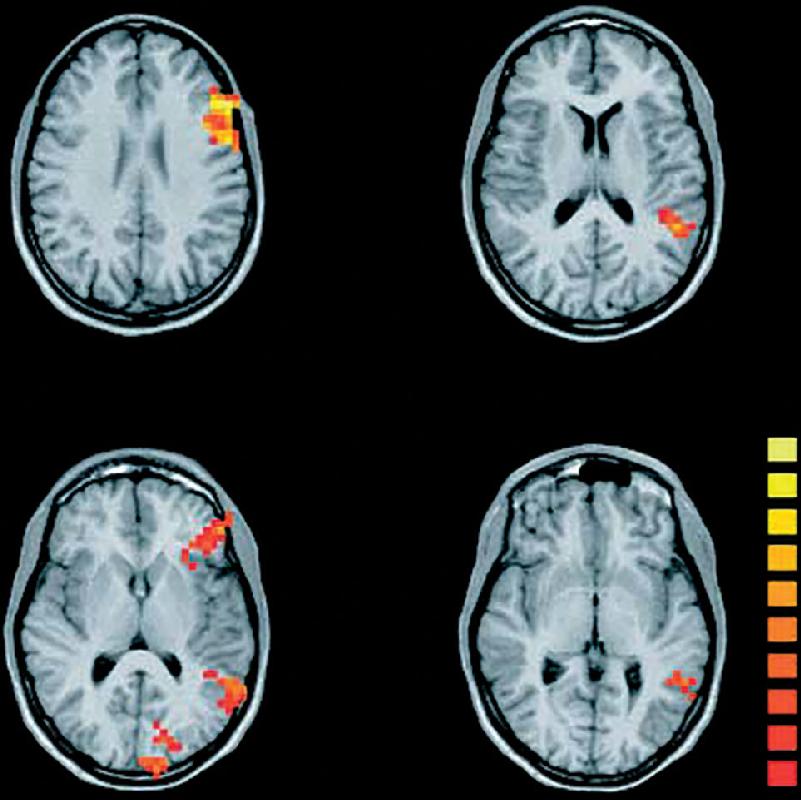
We do not yet know the brain mechanisms by which infants learn to distinguish and speak words. However, Ghislaine Dehaene-Lambertz et al. have found using fMRI that even at the age of 3 months, the brain response to spoken words is distributed in a manner similar to that in adults (Figure 20.4). Listening to speech activates extensive areas in the temporal lobe, with the activation strongly biased toward the left hemisphere. These findings do not show that the infant brain processes language the same as the adult brain, but they indicate a similar early organization of auditory areas and lateralization for language.

FIGURE 20.4 Brain activity in a 3-month-old infant listening to speech. Horizontal sections show the planum temporale, superior temporal gyrus, and the pole of the temporal lobe that were all significantly activated when the infant listened to speech. In the fMRI images, red, orange, and yellow indicate increasingly greater brain activity. (Source: Dehaene-Lambertz et al., 2002.)
Speech and language disorders run in families and are more likely to co-occur in identical twins than in fraternal twins. These observations suggest that genetic factors play an important role in the susceptibility to language disorders. However, for many years the complex patterns of inheritance in language disorders made it difficult to implicate particular genes.
FOXP2 and Verbal Dyspraxia. The view of how genetics may affect language changed dramatically in 1990 with the first publications describing a British family known only as KE. In three generations of the KE family, about half had verbal dyspraxia, an inability to produce the coordinated muscular movements needed for speech (Figure 20.5a). Their speech was largely unintelligible to both the general public and family members, and they developed hand signs to supplement spoken language. In addition to dyspraxia, the affected members of the KE family had broader difficulties involving grammar and language and a lower IQ than unaffected members. The deficit was thought to be language-specific rather than a more general cognitive impairment because the language problems were observed even in affected family members with normal IQs. Brain scans revealed that affected members of the KE family had structural abnormalities in a variety of motor structures, including the motor cortex, cerebellum, and striatum (caudate and putamen) compared to unaffected members (Figure 20.5b).

FIGURE 20.5 FOXP2 mutations in the KE family. (a) Inheritance of language deficits in three generations of the KE family. (b) In affected KE family members, reduced gray matter was found in the caudate nucleus (upper left), cerebellum (upper right), and Broca’s area in the frontal lobe (bottom). (Sources: a adapted from Watkins et al., 2002; b adapted from Vargha-Khadem et al., 2005.) Description
What is known about the genetics underlying this striking familial language disorder? The first thing to note is that unlike previously observed inherited language disorders that appeared to involve multiple genes, the inheritance pattern seen in the KE family was consistent with the mutation of a single gene. This gene appears to affect the development of the motor cortex, cerebellum, and striatum; there are particular deficits in the muscular control of the lower face. The hunt for the culprit gene was aided by the discovery of an unrelated boy, known as CS, who had a language disorder resembling that in the KE family. Combining what was known about CS and the KE family, the mutated gene was eventually identified as FOXP2, which codes a transcription factor responsible for turning other genes on and off. It would be incorrect to call FOXP2 the language gene, but it does appear to be one critical gene involved in language. From our two parents, we all have two copies of the FOXP2 gene, but a mutation in either one is sufficient to produce severe language deficits. It is a striking finding that the change in a single gene can affect a complex behavior such as speech. It should be recognized, however, that through its action as a transcription factor, FOXP2 is capable of influencing hundreds of other genes that might be involved in language.
Versions of FOXP2 are found in many animals. It is interesting to note that in highly vocal songbirds, FOXP2 is strongly expressed in brain areas involved in song learning. An important question is what might be special in humans that conceivably underlies our extensive language abilities compared to those of nonhuman primates. Only two amino acids distinguish the human form of the FOXP2 protein from the chimpanzee, gorilla, and rhesus monkey form. The evolutionary paths leading to humans and chimpanzees diverged about six million years ago, but it has been estimated that the mutations differentiating human and nonhuman primate FOXP2 genes occurred about 200,000 years ago. An exciting speculation is that a small and relatively recent mutation in the FOXP2 gene set humans on a path toward developing language that was needed for higher cognitive function and the development of human culture.
Genetic Factors in Specific Language Impairment and Dyslexia. After the KE family was studied and the FOXP2 gene implicated in their verbal dyspraxia, other individuals unrelated to the KE family were identified who had various mutations of FOXP2. Their speech impairments were consistent with the idea that a mutation in the FOXP2 gene alone can disrupt the normal development of speech. Affected individuals also exhibited other grammatical and cognitive deficits, but it is not clear if these are distinct deficits or somehow related to the verbal dyspraxia.
Spurred on by the FOXP2 findings, growing numbers of genes have been identified that are potentially involved in common language disorders. For example, specific language impairment (SLI) is found in about 7% of all 6-year-olds in the United States. This condition consists of a developmental delay in the mastery of language that may persist into adulthood and that is not associated with hearing difficulty or more general developmental delays. These children have difficulty learning and using words, especially verbs. Because more than 50% of children with SLI have a parent or sibling with the condition, there appears to be a strong genetic component.
Genetic studies of children with SLI have identified a handful of genes that may be involved. Often mentioned, in addition to FOXP2, are the genes CNTNAP2 and KIAA0319. More interesting than the long acronyms denoting these genes are their functions. CNTNAP2 codes a neurexin protein; these are proteins on the presynaptic side of synapses that serve to hold the presynaptic and postsynaptic elements together. The CNTNAP2 neurexin plays an important role in brain development; it appears to be involved in properly locating potassium channels in developing neurons. KIAA0319 is thought to be critical for neuronal migration during neocortical development as well as normal function of adult neurons. We have not reached the point of knowing the specific neural abnormalities underlying SLI, but the candidate genes have focused attention on key aspects of neuronal migration and development.
Another common disorder associated with language is dyslexia, which involves a difficulty learning to read despite normal intelligence and training. Estimates are that dyslexia occurs in 5–10% of people and is somewhat more common in males than in females. The disorder appears to have a strong genetic link, as children of a dyslexic parent have a roughly 30% chance of being dyslexic, and 30–50% of the siblings of a dyslexic person also have dyslexia. A gene often associated with dyslexia is KIAA0319, one of the genes thought to be involved in specific language impairment. Interestingly, dyslexia is quite often found in individuals with SLI. Comorbidity of the disorders is about 40–50%, suggesting that they may have similar causes or be different manifestations of the same deficit. Like SLI, dyslexia appears to involve deviations from the normal pattern of neocortical development.
THE DISCOVERY OF SPECIALIZED LANGUAGE AREAS IN THE BRAIN
As in many other areas of neuroscience, only in the last century have we come to understand a clear relationship between language and the brain. Much of what we know about the importance of certain brain areas is derived from studies of aphasia. Aphasia is the partial or complete loss of language abilities following brain damage, often without the loss of cognitive faculties or the ability to move the muscles used in speech.
During the Greek and Roman Empires, it was commonly thought that the tongue controls speech and that speech disorders originate there rather than in the brain. If a head injury resulted in a loss of speech, the treatment involved special gargles or massage of the tongue. By the sixteenth century, it had been noted that a person could suffer speech impairment without paralysis of the tongue. However, in spite of this step forward, treatment still included such procedures as cutting the tongue, drawing blood, and applying leeches.
Around 1770, Johann Gesner published a relatively modern theory of aphasia, describing it as the inability to associate images or abstract ideas with their expressive verbal symbols. He attributed this loss to brain damage resulting from disease. Gesner’s definition makes the important observation that in aphasia, cognitive ability may remain intact but some function specific to verbal expression is lost. Despite the incorrect association Franz Joseph Gall and later phrenologists made between skull shape and brain function (see Chapter 1), they too made an important observation about aphasia. They reasoned that cases of brain lesions in which speech is lost but other mental faculties are retained suggest that there is a specific region of the brain used for speech.
In 1825, based on many case studies, the French physician Jean-Baptiste Bouillaud proposed that speech is specifically controlled by the frontal lobes. But it took another four decades before this idea was generally accepted. In 1861, Simon Alexandre Ernest Aubertin, Bouillaud’s son-in-law, described the case of a man who had shot away his frontal skull bone in a failed suicide attempt. While treating this man, Aubertin discovered that if a spatula was pressed against the exposed frontal lobe while the man was speaking, his speech immediately halted and did not resume until the pressure was released. He inferred that the pressure on the brain interfered with the normal function of a cortical area in the frontal lobe.
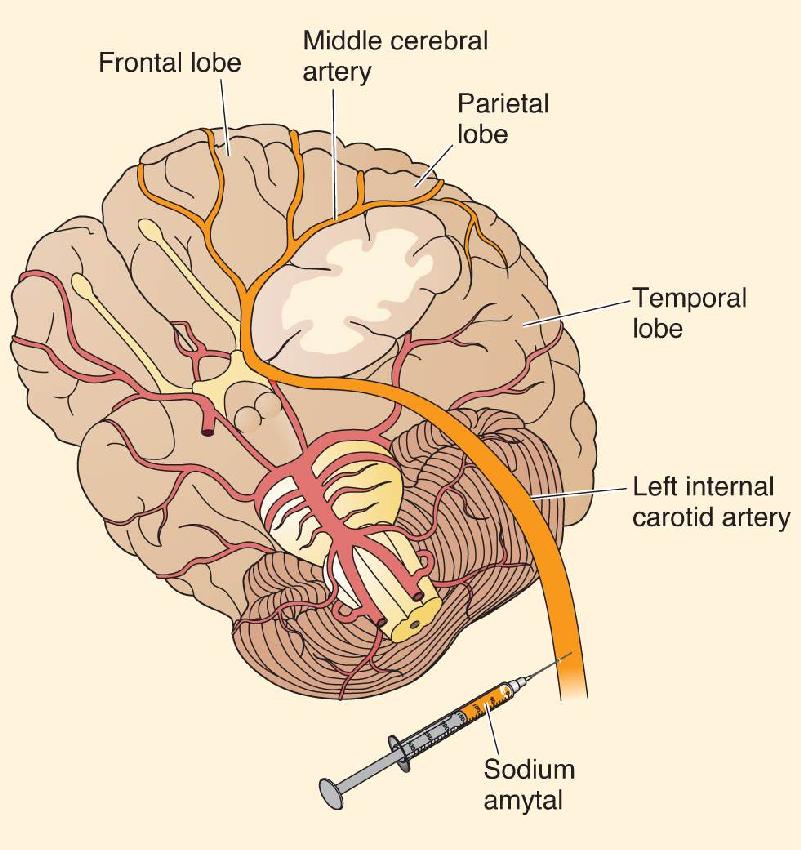
Also in 1861, French neurologist Paul Broca had a patient who was almost entirely unable to speak (the man was called Tan because “tan” was the only sound he could produce). The patient died shortly after Broca met him, and at the postmortem Broca found a lesion in the frontal lobes. Perhaps because of a change in the scientific climate, Broca’s case study appears to have swung popular opinion around to the idea that there is a language center in the brain. In 1863, Broca published a paper describing eight cases in which language was disturbed by damage to the frontal lobe in the left hemisphere. Additional similar cases, along with reports that speech was not disturbed by right hemispheric lesions, led Broca in 1864 to propose that language expression is controlled by only one hemisphere, almost always the left. This view is supported by results from the Wada procedure, in which a single hemisphere of the brain is anesthetized. In most cases, anesthesia of the left hemisphere, but not the right, disrupts speech. In the 1990s, functional brain imaging began to replace the Wada procedure for assessing the dominant hemisphere for language, and the findings are the same (Box 20.2).
The first reports that one of the cerebral hemispheres is dominant for language came from studies of brain-damaged patients. A simple procedure used for studying the function of a single cerebral hemisphere in people without brain damage is the Wada procedure, developed by Japanese-Canadian neurologist Juhn Wada. A fast-acting barbiturate, such as sodium amytal, is injected into the carotid artery on one side of the neck (Figure A). The drug is preferentially carried in the bloodstream to the hemisphere ipsilateral to the injection, where it acts as an anesthetic for about 10 minutes. The effects are sudden and dramatic. Within a matter of seconds, the limbs on the side of the body contralateral to the injection become paralyzed along with loss of somatic sensation.

By asking the patient to answer questions, one can assess his or her ability to speak. If the injected hemisphere is dominant for speech, the patient will be completely unable to talk until the anesthesia wears off. If the injected hemisphere is not dominant, the person can continue to speak throughout the procedure.
Table A shows that in 96% of right-handed people and 70% of left-handed people, the left hemisphere is dominant for speech. Because 90% of all people are right-handed, this means that the left hemisphere is dominant for language in roughly 93% of people. While small but significant numbers of people with either handedness have a dominant right hemisphere, only in left-handers are bilateral representations of speech seen. In the Wada procedure, this is indicated when an injection into either hemisphere has some disruptive effect on speech, although specifics of the disruption may be different for the two hemispheres.
Hemispheric Control of Speech in Relation to Handedness
More recently, fMRI has been used to assess hemispheric dominance for language. As fMRI is not invasive and it is not limited by the short duration of the anesthetic effect of sodium amytal, it has advantages over the Wada procedure. The brain images in Figure B were collected while a subject was given a word and asked to select a synonym from four word options. The brain scans show that frontal, temporal, and parietal areas are activated exclusively in the left hemisphere, which is thus dominant for language in this person. (Note that the convention with MRI images is to show the left hemisphere on the right side of the image.)

Figure B (Source: Spreer et al., 2002, Fig. 4.) Description
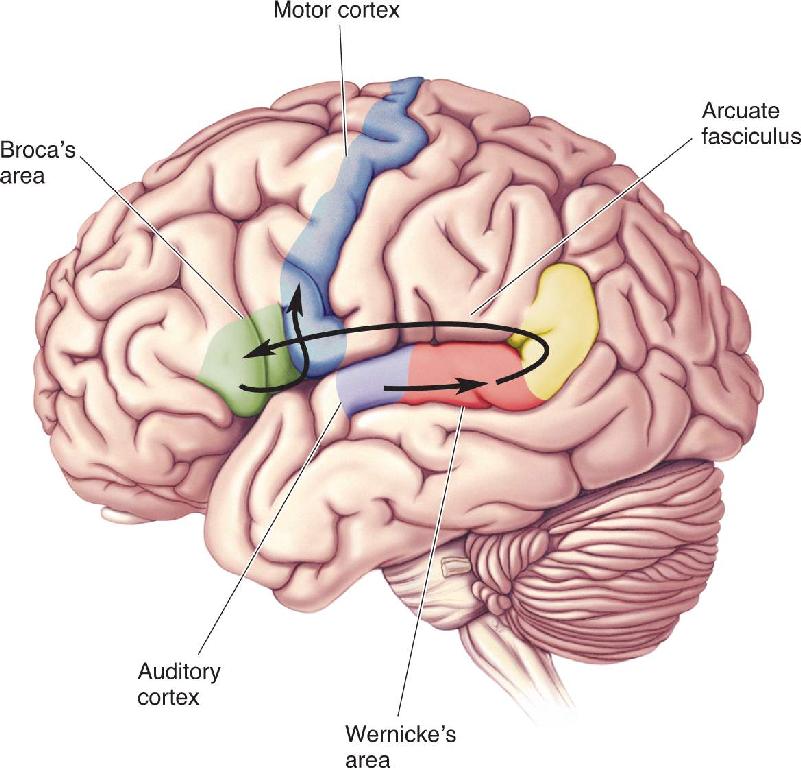
If one hemisphere is thought to be more heavily involved in a particular task, it is said to be dominant. The region of the dominant left frontal lobe that Broca identified as critical for articulate speech has come to be called Broca’s area (Figure 20.6). Broca’s work has considerable significance as the first clear demonstration that brain functions can be anatomically localized.

FIGURE 20.6 Key components of the language system in the left hemisphere. In the frontal lobe, Broca’s area lies next to the area that controls the mouth and lips in the motor cortex. Wernicke’s area, on the superior surface of the temporal lobe, is situated between the auditory cortex and the angular gyrus. Description
In 1874, German neurologist Karl Wernicke reported that lesions in the left hemisphere, in a region distinct from Broca’s area, also disrupt normal speech. Located on the superior surface of the temporal lobe between the auditory cortex and the angular gyrus, this region is now commonly called Wernicke’s area (see Figure 20.6). The nature of the aphasia Wernicke observed is different from that associated with damage to Broca’s area. Having established that there are two language areas in the left hemisphere, Wernicke and others proceeded to construct maps of language processing in the brain. Interconnections between the auditory cortex, Wernicke’s area, Broca’s area, and the muscles required for speech were hypothesized, and different types of language disabilities were attributed to damage in different parts of this system.
Although the terms Broca’s area and Wernicke’s area are still commonly used, the boundaries of these areas are not clearly defined, and they appear to be quite variable from one person to the next. We will also see that each area may be involved in more than one language function, although that more recent finding will make sense only after we look at the aphasias produced by damage to Broca’s and Wernicke’s areas.
As in the studies of Broca and Wernicke, the oldest technique for studying relationships between language and the brain involves correlating functional deficits with lesions in particular brain areas. The occurrence of distinct types of aphasia, as shown in Table 20.1, suggests that language is processed in several stages at different locations in the brain. By examining language deficits that result from damage to different areas of the brain, Nina Dronkers at the University of California at Davis, has clarified much of the neural machinery of language (Box 20.3).

My passion for clinical neuroscience began one day in a class at the University of California at Berkeley where I was pondering my “senioritis” and what to do with my life after college. My professor showed a video of a man who couldn’t read a handwritten message. Paradoxically, the man had just written the message himself! Such cases of language disturbance after brain injury were what sparked my interest in how the brain processes language and continued to fascinate me for the next 30 years.
In working with individuals who have sustained brain injuries, I have had the unique opportunity to evaluate the relationships between affected areas of the brain (as imaged with brain scans) and the speech and language deficits (aphasia) that result from the injury. The first thing that struck me in working with individuals who have aphasia was that the classic relationship between aphasia syndromes and injury to certain language areas was not always as I had learned. I saw patients with Broca’s aphasia who did not necessarily have lesions in Broca’s area, and those with lesions in Broca’s area did not always have Broca’s aphasia. The same discrepancy held for other aphasia syndromes as well. Soon, my colleagues and I realized that some deficits could still be “localized,” but that these needed to be narrowed down into smaller components of the speech and language system, rather than by entire syndromes. Deficits such as coordinating complex articulatory movements could be related to lesions in a small part of the insula, problems with the verbatim repetition of low-frequency sentences were seen after injury to the posterior superior temporal gyrus, and difficulty recognizing the syntactic structure of a sentence could be related to the lesions in the anterior superior temporal gyrus. We found that fiber pathways in the brain also play an important role in language production and comprehension. Destruction of the arcuate fasciculus, for example, can lead to a severe speech production disorder. It became clear that while certain individual brain structures can play a specific role in speech or language functions, aphasia syndromes are caused by injury to large swaths of brain tissue as well as the fiber pathways that connect them. In the normal brain, all of these structures work together in a complex network that helps to support the extraordinary language functions we all take for granted.
A particularly exciting time of my life occurred in Paris, France, where I had the extraordinary opportunity to study the brains of Paul Broca’s first two patients. These were the cases of aphasia he had examined as a surgeon in 1861 and whose deficits led Broca to believe that the inferior part of the frontal lobe was important for spoken language. Since so much had been written about Broca’s area, particularly with the growth of functional neuroimaging with PET and fMRI in cognitive neuroscience, it was clearly necessary to revisit these cases and see which anatomical areas were actually affected in these historic brains. Luckily, the brains had never been dissected or discarded, and my colleague, Odile Plaisant, and I were tremendously fortunate in being able to examine these brains more closely. We could see right away that the area we now think of as Broca’s area was only partially affected in these two cases. We were curious to know how deeply the injuries extended, and with the expertise of Marie-Thérèse Iba-Zizen and Emanuel Cabanis, both neuroradiologists, we were actually able to scan these two important brains in an MRI scanner and acquired high-resolution images with exquisite detail.
What astonished us was the degree of involvement of other regions of the brain, particularly in the insula and in the fiber tracts that travel throughout the brain. The case of Mr. Leborgne, or “Tan,” Broca’s first and most famous case, had extensive involvement of the insula but only a portion of what we now call Broca’s area. In addition, the major fiber bundles, including the arcuate and superior longitudinal fasciculi that travel between the frontal and posterior parts of the brain, were completely destroyed. Broca’s second case, Mr. Lelong, had atrophy in the insula, but when the scanner advanced into the deeper parts of the brain, we saw several small lesions again in the arcuate and superior longitudinal fasciculi. This had never been seen before, and we were quite thrilled to see it unfold as we watched. Thus, we saw that the damage that produced the aphasia in these two cases was more extensive than previously thought but, in fact, consistent with what we see in our current cases of severe Broca’s aphasia.
As a neuroscientist, I feel extraordinarily lucky to be able to work with individuals with aphasia who have taught us so much about this remarkable part of the body. The brain is, in many ways, still an open frontier, with much to be learned about its functions, its mechanisms, and its potential for recovery. The next generations of neuroscientists will have much to contribute to our knowledge of the brain and will surely experience the same excitement of discovery as those who came before them.
The syndrome called Broca’s aphasia is also known as motor or nonfluent aphasia, because the person has difficulty speaking even though he or she can understand language heard or read. The case of David Ford is typical. Ford was a radio operator in the Coast Guard when, at age 39 years, he suffered a stroke. He remained an intelligent man, but he had little control over his right arm and leg (demonstrating that his lesion was in the left hemisphere). His speech was also abnormal, as the following discussion with psychologist Howard Gardner illustrates:
“I asked Mr. Ford about his work before he entered the hospital.
I’m a sig. . . no. . . man. . . uh, well,. . . again.” These words were emitted slowly, and with great effort. The sounds were not clearly articulated; each syllable was uttered harshly, explosively, in a throaty voice. With practice, it was possible to understand him, but at first I encountered considerable difficulty in this.
“Let me help you,” I interjected. “You were a signal. . . .”
“A signal man. . . right,” Ford completed my phrase triumphantly.
“No, er, yes, yes. . . ship. . . Massachu. . . chusetts. . . Coastguard. . .years.” He raised his hands twice, indicating the number “nineteen.”
“Could you tell me, Mr. Ford, what you’ve been doing in the hospital?”
“Yes, sure. Me go, er, uh, P.T. nine o’cot, speech. . . two times. . . read. . . wr. . . ripe, er, rike, er, write. . . practice. . . get-ting better.”
“Why, yes. . . Thursday, er, er, er, no, er, Friday. . . Bar-ba-ra. . . wife. . . and, oh, car. . . drive. . . purnpike. . . you know. . . rest and. . . tee-vee.”
“Are you able to understand everything on television?”
“Oh, yes, yes. . . well. . . almost.” Ford grinned a bit. (Gardner, 1974, pp. 60–61)
People with Broca’s aphasia have difficulty saying anything, often pausing to search for the right word. The inability to find words is called anomia (literally meaning “no name”). Interestingly, there are certain “overlearned” things Broca aphasics can say without much hesitation, such as the days of the week and the American Pledge of Allegiance. The hallmark of Broca’s aphasia is a telegraphic style of speech, in which mainly content words (nouns, verbs, and adjectives carrying content specific to the sentence) are used. For instance, when Mr. Ford was asked about being in the Coast Guard, his answer contained the words “ship,” “Massachusetts,” “Coast Guard,” and “years,” but little else. Many function words (articles, pronouns, and conjunctions connecting the parts of the sentence grammatically) are left out (there are no ifs, ands, or buts). As well, verbs are frequently not conjugated. In the jargon of aphasia deficits, the inability to construct grammatically correct sentences is called agrammatism. There are some peculiar nuances to the agrammatical tendencies in Broca’s aphasia. Ford, for example, could read and use the words “bee” and “oar” but had difficulty with the more common words “be” and “or.” This problem is related not to the word’s sound but to whether or not it is a noun. In a similar vein, Broca aphasics have difficulty repeating things spoken to them, although they tend to be better with familiar nouns such as “book” and “nose.” Sometimes they substitute incorrect sounds or words (Ford said “purnpike” for “turnpike”); these are called paraphasic errors.
In contrast to the speech difficulties in Broca’s aphasia, comprehension is generally quite good. In the dialogue above, Ford seemed to understand the questions asked of him, and for the most part, he said he understood what he saw on television. In Gardner’s study, Ford was able to answer simple questions, such as “Does a stone float on water?” However, more difficult questions demonstrated that he did not have completely normal comprehension abilities. If he was told “The lion was killed by the tiger; which animal is dead?” or “Put the cup on top of the fork and place the knife inside the cup,” he had difficulty understanding. This was probably related to the fact that he generally had trouble with the function words “by” in the first example and “on top of” in the second example.
Because the most obvious difficulty is in producing speech, Broca’s aphasia is considered a language disturbance toward the motor end of the language system. Language is understood but not easily produced. While it is true that Broca aphasics are worse at speech than other types of aphasics, several characteristics suggest that there is more to the syndrome. As pointed out earlier, comprehension is generally good, but comprehension deficits can be demonstrated by tricky questions. Also, a simple motor deficit would not explain the ability to say “bee” but not “be.” Finally, patients sometimes have considerable anomia, suggesting that they have problems “finding” words as well as making the appropriate sounds.
Wernicke suggested that the area damaged in Broca’s aphasia contains memories for the fine series of motor commands required for articulating word sounds. Because Broca’s area is near the part of the motor cortex that controls the mouth and lips, there is an appealing logic to this idea. Wernicke’s theory is still held by some, but there are other ways of looking at the problem. For instance, the difference in the aphasic’s ability to use content words and function words suggests that Broca’s area and nearby cortex may be specifically involved in making grammatical sentences out of words. This might explain why Mr. Ford could produce sounds such as “bee” and “oar” when they represent content words but not when the sounds represent the function words “be” and “or.”
When Wernicke noted that superior temporal lesions could lead to aphasia, the syndrome he observed was quite distinct from Broca’s aphasia. Indeed, Wernicke suggested there were two general types of aphasia. In Broca’s aphasia, speech is disturbed but comprehension is relatively intact. In Wernicke’s aphasia, speech is fluent but comprehension is poor. (Although these descriptions are oversimplified, they are useful for remembering these syndromes.)
Let’s consider the case of Philip Gorgan, another patient studied by Gardner.
“What brings you to the hospital?” I asked the 72-year-old retired butcher 4 weeks after his admission to the hospital.
“Boy, I’m sweating, I’m awful nervous, you know, once in a while I get caught up, I can’t mention the tarripoi, a month ago, quite a little, I’ve done a lot well, I impose a lot, while, on the other hand, you know what I mean, I have to run around, look it over, trebbin and all that sort of stuff.”
I attempted several times to break in, but was unable to do so against this relentlessly steady and rapid outflow. Finally, I put up my hand, rested it on Gorgan’s shoulder, and was able to gain a moment’s reprieve.
“Oh sure, go ahead, any old think you want. If I could I would. Oh, I’m taking the word the wrong way to say, all of the barbers here whenever they stop you it’s going around and around, if you know what I mean, that is tying and tying for repucer, repuceration, well, we were trying the best that we could while another time it was with the beds over there the same thing. . . .” (Gardner, 1974, pp. 67–68)
Clearly, Mr. Gorgan’s speech is altogether different from that of Mr. Ford. Gorgan’s speech is fluent, and he has no trouble using function words as well as content words. If you didn’t understand English, his speech would probably sound normal because of its fluency. However, the content does not make much sense. It is a strange mixture of clarity and gibberish. Along with their far greater output of speech compared to Broca aphasics, Wernicke aphasics also make far more paraphasic errors. Gorgan would sometimes use the correct sounds but in an incorrect sequence, such as “plick” instead of “clip.” Occasionally, he would stumble around the correct sound or word, as when, in another conversation, he called a piece of paper “piece of handkerchief, pauper, hand pepper, piece of hand paper.” Interestingly, he would sometimes use an incorrect word but one categorically similar to the correct word, such as “knee” instead of “elbow.”
Because of the stream of unintelligible speech, it is difficult to assess with speech alone whether Wernicke aphasics comprehend what they hear or read. Indeed, one of the intriguing things about Wernicke aphasics is that they frequently appear undisturbed by the sound of their own speech and the speech of others, even though they probably don’t understand either. Comprehension is usually assessed by asking the patient to respond in a nonverbal manner. For instance, the patient could be asked to put object A on top of object B. Questions and commands of this sort quickly lead to the conclusion that Wernicke aphasics do not understand most instructions. They are completely unable to comprehend questions of the sort understood by Broca aphasics. When Gorgan was presented with commands written on cards (“Wave goodbye,” “Pretend to brush your teeth”), he was often able to read the words but never acted as if he understood what they meant.
Gorgan’s strange speech was mirrored in his writing and his ability to play music. When Gardner gave him a pencil, he spontaneously took it and wrote “Philip Gorgan. This is a very good beautifyl day is a good day, when the wether has been for a very long time in this part of the campaning. Then we want on a ride and over to for it culd be first time. . . .” (p. 71). Likewise, when he sang or played the piano, pieces of the appropriate song were intermixed with musical gibberish, and he had a difficult time ending, just as in his speech.
Insight about the possible function of Wernicke’s area is provided by its location on the superior temporal gyrus near the primary auditory cortex. Wernicke’s area may play a critical role in relating incoming sounds to their meaning. In other words, it is an area specialized for storing memories of the sounds that make up words. Some have suggested that Wernicke’s area is a high-order area for sound recognition in the same sense that the inferior temporal cortex is thought to be a high-order area for visual recognition. A sound recognition deficit would explain why Wernicke aphasics don’t comprehend speech well. However, there must be more to Wernicke’s area to account for the odd speech patterns and comprehension deficits. Speech in Wernicke’s aphasia suggests that Broca’s area and the system responsible for speech production are running without control over content. The speech zooms along, swerving in every direction like a car with a sleepy driver at the wheel.
The Wernicke–Geschwind Model of Language and Aphasia
Shortly after making his observations about what came to be called Wernicke’s aphasia, Wernicke proposed a model for language processing in the brain. Later extended by Norman Geschwind at Boston University, this model is known as the Wernicke–Geschwind model. The key anatomical elements in the system are Broca’s area, Wernicke’s area, the arcuate fasciculus (a bundle of axons connecting the two cortical areas), and the angular gyrus. The model also includes sensory and motor areas involved in receiving and producing language. To understand what the model entails, we’ll consider the performance of two tasks.
The first task is the repetition of spoken words (Figure 20.7). When incoming speech sounds reach the ear, the auditory system processes the sounds, and neural signals eventually reach the auditory cortex. According to the model, the sounds are not understood as meaningful words until they are processed in Wernicke’s area. For the person to be able to repeat the words, word-based signals are passed to Broca’s area from Wernicke’s area via the arcuate fasciculus. In Broca’s area, the words are converted to a code for the muscular movements required for speech. Output from Broca’s area is sent to the nearby motor cortical areas responsible for moving the lips, tongue, larynx, and so on.

FIGURE 20.7 The pathway involved in repeating a spoken word, according to the Wernicke–Geschwind model. Description
The second task we’ll consider is reading written text aloud (Figure 20.8). In this case, the incoming information is processed by the visual system through the striate cortex and higher-order visual cortical areas. The visual signals are then passed to the angular gyrus at the junction of the occipital, parietal, and temporal lobes. In the cortex of the angular gyrus, it is assumed that a transformation occurs so that the output evokes the same pattern of activity in Wernicke’s area as if the words were spoken rather than written. From this point, the processing follows the same progression as in the first example: Wernicke’s area to Broca’s area to motor cortex.

FIGURE 20.8 The pathway involved in repeating aloud a written word, according to the Wernicke–Geschwind model. Description
This model offers simple explanations for key elements of both Broca’s and Wernicke’s aphasia. A lesion in Broca’s area seriously interferes with speech production because the proper signals can no longer be sent to the motor cortex. On the other hand, comprehension is relatively intact because Wernicke’s area is undisturbed. A lesion in Wernicke’s area produces great comprehension problems because this is the site of the transformation of sounds into words. The ability to speak is unaffected because Broca’s area is still able to drive the muscles required for speech.
The Wernicke–Geschwind model has several errors and oversimplifications, however. For example, words read do not have to be transformed into a pseudo-auditory response, as suggested in the reading task described earlier. In fact, visual information can reach Broca’s area from the visual cortex without making a stop at the angular gyrus. One of the dangers inherent in any model is overstating the significance of a given cortical area for a particular function. It has been found that the severity of Broca’s and Wernicke’s aphasias depends on how much cortex is damaged beyond the limits of Broca’s and Wernicke’s areas. Also, aphasia is influenced by damage to subcortical structures such as the thalamus and caudate nucleus, which are not in the model. In surgical cases where parts of cortex are removed, the resulting language deficits are usually milder than the deficits resulting from stroke, which affects both cortical and subcortical structures.
Another important factor is that there is often significant recovery of language function after a stroke, and it appears that other cortical areas can sometimes compensate for what is lost. As with many neurological syndromes, young children generally recover very well, but even adults, especially left-handers, can show good recovery of function.
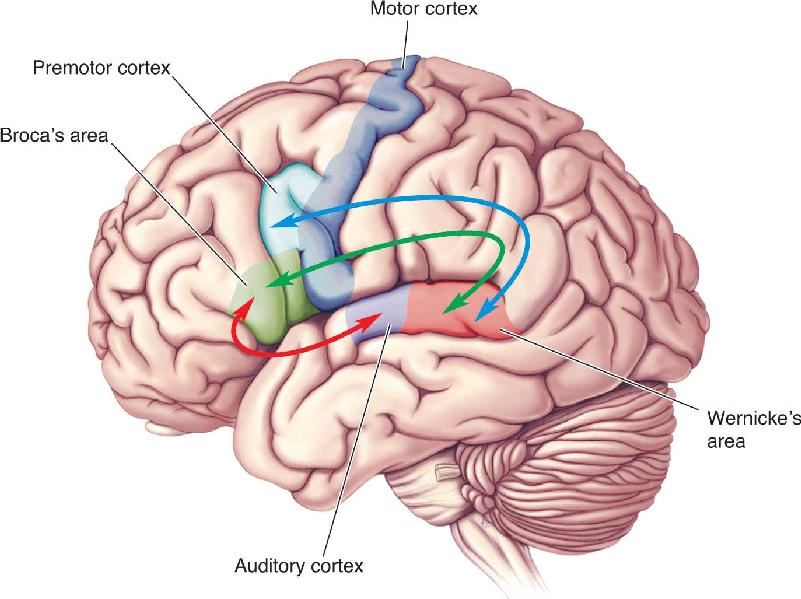
A final problem with the Wernicke–Geschwind model is that most aphasias involve both comprehension and speech deficits. Mr. Ford, with Broca’s aphasia, had good comprehension but was confused by complex questions. Conversely, Mr. Gorgan, with Wernicke’s aphasia, had several speech abnormalities in addition to a severe lack of comprehension. Therefore, in cortical processing, the sharp functional distinctions between regions as implied by the model do not exist. Despite its problems, the Wernicke–Geschwind model continues to be of clinical use because of its simplicity and approximate validity. In the latter half of the twentieth century, numerous more elaborate language models were developed to account for the complexities of language and brain processing as well as the shortcomings of the Wernicke–Geschwind model. Similar to parallel pathways that have been outlined in the visual system, these models involve a variable number of parallel pathways with distinct but interacting functions (Figure 20.9).

FIGURE 20.9 Parallel language pathways. Current models of language processing emphasize multiple streams of processing, much like the dorsal and ventral streams described for vision. The model here includes two dorsal pathways and a ventral pathway. Note that unlike in the Wernicke–Geschwind model, language here is not based on a single stream connecting Wernicke’s area with Broca’s area via the arcuate fasciculus. One dorsal pathway (blue) connects the superior temporal gyrus (Wernicke’s and auditory areas) with premotor cortex and is involved in speech production and repeating words. The other dorsal pathway (green) connects the superior temporal gyrus with Broca’s area and is thought to be involved in processing complex syntactic structure—that is, the analysis of words arranged according to a grammar system. The ventral stream (red) takes the sounds of speech and extracts their meaning. (Adapted from Berwick et al., 2013, Fig. 2.) Description
The value of any model involves not only its ability to account for previous observations but also its ability to predict. Based on his observation that distinct forms of aphasia result from lesions in areas of frontal and superior temporal cortex, Wernicke predicted that a unique form of aphasia should result from a lesion that disconnects these two brain areas but leaves the areas themselves intact. In the Wernicke–Geschwind model, a lesion in the fibers composing the arcuate fasciculus could accomplish this. In reality, such disconnection lesions usually involve damage to parietal cortex in addition to the arcuate fasciculus, but Broca’s and Wernicke’s areas are spared.
Wernicke’s prediction turned out to be correct; aphasia from a disconnection lesion was demonstrated and is now known as conduction aphasia. As the model predicts, based on the preservation of Broca’s and Wernicke’s areas, comprehension is good and speech is fluent. Patients are typically able to express themselves through speech without difficulty. The deficit that chiefly characterizes conduction aphasia is difficulty in repeating words. In response to hearing a few words, the patient may attempt to repeat what was said, but the repetition will substitute words, omit words, and include paraphasic errors. Repetition is usually best with nouns and short common expressions, but it may fail entirely if the spoken words are function words, polysyllabic words, or nonsense sounds. Interestingly, a person with conduction aphasia comprehends sentences he or she reads aloud, even though what is said aloud contains many paraphasic errors. This is consistent with the idea that comprehension is good and the deficit occurs between the regions involved in comprehension and speech.
One of the sad yet fascinating things about aphasia is the diversity of syndromes that occur following strokes. While these syndromes challenge language models, each one offers a clue to our understanding of language processing. Characteristics of a few other aphasias are listed in Table 20.1.
Cases of aphasia in bilingual people and deaf people provide fascinating insight about language processing in the brain. Suppose a person knows two languages before having a stroke. Does the stroke produce aphasia in one language and not the other, or do both languages suffer equally? The answer depends on several factors, including the order in which the languages were learned, the fluency achieved in each language, and how recently the language was used. The consequences of a stroke are not always predictable, but the language learned more fluently and earlier in life tends to be relatively more preserved. If the person learned two languages at the same time to equivalent levels of fluency, a lesion will probably produce similar deficits in both languages. If the languages were learned at different times in life, it is likely that one language will be affected more than the other. The implication is that the second language may make use of different, although overlapping, populations of neurons from those used by the first.
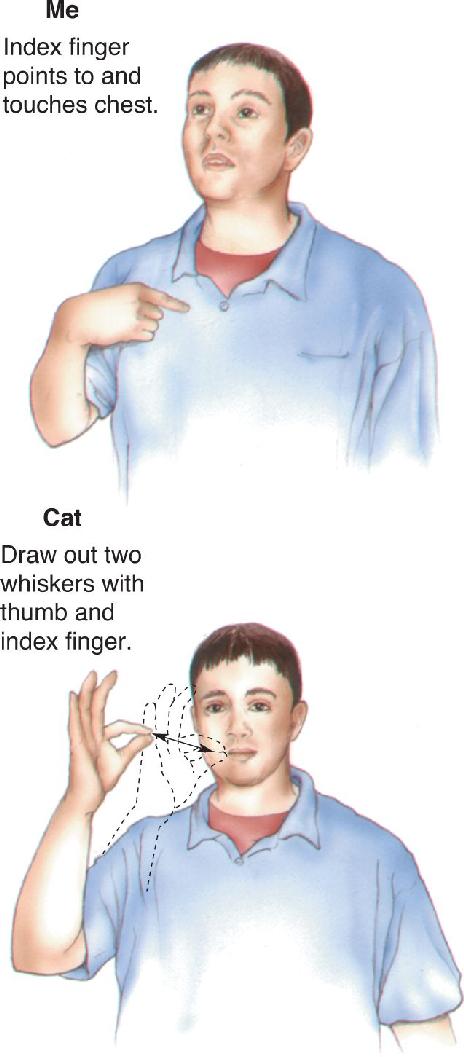
The study of language deficits in those who are deaf and/or know sign language suggests that there is some universality to language processing in the brain. American Sign Language uses hand gestures to express the ideas and emotions most of us convey with spoken language (Figure 20.10). Left hemispheric lesions in people who use sign language appear to cause language deficits similar to those occurring in verbal aphasics. There are cases analogous to Broca’s aphasia in which comprehension is good but the ability to “speak” through sign language is severely impaired. Importantly, the ability to move the hands is not impaired (i.e., the problem is not with motor control). Rather, the deficit is specific to the use of hand movements for the expression of language.

FIGURE 20.10 “Speaking” in American Sign Language. Description
There are also sign language versions of Wernicke’s aphasia in which the patient signs fluently but with many mistakes while also having difficulty comprehending the signing of others. In one unusual case, a hearing man who was the child of deaf parents learned both sign language and verbal language. A left hemispheric stroke initially gave him global aphasia, but his condition significantly improved with time. An important observation was that his verbal and sign languages recovered together, as if overlapping brain areas were used. While there do appear to be aphasias in those using sign language analogous to speech aphasias, there is also evidence that signing aphasia and speaking aphasia can be produced by left hemispheric lesions in somewhat different locations.
ASYMMETRICAL LANGUAGE PROCESSING IN THE TWO CEREBRAL HEMISPHERES
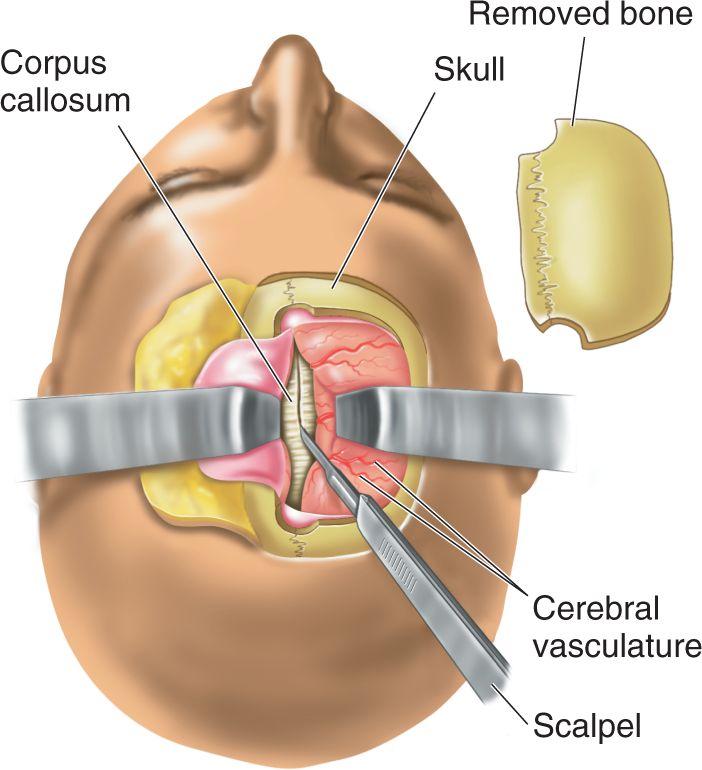
We have seen that damage to certain parts of the brain leads to a variety of aphasias. As the early work of Broca indicated, language is usually not handled equally by the two cerebral hemispheres. Some of the most valuable and fascinating findings on the language differences of the two hemispheres come from split-brain studies in which the hemispheres are surgically disconnected. Communication between the cerebral hemispheres is normally served by several bundles of axons known as commissures. Recall from Chapter 7 that the largest of these is the great cerebral commissure, also called the corpus callosum (Figure 20.11). The corpus callosum consists of about 200 million axons crossing between the hemispheres. Surely such a huge bundle of fibers must be of great importance. Surprisingly, until about 1950, researchers had been unable to demonstrate any important role of the corpus callosum.

FIGURE 20.11 The corpus callosum. The corpus callosum is the largest bundle of axons providing communication between the cerebral hemispheres.
In split-brain procedures, the skull is opened and the axons making up the corpus callosum are severed (Figure 20.12). The hemispheres may retain some communication via the brain stem or smaller commissures (if they aren’t also severed), but most of the intercerebral communication is lost. In the 1950s, Roger Sperry and his colleagues at the University of Chicago and later at the California Institute of Technology performed a series of experiments, using split-brain animals, to explore the function of the corpus callosum and the separated cerebral hemispheres. Sperry’s group confirmed earlier reports that cutting the corpus callosum in a cat or monkey has no noticeable effect on the animal’s behavior. Temperament is unchanged, and the animals appeared to be normal in coordination, reaction to stimuli, and ability to learn. However, in cleverly devised experiments, Sperry’s group showed that the animals sometimes acted as if they had two separate brains. For example, in one experiment, circle and cross visual stimuli were both shown to a monkey’s left eye and the animal was trained to select the circle. In alternate trials, it was trained to choose the cross when the same visual stimuli were presented to the right eye. With both eyes open, it is not possible for the monkey (or humans) to know which eye sees a stimulus, so it appeared that the two cerebral hemispheres were learning opposite discriminations at the same time—thinking with two brains. You might wonder what happens if both eyes simultaneously see the stimuli. The answer is that the monkey hesitates and then chooses either the cross or the circle and sticks with this choice for a series of trials before switching to the opposite stimulus. The scientists speculated that the two cerebral hemispheres competed and, on any given trial, one won.

FIGURE 20.12 Split-brain surgery in a human. To reach and cut the corpus callosum, a portion of the skull is removed and the cerebral hemispheres are retracted. Description
Because split-brain monkeys did not appear to have any major deficits, surgeons felt they were justified in cutting the corpus callosum as a last resort in treating certain types of severe epilepsy in humans. They hoped to prevent the spread of epileptic activity from one hemisphere to the other. It may seem questionable to cut 200 million axons on the assumption that they are not very important, but the surgery is often beneficial in restoring a seizure-free life. Michael Gazzaniga, then at New York University, studied a number of these people. Gazzaniga had initially worked with Sperry, and his techniques were modifications of those used with experimental animals.
One key methodological feature of studying split-brain humans involves careful control to present visual stimuli to only one cerebral hemisphere. Gazzaniga did this by taking advantage of the fact that only the right hemisphere sees objects to the left of the point of fixation, and only the left hemisphere sees objects to the right (see Figure 10.3), as long as the eyes can’t move to bring the image onto the fovea (Figure 20.13). Pictures or words were flashed on for a fraction of a second using a device with a camera-like shutter. Note that when a shutter opens it does not present a stimulus to one eye and not the other; instead, it presents a stimulus to both eyes in such a way that only one cerebral hemisphere “sees” the stimulus. Because the images were presented for a shorter time than that required to move the eyes, the images were seen by only one hemisphere.

FIGURE 20.13 Visual stimulation of one hemisphere in humans. A visual stimulus is briefly flashed to the left or right visual field by means of a shutter. Each projector shows an image to both eyes in such a way that only one cerebral hemisphere “sees” the image. The display time is shorter than the time needed to generate a saccadic eye movement, thus assuring that only one hemisphere sees the stimulus. Description
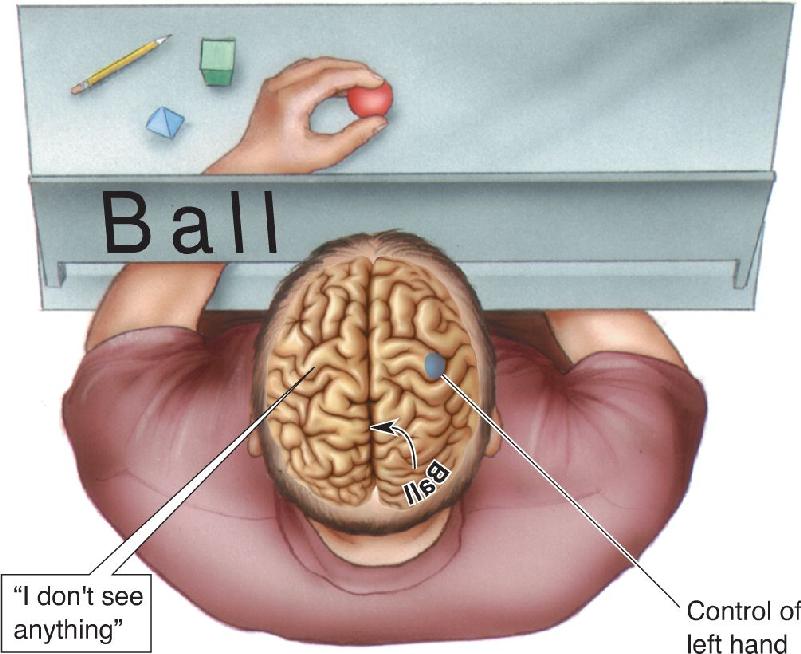
Left Hemisphere Language Dominance. Although split-brain humans are normal in most every way, there is a striking asymmetry in their ability to verbalize answers to questions posed separately to the two hemispheres. For instance, they can repeat or describe numbers, words, and pictures visually presented only in the right visual field because the left hemisphere is usually dominant for language. Likewise, they can describe objects being manipulated only by the right hand (out of view of both eyes). These findings would be entirely unremarkable except for the fact that such simple verbal descriptions of sensory input are impossible for the right hemisphere.
If an image is shown only in the left visual field or an object is felt only by the left hand, a split-brain person cannot describe it and usually says that nothing is there (Figure 20.14). An object could be covertly placed in a patient’s left hand, and there would be no verbal indication of even noticing. This absence of response is a consequence (and demonstration) of the fact that the left hemisphere controls speech in most people. If you think about the implications for split-brain people, you’ll realize that they have an unusual existence. Following their surgery, they are unable to describe anything to the left of their visual fixation point: the left side of a painting, the left side of a room, and so on. What is startling is that this doesn’t seem to disturb the patients.

FIGURE 20.14 Demonstrating language comprehension in the right hemisphere. A split-brain person shown a word in the left visual field will say nothing is seen. This is because the left hemisphere, which usually controls speech, did not see the word, and the right hemisphere, which saw the word, cannot speak. Information about the word cannot be transferred to the verbal left hemisphere without the corpus callosum. However, the left hand, which is controlled by the right hemisphere, can pick out the object corresponding to the word by touch alone.
Language Functions of the Right Hemisphere. While there is a dramatic inability of the right hemisphere to speak, this does not mean it knows nothing of language. It can be demonstrated that the right hemisphere can read and understand numbers, letters, and short words as long as the required response is nonverbal. In one experiment, the right hemisphere is shown a word that is a noun. As already mentioned, the person will say nothing is seen. Of course, that’s the talkative left hemisphere speaking, and it didn’t see anything. But if the person is urged to use the left hand to select a card containing a picture corresponding to the word shown, or pick out an object by touch, the person can do it (see Figure 20.14). The right hemisphere cannot do this with more complex words or sentences, but the results clearly imply that the right hemisphere does have language comprehension.
A study conducted by Kathleen Baynes, Michael Gazzaniga, and their colleagues then at the University of California at Davis suggests that the right hemisphere may sometimes also be able to write even if it can’t speak. In most people, reading, speaking, and writing are all controlled by the left hemisphere. However, in a split-brain woman known as V.J., this was found not to be true. Words were flashed to either her left or her right hemisphere. Words seen by her left hemisphere could be spoken but not written. Conversely, she could write but not speak words shown to her right hemisphere. While this separation of function may be abnormal, the case of V.J. indicates that there isn’t necessarily a single brain system for all aspects of language located in one hemisphere.
Evidence also suggests that the right hemisphere understands complex pictures despite its inability to say so. In one experiment, a subject was shown a series of pictures in her left visual field, and at one point a nude photo appeared in the series. When the researcher asked what she saw, she said “Nothing,” but then she began to laugh. She told the researcher that she didn’t know what was funny, but that perhaps it was the machine used in the experiment.
The right hemisphere appears to be more skilled at certain tasks than the left hemisphere. For instance, even though the split-brain patients were right-handed and thus their left hemispheres were much more practiced at drawing, the left hand controlled by the right hemisphere was better at drawing or copying figures containing three-dimensional perspective. These patients were also better at solving complex puzzles with their left hand. It has also been reported that the right hemisphere is somewhat better at perceiving nuances in sound.
In some split-brain studies, the two hemispheres initiated conflicting behaviors, apparently because they were thinking differently. In one task, a patient was asked to arrange a group of blocks to match a pattern on a small card. He was told to do this using only his right hand (left hemisphere), which is not generally good at this type of task. As the right hand struggled to arrange the blocks, the left hand (right hemisphere), which knew how to do it, reached in to take over. Only the restraint of the experimenter kept the left hand from pushing the right one out of the way to solve the puzzle. Another patient studied by Gazzaniga would sometimes find himself pulling his pants down with one hand while pulling them up with the other. These bizarre behaviors make a strong case that there are two independent brains controlling the two sides of the body.
The results of these split-brain studies demonstrate that the two hemispheres can function as independent brains and that they have different language abilities. Although the left hemisphere is usually dominant for language, the right hemisphere has significant skills in comprehending language. It is important to keep in mind that the split-brain studies test the ability of each hemisphere to perform on its own. Presumably in the intact brain, the callosum allows for synergistic interactions between the hemispheres for language and other functions.
In the nineteenth century, there were reports of anatomical differences between the two hemispheres. For example, it was noted that the left Sylvian fissure is longer and less steep than the right (Figure 20.15). However, as recently as the 1960s, there was considerable doubt about the existence of significant cortical asymmetries. Because of the strikingly asymmetrical control of speech demonstrated by the Wada procedure, it would be interesting to know if the two cerebral hemispheres are anatomically different. Some of the first good quantitative data demonstrating hemispheric differences came from the work of Geschwind and his colleague Walter Levitsky. Initial observations were made of postmortem brains, but more recently, the results were confirmed with magnetic resonance imaging (MRI) (see Box 7.2).

FIGURE 20.15 Asymmetry of the Sylvian fissure. In most right-handed people, the Sylvian fissure in the left hemisphere is longer and runs at a shallower angle than the fissure in the right hemisphere. (Source: Adapted from Geschwind, 1979, p. 192.)
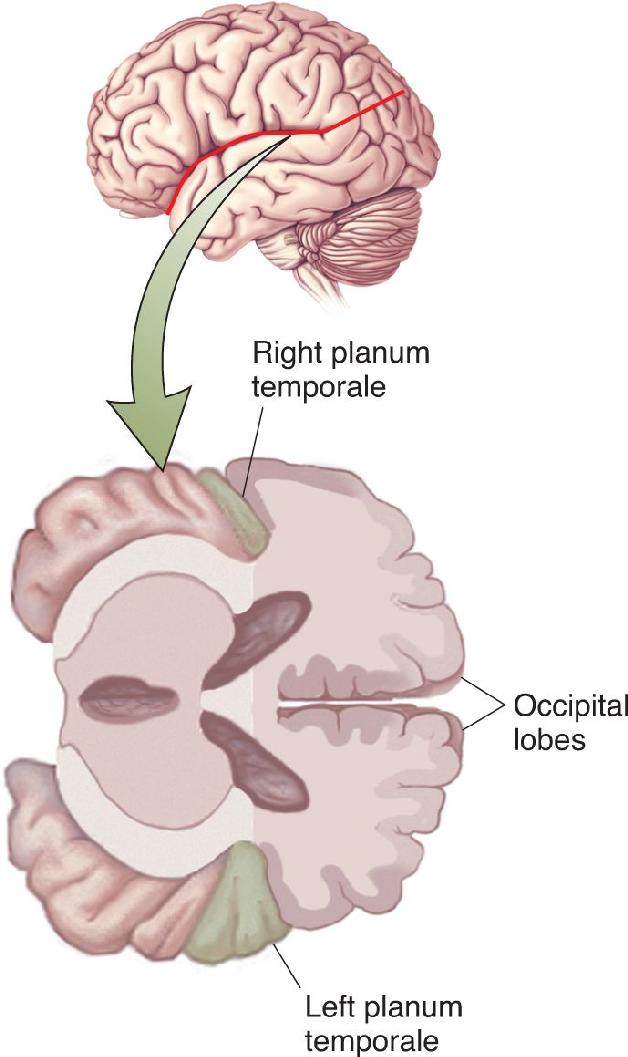
The most significant difference seen was in a region called the planum temporale, which is a part of Wernicke’s area on the superior surface of the temporal lobe (Figure 20.16). Based on measurements of 100 brains, Geschwind and Levitsky found that in about 65% of the brains, the left planum temporale was larger than the right, whereas in only about 10%, the right was larger. In some instances, the left area was more than five times larger than the right. Interestingly, the asymmetry in this area is seen even in the human fetus, suggesting that it is not a developmental consequence of the use of the left hemisphere for speech. Indeed, apes also have a larger left planum temporale. This suggests the possibility that speech became dominant in the left hemisphere because of a preexisting size difference. Other studies found that a portion of Broca’s area also tends to be larger in the left hemisphere. Are these larger areas in the left hemisphere related to the common dominance of the left hemisphere for speech?

FIGURE 20.16 Asymmetry of the planum temporale. This region on the superior temporal lobe is usually significantly larger in the left hemisphere. (Source: Adapted from Geschwind and Levitsky, 1968, Fig. 1.)
More recent investigations of gray matter volume have been made using MRI in living subjects, making it possible to test correlations among brain anatomy, asymmetries, and language dominance. One challenge in these studies is finding enough people who are right hemisphere dominant for language. Several language areas, including the planum temporale, Broca’s area, and the insula, are generally larger in the left hemisphere than the right, and this is true in people with a left or right hemisphere dominant for language. The big question is whether the hemisphere dominant for language can be predicted from the degree to which the structure on the left is larger than on the right. Perhaps some structure on the left side is much bigger in a person with a dominant left hemisphere but only a little bigger or smaller in a person with a dominant right hemisphere.
There have been mixed reports about a correlation between the size of the left and right planum temporale and the hemisphere dominant for language. There have also been reports about a correlation between the language-dominant hemisphere and the relative size of Broca’s area on each side of the brain. At present, it appears there may be some correlation between the asymmetric sizes of Broca’s area and the planum temporale with the hemisphere dominant for language, but the correlation is not strong enough to allow one to predict the language-dominant hemisphere from anatomical measurements alone. The brain region that presently appears to best predict which hemisphere is dominant for language is the insula, which is the cerebral cortex within the lateral sulcus that is between the temporal and parietal lobes (Figure 20.17). Even though the insula has been thought to be involved in human language for some time, the relationship between its size and language lateralization is somewhat surprising, as the language functions of the insula have been studied less and are less well understood than other language areas in the brain. Also, the insula appears to be involved in numerous brain functions ranging from taste to emotion. Further research is needed to clarify its role in language and its relationship with hemispheric dominance.

FIGURE 20.17 The insula. The insula, also called insular cortex, lies within the lateral sulcus between the temporal and parietal lobes.
It has probably occurred to you that a functional human asymmetry more obvious than language is handedness. More than 90% of humans are right-handed and usually relatively uncoordinated with their left hand, implying that in some way the left hemisphere is specialized for fine motor control. Is this related to the left hemispheric dominance for language? The answer is not known, but it is interesting that humans are different from nonhuman primates in handedness as well as language. While animals of many species show a consistent preference for using one hand, there are typically equal numbers of left-handers and right-handers.
LANGUAGE STUDIES USING BRAIN STIMULATION AND HUMAN BRAIN IMAGING
Until late in the twentieth century, language processing in the brain was examined mainly by correlating language deficits with postmortem analysis of brain damage. But now, aspects of language processing have been revealed by electrical brain stimulation and brain imaging with fMRI and PET in living humans.
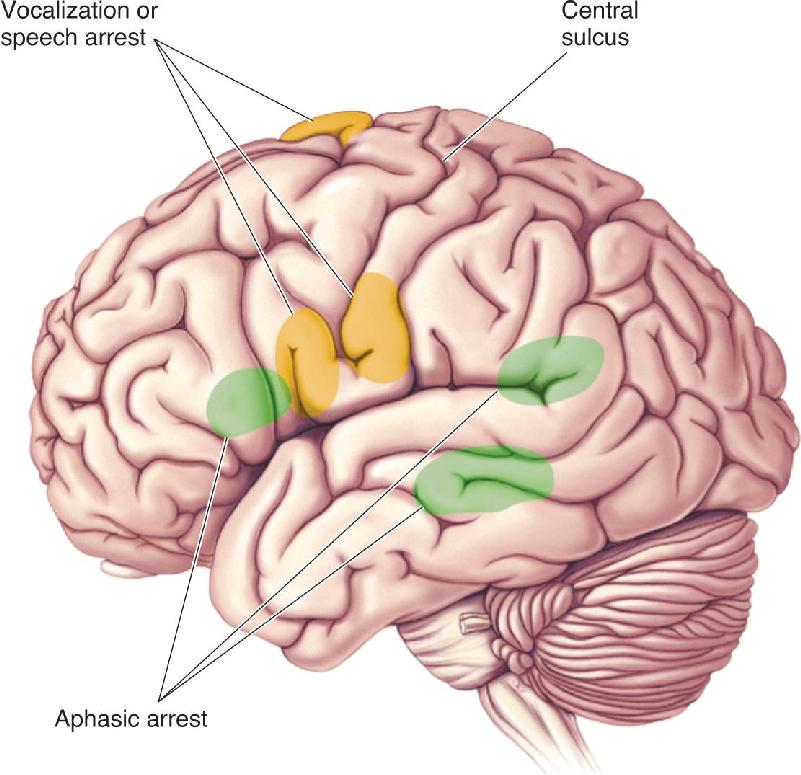
At several points in this book, we have discussed the electrical brain stimulation studies of Wilder Penfield. Without general anesthesia, patients could report the effects of stimulation at different cortical sites. In these experiments, Penfield noted that stimulation at certain locations affected speech. These effects occurred in three main categories: vocalizations, speech arrest, and speech difficulties similar to aphasia.
Stimulation of motor cortex in the area that controls the mouth and lips caused immediate speech arrest (Figure 20.18). Such a response is logical because the activated muscles sometimes pulled the mouth to one side or clenched the jaw shut. Stimulation of motor cortex occasionally evoked cries or rhythmic vocalizations; importantly, these effects occurred with electrical stimulation of motor cortex on either side of the brain. Penfield found three other areas where electrical stimulation interfered with speech, but these were only in the dominant left hemisphere. One of these areas appeared to correspond to Broca’s area. If this area was stimulated while a person was speaking, speech either stopped entirely (with strong stimulation) or became hesitant (with weaker stimulation). Some patients were unable to name objects they could name before and after the brain stimulation. Occasionally, they substituted an incorrect word. They apparently experienced a mild transient form of anomia. In some subjects, word confusion and speech arrest also resulted from stimulation at two other sites, one in the posterior parietal lobe near the Sylvian fissure and the other in the temporal lobe. These two areas were in the vicinity of the arcuate fasciculus and Wernicke’s area, although not perfectly aligned with estimated locations of those areas.

FIGURE 20.18 Sites where electrical brain stimulation affects language. Stimulation of motor cortex causes vocalizations or speech arrest by activating facial muscles. At other sites, stimulation causes an aphasic arrest in which language is agrammatical or anomia is observed. (Source: Adapted from Penfield and Rasmussen, 1950, Fig. 56.)
It is reassuring to find that electrical stimulation selectively affects speech in the brain areas roughly corresponding to those responsible for aphasia. However, the results of stimulation vary surprisingly between nearby cortical sites and between subjects. In studies similar to those of Penfield, neurosurgeon George Ojemann, at the University of Washington, found that the effects of stimulation are sometimes quite specific. For example, stimulation of small parts of cortex at different locations can interfere with naming, reading, or repetition of facial movements (Figure 20.19). The data support several important conclusions. First, across different people there is clearly considerable variability in the brain areas at which electrical stimulation affects language. Second, between patches of cortex involved in different aspects of language are other areas not affected by stimulation. We don’t know if further testing would have revealed that these in-between patches serve some untested role in language or whether these areas are actually not involved in language. Third, electrical stimulation at nearby sites can evoke quite different effects, and conversely, stimulation at distant sites can have the same effect. These findings suggest that language areas in the brain are much more complex than is implied by the Wernicke–Geschwind model. Areas involved in language are also more extensive than simply Broca’s and Wernicke’s areas, as they have been found to include other cortical areas as well as parts of the thalamus and striatum. Within Broca’s area and Wernicke’s area there may be specialized regions, possibly on the scale of functional columns in somatosensory cortex or ocular dominance columns in visual cortex. It appears that the large language areas identified on the basis of aphasic syndromes may well encompass a good deal of finer structure.

FIGURE 20.19 The effects of brain stimulation in three patients being treated for epilepsy. Patients were awake, and difficulties in speaking or reading were noted. N, naming difficulty with intact speech (anomia); A, arrest of speech; G, grammatical errors; J, jargon (fluent speech with frequent errors); R, failure to read; M, facial movement errors. (Source: Adapted from Ojemann and Mateer, 1979, Fig. 1.) Description
With the advent of modern imaging techniques, it has become possible to observe language processing in the human brain. With positron emission tomography (PET) and functional magnetic resonance imaging (fMRI), the level of neural activity in different parts of the brain is inferred from regional blood flow (see Box 7.3). In many ways, brain imaging confirms what was already known about language areas in the brain. For example, various language tasks make different portions of cerebral cortex active, and the activated areas are generally consistent with areas implicated by studies of aphasia.
However, brain imaging suggests that language processing is more complex. In an experiment conducted by Lehericy and associates, brain activity was recorded while subjects performed three different language tasks (Figure 20.20). In the first task, subjects were instructed to generate as many words as possible in a particular category such as fruits or animals (see Figure 20.20a). In the second task, subjects simply listened to a story being read aloud (see Figure 20.20b). The third task required that they silently repeat to themselves a sentence they previously heard read aloud. (see Figure 20.20c). Note that the location of activated brain areas is roughly consistent with the temporal and parietal language areas discovered by examining aphasia resulting from brain lesions. More surprising is the extent to which the brain activation is bilateral. Based on a Wada procedure for language lateralization, the subject shown in Figure 20.20 had a strongly dominant left hemisphere. The fMRI results suggest that there is more going on in the nondominant hemisphere than the Wada procedure suggests. Significant bilateral activation is commonly observed in fMRI studies, with ongoing debate about its significance. Recent PET and fMRI studies also suggest fascinating similarities and differences in language processing of spoken language, sign language, and Braille (Box 20.4).

FIGURE 20.20 Bilateral brain activation shown by fMRI. Based on a Wada procedure, the subject illustrated here had a strongly dominant left hemisphere for language. fMRI shows significant bilateral activation of language areas on a (a) word generation task, (b) passive story listening, and (c) silent sentence repetition. (Source: Adapted from Lehericy et al., 2000, Fig 1.)
The human brain is a remarkably adaptable organ, and some of the most dramatic examples of brain reorganization come from studies of human language processing. The top panel of Figure A, from an fMRI study, shows brain areas activated when normally hearing English speakers read sentences in English. Areas in red are those most highly specifically activated by language, and areas in yellow are somewhat less activated. (Visual activity not associated with language has been subtracted out.) There is significant activity in the standard language areas of the left hemisphere, including Broca’s area and Wernicke’s area, but little activity in the right hemisphere.

Figure A Top: Written English, read by hearing subjects. Center: ASL, seen by English-speaking hearing subjects. Bottom: ASL, seen by deaf subjects. (Source: Neville et al., 1998.)
The center panel of Figure A is the activity seen when hearing English speakers saw sentences in American Sign Language (ASL). These people who did not understand ASL showed no specific activity after their response to meaningless hand signs was subtracted. On the other hand, the bottom panel of Figure A shows the response to ASL in deaf subjects who were raised with it as their sole language. The activation includes Broca’s area and Wernicke’s area in the left hemisphere, indicating that ASL activates the same key language areas activated by spoken English in hearing subjects. Even more surprising is the great deal of activity in the right hemisphere. Remarkably, the superior temporal gyrus is active in response to ASL in deaf subjects, an area that would normally respond to spoken language in hearing subjects. As these areas are also active in hearing subjects who are bilingual in ASL, it appears that something about the ASL language recruits right hemisphere auditory areas in addition to the usual left hemisphere language areas.
A different form of brain reorganization is seen in blind subjects who read Braille. A system of writing that uses patterns of small bumps on paper to represent letters, Braille is read by scanning the fingertips across the bumps. As you would expect, Braille reading activates the somatosensory cortex, but other brain areas also activated are a real surprise. Figure B shows a PET image obtained from a person reading Braille. There is significant activity at the occipital pole of the brain (yellow)—unmistakably portions of visual cortex. Through a process of brain reorganization, these blind subjects have a brain that uses traditional visual areas to process Braille, similar in spirit to the auditory areas used for ASL in deaf subjects. (The chapters in Part IV explore the mechanisms by which sensory experience can affect brain organization and lead to learning and memory.)
Figure B Reading Braille. (Source: Sadato et al., 1996).
In another study, researchers used PET imaging to observe differences in brain activity between the sensory responses to words and the production of speech. They began by measuring cerebral blood flow with the subject at rest. They then had the person either look at words presented on a computer display (“seeing words”) or listen to words being read aloud (“hearing words”). By subtracting the levels of blood flow at rest from the levels during hearing or seeing, blood flow levels were obtained specifically corresponding to the activity evoked by the sensory input. The results are shown in the top half of Figure 20.21. Not surprisingly, the visual stimuli evoked increased brain activity in striate cortex and extrastriate cortex, and the auditory stimuli elicited activity in primary and secondary auditory cortex. However, the areas activated in extrastriate cortex and secondary auditory cortex did not respond to visual and auditory stimuli that were not words. These areas may be specialized for the coding of words either seen or heard. The visual stimuli did not evoke noticeably increased activity in the area of the angular gyrus and Wernicke’s area, as one would expect based on the Wernicke–Geschwind model.

FIGURE 20.21 PET imaging of sensation and speech. Relative levels of cerebral blood flow are color coded. Red indicates the highest levels, and progressively lower levels are represented by orange, yellow, green, and blue. (Source: Posner and Raichle, 1994, p. 115.)
Another task studied with PET imaging was repeating words. To know what words to repeat, subjects must perceive and process the words by either the visual or the auditory system. Thus, the brain activity seen in the repetition task should include a component associated with the basic perceptual process as well as a component associated with speech. To isolate the speech component, the response pattern previously obtained in the simple sensory task was subtracted out. In other words, the image shown for “speaking words” equals an image corresponding to “repeating spoken words” minus an image corresponding to “listening to words.” After the subtraction, the blood flow pattern indicated high levels of activity in primary motor cortex and the supplementary motor area (Figure 20.21, bottom left). There was also increased blood flow around the Sylvian fissure near Broca’s area. However, the PET images showed such activity bilaterally, and it was even observed when the subjects were instructed to move their mouth and tongue without speaking. Since there is good evidence that Broca’s area is unilateral, it may not be showing up in these images, for reasons unknown.
The final task required the subjects to think a little. For each word presented, the subject had to state a use for the word (e.g., if “cake” was presented, the subject said “eat”). To isolate activity specific to this noun–verb association task (“generating words”), the blood flow pattern obtained previously for speaking words was subtracted. Areas that were activated in the association task are located in the left inferior frontal area, the anterior cingulate gyrus, and the posterior temporal lobe (see Figure 20.21, bottom right). The activity in frontal and temporal cortex is believed to be related to the performance of the word-association task, whereas the activity in cingulate cortex may be related to attention.
There is also evidence from numerous PET and fMRI studies that distributed and distinct brain areas store information about different categories of objects. These reports appear consistent with observations showing that brain damage sometimes leads to category-specific losses in the ability to name objects. For example, with a brain lesion a person may be able to name tools and some living things such as fruits and vegetables but show a large deficit in the ability to name animals. One such patient called a giraffe a kangaroo and a goat a chicken. In a PET imaging experiment, different regions of the temporal lobe were more active when a person was asked to name people, animals, and tools. In other studies, overlapping but different brain activity patterns were found for concrete nouns (e.g., “door”), abstract nouns (e.g., “despair”), concrete verbs (e.g., “speak”), and abstract verbs (e.g., “suffer”). The findings in these experiments raise many questions for further research to answer. How might the brain process different categories of words differently and integrate the results in a unified understanding? What is the distinction between brain areas involved in recognition of sensory input and areas that assign names or meaning to objects perceived?
Language was one of the most important steps in human evolution. Communication between people is such a fundamental aspect of being human that it is difficult to imagine life without language. Current estimates are that the human capacity for language evolved relatively recently, around 100,000 years ago. While animals use a great diversity of sounds and behaviors to communicate, none of these come close to the elaborate and flexible system of language and speech used by humans. Aspects of language acquisition and use have been fruitfully studied in songbirds and nonhuman primates, but unlike other brain systems, the study of human language requires experiments and observations in humans. For this reason, experimental approaches are largely limited to behavioral studies of language acquisition and function, the consequences of lesions, effects of brain stimulation, and brain imaging with fMRI and PET. Even with these few techniques, however, a great deal has been learned. Consistent with the locations of sensory and motor areas in the brain, the basics of language organization can be understood. Early studies focused attention on Broca’s area, which is near the motor cortex and associated with speech production, and Wernicke’s area, which is near the auditory cortex and associated with speech comprehension. These observations are still of clinical use today.
More recent research has shown that language processing is far more complex and engages far more of the brain, than implied by the Wernicke–Geschwind model. Brain imaging and stimulation studies have revealed widespread brain areas in both hemispheres that are involved in language and that vary from one person to the next. From our perspective today, the complexity of language and its extensive representation in the brain are not so surprising because language involves many different components, such as understanding the basis of words in sound, the meanings of words, the grammar used to combine words into meaningful statements, naming objects, producing speech, and on and on. As in research on other brain systems for sensation, motor output, emotion, and so on, we are interested in the extent to which language processing involves a collection of interacting subsystems for different language skills. There is clearly still much to be learned. Further brain imaging studies will hopefully clarify the organization of language systems in the brain at a finer scale than was possible from studying the consequences of brain lesions and perhaps identify distinct circuits that serve different functions.
The Discovery of Specialized Language Areas in the Brain
Asymmetrical Language Processing in the Two Cerebral Hemispheres
1. How is it possible for a split-brain human to speak intelligibly if the left hemisphere controls speech? Isn’t this inconsistent with the fact that the left hemisphere must direct motor cortex in both hemispheres to coordinate movements of the mouth?
2. What can you conclude about the normal function of Broca’s area from the observation that there are usually some comprehension deficits in Broca’s aphasia? Must Broca’s area itself be directly involved in comprehension?
3. Pigeons can be trained to press one button when they want food and to press other buttons when they see particular visual stimuli. This means the bird can “name” things it sees. How would you determine whether or not the pigeon is using a new language—“button-ese”?
4. What does the Wernicke–Geschwind language processing model explain? What data are inconsistent with this model?
5. In what ways is the left hemisphere usually language dominant? What does the right hemisphere contribute?
6. What evidence is there that Broca’s area is not simply a premotor area for speech?
Berwick RC, Friederici AD, Chomsky N, Bolhuis JJ. 2013. Evolution, brain, and the nature of language. Trends in Cognitive Sciences 17:89–98.
Bookeheimer S. 2002. Functional MRI of language: new approaches to understanding the cortical organization of semantic processing. Annual Review of Neuroscience 25:51–188.
Friederici, AD. 2012. The cortical language circuit: from auditory perception to sentence comprehension. Trends in Cognitive Sciences 16:262–268.
Graham SA, Fisher SE. 2013. Decoding the genetics of speech and language. Current Opinion in Neurobiology 23:43–51.
Kuhl PK. 2010. Brain mechanisms in early language acquisition. Neuron 67:713–727.
Saffran EM. 2000. Aphasia and the relationship of language and brain. Seminars in Neurology 20:409–418.
Scott SK, Johnsrude IS. 2002. The neuroanatomical and functional organization of speech perception. Trends in Neurosciences 26:100–107.
Vargha-Khadem F, Gadian DG, Copp A, Mishkin M. 2005. FOXP2 and the neuroanatomy of speech and language. Nature Reviews Neuroscience 6:131–138.
Additional figures