Memory Systems
BOX 24.1 OF SPECIAL INTEREST: Extraordinary Memory
Electrical Stimulation of the Human Temporal Lobes
Neural Recordings from the Human Medial Temporal Lobe
BOX 24.2 OF SPECIAL INTEREST: Korsakoff’s Syndrome and the Case of N.A.
BOX 24.3 PATH OF DISCOVERY: How the Brain Makes Maps, by Edvard and May-Britt Moser
Standard and Multiple Trace Models of Consolidation
BOX 24.4 OF SPECIAL INTEREST: Introducing False Memories and Erasing Bad Memories
The brain has numerous systems for performing functions related to sensation, action, and emotion, and each system contains billions of neurons with enormous numbers of interconnections. In Chapter 23, we explored the mechanisms that guide the construction of these systems during brain development. But as impressive and orderly as prenatal development is, no one would confuse a newborn baby and a Nobel Prize winner. Much of the difference between the two comes down to what has been learned and remembered. From the moment we take our first breath, and possibly before, the sensory stimuli we experience modify our brain and influence our behavior. We learn an enormous number of things, some straightforward (e.g., snow is cold), and others more abstract (e.g., an isosceles triangle has two sides of equal length). Some of the things we learn are easily stated facts, while others, such as driving or playing soccer, involve ingrained motor patterns. We will see that brain lesions differentially affect different types of remembered information, suggesting that there is more than one memory system.
There is a close relationship between what we called experience-dependent brain development in Chapter 23 and what we call learning in this chapter. Visual experience during infancy is essential for the normal development of the visual cortex, but it also allows us to recognize an image of our mother’s face. Visual development and learning probably involve similar mechanisms, but at different times and in different cortical areas. Understood in this way, learning and memory are lifelong adaptations of brain circuitry to the environment. They enable us to respond appropriately to situations we have experienced before.
In this chapter, we discuss the anatomy of memory—the different parts of the brain involved in storing particular types of information. Chapter 25 will then focus on the elementary molecular mechanisms that can store information in the brain.
Learning is the acquisition of new knowledge or skills. Memory is the retention of learned information. We learn and remember lots of different things, and it is important to appreciate that these various things might not be processed and stored by the same neural hardware. No single brain structure or cellular mechanism accounts for all learning. Moreover, the way in which information of a particular type is stored may change over time.
Psychologists have studied learning and memory extensively and have distinguished what appear to be different types. Useful for our purposes is the distinction between declarative memory and nondeclarative memory.
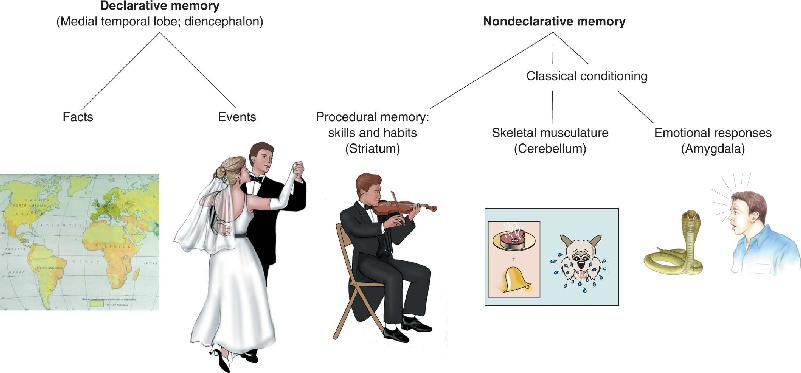
During the course of our lives, we learn many facts (e.g., the capital of Thailand is Bangkok, Darth Vader is Luke Skywalker’s father). We also store memories of life’s events (e.g., “Yesterday’s neuroscience exam was fun!” or “I went swimming with my pet dog named Axon when I was five years old.”) Memory of facts and events is called declarative memory (Figure 24.1). A declarative memory distinction we will examine later on is between episodic memory for autobiographical life experiences and semantic memory for facts. Declarative memory is what people usually mean in everyday uses of the word “memory,” but we actually remember many other things too. These nondeclarative memories fall into several categories. The type we are most concerned with here is procedural memory, or memory for skills, habits, and behaviors. We learn to play the piano, throw a Frisbee, or tie our shoes, and somewhere that is stored in our brain.
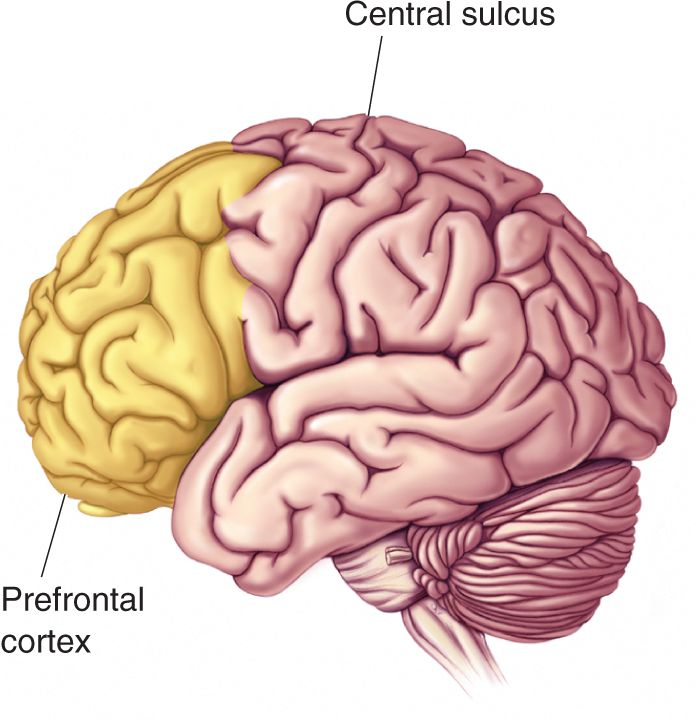

FIGURE 24.1 Types of declarative and nondeclarative memory. Brain structures thought to be involved in each type of memory are indicated. (Note that this is not a complete representation of all types of memory.) Description
Generally, declarative memories are accessed for conscious recollection, whereas the tasks we learn, as well as the reflexes and emotional associations we have formed, operate smoothly without conscious recollection. As the old saying goes, you never forget how to ride a bicycle. You may not explicitly remember the day you first rode a two-wheeler on your own (the declarative part of the memory), but your brain remembers what to do when you’re on one (the procedural part of the memory). Nondeclarative memory is also frequently called implicit memory, because it results from direct experience, and declarative memory is often called explicit memory, because it results from more conscious effort.
Another distinction is that declarative memories are often easy to form and are easily forgotten. In contrast, forming nondeclarative memories usually require repetition and practice over a longer period of time, but these memories are less likely to be forgotten. Think of the difference between memorizing the names of people you meet at a party (declarative) and learning to ski (nondeclarative). While there is no clear limit to the number of declarative memories the brain can store, there is great diversity in the ease and speed with which such new information is acquired. Studies of humans with abnormally good memories suggest that the limit on the storage of declarative information is remarkably high (Box 24.1).
Some people have astonishing memory abilities, and these cases suggest that human memory capacity may, in general, be incredibly large. For example, the British artist Stephen Wiltshire draws enormous cityscapes from memory; his most elaborate is a 10-meter-wide accurate drawing of Tokyo that he drew in 7 days after only a 30-minute helicopter ride over the city. In rare cases of hyperthymesia, also known as superior autobiographical memory, people have explicit memories for almost every day of their lives. The American actress Marylu Henner has this ability.
One of the oldest and best documented cases of an extraordinary memory was documented by the Russian psychologist Alexander Luria. In the 1920s, a man named Solomon Shereshevsky came to see Luria, beginning a 30-year study of the uncommon memory of this man Luria referred to simply as S. Luria published his fascinating description of this study in his book, The Mind of a Mnemonist. Luria initially studied S. by giving him conventional tests, such as memorizing lists of words, numbers, or nonsense syllables. He’d read the list once and then ask S. to repeat it. Much to Luria’s surprise, he couldn’t come up with a test that S. could not pass. Even when 70 words were read in a row, S. could repeat them forward, backward, and in any other order. During the many years they worked together, Luria never found a limit to S.’s memory. In tests of his retention, S. demonstrated that he remembered lists he had previously seen even 15 years earlier!
How did he do it? S. described several factors that may have contributed to his great memory. One was his unusual sensory response to stimuli; he retained vivid images of things he saw. When shown a table of 50 numbers, he claimed it was easy to later read off numbers in one row or along the diagonal because he simply called up a visual image of the entire table. Interestingly, when he occasionally made errors in recalling tables of numbers written on a chalkboard, they appeared to be “reading” errors rather than memory errors. For instance, if the handwriting was sloppy, he would mistake a 3 for an 8 or a 4 for a 9. It was as if he were seeing the chalkboard and numbers all over again when he was recalling the information.
Another interesting aspect of S.’s sensory response to stimuli was a powerful form of synesthesia. Synesthesia is a phenomenon in which sensory stimuli evoke sensations usually associated with stimuli of a different sense or different stimuli in the same sense. For example, when S. heard a sound, in addition to hearing, he would see splashes of colored light and perhaps have a certain taste in his mouth. The multimodal response to sensory input may have caused the brain to form particularly strong memory traces.
After learning that his memory was unusual, S. left his job as a reporter and became a professional stage performer—a mnemonist. In order to remember huge lists of numbers or tables of words given by members of the audience trying to stump him, he complemented his lasting sensory responses to stimuli and his synesthesia with memory “tricks.” To remember a long list of items, he made use of the fact that each item evoked some sort of visual image. As the list was read or written, S. imagined himself walking through his home town; as each item was given, he placed its evoked image along his walk—the image evoked by item 1 by the mailbox, the image for item 2 by a bush, and so on. To later recall the items, he walked the same route and picked up the imaginary items he had put down. Though we may not have the complex synesthetic sensations of S., this ancient technique of making associations with familiar objects is one we all can use.
But not everything about S.’s memory was to his advantage. While the complex sensations evoked by stimuli helped him remember lists of numbers and words, they interfered with his ability to integrate and remember more complex things. He had trouble recognizing faces because each time a person’s expression changed, he would also “see” changing patterns of light and shade, which would confuse him. He also wasn’t very good at following a story read to him. Rather than ignoring the exact words and focusing on the important ideas, S. was overwhelmed by an explosion of sensory experiences. Imagine how bewildering it would be to be bombarded by constant visual images evoked by each word, plus sounds and images evoked by the tone of voice of the person reading the story.
S. also experienced the inability to forget. This became a particular problem when performing as a professional mnemonist and he was asked to remember things written on a chalkboard. He would see things that had been written there on many different occasions. Although he tried various tricks to try to forget old information, such as mentally erasing the board, nothing worked. Only by the strength of his attention, and by actively telling himself to let information slip away, was he able to forget. It was as if the effort most of us use for remembering and the ease with which we forget were reversed for S.
We don’t know the neural basis for S.’s remarkable memory. Perhaps he lacked the segregation most of us have between sensations in different sensory systems. This may have contributed to an uncommonly strong multimodal coding of memories. Maybe his synapses were more malleable than normal. Unfortunately, we’ll never know.
The type of nondeclarative memory we will focus on is procedural memory, which involves learning a motor response (procedure) in reaction to a sensory input. The formation of procedural memories occurs through two categories of learning: nonassociative learning and associative learning.
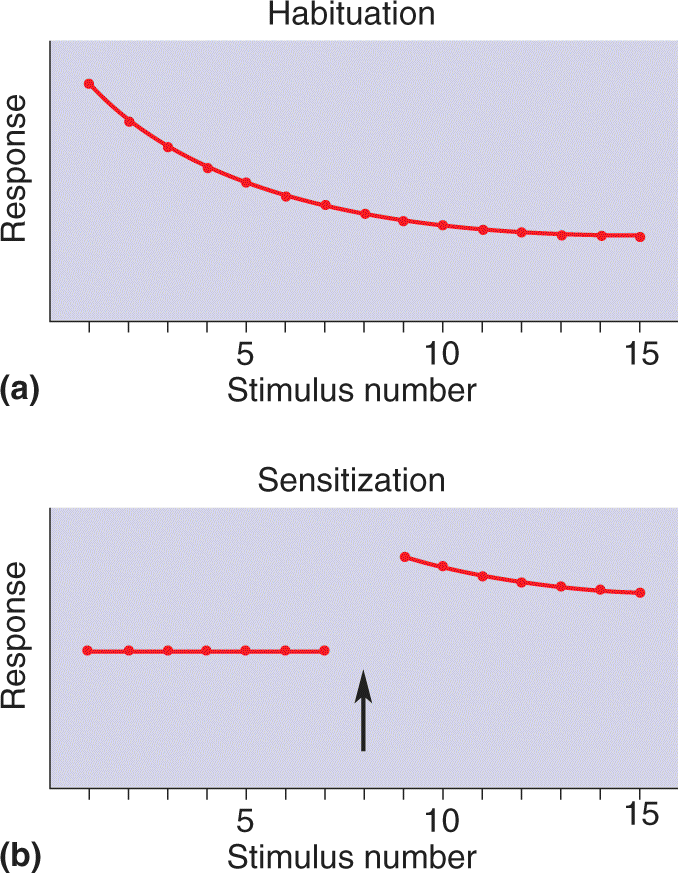
Nonassociative Learning. Nonassociative learning describes a change in behavioral response that occurs over time in response to a single type of stimulus. There are two types: habituation and sensitization.
Suppose you live in a house with a single telephone. When the phone rings, you run to answer it, but every time the call is for someone else. Over time, you stop reacting to the ringing of the phone and eventually no longer even notice it. This type of learning, habituation, is learning to ignore a stimulus that lacks meaning (Figure 24.2a). You are habituated to a lot of stimuli. Perhaps as you read this sentence, cars and trucks are passing by outside, a dog is barking, your roommate is playing the same tune for the hundredth time—and all this goes on without your really noticing. You have habituated to these stimuli.

FIGURE 24.2 Types of nonassociative learning. (a) In habituation, repeated presentation of the same stimulus produces a progressively smaller response. (b) In sensitization, a strong stimulus (arrow) results in an exaggerated response to all subsequent stimuli. Description
Now suppose you’re walking down the sidewalk on a well-lit city street at night, and suddenly there is a blackout. You hear footsteps behind you, and though this wouldn’t normally disturb you, now you nearly jump out of your skin. Car headlights appear, and you react by side-stepping away from the street. The strong sensory stimulus (the blackout) caused sensitization, a form of learning that intensifies your response to all stimuli, even ones that previously evoked little or no reaction (Figure 24.2b).
Associative Learning. In associative learning behavior is altered by the formation of associations between events; this is in contrast to a changed response to a single stimulus in nonassociative learning. Two types of associative learning are usually distinguished: classical conditioning and instrumental conditioning.
Classical conditioning was discovered and characterized in dogs by the famous Russian physiologist Ivan Pavlov around the turn of the nineteenth century. Classical conditioning involves associating a stimulus that evokes a measurable response with a second stimulus that normally does not evoke this response. The first type of stimulus, the one that normally evokes the response, is called the unconditional stimulus (US) because no training (conditioning) is required for it to yield a response. In Pavlov’s experiments, the US was the sight of a piece of meat, and the dog’s response was salivation. The second type of stimulus, the one that normally does not evoke this same response, is called the conditional stimulus (CS) because this one requires training (conditioning) before it will yield this response. In Pavlov’s experiments, the CS was an auditory stimulus, such as the sound of a bell. Training consisted of repeatedly pairing the presentation of the meat with the sound of the bell (Figure 24.3a). After many of these pairings the meat was withheld, and the animal salivated to the sound alone. The dog had learned an association between the sound (CS) and the presentation of meat (US) (Figure 24.3b). The learned response to the conditioned stimulus is called the conditioned response (CR).

FIGURE 24.3 Classical conditioning. (a) Before conditioning, the sound of a bell (the conditional stimulus, CS) elicits no response, in sharp contrast to the response elicited by the sight of a piece of meat (the unconditional stimulus, US). (b) Conditioning entails pairing the sound of the bell with the sight of the meat. The dog learns to associate the sound of the bell with the meat and after conditioning will salivate when the bell rings without the meat. Description
Instrumental conditioning was discovered and studied by Columbia University psychologist Edward Thorndike early in the last century. In instrumental conditioning, an individual learns to associate a response, a motor act, with a meaningful stimulus, typically a reward such as food. For example, consider what happens when a hungry rat is placed in a box with a lever that dispenses food. In the course of exploring the box, the rat bumps the lever and out pops a piece of food. After this happy accident occurs a few more times, the rat learns that pressing the lever leads to a food reward. The rat will then work the lever (and eat the food) until it is no longer hungry. As in classical conditioning, a predictive relationship is learned during instrumental conditioning. In classical conditioning, the subject learns that one stimulus (CS) predicts another stimulus (US). In instrumental conditioning, the subject learns that a particular behavior is associated with a particular consequence. Because motivation plays such a large part in instrumental conditioning (after all, only a hungry rat will lever-press for a food reward), the underlying neural circuits are considerably more complex than those involved in simple classical conditioning.
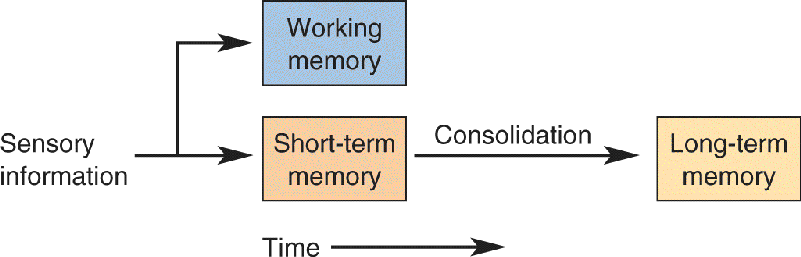
From daily experience we know that some memories last longer than others. Long-term memories are those that you can recall days, months, or years after they were originally stored. The information that makes it into long-term memory, of course, represents only a fraction of what we experience every day. Most information is held by the brain only temporarily, on the order of hours. These short-term memories have in common the property that they are vulnerable to disruption. For example, short-term memory can be erased by head trauma or electroconvulsive therapy (ECT) used to treat psychiatric illness. But the same trauma and ECT do not affect long-term memories, which were stored long ago (e.g., childhood memories). These observations have led to the idea that facts and events are stored in short-term memory and a subset are converted into long-term memories via a process called memory consolidation (Figure 24.4).

FIGURE 24.4 Memory Consolidation. Sensory information can be temporarily stored in short-term memory that is susceptible to disruption. Stable long-term memories are formed by consolidation. Another type of memory, working memory, is used to hold information “in mind.” Description
A second, entirely distinct form of temporary storage, lasting on the order of seconds, is working memory. Unlike the short-term memory discussed above, working memories are sharply limited in capacity and require rehearsal. It is often said that working memory is information held “in mind.” When someone tells you his or her phone number, you can retain it for a limited period of time by repeating the number to yourself. Keeping a memory alive through repetition is a hallmark of working memory. If the number is too long (e.g., a phone number with extra numbers for a foreign country), you may have trouble remembering the number at all. Eventually the number may be consolidated into a long-term memory. Working memory is commonly studied by measuring a person’s digit span, the maximum number of randomly chosen numbers a person can repeat back after hearing a list read. The normal digit span is seven plus or minus two. Working memory is distinguished from short-term memory by the very limited capacity, the need for repetition, and the very short duration.
Interestingly, there are reports of humans with cortical lesions who have normal memory for information coming from one sensory system (e.g., they can remember the same number of visually seen numbers as other people) but a profound deficit when information comes from another sensory system (e.g., they cannot remember more than one number spoken to them). These different digit spans in different modalities are consistent with the notion of multiple temporary storage areas in the brain.
As we all know, in daily life, forgetting happens nearly as often as learning. Less commonly, certain diseases and injuries to the brain cause a serious loss of memory and/or the ability to learn called amnesia. Concussion, chronic alcoholism, encephalitis, brain tumor, and stroke can all disrupt memory. You’ve probably seen a movie or television show in which a person experiences some trauma and wakes up the next day not knowing who he or she is and not remembering the past. That kind of absolute amnesia for past events and information is actually extremely rare. It is more common for trauma to cause limited amnesia along with other nonmemory deficits. If amnesia is not accompanied by any other cognitive deficit, it is known as dissociated amnesia (i.e., the memory problems are dissociated from any other problems). We will focus on cases of dissociated amnesia because a clear relationship can be drawn between memory deficits and brain injury.
Following trauma to the brain, two different types of memory loss may occur: retrograde amnesia and anterograde amnesia (Figure 24.5). Retrograde amnesia is characterized by memory loss for events prior to the trauma; you forget things you already knew. In severe cases, there might be complete amnesia for all declarative information learned before the trauma. More often, retrograde amnesia follows a pattern in which events of the months or years preceding the trauma are forgotten, but memory is increasingly strong for older memories. The graded loss of old memories across time apparently reflects the changing nature of memory storage, a topic we explore in Chapter 25. Anterograde amnesia is an inability to form new memories following brain trauma. If the anterograde amnesia is severe, a person might be completely incapable of learning and remembering anything new. In milder cases, learning may be slower and require more repetition than normal. In clinical cases, there is often a mixture of retrograde and anterograde amnesias of different degrees of severity.

FIGURE 24.5 Amnesia produced by trauma to the brain. (a) In retrograde amnesia, events for a period of time prior to the trauma are forgotten, but memories from the distant past and the period following the trauma are intact. (b) In anterograde amnesia, events prior to the trauma can be remembered, but there are no memories for the period following the trauma. Description
An example will help clarify. Suppose that on the last day of freshman year in college you are walking past a friend’s dormitory. In a fit of excitement about the end of the semester, your friend throws her books out the window, crashing on your head. If this trauma causes you to have retrograde amnesia, you may be unable to remember the final exam you took the day before or, in a more serious case, any of the courses you took freshman year. If you experience anterograde amnesia, you may recall the exams you took before the accident, but when you graduate from college you might be unable to recall the ambulance ride to the hospital after the accident, your friend’s endless apologies, or even the summer you spent recovering after freshman year.
A form of amnesia that involves a much shorter period of time is called transient global amnesia. This is a sudden onset of anterograde amnesia that lasts for only a period of minutes to days, often accompanied by retrograde amnesia for recent events preceding the attack. During the spell, the person may appear disoriented and ask the same questions repeatedly, but he or she is conscious, and measures of working memory, such as digit span, are normal. In a matter of hours the attack usually subsides, and the person is left with a permanent memory gap.
Transient global amnesia can be frightening to both the person experiencing it and those witnessing it. Although the cause has not been clearly established, brief cerebral ischemia, in which the blood supply to the brain is temporarily reduced, or concussion to the head from trauma, such as a car accident or a hard blow while playing football, might be implicated. There have been reports of transient global amnesia brought on by seizures, physical stress, drugs, cold showers, and even sex, presumably because all of these affect cerebral blood flow. Many cases were linked to use of the antidiarrheal drug clioquinol (which was taken off the market). While we don’t know exactly what causes transient global amnesia, it may be a consequence of temporary blood deprivation to structures essential for learning and memory. Other forms of temporary amnesia can be caused by disease, brain trauma, and environmental toxins.
Our brains acquire all kinds of information through our sensory systems, but as discussed in Chapter 21, we pay attention to only a fraction of it. To serve immediate behavioral needs, some of this sensory information is “held in mind” by working memory, such as a phone number that we must remember in order to call. Unlike long-term memory, working memory has a very small capacity, as shown by the digit span described earlier. However, there are subtleties to the quantification of working memory capacity. For example, more words can be held in memory if they are short common words. Also, more words and numbers can be held in working memory if they can be chunked into meaningful groups (e.g., a 12-digit number is easily held when chunked into three years, such as 1945 1969 2001). Working memory can be thought of as a limited resource that can be used in a variety of ways; there are tradeoffs in the amount and precision of stored information that are influenced by the behavioral significance of the information.
Information held in working memory might be converted into long-term memories, but most of it is discarded when no longer needed. How is information retained in the brain by working memory long enough to be useful? Research in both animals and humans suggests that, rather than a single system, working memory is a capability of neocortex found in numerous locations in the brain. To illustrate, we look at a couple of examples of working memory in the frontal and parietal cortex.
One of the most obvious anatomical differences between primates (especially humans) and other mammals is that primates have a large frontal lobe. The rostral end of the frontal lobe, the prefrontal cortex, is particularly highly developed (Figure 24.6). Compared to the sensory and motor cortical areas, the function of the prefrontal cortex is relatively poorly understood. But because it is so well developed in humans, the prefrontal cortex is often assumed to be involved in those characteristics that distinguish us from other animals, such as self-awareness and the capacity for complex planning and problem solving.

FIGURE 24.6 Prefrontal cortex. The brain rostral to the central sulcus is the frontal lobe. The prefrontal cortex is the anterior portion of the frontal lobe that receives afferents from the medial dorsal nucleus of the thalamus.
Some of the first evidence suggesting that the frontal lobe is important for learning and memory came from experiments performed in the 1930s using a delayed-response task. A monkey was first shown food being placed in a well below one of two identical covers in a table. A delay period followed, during which the animal could not see the table. Finally, the animal was allowed to see the table again and received the food as a reward if it chose the correct well. Large prefrontal lesions seriously degraded performance in this delayed-response task, as well as other tasks including a delay period. Moreover, the monkeys performed increasingly poorly as the delay period was lengthened. These results imply that the prefrontal cortex may normally be involved in retaining information in working memory.
Experiments conducted more recently suggest that the prefrontal cortex is involved with working memory for problem solving and the planning of behavior. One piece of evidence comes from the behavior of humans with lesions in the prefrontal cortex. Recall the case of Phineas Gage, discussed in Chapter 18. Having sustained severe frontal lobe damage by an iron bar passing through the head, Gage had a difficult time maintaining a course of behavior. Although he could carry out behaviors appropriate for different situations, he had difficulty planning and organizing these behaviors, perhaps because of the damage to his frontal lobe.
The Wisconsin card-sorting test can demonstrate problems associated with prefrontal cortical damage. A person is asked to sort a deck of cards with a variable number of colored geometric shapes (Figure 24.7). The cards can be sorted by color, shape, or number of symbols, but at the beginning of the test, the subject isn’t told which category to use. The subject begins putting cards into stacks and is informed when errors occur, by which the subject learns what sorting category is to be used. Then, after ten correct card placements have been made, the sorting category is changed, and the subject starts over again. To perform well on this test, a person must use memory of previous cards and errors in order to plan the next card placement. People with prefrontal lesions have great difficulty on this task when the sorting category is changed; they continue to sort according to a rule that no longer applies. It appears that they have a working memory deficit that limits their ability to make use of recent information to change their behavior.

FIGURE 24.7 The Wisconsin card-sorting test. Cards containing various numbers of colored symbols must first be sorted by color. After a string of correct responses is made, the sorting category is changed to shape.
The same kind of deficit is seen in other tasks. For example, a person with a prefrontal lesion might be asked to trace a path through a maze drawn on a piece of paper. Although the patient understands the task, he or she will repeatedly make the same mistakes, returning to blind alleys. In other words, these patients do not learn from their recent experience in the same way as a normal person, suggesting a working memory deficit.
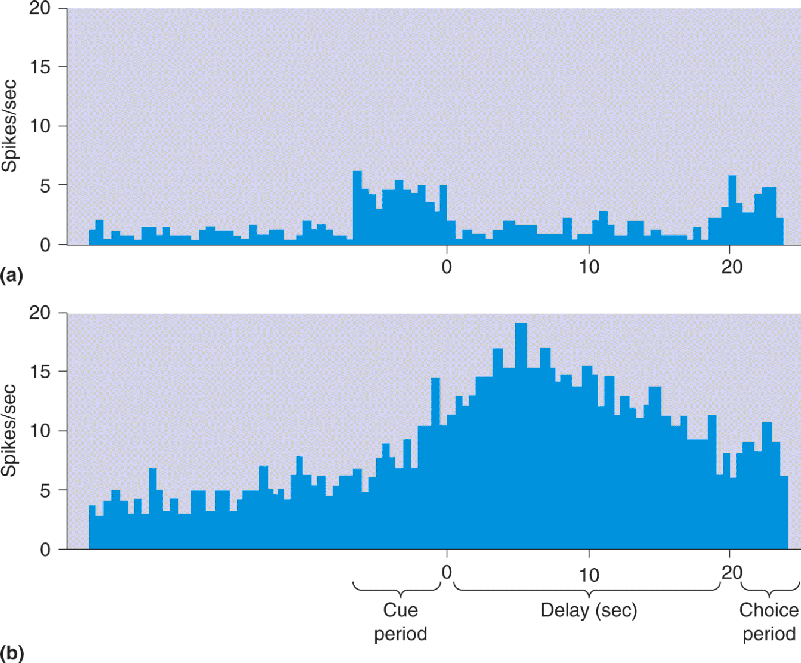
The neurons in prefrontal cortex have a variety of response types, some of which may reflect a role in working memory. Figure 24.8 shows two response patterns obtained while a monkey performed a delayed-response task. The neuron in the top trace responded while the animal first saw the food wells, was unresponsive during the delay interval, and responded again when the animal saw the food wells again (Figure 24.8a). The response of the neuron simply correlates with visual stimulation. More interesting is the response pattern of the other neuron, which fired only during the delay interval (Figure 24.8b). This cell was not directly activated by the stimuli in the first or second interval in which the monkey saw the food wells. The increased activity during the delay period may be related to the retention of information needed to make the correct choice after the delay (i.e., working memory).

FIGURE 24.8 Working memory activity in monkey prefrontal cortex. The two histograms show the activity of cells in prefrontal cortex recorded while the animal performed a delayed-response task. During a cue period of 7 seconds, food is placed into one of two wells within the monkey’s view. During the delay period, the animal cannot see the food wells; after the delay, it is allowed to choose a well to receive a food reward (the choice period). (a) This cell responds when the animal first sees the food wells and when it sees them again after the delay period. (b) This cell responds strongest during the delay period, when there is no visual stimulus. (Source: Adapted from Fuster, 1973, Fig. 2.)
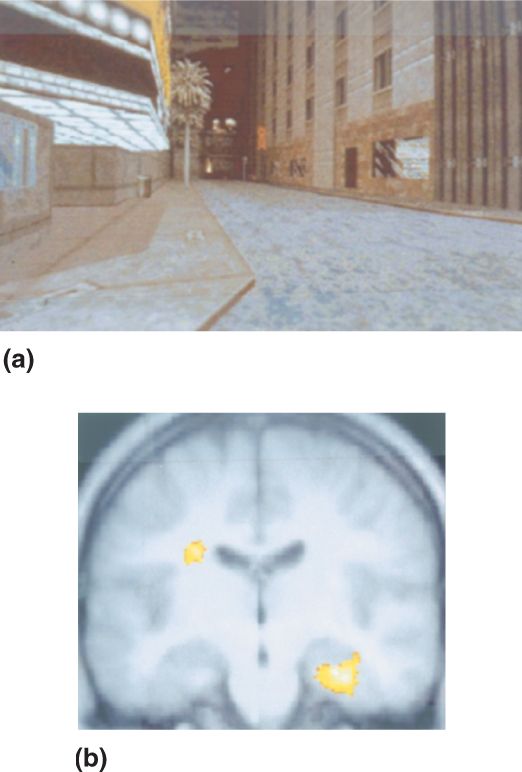
Imaging Working Memory in the Human Brain. Human brain imaging experiments suggest that numerous brain areas in the prefrontal cortex are involved in working memory. In one study by Courtney et al., brain activity was recorded by positron emission tomography (PET) while subjects performed two working memory tasks. In the identity task, three face photographs were briefly shown in succession; each image was at a different location and the subject looked at each face to memorize it. In the test phase, a face picture was shown at a new location and the subject indicated whether the face was the same as one of the memorized faces (Figure 24.9a). In the location task a similar paradigm was used, but the subject’s task was to memorize the locations of the three faces presented before the delay, the identities of the faces were irrelevant. In the test phase a fourth face was shown and the subject answered whether that face was at the same location as one of the memorized locations (Figure 24.9b). Both experiments looked for brain activity during the delay interval between the memorization and test phases during which the subject had to hold information in mind. In the first experiment, this was information about faces; in the second experiment, it was information about spatial locations.

FIGURE 24.9 Human brain activity in two working memory tasks. (a) In an identity task, subjects saw three photographs of faces in succession (shown together in the figure). The faces were memorized, and after a delay, a test face was presented at a new location. Subjects indicated whether the test face was the same as one of the memorized faces. (b) In a location task, three faces were shown in succession and the subjects memorized their locations on the display. In the test phase, a fourth face was shown and subjects indicated whether it appeared at the same location as one of the faces in the memorization phase. (c) Lateral and (d) medial views of brain activation in the two tasks. Six areas in the frontal lobe showed sustained activity correlated with working memory. The three blue areas showed greater activity in the facial identity task, the two green areas were equally active in facial identity and spatial location tasks, and the red area was more active in the location task. (Source: Adapted from Haxby et al., 2000, Fig. 5.) Description
The brain areas that demonstrated significant working memory activity in these experiments are shown in Figure 24.9c and Figure 24.9d. Six areas in the frontal lobe showed significant sustained activity during the delay period, suggesting a role in working memory. Three areas exhibited stronger sustained activity for facial identity than spatial location, one area was more responsive to spatial memory, and two areas were active equally in facial and spatial memory tasks. An interesting unanswered question is whether working memory for other types of information is held in the same or different brain areas.
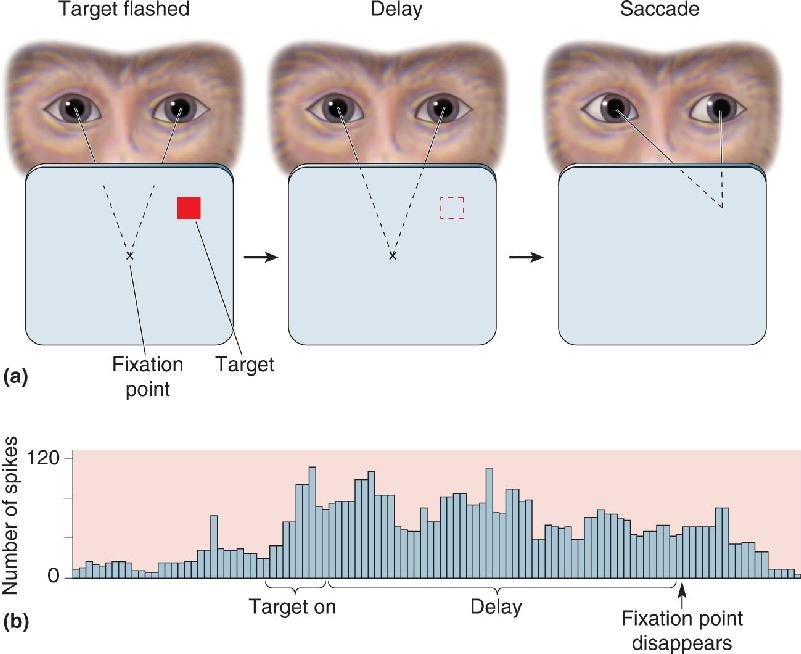
Cortical areas outside the frontal lobe have also been found to contain neurons that appear to retain working memory information. In Chapter 14, we saw an example in area 6 (see Figure 14.9). Another example is provided by the lateral intraparietal cortex (area LIP), buried in the intraparietal sulcus (see Figure 21.18). Area LIP is thought to be involved in guiding eye movements because electrical stimulation here elicits saccades in monkeys. The responses of many neurons in area LIP of monkeys suggest that they are also involved in a type of working memory. This pattern is evident in a delayed-saccade task, in which the animal fixates on a point on a computer screen and a target is briefly flashed at a peripheral location (Figure 24.10a). After the target goes off, there is a variable length delay. At the end of the delay period, the fixation point disappears, and the animal’s eyes make a saccadic movement to the remembered location of the target. The response of an LIP neuron while a monkey performs this task is shown in Figure 24.10b. The neuron begins firing shortly after the peripheral target is presented; this seems like a normal stimulus-evoked response. But the cell keeps firing throughout the delay period in which there is no stimulus. The neuron stops firing only after the saccadic eye movement begins. Further experiments using this delayed-saccade task suggest that the response of the LIP neuron is temporarily holding information that will be used to produce the saccade.

FIGURE 24.10 The delayed-saccade task. (a) To obtain a juice reward, the monkey is trained to perform the following actions. First, the animal fixates a central point while a peripheral target flashes on and off. During a delay period after the target goes off, the monkey continues to fixate the central point (the dashed square shows the remembered location where the target had been). At the end of the delay, the fixation point disappears, and the animal saccades to the remembered location of the target. (b) The histogram shows the response of an LIP neuron. The neuron begins firing when the target is presented and continues firing through the delay period until after the fixation point is gone and the saccadic eye movement begins. (Source: Adapted from Goldman-Rakic, 1992, Fig. p. 113, and Gnadt and Andersen, 1988, Fig. 2.) Description
Other areas in the parietal and temporal cortex have been shown to have analogous working memory responses. These areas seem to be modality-specific, just as the responses in area LIP are specific to vision. This is consistent with the clinical observation that there are distinct auditory and visual working memory deficits in humans produced by cortical lesions.
We’ve seen that sensory information can be temporarily held in mind by working memory, but how does the brain retain information for a longer time? Even before humans evolved to the point that we could cram for neuroscience exams by drawing cartoons of the brain, we needed to remember many things—the location of the river to drink from, where to find food, which cave to call home. To understand the neural basis of declarative memory storage, we first need to examine where in the brain it is stored. In other words, we must explore the location of a memory, known as an engram or memory trace. For example, when you learn the meaning of a word in a foreign language, where in your brain is this information stored; where is the engram?
In the 1920s, American psychologist Karl Lashley conducted experiments to study the effects of brain lesions on learning in rats. Well aware of the cytoarchitecture of the neocortex, Lashley set out to determine whether engrams resided in particular association areas of cortex (see Chapter 7), as was widely believed at the time.
In a typical experiment, he trained a rat to run through a maze to get a food reward. On the first trial, the rat was slow getting to the food because it would enter blind alleys and have to turn around. After running through the same maze repeatedly, the rat learned to avoid blind alleys and go straight to the food. Lashley was investigating how performance on this task was affected by different lesions in the rat’s cortex. He found that a rat given a brain lesion after it had learned to run the maze then made mistakes and went down blind alleys it had previously learned to avoid. Apparently the lesion damaged or destroyed the memory for how to reach the food.
How did the size and location of lesions affect learning and memory? Interestingly, Lashley found that the severity of the deficits caused by the lesions (both learning and remembering) correlated with the size of the lesions but was apparently unrelated to the location of the lesion within the cortex. Based on these findings, he speculated that all cortical areas contribute equally (are equipotential) to learning and memory; it was simply a matter of performance on the maze task becoming poorer as the lesion became bigger and the ability to remember the maze worsened. If true, this would be a very important finding because it implied that engrams are based on neural changes spread throughout the cortex rather than being localized to one area. The problem with this interpretation was that the Lashley’s lesions were large, each damaging multiple brain areas possibly involved in learning or remembering the maze task. Another problem was that the rats may have solved the maze in several different ways—by sight, feel, and smell—and the loss of one memory might have been compensated for by another.
Subsequent research has proven Lashley’s conclusions to be incorrect. All cortical areas do not contribute equally to all memories. Nonetheless, his conclusions that all of the cortex participates in memory storage, and that an engram can be widely distributed in the brain, are correct and important. Lashley had a major impact on the study of learning and memory because he led other scientists to consider ways in which memories might be distributed among the vast number of neurons of the cerebral cortex.
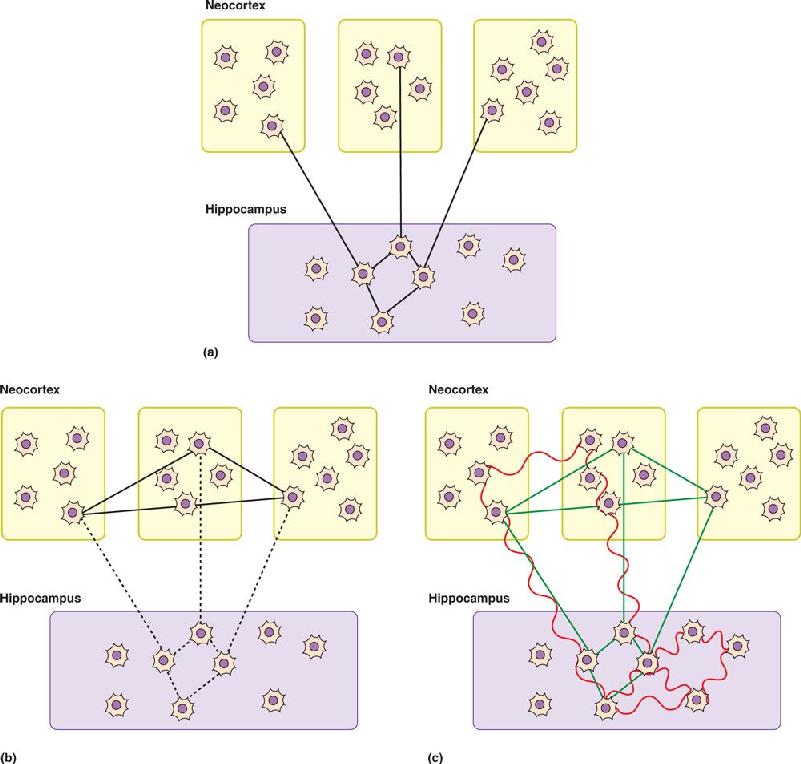
Hebb and the Cell Assembly. Lashley’s most famous student was Donald Hebb, introduced in Chapter 23. Hebb reasoned that it was crucial to understand how external events are represented in the activity of the brain before one can hope to understand how and where these representations are stored. In his remarkable book The Organization of Behavior, published in 1949, Hebb proposed that the internal representation of an object consists of all the cortical cells that are activated by the external stimulus (e.g., the circle in Figure 24.11). Hebb called this group of simultaneously active neurons a cell assembly (Figure 24.11a). Hebb imagined that all these cells were reciprocally interconnected. The internal representation of the object was held in working memory as long as activity reverberated through the connections of the cell assembly. Hebb further hypothesized that if activation of the cell assembly persisted long enough, consolidation would occur by a “growth process” that made these reciprocal connections more effective; neurons that fired together would wire together (Figure 24.11b). Subsequently, if only a fraction of the cells of the assembly were activated by a later stimulus (e.g., segments of a circle), the now-powerful reciprocal connections would cause the whole assembly to become active again, thus recalling the entire internal representation of the external stimulus—in this case, the circle (Figure 24.11c).

FIGURE 24.11 Hebb’s cell assembly and memory storage. Description
Hebb’s important message about the engram was twofold: (1) It could be widely distributed among the connections that link the cells of the assembly, and (2) it could involve the same neurons that are involved in sensation and perception. Destruction of only a fraction of the cells of the assembly would not be expected to eliminate the memory, possibly explaining Lashley’s results. Hebb’s ideas stimulated the development of neural network computer models. Although his original assumptions had to be modified slightly, we will see in Chapter 25 that these models have successfully reproduced many features of human memory.
Where is the engram for a foreign language? Look to the regions of the brain in the temporal and parietal lobes that normally process language. A lesion here can disrupt your memory of a foreign word but leave intact the memory of your foreign-born grandmother’s face. However, although declarative memories may finally reside in many areas of the neocortex, decades of research indicate that to get there, they must pass through structures in the medial temporal lobes. Let’s explore the evidence.
A variety of experiments indicate that structures in the medial temporal lobe are particularly important for the consolidation and storage of declarative memories. Examples are experiments that used electrical stimulation or neural recordings from the temporal lobe. Further evidence came from cases of amnesia that resulted from damage to the temporal lobes. Before considering the studies that suggest that memory storage involves the medial temporal lobes, let’s look at the anatomy of this brain region.
Anatomy of the Medial Temporal Lobe. The temporal lobe is located under the temporal bone, so named because the hair of the temples is often the first to go gray with the passage of time (tempus is Latin for “time”). The association of the temporal lobe with time was fortuitous as this region of the brain is important for recording past events. The medial portion of the temporal lobe contains the temporal neocortex, which may be a site of long-term memory storage, and a group of structures interconnected with neocortex that are critical for the formation of declarative memories.
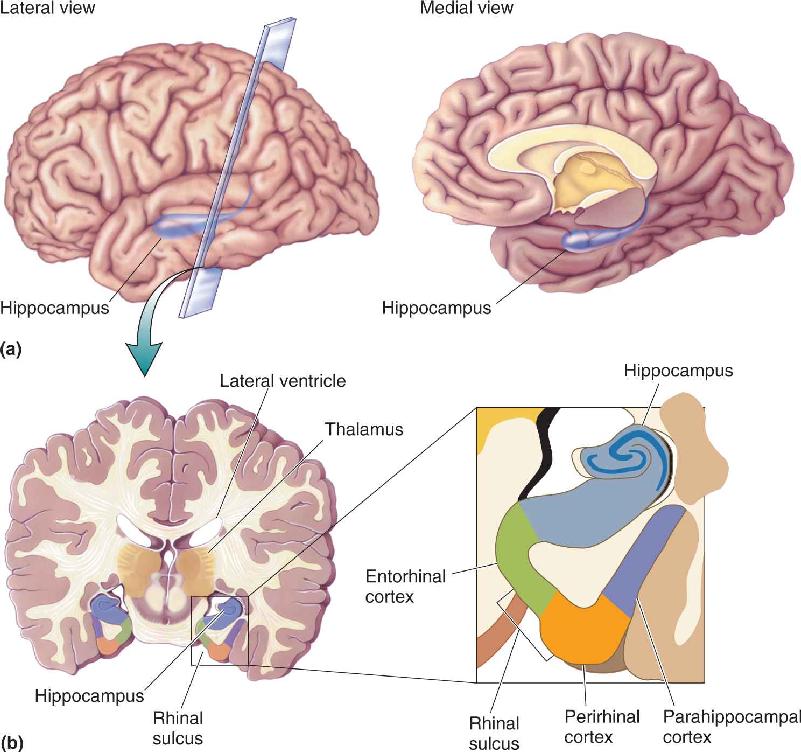
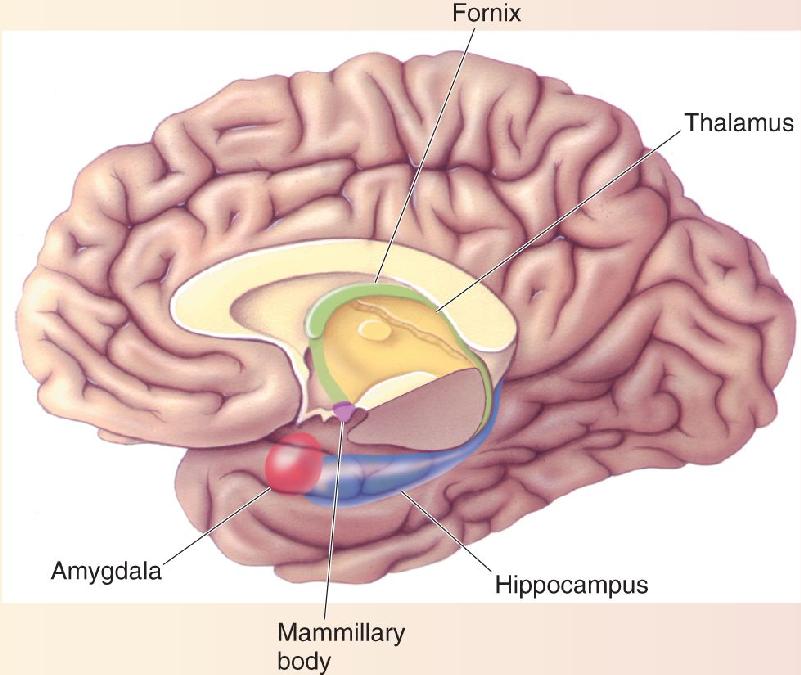
The key structures are the hippocampus, the nearby cortical areas, and the pathways that connect these structures with other parts of the brain (Figure 24.12). As we saw in Chapter 7, the hippocampus is a folded structure situated medial to the lateral ventricle. The name means “seahorse,” a resemblance you can see in Figure 24.13. Ventral to the hippocampus are three important cortical regions that surround the rhinal sulcus: the entorhinal cortex, which occupies the medial bank of the rhinal sulcus; the perirhinal cortex, which occupies the lateral bank; and the parahippocampal cortex, which lies lateral to the rhinal sulcus. (We’ll refer to entorhinal cortex and perirhinal cortex collectively as rhinal cortex.)

FIGURE 24.12 Structures in the medial temporal lobe involved in declarative memory formation. (a) Lateral and medial views show the location of the hippocampus in the temporal lobe. (b) The brain is sectioned coronally to show the hippocampus and cortex of the medial temporal lobe. Description
FIGURE 24.13 The hippocampus. In Greek mythology, the hippocampus was a creature combining the front of a horse with the hind end of a dolphin or fish. Here a dissected hippocampus is shown next to a seahorse. (Source: Laszlo Seress/Wikimedia Commons.) Description
Inputs to the medial temporal lobe come from the association areas of the cerebral cortex, containing highly processed information from all sensory modalities (Figure 24.14). For instance, inferotemporal visual cortex (area IT) projects to the medial temporal lobe, but low-order visual areas such as striate cortex do not. This means that the input contains complex representations, perhaps of behaviorally important sensory information, rather than responses to simple features such as light–dark borders. Input first reaches the rhinal and parahippocampal cortex before being passed to the hippocampus. A major output pathway from the hippocampus is the fornix, which loops around the thalamus before terminating in the hypothalamus.

FIGURE 24.14 Information flow through the medial temporal lobe. Description
Electrical Stimulation of the Human Temporal Lobes. One of the most intriguing and controversial studies implicating the neocortex of the temporal lobe in the storage of declarative memory traces involved electrical stimulation of the human brain. In Chapters 12 and 14 we discussed the work of Wilder Penfield, in which patients’ brains, as part of surgical treatment for severe epilepsy, were electrically stimulated at numerous locations prior to ablation of the seizure-prone region. Stimulation of somatic sensory cortex caused the patient to experience tingling sensations in regions of skin, whereas stimulation of motor cortex caused a certain muscle to twitch.
Electrical stimulation of the temporal lobe occasionally produced more complex sensations than stimulation in other brain areas. In a number of cases, Penfield’s patients described sensations that sounded like hallucinations or recollections of past experiences. This is consistent with reports that epileptic seizures of the temporal lobes can evoke complex sensations, behaviors, and memories. Here is part of Penfield’s account of one operation:
At the time of operation, stimulation of a point on the anterior part of the first temporal convolution on the right caused him [the patient] to say, “I feel as though I were in the bathroom at school.” Five minutes later, after negative stimulations elsewhere, the electrode was reapplied near the same point. The patient then said something about “street corner.” The surgeon asked him, “where” and he replied, “South Bend, Indiana, corner of Jacob and Washington.” When asked to explain, he said he seemed to be looking at himself—at a younger age. (Penfield, 1958, p. 25.)
Another patient reported a similar sense of experiencing flashbacks. When her temporal cortex was stimulated, she said, “I think I heard a mother calling her little boy somewhere. It seemed to be something that happened years ago.” With stimulation at another location, she said, “Yes, I hear voices. It is late at night around the carnival somewhere—some sort of traveling circus. . . I just saw lots of big wagons that they use to haul animals in.”
Are these people reexperiencing events from earlier in their life because memories are evoked by the electrical stimulation? Does this mean that memories are stored in the neocortex of the temporal lobe? Those are tough questions! One interpretation is that the sensations are recollections of past experiences. The fact that such elaborate sensations resulted only when the temporal lobe was stimulated suggests that the temporal lobe may play a special role in memory storage. However, other aspects of the findings do not clearly support the hypothesis that engrams are being electrically activated. For instance, some brain-stimulated patients said they saw themselves, something that we cannot normally experience. Also, it is important to appreciate that complex sensations were reported only by a minority of the patients, and all of these patients had an abnormal cortex associated with their epilepsy.
There is no way to prove whether the complex sensations evoked by temporal lobe stimulation are recalled memories. However, it is clear that the consequences of temporal lobe stimulation and temporal lobe seizures can be qualitatively different from stimulation of other areas of the neocortex.
Neural Recordings from the Human Medial Temporal Lobe. Electrical stimulation of the temporal lobe sometimes produces memory-like experiences, and as we will see, lesions here disrupt memory. But what are the medial temporal neurons normally doing? A glimpse at the normal function of these neurons comes from studies using microelectrodes to record from the brains of living humans who suffered from epileptic seizures that were not alleviated by drugs. As in Penfield’s studies, plans were made to surgically remove the abnormally active part of the brain after localizing the area with electrode recordings. Because temporal lobe seizures are common, the electrodes were often placed in the hippocampus and surrounding structures. In some ways such recordings are reminiscent of studies of visual neurons in monkey inferotemporal cortex. As seen in monkeys, neurons were found that preferentially responded to categories of objects, including faces, household objects, and outdoor scenes (recall face-selective neurons in Chapter 10). These neurons are said to be invariant because they responded to quite a variety of visual images that are structurally or conceptually related.
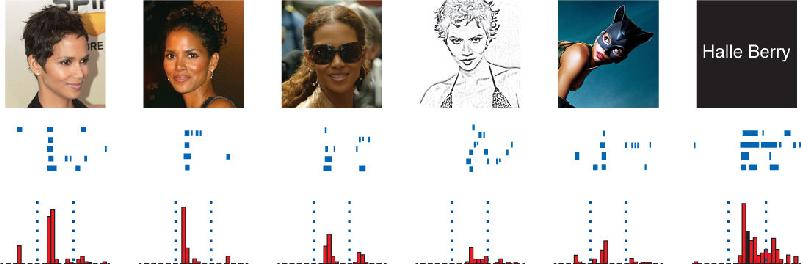
In further studies, even more selectivity was discovered in a small percentage of the neurons examined. For example, individual hippocampal neurons were found in one patient that responded selectively to pictures of the actress Jennifer Aniston or the basketball player Michael Jordan. Figure 24.15 shows a neuron in the hippocampus that responded to various stimuli associated with the actress Halle Berry. The diversity of the effective stimuli was striking, including photographs of Ms. Berry, a drawing of her face, and even her printed name. The neuron was activated by pictures of Ms. Berry dressed for her role in the movie Catwoman, yet it did not respond to other women similarly dressed. Other neurons were selective for landmarks such as the Eiffel Tower and the Leaning Tower of Pisa.

FIGURE 24.15 A patient’s hippocampal neuron selectively responds to actress Halle Berry. This neuron responded to photos and drawings of Halle Berry as well as her written name. The cell responded less or not at all to photos, drawings, and written names of other people. (Adapted from: Quiroga et al., 2005, Fig. 2.)
What are we to make of these neurons? One way to think about them is that they are somewhere on a continuum between purely visual coding in the lateral temporal lobe and memory coding in the medial temporal lobe. We can’t be sure, but such neurons are probably not essential for recognition because common objects and famous faces are recognized even after lesions to the hippocampus; even H.M. recognized people and things that existed before his surgery. Recognition may rely on portions of the temporal lobe that are more lateral and posterior. The highly selective neurons in the hippocampus may serve a role in the formation of new memories of people and things we already recognize, such as the patient’s memory of Halle Berry. Many questions remain. Might less specific responses have been found in these experiments if more stimuli were used (a cell that responds to Justin Timberlake, canned peas, and doorknobs)? Are there neurons selectively activated by every object that we recognize, or are the examples we’ve discussed rare cases related to repeated exposure of very famous people or things? Do these findings even apply to normal brains, since it is conceivable that the seizure-prone brains were abnormally organized and responsive?
If the temporal lobe is particularly important for learning and memory, one would expect that removing both temporal lobes would have a profound effect on these functions. Studies in both humans and animals show that this is indeed the case.
The Case of H.M.: Temporal Lobectomy and Amnesia. A renowned case of amnesia resulting from temporal lobe damage provides further evidence for the importance of this region in memory. This case concerns the memory of Henry Molaison, whose name was made public only after his death in 2008 (Figure 24.16). For the previous half century, studies referred to him as H.M., probably the most famous initials in the history of neuroscience. H.M. experienced minor epileptic seizures beginning around age 10, and as he aged, they became more serious generalized seizures involving convulsions, tongue biting, and loss of consciousness. Although the cause of the seizures is not known, they may have resulted from damage sustained in a bicycle accident at the age of 9 that left him unconscious for 5 minutes. After graduating from high school he got a job, but despite heavy medication with anticonvulsants, his seizures increased in frequency and severity to the point that he was unable to work. In 1953, at the age of 27, H.M. had an operation in which an 8 cm length of medial temporal lobe was bilaterally excised, including cortex, the underlying amygdala, and the anterior two-thirds of the hippocampus, in a last-ditch attempt to assuage the seizures. The surgery was successful in alleviating the seizures.

FIGURE 24.16 The brain lesion in patient H.M. that produced profound anterograde amnesia. (a) The medial temporal lobe was removed from both hemispheres in H.M.’s brain to alleviate epileptic seizures. (b) A normal brain, showing the location of the hippocampus and cortex that were removed from H.M.’s brain. (Source: Adapted from Scoville and Milner, 1957, Fig. 2.) (c) Henry Molaison as a high school student before the surgery. (Source: Photo courtesy of Suzanne Corkin. Copyright © by Suzanne Corkin used by permission of The Wylie Agency LLC.) Description
The removal of much of the temporal lobes had little effect on H.M.’s perception, intelligence, or personality, but the surgery left him with profound and debilitating anterograde amnesia. Drs. Brenda Milner and Suzanne Corkin, initially at the Montreal Neurological Institute, worked with H.M. for 50 years, but, incredibly, they had to introduce themselves to him every time they met. They found that H.M. would forget events almost as quickly as they occurred. With repetition he could remember a number for a short time, but if he was distracted he would not only forget the number, he would also forget that he had even been asked to remember one.
To be clear about the nature of H.M.’s amnesia, we must contrast what was lost with what was retained. In addition to anterograde amnesia, he had some degree of retrograde amnesia. He retained some memories of his childhood but little or no memory for events just before his surgery. Testing shortly after the surgery suggested that H.M. had retrograde amnesia for events extending back several years before his surgery. Later studies suggested that his retrograde amnesia might extend back decades. H.M.’s working memory was largely normal. For instance, with constant rehearsal he could remember a list of six numbers, although any interruption would cause him to forget. H.M. was actually able to learn a very small number of declarative facts after his surgery. For example, he could recognize and name a few people who became famous after his surgery, such as U.S. President John Kennedy. He also learned the floor plan of a home he moved to after the surgery. These rare remembered facts probably resulted from extensive daily repetition. H.M. was also able to learn new tasks (i.e., form new procedural memories). For example, he was taught to draw by looking at his hand in a mirror, a task that takes a good deal of practice for anyone. The odd thing is that he learned to perform new tasks despite the fact that he had no recollection of the specific experiences in which he was taught to do them (the declarative component of the learning).
To appreciate the significance of H.M., realize that prior to his surgery, little was known about the function of the hippocampus and surrounding structures. Considering H.M.’s amnesia in the context of earlier research, we can conclude that the medial temporal lobe is critical for memory consolidation but not for the retrieval of memories. Although there is some controversy about the temporal extent of H.M.’s retrograde amnesia, he clearly retained declarative memories for many things from before the surgery, such as famous faces and the meanings of words. This implies that medial temporal structures do not store all memories even though engrams for some things may be located there. The fact that his working memory was largely intact means this does not rely on the medial temporal lobe. Finally, H.M’s amnesia indicates that the formation and retention of procedural memories use brain structures distinct from those involved in declarative memory consolidation and perhaps storage.
An Animal Model of Human Amnesia. The amnesia of H.M. makes a strong case that one or more structures in the medial temporal lobe are essential for the formation of declarative memories. If these structures are damaged, severe anterograde amnesia results. Experiments have mostly used the experimental ablation technique to assess whether the removal of various parts of the temporal lobe affects memory.
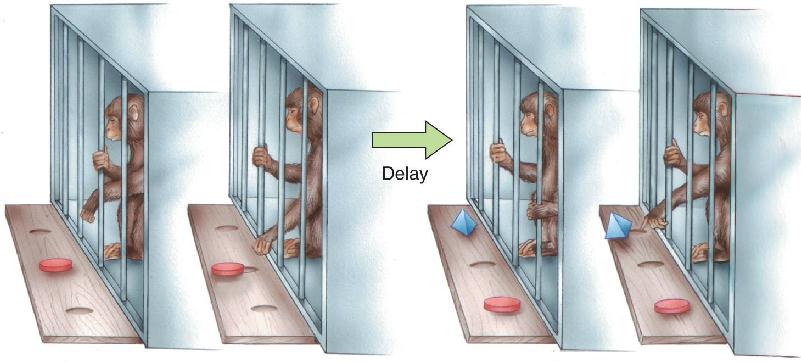
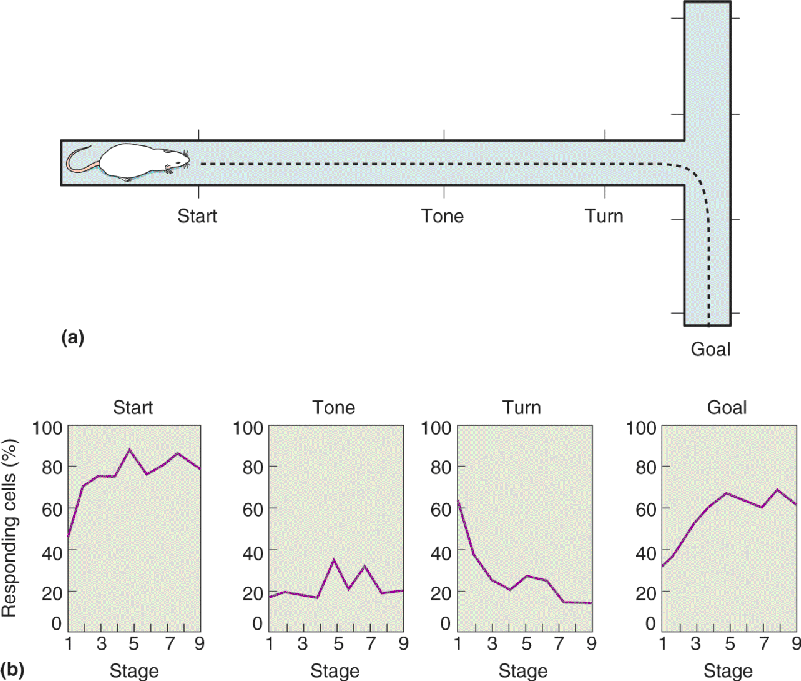
Because the macaque monkey brain is similar in many ways to the human brain, macaques are frequently studied to further our understanding of human amnesia. The monkeys are often trained to perform tasks called delayed match-to-sample and delayed non-match to sample (DNMS) (Figure 24.17). In this type of experiment, a monkey faces a table that has several small wells in its surface. It first sees the table with one object on it covering a well. The object might be a wooden block or a chalkboard eraser (the sample stimulus). The monkey is trained to displace the object so that it can grab a food reward in the well under the object. After getting the food, a screen prevents the monkey from seeing the table for some period of time (the delay interval). Finally, the animal gets to see the table again, but now there are two objects on it: One is the same as before, and another is new. If a match-to-sample experiment is being conducted, the animal must displace the object it recognizes to get a food reward. In DNMS, the monkey’s task is to displace the new object (the non-matching object) in order to get a food reward in the well below it. Normal monkeys are relatively easy to train on the non-matching task and get very good at it, perhaps because it exploits their natural curiosity for novel objects. With delays between the two stimulus presentations of anywhere from a few seconds to 10 minutes, the monkey correctly displaces the non-matching stimulus on about 90% of the trials. Memory required in the DNMS task has been called recognition memory because it involves the ability to judge whether a stimulus has been seen before.

FIGURE 24.17 The delayed non-match to sample (DNMS) task. A monkey first displaces a sample object to obtain a food reward. After a delay, two objects are shown, and recognition memory is tested by having the animal choose the object that does not match the sample. (Source: Adapted from Mishkin and Appenzeller, 1987, p. 6.)
In the early 1980s, experiments performed by Mortimer Mishkin and his colleagues at the National Institute of Mental Health, and Larry Squire and his coworkers at the University of California, San Diego, demonstrated that severe deficits on the DNMS task result from bilateral medial temporal lesions in macaque monkeys. Performance was close to normal if the delay between the sample stimulus and the two test stimuli was short (a few seconds). This is important because it indicates that the monkey’s perception was still intact after the ablation and the animal remembered the DNMS procedure. But when the delay was increased from a few seconds to a few minutes, the monkey made increasingly more errors choosing the non-matching stimulus (Figure 24.18). With the lesion, the animal was no longer as good at remembering what the sample stimulus was in order to choose the other object. This behavior suggests that it forgot the sample stimulus if the delay was too long. The deficit in recognition memory produced by the lesion was not specific to the visual modality, since this deficit was also observed if the monkey was allowed to touch but not see the objects.

FIGURE 24.18 The effect of medial temporal lesions on DNMS performance. The Y axis shows the percentage of correct choices made by monkeys as a function of the length of the delay interval. The performance of normal monkeys is compared to that of monkeys with large bilateral medial temporal lesions. (Source: Adapted from Squire, 1987, Fig. 49.) Description
The monkeys with medial temporal lesions appeared to provide a good model of human amnesia. As with H.M., the amnesia was anterograde, it involved declarative rather than procedural memory, working memory was intact, and consolidation was severely impaired. Note that the surgical lesions that produced recognition memory deficits in these monkeys were quite large. They included the hippocampus, amygdala, and rhinal cortex. At one time it was thought that the key structures damaged in such lesions were the hippocampus and amygdala. Recall from Chapter 18 that the amygdala plays a special role in memory for emotional experiences. However, research has now shown that selective amygdala lesions have no effect on recognition memory, and lesions of the hippocampus alone produce only relatively mild amnesia. For example, Squire studied a man known as R.B. who had bilateral hippocampal damage as a result of oxygen deprivation during surgery. Although R.B. clearly had difficulty forming new memories, his anterograde amnesia was not nearly as severe as that seen in H.M. The most severe memory deficits result from damage to the perirhinal cortex. The anterograde amnesia resulting from perirhinal lesions is not specific to information from a particular sensory modality, reflecting the convergence of input from association cortex of multiple sensory systems.
Together with the hippocampus, the cortex in and around the rhinal sulcus evidently performs a critical transformation of the information coming from association cortex. Some studies suggest that the hippocampus and rhinal cortex are involved in different facets of memory; the hippocampus may signal that a particular object had been seen before (“I remember that object”), whereas the perirhinal cortex may be involved more in signaling familiarity (“That object looks familiar but I don’t remember it specifically”). Still, such distinctions remain controversial. At any rate, it appears that, collectively, medial temporal structures are critical for the consolidation of memory. They may also have an essential intermediate processing role that involves something more than consolidation. H.M. and possibly R.B. had some retrograde amnesia. Perhaps in addition to consolidation, medial temporal structures play a role in the storage of memories for a long or short time (depending on which expert you ask). Our discussion of brain areas involved in anterograde amnesia has focused on structures in the medial temporal lobe, but it is important to note that lesions to interconnected areas elsewhere in the brain also produce amnesia (Box 24.2).
In Chapter 18 we learned about the Papez circuit, a series of strongly interconnected structures that ring the diencephalon. A major component of this circuit is a massive bundle of axons called the fornix that connects the hippocampus with the mammillary bodies in the hypothalamus (Figure A). The mammillary bodies, in turn, send a strong projection to the anterior nuclei of the thalamus. The dorsomedial nucleus of the thalamus also receives input from temporal lobe structures, including the amygdala and inferotemporal neocortex, and it projects to virtually all of the frontal cortex.

Figure A Midline brain structures involved in memory. Temporal lobe structures including the hippocampus, amygdala, and inferotemporal cortex project to the thalamus and hypothalamus, including the mammillary bodies, in the diencephalon.
Considering the central role of the temporal lobes in memory processing, it is perhaps not surprising that damage to these connected diencephalic structures can also cause amnesia.
A particularly dramatic example of the amnesic effects of damage to the diencephalon in humans is the case of a man known as N.A. In 1959 at the age of 21, N.A. was a radar technician in the U.S. Air Force. One day he was sitting down assembling a model in his barracks, while behind him a roommate played with a miniature fencing foil. N.A. turned at the wrong moment and was stabbed. The foil went through his right nostril and took a leftward course into his brain. Many years later when a CT scan was performed, the only obvious damage was a lesion in his left dorsomedial thalamus, though there may have been other damage.
After his recovery, N.A.’s cognitive ability was normal, but his memory was impaired. He had relatively severe anterograde amnesia as well as retrograde amnesia for a period of about 2 years preceding the accident. While he could remember some faces and events from the years following his accident, even these memories were sketchy. He had difficulty watching television because during commercials, he’d forget what had previously happened. In a sense, he lived in the past and preferred to wear old familiar clothes and keep his hair in an older style.
Although N.A.’s amnesia was less severe than H.M.’s, its quality was strikingly similar. He had preservation of short-term memory, recollection of old memories, and general intelligence. Along with difficulty forming new declarative memories, he had retrograde amnesia for 2 years preceding the accident that produced the amnesia. The similarities in the effects of medial temporal and diencephalic lesions suggests that these interconnected areas are part of a system serving the common function of memory consolidation.
Further support for a role of the diencephalon in memory comes from studies of Korsakoff’s syndrome. Usually resulting from chronic alcoholism, Korsakoff’s syndrome is characterized by confusion, confabulations, severe memory impairment, and apathy. As a result of poor nutrition, alcoholics may develop a thiamin deficiency, which can lead to such symptoms as abnormal eye movements, loss of coordination, and tremors. This condition can be treated with supplemental thiamin. If left untreated, however, thiamin deficiency can lead to structural brain damage that produces Korsakoff’s syndrome. Although all cases of Korsakoff’s syndrome are not associated with damage to the same parts of the brain, there are usually lesions in the dorsomedial thalamus and the mammillary bodies.
In addition to anterograde amnesia, Korsakoff’s syndrome can involve more severe retrograde amnesia than observed in N.A. and H.M. No strong correlation exists between the severities of anterograde amnesia and retrograde amnesia in Korsakoff’s syndrome. This is consistent with the other studies of amnesia we’ve discussed, suggesting that the mechanisms involved in consolidation (disrupted in anterograde amnesia) are largely distinct from the processes used to recall memories (disrupted in retrograde amnesia). Based on a small number of cases such as that of N.A., researchers suspect that anterograde amnesia associated with diencephalic lesions results from damage to the thalamus and mammillary bodies. Although it is not clear which damage causes the retrograde amnesia, in addition to diencephalic lesions, Korsakoff’s patients sometimes have damage to the cerebellum, brain stem, and neocortex.
Memory formation, retention, and retrieval involve a system of interconnected brain areas. Considerable evidence points to the importance of the medial temporal lobe for declarative memory, and within this region of the brain the hippocampus has received the greatest attention. It’s not simple to pin down just what the hippocampus does, however, as it is involved in various memory functions at multiple time scales. To avoid getting lost, keep a few basic points in mind. First, the hippocampus appears to play a critical role in binding sensory information for the purpose of memory consolidation. Second, a lot of research, particularly in rodents, has shown that the hippocampus supports spatial memory of the location of objects of behavioral importance. This may be one of the specialized functions of the hippocampus, or it might be an example of the binding of sensory information. Finally, the hippocampus is involved in the storage of memories for some length of time, though the time duration is controversial.
The Effects of Hippocampal Lesions in Rats. Rodents have played an important role elucidating the memory functions of the hippocampus. In one type of experiment, rats are trained to get food in a radial arm maze, a device devised by David Olton and his colleagues at Johns Hopkins University. This apparatus consists of arms, or passageways, radiating from a central platform (Figure 24.19a). If a normal rat is put in such a maze, it will explore until it finds the food at the end of each arm. With practice, the rat becomes efficient at finding all the food, going down each arm of the maze just once (Figure 24.19b). To run through the maze without going twice into any of the arms, the rat uses visual or other cues around the maze to remember where it has already been. Working memory is presumably used to retain information about which arms have been visited.

FIGURE 24.19 Following a rat through a radial arm maze. (a) An eight-arm radial maze. (b) The path of a rat through a maze in which all the arms contain food. (c) If a rat learns that four of the eight arms never contain food, it will ignore these and follow a path to only the baited arms. (Source: Parts b and c, adapted from Cohen and Eichenbaum, 1993, Fig. 7.4.) Description
If the hippocampus is destroyed before the rat is put in the maze, its performance will differ from normal behavior in an interesting way. In one sense, rats with lesions seem normal; they learn to go down the arms of the maze and eat the food placed at the end of each arm. But unlike normal rats, they never learn to do this efficiently. Rats with hippocampal lesions go down the same arms more than once, only to find no food after the first trip, and they leave other arms containing food unexplored for an abnormally long time. It appears that the rats can learn the task in the sense that they go down the arms in search of food (the procedural memory). But they cannot seem to remember which arms they’ve already visited.
A variation on the radial arm experiment illustrates an important subtlety in the deficit produced by destroying the hippocampus. Instead of placing food at the end of all the arms of the maze, food is placed only at the ends of certain arms and never in the other ones. After a bit of practice, a normal rat learns to avoid going down the arms that never contain food (Figure 24.19c). At the same time, the rat learns to efficiently get the food in the other arms, entering each arm just once. How do rats with hippocampal lesions perform on this task? Interestingly, just like normal rats, they are able to learn to avoid the arms that never contain food. But they still are not able to get the food from the other arms without wasting time going down the same arms more than once. So how can we argue that the lesion disrupts the ability to learn the locations of arms that have already been entered, even though the rat can learn to avoid arms that never contain food? Evidently, the key to making sense of these findings is that the information about the no-food arms is always the same each time the rat goes in the maze (i.e., no-food arms are memorized as part of the “procedure”), whereas the information about which arms the rat has already entered requires working memory and varies from one trial to the next.
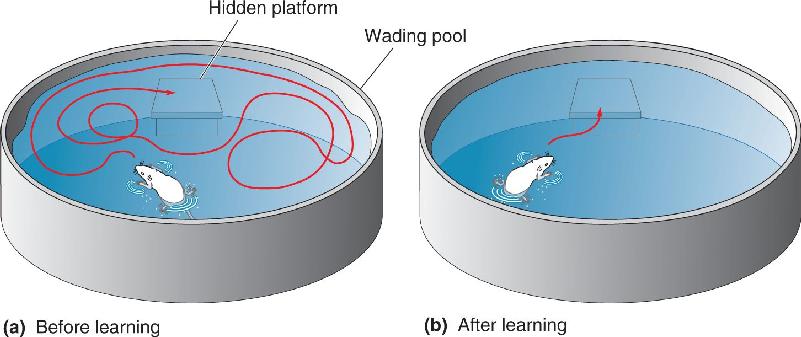
Spatial Memory, Place Cells, and Grid Cells. Several lines of evidence suggest that the hippocampus is particularly important for spatial memory. The Morris water maze, a commonly used test of spatial memory in rats, was devised by Richard Morris of the University of Edinburgh. In this test, a rat is placed in a pool filled with cloudy water (Figure 24.20). Submerged just below the surface in one location is a small platform that allows the rat to escape the water. A rat placed in the water the first time will swim around until it bumps into the hidden platform, and then it will climb onto it. Normal rats quickly learn the spatial location of the platform and on subsequent trials waste no time swimming straight to it. Moreover, once they have figured out what to search for, rats put in a maze with the platform at a different location learn the task much faster. But rats with bilateral hippocampal damage never seem to figure out the game or remember the location of the platform.

FIGURE 24.20 The Morris water maze. (a) The trajectory a rat might take to find a hidden platform the first time the rat is placed in the pool. (b) After repeated trials, the rat knows where the platform is and swims straight to it. Description
What properties of hippocampal neurons aid them in their spatial navigation and memory? In a fascinating series of experiments begun in the early 1970s, John O’Keefe and his colleagues at University College London showed that many neurons in the hippocampus selectively respond when a rat is in a particular location in its environment. Suppose we have a microelectrode implanted in the hippocampus of a rat while it scurries about inside a large box. At first the cell is quiet, but when the rat moves into the northwest corner of the box, the cell starts firing. When it moves out of the corner, the firing stops; when it returns, the cell starts firing again. The cell responds only when the rat is in that one portion of the box (Figure 24.21a). This location, which evokes the greatest response, is called the neuron’s place field. We try recording from another hippocampal cell and it too has a place field, but this one fires only when the rat goes to the center of the box. For obvious reasons, these neurons are called place cells.

FIGURE 24.21 Place cells in the hippocampus. A rat explores a small box for 10 minutes (left panels). Then the partition is removed, so the rat can explore a larger area (center and right panels). (a) Color coding indicates the area in the box where one place cell in the hippocampus responds: red, large response; yellow, moderate response; blue, no response. This cell has a place field in the smaller upper box; when the partition is removed, it stays in the same location. (b) In this case, an electrode is next to a cell in the hippocampus that does not respond when the rat is in the smaller upper box (left). In the first 10 minutes after the partition is removed, the cell also does not respond (center). But after another 10 minutes, a place field develops in the new larger box (right). (Source: Adapted from Wilson and McNaughton, 1993, Fig. 2.) Description
In some ways, place fields are similar to the receptive fields of neurons in the sensory systems. For instance, the location of the place field is related to sensory input such as visual stimuli in the environment. In our experiment with the rat in the box, we could paint images above the four corners, such as a star above the northwest corner, a triangle above the southeast corner, and so on. Consider a cell that responds only when the rat is in the northwest corner of the box, near the painted star. Suppose we take the rat out of the box and blindfold it. We then secretly go back and rotate the box 180° so that now the northwest corner has the triangle and the southeast corner has the star. Will the cell we were previously studying respond when the animal is in the northwest corner, or will it respond in the corner where the star is now located (the southeast corner)? We put the rat back in the box and take off the blindfold. It starts exploring, and the neuron becomes active when the rat goes to the corner near the star. This demonstrates that, at least under some conditions, the response is based on visual input.
While place cells are similar to receptive fields in some ways, there are also major differences. For instance, once the animal has become familiar with the box with the images painted in each corner, a neuron will continue to fire when the rat goes into the northwest corner, even if we turn off the lights so it cannot see the location markers. Evidently, the responses of place cells are related to where the animal thinks it is. If there are obvious visual cues (such as the star and triangle), the place fields will be based on these cues. But if there are no cues (e.g., because the lights are out), the place cells will still be location-specific as long as the animal has had enough time to explore the environment and develop a sense of where it is.
Performance in the radial arm maze, discussed above, may utilize these place cells that code for location. Of particular importance in this regard is the finding that place fields are dynamic. For instance, if the box the animal is in is stretched along one axis, place fields will stretch in the same direction. In another manipulation, we first let a rat explore a small box and determine the place fields of several cells. Then we cut a hole in a side of the box so the animal can explore a larger area. Initially, there are no place fields outside the smaller box, but after the rat has explored its new expanded environment, some cells will develop place fields outside the smaller box (Figure 24.21b). These cells seem to learn in the sense that they alter their receptive fields to suit behavioral needs in the new larger environment. It’s easy to imagine how these sorts of cells could be involved in remembering arms already visited in the radial arm maze, just as you might return from a long hike by following markers you left when you first walked through the woods. If hippocampal place cells are involved in running the maze, it makes sense that performance is degraded by destroying the hippocampus.
Whether or not there are place cells in the human brain is not known. However, PET imaging studies show that the human hippocampus is activated in situations involving virtual or imagined navigation through the environment. In one experiment, subjects were positioned in a PET machine while playing a video game. They could navigate a virtual town on the game monitor by using buttons for forward movement, backward movement, and turning (Figure 24.22a). After learning their way around the virtual town, their brain activity was recorded while they navigated from an arbitrary starting point to a chosen destination. In a control condition, subjects moved through the virtual environment from the same start to finish locations, but arrows in the town always pointed them in the correct direction. In this condition, they did not have to think about how to navigate.

FIGURE 24.22 Activity in the human brain related to spatial navigation. (a) A virtual town was shown on a computer monitor, and subjects in a PET imaging machine used buttons to navigate the virtual environment. (b) Increased brain activity associated with spatial navigation was observed in the right hippocampus and left tail of the caudate (yellow). (Source: Maguire et al., 1998, Fig. 1.) Description
Figure 24.22b shows the difference in brain activity between the navigation condition and the control condition with directional arrows. When the person had to navigate the environment, there was increased activation of the right hippocampus and the left tail of the caudate. The asymmetry in the activation of the left and right hemispheres is an interesting frequent observation, but our primary point here is that the hippocampus is particularly active in this spatial navigation task with humans, just as it is in rats. The caudate activation is thought to reflect movement planning.
The hippocampus has also been studied in London taxi drivers who must learn the locations of innumerable city sites and roughly 25,000 streets to pass rigorous exams and acquire a license. One study showed that, compared to a control group, taxi drivers have a larger posterior and smaller anterior hippocampus. The size of the posterior hippocampus also seems to correlate with the length of experience as a taxi driver.
If the human hippocampus is used for spatial navigation, do hippocampal lesions impair navigation? An interesting case is a man known as T.T. who suffered bilateral hippocampal damage from encephalitis after a career of nearly 40 years as a London taxi driver. After his hippocampal damage, T.T. was very good at recognizing city landmarks and their topographical layout. A virtual reality simulation of driving through London was used to test his navigation skills. The researchers found that sometimes T.T. could “drive” efficiently from one point to another in the city while at other times he deviated from the ideal route. They found that T.T. did a good job when he could stick to main roads but got lost when smaller roads were required, as if he had lost the fine-grained knowledge of city topography he once had. The studies of humans navigating space in video games and the streets of London suggest that the human hippocampus is important for spatial memory, a finding reminiscent of the rat lesion experiments.
In addition to place cells in the hippocampus, recordings in rodents have identified neurons in the entorhinal cortex that are called grid cells. These cells, discovered by Edvard and May-Britt Moser and their colleagues at the Norwegian University of Science and Technology (Box 24.3), are also spatially selective. However, unlike place cells, grid cells respond when the animal is at multiple locations that form a hexagonal grid (Figure 24.23). Cells in different portions of entorhinal cortex differ in the spacing between “hotspots” in the grid, but the sensitivity grid for each cell tiles the environment the rodent is in.

FIGURE 24.23 A rat place cell and a grid cell. The black lines show the path a rat took through a square enclosure. Red spots indicate locations of the rat associated with neural activity. (a) A place cell in the hippocampus responds when the rat is in one region of the enclosure. This is the cell’s place field. (b) A grid cell in entorhinal cortex is active when the rat is at multiple locations that form a grid pattern. (Source: Moser et al., 2008, Fig. 1.) Description
We both grew up on remote islands off the west coast of Norway, a couple of hundred miles north of Bergen. This was not exactly a center of academic ferment or intellectual competition. Still, our scientific interests were nurtured by parents who’d had no opportunity to get an education themselves. We went to the same high school but didn’t really get to know each other until we met again at the University of Oslo in the 1980s.
With no clear career plan, and with different scientific backgrounds, we met in an undergraduate course in psychology. Psychology kindled and reinforced our fascination with the brain, and we jointly decided to learn more about the neural basis of behavior. The university had no neuroscience curriculum at the time, but Carl-Erik Grenness, who taught an undergraduate course on behavior analysis, alerted us to the pioneering work on brain–behavior relationships taking place at this time. He also gave us a copy of a special issue about the brain published by Scientific American in 1979. During our wandering in wilderness, this was like manna from heaven. The issue conveyed the enthusiasm of the field and strongly attracted us to this evolving scientific discipline. Among the advances reviewed there were Kandel’s demonstration of synaptic mechanisms of memory in Aplysia californica and Hubel and Wiesel’s characterization of the mechanism for feature analysis in the visual cortex.
Grenness also sent us to Terje Sagvolden, the only psychologist at the university with research projects in neuroscience at that time. We worked on neurochemical mechanisms of attention deficit disorder for two years, in parallel with studies in psychology, and learned the basics of animal behavior and experimental design. This triggered our interest in animal learning. That brought us to visit Per Andersen, the grand neurophysiologist of Norway. We sat there for hours, trying to persuade him to take us on as graduate students. He really couldn’t get us out of his office, and we wouldn’t take no for an answer. In the end, he yielded to our combination of furious curiosity and unwavering determination and took us on.
Per Andersen became our Ph.D. supervisor and introduced us to the mysteries of the brain. We learned to focus on basic questions with broad implications. Through Per, we came in touch with Richard Morris at the University of Edinburgh and John O’Keefe at University College London. Richard and John were the best mentors we could have had. They guided us into the mysteries at the intersection of behavior and neuroscience. During our studies we visited Richard several times to participate in work on the functions of the hippocampus and the role of hippocampal long-term potentiation in memory formation. After our Ph.D. defense late in 1995, we spent a few very rewarding months with John to learn place cell recording in the hippocampus. This was probably the most intense learning experience in our lives. But then, in 1996, we were almost ambushed with a job offer in Trondheim. We could not move there if only one of us got a job, so we negotiated two jobs as well as equipment to start up a new lab. We literally started our lab in a bomb shelter in a basement at the university. Our few months of postdoctoral experience had been brief, but with a decent start-up package we now had the opportunity to combine what we had learned about animal behavior and neurophysiology, fulfilling our dream from the early 1980s. We began recording the activity of individual cells in the hippocampus, with electrodes implanted in the brains of rats as they roamed a square black box.
Our start-up in Trondheim was tough but enjoyable. There were no animal housing facilities, no workshops, and no technicians. We did all the work on our own; we cleaned rat cages, changed bedding, sliced brains, and repaired cables. Starting from scratch gave us the opportunity to shape the lab exactly as we wanted it.
As we began, we got a grant from the European Commission to coordinate a consortium of seven groups that collectively aimed to perform one of the first integrated neural network studies of hippocampal memory. This was virgin territory in the late 1990s. One of the aims was to determine how the position code of the hippocampus is computed. It had been known since John O’Keefe’s studies in 1971 that the hippocampus has place cells that fire if and only if an animal is in a certain place. But it was unclear whether those place signals originated in the hippocampus itself or came from the outside. To address this question, we made intrahippocampal lesions that disconnected the output stage of the circuit—CA1—from the earlier stages. To our surprise, this did not abolish place coding in CA1. Then we had to grapple with the idea that the spatial signal might originate from somewhere else, most likely the surrounding cortex, through connections that went around the intrahippocampal circuit. The most likely candidate was the entorhinal cortex, a cortical region with major direct connections to the CA1 area of the hippocampus.
We started recording in this region, with invaluable help from Menno Witter, a neuroanatomist who was then located at the Free University of Amsterdam but later moved to become a part of the Kavli Institute in Trondheim. Witter had by that time worked out much of the connectivity between the entorhinal cortex and hippocampus and helped us in the delicate task of guiding electrodes to the right spot. By 2002, our research group had grown, and we now had an outstanding team of students working side by side with us in the lab and on the computer.
Sometimes scientific discoveries are portrayed as “Eureka” moments, where the researcher suddenly understands the significance of what he or she has found. In our case, it didn’t quite work that way: We didn’t immediately realize that the cells we recorded from were grid cells. At first we noticed that many entorhinal cells spiked every time a rat went to a particular spot, like the place cells in the hippocampus. However, each cell had multiple firing locations. After seeing the firing locations in sufficiently large environments, we became convinced that those locations formed a peculiarly regular pattern—a hexagonal grid—much like the arrangement of marbles in a Chinese checker board. Every cell did it this way, with actual firing locations differing between cells. The cells were organized topographically in the sense that the size of and distance between grid fields increased from dorsal to ventral. Moreover, cells maintained firing relationships from one environment to the next, suggesting that we were on track of a universal type of spatial map, a map whose activity pattern in many ways disregarded the fine details of the environment. With their strict regularity, the cells had the metrics of the spatial map that had not been found in the hippocampus.
These discoveries were published in a series of papers that began in 2004, only two years after we published the hippocampal disconnection study. The grid pattern itself was published in 2005. Since then we have continued to explore how grid cells operate, how they are generated, and how they interact with other spatial cell types. There is still a lot to find out. Grid cells have helped us better understand the neural representation of space, but they also provide a window into some of the innermost workings of the brain. Perhaps the most fascinating thing is that the hexagonal pattern is generated by the cortex itself. There is no grid pattern in the outside world; this is made by the brain alone. Because the pattern is so reliable and so regular, it may put us on the track of understanding the fundamental computations of the cortex.
A recent experiment suggests that there may also be grid cells in human entorhinal cortex. If you imagine drawing lines through the centers of grid cell hotspots in Figure 24.23, you will notice that along some axes you can connect many of the hotspots, such as with diagonal lines from the lower left to the upper right. If the drawn line is then rotated around the clock, there will be a periodic variation in the number of hotspots connected. This suggests that if a rat or human were to walk in various directions, grid cells would be activated more often and there would be more overall activity in the entorhinal cortex in some directions than others. This idea was tested by Christian Doeller, Caswell Barry, and Neil Burgess at University College London by having human subjects navigate a virtual reality game while fMRI images of their brain activity were recorded. The recordings made in this clever experiment showed a sinusoidal variation in the size of the fMRI signal from the entorhinal cortex when subjects navigated in different directions around the clock. This implies the presence of human grid cells that have their hotspots aligned similarly in space.
Recall that the entorhinal cortex provides input to the hippocampus. Models suggest that place fields in the hippocampus may result from summation of inputs from grid cells. The single place field of a hippocampal neuron would be the location at which the grid locations of multiple input grid cells align. Like place cells, grid cells continue to fire when the animal is at the same grid locations even when the lights are turned off. This suggests that rather than being simply a sensory receptive field, the cell’s response is based on where the animal thinks it is. Taken together, place cells, grid cells, and other hippocampal system neurons showing sensitivity for head direction make a compelling case that this brain region is highly specialized for spatial navigation.
Hippocampal Functions Beyond Spatial Memory. From our discussion of the hippocampus to this point, it may seem that its role is easily defined. First, we saw that performance in a radial arm maze, which requires memory for the locations of arms already explored, is disrupted by hippocampal lesions. Second, the responses of place cells in the hippocampus, considered along with grid cells in the entorhinal cortex, suggests that these neurons are specialized for location memory. This is consistent with the cognitive map theory proposed by O’Keefe and his colleague Lynn Nadel, which states that the hippocampus is specialized for creating a spatial map of the environment. In one sense it is undeniable that the hippocampus, at least in rats, seems to play an important role in spatial memory.
Others argue, however, that this is not the only or best description of what the hippocampus does. In his original studies using the radial arm maze, Olton described the result of hippocampal lesions as a deficit in working memory. The rats were not able to retain recently acquired information concerning arms already explored. Thus, working memory might be one aspect of hippocampal function. This would explain why the rats with lesions could avoid going down arms that never contained food but still not remember which arms they had recently visited. Presumably after training, the information about no-food arms was saved in long-term memory, but working memory was still required to avoid the arms where food had already been retrieved.
Other theories are built on observations that the hippocampus integrates or associates sensory input of behavioral importance. For example, as you read this book, you may form memories relating multiple things: specific facts, illustrations that catch your eye, interesting passages, the arrangement of material on the page, and information about the sounds or events going on around you as you read. Maybe you’ve tried finding a passage in a book by searching for a page that looks a particular way. Another common example is how remembering one thing, such as the theme song to an old television show, can bring back a flood of related associations—the characters in the show, your living room at home, the friends you watched with, and so on. Interconnectedness is a key feature of declarative memory storage.
Odor discrimination provides an example of the involvement of the hippocampus in tasks that are not entirely based on spatial memory. In one such task, a rat’s cage had two ports at which the animal smelled two different odors (Figure 24.24). For each pair of odors, the animal was trained to go toward the port releasing one odor and to avoid the other port. The researchers found that some neurons in the hippocampus became selectively responsive for certain pairs of odors. Moreover, the neurons were particular about which odor was at which port; they would respond strongly with odor 1 at port A and odor 2 at port B, but not with the odors switched to the opposite ports. This indicates that the response of the hippocampal neurons related the specific odors, their spatial locations, and whether they were presented separately or together. It was also shown that hippocampal lesions produced deficits on this discrimination task.

FIGURE 24.24 An odor discrimination experiment used to study relational memory. For various combinations of odors, rats were trained to move toward a port emitting one odor and avoid the other port. (Source: Adapted from Eichenbaum et al., 1988, Fig. 1.)
Let’s summarize the diverse studies of the hippocampus we’ve discussed. First, research since the time of H.M. indicates that the hippocampus is critical for memory consolidation of facts and events. Strong evidence suggests that in rodents, and in people, the hippocampus is especially important for spatial memory. In recordings from human hippocampal neurons there is sometimes surprising selectivity for people or objects we are familiar with. Finally, hippocampal cells appear to form associations between sensory stimuli even when the information is not about space. A thread that runs through these various studies is that the hippocampus links different experiences together. It receives a huge spectrum of sensory inputs and may construct new memories by integrating the varied sensory experiences associated with an event (e.g., the theme music of a television show is integrated with memories of people and place). The hippocampus may also be essential for building or enhancing memories by connecting new sensory input with existing knowledge. It has been suggested that the input from the grid cells in the entorhinal cortex provides “where” information to the hippocampus, while other input contains information about “what.” The neural associations constructed and then consolidated in the hippocampus might effectively establish memories for “what happened where.”
There is compelling evidence that declarative memory formation involves a system of interconnected brain structures that take in sensory information, make associations between related information, consolidate learned information, and store engrams for later recall. Components of this system include the hippocampus, cortical areas around the hippocampus, the diencephalon, neocortex, and more. The questions we want to look at now concern the timing of these things: When and where are memories stored in a permanent form? How long does it take memories to become “permanent”? Do engrams change location over time? Can subsequent experiences alter, enhance, or degrade memories?
Standard and Multiple Trace Models of Consolidation. Since the time of H.M., a view of memory consolidation and storage has developed that has come to be called the standard model of memory consolidation. In this model, information comes through neocortex areas associated with sensory systems and is then sent to the medial temporal lobe for processing (especially the hippocampal system). As we will discuss in more detail in Chapter 25, changes in synapses create a memory trace via a process sometimes called synaptic consolidation (Figure 24.25a). After synaptic consolidation, or perhaps overlapping with it in time, systems consolidation occurs in which engrams are moved gradually over time into distributed areas of the neocortex (Figure 24.25b). It is in a variety of neocortical areas that permanent engrams are stored. Before systems consolidation, memory retrieval requires the hippocampus, but after systems consolidation is complete, the hippocampus is no longer needed.

FIGURE 24.25 Two models of memory consolidation. (a) In both standard and multiple trace models, memory formation is initiated by synaptic changes in the hippocampus. In this schematic, the hippocampal neurons interact with neurons in three distributed areas of neocortex. (b) In the standard model, a temporary memory trace is formed in the hippocampus through synaptic consolidation and engrams later develop in neocortex through systems consolidation. Over time, memory depends more on connections in neocortex (solid lines) and less on the hippocampus (dashed lines). (c) In the multiple trace model, engrams for episodic memories always involve both the hippocampus and neocortex (all lines are solid). The red and green lines indicate two traces for the same memory that were made in somewhat different sensory contexts. Description
Many observations about memory formation are consistent with the standard model, but questions have been raised about whether it provides the most accurate account of consolidation. A key point is the duration of retrograde amnesia. For example, early accounts of H.M.’s amnesia reported that his retrograde amnesia extended back a few years. An interpretation of this observation is that synaptic consolidation is complete quickly but systems consolidation takes years to complete and the retrograde memories H.M. lost were ones not “fully baked” (i.e., engrams still dependent on the hippocampus). Later studies examined H.M.’s retrograde amnesia in more detail and found that it extended back decades. Perhaps systems consolidation is a very slow process that takes decades. However, some scientists have asked whether this makes sense in a species whose individuals, not so long ago, lived only for a few decades. As if that question isn’t confusing enough, it should be noted that later studies of H.M. suggested that he had retrograde amnesia for episodic memories that extended back virtually his entire life. This implies that the hippocampus, perhaps in concert with other medial temporal structures, may be involved with memories for a lifetime.
Alternatives to the standard model have been proposed, most notably the multiple trace model of consolidation proposed by Lynn Nadel of the University of Arizona and Morris Moscovitch of the University of Toronto. The multiple trace model was proposed as a way to avoid the necessity of the decades-long systems consolidation process the standard model needs to account for extended retrograde amnesia. If hippocampal damage disrupts episodic memories going back decades or a lifetime, perhaps the hippocampus is always involved in memory storage. In other words, systems consolidation does not ever relinquish engrams entirely to neocortex.
According to this theory, engrams involve neocortex, but even old memories also involve the hippocampus (Figure 24.25c). The term “multiple trace” refers to the way the model allows for retrograde amnesia resulting from hippocampal damage to sometimes be graded in time. The hypothesis is that each time an episodic memory is retrieved, it occurs in a context different from the initial experience and the recalled information combines with new sensory input to form a new memory trace involving both the hippocampus and neocortex. This creation of multiple memory traces presumably gives the memory a more solid foundation and makes it easier to recall. Because retrieval requires the hippocampus, complete loss of the hippocampus should cause retrograde amnesia for all episodic memories no matter how old. If there is partial damage, then the memories that are intact would be the ones with multiple traces. To the extent that older memories would have been recalled more times than recent memories, they would be more likely to survive hippocampal damage, and this would give rise to a temporal gradient in retrograde amnesia. Suffice it to say that at present experts disagree about assessments of gradients in retrograde amnesia and the validity of various models of consolidation.
Reconsolidation. In 1968 a report published by James Misanin, Ralph Miller, and Donald Lewis of Rutgers University made the surprising claim that memories can be altered or selectively erased even after they are consolidated. From the perspective of the standard model, this is unexpected because memories should be sensitive to disruption, such as by electrical convulsive shock (ECS), only before consolidation. After consolidation, memories should be stable and fixed. Indeed, varying the time between an experience and ECS has been used to measure how long consolidation takes. In an experiment by Misanin and colleagues, rats were trained by presenting a loud noise followed by a foot shock (don’t confuse the foot shock with the ECS to the head). This is an example of classical conditioning in which the foot shock is the unconditional stimulus (US) and the loud noise is the conditional stimulus (CS). Trained rats subsequently fear a shock when they hear the noise. After the loud noise, the unconditioned rats licked rapidly from a drinking tube, but the trained animals drank much slower, an indication that they feared the foot shock. If an animal was conditioned but immediately given electroconvulsive shock, it would lick quickly the next day, just as if it had never been conditioned. This is evidence of retrograde amnesia for the conditioning. If there was a delay of 24 hours between the conditioning and the electroconvulsive shock, there was no fear attenuation. Apparently by 24 hours the fear memory had been consolidated and amnesia for the conditioning could no longer be caused by ECS.
On the second day of the experiment, presumably after consolidation of the fear memory, some of the conditioned animals were presented the loud noise immediately followed by the ECS. On the third day of the experiment the rats were tested once more. Surprisingly, the rats that received the noise–foot shock combination on the second day licked as much as rats that had received ECS on the first day immediately after training (i.e., the animals with amnesia that didn’t fear the loud noise). They also licked much more than rats given ECS on the second day but without the loud noise. What these results suggest is that the sound on the second day reactivated the fear memory, and, once reactivated, this memory was wiped out by ECS. If the memory was not reactivated (the animals that got ECS on the second day but without the loud sound), then ECS had no effect. The implication of the study is very important because it suggests that reactivating a memory makes it sensitive just as it had been immediately after the memory was first formed (before consolidation). For this reason the reactivation effect is called reconsolidation.
In recent years there has been increased interest in reconsolidation, which appears to also occur in human memory. The original rat experiment on reconsolidation involved procedural learning (classical conditioning), but it now appears that reconsolidation is also seen with human episodic memory. In one experiment, college students were instructed to memorize 20 objects (e.g., balloon, crayon, spoon) that they were shown one at a time and then placed in a basket. Each student practiced naming all the items until they remembered at least 17 of them from this day 1 list. One day later, presumably after memory consolidation, some of the students were given a reminder by showing them the empty basket and asking them to describe what happened the day before without naming the objects. The aim was to subtly reactivate the memories, possibly opening them up to reconsolidation. A control group of students did not receive this reminder, with the hope of not reactivating their day 1 memories. Both the reminded and the not-reminded groups were then asked to memorize a second group of 20 objects on day 2. Finally, on day 3 of the experiment, all the subjects were asked to recall the objects from the first group on day 1 of the study.
On average, subjects recalled about eight of the objects from day 1 regardless of whether they had received a reminder on day 2. The interesting finding was that the subjects given the reminder on day 2 “accidentally” included numerous objects from day 2 in their recall (five objects on average), whereas these intrusions rarely occurred in subjects not given the reminder on day 2. Based on that experiment and several other variations, it appeared that the reminder may have served to reactivate the consolidated day 1 memories, making them labile again. These memories were then reconsolidated on day 2, erroneously mixing in new sensory information associated with the day 2 objects. A variety of experiments looking at reconsolidation in humans have been performed and have even shown hippocampal activity in the reactivation condition. Evidently, when we recall a memory, it becomes susceptible to change and reconsolidation. This fascinating finding has profound implications for the treatment of post-traumatic stress associated with unpleasant memories and even for the reliability of our normal recollections (Box 24.4).
Introducing False Memories and Erasing Bad Memories
If reconsolidation can alter existing memories, how can we ever be sure what we remember is correct? It may sound like science fiction, but it has been reported that memories already consolidated can be altered and memories for things that never happened can be introduced into the brain.
Experiments testing the ability to tinker with memories have largely been conducted with mice. In a frightening situation a mouse will “freeze” in place, a reaction that is presumably a form of “playing dead” to avoid detection or consumption by a predator. In a laboratory experiment a mouse is placed in a box that the animal can recognize based on its visual appearance and smell. When the mouse enters the box it receives an electrical foot shock. If the mouse is removed from the box and returned to it the next day, it will freeze when it recognizes the box, in anticipation of a foot shock even when none is given. The animal does not freeze when placed in a distinctly different box, indicating that the animal has formed a memory associating the shock with only the box where it received the shock.
A team of scientists at the Massachusetts Institute of Technology recently used transgenic mice in a clever variation on this fear-conditioning experiment to study the malleability of memories. The mice explored a box, and a small percentage of hippocampal neurons became activated by the visual and olfactory experiences associated with the box. Critically, in these transgenic animals the scientists had the ability to essentially turn a switch on and off chemically so that active neurons would or would not express channelrhodopsin-2 (ChR2), as described in Chapter 4. Neurons with ChR2 could then be activated at a later time by exposing them to blue light. With these abilities in place, the experimental procedure went like this (Figure A):

- On day 1, expose animals to box A with the label-active-cells switch on so that neurons activated by the box A sensory stimulation express ChR2. No foot shock is given, and animals do not freeze in box A.
- On day 2, expose animals to the different sensory context of box B. On this day the cell label switch is off, so ChR2 is not expressed in the hippocampal neurons activated by the box B context. While the animals are in box B, blue light was delivered to the hippocampus through a fine fiber optic cable, which reactivated the neurons from the previous day that encoded sensory information associated with box A. At the same time, a foot shock was delivered. Remember, this all occurs when the animal is in box B. The hypothesis was that the reactivated memory of box A would be reconsolidated on the second day in a manner that links in the painful foot shock.
- The moment of truth! On day 3 the mouse is put back in box A, and as predicted, it freezes even though it has never actually experienced a foot shock in box A. When the animal is placed in an unfamiliar context, box C, it does not freeze.
It appears that a false memory has been created such that the animal fears box A even though the electrical shock occurred only in box B. The lack of freezing behavior in box C indicates that the false memory is specific to box A, presumably because neurons encoding box A information were reactivated by the blue light while the animal was in box B. You have probably heard of people convicted of crimes based on eyewitness testimony and the convicted person is later released from prison based on DNA evidence proving their innocence. Evidently the memory of the eyewitness had been wrong. Might this happen in some cases because witnesses are coached and the coached information interacts and reconsolidates with the events of the recalled crime? Ongoing research is examining the conditions under which reconsolidation can occur; the results have important implications for the judicial system and our ability to trust our own memories.
If we are able to alter memories after they have been consolidated, perhaps a process can be devised to treat people with memories that haunt them. We all have embarrassing moments we would like to forget, but some people have memories that are so disturbing that they interfere with daily life. In post-traumatic stress disorder (PTSD), an earlier traumatic event has serious deleterious effects on later behavior, mood, and social interactions, even in nonthreatening situations. An example is a war veteran who experiences stress and fear in daily life long after the war is over. What if there was a way to erase or at least weaken such unpleasant memories? Studies exploring this question suggest that it might be possible.
One approach takes advantage of the observation that administration of the beta-adrenergic antagonist propranolol shortly after a traumatic event will reduce physiological responses (e.g., heart rate) to later recollection of the event. It is thought that propranolol may counter the effect of stress hormones usually activated by the fearful experience. Unfortunately, it is generally not possible to mediate with someone immediately after a traumatic experience. The important question for a potential PTSD treatment is whether it is possible to make use of reconsolidation to weaken the traumatic memory well after the event. In one study looking into this question, people suffering from chronic PTSD were instructed to describe the trauma they experienced. They were simultaneously given either propranolol or a placebo. One week later when they were asked to recall their traumatic event, physiological responses were lower in the people given propranolol than in the placebo group. Perhaps the drug administration at the time of memory reactivation led to a reconsolidated memory with weakened emotional impact. Note that in this case, the propranolol treatment affected the emotional weight of the memory but not the declarative memory itself.
We don’t know if mice experience PTSD, but a recent study by Tsai and her colleagues at MIT attempted to reduce an unpleasant memory in mice by targeting neural plasticity rather than affecting whole-body physiology as with propranolol. As in experiments we have already examined, mice are made to fear a loud sound by pairing the sound with an electrical foot shock. When mice later hear the sound they freeze in fear even if the foot shock is not given. The usual way to decrease the strength of the fear reaction is to repeatedly expose the mice to the sound without giving an electrical shock (similar to the treatment humans with PTSD are given in which they recall a traumatic memory in a safe environment). This extinction therapy in the mice is found to reduce or eliminate fear associated with the chamber if the therapy is started one day after the traumatic experience but not 30 days later. With an eye toward the delayed treatment of PTSD in humans, Tsai et al aimed to reduce the memory in mice a month after the electrical shock, a time at which extinction therapy alone is ineffective. This was accomplished by combining the fear-inducing sound with administration of a drug that inhibits the HDAC2 (histone deacetylase 2) enzyme. This enzyme, which switches neuroplasticity genes off in the nucleus of neurons (discussed in Chapter 25), was found to be inactive the day after the electrical shock but active a month later. By inhibiting HDAC2, the plasticity genes were activated at the later time. With the genes active and the traumatic memories revived by the loud sound, it was possible to reconsolidate the memory in a less fearful form. After just one dose, the mice no longer froze when the sound was heard. We don’t know if this or a related approach will work to treat human PTSD, but there is hope that memory reconsolidation might be of use in alleviating this disorder.
Brunet A, Orr, SP, Tremblay J, Robertson K, Nader K, Pitman RK. 2008. Effect of post-retrieval propranolol on psychophysiologic responding during subsequent script-driven traumatic imagery in post-traumatic stress disorder. Journal of Psychiatric Research 42:503–506.
Graff J, Joseph NF, Horn, ME, Samiei A, Meng J, Seo J, et al. 2014. Epigenetic priming of memory updating during reconsolidation to attenuate remote fear memories. Cell 156:261–276.
Ramirez S, Liu X, Lin P, Suh J, Pignatelli M, Redondo RL, et al. 2013. Creating a false memory in the hippocampus. Science 341:387–391.
Thus far, we have focused on the brain systems involved in the formation and retention of declarative memories, partly because declarative information is what we usually mean when we say we remember something. In addition, however, the neural basis of nondeclarative memory is complex because different types seem to involve different brain structures. As indicated in Figure 24.1, various kinds of nondeclarative memory are thought to involve different parts of the brain. As an example of nondeclarative memory, we will take a look at evidence supporting the involvement of the striatum in habit learning and procedural memory.
Recall from Chapter 14 that the basal ganglia are important for the control of voluntary movements. Two elements of the basal ganglia are the caudate nucleus and the putamen, together called the striatum. The striatum sits at a key location in the motor loop, receiving input from frontal and parietal cortex and sending output to thalamic nuclei and cortical areas involved in movement. Several lines of evidence in studies of rodents and humans suggest that the striatum is critical for the procedural memory involved in forming behavioral habits.
The amnesia experienced by H.M. was surprising, in part, because he was able to learn new habits despite his complete inability to form new declarative memories. Indeed, this is one of the most compelling reasons for hypothesizing that procedural memory uses distinct circuitry. In the monkey model of amnesia, we saw that the formation of new declarative memories could be disrupted by making small lesions in the rhinal cortex of the medial temporal lobe. Such a lesion had relatively little effect on procedural memory, which raises an obvious question: Are there comparable lesions that disrupt procedural memory without affecting declarative memory? In rodents, lesions to the striatum have this effect.
In one study, rats had to learn two versions of the radial arm maze task. The first was the standard version described earlier, in which the rat learned to move as efficiently as possible when retrieving the food from each of the baited arms of the maze. In the second version, small lights were illuminated above one or more arms containing food, and the unlit arms had no food. The lights could be turned on or off at any time. In this case, optimal performance meant that the animal kept returning to retrieve food from lit arms as long as they were lit, and avoided arms that were never lit. The standard maze task was designed to require the use of declarative memory. The “light” version of the task was intended to draw on procedural memory because of the consistent association between the presence of food and the illuminated lights. The rat did not have to remember which arms it had already explored; it simply had to form a habit based on the association of the light with food. The rat’s performance on the light task was analogous to the habits H.M. was able to form, such as mirror drawing.
Performance on the two versions of the radial arm maze task were affected in markedly different ways by two types of brain lesions. If the hippocampal system was damaged (in this case, with a lesion to the fornix that sends hippocampal output), performance was degraded on the standard maze task but was relatively unaffected on the light version. Conversely, a lesion in the striatum impaired performance of the light task but had little effect on the standard task. This “double dissociation” of the lesion site and the behavioral deficit suggests that the striatum is part of a procedural memory system but is not crucial for the formation of declarative memories.
Recordings made from the rat striatum in other experiments showed that neural responses changed as the animals learned a procedure associated with a food reward. In a simple T-maze task, rats were placed at the end of one arm of the T and, as they moved away from the arm’s end, a tone sounded (Figure 24.26a). A low tone was used to train the animal to turn left into the next arm of the T to get a chocolate snack, and a high tone was used to train the animal to turn right for the reward. Figure 24.26b shows the percentage of neurons that responded when the animal was in several phases of the task: start position, tone sounding, turning into reward arm, and reaching the reward.

FIGURE 24.26 Changing responses in rat striatum during the learning of a habit. (a) The rat began at the end of the long arm of a T-maze and turned left or right depending on the pitch of a tone. (b) The percentage of neurons that responded during several phases of the task: start position, tone sounding, turning into reward arm, and reaching the reward (goal). Each stage of training and testing consisted of 40 runs through the maze. Over the stages of learning and mastering the maze, more cells responded to the start and the goal and fewer to the turn. (Source: Adapted from Jog et al., 1999, Figs. 1 and 2.) Description
When the rats first performed the task (stage 1), the highest percentage of neurons responded when the animals turned into the reward arm. However, in later stages of the experiment, as training and testing progressed, this percentage decreased significantly. As the rats mastered the procedure, increasingly more neurons became responsive at the start and completion of the task. Also, increasing numbers of neurons responded during more than one stage of the task. One possible interpretation of these changes in response patterns is that they reflect the formation of a habit for which the striatum codes a sequence of behaviors initiated in the T-maze situation. At present this is only a hypothesis, but it is intriguing because of the connectivity of the striatum—taking in highly processed sensory information and sending out signals involved in motor responses.
Studies with monkeys indicate that the effects of selective brain lesions on memory are comparable in rodents and primates. In primates, there is a similar dissociation between the effects of lesions to the hippocampal system and the striatum. As we have already seen, lesions to the medial temporal lobe significantly impair performance on the delayed non-match to sample task that uses declarative memory. However, consider another task in which the animal repeatedly sees two visual stimuli, such as a square and a cross, and must learn to associate a food reward with only the cross (i.e., instrumental conditioning). This kind of habit learning is relatively unaffected by medial temporal lesions. Such preservation of habit learning in monkeys is analogous to the rat’s ability to retrieve food consistently associated with a light above an arm of a maze, even after a fornix lesion.
In monkeys, lesions that involve the striatum or connections to it have quite different effects from medial temporal lesions. When the striatum is damaged, there is no effect on the performance of the DNMS task, demonstrating both that declarative memory formation is still possible and that the animal can discriminate visual stimuli. But when the striatum is damaged, the animal is unable to form the habit of always retrieving food associated with one visual stimulus rather than another. Repeated exposure to this fixed stimulus–reward situation just doesn’t seem to sink in. Thus, there appear to be somewhat distinct anatomical systems for declarative memory and procedural memory, and behaviors such as learned habits utilize the striatum.
Several diseases in humans attack the basal ganglia, and certain of their effects on memory appear consistent with the striatum’s role in procedural memory. For example, Huntington’s disease kills neurons throughout the brain, but the striatum is a focus of the attack. Huntington’s patients have been shown to have difficulty learning tasks in which a motor response is associated with a stimulus. Although these people generally have motor dysfunction, difficulty in learning the stimulus–response habit does not correlate with the severity of the motor deficits, suggesting that it is an independent consequence of the disease.
Further evidence for the involvement of the striatum in habit learning comes from comparisons of patients with Parkinson’s disease with amnesic patients. As we saw in Chapter 14, Parkinson’s disease is characterized by degeneration of the substantia nigra inputs to the striatum. In one study, people were tested on two tasks. In the first task, patients saw one, two, or three out of four possible cues in one of 14 possible combinations. They then had to guess whether this combination was arbitrarily associated with a prediction of sunny or rainy weather (Figure 24.27a). For each patient, the experimenter assigned different probabilities that various cues were associated with sun or rain. By being told when they guessed correctly or incorrectly about the predicted weather, the patients slowly built up an association between the cues and the weather. The idea behind this task was that it draws on the formation of a stimulus–response habit. In the second task, declarative memory was tested by having patients answer multiple-choice questions about the appearance of the cues and the computer screen.

FIGURE 24.27 The performance of patients with amnesia and Parkinson’s disease on two memory tasks. (a) Four cue cards were presented in various combinations associated with the icons indicating sun or rain. Based on repeated exposure to the combinations, patients had to learn to predict sun or rain by inferring the associations. (b) With successive trials, control subjects and amnesic patients improved on the association task. Parkinson’s patients showed little improvement. (c) On a test of declarative memory formation (a questionnaire), Parkinson’s patients performed similar to control subjects, and amnesia patients were greatly impaired. (Source: Adapted from Knowlton et al., 1996, Figs. 1 and 2.) Description
The Parkinson’s patients had great difficulty learning the weather forecasting task (Figure 24.27b) but performed at normal levels on the declarative memory questionnaire (Figure 24.27c). Conversely, amnesic patients had no trouble learning the weather classification, but they performed significantly worse than either Parkinson’s patients or normal controls on the questionnaire. These results suggest that the striatum in humans may play a role in procedural memory as part of a system distinct from the medial temporal system used for declarative memory.
Far from being a computer with fixed connections, the human brain is constantly changing as a result of experience. We use working memory to temporarily hold onto information, and the patterns of sensory input from some of our experiences are assembled into permanent engrams. As a child you learned to do a summersault, and the sequence of movements was unconsciously stored for use time after dizzying time. You learned the structure of the brain and are able to impress Aunt Tilly by making a sketch showing the location of the medulla oblongata. We cannot identify the precise neurons and synapses involved in storing nondeclarative or declarative memories, but research is moving us closer to such an understanding. We know that learning and memory involve changes widely across the brain. Structures in the medial temporal lobe and diencephalon are critical for memory consolidation, and engrams are stored in the neocortex through interactions with the hippocampus and other structures. Specifying precisely what each brain structure contributes to learning and memory continues to challenge researchers.
We have seen that memories can be classified based on duration, the kind of information stored, and the brain structures involved. Early brain research relied on interpreting the effects of brain lesions on amnesia. From the case of H.M. alone, a tremendous amount was learned about memory in the human brain. The distinct types of memory, and the fact that one type can be disrupted without affecting others in amnesia, indicate that multiple brain systems are used for memory storage. More recent research uses human brain imaging and molecular genetic techniques to examine memory formation and sort out the temporal processes and multiple systems. There is even hope that one day there will be a treatment to significantly reduce the deleterious consequences of traumatic memories.
In this chapter we have focused on questions about where memories are stored and how different brain structures interact. But what is the physiological basis for the memory storage? When we try to remember a phone number, an interruption can make us forget, suggesting that memories are initially held in a particularly fragile form. Long-term memory is much more robust; it can survive interruption, anesthesia, and the normal bumps and traumas of life. Because of this robustness, it is thought that memories are ultimately stored in structural brain changes. The nature of these structural changes in the brain is the subject of Chapter 25.
1. If you try to recall how many windows there are in your house by mentally walking from room to room, are you using declarative memory, procedural memory, or both?
2. What kind of experiment might you conduct to find the place in the brain that people use to hold a phone number in mind?
3. In what brain areas have neural correlates of working memory been observed?
4. What structures in the medial temporal lobe are thought to be involved in memory?
5. Why did Lashley conclude that all cortical areas contribute equally to learning and memory? Why was this conclusion later called into question?
6. What arguments can you think of for and against the idea that Wilder Penfield’s electrical brain stimulation evoked memories?
7. What evidence is there that declarative and nondeclarative memory use distinctly different circuits?
8. In the famous amnesic known as H.M., what types of memory were lost after temporal lobe surgery? What kinds were preserved?
9. What are place cells and grid cells? Where have they been observed?
10. What evidence indicates that long-term memories are stored in the neocortex?
11. The multiple trace model of memory consolidation was proposed to deal with what concern(s) about the standard model of memory consolidation?
12. Where is it thought that procedural memories are stored?
Corkin S. 2013. Permanent Present Tense: The Unforgettable Life of the Amnesic Patient H.M. New York: Basic Books.
Kandel ER, Dudai Y, Mayford MR. 2014. The molecular and systems biology of memory. Cell 157:163–186.
Ma WJ, Husain M, Bays PM. 2014. Changing concepts of working memory. Nature Neuroscience 17:347–356.
McKenzie S, Eichenbaum H. 2011. Consolidation and reconsolidation: two lives of memories? Neuron 71:224–233.
Moser EI, Kropff E, Moser M. 2008. Place cells, grid cells, and the brain’s spatial representation system. Annual Review of Neuroscience 31:69–89.
Nadel L, Hardt O. 2011. Update on memory systems and processes. Neuropsychopharmacology 36:251–273.
Quiroga RQ, Kreiman G, Koch C, Fried I. 2008. Sparse but not “grandmother-cell” coding in the medial temporal lobe. Trends in Cognitive Sciences 12:87–91.
Squire LR, Wixted JT. 2011. The cognitive neuroscience of human memory since H.M. Annual Review of Neuroscience 34:259–288.
Wang S, Morris RGM. 2010. Hippocampal-neocortical interactions in memory formation, consolidation, and reconsolidation. Annual Review of Psychology 61:49–79.
Additional figures