Introduction
Chapter 2 Chemical Foundations Model of sodium chloride (NaCl) crystal. This photograph shows the largest (3.1 m tall) balland-stick physical model of a crystalline salt ever constructed, built from 40,110 balls by Robert Krickl.

2.4 Biochemical Energetics The life of a cell depends on thousands of chemical interactions and reactions exquisitely coordinated with one another in time and space, influenced by the cell’s genetic instructions and its environment. By understanding these interactions and reactions at a molecular level, we can begin to answer fundamental questions about cellular life: How does a cell extract nutrients and information from its environment? How does a cell convert the energy stored in nutrients into the work of movement or metabolism? How does a cell transform nutrients into the cellular components required for its survival? How does a cell transmit its genetic information to its progeny and use this information to survive and thrive? How does a cell link itself to other cells to form a tissue? How do cells communicate with one another so that a complex, efficiently functioning organism can develop and thrive? One of the goals of this textbook is to answer these and other questions about the structure and function of cells and organisms in terms of the properties of individual molecules and ions. For example, the properties of one such molecule, water, control the evolution, structure, and function of all cells. An understanding of biology
is not possible without appreciating how the properties of water control the chemistry of life. Life first arose in a watery environment, and water constitutes 70–80 percent of most cells by weight. In fact, water is the most abundant molecule in biological systems. The remaining mass of organisms is made up of small molecules and ions and of combinations of these that form the larger macromolecules and macromolecular assemblies that make up a cell’s machinery and architecture. The small molecules typically have molecular weights of less than 1000 daltons and include amino acids (the building blocks of proteins), nucleotides (the building blocks of DNA and RNA), lipids (the building blocks of biomembranes), and sugars (the building blocks of complex carbohydrates). The small molecules and ions together make up about 7 percent of the weight of living matter. Many of the cell’s biomolecules (such as sugars) readily dissolve in water; these molecules are referred to as hydrophilic (“water liking”). Others (such as cholesterol) are oily, fatlike substances that shun water; these molecules are said to be hydrophobic (“water fearing”). Still other biomolecules (such as phospholipids) contain both hydrophilic and hydrophobic regions; these molecules are said to be amphipathic or amphiphilic (“both liking”). The smooth functioning of cells, tissues, and organisms depends on all these molecules, from the smallest to the largest. Indeed, the chemistry of the simple proton can be as important to the survival of a human cell as that of each gigantic DNA molecule (the mass of the DNA molecule in human chromosome 1 is times that of a proton!). The chemical interactions of all these molecules, large and small, with water and with one another define the nature of life.
Luckily, although many types of biomolecules interact and react in numerous and complex pathways to form functional cells and organisms, a relatively small number of chemical principles are necessary to understand cellular processes at the molecular level (Figure 2-1).
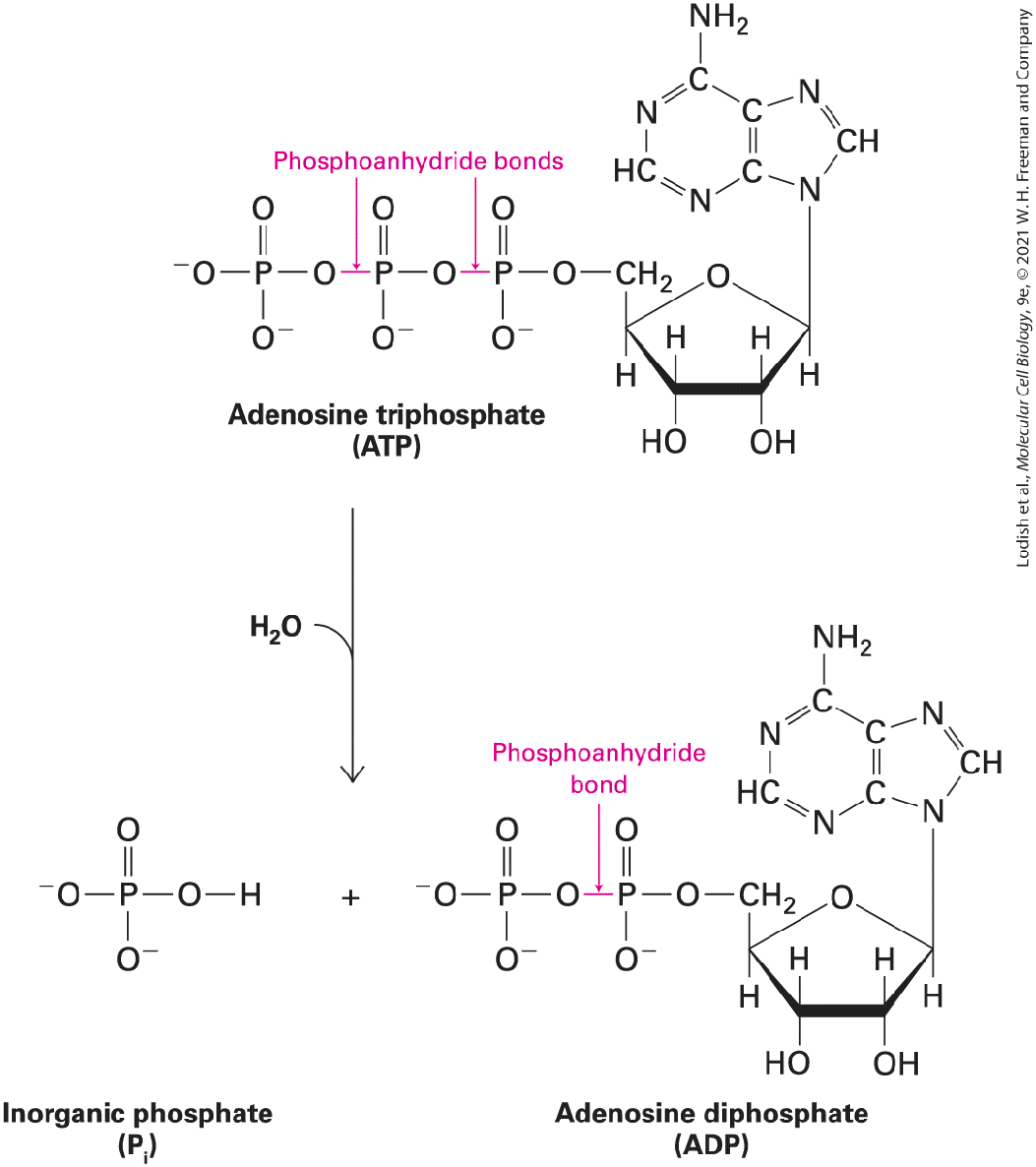
FIGURE 2-1 Chemistry of life: four key concepts. (a) Molecular complementarity lies at the heart of all biomolecular interactions (see Section 2.1), as when two proteins with complementary shapes and chemical properties come together to form a tightly bound complex. (b) Small molecules serve as building blocks for larger structures (see Section 2.2). For example, to generate the information-carrying macromolecule DNA, four small nucleotide building blocks are covalently linked into long strings (polymers), which then wrap around each other to form the double helix. (c) Chemical reactions are reversible, and the distribution of the chemicals between starting reactants (left) and the products of the reactions (right) depends on the rate constants of the forward ( , upper arrow) and reverse ( , lower arrow) reactions. The ratio of these, , provides an informative measure of the relative amounts of products and reactants that will be present at equilibrium (see Section 2.3). (d) In many cases, the source of energy for chemical reactions in cells is the hydrolysis of the molecule ATP (see Section 2.4). This energy is released when a high-
energy phosphoanhydride bond linking the β and γ phosphates in the ATP molecule (red) is broken by the addition of a water molecule, forming ADP and . Description The illustration labeled A molecular complexity, shows non-covalent interactions between proteins. The hydroxyl group of protein A is bonded to the carboxyl group of protein B, and vice versa. The illustration labeled B chemical building blocks, shows nucleotides arranging themselves to form a D N A macromolecule strand through the process of polymerization. The illustration labeled C chemical equilibrium, shows a chemical reaction in which several large and small molecules combine together. The equilibrium constant for the forward and reverse reactions is k subscript f and k subscript r, respectively. The overall equilibrium constant k subscript e q equals k subscript f divided by k subscript r. The illustration labeled D chemical bond energy, shows a 3-dimensional model of adenosine triphosphate (A T P). The bonds between alpha and beta, and beta and gamma phosphates in an A T P molecule are labeled highenergy phosphoanhydride bonds. This A T P molecules breaks to yield A D P plus P subscript i plus energy. First, biological molecules are able to bind together where their surfaces have complementary shapes and chemical properties — negatively charged surfaces with positively charged surfaces, for example (Figure 21a). Second, as we have seen, chemical building blocks assemble to form macromolecules (Figure 2-1b). Third, these assemblages are possible because molecules are able to react to form new products in chemical reactions, which can proceed to chemical equilibrium (Figure 2-1c) or steady state. Finally, molecules release the energy stored in chemical bonds in a controlled fashion to power the work of the cell (Figure 2-1d). In this chapter, we review these key principles, some of which you already know well, because a little bit of chemistry can go a very long way in helping us to understand molecular cell biology. We begin with the
covalent bonds that connect atoms into molecules and the noncovalent interactions that stabilize groups of atoms within and between molecules. We then consider the basic chemical building blocks of macromolecules and macromolecular assemblies. After reviewing those aspects of chemical equilibrium that are most relevant to biological systems, we end the chapter with the basic principles of biochemical energetics, including the central role of ATP (adenosine triphosphate) in capturing and transferring energy in cellular metabolism.
The Electronic Structure of an Atom Determines the Number and Geometry of the Covalent Bonds It Can Make
2.1 Covalent Bonds and Noncovalent Interactions Underpinning all of biology is the assembly of atoms into molecules that in turn interact with one another. Strong and weak attractive forces between atoms are the glue that holds individual molecules together and permits interactions between different molecules. When two atoms share a single pair of electrons, the result is a covalent bond — a type of strong force that links atoms together in molecules. When two atoms share multiple pairs of electrons, the result is multiple covalent bonds (e.g., double or triple bonds). The weak attractive forces of noncovalent interactions are equally important in determining the properties and functions of biomolecules such as proteins, nucleic acids, carbohydrates, and lipids. In this section, we first review covalent bonds and then discuss the four major types of noncovalent interactions: ionic bonds, hydrogen bonds, van der Waals interactions, and the hydrophobic effect. The Electronic Structure of an Atom Determines the Number and Geometry of the Covalent Bonds It Can Make
Hydrogen, oxygen, carbon, nitrogen, phosphorus, and sulfur are the most abundant elements in biological molecules. These atoms, which rarely exist as isolated entities, readily form covalent bonds, sharing electrons in the outermost electron orbitals surrounding their nuclei (Figure 2-2). As a rule, each type of atom forms a characteristic number of covalent bonds with other atoms. These bonds have a well-defined geometry determined by the atom’s size and by both the distribution of electrons around the nucleus and the number of electrons it can share. In some cases, the number of stable covalent bonds an atom can make is fixed; carbon, for example, always forms four covalent bonds. In other cases, the number of stable covalent bonds can vary; for example, sulfur can form two, four, or six stable covalent bonds.
FIGURE 2-2 Covalent bonds form by the sharing of electrons. Covalent bonds, the strong forces that hold atoms together in molecules, form when atoms share electrons from their outermost electron orbitals. Each atom forms a defined number of covalent bonds having a distinct geometry.
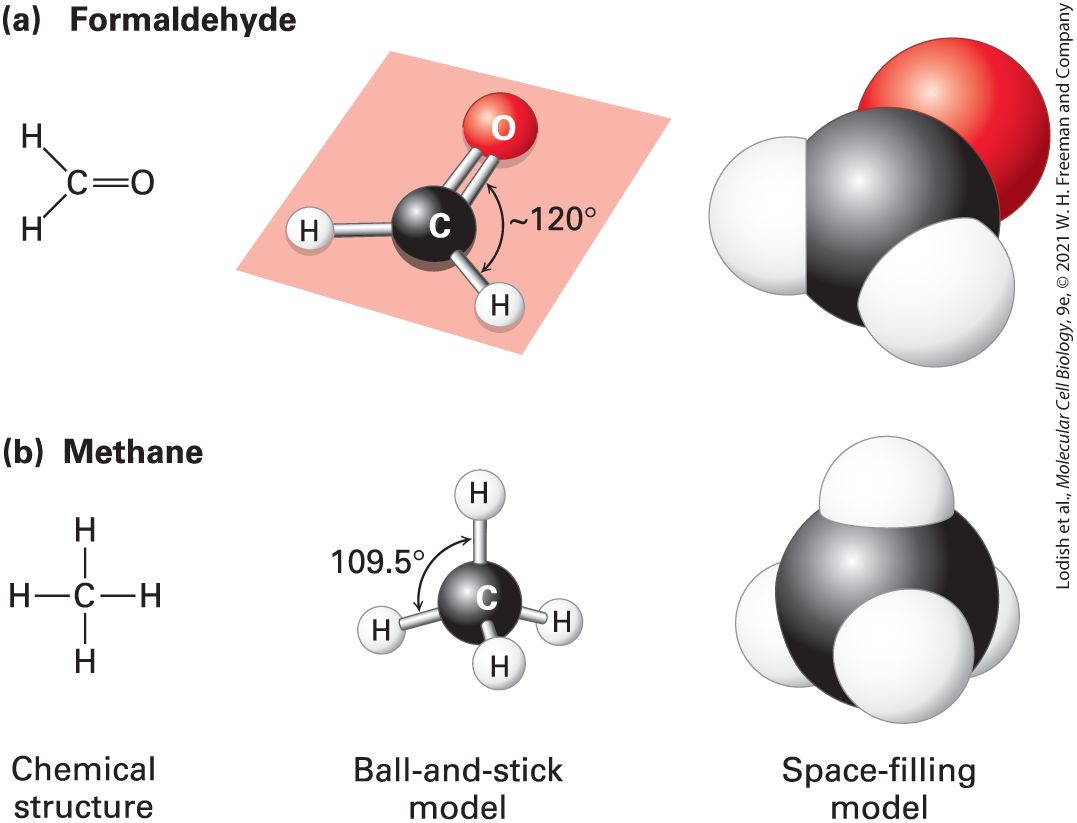
Description The illustration shows a 3-D orbital of a carbon atom with four electrons on each side combines with four hydrogen atoms with one electron in each to form methane. The carbon atom shares one electron with each hydrogen atom. A covalent bond thus forms between the carbon atom and the hydrogen atom. All the biological building blocks are organized around the carbon atom. In these organic biomolecules, each carbon usually bonds to three or four other atoms. [Carbon can also bond to two other atoms, as in the linear molecule carbon dioxide, , which has two carbon-oxygen double bonds ; however, such bond arrangements of carbon are not found in biological building blocks.] As illustrated in Figure 2-3a for formaldehyde, carbon can bond to three atoms, all in a common plane. The carbon atom forms two single bonds with two atoms and one double bond (two shared electron pairs) with the third atom. In the absence of other constraints, atoms joined by a single bond generally can rotate freely about the bond axis, whereas those connected by a double bond cannot. The rigid planarity imposed by double bonds has enormous significance for the shapes and flexibility of biomolecules such as phospholipids, proteins, and nucleic acids.
FIGURE 2-3 Geometry of bonds when carbon is covalently linked to three or four other atoms. (a) A carbon atom can be bonded to three atoms, as in formaldehyde . In this case, the carbon-bonding electrons participate in two single bonds and one double bond. All bonds lie in the same plane. Unlike atoms connected by a single bond, which usually can rotate freely about the bond axis, those connected by a double bond cannot. (b) When a carbon atom forms four single bonds, as in methane the bonded atoms (all H in this case) are oriented in space in the form of a tetrahedron. The letter representations on the left clearly indicate the atomic composition of each molecule and its bonding pattern. The ball-and-stick models in the center illustrate the geometric arrangement of the atoms and bonds, but the diameters of the balls representing the atoms and their nonbonding electrons are unrealistically small compared with the bond lengths. The sizes of the electron clouds in the space-filling models on the right more accurately represent the structure in three dimensions. Description
The illustration labeled A shows the chemical structure of formaldehyde with a carbonyl group bonded two hydrogen atoms. The ball-and-stick model has a black sphere representing carbon atom double bonded to a red sphere representing an oxygen atom, and single bonded to two white spheres each representing hydrogen atoms. The angle between the oxygen atom and the hydrogen atom is approximately 120 degrees. The space-filling model shows a black sphere fused with a red sphere at top right, a white sphere at bottom right, and another white sphere at the left. The illustration labeled A shows the chemical structure of methane with a carbon atom single bonded to four hydrogen atoms one each at the top, the bottom, the left, and the right. The ball-and-stick model has a black sphere representing a carbon atom bonded to four white spheres representing hydrogen atoms at the top, the front, the back, and the bottom left, each. The angle between the top hydrogen atom and the bottom left hydrogen atom is 109.5 degrees. The space-filling model shows a black sphere fused with four white spheres at the top, at the front, at the right, and at the left. Carbon can also bond to four rather than three atoms. As illustrated by methane when carbon is bonded to four other atoms, the angle between any two bonds is , and the positions of bonded atoms define the four points of a tetrahedron (Figure 2-3b). This geometry defines the structures of many biomolecules. A carbon (or any other) atom bonded to four dissimilar atoms or groups in a nonplanar configuration is said to be asymmetric. The four atoms connected to an asymmetric carbon atom in a tetrahedral orientation can be arranged in three-dimensional space in two different ways, producing molecules that are mirror images of each other, a property called chirality (“handedness,” from the Greek word cheir, meaning “hand”) (Figure 2-4). Such molecules are called optical isomers, or stereoisomers. Many molecules in cells contain at least one asymmetric carbon atom, often called a chiral carbon atom. The different stereoisomers of a molecule usually have completely different biological
activities because the arrangement of atoms within their structures, and thus their ability to interact with other molecules, differs.
FIGURE 2-4 Stereoisomers. Many molecules in cells contain at least one asymmetric carbon atom. The tetrahedral geometry of bonds formed by an asymmetric carbon atom can be arranged in three-dimensional space in two different ways, producing molecules that are mirror images, or stereoisomers, of each other. Shown here is the common structure of an amino acid, with its central asymmetric carbon and four attached groups, including the R group, discussed in Section 2.2. Amino acids can exist in two mirror-image forms, designated L and D. Although the chemical properties of such stereoisomers are identical, their biological activities are distinct. Only L amino acids are found in proteins. Description The amino acid model shows a black sphere representing an alpha carbon bonded to a red sphere representing a carboxylate ion at the top, a blue sphere representing an ammonia cation at the right, a green sphere representing an R group at the bottom, and a white sphere representing a hydrogen atom at the left. This structure is labeled D
isomer. Its mirror image labeled L isomer shows a non-superimposable structure of a reversed copy of this amino acid. Some drugs are mixtures of the stereoisomers of a small molecule in which only one stereoisomer has the biological activity of interest, for example, inhibiting the activity of a specific protein molecule. Unfortunately, both the stereoisomer that has the biological activity of interest and the one without biological activity might have side effects that could be harmful. The use of a pure, single stereoisomer of the chemical in place of the mixture may result in a more potent drug with fewer side effects. For example, one stereoisomer of the antidepressant drug citalopram (Celexa) is 170 times more potent than the other. Some stereoisomers have very different activities. Darvon is a pain reliever, whereas its stereoisomer, Novrad (Darvon spelled backward), is a cough suppressant. One stereoisomer of ketamine is an anesthetic, whereas the other causes hallucinations. The typical numbers of covalent bonds formed by atoms common in biomolecules are shown in Table 2-1. A hydrogen atom forms only one covalent bond. An oxygen atom usually forms only two covalent bonds but has two additional pairs of electrons (indicated as dots in the table) that can participate in noncovalent interactions. These extra electrons that are not shared in a covalent bond are also called nonbonding electrons. As we will see in later chapters, the nonbonding electrons can play important roles in the chemical reactions between biomolecules. A sulfur atom forms two covalent bonds in hydrogen sulfide but can accommodate
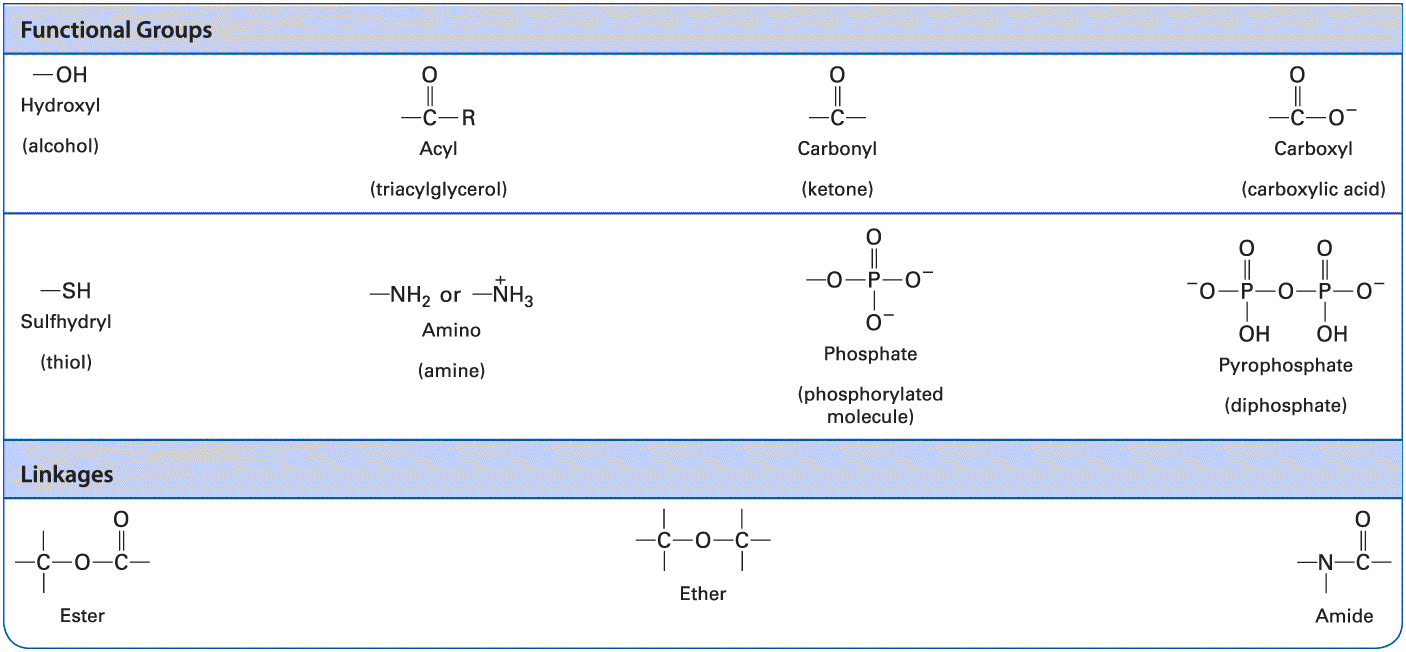
six covalent bonds in sulfuric acid and its sulfate derivatives. Nitrogen and phosphorus atoms each have five electrons to share. In ammonia the nitrogen atom forms three covalent bonds; the pair of electrons around the atom not involved in a covalent bond can take part in noncovalent interactions. In the ammonium ion the nitrogen atom forms four covalent bonds, which have a tetrahedral geometry. A phosphorus atom commonly forms five covalent bonds, as in phosphoric acid and its phosphate derivatives, which form the backbone of nucleic acids. Phosphate groups covalently attached to proteins play a key role in regulating the activity of many proteins, and the central molecule in cellular energetics, ATP, contains three phosphate groups (see Section 2.4). A summary of common covalent linkages and functional groups, which confer distinctive chemical properties on the molecules of which they are a part, is provided in Table 2-2.
TABLE 2-1 • Bonding Properties of Atoms Most Abundant in Biomolecules
Description The column headers are as follows: atom and outer electrons; usual number of covalent bonds; and typical bond geometry. The row entries are as follows: Row 1: Atom and outer electrons, hydrogen atom having a valence electron. Usual number of covalent bonds, 1. typical bond geometry, hydrogen atom with a single open bond at the bottom left. Row 2: Atom and outer electrons, oxygen atom having two pairs of lone electrons and two valence electrons. Usual number of covalent bonds, 2. typical bond geometry, oxygen atom having two lone pairs of electrons with two single open bonds at the bottom left and at the bottom right. Row 3: Atom and outer electrons, sulfur atom having two pairs of lone electrons and two valence electrons. Usual number of covalent bonds, 2; 4; or 6. typical bond geometry, sulfur atom having two lone pairs of electrons with two single open bonds at the bottom left and at the bottom right. Row 4: Atom and outer electrons, nitrogen atom having a lone pair of electron and three valence electrons. Usual number of covalent bonds, 3 or 4. typical bond geometry, nitrogen atom having a lone electron pair with two single open bonds at the bottom left and at the bottom right.
Row 5: Atom and outer electrons, phosphorus having a lone pair of electron and three valence electrons. Usual number of covalent bonds, 5. typical bond geometry, phosphorus atom with three single bonds each at the bottom left, at the bottom right, and at the bottom, and a double bond at the top. Row 6: Atom and outer electrons, carbon atom having four valence electrons. Usual number of covalent bonds, 4. typical bond geometry, carbon atom with four single bonds one each at the bottom left, bottom right, bottom, and the top.
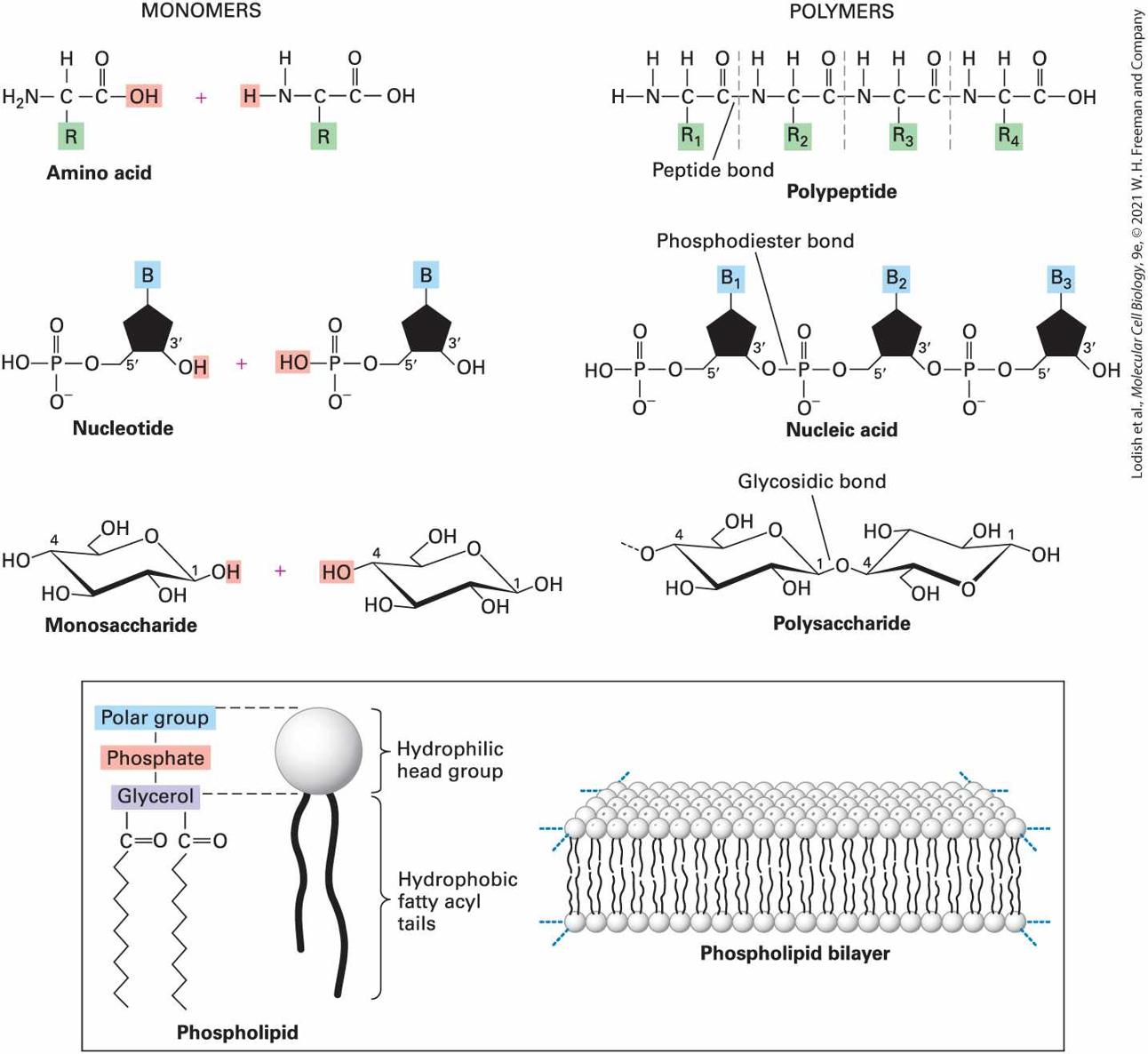
TABLE 2-2 • Common Functional Groups and Linkages in Biomolecules Description The functional groups include hydroxyl or alcohol groups. Acyl or triacylglycerol group; where a carbonyl group with an open bond is bonded to an R group. Carbonyl or ketone group. Carboxyl or carboxylic acid group, where a carbonyl group with an open bond is bonded to an oxygen anion. Sulfhydryl or thiol group. Amino or amine group. Phosphate or phosphorylated molecule, where phosphorus is single bonded to three oxygen anions and double bonded to an oxygen atom, and pyrophosphate or diphosphate group, where an oxygen atom is bonded to two phosphorus atoms. Each phosphorus atom is further bonded to a hydroxyl group, an oxygen anion and double
All Covalent Bonds Are Not Equal: Electrons May Be Shared Equally or Unequally in Covalent Bonds
bonded to another oxygen atom. The linkages include ester, ether, and amide. Ester has an oxygen atom bonded to a carbon atom with three open bonds and a carboxyl group with an open bond. Ether has an oxygen atom bonded to two carbon atoms each having three open bonds. Amide has a nitrogen atom with two open bonds bonded to a carboxyl group having an open bond. All Covalent Bonds Are Not Equal: Electrons May Be Shared Equally or Unequally in Covalent Bonds The extent of an atom’s ability to attract an electron is called its electronegativity. In a bond between atoms with identical or similar electronegativities, the bonding electrons are essentially shared equally between the two atoms, as is the case for most carbon-carbon single bonds and carbon-hydrogen single bonds . Such bonds are called nonpolar. In many molecules, however, the covalently bonded atoms sharing electrons have different electronegativities, resulting in unequal sharing of electrons. The bond between atoms that share electrons unequally is said to be polar. One end of a polar bond has a partial negative charge and the other end has a partial positive charge . In an bond, for example, the oxygen atom is more electronegative than the hydrogen atom. As a result, the electrons forming the covalent bond spend more time around the oxygen atom than around the hydrogen. Thus the bond possesses an electric dipole, a positive charge separated from an equal but opposite
negative charge. The amount of charge on the oxygen atom of an dipole is approximately 25 percent that of an electron, and there is an equivalent and opposite charge on the H atom. A common quantitative measure of the extent of charge separation, or strength, of a dipole is called the dipole moment, . For a chemical bond, the dipole moment is the product of the partial charge on each atom and the distance between the two atoms. Many molecules have multiple dipoles. In these cases, the amount of charge separation for the molecule as a whole — the molecule’s dipole moment — depends in part on the dipole moments of all of its individual chemical bonds and in part on the geometry of the molecule (the relative orientations of the individual bond dipole moments). Consider the example of water . A water molecule has two bonds and thus two individual bond dipole moments. If water were a linear molecule with the two bonds on exact opposite sides of the O atom, the two dipoles on each end of the molecule would be identical in strength but would be oriented in opposite directions. The two dipole moments would cancel each other, and the dipole moment of the molecule as a whole would be zero. However, water is a V-shaped molecule, with the individual dipoles of its two bonds both pointing toward the oxygen. One end of the water molecule (the end with the oxygen atom) has a partial negative charge and the other end (the one with the two hydrogen atoms) has a partial positive charge. As a consequence, the molecule as a whole is a dipole with a well-defined dipole moment (Figure 2-5). This dipole moment and the electronic properties of the oxygen and hydrogen atoms
allow water to form electrostatic, noncovalent interactions with other water molecules and with molecules of other types. These interactions play a critical role in almost every biochemical interaction in cells and organisms, as we will see shortly.
FIGURE 2-5 The dipole nature of a water molecule. The symbol represents a partial charge (a weaker charge than the one on an electron or a proton). Because of the difference in the electronegativities of H and O, each of the polar bonds in water is a dipole. The sizes and directions of the dipoles of each of the bonds determine the net distance between charges and amount of charge separation in the molecule — its dipole moment. Description The model shows a red sphere representing an oxygen atom bonded to two hydrogen atoms represented by two white spheres. The angle between the two hydrogen atoms is 104.5 degrees. Both hydrogen atoms are labeled delta positive while the oxygen atom is labeled delta negative near the bond. To the right of the molecule, an upward arrow labeled positive at the bottom and negative at the top arrowhead indicate a dipole moment. Another important example of polarity is the double bond in . In the structure of shown on the top of the next page, lines
represent single and double bonds and nonbonding electrons are shown as pairs of dots (each dot represents one electron): Description The illustration shows two resonance structures of phosphate, with lines representing single and double bonds, and nonbonding electrons represented as pairs of dots, with each dot representing one electron. A double-headed arrow connects the neutral phosphate and a resonance structure, which is overall neutral, but has a negatively charged oxygen and a positively charged phosphorus. Because of the polarity of the double bond, can also be represented by the structure on the right, in which one of the electrons from the double bond has accumulated around the O atom, giving it a negative charge and leaving the P atom with a positive charge. These charges are important in noncovalent interactions. Neither of these two models precisely describes the electronic state of . The actual structure can be considered to be an intermediate, or hybrid, between these two representations, as indicated by the double-headed arrow between them. Such intermediate structures are called resonance hybrids.
Covalent Bonds Are Much Stronger and More Stable Than Noncovalent Interactions
Covalent Bonds Are Much Stronger and More Stable Than Noncovalent Interactions Covalent bonds are considered to be strong because the energies required to break them are much greater than the thermal energy available at room temperature or body temperature . As a consequence, covalent bonds are stable at these temperatures. For example, the thermal energy available at is approximately 0.6 kilocalorie per mole whereas the energy required to break the bond in ethane is about 140 times larger (Figure 2-6). Consequently, at room temperature fewer than 1 in ethane molecules are broken into pairs of molecules, each containing an unpaired, nonbonding electron (called a radical).
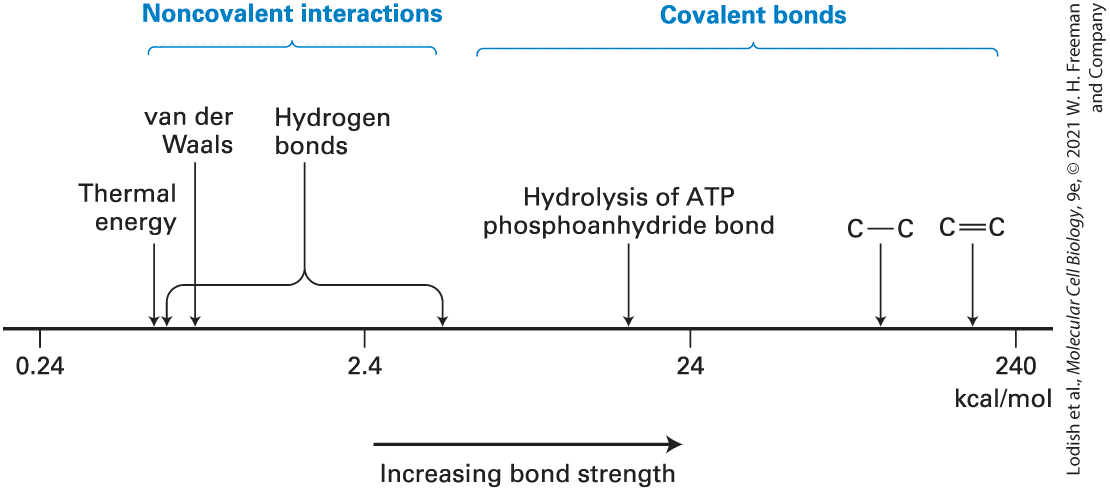
FIGURE 2-6 Relative energies of covalent bonds and noncovalent interactions. Bond energies are defined as the energy required to break a particular type of linkage. Shown here are the energies required to break a variety of linkages, arranged on a log scale.
Covalent bonds, including single and double carbon-carbon bonds, are one to two powers of 10 stronger than noncovalent interactions. Noncovalent interactions have energies somewhat greater than the thermal energy of the environment at normal room temperature . Many biological processes are coupled to the energy released during hydrolysis of a phosphoanhydride bond in ATP. Description A chart shows a horizontal scale running from 0.24 to 240 in units of kilocalories per mole. An arrow pointing from the left to right indicates increasing bond strength and from lower energies to higher energies. Noncovalent interactions, including thermal energy, van der Waals, and hydrogen bonds, lie between 0.24 and more than 2.4 kilocalories per mole. Hydrogen bonds are spread over a wide range of energies. The energies of several covalent bonds are also listed, such as the bond energy of an A T P phosphoanhydride bond, close to 24 kilocalories per mole, and carbon-carbon single and double bonds, at several hundred kilocalories per mole. Covalent single bonds in biological molecules have energies similar to the energy of the bond in ethane. Because more electrons are shared between atoms in double bonds, they require more energy to break than single bonds. For instance, it takes to break a single bond but to break a double bond. The most common double bonds in biological molecules are and . In contrast, the energy required to break noncovalent interactions is only much less than the bond energies of covalent bonds (see
Figure 2-6). Indeed, noncovalent interactions are weak enough that they are constantly being formed and broken at room temperature. Although these weak interactions have a transient existence at physiological temperatures multiple noncovalent interactions can, as we
Ionic Bonds Are Noncovalent Interactions Formed by the Electrostatic Attractions Between Oppositely Charged Ions
will see, act together to produce highly stable and specific associations between different parts of a large molecule or between different macromolecules. Protein-protein and protein-nucleic acid interactions are good examples of such noncovalent interactions. Below, we review the four main types of noncovalent interactions and then consider their roles in the binding of biomolecules to one another and to other molecules. Ionic Bonds Are Noncovalent Interactions Formed by the Electrostatic Attractions Between Oppositely Charged Ions Ionic bonds (also called ionic interactions) result from the attraction between a positively charged ion — a cation — and a negatively charged ion — an anion. In sodium chloride , for example, the bonding electron contributed by the sodium atom is completely transferred to the chlorine atom (Figure 2-7a). Unlike covalent bonds, ionic interactions do not have fixed or specific geometric orientations because the electrostatic field around an ion — its attraction for an opposite charge — is uniform in all directions. In solid , oppositely charged ions pack tightly together in an alternating pattern, forming the highly ordered crystalline array, or lattice, that is typical of salt crystals (Figure 2-7b, and see the opening figure for this chapter). The energy required to break an ionic interaction depends on the distance between the ions and the electrical properties of the environment of the ions.
FIGURE 2-7 Electrostatic interactions of the oppositely charged ions of salt (NaCl) in crystals and in aqueous solution. (a) In crystalline table salt, sodium atoms are positively charged ions due to the loss of one electron each, whereas chloride atoms are correspondingly negatively charged due to the gain of one electron each. (b) In solid form, ionic compounds form neatly ordered arrays, or crystals, of tightly packed ions in which the positively and negatively charged ions counterbalance each other. (c) When the crystals are dissolved in water, the ions separate, and their charges, no longer balanced by immediately adjacent ions of opposite charge, are stabilized by interactions with polar water. The water molecules and the ions are held together by electrostatic interactions between the charges on the ion and the partial charges on the water’s oxygen and hydrogen atoms. In aqueous solutions, all ions are surrounded by a hydration shell of water molecules. Description The illustration labeled A shows two sodium atoms represented by purple spheres donate an electron each to two chlorine atoms represented by green spheres to form sodium cations and chloride anions. This leads to the formation of solid, crystalline sodium chloride. In illustration B, the crystalline structure has sodium and chloride ions arranged in order to form a cube. Four space-filling models of water are present surrounding the crystal structure. The space-filling structure of water is made of a red sphere fused with two white hemispheres. In illustration C, hydration occurs in water, where the cations and anions are solvated by water molecules. There are two sodium cations and two chlorine anions each bonded to four water molecules. A reversible arrow indicates the two processes, dissolution and crystallization.
Hydrogen Bonds Are Noncovalent Interactions That Determine the Properties of Water and the Water Solubility of Uncharged Molecules
When solid salts dissolve in water, the ions separate from one another and are stabilized by their interactions with water molecules. In aqueous solutions, simple ions of biological significance, such as , and , are hydrated, surrounded by a stable shell of water molecules held in place by electrostatic interactions between the ion at the center and the oppositely charged ends of the water molecules, which are dipoles (Figure 2-7c). Energy is released when ions tightly bind water molecules and spread out in an aqueous solution, due to the increase in disorder, or entropy (see Section 2.4). Most ionic compounds dissolve readily in water because this energy of hydration is greater than the lattice energy that stabilizes the crystal structure. Parts or all of the aqueous hydration shell must be removed from ions in solution when they interact directly with proteins. For example, water of hydration is lost when ions pass through protein pores in the cell membrane during nerve conduction. The relative strength of the interaction between two oppositely charged ions, and , depends on the concentration of other ions in a solution. The higher the concentration of other ions (e.g., and ), the more opportunities and have to interact ionically with those other ions, and thus the lower the energy required to break the interaction between and . As a result, increasing the concentrations of salts such as in a solution of biological molecules can weaken and even disrupt the ionic interactions holding the biomolecules together. This principle can be exploited to separate complex mixtures of interacting molecules, such as proteins, into their individual, pure components.
Hydrogen Bonds Are Noncovalent Interactions That Determine the Properties of Water and the Water Solubility of Uncharged Molecules A hydrogen bond is the interaction of a hydrogen atom with a partial positive charge in a dipole, such as water, with unpaired electrons from another atom, either in the same or in a different molecule. Normally, a hydrogen atom forms a covalent bond with a single other atom. However, a hydrogen atom covalently bonded to an electronegative donor atom D may form an additional weak association, the hydrogen bond, with an acceptor atom A, which must have a nonbonding pair of electrons available for the interaction: The covalent bond is a bit longer than it would be if there were no hydrogen bond because the acceptor pulls the hydrogen away from the donor. An important feature of all hydrogen bonds is directionality. In the strongest hydrogen bonds, the donor atom, the hydrogen atom, and the acceptor atom all lie in a straight line. Nonlinear hydrogen bonds are weaker than linear ones; still, multiple nonlinear hydrogen bonds help to stabilize the three-dimensional structures of many proteins.
Hydrogen bonds are both longer and weaker than covalent bonds between the same atoms. In water, for example, the distance between the nuclei of the hydrogen and oxygen atoms forming a hydrogen bond connecting two adjacent water molecules is about , about twice the length of the covalent bonds within a single water molecule (Figure 2-8a). A hydrogen bond between water molecules (approximately ) is much weaker than a covalent bond (roughly ), although it is stronger than many other hydrogen bonds in biological molecules . Extensive hydrogen bonding between water molecules accounts for many of water’s key properties, including its unusually high melting and boiling points and its ability to dissolve many other molecules.
FIGURE 2-8 Hydrogen bonding of water with itself and with other compounds. Each pair of nonbonding outer electrons in an oxygen or a nitrogen atom can accept a hydrogen atom in a hydrogen bond. The hydroxyl and the amino groups can also form hydrogen bonds with water. (a) In liquid water, each water molecule forms transient hydrogen bonds with several others, creating a dynamic network of hydrogen-bonded molecules. (b) Water can also form hydrogen bonds with alcohols and amines, which accounts for the high solubility of these compounds. (c) The peptide group and the ester group, which are present in many biomolecules, commonly participate in hydrogen bonds with water or polar groups in other molecules.
Description Illustration A, Hydrogen bonding in water: Five molecules of water are dot bonded to each other to form a square network. Each oxygen atom of the water molecules has two lone pairs of electrons. The dot bonds represent hydrogen bonds. Illustration B, Hydrogen bonding between water and alcohol: Methanol ( C H subscript 3 O H) is dot bonded to two water molecules. Each oxygen atom of the water molecules and methanol has two lone pairs of electrons. The dot bonds represent hydrogen bonds. Hydrogen bonding between amine and water: A methylamine group (C H subscript 3 end subscript N H subscript 2) is dot bonded to two water molecules. Each oxygen atom of the water molecules has two lone pairs of electrons. The dot bonds represent hydrogen bonds. Illustration C, Hydrogen bonding between peptide group and water: A peptide group (C O N H) is dot bonded to two water molecules. Each oxygen atom of the water molecules and the peptide group has two lone pairs of electrons. The dot bonds represent hydrogen bonds. Hydrogen bonding between ester and water: An ester group (C O O) is dot bonded to a water molecule. Each oxygen atom of the water molecules and the ester group has two lone pairs of electrons. The solubility of uncharged substances in an aqueous environment depends largely on their ability to form hydrogen bonds with water. For instance, the hydroxyl group in alcohols and the amino group in amines can form several hydrogen bonds with water, which allows these molecules to dissolve in water at high concentrations (Figure 2-8b). In general, molecules with polar bonds that easily form hydrogen bonds with water, as well as charged molecules and ions that interact with the dipole in water, can readily dissolve in water; that is, they are hydrophilic. In addition to hydroxyl and amino groups, many biological molecules contain peptide and ester groups, which form hydrogen bonds with water via otherwise nonbonded electrons on their
carbonyl oxygens (Figure 2-8c). X-ray crystallography combined with computational analysis permits an accurate depiction of the distribution of the outermost unbonded electrons of atoms that can participate in hydrogen bonds as well as the electrons in covalent bonds, as illustrated in
FIGURE 2-9 Distribution of bonding and outer nonbonding electrons in the peptide group. Shown here is a peptide bond linking two amino acids within a protein called
Van der Waals Interactions Are Weak Attractive Interactions Caused by Transient Dipoles
crambin. No protein has been structurally characterized at higher resolution than crambin. The black lines represent the covalent bonds between atoms. The red (negative) and blue (positive) lines represent contours of charge determined using x-ray crystallography and computational methods. The greater the number of contour lines, the higher the charge. The high density of red contour lines between atoms represents the covalent bonds (shared electron pairs). The two sets of red contour lines emanating from the oxygen (O) and not falling on a covalent bond (black line) represent the two pairs of nonbonding electrons on the oxygen that are available to participate in hydrogen bonding. The high density of blue contour lines near the hydrogen (H) bonded to nitrogen (N) represents a partial positive charge, indicating that this H can participate in hydrogen bonding. [From Proc. Nat’l. Acad. Sci. USA, 2000, 97(7):3171–3176, Fig. 3A. Accurate protein crystallography at ultra-high resolution: Valence electron distribution in crambin, by Christian Jelsch et al., copyright (2000) National Academy of Sciences, USA.] Description The illustration shows a planar peptide bond (C O N H) between C subscript alpha (crambin) and two amino acids. Areas of high electron density are indicated by a high density of red contour lines. Low electron density is shown by sparse blue contours. Black lines indicate covalent bonds. Labels include H (hydrogen), N (nitrogen), O (oxygen), and C subscript alpha (crambin). At the right, around the oxygen, an area of dense red contour lines is labeled Nonbonded electrons. Areas of high electron density correspond to the regions between atoms, the bonds, and the lone pairs of the carbonyl oxygen atoms. Van der Waals Interactions Are Weak Attractive Interactions Caused by Transient Dipoles
When any two atoms approach each other closely, they create a weak attractive force called a van der Waals interaction. These interactions result from the momentary random fluctuations in the distribution of the electrons of any atom. The random fluctuations give rise to a transient unequal distribution of electrons, creating a temporary dipole. If two noncovalently bonded atoms are close enough, electrons of one atom will perturb the electrons of the other. This perturbation generates a transient dipole in the second atom, and the two dipoles attract each other weakly (Figure 2-10). Similarly, a polar covalent bond in one molecule attracts an oppositely oriented dipole in another.
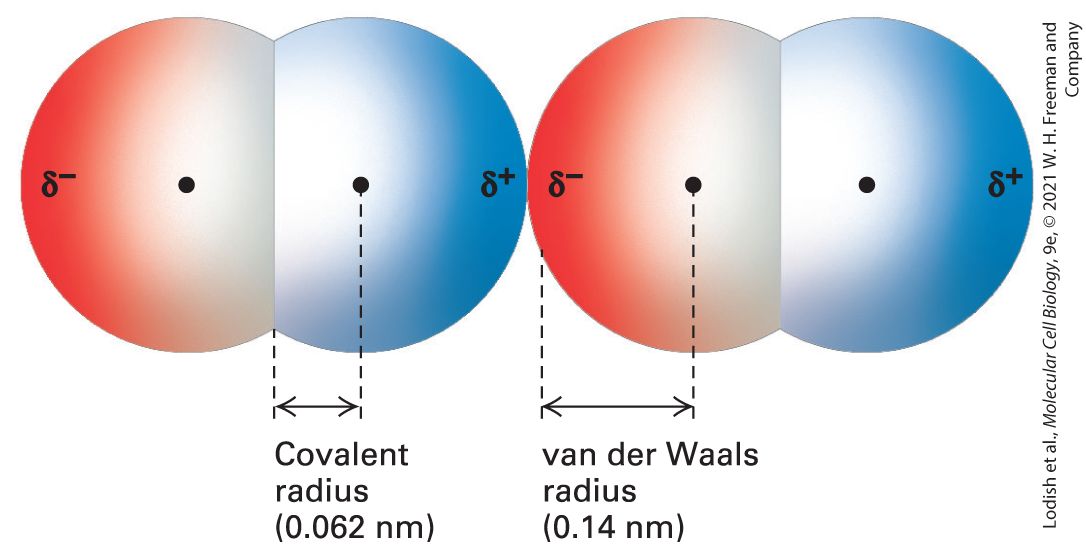
FIGURE 2-10 Two oxygen molecules in van der Waals contact. In this model, red indicates negative charge and blue indicates positive charge. Transient dipoles in the electron clouds of all atoms give rise to weak attractive forces, called van der Waals interactions. Each type of atom has a characteristic van der Waals radius at which van der Waals interactions with other atoms are optimal. Because atoms repel one another if they are close enough together for their outer electrons to overlap without being shared in a covalent bond, the van der Waals radius is a measure of the size of the electron cloud surrounding an
atom. The covalent radius indicated here is for the double bond of ; the single-bond covalent radius of oxygen is slightly longer. Description Two space-filling models of diatomic oxygen molecules are present next to each other. Their nuclei are illustrated by points and the atoms are represented as hemispheres with their diameters equal to the van der Waals radii. The terminal ends of each of the molecules are labeled delta superscript negative on the left and delta superscript positive on the right. The van der Waals radius between the nucleus and delta superscript negative is indicated as 0.14 nanometers. The covalent radius, which is the distance between the nucleus and overlapping region, is indicated as 0.062 nanometers. Van der Waals interactions, involving either transient or permanent dipoles, occur in all types of molecules, both polar and nonpolar. In particular, van der Waals interactions are responsible for the cohesion between nonpolar molecules such as heptane, , that cannot form hydrogen bonds or ionic interactions with each other. The strength of van der Waals interactions decreases rapidly with increasing distance; thus these noncovalent interactions can form only when atoms are quite close to one another. However, if atoms get too close together, the negative charges of their electrons create a repulsive force. When the van der Waals attraction between two atoms exactly balances the repulsion between their two electron clouds, the atoms are said to be in van der Waals contact. The strength of the van der Waals interaction is about so it is weaker than typical hydrogen bonds, and its energy is only slightly higher than the average thermal energy of molecules at . Thus multiple van der Waals interactions, a van der Waals interaction together with other noncovalent interactions, or both are
The Hydrophobic Effect Causes Nonpolar Molecules to Adhere to One Another
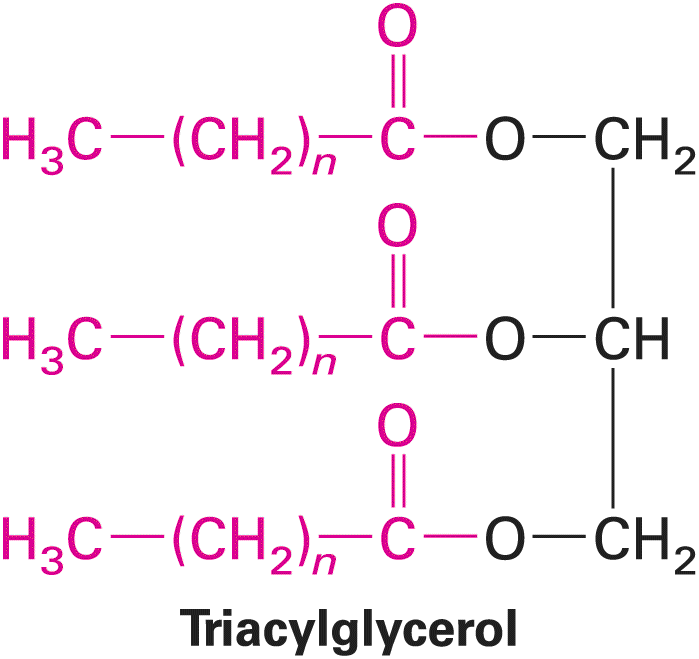
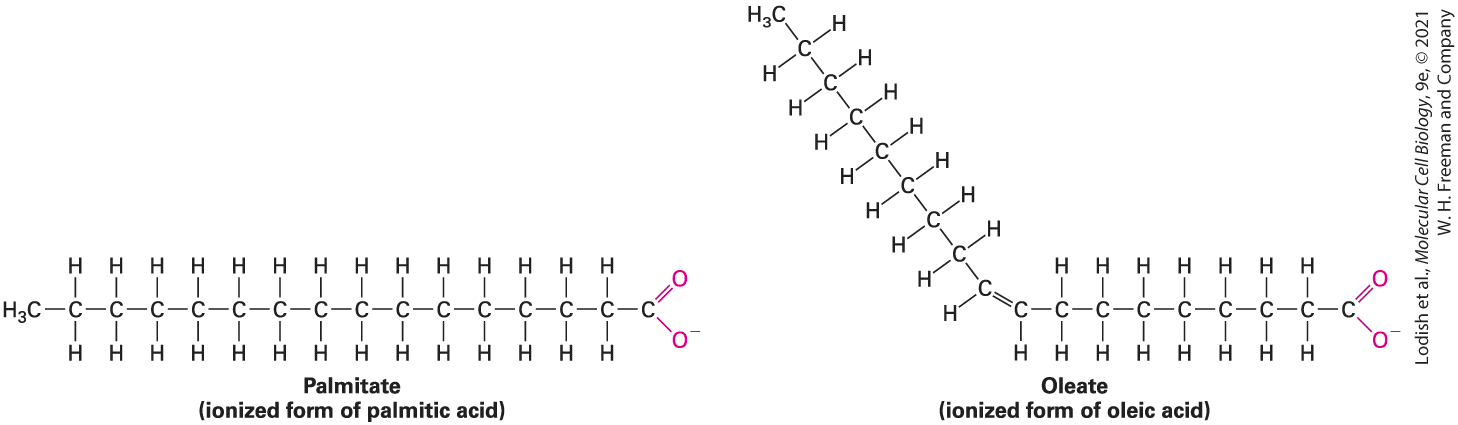
required to form van der Waals–mediated, stable attractions within and between molecules. The Hydrophobic Effect Causes Nonpolar Molecules to Adhere to One Another Because nonpolar molecules do not contain charged groups, do not possess a dipole moment, and do not become hydrated, they are insoluble, or almost insoluble, in water; that is, they are hydrophobic. The covalent bonds between two carbon atoms and between carbon and hydrogen atoms are the most common nonpolar bonds in biological systems. Hydrocarbons — molecules made up only of carbon and hydrogen — are virtually insoluble in water. Large triacylglycerols (also known as triglycerides), which make up animal fats and vegetable oils, also essentially are insoluble in water. As we will see later, the major part of these molecules consists of long hydrocarbon chains. After being shaken in water, triacylglycerols form a separate phase. A familiar example is the separation of oil from the water-based vinegar in an oil-and-vinegar salad dressing. Nonpolar molecules or nonpolar parts of molecules tend to aggregate in water owing to a phenomenon called the hydrophobic effect. Because water molecules cannot form hydrogen bonds with nonpolar substances, they tend to form cages of relatively rigid hydrogen-bonded pentagons and hexagons around nonpolar molecules (Figure 2-11, left). This state is
energetically unfavorable because it decreases the entropy, or randomness, of the population of water molecules. (The role of entropy in chemical systems is discussed in Section 2.4.) If nonpolar molecules in an aqueous environment aggregate with their hydrophobic surfaces facing each other, the net hydrophobic surface area exposed to water is reduced (Figure 2-11, right). As a consequence, fewer water molecules are needed to form the ordered cages surrounding the nonpolar molecules, entropy increases relative to the unaggregated state, and an energetically more favorable state is reached. In a sense, then, water squeezes the nonpolar molecules into aggregates. Rather than constituting an attractive force, as in hydrogen bonds, the hydrophobic effect results from an avoidance of an unstable state — that is, extensive water cages around individual nonpolar molecules.
FIGURE 2-11 Schematic depiction of the hydrophobic effect. Cages of water molecules form around nonpolar molecules in solution. These water molecules in cages are more ordered than water molecules in the surrounding bulk liquid. Aggregation of nonpolar molecules reduces the number of water molecules involved in forming highly ordered cages, resulting in a higher entropy, more energetically favorable state (right) compared with the unaggregated state (left). Description Two nonpolar molecules are illustrated as ovals surrounded by cages of highly ordered water molecules. A label states that this situation has lower entropy than the system after hydrophobic aggregation. After aggregation, the two nonpolar molecules are closer together, the cage of water is larger but contains fewer ordered water molecules, the excess water molecules no longer required have been released, and the entropy of the system is larger.
Nonpolar molecules can also associate, albeit weakly, through van der Waals interactions. The net result of the hydrophobic effect and van der Waals interactions is a very powerful tendency for hydrophobic molecules in water to interact with one another, not with water molecules. Simply put, like dissolves like. Polar molecules dissolve in polar solvents such as water; nonpolar molecules dissolve in nonpolar solvents such as hexane. One well-known hydrophobic molecule is cholesterol (see Section 2.2). Cholesterol, triglycerides, and other poorly water-soluble molecules are called lipids. Unlike hydrophilic molecules such as glucose or amino acids, lipids cannot readily dissolve in the liquid portion of the blood, the aqueous circulatory system that transports molecules and cells throughout the body. Instead, lipids, such as cholesterol, must be packaged into special hydrophilic carriers, called lipoproteins, that can themselves dissolve in the blood and be transported throughout the body. There can be hundreds to thousands of lipid molecules packed into the center, or core, of each lipoprotein. The hydrophobic core is surrounded by amphipathic molecules that have hydrophilic parts that interact with water and hydrophobic parts that interact with one another and the core. The packaging of lipids into lipoproteins (discussed in Chapter 14) permits their efficient transport in blood and is reminiscent of the containerization of cargo for efficient, long-distance transport via cargo ships, trains, and trucks. High-density lipoprotein (HDL) and low-density lipoprotein (LDL) are two such lipoprotein carriers. HDL is associated with reduced heart
Molecular Complementarity Due to Noncovalent Interactions Leads to a Lock-and-Key Fit Between Biomolecules
disease, and LDL with increased heart disease, and they are therefore often referred to as good and bad cholesterol, respectively. Actually, the cholesterol molecules and their derivatives that are carried by both HDL and LDL are essentially identical and in themselves are neither good nor bad. However, HDL and LDL have different effects on cells. The bad cholesterol, LDL, contributes to clogging of the arteries (known as atherosclerosis) and consequent heart disease and stroke. The mechanisms linking HDL to reduced risk for heart attacks are unclear. Molecular Complementarity Due to Noncovalent Interactions Leads to a Lock-and-Key Fit Between Biomolecules Both inside and outside cells, ions and molecules constantly collide. The higher the concentration of any two types of molecules, the more likely they are to encounter each other. When two molecules do encounter each other, they are most likely to simply bounce apart because the noncovalent interactions that would bind them together are weak and have a transient existence at physiological temperatures. However, two distinct molecules can exhibit molecular complementarity, a lock-and-key kind of fit between their shapes, charges, or other physical properties. Two such structurally complementary molecules can form multiple noncovalent interactions at close range. When two such molecules bump into each
other, these multiple interactions can result in their sticking together, or binding.
Figure 2-12 illustrates how a variety of multiple weak interactions can cause two hypothetical proteins to bind together tightly. Numerous examples of such protein-to-protein molecular complementarity may be found throughout this book (see, e.g., Figures 16-9 and 16-18b). Almost any other arrangement of the same groups of atoms on the two surfaces would not allow the molecules to bind so tightly. Such molecular complementarity between regions within a protein molecule allow it to fold into a unique three-dimensional shape (see Chapter 3); it is also what holds the two chains of DNA together in a double helix (see Chapter 5). Similar interactions underlie the association of groups of molecules into multimolecular assemblies, or complexes, leading to the formation of muscle fibers, to the gluelike associations between cells in solid tissues, and to numerous other cellular structures. The antibodies that help neutralize pathogens (see Chapter 24) bind to them using similar principles of complementary molecular shapes.
FIGURE 2-12 Molecular complementarity permits tight protein bonding via multiple noncovalent interactions. The complementary shapes, charges, polarity, and hydrophobicity of two protein surfaces permit multiple weak interactions, which in combination produce a strong interaction and tight binding. Because deviations from molecular complementarity substantially weaken binding, a particular surface region of any given biomolecule usually can bind tightly to only one or a very limited number of other molecules. The complementarity of the two protein molecules on the left permits them to bind much more tightly than the two noncomplementary proteins on the right. Description The first illustration has space-filling models of proteins A and B that show complementary shapes. They have various complementary interactions, including ionic bonds, hydrogen bonds, and hydrophobic and van der Waals interactions. A hydroxyl group on protein A is dot bonded to an oxygen atom further bonded to a carbon atom in protein B at the top. The ionic bond is represented by protein A having a positive charge and protein B having a negative charge. The hydrogen bond is represented a carbonyl group of protein A dot bonded to an amide group of protein B. Three methyl groups are present on both proteins A and B in the middle. Protein A has two negative
charges and protein B has two positive charges towards the bottom end. A carbonyl group of protein A is dot bonded to a hydroxyl group of protein B at the bottom. Both proteins A and B form a stable complex. The second illustration has space-filling models of proteins A and C. They have very little shape complementarity and fewer intermolecular interactions as compared to the first illustration. Both proteins A and B form a less stable complex. Depending on the number and strength of the noncovalent interactions between the two molecules and on their environment, their binding may be tight or loose and, as a consequence, either lasting or transient. The higher the affinity of two molecules for each other, the better the molecular fit between them, the more noncovalent interactions can form, and the more tightly they can bind together. An important quantitative measure of affinity is the binding dissociation constant , described in Section 2.3. In many cases in cell biology, molecules must bind to one another but also be able to subsequently separate (dissociate). In other words, the binding needs to be reversible. Thus the binding cannot be too tight, or the molecules would not be able to dissociate when necessary. It is important to note that many large biological molecules are not hard, rigid structures, but rather can be somewhat malleable. Thus the binding of a molecule to another has the potential to induce a change in the shape of its binding partner. When the molecular complementarity increases after such interactions, the process is called induced fit. As we discuss in Chapter 3, nearly all the chemical reactions that occur in cells depend on the binding properties of enzymes. These proteins not only speed up, or catalyze, reactions, but do so with a high degree of specificity
because they are able to bind tightly to only one or a few related molecules. The specificity of intermolecular interactions and reactions, which depends on molecular complementarity, is essential for many processes critical to life. KEY CONCEPTS OF SECTION 2.1 Covalent Bonds and Noncovalent Interactions The terms hydrophilic, hydrophobic, and amphipathic/amphiphilic refer to the tendency of molecules to be water-loving, incapable of interacting with water, and having features of or being tolerant of both, respectively. Hydrophilic molecules typically dissolve readily in water, whereas hydrophobic molecules are poorly soluble or insoluble in water. Covalent bonds consist of pairs of electrons shared by two atoms. Covalent bonds arrange the atoms of a molecule into a specific geometry. Many molecules in cells contain at least one asymmetric carbon atom, which is bonded to four dissimilar atoms. Such molecules can exist as stereoisomers (mirror images), designated D and L (see Figure 2-4), which have different biological activities. Nearly all amino acids are L isomers. Electrons may be shared equally or unequally in covalent bonds. Atoms that differ in electronegativity form polar covalent bonds, in which the bonding electrons are distributed unequally. One end of a polar bond has a partial positive charge and the other end has a partial negative charge (see Figure 2-5). Covalent bonds are stable in biological systems because the relatively high energies required to break them are much larger than the thermal kinetic energy available at room or body in mammals) temperatures. Noncovalent interactions between atoms are considerably weaker than covalent bonds, with energies ranging from about (see Figure 2-6). Four main types of noncovalent interactions occur in biological systems: ionic bonds, hydrogen bonds, van der Waals interactions, and interactions due to the hydrophobic effect. Ionic bonds result from the electrostatic attraction between the positive and negative charges of ions. In aqueous solutions, all cations and anions are surrounded by a shell of bound water molecules (see Figure 2-7c). Increasing the salt (e.g., )
concentration of a solution can weaken the relative strength of and even break the ionic bonds between biomolecules. In a hydrogen bond, a hydrogen atom covalently bonded to an electronegative atom associates with an acceptor atom whose nonbonding electrons attract the hydrogen (see Figure 2-8). Weak van der Waals interactions result from the attraction between transient dipoles associated with all molecules. They can form when two atoms approach each other closely (see Figure 2-10). In an aqueous environment, nonpolar molecules or nonpolar parts of larger molecules are driven together by the hydrophobic effect, thereby reducing the extent of their direct contact with water molecules (see Figure 2-11). Molecular complementarity is the lock-and-key fit between molecules whose shapes, charges, and other physical properties are complementary. Multiple noncovalent interactions can form between complementary molecules, causing them to bind tightly (see Figure 2-12), but cannot form between molecules that are not complementary. The high degree of binding specificity that results from molecular complementarity is one of the features that underlies intermolecular interactions in biology and thus is essential for many processes critical to life.
2.2 Chemical Building Blocks of Cells
2.2 Chemical Building Blocks of Cells A common theme in biology is the construction of large macromolecules and macromolecular complexes (clusters of macromolecules) out of smaller molecular subunits, which can be thought of as building blocks. Often these subunits are similar or identical. The three main types of biological macromolecules — proteins, nucleic acids, and polysaccharides — are all polymers composed of multiple covalently linked small molecules, or monomers (Figure 2-13). Proteins are linear polymers containing up to several thousand amino acids linked by peptide bonds. Nucleic acids are linear polymers containing dozens to millions of nucleotides linked by phosphodiester bonds. Polysaccharides are linear or branched polymers of monosaccharides (sugars) such as glucose linked by glycosidic bonds.
FIGURE 2-13 Overview of the cell’s principal chemical building blocks. Top: The three major types of biological macromolecules are each assembled by the polymerization of multiple small molecules (monomers) of a particular type: proteins from amino acids (see Chapter 3), nucleic acids from nucleotides (see Chapter 5), and polysaccharides from monosaccharides (sugars). Each monomer is covalently linked into the polymer by a reaction whose net result is loss of a
water molecule (dehydration). Bottom: In contrast, phospholipid monomers noncovalently assemble into a bilayer structure, which forms the basis of all cellular membranes (see Chapter 10). Description Monomers: Two amino acid monomers are illustrated. The first amino acid has a central carbon bonded to a hydrogen atom at the top, an amide group on the left, R group (highlighted) at the bottom, and a carbonyl group on the right which is further bonded to a hydroxyl group (highlighted). The first amino acid combines with another amino acid having the same structure except one of the hydrogen atoms of the amide is highlighted. Polymers: A polypeptide shows four amino acids bound together by a peptide bond. The first amino acid has a central carbon bonded to a hydrogen atom at the top, an amide group on the left, R subscript 1 group (highlighted) at the bottom, and a carbonyl group on the right which is further bonded to the second amino acid. The second and third amino acids have similar structures where they have a central carbon atom bonded to a hydrogen atom at the top, an amine group on the left, R group (highlighted) at the bottom, and a carbonyl group on the right. The second amino acid is bonded to an R subscript 2 group whereas the third amino acid is bonded to an R subscript 3 group. The fourth amino acid is similar to that of the third except that the central carbon atom is bonded to an R subscript 4 group, and the carbonyl group is bonded to a hydroxyl group. Dotted vertical lines between the amino acids represent the peptide bond. Monomers: A nucleotide has a pentose sugar represented by a cyclopentane ring. C 1 is bonded to a B group (highlighted) that represents nitrogenous bases. C 3 is bonded to a hydroxyl group (where its hydrogen atom is highlighted) and is labeled 3 prime. C 4 labeled 5 prime is bonded to an oxygen atom on the left which is further bonded to a phosphorus atom bonded to an oxygen anion, a hydroxyl group, and double bonded to another oxygen atom. This nucleotide combines with another nucleotide that has the same structure except that the hydroxyl group of the phosphate group is highlighted. Polymers: Three nucleotides are bonded to each other by phosphodiester bonds to form a nucleic acid. Each nucleotide is bonded to B subscript 1, B subscript 2, and B subscript 3 bases respectively. Monomers: A chain conformation of allose represents a monosaccharide. C 1 is bonded to a hydroxyl group (where its hydrogen atom is highlighted). C 2, C 3, C 4, and C 5 are each bonded to hydroxyl groups. Another chain conformation of allose is present with its C 4 hydroxyl group highlighted. Polymers: Two chain conformations of monosaccharides are bonded to each other by a glycosidic bond to form a polysaccharide. C 1 of the first monosaccharide is bonded to an oxygen atom which is further bonded to C 4 of the second monosaccharide. The structure of a phospholipid shows a hydrophilic head group, indicated by a sphere, with two hydrophobic fatty acyl tails attached to the sphere. A label indicates that the hydrophilic head group has a polar group, a phosphate, and glycerol, followed by the attachment of the fatty acid chains. A schematic representation of the phospholipid bilayer shows two layers of phospholipids with hydrophilic portions facing outward, and the hydrophilic portions mix together in the interior of the bilayer. Although the actual mechanisms of covalent bond formation between monomers are complex, as we will see, the formation of a covalent bond between two monomers usually results in the net loss of a hydrogen from one monomer and a hydroxyl from the other monomer — or the net
Amino Acids Differing Only in Their Side Chains Compose Proteins
loss of one water molecule — and can therefore be thought of as a dehydration reaction. The cleavage of a covalent bond in a polymer is the result of the reverse reaction, or the addition of water, called hydrolysis. Hydrolysis can release a single monomer from the end of the polymer or can break the polymer into two shorter polymers, usually of unequal length, when the break occurs somewhere within the polymer chain. The covalent bonds that link monomers together are usually stable under normal biological conditions (e.g., neutral ), so these biopolymers are stable and can perform a wide variety of jobs in cells, such as storing information, catalyzing chemical reactions, serving as structural elements that define cellular shape and movement, and many others. Macromolecular complexes can also be assembled using noncovalent interactions. The two-ply, or bilayer, structure of cellular membranes is built up by the noncovalent assembly of many thousands of small molecules called phospholipids (see Figure 2-13). In this chapter, we focus on the chemical building blocks making up cells — amino acids, nucleotides, sugars, and phospholipids. The structure, function, and assembly of proteins, nucleic acids, polysaccharides, and biomembranes are discussed in subsequent chapters. Amino Acids Differing Only in Their Side Chains Compose Proteins The monomeric building blocks of proteins are 20 amino acids, which — when incorporated into a protein polymer — are sometimes called residues. All amino acids have a characteristic structure consisting of a central alpha carbon atom bonded to four different chemical groups (Figure 2-14): an amino group, a carboxyl or carboxylic acid group (hence the name amino acid), a hydrogen atom, and one variable group, called a side chain or R group. The carbon in all amino acids is asymmetric with the exception of glycine, whose R group is a hydrogen. Because they are asymmetric, these molecules can exist in two mirror-image forms, called by convention the D (dextro) and the L (levo) isomers (see Figure 2-4). The two isomers cannot be interconverted (one made identical to the other) without breaking and then re-forming a chemical bond in one of them. With rare exceptions, only the L forms of amino acids are found in proteins. However, D amino acids are prevalent in bacterial cell walls and other microbial products.
FIGURE 2-14 The 20 common amino acids used to build proteins. The side chain (R group; red) determines the distinctive properties of each amino acid and is the basis for grouping amino acids into three main categories: hydrophobic, hydrophilic, and special. Shown are the ionized forms that exist at the of the cytosol. In parentheses are the three-letter and one-letter abbreviations for each amino acid. Description Hydrophobic amino acids: Alanine (Ala or A): the bond-line structure of alanine has a central carbon atom bonded to a hydrogen atom on the right, a methyl group at the bottom, an alpha amino group (N H subscript 3 end subscript superscript positive) on the left and a carboxylate anion at the top. Valine (Val or V): the bond-line structure of valine has a central carbon atom bonded to a hydrogen atom on the right, a methine group at the bottom which is bonded to two methyl groups, an alpha amino group (N H subscript 3 end subscript superscript positive) on the left and a carboxylate anion at the top. Isoleucine (Ile or I): the bond-line structure of isoleucine has a central carbon atom bonded to a hydrogen atom on the right, another carbon atom at the bottom which is bonded to a hydrogen atom on the left, a methyl group on the right and a methylene group at the bottom further which is bonded to a methyl group, an alpha amino group (N H subscript 3 end subscript superscript positive) on the left and a carboxylate anion at the top. Leucine (Leu or L): the bond-line structure of leucine has a central carbon atom bonded to a hydrogen atom on the right, a methylene group at the bottom which is bonded to a methine group further bonded to two methyl groups, an alpha amino group (N H subscript 3 end subscript superscript positive) on the left and a carboxylate anion at the top.
Methionine (Met or M): The bond-line structure of methionine has a central carbon atom bonded to a hydrogen atom on the right, a four carbon chain at the bottom where C 1 and C 2 are methylene groups; C 3 is replaced by a sulfur atom and C 4 is a methyl group, an alpha amino group (N H subscript 3 end subscript superscript positive) on the left and a carboxylate anion at the top. Phenylalanine (Phe or F): The bond-line structure of methionine has a central carbon atom bonded to a hydrogen atom on the right, a methylene group at the bottom further bonded to a benzene ring, an alpha amino group (N H subscript 3 end subscript superscript positive) on the left and a carboxylate anion at the top. Tyrosine (Tyr or Y): The bond-line structure of tyrosine is similar to that of phenylalanine except that, the C 4 of the benzene ring is bonded to a hydroxyl group. Tryptophan (Trp or W): The bond-line structure of tryptophan has a central carbon atom bonded to a hydrogen atom on the right, a methylene group at the bottom which is bonded to an indole ring, an alpha amino group (N H subscript 3 end subscript superscript positive) on the left and a carboxylate anion at the top. Hydrophilic amino acids: Basic amino acids: Lysine (Lys or K): The bond-line structure of lysine has a central carbon atom bonded to a hydrogen atom on the right, a four carbon chain at the bottom where C 1, C 2, C 3, and C 4 are methylene groups; C 4 is further bonded to an amine group (N H subscript 3 end subscript superscript positive), an alpha amino group (N H subscript 3 end subscript superscript positive) on the left and a carboxylate anion at the top. Arginine (Arg or R): The bond-line structure of arginine has a central carbon atom bonded to a hydrogen atom on the right, a five carbon chain at the bottom where C 1, C 2, and C 3 are methylene groups; C 4 is replaced by an amine (N H) group; C 5 is a carbon atom which is double bonded to an amide cation and single bonded to an amide group, an alpha amino group (N H subscript 3 end subscript superscript positive) on the left and a carboxylate anion at the top. Histidine (His or H): The bond-line structure of histidine has a central carbon atom bonded to a hydrogen atom on the right, a methylene group at the bottom bonded to a protonated imidazole ring, an alpha amino group (N H subscript 3 end subscript superscript positive) on the left and a carboxylate anion at the top. Acidic amino acids: Aspartate (Asp or D): The bond-line structure of aspartate has a central carbon atom bonded to a hydrogen atom on the right, a methylene group at the bottom which is further bonded to a carboxylate anion, an alpha amino group (N H subscript 3 end subscript superscript positive) on the left and a carboxylate anion at the top. Glutamate (Glu or E): The bond-line structure of glutamate has a central carbon atom bonded to a hydrogen atom on the right, two methylene groups at the bottom which is further bonded to a carboxylate anion, an alpha amino group (N H subscript 3 end subscript superscript positive) on the left and a carboxylate anion at the top. Polar amino acids with uncharged R groups: Serine (Ser or S): The bond-line structure of serine has a central carbon atom bonded to a hydrogen atom on the right, a methylene group at the bottom which is further bonded to a hydroxyl group, an alpha amino group (N H subscript 3 end subscript superscript positive) on the left and a carboxylate anion at the top. Threonine (Thr or T): The bond-line structure of threonine has a central carbon atom bonded to a hydrogen atom on the right, another carbon atom at the bottom which is further bonded to a hydrogen atom on the left, a hydroxyl group
on the right and a methyl group at the bottom, an alpha amino group (N H subscript 3 end subscript superscript positive) on the left and a carboxylate anion at the top. Asparagine (Asn or N): The bond-line structure of asparagine has a central carbon atom bonded to a hydrogen atom on the right, a methylene group at the bottom which is further bonded to a carbonyl group further bonded to an amide group, an alpha amino group (N H subscript 3 end subscript superscript positive) on the left and a carboxylate anion at the top. Glutamine (Gln or Q): The bond-line structure of glutamine has a central carbon atom bonded to a hydrogen atom on the right, two methylene groups at the bottom which is further bonded to a carbonyl group further bonded to an amide group, an alpha amino group (N H subscript 3 end subscript superscript positive) on the left and a carboxylate anion at the top. Special amino acids: Cysteine (Cys or C): The bond-line structure of Cysteine has a central carbon atom bonded to a hydrogen atom on the right, a methylene group at the bottom which is further bonded to a thiol group, an alpha amino group (N H subscript 3 end subscript superscript positive) on the left and a carboxylate anion at the top. Glycine (Gly or G): The bond-line structure of glycine has a central carbon atom bonded to a hydrogen atom on the right, a hydrogen atom at the bottom, an alpha amino group (N H subscript 3 end subscript superscript positive) on the left and a carboxylate anion at the top. Proline (Pro or P): Proline has a pyrrolidine ring where its amine group (N H) is replaced by an amide cation, C 2 is bonded to a hydrogen atom and a carboxylate anion. C 3, C 4, and C 5 are methylene groups. To understand the three-dimensional structures and functions of proteins, discussed in detail in
Chapter 3, you must be familiar with some of the distinctive properties of amino acids. These properties are determined in part by their side chains. You need not memorize the detailed structure of each type of side chain to understand how proteins work because amino acids can be classified into several broad categories based on the size, shape, charge, hydrophobicity (a measure of water solubility), and chemical reactivity of their side chains (Figure 2-14). Amino acids with nonpolar side chains, called hydrophobic amino acids, are poorly soluble in water. The larger the nonpolar side chain, the more hydrophobic the amino acid. The side chains of alanine, valine, leucine, and isoleucine are linear or branched hydrocarbons that do not form a ring, and they are therefore called aliphatic amino acids. These amino acids are all nonpolar, as is methionine, which is similar to them except that it contains one sulfur atom. Phenylalanine, tyrosine, and tryptophan have large, hydrophobic, aromatic rings in their side chains. In later chapters, we will see in detail how hydrophobic side chains under the influence of the hydrophobic effect often pack into the interior of proteins or line the surfaces of proteins that are embedded within hydrophobic regions of biomembranes.
Amino acids with polar side chains are called hydrophilic amino acids. The most hydrophilic of these amino acids is the subset with side chains that are charged (ionized) at the typical of biological fluids both inside and outside the cell (see Section 2.3). Arginine and lysine have positively charged side chains and are called basic amino acids; aspartic acid and glutamic acid have negatively charged side chains due to the carboxylic acid groups in their side chains (their charged forms are called aspartate and glutamate) and are called acidic amino acids. A fifth amino acid, histidine, has a side chain containing a ring with two nitrogens, called imidazole. The imidazole ring can shift from being positively charged to uncharged in response to small changes in the acidity of its environment (defined as , see Section 2.3): Description At p H 5.8, the imidazole ring is a five-member ring where C 1 is replaced by a nitrogen atom bonded to a hydrogen atom, C 2 is bonded to a hydrogen atom, and is double bonded to C 3. C 3 is replaced by a nitrogen atom which is positively charged and bonded to a hydrogen atom. C 4 is bonded to a hydrogen atom and is double bonded to C 5. C 5 is bonded to a methylene group with an open bond. At p H 7.8 the imidazole structure is the same except N 3 is uncharged and is not bonded to a hydrogen atom. The activities of many proteins are modulated by shifts in environmental acidity through protonation or deprotonation of histidine side chains. Asparagine and glutamine are uncharged amino acids but have polar side chains containing amide groups with extensive hydrogen-bonding capacities. Similarly, serine and threonine are uncharged but have polar hydroxyl groups, which also participate in hydrogen bonds with other polar molecules.
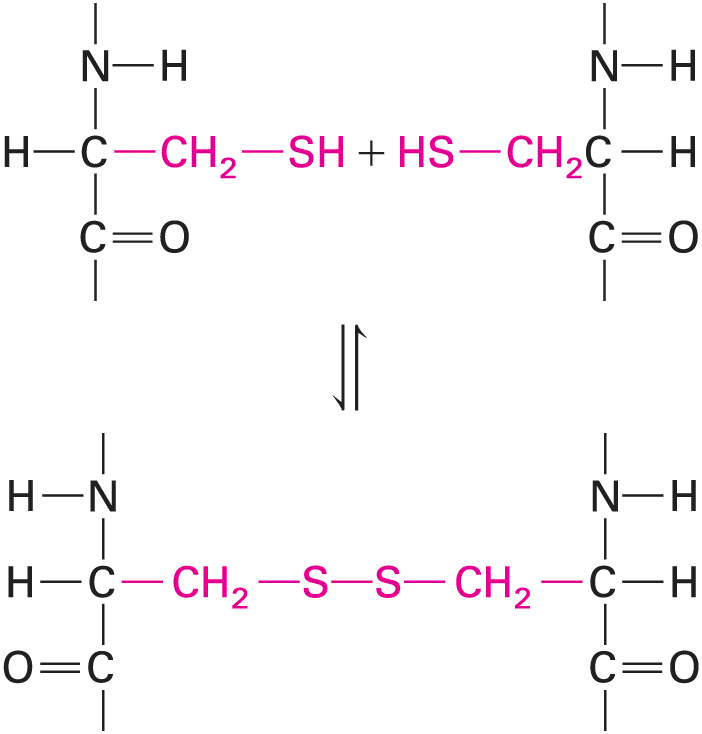
Finally, cysteine, glycine, and proline play special roles in proteins because of the unique properties of their side chains. The side chain of cysteine contains a reactive sulfhydryl group . On release of a proton a sulfhydryl group is converted into a thiolate anion . Thiolate anions in certain enzymes can play important roles in catalysis, notably in some proteases that destroy proteins. In many proteins, especially those secreted from cells or that extend outward from the plasma membrane into the extracellular fluid, each of two adjacent sulfhydryl groups can be oxidized, each releasing a proton and an electron, to form a covalent disulfide bond : Description The structure of the first reactant has a central carbon atom bonded to a hydrogen atom on the left, a nitrogen atom having an open bond at the top further bonded to a hydrogen atom, a methylene group on the right further bonded to a thiol group, and a carbonyl group with an open bond at the bottom. The second reactant is the mirror image of the first reactant. A disulfide bond is formed between these two reactants. The structure formed has two sulfur atoms bonded to each other. Each sulfur atom is further bonded to a methylene group, which is further bonded to a carbon atom. Each carbon atom is bonded to a hydrogen atom, a nitrogen atom having an open bond at the top further bonded to a hydrogen atom, and a carbonyl group with an open bond at the bottom.
Disulfide bonds serve to cross-link regions within a single polypeptide chain (intramolecular cross-linking) or between two separate chains (intermolecular cross-linking). Disulfide bonds stabilize the folded structure of some proteins. The smallest amino acid, glycine, has a single hydrogen atom as its R group. Its small size allows it to fit into tight spaces. We will see an example of this tight fit when we examine the structure of the most abundant protein in humans, collagen (see Chapter 20). Unlike the side chains of the other common amino acids, the side chain of proline (pronounced pro-leen) bends around to form a ring by covalently bonding to the nitrogen atom in the amino group attached to the . As a result, proline is very rigid, and its amino group is not available for typical hydrogen bonding. The presence of proline in a protein creates a fixed kink in the polymer chain, limiting how it can fold in the vicinity of the proline residue. Some amino acids are more abundant in proteins than others. Cysteine, tryptophan, and methionine are not common amino acids: together, they constitute approximately 5 percent of the amino acids in a typical protein. Four amino acids — leucine (hydrophobic), serine (polar), lysine (positively charged), and glutamic acid (negatively charged) — are the most abundant amino acids, constituting 32 percent of all the residues in a typical protein. However, the amino acid compositions of particular proteins may vary widely from these values. Humans and other mammals can synthesize 11 of the 20 amino acids. The other nine are called essential amino acids and must be included in the diet to permit normal protein production. These essential amino acids are phenylalanine, valine, threonine, tryptophan, isoleucine, methionine, leucine, lysine, and histidine. Adequate provision of these essential amino acids in feed is key to the livestock industry. Indeed, a genetically engineered variety of corn with a high lysine content is now in use as an enhanced feed to promote the growth of animals. Although cells use the 20 amino acids shown in Figure 2-14 in the initial synthesis of proteins, analysis of cellular proteins reveals that they contain over 100 different amino acids. The difference is due to the covalent modification by the addition of a variety of chemical groups to some amino acids after they are incorporated into proteins (Figure 2-15). One important modification is the addition of acetyl groups to amino acids, a process known as
acetylation. Acetylation of the amino group of the N-terminal residue is the most common form of amino acid covalent modification, affecting an estimated 80 percent of all proteins: Description The structure has a methyl group bonded to a carbonyl group which is further bonded to an amine group (N H). The amine group is bonded to a carbon atom bonded to a hydrogen atom at the bottom, R group at the top, and a carbonyl group with an open bond on the right. This modification may play an important role in controlling the life span of proteins within cells because many non-acetylated proteins are rapidly degraded.
FIGURE 2-15 Common modifications of amino acid side chains in proteins. These modified residues and numerous others are formed by addition of various chemical groups (red) to the amino acid side chains during or after synthesis of a polypeptide chain. Description Acetyl lysine: The bond-line structure of acetyl lysine has a methyl group bonded to a carbonyl group (acetyl group) which is further bonded to a nitrogen atom bonded to a hydrogen atom at the top. The nitrogen atom is further bonded to four methylene units which are further bonded to a methine group. The methine group is bonded to an alpha amino group and a carboxylate anion. The acetyl group is highlighted. Phosphoserine: The bond-line structure of
phosphoserine has a methine group bonded to a methylene group on the left further bonded to an oxygen atom which is further bonded to a phosphorus atom. The phosphorus atom is single bonded to two oxygen anions and double bonded to another oxygen atom (phosphate group), an alpha amino group at the bottom and a carboxylate anion on the right. The phosphate group is highlighted. Phosphotyrosine: The bond-line structure of phosphotyrosine has a methine group bonded to a methylene group on the left further bonded to a benzene ring further bonded to an oxygen atom which is further bonded to a phosphorus atom. The phosphorus atom is single bonded to two oxygen anions and double bonded to another oxygen atom (phosphate group), an alpha amino group at the bottom and a carboxylate anion on the right. The phosphate group is highlighted. Phosphothreonine: The bond-line structure of phosphothreonine has a methine group bonded to a methylene group on the left further bonded to a methyl group at the top and an oxygen atom at the left which is further bonded to a phosphorus atom. The phosphorus atom is single bonded to two oxygen anions and double bonded to another oxygen atom (phosphate group), an alpha amino group at the bottom and a carboxylate anion on the right. The phosphate group is highlighted. 3-Hydroxyproline: 3-Hydroxyproline has a pyrrolidine ring where its amine group (N H) is replaced by an amide cation; C 2 is bonded to a hydrogen atom which is further bonded to a carboxylate anion. C 3 is a methine group bonded to a hydroxyl group (highlighted). C 4 and C 5 are methylene groups. 3-Methylhistidine: The bond-line structure of 3-Methylhistidine has a methine group bonded to an alpha amino group (N H subscript 3 end subscript superscript positive) at the bottom, a carboxylate anion on the right, a methylene group on the left which is further bonded to a protonated imidazole ring. N 3 of the imidazole ring is bonded to a methyl group (highlighted). Gamma-carboxyglutamate: The bond-line structure of gamma-carboxyglutamate has a methine group bonded to a methylene group on the left further bonded to a methine group further bonded to two carboxylate anions one of which is highlighted, an alpha amino group at the bottom and a carboxylate anion on the right. The phosphate group is highlighted. O-GlcNAc-threonine: O-GlcNAc-threonine has a methine group bonded to an alpha amino group at the bottom, a carboxylate anion on the right, another methine group on the left further bonded to an oxygen atom at the left which is further bonded to C 1 of the chair conformation of N-Acetylglucosamine. C 2 is bonded to an amine (N H) group which is further bonded to a carbonyl group bonded to a methyl group. C 3, C 4, and C 5 are each bonded to a hydroxyl group. Another important modification is the addition of a phosphate group to hydroxyl groups in serine, threonine, and tyrosine residues, a process known as phosphorylation. We will encounter numerous examples of proteins whose activity is regulated by reversible phosphorylation and dephosphorylation. A well-known site of phosphorylation in bacteria, fungi, and plants is the nitrogen in the side chain of histidine, but the prevalence and functional significance of this modification in higher eukaryotes, such as mammals, has not been well-defined. Like phosphorylation and dephosphorylation, controlled methylation and demethylation of arginine and lysine side chains on proteins are important regulatory processes. For example, methylation of these side chains on proteins called histones is an important regulator of gene expression in
Five Different Nucleotides Are Used to Build Nucleic Acids
eukaryotes (see Chapter 8). The side chains of asparagine, serine, and threonine are sites for glycosylation, the attachment of linear and branched carbohydrate chains. Many secreted proteins and membrane proteins contain glycosylated residues, and the reversible modification of hydroxyl groups on specific serines and threonines by a sugar called N-acetylglucosamine also appears to regulate protein activities inside cells. Other amino acid modifications found in selected proteins include the hydroxylation of proline and lysine residues in collagen (see Chapter 20), the methylation of histidine residues in membrane receptors, and the γ-carboxylation of glutamate in blood-clotting factors such as prothrombin. Deamidation of asparagine and glutamine to form the corresponding acidic amino acids, aspartate and glutamate, also occurs. Five Different Nucleotides Are Used to Build Nucleic Acids Two types of chemically similar nucleic acids, DNA (deoxyribonucleic acid) and RNA (ribonucleic acid), are the cell’s principal molecules that carry genetic information. RNAs can also help catalyze chemical reactions. DNA and RNA polymers are built from monomers called nucleotides. All nucleotides have a common three-part structure: a phosphate group is linked by a phosphoester bond to a pentose (five-carbon) sugar, and the pentose sugar in turn is linked to a nitrogen- and carbon-containing ring structure commonly referred to as a base (Figure 2-16a). In RNA, the pentose is ribose; in DNA, it is deoxyribose, which has a proton, rather than a hydroxyl group, at position (Figure 2-16b). (We describe the structures of sugars in more detail below) The bases adenine, guanine, and cytosine (Figure 2-17) are found in both DNA and RNA; thymine is found only in DNA, and uracil is found only in RNA. The bases are often abbreviated A, G, C, T, and U, respectively; these same single-letter abbreviations are also commonly used to denote all the nucleotides in nucleic acid polymers.
FIGURE 2-16 Common structure of nucleotides. (a) Adenosine -monophosphate (AMP), a nucleotide present in RNA. By convention, the carbon atoms of the pentose sugar in nucleotides are numbered with primes. In natural nucleotides, the carbon is joined by a β linkage to the base (in this case, adenine); both the base (blue) and the phosphate on the hydroxyl (red) extend above the plane of the sugar ring. (b) Ribose and deoxyribose, the pentoses in RNA and DNA, respectively. Description The illustration labeled A shows the structure of adenosine 5 prime-monophosphate: A Haworth projection of ribose has a pyranose ring where C 1 prime is bonded to N 9 of the adenine group at the top and a hydrogen atom at the bottom. C 2 prime and C 3 prime are each bonded to a hydrogen atom at the top and a hydroxyl group at the bottom. C 4 prime and is bonded to a methylene group on top and a hydrogen atom at the bottom. The carbon atom of the methylene is labeled 5 prime and is further bonded to the oxygen atom of a phosphate group. The phosphate group has a central phosphorus atom bonded to three oxygen atoms, two of which are anions; and double bonded to another oxygen atom. The phosphate group is highlighted. Adenine has a cyclohexane ring fused with a cyclopentane ring. C 1, C 3, C 7 and C 9 are each replaced by a nitrogen atom. C 2 and C 8 are each bonded to a hydrogen atom. C 6 is bonded to an amide group. Double bonds are present between C 2 and C 3, C 4 and C 5, and N 7 and C 8. Adenine is highlighted. The illustration labeled B shows the structure of ribose and two prime deoxyribose. Ribose: A Haworth projection of ribose has a pyranose ring where C One prime is bonded to a hydroxyl group at the top and a hydrogen atom at the bottom. C 2 prime and C 3 prime are each bonded to a hydrogen atom at the top and a hydroxyl group at the bottom. The hydroxyl group bonded to C 2 prime is highlighted. C 4 prime is bonded to a hydroxymethyl group. The carbon atom of the hydroxymethyl group is labeled 5 prime. Two prime deoxyribose: A Haworth projection of 2 prime deoxyribose has a pyranose ring where C 1 prime is bonded to a hydroxyl group at the top and a hydrogen atom at the bottom. C 2 prime and C 3 prime are each bonded to a hydrogen atom at the top and a hydroxyl group at the bottom. The hydroxyl group bonded to C 2 prime is highlighted. C 4 prime is bonded to a hydroxymethyl group. The carbon atom of the hydroxymethyl group is labeled 5 prime.
FIGURE 2-17 Chemical structures of the principal bases in nucleic acids. In nucleic acids and nucleotides, nitrogen 9 of purines and nitrogen 1 of pyrimidines (red) are bonded to the carbon of ribose or deoxyribose. U is found only in RNA, and T is found only in DNA. Both RNA and DNA contain A, G, and C. Description Purines: Adenine (A): Adenine has a cyclohexane ring fused with a cyclopentane ring. C 1, C 3, C 7 and C 9 are each replaced by a nitrogen atom. C 2 and C 8 are each bonded to a hydrogen atom. C 6 is bonded to an amide group. Double bonds are present between C 2 and C 3, C 4 and C 5, and N 7 and C 8. N 9 is highlighted. Guanine (G): Guanine has a cyclohexane ring fused with a cyclopentane ring. C 1, C 3, C 7 and C 9 are each replaced by a nitrogen atom. N 1 and N 9 are each bonded to a hydrogen atom. C 2 is bonded to an amide group. C 6 is double bonded to an oxygen atom. C 8 is a methine group. Double bonds are present between C 2 and C 3, C 4 and C 5, and N 7 and C 8. N 9 is highlighted. Pyrimidine: Uracil (U): Uracil has a cyclohexane ring where C 1 and C 3 are each replaced by a nitrogen atom bonded to a hydrogen atom. C 2 and C 4 are carbonyl groups. C 5 and C 6 are methine groups. C 5 is double bonded to C 6.
Thymine (T): Thymine has a similar structure to that of uracil except, the C 5 carbon atom is bonded to a methyl group. Cytosine (C): Cytosine has a similar structure to that of uracil except that, N 3 is not bonded to a hydrogen group and it is double bonded to C 4 and C 4 is bonded to an amide group. Adenine and guanine are purines, bases that contain a pair of fused rings; cytosine, thymine, and uracil are pyrimidines, bases that contain a single ring (see Figure 2-17). In nucleotides, the carbon atom of the sugar (ribose or deoxyribose) is attached to the nitrogen atom at position 9 of a purine or at position 1 of a pyrimidine . For example, the connection of deoxyribose to a purine base in the nucleotide adenosine -monophosphate (AMP) is shown in Figure 2-16a. Due to the phosphate group, nucleotides are acidic: under normal intracellular conditions the phosphate group releases hydrogen ions becoming negatively charged (see Figure 2-16a). Most nucleic acids in cells are associated with proteins, which form both ionic interactions with the negatively charged phosphates and other noncovalent interactions with other portions of the nucleic acid. Cells and extracellular fluids in organisms contain small concentrations of nucleosides, combinations of a base and a sugar without a phosphate group. Nucleotides are nucleosides that have one, two, or three phosphate groups covalently attached to the ribose via an ester bond. An ester bond is a covalent bond linking an acid, such as a carboxylic acid or a phosphoric acid, with an alcohol. The formation of such a bond, called esterification, is accompanied by the release of a hydroxyl group from the acid and a proton from the hydroxyl group on the alcohol, which together form a water molecule. In a nucleotide, a phosphoric acid is esterified with the hydroxyl group of the ribose (see Figure 2-16a). Nucleoside monophosphates have a single esterified phosphate (see Figure 2-16a); nucleoside diphosphates contain a pyrophosphate group:
Description Pyrophosphate has a central oxygen atom which is bonded to two phosphorus atoms on the left and right, each. Each phosphorus atom is further bonded to two oxygen anions and double bonded to another oxygen atom. Nucleoside triphosphates have a third phosphate. Table 2-3 lists the names of the nucleosides and nucleotides in nucleic acids and the various forms of nucleoside phosphates. Later in this chapter (Section 2.4), we will consider in detail the structure of the nucleoside triphosphate ATP (see Figures 2-1d and 2-31). The nucleoside triphosphates are used in the synthesis of nucleic acids, as we explain in Chapter 5. Among their other functions in the cell, GTP participates in intracellular signaling and acts as an energy reservoir, particularly in protein synthesis, and ATP is the most widely used biological energy carrier.
TABLE 2-3 • Terminology of Nucleosides and Nucleotides Bases Purines Pyrimidines Adenine (A) Guanine (G) Cytosine (C) Uracil (U) Thymine (T) Adenosine Guanosine Cytidine Uridine Deoxyadenosine Deoxyguanosine Deoxycytidine Deoxythymidine Adenylate Guanylate Cytidylate Uridylate Deoxyadenylate Deoxyguanylate Deoxycytidylate Deoxythymidylate
Monosaccharides Covalently Assemble into Linear and Branched Polysaccharides
Nucleoside monophosphates AMP GMP CMP UMP Nucleoside diphosphates ADP GDP CDP UDP Nucleoside triphosphates ATP GTP CTP UTP Deoxynucleoside mono-, di-, and triphosphates dAMP, etc. dGMP, etc. dCMP, etc. dTMP, etc. Monosaccharides Covalently Assemble into Linear and Branched Polysaccharides The building blocks of the polysaccharides are the simple sugars, or monosaccharides. Monosaccharides are carbohydrates, which are literally covalently bonded combinations of carbon atoms (“carbo”) and water molecules (“hydrate”) in a one-to-one ratio , where n equals 3, 4, 5, 6, or 7. Hexoses and pentoses are the most common monosaccharides. All monosaccharides contain hydroxyl groups and either an aldehyde or a keto group: Description An aldehyde has a carbonyl group bonded to a carbon atom having three open bonds on the left and a hydrogen atom on the right. A keto group has a carbonyl group bonded to two carbon atoms each having three open bonds. Many biologically important sugars are hexoses, including glucose, mannose, and galactose (Figure 2-18). Mannose is identical to glucose except that the orientation of the groups bonded to carbon 2 is reversed — they are stereoisomers (also called epimers). Similarly, galactose, another hexose, differs from glucose only in the orientation of the groups attached to carbon 4.
Interconversion of glucose and mannose or galactose requires the breaking and making of covalent bonds; such reactions are carried out by enzymes called epimerases.
FIGURE 2-18 Chemical structures of hexoses. All hexoses have the same chemical formula and contain an aldehyde or a keto group. (a) The ring forms of D-glucose are generated from the linear molecule by reaction of the aldehyde at carbon 1 with the hydroxyl on carbon 5 or carbon 4. The three forms are readily interconvertible, although the pyranose form (right) predominates in biological systems. (b) In D-mannose and D-galactose, the configuration of the H (green) and OH (blue) bound to one carbon atom differs from that in glucose. These sugars, like glucose, exist primarily as pyranoses (six-member rings). Description In illustration A, D-Glucose forms two ring structures, D-Glucofuranose (rare) and D-Glucopyranose (common). D-Glucose: D-glucose has a six-carbon chain where C 1 is a carbonyl group bonded to a hydrogen atom. C 2, C 4, and C 5 are each bonded to a hydrogen atom on the left and a hydroxyl group on the right. The hydrogen atom and the hydroxyl group bonded to C 2 and C 4 are highlighted. C 3 is bonded to a hydroxyl group on the left and a hydrogen atom on the right. C 6 is a hydroxymethyl group. D-Glucofuranose (rare): The Haworth projection of D-Glucofuranose has a furanose ring where C 1 and C 2 are each bonded to a hydrogen atom at the top and a hydroxyl group at the bottom. C 3 is bonded to a hydroxyl group at the
top and a hydrogen atom at the bottom. C 4 is bonded to C 5 of the formaldehyde group at the top and a hydrogen atom at the bottom. C 5 is bonded to C 6 of the hydroxymethyl group. D-Glucopyranose (common): The Haworth projection of D-Glucopyranose has a pyranose ring where C 1, C 2, and C 3 are each bonded to a hydrogen atom at the top and a hydroxyl group at the bottom. C 3 is bonded to a hydroxyl group at the top and a hydrogen atom at the bottom. C 5 is bonded to a hydroxymethyl group at the top and a hydrogen atom at the bottom. Illustration B shows D-Mannose: D-Mannose has a six-carbon chain where C 1 is a carbonyl group bonded to a hydrogen atom. C 2 and C 3 are each bonded to a hydroxyl group on the left and a hydrogen atom on the right. The hydrogen atom and the hydroxyl group bonded to C 2 are highlighted. C 4 and C 5 are each bonded to a hydrogen atom on the left and a hydroxyl group on the right. C 6 is a hydroxymethyl group. D-Galactose: D-galactose has a six-carbon chain where C 1 is a carbonyl group bonded to a hydrogen atom. C 3 and C 4 are each bonded to a hydroxyl group on the left and a hydrogen atom on the right. The hydrogen atom and the hydroxyl group bonded to C 4 are highlighted. C 2 and C 5 are each bonded to a hydrogen atom on the left and a hydroxyl group on the right. C 3 is bonded to a hydroxyl group on the left and a hydrogen atom on the right. C 6 is a hydroxymethyl group. D-Glucose is one of the principal external sources of energy for most cells in complex multicellular organisms. It can exist in three different forms: a linear structure and two different hemiacetal ring structures (Figure 2-18a). If the aldehyde group on carbon 1 combines with the hydroxyl group on carbon 5, the result is the formation of D-glucopyranose, a hemiacetal containing a six-atom (usually called a six-membered) ring. In the anomer of D-glucopyranose, the hydroxyl group attached to carbon 1 points downward from the ring, as shown in Figure 2-18a; in the anomer, this hydroxyl points upward. In aqueous solution, the and anomers readily interconvert spontaneously; at equilibrium there is about one-third anomer and two-thirds , with very little of the open-chain form. Because enzymes can distinguish between the and anomers of D-glucose, these forms have distinct biological roles. Condensation of the hydroxyl group on carbon 4 of the linear glucose with its aldehyde group results in the formation of Dglucofuranose, a hemiacetal containing a five-membered ring. Although all three forms of Dglucose exist in biological systems, the pyranose (six-membered ring) form is by far the most abundant. The pyranose ring in Figure 2-18a is depicted as planar. In fact, because of the tetrahedral geometry of each of the five carbon atoms, the most stable conformation of a pyranose ring has a nonplanar, chairlike shape. In this conformation, each bond from a ring carbon to a nonring atom (e.g., H or O) is either nearly perpendicular to the ring — referred to as axial (a) — or nearly in the plane of the ring — referred to as equatorial (e):
Description Pyranoses: The chair conformation of pyranoses where C 1, C 3, and C 5 are each bonded to e and dot bonded to a. C 2 and C 4 are each bonded to a and dot bonded to e. a and e are highlighted. Alpha-D-glucopyranose: The chair conformation of alpha-D-glucopyranose where C 1, C 2, and C 4 are each bonded to a hydrogen atom and dot bonded to a hydroxyl group. C 3 is bonded to a hydroxyl group and dot bonded to a hydrogen atom. C 5 is bonded to C 6 of the hydroxymethyl group and dot bonded to a hydrogen atom. All hydrogen atoms, hydroxyl groups, and hydroxymethyl group is highlighted. Disaccharides, formed from two monosaccharides, are the simplest polysaccharides. The disaccharide lactose, composed of galactose and glucose, is the major sugar in milk; the disaccharide sucrose, composed of glucose and fructose, is a principal product of plant photosynthesis and is refined into common table sugar (Figure 2-19).
FIGURE 2-19 Formation of the disaccharides lactose and sucrose. In any glycosidic linkage, the anomeric carbon of one sugar molecule (in either the or conformation) is linked to a hydroxyl oxygen on another sugar molecule. The linkages are named accordingly; thus lactose contains a bond, and sucrose contains an bond. Description Galactose and glucose combine to form lactose. A molecule of water is eliminated in this reaction. A bond is formed between the first beta carbon of galactose and the fourth carbon of glucose.
The Haworth projection of galactose has a pyranose ring where C 1, C 3, and C 4 are each bonded to a hydroxyl group at the top and a hydrogen atom at the bottom. C 2 is bonded to a hydrogen atom at the top and a hydroxyl group at the bottom. C 5 is bonded to a hydroxymethyl group at the top and a hydrogen atom at the bottom. C 1 is labeled beta and the hydroxyl group bonded to it is highlighted. The Haworth projection of glucose has a pyranose ring where C 1 and C 3 are each bonded to a hydroxyl group at the top and a hydrogen atom at the bottom. C 2 and C 4 are each bonded to a hydrogen atom at the top and a hydroxyl group at the bottom. The hydroxyl group bonded to C 4 is highlighted. C 5 is bonded to a hydroxymethyl group at the top and a hydrogen atom at the bottom. The Haworth projection of fructose has the Haworth projections of galactose and glucose bonded together. C 1 beta of galactose is bonded to an oxygen atom which is further bonded to C 4 of glucose. This bond is highlighted. Glucose and fructose combine to form sucrose. A molecule of water is eliminated in this reaction. A bond is formed between the first alpha carbon of glucose and the fourth carbon of glucose. The Haworth projection of glucose has a pyranose ring where alpha C 1, C 2, and C 3 are each bonded to a hydrogen atom at the top and a hydroxyl group at the bottom. The hydroxyl group bonded to alpha C 1 is highlighted. C 3 is bonded to a hydroxyl group at the top and a hydrogen atom at the bottom. C 5 is bonded to a hydroxymethyl group at the top and a hydrogen atom at the bottom. The Haworth projection of fructose has a furanose ring where C 1 is a hydroxymethyl group bonded to C 2 which is further bonded to a hydroxyl group (highlighted). C 3 is bonded to a hydrogen atom at the top and a hydroxyl group at the bottom. C 4 is bonded to a hydroxyl group at the top and a hydrogen atom at the bottom. C 5 is bonded to a hydrogen atom at the top and a hydroxymethyl group at the bottom. The Haworth projection of sucrose has the Haworth projections of glucose and fructose bonded together. Alpha C 1 of glucose is bonded to an oxygen atom which is further bonded to C 2 of fructose. This bond is highlighted. Larger polysaccharides contain dozens to hundreds of monosaccharide units. They can function as reservoirs for glucose, as structural components, or as adhesives that help hold cells together in tissues. The most common storage carbohydrate in animal cells is glycogen, a very long, highly branched polymer of glucose. As much as 10 percent of the liver by weight can be glycogen. The primary storage carbohydrate in plant cells, starch, is also a glucose polymer. It occurs in an unbranched form (amylose) and a lightly branched form (amylopectin). Both glycogen and starch are composed of the anomer of glucose. In contrast, cellulose, the major constituent of plant cell walls, is an unbranched polymer of the anomer of glucose. Cellulose confers stiffness to many plant structures (see Chapter 20). Human digestive enzymes can hydrolyze the glycosidic bonds in starch but not the glycosidic bonds in cellulose. Many species of plants, bacteria, and molds produce cellulose-degrading enzymes. Cows and termites can break down cellulose because they harbor cellulose-degrading bacteria in their gut. Bacterial cell walls consist of peptidoglycan rather than cellulose. A peptidoglycan has a polysaccharide chain cross-linked by peptide cross-
bridges, which confer rigidity and cell shape. Human tears and gastrointestinal fluids contain lysozyme, an enzyme capable of hydrolyzing peptidoglycan in the bacterial cell wall. The enzymes that make the glycosidic bonds, linking monosaccharides into polysaccharides, are specific for the or anomer of one sugar and a particular hydroxyl group on the other. In principle, any two sugar molecules can be linked in a variety of ways because each monosaccharide has multiple hydroxyl groups that can participate in the formation of glycosidic bonds. Furthermore, any one monosaccharide has the potential to be linked to more than one other monosaccharide, thus generating a branch point and nonlinear polymers. Glycosidic bonds are usually formed between the growing polysaccharide chain and a covalently modified form of a monosaccharide. Such modifications include the addition of a phosphate (e.g., glucose 6phosphate) or a nucleotide (e.g., UDP-galactose): Description Glucose 6-phosphate: The Haworth projection of glucose 6-phosphate has a pyranose ring where C 1, C 2, and C 3 are each bonded to a hydrogen atom at the top and a hydroxyl group at the bottom. C 3 is bonded to a hydroxyl group at the top and a hydrogen atom at the bottom. C 5 is bonded to C 6 of a methyl group at the top which is further bonded to a phosphate group and a hydrogen atom at the bottom. The phosphate group is highlighted. U D P-galactose: The Haworth projection of U D P-galactose has a pyranose ring where C 1 is bonded to a hydrogen atom at the top and two phosphate groups which are further bonded to a uridine group at the bottom. The phosphate group has a central phosphorus atom bonded to three oxygen atoms of which one is an oxygen anion; and is double bonded to another oxygen atom. The phosphate groups bonded to uridine is highlighted. C 2 is bonded to a hydrogen atom at the top and a hydroxyl group at the bottom. C 3 and C 4 are each bonded to a hydroxyl group at the top and a hydrogen atom at the bottom. C 5 is bonded to C 6 of a hydroxymethyl group. The epimerase enzymes that interconvert different monosaccharides often do so using the nucleotide sugars rather than the unmodified, or free, sugars.
Phospholipids Associate Noncovalently to Form the Basic Bilayer Structure of Biomembranes
Many complex polysaccharides contain modified sugars that are covalently linked to various small groups, particularly amino, sulfate, and acetyl groups. Such modifications are abundant in glycosaminoglycans, major polysaccharide components of the extracellular matrix that we describe in Chapter 20. As you can see here and in many examples throughout this book, a relatively simple set of monosaccharides can be assembled into a wide variety of polysaccharides with dramatically different biological functions. Phospholipids Associate Noncovalently to Form the Basic Bilayer Structure of Biomembranes Biomembranes are large, flexible sheets with a two-ply, or bilayer, structure. They serve as the boundaries of cells and their intracellular organelles and form the outer surfaces of some viruses. Membranes literally define what is a cell (the outer membrane and the contents within the membrane) and what is not (the extracellular space outside the membrane). Unlike proteins, nucleic acids, and polysaccharides, membranes are assembled by the noncovalent association of their component building blocks. The primary building blocks of all biomembranes are phospholipids, whose physical properties are responsible for the formation of the sheet-like bilayer structure of membranes. In addition to phospholipids, biomembranes can contain a variety of other molecules, including cholesterol, glycolipids, and proteins. The structure and function of biomembranes will be described in detail in Chapter 10. Here we will focus on the phospholipids in biomembranes. To understand the structure of a phospholipid molecule, we have to understand each of its component parts and how they are assembled. As we will see shortly, a phospholipid molecule consists of two long-chain, nonpolar fatty acid groups linked (usually by an ester bond) to small, highly polar groups. These polar groups typically include a short organic molecule such as glycerol (trihydroxy propane), a phosphate, and typically a small organic molecule (Figure 2-20).
FIGURE 2-20 Phosphatidylcholine, a typical phosphoglyceride. All phosphoglycerides are amphipathic phospholipids, having a hydrophobic tail (yellow) and a hydrophilic head (blue) in which glycerol is linked via a phosphate group to an alcohol. Either or both of the fatty acyl side chains in a phosphoglyceride may be saturated or unsaturated. In phosphatidic acid (red), the simplest phospholipid, the phosphate is not linked to an alcohol. Description Hydrophilic head: Choline has a central nitrogen cation bonded to three methyl groups and two methylene units. One of the methylene units of choline is bonded to one of the oxygen atoms of a phosphate group. The phosphate group has a central phosphorus atom bonded to three oxygen atoms of which one is an oxygen anion; and is double bonded to another oxygen atom. The other oxygen atom of phosphate is bonded to a methylene group which is further bonded to a methine group which is further bonded to another methylene group. The methine and the methylene groups are each further bonded to an oxygen atom which is further bonded to a carbonyl group. Each carbonyl group are further bonded to a long fatty acid chain. The fatty acid chain makes up the hydrophobic tail. Fatty acids consist of a hydrocarbon chain attached to a carboxyl group . Like glucose, fatty acids are an important energy source for many cells (see Chapter 12). They differ in length, although the predominant fatty acids in cells have an even number of carbon atoms, usually 14, 16, 18, or 20. The major fatty acids in phospholipids are listed in Table 2-4. Fatty acids are often designated by the abbreviation Cx:y, where x is the number of carbons in the chain and y is the number of double bonds. Fatty acids containing 12 or more carbon atoms are nearly insoluble in aqueous solutions because of their long hydrophobic hydrocarbon chains.
TABLE 2-4 • Fatty Acids That Predominate in Phospholipids Common Name of Acid (ionized form in parentheses) Abbreviation Chemical Formula Saturated Fatty Acids Myristic (myristate) C14:0 Palmitic (palmitate) C16:0 Stearic (stearate) C18:0 Unsaturated Fatty Acids Oleic (oleate) C18:1 Linoleic (linoleate) C18:2 Arachidonic (arachidonate) C20:4
Fatty acids in which all the carbon-carbon bonds are single bonds — that is, the fatty acids have no carbon-carbon double bonds — are said to be saturated; those with at least one carbon-carbon double bond are called unsaturated. Unsaturated fatty acids with more than one carbon-carbon double bond are referred to as polyunsaturated. Two essential polyunsaturated fatty acids, linoleic acid (C18:2) and linolenic acid (C18:3), cannot be synthesized by mammals and must be supplied in their diet. Mammals can synthesize other common fatty acids. In phospholipids, fatty acids are covalently attached to another molecule by esterification. In the combined molecule formed by this reaction, the part derived from the fatty acid is called an acyl group, or fatty acyl group. The most common phospholipids are the phosphoglycerides, which contain two acyl groups attached to two of the three hydroxyl groups of glycerol (see Figure 2-20). Other types of phospholipids are described in Chapter 10 (also see Figure 10-8). In phosphoglycerides, one hydroxyl group of the glycerol is esterified to phosphate while the other two are normally esterified to fatty acids. The simplest phospholipid, phosphatidic acid, contains only these components. Phospholipids such as phosphatidic acids are not only membrane building blocks but also important signaling molecules. Lysophosphatidic acid, for example, can be a potent inducer of cell division (called a mitogen). In this phospholipid, the acyl chain at the 2 position (attached to the hydroxyl group on the central carbon of the glycerol) has been removed, so the molecule is relatively water soluble. In most phospholipids found in membranes, the phosphate group is also esterified to a hydroxyl group on another hydrophilic compound. In phosphatidylcholine, for example, the phosphate is covalently attached to the small molecule called choline (see Figure 2-20). The negatively charged phosphate, as well as the charged or polar groups esterified to it, can interact strongly with water. The phosphate and its associated esterified group constitute the head group of a phospholipid, which is hydrophilic, whereas the fatty acyl chains, the tails, are hydrophobic. Other common phosphoglycerides and associated head groups are shown in Table 25. Molecules such as phospholipids that have both hydrophobic and hydrophilic regions are called amphipathic. In Chapter 10, we will see how the amphipathic properties of phospholipids allow their assembly into sheet-like bilayers in which the fatty acyl tails point into the center of the sheet and the head groups point outward toward the aqueous environment (see Figure 2-13 bottom).
TABLE 2-5 • Common Phosphoglycerides and Head Groups Common Phosphoglycerides Head Group
Phosphatidylcholine Phosphatidylethanolamine Phosphatidylserine Phosphatidylinositol Fatty acyl groups also can be covalently linked in other fatty molecules, including triacylglycerols (or triglycerides), which contain three acyl groups esterified to glycerol:
Description Triacylglycerol has a methylene group bonded to a methine group bonded to another methylene group. Each of these groups are further bonded to fatty acid chains made of an oxygen atom which are further bonded to a carbonyl group, further bonded to n units of methylene units which is further bonded to a methyl group. The fatty acid chains are highlighted. They can also be covalently attached to the very hydrophobic molecule cholesterol, an alcohol, to form cholesteryl esters:
Description Cholesterol: It has four fused rings in which the first, second, and third are six membered and the fourth ring is five membered. C 3 is single bonded to a hydroxyl group. C 5 is double bonded to C 6. C 10 and C 13 are each bonded to a methyl group. C 17 is bonded to a carbon atom. The carbon atom is bonded to a methyl group and to a five-carbon chain. C 4 and C 5 of the five carbon chain are each bonded to a methyl group. Cholesterol ester: It has a cholesterol molecule where C 3 is bonded to an oxygen atom which is further bonded to a carbonyl group and a 17 membered carbon chain. Triglycerides and cholesteryl esters are extremely water-insoluble molecules in which fatty acids and cholesterol are either stored or transported. Triglycerides are the storage form of fatty acids in the fat cells of adipose tissue and are the principal components of dietary fats. Cholesteryl esters and triglycerides are transported between tissues through the bloodstream in specialized carriers called lipoproteins (see Chapter 14). We saw previously that the fatty acids, which are key components of both phospholipids and triglycerides, can be either saturated or unsaturated. An important consequence of the carboncarbon double bond in an unsaturated fatty acid is that two stereoisomeric configurations, cis and trans, are possible around each of these bonds:
Description The cis form has two carbon atoms double bonded to each other. Each carbon atom is bonded to a methylene group with an open bond at the top and a hydrogen atom at the bottom. The trans form has two carbon atoms double bonded to each other. The first carbon atom is bonded to a methylene group with an open bond at the top and a hydrogen atom at the bottom. The second carbon atom is bonded to a hydrogen atom at the top and a methylene group with an open bond at the bottom. A cis double bond introduces a rigid kink in the otherwise flexible straight acyl chain of a saturated fatty acid (Figure 2-21). In general, the unsaturated fatty acids in biological systems contain only cis double bonds. Saturated fatty acids without the kink can pack together tightly and so have higher melting points than unsaturated fatty acids. The main fatty molecules in butter are triglycerides with saturated fatty acyl chains, which is why butter is usually solid at room temperature. Unsaturated fatty acids or fatty acyl chains with the cis double bond kink cannot pack as closely together as saturated fatty acyl chains. Thus vegetable oils, composed of triglycerides with unsaturated fatty acyl groups, usually are liquid at room temperature. Vegetable and similar oils may be partially hydrogenated to convert some of their unsaturated fatty acyl chains to saturated fatty acyl chains. As a consequence, the hydrogenated vegetable oil can be molded into solid sticks of margarine. A by-product of the hydrogenation reaction is the conversion of some of the fatty acyl chains into trans fatty acids, popularly called trans fats. These trans fats, found in partially hydrogenated margarine and other food products, are not natural. Saturated and trans fatty acids have similar physical properties; for example, they tend to be solids at room temperature. Their consumption, relative to the consumption of unsaturated fats, is associated with increased plasma cholesterol levels and is discouraged by some nutritionists.
FIGURE 2-21 The effect of a double bond on the shape of fatty acids. Shown are chemical structures of the ionized form of palmitic acid, a saturated fatty acid with 16 C atoms, and oleic acid, an unsaturated one with 18 C atoms. In saturated fatty acids, the hydrocarbon chain is often linear; the cis double bond in oleate creates a rigid kink in the hydrocarbon chain. Description Palmitate: The bond-line structure of palmitate has a 15 carbon chain where C 1 is bonded to a methyl group on the left, and a hydrogen atom each on the top and bottom. C 2 to C 14 are each bonded to one hydrogen atom on the top and one hydrogen atom at the bottom. C 15 is double bonded to an oxygen atom and single bonded to an oxygen anion. Oleate: The bond-line structure of oleate has an 18 carbon chain where C 1 is double bonded to an oxygen atom and single bonded to an oxygen anion. C 2 to C 8 and C 11 to C 17 are each bonded to one hydrogen atom on the top and one hydrogen atom at the bottom. C 9 and C 10 are each bonded to a hydrogen atom and are double bonded to each other. C 18 is a methyl group. KEY CONCEPTS OF SECTION 2.2 Chemical Building Blocks of Cells Macromolecules are polymers of monomer subunits linked together by covalent bonds via dehydration reactions. Three major types of macromolecules are found in cells: proteins, composed of amino acids linked by peptide bonds; nucleic acids, composed of nucleotides linked by phosphodiester bonds; and polysaccharides, composed of monosaccharides (sugars) linked by glycosidic bonds (see Figure 2-13). Phospholipids, the fourth major chemical building block, assemble noncovalently into biomembranes. Differences in the size, shape, charge, hydrophobicity, and reactivity of the side chains of the 20 common amino acids determine the chemical and structural properties of proteins (see Figure 2-14). The three general categories into which the side chains fall are hydrophobic, hydrophilic (basic, acidic, polar), and special (see Figure 2-14). It is helpful to remember which amino acids fall into each of these categories. The bases in the nucleotides composing DNA and RNA are carbon- and nitrogen-containing rings attached to a pentose sugar. They form two groups: the purines, with two rings — adenine (A) and guanine (G) — and the pyrimidines, with one ring — cytosine (C), thymine (T), and uracil (U) (see Figure 2-17). A, G, T, and C are found in DNA, and A, G, U, and C are found in RNA. Glucose and other hexoses can exist in three forms: an open-chain linear structure, a six-member (pyranose) ring, and a five-member (furanose) ring (see Figure 2-18). In biological systems, the pyranose form of D-glucose predominates. Glycosidic bonds are formed between either the or the anomer of one sugar and a hydroxyl group on another sugar, leading to formation of disaccharides and other polysaccharides (see Figure 2-19).
Phospholipids are amphipathic molecules with a hydrophobic tail (often two fatty acyl chains) connected by a small organic molecule (often glycerol) to a hydrophilic head (see Figure 2-20). The long hydrocarbon chain of a fatty acid may be saturated (containing no carbon-carbon double bonds) or unsaturated (containing one or more double bonds). Fatty substances such as butter that have primarily saturated fatty acyl chains tend to be solid at room temperature, whereas unsaturated fats with cis double bonds have kinked chains that cannot pack closely together and so tend to be liquids at room temperature.
A Chemical Reaction Is in Equilibrium When the Rates of the Forward and Reverse Reactions Are Equal
2.3 Chemical Reactions and Chemical Equilibrium We now shift our discussion to chemical reactions in which bonds, primarily covalent bonds in reactant chemicals, are broken and new bonds are formed to generate reaction products. At any one time, several hundred different kinds of chemical reactions are occurring simultaneously in every cell, and many chemicals can, in principle, undergo multiple chemical reactions. Both the extent to which reactions can proceed and the rate at which they take place determine the chemical composition of cells. In this section, we discuss the concepts of equilibrium and steady state as well as dissociation constants and . These concepts will arise again and again throughout this textbook, so it is important for you to be familiar with them. In Section 2.4, we discuss how energy influences the extents and rates of chemical reactions. A Chemical Reaction Is in Equilibrium When the Rates of the Forward and Reverse Reactions Are Equal When reactants first mix together — before any products have been formed — the rate of the forward reaction to form products is determined
in part by the reactants’ initial concentrations, which determine the likelihood of reactants bumping into one another and reacting (Figure 222). As the reaction products accumulate, the concentration of each reactant decreases, and so does the forward reaction rate. Meanwhile, some of the product molecules begin to participate in the reverse reaction, which re-forms the reactants. The ability of a reaction to go backward is called microscopic reversibility. The reverse reaction is slow at first but speeds up as the concentration of product increases. Eventually, the rates of the forward and reverse reactions become equal, so that the concentrations of reactants and products stop changing. The system is then said to be in chemical equilibrium (plural, equilibria).
The Equilibrium Constant Reflects the Extent of a Chemical Reaction
FIGURE 2-22 Time dependence of the rates of a chemical reaction. The forward and reverse rates of a reaction depend in part on the initial concentrations of reactants and products. The net forward reaction rate slows as the concentration of reactants decreases, whereas the net reverse reaction rate increases as the concentration of products increases. At equilibrium, the rates of the forward and reverse reactions are equal, and the concentrations of reactants and products remain constant. Description The vertical axis represents time and the horizontal axis represents reaction rate. Two curves are shown, the rate of reverse reaction (in blue) and the rate of forward reaction (in purple). Both lines converge with time as the system reaches chemical equilibrium, where the forward and reverse rates are equal and there is no change in the concentration of reactants and products. A label states that the beginning of the time axis is the point of first mixing of the reactants, where the initial concentration of products is zero. The rate of reverse reaction rises rapidly as the concentration of products increases, before plateauing. The rate of forward reaction decreases as the concentration of reactants decreases. The ratio of the concentrations of the products to the concentrations of the reactants when they reach equilibrium, called the equilibrium constant , is a fixed value. Thus provides a measure of the extent to which a reaction occurs by the time it reaches equilibrium. The rate of a chemical reaction can be increased by a catalyst, a chemical that accelerates the making and breaking of covalent bonds in other molecules but itself is not permanently changed during a reaction. Although a catalyst increases the reaction rate, it does not change the equilibrium constant (see Section 2.4).
The Equilibrium Constant Reflects the Extent of a Chemical Reaction For any chemical reaction, depends on the chemical nature of the reactants and products, the temperature, and the pressure (particularly in reactions involving gases). Under standard physical conditions ( and 1 atm pressure for biological systems), is always the same for a given reaction, whether or not a catalyst is present. Consider the general reaction with three reactants and three products, (2-1) where capital letters represent particular molecules or atoms and lowercase letters represent the number of each in the reaction. For this reaction, the formula for the equilibrium constant is given by (2-2) where brackets denote the concentrations of the molecules. In Equation 22, the concentrations of reactants and products are those present at equilibrium. The rate of the forward reaction (left to right in Equation 2-1) is
Chemical Reactions in Cells Are at Steady State
where is the rate constant for the forward reaction. Similarly, the rate of the reverse reaction (right to left in Equation 2-1) is where is the rate constant for the reverse reaction. These reaction rate equations apply whether or not the reaction has reached equilibrium. It is important to remember that the forward and reverse rates of a reaction can change because of changes in reactant or product concentrations, yet at the same time the forward and reverse rate constants do not change; hence the name constant. Confusing rates and rate constants is a common error. At equilibrium the forward and reverse rates are equal, so . By rearranging these equations, we can express the equilibrium constant as the ratio of the rate constants: (2-3) The concept of is particularly helpful when we want to think about the energy that is released or absorbed when a chemical reaction occurs. We will discuss this concept in considerable detail in Section 2.4. Chemical Reactions in Cells Are at Steady State Under appropriate conditions and given sufficient time, a single biochemical reaction carried out in a test tube eventually reaches
equilibrium, at which the concentrations of reactants and products do not change with time because the rates of the forward and reverse reactions are equal. Within cells, however, many reactions are linked in pathways in which the fate of the product of one reaction is not simply to remain unchanged or to be reconverted via a reverse reaction to the reactants. For example, the product of one reaction might serve as a reactant in another reaction, or it might be pumped out of the cell. In this more complex situation, the original reaction can never reach equilibrium because some of the product molecules do not have a chance to be converted back to reactants. Nevertheless, in such non-equilibrium conditions, the rate of formation of a substance can be equal to the rate of its consumption, and as a consequence, the concentration of that substance remains constant over time. In such circumstances, the system of linked reactions for producing and consuming that substance is said to be in a steady state (Figure 2-23). One consequence of such linked reactions is that they prevent the accumulation of excess intermediates, protecting cells from the harmful effects of intermediates that are toxic at high concentrations. When the concentration of a product of an ongoing reaction is not changing over time, it might be a consequence of a state of equilibrium, or it might be a consequence of a steady state. In biological systems, when metabolite concentrations, such as blood glucose levels, are not changing with time — a condition called homeostasis (from the Greek homoios meaning “similar” and stasis meaning “standing still or stability”) — it is a consequence of a steady state rather than equilibrium.
Dissociation Constants of Binding Reactions Reflect the Affinity of Interacting Molecules
FIGURE 2-23 Comparison of reactions at equilibrium and at steady state. (a) In the test tube, a biochemical reaction eventually reaches equilibrium, at which the rates of the forward and reverse reactions are equal (as indicated by the reaction arrows of equal length), and the relative concentrations of the reactants (A) and products (B) do not change (in this example, the B:A ratio at equilibrium is 9:3). (b) In metabolic pathways within cells, the product B is commonly consumed — in this example, by conversion to C. A pathway of linked reactions is at steady state when the rate of formation of the intermediates (e.g., B) equals its rate of consumption. As indicated by the unequal length of the arrows, the individual reversible reactions constituting a metabolic pathway do not reach equilibrium. Moreover, the concentrations of the intermediates at steady state can differ from what they would be at equilibrium. Description The reversible arrows are of equal length. There are 3 As in a row and 3 rows of 3 Bs each. In both cases, the forward reaction has a longer arrow than that of the reverse reaction. There are 2 As, 2 rows of 3 Bs each, and 2 rows of 2 Cs each.
Dissociation Constants of Binding Reactions Reflect the Affinity of Interacting Molecules The concept of equilibrium also applies to the binding of one molecule to another even when there are no covalent changes to either molecule. Many important cellular processes depend on such binding reactions, which involve the making and breaking of various noncovalent interactions rather than covalent bonds, as discussed above. In some cases, the noncovalent binding can be relatively weak, or loose; in other cases the binding can be very strong, or tight. A common example is the binding of a ligand (e.g., the hormone insulin or adrenaline) to its receptor on the surface of a cell, which triggers an intracellular signaling pathway (see
Chapter 15). Another example is the binding of a protein to a specific sequence of bases in a molecule of DNA, which frequently causes the expression of a nearby gene to increase or decrease (see Chapter 8). Here we will review the most common approach used to think about how tightly or loosely two molecules bind to one another. To illustrate the general approach, let’s calculate the extent to which a protein (P) binds to a particular, short sequence of As, Ts, Gs, and Cs present in a relatively long DNA molecule (D) at equilibrium, forming a protein-DNA complex (PD):
Most commonly, binding reactions are described in terms of the dissociation constant which is the reciprocal of the equilibrium constant . For this binding reaction, the dissociation constant is calculated from the concentrations of the three components when they are at equilibrium by (2-4) It is worth noting that in such a binding reaction, when half of the DNA is bound to the protein (PD) and the other half is unbound (D), then the concentration of PD is equal to that of D, , and the concentration of P ([P]) is equal to . The lower the , the tighter that P binds to D — often referred to as a higher binding affinity of P for D — and thus the lower the concentration of P needed to bind to half of D. This can sometimes lead to a bit of confusion. Remember, the lower the , the higher the binding affinity, and the tighter the binding. Typically, a protein’s binding to a specific DNA sequence exhibits a of where M symbolizes molarity, or moles per liter . To relate the magnitude of this dissociation constant to the intracellular ratio of bound to unbound DNA, let’s consider the simple example of a bacterial cell having a volume of and containing 1 molecule of DNA and 10 molecules of the DNA-binding protein P. In this case, given a of and the total concentration of the P in the cell ( about a hundredfold higher than the ), 99 percent of the time the specific DNA sequence to which P binds will have a molecule of protein P
bound to it and 1 percent of the time it will not, even though the cell contains only 10 molecules of the protein! Clearly P and D have a high affinity for each other and bind tightly, as reflected by the low value of the dissociation constant for their binding. For protein-protein and proteinDNA binding, values of (nanomolar) or less are considered to be tight, (micromolar) modestly tight, and (millimolar) relatively weak. A large biological macromolecule, such as a protein, can have multiple binding surfaces for binding several molecules simultaneously (Figure 224). In some cases, these binding reactions are independent, each with its own distinct value that is independent of the others. In other cases, binding of a molecule at one site on a macromolecule can change the three-dimensional shape, or conformation, of a distant site (a phenomenon called allostery, see Chapter 3). That change in conformation alters the site’s binding interactions with some other molecule (altering the ). The covalent modifications of amino acid side chains — mentioned previously — often influence the three-dimensional shapes of binding sites required for such binding interactions. These covalent and noncovalent binding reactions are important mechanisms by which one molecule can alter, and thus regulate, the structure and binding activity of another. We examine this regulatory mechanism in more detail in Chapter 3.
FIGURE 2-24 Macromolecules can have distinct binding sites for multiple ligands. A large macromolecule (e.g., a protein, purple) with three distinct binding sites (yellow, A–C) is shown; each of the three binding sites exhibit molecular complementarity to three different binding partners (ligands A–C) with distinct dissociation constants . Description A three dimensional model shows a large macromolecule in purple color. Ligand A is represented by a small 3-D model colored green. The dissociation constant of the binding site A is K subscript d A. Ligand B is represented by a space-filling model made up of carbon, oxygen, and nitrogen atoms. The dissociation constant of the binding site B is K subscript d B. Ligand C is a polysaccharide which is represented by a stick-model made of carbon, hydrogen, and oxygen atoms. The dissociation constant of the binding site C is K subscript d C.
Biological Fluids Have Characteristic pH Values
Biological Fluids Have Characteristic Values The solvent inside cells and in all extracellular fluids is water. An important characteristic of any aqueous solution is the concentration of positively charged hydrogen ions and negatively charged hydroxyl ions . Because these ions are the dissociation products of they are constituents of all living systems, and they are liberated by many reactions that take place between molecules within cells. These ions can also be transported into or out of cells, as when highly acidic gastric juice is secreted by cells lining the walls of the stomach. When a water molecule dissociates, one of its polar bonds breaks. The resulting hydrogen ion, referred to as a proton, has a short lifetime as a free ion and quickly combines with a water molecule to form a hydronium ion . For convenience, we refer to the concentration of hydrogen ions in a solution, , even though this quantity really represents the concentration of hydronium ions, . Dissociation of generates one ion along with each . The dissociation of water is a reversible reaction: At , so that in pure water, .
The concentration of hydrogen ions in a solution is expressed conventionally as its pH, defined as the negative log of the hydrogen ion concentration. The of pure water at is 7: It is important to keep in mind that a one-unit difference in represents a tenfold difference in the concentration of protons. On the scale, 7.0 is considered neutral: values below 7.0 indicate acidic solutions (higher ), and values above 7.0 indicate basic, or alkaline, solutions (Figure 225). For instance, gastric juice, which is rich in hydrochloric acid , has a of about 1. Its is roughly 1-million-fold greater than that of cytoplasm, which has a of about 7.2–7.4.
Hydrogen Ions Are Released by Acids and Taken Up by Bases
FIGURE 2-25 Some values for common solutions. The of an aqueous solution is the negative log of the hydrogen ion concentration. The values for most intracellular and extracellular biological fluids are near 7 and are carefully regulated to permit the proper functioning of cells, organelles, and cellular secretions. The values for solutions of ammonia and hydrochloric acid are for one molar (1 M) solutions. Description The p H scale ranges from 0 to 14, 0 being acidic, 7 neutral, and 14 basic. Several examples are given, from bottom (most acid) to top (most basic): hydrochloric acid, p H 0; gastric juice, p H 1; grapefruit juice, p H 3; the interior of a lysosome, p H 4; the p H of urine, p H 6; an unfertilized egg, around pH 7; a fertilized egg, slightly above p H 7; the interior of a cell, slightly below p H 8; seawater, p H 9 to 10; household, p H 12; and sodium hydroxide, p H 14. Although the cytosol of cells normally has a of about 7.2, the interior of certain organelles in eukaryotic cells (see Chapter 1) can have a much lower . The internal (luminal) fluid in lysosomes, for example, has a of about 4.5. The many degradative enzymes within lysosomes function optimally in an acidic environment, whereas their action is inhibited in the near neutral environment of the cytoplasm. As this example illustrates, maintenance of a particular is essential for the proper functioning of some cellular structures. On the other hand, dramatic shifts in cellular may play an important role in controlling cellular activity. For example, the of the cytoplasm of an unfertilized egg of the sea urchin, an aquatic animal, is 6.6. Within 1 minute of fertilization, however, the rises to 7.2; that is, the decreases to about one-fourth its original value, a change that is necessary for subsequent growth and division of the egg.
Hydrogen Ions Are Released by Acids and Taken Up by Bases In general, an acid is any molecule, ion, or chemical group that tends to release a hydrogen ion . Examples are the carboxyl group , which tends to dissociate to form the negatively charged carboxylate ion and a hydrogen ion, and hydrochloric acid . Conversely, a base is any molecule, ion, or chemical group that readily combines with an . Examples are the hydroxyl ion ; ammonia which forms an ammonium ion ; and the amino group . When an acid is added to an aqueous solution, the increases, and the goes down. Conversely, when a base is added to an aqueous solution, the decreases, and the goes up. Because , any increase in is coupled with a commensurate decrease in , and vice versa. Many biological molecules contain both acidic and basic groups. For example, in neutral solutions many amino acids exist predominantly in the doubly ionized form, in which the carboxyl group has lost a proton and the amino group has accepted one:
Description A central carbon atom is bonded to a hydrogen atom on the left, an R group at the bottom, a carboxylate anion on the right, and an ammonium cation at the top. where R represents the uncharged side chain. Such a molecule, containing an equal number of positive and negative ions, is called a zwitterion. Zwitterions, having no net charge, are neutral. At extreme values, only one of these two ionizable groups of an amino acid is charged: the at low or the at high . The dissociation reaction for an acid (or acid group in a larger molecule), , can be written as . The equilibrium constant for this reaction, denoted (the subscript a stands for “acid”), is defined as . Taking the logarithm of both sides and rearranging the result yields a very useful relation between the equilibrium constant and :
(2-5) where equals . From this expression, commonly known as the Henderson–Hasselbalch equation, it can be seen that the of any acid is equal to the at which half the molecules are dissociated and half are neutral (undissociated). This is because when , then , and thus . The Henderson–Hasselbalch equation allows us to calculate the degree of dissociation of an acid — that is, the ratio of dissociated and undissociated forms — if both the of the solution and the of the acid are known. Experimentally, by measuring the and as a function of the solution’s , one can calculate the of the acid and thus the equilibrium constant for the dissociation reaction (Figure 2-26). Knowing the of a molecule not only provides an important description of its properties, but also allows us to exploit these properties to manipulate the acidity of an aqueous solution and to understand how biological systems control this critical characteristic of their aqueous fluids.
FIGURE 2-26 The relationship between , , and the dissociation of an acid. As the of a solution of carbonic acid rises from 0 to 8.5, the percentage of the compound in the undissociated, or un-ionized, form decreases from 100 percent and that of the ionized form increases from 0 percent. When the is equal to the acid’s , half of the carbonic acid has ionized. When the rises to above 8, virtually all of the acid has ionized to the bicarbonate form . Description The vertical axis represents p H ranging from 0 to 8, in increments of 2 (7.4 is also labeled). The horizontal axis represents percentage of carbonic acid or bicarbonate molecules ranging from 0 to 100, in increments of 50. A blue curve shows the results for bicarbonate and a purple curve shows the results for hydrogen carbonate. The percentage of hydrogen carbon remains constant until about pH 4, then falling to zero at about pH 8. The curve for bicarbonate is zero until pH 4, where it begins to rise, crossing the curve for hydrogen carbonate at pH 6.4, and rising to 100% at pH 8. The p
Buffers Maintain the pH of Intracellular and Extracellular Fluids
K a is listed as equal to the intersection of these curves, i.e., the p K a of hydrogen carbonate is 6.4. The graph also has the following reaction. Carbonic acid yields bicarbonate anion and a proton. Buffers Maintain the of Intracellular and Extracellular Fluids A living, actively metabolizing cell must maintain a constant in the cytoplasm of about 7.2–7.4, and it must do so even as its metabolism is producing many acids. Cells have a reservoir of weak bases and weak acids, called buffers, which ensure that the cell’s cytoplasmic remains relatively constant despite small fluctuations in the amounts of or being generated by metabolism or by the uptake or secretion of molecules and ions by the cell. Buffers maintain a constant by soaking up excess or when these ions are added to the cell or are produced by metabolism. As we shall see, buffers are most effective at preventing changes in when the of the solution is similar to the of the buffer. Consider a buffered solution whose is equal to the of the buffer . If additional acid (or base) is added to this solution, the of the solution changes, but it changes less than it would if the buffer had not been present. This is because protons released by the added acid are taken up by the ionized form of the buffer ; likewise, hydroxyl ions released by the added base are neutralized by protons released by the undissociated buffer . The capacity of a buffer or any other substance
to release hydrogen ions or take them up depends partly on the extent to which the substance has already taken up or released protons, which in turn depends on the of the solution relative to the of the substance. The ability of a buffer to minimize changes in , its buffering capacity, depends on the concentration of the buffer and the relationship between its value and the , which is expressed by the Henderson–Hasselbalch equation. The titration curve for acetic acid shown in Figure 2-27 illustrates the effect of on the fraction of molecules in the un-ionized and ionized forms. When the is equal to the , half of the acetic acid is dissociated (dashed lines). At one unit below the of an acid, 91 percent of the molecules are in the form; at one unit above the , 91 percent are in the form. At values more than one unit above or below the (unshaded regions in Figure 2-27), the buffering capacity of weak acids and bases declines rapidly. In other words, the addition of the same number of moles of base — for example, hydroxyl ions added as sodium hydroxide — to a solution containing a mixture of and that is at a near the will cause less of a change than it would if the and were not present or if the were far from the value.
FIGURE 2-27 The titration curve of the buffer acetic acid . The for the dissociation of acetic acid to hydrogen and acetate ions is 4.75. At this , half the acetic acid molecules are dissociated. Because is measured on a logarithmic scale, the solution changes from 91 percent at to 9 percent at . The acid has maximum buffering capacity in this range. Description The vertical axis of the graph represents p H values ranging from 0 to 8, in increments of 1. The horizontal axis represents fraction of dissociated acetic acid and 'added O H anion'. It ranges from 0 to 1, in increments of 0.1. A sigmoidal curve is shown, starting at zero on the horizontal axis at p H 3 rising and passing through an inflection point, the half-dissociated point at p H 4.75, and then rising until full dissociation. The P K subscript a is 4.75. Acetic acid yields an acetate ion and a proton.
All biological systems contain one or more buffers. Phosphate ions, the ionized forms of phosphoric acid, are present in considerable quantities in cells and are an important buffer in maintaining, or buffering, the of the cytoplasm. Phosphoric acid has three protons that are capable of dissociating, but they do not dissociate simultaneously. Loss of each proton can be described by a discrete dissociation reaction and , as shown in Figure 2-28. When hydroxyl ions are added to a solution of phosphoric acid, the change is much less steep at values near the three values (shaded regions) than when the of the solution is not similar to any of the . The titration curve for phosphoric acid shows that the for the dissociation of the second proton is 7.2. Thus at , about 50 percent of cellular phosphate is and about 50 percent is according to the Henderson–Hasselbalch equation. For this reason, phosphate is an excellent buffer at values around 7.2, the approximate of the cytoplasm of cells, and at , the of human blood. The amino (lysine), guanidinium (arginine), and carboxylate (aspartate, glutamate) portions of amino acid side chains of proteins as well as the amino and carboxylate groups at the N- and C-termini of proteins can also bind and release protons. Thus proteins that are present in high concentrations inside cells and in many extracellular fluids can themselves serve as buffers.
FIGURE 2-28 The titration curve of phosphoric acid , a common buffer in biological systems. This biologically ubiquitous molecule has three hydrogen atoms that dissociate at different values; thus phosphoric acid has three values, as noted on the graph. The shaded areas denote the ranges — within one unit of the three values — where the buffering capacity of phosphoric acid is high. In these regions, the addition of an acid (or base) will cause relatively small changes in the . Description The vertical axis of the graph represents p H, ranging from 0 to 14; in increments of 2. The horizontal axis represents added O H anion. The curve rises sharply before leveling out and passing through an inflection point at the first p K subscript a, the loss of the first proton from phosphoric acid, at pH 2.1. This pattern of rising sharply and passing through an inflection point continues twice more, at pH 7.2 and 12.7, corresponding to losses of the second and third protons from phosphoric acid. The pH
ranges where the pH buffering action is most pronounced lies within 1 pH unit either side of each p K A value. KEY CONCEPTS OF SECTION 2.3 Chemical Reactions and Chemical Equilibrium A chemical reaction is at equilibrium when the rate of the forward reaction is equal to the rate of the reverse reaction, and thus there is no net change over time in the concentration of the reactants or products. The equilibrium constant of a reaction reflects the ratio of products to reactants at equilibrium and thus is a measure of the extent of the reaction and the relative stabilities of the reactants and products. The depends on the temperature, pressure, and chemical properties of the reactants and products but is independent of the reaction rate and of the initial concentrations of reactants and products. For any reaction, the equilibrium constant equals the ratio of the forward rate constant to the reverse rate constant . The rates of conversion of reactants to products and vice versa depend on the rate constants and the concentrations of the reactants or products. Within cells, the linked reactions in metabolic pathways generally are not at equilibrium, but rather at steady state, at which the rate of formation of the intermediates equals their rate of consumption (see Figure 2-23), and thus the concentrations of the intermediates are not changing. The dissociation constant for the noncovalent binding of two molecules is a measure of the tightness of binding or the stability of the complex formed between the molecules (e.g., ligand-receptor or protein-DNA complexes). The lower the , the higher the binding affinity, and the tighter the binding. values of (nanomolar) are considered to be tight, (micromolar) modestly tight, and (millimolar) relatively weak. The is the negative logarithm of the concentration of hydrogen ions . The of the cytoplasm is normally about 7.2–7.4, whereas the interior of lysosomes has a of about 4.5. Acids release protons and bases bind them. Buffers are mixtures of a weak acid and its corresponding base form , which minimize the change in of a solution when an acid or base is added.
Biological systems use various buffers to maintain their within a very narrow range to permit the optimal functioning of biomolecules.
Several Forms of Energy Are Important in Biological Systems
2.4 Biochemical Energetics Obtaining, storing, and using energy are central to cellular life. Energy may be defined as the ability to do work, a concept that is as applicable to cells as to automobile engines and electric power plants. The energy stored within chemical bonds can be harnessed to support chemical work and the physical movements of cells. In this section, we review how energy influences the extents of chemical reactions (chemical thermodynamics) and the rates of chemical reactions (chemical kinetics). Several Forms of Energy Are Important in Biological Systems There are two principal forms of energy: kinetic and potential. Kinetic energy is the energy of movement — the motion of molecules, for example. Potential energy is stored energy — the energy stored in covalent bonds, for example. Potential energy plays a particularly important role in the energy economy of cells. Thermal energy, or heat, is a form of kinetic energy — the energy of the motion of molecules. For heat to do work, it must flow from a region of higher temperature, where the average speed of molecular motion is greater, to one of lower temperature. Although differences in temperature can exist between the internal and external environments of cells, these
thermal gradients do not usually serve as the source of energy for cellular activities. The thermal energy in warm-blooded animals, which have evolved a mechanism for thermoregulation, is used chiefly to maintain constant body temperature. This is an important homeostatic function because the rates of many cellular activities are temperature dependent. For example, cooling mammalian cells from their normal body temperature of to can almost completely freeze or stop many cellular processes (e.g., intracellular membrane movements). Overheating can be lethal because heat disrupts the noncovalent interactions that are required to maintain the structures and functions of biological macromolecules and macromolecular complexes. Radiant energy is the kinetic energy of photons, or waves of light. This form of kinetic energy is critical to biology. Radiant energy can be converted to thermal energy, for instance, when light is absorbed by molecules and the energy is converted to molecular motion. Radiant energy absorbed by molecules can also change the electronic structure of the molecules by moving electrons into higher energy orbitals, from whence the energy can later be recovered to perform work. For example, during photosynthesis, light energy absorbed by pigment molecules such as chlorophyll is subsequently converted into the energy of chemical bonds (see Chapter 12). Mechanical energy, a major form of kinetic energy in biology, usually results from the conversion of stored chemical energy. For example, changes in the lengths of cytoskeletal filaments powered by chemical
energy generate mechanical forces that push or pull on membranes and organelles (see Chapters 17 and 18). Electric energy — the energy of moving electrons or other charged particles — is yet another major form of kinetic energy. Electric energy has particular importance to membrane function, as in electrically active neurons (see Chapters 11, 12, and 23). Several forms of potential energy are biologically significant. Central to biology is chemical potential energy, the energy stored in the bonds connecting atoms in molecules. Indeed, most of the biochemical reactions described in this book involve the making or breaking of at least one covalent chemical bond. In general, energy must be expended to make covalent bonds in typical biomolecules, and energy is released when those bonds are broken. For example, the high potential energy in the covalent bonds of glucose can be released by controlled enzymatic combustion in cells (see Chapter 12). This energy is harnessed by the cell to do many kinds of work. A second biologically important form of potential energy is the energy in a concentration gradient. When the concentration of a substance on one side of a barrier, such as a membrane, is different from that on the other side, a concentration gradient exists. All cells form concentration gradients between their interior and the external fluids by selectively exchanging nutrients, waste products, and ions with their surroundings. Furthermore, the fluids within organelles in cells (e.g., mitochondria, lysosomes) frequently contain different concentrations of ions and other
Cells Can Transform One Type of Energy into Another
molecules than those in the cytosol; the concentration of protons within a lysosome as we saw in the last section, is about 500 times that in the cytosol . Concentration gradients of protons across membranes are an important driver of energy production in mitochondria (see Chapter 12). A third form of potential energy in cells is an electric potential — the energy of charge separation. For instance, there is a gradient of electric charge of about 200,000 volts per centimeter across the plasma membranes of virtually all cells. In Chapter 11, we discuss how concentration gradients and electric potential gradients are generated and maintained. In Chapter 12, we cover how they are converted to chemical potential energy. Cells Can Transform One Type of Energy into Another According to the first law of thermodynamics, energy is neither created nor destroyed, but can be converted from one form to another. (In nuclear reactions, mass is converted to energy, but this is irrelevant in biological systems.) Energy conversions are very important in biology. In photosynthesis, for example, the radiant energy of light is transformed into the chemical potential energy of the covalent bonds between the atoms in a sucrose or starch molecule. In muscles and nerves, chemical potential energy stored in covalent bonds is transformed, respectively, into the kinetic energy of muscle contraction and the electric energy of neural
The Change in Free Energy Determines If a Chemical Reaction Will Occur Spontaneously
transmission. In all cells, potential energy — released by breaking certain chemical bonds — is used to generate potential energy in the form of concentration and electric potential gradients. Similarly, energy stored in chemical concentration gradients or electric potential gradients is used to synthesize chemical bonds or to transport molecules from one side of a membrane to another to generate a concentration gradient. The latter process occurs during the transport of nutrients such as glucose into certain cells and the transport of many waste products out of cells. Because all forms of energy are interconvertible, they can be expressed in the same units of measurement. Although the standard unit of energy is the joule, biochemists have traditionally used an alternative unit, the calorie . A calorie is the amount of energy required to raise the temperature of one gram of water by . Throughout this book, we use the kilocalorie to measure energy changes . When you read or hear about the “Calories” in food (note the capital C), the reference is almost always to kilocalories as defined here. The Change in Free Energy Determines If a Chemical Reaction Will Occur Spontaneously Chemical reactions can be divided into two types, depending on whether energy is absorbed or released in the process. In an exergonic (“energyreleasing”) reaction, the products contain less energy than the reactants.
Exergonic reactions take place spontaneously. The liberated energy is usually released as heat (the energy of molecular motion) and generally results in a rise in temperature, as in the oxidation (burning) of wood. In an endergonic (“energy-absorbing”) reaction, the products contain more energy than the reactants, and energy is absorbed from the surroundings during the reaction. If there is no external source of energy to drive an endergonic reaction, it cannot take place. Endergonic reactions are responsible for the ability of the instant cold packs often used to treat injuries to rapidly cool below room temperature. Crushing the pack mixes the reactants, initiating the reaction. A fundamentally important concept in understanding if a reaction is exergonic or endergonic, and therefore if it occurs spontaneously or not, is free energy (G), or Gibbs free energy, named after J. W. Gibbs. Gibbs, who received the first PhD in engineering in America in 1863, showed that “all systems change in such a way that free energy [G] is minimized.” In other words, a chemical reaction occurs spontaneously when the free energy of the products is lower than the free energy of the reactants. In the case of a chemical reaction, , the free-energy change, , is given by The relation of to the direction of any chemical reaction can be summarized in three statements: If is negative, the forward reaction (generation of products) will tend to occur spontaneously, and energy usually will be released as
the reaction takes place (exergonic reaction) (Figure 2-29a). A reaction with a negative is referred to as thermodynamically favorable. If is positive, the forward reaction will not occur spontaneously; energy will have to be added to the system in order to force the reactants to become products (endergonic reaction, Figure 2-29b). If is zero, both forward and reverse reactions will occur without a gain or loss of energy, and there will be no spontaneous net conversion of reactants to products, or vice versa; the system is at equilibrium.
FIGURE 2-29 Changes in the free energy of exergonic and endergonic reactions. (a) In exergonic reactions, the free energy of the products is less than that of the reactants. Consequently, these reactions occur spontaneously, and energy is released as the
reactions proceed. (b) In endergonic reactions, the free energy of the products is greater than that of the reactants, and these reactions do not occur spontaneously. An external source of energy must be supplied if the reactants are to be converted into products. Description In graph A, the vertical axis represents free energy, G and the horizontal axis represents progress of reaction. The reactants are at higher free energy than the products. The difference is labeled as the free energy difference, delta G, and it is less than zero. In graph B, the vertical axis represents free energy, G and the horizontal axis represents progress of reaction. The reactants are at lower free energy than the products. The difference is labeled as the free energy difference, delta G, and it is greater than zero. By convention, the standard free-energy change of a reaction is the value of the change in free energy at 1 atm pressure, (as in pure water), and initial concentrations of 1 M for all reactants and products except protons, which are kept at . The conditions of most biological reactions differ from these standard conditions, particularly in the concentrations of reactants, which are normally less than , and in the temperature of mammals . The free energy of a chemical system can be defined as , where H is the bond energy, or enthalpy (H), of the system; T is its temperature in degrees Kelvin (K); and S is the entropy (S), a measure of its randomness or disorder. According to the second law of thermodynamics, the natural tendency of any isolated system is to become more disordered — that is, for entropy to increase. For example, a drop of ink placed in a large container of water will spontaneously spread out (become less ordered) rather than remain in a concentrated drop in the
container. A reaction can occur spontaneously only if the combined effects of changes in enthalpy and entropy lead to a lower . That is, if temperature remains constant, a reaction proceeds spontaneously only if the free-energy change, , in the following equation is negative: (2-6) In an exothermic (“heat-releasing”) chemical reaction, is negative. In an endothermic (“heat-absorbing”) reaction, is positive. The combined effects of the changes in the enthalpy and entropy determine if the for a reaction is positive or negative and thus if the reaction occurs spontaneously. An exothermic reaction in which entropy increases occurs spontaneously . An endothermic reaction will occur spontaneously if increases enough so that the term can overcome the positive . Many biological reactions lead to an increase in order and thus a decrease in entropy . An obvious example is the reaction that links amino acids to form a protein. A solution of protein molecules has a lower entropy than does a solution of the same amino acids unlinked because the free movement of any amino acid is more restricted (greater order) when it is bound into a long chain than when it is not. Thus when cells synthesize polymers such as proteins from their constituent monomers, the polymerizing reaction will be spontaneous only if the cells can efficiently transfer energy to both generate the bonds that hold the monomers together and overcome the loss in entropy that accompanies polymerization. Often cells accomplish this feat by coupling such
synthetic, entropy-lowering reactions with independent reactions that have a very highly negative , such as the hydrolysis of nucleoside triphosphates (discussed later in this section). In this way, cells can convert sources of energy in their environment into the highly organized structures and metabolic pathways that are essential for life. The actual change in free energy during a reaction is influenced by temperature, pressure, and the initial concentrations of reactants and products, so it usually differs from the standard free-energy change . Most biological reactions — like others that take place in aqueous solutions — are also affected by the of the solution. We can estimate free-energy changes for temperatures and initial concentrations that differ from the standard conditions by using the equation (2-7) where R is the gas constant of , T is the temperature (in degrees Kelvin), and Q is the initial ratio of products to reactants. For a reaction , in which two molecules combine to form a third, Q in Equation 2-7 equals . In this case, an increase in the initial concentration of either or will result in a larger negative value for and thus drive the reaction toward spontaneous formation of C. Regardless of the of a particular biochemical reaction, it will proceed spontaneously within cells only if is negative given the intracellular concentrations of reactants and products. For example, the
The ΔG°′ of a Reaction Can Be Calculated from Its Keq
conversion of glyceraldehyde 3-phosphate (G3P) to dihydroxyacetone phosphate (DHAP), two intermediates in the cellular breakdown of glucose (see Chapter 12), has a of . If the initial concentrations of G3P and DHAP are equal, then because ; in this situation, the reversible reaction will proceed spontaneously in the direction of DHAP formation until equilibrium is reached. However, if the initial [DHAP] is and the initial [G3P] is , with other conditions standard, then Q in Equation 2-7 equals , giving a of . Under these conditions, the reaction will proceed in the direction of formation of G3P. The of a reaction is independent of the reaction rate. Indeed, under normal physiological conditions, few, if any, of the biochemical reactions needed to sustain life would occur quickly enough without some mechanism for increasing reaction rates. As we describe below and in more detail in Chapter 3, the rates of reactions in biological systems are usually determined by the activity of enzymes, the protein catalysts that accelerate the formation of products from reactants without altering the value of . The ΔG° of a Reaction Can Be Calculated from Its ′
The Rate of a Reaction Depends on the Activation Energy Necessary to Energize the Reactants into a Transition State
A chemical mixture at equilibrium is in a stable state of minimal free energy. For a system at equilibrium under standard conditions, we can write (2-8) (note the change to base 10 logarithms). Thus if we determine the concentrations of reactants and products at equilibrium (i.e., the ), we can calculate the value of . For example, the for the interconversion of glyceraldehyde 3-phosphate to dihydroxyacetone phosphate is 22.2 under standard conditions. Substituting this value into Equation 2-8, we can easily calculate the for this reaction as . By rearranging Equation 2-8 and taking the antilogarithm, we obtain (2-9) From this expression, it is clear that if is negative, the exponent will be positive, and hence will be greater than 1. Therefore, at equilibrium there will be more products than reactants; in other words, the formation of products from reactants is favored. Conversely, if is positive, the exponent will be negative, and will be less than 1. The relationship between and further emphasizes the influence of the relative free energies of reactants and products on the extent to which a reaction will occur spontaneously.
The Rate of a Reaction Depends on the Activation Energy Necessary to Energize the Reactants into a Transition State As a chemical reaction proceeds, reactants approach each other; some bonds begin to form while others begin to break. One way to think of the state of the molecules during this transition is that there are strains in the electronic configurations of the atoms and their bonds. The collection of atoms moves from the relatively stable state of the reactants to a transient, intermediate, and higher energy state during the course of the reaction (Figure 2-30). The state during a chemical reaction at which the system is at its highest energy level is called the transition state, and the collection of reactants in that state is called the transition-state intermediate. The energy needed to excite the reactants to this higher energy state is called the activation energy of the reaction. The activation energy is the difference in free energy between the reactants and the transition-state intermediate. The activation energy is usually represented by , which is analogous to the representation of the change in Gibbs free energy already discussed. From the transition state, the collection of atoms can either release energy as the reaction products are formed (reaction proceeds to the right in Figure 2-30) or release energy as the atoms go backward (leftward in Figure 2-30) and re-form the original reactants.
FIGURE 2-30 Activation energy of uncatalyzed and catalyzed chemical reactions. This hypothetical reaction pathway (blue) depicts the changes in free energy, G, as a reaction proceeds. A reaction will take place spontaneously if the free energy (G) of the products is less than that of the reactants . However, all chemical reactions proceed through one (shown here) or more high-energy transition states, and the rate of a reaction is inversely proportional to the activation energy , which is the difference in free energy between the reactants and the transition state. In a catalyzed reaction (red), the free energies of the reactants and products are unchanged, but the free energy of the transition state is lowered, thus increasing the velocity of the reaction. Description The vertical axis of the graph represents free energy, G. The horizontal axis represents progress of reaction. No units are given. Two curves are shown. In both, the reactants and products of each have the same free energy, and the reactants are higher in energy than the products. Both curves rise in free energy, reaching a maximum at a transition state, and then falling to the energy of the products. The change in free energy is given by the difference between the energy of the reactants and the energy of the transition
state. The change in free energy of the catalyzed reaction is lower than that of the uncatalyzed reaction. The velocity (V) at which products are generated from reactants during the reaction under a given set of conditions (temperature, pressure, reactant concentrations) will depend on the concentration of material in the transition state. That concentration will depend on (1) the rate at which reactants are converted into the transition state, which depends on the activation energy, and (2) the characteristic rate constant (v) at which the material in the transition state is converted to products. The higher the activation energy, the lower the fraction of reactants that reach the transition state, and the slower the overall rate of the reaction. The relationship between the concentration of reactants, v, and V is From this equation, we can see that lowering the activation energy — that is, decreasing the difference in free energy between the reactants and the transition state — leads to an acceleration of the overall reaction rate V. A reduction in of leads to a tenfold increase in the rate of the reaction, whereas a reduction increases the rate a hundredfold. Thus relatively small changes in can lead to large changes in the overall rate of the reaction. Catalysts such as enzymes (discussed further in Chapter 3) accelerate reaction rates by lowering the relative energy of the transition state and thus the activation energy required to reach it (see Figure 2-30). The
Life Depends on the Coupling of Energetically Unfavorable Chemical Reactions with Energetically Favorable Ones
relative energies of reactants and products determine if a reaction is thermodynamically favorable (negative ), whereas the activation energy determines how rapidly products form — that is, the reaction kinetics. Thermodynamically favorable reactions will not occur at appreciable rates if the activation energies are too high. Life Depends on the Coupling of Energetically Unfavorable Chemical Reactions with Energetically Favorable Ones Many processes in cells are energetically unfavorable and will not proceed spontaneously. Examples include the synthesis of proteins from amino acids and the transport of a substance across the plasma membrane from a lower to a higher concentration. Cells can carry out an energy-requiring, or endergonic, reaction by coupling it to an energy-releasing, or exergonic, reaction if the sum of the two reactions has an overall net negative . Suppose, for example, that the reaction has a of and that the reaction has a of :
Hydrolysis of ATP Releases Substantial Free Energy and Drives Many Cellular Processes
In the absence of the second reaction, there would be much more A than B at equilibrium. However, because the conversion of X to is such a favorable reaction, it will pull the first process toward the formation of B and the consumption of A. Energetically unfavorable reactions in cells are often coupled to the energy-releasing hydrolysis of ATP. Hydrolysis of ATP Releases Substantial Free Energy and Drives Many Cellular Processes In almost all organisms, the nucleoside triphosphate adenosine triphosphate, or ATP (Figures 2-1d and 2-31), is the most important molecule for capturing, transiently storing, and subsequently transferring energy to perform work such as biosynthesis or mechanical motion. Commonly referred to as a cell’s energy currency, ATP is a type of usable potential energy that cells can spend in order to power their activities. The storied history of ATP begins with its discovery in 1929, apparently simultaneously by Kurt Lohmann, who was working with the famous biochemist Otto Meyerhof in Germany and who published first, and by the less well-known Cyrus Fiske and Yellapragada SubbaRow in the United States. [It is noteworthy that SubbaRow, who died in 1948 at the age of 53, also made key contributions to treatments for major diseases, including anemia (folic acid), cancer and rheumatoid arthritis (methotrexate), and bacterial infections (first tetracycline antibiotic).] Muscle contractions were shown to depend on ATP in the 1930s. The proposal that ATP is the main intermediary for the transfer of energy in cells is credited to Fritz
Lipmann around 1941. Many Nobel Prizes have been awarded for the study of ATP and its role in cellular-energy metabolism, and its importance in understanding molecular cell biology cannot be overstated.
FIGURE 2-31 Hydrolysis of adenosine triphosphate (ATP). The two phosphoanhydride bonds (red) in ATP (top), which link the three phosphate groups, each have a of
about for hydrolysis. Hydrolysis of the terminal phosphoanhydride bond by the addition of water results in the release of phosphate and generation of ADP. Hydrolysis of the phosphoanhydride bonds of ATP is exergonic (Figure 2-29a), especially the terminal one, and the hydrolysis of these bonds is the source of energy that drives many energy-requiring reactions in biological systems. Description The illustration shows the chemical structure of adenosine triphosphate. Upon the addition of water, a phosphoanhydride bond is broken, yielding inorganic phosphate and adenosine diphosphate. Adenosine triphosphate (A T P): A T P has a Haworth projection of ribose which has a pyranose ring where C 1 is bonded to N 9 of the adenine group at the top and a hydrogen atom at the bottom. C 2 and C 3 are each bonded to a hydrogen atom at the top and a hydroxyl group at the bottom. C 4 and is bonded to a methylene group on top and a hydrogen atom at the bottom. The methylene group is further bonded to the oxygen atom of a phosphate group further bonded to two more phosphate groups. Each phosphate group has a central phosphorus atom bonded to three oxygen atoms, two of which are anions; and double bonded to another oxygen atom. The phosphate groups have phosphoanhydride bonds. Adenine has a cyclohexane ring fused with a cyclopentane ring. C 1, C 3, C 7 and C 9 are each replaced by a nitrogen atom. C 2 and C 8 are each bonded to a hydrogen atom. C 6 is bonded to an amide group. Double bonds are present between C 2 and C 3, C 4 and C 5, and N 7 and C 8. Inorganic phosphate (P subscript i): Inorganic phosphate has a central phosphorus atom bonded to three oxygen atoms, two of which are anions; and double bonded to another oxygen atom. One of the oxygen atoms is bonded to a hydrogen atom. Adenosine diphosphate (ADP): A D P has a Haworth projection of ribose which has a pyranose ring where C 1 is bonded to N 9 of the adenine group at the top and a hydrogen atom at the bottom. C 2 and C 3 are each bonded to a hydrogen atom at the top and a hydroxyl group at the bottom. C 4 and is bonded to a methylene group on top and a hydrogen atom at the bottom. The methylene group is further bonded to the oxygen atom of a phosphate group further bonded to one more phosphate group. Each phosphate group has a central phosphorus atom bonded to three oxygen atoms, two of which are anions; and double bonded to another oxygen atom. The phosphate groups
have phosphoanhydride bonds. Adenine has a cyclohexane ring fused with a cyclopentane ring. C 1, C 3, C 7 and C 9 are each replaced by a nitrogen atom. C 2 and C 8 are each bonded to a hydrogen atom. C 6 is bonded to an amide group. Double bonds are present between C 2 and C 3, C 4 and C 5, and N 7 and C 8. The useful energy in an ATP molecule is contained in phosphoanhydride bonds. These are covalent bonds formed from the condensation of two molecules of phosphate by the loss of water: Description A phosphate group has a central phosphorus atom double bonded to an oxygen atom at the top, a hydroxyl group on the right, an oxygen anion at the bottom and another oxygen anion on the left. The other phosphate group has a central phosphorus atom double bonded to an oxygen atom at the top, a hydroxyl group on the left, an oxygen anion at the bottom and another oxygen anion on the right. The hydrogen atom of the hydroxyl group the first phosphate and the hydroxyl group of the second phosphate are highlighted. With the liberation of a water molecule, phosphoanhydride bond is formed. The structure formed has a central oxygen atom bonded to two phosphorus atoms, one on the left and one on the right. Each phosphorus atom is further bonded to two oxygen anions and double bonded to an oxygen atom.
As shown in Figure 2-31, an ATP molecule has two key phosphoanhydride (also called phosphodiester) bonds. Forming these bonds (represented here by the symbol ~) in ATP requires an input of energy. When these bonds are hydrolyzed, or broken by the addition of water, that energy is released. Hydrolysis of a phosphoanhydride bond in each of the following reactions has a highly negative of about : Description First reaction: A p tilde symbol p tilde symbol p plus H subscript 2 O (A T P) gives A p tilde symbol p plus p i (inorganic phosphate) plus H superscript plus (A D P). Second reaction: A p tilde symbol p tilde symbol p plus H subscript 2 O (A T P) gives A p plus p p i (inorganic pyrophosphate) plus H superscript plus (A M P). Third reaction: A p tilde symbol p plus H subscript 2 O (A D P) gives A p plus p i (inorganic phosphate) plus H superscript plus (A M P).
stands for inorganic phosphate and for inorganic pyrophosphate, two phosphate groups linked by a phosphoanhydride bond. As the top two reactions show, the removal of a phosphate group from ATP leaves adenosine diphosphate (ADP), and the removal of a pyrophosphate group from ATP leaves adenosine monophosphate (AMP). A phosphoanhydride bond or other high-energy bond (commonly denoted by ~) is not intrinsically different from other covalent bonds. High-energy bonds simply release substantial amounts of energy when hydrolyzed. For instance, the for hydrolysis of a phosphoanhydride bond in ATP is more than three times the for hydrolysis of the phosphoester bond (red) in glycerol 3-phosphate : Description Glycerol 3-phosphate has a methine group bonded to a hydroxymethyl on the right, a hydroxyl group at the top, a methylene group on the left further bonded to an oxygen atom of a phosphate group.
A principal reason for this difference is that ATP and its hydrolysis products, ADP and , are charged at neutral . During synthesis of ATP, a large amount of energy must be used to force the negative charges in ADP and together. Conversely, this energy is released when ATP is hydrolyzed to ADP and . In comparison, formation of the phosphoester bond between an uncharged hydroxyl in glycerol and requires less energy, and less energy is released when this bond is hydrolyzed. Cells have evolved mechanisms by which enzymes transfer the free energy released by hydrolysis of phosphoanhydride bonds to other molecules, thereby driving reactions that would otherwise be energetically unfavorable. For example, if the for the reaction is positive but less than the absolute value of the for hydrolysis of ATP, the reaction can be driven to the right by coupling it to hydrolysis of the terminal phosphoanhydride bond in ATP. In one common mechanism of such energy coupling, some of the energy stored in this phosphoanhydride bond is transferred to one of the reactants (here, B) by the breaking of the bond in ATP and the formation of a covalent bond between the released phosphate group and that reactant. The phosphorylated intermediate generated in this way can then react with reactant C to form product in a reaction that has an overall negative : The overall reaction
is energetically favorable . Similarly, hydrolysis of GTP to GDP can provide energy to perform work, including the synthesis of ATP (see
Chapter 12), but most often GTP hydrolysis is used to control cellular systems (e.g., protein synthesis, hormonal signaling) rather than as a source of energy (see Chapter 15). An alternative mechanism of energy coupling is to use the energy released by ATP hydrolysis to change the conformation of a molecule to an energyrich stressed state. In turn, the energy stored as conformational stress can be released as the molecule relaxes back into its unstressed conformation. If this relaxation process can be coupled to another reaction, the released energy can be harnessed to drive cellular processes. As with many biosynthetic reactions, transport of molecules into or out of the cell often has a positive and thus requires an input of energy to proceed. Such simple transport reactions do not directly involve the making or breaking of covalent bonds; thus their is 0. In the case of a substance moving into a cell, Equation 2-7 becomes (2-10) where is the initial concentration of the substance inside the cell and is its concentration outside the cell. We can see from Equation 2-10 that is positive for transport of a substance into a cell against its concentration gradient (when ); the energy to drive such uphill transport is often supplied by the hydrolysis of ATP. Conversely,
ATP Is Generated During Photosynthesis and Respiration
when a substance moves down its concentration gradient , is negative. Such downhill transport releases energy that can be coupled to an energy-requiring reaction, such as the movement of another substance uphill across a membrane or the synthesis of ATP itself (see Chapters 11 and 12). ATP Is Generated During Photosynthesis and Respiration ATP is continuously being hydrolyzed to provide energy for many cellular activities. Some estimates suggest that humans daily hydrolyze a mass of ATP equal to their entire body weight. Clearly to continue functioning, cells must constantly replenish their ATP supply. Therefore, cells must obtain energy from their environment that can be used to resynthesize the ATP. For nearly all cells, the ultimate source of energy used to make ATP is sunlight. Some organisms can use sunlight directly. Through the process of photosynthesis, plants, algae, and certain photosynthetic bacteria trap the energy of sunlight and use it to synthesize ATP from ADP and . Much of the ATP produced in photosynthesis is hydrolyzed to provide energy for the conversion of carbon dioxide to six-carbon sugars, a process called carbon fixation:
The sugars made during photosynthesis are a source of food, and thus energy, for the photosynthetic organisms making them and for the nonphotosynthetic organisms, such as animals, that consume the plants either directly or indirectly by eating other animals that have eaten the plants. In this way, sunlight is the direct or indirect source of energy for most organisms (see Chapter 12). In plants, animals, and nearly all other organisms, the free energy in sugars and other molecules derived from food is released in the processes of glycolysis and cellular respiration. During cellular respiration, energyrich molecules in food (e.g., glucose) are oxidized to carbon dioxide and water. The complete oxidation of glucose, has a of and is the reverse of photosynthetic carbon fixation. Cells employ an elaborate set of protein-mediated reactions to couple the oxidation of 1 molecule of glucose to the synthesis of as many as 30 molecules of ATP from 30 molecules of ADP. This oxygendependent (aerobic) degradation (catabolism) of glucose is the major pathway for generating ATP in all animal cells, all non-photosynthetic plant cells, and many bacterial cells. Catabolism of fatty acids can also be an important source of ATP. We discuss the mechanisms of photosynthesis and cellular respiration in Chapter 12. Although light energy captured in photosynthesis is the primary source of chemical energy for cells, it is not the only source. Certain microorganisms that live in or around deep-sea hydrothermal vents, where
NAD+ and FAD Couple Many Biological Oxidation and Reduction Reactions
adequate sunlight is unavailable, derive the energy for converting ADP and into ATP from the oxidation of reduced inorganic compounds. These reduced compounds originate deep in the earth and are released at the vents. and FAD Couple Many Biological Oxidation and Reduction Reactions In many chemical reactions, electrons are transferred from one atom or molecule to another; this transfer may or may not accompany the formation of new chemical bonds or the release of energy that can be coupled to other reactions. The loss of electrons from an atom or a molecule is called oxidation, and the gain of electrons by an atom or a molecule is called reduction. An example of oxidation is the removal of electrons from the sulfhydryl group–containing side chains of two cysteine amino acids to form a disulfide bond, described in Section 2.2. Electrons are neither created nor destroyed in a chemical reaction, so if one atom or molecule is oxidized, another must be reduced. For example, oxygen draws electrons from (ferrous) ions to form (ferric) ions, a reaction that occurs as part of the process by which carbohydrates are degraded in mitochondria. Each oxygen atom receives two electrons, one from each of two ions:
Thus is oxidized and is reduced. Such reactions in which one molecule is reduced and another is oxidized are often referred to as redox reactions. Oxygen is an electron acceptor in many redox reactions in cells under aerobic conditions. Many biologically important oxidation and reduction reactions involve the removal or addition of hydrogen atoms (protons plus electrons) rather than the transfer of isolated electrons on their own. The oxidation of succinate to fumarate, which occurs in mitochondria, is an example (Figure 2-32). Protons are soluble in aqueous solutions (as ), but electrons are not, so they must be transferred directly from one atom or molecule to another without a water-dissolved intermediate. In this type of oxidation reaction, electrons are often transferred to small, electron-carrying molecules, sometimes referred to as coenzymes. The most common of these electron carriers are (nicotinamide adenine dinucleotide), which is reduced to NADH, and FAD (flavin adenine dinucleotide), which is reduced to (Figure 2-33). The reduced forms of these coenzymes can transfer protons and electrons to other molecules, thereby reducing them.
FIGURE 2-32 Conversion of succinate to fumarate. In this oxidation reaction, which occurs in mitochondria as part of the citric acid cycle, succinate loses two electrons and two protons. These protons and electrons are transferred to FAD, reducing it to . Description Succinate has a four-carbon chain where C 1 and C 4 are each a carbonyl group bonded to an oxygen anion. C 2 and C 3 are each bonded to a hydrogen atom on the left and a hydrogen atom on the right. Fumarate has a four-carbon chain where C 1 and C 4 are each a carbonyl group bonded to an oxygen anion. C 3 and C 4 are each bonded to a hydrogen atom on the right.
FIGURE 2-33 The electron-carrying coenzymes and FAD. (a) (nicotinamide adenine dinucleotide) is reduced to NADH by the addition of two electrons and one proton simultaneously. In many biological redox reactions, a pair of hydrogen atoms (two protons and two electrons) is removed from a molecule. In some cases, one of the protons and both electrons are transferred to ; the other proton is released into solution. (b) FAD (flavin adenine dinucleotide) is reduced to by the addition of two electrons and two protons, as occurs when succinate is converted to fumarate (see
Figure 2-32). In this two-step reaction, addition of one electron together with one proton first generates a short-lived semiquinone intermediate (not shown), which then accepts a second electron and proton. Description In the illustration labeled A, N A D superscript plus is reduced to N A D H by the addition of a proton and two electrons. N A D has a nicotinamide group bonded to ribose, two phosphates and adenosine. Nicotinamide has a benzene ring where C 1 is replaced by a nitrogen cation, C 3 is bonded to a carbonyl group further bonded to an amide group, C 4 is bonded to a hydrogen atom. Double bonds are present between C 2 and C 3, C 4 and C 5, and C 1 and C 6. N A D H has a structure similar to that of N A D except the N 1 has a lone pair of electron and C 4 is bonded to two hydrogen atoms of which one is highlighted. Double bonds are now present between C 2 and C 3, and C 5 and C 6. In the illustration labeled B, F A D; with the addition of two protons and two electrons yield F A D H subscript 2.
F A D has a flavin group bonded to ribitol, two phosphates and adenosine. Flavin has three cyclohexane rings fused together. C 1, C 3, C 5, and C 10 are each replaced by a single nitrogen atom. N 3, C 6, and C 9 are each bonded to a single hydrogen atom. C 2 and C 4 are carbonyl groups. C 7 and C 8 are each bonded to a methyl group. Double bonds are present between N 1 and C 10, C 4 and N 5, C 6 and C 7, C 8 and C 9, and C 5 and C 10. F A D H subscript 2 has a structure similar to F A D except that N 1 and N 5 are each bonded to a single hydrogen atom. To describe redox reactions, such as the reaction of ferrous ion and oxygen it is easiest to divide them into two half-reactions: In this case, the reduced oxygen readily reacts with two protons to form one water molecule . The readiness with which an atom or a molecule gains an electron is its reduction potential (E). The tendency to lose electrons, the oxidation potential, has the same magnitude as the reduction potential for the reverse reaction, but has the opposite sign. Reduction potentials are measured in volts (V) from an arbitrary zero point set at the reduction potential of the following half-reaction under standard conditions ( 1 atm, and reactants at ):
The value of the reduction potential (E) for a molecule or an atom under standard conditions is its standard reduction potential, . A molecule or an ion with a positive has a higher affinity for electrons than the ion does under standard conditions. Conversely, a molecule or ion with a negative has a lower affinity for electrons than the ion does under standard conditions. Like the values of , standard reduction potentials may differ somewhat from those found under the conditions in a cell because the concentrations of reactants in a cell are not 1 M. In a redox reaction, electrons move spontaneously toward atoms or molecules having more positive reduction potentials. In other words, a molecule having a more negative reduction potential can transfer electrons spontaneously to, or reduce, a molecule with a more positive reduction potential. In this type of reaction, the change in electric potential is the sum of the reduction and oxidation potentials for the two halfreactions. The for a redox reaction is related to the change in free energy by the following expression: (2-11) where n is the number of electrons transferred. Note that a redox reaction with a positive value will have a negative and thus will tend to proceed spontaneously from left to right. KEY CONCEPTS OF SECTION 2.4 Biochemical Energetics
The change in free energy, , is the most useful measure for predicting the potential of chemical reactions to occur spontaneously in biological systems. Chemical reactions tend to proceed spontaneously in the direction for which is negative. The magnitude of is independent of the reaction rate. A reaction with a negative is referred to as thermodynamically favorable. The chemical free-energy change, , equals . Thus the value of can be calculated from the experimentally determined concentrations of reactants and products at equilibrium. The rate of a reaction depends on the activation energy needed to energize reactants to a transition state. Catalysts such as enzymes speed up reactions by lowering the activation energy of the transition state. A chemical reaction having a positive can proceed spontaneously if it is coupled with a reaction having a negative of larger magnitude. Many otherwise energetically unfavorable cellular processes are driven by the hydrolysis of phosphoanhydride bonds in ATP (see Figure 2-31). Directly or indirectly, light energy captured by photosynthesis in plants, algae, and photosynthetic bacteria is the ultimate source of chemical energy for nearly all cells on Earth. An oxidation reaction (loss of electrons) is always coupled with a reduction reaction (gain of electrons). Biological oxidation and reduction reactions are often coupled by electron-carrying coenzymes such as and FAD (see Figure 2-33). Oxidation-reduction reactions with a positive have a negative and thus tend to proceed spontaneously.
Key Terms
End of Chapter Visit Achieve to access study tools and to learn more about the content in this chapter: Analyze the Data Chapter References Additional study tools, including videos, animations, and quizzes Key Terms acid activation energy adenosine triphosphate (ATP) aerobic allostery alpha carbon atom amino acid amphipathic (or amphiphilic) anion base buffer carbohydrate catabolism catalyst
cation cellular respiration cellulose chemical equilibrium chemical potential energy concentration gradient covalent bond dipole dipole moment disaccharide dissociation constant (K ) DNA (deoxyribonucleic acid) electric potential endergonic endothermic enthalpy (H) entropy (S) enzyme equilibrium constant (K ) exergonic exothermic FAD (flavin adenine dinucleotide) fatty acids free-energy change (ΔG) glycogen glycolysis glycosaminoglycan glycosidic bond d eg
hexose hydrocarbon hydrogen bond hydrophilic hydrophobic hydrophobic effect ionic bond (ionic interaction) kinetic energy ligand macromolecule mitogen molecular complementarity monomer monosaccharide (nicotinamide adenine dinucleotide) noncovalent interactions nonpolar nucleic acid nucleoside nucleotide oxidation oxidation potential pentose peptide bond peptidoglycan pH phosphoanhydride bond phosphodiester bond
phosphoglyceride phospholipid photosynthesis polar polymer polysaccharide polyunsaturated potential energy protein purine pyrimidine rate constant receptor redox reaction reduction reduction potential (E) residue RNA (ribonucleic acid) saturated starch steady state stereoisomer transition state transition-state intermediate triacylglycerol (triglyceride) unsaturated van der Waals interaction
Review the Concepts
Review the Concepts 1. The gecko is a reptile with an amazing ability to climb smooth surfaces, including glass. Geckos appear to stick to smooth surfaces via van der Waals interactions between septa on their feet and the smooth surface. How is this method of stickiness advantageous over covalent interactions? Given that van der Waals forces are among the weakest molecular interactions, how can the gecko’s feet stick so effectively? 2. The channel is an example of a transmembrane protein (a protein that spans the phospholipid bilayer of the plasma membrane). What types of amino acids are likely to be found (a) lining the channel through which passes, (b) in contact with the hydrophobic core of the phospholipid bilayer containing fatty acyl groups, (c) in the cytosolic domain of the protein, and (d) in the extracellular domain of the protein? 3. V-M-Y-F-E-N: This is the single-letter amino acid abbreviation for a peptide. What is the net charge of this peptide at ? An enzyme called a protein tyrosine kinase can attach a phosphate to the hydroxyl group of a tyrosine (Y). What is the net charge of the peptide at after it has been phosphorylated by a tyrosine kinase? What is the likely source of phosphate used by the kinase for this reaction? 4. Disulfide bonds help to stabilize the three-dimensional structure of proteins. What amino acids are involved in the formation of disulfide bonds? Does the formation of a disulfide bond increase or decrease entropy ?
5. In the 1960s, the drug thalidomide was prescribed to pregnant women to treat morning sickness. However, thalidomide caused severe limb defects in the children of some women who took the drug, and its use for morning sickness was discontinued. It is now known that thalidomide was administered as a mixture of two stereoisomeric compounds, one of which relieved morning sickness and the other of which was responsible for the birth defects. What are stereoisomers? Why might two such closely related compounds have such different physiological effects? 6. Name the compound shown below. Description The structure has a Haworth projection which has a pyranose ring where C 1 is bonded to N 9 of the adenine group at the top and a hydrogen atom at the bottom. C 2 and C 3 are each bonded to a hydrogen atom at the top and a hydroxyl group at the bottom. C 4 and is bonded to C 5 of methylene group on top and a hydrogen atom at the bottom. The
methylene group is further bonded to the oxygen atom of a phosphate group further bonded to two more phosphate groups. Each phosphate group has a central phosphorus atom bonded to three oxygen atoms, two of which are anions; and double bonded to another oxygen atom. Adenine has a cyclohexane ring fused with a cyclopentane ring. C 1, C 3, C 7 and C 9 are each replaced by a nitrogen atom. C 2 is bonded to an amide group. C 8 is bonded to a hydrogen atom. C 6 is a carbonyl group. Double bonds are present between C 2 and C 3, C 4 and C 5, and N 7 and C 8. Is this nucleotide a component of DNA, RNA, or both? Name one other function of this compound. 7. The chemical basis of blood-group specificity resides in the carbohydrates displayed on the surfaces of red blood cells. Carbohydrates have the potential for great structural diversity. Indeed, the structural complexity of the oligosaccharides that can be formed from four sugars is greater than that of the oligopeptides that can be formed from four amino acids. What properties of carbohydrates make this great structural diversity possible? 8. Calculate the of of pure water at equilibrium. How will the change after of the strong base sodium hydroxide are dissolved in the water? Now, calculate the of a aqueous solution of the weak acid 3-(N-morpholino) propane-1-sulfonic acid (MOPS) in which 61 percent of the solute is in its weak acid form and 39 percent is in the form of MOPS’s corresponding base (the for MOPS is 7.20). What is the final after of are added to of this MOPS buffer? 9. Ammonia is a weak base that under acidic conditions becomes protonated to the ammonium ion in the following
reaction: freely permeates biological membranes, including those of lysosomes. The lysosome is a subcellular organelle with a of about 4.5–5.0; the of cytoplasm is about 7.0. What is the effect on the of the fluid content of lysosomes when cells are exposed to ammonia? Note: Ammonium does not diffuse freely across membranes. 10. Consider the binding reaction where L is a ligand and R is its receptor. When of L is added to a solution containing of R, 90 percent of the L binds to form LR. What is the of this reaction? How will the be affected by the addition of a protein that facilitates (catalyzes) this binding reaction? What is the dissociation equilibrium constant ? 11. What is the ionization state of phosphoric acid in the cytoplasm? Why is phosphoric acid such a physiologically important compound? 12. The for the reaction is . What is the at starting with each of X, Y, and XY? Suggest two ways one could make this reaction energetically favorable. 13. According to health experts, saturated fatty acids, which come from animal fats, are a major factor contributing to coronary heart disease. What distinguishes a saturated fatty acid from an unsaturated fatty acid, and to what does the term saturated refer? Recently, trans unsaturated fatty acids, or trans fats, which raise
total cholesterol levels in the body, have also been implicated in heart disease. How does the cis stereoisomer differ from the trans configuration, and what effect does the cis configuration have on the structure of the fatty acid chain? 14. Chemical modifications of amino acids contribute to the diversity and function of proteins. For instance, γ-carboxylation of specific amino acids is required to make some proteins biologically active. What particular amino acid undergoes this modification, and what is its biological relevance? Warfarin, a derivative of coumarin, which is present in many plants, inhibits γ-carboxylation of this amino acid and was used in the past as a rat poison. At present, it is also used clinically in humans. What patients might be prescribed warfarin and why?