Introduction
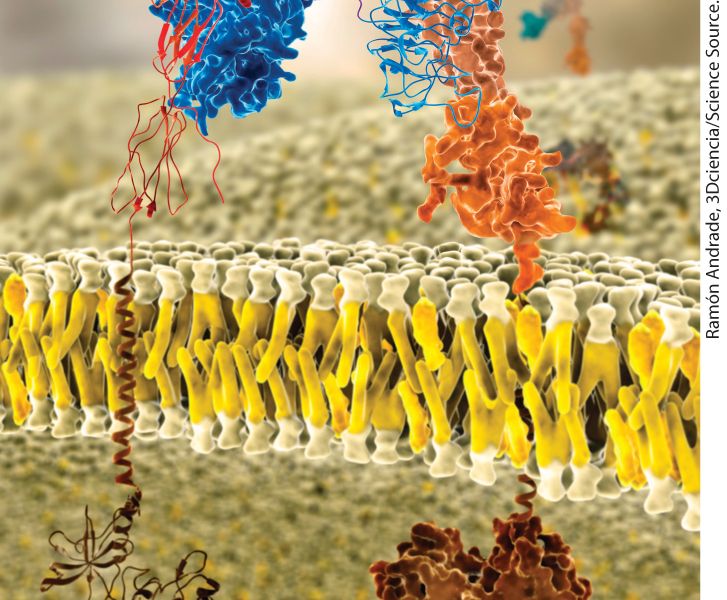
Chapter 10 Biomembrane Structure Molecular model of a lipid bilayer with embedded membrane proteins. Integral membrane proteins have distinct exoplasmic, cytosolic, and membrane-spanning domains. Shown here are portions of the insulin receptor, which regulates cell metabolism.
10.1 The Lipid Bilayer: Composition and Structural Organization
10.2 Membrane Proteins: Structure and Basic Functions
10.3 Phospholipids, Sphingolipids, and Cholesterol: Synthesis and Intracellular Movement Membranes participate in many aspects of cell structure and function. The plasma membrane defines the cell and separates the inside from the outside. In eukaryotes, membranes also define intracellular organelles such as the nucleus, mitochondrion, and lysosome. These biomembranes all have the same basic architecture: a phospholipid bilayer in which proteins are embedded (Figure 10-1). By preventing the unassisted movement of most water-soluble substances across the membrane, the phospholipid bilayer serves as a permeability barrier, helping to maintain the characteristic differences between the inside and the outside of the cell or organelle. In turn, the embedded proteins endow the membrane with specific functions, such as regulated transport of substances from one side to the other. Each type of cellular membrane has its own set of proteins that allow it to carry out many different functions.
FIGURE 10-1 Fluid mosaic model of biomembranes. A bilayer of phospholipids about 3– 4 nm thick provides the basic architecture of all cellular membranes; membrane proteins give each cellular membrane its unique set of functions. Individual phospholipids can move laterally and spin within the plane of the membrane, giving the membrane a fluidlike consistency similar to that of olive oil. Noncovalent interactions between phospholipids, and between phospholipids and proteins, lend strength and resilience to the membrane, while the hydrophobic core of the bilayer prevents the unassisted movement of watersoluble substances from one side to the other. Integral membrane proteins (transmembrane proteins) span the bilayer and often form dimers and higher order oligomers. Lipidanchored proteins are tethered to one leaflet by a covalently attached hydrocarbon chain. Peripheral membrane proteins associate with the membrane primarily by specific noncovalent interactions with integral membrane proteins or membrane lipids. See D. Engelman, 2005, Nature 438:578–580.] Description The exterior and cytosol are labeled. Embedded in the phospholipid plasma membrane are several proteins such as lipid anchored proteins, small green structures embedded in the exterior layer of the membrane, integral membrane proteins that run all the way through the membrane, and peripheral membrane proteins that can be positioned on the interior or exterior of the cell and interact with embedded proteins. The sandwich-like
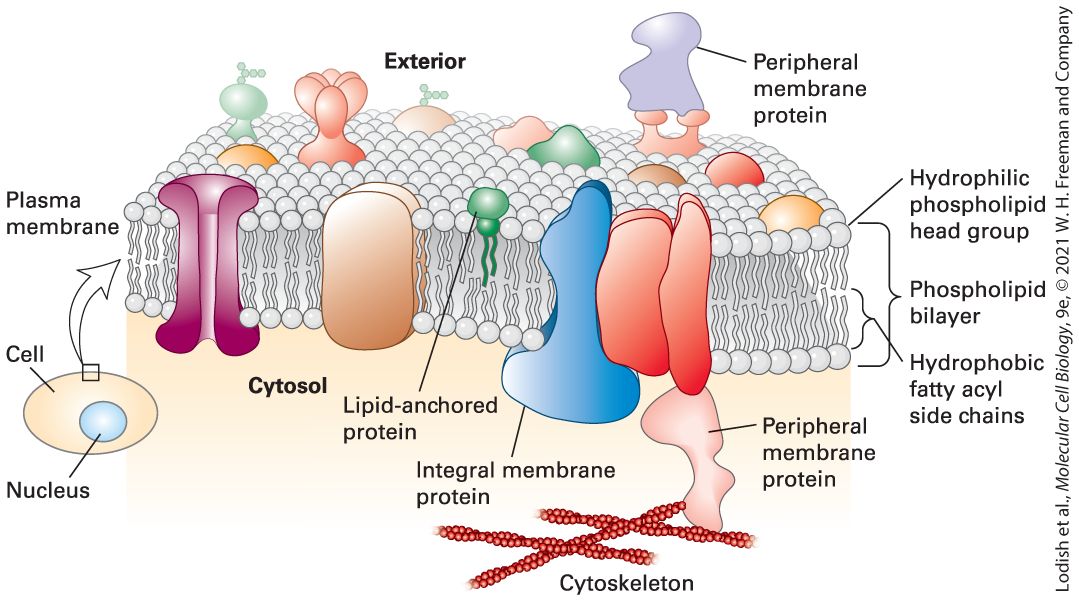
structure of the plasma membrane is highlighted, with the hydrophilic phospholipid head groups facing the exterior and the cytosol, and the hydrophobic fatty acyl side chains in the interior of the membrane. Prokaryotes, the simplest and smallest cells, are about in length and are usually surrounded by a single plasma membrane; in most cases, they contain no internal membrane-limited subcompartments (see Figure 1-12). However, this single plasma membrane contains hundreds of different types of proteins that are integral to the function of the cell. Some of these proteins catalyze ATP synthesis and initiation of DNA replication, for instance. Others include the many types of membrane transport proteins that enable specific ions, sugars, amino acids, and vitamins to cross the otherwise impermeable phospholipid bilayer to enter the cell and that allow specific metabolic products to exit. Receptor proteins in the plasma membrane are proteins that allow the cell to recognize chemical signals present in its environment and adjust its metabolism or pattern of gene expression in response. Peripheral membrane proteins, discussed in Section 10.2, associate with the membrane primarily by specific noncovalent interactions with integral membrane proteins or membrane lipids. Eukaryotes also have a plasma membrane studded with many different proteins that perform a variety of functions, including membrane transport, cell signaling, and connecting cells into tissues. In addition, eukaryotic cells — which are generally much larger than prokaryotes — have a variety of internal membrane-bounded organelles (see Figure 1-13). Each type of organelle membrane has a unique complement of proteins
that enable it to carry out its characteristic cellular functions, such as ATP generation (in mitochondria) and DNA synthesis (in the nucleus). Many plasma membrane proteins also bind components of the cytoskeleton, a dense network of protein filaments that crisscrosses the cytosol to provide mechanical support for cellular membranes. These interactions are essential for the cell to assume and maintain its specific shape and for many types of cell movements. Despite playing a structural role in cells, membranes are not rigid structures. They can bend and flex in three dimensions while still maintaining their integrity, in part because of the abundant noncovalent interactions that hold the lipids and proteins together. Moreover, there is considerable mobility of individual lipids and proteins within the plane of the membrane. According to the fluid mosaic model of biomembranes, first proposed by researchers in the 1970s, the phospholipid bilayer behaves in some respects like a two-dimensional fluid, with individual lipid molecules able to move past one another as well as spin in place. Such fluidity and flexibility not only allow organelles to assume their typical shapes but also confer on the membrane the dynamic property that enables membrane budding and fusion/fission, as occurs when viruses enter a cell or are released from an infected cell (Figure 10-2a). Internal cellular membranes of the Golgi complex bud into vesicles in the cytosol (Figure 10-2b) and then fuse with other membranes to transport their contents from one organelle to another (see Chapter 14).
FIGURE 10-2 Eukaryotic cellular membranes are dynamic structures. (a) An electron micrograph of the plasma membrane of an HIV-infected cell, showing HIV particles
budding into the culture medium. As the virus core buds from the cell, it becomes enveloped by a membrane, derived from the cell’s plasma membrane, that contains specific viral proteins. (b) Stacked membranes of the Golgi complex with budding vesicles. Note the irregular shape and curvature of these membranes. Description (a) The micrograph shows H I V particles as small rounded structures separating from the plasma membrane. (b) The micrograph shows a stack of membranes of the Golgi complex. We begin our examination of biomembranes by considering their lipid components. These molecules not only affect membrane shape and function but also help anchor proteins to the membrane, modify membrane protein activities, and transduce signals to the cytoplasm. We then consider the structure of membrane proteins. Many of these proteins have large segments that are embedded in the hydrocarbon core of the phospholipid bilayer, and we focus on the principal classes of such transmembrane proteins. Finally, we consider how lipids such as phospholipids and cholesterol are synthesized in cells and distributed to their many membranes and organelles.
Phospholipids Spontaneously Form Bilayers
10.1 The Lipid Bilayer: Composition and Structural Organization In Chapter 2, we learned that phospholipids are the principal building blocks of biomembranes. There are multiple types of phospholipids, but the most common phospholipids in membranes are the phosphoglycerides (see Figure 2-20). All phospholipids are amphipathic molecules, meaning that they consist of two segments with very different chemical properties: a fatty acid–based (fatty acyl) hydrocarbon “tail” that is hydrophobic (“water fearing”) and partitions away from water, and a polar “head group” that is strongly hydrophilic (“water loving”) and tends to interact with water molecules. The interactions of phospholipids with one another and with water largely determine the structure of biomembranes. Besides phospholipids, biomembranes contain smaller amounts of other amphipathic lipids, such as glycolipids and cholesterol, which contribute to membrane function in important ways. We first consider the structure and properties of pure phospholipid bilayers and then discuss the composition and behavior of natural cellular membranes. Then we consider how the precise lipid composition of a given membrane influences its physical properties.
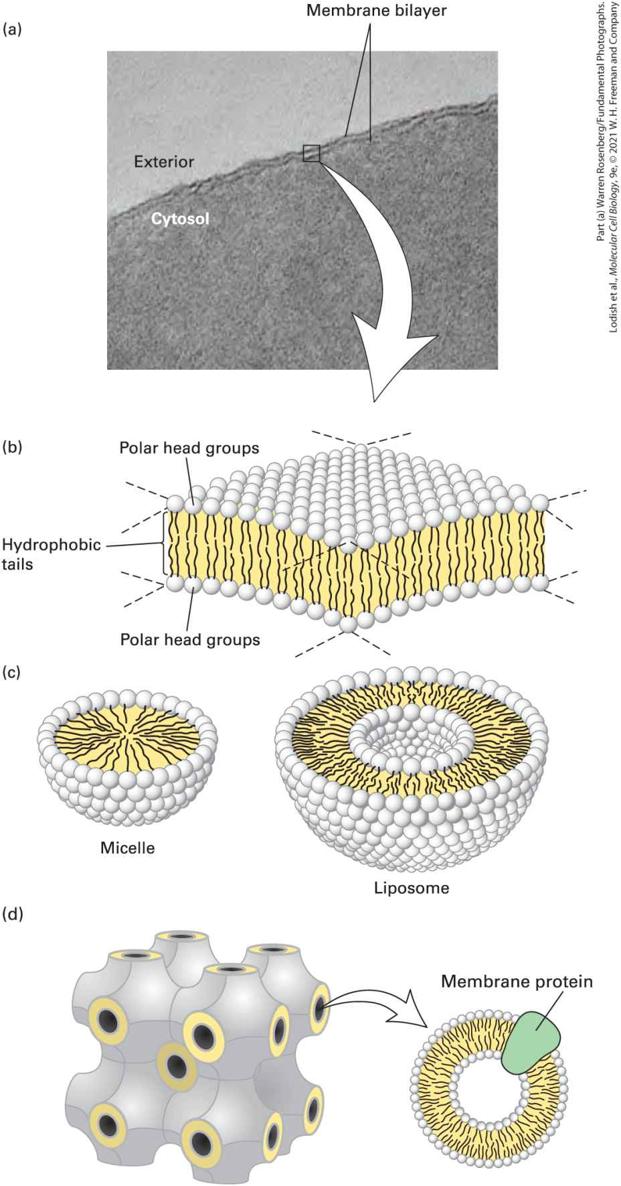
Phospholipids Spontaneously Form Bilayers The amphipathic nature of phospholipids, which governs their interactions, is critical to the structure of biomembranes. When a suspension of phospholipids is mechanically dispersed (e.g., by ultrasound, or “sonication”) in an aqueous solution, the phospholipids aggregate into one of three structures: spherical micelles or liposomes, or sheet-like phospholipid bilayers that are two molecules thick (Figure 103). The type of structure formed by pure phospholipids or a mixture of phospholipids in the absence of added proteins depends on several factors, including the length of the fatty acyl chains in the hydrophobic tails, their degree of saturation (i.e., the number of and bonds), and temperature. In all three structures, the hydrophobic effect (see Chapter 2) causes the fatty acyl chains to aggregate and exclude water molecules from the core of the structure. Micelles are rarely formed from natural phospholipids, whose fatty acyl chains are generally too bulky to fit into the interior of a micelle. Micelles are formed, however, if one of the two fatty acyl chains that make up the tail of a phospholipid is removed by hydrolysis to form a lysophospholipid, as occurs upon treatment with the enzyme phospholipase. In aqueous solutions, common detergents and soaps form micelles that behave like the balls in tiny ball bearings, thus giving soap solutions their slippery feel and lubricating properties.
FIGURE 10-3 The bilayer structure of biomembranes. (a) Electron micrograph of a thin section through an erythrocyte membrane stained with osmium tetroxide. The characteristic “railroad track” appearance of the membrane indicates the presence of two polar layers, consistent with the bilayer structure of phospholipid membranes. (b) Schematic interpretation of the phospholipid bilayer. Polar groups face outward to shield the hydrophobic fatty acyl tails from water. The hydrophobic effect and van der Waals interactions between the fatty acyl tails drive the assembly of the bilayer (see Chapter 2). (c) Cross-sectional views of two other structures formed by dispersal of phospholipids in water. A spherical micelle has a hydrophobic interior composed entirely of fatty acyl chains; a spherical liposome consists of a phospholipid bilayer surrounding an aqueous center. (d) Under certain circumstances, lipids can assume yet other forms of organization. Shown here is the cubic phase of lipids, a highly regular recurring structure that has helped the formation of crystals of membrane proteins that were otherwise difficult to crystallize. Description (a) The electron micrograph of a biomembrane with a white arrow pointing to the schematics below. (b) A schematic of the sandwich like membrane with the polar head groups labeled. The head groups point outward from the center of the membrane, and the hydrophilic tails point inward. (c) Schematics of a liposome and a micelle with the same type of layers, (d) a cubic formation of lipids, with the membrane protein labeled. Phospholipid mixtures of the composition present in cells spontaneously form a symmetric phospholipid bilayer. Each phospholipid layer in this lamellar structure is called a leaflet. The hydrophobic fatty acyl chains in each leaflet minimize their contact with water by aligning themselves tightly together in the center of the bilayer, forming a hydrophobic core that is about 3–4 nm thick (see Figure 10-3b). The close packing of these nonpolar tails is stabilized by van der Waals interactions between the hydrocarbon chains. Ionic and hydrogen bonds stabilize the interactions of the polar head groups with one another and with water. Electron
Phospholipid Bilayers Form a Sealed Compartment Surrounding an Internal Aqueous Space
microscopy of thin sections of cells stained with osmium tetroxide, which binds strongly to the polar head groups of phospholipids, shows the bilayer structure (see Figure 10-3a). A cross section of a single membrane stained with osmium tetroxide looks like a railroad track: two thin dark lines (the stained head group complexes) with a uniform light space of about 2 nm between them (the hydrophobic tails). A phospholipid bilayer can be of almost unlimited size — from micrometers (μm) to millimeters (mm) in length or width — and can contain tens of millions of phospholipid molecules. The phospholipid bilayer is the basic structural unit of nearly all biological membranes. Its hydrophobic core prevents most water-soluble substances from crossing from one side of the membrane to the other. Although biomembranes contain other molecules (e.g., cholesterol, glycolipids, proteins), it is the phospholipid bilayer that separates two aqueous solutions and acts as a permeability barrier. The lipid bilayer thus defines cellular compartments and separates the cell’s interior from the outside world. The three structures mentioned above are not the only forms that lipids can assume in an aqueous environment. Unusual configurations of lipids have been instrumental in enforcing order on otherwise difficult-tocrystallize membrane proteins, including G protein–coupled receptors, enabling crystallographic analysis of membrane proteins in a true lipid environment (Figure 10-3d).
Phospholipid Bilayers Form a Sealed Compartment Surrounding an Internal Aqueous Space Phospholipid bilayers can be generated in the laboratory by simple means, using either chemically pure phospholipids or lipid mixtures of the composition found in cellular membranes (Figure 10-4). Such synthetic bilayers possess three important properties. First, they are virtually impermeable to water-soluble (hydrophilic) solutes, which do not readily diffuse across the bilayer. These solutes include salts, sugars, and most other small hydrophilic molecules — including water itself. The second property of a bilayer is its stability. The hydrophobic and van der Waals interactions between the fatty acyl chains maintain the integrity of the interior of the bilayer structure. Even though the exterior aqueous environment may vary widely in ionic strength and pH, the bilayer has the strength to retain its characteristic architecture. Third, all synthetic phospholipid bilayers can spontaneously form sealed compartments in which the aqueous space on the inside is separated from the exterior environment. An edge of a phospholipid bilayer, as depicted in Figure 103b, with the hydrocarbon core of the bilayer exposed to an aqueous solution, would be unstable; the exposed fatty acyl chains would be in an energetically much more stable state if they were not adjacent to water molecules but rather surrounded by other fatty acyl chains. Thus, in an aqueous solution, sheets of phospholipid bilayers spontaneously seal their edges, forming a spherical bilayer that encloses an aqueous central
compartment. The liposome depicted in Figure 10-3c is an example of such a structure viewed in cross section. Phospholipid bilayers can even be formed and immobilized on a solid support such as a microscope slide, which allows the incorporation of membrane proteins whose behavior in the plane of the membrane can then be studied by various microscopical techniques.
EXPERIMENTAL FIGURE 10-4 Formation and study of pure phospholipid bilayers. Top: A preparation of biological membranes is treated with an organic solvent, such as a mixture of chloroform and methanol (3:1), which selectively solubilizes the phospholipids and cholesterol. Proteins and carbohydrates remain in an insoluble residue. The solvent is removed by evaporation. Bottom left: If the extracted materials are mechanically dispersed in water, they spontaneously form a liposome, shown in cross section, with an internal aqueous compartment. Bottom right: A planar bilayer, also shown in cross section, can form over a small hole in a partition separating two aqueous phases; such a system can be used to study the physical properties of bilayers, such as their permeability to solutes. Description The schematic shows the phospholipid bilayer with embedded proteins and oligosaccharides labeled. The following treatment is depicted: 1. The bilayer is treated with an organic solvent, and the proteins and oligosaccharides form insoluble residues that are removed, leaving the phospholipids in solution. 2. The solvent is evaporated. 3. There are now two options. The phospholipids can be dispersed in water, forming liposomes, or dissolve the phospholipids in solvent and apply them to a small whole in a plastic partition separating to aqueous portions. This physical chemical property of a phospholipid bilayer has important implications for cellular membranes: no membrane in a cell can have an edge with exposed hydrocarbon fatty acyl chains. All membranes form closed compartments, similar in basic architecture to liposomes. Because all cellular membranes enclose an entire cell or an internal compartment, they have an internal face (the surface oriented toward the interior of the compartment) and an external face (the surface presented to the environment). More commonly, we designate the two surfaces of a cellular
membrane as the cytosolic face and the exoplasmic face. This nomenclature is useful in highlighting the topological equivalence of the faces in different membranes, as diagrammed in Figures 10-5 and 10-6. For example, the exoplasmic face of the plasma membrane is directed away from the cytosol, toward the extracellular space or external environment, and defines the outer limit of the cell. The cytosolic face of the plasma membrane faces the cytosol.
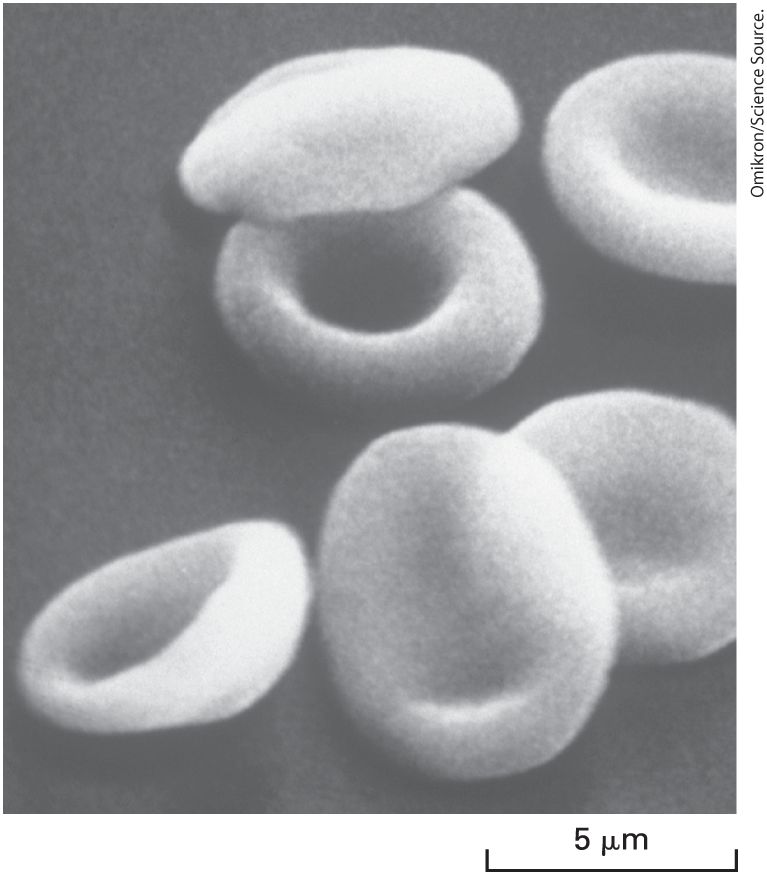
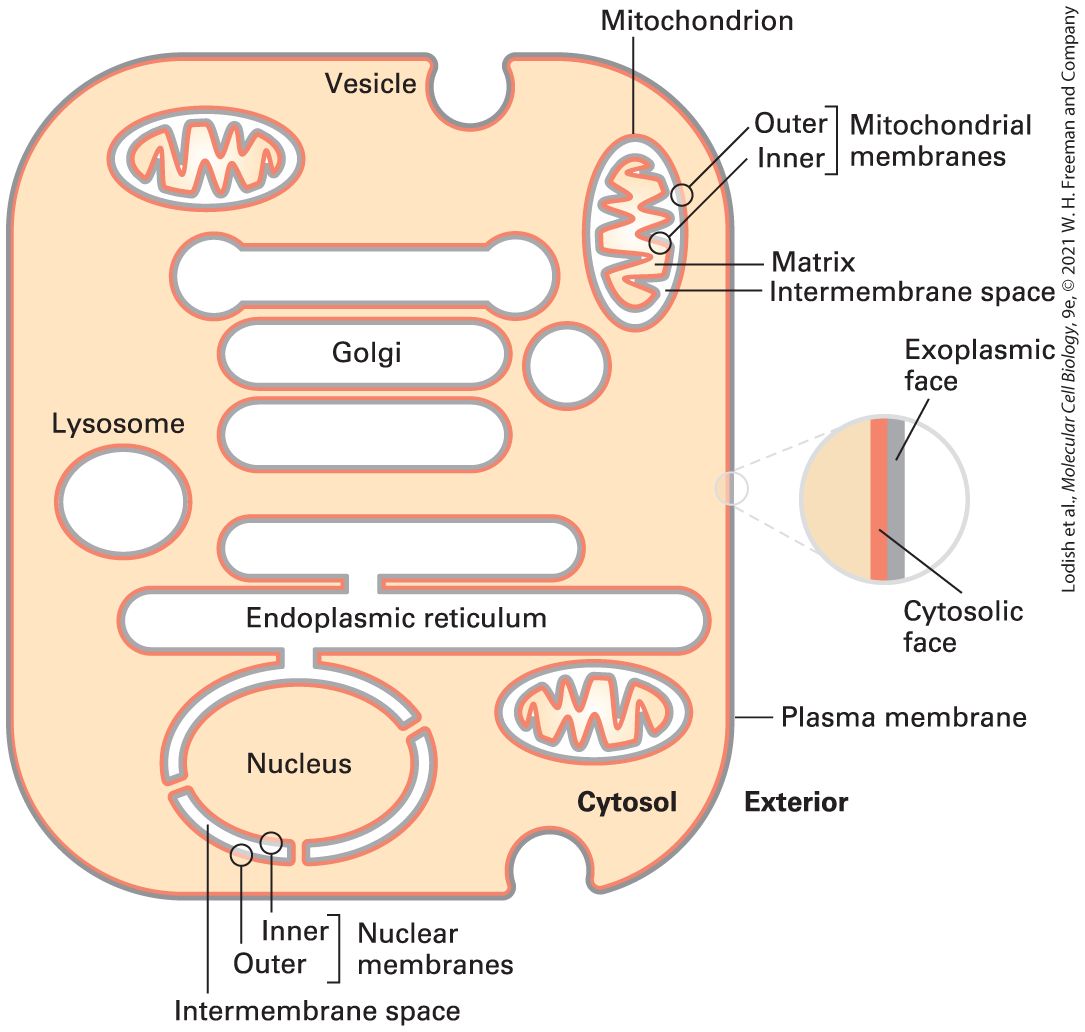
FIGURE 10-5 The faces of cellular membranes. The plasma membrane, a single bilayer, encloses the cell. In this highly schematic representation, internal cytosol (tan) and external environment (white) define the cytosolic (red) and exoplasmic (gray) faces of the bilayer. Vesicles and some organelles have a single membrane, and their internal aqueous space (white) is topologically equivalent to the outside of the cell. Three organelles — the nucleus, mitochondrion, and chloroplast (not shown) — are enclosed by two membranes separated by a small intermembrane space. The exoplasmic faces of the inner and outer membranes around these organelles border the intermembrane space between them. For simplicity, the hydrophobic membrane interior is not indicated in this diagram. Description Several structures are present in the cytosol of cells. These include vesicles, mitochondria, the nucleus, the endoplasmic reticulum, lysosomes, and the Golgi complex. Each has a red and a gray layer indicating the outer layer and inner layer of each membrane. An inset to the right shows labels for the red (cytosolic face) and the gray (exoplasmic face).
FIGURE 10-6 The faces of cellular membranes are conserved during membrane budding and fusion. Red membrane surfaces are cytosolic faces; gray membrane surfaces are exoplasmic faces. During endocytosis, a segment of the plasma membrane buds inward toward the cytosol and eventually pinches off a separate vesicle. During this process, the cytosolic face of the plasma membrane remains facing the cytosol, and the exoplasmic face of the new vesicle membrane faces the vesicle lumen. During exocytosis, an intracellular vesicle fuses with the plasma membrane, and the lumen of the vesicle (exoplasmic face) connects with the extracellular medium. Proteins that span the membrane retain their asymmetric orientation during vesicle budding and fusion; in particular, the same segment always faces the cytosol. Description The outer surface of the plasma membrane is labeled exoplasmic face and the inner surface is labeled cytosolic face. A vesicle is shown forming, and the schematic indicates how the exoplasmic face becomes the inner surface of the vesicle. Membrane proteins are embedded in the membrane, and a membrane protein with exoplasmic and cytosolic segments is illustrated being absorbed into a vesicle, the exoplasmic segment
is maintained on the exoplasmic face of the membrane at all times. The process of forming a vesicle is labeled endocytosis, whereas that of fusing a vesicle to the membrane is labeled exocytosis. Similarly for organelles and vesicles surrounded by a single membrane, the cytosolic face is directed toward the cytosol. The exoplasmic face is always directed away from the cytosol, and in this case it is on the inside of the organelle, in contact with the internal aqueous space, or lumen. The lumen of a vesicle is topologically equivalent to the extracellular space, a concept most easily understood for vesicles that arise by invagination (endocytosis) of the plasma membrane. The external face of the plasma membrane becomes the internal face of the vesicle membrane, while in the vesicle the cytosolic face of the plasma membrane still faces the cytosol (see Figure 10-6). Three organelles — the nucleus, mitochondrion, and chloroplast — are surrounded not by a single membrane, but by two. The exoplasmic surface of each membrane faces the space between the two membranes (the intermembrane space). This relationship can perhaps be best understood by reference to the endosymbiont hypothesis, discussed in Chapter 12, which posits that mitochondria and chloroplasts arose early in the evolution of eukaryotic cells by the engulfment of bacteria capable of oxidative phosphorylation or photosynthesis, respectively (see Figure 127). Gram-negative bacteria such as Escherichia coli are likewise surrounded by two membranes (see Figure 1-12).
Natural membranes from different cell types exhibit a variety of shapes, which complement a cell’s function. The smooth, flexible surface of the erythrocyte plasma membrane allows the cell to squeeze through narrow blood capillaries (Figure 10-7). Some cells have a long, slender extension of the plasma membrane, called a cilium or flagellum, which beats in a whiplike manner (see Figure 1-15). This motion causes fluid to flow across the surface of a sheet of cells or a sperm cell to swim toward an egg. The differing shapes and properties of biomembranes raise a key question in cell biology, namely, how the composition of biological membranes is regulated to establish and maintain the identity of the different membrane structures and membrane-limited compartments. We return to this question in Section 10.3 and in Chapter 14.
Biomembranes Contain Three Principal Classes of Lipids
FIGURE 10-7 Variation in biomembranes in different cell types. A smooth, flexible membrane covers the surface of the discoid erythrocyte cell, as seen in this scanning electron micrograph.
Biomembranes Contain Three Principal Classes of Lipids The term phospholipid refers to any amphipathic lipid with a phosphatebased head group and a two-chain hydrophobic tail. A typical biomembrane is not composed of phospholipids alone, but actually contains three classes of amphipathic lipids: phosphoglycerides, sphingolipids, and sterols, which differ in their chemical structures, abundance, and functions in the membrane (Figure 10-8). While all phosphoglycerides are phospholipids, only certain sphingolipids are, and no sterols are.
FIGURE 10-8 Three classes of membrane lipids. (a) Most phosphoglycerides are derivatives of glycerol 3-phosphate (red), which contains two esterified fatty acyl chains that constitute the hydrophobic “tail” and a polar “head group” esterified to the phosphate. The fatty acids can vary in length and be saturated (no double bonds) or unsaturated (one, two, or three double bonds). In phosphatidylcholine (PC), the head group is choline. Also
shown are the molecules attached to the phosphate group in three other common phosphoglycerides: phosphatidylethanolamine (PE), phosphatidylserine (PS), and phosphatidylinositol (PI). Plasmalogens contain one fatty acyl chain attached to glycerol by an ester linkage and one attached by an ether linkage; they contain the same head groups as other phosphoglycerides. (b) Sphingolipids are derivatives of sphingosine (red), an amino alcohol with a long hydrocarbon chain. Various fatty acyl chains are connected to sphingosine by an amide bond. The sphingomyelins (SM), which contain a phosphocholine head group, are phospholipids. Other sphingolipids are glycolipids in which a single sugar residue or branched oligosaccharide is attached to the sphingosine backbone. For instance, the simple glycolipid glucosylcerebroside (GlcCer) has a glucose head group. (c) The major sterols in animals, fungi, and plants differ slightly in structure, but all serve as key components of cellular membranes. The basic structure of sterols is a four-ring hydrocarbon (yellow). Like other membrane lipids, sterols are amphipathic. The single hydroxyl group is equivalent to the polar head group in other lipids; the conjugated ring and short hydrocarbon chain form the hydrophobic tail. See H. Sprong, P. van der Sluijs, and G. van Meer, 2001, Nature Rev. Mol. Cell Biol. 2:504–513. Description (a) Phosphoglyceride - An illustration shows a rounded head with two tails. The chemical structure below shows a variable portion of the head group and the hydrophobic tail. Another chemical structure below labeled plasmalogen shows a hydrophobic tail with a variable head group (b) sphingolipids - the chemical structure highlights the tail formation flowing into the chemical diagram for the head, with only two variations. (c) Three chemical diagrams for three sterols: cholesterol (animal) at left, ergosterol (fungi) in center and stigmasterol (plant) at right. All of these share the same central ring configuration which are highlighted in yellow. Phosphoglycerides, the most abundant class of phospholipids in most membranes are derivatives of glycerol 3-phosphate (see Figure 10-8a). A typical phosphoglyceride molecule consists of a hydrophobic tail composed of two fatty acyl chains esterified to the two hydroxyl groups in glycerol phosphate and a polar head group attached to the phosphate
group. The structure comprising the glycerol moiety and the two fatty acyl chains is referred to as a diacylglycerol. The two fatty acyl chains may differ in the number of carbons that they contain (commonly 16 or 18) and their degree of saturation (0, 1, or 2 double bonds). A phosphoglyceride is classified according to the nature of its head group. In phosphatidylcholines, the most abundant phospholipids in the plasma membrane, the head group consists of choline, a positively charged alcohol, esterified to the negatively charged phosphate. In other phosphoglycerides, an OH-containing molecule such as ethanolamine, serine, or the sugar derivative inositol is linked to the phosphate group. In the case of the inositol head group, the hydroxyl groups may be further modified with phosphates, yielding a class of phospholipids called phosphoinositides. The phosphoinositides fulfill an important function in signal transduction, as discussed in Chapters 15 and 16. The negatively charged phosphate group and the positively charged groups or hydroxyl groups on the head group interact strongly with water. At neutral pH, some phosphoglycerides (e.g., phosphatidylcholine and phosphatidylethanolamine) carry no net electric charge, whereas others (e.g., phosphatidylinositol and phosphatidylserine) carry a single net negative charge. Nonetheless, the polar head groups in all these phospholipids can pack together into the characteristic bilayer structure. When phospholipases act on phosphoglycerides, they produce lysophospholipids, which lack one of the two acyl chains. Lysophospholipids are not only important signaling molecules, released from cells and recognized by specific receptors; their presence can also affect the physical properties of the membranes in which they reside.
The plasmalogens are a group of phosphoglycerides that contain one fatty acyl chain attached to carbon 2 of glycerol by an ester linkage and one long hydrocarbon chain attached to carbon 1 of glycerol by an ether ( ) rather than an ester linkage. Plasmalogens are particularly abundant in human brain and heart tissue. The greater chemical stability of the ether linkage in plasmalogens compared with the ester linkage, and the subtle differences in three-dimensional structure between plasmalogens and other phosphoglycerides, may have as yet unrecognized physiological significance. A second class of membrane lipids is the sphingolipids. All these compounds are derived from sphingosine, an amino alcohol with a long hydrocarbon chain. Sphingolipids contain a long-chain fatty acid attached in amide linkage to the sphingosine amino group (see Figure 10-8b). Like phosphoglycerides, some sphingolipids have a phosphate-based polar head group. In sphingomyelin, the most abundant sphingolipid, phosphocholine is attached to the terminal hydroxyl group of sphingosine (see Figure 108b; SM). Thus sphingomyelin is a phospholipid, and its overall structure is quite similar to that of phosphatidylcholine. Sphingomyelins are similar in shape to phosphoglycerides and can form mixed bilayers with them. Other sphingolipids are amphipathic glycolipids whose polar head groups are sugars that are not linked to the tails via a phosphate group (so they are not phospholipids). Glucosylcerebroside, the simplest glycosphingolipid, contains a single glucose unit attached to sphingosine. In the complex glycosphingolipids called gangliosides, one or two branched sugar chains (oligosaccharides) containing sialic acid groups are attached to
sphingosine. Glycolipids constitute 2–10 percent of the total lipid content of plasma membranes; they are most abundant in nervous tissue. Cholesterol and its analogs constitute the third important class of membrane lipids, the sterols. The basic structure of sterols is a four-ring isoprenoid-based hydrocarbon. The structures of the principal yeast sterol (ergosterol) and plant phytosterols (e.g., stigmasterol) differ slightly from that of cholesterol, the major animal sterol (see Figure 10-8c). The small differences in the biosynthetic pathways and structures of fungal and animal sterols are the basis of most antifungal drugs currently in use. Cholesterol, like the two other sterols, has a hydroxyl substituent on one ring. Although cholesterol is almost entirely hydrocarbon in composition, it is amphipathic because its hydroxyl group can interact with water. Because it lacks a phosphate-based head group, it is not a phospholipid. Cholesterol is especially abundant in the plasma membranes of mammalian cells but is absent from most prokaryotic and all plant cells. As much as 30–50 percent of the lipids in plant plasma membranes consist of certain steroids unique to plants. Between 50 and 90 percent of the cholesterol in most mammalian cells is present in the plasma membrane and associated vesicles. Cholesterol and other sterols are too hydrophobic to form a bilayer structure on their own. Instead, at the concentrations found in natural membranes, these sterols must intercalate between phospholipid molecules to be incorporated into biomembranes. When so intercalated, sterols provide structural support to membranes, preventing too close a packing of the phospholipids’ acyl chains to maintain a significant measure of membrane fluidity and at the same time conferring
Most Lipids and Many Proteins Are Laterally Mobile in Biomembranes
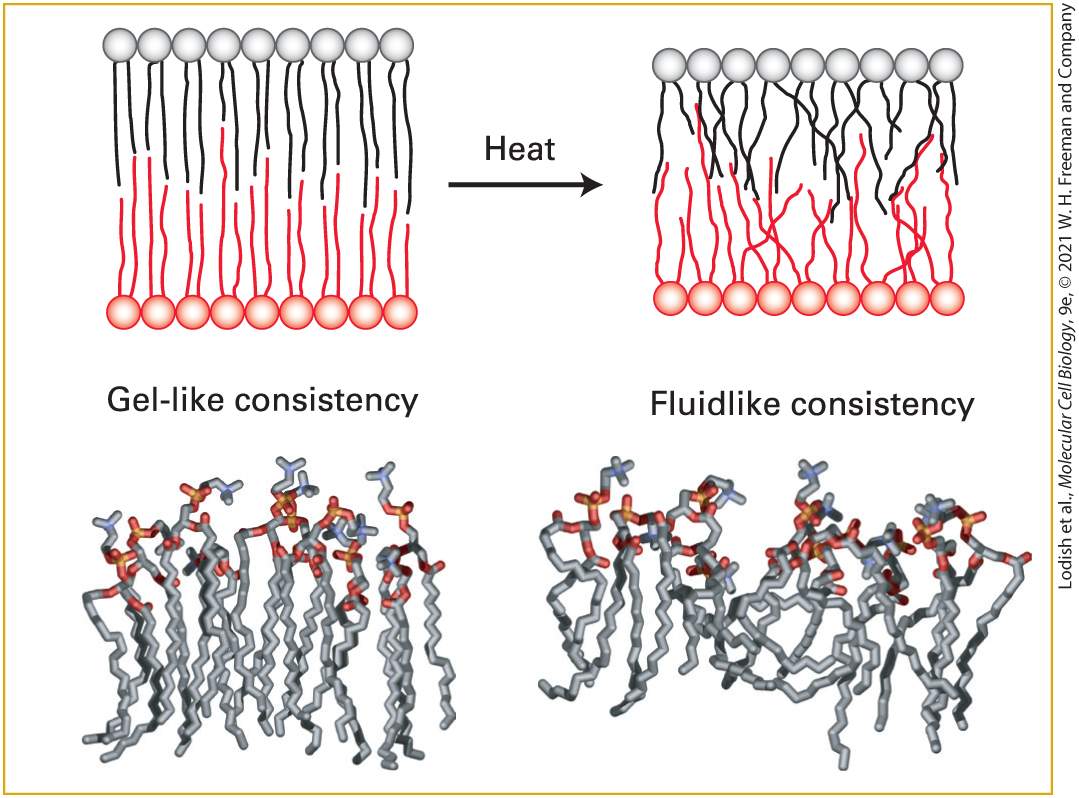
the necessary rigidity required for mechanical support. Some of these effects can be highly local, as in the case of lipid rafts, discussed below. In addition to its structural role in membranes, cholesterol is the precursor for several important bioactive molecules. They include bile acids, which are made in the liver and help emulsify dietary fats for digestion and absorption in the intestines; steroid hormones produced by endocrine cells (e.g., adrenal gland, ovary, testes); and vitamin D produced in the skin and kidneys. Another critical function of cholesterol is its covalent addition to Hedgehog protein, a key signaling molecule in embryonic development (see Chapter 16). Most Lipids and Many Proteins Are Laterally Mobile in Biomembranes In the two-dimensional plane of a bilayer, thermal motion permits lipid molecules to rotate freely around their long axes and to diffuse laterally within each leaflet. Because such movements are lateral or rotational, the fatty acyl chains remain in the hydrophobic interior of the bilayer. In both natural and artificial membranes, a typical lipid molecule exchanges places with its neighbors in a leaflet about times per second and diffuses several micrometers per second at 37 °C. These diffusion rates indicate that the bilayer is 100 times more viscous than water — about the same as the viscosity of olive oil. Even though lipids diffuse more slowly in the bilayer than in an aqueous solvent, a membrane lipid could diffuse the length of a typical bacterial cell (1 μm) in only 1 second and the length
of an animal cell in about 20 seconds. When artificial pure phospholipid membranes are cooled below 37 °C, the lipids can undergo a phase transition from a liquid-like (fluid) state to a gel-like (semisolid) state, analogous to the liquid-solid transition when molten butter solidifies upon cooling or liquid water freezes (Figure 10-9). Below the phase-transition temperature, the rate of diffusion of the lipids drops precipitously. At usual physiological temperatures, the hydrophobic interior of natural membranes generally has a low viscosity and a fluidlike consistency, in contrast to the gel-like consistency observed at lower temperatures.
FIGURE 10-9 Gel-like and fluidlike forms of the phospholipid bilayer. Top: Depiction of gel-to-fluid transition. Phospholipids with long saturated fatty acyl chains tend to assemble into a highly ordered, gel-like bilayer in which there is little overlap of the nonpolar tails in
the two leaflets. Heat disorders the nonpolar tails and induces a transition from a gel to a fluid within a temperature range of only a few degrees. As the chains become disordered, the bilayer also decreases in thickness. Bottom: Molecular models of phospholipid monolayers in gel-like and fluidlike states, as determined by molecular dynamics calculations. [Data from H. Heller, M. Schaefer, and K. Schulten, 1993, J. Phys. Chem. 97:8343.] In pure lipid bilayers (i.e., in the absence of protein), phospholipids and sphingolipids rotate and move laterally, but they do not usually spontaneously migrate, or flip-flop, from one leaflet to the other. The energetic barrier is too high; migration would require moving the polar head group from its aqueous environment through the hydrocarbon core of the bilayer to the aqueous solution on the other side. Special membrane proteins discussed in Chapter 11 are required to flip membrane lipids and other polar molecules from one leaflet to the other. The lateral movements of specific plasma-membrane proteins and lipids can be quantified by a technique called fluorescence recovery after photobleaching (FRAP). Phospholipids containing a fluorescent substituent are used to monitor lipid movement. For proteins, a fragment of a monoclonal antibody (see Chapter 23) that is specific for the exoplasmic domain of the desired protein and that has only a single antigen-binding site is tagged with a fluorescent dye (alternatively, a genetic fusion of the desired protein with fluorescent green protein can accomplish the same goal). With this method, described in Figure 10-10, the rate at which membrane molecules move — the diffusion coefficient
— can be determined, as well as the proportion of the molecules that are laterally mobile. EXPERIMENTAL FIGURE 10-10 Fluorescence recovery after photobleaching (FRAP) experiments can quantify the lateral movement of proteins and lipids within the plasma membrane. (a) Experimental protocol. Step 1 : Cells are first labeled with a fluorescent reagent that binds uniformly to a specific membrane lipid or protein. Genetic fusion of the desired protein with green fluorescent protein (GFP) can accomplish the same goal. Step 2 : A laser light is then focused on a small area of the cell surface, irreversibly bleaching the bound reagent (or GFP) and thus reducing the fluorescence in the illuminated area. Step 3 : In time, the fluorescence of the bleached patch increases as unbleached fluorescent surface molecules diffuse into it and bleached ones diffuse outward. The extent of recovery of fluorescence in the bleached patch is proportional to the fraction of labeled molecules that are mobile in the membrane. (b) Results of a FRAP experiment with human hepatoma cells treated with a fluorescent antibody specific for the asialoglycoprotein receptor protein. The finding that 50 percent of the fluorescence returned to the bleached area indicates that 50 percent of the receptor molecules in the illuminated membrane patch were mobile and 50 percent were immobile. Because the rate of fluorescence recovery is proportional to the rate at which labeled molecules move into the bleached region, the diffusion coefficient of a protein or lipid in the membrane can be calculated from such data.
Lipid Composition Influences the Physical Properties of Membranes
[Data from Y. I. Henis et al., 1990, J. Cell Biol. 111:1409.] Description (a) The several steps in a fluorescence recovery after photo bleaching. A cell with a membrane containing membrane proteins is depicted as a circle. 1. The proteins are labeled with a fluorescent reagent, attached to each protein. 2. A region is bleached with a laser, shown as a patch of bleached fluorescent markers. 3. The recovery is shown as the fluorescent markers move around the membrane. (b) A graph showing fluorescence intensity over time. To the left top, a series of blue dots shows fluorescence before bleaching and then the main graph is a scatterplot with a quick rise followed by a steady horizontal flow near the 2000 mark. The results of FRAP studies with fluorescence-labeled phospholipids have shown that in fibroblast plasma membranes, all phospholipids are freely mobile over distances of about 0.5 μm, but most cannot diffuse over much longer distances. These findings suggest that protein-rich regions of the plasma membrane about 1 μm in diameter separate lipid-rich regions containing the bulk of the membrane phospholipids. Phospholipids are free to diffuse within such regions, but not from one lipid-rich region to an adjacent one. Furthermore, the rate of lateral diffusion of lipids in the plasma membrane is nearly an order of magnitude slower than in pure phospholipid bilayers: diffusion constants of per second and per second are characteristic of the plasma membrane and a pure phospholipid bilayer, respectively. This difference suggests that lipids may be tightly but not irreversibly bound to certain integral proteins in some membranes, as indeed has recently been demonstrated (see the discussion of annular phospholipids below).
Lipid Composition Influences the Physical Properties of Membranes A typical cell contains many different types of membranes, each with unique properties derived from its particular mix of lipids and proteins. The data in Table 10-1 illustrate the variation in lipid composition in different biomembranes. Several phenomena contribute to these differences. For instance, the relative abundances of phosphoglycerides and sphingolipids differ between membranes in the endoplasmic reticulum (ER), where phospholipids are synthesized, and the Golgi complex, where sphingolipids are synthesized. The proportion of sphingomyelin as a percentage of total membrane lipid phosphorus is about six times as high in Golgi membranes as it is in ER membranes. In other cases, the movement of membranes from one cellular compartment to another can selectively enrich certain membranes in lipids such as cholesterol. In responding to differing environments throughout an organism, different types of cells generate membranes with differing lipid compositions. In the cells that line the intestinal tract, for example, the membranes that face the harsh environment in which dietary nutrients are digested have a sphingolipid-to-phosphoglyceride-to-cholesterol ratio of 1:1:1, rather than the 0.5:1.5:1 ratio found in cells subject to less stress. The relatively high concentration of sphingolipids in these intestinal membranes may increase their stability because of extensive hydrogen bonding by the free group in the sphingosine moiety (see Figure 10-8).
TABLE 10-1 • Major Lipid Components of Selected Biomembranes
Composition (mol %) Source/Location PC PE + PS SM Cholesterol Plasma membrane (human erythrocytes) Myelin membrane (human neurons) Plasma membrane (mung bean) Inner mitochondrial membrane (cauliflower) Outer mitochondrial membrane (cauliflower) Plasma membrane (E. coli) Endoplasmic reticulum membrane (rat) Golgi membrane (rat) Inner mitochondrial membrane (rat) Outer mitochondrial membrane (rat) Primary leaflet location Exoplasmic Cytosolic Exoplasmic Both PC = phosphatidylcholine; PE = phosphatidylethanolamine; PS = phosphatidylserine, SM = sphingomyelin. SOURCE: Data from S. E. Horvath and G. Daum, 2013, “Lipids of Mitochondria,” Prog. Lipid Res. 52:590–614.
The degree of bilayer fluidity depends on lipid composition, the structure of the phospholipid hydrophobic tails, and temperature. As already noted, van der Waals interactions and the hydrophobic effect cause the nonpolar tails of phospholipids to aggregate. Long, saturated fatty acyl chains have the greatest tendency to aggregate, packing tightly together into a gel-like state. Phospholipids with short fatty acyl chains, which have less surface area and therefore fewer van der Waals interactions, form more fluid bilayers. Likewise, the kinks in cis-unsaturated fatty acyl chains (see
Chapter 2) result in their forming less stable van der Waals interactions with other lipids, and hence more fluid bilayers, than do straight saturated chains, which can pack more tightly together. Cholesterol is important in maintaining the appropriate fluidity of natural membranes, a property that appears to be essential for normal cell growth and reproduction. Cholesterol restricts the random movement of phospholipid head groups at the outer surfaces of the leaflets, but its effect on the movement of long phospholipid tails depends on its concentration. At the cholesterol concentrations normally present in the plasma membrane, the interaction of the steroid ring with the long hydrophobic tails of phospholipids tends to immobilize those lipids and thus decreases biomembrane fluidity. It is this property that can help organize the plasma membrane into discrete subdomains of unique lipid and protein composition. At lower cholesterol concentrations, however, the steroid ring separates and disperses phospholipid tails, causing the inner regions of the membrane to become slightly more fluid.
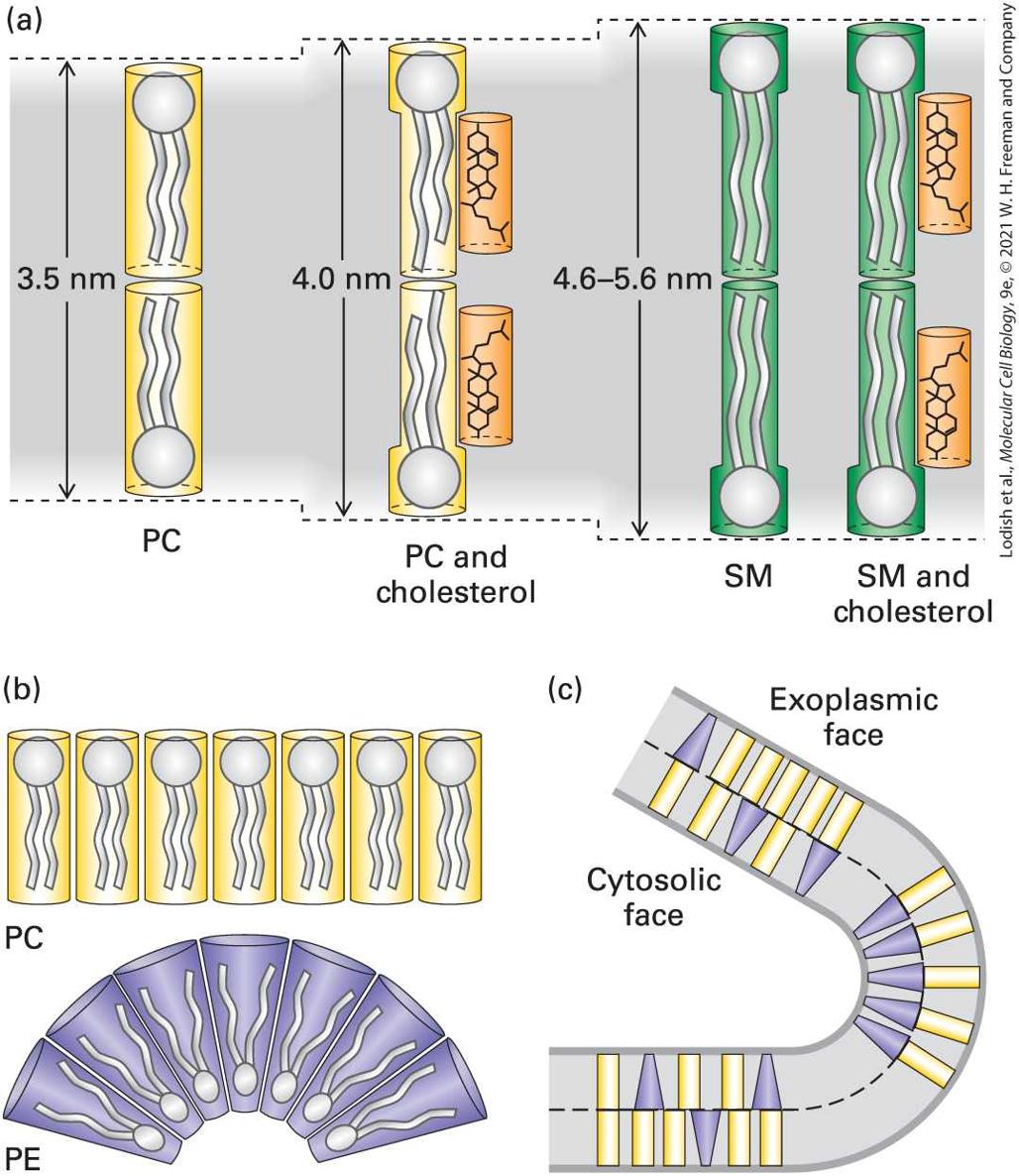
The lipid composition of a bilayer also influences its thickness, which in turn may influence the distribution of other membrane components, such as proteins, in a particular membrane. It has been argued that relatively short transmembrane segments of certain Golgi-resident enzymes (glycosyltransferases) are an adaptation to the lipid composition of the Golgi membrane and contribute to the retention of these enzymes in the Golgi apparatus. The results of biophysical studies on artificial membranes demonstrate that sphingomyelin associates into a more gellike and thicker bilayer than phosphoglycerides do (Figure 10-11a). Cholesterol and other molecules that decrease membrane fluidity also increase membrane bilayer thickness. Because sphingomyelin tails are already optimally stabilized, the addition of cholesterol has no effect on the thickness of a sphingomyelin bilayer.
FIGURE 10-11 Effect of lipid composition on bilayer thickness and curvature. (a) A pure sphingomyelin (SM) bilayer is thicker than one formed from a phosphoglyceride such as phosphatidylcholine (PC). Cholesterol has a lipid-ordering effect on phosphoglyceride bilayers that increases their thickness, but it does not affect the thickness of the more ordered SM bilayer. (b) Phospholipids such as PC have a cylindrical shape and form essentially flat monolayers, whereas those with smaller head groups, such as
phosphatidylethanolamine (PE), have a conical shape. (c) A bilayer enriched with PC in the exoplasmic leaflet and with PE in the cytosolic face, as in many plasma membranes, would have a natural curvature. [Information from H. Sprong et al., 2001, Nat. Rev. Mol. Cell Biol. 2:504–513.] Description (a) The effect of phospholipid on membrane curvature. Phosphatidylcholine (known as P C) forms a linear membrane, but phosphatidylethanolamine (known as P E) forms a curved membrane. (b) A membrane comprising P E and P C shows portions that are straight, where P C is dominant and the mixing is random, and curved portions, where the distribution of P E on the cytosolic face induces curvature in the membrane. (c) Chemical diagram of a phospholipid shows the ester bonds targeted by phospholipases. Another property dependent on the lipid composition of a bilayer is its curvature, which depends on the relative sizes of the polar head groups and nonpolar tails of its constituent phospholipids. Lipids with long tails and large head groups are cylindrical in shape; those with small head groups are cone-shaped (Figure 10-11b). As a result, bilayers composed of cylindrical lipids are relatively flat, whereas those containing large numbers of cone-shaped lipids form curved bilayers (Figure 10-11c). This effect of lipid composition on bilayer curvature may play a role in the formation of highly curved membranes, such as sites of viral budding (see
Figure 10-2) and of formation of internal vesicles from the plasma membrane (see Figure 10-6), and in specialized stable membrane structures such as microvilli. Several proteins, including reticulons (RTNs in vertebrates; reticulon-like proteins or RTNLs in other eukaryotes) bind to the surface of a phospholipid bilayer and cause the membrane to curve;
Lipid Composition Is Different in the Exoplasmic and Cytosolic Leaflets
such proteins are important in formation of transport vesicles that bud from a donor membrane (see Chapter 14). Lipid Composition Is Different in the Exoplasmic and Cytosolic Leaflets A characteristic of all biomembranes is an asymmetry in lipid composition across the bilayer. Although most phospholipids are present in both membrane leaflets, some are more abundant in one or the other leaflet. For instance, in plasma membranes from human erythrocytes and Madin-Darby canine kidney (MDCK) cells grown in culture, almost all the sphingomyelin and phosphatidylcholine, both of which form less fluid bilayers, are found in the exoplasmic leaflet. In contrast, phosphatidylethanolamine, phosphatidylserine, and phosphatidylinositol, which form more fluid bilayers, are preferentially located in the cytosolic leaflet. Because phosphatidylserine and phosphatidylinositol carry a net negative charge, the stretch of amino acids on the cytosolic face of a membrane protein with a single transmembrane segment is often enriched in positively charged (Lys, Arg) residues in close proximity to the transmembrane segment (this distribution of charged amino acids is referred to as the “inside positive” rule). This segregation of lipids across the bilayer may influence membrane curvature (see Figure 10-11c). Unlike particular phospholipids, cholesterol is relatively evenly distributed in both leaflets of cellular membranes.
The relative abundances of a particular phospholipid in the two leaflets of a plasma membrane can be determined experimentally on the basis of the susceptibility of phospholipids to hydrolysis by phospholipases, enzymes that cleave the ester bonds via which acyl chains and head groups are connected to the lipid molecule (Figure 10-12). When added to the external medium, phospholipases cannot cross the membrane, and thus they cleave off the head groups of only those lipids present in the exoplasmic face; phospholipids in the cytosolic leaflet are resistant to hydrolysis because the enzymes cannot penetrate to the cytosolic face of the plasma membrane.
FIGURE 10-12 Specificity of phospholipases. Each type of phospholipase cleaves one of the susceptible bonds shown in red. The glycerol carbon atoms are indicated by small numbers. In intact cells, only phospholipids in the exoplasmic leaflet of the plasma membrane are cleaved by phospholipases in the surrounding medium. Phospholipase C, a cytosolic enzyme, cleaves certain phospholipids in the cytosolic leaflet of the plasma membrane.
How the asymmetric distribution of phospholipids in membrane leaflets arises and is maintained is still unclear. As noted, in pure bilayers, phospholipids do not spontaneously migrate, or flip-flop, from one leaflet to the other. In part, the asymmetry in phospholipid distribution may reflect where these lipids are synthesized in the ER and Golgi. Sphingomyelin is synthesized on the luminal (exoplasmic) face of the Golgi, which becomes the exoplasmic face of the plasma membrane. In contrast, phosphoglycerides are synthesized on the cytosolic face of the ER membrane, which is topologically equivalent to the cytosolic face of the plasma membrane (see Figure 10-5). Clearly, however, this explanation does not account for the preferential location of phosphatidylcholine (a phosphoglyceride) in the exoplasmic leaflet. Movement of this phosphoglyceride, and perhaps others, from one leaflet to the other in some natural membranes is most likely catalyzed by ATPpowered transport proteins called flippases, which are discussed in
Chapter 11. The preferential location of lipids on one face of the bilayer is necessary for a variety of membrane-based functions. For example, the head groups of all phosphorylated forms of phosphatidylinositol (see Figure 10-8b; PI), an important source of second messengers, face the cytosol. Stimulation of many cell-surface receptors by their corresponding ligands results in activation of the cytosolic enzyme phospholipase C, which can then hydrolyze the bond within PI connecting the phosphoinositols to the diacylglycerol. As we will see in Chapter 15, both water-soluble phosphoinositols and membrane-embedded diacylglycerol participate in intracellular signaling pathways that affect many aspects of cellular
Cholesterol and Sphingolipids Cluster with Specific Proteins in Membrane Microdomains
metabolism. Phosphatidylserine, too, is normally most abundant in the cytosolic leaflet of the plasma membrane. In the initial stages of platelet stimulation by serum, phosphatidylserine is briefly translocated to the exoplasmic face, presumably by a flippase enzyme, where it activates enzymes participating in blood clotting. When cells die, lipid asymmetry is no longer maintained, and phosphatidylserine, normally enriched in the cytosolic leaflet, is increasingly found in the exoplasmic one. This increased exposure is detected experimentally by use of a labeled version of annexin V, a protein that specifically binds to phosphatidylserine, to measure the onset of programmed cell death (apoptosis). As discussed in
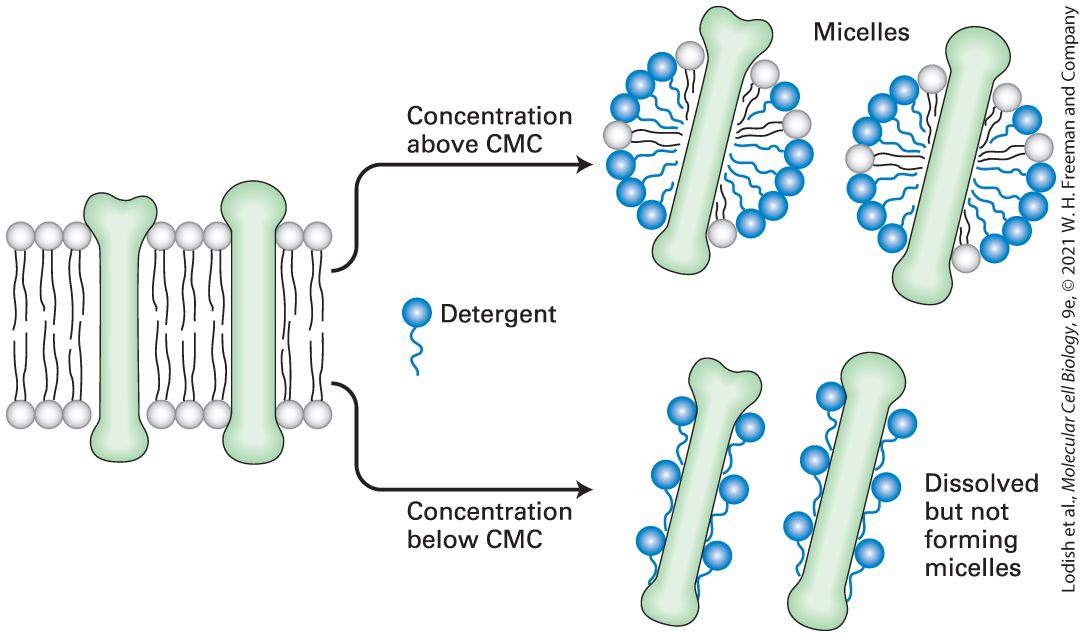
Chapter 22, the increased exposure of phosphatidylserine on dying or dead cells is recognized by phagocytic cells, which initiate engulfment of such apoptotic bodies and thus ensure timely and safe disposal of cell remnants. Cholesterol and Sphingolipids Cluster with Specific Proteins in Membrane Microdomains Membrane lipids are not randomly distributed (evenly mixed) in each leaflet of a bilayer. One hint that lipids may be organized within the leaflets was the discovery that the lipids remaining after the extraction (solubilization) of plasma membranes with non-ionic detergents such as Triton X-100 predominantly contain two species: cholesterol and sphingomyelin. Because these two lipids are found in more ordered, less fluid bilayers, researchers hypothesized that they form microdomains, termed lipid rafts, surrounded by other, more fluid phospholipids that are
Cells Store Excess Lipids in Lipid Droplets
more readily extracted by non-ionic detergents. (We discuss more fully the role of ionic and non-ionic detergents in extracting membrane proteins in Section 10.2.) Some biochemical and microscopic evidence supports the existence of lipid rafts, which in natural membranes are typically 50 nm in diameter. Rafts can be disrupted by methyl-β-cyclodextrin, which specifically extracts cholesterol from membranes, or by antibiotics such as filipin that sequester cholesterol into aggregates within the membrane. Such findings indicate the importance of cholesterol in maintaining the integrity of lipid rafts. These raft fractions, defined by their insolubility in non-ionic detergents, contain a subset of plasma-membrane proteins, many of which are implicated in sensing extracellular signals and transmitting them into the cytosol. Because raft fractions are enriched in glycolipids, an important tool for microscopic visualization of raft-type structures in intact cells is the use of fluorescently labeled cholera toxin, a protein that specifically binds to certain gangliosides. By bringing many key proteins into close proximity and stabilizing their interactions, lipid rafts may facilitate signaling by cell-surface receptors and the subsequent activation of cytosolic events. However, much remains to be learned about the structure and biological function of lipid rafts. The unique properties of some of the raft-associated lipids, such as glycolipids, may permit interactions of their tails across the hydrophobic core and help organize lipids of the cytosolic leaflet in the formation of signaling platforms.
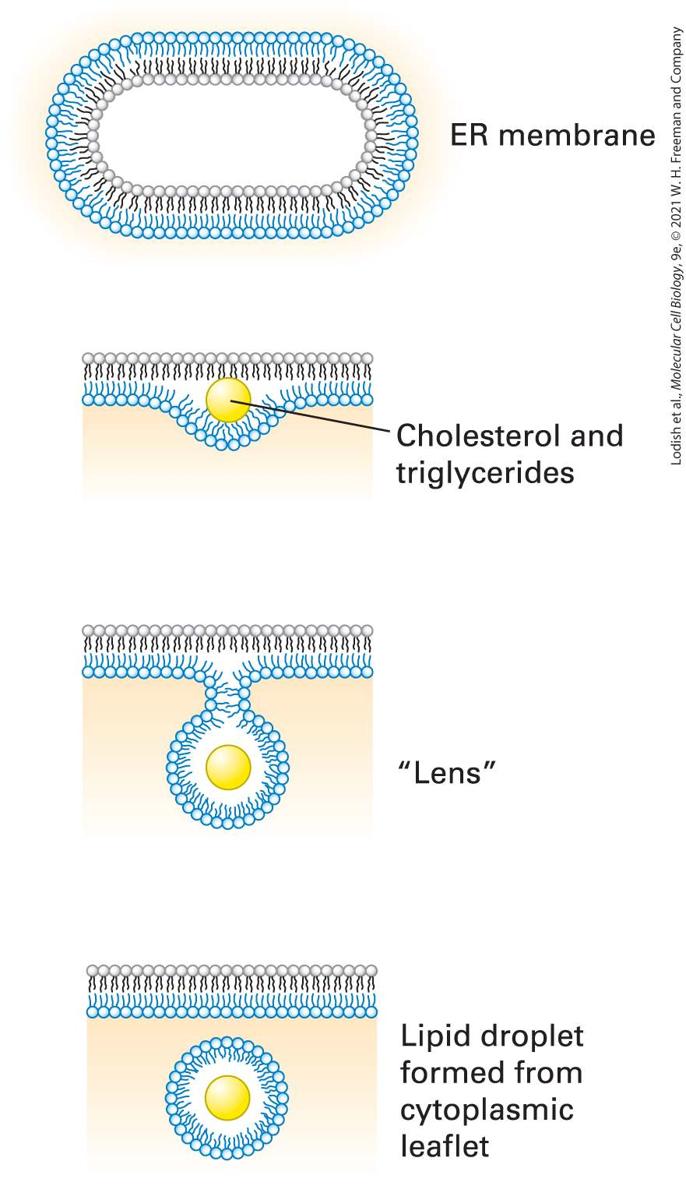
Cells Store Excess Lipids in Lipid Droplets Lipid droplets are vesicular structures, composed of triglycerides and cholesterol esters, that originate from the ER and serve a lipid-storage function. When a cell’s supply of lipids exceeds the immediate need for membrane construction, excess lipids are relegated to these lipid droplets, readily visualized in live cells by staining with a lipophilic dye such as Congo red. Feeding cells with oleic acid, a type of fatty acid, enhances lipid droplet formation. Lipid droplets are not only storage compartments for triglycerides and cholesterol esters, but may also serve as platforms for storage of proteins targeted for degradation. The biogenesis of lipid droplets starts with delamination of the lipid bilayer of the ER through insertion of triglycerides and cholesterol esters that are more hydrophobic than cholesterol itself (Figure 10-13). The lipid “lens” continues to grow by insertion of more lipid, until finally a lipid droplet is hatched by scission from the ER. The resulting cytoplasmic droplet is thereby surrounded by a phospholipid monolayer. The details of lipid droplet biogenesis, as well as its functions, remain to be defined more clearly.
FIGURE 10-13 Lipid droplets form by budding and scission from the ER membrane. Lipid droplet formation begins with the accumulation of cholesterol esters and triglycerides within the hydrophobic core of the lipid bilayer. The resulting delamination of the two lipid monolayers causes a “lens” to form, the further growth of which creates a spherical droplet that is then released by scission at the neck. The newly formed droplet is surrounded by a lipid monolayer, derived from the cytosolic leaflet of the ER membrane. Description An illustration shows an oval representation of the endoplasmic reticulum membrane. The following steps occur in the formation of a lipid droplet. 1. Cholesterol and triglycerides collect together in the hydrophobic core. 2. The cytosolic portion begins to bud, closing around the yellow lipid droplet, forming a lens-like structure. 3. The lipid droplet forms and separates from the membrane. KEY CONCEPTS OF SECTION 10.1 The Lipid Bilayer: Composition and Structural Organization Membranes are crucial to cell structure and function. The eukaryotic cell is demarcated from the external environment by the plasma membrane and organized into membrane-limited subcompartments (organelles and vesicles). The phospholipid bilayer, the basic structural unit of all biomembranes, is a lipid sheet two molecules thick, with hydrophilic faces and a hydrophobic core that is impermeable to water-soluble molecules and ions. Proteins embedded in the bilayer endow the membrane with specific functions (see Figure 10-1). Phospholipids spontaneously form bilayers and sealed compartments surrounding an aqueous space (see Figure 10-3). As bilayers, all biological membranes have an internal (cytosolic) face and an external (exoplasmic) face (see Figure 10-5). Some organelles, such as E. coli, are surrounded by two, rather than one, membrane bilayers. The primary lipid components of biomembranes are phosphoglycerides, sphingolipids, and sterols such as cholesterol. The term “phospholipid” applies to any amphipathic lipid molecule with a fatty acyl hydrocarbon tail and a phosphate-based polar head group (see Figure 10-8).
Biomembranes can undergo phase transitions from fluidlike to gel-like states depending on the temperature and the composition of the membrane (see Figure 109). Most lipids and many proteins are laterally mobile in biomembranes (see Figure 1010). Different cellular membranes vary in lipid composition (see Table 10-1). Phospholipids and sphingolipids are asymmetrically distributed in the two leaflets of the bilayer, whereas cholesterol is fairly evenly distributed in both leaflets. Natural biomembranes generally have a viscous consistency with fluidlike properties. In general, membrane fluidity is decreased by sphingolipids and cholesterol and increased by phosphoglycerides. The lipid composition of a membrane also influences its thickness and curvature (see Figure 10-11). Lipid rafts are microdomains containing cholesterol, sphingolipids, and certain membrane proteins that form in the plane of the bilayer. These lipid-protein aggregates might facilitate signaling by certain plasma-membrane receptors. Lipid droplets are storage vesicles for lipids, originating in the ER (see Figure 10-13).
10.2 Membrane Proteins: Structure and Basic Functions
10.2 Membrane Proteins: Structure and Basic Functions Membrane proteins are defined by their location within or at the surface of a phospholipid bilayer. Although every biological membrane has the same basic bilayer structure, in which the proteins associated with a particular membrane are responsible for its distinctive activities. The kinds and amounts of proteins associated with biomembranes vary depending on cell type and subcellular location. For example, the inner mitochondrial membrane is 76 percent protein, but the myelin membrane that surrounds nerve axons is only 18 percent. The high phospholipid content of myelin allows it to electrically insulate the nerve from its environment, as we discuss in Chapter 23. The importance of membrane proteins is evident from the finding that approximately a third of all yeast genes encode a membrane protein. The relative abundance of genes for membrane proteins is even greater in multicellular organisms, in which they have additional functions in cell adhesion and in communication between different cell types (cell-cell interactions). Indeed, of the ~20,000 genes in the human genome, ~7000 encode membrane proteins. The lipid bilayer presents a distinctive two-dimensional hydrophobic environment for membrane proteins. Some proteins contain segments that are embedded within the hydrophobic core of the phospholipid bilayer; other proteins are associated with the exoplasmic or cytosolic leaflet of
Proteins Interact with Membranes in Three Different Ways
the bilayer. Protein domains on the extracellular surface of the plasma membrane generally bind to extracellular molecules, including external signaling proteins, ions, and small metabolites (e.g., glucose, fatty acids), as well as proteins on other cells or in the external environment. Segments of proteins within the plasma membrane perform multiple functions, such as forming the channels and pores through which molecules and ions move into and out of cells. These intramembrane segments also serve to organize multiple membrane proteins into larger assemblies within the plane of the membrane. Domains lying along the cytosolic face of the plasma membrane have a wide range of functions, from anchoring cytoskeletal proteins to the membrane to triggering intracellular signaling pathways or synthesis of ATP. In many cases, the function of a membrane protein and the topology of its polypeptide chain in the membrane can be predicted on the basis of its similarity with other well-characterized proteins. In this section, we examine the characteristic structural features of membrane proteins and some of their basic functions. We describe the structures of several proteins to help you get a feel for how membrane proteins interact with membranes. More complete characterizations of the properties of various types of membrane proteins are presented in later chapters that focus on their structures and activities in the context of their cellular functions. Proteins Interact with Membranes in Three Different Ways
Membrane proteins can be classified into three categories — integral, lipid-anchored, and peripheral — on the basis of their position with respect to the membrane (see Figure 10-1). Integral membrane proteins, also called transmembrane proteins, span a phospholipid bilayer and comprise three domains. The cytosolic and exoplasmic domains have hydrophilic exterior surfaces that interact with the aqueous environment on the cytosolic and exoplasmic faces of the membrane. These domains resemble segments of other water-soluble proteins in their amino acid composition and structure. In contrast, the membrane-spanning segments usually contain many hydrophobic amino acids whose side chains protrude outward and interact with the hydrophobic hydrocarbon core of the phospholipid bilayer. In all transmembrane proteins examined to date, the membrane-spanning domains consist of one or more α helices or of multiple β strands. We discuss the ribosomal synthesis and posttranslational processing of soluble cytosolic proteins in Chapters 5 and 9; the process by which integral membrane proteins are inserted into membranes as part of their synthesis is discussed in Chapter 13. Lipid-anchored membrane proteins are bound covalently to one or more lipid molecules. The hydrophobic tail of the attached lipid is embedded in one leaflet of the membrane and anchors the protein to the membrane. The polypeptide chain itself does not enter the phospholipid bilayer. Peripheral membrane proteins (see Figure 10-1) do not directly contact the hydrophobic core of the phospholipid bilayer. Instead, they are bound to the membrane either indirectly by interactions with integral or lipidanchored membrane proteins or directly by interactions with lipid head
Most Transmembrane Proteins Have Membrane-Spanning α Helices
groups. Peripheral proteins can be bound to either the cytosolic or the exoplasmic face of the plasma membrane. In addition to these proteins, which are closely associated with the bilayer, cytoskeletal filaments can be more loosely associated with the cytosolic face, usually through one or more peripheral adapter proteins. Such associations with the cytoskeleton provide support for various cellular membranes, helping to determine the cell’s shape and mechanical properties, and play a role in the two-way communication between the cell interior and the exterior, as we explore in
Chapter 17. Finally, peripheral proteins on the outer surface of the plasma membrane and the exoplasmic domains of integral membrane proteins are often attached to components of the extracellular matrix or to the cell walls surrounding bacterial and plant cells, providing a crucial interface between the cell and its environment. Most Transmembrane Proteins Have Membrane-Spanning α Helices Soluble proteins exhibit hundreds of distinct localized folded structures, or motifs (see Figure 3-7). In comparison, the repertoire of folded structures in the transmembrane domains of integral membrane proteins is quite limited, with the hydrophobic α helix predominating. Proteins containing membrane-spanning α-helical domains are stably embedded in membranes. This is due to energetically favorable hydrophobic and van der Waals interactions of the hydrophobic side chains in the domain with specific lipids and probably also because of ionic interactions with the polar head groups of the phospholipids.
A single α-helical domain is sufficient to incorporate an integral membrane protein into a membrane. However, many such proteins have more than one transmembrane α helix. Typically, a membrane-embedded α helix is composed of a continuous segment of 20–25 hydrophobic (uncharged) amino acids (see Figure 2-14). The predicted length of such an α helix (3.75 nm) is just sufficient to span the hydrocarbon core of a phospholipid bilayer. In many membrane proteins, these helices are perpendicular to the plane of the membrane, whereas in others, the helices traverse the membrane at an oblique angle. The hydrophobic side chains protrude outward from the helix and form van der Waals interactions with the fatty acyl chains in the bilayer. In contrast, the hydrophilic amide peptide bonds are in the interior of the α helix (see Figure 3-4); each carbonyl ( ) group forms a hydrogen bond with the amide hydrogen atom of the amino acid four residues toward the C-terminus of the helix. These polar groups are shielded from the hydrophobic interior of the membrane. To help you get a better sense of the structures of proteins with α-helical domains, we will briefly discuss four different kinds of such proteins: glycophorin A, G protein–coupled receptors, aquaporins (water/glycerol channels), and the T-cell receptor for antigen. Glycophorin A, the major protein in the erythrocyte plasma membrane, is a representative single-pass transmembrane protein, which contains only one membrane-spanning α helix (Figure 10-14a). The 23-residue membrane-spanning α helix is composed of amino acids with hydrophobic (uncharged) side chains, which interact with the fatty acyl chains in the
surrounding bilayer. In cells, glycophorin A typically forms dimers: the transmembrane helix of one glycophorin A polypeptide associates with the corresponding transmembrane helix in a second glycophorin A to form a coiled-coil structure (Figure 10-14b). Such interactions of membranespanning α helices are a common mechanism for creating dimeric membrane proteins, and many membrane proteins form oligomers (two or more polypeptides bound together noncovalently) by interactions between their membrane-spanning helices.
FIGURE 10-14 Structure of glycophorin A, a typical single-pass transmembrane protein. (a) Diagram of dimeric glycophorin, showing its major sequence features and its relation to the membrane. The single 23-residue membrane-spanning α helix in each monomer is composed of amino acids with hydrophobic (uncharged) side chains (red and green spheres). By binding negatively charged phospholipid head groups, the positively charged arginine and lysine residues (blue spheres) near the cytosolic side of the helix help anchor glycophorin in the membrane. Both the extracellular and the cytosolic domains are rich in charged residues and polar uncharged residues; the extracellular domain is heavily
glycosylated, with carbohydrate chains (green diamonds) attached to specific serine, threonine, and asparagine residues. (b) Molecular model of the transmembrane domain of dimeric glycophorin A corresponding to residues 73–96. The hydrophobic side chains of the α helix in one monomer are shown in pink; those of the other monomer, in green. Residues depicted as space-filling structures participate in van der Waals interactions that stabilize the coiled-coil dimer. Note how the hydrophobic side chains project outward from the helix, toward what would be the surrounding fatty acyl chains. [Part (b) Data from K. R. MacKenzie, J. H. Prestegard, and D. M. Engelman, 1997, Science 276:131, PDB ID 1afo.] Description (a) An illustration shows the glycophorin A dimer. The protein spans the plasma membrane, the c terminus being within the cytosol and the n terminus being in the extracellular domain. A 23 residue hydrophobic alpha helix spans the residue, from residues 73 to 96. The extracellular domain of the protein contains many glycosylated regions. (b) The space filling model highlights the coiled-coil dimer stabilized by van der Waals interactions between adjacent side chains. A large and important group of integral membrane proteins is defined by the presence of seven membrane-spanning α helices. This group includes the large family of G protein–coupled cell-surface receptors discussed in
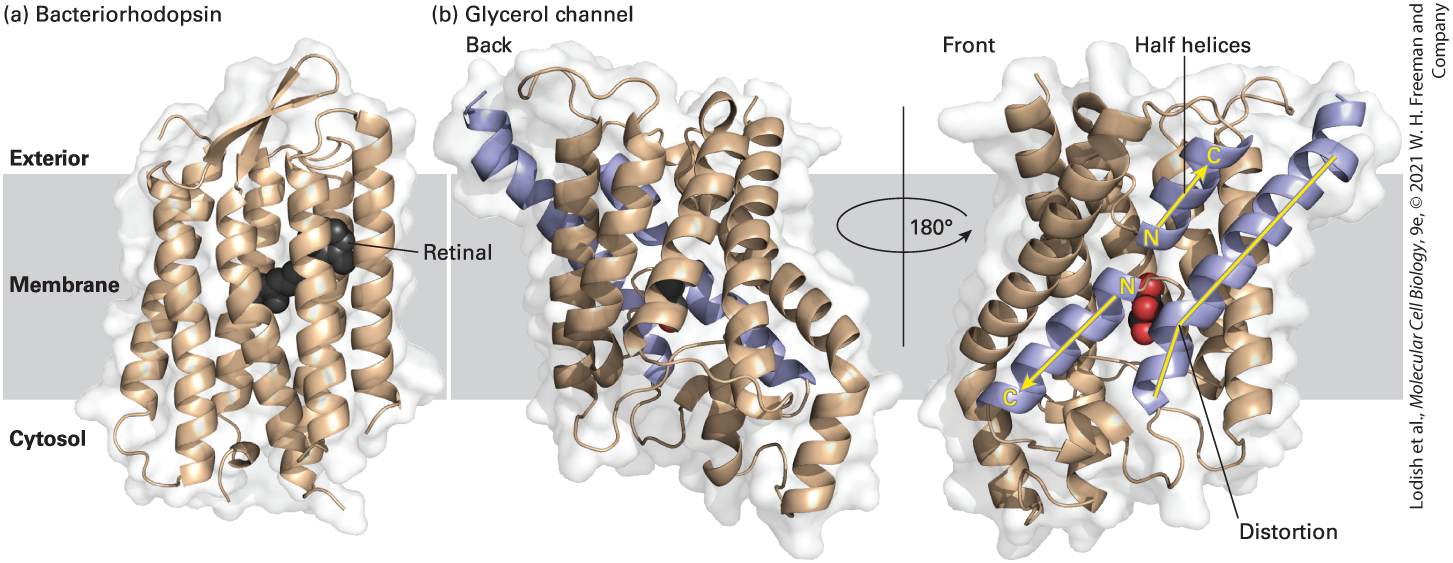
Chapter 15, many of which have been crystallized. One such multipass transmembrane protein of known structure is bacteriorhodopsin, a protein found in the membranes of certain photosynthetic bacteria; it illustrates the general structure of all these proteins (Figure 10-15a). Absorption of light by the retinal group covalently attached to this protein causes a conformational change in the protein that results in the pumping of protons from the cytosol across the bacterial membrane to the extracellular space. The proton concentration gradient thus generated
across the membrane is used to synthesize ATP during photosynthesis (see
Chapter 12). In the high-resolution structure of bacteriorhodopsin, the positions of all the individual amino acids, the retinal group, and the surrounding lipids are clearly defined. As might be expected, virtually all the amino acids on the exterior of the membrane-spanning segments of bacteriorhodopsin are hydrophobic, permitting energetically favorable interactions with the hydrocarbon core of the surrounding lipid bilayer.
FIGURE 10-15 Structural models of two multipass transmembrane proteins. (a) The seven hydrophobic α helices in bacteriorhodopsin (a photoreceptor in certain bacteria) traverse the lipid bilayer roughly perpendicular to the plane of the membrane. A retinal molecule (black) covalently attached to one helix absorbs light. The large class of G protein–coupled receptors in eukaryotic cells also has seven membrane-spanning α helices; their three-dimensional structure is similar to that of bacteriorhodopsin. (b) Two views of the glycerol channel Glpf, rotated 180° with respect to each other along an axis perpendicular to the plane of the membrane. Note the several membrane-spanning α helices that are at oblique angles, the two helices that penetrate only halfway through the membrane (purple with yellow arrows), and the one long membrane-spanning helix with a “break” or distortion in the middle (purple with yellow line). The glycerol molecule in the hydrophilic “core” is colored red. The protein structure was approximately positioned in the hydrocarbon core of the membrane by finding the most hydrophobic 3-μm slab of the protein perpendicular to the membrane plane.
[Part (a) Data from H. Luecke et al., 1999, J. Mol. Biol. 291:899. Part (b) Data from J. Bowie, 2005, Nature 438:581–589, PDB ID 1c3w; and D. Fu et al., 2000, Science 290:481–486, PDB ID 1fx8.] Description (a) A 3-D model shows alpha helices in bacteriorhodopsin, a transmembrane protein. The model shows a retinal molecule docked in the center of the protein. Two 3-D views of the glycerol channel are depicted in section b, comprising alpha helices and halfhelices. Glycerol is docked in the middle of the protein, close to the site of the distortion in a membrane spanning helix containing a distortion. The second view is a 180 degree turn from the first view so oblique angles can be presented. The aquaporins are a large family of highly conserved proteins that transport water, glycerol, and other hydrophilic molecules across biomembranes. They illustrate several aspects of the structure of multipass transmembrane proteins. Aquaporins are tetramers of four identical subunits. Each of the four subunits has six membrane-spanning α helices, some of which traverse the membrane at oblique angles rather than perpendicularly. Because all aquaporins have similar structures, we will focus on one, the glycerol channel Glpf, whose structure has been especially well defined by x-ray diffraction studies (Figure 10-15b). This aquaporin has one long transmembrane helix with a bend in the middle, and more strikingly, there are two α helices that penetrate only halfway through the membrane. The N-termini of these helices face each other (yellow N’s in the figure), and together they span the membrane at an oblique angle. Thus some membrane-embedded helices — and other, nonhelical, structures we will encounter later — do not traverse the entire bilayer. As we will see in Chapter 11, these short helices in aquaporins
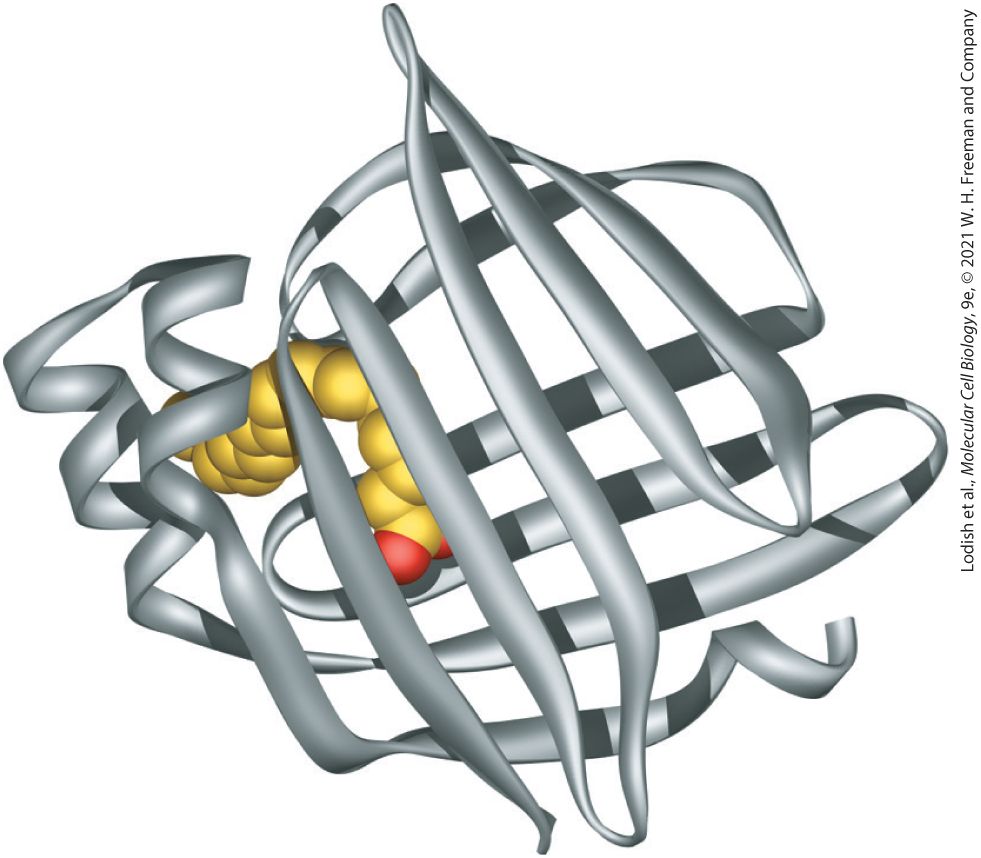
form part of the glycerol/water-selective pore in the middle of each subunit (see Figure 11-8). This structure highlights the considerable diversity in the ways membrane-spanning α helices interact with the lipid bilayer and with other segments of the protein. The specificity of phospholipid-protein interactions is evident from the structure of a different aquaporin, aquaporin 0 (Figure 10-16). Aquaporin 0 is the most abundant protein in the plasma membrane of the fiber cells that make up the bulk of the lens of the mammalian eye. Like other aquaporins, it is a tetramer of identical subunits. The protein’s surface is not covered by a set of uniform binding sites for phospholipid molecules. Instead, fatty acyl chains pack tightly against the irregular hydrophobic outer surface of the protein. The lipids involved in this interaction are referred to as annular phospholipids because they form a tight ring (annulus) of lipids around the protein that are not easily exchanged with bulk phospholipids in the bilayer. Some of the fatty acyl chains are straight, in the trans conformation (see Chapter 2), whereas others are kinked in order to interact with bulky hydrophilic side chains on the surface of the protein. Some of the lipid head groups are parallel to the surface of the membrane, as is the case in pure phospholipid bilayers. Others, however, are oriented almost at right angles to the plane of the membrane. Thus there can be specific interactions between phospholipids and membrane-spanning proteins, and the function of many membrane proteins can be affected by the specific types of phospholipids present in the bilayer.
FIGURE 10-16 Annular phospholipids. Side view of the three- dimensional structure of one subunit of the lens-specific aquaporin 0 homotetramer, crystallized in the presence of the phospholipid dimyristoylphosphatidylcholine, a phospholipid with 14 carbon-saturated fatty acyl (myristic acid) chains. Note the lipid molecules forming a bilayer shell around the protein, which is shown as a surface plot (the lighter background molecule). The lipid molecules are shown in space-filling format; the polar lipid head groups (gray and red) and the lipid fatty acyl chains (the extended black and gray structures) form a bilayer with almost uniform thickness around the protein. Presumably, in the membrane, lipid fatty acyl chains cover the whole of the hydrophobic surface of the protein; only the most ordered of the lipid molecules would be resolved in the crystallographic structure. [Data from A. Lee, 2005, Nature 438:569–570; and T. Gonen et al., 2005, Nature 438:633– 688, PDB ID 2b6o.] Description
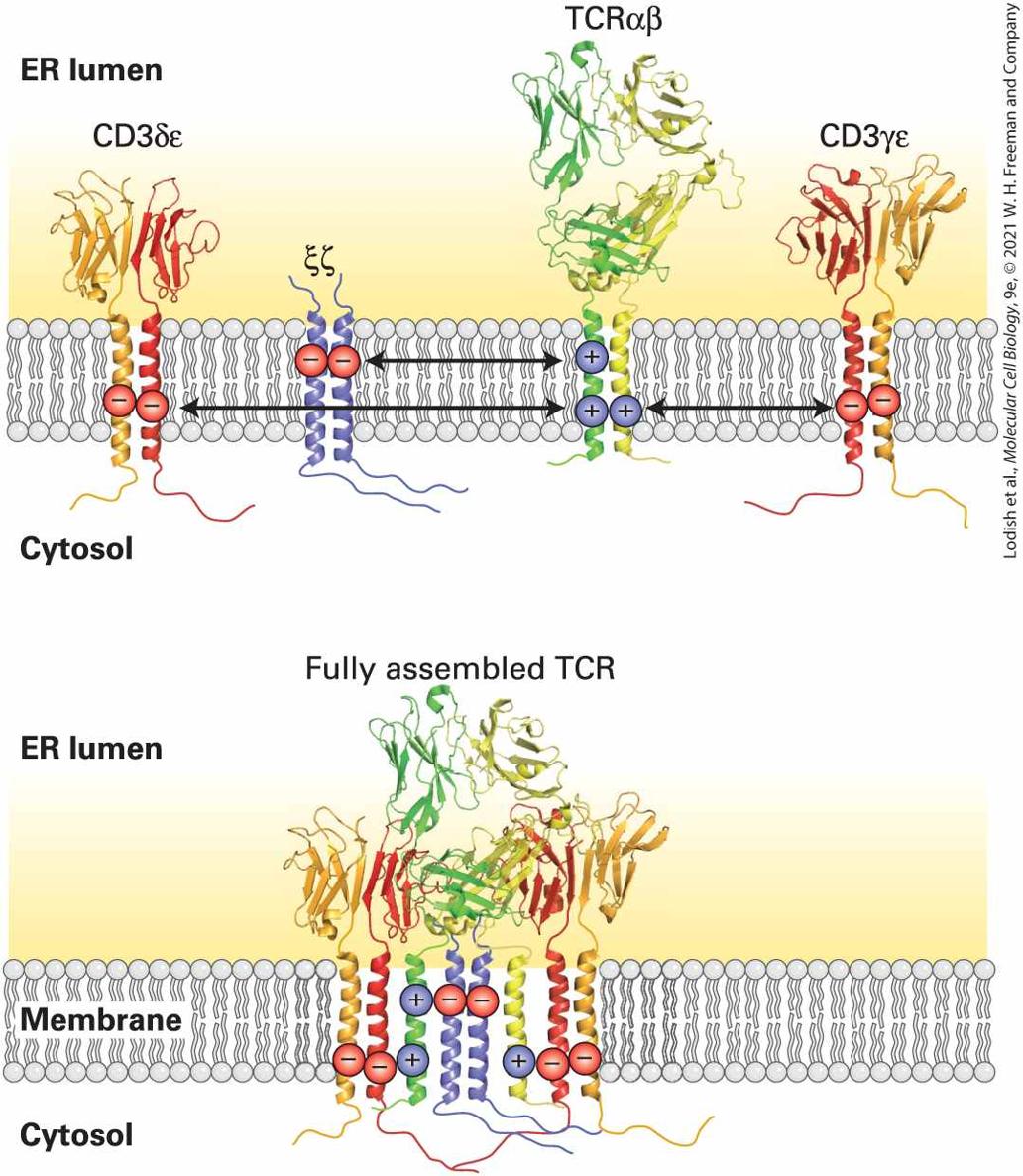
The 3-D model of the aquaporin tetramer, determined by X-ray crystallography is presented in surface view. The transmembrane protein is covered in phospholipid, the heads being oriented at the exterior and cytosolic ends of the protein. In addition to the predominantly hydrophobic (uncharged) residues that serve to embed integral membrane proteins in the bilayer, many α-helical transmembrane segments contain polar or charged residues. Their amino acid side chains can be used to guide the assembly and stabilization of multimeric membrane proteins. The T-cell receptor for antigen is a case in point: it is composed of four separate dimers, the interactions of which are driven by charge-charge interactions between α helices at the appropriate “depth” in the hydrocarbon core of the lipid bilayer (Figure 10-17). The electrostatic attraction of positive and negative charges on each dimer helps the dimers to find each other. Thus charged residues in otherwise hydrophobic transmembrane segments can help guide assembly of multimeric membrane proteins.
FIGURE 10-17 Charged residues can orchestrate the assembly of multimeric membrane proteins. The T-cell receptor (TCR) for antigen is composed of four separate dimers: an αβ pair directly responsible for antigen recognition and accessory subunits collectively referred to as the CD3 complex. These accessory subunits include the γ, δ, ε, and ζ subunits. The ζ subunits form a disulfide-linked homodimer. The γ and δ subunits occur in complex with an ε subunit to generate a γε and a δε pair. The transmembrane
Multiple β Strands in Porins Form Membrane-Spanning “Barrels”
segments of the TCR α and β chains each contain positively charged residues (blue). These residues allow recruitment of corresponding δε and γε heterodimers, which carry negative charges (red) at the appropriate depth in the hydrophobic core of the bilayer. The ζ homodimer docks onto the charges in the TCR α chain (dark green), while the γε and δε subunit pairs find their corresponding partners deeper down in the hydrophobic core on both the TCR α and TCR β chain (light green). Charged residues in otherwise nonpolar transmembrane segments can thus guide assembly of higher order structures. [Data from K. W. Wucherpfennig et al., 2010, Cold Spring Harb. Perspect. Biol. 2:a005140, PDB ID 1xmw; M. E. Call et al., 2006, Cell 127:355, PDB ID 2hac; and L. KjerNielsen et al., 2003, Immunity 18:53, PDB ID 1mi5.] Description The first illustration shows four transmembrane proteins for the T cell receptor complex embedded in the endoplasmic reticulum membrane. The hydrophobic portion of each membrane is indicated as charged, and the attraction between opposing charged proteins is indicated by double headed arrows. In the second illustration, the proteins have formed a fully assembled T cell receptor complex due to opposing charges. Multiple β Strands in Porins Form Membrane-Spanning “Barrels” The porins are a class of transmembrane proteins whose structure differs radically from that of other integral membrane proteins based on α-helical transmembrane domains. (Note that the aquaporins discussed above, despite their name, are not porins and contain multiple transmembrane α helices.) Several types of porins are found in the outer membranes of gram-negative bacteria such as E. coli (see Figure 1-12a) and in the outer membranes of mitochondria and chloroplasts. The outer membrane
protects an intestinal bacterium from harmful agents (e.g., antibiotics, bile salts, and proteases) but permits the uptake and disposal of small hydrophilic molecules, including nutrients and waste products. Different types of porins in the outer membrane of an E. coli cell provide channels for the passage of specific types of disaccharides or other small molecules as well as for ions such as phosphate. The amino acid sequences of porins contain none of the long, continuous hydrophobic segments typical of integral membrane proteins with α-helical membrane-spanning domains. Rather, the entire outer surface of the fully folded porin displays its hydrophobic character to the hydrocarbon core of the lipid bilayer. X-ray crystallography shows that porins are trimers of identical subunits. In each subunit, 16 β strands form a sheet that twists into a barrel-shaped structure with a pore in the center (Figure 10-18). Unlike a typical water-soluble globular protein, a porin has a hydrophilic interior and a hydrophobic exterior; in this sense, porins are inside out. In a porin monomer, the outward-facing side chains on each of the β strands are hydrophobic and form a nonpolar ribbonlike band that encircles the outside of the barrel. This hydrophobic band interacts with the fatty acyl groups of the membrane lipids or with other porin monomers. The side chains facing the inside of a porin monomer are predominantly hydrophilic; they line the pore through which small water-soluble molecules cross the membrane.
Covalently Attached Lipids Anchor Some Proteins to Membranes
FIGURE 10-18 Structural model of one subunit of OmpX, a porin found in the outer membrane of E. coli. All porins are trimeric transmembrane proteins. Each subunit is barrel shaped, with β strands forming the wall and a transmembrane pore in the center. A band of aliphatic (hydrophobic and noncyclic) side chains (yellow) and a border of aromatic (ringcontaining) side chains (red) position the protein in the bilayer. [Data from J. Vogt and G. E. Schulz, 1999, Structure 7:1301, PDB ID 1qj8.] Recent advances in structural biology, notably the application of cryoelectron microscopy (see Chapter 3) have made it possible to study the structure of membrane proteins, including their transmembrane segments, at near-atomic resolution. Examples include members of the ATP-binding cassette family of transporters (ABC transporters). Covalently Attached Lipids Anchor Some Proteins to Membranes In eukaryotic cells, covalently attached lipids anchor some otherwise typically water-soluble proteins to one or the other leaflet of the membrane. In such lipid-anchored proteins, the hydrocarbon chains of the lipid anchor are embedded in the bilayer, but the protein itself does not enter the bilayer. The lipid anchors used to anchor proteins to the cytosolic face are not used for the exoplasmic face, and vice versa. One group of cytosolic proteins is anchored to the cytosolic face of a membrane by a fatty acyl group (e.g., myristate or palmitate) covalently attached to an N-terminal glycine residue, a modification called acylation
(Figure 10-19a). Retention of such proteins at the membrane by the N-terminal acyl anchor may play an important role in a membraneassociated function. For example, v-Src, a mutant form of a cellular tyrosine kinase, induces abnormal cellular growth that can lead to cancer, but does so only when it has a myristylated N-terminus (see Chapter 25). A second group of cytosolic proteins are anchored to membranes by a hydrocarbon chain attached to a cysteine residue at or near the C-terminus, a modification called prenylation (Figure 10-19b). Prenyl anchors are built from 5-carbon isoprene units, which, as detailed in Section 10.3, are also used in the synthesis of cholesterol. In prenylation, a 15-carbon farnesyl or 20-carbon geranylgeranyl group is bound by a thioether bond to the group of a C-terminal cysteine residue of the protein, usually part of a C-terminal Cys-Ala-Ala-X (X = any of a number of amino acids) or CAAX box. Once prenylation has occurred, the C-terminal Ala-Ala-X motif may be removed by proteolysis. In some cases, a second geranylgeranyl group or a fatty acyl palmitate group is linked to a nearby cysteine residue. The additional hydrocarbon anchor is thought to reinforce the attachment of the protein to the membrane. For example, Ras, a GTPase superfamily protein that functions in intracellular signaling (see Chapter 16), is recruited to the cytosolic face of the plasma membrane by such a double anchor. Rab proteins, which also belong to the GTPase superfamily, are similarly bound to the cytosolic surface of intracellular vesicles by prenyl anchors; these proteins are required for the fusion of vesicles with their target membranes (see Chapter 14). Yet other proteins are palmitoylated on membrane-proximal cysteine residues in the absence of other lipid modifications.
FIGURE 10-19 Anchoring of plasma-membrane proteins to the phospholipid bilayer by covalently linked hydrocarbon groups. (a) Cytosolic proteins such as v-Src are associated with the plasma membrane through a single fatty acyl chain attached to the N-terminal glycine (Gly) residue of the polypeptide. Myristate (C14) and palmitate (C16) are common acyl anchors. (b) Other cytosolic proteins (e.g., Ras and Rab proteins) are anchored to the membrane by prenylation of one or two cysteine (Cys) residues at or near the C-terminus. The anchors are farnesyl (C15) and geranylgeranyl (C20) groups, both of which are unsaturated. (c) The lipid anchor on the exoplasmic surface of the plasma membrane is
glycosylphosphatidylinositol (GPI). The phosphatidylinositol part (red) of this anchor contains two fatty acyl chains that extend into the bilayer. The phosphoethanolamine unit (purple) in the anchor links it to the protein. The two green hexagons represent sugar units, which vary in number, nature, and arrangement in different GPI anchors. The complete structure of a yeast GPI anchor is shown in Figure 13-15. [Information from H. Sprong et al., 2001, Nature Rev. Mol. Cell Biol. 2:504–513.] Description The three types of lipid anchors are as follows. (a) acylation is depicted with a C O O-at the cytosol end attached to the membrane by a glycine. (b) prenylation with a N H subscript 3 at the cytosol end and attached at the membrane by cysteine. (c) G P I anchor showing a red hexagon and two green hexagon and a blue square attached in the exterior to H subscript 3 N. The red Hexagon is attached to the exterior of the membrane. Some cell-surface proteins, and some specialized proteins with distinctive covalently attached polysaccharides called proteoglycans (see Chapter 20), are bound to the exoplasmic face of the plasma membrane by a third anchor group, glycosylphosphatidylinositol (GPI) anchors. The exact structures of GPI anchors vary greatly among different cell types, but they always contain phosphatidylinositol, whose two fatty acyl chains extend into the lipid bilayer just like those of typical membrane phospholipids; phosphoethanolamine, which covalently links the anchor to the C-terminus of a protein; and several sugar residues (Figure 10-19c). Therefore, GPI anchors are glycolipids. The GPI anchor is both necessary and sufficient for binding proteins to the membrane. For instance, treatment of cells with phospholipase C, which cleaves the phosphate-glycerol bond in phospholipids and in GPI anchors (see Figure 10-12), releases GPI-
All Transmembrane Proteins and Glycolipids Are Asymmetrically Oriented in the Bilayer
anchored proteins such as Thy-1 and placental alkaline phosphatase (PLAP) from the cell surface. All Transmembrane Proteins and Glycolipids Are Asymmetrically Oriented in the Bilayer Every type of transmembrane protein has a specific orientation, known as its topology, with respect to the membrane faces. Its cytosolic segments always face the cytosol, and its exoplasmic segments always face the opposite side of the membrane. This asymmetry in protein orientation gives the two membrane faces their different properties. The orientations of different types of transmembrane proteins are established during their synthesis, as we will see in Chapter 13. Membrane proteins have never been observed to flip-flop across a membrane; this would require a transient movement of hydrophilic amino acid residues through the hydrophobic interior of the membrane, which would be energetically unfavorable. Accordingly, the asymmetric topology of a transmembrane protein is maintained throughout the protein’s lifetime. As Figure 10-6 shows, membrane proteins retain their asymmetric orientation during membrane budding and fusion events; the same segment always faces the cytosol and the same segment is always exposed to the exoplasmic face. In multipass membrane proteins, the orientation of individual transmembrane segments can be affected by changes in the membrane’s phospholipid composition.
Many transmembrane proteins contain carbohydrate chains covalently linked to serine, threonine, or asparagine side chains of the polypeptide. Such transmembrane glycoproteins are always oriented so that all the carbohydrate chains are in the exoplasmic domain (see Figure 10-14 for the example of glycophorin A). Likewise, glycolipids, in which a carbohydrate chain is attached to the glycerol or sphingosine backbone of a membrane lipid, are always located in the exoplasmic leaflet, with the carbohydrate chain protruding from the membrane surface. The biosynthetic basis for the asymmetric glycosylation of proteins is described in Chapters 13 and 14. Both glycoproteins and glycolipids are especially abundant in the plasma membranes of eukaryotic cells and in the membranes of the intracellular compartments that establish the secretory and endocytic pathways; they are absent from the inner mitochondrial membrane, chloroplast lamellae, and several other intracellular membranes. Because the carbohydrate chains of glycoproteins and glycolipids in the plasma membrane extend into the extracellular space, they are available to interact with components of the extracellular matrix as well as with lectins (proteins that bind specific sugars), growth factors, and antibodies. One important consequence of interactions involving membrane glycoproteins and glycolipids is illustrated by the ABO blood-group antigens. These three structurally related oligosaccharide components of certain glycoproteins and glycolipids are expressed on the surfaces of human red blood cells and many other cell types (Figure 10-20). All humans have the enzymes for synthesizing O antigen. Persons with type A
blood also have a glycosyltransferase enzyme that adds an extra modified monosaccharide called N-acetylgalactosamine to O antigen to form A antigen. Those with type B blood have a different transferase that adds an extra galactose to O antigen to form B antigen. People with both transferases produce both A and B antigen (AB blood type); those who lack these transferases produce only O antigen (O blood type).
FIGURE 10-20 Human ABO blood-group antigens. These antigens are oligosaccharide chains covalently attached to glycolipids or glycoproteins in the plasma membrane. The terminal oligosaccharide sugars distinguish the three antigens. The presence or absence of
the glycosyltransferases that add galactose (Gal) or N-acetylgalactosamine (GalNAc) to O antigen determine a person’s blood type. Description The illustration uses hexagons to show chains attached to lipids or proteins to form blood antigens. The top one is labeled A antigen. The lipid or protein is at left, with hexagons labeled G l c, G a l, G l c N A c, G a l, F u c, G a l N A c. The second diagram is labeled O antigen. It shows the lipid or protein at left, followed by hexagons labeled G l c, G a l, G l c N A c, G a l, F u c. The last diagram is labeled B antigen. It starts with the lipid or protein, then hexagons for G l c, G a l, G l c N A c, G a l, F u c, G a l. A key to the abbreviations is at the bottom. The key reads as follows: G l cGlucose; Gal- Galactose; G l c N a C-N-Acetylglucosamine; G a l N A c - N-Acetylgalactosamine; F u c- Fucose. People whose erythrocytes lack the A antigen, the B antigen, or both on their surface normally have antibodies against the missing antigen(s) in their serum. Thus if a type A or O person receives a transfusion of type B blood, antibodies against the B antigen will bind to the introduced red cells and trigger their destruction. To prevent such harmful reactions, blood-group typing and appropriate matching of blood donors and recipients are required in all transfusions (Table 10-2).
TABLE 10-2 • ABO Blood Groups Blood Group Antigens on RBCs Serum Antibodies Can Receive Blood Types A A Anti-B A and O B B Anti-A B and O AB A and B None All i
Lipid-Binding Motifs Help Target Peripheral Proteins to the Membrane
O O Anti-A and anti-B O See Figure 10-20 for antigen structures. Lipid-Binding Motifs Help Target Peripheral Proteins to the Membrane Many water-soluble enzymes whose substrates are phospholipids must bind to membrane surfaces. Phospholipases, for example, hydrolyze various bonds in the head groups of phospholipids (see Figure 10-12) and thereby play a variety of roles in cells — helping to degrade damaged or aged cellular membranes, generating precursors to signaling molecules, and even serving as the active components in many snake venoms. Many peripheral proteins, including phospholipases, initially bind to the polar head groups of membrane phospholipids to carry out their catalytic functions. The mechanism of action of phospholipase illustrates how such enzymes can reversibly interact with membranes and catalyze reactions at the interface of an aqueous solution and a lipid surface. When this enzyme is in aqueous solution, its -containing active site is buried in a channel lined with hydrophobic amino acids. The enzyme binds with greatest affinity to bilayers composed of negatively charged phospholipids (e.g., phosphatidylserine). This observation suggests that the rim of positively charged lysine and arginine residues around the entrance to the catalytic channel is particularly important in binding (Figure 10-21a) and constitutes a lipid binding motif. Binding induces a conformational change in phospholipase that strengthens its binding to i
the phospholipid heads and opens the hydrophobic channel. As a phospholipid molecule moves from the bilayer into the channel, the enzyme-bound binds to the phosphate in the head group, thereby positioning the ester bond to be cleaved in the catalytic site (Figure 1021b), releasing the acyl chain.
FIGURE 10-21 Lipid-binding surface and mechanism of action of phospholipase . (a) A structural model of the enzyme, showing the surface that interacts with a membrane.
Proteins Can Be Removed from Membranes by Detergents or High-Salt Solutions
This lipid-binding surface contains a rim of positively charged arginine and lysine residues (blue), surrounding the cavity of the catalytic active site, in which a substrate lipid (red balland-stick structure) is bound. (b) Diagram of catalysis by phospholipase . When docked on a lipid membrane, positively charged residues of the binding site bind to negatively charged polar groups at the membrane surface. This binding triggers a small conformational change, opening a channel lined with hydrophobic amino acids that leads from the bilayer to the catalytic site. As a phospholipid moves into the channel, an enzyme-bound ion (green) binds to the head group, positioning the ester bond to be cleaved (red) next to the catalytic site. See D. Blow, 1991, Nature 351:444; and M. H. Gelb, W. Cho, and D. C. Wilton, 1999, Curr. Opin. Struct. Biol. 9:428. [Part (a) Data from D. L. Scott et al., 1990, Science 250:1563, PDB ID 1poc.] Description A 3-D model shows phospholipase A. The active site is indicated and colored in red. A schematic shows that, on docking with the membrane, lipids enter the active site. A zoomed out view of the active site shows a chemical diagram which depicts the interaction of phosphate and carboxylates in the polar head group with a calcium cation of the active site. Proteins Can Be Removed from Membranes by Detergents or HighSalt Solutions Membrane proteins are often difficult to purify and study, mostly because of their tight association with membrane lipids and other membrane proteins. Detergents, which are amphipathic molecules that disrupt membranes by intercalating into phospholipid bilayers, can be used to solubilize lipids and many membrane proteins. The hydrophobic part of a
detergent molecule is attracted to the phospholipid hydrocarbons and mingles with them readily; the hydrophilic part is strongly attracted to water. Some detergents, such as bile salts, are natural products, but most are synthetic molecules developed for cleaning and for dispersing mixtures of oil and water in the food industry (e.g., to keep salad dressings from separating). Ionic detergents, such as sodium deoxycholate (a bile salt) and sodium dodecylsulfate (SDS), contain a charged group; non-ionic detergents, such as Triton X-100 and octylglucoside, lack a charged group (Figure 10-22). At very low concentrations, detergents dissolve in pure water as isolated molecules. As the concentration increases, the molecules begin to form micelles — small spherical aggregates in which the hydrophilic parts of the molecules face outward and the hydrophobic parts cluster in the center (see Figure 10-3c). The critical micelle concentration (CMC) at which micelles form is characteristic of each detergent and is a function of the structures of its hydrophobic and hydrophilic parts.
FIGURE 10-22 Structures of four common detergents. The hydrophobic part of each molecule is shown in yellow; the hydrophilic part, in blue. The bile salt sodium deoxycholate is a natural product; the others are synthetic. Although ionic detergents commonly cause denaturation of proteins, non-ionic detergents do not and are thus useful in solubilizing integral membrane proteins. Description In each chemical structure, the hydrophobic part is highlighted in yellow and the hydrophilic portion is highlighted in blue. The ionice detergents are Sodium Deoxycholate, Sodium Dodecylsulfate (S D S). The non-ionic detergents are Triton X100 (polyoxyethylene(9.5) p-t-octylphenol) and Octylglucoside (octyl-beta-D- glucopyranoside) Ionic and non-ionic detergents interact differently with proteins and have different uses in the lab. Ionic detergents bind to the exposed hydrophobic regions of membrane proteins as well as to the hydrophobic cores of water-soluble proteins. Because of their charge, these detergents can disrupt ionic and hydrogen bonds. At high concentrations, sodium dodecylsulfate, for example, completely denatures proteins by binding to every side chain, a property that is exploited in SDS gel electrophoresis (see Figure 3-41). Non-ionic detergents generally do not denature proteins and are thus useful in extracting proteins in their folded and active form from membranes before the proteins are purified. Protein-protein interactions, especially the weaker ones, can be sensitive to both ionic and non-ionic detergents. At high concentrations (above the CMC), non-ionic detergents solubilize biological membranes by forming mixed micelles of detergent,
phospholipids, and integral membrane proteins — bulky hydrophobic structures that do not dissolve in water (Figure 10-23, top). At low concentrations (below the CMC), these detergents bind to the hydrophobic regions of most integral membrane proteins, but without forming micelles, allowing the proteins to remain soluble (Figure 10-23, bottom). Creating such an aqueous solution of integral membrane proteins is a necessary first step in protein purification.
FIGURE 10-23 Solubilization of integral membrane proteins by non-ionic detergents. At a concentration higher than its critical micelle concentration (CMC), a detergent solubilizes lipids and integral membrane proteins, forming mixed micelles containing detergent, protein, and lipid molecules. At concentrations below the CMC, non-ionic detergents (e.g., octylglucoside, Triton X-100) can dissolve membrane proteins without forming micelles by coating the membrane-spanning regions. Description
A membrane is depicted with embedded proteins. A detergent, represented by a blue head and tail figure, is added at two different concentrations, above the critical micelle concentration and below the critical micelle concentration. Above the C M C, micelles form around the proteins and are dissolved in the solvent. Below the C M C, the detergent molecules do not form micelles, but coat the proteins allowing their dissolution in the solvent. Treatment of cultured cells with a buffered salt solution containing a nonionic detergent such as Triton X-100 extracts water-soluble proteins as well as integral membrane proteins from cellular membranes. As noted earlier, the exoplasmic and cytosolic domains of integral membrane proteins are generally hydrophilic and soluble in water. The membranespanning domains, however, are rich in hydrophobic and uncharged residues (see Figure 10-14). When separated from membranes, these exposed hydrophobic segments tend to interact with one another, causing the protein molecules to aggregate and precipitate from aqueous solutions. The hydrophobic parts of non-ionic detergent molecules preferentially bind to the hydrophobic segments of transmembrane proteins, preventing protein aggregation and allowing the proteins to remain in aqueous solution. Detergent-solubilized transmembrane proteins can then be purified by affinity chromatography and other techniques used in purifying water-soluble proteins (see Chapter 3). As discussed previously, most peripheral membrane proteins are bound to specific transmembrane proteins or membrane phospholipids by ionic or other weak noncovalent interactions. Generally peripheral proteins can be removed from the membrane by solutions of high ionic strength (high salt concentrations), which disrupt ionic bonds, or by chemicals that bind
divalent cations such as . Unlike integral membrane proteins, most peripheral proteins are soluble in water and do not need to be solubilized by non-ionic detergents. The localization of membrane proteins to the phospholipid bilayer is not necessarily permanent. Phosphatidylinositide-specific phospholipase C hydrolyzes the membrane anchor of proteins attached to the membrane via this hydrophobic tail. Similarly, proteases can “shave off” the extracellular portions of single-pass membrane proteins. This can be done experimentally, for example, by exposing cells or membranes derived from them to a protease such as papain, and often allows recovery of the released extracellular portion for structural and functional studies. These and similar methods are commonly used to detach cultured mammalian cells from the solid supports on which they are often grown or to detach cells from tissues to generate single cell suspensions. However, many single-pass membrane proteins are natural substrates for sheddases: members of the metalloprotease family of proteases that are discussed in
Chapter 16. The extracellular domains released in this fashion may be found in the circulation and exert biological functions distinct from their membrane-anchored equivalents. KEY CONCEPTS OF SECTION 10.2 Membrane Proteins: Structure and Basic Functions Biological membranes usually contain integral (transmembrane) proteins as well as lipid-anchored proteins and peripheral membrane proteins, which do not enter the hydrophobic core of the bilayer (see Figure 10-1).
Most integral membrane proteins contain one or more membrane-spanning hydrophobic α helices, which are bracketed by hydrophilic domains that extend into the aqueous environment surrounding the cytosolic and exoplasmic faces of the membrane (see Figures 10-14, 10-15, and 10-17). Fatty acyl side chains as well as the polar head groups of membrane lipids pack tightly and irregularly around the hydrophobic segments of integral membrane proteins (see Figure 10-16). The porins, unlike other integral membrane proteins, contain membrane-spanning β sheets that form a barrel-like channel through the bilayer (see Figure 10-18). Lipids attached to certain amino acids anchor some proteins to one or the other membrane leaflet (see Figure 10-19). All transmembrane proteins and glycolipids are asymmetrically oriented in the bilayer. Invariably, carbohydrate chains are present only on the exoplasmic surface of a glycoprotein or glycolipid. Many water-soluble enzymes (e.g., phospholipases) whose substrates are phospholipids must bind to the membrane surface to carry out their function. Such binding is often due to the attraction between positive charges on lysine and arginine residues in the protein and negative charges on phospholipid head groups in the bilayer. Transmembrane proteins can be selectively extracted from membranes with the use of non-ionic detergents.
10.3 Phospholipids, Sphingolipids, and Cholesterol: Synthesis and Intracellular Movement
10.3 Phospholipids, Sphingolipids, and Cholesterol: Synthesis and Intracellular Movement In this section, we consider some of the special challenges that a cell faces in synthesizing and transporting lipids, which are poorly soluble in the aqueous interior of a cell. The focus will be the biosynthesis and movement of the major lipids found in cellular membranes — phospholipids, sphingolipids, and cholesterol — and their precursors. In lipid biosynthesis, water-soluble precursors are assembled into membraneassociated intermediates that are then converted into membrane lipid products. The movement of these lipids, especially membrane components, between different organelles is critical for maintaining the proper composition and properties of membranes and overall cell structure. A fundamental principle of membrane biosynthesis is that cells synthesize new membranes only by the expansion of existing membranes. [The one exception may be autophagy, in which new membrane is formed first through the formation of an autophagic crescent, the construction of which involves modification of phosphatidylethanolamine with the ubiquitin-like modifier Atg8 (see Figure 14-34).] Although some early steps in the synthesis of membrane lipids take place in the cytosol, the final steps are catalyzed by enzymes bound to preexisting cellular membranes, and the
Fatty Acids Are Assembled from Two-Carbon Building Blocks by Several Important Enzymes
products are incorporated into the membranes as they are generated. Evidence for this process is seen when cells are briefly exposed to radioactively labeled precursors (e.g., phosphates or fatty acids): all the phospholipids and sphingolipids incorporating these labeled precursors are associated with intracellular membranes; as expected from the hydrophobicity of the fatty acyl chains, none are found free in the cytosol. After they are formed, membrane lipids must be distributed appropriately both between the leaflets of a given membrane and among the independent membranes of different organelles in eukaryotic cells, as well as the plasma membrane. Here we consider how this precise lipid distribution is accomplished; in Chapters 13 and 14, we discuss how membrane proteins are inserted into cellular membranes and trafficked to their appropriate location within the cell. Fatty Acids Are Assembled from TwoCarbon Building Blocks by Several Important Enzymes Fatty acids (see Chapter 2) play a number of important roles in cells. In addition to being a cellular fuel source (see the discussion of aerobic oxidation in Chapter 12), fatty acids are key components of both the phospholipids and the sphingolipids that make up cellular membranes; they also anchor some proteins to cellular membranes (see Figure 10-19). Thus the regulation of fatty acid synthesis plays a key role in the regulation of membrane synthesis as a whole. The major fatty acids in
phospholipids contain 14, 16, 18, or 20 carbon atoms and include both saturated and unsaturated chains. The fatty acyl chains found on sphingolipids can be longer than those in the phosphoglycerides, containing up to 26 carbon atoms, and may bear other chemical modifications (e.g., hydroxylation) as well. Fatty acids are synthesized from the two-carbon building block acetate ( ). In cells, both acetate and the intermediates in fatty acid biosynthesis are esterified to a large water-soluble molecule called coenzyme A (CoA), as exemplified by the structure of acetyl CoA shown here. Acetyl CoA is an important intermediate in the metabolism of glucose, fatty acids, and many amino acids, as detailed in Chapter 12. It also contributes acetyl groups in many biosynthetic pathways. Saturated fatty acids (with no carbon-carbon double bonds) containing 14 or 16 carbon atoms are made from acetyl CoA by two enzymes, acetyl-CoA carboxylase and fatty acid synthase. In animal cells, these enzymes are found in the cytosol; in plants, they are found in chloroplasts. Palmitoyl CoA (a 16carbon fatty acyl group linked to CoA) can be elongated to 18–24 carbons by the sequential addition of two-carbon units in the endoplasmic reticulum (ER) or sometimes in the mitochondrion. Desaturase enzymes, also located in the ER, introduce double bonds at specific positions in
Small Cytosolic Proteins Facilitate Movement of Fatty Acids
some fatty acids, yielding unsaturated fatty acids. Oleyl CoA (oleate linked to CoA; see Table 2-4), for example, is formed by removal of two H atoms from stearyl CoA. In contrast to free fatty acids, fatty acyl CoA derivatives are soluble in aqueous solutions because of the hydrophilicity of the CoA segment. Small Cytosolic Proteins Facilitate Movement of Fatty Acids In order to be transported through the cell cytosol, free, or unesterified, fatty acids (those unlinked to CoA) are commonly bound by fatty acid– binding proteins (FABPs), which belong to a group of small cytosolic proteins that act as chaperones to facilitate the intracellular movement of many lipids. These proteins contain a hydrophobic pocket lined by β sheets (Figure 10-24). A long-chain fatty acid can fit into this pocket and interact noncovalently with the surrounding protein.
FIGURE 10-24 Binding of a fatty acid to the hydrophobic pocket of a fatty acid–binding protein (FABP). The crystal structure of adipocyte FABP (ribbon diagram) shows that the hydrophobic binding pocket is generated from two β sheets that are nearly at right angles to each other, forming a clam shell–like structure. A fatty acid (carbons yellow; oxygens red) interacts noncovalently with hydrophobic amino acid residues within this pocket. [Data from Z. Xu, D. A. Bernlohr, and L. J. Banaszak, 1993, J. Biol. Chem. 268:7874, PDB ID 1lid.] The expression of cellular FABPs is regulated coordinately with cellular requirements for the uptake and release of fatty acids. Thus FABP levels are high in active muscles that are using fatty acids for generation of ATP,
Fatty Acids Are Incorporated into Phospholipids Primarily on the ER Membrane
and in adipocytes (fat-storing cells) when they are either taking up fatty acids to be stored as triglycerides or releasing fatty acids for use by other cells. The importance of FABPs in fatty acid metabolism is highlighted by the observations that they can compose as much as 5 percent of all cytosolic proteins in the liver and that genetic inactivation of cardiac muscle FABP converts the heart from a muscle that primarily burns fatty acids for energy into one that primarily burns glucose. Fatty Acids Are Incorporated into Phospholipids Primarily on the ER Membrane Fatty acids are not directly incorporated into phospholipids; rather, in eukaryotic cells, they are first converted into CoA esters. The subsequent synthesis of phospholipids such as the phosphoglycerides is carried out by enzymes associated with the cytosolic face of the ER membrane, usually the smooth ER, in animal cells; through a series of steps, fatty acyl CoAs, glycerol 3-phosphate, and polar head group precursors are linked together and then inserted into the ER membrane (Figure 10-25). The fact that the enzymes involved in this process are located on the cytosolic side of the membrane means that there is an inherent asymmetry in membrane biogenesis: new membranes are initially synthesized only on one leaflet — a fact with important consequences for the asymmetric distribution of lipids in membrane leaflets. Once synthesized on the ER, phospholipids are transported to other organelles and to the plasma membrane.
Mitochondria synthesize some of their own membrane lipids and import others.
FIGURE 10-25 Phospholipid synthesis in the ER membrane. Because phospholipids are amphipathic molecules, the last steps of their multistep synthesis take place at the interface between a membrane and the cytosol and are catalyzed by membrane-associated enzymes. Step 1 : Two fatty acids from fatty acyl CoA are esterified to the phosphorylated glycerol backbone, forming phosphatidic acid, whose two long hydrocarbon chains anchor the molecule to the membrane. Step 2 : A phosphatase converts phosphatidic acid into diacylglycerol. Step 3 : A polar head group (e.g., phosphorylcholine) is transferred from cytosine diphosphocholine (CDP-choline) to the exposed hydroxyl group. Step 4 : Flippase proteins catalyze the movement of phospholipids from the cytosolic leaflet in which they are initially formed to the exoplasmic leaflet. Description A schematic shows an E R membrane with cytosol labeled at the top, and Lumen at the bottom of the membrane. The chemical diagram for Acetyl CoA is shown at left above the membrane. At number 1, a G P A T and A L P A A T are shown in the membrane with a phosphatidic acid next to it. At number 2, a phosphatase, as a blue circle is shown at the top of the membrane. At number 3 a choline phosphotransferase structure
Flippases Move Phospholipids from One Membrane Leaflet to the Opposite Leaflet
is in the membrane. At number 4, the choline has gone into the E R and there is a blue structure with the label flippase. Sphingolipids are also synthesized indirectly from multiple precursors. Sphingosine, the building block of these lipids, is made in the ER, beginning with the coupling of a palmitoyl group from palmitoyl CoA to serine; the subsequent addition of a second fatty acyl group to form N-acyl sphingosine (ceramide) also takes place in the ER. Later, in the Golgi, a polar head group is added to ceramide, yielding sphingomyelin, whose head group is phosphorylcholine, and various glycosphingolipids, in which the head group may be a monosaccharide or a more complex oligosaccharide (see Figure 10-8b). Some sphingolipid synthesis can also take place in mitochondria. In addition to serving as the backbone for sphingolipids, ceramide and its metabolic products are important signaling molecules that can influence cell growth, proliferation, endocytosis, resistance to stress, and programmed cell death (apoptosis). After their synthesis is completed in the Golgi, sphingolipids are transported to other cellular compartments through vesicle-mediated mechanisms similar to those for the transport of proteins, discussed in
Chapter 14. Any type of vesicular transport results in movement not only of the protein payload but also of the lipids that compose the vesicular membrane. Phospholipids such as phosphoglycerides, as well as cholesterol, can move between organelles by additional mechanisms, described below.
Cholesterol Is Synthesized by Enzymes in the Cytosol and ER Membrane
Flippases Move Phospholipids from One Membrane Leaflet to the Opposite Leaflet Even though phospholipids are initially incorporated into the cytosolic leaflet of the ER membrane, various phospholipids are asymmetrically distributed in the two leaflets of the ER membrane and of other cellular membranes. As noted above, phospholipids do not readily flip-flop from one leaflet to the other. For the ER membrane to expand by growth of both leaflets and to have asymmetrically distributed phospholipids, its phospholipid components must be able to move from one leaflet to the other. Although the mechanisms employed to generate and maintain membrane phospholipid asymmetry are not well understood, it is clear that flippases play a key role. As will be described in Chapter 11, these integral membrane proteins use the energy of ATP hydrolysis to facilitate the movement of phospholipid molecules from one leaflet to the other. When cells die, this lipid asymmetry is lost and results in increased surface exposure of those phospholipids mostly confined to the cytoplasmic leaflet, such as phosphatidylserine. Cholesterol Is Synthesized by Enzymes in the Cytosol and ER Membrane
Next we focus on cholesterol, the principal sterol in animal cells. Cholesterol is synthesized mainly in the liver. The first steps of cholesterol synthesis (Figure 10-26) — conversion of three acetyl groups linked to CoA (acetyl CoA) forming the six-carbon molecule β-hydroxy-β- methylglutaryl linked to CoA (HMG-CoA) — take place in the cytosol. The conversion of HMG-CoA into mevalonate, the key rate-controlling step in cholesterol biosynthesis, is catalyzed by HMG-CoA reductase, an ER integral membrane protein, even though both its substrate and its product are water soluble. The water-soluble catalytic domain of HMGCoA reductase extends into the cytosol, but its eight transmembrane α helices firmly embed the enzyme in the ER membrane. Five of the transmembrane α helices compose the so-called sterol-sensing domain and regulate enzyme stability. When levels of cholesterol in the ER membrane are high, binding of cholesterol to this domain causes the enzyme to bind to two other integral ER membrane proteins, Insig-1 and Insig-2. This binding, in turn, induces ubiquitylation (see Figure 3-32) of HMG-CoA reductase and its degradation by the proteasome pathway, reducing the production of mevalonate, the key intermediate in cholesterol biosynthesis.
FIGURE 10-26 Cholesterol biosynthetic pathway. The regulated rate-controlling step in cholesterol biosynthesis is the conversion of β-hydroxy-β-methylglutaryl CoA (HMG-CoA) into mevalonate by HMG-CoA reductase, an ER-membrane protein. Mevalonate is then converted into isopentenyl pyrophosphate (IPP), which has the basic five-carbon isoprenoid structure. IPP can be converted into cholesterol and many other lipids, often through the polyisoprenoid intermediates shown. Some of the numerous compounds derived from isoprenoid intermediates and cholesterol itself are indicated. Description The steps read as follows. 1. acetyl coenzyme A plus acetoacetyl coenzyme a yield 2. HMG-coenzyme A, which is reduced by HMG-Co-A reductase to 3. mevalonate.
4.Isopentenyl pyrophosphate (I P P) is shown with two choice arrows, isopentenyl adenosine and many other isoprenoids, 5. Farnesyl pyrophosphate is shown with 6 choices: dolichol, heme, ubiquinone, vitamins A,E, K, chlorophyll, Lipid anchored proteins (Ras) 6. squalene, and lastly cholesterol with 5 choices: vitamin D, bile acids, steroid hormones, cholesterol esters, modified proteins (hedgehog). Atherosclerosis, frequently called cholesterol-dependent clogging of the arteries, is characterized by the progressive deposition of cholesterol and other lipids, cells, and extracellular matrix material in the inner layer of the wall of an artery. The resulting distortion of the artery’s wall can lead, either alone or in combination with a blood clot, to major blockage of blood flow. Atherosclerosis accounts for 75 percent of deaths due to cardiovascular disease in the United States. Perhaps the most successful anti-atherosclerosis medications are the statins. These drugs bind to HMG-CoA reductase and directly inhibit its activity, thereby lowering cholesterol biosynthesis. As a consequence, the concentration of low-density lipoproteins (see Figure 14-27) — the small, membrane-enveloped particles containing cholesterol esterified to fatty acids that often and rightly are called “bad cholesterol” — drops in the blood, reducing the formation of atherosclerotic plaques. Mevalonate, the six-carbon product formed by HMG-CoA reductase, is converted in several steps into the five- carbon isoprenoid compound isopentenyl pyrophosphate (IPP) and its stereoisomer, dimethylallyl pyrophosphate (DMPP) (see Figure 10-26). These reactions are catalyzed by cytosolic enzymes, as are the subsequent reactions in the cholesterol
Cholesterol and Phospholipids Are Transported Between Organelles by Several Mechanisms
synthesis pathway, in which six IPP units condense to yield squalene, a branched-chain 30-carbon intermediate. Enzymes bound to the ER membrane catalyze the multiple reactions that convert squalene into cholesterol in mammals or into related sterols in other species. One of the intermediates in this pathway, farnesyl pyrophosphate, is the precursor of the prenyl lipid that anchors Ras and related proteins to the cytosolic surface of the plasma membrane (see Figure 10-19) as well as other important biomolecules (see Figure 10-26). Cholesterol and Phospholipids Are Transported Between Organelles by Several Mechanisms As already noted, the final steps in the synthesis of cholesterol and phospholipids take place primarily in the ER. Thus the plasma membrane and the membranes bounding other organelles must obtain these lipids by means of one or more intracellular transport processes. Membrane lipids can and do accompany both soluble and membrane proteins along the secretory pathway described in Chapter 14: membrane vesicles bud from the ER and fuse with membranes in the Golgi complex, and other membrane vesicles bud from the Golgi complex and fuse with the plasma membrane (Figure 10-27a). However, evidence suggests that there is substantial inter-organelle movement of cholesterol and phospholipids through other mechanisms. For example, chemical inhibitors of the classic secretory pathway and mutations that impede vesicular traffic along this
pathway do not prevent cholesterol or phospholipid transport between membranes.
FIGURE 10-27 Proposed mechanisms of transport of cholesterol and phospholipids between membranes. In mechanism (a), vesicles transfer lipids between membranes. In mechanism (b), lipid transfer is a consequence of direct contact between membranes that is mediated by membrane-embedded proteins. In mechanism (c), transfer is mediated by small, soluble lipid-transfer proteins. [Information from F. R. Maxfield and D. Wustner, 2002, J. Clin. Invest. 110:891.] Description (a) In the vesicle mechanism, vesicles containing cholesterol are budded from the membrane, travel across the cytosol, and fuse with the destination membrane. In lipid transfer, membranes are brought close by hypothetical proteins, allowing exchange of cholesterol. (c) In the binding protein model, cholesterol binds to proteins and is transferred through the cytoplasm to the destination. A second proposed mechanism of lipid movement entails direct proteinmediated contact of ER or ER-derived membranes with the membranes of other organelles (Figure 10-27b). In a third proposed mechanism, small lipid-transfer proteins facilitate the exchange of phospholipids or cholesterol between different membranes (Figure 10-27c). Although such
lipid-transfer proteins have been identified in assays in vitro, their role in the intracellular movements of most phospholipids is not well defined. For instance, mice with a knockout mutation in the gene encoding the phosphatidylcholine-transfer protein appear to be normal in most respects, indicating that this protein is not essential for cellular phospholipid metabolism. As noted earlier, the lipid compositions of different organelle membranes vary considerably (see Table 10-1). Some of these differences are due to different sites of synthesis. For example, a phospholipid called cardiolipin, which is localized to the mitochondrial membrane, is made only in mitochondria, and little of it is transferred to other organelles. Differential transport of lipids also plays a role in determining the lipid compositions of different cellular membranes. For instance, even though cholesterol is made in the ER, the cholesterol concentration (cholesterolto-phospholipid molar ratio) is 1.5–13-fold higher in the plasma membrane than in other organelles (ER, Golgi, mitochondrion, lysosome). Although the mechanisms responsible for establishing and maintaining these differences are not well understood, we have seen that the distinctive lipid composition of each membrane has a major influence on its physical and biological properties. KEY CONCEPTS OF SECTION 10.3 Phospholipids, Sphingolipids, and Cholesterol: Synthesis and Intracellular Movement Saturated and unsaturated fatty acids of various chain lengths are components of phospholipids and sphingolipids.
Fatty acids are synthesized from acetyl CoA by water-soluble enzymes and modified by elongation and desaturation in the endoplasmic reticulum (ER). Free fatty acids are transported within cells by fatty acid–binding proteins (FABPs). Fatty acids are incorporated into phospholipids through a multistep process. The final steps in the synthesis of phosphoglycerides and sphingolipids are catalyzed by membrane-associated enzymes primarily on the cytosolic face of the ER (see Figure 10-25). Each type of newly synthesized lipid is incorporated into the preexisting membranes on which it is made; thus membranes are themselves the platform for the synthesis of new membrane material. Most membrane phospholipids are preferentially distributed in either the exoplasmic or the cytosolic leaflet. This asymmetry results in part from the action of flippases, which flip phospholipids from one leaflet to the other. The initial steps in cholesterol biosynthesis take place in the cytosol, whereas the last steps are catalyzed by enzymes associated with the ER membrane. The rate-controlling step in cholesterol biosynthesis is catalyzed by HMG-CoA reductase, whose transmembrane segments are embedded in the ER membrane and contain a sterol-sensing domain. Considerable evidence indicates that vesicular transport, direct protein-mediated contacts between different membranes, soluble lipid-transfer proteins, or all three may account for some inter-organelle transport of cholesterol and phospholipids (see
Key Terms
End of Chapter Visit Achieve to access study tools and to learn more about the content in this chapter. Perspectives for the Future Analyze the Data Chapter References Additional study tools, including videos, animations, and quizzes Key Terms acetyl CoA amphipathic aquaporin cholesterol cilium cytoskeleton cytosolic face exoplasmic face flagellum glycolipid glycoprotein hydrophilic hydrophobic integral membrane protein
Review the Concepts
lectin lipid-anchored membrane protein lipid raft liposome membrane transport protein micelle peripheral membrane protein phosphoglyceride phospholipase phospholipid bilayer plasma membrane porin receptor protein saturated sphingolipid unsaturated Review the Concepts 1. When viewed by electron microscopy, the lipid bilayer is often described as looking like a railroad track. Explain how the structure of the bilayer creates this image. 2. Explain the following statement: The structure of all biomembranes depends on the chemical properties of phospholipids, whereas the function of each specific biomembrane depends on the specific proteins associated with that membrane.
3. Biomembranes contain many different types of lipid molecules. What are the three main types of lipid molecules found in biomembranes? How are the three types similar, and how are they different? 4. Lipid bilayers are said to behave like two-dimensional fluids. What does this mean? What drives the movement of lipid molecules and proteins within the bilayer? How can such movement be measured? What factors affect the degree of membrane fluidity? 5. Why are water-soluble substances unable to freely cross the lipid bilayer of the plasma membrane? How does the cell overcome this permeability barrier? 6. Name the three groups into which membrane-associated proteins may be classified. Explain the mechanism by which each group associates with a biomembrane. 7. Identify the following membrane-associated proteins based on their structure: (a) tetramers of identical subunits, each with six membrane-spanning α helices; (b) trimers of identical subunits, each with 16 β sheets forming a barrel-like structure. 8. Proteins may be bound to the exoplasmic or cytosolic face of the plasma membrane by covalently attached lipids. What are the three types of lipid anchors responsible for tethering proteins to the plasma-membrane bilayer? Which type is used by cellsurface proteins that face the external medium? By glycosylated proteoglycans? 9. Although both faces of a biomembrane are composed of the same general types of macromolecules, principally lipids and
proteins, the two faces of the bilayer are not identical. What accounts for the asymmetry between the two faces? 10. What are detergents? How do ionic and non-ionic detergents differ in their ability to disrupt biomembrane structure? 11. What is the likely identity of these membrane-associated proteins: (a) a protein that is released from a membrane treated with a high-salt solution, which causes disruption of ionic linkages; (b) a protein that is not released from the membrane upon its exposure to a high-salt solution alone, but is released when the membrane is incubated with an enzyme that cleaves phosphate-glycerol bonds and covalent linkages are disrupted; (c) a protein that is not released from the membrane upon exposure to a high-salt solution, but is released after the addition of the detergent sodium dodecylsulfate (SDS). Will the activity of the protein released in part (c) be preserved following its release? 12. Following the production of membrane extracts using the nonionic detergent Triton X-100, you analyze the membrane lysates via mass spectrometry and note a high content of cholesterol and sphingolipids. Furthermore, biochemical analysis of the lysates shows potential kinase activity. What have you probably isolated? 13. Phospholipid biosynthesis at the interface between the endoplasmic reticulum (ER) and the cytosol presents a number of challenges that must be solved by the cell. Explain how each of the following is handled. a. The substrates for phospholipid biosynthesis are all water soluble, yet the end products are not.
b. The immediate site of incorporation of all newly synthesized phospholipids is the cytosolic leaflet of the ER membrane, yet phospholipids must be incorporated into both leaflets. c. Many membrane systems in the cell, such as the plasma membrane, are unable to synthesize their own phospholipids, yet these membranes must also expand if the cell is to grow and divide. 14. What are the common fatty acid chains in phosphoglycerides, and why do these fatty acid chains differ in their number of carbon atoms by multiples of 2? 15. Fatty acids must associate with lipid chaperones in order to move within the cell. Why are these chaperones needed, and what is the name given to a group of proteins that are responsible for this intracellular trafficking of fatty acids? What is the key distinguishing feature of these proteins that allows fatty acids to move within the cell? 16. What is the key enzyme in the highly regulated process of cholesterol biosynthesis? This enzyme is subject to feedback inhibition. What is feedback inhibition? How does this enzyme sense cholesterol levels in a cell? 17. Phospholipids and cholesterol must be transported from their site of synthesis to various membrane systems within cells. One way of doing this is through vesicular transport, as is the case for many proteins in the classic secretory pathway (see Chapter 14). However, phospholipid and cholesterol membrane-to-membrane transport in cells does not occur solely by vesicular transport.
What is the evidence for this? What appear to be the major mechanisms for phospholipid and cholesterol transport? 18. Explain the mechanism by which statins lower “bad” cholesterol.