11 Population Distributions
11Population Distributions A male collared lizard. Males of this species have a bright orange throat that intimidates other males and attracts females. Bringing Back the Mountain Boomer When Alan Templeton was a Boy Scout in 1960, he encountered his first collared lizards (Crotaphytus collaris), also known as mountain boomers, in the Ozark Mountains of Missouri. He was struck by the brightly colored males that ran around in the forest openings that dotted the mountains. Two decades later, as a biology professor at Washington University in St. Louis, he returned to the Ozark Mountains and was shocked to find that most of the lizards were gone. He began a course of research to identify the causes of the decline and to determine the steps that could restore the lizard population. The collared lizard is a fascinating animal that feeds on insects such as grasshoppers. It prefers to live in dry, open areas and has a geographic range that spans from Kansas to Mexico; Missouri lies on

the eastern edge of this range. Although much of the Ozark Mountains are forested, there are openings in the forest, known as glades, that contain exposed bedrock with the hot and dry conditions that provided suitable habitat for the lizards. These small, patchy habitats were once surrounded by savannas and forests with open understories. Over time, these habitats began to change. In the Ozark Mountains, forest fires were historically common. These fires removed the small understory trees and accumulating leaves scattered across the forest floor. Forest fires were also important for maintaining the glades as open, sunny areas. However, beginning in the 1940s, a national campaign to suppress forest fires caused the glade habitats to be invaded by eastern red cedar trees (Juniperus virginiana). The cedar trees shaded the glades and made them cooler, which is not conducive to the growth of an ectotherm such as the collared lizard. The shade also reduced the number of insect prey, including grasshoppers. With the passage of time, only a few glades continued to support lizard populations and the geographic range shrank. Moreover, there appeared to be no movement of the lizards from these suitable glades to other suitable glades that were unoccupied. “The great success in restoring the collared lizard required researchers to understand its habitat needs and to recognize that the regional population was actually composed of many small populations interconnected over a large spatial area.” Templeton and his colleagues began their research in 1979. They discovered that many glades once occupied by lizards no longer had any lizard populations. They took several steps to restore those populations. First, they cut down the cedar trees in several glades to make the habitat more suitable. Next, they reintroduced 28 lizards to
three sites. They marked the animals to estimate the abundance and density of each population and to determine whether the lizards they introduced would disperse to colonize the neighboring glades. Dispersal events were rare and no neighboring glades were colonized, despite the fact that some were only 50 m away. Although several glades had been cleared of cedar trees, conditions were still not suitable; the thick forest understory and accumulating leaf litter shaded much of the ground, which made the forest cooler and caused lower abundances of insect prey. At this point, the researchers realized that they had two tasks. They needed to improve both the habitat of the glades, where the lizards spent most of their time, and the forested habitat between the glades through which the lizards traveled when dispersing from one glade to another. In cooperation with the Missouri Department of Conservation, the team began using controlled fires in 1994 to burn away the cedars in many glades as well as the small understory trees and leaves in the forest habitat between the glades. The lizards responded so well that 2 months after the burn, they began to colonize new glades. Two decades later, Templeton and colleagues reported that from the original three glades where lizards were reintroduced, there were more than 500 lizards spread across more than 140 glades. Burning the forested habitat allowed more sunlight to come in and dramatically increased the number of grasshoppers for the lizards to eat when they traveled between glades.
A glade in the Ozarks. The population increase and enlarged geographic range also improved the genetic diversity of the lizards. Prior to the controlled burning of the glades, the lizard populations remained small and contained, and their genetic diversity remained low due to the founder effect and genetic drift, which we discussed in Chapter 7. Today, the future of the collared lizard looks bright in Missouri as the use of controlled fire has brought back the required habitat, the genetic diversity is higher, and dispersal among glades allows any local populations that might go extinct to be recolonized by new dispersers. The great success in restoring the collared lizard required researchers to understand its habitat needs and to recognize that the regional population was actually composed of many small populations interconnected over a large spatial area. In this chapter, we will explore how spatial distribution and the movement of individuals among habitats influence the long-term persistence of species.
Neuwald, J. L., and A. R. Templeton. 2013. Genetic restoration in the eastern collared lizard under prescribed woodland burning. Molecular Ecology 22:3666–3479. Templeton, A. R. 2011. The transition from isolated patches to a metapopulation in the eastern collared lizard in response to prescribed fires. Ecology 92:1736–1747.
Restoration as science: The case of the collared lizard. 2011. Science Daily, August 22. http://www.sciencedaily.com/releases/2011/08/110822091918.htm.
Learning Objectives
After reading this chapter, you should be able to:
11.1 Explain why the distribution of populations is limited to ecologically suitable habitats.
11.2 Give the five important characteristics of population distributions.
11.3 Describe how distribution properties of populations can be estimated.
11.4 Recognize that population abundance and density are related to geographic range and adult body size.
11.5 Explain why dispersal is essential to colonizing new areas.
11.6 Describe how populations commonly live in distinct patches of habitat. The story of the collared lizard demonstrates that studying ecology at the population level is fascinating and can have real-world applications, just as the prior chapters on ecology at the individual level showed. In this chapter, we will focus on the spatial structure of populations, which is defined as the pattern of density and spacing of individuals. We will begin by examining how the distribution of suitable habitat affects the distribution of populations. We will then discuss the many properties of population distributions and examine how we estimate these properties. Next, we will investigate the importance of individuals moving between patches of habitats. Finally, we will learn how providing strips of favorable habitat can facilitate the movement of individuals between habitats to help ensure the persistence of populations over time. Spatial structure
The pattern of density and spacing of individuals in a population.
#### 11.1 the Distribution of Populations Is Limited to Ecologically Suitable Habitats
11.1 The distribution of populations is limited to ecologically suitable habitats At the beginning of this chapter, we saw that collared lizards inhabited habitat patches, known as glades, interspersed within a forest. As these glades became invaded by cedar trees, however, they became less suitable for the collared lizards. In this section, we will explore how ecologists determine the suitability of habitats. We will see that understanding habitat suitability is critical for understanding where a species is capable of living and the extent to which a species can expand its geographic range.
Determining Suitable Habitats
In Chapter 1, we mentioned that a species niche includes the range of abiotic and biotic conditions it can tolerate. We are now ready to elaborate on this point. To explore the concept of niche, it is useful to draw a distinction between the fundamental niche and the realized niche of a species. The fundamental niche of a species is the range of abiotic conditions under which species can persist. This includes the range of temperature, humidity, and salinity conditions that allow a population to survive, grow, and reproduce. Fundamental niche The range of abiotic conditions under which species can persist. Although a species can potentially live under the conditions of its fundamental niche, many favorable locations can remain unoccupied because of other species in those locations. For example, the presence of competitors, predators, and pathogens can often prevent a population from persisting in an area, despite the existence of favorable abiotic conditions. The range of abiotic and biotic conditions under which a species persists is its realized niche. The realized niche determines the geographic range of a species or of various populations that compose a species. The geographic range is a measure of the total area covered by a population. For example, the American chestnut tree (Castanea dentata) was once a very common tree in the eastern forest of the United States because it grew and reproduced well under the abiotic and biotic conditions that existed in this region for thousands of years.
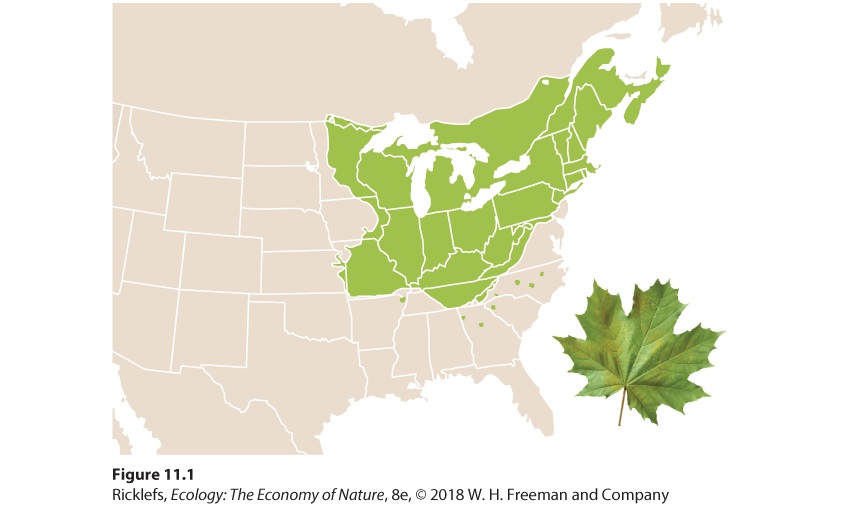
Around 1900, however, a fungus (Cryphonectria parasitica) introduced from Asia caused a deadly disease known as chestnut blight. The fungus spread rapidly throughout the eastern forests and killed billions of trees. As a result of this biotic interaction between the chestnut trees and the fungus, few places remain where adult trees can persist. In short, the fungus has caused a major reduction in the realized niche of the American chestnut. Realized niche The range of abiotic and biotic conditions under which a species persists. Geographic range A measure of the total area covered by a population. When we think about the geographic range of a species or population, we need to realize that individuals often do not occupy every location within the range. This is because climate, topography, soils, vegetation structure, and other factors influence the abundance of individuals. Consider the case of the sugar maple tree. As shown in Figure 11.1, its geographic range includes the midwestern United States, the northeastern United States, and southeastern Canada. Its distribution is limited by cold winter temperatures at the northern extent of its range, by summer droughts at the western extent, and by hot summer temperatures at the southern extent. Throughout this entire range, however, the sugar maple does not live everywhere. For instance, it cannot live in marshes, newly formed sand dunes, recently burned areas, and a variety of other habitats that lie outside its fundamental niche. As a result, the geographic range of the sugar maple is actually composed of a patchwork of smaller occupied and unoccupied areas.
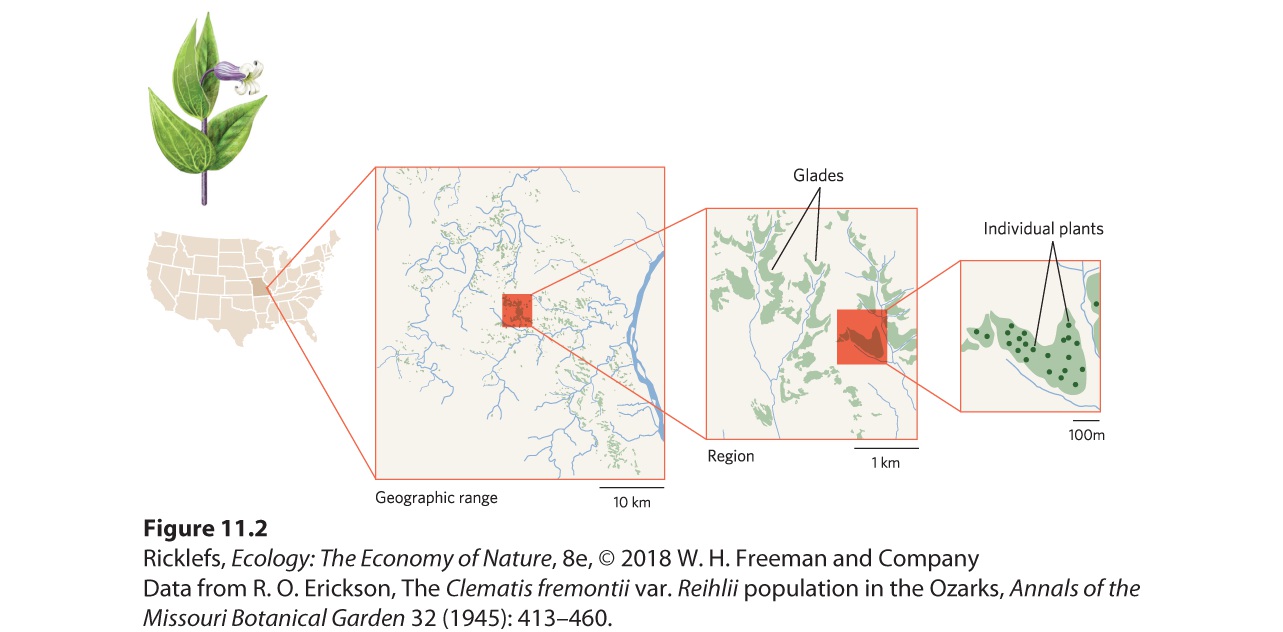
Figure 11.1 The geographic range of the sugar maple. The range is limited in the north by low winter temperatures, to the south by high summer temperatures, and to the west by droughts. The distribution of a shrub known as Fremont’s leather flower (Clematis fremontii) provides an excellent example of how small-scale variation in the environment can create geographic ranges that are composed of small patches of suitable habitat. As illustrated in Figure 11.2, the plant’s geographic range is just three counties in the state of Missouri. This small range is thought to be the result of climatic conditions and competitive interactions with ecologically similar plants. Within its geographic range, the plant is restricted to dry, rocky soils on outcroppings of limestone, known as limestone glades, which are similar to the glades frequented by the collared lizard discussed at the beginning of this chapter. Small variations in elevation and soil quality further confine these plants within each limestone glade to sites with suitable soil structure, moisture, and nutrients. Local aggregations occurring on each of these sites consist of individuals that are fairly evenly distributed in space. In other words, while Fremont’s leather flower has a geographic range that includes three counties, its distribution is spotty in this region due to its narrow habitat requirements.
Figure 11.2 The geographic range of Fremont’s leather flower. A survey found that this shrub species only existed in three counties in the state of Missouri. Within this range, individuals were only found living in the outcroppings of limestone, known as glades. Although patterns of distribution would suggest that only certain habitats are suitable, we can test whether this is the case. Consider the case of two species of wildflowers that live at distinct elevations in the Sierra Nevada of California. One species, known as Lewis’ monkeyflower (Mimulus lewisii), lives at higher elevations. The other species, known as the scarlet monkeyflower (Mimulus cardinalis), lives at lower elevations. Both species occur at mid-elevations. To determine if environmental conditions cause these different species distributions, researchers planted the two species at locations within and outside of the elevations where they grow in nature. The results of this experiment are shown in Figure 11.3. If we examine plant survival, we see that Lewis’ monkeyflower survives well at high elevations but survives poorly at low elevations. The opposite is true for the scarlet monkeyflower. If we examine plant growth, a similar pattern emerges. Lewis’ monkeyflower grows better as elevation increases, although growth declines under the extreme conditions of the highest elevation. The scarlet monkeyflower grows the best at low elevations and its growth declines with each increase in elevation. For each species, the survival and growth of the transplanted populations were the highest when grown within its normal elevation range. When grown outside this range, both species experienced lower survival and slower growth.
Figure 11.3 The distribution of two monkeyflower species. The scarlet monkeyflower is found at low elevations, whereas Lewis’ monkeyflower grows at high elevations, as shown in the cross-section of the mountain. When transplanted to gardens at different elevations, the scarlet monkeyflower survives best at low elevations, while Lewis’ monkeyflower survives best at high elevations. Similar patterns occur for plant growth. Error bars are standard errors.
Ecological Niche Modeling
As a general rule, the more suitable the habitat, the larger a population can grow within that habitat. This fundamental relationship allows ecologists to predict the actual or potential distributions of species, which has a number of important applications. For example, to bring species back from the brink of extinction, we need to know the habitat conditions that the species requires. This knowledge is used to determine the locations that would provide the highest probability of successful reintroductions. Similarly, if a new pest species is accidentally introduced to a continent, its suitable habitats must be assessed to predict the area over which it might spread and the extent of the damage it might cause. Predicting the potential geographic range of a single population or of all populations of a given species is a major challenge when there are few individuals living in the wild. One way to overcome this challenge is by using historic data on the distributions of populations. Such data are often

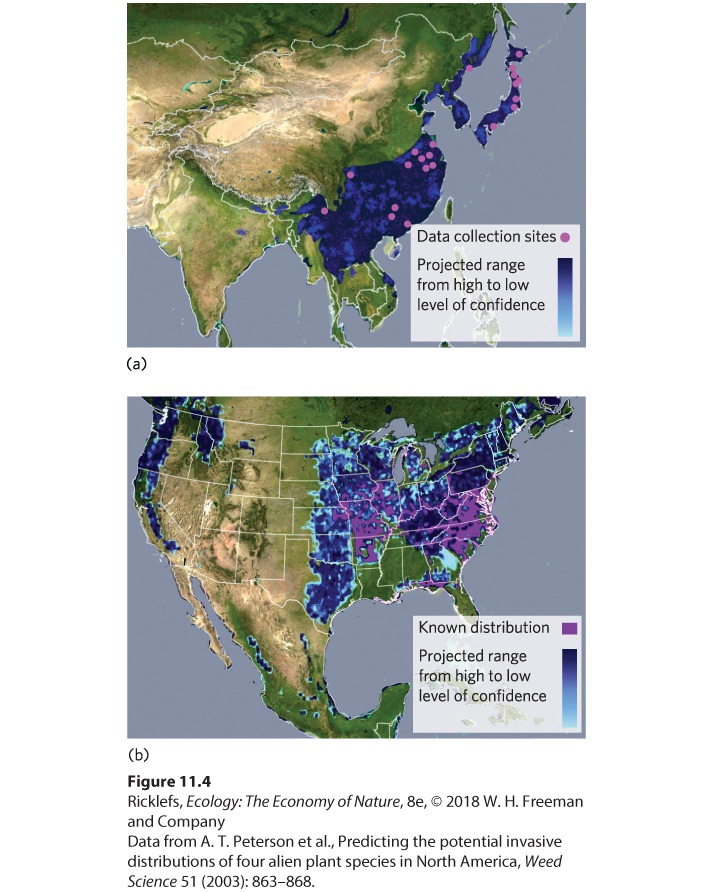
available from collections of preserved organisms stored in museums and herbariums. In addition, when species have been introduced from other continents, researchers can try to determine the suitable habitat conditions found on the originating continent. The process of determining the suitable habitat conditions for a species is known as ecological niche modeling. Because temperature and precipitation have a dominant influence on the distribution of biomes, modelers often begin by mapping the known locations of a species and then by quantifying the ecological conditions at the locations where the species has been recorded. The modeler can potentially include many additional variables such as different soil types and the presence of potential predators, competitors, and pathogens that might limit the population’s distribution. The range of ecological conditions that are predicted to be suitable for a species is the ecological envelope of the species. The concept of the ecological envelope is similar to the realized niche, but the realized niche includes the conditions under which a species currently lives, whereas the ecological envelope is a prediction of where a species could potentially live. Ecological niche modeling The process of determining the suitable habitat conditions for a species. Ecological envelope The range of ecological conditions that are predicted to be suitable for a species. Modeling the Spread of Invasive Species Ecological niche modeling can be a useful way to predict the expansion of pest species introduced to a continent where they have not previously lived. One example is the Chinese bushclover (Lespedeza cuneata), which is native to eastern Asia. In the late 1800s, this species of bushclover was brought to North Carolina to help control erosion, reclaim land that had been mined, and provide food for cattle. Over time, though, the plant quickly spread into the Great Smoky Mountains National Park and throughout the grasslands of the United States, where it displaced native plants. To determine the likely extent of its future spread, ecologists gathered data on the environmental conditions under which the bushclover lived in 28 locations in Asia. As illustrated in
Figure 11.4a, they then quantified the ecological envelope for the clover to predict the entire geographic range in Asia. They used these data to predict
the potential geographic range in North America. As you can see in Figure 11.4b, they successfully predicted all locations where the clover had already spread. The model also predicted that the clover has the potential to live in many other locations, with widespread negative effects on the plant communities. The lack of Chinese bushclover in these areas suggests either that the model doesn’t include additional ecological factors important to the clover or that the clover simply has not had enough time to disperse to these more distant locations.
Figure 11.4 Ecological modeling of an invasive species. (a) Researchers collected data on the environmental conditions where the Chinese bushclover lives in Asia and used these data to predict the entire native range of the plant in Asia. (b) They then used the data to
predict the future geographic range of Chinese bushclover in North America, where the plant was introduced 100 years ago and has continued to spread slowly.
Habitat Suitability and Global Warming
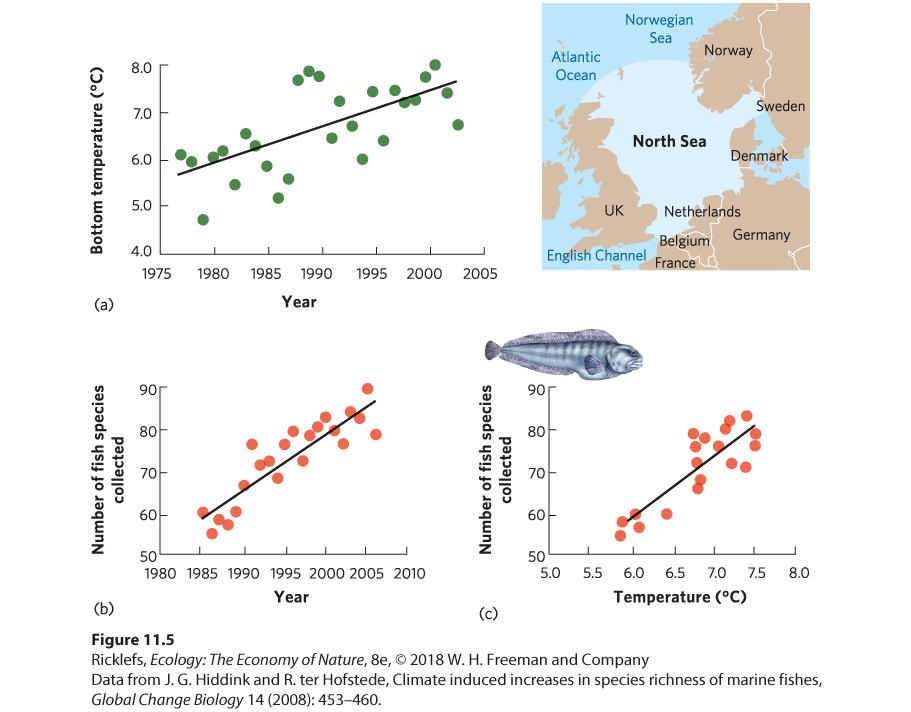
Knowledge of the environmental conditions that make a habitat suitable can also be used to understand the shift in the geographic ranges of species as the environmental conditions of the world change. During the past century, for example, the average temperature of Earth has increased by 0.9 °C. Some regions of the world—such as Alaska and northern Canada—have warmed as much as 4 °C. In the relatively shallow North Sea, situated between the United Kingdom and Norway, temperatures in the bottom waters have increased more than 2 °C since the 1970s, as shown in Figure 11.5a. Given that most species of fish have optimal temperature ranges, we might predict that warming ocean waters could cause southern fish species, which live in warm waters, to move north.
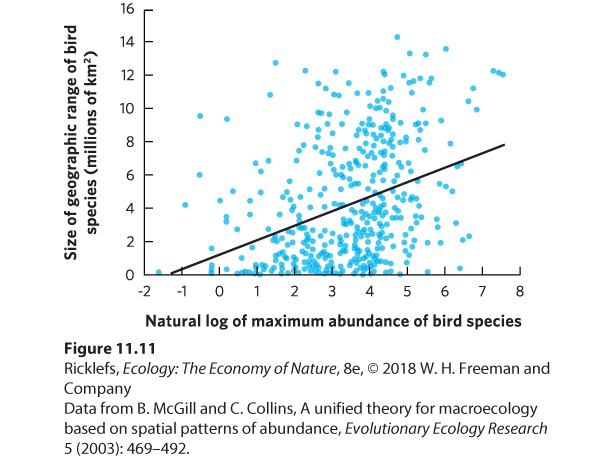
Figure 11.5 Changes in the distribution of fish in the North Sea. (a) From 1977 to 2003, the temperature of the water at the bottom of the North Sea increased by 2 °C. (b) Surveys of fish trawls from 1985 to 2006 found that the total number of fish species collected per year in the North Sea increased from 60 to approximately 90. (c) The total number of fish species collected per year was positively correlated with the average sea temperatures during the previous 5 years.
During the same period that temperatures in the North Sea were being monitored, the International Council for the Exploration of the Sea (ICES) compiled data on fish distributions by pulling large nets (called “trawls”) along the ocean floor. From 1985 to 2006, scientists from six countries worked together to fish the bottom of the ocean at 300 locations distributed throughout the North Sea. Based on 7,000 trawl samples, the researchers reported that fish species richness in the North Sea had increased steadily over 22 years. As you can see in Figure 11.5b, there were about 60 species in the mid-1980s, but this grew to nearly 90 species 2 decades later. This list of species included dozens of more southerly species that had expanded their ranges northward. The increase in species richness was positively correlated with the increase in bottom-water temperatures in the North Sea. This correlation—shown in
Figure 11.5c—suggests that warmer temperatures are more hospitable to a greater variety of species, and that the warming of the North Sea has allowed more southerly species to expand the northern edges of their ranges into the area. Hence, this is a case of global warming increasing the diversity of species in a region. The increase in the diversity of species in the North Sea not only is dramatically changing the fish community but also may affect the important commercial fisheries that depend on this community. For example, the three species whose ranges have contracted as the North Sea became warmer—the wolffish (Anarhichas lupus), the spiny dogfish (Squalus acanthias), and the ling (Molva molva)—are all commercially important, whereas more than half of the southerly species with expanded ranges have little or no commercial value. As a result, while these shifts in species distributions with warming temperatures may increase fish diversity, it could actually decrease the value of North Sea commercial fisheries.
Concept Check
1. What is a species’ realized niche? 2. Why are not all locations in a species’ geographic range occupied by the species? 3. What two environmental factors are commonly used when

#### 11.2 Population Distributions Have Five Important Characteristics
11.2 Population distributions have five important characteristics To study the distribution of a population like the collared lizard in the Ozark Mountains, several characteristics of the population must be considered, including geographic range, abundance, density, dispersion, and dispersal. As we will see, each of these properties tells us something important about how individuals are distributed.
Geographic Range
We saw an example of geographic range in our discussion of the sugar maple tree (see Figure 11.1). The geographic range of a species includes all the areas its members occupy during their life. For example, the geographic range of sockeye salmon includes not only the rivers of western North America and eastern Asia, which are their spawning grounds, but also vast areas of the North Pacific Ocean, where individuals grow to maturity before making the long migration back to their birthplace. The geographic range is an important measure because it tells us how large an area a population occupies. If a population is restricted to a small area, for example, it may be very susceptible to a natural disaster such as a hurricane or a fire that can wipe it out. This is a serious challenge for endemic species, which live in a single, often isolated, location, such as the Galápagos finches that live on islands off the coast of South America. Populations with a larger geographic range are less vulnerable to such events because much of the population would remain unaffected. Species with very large geographic ranges that can span several continents are known as cosmopolitan species. Endemic species Species that live in a single, often isolated, location. Cosmopolitan Species with very large geographic ranges that can span several continents.
Abundance
The abundance of a population is the total number of individuals that exist within a defined area. For example, we might count the total number of
lizards on a mountain, the number of sunfish in a lake, or the number of coconut trees on an island. The total abundance of a population is important because it provides a measure of whether a population is thriving or on the brink of extinction. Abundance The total number of individuals in a population that exist within a defined area.
The density of a population is the number of individuals in a quantified area or volume. If we know the abundance of a population in a given area and we know the size of the area, then we can calculate density by dividing the abundance by the area. Examples of density include the number of bears per square kilometer in Alaska, the number of cattails per square meter in a pond, or the number of bacteria per milliliter of water. Density is a valuable measure because it tells ecologists how many individuals are packed into a particular area. If a habitat can support a higher density than currently exists, the population can continue to grow in the area. If the population density is greater than what the habitat can support, either some individuals will have to leave the area or the population will experience lower growth and survival. Density In a population, the number of individuals in a quantified area or volume. Although individuals live only in suitable habitats, not all habitats are equal in quality because the environment is inherently variable. Some habitat patches have abundant resources that support a large number of individuals, while others have scarce resources and can support only a few individuals. Across a large geographic area, the highest concentrations of individuals are typically near the center of a population’s geographic range. As one moves closer to the periphery of the geographic range, biotic and abiotic conditions become less ideal and support fewer individuals. Consider, for example, the geographic range of the dickcissel (Spiza americana), a small songbird related to the cardinal and found in North American prairies and grasslands. As illustrated in Figure 11.6, this bird has its highest densities in the center of its geographic range and its lowest densities near the periphery. However,
because environmental conditions do not vary smoothly, the pattern of the dickcissel’s preferred habitat is highly irregular.
Figure 11.6 Densities across a geographic range. In the dickcissel bird, a relative of the cardinal, the highest densities are near the center of its geographic range and the lowest densities are near the periphery of its geographic range.
Dispersion
Dispersion of a population describes the spacing of individuals with respect to one another within the geographic range of a population. As shown in
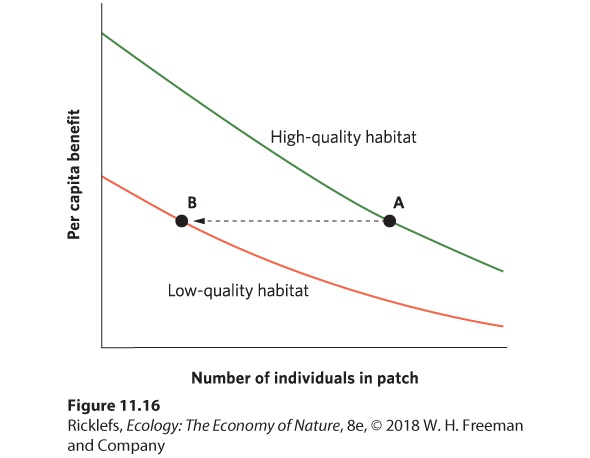
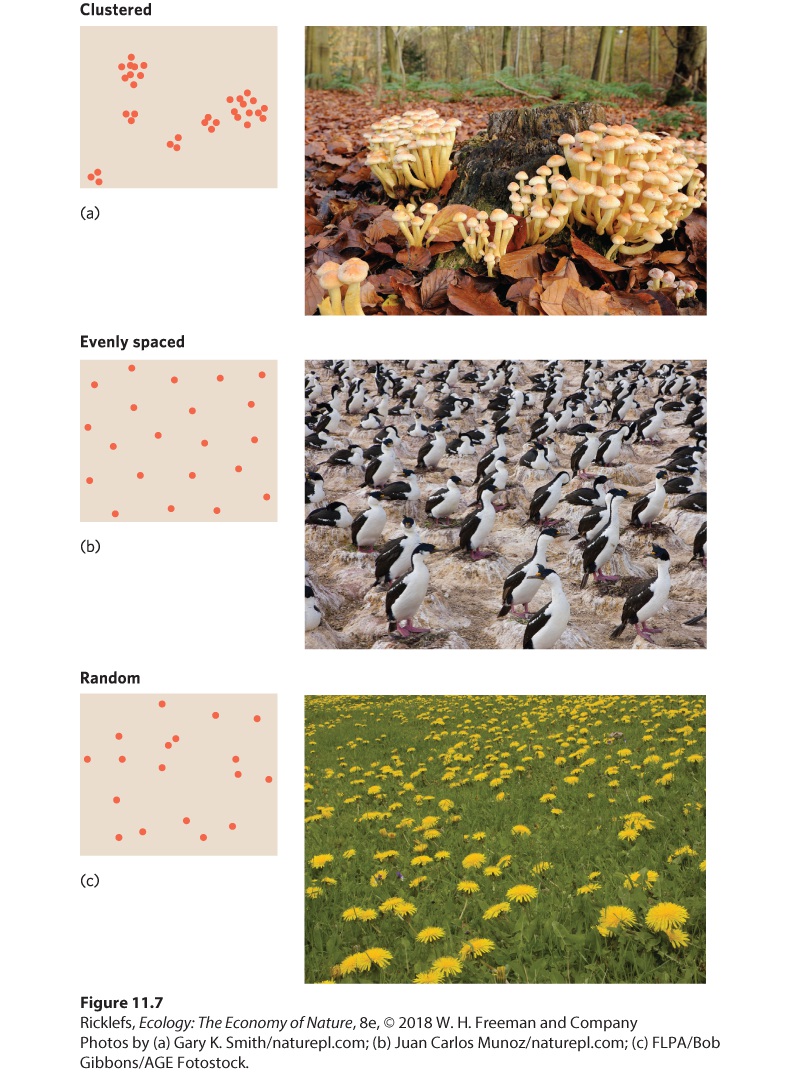
Figure 11.7, dispersion patterns can either be clustered, evenly spaced, or random.
Figure 11.7 Three types of dispersion patterns. (a) Clustered dispersion is characterized by individuals that are aggregated. For example, sulphur tuft fungi (Hypholoma fasciculare) grow in clusters anywhere there is a rotting stump, such as this site in the United Kingdom. (b) Evenly spaced dispersion is characterized by each individual maintaining a minimum distance between neighbors, as is the case for these blue-eyed cormorants (Phalacrocorax atriceps) nesting in Patagonia, Argentina. (c) Random dispersion is characterized by each individual’s position being independent of the location of other individuals, such as these dandelions growing in a pasture in Bulgaria. Dispersion The spacing of individuals with respect to one another within the geographic range of a population.
Clustered Dispersion In clustered dispersion, individuals are aggregated in discrete groups. Some clustered dispersions result from individuals living in social groups, as we discussed in Chapter 10. For instance, some birds live in flocks and some fish live in schools. Other clustered dispersions occur because individuals stay near clustered resources. For example, salamanders and sow bugs aggregate under logs because individuals of both species are attracted to dark, moist places. Another cause of clustered dispersions is offspring that remain close to their parents. Some species of trees, such as the quaking aspen, form clusters of stems because a parent tree gives rise to offspring by sending up new stems from its roots, a form of vegetative reproduction. As a result, we commonly see clusters of aspen trees that are composed of a parent tree surrounded by its offspring. Within such a cluster, however, the stems tend to be evenly spaced. As a result, a population can exhibit one pattern of dispersion at a large scale but a different pattern of dispersion at a smaller scale. Clustered dispersion A pattern of population dispersion in which individuals are aggregated in discrete groups. Evenly Spaced Dispersion In evenly spaced dispersion, each individual maintains a uniform distance between itself and its neighbors. In agricultural settings, we can observe even spacing in crops such as corn or apple trees because farmers want each plant to have sufficient resources to maximize crop production. In natural settings, even spacing most commonly arises from direct interactions between individuals. For example, plants positioned too close to larger neighbors often suffer from shading and root competition. In addition, some plants can emit chemicals from their leaves and roots that inhibit the growth of other plants around them. As these crowded individuals die, the remaining individuals become more evenly spaced. We can also observe evenly spaced distributions in animals that defend territories, such as birds and lizards. Because territory size commonly depends on the amount of resources available, neighboring territory holders commonly defend areas that are similar in size, which causes the territory holders to be evenly spaced.
Evenly spaced dispersion A pattern of dispersion of a population in which each individual maintains a uniform distance between itself and its neighbors. Random Dispersion In random dispersion, the position of each individual is independent of the position of other individuals in the population. Random dispersions are not common in nature, largely because abiotic conditions, resources, and interactions with other species are not randomly distributed. However, if we wished to know whether the dispersion pattern of a population at a particular spatial scale is clustered or evenly spaced, we would have to demonstrate statistically that its distribution was significantly different from random. Random dispersion A pattern of dispersion of a population in which the position of each individual is independent of the position of other individuals in the population.
Dispersal
Dispersal is the movement of individuals from one area to another. Dispersal is distinct from migration, which is the seasonal movement of individuals back and forth between habitats, such as birds flying north and south with the changing seasons. In contrast, dispersal involves individuals leaving their habitat of origin—where a seed was made or where a squirrel was born—and typically not returning. As we saw at the beginning of this chapter, dispersal is of great interest to ecologists because it is the mechanism by which individuals can move between suitable habitats and, in some cases, colonize suitable habitats that are not already inhabited by the species. Dispersal can also be a way to avoid areas of high competition or high predation risk. For example, when a fish arrives in a section of a stream, many of the aquatic insects in the stream disperse by floating downstream at night to avoid being eaten. Dispersal The movement of individuals from one area to another.
Concept Check
1. What mechanisms could cause individuals in a population distribution to be either evenly spaced or clustered? 2. Why are endemic species at a high risk of extinction? 3. What is the difference between dispersal and dispersion when describing population distributions?
#### 11.3 the Distribution Properties of Populations Can Be Estimated
11.3 The distribution properties of populations can be estimated Thus far, we have focused on the conceptual basis of the five properties— abundance, density, geographic range, dispersion, and dispersal. For a more complete understanding of how populations are distributed, quantitative measures must also be considered.
Quantifying the Location and Number of Individuals
One way to determine the number of individuals in an area is to conduct a census, which means counting every individual in a population. Every 10 years, for example, the United States government conducts a census with the goal of counting every person living in the country. For most species, however, it is not feasible to count every individual in the population. As a result, scientists must conduct a survey, in which they count a subset of the population. Using these samples, they estimate the abundance, density, geographic range, and distribution of the population. Scientists have developed a variety of ways to make these estimates, including area- and volume-based surveys, line-transect surveys, and mark-recapture surveys (Figure 11.8).
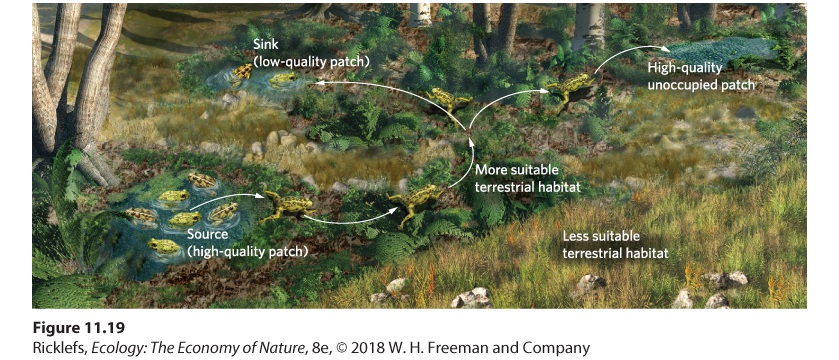
Figure 11.8 Estimating population abundance, density, and distribution. (a) Area-based studies count the number of individuals within a fixed area, such as this researcher working on the Dendles National Natural Reserve in England. (b) Linear-transect studies, including this one at the Great Barrier Reef in Australia, count the number of individuals that are observed along a predefined line. (c) Mark-recapture studies collect a sample of the population, mark
them, and then return them. For example, horseshoe crabs (Limulus polyphemus) in Delaware are marked with small circular tags. A short time later, a second sample is collected to determine the proportion of marked animals in the population, which can be used to estimate the size of the total population. Census A count of every individual in a population. Survey Counting a subset of the population. Area- and Volume-Based Surveys Area- and volume-based surveys define the boundaries of an area or volume and then count all the individuals within that space. The size of the defined space is typically related to the abundance and density of the population. For example, researchers who wish to know the number of bacteria in the soil might collect samples of soil that are only a few cubic centimeters in volume. In contrast, researchers who wish to know the number of individual corals on a coral reef might sample areas that are 1 m2. At the most extreme, researchers who want to estimate the abundance, density, and distribution of large mammals might count the number of individuals in aerial photos that cover hundreds of square meters. By taking multiple samples, scientists can determine how many individuals are present in an average sample area of land or volume of soil or water. Area- and volume-based surveys Surveys that define the boundaries of an area or volume and then count all the individuals in the space. Line-Transect Surveys Line-transect surveys count the number of individuals observed as one moves along a line. There are many variations on this technique. For example, researchers might survey small plants in a field or forest by tying a long string between two fixed points and counting the number of individuals the string crosses. From this, we can determine the abundance in a given linear distance. Alternatively, researchers might count all individuals that are observed within a fixed distance of a line, such as the number of trees on a savanna located within 100 m of a line, which would provide us with an
estimate of abundance in a given area. A similar approach has been used in surveys of amphibians. In this case, observers count the number of frogs that can be heard along a predetermined path. If we know how far, on average, a person can hear a frog call, we can estimate the number of frogs calling in an area that includes both sides of the path. Such line-transect data can be converted into area estimates. Line-transect surveys Surveys that count the number of individuals observed as one moves along a line. One of the most famous line-transect studies is the annual Christmas bird count. The bird count began in 1900 when 27 volunteers from the Audubon Society positioned themselves at different locations in North America and counted all the birds they saw in one day. Today, tens of thousands of volunteers go outside during their winter holidays and follow a predetermined path that covers a 24-km circle. Throughout the day, the volunteers count the number of individuals of every bird species they can see or hear within this circle. This long-term survey of birds in North America has provided incredibly valuable data that has helped scientists determine which species of birds have populations that are increasing, stable, or declining. Mark-Recapture Surveys Area- and volume-based and line-transect studies are very useful for organisms that do not move—such as plants and corals—or animals that are not easily disturbed—such as snails—and therefore less likely to leave the area during the survey. Some animals, however, are very sensitive to the presence of researchers and will leave the area, while other species are well camouflaged and difficult to find. Both situations can cause us to underestimate the number of individuals in a population. For these situations, we need a different type of sampling technique. One effective method is the use of mark-recapture surveys. As the name implies, mark-recapture surveys collect a number of individuals from a population and mark them. These individuals are then returned to the population. Once enough time has passed for the marked individuals to mingle throughout the population, a second sample of the population is made. Based on the number originally
marked, the total number collected the second time, and the number of marked animals collected the second time, we can estimate the size of the population. Mark-recapture studies are commonly conducted on birds, fish, mammals, and highly mobile invertebrates. The actual calculations for arriving at this estimate are discussed in “Analyzing Ecology: MarkRecapture Surveys.” Mark-recapture survey A method of population estimation in which researchers capture and mark a subset of a population from an area, return it to the area, and then capture a second sample of the population after some time has passed.
Analyzing Ecology
Mark-Recapture Surveys To estimate the number of individuals in a population using mark-recapture surveys, we need to know how many individuals were initially sampled and marked. For example, let’s imagine that crayfish researchers collected 20 crayfish from a 300-m2 stretch of stream and marked them with a dot of red fingernail polish. Once the fingernail polish was dry, the crayfish were returned to the stream. After waiting one day for the marked crayfish to move around the stream, the researchers collected another sample of crayfish. In this second sample, they captured 30 crayfish, 12 of which were marked. Based on these data, how many crayfish were in the 300 m2 of stream? To find out, we can use an equation that considers how many individuals are marked and—after the marked individuals are released back into the population—the ratio of total individuals to marked individuals in the entire population. Let’s look at how we estimate the size of the population. First, note that that we capture and mark a number of individuals (M) from an entire population whose size is defined as N. Therefore, the fraction of marked individuals in the entire population is MN . When we go back and capture individuals the second time, we record

the number of individuals in the second capture (C) and the number of marked individuals that are recaptured (R). The fraction of marked individuals in the recaptured population is RC . The fraction RC and our first fraction, MN , actually represent the same fraction, which is the proportion of marked individuals in a sample. Given that these two fractions should represent the same number, we can set the two ratios equal to each other and solve for the unknown variable (N), which is the total size of the population: MN=RCN=M×C÷R Applying this equation to our crayfish data, the estimated number of crayfish in the stream is N=20×30÷12N=50 YOUR TURN What is the estimated size of the crayfish population if the second sample collected 48 crayfish and the number recaptured in that sample was 24? Based on your estimate of crayfish abundance and the data provided on stream area, what is your estimate of crayfish density?
Quantifying the Dispersal of Individuals
Quantifying the dispersal of individuals from a population requires identifying the source of individuals. As we will see, this can be done by ensuring that there is only one possible source of individuals and then determining how far individuals disperse from this single location. In other cases, individuals are marked and then observed or recaptured at some later time to determine how far they moved from the location where they were marked (Figure 11.9). In animal studies, possible marks include ear tags, radio transmitters, or leg bands. In plant studies, researchers can mark pollen with fluorescent powders and then examine surrounding flowers to determine how far the pollen grains have been moved either by the wind or by pollinators.
Figure 11.9 Measuring dispersal. This California condor (Gymnogyps californianus), which was captured in the Grand Canyon National Park in Arizona, has been marked using a wing band that has a unique number to identify where it hatched and how far it dispersed. A common measure of dispersal is the lifetime dispersal distance, which is the average distance that an individual moves from where it begins its life to where it reproduces. By knowing the lifetime dispersal distance, we can also estimate how rapidly a growing population can increase its geographic range. For example, when researchers marked eight species of songbirds with leg bands, they found that lifetime dispersal distances averaged between 344 and 1,681 m. A lifetime dispersal distance of about 1 km per generation is therefore not unusual for populations of songbirds. At this rate, the descendants of an average individual might traverse an entire continent in a thousand generations or so. Lifetime dispersal distance The average distance an individual moves from where it was hatched or born to where it reproduces. These calculations suggest that it will take an average species of songbird more than 1,000 generations of dispersal to travel across a continent. However, it can actually happen much faster because a few individuals in a population can disperse much farther than the
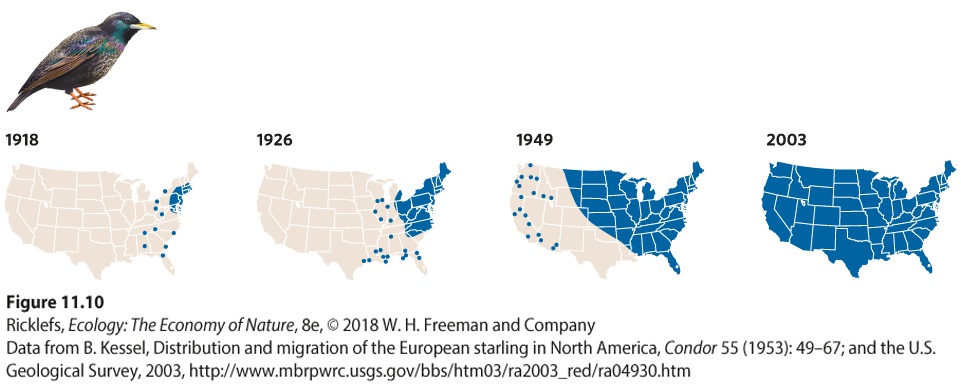
average bird in the population. An excellent example of this occurred following the introduction of the European starling to the United States. In 1890 and 1891, 160 European starlings were released in the vicinity of New York City. Within 60 years, the population had spread 4,000 km from New York to California, at an average rate of about 67 km per year. You can see this rapid spread of starlings in Figure 11.10. The expansion occurred rapidly because a few individuals dispersed much longer distances than the average and established new populations beyond the range boundary of the species. Although the few individuals that might move over such long distances are rare, they can have large effects on population distributions. Today, the starling lives throughout most of the United States.
Figure 11.10 The rapid dispersal of European starlings across the United States. After being introduced near New York City in 1890 and 1891, the bird rapidly spread across the United States over a 60-year period. The blue-shaded area represents the range of breeding populations. The blue dots indicate detections of starlings that exhibited unusually long dispersal, thereby facilitating the rapid spread of the species. Today, the starling population covers more than 7 million km2 of North America from coast to coast.
1. Why are surveys, rather than censuses, used to quantify the abundance of many animals? 2. How can line-transect surveys be used to estimate the number of animals per unit area? 3. Why is the average dispersal distance a misleading estimate of how rapidly a population can move across very long distances over time?
#### 11.4 Population Abundance and Density Are Related to Geographic Range and Adult Body Size
11.4 Population abundance and density are related to geographic range and adult body size Given the five properties of population distributions, all of which can span a wide range of values, ecologists often search for relationships that might explain the underlying causes of variation in these values. Common patterns include the relationship between population abundance and geographic range and the relationship between population density and adult body size.
Population Abundance and Geographic Range
Ecologists have consistently found that populations with high abundance also have a large geographic range. This pattern has been observed in plants, mammals, birds, and protists. Consider the data on the birds of North America in Figure 11.11. We can see a positive relationship; species with the highest abundances also are the species with the largest range sizes. This same pattern was observed in collared lizards; as their abundance declined, fewer glades were occupied and the geographic range of the population shrank. In the bird data, however, there is a great deal of variation around the regression line. If we were to quantify the coefficient of determination, we would find that abundance of the populations only explained 13 percent of the variation in the geographic ranges of the populations. To review the concept of coefficient of determination, see “Analyzing Ecology: Coefficients of Determination” on page 182 in Chapter 8.
Figure 11.11 Abundance and range size of birds. Data from 457 species of North American birds show that bird species with higher abundances are generally more widely distributed. However, this relationship contains a great deal of variation around the line of best fit due, in part, to annual variations in abundance and geographic ranges. The causes of the range‒abundance relationship are still widely debated. There is general consensus that resource availability plays an important role. For example, if a species relies on resources that are only available in a small geographic area, the species will only inhabit a small geographic range. If resources are abundant over a large geographic area, we might expect that species to cover a large geographic range and to be abundant. In short, the distribution of resources should cause a positive relationship between abundance and geographic range. Although we do see this positive relationship, we are left to wonder about all the unexplained variation in the relationship shown in Figure 11.11. In some cases, we might be observing fluctuations within a geographic range. For example, individuals might disperse to more marginal habitats during years that favor especially high reproduction and survival. The range would then contract during years that have low reproduction and survival. Such year-to-year fluctuations can cause the data on the abundance and geographic range of a population to vary a great deal. The relationship between population abundance and distribution suggests
that reducing the geographic range of a population—for example, by converting habitat to agricultural purposes or to housing—will also reduce the size of the population. Similarly, factors that reduce the overall size of the population will simultaneously reduce the geographic range of the population because marginal habitats will no longer receive as many dispersing individuals.
Population Density and Adult Body Size
Another common pattern across species is the relationship between population density and body size. Generally, the density of a population is negatively correlated to the body size of the species. Figure 11.12 shows that in herbivorous animals, the smallest-bodied species—such as mice—live at the highest densities, and the largest-bodied species—such as elephants—live at the lowest densities. Part of this relationship is a matter of body size relative to space. A square meter of soil harbors hundreds of thousands of small arthropods, whereas a single elephant simply would not fit into this space. Even if it could be squeezed in, a square meter of soil would not produce enough food to sustain the elephant. A large individual requires more food and other resources than a small one. As a result, we expect a given plot of habitat to support fewer large individuals than it does small individuals.
Figure 11.12 Body mass and population density. Across 200 species of herbivorous mammals, population density declines as a function of adult body mass.
Concept Check
1. What is the relationship between population abundance and geographic range across many species? 2. What is the relationship between population density and adult body size across many species?
#### 11.5 Dispersal Is Essential to Colonizing New Areas
11.5 Dispersal is essential to colonizing new areas As we mentioned earlier in this chapter, dispersal plays a key role in allowing individuals to move between patches of habitat and to colonize suitable habitats that are not inhabited. In this section, we will discuss how suitable habitats may remain unoccupied due to dispersal limitation; we will also examine how habitat corridors can play an important role in facilitating dispersal.
Dispersal Limitation
In some cases, as we saw with the Chinese bushclover, not enough time has passed since the introduction of a species for individuals to spread to every suitable habitat. In other cases, such as the collared lizards, there can be substantial barriers that prevent dispersal between suitable habitats. A common barrier to dispersal is the presence of large expanses of inhospitable habitat that an organism cannot cross, such as an ocean that a plant seed cannot cross or a large desert that an amphibian cannot cross. Recall that the sugar maple has a preferred range of temperatures and precipitation that favors its presence in the northern temperate forests of North America. Although there are no other suitable areas for sugar maples to grow in North America, there is an abundance of suitable habitat in Europe and Asia. Indeed, several other species of the maple genus (Acer) live in habitats that should be suitable for the sugar maple. Sugar maple does not exist in Europe and Asia because its seeds are not capable of dispersing from North America across the oceans to these distant regions. Sometimes the inhospitable habitat is not particularly expansive but is still an effective barrier to dispersal, as was the case with fire-suppressed forests that surrounded glades and prevented collared lizard dispersal. The absence of a population from suitable habitat because of barriers to dispersal is called dispersal limitation. Dispersal limitation The absence of a population from suitable habitat because of barriers to dispersal. Occasionally, individuals cross formidable barriers and disperse long distances without assistance from humans. We know this because many species of plants and animals have populated remote islands, such as the
Hawaiian Islands, before humans arrived. However, humans have affected the dispersal of many species. For example, human activities such as road building and forest clearing have created barriers to dispersal for some species. Humans have also assisted in the dispersal of plants and animals for thousands of years. For instance, Aboriginal peoples brought dogs to Australia and Polynesians distributed pigs and rats throughout the small islands of the Pacific. In more recent times, foresters have transplanted fastgrowing eucalyptus trees from Australia and pines from California to points all over the world for timber and fuel wood. Other species have been intentionally moved to assist them in dispersing over barriers created by humans, such as roads. Still other species have been accidentally moved over large distances by riding along in the ballast water of cargo ships or by attaching themselves to the outside of ship hulls. In most cases, individuals are introduced into a new area but never establish a viable population. In some cases, however, the introduced individuals are able to develop into a population that can grow and expand its geographic range over time. We’ll have much more to say about such invasive organisms later in this book, but their success in many places outside their native ranges emphasizes the role of barriers to dispersal in limiting species’ distributions.
Habitat Corridors
In some landscapes, dispersal is helped when strips of favorable habitat known as habitat corridors are located between large patches of habitat. For example, two forests might be separated by an open field with a stream that has a narrow band of trees along its banks. This band of trees along the stream can serve as a corridor of suitable habitat between the forests and allow individuals to easily disperse between them. Habitat corridor A strip of favorable habitat located between two large patches of habitat that facilitates dispersal. In recent years, large manipulative experiments have tested the importance of habitat corridors. For example, in the pine forests of South Carolina, storms and fires often create cleared areas that are suitable habitat for many species of understory plants and animals. To test the importance of habitat corridors in facilitating dispersal between these cleared areas, researchers
cleared large (1.375 ha) patches of forest. One of the five cleared areas, called the central patch, was the source of dispersers. Of the remaining four patches, one was connected to the central patch with a cleared path that could serve as a corridor. The other three outer patches included a simple rectangularshaped patch and two patches with “wings,” meaning that they were patches with corridors but did not connect to other patches (Figure 11.13). The researchers replicated this experimental design eight times.
Figure 11.13 Manipulating habitat corridors. Researchers cleared patches in the pine forests of South Carolina. Using groups of five patches, the central patch served as a source of dispersers. The outer patches included one patch that was connected to the central patch by a cleared habitat corridor, while the other three were not connected. One unconnected patch was a simple rectangle. The other two patches had cleared corridor paths, termed “wings,” that were not connected to another patch. A number of methods were used to measure dispersal. The movements of the common buckeye butterfly (Junonia coenia) were followed by marking individuals in the central patch and recapturing them in the four outer patches. To track pollen movement, eight male winterberry plants (Ilex verticillata) were planted in each central patch and three mature female plants were planted in each of the four outer patches. Fertilized flowers producing fruits in the outer patches would indicate that pollen had moved from the central patch. To track seed and fruit movements, a local holly (Ilex vomitoria) and wax myrtle (Myrica cerifera) were planted in the central patch. In some cases, the fruits in the central patch were dusted with colored fluorescent powder. The researchers then collected samples of bird droppings in traps placed under artificial perches in each outer patch. Fecal matter that
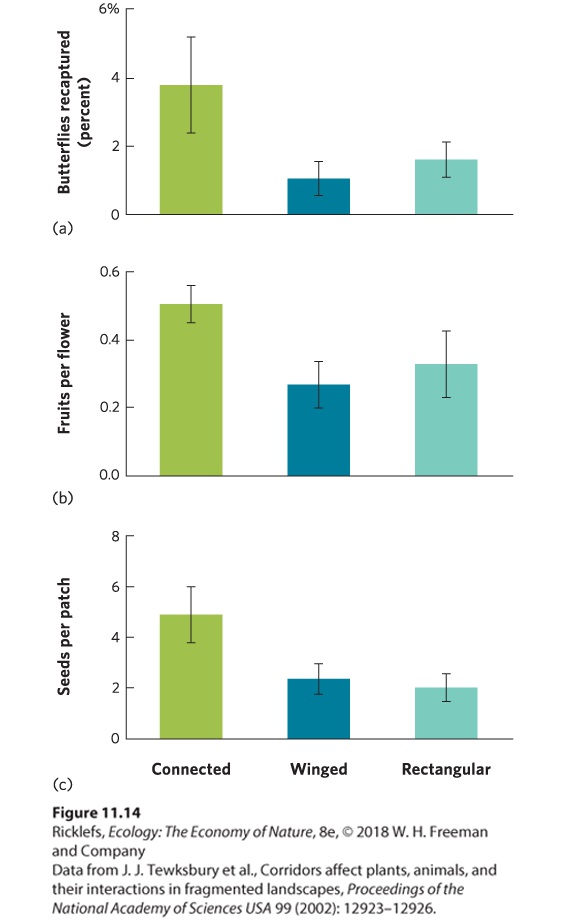
fluoresced under ultraviolet light indicated that birds had consumed fruits in the central patch and dispersed them to the outer patches. The results of these experiments are shown in Figure 11.14. The outer patch connected by a habitat corridor had much more dispersal from the central patch, including more dispersal by butterflies, more traffic by insect pollinators—which caused more fruits to be produced—and greater movement of fruits and seeds found in bird droppings. Indeed, the movement of fruits and seeds through the corridors was so frequent that the number of species of herbs and shrubs increased faster in the connected patches.
Figure 11.14 The effects of manipulated corridors on dispersal. Compared to unconnected patches that were either winged or rectangular in shape, patches connected by a habitat corridor experienced (a) a higher number of recaptured butterflies, (b) greater pollination of plants, leading to more fruits per flower, and (c) greater seed dispersal by birds. Error bars are standard errors.
Experiments such as these confirm the importance of habitat corridors. Indeed, conservation efforts have increasingly considered the preservation of corridor habitats. Along the Rio Grande in Texas, for example, state and federal biologists, in collaboration with conservation organizations, have pushed to protect river-side habitats that would allow species to move easily among large patches of protected land. As you can see in Figure 11.15, in the United States, this land includes Big Bend National Park, Big Bend Ranch State Park, and the Black Gap Wildlife Management Area. Across the river in Mexico the protected lands include the Cañon de Santa Elena Flora and Fauna Protected Area, the Ocampo Flora and Fauna Protected Area, and the Maderas del Carmen Flora and Fauna Protected Area.
Figure 11.15 Conservation of habitat corridors. On the border of Texas and Mexico, state and national governments have protected a massive corridor of more than 1.3 million ha of land along the Rio Grande.
1. Why are many species absent from continents that contain suitable habitat? 2. How do habitat corridors facilitate dispersal?
#### 11.6 Many Populations Live in Distinct Patches of Habitat
11.6 Many populations live in distinct patches of habitat In our discussion of the collared lizard, we saw that individuals in the region live in many small populations, with each group restricted to a particular glade. The lizards can also move between habitats, but such movement depends on the quality of the forested habitat connecting the glades. When we want to understand the movement of individuals among patches of habitats and how this affects the abundance of animals in each patch of habitat, we need to consider the quality of the habitat. We also need to consider the difficulty in reaching the habitat, which is determined by the distance to the habitat and the difficulty in crossing the area between habitat patches. In this section, we will explore how habitat quality affects the distribution of individuals among habitats and how ecologists have conceptualized the distribution and movement of individuals among suitable patches of habitat.
The Ideal Free Distribution Among Habitats
When habitats differ in quality and individuals can move easily among habitat patches, natural selection should favor those individuals that can choose the habitat that provides them with the most energy; this would improve their fitness. If all individuals are able to distinguish between highand low-quality habitats, and assess the benefit of living in a particular habitat, then all individuals should move to the high-quality habitats, which are represented by a green line in Figure 11.16. However, as more and more individuals choose the high-quality habitat, the resources available must be divided among many individuals. This reduces the resources available to each individual, also known as the per capita benefit. At some point, the per capita benefit in the high-quality habitat falls so low that an individual would be better off if it moved over to the low-quality habitat, which is indicated by the orange line in Figure 11.16. Continued increases in the population size would continue to add individuals to both the high- and low-quality habitats such that individuals in both habitats would experience the same per capita benefit. When all individuals have perfect knowledge of habitat variation and they distribute themselves in a way that allows them to have the same per capita benefit, we call it the ideal free distribution.
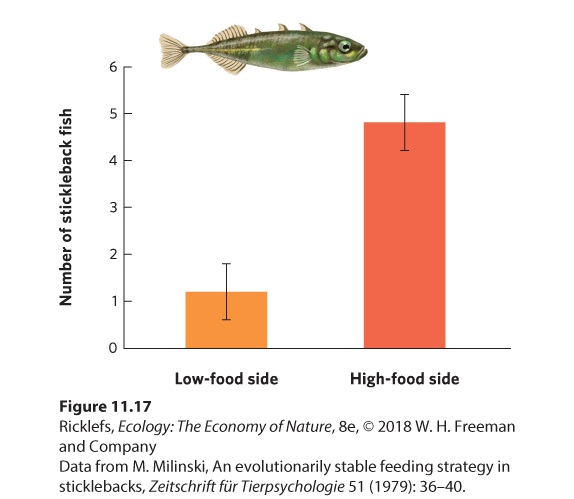
Figure 11.16 The ideal free distribution. Given the existence of high- and low-quality habitats, the first individual to arrive should select the high-quality habitat. As more individuals choose the high-quality habitat, however, the per capita benefit of the habitat declines. At some point (point A), it is equally beneficial for an individual to move to the low-quality habitat (point B) because the per capita benefit equals that experienced in the high-quality habitat. Although the low-quality habitat will have fewer individuals than the high-quality habitat, all individuals will experience the same per capita benefits. Ideal free distribution When individuals distribute themselves among different habitats in a way that allows them to have the same per capita benefit. An example of ideal free distribution can be seen in an experiment in which stickleback fish (Gasterosteus aculeatus) were presented with highand low-quality habitats. Researchers placed different numbers of water fleas (Daphnia magna), which are a type of plankton that is a food source for stickleback fish, at opposite ends of an aquarium, thereby creating habitats of different quality at each of its ends. When the first fish were placed into the aquarium with no water fleas present, the fish were distributed equally between the two halves of the aquarium. Then researchers added 30 water fleas per minute to one end of an aquarium and 6 per minute to the other, a ratio of five to one. The results of this experiment are shown in Figure 11.17. Within 5 minutes, the fish had distributed themselves between the two halves
in a ratio that was approximately four to one, which is close to the five to one ratio predicted by an ideal free distribution. When the provisioning ratio was changed—that is, the high- and low-quality patches were switched between ends of the aquarium—the fish quickly adjusted their distribution ratio. How they achieved this ideal free distribution was not determined, but they may have used the rate at which they encountered food items, or perhaps the number of other fish close by, as cues to patch quality.
Figure 11.17 The ideal free distribution in stickleback fish. When six fish were placed in an aquarium with the two ends receiving a five-to-one difference in food rations, the fish distributed themselves in a four-to-one ratio. As a result, each fish received a similar per capita amount of food. Error bars are standard deviations. The ideal free distribution tells us how individuals should distribute themselves among habitats of differing quality, but individuals in nature rarely match the ideal expectations. In some cases, individuals may not be aware that other habitats exist. Also, the fitness of an individual is not determined solely by maximizing its resources. Other factors that influence an individual’s use of a particular habitat include the presence of predators or of a territory owner that precludes moving into a high-quality habitat.
When reproductive success has been measured in the field, ecologists commonly find that individuals living in high-quality habitats have higher reproductive success, while individuals in the poorest habitats do not produce enough offspring to replace themselves. As a result, the populations living in the high-quality habitats are a source of dispersing offspring that move to low-quality habitats, which allows populations in low-quality habitats to persist. A study of the blue tit (Parus caeruleus), a small songbird from southern Europe, helps to illustrate this point. The blue tit breeds in two kinds of forest habitat, one dominated by the deciduous downy oak (Quercus pubescens) and the other by the evergreen holm oak (Quercus ilex). Long-term studies in southern France have revealed that the downy oak habitat produces more caterpillars, an important food item for the blue tits. This difference in caterpillar availability is reflected in the population densities of the birds. As you can see in Figure 11.18, the downy oak forest supports more than six times as many adult breeding pairs as the holm oak forest. Moreover, each breeding pair produces about 60 percent more offspring per year in the downy oak forest than in the holm oak forest. If we assume that juvenile survival is 20 percent in the first year and that annual adult survival is 50 percent—values that are typical of temperate songbirds—the population in the downy oak habitat would experience an annual growth of 9 percent per year if the surplus individuals did not disperse out of the habitat. At the same time, the population in the low-quality, holm oak habitat would experience an annual decline of 13 percent per year if no birds emigrated from the high-quality habitat. This would cause the population in the low-quality habitat to rapidly go extinct. In reality, the populations persist over time because the lowquality habitat receives surplus individuals that disperse in from the highquality habitat.
Figure 11.18 The effects of habitat quality. In blue tit songbirds, deciduous downy oak forests support many more adult breeding pairs than evergreen holm oak forests. In addition, birds living in the downy oak forest produce about 60 percent more fledglings per nest.
Conceptual Models of Spatial Structure
The ideal free distribution describes how individuals should distribute themselves when they are aware of the quality of different habitats and can freely move among them. However, as mentioned, it is not always easy to move from one patch of habitat to another. In such scenarios, most individuals in the patch stay put, only occasionally dispersing to other habitat patches. As a result, the larger population is broken up into smaller groups of conspecifics that live in isolated patches, called subpopulations. Subpopulations When a larger population is broken up into smaller groups that live in isolated patches. When individuals frequently disperse among subpopulations, the whole population functions as a single structure and they all increase and decrease in abundance synchronously. When dispersal is infrequent, however, the abundance of individuals in each subpopulation can fluctuate independently of one another. In considering the spatial structure of subpopulations, ecologists have devised three models: the basic metapopulation model, the source–sink metapopulation model, and the landscape metapopulation model. We will explore these models in more detail in Chapter 13. The Basic Metapopulation Model The basic metapopulation model describes a scenario in which there are
patches of suitable habitat embedded within a matrix of unsuitable habitat. All suitable patches are assumed to be of equal quality. Some suitable patches are occupied while others are not, although the unoccupied patches can be colonized by dispersers from occupied patches. The basic metapopulation models emphasize how colonization and extinction events can affect the proportion of total suitable habitats that are occupied. Basic metapopulation model A model that describes a scenario in which there are patches of suitable habitat embedded within a matrix of unsuitable habitat. The Source–Sink Metapopulation Model The source–sink metapopulation model builds on the basic metapopulation model and adds the reality that different patches of suitable habitat are not of equal quality. As we saw in the study of blue tits, it is common that occupants of high-quality habitats are a source of dispersers. Such populations are referred to as source subpopulations. At the same time, there can be lowquality habitats that rarely produce enough offspring to produce any dispersers. These habitats depend on outside dispersers to maintain the subpopulation. These subpopulations are known as sink subpopulations. Source–sink metapopulation model A population model that builds on the basic metapopulation model and accounts for the fact that not all patches of suitable habitat are of equal quality. Source subpopulations In high-quality habitats, subpopulations that serve as a source of dispersers within a metapopulation. Sink subpopulations In low-quality habitats, subpopulations that rely on outside dispersers to maintain the subpopulation within a metapopulation. The Landscape Metapopulation Model The landscape metapopulation model is even more realistic than the source–sink model because it incorporates differences in the quality of the suitable patches and the quality of the surrounding matrix. As we discussed earlier in this chapter, the habitat in the surrounding matrix can vary in quality for dispersing organisms. For example, for a regional population of
collared lizards to persist and grow requires both high-quality glade habitats and a high-quality matrix of open forest that allow for dispersal. Similarly, consider the challenge that metamorphosing frogs encounter when they move from their natal pond. They face risks of both predation and desiccation. Moving through a grassy field poses a much higher risk of predation and drying out than moving through a humid forest. Although neither field nor forest is a suitable habitat for the frog to reproduce, each is a dispersal barrier. The landscape model, which is illustrated in Figure 11.19, represents the most realistic, yet also the most complex, spatial structure of populations.
Figure 11.19 The landscape metapopulation model. The landscape model contains the most realistic conditions of habitat patches that differ in quality and a surrounding matrix that is variable in its suitability for dispersal and the presence of dispersal barriers. Landscape metapopulation model A population model that considers both differences in the quality of the suitable patches and the quality of the surrounding matrix. Throughout this chapter, we have examined the spatial structure of populations by considering the characteristics of abundance, density, and geographic range. We have also focused on the importance of suitable habitats and dispersal and how ecological niche modeling can help us predict the distributions of populations in the future. In doing so, we have largely ignored the fact that populations increase and decrease in abundance, but in the next chapter, we will take an in-depth look at how populations grow and how this growth is regulated.
Concept Check
1. Why do many species fail to exhibit an ideal free distribution? 2. What reality does the source–sink metapopulation model involve that the basic metapopulation does not? 3. What reality does the landscape metapopulation model include that the source-sink metapopulation does not?
Concepts
The Invasion of the Emerald Ash Borer The emerald ash borer. After being accidentally introduced to the United States in the 1990s, the insect has decimated ash tree populations. In 2002, a beautiful green insect, the emerald ash borer (Agrilus planipennis), was observed for the first time in North America. A native of eastern Asia, it had never lived
in North America and was unfamiliar to most people. Over the next decade, its population grew and the emerald ash borer became one of the most numerous invasive pests on the continent. The expanse of inhospitable oceans had previously prevented the emerald ash borer from dispersing to North America. Researchers suspect that the insect arrived in the 1990s from Asia by accidentally hitching a ride inside wooden crates that were shipped from Asia to Detroit, Michigan. Once the insects arrived in Michigan, they found an abundant supply of their favorite food: ash trees. The adult insect does little harm to ash trees, but it lays eggs in the cracks of the tree’s bark. When the eggs hatch, the larvae live in clustered distributions and they consume the underlying cambium and phloem of the trees. Because the cambium is essential to tree growth and the phloem is essential for transporting nutrients, the consumption of this tissue by the larvae causes an ash tree to die within 2 to 3 years. Mature larvae metamorphose into adult beetles and the cycle starts all over again.
American ash tree. The larvae of the emerald ash borer consume the underlying cambium and phloem under the bark of the trees, causing the trees to die. Midwestern forests have an abundance of ash trees, and the beetle population grew rapidly in just a few years. However, the beetle has a fairly short dispersal distance, rarely moving more than 100 m from where it first emerges as an adult, so one would expect the geographic range of this pest to expand slowly due to its dispersal limitation. But the beetle’s range expanded rapidly through unintentional human assistance. As ash trees were dying,
many of them were cut down for firewood. Much of this firewood was carried long distances for events such as camping trips, which allowed the beetle to colonize new subpopulations. In response, many states are now requiring that firewood not be transported long distances. The impact of this assisted dispersal has been dramatic. In 2012, only 10 years after its initial discovery, the emerald ash borer spread to a geographic range from Ontario and Quebec to Missouri and Tennessee and from Wisconsin and Iowa to Virginia and Pennsylvania. By 2017, the beetle had spread from Quebec and Ontario all the way down to Florida and dispersed to the west as far away as Texas, Colorado, and Minnesota. In all these states and provinces, biologists have set up traps containing beetle sex attractants to estimate the abundance and density of beetles in the area. The emerald ash borer has already killed hundreds of millions of ash trees; the financial impact is in the hundreds of millions of dollars. As a result, biologists from around North America are working rapidly to determine how far the insect may potentially spread and how it might be controlled. The estimates of its spread are being made using ecological niche modeling based on the environments that the insect inhabits in Asia. Unfortunately, current models, based on the fundamental niche of the pest, predict that the beetle will be able to live across a very large geographic range that includes most of North America. However, there are several efforts to reduce this range by introducing natural enemies of the beetle from Asia, including a parasitoid wasp and a pathogenic fungus. The hope is that these enemies of the beetle will be successful in reducing its range to a much
smaller realized niche. The invasion of the emerald ash borer will likely remain a major killer of ash trees in North America for many years to come, but an understanding of its population structure has helped biologists predict its spread and develop strategies to reduce its future impact on the forest. SOURCES: Kovacs, K. F., et al. 2011. The influence of satellite populations of emerald ash borer on projected economic costs in U.S. communities, 2010–2020. Journal of Environmental Management 92: 2170–2181. Emerald Ash Borer Information Network: http://www.emeraldashborer.info/index.cfm.
Summary of Learning Objectives
11.1 The distribution of populations is limited to ecologically suitable habitats. The range of suitable abiotic conditions where individuals can persist represents the fundamental niche of a species. The subset of conditions where a species actually persists due to biotic interactions is known as the realized niche. With an understanding of the realized niche, ecologists can use ecological niche modeling to predict the areas in which a species could persist if it were to be introduced. Key Terms: Spatial structure, Fundamental niche, Realized niche, Geographic range, Ecological niche modeling, Ecological envelope
11.2 Population distributions have five important characteristics. The geographic range of a population is a measure of the total area it covers. The abundance of a population is the total number of individuals that exist within a defined area. The density of a population is the number of individuals per unit area or volume. Dispersion of a population describes the spacing of individuals with respect to one another. Dispersal is the movement of individuals from one area to another. Key Terms: Endemic, Cosmopolitan, Abundance, Density, Dispersion, Clustered dispersion, Evenly spaced dispersion, Random dispersion, Dispersal
11.3 The distribution properties of populations can be estimated. These properties are typically measured by using a variety of survey techniques, including area- and volume-based studies, line-transect studies, and mark-recapture studies. Key Terms: Census, Survey, Area- and volume-based surveys, Line-transect surveys, Mark-recapture survey, Lifetime dispersal
11.4 Population abundance and density are related to geographic range and adult body size. There is generally a positive relationship between the abundance of a population and the size of its geographic range, although many other biotic and abiotic factors play a role in determining geographic range size. There is commonly a negative relationship between adult body size and the density of a population because larger individuals require more energy.
11.5 Dispersal is essential to colonizing new areas. Many populations do not inhabit suitable habitats because they are dispersal limited. One of the key ways to facilitate dispersal is through the creation of habitat corridors. Key Terms: Dispersal limitation, Habitat corridor
11.6 Many populations live in distinct patches of habitat. The ideal free distribution makes predictions about how individuals should distribute themselves if they were to equalize the per capita benefits, although this is rarely observed in nature due to the importance of other factors, such as predators and territoriality. Ecologists have used three types of population structure models: the metapopulation model, the source–sink model, and the landscape model. Key Terms: Ideal free distribution, Subpopulations, Basic metapopulation model, Source–sink metapopulation model, Source subpopulations, Sink subpopulations, Landscape metapopulation model
Critical Thinking Questions
1. Why is the realized niche considered to be a subset of the fundamental niche? 2. How might we use an ecological niche model to predict the future spread of the emerald ash borer? 3. The American bullfrog is native to eastern North America, but it has been moved by humans and now thrives in western North America. What does this suggest about the cause of the bullfrog’s historical geographic range? 4. With continued global warming, what dispersal barriers might arise for reptiles that try to move to higher latitudes? 5. In a forest, why might you observe young maple trees exhibit a clustered pattern of dispersion? 6. Suppose that 100 cattle were allowed to graze in either one of two pastures. If the grass was three times more productive in pasture A than in pasture B, how many cattle would be in each pasture if they followed an ideal free distribution? What might prevent this distribution of cattle from happening? 7. How does knowledge of the landscape metapopulation model provide guidance in how we need to preserve habitat for species? 8. You gather a sample of 20 horseshoe crabs, and mark release them. If you return the following week and collect 30 horseshoe crabs and 6 possess marks, how large would you estimate the crab population to be? 9. Why do birds have their highest densities near the center of their geographic range? 10. Why might you expect to find a negative correlation between adult body size and population density in fish?
#### Graphing the Data: an Ideal Free Distribution
GRAPHING THE DATA An Ideal Free Distribution Using the data for high- and low-quality habitats, create a graph that represents how per capita benefit changes as a function of the number of individuals in the patch. Based on these data, how many individuals must move to the highquality patch before an individual moves to the low-quality patch? If there were 12 individuals and they followed an ideal free distribution, approximately how many individuals would be in each habitat? Number of Individuals High-Quality Patch Per Capita Benefit Low-Quality Patch Per Capita Benefit 10.0 5.0 7.9 4.3 6.5 3.7 5.4 3.2 4.5 2.9 3.9 2.6 3.4 2.4 3.0 2.2 2.7 2.0 2.4 1.9