13 Population Dynamics over Space and Time
13Population Dynamics over Space and Time The wolves and moose of Isle Royale in Lake Superior. For more than 50 years, researchers have tracked large swings in moose and wolf population sizes. Monitoring Moose in Michigan Isle Royale, which is part of the state of Michigan and located off the northern shore of Lake Superior, has long served as a natural laboratory for ecologists. In the early 1900s, the island was colonized by moose from the mainland. This unlikely feat probably occurred during winter when the lake was frozen and the moose could travel the 40 km from the shores of the province of Ontario in Canada. The moose found an abundance of food on the island, and during subsequent decades the population grew rapidly. For more than 100 years, ecologists have watched the number of moose fluctuate widely due to changes in food, predators, and pathogens.

The initial changes in the population size of moose were driven by changes in the abundance of food. The population grew to approximately 3,000 animals by the early 1930s, but in 1934, when the population finally exceeded the island’s carrying capacity, the moose suffered starvation and many died. In 1936, the island experienced extensive forest fires that stimulated new plant growth and, as a result, the moose population began to increase. By the late 1940s, the moose once again exceeded their carrying capacity and experienced another decline due to starvation. The population would probably have continued this cycle of growth and decline, but sometime in the late 1940s, gray wolves (Canis lupus) arrived on Isle Royale by crossing the ice during the winter. Beginning in 1958, ecologist David Mech and his colleagues started estimating the number of moose and the number of wolves on the island. This was a unique research opportunity. Because both immigration and emigration of the wolves and moose were extremely rare, the researchers could watch the dynamics of the two populations unfold over time. They have estimated the wolf and moose populations every year since 1958 and have continued to make new discoveries. During the 1960s and early 1970s, there were about 24 wolves on the island while the moose population increased from about 600 to 1,500. In the 1970s, there was a series of winters with deep snow that made it difficult for the moose to move around and locate food. At this time, the population of wolves grew to 50 animals. These two factors caused a sharp drop in the moose population. “There has been tremendous public debate regarding what to do about the rapidly declining wolf population on Isle Royale.” In 1981, a lethal virus (canine parvovirus) arrived on the island, probably carried to the island by a visitor’s domestic dog. The virus
caused the wolf population to experience a major decline, dropping from approximately 50 to 14 animals. With fewer wolves on the island, the moose population rebounded and it continued to grow. In 1996, the moose once again exceeded their carrying capacity and experienced widespread starvation, causing the population to drop from approximately 2,400 to 500 individuals. From 1997 to 2011, the number of moose fluctuated between 500 and 1,000, while the number of wolves fluctuated between 15 and 30. By 2016, only two wolves remained. During the past few years, there has been tremendous public debate regarding what to do about the rapidly declining wolf population on Isle Royale. Some people have argued that we should let nature take its course. Their position is that wolves were not originally on the island, so it is acceptable if wolves become extinct on the island. Other people argue that global warming has made it less common for ice to form between the mainland and the island, so the wolves are less likely to naturally recolonize. This group believes that because the problem was caused by humans, humans should solve it by introducing more wolves to the island. Still others argue that our attention needs to be on the whole ecosystem; bringing in more wolves will help keep the moose population in check and prevent the moose from decimating the plants on the island. After considering all these positions, the National Park Service, which manages the island, announced in 2016 that they would introduce 20 to 30 wolves to the island over a period of 3 years. The story of the moose and wolf populations on Isle Royale during the past 100 years highlights the fact that populations in nature often experience large fluctuations in size. These fluctuations can have many different causes that operate either separately or together. In this chapter, we will examine how populations fluctuate over space and time. Understanding the causes of natural fluctuations of organisms in nature enables us to predict the future abundances of populations and the species with which they interact. SOURCES: Daley, J. 2016, Park Service may boost wolf pack on Isle Royale. Smithsonian, December 23.
http://www.smithsonianmag.com/smart-news/park-service-may-boost-wolf-pack-isle-royale- 180961562/. Peterson, R. O. 1999. Wolf-moose interaction on Isle Royale: The end of natural regulation? Ecological Applications 9: 10–16. Vucetich, J. A., and R. O. Peterson. 2004. The influence of top-down, bottom-up and abiotic factors on the moose (Alces alces) population of Isle Royale. Proceedings of the Royal Society of London-Series B 271: 183–189. Vucetich, J. A., and R. O. Peterson. 2011. Ecological Studies of Wolves on Isle Royale: Annual Report 2010–2011, School of Forest Resources and Environmental Science, Michigan Technological University.
Learning Objectives
After reading this chapter, you should be able to:
13.1 Recognize that population size fluctuates naturally over time.
13.2 Explain how density dependence with time delays can cause population size to be inherently cyclic.
13.3 Describe how chance events can cause small populations to go extinct.
13.4 Illustrate how metapopulations are composed of subpopulations that can experience independent population dynamics across space. In the previous chapter, we saw that populations can be regulated by densitydependent and density-independent factors. In this chapter, we look at the causes of variation in population size, an issue that has become increasingly relevant as many species dwindle toward extinction and human activities fragment habitats into ever smaller, more isolated patches. Because these fluctuations include both random and cyclical changes over time, we will incorporate them into population models. We will also investigate how population fluctuations can cause small populations to be more prone to extinction. With this understanding of population variation over time, we will move on to examine population fluctuations over space by expanding our
discussion of metapopulations from Chapter 12 to include metapopulation models and current research in metapopulations.
#### 13.1 Populations Fluctuate Naturally over Time
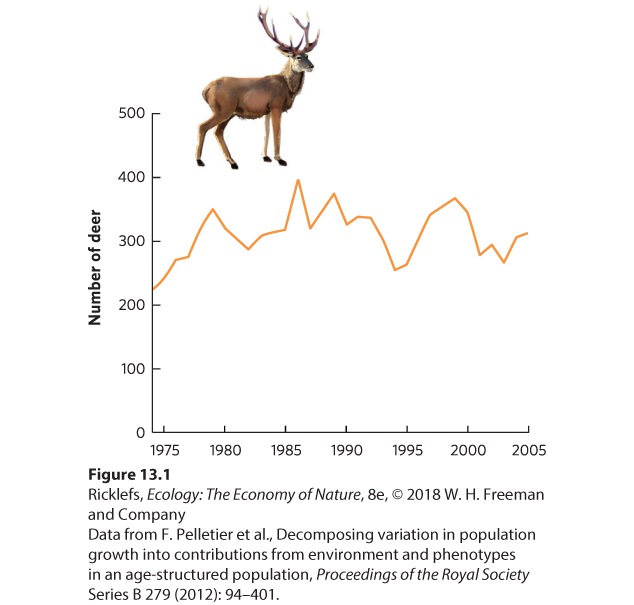
13.1 Populations fluctuate naturally over time All populations experience fluctuations in size over time. There are many reasons for these fluctuations, such as changes in the availability of food and nesting sites, predation, competition, disease, parasites, weather, and climate. Despite these fluctuations, some populations tend to remain relatively stable over long periods. For example, fluctuations in a population of red deer—a close relative of the North American elk—on the Isle of Rum in Scotland have been followed for over 30 years. During that time, the population has been relatively stable, fluctuating between approximately 200 and 400 individuals, as shown in Figure 13.1.
Figure 13.1 Population size fluctuations in red deer on the Isle of Rum. From 1974 to 2005, the population of the deer herd was relatively stable, ranging from approximately 200 to 400 individuals. Other populations exhibit much wider fluctuations. In Lake Erie, for
example, researchers monitored how the numbers of waterborne algal cells, known as phytoplankton, fluctuate throughout the year. As Figure 13.2 shows, the algae population fluctuated widely from nearly 0 cells per cubic centimeter in June to more than 7,000 cells per cubic centimeter in September. These wide fluctuations occurred across scales of days, weeks, and months.
Figure 13.2 Population size fluctuations in the algae of Lake Erie. The number of algal cells in the water has fluctuated widely over time. The populations of green algae and diatoms that compose the phytoplankton varied by several orders of magnitude on a scale of days, weeks, and months. Red deer populations are inherently more stable than algae. We can explain this by looking at differences in body size, population response time, and sensitivity to environmental change. Small organisms such as algae can reproduce in a matter of hours, which means that their populations respond very quickly to both favorable and unfavorable environmental conditions. Their small bodies and the associated high surface-area-to-volume ratio cause them to be much more affected by unfavorable environmental changes, including abiotic conditions such as temperature. As a result, their rates of survival and reproduction can decline quickly. With larger animals that live
for several years and have longer generation times, a population at a given time includes individuals born over a long period, which tends to even out the effects of short-term fluctuations in birth rate. In addition, organisms with large body sizes can maintain homeostasis in the face of unfavorable environmental changes and therefore have higher survival rates.
Fluctuations in Age Structure
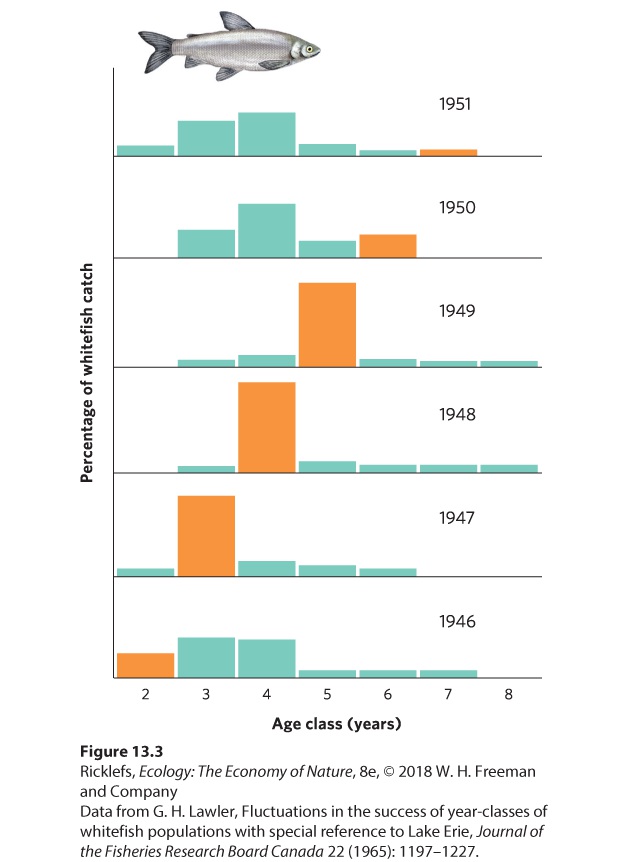
We have observed population fluctuations over time by looking at surveys of population size. In some cases, however, we can detect fluctuations over time by examining a population’s age structure. When a certain age group contains an unusually high or low number of individuals, it suggests that the population experienced unusually high birth or death rates in the past. A classic example of age structure as an indication of population fluctuations comes from data of the commercial harvest of whitefish (Coregonus clupeaformis) in Lake Erie from 1945 to 1951. Biologists determined the age of each fish by examining the scales of the harvested fish. As Figure 13.3 shows, in 1947, there was a particularly large number of 3year-old whitefish. This suggests that in 1944 the whitefish population experienced a very high rate of reproduction. This cohort continued to dominate the population’s age structure in subsequent years, with high numbers of 4-year-old fish in 1948 and 5-year-old fish in 1949. Even in 1950, the 6-year-old age class contained more individuals than had been observed in that age class in previous years. From these data, we can see that a year of high birth or low death continues to show up as an abundant age class for many years.
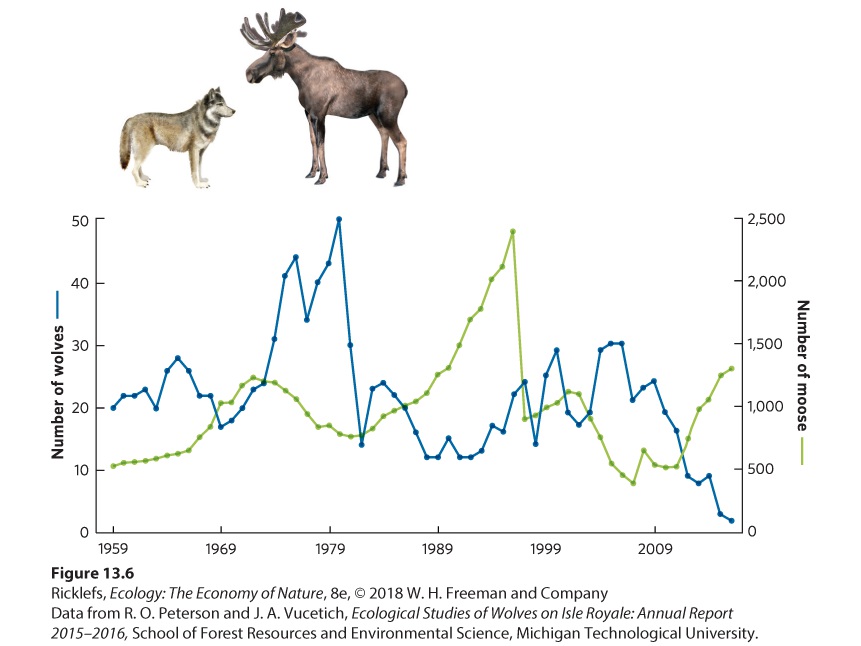
Figure 13.3 Whitefish age structure. The whitefish population in Lake Erie experienced an unusually high amount of reproduction in 1944. However, young fish are not captured by fishing nets until they are 2 years old, so the large 1944 cohort was not detected until 1946. This large reproduction event led to age structures that were dominated by that cohort of fish in subsequent years. The age structure of a forest is also readily analyzed. As you may know, the age of a tree can be determined by counting the number of rings in its trunk; under most circumstances, one ring is added each year. To examine how fluctuations affect the age structure of forest populations, researchers
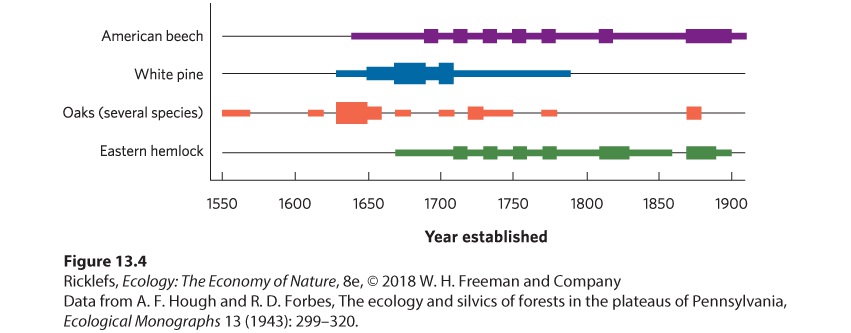
work at sites that have not been logged, because logging alters the age structure of the current tree population. In one such study, a small area of oldgrowth forest in Pennsylvania, named Hearts Content, has been protected from logging for more than 400 years. Researchers drilled into the trunks of trees to remove a sample of wood that contained tree rings. Using these samples, they determined the age of each tree and, therefore, the time when each tree started its life. The data from this survey of trees are illustrated in Figure 13.4. The data revealed that in the 1500s the forest was largely composed of several species of oak trees. A fire and drought in the mid-1600s created openings in much of the forest. Following the fire, oak seedlings reestablished themselves in the forest, although there was also an increase in the proportion of new white pines (Pinus strobis), which grow well under low-shade conditions. As the white pines grew large, however, they cast so much shade that new white pine seedlings had a hard time surviving, leading to a decline in the recruitment of white pine during the late 1700s. As a result, most white pine trees in the modern forest dated from the late 1600s and early 1700s. In contrast, American beech trees (Fagus grandifolia) and eastern hemlock trees (Tsuga canadensis) are very tolerant of high-shade environments; new individuals of these species began to grow as the white pines came to dominate and shade the forest. These two species continued to recruit new individuals over time, which caused beech and hemlock to have a population age structure that was more evenly distributed than that of white pine.
Figure 13.4 Age structure in an old-growth forest. By dating the ages of existing trees in an old-growth forest in Pennsylvania, researchers could determine the years in which populations added individuals. For simplicity, not all species of trees are shown. The thickness of the bars indicates the relative abundance of each species.
Overshoots and Die-Offs
In Chapter 12, we saw that populations experiencing density dependence grow quite rapidly at first, but then the growth rate slows as the population reaches its carrying capacity. However, populations in nature rarely follow a smooth approach to their carrying capacity. In many cases, they grow beyond their carrying capacity, a phenomenon referred to as an overshoot. An overshoot can occur when the carrying capacity of a habitat decreases from one year to the next. For example, if one year has abundant rainfall and the next year has a drought, the habitat will produce less plant mass in the second year. As a result, the carrying capacity for herbivores that rely on the plants for food will be reduced. A population that overshoots its carrying capacity is living at a density that cannot be supported by the habitat. Such populations experience a die-off, which is a substantial decline in population density that typically goes well below the carrying capacity. Figure 13.5 illustrates an overshoot and die-off in a population of a hypothetical organism. The population begins small but grows at such a rapid rate that it exceeds the carrying capacity. Following the overshoot, the population experiences a rapid die-off to a point that is below the carrying capacity. We saw an example of this in the story at the beginning of this chapter that described wolves and moose on Isle Royale. Over time, the moose exceeded their carrying capacity and the population underwent massive die-offs from starvation. You can see these data in Figure 13.6.
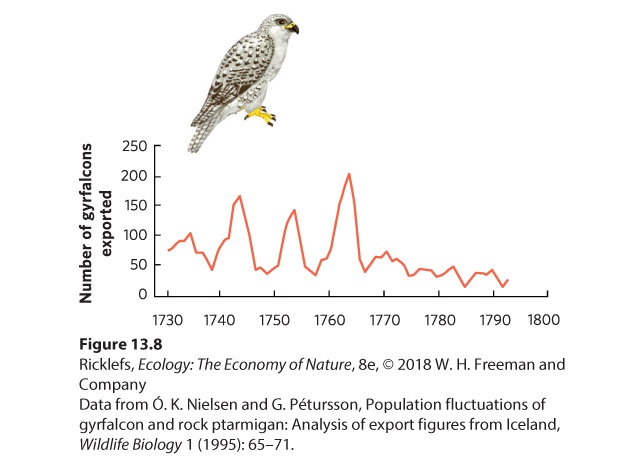
Figure 13.5 Population overshoots and die-offs. Some populations can overshoot their carrying capacity either because the carrying capacity has decreased or because the population can increase by large amounts in a single breeding season. Populations that overshoot their carrying capacity subsequently experience a die-off that causes a rapid decline in the population.
Figure 13.6 The population dynamics of wolves and moose on Isle Royale. Population estimates from 1959 to 2016 indicated that moose and wolf populations both experience wide fluctuations.
Overshoot When a population grows beyond its carrying capacity. Die-off A substantial decline in density that typically goes well below the carrying capacity. An experiment with reindeer in Alaska is another good example of overshoots and die-offs. In 1911, the U.S. government introduced 25 reindeer to St. Paul Island, Alaska, to provide a source of meat for the local population. The island contained no predators of reindeer, and the reindeer population quickly began to reproduce. The reindeer fed on a variety of items during the spring, summer, and fall, but they relied on lichens to get them through the winter. By 1938, the reindeer population had swelled to more than 2,000 individuals. As you can see in Figure 13.7, this rate of growth followed a J-shaped growth curve that indicates exponential growth. As the population grew in the early years, the lichens they consumed in the winter remained abundant. However, with continued growth of the reindeer population, the lichens became rare, which suggests that the reindeer had far exceeded the carrying capacity of the island. Following a peak in 1938, the reindeer population began to experience a massive die-off, probably from a combination of scarce winter food and unusually cold winters. In 1940 and 1941, the government culled several hundred reindeer in an attempt to reduce the size of the herd to get it closer to the island’s now much-reduced carrying capacity. Despite this effort, the population continued to decline and, by 1950, only 8 individuals remained. In 1951, 31 new reindeer were brought to the island to supplement the population. Since 1980, the population has been managed by the St. Paul Tribal Government, which maintains the population at just a few hundred animals. In 2017, the population was estimated to be 400 reindeer. The reindeer, maintained at a sustainable population size, can now continue to provide valuable and affordable meat to the local residents.
Figure 13.7 The overshoot and die-off of a reindeer population. A herd of 25 reindeer was introduced to St. Paul Island in Alaska in 1911. The population experienced a rapid increase in size that approximates an exponential J-shaped growth curve, shown by the dashed black line. After growing to nearly 2,000 animals in 1938, the population crashed, probably because the animals exhausted the food supply.
Concept Check
1. What is the relationship between species’ life spans and the degree to which their population size fluctuates over time? 2. What insights can we gain about population dynamics when we examine fluctuations in age structure? 3. What are the causes of population overshoots and die-offs?
#### 13.2 Density Dependence with Time Delays Can Cause Population Size to Be Inherently Cyclic
13.2 Density dependence with time delays can cause population size to be inherently cyclic As we have just seen, populations can experience large fluctuations over time, including overshoots and die-offs. But regular patterns known as population cycles can be seen in some populations studied over many decades. Population cycles Regular oscillation of population size over a long period of time. An interesting example of a population cycle comes from long-term records on the gyrfalcon. Falconry was a popular pastime among European nobility during the seventeenth and eighteenth centuries, and the gyrfalcon was especially prized because it is one of the largest and most beautiful falcons. During this time, Danish royalty imported gyrfalcons from Danish territories in Iceland, where the falcons were trapped and then transported to Copenhagen. The Danish royalty would then present the falcons as diplomatic gifts to the royal courts of Europe. The governor of Iceland wrote the export permits, which provide a detailed historical record of gyrfalcon exports over several decades. As shown in Figure 13.8, the number of gyrfalcons exported from Iceland between 1731 and 1793 reveals 10-year cycles of abundance, which reflects natural fluctuations in the abundance of the gyrfalcons in nature. After 1770, falconry became less popular, so exports declined to very low levels regardless of the bird’s abundance in nature.
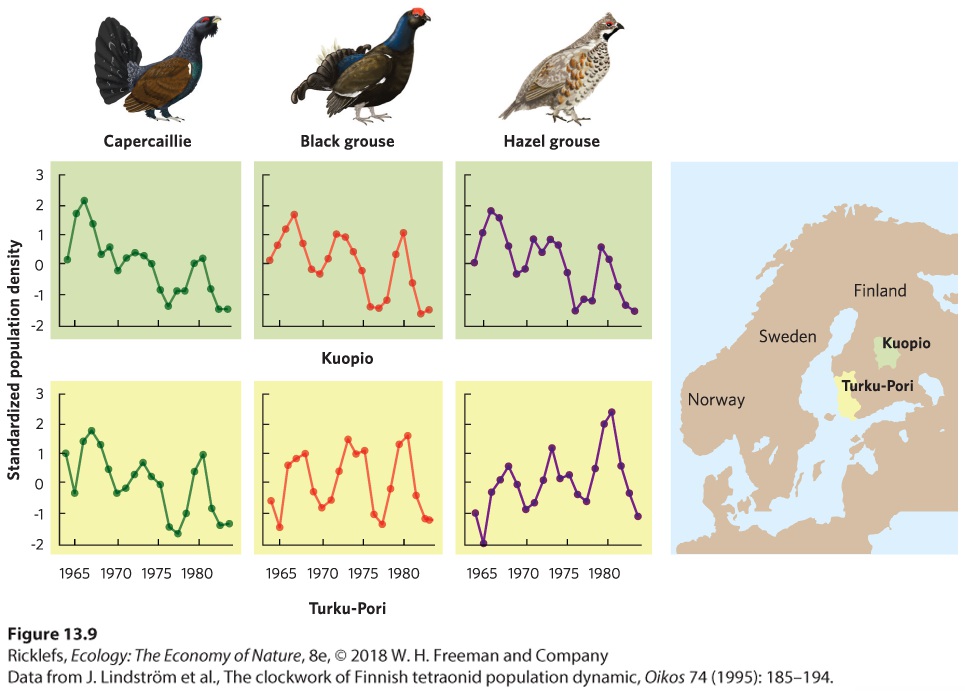
Figure 13.8 Cyclic fluctuations in gyrfalcon’s population size. From 1730 to 1770, falconry was popular in Denmark and the number of falcons captured and exported from Iceland showed regular cycles that occurred approximately every 10 years. After 1770, the popularity of falconry declined and the low demand for falcons could be met even in lowpopulation years. In some cases, cyclic population fluctuations occur among species and across large geographic areas. In Finland, for example, biologists conducted annual surveys in 11 provinces to determine the abundance of three species of grouse: capercaillie (Tetrao urogallus), black grouse (Tetrao tetrix), and hazel grouse (Bonasa bonasia). After monitoring the birds for 20 years and graphing the data—as shown in Figure 13.9—they found that all three species experienced natural population cycles every 6 to 7 years. Moreover, these three species appear to exhibit high and low populations at the same time and across all the provinces. This suggests that the drivers of natural population cycles can happen over large areas.
##### The Cycling of Populations Around Their Carrying Capacities
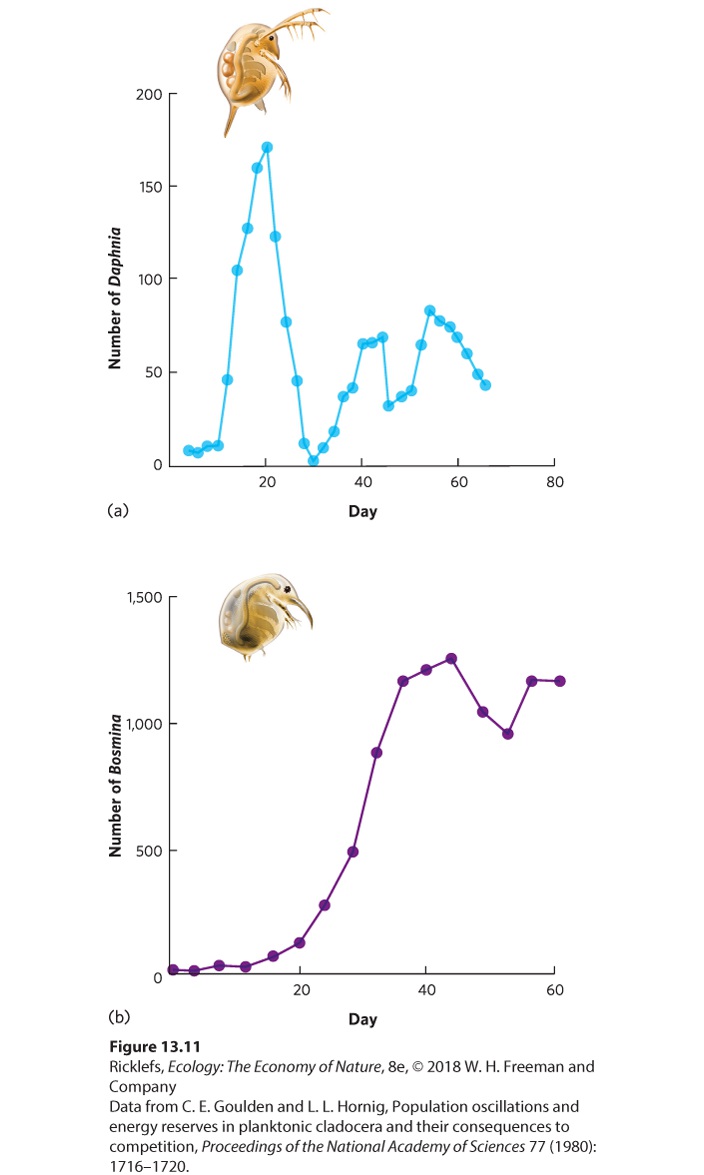
Figure 13.9 Cyclical fluctuations of population size in grouse species in two different provinces of Finland. The three species of grouse experience cycles every 6 to 7 years and appear to fluctuate in synchrony with each other and across the provinces. These examples demonstrate that some populations can exhibit remarkably regular fluctuations. The cause of such cycles, and of their synchrony, has been an interesting and persistent question in ecology. In this section, we will examine the inherent cycling behavior of populations that can be caused by fluctuating resources. In doing so, we will apply a modified version of the logistic growth equation. In subsequent chapters, we will look at how interactions with other species also influence population cycles.
Capacities
As we have just seen, populations have an inherent periodicity and tend to fluctuate up and down, although the time required to complete a cycle differs among species. To help us understand such behavior, we can think of a population as being analogous to a swinging pendulum. We know that a pendulum is stable when hanging straight up and down. The pull of gravity will cause any movement of the pendulum to the right or left to move back toward the center. However, since this movement back to the center has momentum, the pendulum overshoots the stable, center position and swings

to the other side. Gravity then pulls it back toward the center, where momentum once again causes it to overshoot the stable, center position. Populations behave like the pendulum; the momentum of increases and decreases in a population causes it to oscillate. Populations are stable at their carrying capacity. Whenever the carrying capacity increases or the size of the population decreases due to predation, disease, or a density-independent event, the population responds by growing. If the growth is sufficiently rapid, the population can grow beyond its carrying capacity. We see this phenomenon when there is a delay between the initiation of breeding and the time that offspring are added to the population. Populations that overshoot their carrying capacity subsequently experience a die-off that causes the population to swing back toward its carrying capacity. Because death rates are high and birth rates are low, the population can experience a large reduction and undershoot its carrying capacity.
Delayed Density Dependence
We can model population cycles by starting with the logistic growth model introduced in Chapter 12. You may recall that this model incorporates density dependence, which causes population growth rates to slow as the population increases in size. When density dependence occurs based on a population density at some time in the past, it is called delayed density dependence. The key to modeling delayed density-dependent populations cycles is to incorporate a delay between a change in carrying capacity and the time the population reproduces. Delayed density dependence When density dependence occurs based on a population density at some time in the past. Delayed density dependence can be caused by any number of factors. For example, large herbivores such as moose often breed in the fall but do not give birth until the following spring. If food is abundant in the fall, the carrying capacity is high, but by the time the offspring are born in the spring, the carrying capacity of the habitat could be much lower. The offspring will still be born, but the population will now exceed the carrying capacity of the habitat because the amount of reproduction was based on the earlier environmental conditions. We can also think about time delays for predators.
When predators experience an increase of prey, their carrying capacity increases. However, it may take weeks or months for the predators to convert abundant prey into higher reproductive rates. By this time, the prey may no longer be abundant. The lack of prey will cause the carrying capacity of the predator to decline just as the predator population is increasing. In both scenarios, the population experiences a time delay in density dependence. Modeling Delayed Density Dependence Delayed density dependence can be modeled with a modified form of the logistic growth model equation. As you may recall, the logistic model uses the following equation: dNdt=rN(1-NtK) where dNdt is the rate of change in the population size, r is the intrinsic growth rate, Nt is the current size of the population at time t, and K is the carrying capacity. To incorporate a time delay, we begin by defining the amount of time delay as τ, which is the Greek letter tau. Now we can rewrite the logistic growth equation by making the density-dependent part of the equation (i.e., the portion of the equation in parentheses) based on the population’s size (Ntτ) at τ time units in the past: dNdt=rN(1-Nt-τK) In words, this equation tells us that the population slows its growth when the population’s size, at τ time units in the past, approaches the carrying capacity. Whether a population cycles above or below the carrying capacity depends on both the magnitude of the time delay and the magnitude of the intrinsic growth rate. As the time delays increase, density dependence is further delayed, making the population more prone to overshooting and undershooting the carrying capacity. In addition, having a high intrinsic rate of growth allows a population to grow more rapidly in a given amount of time, making it more likely that the population will overshoot the carrying capacity.
Population modelers have determined that the amount of cycling in a population experiencing delayed density dependence depends on the product of r and τ, as illustrated in Figure 13.10. As you can see in Figure 13.10a, when this product is a low value (rτ < 0.37), the population approaches the carrying capacity without any oscillations. If this product is an intermediate value (0.37 < rτ < 1.57), as shown in Figure 13.10b, the population initially oscillates, but the magnitude of the oscillations declines over time, a pattern known as damped oscillations. When the product is a high value (rτ > 1.57), as shown in Figure 13.10c, the population continues to exhibit large oscillations over time, a pattern known as a stable limit cycle.
Figure 13.10 Population size cycles in models containing delayed density dependence. (a) In population models where the product of rτ is a low value (rτ < 0.37), the population approaches the carrying capacity without any oscillations. (b) When the product of rτ is an intermediate value (0.37 < rτ < 1.57), the population will exhibit damped oscillations. (c) When the product is a high value (rτ > 1.57), the population will oscillate over time as a stable limit cycle. Damped oscillations A pattern of population growth in which the population size initially oscillates, but the magnitude of the oscillations declines over time.
Stable limit cycle A pattern of population growth in which the population size continues to exhibit large oscillations over time.
Population Sizes Cycle in Laboratory Populations
Population models that incorporate delayed density dependence help us understand how time delays cause populations to oscillate in regular cycles. Although the models do not identify specific mechanisms by which time delays occur, ecologists have investigated real populations using laboratory experiments, where it is easier to observe the cycles and identify the underlying mechanisms that drive the cycles In some cases, delayed density dependence occurs because the organism can store energy and nutrient reserves. The water flea Daphnia galeata, for example, is a tiny zooplankton species that lives in lakes throughout the Northern Hemisphere. When the population is low and there is an abundance of food in a laboratory experiment, individuals can store surplus energy in the form of lipid droplets. As the population grows over time to the carrying capacity and food becomes scarce, adults with stored energy can continue to reproduce. Daphnia mothers can also transfer some of these lipid droplets to their eggs, which allows their offspring to grow well even if the carrying capacity of the lake has been exceeded. Eventually, the stored energy is used up and the Daphnia population crashes to low numbers. When the population is low, the food can once again become abundant and the cycle begins once more. You can see these oscillations in Figure 13.11a.
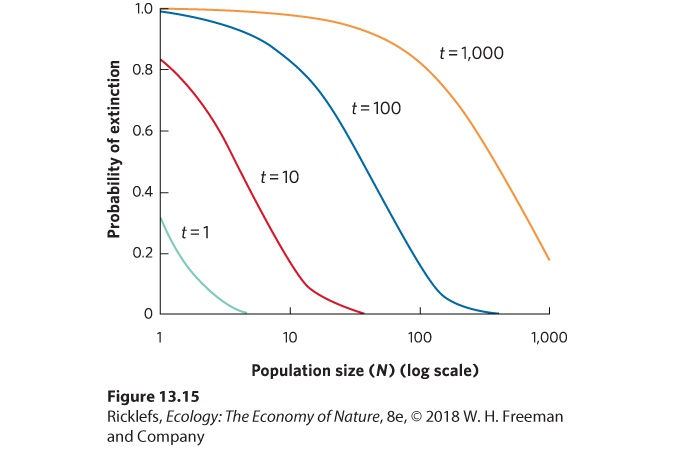
Figure 13.11 The importance of energy reserves in causing population size cycling. (a) Daphnia galeata water fleas can store high amounts of energy, which allows them to survive and reproduce even after reaching carrying capacity. When energy reserves run out, the population crashes to very low numbers and then rebounds and continues to oscillate. (b)
##### Analyzing Ecology: Delayed Density Dependence in the Flixweed
Bosmina longirostris water fleas can only store a low amount of energy, so as the population nears carrying capacity, they experience reduced survival and reproduction. As a result, the population remains near its carrying capacity and oscillates much less.
Analyzing Ecology
Delayed Density Dependence in the Flixweed Flixweed (Descurainia sophia) is a weed that is native of Europe but has been introduced into North America. Studies of this weed show that the number of plants per square meter of soil fluctuates in a cyclical manner over time. The population grows according to a delayed density dependence model, where K = 100, r = 1.1, and τ = 1. The flixweed, a common plant in Europe and North America. From plant surveys, we know that there were 10 plants per square meter in year 1 and 20 plants per square meter in year 2. Based on these data, we can calculate the expected change in population size in year 3:

dNdt=rN2(1-N1K)dNdt=(1.1)(20)(1-10100)dNdt=20 Rounded off to the nearest whole number, the flixweed will add 20 individuals to the population in year 3. Given that the population in year 2 is 20 individuals, adding 20 more individuals in year 3 will produce a total population size of 40 individuals. We can continue the calculations to determine the population size in year 4: dNdt=rN3(1-N2K)dNdt=(1.1)(40)(1-20100)dNdt=35 As we can see, the flixweed population will increase by another 35 individuals in year 4, producing a total population size of 75 individuals. YOUR TURN Using the data provided, calculate the population sizes of the plant from year 5 through year 15. Based on the product of rτ, what type of oscillating behavior do you expect to see in this population even before doing the calculations? Graph the results to confirm your prediction. We can contrast the story of the Daphnia water flea with the story of another species of water flea, Bosmina longirostris. Bosmina do not store as many lipid droplets as Daphnia, so when their population approaches the lake’s carrying capacity, they have little energy to buffer the reduction in food. As a result, Bosmina populations do not exhibit large oscillations. Instead, as illustrated in Figure 13.11b, they grow to their carrying capacity and remain there. Delayed density dependence can also occur when there is a time delay in development from one life stage to another. In a classic study of developmental delays, A. J. Nicholson examined the effect of time delays between the larval and adult stages of the sheep blowfly (Lucilia cuprina), an insect that feeds on the flesh of domestic sheep.
In his first laboratory experiment, Nicholson fed the larvae a fixed amount of food—thereby setting a carrying capacity for the larvae—but he fed the adults an unlimited amount of food. At the start of the experiment, the larvae began to metamorphose into the first set of adults, as shown in the orange line in Figure 13.12a. These adults then laid eggs that hatched into more larvae that eventually became adults. The adult fly population rapidly increased to more than 4,000 individuals.
Figure 13.12 Population size cycling in sheep blowflies. (a) When researchers limited food for larvae but not adults, they observed a delay between the time that the adults produced a large number of eggs and the time these eggs hatched into larvae, died from high larval competition, and failed to produce new adults. As a result, the adult population experienced regular cycles. (b) When adults were initially raised with unlimited food, but then given limited food halfway through the experiment, they began to experience density dependence without a time delay. As a result, the adult population still fluctuated but no longer experienced regular cycles. As the adult population increased, the unlimited food supply allowed them to continue laying eggs. The large number of eggs hatched into a large number of larvae, but because the larvae had a limited food supply, they did not grow well enough to metamorphose into adults. The larvae died and no new adults were produced. Eventually, the adult population crashed. However, before the last few adults died, they laid a small number of eggs.
When these eggs hatched, the fixed food supply provided an abundance of food for the low number of larvae. As a result, the larvae had a high rate of survival and most of them metamorphosed into adults. These new adults then laid a large number of eggs and the cycle began again. In short, there was a delay between the time that the adults produced a large number of eggs and the time these eggs hatched into larvae, died from high larval competition, and failed to produce new adults. This time delay appeared to cause the adult population cycles. Nicholson then reasoned that if limited food for offspring caused the time delay, limiting the food for the adults should eliminate the time delay and reduce the extreme fluctuations in the adult population. To test this hypothesis, he ran the experiment again by starting with unlimited adult food, but halfway through the experiment, he limited the adult food. Under these conditions, the adults experienced density dependence without delay. As you can see in Figure 13.12b, although the abundance of the adult population still fluctuated, it no longer exhibited regular population cycles. These laboratory studies confirmed that time delays between life stages caused population cycles.
Concept Check
1. Why does delayed density dependence cause population size to cycle? 2. How do we adjust the population growth model to account for delayed density dependence? 3. Why does the ability to store large amounts of energy reserves lead to delayed density dependence?

#### 13.3 Chance Events Can Cause Small Populations to Go Extinct
13.3 Chance events can cause small populations to go extinct In our models of population growth, we have seen that when populations are large, density-dependent factors cause slower growth, and when populations are small, density-dependent factors cause faster growth. Given these outcomes, it is hard to see how populations could go extinct, yet we know that extinction does occur in real populations in nature. In this section, we will explore the relationship between population size and the probability of extinction. We will then examine the underlying causes of this relationship.
Extinction in Small Populations
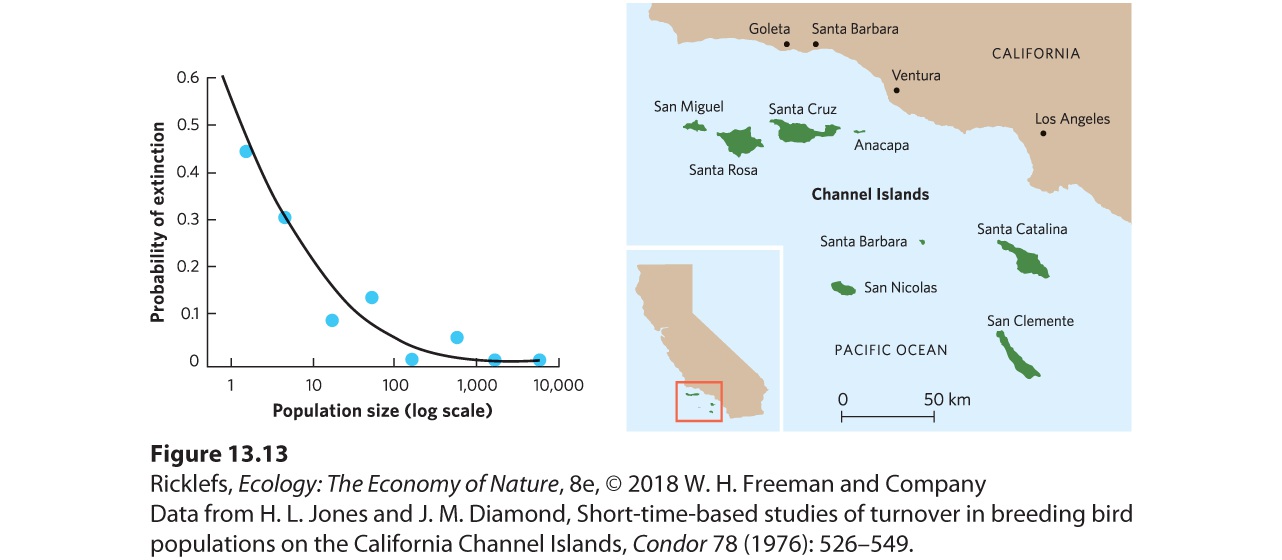
In nature, we find that smaller populations are more vulnerable to extinction than larger populations. To study this phenomenon, biologists conducted surveys of birds on the Channel Islands, which are located off the coast of California and range in size from 2.6 to 249 km2. At different times over a period of approximately 80 years, researchers studied the number of breeding pairs of different species and the extinction rates of populations on particular islands. They counted the number of breeding pairs and then determined the extinction probability for the populations. As shown in Figure 13.13, the smallest populations experienced the highest probability of extinction and the largest populations experienced the lowest probability of extinction. Similar patterns have been observed for many other groups of animals, including mammals, reptiles, and amphibians.
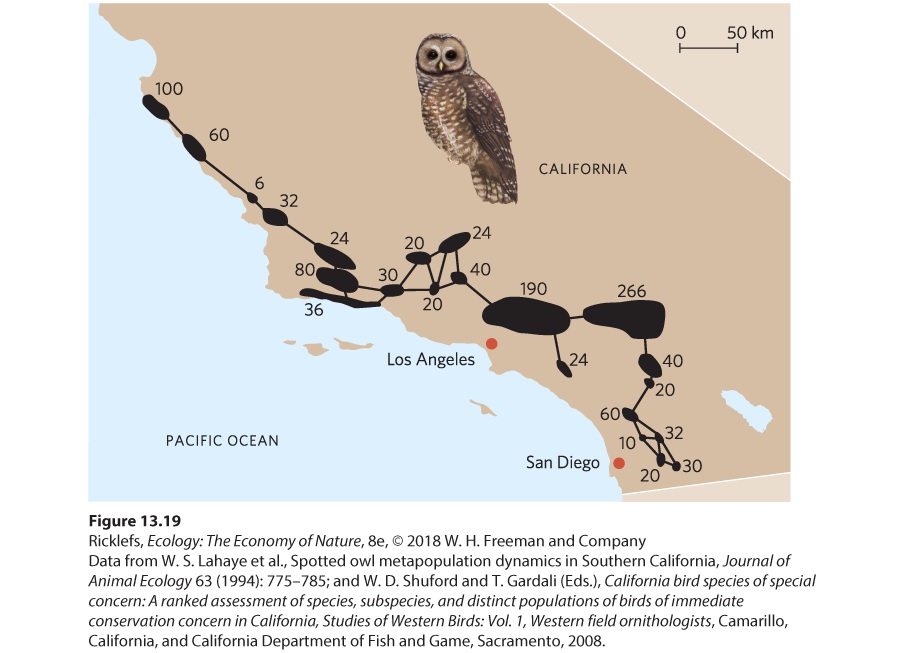
Figure 13.13 Smaller populations and the likelihood of extinction. Bird populations of the Channel Islands off the coast of California were measured in terms of the number of breeding
##### Extinction Due to Variation in Population Growth Rates
pairs. Islands with larger population sizes had a reduced probability of going extinct over a period of 80 years. The increased extinction rate of small populations is also seen in plants. For example, researchers in Germany examined the persistence of 359 populations of plants that spanned eight different species. In 1986, they counted the number of individuals in each population. They returned 10 years later and found that 27 percent of the populations had gone extinct. As you can see in Figure 13.14, when the researchers placed the populations into one of six population size categories and averaged the extinction rates across the eight species, they found that the probability of extinction was high for smaller population sizes and low for larger populations.
Figure 13.14 The probability of plant populations going extinct. Researchers surveyed 359 plant populations in Germany in 1986 and then again in 1996. They subsequently placed the populations into six size categories and averaged extinction rates across the eight species. They found that populations with the lowest number of individuals experienced the highest probability of extinction over a 10-year period.
Extinction Due to Variation in Population Growth
You have seen evidence that small populations are more likely to go extinct, yet the density-dependent population models we have examined show that small populations have more rapid growth rates than large populations. This suggests that a small population should quickly rebound and grow into a
larger population. By this reasoning, small populations should be resistant to extinction. How do we resolve the fundamental difference between these models and our observations of actual populations? The models that we have studied to this point have assumed a single birth rate and death rate for each individual in the population. When a model is designed to predict a result without accounting for random variation in population growth rate, we say it is a deterministic model. Although deterministic models are simpler to work with, not every individual in the real world has the same birth rate or the same probability of dying. Rather, because we see random variation among individuals in a population, the growth rate is not constant; it can vary over time. Models that incorporate random variation in population growth rate are known as stochastic models. When random variation in birth rates and death rates is due to differences among individuals and not due to changes in the environment, we call it demographic stochasticity. In contrast, when random variation in birth rates and death rates is due to changes in the environmental conditions, we call it environmental stochasticity. Examples of environmental stochasticity include changes in the weather, which can cause small changes in the growth rate of a population, and natural disasters, which can cause large changes in the growth rate of a population. Deterministic model A model that is designed to predict a result without accounting for random variation in population growth rate. Stochastic model A model that incorporates random variation in population growth rate. Demographic stochasticity Variation in birth rates and death rates due to random differences among individuals. Environmental stochasticity Variation in birth rates and death rates due to random changes in environmental conditions. When we use stochastic models, there is an average growth rate with some variation around that average. The actual growth rate experienced by the modeled population can take any of the values within the range of this variation. As a result, a population’s growth rate may be above or below the average growth rate. If a population experiences a string of years with above-
average growth rates, it will have faster growth. If the population happens to have a string of years with below-average growth rates, it will have slower growth. Which outcome actually occurs is determined strictly by chance. We can model stochastic population growth by modifying the exponential growth model discussed in Chapter 12. For example, we can set the birth rate and death rate to 0.5, which are reasonable values for adult mortality and recruitment in a population of terrestrial vertebrates. We can let the values for average birth and death rates vary at random. When a population randomly experiences years of low birth rates or high death rates, it is more likely to go extinct. A string of bad years can drive the population extinct faster in small populations than in large populations. Moreover, at a given population size, as time passes, there is an increased chance that a population will have a string of bad growth years and go extinct. You can see the effect of population size and time on the average probability of extinction in Figure 13.15. For example, in a population with 10 individuals, the average probability of extinction within 10 years is 0.16, the average probability of extinction within 100 years is 0.82, and extinction becomes virtually certain (0.98) within 1,000 years. In contrast, a population with an initial size of 1,000 has an average probability of extinction of only 0.18 within 1,000 years. By adding the reality of stochasticity to our population models, we see that although populations of any size eventually have some chance of going extinct, the smallest populations face the greatest risk of extinction.
Figure 13.15 Population models predict the probability of extinction. Using an exponential growth model and stochastic rates of birth and death, the average probability of extinction increases with decreasing population sizes (N) and longer times into the future (t).
1. What is the relationship between population size and probability of extinction? 2. What is the difference in approach between deterministic and stochastic population growth models? 3. What is the difference between demographic stochasticity and environmental stochasticity?
#### 13.4 Metapopulations Are Composed of Subpopulations That Can Experience Independent Population Dynamics Across Space
13.4 Metapopulations are composed of subpopulations that can experience independent population dynamics across space We have considered population dynamics over time, but population dynamics also occur over space. Whether on land or in water, species that have a particular geographic range are commonly subdivided into smaller subpopulations. This happens because preferred habitat is not continuous but occurs as patches of suitable habitat that are surrounded by a matrix of unsuitable habitat. When individuals are able to move between patches, we consider the collection of subpopulations to be a metapopulation, a concept first introduced in Chapter 11. As you may recall, the example of the collared lizards in Chapter 11 involved a metapopulation that did not live in the entire forest, but rather in glades, where temperatures were more favorable and the food supply was more abundant. In this section, we will examine why populations are broken up into metapopulations and review the three types of metapopulation models. We will then examine studies of real metapopulations in nature.
The Fragmented Nature of Habitats
Metapopulations occur when a habitat is fragmented. A good example of a fragmented habitat is the wetlands that are dotted across the landscape of many parts of North America (Figure 13.16). Each wetland can contain populations of many different species of amphibians, crustaceans, snails, and aquatic plants. The terrestrial habitat located between these wetlands is generally inhospitable to these organisms, yet individuals of many species are able to disperse across these intervening regions to arrive at another wetland. As you might guess, the amount of movement among wetlands depends on the distance between neighboring wetlands and how far and fast an individual can move.
Figure 13.16 Wetlands within a terrestrial landscape. In many regions of the world, such as this site in North Dakota, wetlands provide patches of habitat that are hospitable to aquatic organisms such as amphibians, crustaceans, snails, and aquatic plants. However, the terrestrial habitat between the wetlands is often inhospitable to these organisms. Metapopulations also occur as a result of human activities such as clearing forests, draining large wetlands, and constructing roads, housing developments, and commercial properties—all of which contribute to breaking up large habitats into a number of smaller habitats (Figure 13.17). Because the small habitats represent only fragments of the original habitat, this process is called habitat fragmentation. Small habitats typically support small populations, which, as we have seen, are more prone to extinction. However, a group of small populations that is interconnected by occasional dispersal have a unique dynamic in that dispersers can form new subpopulations. This balance of extinctions and colonizations allows the metapopulation to persist over time.
Figure 13.17 Habitat fragmentation due to human activities. Activities such as construction, harvesting of lumber, and agriculture have created fragmented habitats throughout the world. For example, the originally forested landscape on this site in Exmoor, England, is currently composed of cleared fields with fragments of forests between the fields. Habitat fragmentation The process of breaking up large habitats into a number of smaller habitats. In Chapter 11, we examined three different types of metapopulation models. The basic metapopulation model assumes that all habitat patches are equal in quality and that the habitat matrix between patches is inhospitable. The source–sink metapopulation model builds on the basic model but incorporates the fact that habitat patches differ in quality. High-quality patches are known as sources because they produce high numbers of individuals that can disperse to other patches. In contrast, low-quality patches are known as sinks because they produce few individuals and rely on dispersers coming in from other patches to keep the subpopulation from going extinct. The landscape metapopulation model is even more realistic because it acknowledges that both the patches and the habitat matrix can vary in quality. The dynamics of a metapopulation have a range of possible outcomes. If the subpopulations rarely exchange individuals, the fluctuations in abundance
will be independent among subpopulations. Some subpopulations will increase over a period of time, while others will decrease or stay relatively constant. At the other extreme, if subpopulations are highly connected by individuals frequently moving among habitat patches, the subpopulations will act as one large population, with all experiencing the same fluctuations. Between these two extremes is the scenario in which individuals occasionally move between habitat patches, such as when juvenile animals disperse away from their family to find a mate. In this case, fluctuations in abundance in one subpopulation can influence the abundance of other subpopulations.
The Basic Model of Metapopulation Dynamics
Having reviewed the spatial structure of metapopulations, we can now examine the population dynamics that occur in metapopulations. We will start with the basic metapopulation model that contains a number of simplifying assumptions. Although these assumptions are not realistic, they can help us understand the basic dynamics of a metapopulation. In this model, we begin with a population that is divided into several subpopulations, each occupying a distinct patch of habitat. We assume that these habitat patches are of equal quality, each occupied patch has the same subpopulation size, and each subpopulation supplies the same number of dispersers to other habitat patches. For the entire collection of habitat patches that exist, we will assume that some fraction of them is occupied, which we denote as p. We will also assume that there is a fixed probability of each patch becoming unoccupied— that is, that subpopulation going extinct—in a given amount of time, which we shall refer to as e. Finally, we will assume that there is a fixed probability that each unoccupied patch could be colonized, which we will denote as c. Using these variables, the proportion of occupied patches when at equilibrium, as indicated by pˆ , is given by the following equation: pˆ=1-ec This basic model indicates how the number of occupied habitat patches could increase, which would increase the total number of individuals in the metapopulation (assuming that each occupied patch has the same number of individuals). One way would be to provide corridors between neighboring
populations, thereby increasing the rate of colonization, as we saw in the case of the collared lizards. A second way would be to decrease the rates of extinction by reducing the major causes of population decline in subpopulations. The concept of the metapopulation is important for the conservation of species around the world as humans continue to fragment terrestrial and aquatic habitats. We know that a key to preserving populations is to maintain large fragments of habitat whenever possible because populations in large habitats are less likely to go extinct. When we can only preserve small fragments, we must ensure that individuals can disperse to and from them so they can be colonized, which will help prevent small, declining subpopulations from going extinct.
Observing Metapopulation Dynamics in Nature
The basic metapopulation model indicates that a metapopulation persists because of a balance between extinction of the subpopulation in some habitat patches and the colonization of others. Although these are the predictions of the model, we need to see if these processes occur in nature. One of the most extensive studies of metapopulations has been conducted on the Glanville fritillary butterfly (Melitaea cinxia) by ecologist Illka Hanski and colleagues. On the Åland Islands of Finland, this butterfly lives in isolated patches of dry meadows (Figure 13.18). The researchers found 1,600 suitable meadows on the islands, but determined that only 12 to 39 percent of these patches were occupied in any given year. Over a 9-year period, they observed that more than 100 of the occupied patches experienced extinction each year and more than 100 of the unoccupied patches experienced colonization. Based on their observations, it became clear that no single patch was safe from extinction, but the metapopulation persisted because patch extinctions were offset by continued patch colonizations.
Figure 13.18 Metapopulation dynamics of a butterfly. The Glanville fritillary butterfly lives in isolated meadows on the Åland Islands of Finland. Here, the butterflies exist as a metapopulation.
The Importance of Patch Size and Patch Isolation
We have noted that the basic model of metapopulation dynamics does not consider variations found in nature. For example, habitat patches are rarely equal in quality. Some patches are larger or contain a higher density of needed resources. In Southern California, for example, the California spotted owl (Strix occidentalis occidentalis) lives along the coast in small habitat fragments. As shown in Figure 13.19, these fragments are of very different sizes, and the estimates of how many owls could be supported in each fragment range from 6 to 266. Moreover, not all patches are equally distant from all other patches.
Figure 13.19 A metapopulation of the California spotted owl. Along the coast of Southern California, the owl lives in small fragments of forested habitat, as indicated by the black regions. The lines connecting the habitat patches indicate potential dispersal paths of owls between the patches of forest. The numbers next to each patch indicate researcher estimates of how many owls could live in each of the patches. When each patch in a metapopulation supports a different number of individuals, we expect the small patches to experience higher rates of extinction. As a result, small patches are less likely to be occupied than large patches. At the same time, we might also predict that more distant patches will have a lower probability of being occupied than closer patches. We make this prediction because successful dispersal is a function of the distance an individual has to travel. Therefore, unoccupied patches that are close to occupied patches have a better chance of being colonized. Moreover, subpopulations on the brink of extinction can be supplemented by the arrival of dispersers from other subpopulations. The phenomenon of dispersers supplementing a declining subpopulation that is headed toward extinction is called the rescue effect. These two mechanisms of colonization and the rescue effect should result in a higher probability that less isolated patches will be occupied. Rescue effect The phenomenon of dispersers supplementing a declining subpopulation that is headed toward extinction.
The effect of patch size and patch isolation on patch colonization has been tested by studying populations of the common shrew (Sorex araneus) on a series of islands in two lakes in Finland. The islands vary in size from about 0.1 to 1,000 ha, and they vary in isolation from less than 0.1 to more than 2 km from other islands or from the shore of the lake. As shown in Figure 13.20, the researchers found that shrews were much less likely to occupy the smaller islands and those that were more isolated.
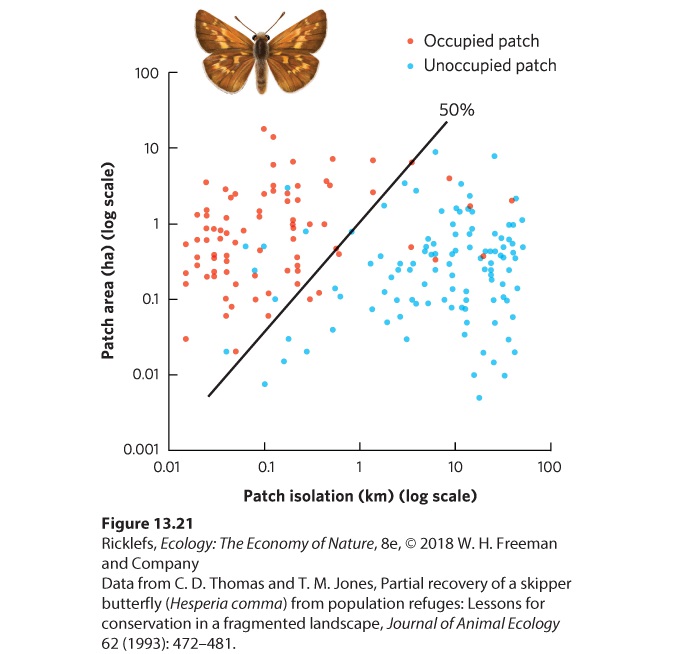
Figure 13.20 Effects of patch size and patch isolation on patch occupancy by the common shrew. The shrew was much less likely to occupy patches that were small or more isolated. Similar patterns have been found in other species and other areas of the world. In Britain, for example, skipper butterflies (Hesperia comma) prefer to live in calcareous grasslands that are heavily grazed by rabbits. The grassland patches varied from about 0.01 to 10 ha in area and the distances between patches ranged from 0.02 to 100 km. As shown in Figure 13.21, the researchers found that the largest and least isolated patches were occupied, whereas the smallest and most isolated patches were not occupied. As we will see in
Chapter 22, patch size and isolation are not only important to the metapopulation of one species but also to the total number of species that live on habitat islands.
Figure 13.21 Butterfly occupation of habitat patches that differ in size and isolation. The skipper butterfly in Britain occupies the largest patches of grasslands and those that are the least isolated. The line indicates the combinations of patch area and patch isolation that correspond to 50 percent probability of patch occupancy. Throughout this chapter, we have examined how populations vary over time and space. Fluctuations over time are quite common in nature, and some of these fluctuations exhibit cyclic patterns that are due to delayed density dependence. All populations have some chance of going extinct given enough time, but the smallest populations face a much higher risk of extinction due to demographic and environmental stochasticity. Population dynamics over time can be offset by population dynamics over space, as observed in metapopulations in which habitat fragments can experience extinctions and colonizations that allow the entire metapopulation to persist over time. As we
will see in “Ecology Today: The Recovery of the Black-Footed Ferret,” understanding this interplay of population dynamics over time and space can be critical in bringing species back from the brink of extinction.
Concept Check
1. How are human activities causing large populations to be broken up into metapopulations? 2. Why is the proportion of occupied patches in a metapopulation at equilibrium determined by the rates of patch colonization and extinction? 3. How does distance between habitable patches affect the rate of patch colonization?
Concepts
The Recovery of the Black-Footed Ferret Black-footed ferrets. An understanding of how populations fluctuate over time and space has allowed scientists to facilitate the
recovery of the black-footed ferret. The black-footed ferret (Mustela nigripes) is an elusive animal of the American West. As a nocturnal predator and a member of the weasel family, it feeds almost entirely on prairie dogs. In fact, an average ferret can consume 125 to 150 prairie dogs per year. The ferret’s life is so tightly connected to prairie dogs that it lives in old prairie dog burrows within aggregations of prairie dogs known as “prairie dog towns.” Historically, the ferret is thought to have had a geographic range that spanned from Texas to Arizona and north all the way up to the Canadian border. No data exist on historic population sizes, but their numbers were probably in the tens of thousands and likely varied in relation to the fluctuating abundance of prairie dogs. By the 1980s, however, the ferret was thought to have gone extinct in the wild. The decline of the black-footed ferret occurred over the course of more than a century. With settlement of the West in the 1800s, farmers converted a large portion of the land occupied by prairie dogs to cropland. This loss of habitat substantially reduced the carrying capacity of rangelands for prairie dogs, which in turn caused a large decline in the carrying capacity of ferrets. By the 1920s, a campaign of poisoning prairie dog towns was initiated to reduce the number of prairie dogs, which were thought to compete with cattle and sheep for rangeland plants. The poisons killed many of the ferrets, but they also died from canine distemper and sylvatic plague. Canine distemper is a viral disease that is native to North America and deadly to members of the weasel family. Sylvatic plague is a disease introduced to North
America from Asia in 1900 and is highly lethal to ferrets. A combination of habitat loss, poisoning, and disease caused prairie dog and ferret populations to decline rapidly. The situation of the ferret started to receive attention in the 1960s when a single, small population was discovered in South Dakota. In a scramble to save the species from extinction, it was classified as an endangered species in 1967, which meant that efforts would be taken to promote the recovery of the species. Nine animals were brought into captivity for a breeding program. This initial attempt at captive breeding was not very successful; a few offspring were born but none survived beyond a few days. At the same time, the habitat at the South Dakota site continued to become more fragmented. As the ferret populations became smaller, the ferret faced a higher risk of extinction due to stochastic factors. Indeed, the small ferret population in nature went extinct in 1974 and the last ferret in captivity died in 1979. Everyone thought the ferret was extinct. Two years later, a report that a rancher’s dog killed a ferret near the town of Meeteetse, Wyoming, led to the discovery of a population of more than 120 ferrets living in a nearby prairie dog town. However, in 1985, this population was infected by canine distemper and sylvatic plague and began a rapid decline. Because small populations are prone to extinction from stochastic factors, biologists captured the remaining 24 ferrets and brought them into a new captive breeding program in Wyoming. Although only 18 of the 24 ferrets survived, this time the captive breeding program was so successful that additional programs were started in other locations. These locations included several zoos to ensure against a stochastic
catastrophe at the Wyoming facility. Within 6 years of bringing ferrets into captivity, biologists began to re-introduce animals into the wild where prairie dogs were abundant. By this time, because the concept of metapopulation was much better understood, they chose to conduct several re-introductions throughout western North America. This strategy accounted for the possibility that should some subpopulations go extinct due to habitat loss, disease, or other stochastic factors, other subpopulations could persist and serve as future sources of new re- introductions. The recovery program has been an incredible success. As of 2016, ferrets have been introduced to 28 sites throughout the United States, Canada, and Mexico. In fact, the 28th site of re-introduction was in Meeteetse, Wyoming, where the last remaining population was discovered. Across their native range, their numbers have grown from the original 18 individuals that survived in captivity to hundreds of individuals today. They still face challenges of drought years causing a decline in food and wet years causing increases in fleas that carry the lethal sylvatic plague. However, the success of the recovery program reflects a tremendous effort by scientists working to identify and improve the factors that determine ferret population abundance over time and space. SOURCES: Robbins, J. 2008. Efforts on 2 fronts to save a population of ferrets. New York Times, July 15. Black-footed ferret recovery program. http://blackfootedferret.org/. Roelle, J. E., et al. (Eds.). 2006. Recovery of the black-footed ferret —progress and continuing challenges. U.S.G.S. Scientific Investigations Report 2005–5293.
Rogers, N. 2016. Black-footed ferret recovery comes full circle. http://wildlife.org/black-footed-ferret-recovery-comes-full-circle/.
Summary of Chapter Concepts
13.1 Population size fluctuates naturally over time. This phenomenon occurs because density-dependent and densityindependent factors can change from year to year and from place to place. The magnitudes of the fluctuations are often related to the ability of a species to resist changes in the environment and to differences in their life histories, including reproductive rates and life span. In some species, the population can overshoot its carrying capacity and then experience a rapid die-off. In populations that have an age structure, fluctuations in population size over time can be detected by disproportionate numbers of individuals in particular age classes. Many species experience cyclic fluctuations in population size. Key Terms: Overshoot, Die-off, Population cycles
13.2 Density dependence with time delays can cause population size to be inherently cyclic. Delayed density dependence allows populations to fluctuate above and below their carrying capacity. Delayed density dependence can be incorporated into our population models by having the population’s growth rate depend on the population density that occurred at some time in the past. Using these models, we find that the magnitude of the fluctuations depends on the product of the intrinsic growth rate (r) and the time delay (τ). Increasing values of this product causes the population to shift from experiencing no oscillations to damped oscillations to a stable limit cycle. Experiments have confirmed that time delays due to energy reserves or development times between life stages can cause cyclic fluctuations. Key Terms: Delayed density dependence, Damped oscillations, Stable limit cycle
13.3 Chance events can cause small populations to go extinct.
Smaller populations are more likely to go extinct than large populations. This occurs due to demographic and environmental stochasticity. Key Terms: Deterministic model, Stochastic model, Demographic stochasticity, Environmental stochasticity
13.4 Metapopulations are composed of subpopulations that can experience independent population dynamics across space. Metapopulations exist when a habitat exists in small fragments, either naturally or from human activities. The basic model of metapopulation dynamics informs us that metapopulations persist due to a balance between extinctions in some habitat patches and colonizations that occur in other patches. Although the basic metapopulation model assumes that all patches are equal, in reality, larger patches generally contain larger subpopulations and patches that are less isolated and more likely to be occupied as a result of both the rescue effect and higher rates of recolonization. Key Terms: Habitat fragmentation, Rescue effect
Critical Thinking Questions
1. If you examined a large number of similar species that differed in how much each species stored energy, what would be the likely relationship between the amount of stored energy and the likelihood of the species’ population growth to exhibit delayed density dependence? 2. How does your knowledge of small island populations and the importance of the rescue effect help explain the likelihood of extinction of the wolves on Isle Royale? 3. In models of delayed density dependence, why do r and τ work together to determine the magnitude of population oscillations? 4. When predator and prey populations cycle, what are the likely causes of the cycling for the prey versus the predator? 5. How should the probability of extinction due to stochastic processes vary with population size in the California spotted owl? 6. What are the differences between demographic stochasticity and environmental stochasticity? 7. Given what you now know about the decline in the wolf population on Isle Royale, formulate arguments for and against the reintroduction of wolves. 8. In a metapopulation of the collared lizards discussed in Chapter 11, how would decreasing the distance between habitat patches affect the synchrony of fluctuations among subpopulations? 9. If you were trying to save an endangered species that lives in a metapopulation, how might you attempt to increase the proportion of occupied patches? 10. In the basic model of metapopulation dynamics, how might the rescue effect alter both the probability of colonization and the probability of extinction?
#### Graphing the Data: Exploring the Equilibrium of the Basic Metapopulation Model
GRAPHING THE DATA Exploring the Equilibrium of the Basic Metapopulation Model We have seen that the basic metapopulation model allows us to calculate the proportion of occupied patches based on the probability of extinction (e) and on the probability of colonization (c). 1. Based on the probabilities of extinction and colonization provided in the table below, calculate the rates of extinction and the rates of colonization. 2. Use a line graph to plot the relationship between the proportion of occupied patches and the rate of extinction. 3. On the same graph, plot the relationship between the proportion of occupied patches and the rate of colonization. 4. Based on the two line graphs, where do the extinction rates and colonization rates come into equilibrium?
Colonization C
COLONIZATION (c × p) × (1 − p) 0.1 0.25 0.50 0.2 0.25 0.50 0.3 0.25 0.50 0.4 0.25 0.50 0.5 0.25 0.50 0.6 0.25 0.50 0.7 0.25 0.50 0.8 0.25 0.50 0.9 0.25 0.50 1.0 0.25 0.50