17 Mutualism
17Mutualism A tree shrew feeding on a pitcher plant in Borneo. The tree shrew consumes nectar from the plant and then defecates into the pitcher, which provides nitrogen-rich nutrients to the plant. Bathrooms with Benefits Pitcher plants are famous for trapping insects, which are attracted by the smell and the nectar produced by the plant. Unfortunately for the insects, the cup-shaped pitcher has a slippery rim, making it nearly impossible for insects to escape once they enter. As a result, the insects die and are subsequently digested by the plant. Through this predator–prey relationship, pitcher plants obtain the nitrogen that they need to grow and reproduce, which is critical given that they often live in habitats where soil nitrogen is quite limiting. Knowing this, imagine the surprise of researchers when they discovered that a pitcher plant in the tropical forests of Asia was being used as a toilet by the local tree shrews.

The pitcher plant (Nepenthes lowii) is endemic to the island of Borneo in Southeast Asia. Immature plants live close to the ground and trap ants and other insects, while the mature plants live higher up in the trees. In these mature plants, researchers noted that they rarely observed any trapped insects, but they did notice the presence of feces from a small mammal. To determine the identity of the mammal, they set up video cameras around mature pitcher plants during the daylight hours. When they later checked the cameras, they discovered that mountain tree shrews (Tupaia montana) were visiting the pitcher plants and licking the abundant nectar that the plant produces on the overhanging pitcher lid. To reach the nectar, the shrew has to position itself with its posterior directly over the pitcher, and as it feeds, it defecates into the pitcher. The plant can then digest the shrew feces to obtain nitrogen. In fact, the plant can obtain all its nitrogen from shrew feces. This is highly beneficial to the pitcher plants given that it lives in the mountains of Borneo where insects are not particularly abundant. To encourage shrew visits, the plan has evolved to produce very large amounts of nectar, which is important to the shrew since other sources of nectar are scarce. In addition, rather than growing a slippery cup to trap insects, the rim of the mature pitchers can be easily gripped and the cups are structurally reinforced to hold the weight of the shrew. In short, the shrews and the pitcher plants are involved in a highly unusual mutualism. The initial discovery of shrews feeding on and defecating in a species of pitcher plant in 2009 inspired much more research. In 2010, researchers discovered that two other species of large pitcher plants (N. rajah and N. macrophylla) were also visited by mountain tree shrews, and these other pitcher plants had many of the same characteristics as the first species. The following year, other researchers discovered that the pitchers, which are visited by tree shrews during the day, are also visited by summit rats (Rattus baluensis) during the night and the rats also leave their feces behind. Another group of researchers working in Borneo discovered that a tiny species of bat (Kerivoula hardwickii) was using a related species of pitcher plant (N. hemsleyana) as a roosting site. Bats were not known to roost in pitcher plants, but the structure of this species of pitcher
plant allowed one or two bats to hide inside the cup during the day, while still staying above the level of the fluid inside the cup. In exchange for providing the bats with a safe place to hide, the plants obtained nearly a third of their nitrogen requirement from the feces that the bats left behind. “To reach the nectar, the shrew must position itself with its posterior directly over the pitcher, and as it feeds, it defecates into the pitcher.” Research on the pitcher plants in Borneo has made it clear that we have a great deal to learn about how species have co-evolved mutualisms in nature. This work also underscores the fact that mutualisms are probably much more pervasive than we once thought and that many species depend on each other to ensure their growth and reproduction. SOURCES: Clarke, C. M., et al. 2009. Tree shrew lavatories: A novel nitrogen sequestration strategy in a tropical pitcher plant. Biology Letters 5: 632–635. Chin, L., et al. 2010. Trap geometry in three giant montane pitcher plant species from Borneo is a function of tree shrew body size. New Phytologist 186: 461–470. Wells, K., et al. 2011. Pitchers of Nepenthes rajah collect faecal droppings from both diurnal and nocturnal small mammals and emit fruity odour. Journal of Tropical Ecology 27: 347– 353. Grafe, T. U. 2011. A novel resource-service mutualism between bats and pitcher plants. Biology Letters 7: 436–439.
Learning Objectives
After reading this chapter, you should be able to:
17.1 Describe how mutualisms can provide water, nutrients, and places to live.
17.2 Explain how mutualisms can aid in defense against enemies.
17.3 Illustrate the role that mutualisms play in facilitating pollination and seed dispersal.
17.4 Describe how mutualisms can change when conditions change.
17.5 Explain how mutualisms can affect species distributions, communities, and ecosystems. In Chapter 1, we defined mutualism as a positive interaction between two species in which each species receives benefits that only the other species can provide. Mutualisms are common in nature: Corals live with their symbiotic algae, shrews and pitcher plants exchange nutrients, and, as we saw in
Chapter 10, leaf-cutter ants farm a fungus. In this chapter, we will see how species have evolved to participate in mutualistic interactions for a wide variety of benefits. Some species benefit by obtaining resources, while others benefit by obtaining a place to live, aid in defense, and assist in pollination or seed dispersal. When we consider mutualistic relationships, it is tempting to think that each species is trying to help the other. However, as you might recall from our review of natural selection in Chapter 1, selection favors any strategy that increases the fitness of the individual. Regardless of the specific benefit, there are requirements for the evolution of mutualistic interactions and conditions under which a positive, mutualistic relationship can change to a neutral or negative interaction. Finally, we are interested in mutualisms not only because they are a common interaction in nature, but also because they can affect the abundance of populations, the distribution of species, the diversity of communities, and the functioning of ecosystems.
17.1 Mutualisms for resource acquisition We can categorize mutualisms in several ways. For example, some mutualists are generalists, which means that one species interacts with many other species. Other mutualists are specialists, which means that one species interacts with either one other species or a small number of closely related species. When two species provide fitness benefits to each other and require each other to persist, we call them obligate mutualists. We saw an example of this in Chapter 1 when we discussed the tubeworms and chemosynthetic bacteria that live together near deep-sea hydrothermal vents. Tubeworms provide a place for bacteria to live and bacteria provide food for the tubeworms; neither species can survive without the other. In contrast, facultative mutualists provide fitness benefits to each other, but the interaction is not critical to the persistence of either species. For example, a group of tiny insects known as aphids suck the sap from plants and produce a droplet rich in carbohydrates that is consumed by several species of ants. The ants gain a source of food and in exchange they protect the aphids from predators. Although both groups benefit, it is a facultative mutualism because each can persist without the other. A mutualism between two species can be composed of two obligate mutualists, one obligate and one facultative mutualist, or two facultative mutualists. Generalist A species that interacts with many other species. Specialist A species that interacts with one other species or a few closely related species. Obligate mutualists Two species that provide fitness benefits to each other and require each other to persist. Facultative mutualists Two species that provide fitness benefits to each other, but whose interaction is not critical to the persistence of either species. One of the most common functions of mutualisms is to help species acquire resources they need, such as water, nutrients, and a place to live. In previous chapters, we have discussed a few examples of such mutualisms. In
Chapter 1, we saw that lichens are composed of a fungus living with either green algal cells or cyanobacteria (see Figure 1.13). This fungus provides the
algae with water, CO2 from fungal respiration, and nutrients, and, in exchange, the algae provide the fungus with carbohydrates from photosynthesis. Similarly, in Chapter 2, we discussed how corals provide a home for photosynthetic algae known as zooxanthellae. As you can see in
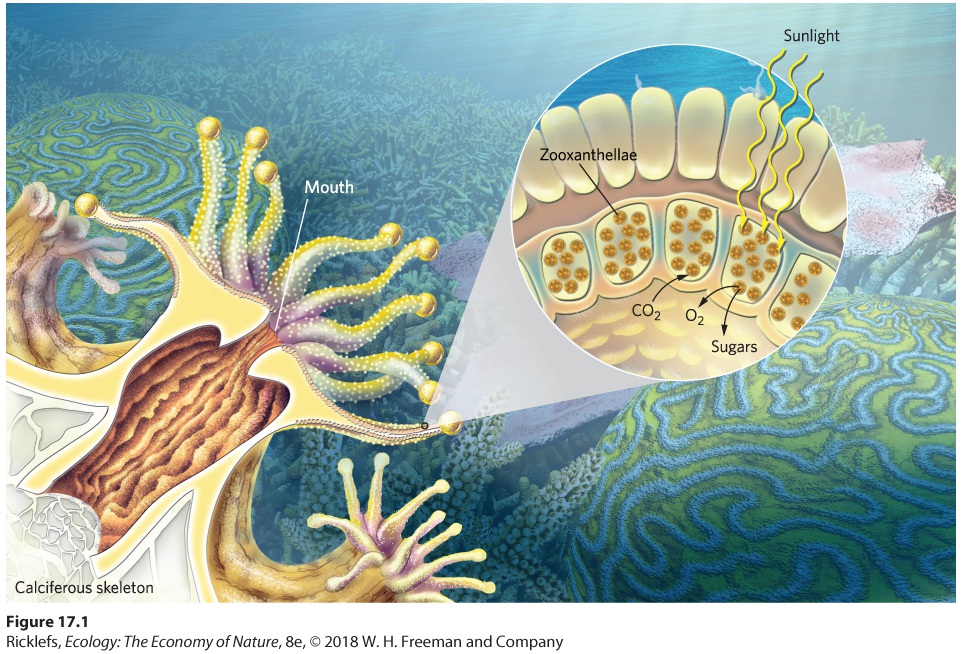
Figure 17.1, the coral catches bits of food with its tentacles and during digestion the coral emits CO2, which the algae use during photosynthesis. The algae then produce sugars and O2, some of which can be consumed by the coral. Other animals also incorporate symbiotic algae into their bodies. Similarly, in Chapter 2, we discussed how the eggs of spotted salamanders incorporate algae in the tissues of the embryo. Similarly, when the leaf sheep sea slug (Costasiella kuroshimae) consumes algae, it stores the chloroplasts from the algae inside its tissues and thus gains most of its energy through photosynthesis (Figure 17.2). In this section, we will review how several species of plants, animals, fungi, and bacteria interact in mutualisms to gain water, nutrients, and places to live.
Figure 17.1 A mutualism between coral and zooxanthellae. A coral grabs food with its tentacles, which contain stinging cells, and pulls the food into its mouth. Zooxanthellae algae live along the surface of a coral’s tentacles where they obtain sunlight for photosynthesis. Corals provide a place for algae to live and emit CO2 that algae use in photosynthesis. As algae photosynthesize, they provide sugars and O2 that corals consume.
Figure 17.2 The leaf sheep sea slug. When the slug first hatches from an egg in the ocean, it is brown. However, as it begins to eat algae, it stores the chloroplasts from the algae in its own tissues. As the slug accumulates a large number of chloroplasts, its body turns green and it is able to acquire most of its energy through photosynthesis rather than through herbivory.
Resource Acquisition in Plants
Although plants obtain water and soil minerals through their root systems, many plants also rely on mutualisms with fungi and bacteria to help them obtain nutrients. Plants and Fungi Fungi that surround plant roots and help plants obtain water and minerals are known as mycorrhizal fungi. The network of fungal hyphae provides plants with minerals, such as nitrogen and phosphorus, and water from the surrounding soil. Plants provide the fungi with the sugars they produce by photosynthesis. Because fungi can increase the amount of minerals obtained by the plants, they are able to increase the plants’ tolerance to both drought and salt stress. They can also help plants combat infections from pathogens. Mycorrhizal fungi

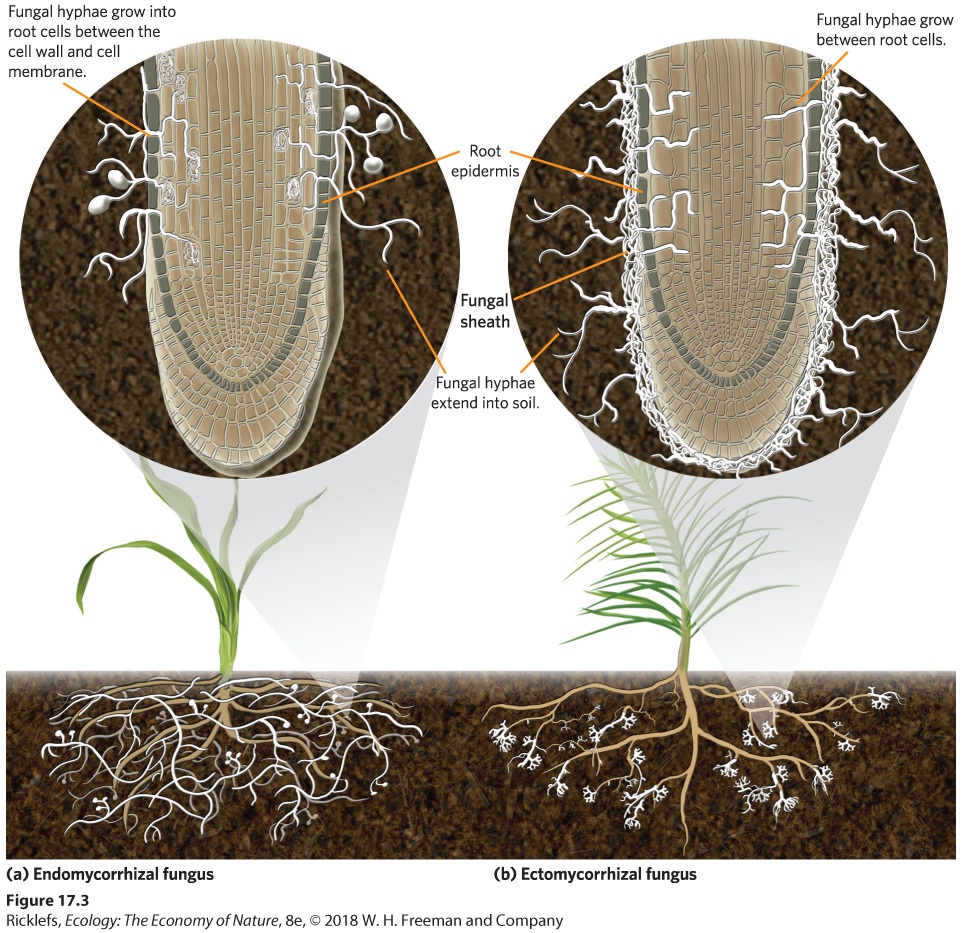
Fungi that surround plant roots and help plants obtain water and minerals. Mycorrhizal fungi can be divided into either endomycorrhizal fungi or ectomycorrhizal fungi. Endomycorrhizal fungi are characterized by hyphal threads that extend far out into the soil and penetrate root cells between the cell wall and the cell membrane (Figure 17.3a). One fungal species can commonly infect multiple plant species. There are several types of endomycorrhizal fungi. The most common type is arbuscular mycorrhizal fungi, which infect a tremendous number of plants, including grasses and apple, peach, and coffee trees. Arbuscules are branching hyphal structures found within plant cells that help the fungus provide nutrients to the plant.
Figure 17.3 Mycorrhizal fungi. Mycorrhizal fungi can be categorized as endomycorrhizal fungi or ectomycorrhizal fungi. (a) Endomycorrhizal fungi have hyphae that penetrate the root cells of plants and reside between the cell wall and cell membrane. (b) Ectomycorrhizal fungi have hyphae that do not penetrate the root cells but instead grow between the root cells of plants.
Endomycorrhizal fungi Fungi characterized by hyphal threads that extend far out into the soil and penetrate root cells between the cell wall and the cell membrane. Arbuscular mycorrhizal fungi A type of endomycorrhizal fungi that infects a tremendous number of plants, including apple trees, peach trees, coffee trees, and grasses. Ectomycorrhizal fungi, illustrated in Figure 17.3b, are characterized by hyphae that surround the roots of plants and enter between root cell walls, but they rarely penetrate between the cell wall and the cell membrane. These fungi are currently known to live only in mutualistic relationships with trees and shrubs. Species of ectomycorrhizal fungi also tend to form mutualistic relationships with fewer plant species than do endomycorrhizal fungi. Ectomycorrhizal fungi Fungi characterized by hyphae that surround the roots of plants and enter between root cells but rarely enter the cells. The mutualistic relationship between plants and mycorrhizal fungi goes back more than 450 mya to the time when plants first evolved to live on land. This ancient interaction between the ancestral plants and fungi probably explains why so many modern species of plants and fungi continue to interact as mutualists. It is estimated that mutualism between plants and fungi involves more than 6,000 species of mycorrhizal fungi and 200,000 species of plants, which is about two-thirds of all plant species. Plants and Bacteria In some cases, mutualistic interactions between plants and bacteria convert unusable forms of minerals into forms that plants can use. One of the bestknown examples is the group of bacteria in the genus Rhizobium that live in a mutualistic relationship with numerous species of legumes, including important crops such as beans, peas (Pisum sativum), and alfalfa (Medicago sativa). When legumes detect the presence of Rhizobium bacteria in the soil or when bacteria enter the plant through an opening in the root, the plant develops small nodules that surround the bacteria on the roots and provide them with a place to live (Figure 17.4). Plants also provide the bacteria with the products of photosynthesis. In exchange, the bacteria do something the
plant cannot do; they convert atmospheric nitrogen—a form of nitrogen plants cannot use—into ammonia, a form of nitrogen plants can readily use. This mutualism can be quite valuable to plants, especially when they are living in areas of low soil fertility. We will discuss this phenomenon in more detail in Chapter 21.
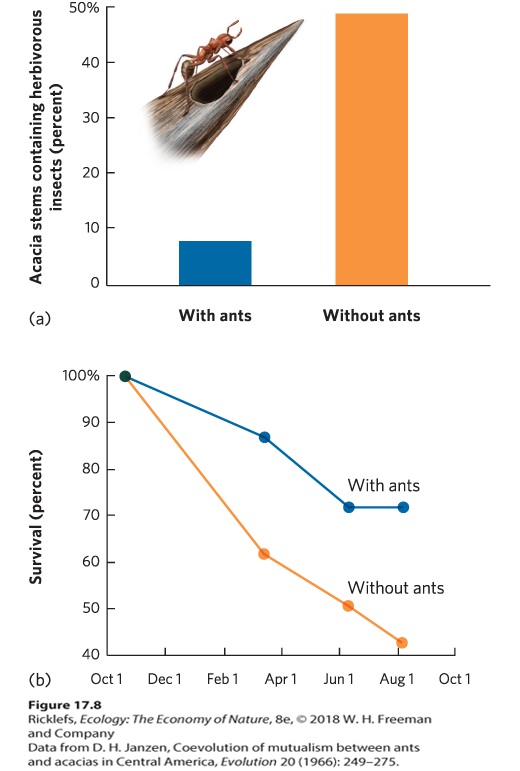
Figure 17.4 Root nodules that contain Rhizobium bacteria. Legumes such as this bean plant can enter into a mutualistic relationship with Rhizobium bacteria. The plant provides the bacteria with sugars from photosynthesis and a root nodule in which the bacteria can grow. In return, the bacteria convert atmospheric nitrogen to ammonia, a form of nitrogen that the plant can use.
Resource Acquisition in Animals
Animals also use a variety of organisms to help them obtain food, water, and habitat. These interactions range from protozoans living in animals to mutualisms between animals. Animals and Protozoans

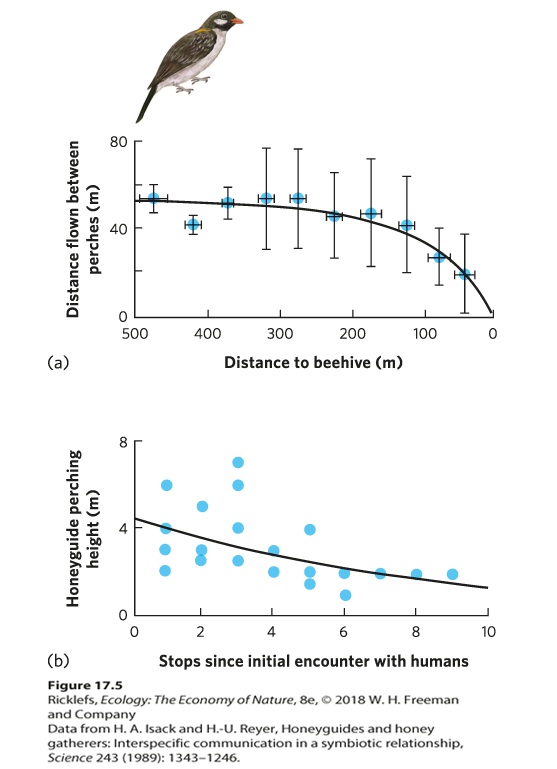
Termites are a group of insects that consume wood, which is difficult to digest since it is composed largely of lignin and cellulose. To assist in this effort, species of protozoa that are able to consume lignin and cellulose live in termite guts. In the gut, the protozoa receive a constant source of food from wood that the termite consumes and, in exchange, the termite receives nutrients from the waste products of protozoan digestion. Many other animals also contain microbes in their digestive system. Humans, for example, host hundreds of species of microbes—bacteria, fungi, and protozoa—that largely seem to be beneficial. In fact, a person’s digestive system contains 10 times more bacterial cells—from more than 500 bacterial species—than the total number of human cells in that person’s body. Animals and Other Animals Mutualisms for acquiring resources can also occur between two species of animals. A fascinating example occurs between humans and a bird known as the greater honeyguide (Indicator indicator). For centuries, people in Africa have consumed the honey produced by bees, but locating the beehives is a challenge. While the greater honeyguide likes to consume bee larvae and bee’s wax, it has a hard time getting into beehives. The local people and the honeyguide both obtain resources by working together. Over time, local people have learned to use calls to attract the attention of the bird and then follow the bird to the beehive. Along the way, the bird stops to perch in nearby trees. As the bird gets closer to the beehive, it flies shorter distances from one perch to the next and perches lower in trees, as shown in Figure 17.5. Local people have learned how to interpret the bird’s behavior and they follow it to the hive. When they find the hive, they scoop out the honey and leave pieces of the honeycomb with beeswax and bee larvae on the ground for the honeyguide to consume. It is thought that the honeyguide may have originally evolved this behavior as a mutualism with other honey-consuming mammals, such as the honey badger (Mellivora capensis).
Figure 17.5 The greater honeyguide. When the greater honeyguide leads humans or other honey-consuming mammals to a beehive, the bird benefits by consuming the discarded honeycombs that contain beeswax and bee larvae. Humans have learned that as the greater honeyguide gets close to a beehive, (a) the bird flies shorter distances between stops and (b) at each stop it perches lower on trees. Error bars are standard deviations. Some animals provide a habitat for other animals in exchange for reciprocal benefits, as we saw in the case of the pitcher plants and the roosting bats. For example, alpheid shrimp live in the ocean and have very poor vision. They burrow into the sand and allow a group of fish known as
gobies to share their burrows. In contrast to shrimp, gobies have excellent vision and are able to see shrimp predators. In exchange for receiving a burrow, a goby allows a shrimp to stay in close contact by permitting the shrimp to place an antenna on it once the goby leaves the burrow. If the goby sees a predator, it warns the shrimp by twitching. The shrimp detects the twitching via its antenna and heads back into the burrow for protection (Figure 17.6).
Figure 17.6 Habitat mutualist. The alpheid shrimp (Alpheus randalli) digs a burrow that it shares with a fish known as the pinkbar goby (Amblyeleotris aurora) in the waters off Indonesia. In exchange for this habitat, the goby allows the shrimp to place an antenna on its body so that the shrimp can feel the goby twitching when it sees an approaching predator.
Concept Check
1. If facultative mutualists do not require another species to help them, why do they engage in mutualism? 2. What benefits do mycorrhizal fungi provide for plants? 3. Why do termites require a mutualism with gut protists?

#### 17.2 Mutualisms Can Aid in Defense Against Enemies
17.2 Mutualisms can aid in defense against enemies As we saw with the alpheid shrimp, mutualisms can help a species defend itself against enemies. To obtain a defense benefit from a mutualistic partner, an organism must provide some type of benefit in return. In this section, we will examine a number of ways that mutualisms have evolved to benefit both the species being defended and the species providing the defense.
Plant Defense
Plants are involved in a number of mutualisms that help defend them from enemies. Two well-known examples are the mutualisms between ants and acacia trees and the mutualisms between fungi and plants. Acacia trees are found in tropical forests throughout the world; they face a variety of herbivores and numerous competitors, including vines that try to wrap around the acacia’s branches. In Central America, acacia trees are commonly inhabited by ants in the genus Pseudomyrmex that constantly patrol the tree branches. The acacia trees provide large thorns with pulpy centers that ants hollow out and convert into nests. The trees also contain nectaries, which produce nectar that the ants consume (Figure 17.7a). In exchange, these ants bite and sting any herbivores—from small insects to large mammals—that attempt to consume the leaves. The ants also eliminate plants that attempt to grow near their home tree by chewing on them until they die (Figure 17.7b).

Figure 17.7 A mutualism between ants and acacia trees. (a) Acacia trees, such as this swollen thorn acacia in Panama, have large brown thorns that the ants can hollow out for nests where they can raise their larvae. The plants also have green nectaries that provide nectar the ants consume. In exchange for these benefits, the ants attack herbivores that try to
eat the acacia tree and they attack vines and other plants that grow near the acacia tree. (b) Because the ants attack encroaching plants, acacia trees with ants are typically surrounded by bare ground. In a classic study, Dan Janzen compared acacia trees that had ants living on them and acacia trees from which he removed the ants. As you can see in
Figure 17.8a, the trees with ants had a lower percentage of herbivorous insects than trees with no ants. In addition, trees with ants grew to be 14 times heavier than trees without ants. When Janzen monitored the trees over a 10month period, he found that trees with ants had much higher survival rates than trees without ants, as shown in Figure 17.8b. In short, Janzen demonstrated that the ants are critical to the survival and growth of the acacia trees.
Figure 17.8 The effect of ants on acacia trees. (a) When ants were removed from acacia trees, the trees experienced a large increase in the number of herbivorous insects feeding on them. (b) As a result, the removal of ants caused much lower survival of the acacia trees. More recently, researchers have discovered that the ant‒acacia mutualism has additional benefits. In 2010, it was reported that the chemicals in the nectaries contain numerous proteins with antibacterial properties. To
determine if ants helped to distribute these chemicals on the leaves, the researchers used two sets of trees: one group had the ants present and the other group had the ants removed for 2 weeks. Researchers also examined the effect of one ant species that acted as a mutualist (P. ferrugineus) and another ant species that did not (P. gracilis). When P. ferrugineus was present, the plant leaves were nearly free of bacteria, but when P. gracilis was present, there was no significant reduction in bacteria. The researchers do not yet know how the P. ferrugineus causes a reduction in leaf bacteria, but it seems likely that P. ferrugineus is distributing some of the antibacterial nectar to the leaves, whereas P. gracilis does not. These results demonstrated that ants in a mutualistic relationship with trees not only defend the tree against herbivores and competitors but appear to also defend the tree against bacteria that act as plant pathogens. You can view the results of the experiment in Figure 17.9.
Figure 17.9 Reduction in bacteria by mutualistic ants. The presence of the mutualistic ant (P. ferrugineus) causes a large reduction in the abundance of bacteria on the leaves of the acacia tree. In contrast, the presence of a nonmutualistic ant (P. gracilis) has no effect on bacterial abundance. Error bars are standard errors. Some plants can defend themselves from herbivores through mutualisms
with fungi, known as endophytic fungi, which live within the plant’s tissues. These fungi produce chemicals that can repel insect herbivores and also provide drought resistance by increasing the concentration of minerals in plant tissues, which increases the plant’s ability to absorb and retain water from the soil. In exchange, the plant supplies the fungi with the products of photosynthesis. While endophytic fungi can be beneficial to plants, some of the chemicals they produce can be quite harmful to herbivores. For example, when the grass known as tall fescue (Festuca arundinacea) contains endophytic fungi, the fungi produce chemicals that are highly toxic to cattle, sheep, goats, and horses. Research is currently focused on identifying alternative strains of fungi that can offer defense against insect herbivores and drought resistance without being toxic to livestock. Endophytic fungi Fungi that live inside a plant’s tissues.
Animal Defense
Animals also participate in mutualisms that aid their defense. An excellent example occurs in a group of fish known as cleaner wrasse. These tiny fish spend their life consuming ectoparasites that are attached to other, much larger fish (Figure 17.10). As the cleaner wrasse approaches, the larger fish opens its mouth and flares its gills to permit access to the many parasites that are attached to its body. The number of parasites removed can be substantial; a single cleaner wrasse can consume more than 1,200 parasites per day. The cleaner wrasse benefits from having a large source of food and the larger fish benefit by having fewer parasites.

Figure 17.10 Cleaner wrasse. Off the coast of Hawaii, the Hawaiian cleaner wrasse (Labroides phthirophagus) removes parasites from a yellowfin goatfish (Mulloidichthys vanicolensis). A similar situation exists for large terrestrial animals in Africa. Two species of birds, the red-billed oxpecker (Buphagus erythrorhynchus) (Figure 17.11). and the yellow-billed oxpecker (B. africanus), perch on the backs of grazing animals such as rhinos and antelopes The birds consume ticks that are attached to the backs of mammals. However, because the birds also peck at the wounds caused by the ticks, scientists wondered whether the birds were mutualists or parasites. If oxpeckers act primarily as mutualists, their preferences for certain species of grazing mammals should be related to the number of ticks carried by each species. Alternatively, if the birds act primarily as parasites that seek to peck at mammal flesh, they should prefer mammals with thinner hides, which their beaks can penetrate more easily. A recent study examined these relationships in both species of oxpeckers using up to 15 species of grazing mammals in Africa. The researchers quantified the oxpecker preferences for different species of mammals by observing grazing mammals of several species and dividing the number of oxpeckers on a given species by the total number of species grazing. Then they quantified the abundance of ticks on an individual of each mammal species. Although they found no relationship between the species preferences of oxpeckers and the thickness of the animals’ hides, they found positive correlations between
the species preferences of oxpeckers and tick abundance, as you can see in
Figure 17.12. These results suggest that the birds are acting primarily as mutualists whose preferences are geared toward feeding on ticks. Thus, the benefits of tick removal for the grazing mammals likely outweigh the cost of having their flesh pecked at by the birds.
Figure 17.11 Oxpeckers. On the African savanna, oxpeckers, such as this red-billed oxpecker in Kruger National Park in South Africa, remove ticks from a variety of grazing mammals, such as this impala (Aepyceros melampus).
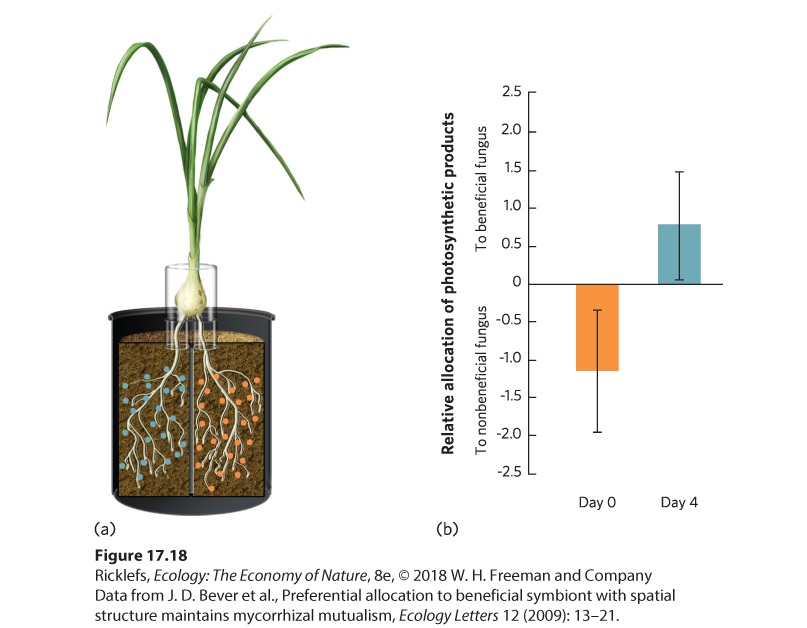
Figure 17.12 Oxpecker preferences for different species of mammals. The preferences of red-billed oxpeckers and yellow-billed oxpeckers are positively correlated to the abundance of ticks on the different mammals. Neither bird showed a preference for mammals with thin hides. These data suggest that oxpeckers seek out particular species of mammals primarily to consume ticks as a mutualist rather than to consume bits of mammal flesh as a parasite.
Concept Check
1. What defense benefits do ants provide to acacia trees? 2. How do endophytic fungi help defend plants against herbivores? 3. What is the evidence that oxpeckers act as mutualists, not as parasites, with large grazing animals?
#### 17.3 Mutualisms Can Aid with Pollination and Seed Dispersal
17.3 Mutualisms can aid with pollination and seed dispersal In addition to providing resources and defense, mutualisms can also provide the valuable services of plant pollination and the dispersal of plant seeds. Indeed, without these services many species could not reproduce or colonize areas throughout their geographic range. In this section, we will examine the various mutualisms that serve these functions.
Pollination
To make seeds, flowering plants need pollen to fertilize their ovules. Some plants, such as grasses, commonly rely on the wind to blow pollen from one plant to another. Other plants rely on animals to carry the pollen. Over evolutionary time, plants have evolved a number of reward mechanisms to entice pollinators to visit their flowers. For example, the common honeybee visits flowers that offer both nectar and pollen. The bees consume both items, but the bees also inadvertently transfer some of the pollen that is attached to their bodies as they move from flower to flower. Many plants appear to have evolved flowers that are specialized in attracting one particular type of pollinator. For example, flowers pollinated by hummingbirds tend to be red —a color hummingbirds favor. Plants pollinated by hummingbirds also tend to have long tubular flowers inaccessible to most insects. However, because hummingbirds have long tongues, they can easily reach the nectar inside. Similarly, many bat-pollinated flowers are large, are often open only at night, and contain high volumes of nectar to attract the large pollinators. In some plant species, the volume of nectar increases during the night when bats are most likely to be foraging (Figure 17.13).
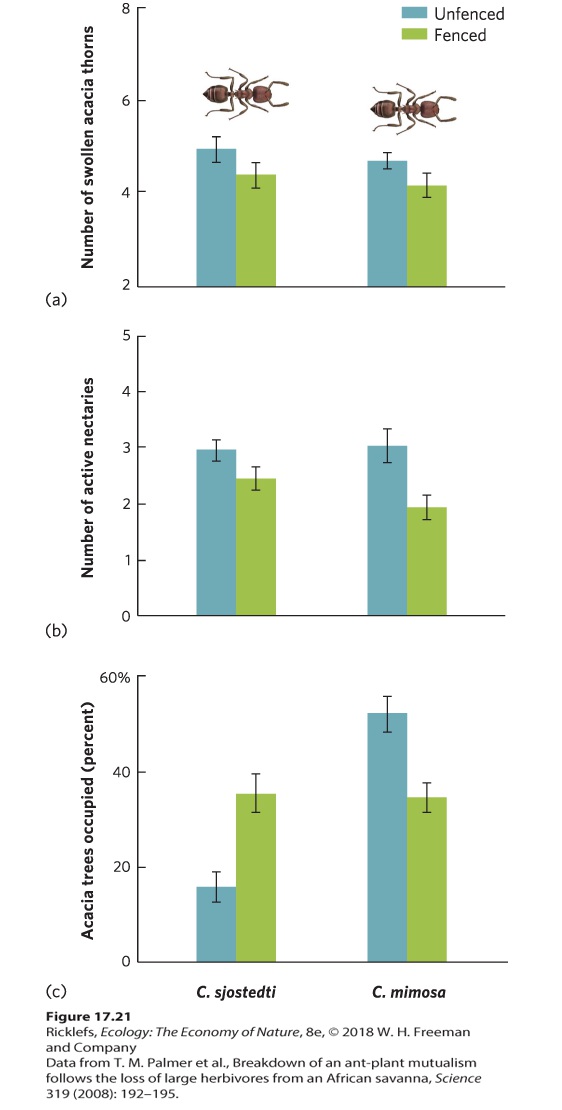
Figure 17.13 Pollination by bats. Flowers that are pollinated by bats are typically large and contain large nectar rewards to entice the bats to visit. Pictured here is a long-nosed bat (Leptonycteris curasoae) pollinating a flower of the saguaro cactus (Carnegiea gigantea). Although flowers in many species of plants can be pollinated by several different pollinator species, some plants, such as the group known as yuccas, have evolved very specific mutualisms with their pollinators. In fact, most species of yucca plants rely on a single species of yucca moth for pollination. Unlike most pollinators, however, a visit from the yucca moth is not a brief affair. A female yucca moth arrives at a yucca flower and lays eggs in the ovaries of the flower (Figure 17.14). To ensure that the flower makes seeds for her offspring to eat, the female moth climbs to the top of the flower and adds pollen grains to the flower’s stigma. These pollen grains produce long tubes that grow down through the style and into the ovary, where they deposit male gametes, which then fuse with the female gametes to produce seeds. The moth eggs hatch into caterpillars that feed on the seeds that the flower forms. As a result, the plant gains a very effective pollinator at the cost of a few
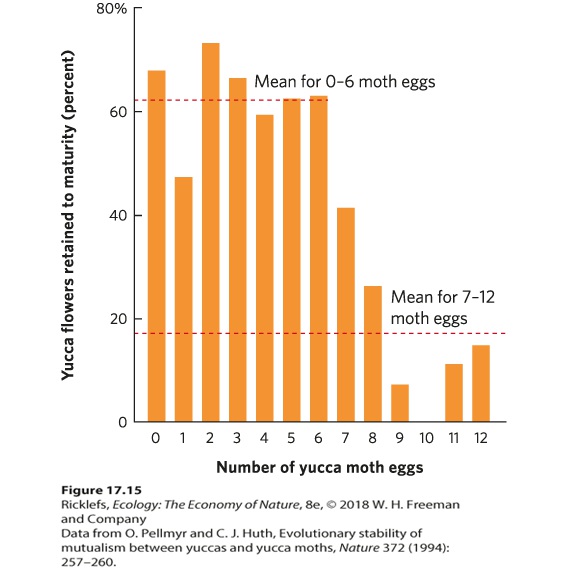
Figure 17.14 Pollination by a yucca moth. A female yucca moth transfers pollen to the style of a yucca flower after she lays her eggs in the ovary of the flower. Despite the consumption of seeds by the caterpillars, enough seeds remain for the yucca to reproduce. For many years researchers wondered what prevented the female moths from laying so many eggs that all the seeds would be consumed, leaving the flower with no fitness. They discovered that the plant is tolerant of up to six moth eggs per flower. Under these conditions, the plant develops 62 percent of its flowers to maturity. However, if there are more than six eggs per flower, some species of yuccas selectively abort most of their flowers and retain only 17 percent of their flowers to maturity, as shown in Figure 17.15. When a flower is aborted, all moth eggs and larvae within the aborted flower die. By aborting flowers whenever the moths lay more than six eggs, the plant favors those moths that do not lay too many eggs in each flower.
Figure 17.15 Controlling consumption of yucca seeds by yucca moth caterpillars. When female yucca moths lay 0 to 6 eggs in a yucca flower, the average plant develops 62 percent of its flowers to maturity. If a moth lays 9 to 12 eggs, the average plant only develops 17 percent of its flowers to maturity; the rest are selectively aborted. This response favors yucca moths that lay fewer eggs.
Seed Dispersal
The seeds and fruits produced by plants span a wide range of sizes, from tiny dandelion seeds that float on the wind to massive fruits such as coconuts. Many of the smallest seeds are easily dispersed by wind, but larger seeds are typically dispersed by animal mutualists. As we have seen, animals contribute to a mutualistic relationship if they receive a benefit in return. The most common benefit is food in the form of seeds or nutritious fruit that surrounds the seeds. When animals eat the fruits of plants, the seeds commonly pass through the digestive system unharmed and are viable after being excreted. However, when animals eat the seeds of plants, the seeds are digested and are no longer viable.

It may seem like a contradiction that plants would depend on seed-eating animals to help disperse their seeds. After all, if the seeds are eaten, there are no seeds left to disperse. However, in most cases, plants produce large numbers of seeds and not all seeds that are carried away get eaten. Many of them are stored in the ground, which means that the seeds are planted and able to germinate if they are not consumed. For example, squirrels store acorns in numerous locations and some of these acorns are never retrieved. Similarly, a single Clark’s nutcracker (Nucifraga columbiana)—a bird that specializes on the seeds of the whitebark pine (Pinus albicaulis)—collects approximately 32,000 pine seeds in a season and stores them in thousands of different locations. Because this number of seeds is three to five times what the bird requires to meet its energy needs, many of the stored seeds are never eaten but rather germinate into new pine trees in locations far from the parent tree. Some species of plants entice animals to disperse their seeds by surrounding them with a fruit that contains substantial nutrients. For example, many forest herbs, including the painted trillium (Trillium undulatum), produce seeds that have a lipid- and protein-rich package—known as an elaiosome—attached to the seed (Figure 17.16). Ants collect the seeds and take them back to their nests. After consuming the elaiosomes, they discard the seeds from the nest. This process disperses the seeds away from the parent plant. The ants receive nutrition and the plants receive dispersal of their seeds.
Figure 17.16 Ants eating elaiosomes. Some forest herbs produce seeds with a packet of lipid- and protein-rich tissue attached that is known as an elaiosome. Shown here are seeds of the bloodroot (Sanguinaria canadensis). The ants carry these seeds to their nest, consume the elaiosome, and discard the seeds outside of the nest where they can subsequently germinate. When plants surround their seeds with a large fruit, the fruit can be a substantial reward to animal dispersers. If the seeds inside the consumed fruit have a hard coat that resists digestion, they pass through the animal’s digestive system and are still able to germinate. One of the most striking examples is the African tree known as Omphalocarpum procerum. The tree produces fruits as large as a person’s head and only the African elephant can break it open. Moreover, the seeds cannot germinate in the soil unless they have passed through an elephant’s digestive system. The tree is highly dependent on elephants to consume and disperse its seeds, but as elephant populations decline, the tree is losing its only seed disperser. A key element to this strategy for seed dispersal is for the seeds to remain inedible or hidden until they are fully developed. As a result, many fruits are green and relatively camouflaged while the seeds are developing. At this stage, the fruit is typically quite unpalatable. However, once the seeds have fully developed, the fruit becomes ripe, the tissues of the fruit become palatable, and it commonly changes color to be highly visible to animal dispersers.
1. Why have some plant species evolved to have flowers of particular sizes and shapes? 2. How might an animal’s storing of a large number of seeds create a mutualism between the animal and the plant? 3. How does the consumption of fruit by an animal result in improved seed dispersal for a plant?
#### 17.4 Mutualisms Can Change When Conditions Change
17.4 Mutualisms can change when conditions change Although individuals of two or more species might interact in a way that allows both to receive fitness benefits, we need to recognize that each individual participates in the mutualism to improve its own fitness and not the fitness of its partner. Therefore, when changing conditions alter the costs and benefits for each species, the interaction can change to something that is no longer a mutualism.
Shifting from Mutualism to Negative Interactions
When one species in a relationship provides a benefit to another species at some cost but no longer receives a benefit in return, the interaction can shift from a positive, mutualistic interaction to a negative interaction such as herbivory, predation, or parasitism. For example, we discussed the role that mycorrhizal fungi play in helping plants obtain nutrients. Such a mutualism should be important to plants when nutrients are rare, but not when nutrients are abundant. In an experiment with citrus trees in highly fertile soils, researchers examined the effect of eliminating the mycorrhizal fungi by treating the soil with a fungicide. They found that eliminating the fungus made the trees grow up to 17 percent faster. Because the soil was fertile, the trees could grow well by collecting nutrients on their own, but when the fungi still existed in the soil, the trees still provided the fungi with the products of photosynthesis. As a result, the normally mutualistic relationship had changed into a parasitic interaction. A similar situation exists for cleaner wrasse fish. You may recall that this fish removes ectoparasites from larger fish. Both species benefit and the interaction is a mutualism. However, it turns out that cleaner wrasse also like to consume the mucus and scales of larger fish, which is harmful to the larger fish because mucus and scales are costly to produce and offer protection against infection. Researchers working on coral reefs in the Caribbean examined whether the feeding decisions of a cleaner wrasse, the Caribbean cleaning goby (Elacatinus evelynae), changed when there were differences in the number of ectoparasites carried by the longfin damselfish (Stegastes diencaeus). They sampled the number of ectoparasites on the damselfish off the coasts of six different islands and then observed the cleaner wrasse to
determine the percentage of mucus and scales in their diet. You can see their data in Figure 17.17. When damselfish populations had a high number of parasites, the cleaner wrasse ingested a small percentage of mucus and scales. However, when the damselfish had a low number of parasites, the cleaner wrasse ingested a much higher percentage of mucus and scales. Therefore, when the parasites are rare on the damselfish, the cleaner wrasse are forced to switch from being mutualistic to predatory and consume a higher percentage of mucus and scales.
Figure 17.17 Switching from mutualism to predation. In populations of damselfish where ectoparasites are abundant, the cleaner fish primarily consume parasites of the damselfish and very little of the damselfish’s mucus and scales. This is a mutualistic interaction. In populations of damselfish where the ectoparasites are not abundant, the diet of the cleaner fish includes a substantial percentage of the mucus and scales from the damselfish. This becomes more of a predatory interaction. Error bars are standard errors.
Dealing with Cheaters in Mutualisms
When a mutualistic relationship changes into a relationship in which one species receives a benefit but does not provide one in return, natural selection should favor mechanisms that enable organisms to defend themselves. We saw an example of this with yuccas and yucca moths. When a moth lays so many eggs that the hatching larvae will eat all the developing yucca seeds,
some species of yucca can respond by aborting the flower, thereby killing the moth larvae. In this way, the yucca punishes any yucca moths that act as cheaters in a mutualism. A similar situation exists in relationships between plants and mycorrhizal fungi. When they operate as mutualists, both species provide a benefit at some cost. If a fungus reduces the benefit it provides to a plant, the plant should respond by providing a smaller benefit to the fungus. In 2009, researchers conducted a study to see if a plant could discriminate between different fungi and send products of photosynthesis to the most beneficial fungal mutualist. To test this question, individual wild onion (Allium vineale) plants were planted in two pots; half of the plant’s roots went into a pot of soil containing a fungal species that helps the plant obtain food and the other half of the roots went into a second pot containing a fungal species that provides no benefit to the plant. This experiment is illustrated in Figure 17.18a. After the onion roots grew for 9 weeks, researchers measured how much photosynthetic product the plant was sending to each fungus. They did this by placing a bag around each plant for 4 days and pumping in a special form of CO2 that contained a rare isotope of carbon known as 14 C. Using this radioactive form of carbon, they could track the movement of carbon during the 4 days from the plant to each fungal species. They found that onion plants sent more products of photosynthesis to the beneficial fungi, as you can see in Figure 17.18b. This means that plants can distinguish among different fungi and preferentially provide greater benefits to the most beneficial fungi.
Figure 17.18 Favoring the most beneficial partner. (a) In an experiment, the wild onion plant was grown with its roots into two separate pots: one contained a beneficial mycorrhizal fungus and the other contained a non-beneficial mycorrhizal fungus. After 9 weeks of growth, researchers were able to determine whether the plant allocates more of its photosynthetic products to the beneficial or non-beneficial fungus. (b) Because the plant can distinguish between the two fungal species, it is able to allocate more of its photosynthetic products to the more beneficial fungus. Error bars are standard errors.
Concept Check
1. How might an increase in soil nutrients alter a mutualism between plants and mycorrhizal fungi? 2. Based on the mutualism between cleaner wrasse and the large species of fish that they clean, what would you predict about the preference of the cleaner wrasse for parasites versus a diet of fish scales and mucus? 3. What is the evidence that plants can detect non-beneficial species of mycorrhizal fungi and respond to them appropriately?
#### 17.5 Mutualisms Can Affect Species Distributions, Communities, and Ecosystems
17.5 Mutualisms can affect species distributions, communities, and ecosystems When we think about mutualisms, we often focus on how the relationship helps each of the interacting species; it often improves the fitness and abundance of each participant. For example, corals cannot survive without zooxanthellae and many plants cannot produce offspring without pollinators. In addition to affecting abundance, mutualisms can also affect other ecological levels. In this section, we will examine how mutualisms, and the interruption of mutualisms, can alter the distributions of species, the diversity of communities, and the functioning of ecosystems.
Effects on Species Distributions
Given the benefits of mutualisms, we might expect natural selection to favor mutualistic relationships among species and to expand the distribution of species engaged in mutualisms. In contrast, we would expect the disruption of a mutualism to cause a decline of the species involved and a reduction in their distribution. We can see an example of this in the plant known as garlic mustard, a member of the mustard family with leaves that smell like garlic when crushed. As we discussed in Chapter 16, garlic mustard was introduced to North America more than a century ago from Native to Europe and Asia. When garlic mustard grows in North American forests, it causes young trees to grow poorly and makes them less likely to reach adult size. For a number of years, researchers sought to understand the mechanism underlying garlic mustard’s harmful effects. In 2006, researchers discovered that garlic mustard was interfering with the mutualism between the forest trees and arbuscular mycorrhizal fungi in the soil. To demonstrate this link, they examined how well three species of trees grew when raised in soil where garlic mustard had invaded and in soil where it had not invaded. As you can see in Figure 17.19a, the increases in biomass of sugar maple, red maple, and white ash trees were many times greater when grown in soil that had not been invaded by garlic mustard. Moreover, when the researchers examined the percentage of tree roots colonized by mycorrhizal fungi, they found that soil collected from forests with garlic
mustard showed little or no colonization by mycorrhizae, as illustrated in
Figure 17.19b. Given that tree species depend on fungal mutualism to different degrees, garlic mustard has the largest negative effect on those tree species that have the greatest dependence on the fungi. Because garlic mustard disrupts vital mutualisms, it has the potential to alter the distribution of a large number of other species as it spreads across North America.
Figure 17.19 Disrupting a mutualism with garlic mustard. When researchers grew three species of trees in soils from forests with and without garlic mustard, they found that soils from forests with garlic mustard caused (a) much smaller increases in biomass and (b) little or no
##### Analyzing Ecology: Comparing Two Groups of Data That Do Not Have Normal Distributions
colonization by mycorrhizal fungi. Error bars are standard errors.
Analyzing Ecology
Comparing Two Groups of Data That Do Not Have Normal Distributions As we saw in the case of the cleaner wrasse, researchers who study mutualisms often need to test whether the mutualistic interaction really provides a benefit to each species in the interaction. To do so, statistical tests are used to compare how each species performs both with and without the presence of the other species. In Chapter 15, we discussed the use of ttests to compare the means of two groups. As you may recall, t-tests require that the data collected follow a normal distribution, which are the bell-shaped curves that we discussed in Chapter 2. In some cases, however, the data from two groups do not have normal distributions, so we cannot use the t-test. In these cases, we need to use the MannWhitney rank sum test, which is named for the statisticians who developed it. The Mann-Whitney rank sum test begins by ranking the data—from low to high—and then taking the sum of those ranks. For example, consider the following set of data for the number of species occurring in eight coral reefs where researchers have removed the cleaner fish and eight coral reefs where researchers have allowed the cleaner fish to remain. If we were to take all 16 numbers and place them in order from the lowest value to the highest value, we could give each value a rank from 1 to 16. Whenever we have more than one occurrence of a value, we assign both values the average rank. Number of Species Observed Without Cleaner Wrasse Ranks Number of Species Observed with Cleaner Wrasse Ranks (Group 1) (Group 1) (Group 2) (Group 2) 6.5
8.5 10.5 4.5 12.5 4.5 12.5 6.5 8.5 10.5 The next step is to sum the ranks for group 1 (denoted as R1) and group 2 (denoted as R2): R1=1+2+3+4.5+4.5+6.5+8.5+10.5=40.5R2=6.5+8.5+10.5+12.5+12.5+14+15+16=95.5 We can then use either value of R to calculate a test statistic known as U. We will use R2: U=R2−(n2×(n2+1)÷2) where n1 is the number of observations in group 1, and n2 is the number of observations in group 2. Using this formula, we get U=95.5−(8×(8+1)÷2)=59.5 Now that we know the value of U, we need to calculate the mean and standard deviation of U for the entire collection of data from both groups. The mean value of U, known as mU, is mU= (n1 n2)÷2mU=(8×8)÷2=32 The standard deviation of U is σU=n1 n2 × (n1+ n2+ 1)12σU=8 × 8 ×(8 + 8 + 1)12=9.52 Using our calculated values of U, mU, and σU, we can calculate z,
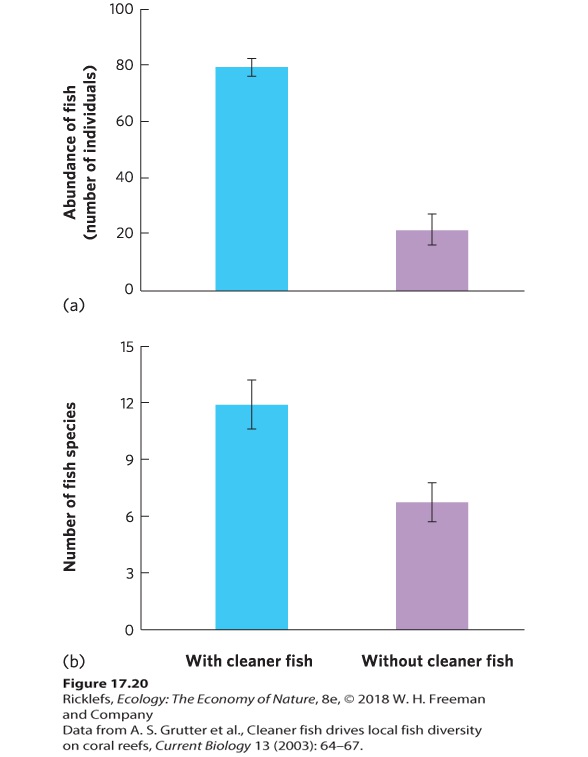
which is defined as z=U - mUσUz=59.5 - 329.52=2.89 We can look up this value in a table of z values in the Statistical Tables appendix, where we find that the probability value is 0.002. Since this probability is less than 0.05, we can conclude that the groups are significantly different from each other. This means that removing the cleaner fish causes a significant decrease in the number of fish species that live on a coral reef. YOUR TURN When we calculate the value of z, we actually use the absolute value of z. To convince yourself that it does not matter whether we use R1 or R2, redo the above calculations based on the formula for U when using R1: U=R1−(n1×(n1+1)÷2) MUTUALISM’S EFFECTS ON COMMUNITIES Mutualisms that alter the abundance and distribution of one or more species can have widespread effects on the rest of the community. A community can be affected in several ways; the mutualism might cause a change in species diversity or it might alter the abundance of individuals within species in the community. Altering Species Diversity A generalist mutualist species interacts with many other species and offers widespread benefits. In the case of the cleaner wrasse fish, for example, a particular species of cleaner wrasse may remove parasites from many different species of larger fish. If this behavior helps species to persist on a coral reef, then the removal of cleaner wrasse should cause a decline in the total number of large fish and the number of species. Researchers working on coral reefs in Australia tested this question by removing a species of cleaner wrasse (Labroides dimidiatus) from nine small reefs. They designated
another nine reefs as controls. After 18 months, they counted both the number of individual fish and the number of fish species on each reef. As you can see in Figure 17.20, removing the cleaner wrasse caused the number of other reef fish to decline by about three-quarters and the number of species to decline by half, which suggests that cleaner wrasse play a critical role in maintaining populations of reef fish.
Figure 17.20 Effects of mutualists on the abundance and diversity of a coral reef fish community. After 18 months of removing cleaner wrasse, (a) the abundance of individual fish declined by about three-quarters and (b) the number of species declined by about half. Error bars are standard errors.
Initiating a Chain Reaction of Species Interactions In some cases, the community does not lose species when a mutualism is disturbed, but the abundance of many species changes through a chain of interactions. Earlier in this chapter, we discussed the role of ants in defending acacia trees against herbivores. We saw that the ants reduce herbivory on acacia trees, which improves tree survival. In return, ants benefit from having a place to make nests and a source of food in the nectaries. However, what would happen if much of the herbivory were removed, causing the ants to no longer provide a benefit to the trees? Researchers examined this question in the savanna region of Kenya. They set aside 12 plots of 4 ha each; half of the plots were fenced to exclude all large herbivores, whereas the other half were left as unfenced controls. The plots contained 40- to 70-year-old acacia trees (Acacia drepanolobum). After 10 years, researchers examined how fencing the plots affected the ant‒acacia mutualism and the rest of the community. You can view their data in Figure 17.21. In the fenced plots that lacked large herbivores, trees produced fewer swollen thorns and nectaries than trees in the control plots, as shown in 17.21a and 17.21b.
Figure 17.21 Community effects of a mutualism. The community effects of the mutualism were discovered when researchers fenced off areas of acacia trees from large mammalian herbivores. Fenced trees that were tended by either C. sjostedti or C. mimosa began to produce (a) fewer swollen thorns and (b) fewer nectaries. (c) In response to the changes in the trees, C. sjostedti began occupying many more acacia trees, while C. mimosa occupied fewer trees. Error bars are standard errors. Changes in the acacia trees caused subsequent changes in the abundances of mutualistic ants. One species of ant, Crematogaster mimosa, relies heavily on the swollen thorns for making its nests and raising its offspring. The reduction of swollen thorns in the fenced plots caused a 30 percent reduction in the proportion of trees occupied by this ant, as illustrated in Figure 17.21c. Moreover, of those trees that were occupied, the average colony of C. mimosa was 47 percent smaller than in the control plots. Another ant species, C. sjostedti, does not use the swollen thorns for nests but instead nests in tree cavities that are excavated in the acacia tree by longhorned beetles. Because this species does not require swollen thorns, the decline in C. mimosa in the fenced plots allowed C. sjostedti to double the proportion of trees that it occupied. This increase in C. sjostedti had further effects on the community. In contrast to C. mimosa, which works to eliminate insect herbivores, C. sjostedti allows long-horned beetles to live on the trees and bore holes into the trunk as it slowly consumes the trees. The beetle receives a food benefit and C. sjostedti receives a nest benefit, so these two species represent another mutualism in the community. However, beetle excavation causes the acacia tree to grow more slowly and die at about twice the rate as trees that are tended by C. mimosa. As a result, although larger mammal herbivores can graze the acacia trees and have negative effects on tree growth, the exclusion of large herbivores turns out to have a much larger negative effect on the trees because the trees are no longer defended from competitors or long-horned beetles. As this example demonstrates, mutualisms affect more than the species involved in the interactions; they can have far-reaching effects throughout the entire community. MUTUALISM’S EFFECTS ON ECOSYSTEM FUNCTION Mutualisms can also have ecological effects at the level of the ecosystem. As we discussed in Chapter 1, researchers working on ecosystems examine the movement of energy and matter among many sources, including both the
biotic and abiotic realms. For example, researchers working with arbuscular mycorrhizal fungi investigated how a grassland ecosystem of 15 species would respond to different numbers of fungal species in the soil. When they examined the total amount of phosphorus that the plants took up from the soil, they found that plants living in soils that contained more fungal species took up a greater amount of phosphorus, as shown in Figure 17.22a. They also quantified the total biomass of roots in the soil and shoots above the soil. They found that a higher number of fungal species in the soil caused substantial increases in the biomass of the roots and shoots in the ecosystem. You can see these data in Figures 17.22b and 17.22c. As illustrated in this example, mutualisms not only affect individual species, but they can also have large effects on the functioning of ecosystems.
Figure 17.22 Ecosystem effects of fungal diversity. When researchers manipulated increases in the species diversity of mycorrhizal fungi, they observed an increase in (a) the total amount of phosphorus in the plants, (b) the total biomass of roots, and (c) the total biomass of shoots. Error bars are standard errors. Throughout this chapter, we have seen how species can interact in a variety of mutualisms. These interactions fulfill a wide range of needs, including resource acquisition, places to live, defense, pollination, and dispersal. These interactions can have important effects on communities and ecosystems and they can even have major conservation implications, as you will see in the “Ecology Today” feature that follows.
Concept Check
1. How is the invasion of garlic mustard altering the distribution of tree species? 2. Why do acacia trees that are no longer experiencing herbivory from large mammals provide smaller rewards for mutualistic ants? 3. Why does increasing the diversity of mycorrhizal fungi lead to greater plant productivity?
Concepts
A substitute mutualist. Aldabra tortoises were introduced onto a small island in the Republic of Mauritius to act as a seed disperser for a population of critically endangered ebony trees. The Republic of Mauritius provides an example of how important mutualisms can be to the persistence of species in nature. Mauritius, a group of islands located in the Indian Ocean southeast of the African continent, is the only place on Earth where the dodo bird (Raphus cucullatus) once lived. A large, flightless bird, the dodo was an easy source of food for the sailors who visited the islands, and was also harmed by the introduction of many non-native species. By the late 1600s, about 200 years after humans first set foot on Mauritius, the dodo was extinct. Numerous other species unique to Mauritius have also been driven extinct, including two species of giant tortoises: the high-backed tortoise (Cylindraspis triserrata) and the domed tortoise (C. inepta). Not only are the islands missing these species, they are also missing the services that these species provided. Over subsequent centuries, as humans settled the islands, they cut down large swaths of tropical forest for
lumber, firewood, and space for sugarcane fields. Today, only about 2 percent of the original forest remains, and some of the tree species continue to decline. For example, Syzygium mamillatum is a critically endangered tree species that consists of just over a hundred individuals. Scientists studying this problem realized that many declining tree species had probably once relied on the dodos and tortoises for seed dispersal. These species consumed the fruit, and the seeds were released after they passed through the animals’ digestive systems. When the animals defecated, typically far away from the trees that produced the fruits, the seeds were ready to germinate. With these mutualists extinct, fewer fruits were consumed and any released seeds remained close to the parent tree. Seeds that geminate near the parent tree compete with the parent, and there are high rates of disease among conspecific trees concentrated in one place. Without dispersers, the seeds of the rare trees cannot disperse and recolonize open areas that have been logged. Researchers wondered if it was possible to bring back the process of seed dispersal even without the animals that originally did the dispersing. The researchers came up with a radical idea hypothesis: perhaps non-native species could be introduced to fill the role of the lost mutualists and help save the endangered trees. To play the role of the dodo, they brought in domestic turkeys (Meleagris gallopavo), which—similar to dodos—have a large gizzard that grinds the fruits they swallow. The thought was that when the dodo consumed the fruits, the grinding of the gizzard broke down the fruits and released the seeds, but maintained the viability of the seeds as they passed through the bird’s digestive system.
To play the role of the two species of extinct tortoises, they brought in giant Aldabra tortoises (Aldabrachelys gigantea) from the nearby Seychelles Islands. Seed-dispersing tortoises. By consuming the large ebony fruits and dispersing the seeds through their feces, the tortoises spread the seeds throughout the island. They may represent the best hope for bringing the ebony tree back from the brink of extinction. To test the potential for these species to act as substitute seed dispersers, fruits of the S. mamillatum tree were fed to the turkeys and Aldabra tortoises. The turkeys' large gizzards not only broke apart the fruit but also ground the seeds into small pieces, which meant that turkeys were not suitable replacements for the dodos. However, when fruits of the tree were fed to the Aldabra tortoises, the researchers found that 16 percent of the seeds inside the fruits remained whole in tortoise feces. Although these seeds exhibited lower germination success than seeds that did not pass through the tortoise’s gut, they produced seedlings that grew taller and had more leaves. This meant that the Aldabra tortoises might be effective substitute dispersers. A follow-up study tested whether the tortoises could be
new mutualists for critically endangered trees if they were released into the wild. To test this idea, Aldabra tortoises were introduced to a small, 25-ha island that is part of the Republic of Mauritius. This island contains another rare tree species, the ebony tree (Diospyros egrettarum), which was once abundant and produces large, 16-g fruits. Logging for firewood had made this tree rare on the island, and large areas contained no new ebony tree seedlings. Without dispersers, the fruits landed near the parent tree and stayed there. After introducing the tortoises into pens in 2000, the researchers confirmed that the tortoises had no negative effects on the plant community. In 2005, 11 tortoises were allowed to roam freely throughout the island. In 2011, the investigators reported some remarkable results. Few fruits were now found near the parent trees because the tortoises were consuming most of them and, because the seeds passed through the tortoises when they defecated, the seeds were relocated to many places around the island. New seedlings showed up everywhere, including open areas where the tree had not existed for many decades. Moreover, ebony seeds that passed through the tortoise gut germinated better than unconsumed seeds. In 2013, the researchers reported the results of releasing tortoises on Round Island, which is another small island off the coast of Mauritius. Introduced rabbits and goats had lived on this island until they were eliminated in the 1970s. The island also had several species of introduced plants that were outcompeting the native plants. The manual labor required to remove the introduced plants was costly, so the researchers proposed using tortoises as an ecological replacement for the extinct tortoises that would
have grazed on many of the plants. The researchers initially introduced Aldabra tortoises and Madagascan radiated (Astrochelys radiata) tortoises to enclosures for 1 year to determine their grazing impacts on the plants. After that, they allowed the tortoises to freely roam around the island and graze on plants. In both situations, the tortoises’ diets were made up of 81 to 93 percent non-native plants. Moreover, the cost of introducing the tortoises over the long term was less than the cost of paying people to control the non-native plants. After more than a decade of research, these results suggest that, although we cannot bring back extinct species, we may be able to substitute some species to resurrect mutualisms so that the original extinctions do not cause subsequent extinctions of their mutualistic partners. SOURCES: Griffiths, C. J., et al. 2011. Resurrecting extinct interactions with extant substitutes. Current Biology 21: 1–4. Griffiths, C. J., et al. 2013. Assessing the potential to restore historic grazing ecosystems with tortoise ecological replacements. Conservation Biology 27: 690–700. Hansen, D. M., et al. 2008. Seed dispersal and establishment of endangered plants on oceanic islands: The Janzen–Connell model, and the use of ecological analogues. PLOS One 3: 1–13. Seddon, P. J., et al. 2014. Reversing defaunation: Restoring species in a changing world. Science 345: 406–411.
Summary of Learning Objectives
17.1 Mutualisms can improve the acquisition of water, nutrients, and places to live. Mutualisms can be categorized as generalists, which interact with many species, or specialists, which interact with few other species. When both species require each other to persist, they are obligate mutualists. When the interaction is beneficial but not critical to the persistence of either species, they are facultative mutualists. Mutualisms for resources include the algae and fungi that compose lichens and the corals and zooxanthellae that build coral reefs. Plants also participate in this type of mutualism by interacting with endomycorrhizal fungi, ectomycorrhizal fungi, and Rhizobium bacteria. In most animals, protists can play an important role in digesting food. Other animals construct habitats that they share with other species in exchange for other benefits. Key Terms: Generalists, Specialists, Obligate mutualists, Facultative mutualists, Mycorrhizal fungi, Endomycorrhizal fungi, Arbuscular fungi, Ectomycorrhizal fungi
17.2 Mutualisms can aid in defense against enemies. Plants make use of defensive mutualisms in a number of ways, including mutualisms with aggressive insects such as ants, and with endophytic fungi that produce chemicals harmful to herbivores. Animals that interact as mutualists to defend against enemies include cleaner fish that remove parasites from large fish and oxpecker birds that remove ticks from mammals. Key Terms: Endophytic fungi
17.3 Mutualisms can facilitate pollination and seed dispersal. Pollinators allow many species of plants to be fertilized, and some plants have evolved traits that favor a particular type of pollinator. When this happens, the plants and the pollinators can co-evolve. Numerous plants also depend on mutualisms to disperse their seeds.
In some cases, seeds are dispersed as the result of animals storing them far from the parent plant. In other cases, animals consume the fruit of plants and the seeds are dispersed after passing through their digestive systems.
17.4 Mutualisms can change when conditions change. Although mutualisms benefit all species in the interaction, a positive mutualism can switch to a neutral or negative interaction when conditions change. In some cases, species can respond to cheaters in a mutualism by only rewarding individuals that provide benefits in return.
17.5 Mutualisms can affect communities. Mutualisms can increase or decrease the abundance of participating species. An absent mutualist can cause another species to be completely eliminated, thereby affecting the distribution of a species. Mutualists can also affect communities either by directly altering the number of species or by initiating a chain of interactions through a community. At the ecosystem level, mutualists can have additional effects, such as moving nutrients into producers and increasing the total biomass of producers.
Critical Thinking Questions
1. Compare and contrast obligate mutualists and facultative mutualists. 2. If garlic mustard were to reduce arbuscular mycorrhizal fungi in an apple orchard, what effect would this have on apple crops? 3. If the mountain tree shrew and summit rat of Borneo were to go extinct, what would likely happen to the pitcher plant that currently lives in a mutualism with these two mammals? 4. If one species provides a habitat as part of a mutualistic relationship, what is the probable effect on the abundance and distribution of the other species? 5. How would you respond to someone who states that a mutualism is favored by natural selection because each species is trying to increase the other species’ fitness? 6. If cleaner wrasse fish consume both parasites and scales from larger fish, what would determine whether the interaction is best categorized as mutualism or parasitism? 7. For a tree that uses a facultative mutualist animal to disperse its seeds, what would be the impact of the animal going extinct? 8. Why should wild strawberries, which have animal-dispersed seeds, not make their fruit very colorful and sweet until the seeds are fully developed? 9. What might prevent an endophytic fungus from reaping the benefits from a grass without providing a benefit in return? 10. When we introduce a non-native herbivore to replace an extinct herbivore that once served as a mutualist, what factors are likely to be important in determining the success at the community level?
#### Graphing the Data: Ecosystem Function of Fungi
GRAPHING THE DATA Ecosystem Function of Fungi Earlier in this chapter, we saw that manipulating the number of mycorrhizal fungal species has several effects on the ecosystem, including an increase in the total phosphorus that accumulates in plants. (See Figure 17.22.) Using the data in the table, calculate the means and standard errors for the amount of phosphorus that the researchers found remaining in the soil for each of the five fungus treatments. Using these calculated data, plot the relationship between the number of fungal species and the amount of soil phosphorus. Number of Fungal Species Phosphorus Remaining in the Soil (mg P/kg Soil)