1 The Microbial World
III Microbial Cultivation Expands the Horizon of Microbiology
IV Molecular Biology and the Unity and Diversity of Life
Microbiology in Motion
The microbial world is strange and fierce. It is teeming with life, ancient, diverse, and constantly changing. Microorganisms are Earth’s life support system, and from our first breath they influence nearly every moment of our lives. Microbes are in our water and our food, and we carry them on us and in us. Indeed, microbes abound in any natural environment that will support life, including many environments too hostile for higher life forms.
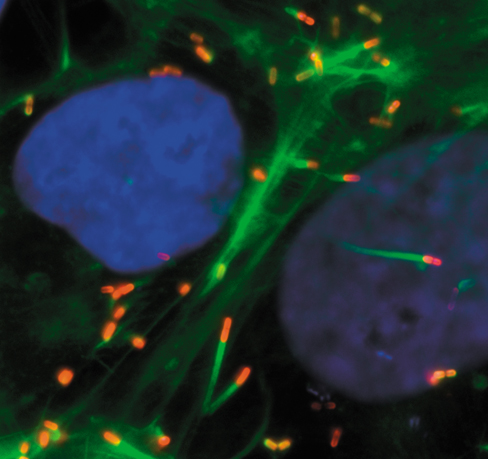
While the microbial world is invisible, we can explore it through the science of microbiology. Microbiology evolves at a breathtaking pace. Even the microscope continues to evolve, providing an ever more detailed picture of the microbial world. The image above was made with a fluorescence microscope that uses lasers, guided by a computer, to map the three-dimensional structure of cells. The image shows neighboring human cells with their nuclei stained blue and actin filaments stained green. These cells are infected with the foodborne bacterial pathogen Listeria monocytogenes, stained red.
Listeria are soil organisms that sometimes find their way into our food. In soils they infect other microbes such as amoebae. Our cells are similar in many ways to those of microscopic organisms, and so Listeria finds itself well adapted to live within us. This bacterium has the unique ability to hijack cellular systems, causing actin to polymerize and propel the cell like a rocket within the host cytoplasm. The force of this propulsion causes Listeria to penetrate adjacent cells (image, lower left), spreading the infection. Listeria can also invade host vacuoles (not shown), where it hides and survives. This persistent state can prolong infection and promote resistance to antibiotic therapy. Research on Listeria has provided new insights on the biology of this pathogen and an ever-changing view of a microbial world in motion.
Source: Kortebi, M., et al. 2017. Listeria monocytogenes switches from dissemination to persistence by adopting a vacuolar lifestyle in epithelial cells. PLoS Pathog. 13: e1006734.
This chapter launches our journey into the microbial world. Here we will begin to discover what the science of microbiology is all about and what microorganisms are, what they do, and how they can be studied. We also place microbiology in historical context, as a process of scientific discovery driven by simple (yet powerful) experiments and insightful minds.
I Exploring the Microbial World
The microbial world consists of microscopic organisms that have defined structures, unique evolutionary histories, and are of enormous importance to the biosphere.
1.1 Microorganisms, Tiny Titans of the Earth
Microorganisms (also called microbes) are life forms too small to be seen by the unaided human eye. These microscopic organisms are diverse in form and function, and they inhabit every environment on Earth that supports life. Many microbes are undifferentiated single-celled organisms, but some can form complex structures, and some are even multicellular. Microorganisms typically live in complex microbial communities (Figure 1.1), and their activities are regulated by interactions with each other, with their environments, and with other organisms. The science of microbiology is all about microorganisms, who they are, how they work, and what they do.
Figure 1.1 Microbial communities.
(a) A bacterial community that developed in the depths of a small Michigan lake, including cells of various phototrophic bacteria. The bacteria were visualized using phase-contrast microscopy. (b) A bacterial community in a sewage sludge sample. The sample was stained with a series of dyes, each of which stained a specific bacterial group. From Journal of Bacteriology 178: 3496–3500, Fig. 2b. © 1996 American Society for Microbiology. (c) Colorized scanning electron micrograph of a microbial community scraped from a human tongue.
Microorganisms were teeming on the land and in the seas for billions of years before the appearance of plants and animals, and their diversity is staggering. Microorganisms represent a major fraction of Earth’s biomass, and their activities are essential to sustaining life. Indeed, the very oxygen (O2) we breathe is the result of microbial activities. Plants and animals are immersed in a world of microbes, and their evolution and survival are heavily influenced by microbial activities, by microbial symbioses, and by pathogens—those microbes that cause disease. Microorganisms are woven into the fabric of human life as well (Figure 1.2), from infectious diseases, to the food we eat, the water we drink, the fertility of our soils, the health of our animals, and even the fuel we put in automobiles. Microbiology is the study of the dominant form of life on Earth, and the effect that microbes have on our planet and all of the living things that call it home.
Figure 1.2 Microbial applications.

Microorganisms have major impacts on the world in which we live. In the chapters that follow we will learn how microorganisms impact our health, the foods we eat, the water we drink, and even the air we breathe. We will learn how microbes can be used to produce valuable products and the many ways in which microorganisms touch our lives.
Microbiologists have many tools for studying microorganisms. Microbiology was born of the microscope, and microscopy is foundational to microbiology. Microbiologists have developed an array of methods for visualizing microorganisms, and these microscopic techniques are essential to microbiology. The cultivation of microorganisms is also foundational to microbiology. A microbial culture is a collection of cells that have been grown in or on a nutrient medium. A medium (plural, media) is a liquid or solid nutrient mixture that contains all of the nutrients required for a microorganism to grow. In microbiology, we use the word growth to refer to the increase in cell number as a result of cell division. A single microbial cell placed on a solid nutrient medium can grow and divide into millions or even billions of cells that form a visible colony (Figure 1.3). The formation of visible colonies makes it easier to see and grow microorganisms. Comprehension of the microbial basis of disease and microbial biochemical diversity has relied on the ability to grow microorganisms in the laboratory.
Figure 1.3 Microbial cells.

(a) Bioluminescent (light-emitting) colonies of the bacterium Photobacterium grown in laboratory culture on a Petri plate. (b) A single colony can contain more than 10 million (107) individual cells. (c) Colorized scanning electron micrograph of cells of Photobacterium.
The ability to grow microorganisms rapidly under controlled conditions makes them highly useful for experiments that probe the fundamental processes of life. Most discoveries relating to the molecular and biochemical basis of life have been made using microorganisms. The study of molecules and their interactions is essential to defining the workings of microbial cells, and the tools of molecular biology and biochemistry are foundational to microbiology. Molecular biology has also provided a variety of tools to study microorganisms without need for their cultivation in the laboratory. These molecular tools have greatly expanded our knowledge of microbial ecology and diversity. Finally, the tools of genomics and molecular genetics are also cornerstones of modern microbiology and allow microbiologists to study the genetic basis of life, how genes evolve, and how they regulate the activities of cells.
In the next section, we explore the basic elements of microbial cell structure and summarize the major physiological activities that take place in all cells, regardless of their structure.
Check Your Understanding
In what ways are microorganisms important to humans?
Why are microbial cells useful for understanding the basis of life?
1.2 Structure and Activities of Microbial Cells
Microbial cells are living compartments that interact with their environment and with other cells in dynamic ways. We purposely exclude viruses in most of this discussion because although they resemble cells in many ways, viruses are not cells but instead a special category of microorganism. We consider the structure, diversity, and activities of viruses in Section 1.4 and in Chapters 5 and 11.
Elements of Microbial Structure
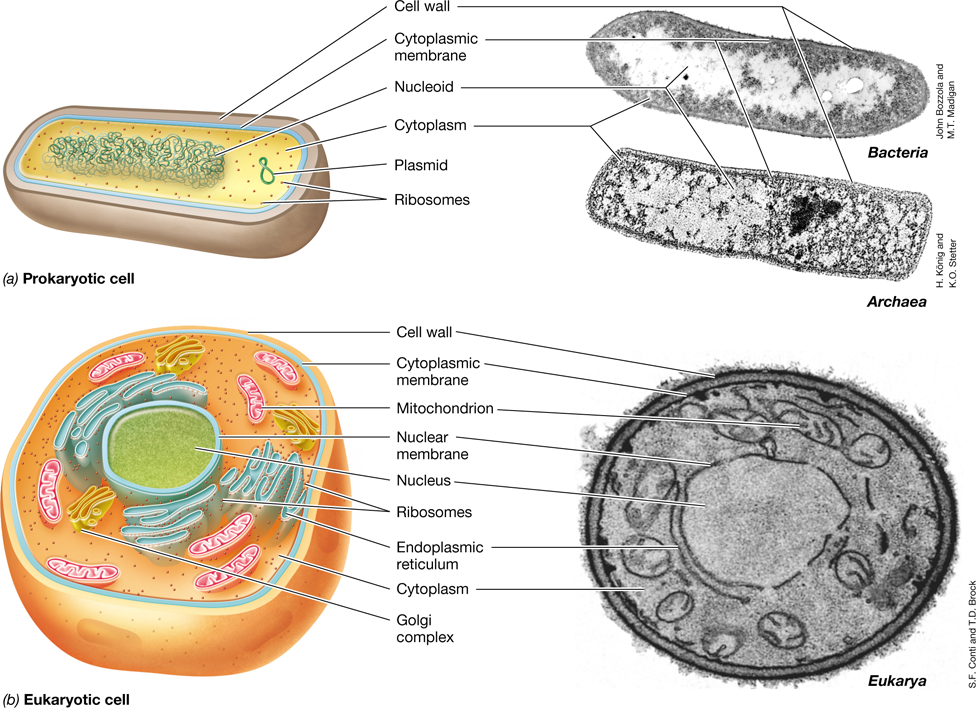
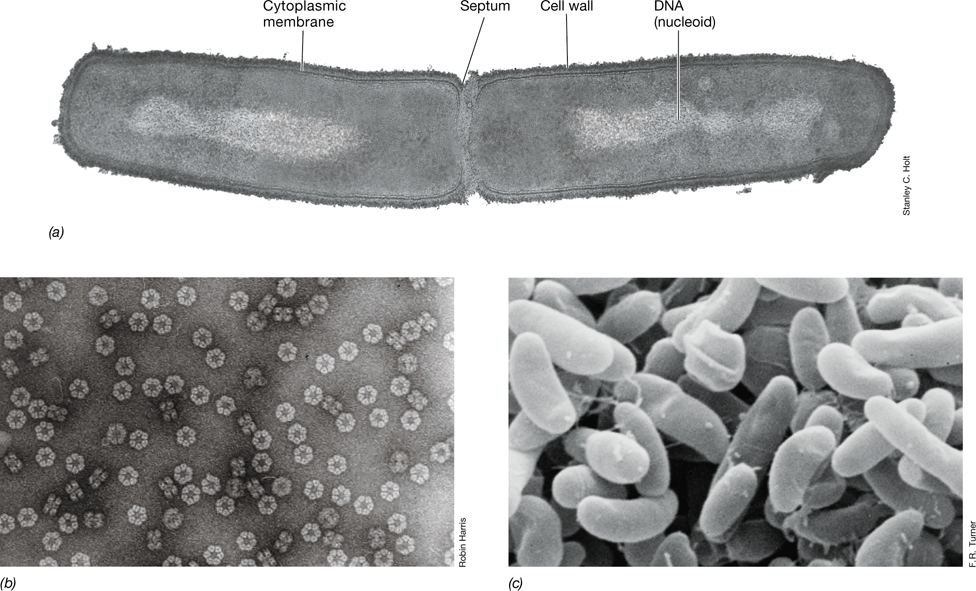
All cells have much in common and contain many of the same components (Figure 1.4). All cells have a permeability barrier called the cytoplasmic membrane that separates the inside of the cell, the cytoplasm, from the outside. The cytoplasm is an aqueous mixture of macromolecules (for example proteins, lipids, nucleic acids, and polysaccharides), small organic molecules (mostly the precursors of macromolecules), various inorganic ions, and ribosomes. All cells also contain ribosomes, which are the structures responsible for protein synthesis. Some cells have a cell wall that lends structural strength to a cell. The cell wall is a relatively permeable structure located outside the cytoplasmic membrane and is a much stronger layer than the membrane itself. Cell walls are typically found in plant cells and most microorganisms but are not found in animal cells.
Figure 1.4 Microbial cell structure.

(a) (Left) Diagram of a prokaryotic cell. (Right) Electron micrograph of Heliobacterium modesticaldum (Bacteria, cell is about 1 μm in diameter) and Thermoproteus neutrophilus (Archaea, cell is about 0.5 μm in diameter). (b) (Left) Diagram of a eukaryotic cell. (Right) Electron micrograph of a cell of Saccharomyces cerevisiae (Eukarya, cell is about 8 μm in diameter). In terms of relative scale, the bacterial cell in a is about the same size as the mitochondria of Saccharomyces in b.
There are two fundamental cell types that differ categorically in cellular organization: those having prokaryotic cell structure, and those having eukaryotic cell structure (Figure 1.4). Cells having eukaryotic cell structure are found in a group of organisms called the Eukarya. This group includes plants and animals as well as diverse microbial eukaryotes such as algae, protozoa, and fungi. Eukaryotic cells contain an assortment of membrane-enclosed cytoplasmic structures called organelles (Figure 1.4b). These include, most prominently, the DNA-containing nucleus but also mitochondria and chloroplasts, organelles that specialize in supplying the cell with energy, and various other organelles.
Mastering Microbiology
Art Activity: Figure 1.3 Common elements of prokaryotic/eukaryotic cells
Prokaryotic cell structure is found within two different groups of organisms we know as Bacteria and Archaea. Prokaryotic cells have few internal structures, they lack a nucleus, and they typically lack organelles (Figure 1.4a). Bacteria and Archaea appeared long before the evolution of eukaryotes (Section 1.5). While all Archaea and Bacteria have prokaryotic cell structure, these two groups diverged very early in the history of life and as a result many of their molecular and genetic characteristics differ at a fundamental level. Indeed, we will see later that in many ways Archaea and Eukarya are more similar to each other than either is to Bacteria.
Genes, Genomes, Nucleus, and Nucleoid
In addition to a cytoplasmic membrane and ribosomes, all cells also possess a DNA genome. The genome is the full set of genes in a cell. A gene is a segment of DNA that encodes a protein or an RNA molecule. The genome is the living blueprint of an organism; the characteristics, activities, and very survival of a cell are governed by its genome.
The genomes of prokaryotic cells and eukaryotic cells are organized into structures called chromosomes. In eukaryotic cells, DNA is present as several linear molecules (each one formed into its own chromosome) within the membrane-enclosed nucleus. By contrast, the genomes of Bacteria and Archaea are typically closed circular chromosomes (though some prokaryotic cells have linear chromosomes). The chromosome aggregates within the prokaryotic cell to form the nucleoid, a mass that is visible in the electron microscope (Figure 1.4a) but which is not enclosed by a membrane. Most prokaryotic cells have only a single chromosome, but many also contain one or more small circles of DNA distinct from that of the chromosome, called plasmids (Figure 1.4a). Plasmids typically contain genes that are not essential but often confer some special property on the cell (such as a unique metabolism, or antibiotic resistance). The genomes of Bacteria and Archaea are typically small and compact, and most contain between 500 and 10,000 genes encoded by 0.5 to 10 million base pairs of DNA. Eukaryotic cells typically have much larger and much less streamlined genomes than prokaryotic cells. A human cell, for example, contains approximately 3 billion base pairs, which encode about 20,000–25,000 genes.
Mastering Microbiology
Art Activity: Figure 1.4 Basic properties of microbial cells
Activities of Microbial Cells
To be competitive in nature, a microorganism must survive and reproduce. Figure 1.5 considers structure and some of the activities that are performed by cells to drive survival and reproduction. All cells show some form of metabolism through which nutrients are acquired from the environment and transformed into new cellular materials and waste products. During these transformations, energy is used to support synthesis of new structures. Production of these new structures culminates in the division of the cell to form two cells. Microbial growth results from successive rounds of cell division.
Figure 1.5 The properties of microbial cells.

While cells are tremendously diverse in form and function, certain properties are shared by all cells.
Genes contain information that is used by the cell to perform the work of metabolism. Genes are decoded to form proteins that regulate cellular processes. Enzymes, those proteins that have catalytic activity, carry out reactions that supply energy and perform biosynthesis within the cell. Enzymes and other proteins are synthesized during gene expression in the sequential processes of transcription and translation. Transcription is the process by which the information encoded in DNA sequences is copied into an RNA molecule, and translation is the process whereby the information in an RNA molecule is used by a ribosome to synthesize a protein (Chapter 6). Gene expression and enzyme activity in a microbial cell are coordinated and highly regulated to ensure that the cell remains optimally tuned to its surroundings. Ultimately, microbial growth requires replication of the genome through the process of DNA replication, followed by cell division. All cells carry out the processes of transcription, translation, and DNA replication.
Microorganisms have the ability to sense and respond to changes in their local environment. Many microbial cells are capable of motility, typically by self-propulsion (Figure 1.5). Motility allows cells to relocate in response to environmental conditions. Some microbial cells undergo differentiation, which may result in the formation of modified cells specialized for growth, dispersal, or survival. Cells respond to chemical signals in their environment, including those produced by other cells of either the same or different species, and these signals often trigger new cellular activities. Microbial cells thus exhibit intercellular communication; that is, they are “aware” of their neighbors and can respond accordingly. Many prokaryotic cells can also exchange genes with neighboring cells, regardless of their species, in the process of horizontal gene transfer.
Evolution (Figure 1.5) results when genes in a population of cells change in sequence and frequency over time, leading to descent with modification. The evolution of microorganisms can be very rapid relative to the evolution of plants and animals. For example, the indiscriminate use of antibiotics in human and veterinary medicine has selected for the proliferation of antibiotic resistance in pathogenic bacteria. The rapid pace of microbial evolution can be attributed in part to the ability of microorganisms to grow very quickly and to acquire new genes though the process of horizontal gene transfer. Not all of the processes depicted in Figure 1.5 occur in all cells. Metabolism, growth, and evolution, however, are universal and will be major areas of emphasis throughout this text.

We now move on to consider the diversity of cell shapes and sizes found in the microbial world.
Check Your Understanding
What structures are universal to all type of cells?
What processes are universal to all types of cells?
What structures can be used to distinguish between prokaryotic cells and eukaryotic cells?
1.3 Cell Size and Morphology
Microscopic examination of microorganisms immediately reveals their morphology, which is defined by cell size and shape. A variety of cell shapes pervade the microbial world, and although microscopic by their very nature, microbial cells come in a variety of sizes. Cell shape can be useful for distinguishing different microbial cells and often has ecological significance. Moreover, the very small size of most microbial cells has a profound effect on their ecology and dictates many aspects of their biology. We begin by considering cell size and then consider cell shape.
The Small World
A micrometer ( μm or micron) is one-millionth of a meter in length. The unaided human eye has difficulty resolving objects that are less than 100 μm in diameter, but this is the scale of the microbial world. Most prokaryotic cells are small, ranging between 0.5 and 10 μm in length, but prokaryotic cells can vary widely in size. For example, the smallest prokaryotic cells are about 0.2 μm in diameter and the largest can be more than 600 μm long (Table 1.1). In contrast, most eukaryotic cells are larger on average than prokaryotic cells, being between 5 and 100 μm in length, but eukaryotic cells can vary widely in size too. For example, the smallest eukaryotic microorganism known is about 0.8 μm in diameter and the largest eukaryotic cells can be many centimeters in length (Section 1.4).
Table 1.1 Cell size and volume of some cells of ***Bacteria***, from the largest to the smallest
aWhere only one number is given, this is the diameter of spherical cells. The values given are for the largest cell size observed in each species. For example, for T. namibiensis, an average cell is only about 200 μm in diameter. But on occasion, giant cells of 750 μm are observed. Likewise, an average cell of S. marinus is about 1 μm in diameter. The species of Beggiatoa here is unclear, and E. fishelsoni, M. bavaricum, and P. ubique are not formally recognized names in taxonomy. For more on ultra-small bacteria, see Explore the Microbial World “Tiny Cells.”
bMycoplasma is a bacterium that lacks a cell wall and can thus take on many shapes (pleomorphic means “many shapes”).
Source: Data obtained from Schulz, H.N., and B.B. Jørgensen. 2001. Annu. Rev. Microbiol. 55: 105–137, and Luef, B., et al. 2015. Nat. Commun. doi:10.1038/ncomms7372.
Cell size is influenced fundamentally by cell structure. Eukaryotic cells, owing to their complex intracellular structure and organelles (Figure 1.4), can actively transport molecules and macromolecules within the cytoplasm. Prokaryotic cells, in contrast, rely on diffusion for transport through the cytoplasm and this limits their size. While diffusion is very fast at small distances, the rate of diffusion increases as the square of the distance traveled. Hence, the metabolic rate in a prokaryotic cell varies inversely with the square of its size. This relationship means that, as cell size increases, it becomes advantageous to have cellular structures that facilitate transport and compartmentalize cellular activities as seen in eukaryotic cells. In contrast, since diffusion is rapid at small spatial scales, high metabolic rates can be maintained in small prokaryotic cells without a need for complex cellular structures.
It is possible, though unusual, for prokaryotic cells to be visible to the human eye; the largest are more than 600 μm (0.6 mm) long. To achieve this size, these bacteria must have traits that allow them to overcome diffusional limitation. The bacterium Epulopiscium fishelsoni (**Figure 1.6*a***; Figure 1.9), which is found in the gut of the surgeonfish, can be more than 75 μm wide and 600 μm long (Table 1.1). One of the traits that allows this bacterium to get so large is that it can have more than 10,000 copies of its genome distributed throughout its cytoplasm, thereby preventing diffusional limitation between the genome and any region of the cytoplasm. Cells of the largest known bacterium, the sulfur-oxidizing chemolithotroph Thiomargarita (Figure 1.6b, Table 1.1), are even larger than those of Epulopiscium, about 750 μm in diameter. Thiomargarita achieves this enormous size by having a large vacuole that fills the center of the cell. Hence, the cytoplasm of Thiomargarita occurs as a thin layer squeezed between the cytoplasmic membrane and this central vacuole. In this way, the cytoplasm is never more than 1 μm from the membrane. In addition, Thiomargarita, like Epulopiscium, also has many copies of its genome, which are distributed throughout its cytoplasm.
Figure 1.6 Two very large ***Bacteria***.

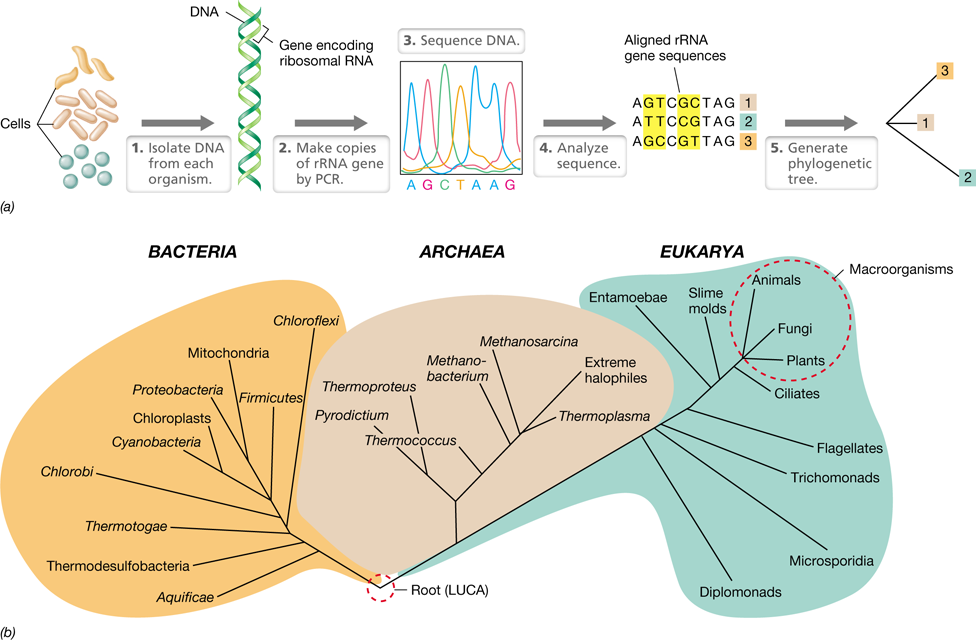
(a) Epulopiscium fishelsoni. The rod-shaped cell is about 600 μm (0.6 mm) long and 75 μm wide and is shown with four cells of the protist Paramecium (a microbial eukaryote), each of which is about 150 μm long. (b) Thiomargarita namibiensis, a large sulfur chemolithotroph and currently the largest known of all prokaryotic cells. Cell widths vary from 400 to 750 μm
At the opposite end of the spectrum from these large prokaryotic cells are very small prokaryotic cells. Exactly how small a cell can be is not precisely known. However, cells 0.2 μm in diameter exist (see Explore the Microbial World, “Tiny Cells”), and the lower limit is probably only a bit smaller than this. Ultimately, the lower limit to cell size is likely a function of the amount of space needed to house the essential biochemical components—proteins, nucleic acids, ribosomes and so on (Section 1.2)—that all cells need to survive and reproduce.
Surface-to-Volume Ratios, Growth Rates, and Evolution
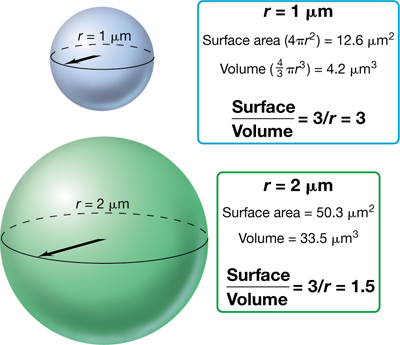
For a cell, there are advantages to being small. Small cells have more surface area relative to cell volume and thus have a higher surface-to-volume ratio than larger cells. To understand this principle, consider a spherical cell. The volume of a sphere is a function of the cube of its radius (V=43πr3), whereas its surface area is a function of the square of the radius (S=4πr2). Therefore, the S/V ratio of a coccus is 3/r (Figure 1.7). As cell size increases, its S/V ratio decreases. To illustrate this, consider the S/V ratio for some of the cells of different sizes listed in Table 1.1: Pelagibacter ubique, 22; Escherichia coli, 4.5; and E. fishelsoni (Figure 1.6a), 0.05. The S/V of a rod-shaped organism can be estimated as if it were a cylinder; hence, the S/V of the cell will decrease as its radius increases.
Figure 1.7 Surface area and volume relationships in cells.

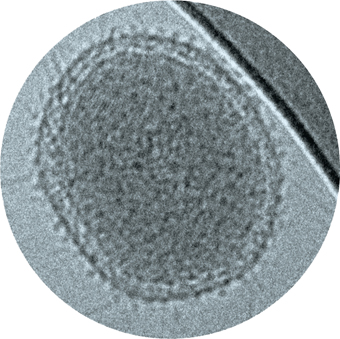
As a cell increases in size, its S/V ratio decreases.
The S/V ratio of a cell controls many of its properties, including how fast it grows (its growth rate) and shape. Cellular growth rate depends in part on the rate at which cells exchange nutrients and waste products with their environment. As cell size decreases, the S/V ratio of the cell increases, and this means that small cells can exchange nutrients and wastes more rapidly (per unit cell volume) than can large cells. As a result, free-living cells that are smaller tend to be more efficient than those that are larger, and any given mass of nutrients will support the synthesis of more small cells than large cells. We will see that cell morphology is also often predicated on the effect of cell shape on S/V ratio. For example, cell shapes that increase the overall membrane area of the cell, such as those having long thin appendages or invaginations, allow bacteria to increase their S/V ratio for a given mass of cytoplasm. We will see that prokaryotic cell morphology is remarkably diverse and different cell shapes can convey different benefits upon the cell.
Major Morphologies of Prokaryotic Cells
Common morphologies of prokaryotic cells are shown in Figure 1.8. A cell that is spherical or ovoid in morphology is called a coccus (plural, cocci). A cylindrically shaped cell is called a rod or a bacillus (plural, bacilli). A spiral-shaped cell is called a spirillum (plural, spirilla). A cell that is slightly curved and comma-shaped is called a vibrio. A spirochete is a special kind of organism (Section 15.17) that has a spiral shape but which differs from spirilla because the cells of spirochetes are flexible, whereas cells of spirilla are rigid. Some bacteria are irregular in shape. Appendages, such as stalks and hyphae, are used by some cells for attachment or to increase surface area. In addition, asymmetrical cell division such as budding can result in irregular and asymmetrical cell shapes.
Figure 1.8 Cell morphologies.

Beside each drawing is a phase-contrast photomicrograph of cells showing that morphology. Coccus (cell diameter in photomicrograph, 1.5 μm); rod (1 μm); spirillum (1 μm); spirochete (0.25 μm); budding (1.2 μm); filamentous (0.8 μm). All photomicrographs are of species of Bacteria. Not all of these morphologies are known among the Archaea, but cocci, rods, and spirilla are common.
Cell division has a major impact on morphology because cells that remain attached to each other can form distinctive shapes. For instance, some cocci occur in pairs (diplococci), some form long chains (streptococci), others occur in three-dimensional cubes (tetrads or sarcinae), and still others occur in grapelike clusters (staphylococci). Filamentous bacteria are long, thin, rod-shaped bacteria that divide terminally and then form long filaments composed of many cells attached end to end.
The cell morphologies described here are representative but certainly not exhaustive; many variations of these morphologies are known. For example, there can be fat rods, thin rods, short rods, and long rods, rods that occur as single cells, as pairs of cells, or rods that form into filaments. As we will see, there are even square bacteria, hexagon-shaped bacteria, and star-shaped bacteria! Cell morphologies thus form a continuum, with some shapes, such as rods and cocci, being very common, whereas others, such as spiral, budding, and filamentous shapes, are less common.
Check Your Understanding
What properties of the cell change as it gets smaller?
Why is it that eukaryotic cells are typically larger than prokaryotic cells?
What traits have allowed the bacteria Epulopiscium and Thiomargarita to have such large cells?
1.4 An Introduction to Microbial Life
As we have seen, microorganisms vary dramatically in size, shape, and structure. In this section we will learn more about different evolutionary (phylogenetic) lineages of cells. All cells fall into one of three major groups: Bacteria, Archaea, or Eukarya. These three major cell lineages are called domains, and all known cellular organisms belong to one of these three domains. In addition, while much of our focus in this chapter is on cellular forms of life, not all microbes form cells. In this section, we will also consider viruses, which are a group of microorganisms that lack a cellular structure. All known microorganisms can be classified into one of these four groups.
Bacteria
Bacteria have a prokaryotic cell structure (Figure 1.4a). Bacteria are often thought of as undifferentiated single cells with a length that ranges from 0.5 to 10 μm. While bacteria that fit this description are common, the Bacteria are actually tremendously diverse in appearance, size, and function (Figure 1.9). Although most bacteria are unicellular, some bacteria can differentiate to form multiple cell types and others are even multicellular (for example, Magnetoglobus, Figure 1.9).
Figure 1.9 Microorganisms vary greatly in size and shape.

The smallest known microbe is the circovirus (20 nm) and the largest shown here is the bacterium Epulopiscium (700 μm), which represents a 35,000-fold difference in length! Certain protozoa can be even larger than Epulopiscium (>2 mm long) and are visible to the unaided eye. Included in the figure are Eukarya: Paramecium (300 μm × 85 μm), diatoms (Navicula, 50 μm × 12 μm), yeast (Saccharomyces, 5 μm), and nanoflagellates (Cafeteria, 2 μm); Bacteria: Epulopiscium (700 μm × 80 μm), cyanobacteria (Oscillatoria, 10 μm-diameter multicellular filaments), Magnetoglobus (multicellular aggregate, 20 μm diameter), Spirochaetes (2−10 μm × 0.25 μm), Flexibacter (5−100 μm × 0.5 μm filaments), Escherichia coli (2 μm × 0.5 μm), Pelagibacter (0.4 μm × 0.15 μm), and Mycoplasma (0.2 μm); Archaea: Giganthauma (10- μm-diameter multicel‑ lular filament), Ignicoccus (6 μm), Nanoarchaeum (0.4 μm), Haloquadratum (2 μm), Methanosarcina (2 μm per cell in packet); and viruses: Pandoravirus (1 μm × 0.4 μm), T4 bacteriophage (200 μm × 90 nm), Influenza A virus (100 nm), Tobacco mosaic virus (300 nm × 20 nm), Circovirus (20 nm).
Among the Bacteria, 30 major phylogenetic lineages (called phyla) have at least one species that has been grown in culture, though many more phyla exist which remain largely uncharacterized. Some of these phyla contain thousands of described species while others contain only a few. More than 90% of cultivated bacteria belong to one of only four phyla: Actinobacteria, Firmicutes, Proteobacteria, and Bacteroidetes. The analyses of environmental DNA sequences provide evidence for the existence of at least 80 bacterial phyla (Section 1.15).
Archaea
Like Bacteria, Archaea also have a prokaryotic cell structure (Figure 1.4a). The domain Archaea consists of five described phyla: Euryarchaeota, Crenarchaeota, Thaumarchaeota, Nanoarchaeota, and Korarchaeota. Archaea have historically been associated with extreme environments; the first isolates came from hot, salty, or acidic sites. But not all Archaea are extremophiles. Archaea are indeed common in the most extreme environments that support life, such as those associated with volcanic systems, and species of Archaea hold many of the records that define the chemical and physical limits of life as we know them. However, in addition to these, Archaea are found widely in nature in nonextreme environments. For example, methane-producing Archaea (methanogens) are common in wetlands and in the guts of animals (including humans) and have a major impact on the greenhouse gas composition of our atmosphere. In addition, species of Thaumarchaeota inhabit soils and oceans worldwide and are important contributors to the global nitrogen cycle.
Archaea are also notable in that this domain lacks any known disease-causing (pathogenic or parasitic) species of plants or animals. Most described species of Archaea fall within the phyla Crenarchaeota and Euryarchaeota while only a handful of species have been described for the Nanoarchaeota, Korarchaeota, and Thaumarchaeota. Analysis of environmental DNA sequences indicate more than 12 archaeal phyla likely exist. We discuss Archaea in detail in Chapter 17.
Eukarya
Plants, animals, and fungi are the most well-known groups of Eukarya. These groups are phylogenetically relatively young compared with Bacteria and Archaea, originating during an evolutionary burst called the Cambrian explosion, which began about 600 million years ago. The first eukaryotes, however, were unicellular microbes. Microbial eukaryotes, which include diverse algae and protozoa, may have first appeared as early as 2 billion years ago, well before the origin of plants, animals, and fungi (Section 1.5). The major lineages of Eukarya are traditionally called kingdoms instead of phyla. There are at least six kingdoms of Eukarya, and this diverse domain contains microorganisms as well as the plants and animals.
Microbial eukaryotes vary dramatically in size, shape, and physiology (Figure 1.9). Among the smallest are the nanoflagellates, which are microbial predators that can be as small as 2 μm long. In addition, Ostreococcus, a genus of green algae that contains species whose cells are only 0.8 μm in diameter, are smaller than many bacteria. The largest single-celled organisms are eukaryotes, but they are hardly microbial. Xenophyophores are amoeba-like, single-celled organisms that live exclusively in the deep oceans and can be up to 10 centimeters in length. In addition, plasmodial slime molds consisting of a single cytoplasmic compartment can be up to 30 cm in diameter. In Chapter 18 we consider microbial eukaryotes in detail.
Viruses
Viruses are not found on the tree of life, and for a variety of reasons, it can be argued that they are not truly alive. Although viruses can replicate—a hallmark of cells—viruses are obligate parasites that can only replicate within the cytoplasm of a host cell. Viruses are not cells, and they lack the cytoplasmic membrane, cytoplasm, and ribosomes found in all forms of cellular life. Viruses do not carry out metabolic processes; instead, they take over the metabolic systems of infected cells and turn them into vessels for producing more viruses. Unlike cells, which all have genomes composed of double-stranded DNA, viruses have genomes composed of DNA or RNA that can be either double- or single-stranded. Viral genomes are often quite small, with the smallest having only three genes. The small size of most viral genomes means that no genes are conserved among all viruses, or between all viruses and all cells.
Although they are not cells, viruses are as diverse as the cells they infect, and different viruses are known to infect cells from all three domains of life. Viruses are often classified on the basis of their structure, genome composition, and host specificity. Viruses that infect bacteria are called bacteriophages (or phages, for short). Bacteriophages have been used as model systems to explore many aspects of viral biology. While most viruses are considerably smaller than bacterial cells (Figure 1.9), there are also unusually large viruses such as the Pandoraviruses, which can be more than 1 micrometer long and have a genome that contains as many as 2500 genes, larger than that of many bacteria! We will learn much more about viruses in Chapters 5 and 11.
Check Your Understanding
How are viruses different from Bacteria, Archaea, and Eukarya?
What four bacterial phyla contain the largest number of well-characterized species?
What phylum of Archaea is common worldwide in soils and in the oceans?
1.5 Microorganisms and the Biosphere
Microbes are the oldest form of life on Earth, and they have evolved to perform critical functions that sustain the biosphere. In this section we will learn how microbes have changed our planet and how they continue to do so.
A Brief History of Life on Earth
Earth is about 4.6 billion years old, and microbial cells first appeared between 3.8 and 4.3 billion years ago (Figure 1.10). During the first 2 billion years of Earth’s existence, its atmosphere was anoxic (O2 was absent), and only nitrogen (N2), carbon dioxide (CO2), and a few other gases were present. Only microorganisms capable of anaerobic metabolism (that is, metabolisms that do not require O2) could survive under these conditions.
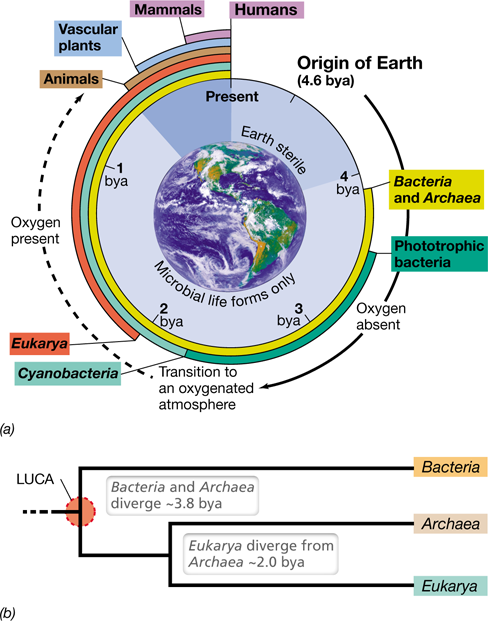
Figure 1.10 A summary of life on Earth through time and origin of the cellular domains.

(a) At its origin, Earth was sterile and anoxic. Cellular life, in the form of Bacteria and Archaea, was present on Earth by 3.8 billion years ago (bya). The evolution of phototrophic bacteria called Cyanobacteria caused Earth’s atmosphere to become oxygenated over time. While the first evidence for oxygen in Earth’s atmosphere appears 2.4 bya, current levels of atmospheric O2 were not achieved until 500–800 million years ago. (b) The three domains of cellular organisms are Bacteria, Archaea, and Eukarya. Bacteria and Archaea appeared first and Eukarya evolved later, diverging from the Archaea. LUCA, last universal common ancestor.
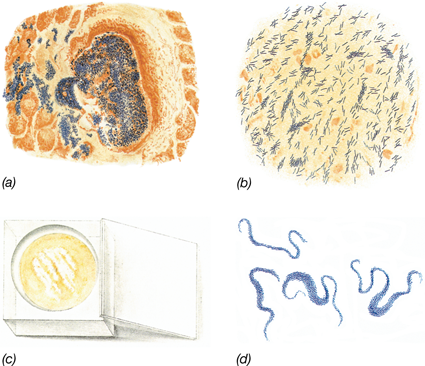
The evolution of phototrophic microorganisms—organisms that harvest energy from sunlight—occurred within 1 billion years of the formation of Earth (Figure 1.10a). The first phototrophs were anoxygenic (non-oxygen-producing), such as the purple sulfur bacteria and green sulfur bacteria we know today (Figure 1.11). Cyanobacteria—oxygen-producing (oxygenic) phototrophs (Figure 1.11f)—evolved nearly a billion years later (Figure 1.10a) and began the slow process of oxygenating Earth’s atmosphere. These early phototrophs lived in structures called microbial mats, which are still found on Earth today (Figure 1.11a–c). After the oxygenation of Earth’s atmosphere, multicellular life forms eventually evolved, culminating in the plants and animals we know today. But plants and animals have only existed for about half a billion years. The timeline of life on Earth (Figure 1.10a) shows that 80% of life’s history was exclusively microbial, and thus in many ways, Earth can be considered a microbial planet.
Figure 1.11 Phototrophic microorganisms.

The earliest phototrophs lived in microbial mats. (a) Microbial mats in the Great Sippewissett Marsh, a salt marsh in Massachusetts, USA. (b) Mats develop a cohesive structure that forms at the sediment surface. (c) A slice through the mat shows colored layers that form due to the presence of photopigments. Cyanobacteria form the green layer nearest the surface, purple sulfur bacteria form the purple and yellow layers below, and green sulfur bacteria form the bottommost green layer. The scale on the knife is in cm. (d) Purple sulfur bacteria, (e) green sulfur bacteria, and (f) cyanobacteria imaged by bright-field and phase-contrast microscopy. Purple and green sulfur bacteria are anoxygenic phototrophs that appeared on Earth long before oxygenic phototrophs (that is, Cyanobacteria) evolved (see Figure 1.10a).
As evolutionary events unfolded, three major lineages of microbial cells—the Bacteria, the Archaea, and the Eukarya (Figure 1.10b)—were distinguished. All cellular organisms share certain characteristics (Figure 1.5) and as a result, certain genes are found in all cells. For example, approximately 60 genes are universally present in cells of all three domains. Examination of these genes reveals that all three domains have descended from a common ancestor, the last universal common ancestor (LUCA, Figure 1.10b). Over enormous periods of time, microorganisms derived from these three domains have evolved to fill every habitable environment on Earth.
Microbial Abundance and Activity in the Biosphere
Microorganisms are present everywhere on Earth that will support life. They constitute a major fraction of global biomass and are key reservoirs of nutrients essential for life. There are an estimated 2×1030 microbial cells on Earth. To put this number in context, the universe in all its vast extent is estimated to contain merely 7×1022 stars. The total amount of carbon present in all microbial cells is a significant fraction of Earth’s biomass (Figure 1.12). Moreover, the total amount of nitrogen and phosphorus (essential nutrients for life) within microbial cells is almost four times that in all plant and animal cells combined. Microbes also represent a major fraction of the total DNA in the biosphere (about 31%), and their genetic diversity far exceeds that of plants and animals.
Figure 1.12 Contribution of microbial cells to global biomass.

Microorganisms comprise a significant fraction of the carbon (C) and a majority of the nitrogen (N) and phosphorus (P) in the biomass of all organisms on Earth. C, N, and P are the macronutrients required in the greatest quantity by living organisms. Animal biomass is a minor fraction (<0.1 %) of total global biomass and is not shown.
Microbes are even abundant in habitats that are much too harsh for other forms of life, such as volcanic hot springs, glaciers and ice-covered regions, high-salt environments, extremely acidic or alkaline habitats, and deep in the sea or deep in the earth at extremely high pressure. Such microorganisms are called extremophiles and their properties define the physiochemical limits to life as we know it (Table 1.2). We will revisit many of these organisms in later chapters and discover the special structural and biochemical properties that allow them to thrive under extreme conditions.
Table 1.2 Classes and examples of extremophilesa

aThe organisms listed are the current “record holders” for growth in laboratory culture at the extreme condition listed.
bAnaerobe showing growth at 122 °C only under several atmospheres of pressure.
cThe permafrost bacterium Planococcus halocryophilus can grow at −15 °C and metabolize at −25 °C. However, the organism grows optimally at 25 °C and grows up to 37 °C and thus is not a true psychrophile.
dP. oshimae is also a thermophile, growing optimally at 60 °C.
eN. gregoryi is also an extreme halophile, growing optimally at 20% NaCl.
fM. yayanosii is also a psychrophile, growing optimally near 4 °C.
All ecosystems are influenced to one extent or another by microbial activities. The metabolic activities of microorganisms can change the habitats in which they live, both chemically and physically, and these changes can affect other organisms. For example, excess nutrients added to a habitat can cause aerobic (O2-consuming) microorganisms to grow rapidly and consume O2, rendering the habitat anoxic (O2-free). Many human activities release nutrients into the coastal oceans, thereby stimulating excessive microbial growth, which can cause enormous anoxic zones in these waters. These “dead zones” cause massive mortality of fish and shellfish in coastal oceans worldwide, because most aquatic animals require O2 and die if it is not available. Only by understanding microorganisms and microbiology can we predict and minimize the effects of human activity on the biosphere that sustains us.
Though diverse habitats are influenced strongly by microorganisms, their contributions are easy to overlook because of their small sizes. Within the human body, for example, more microbial cells can be present than human cells, and more than 200 microbial genes are present for every human gene. These microbes provide benefits and services that are essential to human health. In later chapters, we will return to a consideration of the ways in which microorganisms affect animals, plants, and the entire global ecosystem. This is the science of microbial ecology, perhaps the most exciting subdiscipline of microbiology today. We will see that microbes are important to myriad issues of global importance to humans including climate change, agricultural productivity, and even energy policy.
We focus now on the effects of microbes on humans and human activities.
Check Your Understanding
How old is Earth and when did cells first appear on Earth?
Name the three domains of life. Which of these contain eukaryotic life forms?
Why were cyanobacteria so important in the evolution of life on Earth?
1.6 The Impact of Microorganisms on Human Society
Microbiologists have made great strides in discovering how microorganisms function, and application of this knowledge has greatly advanced human health and welfare. Besides understanding microorganisms as agents of disease, microbiology has made great advances in understanding the important roles microorganisms play in food and agriculture, and microbiologists have exploited microbial activities to produce valuable human products, generate energy, and clean up the environment.
Microorganisms as Agents of Disease
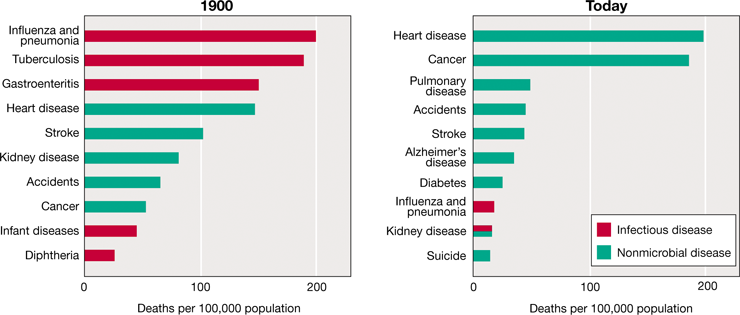
The statistics summarized in Figure 1.13 show how microbiologists and clinical medicine have combined to conquer infectious diseases in the past 120 years. At the beginning of the twentieth century, more than half of all humans died from infectious diseases caused by bacterial and viral pathogens. Today, however, infectious diseases are largely preventable due to advances in our understanding of microbiology. Microbiology has fueled advances in medicine such as vaccination and antibiotic therapy, advances in engineering such as water and wastewater treatment, advances in food safety such as pasteurization, and a better understanding of how microorganisms are transmitted. Infectious diseases now cause fewer than 5% of all deaths in countries where these interventions, made possible by microbiology, are readily available. However, while infectious diseases are preventable, the World Health Organization has documented that they still account for more than a third of all deaths in countries where microbial interventions are less available, such as those having low-income economies. As we will see later in this chapter, the development of microbiology as a science can be traced to pioneering studies of infectious disease.
Figure 1.13 Death rates for the leading causes of death in the United States: 1900 and 2016.

Infectious diseases were the leading causes of death in 1900, whereas today they account for relatively few deaths. Kidney diseases can be caused by microbial infections or systemic sources (diabetes, cancers, toxicities, metabolic diseases, etc.). Data are from the United States National Center for Health Statistics and the Centers for Disease Control and Prevention.
While pathogens and infectious disease remain a major threat to humanity, and combating these harmful organisms remains a major focus of microbiology, most microorganisms are not harmful to humans. In fact, most microorganisms are beneficial, and in many cases are even essential to human welfare and the functioning of the planet. We turn our attention to these microorganisms and microbial activities now.
Microorganisms, Agriculture, and Human Nutrition
Agriculture benefits from nutrient cycling performed by microorganisms, in particular, the cycling of nitrogen, sulfur, and carbon compounds. For example, legumes are a diverse family of plants that include major crop species such as soybeans, peas, and lentils, among others. Legumes live in close association with bacteria that form structures called nodules on their roots. In the nodules, these bacteria convert atmospheric nitrogen (N2) into ammonia (NH3) through the process of nitrogen fixation. NH3 is the major nutrient found in fertilizer and is used as a nitrogen source for plant growth (Figure 1.14). In this way bacteria allow legumes to make their own fertilizer, thereby reducing the need for farmers to apply fertilizers produced industrially. When plants die they are decomposed by bacteria in the soil, and this process produces the nutrients that form the basis of soil fertility. Bacteria regulate nutrient cycles (Figure 1.14), in soils and throughout the biosphere, transforming and recycling the nutrients required by plants and animals.
Figure 1.14 Microorganisms in modern agriculture.

Root nodules on this soybean plant contain bacteria that fix atmospheric nitrogen (N2) to form nitrogenous compounds used by the plant. Ruminant animals such as cows and sheep require rumen microbes to digest cellulose from plants. Plant matter and animal wastes are decomposed in soil to produce nutrients that are the basis of soil fertility and which are required for plant growth.
Also of major agricultural importance are microorganisms that inhabit the rumen of ruminant animals, such as cattle and sheep. Ruminants, like most animals, lack enzymes for breaking down the polysaccharide cellulose, the major component of plant cell walls. The digestive tract of ruminants has a large specialized chamber called the rumen in which cellulose is digested. The rumen contains a dense and diverse community of microorganisms that digest and ferment cellulose. Without these symbiotic microorganisms, ruminants could not digest plant matter like grass and hay, most of which consists of cellulose. Ruminants ultimately get their nutrition by metabolizing the waste products of microbial fermentation and by digesting dead microbial cells. Many domesticated and wild herbivorous mammals—including deer, bison, camels, giraffes, and goats—are also ruminants.
The human gastrointestinal (GI) tract lacks a rumen, but we too rely on microbial partners for our nutrition. Human enzymes lack the ability to break down complex carbohydrates (which can represent 10–30% of food energy) and so we rely on our gut microbiome for this purpose. The colon, or large intestine (Figure 1.15), follows the stomach and small intestine in the human digestive tract, and it contains about 1011 microbial cells per gram of colonic contents. Microbial cell numbers are low in the very acidic (pH 2) stomach (about 104 per gram) but increase to about 108 per gram near the end of the small intestine (pH 4–5) and then reach maximal numbers in the colon (pH 7) (Figure 1.15). The colon contains diverse microbial species that assist in the digestion of complex carbohydrates, and that synthesize vitamins and other nutrients essential to host nutrition. The gut microbiome develops from birth, but it can change over time with the human host. The composition of the gut microbiome has major effects on GI function and human health as we will see in Chapter 24.
Figure 1.15 The human gastrointestinal tract.

(a) Diagram of the human GI tract showing the major organs. (b) Scanning electron micrograph of microbial cells in the human colon (large intestine). Cell numbers in the colon can reach as high as 1011 per gram. As well as high numbers of cells, the microbial diversity in the colon is also quite high.
Microorganisms and Food
Microbes are intimately associated with the foods we eat. Microbial growth in food can cause food spoilage and foodborne disease. The manner in which we harvest and store food (for example, canning, refrigeration, drying, salting, etc.), the ways in which we cook it, and even the spices we use, have all been fundamentally influenced by the goal of eliminating harmful organisms from our food. Microbial food safety and prevention of food spoilage is a major focus of the food industry and a major cause of economic loss every year.
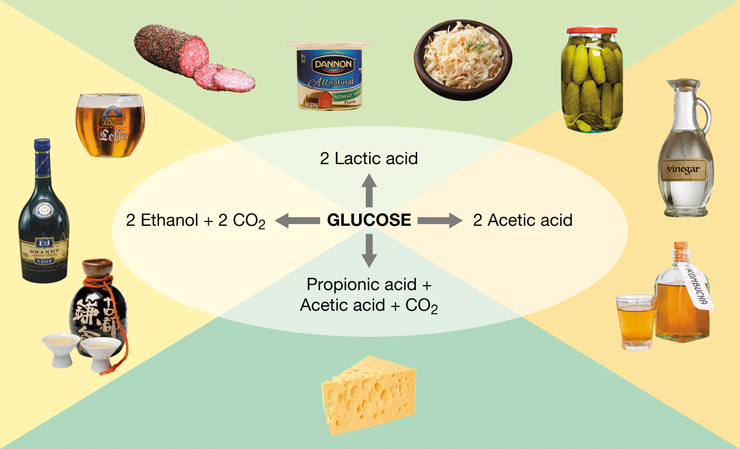
While some microbes can cause foodborne disease and food spoilage, not all microorganisms in foods are harmful. Indeed, beneficial microbes have been used for thousands of years to improve food safety and to preserve foods (Figure 1.16). For example, cheeses, yogurt, and buttermilk are all produced by microbial fermentation of dairy products. Microbial production of lactic acid in these foods improves their shelf life and prevents the growth of foodborne pathogens. Lactic acid–producing bacteria are used to produce a variety of sour-tasting foods, including sauerkraut, kimchi, pickles, and even certain sausages. Even the production of chocolate and coffee rely on microbial fermentation. Moreover, the fermentative activities of yeast are essential for baking (by generating carbon dioxide—CO2—to raise the dough), and for the production of alcoholic beverages (by generating alcohol). The products of microbial fermentation affect the flavor and taste of foods and can prevent spoilage as well as the growth of deleterious organisms.
Figure 1.16 Fermented foods.

Major fermentations in various fermented foods. It is the fermentation product (ethanol, or lactic, propionic, or acetic acids) that both preserves the food and renders in it a characteristic flavor.
Microorganisms and Industry
Microorganisms play important roles in all manner of human activity. The field of industrial microbiology is focused on the use of microorganisms as tools for major industries such as pharmaceuticals and brewing (Figure 1.17). For example, in large industrial settings, naturally occurring microorganisms are grown on a massive scale in bioreactors called fermentors to make large amounts of products, such as antibiotics, enzymes, alcohol, and certain other chemicals, at relatively low cost. By contrast, biotechnology employs genetically engineered microorganisms to synthesize products of high commercial value, such as insulin or other human proteins, usually on a small scale.
Figure 1.17 Industrial microbiology.

Microbes have major impacts on human industry. Microbes can be used to produce valuable products and biofuels and they can also be used to clean up our wastes. Microbial biofilms have major impacts on industry because biofilms can clog and corrode pipelines and holding tanks in factories, in ships, and in the oil industry.
Microorganisms can also be used to produce biofuels (Section 12.19 and Figure 12.33). For example, as previously discussed, natural gas (methane, CH4) is a product of the anaerobic metabolism of methanogenic Archaea. Ethyl alcohol (ethanol) is a major fuel supplement, which is produced by the microbial fermentation of glucose obtained from carbon-rich feedstocks such as sugarcane, corn, or rapidly growing grasses. Microorganisms can even convert waste materials, such as domestic refuse, animal wastes, and cellulose, into ethanol and methane. In producing these biofuels, humans are simply exploiting the metabolic features of particular microbes, but at the same time, are reducing the use of fossil fuels. As we will document in Chapter 21, CO2 levels have been rising rapidly on Earth in the industrial era, and the link between this "greenhouse gas" and Earth’s rising temperatures is firm. Thus, as a sustainable fuel source, biofuels should help cool our planet and are one facet of the "green revolution" many countries support today.
Microorganisms are also used to clean up wastes. Wastewater treatment is essential to sanitation and human health. Wastewater treatment relies on microbes to treat water contaminated with human waste so that it can be reused or returned safely to the environment. Waterborne diseases such as cholera and typhoid (major killers before the blossoming of microbiology: see gastroenteritis, Figure 1.13) can proliferate in the absence of proper wastewater treatment. Microbes can also be used to clean up industrial pollution in a process called bioremediation. In bioremediation, microorganisms are used to transform spilled oil, solvents, pesticides, heavy metals, and other environmentally toxic pollutants into nontoxic forms. Bioremediation accelerates the cleanup process either by adding special microorganisms to a polluted environment or by adding nutrients that stimulate indigenous microorganisms to degrade the pollutants. In either case the goal is to accelerate disappearance of the pollutant.
Microbes can grow in almost any environment containing liquid water, including structures made by humans. For example, microbes often grow on submerged surfaces, forming biofilms. Biofilms that grow in pipes and drains can cause fouling and blockages in factory settings and pipelines, in sewers, and even in water distribution systems. In addition, biofilms that grow on ships’ hulls can cause marked reductions in speed and efficiency. Biofilms can even grow in tanks that store oil and fuel, leading to spoilage of these products. We will learn that biofilms are also of great importance in medicine, as biofilms that form on implanted medical devices (Section 4.9) can cause infections that are extremely difficult to treat.
As these examples show, the influence of microorganisms on humans is great and their activities are essential for the functioning of the planet. Or, as the famous French chemist and early microbiologist Louis Pasteur so aptly put it: “The role of the infinitely small in nature is infinitely large.” Microscopes provide an essential portal through which microbiologists such as Pasteur gazed into the world of microbes. We therefore continue our introduction to the microbial world with an overview of microscopy.
Check Your Understanding
How do microbes contribute to the nutrition of animals such as humans and cows?
Describe several ways in which microorganisms are important in the food and agricultural industries.
What is wastewater treatment and why is it important?
II Microscopy and the Origins of Microbiology
The microscope first revealed the microbial world, and the several different types of microscopes available today remain among the microbiologist’s foremost tools.
Historically, the science of microbiology has taken its greatest leaps forward as new tools are developed and old tools improve. The microscope is the microbiologist’s oldest and most fundamental tool for studying the microbial world. Indeed, microbiology did not exist before the invention of the microscope. Many forms of microscopy are available, and some are extremely powerful. Throughout this text you will see images of microorganisms that were taken through the microscope using a variety of different techniques. So let’s take a moment to explore how microscopy can be used to visualize microbial cells, starting at the very beginning with the invention of the microscope.
1.7 Light Microscopy and the Discovery of Microorganisms
1.7 Light Microscopy and the Discovery of Microorganisms
1.7 Light Microscopy and the Discovery of Microorganisms
Although the existence of creatures too small to be seen with the naked eye had been suspected for centuries, their discovery had to await invention of the microscope. The English mathematician and natural historian Robert Hooke (1635–1703) was an excellent microscopist. In his famous book Micrographia (1665), the first book devoted to microscopic observations, Hooke illustrated many microscopic images including the fruiting structures of molds (Figure 1.18). This was the first known description of microorganisms.
Figure 1.18 Robert Hooke and early microscopy.

A drawing of the microscope used by Robert Hooke in 1664. The lens was fitted at the end of an adjustable bellows (G) and light focused on the specimen by a separate lens (1). Inset: Hooke’s drawing of a bluish mold he found degrading a leather surface; the round structures contain spores of the mold.
The first person to see bacteria, the smallest microbial cells, was the Dutch draper and amateur microscopist Antoni van Leeuwenhoek (1632–1723). Van Leeuwenhoek constructed extremely simple microscopes containing a single lens to examine various natural substances for microorganisms (Figure 1.19). These microscopes were crude by today’s standards, but by careful manipulation and focusing, van Leeuwenhoek was able to see bacteria. He discovered bacteria in 1676 while studying pepper–water infusions and reported his observations in a series of letters to the prestigious Royal Society of London, which published them in English translation in 1684. Drawings of some of van Leeuwenhoek’s “wee animalcules,” as he referred to them, are shown in Figure 1.19b, and a photo taken through a van Leeuwenhoek microscope is shown in Figure 1.19c.
Figure 1.19 The van Leeuwenhoek microscope.

(a) A replica of Antoni van Leeuwenhoek’s microscope. (b) Van Leeuwenhoek’s drawings of bacteria, published in 1684. Even from these simple drawings we can recognize several shapes of common bacteria: A, C, F, and G, rods; E, cocci; H, packets of cocci. (c) Photomicrograph of a human blood smear taken through a van Leeuwenhoek microscope. Red blood cells are clearly apparent.
Van Leeuwenhoek’s microscope was a light microscope, and his design used a simple lens that could magnify an image at least 266 times. In a light microscope the sample is illuminated with visible light. Magnification describes the capacity of a microscope to enlarge an image. All microscopes employ lenses that provide magnification. Magnification, however, is not the limiting factor in our ability to see small objects. It is resolution that governs our ability to see the very small. Resolution is the ability to distinguish two adjacent objects as distinct and separate. The limit of resolution for a light microscope is about 0.2 μm (μm is the abbreviation for micrometer, 10−6 m). What this means is that two objects that are closer together than 0.2 μm cannot be resolved as distinct and separate.
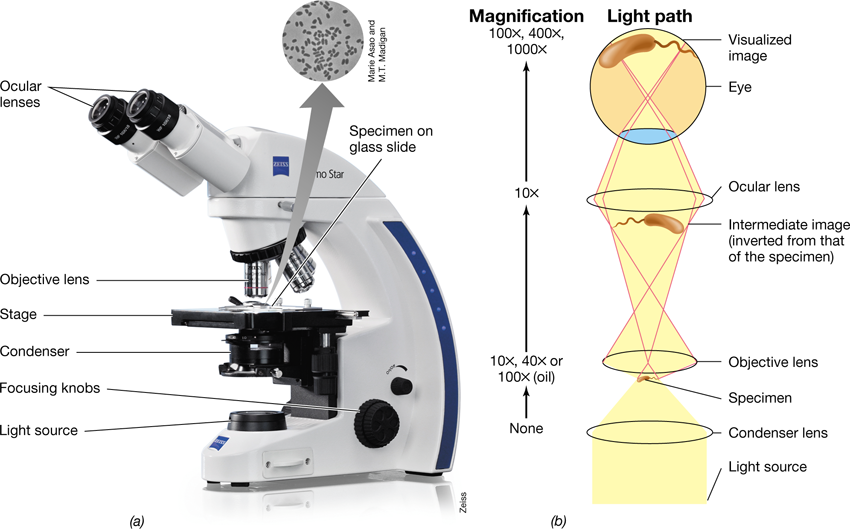
Microscopy has improved remarkably since the days of van Leeuwenhoek. Several types of light microscopy are now available, including bright-field, phase-contrast, differential interference contrast, dark-field, and fluorescence. With the modern compound light microscope, light is focused on the specimen by the condenser (Figure 1.20) and this light passes through the sample and is collected by the lenses. The modern compound light microscope contains two types of lenses, objective and ocular, that function in combination to magnify the image. Microscopes used in microbiology have ocular lenses that magnify 10–30× and objective lenses that magnify 10–100× (Figure 1.20b). The total magnification of a compound light microscope is the product of the magnification of its objective and ocular lenses (Figure 1.20b). Magnification of 1000× is required to resolve objects 0.2 μm in diameter, which is the limit of resolution for most light microscopes (increasing magnification beyond 1000× provides little improvement in the resolution of a light microscope).
Figure 1.20 Microscopy.
(a) A compound light microscope (inset photomicrograph of unstained cells taken through a phase-contrast light microscope). (b) Path of light through a compound light microscope. Figure 1.24 compares cells visualized by bright field with those visualized by phase contrast.
In addition to magnification, the limit of resolution for a light microscope is a function of the wavelength of light used and the light-gathering ability of the objective lens, a property known as its numerical aperture. There is a correlation between the magnification of a lens and its numerical aperture; lenses with higher magnification typically have higher numerical apertures. The diameter of the smallest object resolvable by any lens is equal to 0.5λ/numerical aperture, where λ is the wavelength of light used. With objectives that have a very high numerical aperture (such as the 100× objective), an optical-grade oil is placed between the microscope slide and the objective. Lenses on which oil is used are called oil-immersion lenses. Immersion oil increases the light-gathering ability of a lens, that is, it increases the amount of light that is collected and viewed by the lens.
Mastering Microbiology
In light microscopy, specimens are visualized because of differences in contrast that exist between them and their surroundings (Figure 1.21). In bright-field microscopy, contrast results when cells absorb or scatter light differently from their surroundings. Bacterial cells typically lack contrast, that is, their optical properties are similar to the surrounding liquid, and hence they are difficult to see well with the bright-field microscope. Pigmented microorganisms are an exception because the color of the organism adds contrast, thus improving visualization by bright-field optics (Figure 1.21). For cells lacking pigments there are several ways to boost contrast, and we consider these methods in the next section.
Figure 1.21 Bright-field photomicrographs of pigmented microorganisms.

(a) Purple phototrophic bacteria (Bacteria). The bacterial cells are about 5 μm wide. (b) A green alga (eukaryote). The green structures are chloroplasts. The algal cells are about 15 μm wide. Purple bacteria are anoxygenic phototrophs, whereas algae are oxygenic phototrophs. Both groups contain photosynthetic pigments, but only oxygenic phototrophs produce O2 (Section 1.5 and Figure 1.10a).
Check Your Understanding
What is the limit of resolution for a bright-field microscope? What defines this limit?
1.8 Improving Contrast in Light Microscopy
Contrast is necessary in light microscopy to distinguish microorganisms from their surroundings. Cells can be stained to improve contrast, and staining is commonly used to visualize bacteria with bright-field microscopy. In addition to staining, other methods of light microscopy have been developed to improve contrast with or without staining, and we consider all of these methods here.
Staining: Increasing Contrast for Bright-Field Microscopy
Dyes can be used to stain cells and increase their contrast so that they can be more easily seen in the bright-field microscope. Each class of dye has an affinity for specific cellular materials. Many dyes used in microbiology are positively charged, and for this reason, they are called basic dyes. Examples of basic dyes include methylene blue, crystal violet, and safranin. Basic dyes bind strongly to negatively charged cell components, such as nucleic acids and acidic polysaccharides. These dyes also stain the surfaces of cells because cell surfaces tend to be negatively charged. These properties make basic dyes useful general-purpose stains that nonspecifically stain most bacterial cells.
To perform a simple stain, one begins with dried preparations of cells (Figure 1.22). A clean glass slide containing a dried suspension of cells is flooded for a minute or two with a dilute solution of a basic dye, rinsed several times in water, and blotted dry. Because bacterial cells are so small, it is common to observe dried, stained preparations of those cells with a high-power (oil-immersion) lens.
Figure 1.22 Staining cells for microscopic observation.

Stains improve the contrast between cells and their background. Step 3 lower right: Same cells as shown in Figure 1.20a inset but stained with a basic dye.
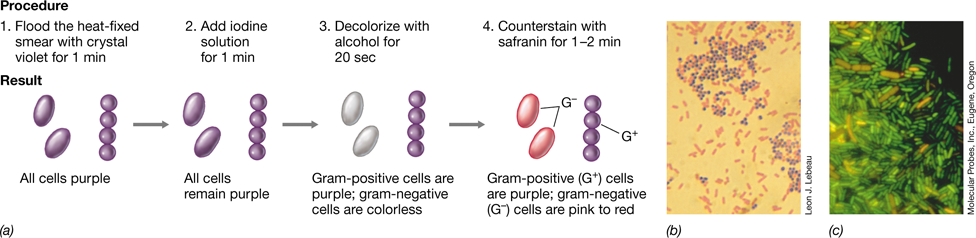
Differential Stains: The Gram Stain
Stains that render different kinds of cells different colors are called differential stains. An important differential-staining procedure used in microbiology is the Gram stain (Figure 1.23). On the basis of their reaction in the Gram stain, bacteria can be divided into two major groups: gram-positive and gram-negative. After Gram staining, gram-positive bacteria appear purple-violet and gram-negative bacteria appear pink (Figure 1.23b). The color difference in the Gram stain arises because of differences in the cell wall structure of gram-positive and gram-negative cells (Section 2.3). Staining with a basic dye such as crystal violet renders cells purple in color. Cells are then treated with ethanol, which decolorizes gram-negative cells but not gram-positive cells. Finally, cells are counterstained with a different-colored stain, typically the red stain safranin. As a result, gram-positive and gram-negative cells can be distinguished microscopically by their different colors (Figure 1.23b).
Figure 1.23 The Gram stain.

(a) Steps in the procedure. (b) Microscopic observation of gram-positive (purple) and gram-negative (pink) bacteria. The organisms are Staphylococcus aureus and Escherichia coli, respectively. (c) Cells of Pseudomonas aeruginosa (gram-negative, green) and Bacillus cereus (gram-positive, orange) stained with a one-step fluorescent staining method. This method allows for differentiating gram-positive from gram-negative bacteria in a single staining step.
The Gram stain is the most common staining procedure used in microbiology, and it is often performed to begin the characterization of a new bacterium. If a fluorescence microscope is available, the Gram stain can be reduced to a one-step procedure; gram-positive and gram-negative cells fluoresce different colors when treated with a special chemical (Figure 1.23c).
Phase-Contrast and Dark-Field Microscopy
Although staining is widely used in light microscopy, staining often kills cells and can distort their features. Two forms of light microscopy improve image contrast of unstained (and thus live) cells. These are phase-contrast microscopy and dark-field microscopy (Figure 1.24). The phase-contrast microscope in particular is widely used in teaching and research for the observation of living preparations.
Figure 1.24 Cells visualized by different types of light microscopy.

The same field of cells of the yeast Saccharomyces cerevisiae visualized by (a) bright-field microscopy, (b) phase-contrast microscopy, and (c) dark-field microscopy. Cells average 8–10 μm wide.
Phase-contrast microscopy is based on the principle that cells differ in refractive index (that is, the ability of a material to alter the speed of light) from their surroundings. Light passing through a cell thus differs in phase from light passing through the surrounding liquid. This subtle difference is amplified by a device in the objective lens of the phase-contrast microscope called the phase ring, resulting in a dark image on a light background (Figure 1.24b; see also inset to Figure 1.20a). The ring consists of a phase plate that amplifies the variation in phase to produce the higher-contrast image.
In the dark-field microscope, light does not pass through the specimen. Instead, light is directed from the sides of the specimen and only light that is scattered when it hits the specimen reaches the lens. Thus, the specimen appears light on a dark background (Figure 1.24c). Dark-field microscopy often has better resolution than light microscopy, and some objects can be resolved by dark-field that cannot be resolved by bright-field or even by phase-contrast microscopes. Dark-field microscopy is a particularly good way to observe microbial motility, as bundles of flagella (the structures responsible for swimming motility) are often resolvable with this technique.
Fluorescence Microscopy
The fluorescence microscope visualizes specimens that fluoresce (emit light). In fluorescence microscopy, cells are made to fluoresce by illuminating them from above with light of a single color. Filters are used so that only fluorescent light is seen, and thus cells appear to glow in a black background (Figure 1.25).
Figure 1.25 Fluorescence microscopy.

(a, b, c, d) Cyanobacteria. The same cells are observed in a and b, and likewise in c and d. The top photos are taken with phase-contrast microscopy and the bottom photos with fluorescence microscopy. The cells fluoresce because they contain chlorophyll a and other pigments. The image in b was generated using a filter specific for the fluorescence of chlorophyll a, while the image in d was generated using a permissive filter that shows fluorescence from a range of pigments that occur naturally in cyanobacteria. (e) Fluorescence photomicrograph of cells of Escherichia coli made fluorescent by staining with the fluorescent dye DAPI, which binds to DNA.
Cells fluoresce either because they contain naturally fluorescent substances such as chlorophyll (autofluorescence, Figure 1.25b, d) or because they have been stained with a fluorescent dye (Figure 1.25e). DAPI (4′,6-diamidino-2-phenylindole) is a widely used fluorescent dye that stains cells bright blue because it complexes with the cell’s DNA (Figure 1.25e). DAPI can be used to visualize cells in their natural habitats, such as soil, water, food, or a clinical specimen. Fluorescence microscopy using DAPI is widely used in clinical diagnostic microbiology and also in microbial ecology for enumerating bacteria in a natural environment or in a cell suspension (Figure 1.25e).
Check Your Understanding
What color will a gram-negative cell be after Gram staining by the conventional method?
What major advantage does phase-contrast microscopy have over staining?
1.9 Imaging Cells in Three Dimensions
Thus far we have only considered forms of microscopy in which the rendered images are two-dimensional. Two methods of light microscopy can render a more three-dimensional image, and in this section we explore these forms of microscopy.
Differential Interference Contrast Microscopy
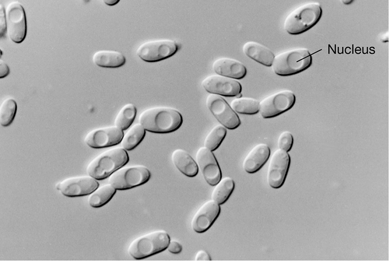
Differential interference contrast (DIC) microscopy is a form of light microscopy that employs a polarizer in the condenser to produce polarized light (light in a single plane). The polarized light then passes through a prism that generates two distinct beams. These beams pass through the specimen and enter the objective lens, where they are recombined into one. Because the two beams pass through substances that differ in refractive index, the combined beams are not totally in phase but instead interfere with each other. This optical effect provides a three-dimensional perspective, which enhances subtle differences in cell structure.
Using DIC microscopy, cellular structures such as the nucleus of eukaryotic cells (Figure 1.26), or various types of inclusions present in some bacterial cells, appear more three-dimensional than in other forms of light microscopy. DIC microscopy is typically used on unstained cells as it can reveal internal cell structures that are nearly invisible by bright-field microscopy without the need for staining (compare Figure 1.24a with Figure 1.26).
Figure 1.26 Differential interference contrast microscopy.

The yeast cells are about 8 μm wide. Note the clearly visible nucleus and compare to the bright-field image of yeast cells in Figure 1.24a.
Confocal Scanning Laser Microscopy
A confocal scanning laser microscope (CSLM) is a computer-controlled microscope that couples a laser to a fluorescence microscope. The laser generates a high-contrast, three-dimensional image and allows the viewer to access several planes of focus in the specimen (Figure 1.27). The laser beam is precisely adjusted such that only a particular layer within a specimen is in perfect focus at one time. Cells struck by the laser fluoresce to generate the image as in fluorescence microscopy (Section 1.8).
Figure 1.27 Confocal scanning laser microscopy.

(a) Confocal image of a microbial biofilm community. The green, rod-shaped cells are Pseudomonas aeruginosa experimentally introduced into the biofilm. Cells of different colors are present at different depths in the biofilm. (b) Confocal image of a filamentous cyanobacterium growing in a soda lake. Cells are about 5 μm wide.
Cells in CSLM preparations can also be stained with fluorescent dyes to make them more distinct (Figure 1.27a). The laser then scans up and down through the layers of the sample, generating an image for each layer. A computer assembles the images to compose the many layers into a single high-resolution, three-dimensional image. Thus, for a relatively thick specimen (such as a bacterial biofilm, Figure 1.27a), not only can cells on the surface of the biofilm be observed, as with conventional light microscopy, but cells in the various layers are also observed by adjusting the plane of focus of the laser beam. CSLM is particularly useful when thick specimens need to be examined.
Check Your Understanding
What structure in eukaryotic cells is more easily seen in DIC than in bright-field microscopy? (Hint: Compare Figures 1.24a and 1.26).
Why is CSLM able to show different layers in a thick preparation while bright-field microscopy cannot?
1.10 Probing Cell Structure: Electron Microscopy
Electron microscopes use electrons instead of visible light (photons) to image cells and cell structures. In the electron microscope, electromagnets function as lenses, and the whole system operates in a vacuum (Figure 1.28). Electron microscopes are fitted with cameras to allow a photograph, called an electron micrograph, to be taken. Two types of electron microscopy are in routine use in microbiology: transmission and scanning.
Figure 1.28 The electron microscope.
This instrument encompasses both transmission and scanning electron microscope functions.
Transmission Electron Microscopy
The transmission electron microscope (TEM) is used to examine cells and cell structure at very high magnification and resolution. The resolving power of a TEM is much greater than that of the light microscope, even allowing one to view structures at the molecular level (Figure 1.29). This is because the wavelength of electrons is much shorter than the wavelength of visible light, and, as we have learned, wavelength affects resolution (Section 1.7). For example, whereas the resolving power of a light microscope is about 0.2 micrometer, the resolving power of a TEM is about 0.2 nanometer, a thousandfold improvement. With such powerful resolution, objects as small as individual protein and nucleic acid molecules can be visualized by transmission electron microscopy (Figure 1.29b).
Figure 1.29 Electron micrographs.

(a) Micrograph of a thin section of a dividing bacterial cell, taken by transmission electron microscopy (TEM). The cell is about 0.8 μm wide. (b) TEM of negatively stained molecules of hemoglobin. Each hexagonal-shaped molecule is about 25 nanometers (nm) in diameter and consists of two doughnut-shaped rings, a total of 15 nm thick. (c) Scanning electron micrograph (SEM) of bacterial cells. A single cell is about 0.75 μm wide.
Unlike photons, electrons are very poor at penetrating; even a single cell is too thick to penetrate with an electron beam. Consequently, to view the internal structure of a cell, thin sections of the cell are needed, and the sections must be stabilized and stained with various chemicals to make them visible. A single bacterial cell, for instance, is cut into extremely thin (20- to 60-nm) slices, which are then examined individually by TEM (Figure 1.29a). To obtain sufficient contrast, the sections are treated with stains such as osmic acid, or permanganate, uranium, lanthanum, or lead salts. Because these substances are composed of atoms of high atomic weight, they scatter electrons well and thus improve contrast. If only the external features of an organism are to be observed, thin sections are unnecessary. Intact cells or cell components can be observed directly in the TEM by a technique called negative staining (Figure 1.29b).
Electron cryotomography (cryoET) is an imaging technique in which TEM is used to obtain three-dimensional images. In cryoET, samples are prepared by rapid freezing to very low temperatures so that they are immobilized in noncrystalline vitreous ice. They are then imaged at very low temperatures (<−150 °C), thereby preserving cell structures in their native states. Finally, samples are tilted as they are being imaged and this series of tilted images is assembled computationally to generate a three-dimensional image of the interior of the cell with a resolution of 4 nm.
Scanning Electron Microscopy
For optimal three-dimensional imaging of cells, a scanning electron microscope (SEM) is used. In scanning electron microscopy, the specimen is coated with a thin film of a heavy metal, typically gold. An electron beam then scans back and forth across the specimen. Electrons scattered from the metal coating are collected and projected on a monitor to produce an image (Figure 1.29c). In the SEM, even fairly large specimens can be observed, and the depth of field (the portion of the image that remains in sharp focus) is extremely good. A wide range of magnifications can be obtained with the SEM, from as low as 15× up to about 100,000×, but only the surface of an object is typically visualized.
Electron micrographs taken by either TEM or SEM are black-and-white images. Although the original image contains the maximum amount of scientific information that is available, color is often added to scanning electron micrographs by manipulating them in a computer. However, such false color does not improve resolution of a micrograph. In this text, false color will be used sparingly in electron micrographs so as to present the micrographs in their original scientific context.
Check Your Understanding
What is an electron micrograph? Why do electron micrographs have greater resolution than light micrographs?
What type of electron microscope would be used to view a cluster of cells? What type would be used to observe internal cell structure?
III: Microbial Cultivation Expands the Horizon of Microbiology
III: Microbial Cultivation Expands the Horizon of Microbiology
III Microbial Cultivation Expands the Horizon of Microbiology
Microbes can be cultured in the laboratory, and microbial cultures have played a major role in unraveling the metabolic diversity and medical importance of the microbial world.
Following the discovery of microorganisms through microscopic methods, advances in microbial cultivation fueled major discoveries in microbiology. Important advances included the development of aseptic technique, a collection of practices that allow for the preparation and maintenance of sterile (that is, without the presence of living organisms) nutrient media and solutions (Chapter 4). Aseptic technique is essential for the isolation and maintenance of pure cultures of bacteria. Pure cultures are those that contain cells from only a single type of microorganism and are of great value for the study of microorganisms. Finally, enrichment culture techniques, which allow for the isolation from nature of microbes having particular metabolic characteristics, facilitate the discovery of diverse microorganisms.
Advances in microbial cultivation are directly responsible for success in fighting infectious disease, the discovery of microbial diversity, and the use of microbes as model systems to discover the fundamental properties of all living cells. Important advances in microbial cultivation occurred in the nineteenth century as microbiologists sought to answer two major questions of that time: (1) Does spontaneous generation occur? (2) What is the nature of infectious disease? Answers to these seminal questions emerged from the work of two giants in the field of microbiology: the French chemist Louis Pasteur and the German physician Robert Koch. We begin with the work of Pasteur.
1.11 Pasteur and Spontaneous Generation
Pasteur was a chemist by training and was one of the first to recognize that many of what were thought to be strictly chemical reactions were actually catalyzed by microorganisms. Pasteur studied the chemistry of crystal formation and he used microscopes to examine crystal structure. His training in chemistry and microscopy prepared him to make a series of foundational discoveries to further the science of microbiology.
The Microbial Basis of Fermentation
Early in his career, Pasteur studied crystals formed during the production of alcohol. Through careful microscopic observation of tartaric acid crystals formed in wine, he observed two types of crystals that had mirror-image structures. He separated these by hand and observed that each type of crystal bent a beam of polarized light in a different direction. In this way he discovered that chemically identical substances can have optical isomers, which have different molecular structures that can influence their properties. Pasteur went on to discover that microorganisms could discriminate between optical isomers. For example, cultures of the mold Aspergillus (Figure 1.30) metabolized exclusively D-tartrate but not its optical isomer, L-tartrate. The fact that a living organism could discriminate between optical isomers led Pasteur to strongly suspect that many reactions previously thought to be abiotic were actually catalyzed by microbes.
Figure 1.30 Louis Pasteur and his observation of isomeric discrimination by ***Aspergillus***.

Light micrograph of cells of the mold Aspergillus.
While a professor of chemistry, Pasteur encountered a local businessman who produced alcohol industrially from beet juice. The businessman was losing money because many of his vats produced, instead of alcohol, a product that smelled like sour milk, which Pasteur determined to be lactic acid. In the mid-nineteenth century the production of alcohol was thought to be solely a chemical process. Pasteur studied the broth with his microscope, but instead of crystals he observed cells. Pasteur observed that the vats that produced alcohol were full of yeast, but the sour vats were full of rod-shaped bacteria. He hypothesized that these were living organisms whose growth produced either alcohol or lactic acid.
Pasteur needed to grow these organisms to prove his hypothesis. He prepared a hot-water extract of yeast cells, deducing that this would contain all of the nutrients that yeast need to grow. He then used a porcelain filter to remove all cells from this yeast extract nutrient medium, rendering it sterile. If he introduced living yeast back into this sterile yeast extract medium, he could observe their growth and show the production of alcohol, but if he instead introduced the small rods, he then observed lactic acid formation. Heating of these cultures eliminated growth and the production of either alcohol or lactic acid. In this way he proved that fermentation is carried out by microorganisms and that different microorganisms perform different fermentation reactions.
During his work on fermentation, Pasteur observed that other organisms would often grow in his yeast extract medium. He deduced that these organisms were being introduced from the air. Pasteur’s work on fermentation had prepared him to conduct a series of classic experiments on spontaneous generation, experiments that are forever linked to his name and which helped establish microbiology as a modern science.
Spontaneous Generation
The concept of spontaneous generation existed for thousands of years and its basic tenet can be easily grasped. If food or some other perishable material is allowed to stand for some time, it putrefies. When examined microscopically, the putrefied material is teeming with microorganisms. From where do these organisms arise? Prior to Pasteur it was common belief that life arose spontaneously from nonliving materials, that is, by spontaneous generation.
Pasteur became a powerful opponent of spontaneous generation. He predicted that microorganisms in putrefying materials were descendants of cells that entered from the air or cells that had initially been present on the decaying materials. Pasteur reasoned that if food were treated in such a way as to destroy all living organisms present—that is, if it were rendered sterile—and if it were kept sterile, it would not putrefy.
Mastering Microbiology
Art Activity: Figure 1.26 Pasteur’s swan-necked flask experiments
Pasteur used heat to kill contaminating microorganisms, and he found that extensive heating of a nutrient solution followed by sealing kept it from putrefying. Proponents of spontaneous generation criticized these experiments by declaring that “fresh air” was necessary for the phenomenon to occur. In 1864 Pasteur countered this objection simply and brilliantly by constructing a swan-necked flask, now called a Pasteur flask (Figure 1.31). In such a flask, nutrient solutions could be heated to boiling and sterilized. After the flask cooled, air could reenter, but the bend in the neck prevented particulate matter (including microorganisms) from entering the nutrient solution and initiating putrefaction. Nutrient solutions in such flasks remained sterile indefinitely. Microbial growth was observed only after particulate matter from the neck of the flask was allowed to enter the liquid in the flask (Figure 1.31c). This experiment settled the spontaneous generation controversy forever.
Figure 1.31 The defeat of spontaneous generation: Pasteur’s swan-necked flask experiment.

In (c) the liquid putrefies because microorganisms enter with the dust. The bend in the flask allowed air to enter (a key objection to Pasteur’s sealed flasks) but prevented microorganisms from entering.
Pasteur’s work on spontaneous generation demonstrated the importance of sterilization and led to the development of effective sterilization procedures that were eventually standardized and applied widely in microbiology, medicine, and industry. For example, the British physician Joseph Lister (1827–1912) deduced from Pasteur’s discoveries that surgical infections were caused by microorganisms. He implemented a range of techniques designed to kill microorganisms and to prevent microbial infection of surgical patients. Lister is credited with the introduction of aseptic techniques for surgeries (1867), and his methods were adopted worldwide; these greatly reduced postoperative infections and greatly improved the survival rate of surgical patients. The food industry also benefited from the work of Pasteur, as his principles were quickly adapted for the preservation of milk and many other foods by heat treatment, which we now call pasteurization.
Other Major Accomplishments of Pasteur
Pasteur went on to many other triumphs in microbiology and medicine. Some highlights include his development of vaccines for the diseases anthrax, fowl cholera, and rabies. Pasteur’s work on rabies was his most famous success, culminating in July 1885 with the first administration of a rabies vaccine to a human, a young French boy named Joseph Meister who had been bitten by a rabid dog. In those days, a bite from a rabid animal was invariably fatal. News spread quickly of the success of Meister’s vaccination, and of one administered shortly thereafter to a young shepherd boy, Jean-Baptiste Jupille (**Figure 1.32*a***). Within a year several thousand people bitten by rabid animals had traveled to Paris to be treated with Pasteur’s rabies vaccine.
Figure 1.32 Louis Pasteur and some symbols of his contributions to microbiology.

(a) A French 5-franc note honoring Pasteur. The shepherd boy Jean-Baptiste Jupille is shown killing a rabid dog that had attacked children. Pasteur’s rabies vaccine saved Jupille’s life. In France, the franc preceded the euro as a currency. (b) Part of the Pasteur Institute, Paris, France. Today this structure, built for Pasteur by the French government, houses a museum that displays some of the original swan-necked flasks used in his experiments and a chapel containing Pasteur’s crypt.
Pasteur’s fame was legendary and led the French government to establish the Pasteur Institute in Paris in 1888 (Figure 1.32b). Originally established as a clinical center for the treatment of rabies and other contagious diseases, the Pasteur Institute today is a major biomedical research center focused on antiserum and vaccine research and production. The medical and veterinary breakthroughs of Pasteur not only were highly significant in their own right but helped solidify the concept of the germ theory of disease, whose principles were being developed at about the same time by a second giant of this era, Robert Koch.
Check Your Understanding
Define the term sterile. What two methods did Pasteur use to make solutions sterile?
How did Pasteur’s experiments using swan-necked flasks defeat the theory of spontaneous generation?
Besides ending the controversy over spontaneous generation, what other accomplishments do we credit to Pasteur?
1.12 Koch, Infectious Diseases, and Pure Cultures
Proof that some microorganisms can cause disease provided the greatest impetus for the development of microbiology as an independent biological science. As early as the sixteenth century it was suspected that some agent of disease could be transmitted from a diseased person to a healthy person. After microorganisms were discovered, a number of individuals proposed that they caused infectious diseases, but skepticism prevailed, and definitive proof was lacking. As early as 1847, the Hungarian physician Ignaz Semmelweis promoted sanitary methods including hand washing as a method for preventing infections. His methods are credited with saving many lives, but he could not prove why these methods worked and his advice was met with scorn by most of the medical community. The work of Pasteur and Lister provided strong evidence that microbes were the cause of infectious disease, but it was not until the work of the German physician Robert Koch (1843–1910) that the germ theory of infectious disease had direct experimental support.
The Germ Theory of Disease and Koch’s Postulates
In his early work Koch studied anthrax, a disease of cattle and occasionally of humans. Anthrax is caused by the bacterium Bacillus anthracis. By careful microscopy and staining, Koch established that the bacteria were always present in the blood of an animal that was succumbing to the disease. However, Koch reasoned that the mere association of the bacterium with the disease was not actual proof of cause and effect, and he seized the opportunity to study cause and effect experimentally using anthrax and laboratory animals. The results of this study formed the standard by which infectious diseases have been studied ever since.
Mastering Microbiology
Art Activity: Figure 1.29 Koch’s Postulates for proving cause and effect in infectious diseases
Koch used mice as experimental animals. Using appropriate controls, Koch demonstrated that when a small drop of blood from a mouse with anthrax was injected into a healthy mouse, the latter quickly developed anthrax. He took blood from this second animal, injected it into another, and again observed the characteristic disease symptoms. However, Koch carried this experiment a critically important step further. He discovered that the anthrax bacteria could be grown in a nutrient medium outside the host and that even after many transfers in laboratory culture, the bacteria still caused the disease when inoculated into a healthy animal.
On the basis of these experiments and others on the causative agent of tuberculosis, Koch formulated a set of rigorous criteria, now known as Koch’s postulates, for definitively linking cause and effect in an infectious disease. Koch’s postulates, summarized in Figure 1.33, stressed the importance of laboratory culture of the putative infectious agent followed by introduction of the suspected agent into virgin animals and recovery of the pathogen from diseased or dead animals. With these postulates as a guide, Koch, his students, and those that followed them discovered the causative agents of most of the important infectious diseases of humans and domestic animals. These discoveries also led to the development of successful treatments for the prevention and cure of many of these diseases, greatly improving the scientific basis of clinical medicine and human health and welfare (Figure 1.13).
Figure 1.33 Koch’s postulates for proving cause and effect in infectious diseases.

Note that following isolation of a pure culture of the suspected pathogen, the cultured organism must both initiate the disease and be recovered from the diseased animal. Establishing the correct conditions for growing the pathogen is essential.
Koch, Pure Cultures, and Microbial Taxonomy
The second of Koch’s postulates states that the suspected pathogen must be isolated and grown away from other microorganisms in laboratory culture (Figure 1.33); in microbiology we say that such a culture is pure. To accomplish this important goal, Koch and his associates developed several simple but ingenious methods of obtaining and growing bacteria in pure culture, and many of these methods are still used today.
Koch started by using natural surfaces such as a potato slice to obtain pure cultures, but he quickly developed more reliable and reproducible growth media employing liquid nutrient solutions solidified with gelatin, and later with agar, an algal polysaccharide with excellent properties for this purpose. Along with his associate Walther Hesse, Koch observed that when a solid surface was incubated in air, masses of microbial cells called colonies developed, each having a characteristic shape and color (Figure 1.34). He inferred that each colony had arisen from a single bacterial cell that had grown to yield the mass of cells (see also Figure 1.3). Koch reasoned that each colony harbored a pure culture (a population of identical cells), and he quickly realized that solid media provided an easy way to obtain pure cultures. Richard Petri, another associate of Koch, developed the transparent double-sided “Petri dish” in 1887, and this quickly became the standard tool for obtaining pure cultures.
Figure 1.34 A hand-colored photograph taken by Walther Hesse of colonies formed on agar.

The colonies include those of molds and bacteria obtained during Hesse’s studies of the microbial content of air in Berlin, Germany, in 1882. From Hesse, W. 1884. “Ueber quantitative Bestimmung der in der Luft enthaltenen Mikroorganismen.” Mittheilungen aus dem Kaiserlichen Gesundheitsamte. 2: 182–207.
Koch was keenly aware of the implications his pure culture methods had for classifying microorganisms. He observed that colonies that differed in color and size (Figure 1.34) bred true and that cells from different colonies typically differed in size and shape and often in their nutrient requirements as well. Koch realized that these differences were analogous to the criteria taxonomists had established for the classification of larger organisms, such as plant and animal species, and he suggested that the different types of bacteria should be considered as “species, varieties, forms, or other suitable designation.” Such insightful thinking was important for the rapid acceptance of microbiology as a new biological science, rooted as biology was in classification during Koch’s era.
Koch and Tuberculosis
Koch’s crowning scientific accomplishment was his discovery of the causative agent of tuberculosis. At the time Koch began this work (1881), one-seventh of all reported human deaths were caused by tuberculosis (Figure 1.13). There was a strong suspicion that tuberculosis was a contagious disease, but the suspected agent had never been seen, either in diseased tissues or in culture. Following his successful studies of anthrax, Koch set out to demonstrate the cause of tuberculosis, and to this end he brought together all of the methods he had so carefully developed in his previous studies with anthrax: microscopy, staining, pure culture isolation, and an animal model system (Figure 1.33).
The bacterium that causes tuberculosis, Mycobacterium tuberculosis, is very difficult to stain because M. tuberculosis cells contain large amounts of a waxlike lipid in their cell walls. Nevertheless, Koch devised a staining procedure for M. tuberculosis cells in lung tissue samples. Using this method, he observed the blue, rod-shaped cells of M. tuberculosis in tubercular tissues but not in healthy tissues (Figure 1.35). Obtaining cultures of M. tuberculosis was not easy, but eventually Koch succeeded in growing colonies of this organism on a solidified medium containing blood serum. Under the best of conditions, M. tuberculosis grows slowly in culture, but Koch’s persistence and patience eventually led to pure cultures of this organism from human and animal sources.
Figure 1.35 Robert Koch’s drawings of ***Mycobacterium tuberculosis***.

(a) Section through infected lung tissue showing cells of M. tuberculosis (blue). (b) M. tuberculosis cells in a sputum sample from a tubercular patient. (c) Growth of M. tuberculosis on a glass plate of coagulated blood serum stored inside a glass box to prevent contamination. (d) M. tuberculosis cells taken from the plate in c and observed microscopically; cells appear as long, cordlike forms (Section 16.11 and Figures 16.32 and 16.33). Original drawings from Koch, R. 1884. “Die Aetiologie der Tuberkulose.” Mittheilungen aus dem Kaiserlichen Gesundheitsamte 2: 1–88.
From this point Koch used his postulates (Figure 1.33) to obtain definitive proof that the organism he had isolated was the cause of the disease tuberculosis. Guinea pigs can be readily infected with M. tuberculosis and eventually succumb to systemic tuberculosis. Koch showed that tuberculous guinea pigs contained masses of M. tuberculosis cells in their lungs and that pure cultures obtained from such animals transmitted the disease to healthy animals. In this way, Koch successfully satisfied all four of his postulates, and the cause of tuberculosis was understood. Koch announced his discovery of the cause of tuberculosis in 1882, and for this accomplishment he was awarded the 1905 Nobel Prize for Physiology or Medicine. Koch had many other triumphs in the growing field of infectious diseases, including the discovery of the causative agent of cholera (the bacterium Vibrio cholerae) and the development of methods to diagnose infection with M. tuberculosis (the tuberculin skin test).
Check Your Understanding
How do Koch’s postulates ensure that cause and effect of a given disease are clearly differentiated?
What advantages do solid media offer for the isolation of microorganisms?
1.13 Discovery of Microbial Diversity
As microbiology entered the twentieth century, its initial focus on basic principles, methods, and medical aspects broadened to include studies of the microbial diversity of soil and water and the metabolic processes that microorganisms carried out in these habitats. Major contributors of this era included the Dutchman Martinus Beijerinck and the Russian Sergei Winogradsky.
Sergei Winogradsky and Chemolithotrophy
Sergei Winogradsky (1856–1953) was interested in the bacterial diversity of soils and waters and was highly successful in isolating several notable bacteria from natural samples. Winogradsky was particularly interested in bacteria that cycle nitrogen and sulfur compounds, such as the nitrifying bacteria and the sulfur bacteria (Chapters 14 and 15). He studied Beggiatoa, which are large sulfur-oxidizing bacteria found in marine sediments. Beggiatoa are morphologically distinctive and readily identified under the microscope (Figure 1.36), but Winogradsky found that they would not grow on the rich nutrient media used by Koch and Pasteur. In order to grow these Beggiatoa, Winogradsky designed a medium that chemically imitated the environment in which Beggiatoa lived. He showed that Beggiatoa are able to grow in the absence of organic nutrients, and that their growth requires only inorganic substances (compounds lacking carbon–carbon bonds). In this way, Winogradsky was the first to define chemolithotrophy, which is any metabolic process in which energy for growth is produced using only inorganic chemical compounds. Winogradsky also revealed that these chemolithotrophic bacteria obtain their carbon from CO2, much like plants, though they get their energy from chemical reactions rather than from light. Winogradsky further showed that these organisms, which he called lithotrophs (meaning, literally, “stone eaters”), are widespread in nature.
Figure 1.36 Sulfur bacteria.

The original drawings were made by Sergei Winogradsky in the late 1880s and then copied and hand-colored by his wife Hèléne. (a) Purple sulfur phototrophic bacteria. Figures 3 and 4 show cells of Chromatium okenii (compare with photomicrographs of C. okenii in Figures 1.1a and 1.11d, and 1.21a). (b) Beggiatoa, a sulfur chemolithotroph (compare with Figure 15.32a).
Martinus Beijerinck, the Enrichment Culture Technique, and Nitrogen Fixation
Martinus Beijerinck (1851–1931) was a professor at the Delft Polytechnic School in Holland and a contemporary of Winogradsky. Beijerinck’s greatest contribution to the field of microbiology was his clear formulation of the enrichment culture technique. Enrichment cultures were used by both Beijerinck and Winogradsky to discover many unique forms of metabolism that we now know to be essential to nutrient cycling in nature.
The growth media used by Pasteur and Koch (for example, yeast extract media) were rich in a wide variety of nutrients. These media support the growth of many microorganisms and are often suitable for growing pathogenic bacteria, but as Winogradsky discovered, most bacteria found in nature do not grow well on such media. Enrichment culture techniques are used to selectively encourage the growth of specific microorganisms. To do this, enrichment cultures employ culture media and selective incubation conditions that are deliberately designed to favor only certain types of microbes.
For example, Winogradsky used enrichment culture to isolate the first nitrogen-fixing bacterium. He devised a liquid growth medium that lacked a source of nitrogen, inoculated this medium with soil, and incubated it in the presence of air. He knew that all living things require nitrogen, and so when he observed bacteria growing in the medium he knew that they must be getting their nitrogen from N2 in the air. In this way he enriched for and ultimately isolated the anaerobic nitrogen-fixing bacterium Clostridium pasteurianum, becoming the first to demonstrate the process of nitrogen fixation. The growth of nitrogen-fixing bacteria was favored in the enrichment culture because only those bacteria that can use nitrogen from the air can grow on media that lack a nitrogen source such as ammonia or nitrate. Beijerinck used a similar technique shortly thereafter to isolate the first aerobic nitrogen-fixing bacterium, Azotobacter (Figure 1.37). Beijerinck also devised a medium to isolate rhizobia and proved that these bacteria cause root nodules to form on legumes and that they perform nitrogen fixation within the nodules (Figure 1.14).
Figure 1.37 Martinus Beijerinck and ***Azotobacter***.

The first aerobic nitrogen-fixing bacterium, Azotobacter chroococcum, was isolated by M. Beijerinck in 1900. The image is a painting by M. Beijerinck’s sister, Henriëtte Beijerinck, showing cells of Azotobacter chroococcum. Beijerinck used such paintings to illustrate his lectures. Compare these drawings of A. chroococcum cells with the photomicrograph of cells of Azotobacter in Figure 15.24.
Lesley Robertson and the Kluyver Laboratory Museum, Delft University of Technology
Using the enrichment culture technique, Beijerinck and Winogradsky isolated the first pure cultures of many important soil and aquatic microorganisms. For example, Winogradsky isolated the first nitrifying bacteria using an enrichment medium that contained ammonium salts and CO2 since these chemolithotrophic bacteria oxidize ammonium as an energy source and are autotrophic. Beijerinck was the first to isolate sulfur-cycling bacteria such as sulfate-reducing bacteria and sulfur-oxidizing bacteria, fermentative bacteria such as the lactic acid bacteria, and many other physiological types of bacteria as well as microbial eukaryotes such as green algae.
Beijerinck was also the first person to observe a virus. While studying “mosaic disease” of tobacco, Beijerinck used selective filters to show that the infectious agent in this disease was smaller than any bacterium and that it somehow became incorporated into cells of the living host plant. In this insightful work, Beijerinck described not only the first virus, but also the basic principles of virology, which we expand upon in Chapters 5 and 11.
Check Your Understanding
What is meant by the term “chemolithotrophy”? In what way are chemolithotrophs like plants?
IV: Molecular Biology and the Unity and Diversity of Life
IV: Molecular Biology and the Unity and Diversity of Life
IV Molecular Biology and the Unity and Diversity of Life
Molecular analyses of microbial cells have unveiled the biochemical principles that govern all life and the evolutionary position of microbes in the tree of life.
The development of aseptic technique and methods for the enrichment, isolation, and propagation of bacteria at the end of the nineteenth century gave rise to explosive growth in the pace of microbiological discovery. Moreover, microbiologists realized that the ability to grow bacteria rapidly and in controlled laboratory conditions made them excellent model systems for exploring the fundamental nature of life.
1.14 Molecular Basis of Life
Experiments with bacterial cultures in the twentieth century were critical in forming the foundations of molecular biology, molecular genetics, and biochemistry. Microbiologists came to realize that while microorganisms were incredibly diverse, all cells appeared to operate on similar basic principles. Thus, the search was on for the overarching molecular processes that define life itself.
Unity in Biochemistry
Albert Jan Kluyver (1888–1956) was Beijerinck’s successor at what was then called the Delft Institute of Technology. Kluyver recognized that though microbial diversity was tremendous, all microorganisms used many of the same biochemical pathways and their metabolic processes were governed by similar thermodynamic constraints. Kluyver promoted the study of comparative biochemistry to identify the unifying features of all cells. He famously proclaimed, “From elephant to butyric acid bacterium—it is all the same!” This was later reformulated by Jacques Monod (1910–1976) into the expression, “What is true for E. coli is also true for the elephant,” a statement that proclaimed the importance of working with bacteria to understand the fundamental principles that govern all living things.
The use of microbes as metabolic model systems led to the discovery that certain macromolecules and biochemical reactions are universal, and that to understand their function in one cell is to understand their function in all cells. We will learn about these fundamental macromolecules and biochemical processes in later chapters. These discoveries were of central importance to understanding microbial evolution, and none were more important than the discovery of DNA as the molecular basis of heredity, a discovery that is less than 80 years old.
Cracking the Code of Life
In the early twentieth century, it was clear that some molecule carried the hereditary information from parent to offspring, but the molecular basis of heredity remained a mystery. Most biologists thought that proteins carried this hereditary information. DNA had been discovered but it was thought to be merely a structural molecule, and too simple in its composition to encode cellular functions. The hunt for the molecular basis of heredity began in earnest with a key experiment by the British bacteriologist Frederick Griffith (1879–1941).
Griffith worked with a virulent strain of Streptococcus pneumoniae, a cause of bacterial pneumonia in both humans and mice. This strain, strain S, produced a polysaccharide coat (that is, a capsule, Section 2.6) that caused cells to form smooth colonies on agar media and conferred the ability to kill infected mice (**Figure 1.38*a***). A related strain, strain R, lacked this polysaccharide and produced “rough” colonies that did not cause disease. However, Griffith observed that strain R could be transformed to type S, forming smooth colonies and causing disease, when it was mixed with the dead remains of cells of strain S (Figure 1.38a). He reasoned that some molecule that contained genetic information must have been transferred from strain R to strain S in this process, and this experiment showed that genetic transfer could be studied in bacteria.
Figure 1.38 Early evidence that DNA is the molecular basis of heredity.

(a) Griffith’s experiment showed that bacteria can transfer genetic information. Streptococcus pneumoniae strain R makes rough colonies on agar media and does not kill mice, but strain S makes smooth colonies and does kill mice. Heat-killed cells of strain S do not cause disease, but if these killed cells are mixed with cells of strain R, then strain R is “transformed” to the S type and begins to make smooth colonies and kill mice. (b) The Avery–MacLeod–McCarty experiment showed that DNA contains genetic information. DNA isolated from strain S can transform strain R to cause disease, though the DNA itself does not cause disease. Degraded DNA (the control in the experiment) lacks the ability to transform strain R.
Later, the Avery–MacLeod–McCarty experiment (1944), named for three scientists at the Rockefeller University, would show that this “transforming principle” was DNA. They treated the dead remains of cells of strain S with chemicals and enzymes that destroyed protein and left behind only DNA. They then repeated Griffith’s experiment with the pure DNA of strain S and showed that this DNA was sufficient to cause transformation, causing strain R cells to become S-type cells and virulent (Figure 1.38b). They also demonstrated that transformation failed if the DNA from strain S was degraded. Collectively, these experiments proved that DNA is the genetic material of cells.
The discovery that DNA is the basis of heredity was followed by intense efforts to understand how this molecule stores genetic information. The structure of DNA was ultimately solved by James D. Watson (1928–) and Francis Crick (1916–2004) using X-ray diffraction images of DNA taken by their colleague Rosalind Franklin (1920–1958). They revealed that DNA is composed of a double helix that contains four nitrogenous bases: guanine, cytosine, adenine, and thymine, which form the genetic code (Section 6.1). Later research would reveal how the genetic code is read from DNA and translated into protein, and these principles are covered in Chapter 6. Once again, however, this research to crack the code of life was enabled by a microbial model system, in this case, the bacterium Escherichia coli.
From DNA to Evolutionary Insight
Not long after the discovery that genetic information is encoded in the sequence of biological molecules, Emile Zuckerkandl (1922–2013) and Linus Pauling (1901–1994) proposed that molecular sequences could be used to reconstruct evolutionary relationships. They recognized that evolution, as described by Darwin, required variation in offspring and that these variations must be caused by changes in molecular sequences. They predicted that these sequence differences occur randomly in a clocklike fashion over time. This led to the conclusion that the evolutionary history of organisms is inscribed in the sequence of molecules such as DNA. Carl Woese seized upon these insights to pursue the ambitious goal of reconstructing the evolutionary history of all cells, and we explore the events that led up to this seminal discovery in evolutionary biology now.
Check Your Understanding
Describe the experiments that proved DNA was the transforming principle described by Griffith.
Why are microbes useful as model organisms to understand basic principles of biology?
1.15 Woese and the Tree of Life
Evolutionary relationships between microorganisms remained a mystery until it was discovered that certain molecular sequences maintain a record of evolutionary history. Here we will examine how the sequence of ribosomal RNA (rRNA) genes, present in all cells, revolutionized the understanding of microbial evolution and made it possible to construct the first universal tree of life. Ribosomal RNAs are components of the ribosome, the biosynthetic machine that all cells use to make proteins (Section 1.2).
Early Attempts to Portray the Evolutionary History of Life
For over a hundred years, following the 1859 publication of Charles Darwin’s On the Origin of Species, evolutionary history was studied primarily with the tools of paleontology (through examining fossils) and comparative biology (through comparing the traits of living organisms). These approaches led to progress in understanding the evolution of plants and animals, but they were powerless to explain the evolution of microorganisms. The vast majority of microorganisms do not leave behind fossils, and their morphological and physiological traits provide few clues about their evolutionary history. Moreover, microorganisms do not share any morphological traits with plants and animals. Thus, it was impossible to create a robust evolutionary framework that included microorganisms.
The first attempt to depict the common evolutionary history of all living cells was published by Ernst Haeckel in 1866 (Figure 1.39). Haeckel correctly suggested that single-cell organisms, which he called Monera (labeled as Moneres near the root of the tree shown in Figure 1.39), were ancestral to other forms of life, but his scheme, which included plants, animals, and protists, did not attempt to resolve evolutionary relationships among microorganisms. The situation was little changed as late as 1969 when Robert Whittaker proposed a five-kingdom classification scheme (Figure 1.40). Whittaker’s scheme distinguished the fungi as a distinct lineage, but it was still largely impossible to resolve evolutionary relationships among most microorganisms. Hence, our understanding of microbial evolution had made little progress since Haeckel’s day.
Figure 1.39 Early efforts to depict the universal tree of life.

A conceptual tree of life proposed by Ernst Haeckel in Generelle Morphologie der Organismen (1866) shortly after Darwin published On the Origin of Species (1859). Haeckel proposed that four main groups of organisms (Plantae, Animalia, Protista, and Moneres) evolved from a common single-celled ancestor. The group that Haeckel proposed as Moneres included all microbes having prokaryotic cell structure, which we know today as Bacteria and Archaea.
Figure 1.40 Later efforts to depict the universal tree of life.

A conceptual tree of life published by Robert H. Whittaker (1969). Whittaker recognized the same groups as Haeckel (Figure 1.39), but he was the first to propose adding Fungi as a distinct kingdom. Compare this tree to Haeckel’s tree and consider how little our knowledge of microbial evolution had changed between 1866 and 1969, prior to the development of nucleic acid sequencing.
Molecular Sequence Data Revolutionized Microbial Phylogeny
Everything changed after the structure of DNA was discovered and it was recognized that evolutionary history is recorded in DNA sequences. Carl Woese (1928–2012), a professor at the University of Illinois (USA), realized in the 1970s that the sequence of ribosomal RNA (rRNA) molecules and the genes that encode them could be used to infer evolutionary relationships between organisms. Woese recognized that genes encoding rRNAs were excellent candidates for phylogenetic analysis because they were (1) present in all cells, (2) functionally constant, (3) highly conserved (slowly changing) in their nucleic acid sequences, and (4) of adequate length to provide a deep view of evolutionary relationships.
Woese compared the sequences of rRNA molecules from many microorganisms. Among the first microbes he examined were methanogens. To his astonishment, he found that the rRNA sequences from methanogens were distinct from those of both Bacteria and Eukarya, the only two domains recognized at that time. He named this new group of prokaryotic cells the Archaea (originally Archaebacteria) and recognized them as the third domain of life alongside the Bacteria and the Eukarya (**Figure 1.41*b***). More importantly, Woese demonstrated that the analysis of rRNA gene sequences could be used to reveal evolutionary relationships between all cells, providing the first effective tool for the evolutionary classification of microorganisms.
Figure 1.41 Evolutionary relationships and the phylogenetic tree of life.

(a) The technology behind ribosomal RNA gene phylogenies. (1) DNA is extracted from cells. (2) Copies of the gene encoding rRNA are made by the polymerase chain reaction (PCR, a technique for making multiple copies of nucleic acid sequences, Section 12.1). (3) The gene is sequenced, and (4) the sequence is aligned and analyzed with sequences of the same gene from other organisms. A computer algorithm makes pairwise comparisons at each base and (5) generates a phylogenetic tree that depicts evolutionary relationships. In the example shown, the sequence differences are highlighted in yellow and are as follows: organism 1 versus organism 2, three differences; 1 versus 3, two differences; 2 versus 3, four differences. Thus organisms 1 and 3 are closer relatives than are 2 and 3 or 1 and 2. (b) The phylogenetic tree of life based on the analysis of rRNA gene sequences as proposed by Carl Woese (adapted from Woese, C.R., et al. PNAS 87: doi.org/10.1073/pnas.87.12.4576). The tree shows the three domains of organisms and a few representative groups in each domain. See detailed discussion of using molecular sequences as phylogenetic tools in Section 13.11.
The Tree of Life Based on rRNA Gene Sequences
The universal tree of life based on rRNA gene sequences (Figure 1.41b) is a genealogy of all life on Earth. It is a true phylogenetic tree, a diagram that depicts the evolutionary history—the phylogeny—of all cells and clearly reveals the three domains. The root of the universal tree represents a point in time when all extant life on Earth shared a common ancestor, the last universal common ancestor, LUCA (Figures 1.10b and 1.41b). From the last universal common ancestor of all cells, evolution proceeded along two paths to form the domains Bacteria and Archaea. At some later time, the domain Archaea diverged to distinguish the Eukarya from the Archaea (Figures 1.10b and 1.41b). The three domains of cellular life are evolutionarily distinct and yet they share features indicative of their common descent from a universal cellular ancestor.
Revealing the Extent of Microbial Diversity
The tools Woese developed to build the tree of life were first used to determine the evolutionary history of microorganisms grown in pure culture (Figure 1.41a). However, Norman Pace (1942–), a professor at the University of Colorado (USA), realized that Woese’s approach could be applied to rRNA molecules isolated directly from the environment as a way to probe the diversity of natural microbial communities without first cultivating their component organisms (Chapter 19). These cultivation-independent methods of rRNA gene analysis pioneered by Pace greatly improved our picture of microbial diversity (Figure 1.42). Despite many advances in the culturing of microorganisms from nature, many microbes have not yet been brought into laboratory culture. Hence, cultivation-independent methods provide us with valuable insights into the diversity and activities of microbes found in nature and oftentimes provide crucial hints for how they can be cultured. We also now know that although much progress has been made in describing microbial diversity, much of this diversity is yet to be explored and thus our work to describe the microbial world has only just begun.
Figure 1.42 Analysis of environmental rRNA genes leads to discovery of new microbial species.

(a) Norman Pace in 1985 described the first approach to sequence rRNA genes obtained directly from the environment without the need to grow microbes by cultivation in the laboratory. (b) Cultivation-independent analysis of microbial communities in environmental samples has revealed a tremendous diversity of microbial species. The application of cultivation-independent methods has been facilitated greatly by improvements in DNA sequencing technology over time. The graph shows the discovery over time of 16S rRNA gene sequences from natural samples that represent unique microbial species (data adapted from Schloss, P.D., et al. mBio 6: doi.org/10.1128/mBio.00201-16).
Improvements in DNA sequencing technology have greatly improved our ability to study Bacteria and Archaea (Figure 1.42b). While the analysis of rRNA genes remains a cornerstone of microbial ecology, we now have the ability to sequence entire microbial genomes instead of just individual genes. Jo Handelsman (1959–), a professor at the University of Wisconsin (USA), was the first to propose metagenomics, a technique in which fragments of microbial genomes (or even entire genomes) can be recovered from a sample of environmental DNA. Metagenomics, the study of genomic information recovered directly from the environment, is currently providing us with profound insights on the evolution and diversity of life and new information on the metabolic potential of the microbial world.
With an evolutionary framework of the microbial world in place, and with powerful new methods in hand to guide future research, advances in microbial diversity are happening quickly. Besides unveiling the previously hidden concept of three evolutionary domains of life, the contributions of Carl Woese and his associates have given microbiologists the tools they need to understand and explore the diversity of our microbial world. In the chapters that follow we will learn how microbiologists have unraveled many of the basic principles that govern living systems, how microorganisms control essential processes that sustain our biosphere, and how we can apply the lessons of microbiology to combat disease and improve our world.
Check Your Understanding
What kinds of evidence support the three-domain concept of life?
List three reasons why rRNA genes are suitable for phylogenetic analyses.
Tiny Cells
Viruses are very small microbes and range in diameter from as small as 20 nm to almost 750 nm. Although no cells exist that are as small as most viruses, the recent discovery of ultra-small bacterial cells1,2 has pushed the lower limits of cell size to what microbiologists feel must be very close to the minimal value. And, because microbiologists today can deduce amazing amounts of information about cells in nature without culturing them, the lack of laboratory cultures of these tiny cells has been only a minor impediment to understanding their biology in detail.
Microbiologists collected groundwater, which travels through Earth’s deep subsurface, from a Colorado (USA) aquifer (Figure 1) and passed it through a membrane filter whose pores were only 0.2 μm in diameter. The liquid that passed through the filter was then subjected to microbiological analyses. Surprisingly, since filters with 0.2-μm pores have been used for decades to remove bacterial cells from solutions to generate “sterile solutions,” prokaryotic cells were present in the groundwater filtrate. In fact, a diverse array of Bacteria were present in the filtrate, revealing that the groundwater was inhabited by a microbial community of tiny cells1 that microbiologists have come to call ultramicrobacteria.
Figure 1 Sampling the anoxic groundwater aquifer that parallels the Colorado River near Rifle, Colorado.

Electron cryotomography, a microscopic technique in which a specimen is examined at extremely cold temperatures without fixation (chemical treatment that can alter a cell’s morphology, see Section 1.10), showed the groundwater ultramicrobacteria to consist primarily of oval-shaped cells about 0.2 μm in diameter (Figure 2). The volume of these cells was calculated to be about 1/200 that of a cell of the bacterium Escherichia coli (see Table 1.1) such that more than 200 of the small cells could fit into one E. coli cell! Each of the tiny cells contained about 50 ribosomes, which is also about 1/100 of the number present in a slowly growing (100-min generation time) cell of E. coli. The very small size of the groundwater ultramicrobacteria gives them an enormous surface-to-volume ratio, and it is hypothesized that this advantage benefits them in extracting resources from their nutrient-deficient habitat.
Figure 2 A tiny bacterial cell from anoxic ground‑water that passed through a filter with 0.2-μm pores.

Despite the fact that the tiny groundwater bacteria have yet to be cultured in the laboratory, much is already known about them because their small genomes—less than 1 megabase (Mb) in size—were obtained and analyzed.2 From a phylogenetic perspective, the different species detected were distantly related to major phyla of Bacteria known from environmental analyses of diverse environments but which have thus far defied laboratory culture. Further analyses showed that genes encoding the enzymes for several core metabolic pathways widely distributed among microorganisms were absent from the genomes of the groundwater ultramicrobacteria. This suggests a metabolically minimalist lifestyle for these tiny cells and a survival strategy of cross-feeding essential nutrients with neighboring species in their microbial community.
A strategy of obtaining nutrients from other organisms is one widely used in the microbial world. As we will see later in this book, many disease-causing (pathogenic or parasitic) bacteria have very small genomes that are missing many key genes otherwise necessary for a free-living lifestyle. However, the pathogenic or parasitic way of life of these microbes lets them "get away" with a minimal genomic complement because any essential molecules they are unable to biosynthesize are supplied by the host.
Although we do not yet know exactly how small a microbial cell can be, microbiologists are closing in on this number from environmental analyses such as the Colorado groundwater study. From the same samples that yielded ultra-small Bacteria in this study, ultra-small Archaea were also detected and found to contain small and highly reduced genomes.2
It is thus likely that a large diversity of very small prokaryotic cells occurs in nature, and from the continued study of these tiny cells, more precise values for both the lower limits to cell size and the minimal genomic requirements for life should emerge. Moreover, theoretical considerations of cell size have shown that DNA and proteins dominate the volume of very small cells and that the theoretical lower limit to cell size agrees closely with the smallest bacteria observed in nature thus far.3
1Luef, B., et al. 2015. Nat. Commun. doi:10.1038/ncomms7372.
2Castelle, C.J., et al. 2015. Curr. Biol. 25: 1–12.
3Kempes, C.P., et al. 2016. ISME J. 10: 2145–2157.
Chapter Review
Go to Mastering Microbiology for videos, animations, practice tests, and more.
I Exploring the Microbial World
1.1 Microorganisms are single-celled microscopic organisms that are essential for the well-being and functioning of other life forms and the planet. The tools of microscopy, microbial cultivation, molecular biology, and genomics are cornerstones of modern microbiology.
Q What are bacterial colonies and how are they formed?
1.2 Prokaryotic and eukaryotic cells differ in cellular architecture, and an organism’s characteristics are defined by its complement of genes—its genome. All cells have a cytoplasmic membrane, a cytoplasm, ribosomes, and a double-stranded DNA genome. All cells carry out activities including metabolism, growth, and evolution.
Q What cellular structures distinguish prokaryotic and eukaryotic cells? What are some differences between a cell wall and a cytoplasmic membrane? In what types of organisms would you expect to find these structures?
1.3 Prokaryotic cells, while typically smaller than eukaryotic cells, can range widely in size and morphology. The most common shapes of prokaryotic cells are rods, cocci, and spirilla, and while most bacteria are 0.5 to 10 μm in length, some can be more than 600 μm long.
**Q How do diffusion and surface-to-volume (S/V) ratio influence the size and shape of prokaryotic cells?**
1.4 Bacteria, Archaea, and Eukarya are the major phylogenetic lineages (domains) of cells. The greatest diversity of microorganisms is found in the Bacteria, while many extremophiles are found within the Archaea. Microbial eukaryotes can vary tremendously in size, with some species being smaller than bacteria. Viruses are acellular and because of this cannot be placed on the tree of life.
**Q What features (or lack of features) can be used to distinguish between viruses, Bacteria, Archaea, and Eukarya?**
1.5 Diverse microbial populations were widespread on Earth for billions of years before plants and animals appeared. Microbes are abundant in the biosphere, and their activities greatly affect the chemical and physical properties of their habitats.
Q How has Earth changed over its history? How have microorganisms contributed to these changes?
1.6 Microorganisms can be both beneficial and harmful to humans, although many more microorganisms are beneficial (or even essential) than are harmful. Agriculture, food, energy, and the environment are all affected in major ways by microorganisms.
Q How would you convince a friend that microorganisms are much more than just agents of disease? What are some of the benefits that microbes provide?
II Microscopy and the Origins of Microbiology
1.7 Microscopes are essential for studying microorganisms. Bright-field microscopy, the most common form of microscopy, employs a microscope with a series of lenses to magnify and resolve the image. The limit of resolution for a light microscope is about 0.2 μm.
Q What is the difference between magnification and resolution? Can either increase without the other?
1.8 An inherent limitation of bright-field microscopy is the lack of contrast between cells and their surroundings. This problem can be overcome by the use of stains or by alternative forms of light microscopy, such as phase contrast or dark field.
Q What is the function of staining in light microscopy? What is the advantage of phase-contrast microscopy over bright-field microscopy?
1.9 Differential interference contrast (DIC) microscopy and confocal scanning laser microscopy allow enhanced three-dimensional imaging or imaging through thick specimens.
Q How is confocal scanning laser microscopy different from fluorescence microscopy? In what ways are they similar? How does differential interference contrast microscopy differ from bright-field microscopy?
1.10 Electron microscopes have far greater resolving power than do light microscopes, the limits of resolution being about 0.2 nm. The two major forms of electron microscopy are transmission, used primarily to observe internal cell structure, and scanning, used to examine the surface of specimens.
Q What are the major differences between electron microscopes and light microscopes? What type of electron microscope would be used to view the three-dimensional features of a cell?
III Microbial Cultivation Expands the Horizon of Microbiology
1.11 Louis Pasteur devised ingenious experiments proving that living organisms cannot arise spontaneously from nonliving matter. Pasteur introduced many concepts and techniques central to the science of microbiology, including sterilization, and developed a number of key vaccines for humans and other animals.
Q Explain the principle behind the Pasteur flask in studies on spontaneous generation. Why were the results of this experiment inconsistent with the theory of spontaneous generation?
1.12 Robert Koch developed a set of criteria called Koch’s postulates for linking cause and effect in infectious diseases. Koch also developed the first reliable and reproducible means for obtaining and maintaining microorganisms in pure culture.
Q What are Koch’s postulates and how did they influence the development of microbiology? Why are Koch’s postulates still relevant today?
1.13 Martinus Beijerinck and Sergei Winogradsky explored soil and water for microorganisms that carry out important natural processes, such as nutrient cycling and the biodegradation of particular substances. Out of their work came the enrichment culture technique and the concepts of chemolithotrophy and nitrogen fixation.
Q What were the major microbiological interests of Martinus Beijerinck and Sergei Winogradsky? It can be said that both men discovered nitrogen fixation. Explain.
IV Molecular Biology and the Unity and Diversity of Life
1.14 All cells share certain characteristics, and microorganisms are used as model systems to explore the fundamental processes that define life. The discoveries of DNA as the molecular basis of heredity, and of its structure and function, paved the way for progress in molecular genetics, microbial phylogeny, and genomics.
Q Describe the experiments that proved DNA to be the molecule at the basis of heredity.
1.15 Carl Woese discovered that ribosomal RNA (rRNA) sequences can be used to determine the evolutionary history of microorganisms, and in so doing, he constructed a modern phylogenetic tree of life that revealed a new domain, the Archaea. Analysis of rRNA sequences from the environment has shown that microbial diversity is exceptional and that the majority of microorganisms have not yet been cultivated.
**Q What insights led to the reconstruction of the tree of life? Which domain, Archaea or Eukarya, is more closely related to Bacteria? What evidence is there to justify your answer?**
Application Questions
Pasteur’s experiments on spontaneous generation contributed to the methodology of microbiology, understanding of the origin of life, and techniques for the preservation of food. Explain briefly how Pasteur’s experiments affected each of these topics.
Describe the lines of proof Robert Koch used to definitively associate the bacterium Mycobacterium tuberculosis with the disease tuberculosis. How would his proof have been flawed if any of the tools he developed for studying bacterial diseases had not been available for his study of tuberculosis?
Imagine that all microorganisms suddenly disappeared from Earth. From what you have learned in this chapter, why do you think that animals would eventually disappear from Earth? Why would plants disappear? By contrast, if all higher organisms suddenly disappeared, what in Figure 1.10a tells you that a similar fate would not befall microorganisms?
Chapter Glossary
a series of steps taken to prevent contamination of laboratory cultures and media Cell wall
a rigid layer present outside the cytoplasmic membrane; it confers structural strength on the cell Chemolithotrophy
a form of metabolism in which energy is generated from the oxidation of inorganic compounds and carbon is obtained typically from CO2 Chromosome
a genetic element, usually circular in prokaryotic cells, carrying genes essential to cellular function Colony
a macroscopically visible population of cells growing on solid medium, arising from a single cell Contrast
the ability to resolve a cell or structure from its surroundings Culture
a collection of microbial cells grown using a nutrient medium Cytoplasm
the fluid portion of a cell, enclosed by the cytoplasmic membrane Cytoplasmic membrane
a semipermeable barrier that separates the cell interior (cytoplasm) from the environment Differentiation
modification of cellular components to form a new structure, such as a spore Domain
one of the three main evolutionary lineages of cells: the Bacteria, the Archaea, and the Eukarya DNA replication
the process by which information from DNA is copied into a new strand of DNA Enrichment culture
a culture that employs highly selective laboratory methods for obtaining microorganisms from natural samples Enzyme
a protein (or in some cases an RNA) catalyst that functions to speed up chemical reactions Eukaryotic
having a membrane-enclosed nucleus and various other membrane-enclosed organelles; cells of Eukarya Evolution
a change over time in gene sequence and frequency within a population of organisms, resulting in descent with modification Extremophiles
microorganisms that inhabit environments characterized by extremes of temperature, pH, pressure, or salinity Genome
the total complement of genes contained in a cell or virus Gram-negative
a bacterial cell with a cell wall containing small amounts of peptidoglycan and an outer membrane Gram-positive
a bacterial cell whose cell wall consists chiefly of peptidoglycan; it lacks the outer membrane of gram-negative cells Gram stain
a differential staining procedure that stains cells either purple (gram-positive cells) or pink (gram-negative cells) Growth
in microbiology, an increase in cell number with time Gut microbiome
the microbial communities present in the animal gastrointestinal tract Horizontal gene transfer
the unidirectional transfer of genes between cells through a process uncoupled from reproduction Intercellular communication
interactions between cells using chemical signals Koch’s postulates
a set of criteria for proving that a given microorganism causes a given disease Macromolecules
a polymer of monomeric units, for example proteins, nucleic acids, polysaccharides, and lipids Magnification
the optical enlargement of an image Medium (plural, media)
in microbiology, the liquid or solid nutrient mixture(s) used to grow microorganisms Metabolism
all biochemical reactions in a cell Microbial community
two or more populations of cells that coexist and interact in a habitat Microbial ecology
the study of the interaction of microorganisms with each other and their environment Microorganism
an organism that is too small to be seen by the unaided human eye Morphology
the physical appearance of a cell as determined by cell shape, for example: rod, coccus, or spirillum Motility
the movement of cells by some form of self-propulsion Nucleoid
the aggregated mass of DNA that makes up the chromosome(s) of prokaryotic cells Nucleus
a membrane-enclosed structure in eukaryotic cells that contains the cell’s DNA genome Organelle
a bilayer-membrane-enclosed structure such as the mitochondrion, found in eukaryotic cells Pathogen
a diagram that depicts the evolutionary history of an organism; consists of nodes and branches Phylogeny
an extrachromosomal genetic element that is usually not essential to the cell Prokaryotic
lacking a membrane-enclosed nucleus and other organelles; cells of Bacteria or Archaea Pure culture
a culture containing a single kind of microorganism Resolution
the ability to distinguish two objects as distinct and separate when viewed under the microscope Ribosomal RNA (rRNA)
the types of RNA found in the ribosome; some participate actively in protein synthesis Ribosome
a cytoplasmic particle composed of ribosomal RNA and protein, whose function is to synthesize proteins Spontaneous generation
the hypothesis that living organisms can originate from nonliving matter Sterile
free of all living organisms (cells) and viruses Transcription
the synthesis of an RNA molecule complementary to one of the two strands of a double-stranded DNA molecule Translation
the synthesis of protein by a ribosome using the genetic information in a messenger RNA as a template