Bacteriophages Mimicking Eukaryotes—Discovery of a Phage-Encoded Nucleus and Spindle
A membrane-bound nucleus is a defining feature of eukaryotic cells that differentiates them from their prokaryotic counterparts. Remarkably, molecular biologists have discovered a group of large Pseudomonas bacteriophages whose genomes encode structures resembling the eukaryotic nucleus and mitotic spindle. The obvious question is why, and to understand these remarkable events, scientists needed to understand the roles of both eukaryotic-like structures.
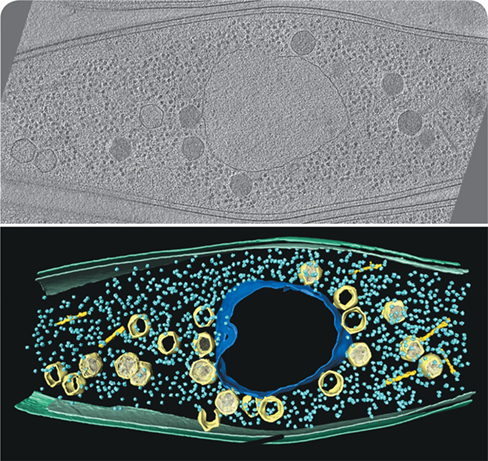
Using fluorescent labeling and cryo-electron microscopy, it was determined that the phage-encoded nucleus contains enzymes specific for replicating and transcribing phage DNA. Once empty phage capsids are assembled at the host membrane, they move rapidly toward the phage nucleus to be filled with phage DNA. The top photo shows a cryo-tomogram slice of a Pseudomonas chlororaphis cell infected with the phage 201ϕ2-1 (phage virions are about 0.1 μm in diameter), and the bottom image highlights the following structures in color: nucleus-like structure (blue), capsids (light yellow), phage tails (bright yellow), cytoplasmic and outer membranes (green), and ribosomes (light blue).
How do the capsids move specifically toward the nucleus? The phage-encoded spindle structure (not shown) consists of tubulin filaments that use a treadmill action to advance the empty capsids to the nucleus for DNA packaging. Thus, the phage-encoded nucleus appears to function as a compartment similar to that of eukaryotic viral factories. Once fully assembled, the mature bacteriophages lyse their bacterial host and hunt for new bacterial prey.
Because of the large genomes of the phages that produce these eukaryotic-like structures, it is possible that the events described here increase the efficiency of large bacteriophage multiplication such that they can better compete with smaller bacteriophages that are completely reliant on the host cell machinery for replication. This discovery also begs one of the most fascinating of viral genomics questions: Was the nucleus actually a viral invention?
Sources: Chaikeeratisak, V., et al. 2017. Assembly of a nucleus-like structure during viral replication in bacteria. Science 335: 194.
Chaikeeratisak, V., et al. 2019. Viral capsid trafficking along treadmilling tubulin filaments in bacteria. Cell 177: 1771.
I Viral Genomes and Classification
Viruses break several of the molecular rules that govern cells, but perhaps the greatest of these is in the nature of their genomes. In some viral genomes, DNA—the genetic blueprint of cells—is replaced by RNA and the double helix is supplanted by a single strand of nucleic acid.
We learned in Chapter 5 that viruses have either DNA or RNA genomes that can be either single-stranded or double-stranded. Thus, compared with cells, viral genomes can create some unusual challenges for genetic information flow. In this chapter, we take a more detailed look at viral biology. We begin by grouping viruses by their genome structure rather than by the hosts they infect, because viruses with the same genome structure face common problems in genetic information flow. We then consider which viruses infect cells in each of the domains of life and conclude with a discussion of infective agents that are neither cells nor viruses.
11.1 Size and Structure of Viral Genomes
Viral genomes vary almost a thousand-fold in size from smallest to largest and are typically smaller than those of cells. DNA viruses exist along this entire gradient from the tiny circovirus, whose 1.75-kilobase single-stranded genome, which contains three genes, pales in comparison to that of the 2.5-megabase-pair double-stranded DNA genome of Pandoravirus, which contains 2500 genes (Figure 11.1). The genome of the latter is over twice that of the previously known largest virus (Mimivirus, see Figure 11.5a) and is larger than the genomes of several species of Bacteria and Archaea (Table 10.1). Pandoravirus infects certain marine amoebae, and with virion dimensions of 1 μm×0.5 μm, it is also larger than some bacterial cells (Figure 11.1).
Figure 11.1 Comparative genomics.

Genome sizes of select viroids, viruses, and archaeal and bacterial cells. Inset: Micrograph of Pandoravirus, ∼1 μm in length. Image courtesy of Chantal Abergel, IGS, UMR7256 CNRS-AMU. Archaeal viruses APBV1 and ATV infect Aeropyrum pernix and Acidianus convivator, respectively. Bacteriophage phi6 and phage G infect Pseudomonas and Bacillus species, respectively; other bacterial viruses infect Escherichia coli.
RNA virus genomes, whether single- or double-stranded, are typically smaller than DNA viruses and contain 2–40 genes. Although some viral genomes are larger than those of some prokaryotic cells, genomes of Bacteria and Archaea are typically much larger than those of viruses (Figure 11.1), and genomes of eukaryotes are the largest of all. Viroids, naked infectious RNAs that cause certain plant diseases (Section 11.12), have the smallest genomes of all microbes (Figure 11.1).
Whether a viral genome is large or small, once a virus has infected its host, transcription of viral genes must occur and new copies of the viral genome must be made. Only later, once viral proteins begin to appear from the translation of viral mRNA, can viral assembly begin. For certain RNA viruses, the genome is also the mRNA. For most viruses, however, viral mRNA must first be made by transcription of the DNA or RNA genome, and we now consider the variations on how this occurs.
The Baltimore Scheme: DNA Viruses
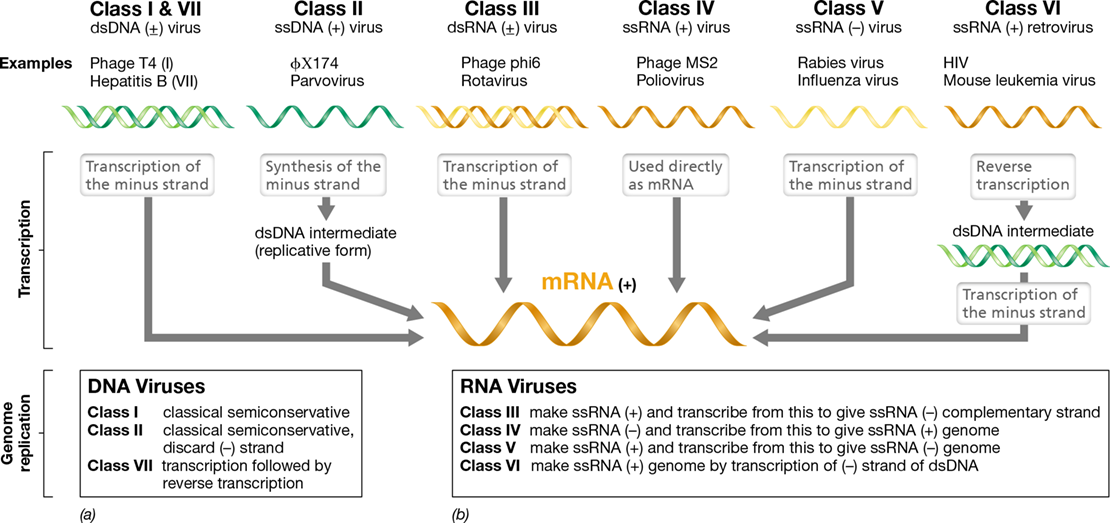
The American virologist David Baltimore, who shared with the American Howard Temin and the Italian American Renato Dulbecco the Nobel Prize for Physiology or Medicine in 1975 for the discovery of retroviruses and their key enzyme, reverse transcriptase, developed a classification scheme for viruses. The scheme is based on the relationship of the viral genome to its mRNA and recognizes seven classes of viruses; three classes have DNA genomes and four have RNA genomes (Figure 11.2). By convention in virology, viral mRNA is always considered to be of the plus (+) configuration. Thus, to understand the molecular biology of a particular class of virus, one must know the nature of the viral genome and the steps necessary to produce (+) mRNA from it (Figure 11.2).
Figure 11.2 The Baltimore classification of viral genomes.

Seven classes of viral genomes are known. The genomes can be either (a) DNA or (b) RNA, and either single-stranded (ss) or double-stranded (ds). With the exception of classes V and VI viruses, where the only known examples infect eukaryotic hosts, the top example listed is a bacterial virus and the bottom example an animal virus. The path each viral genome takes to form its mRNA and the strategy each uses for replication is shown.
Double-stranded DNA viruses are in Baltimore class I. The mechanism of mRNA production and genome replication of class I viruses is the same as that used by cells (Chapter 6). A virus containing a single-stranded genome may be either a positive-strand virus (also called a plus-strand virus) or a negative-strand virus (also called a minus-strand virus). Class II viruses contain single-stranded plus-strand DNA genomes. Transcription of such a genome would yield (−) mRNA, and thus, before (+) mRNA can be produced from class II viruses, a complementary DNA strand must first be made to form a double-stranded DNA intermediate; this is called the replicative form. The latter is used to produce (+) mRNA, and as the source of new genome copies, the plus strand becomes the genome while the minus strand is discarded (Figure 11.2). With only one known exception, all single-stranded DNA viruses are positive-strand viruses.
The Baltimore Scheme: RNA Viruses
The production of mRNA and genome replication will obviously be different for RNA viruses than for DNA viruses. Cellular RNA polymerases do not catalyze the formation of RNA from an RNA template, but instead require a DNA template. Therefore, depending on the virus, RNA viruses must either carry in their virions or encode in their genomes an RNA-dependent RNA polymerase called RNA replicase. RNA replicases replicate the viral RNA genome and produce viral-specific mRNA. With positive-strand RNA viruses (class IV), the genome is also mRNA. But for negative-strand RNA viruses (class V), RNA replicase must synthesize a plus strand of RNA from the negative-strand template, and the plus strand is then used as mRNA. The plus strand is also used as a template to make more negative-strand genomes (Figure 11.2). RNA viruses of class III face a similar problem but start with double-stranded (+/−) RNA instead of only a positive or negative strand.
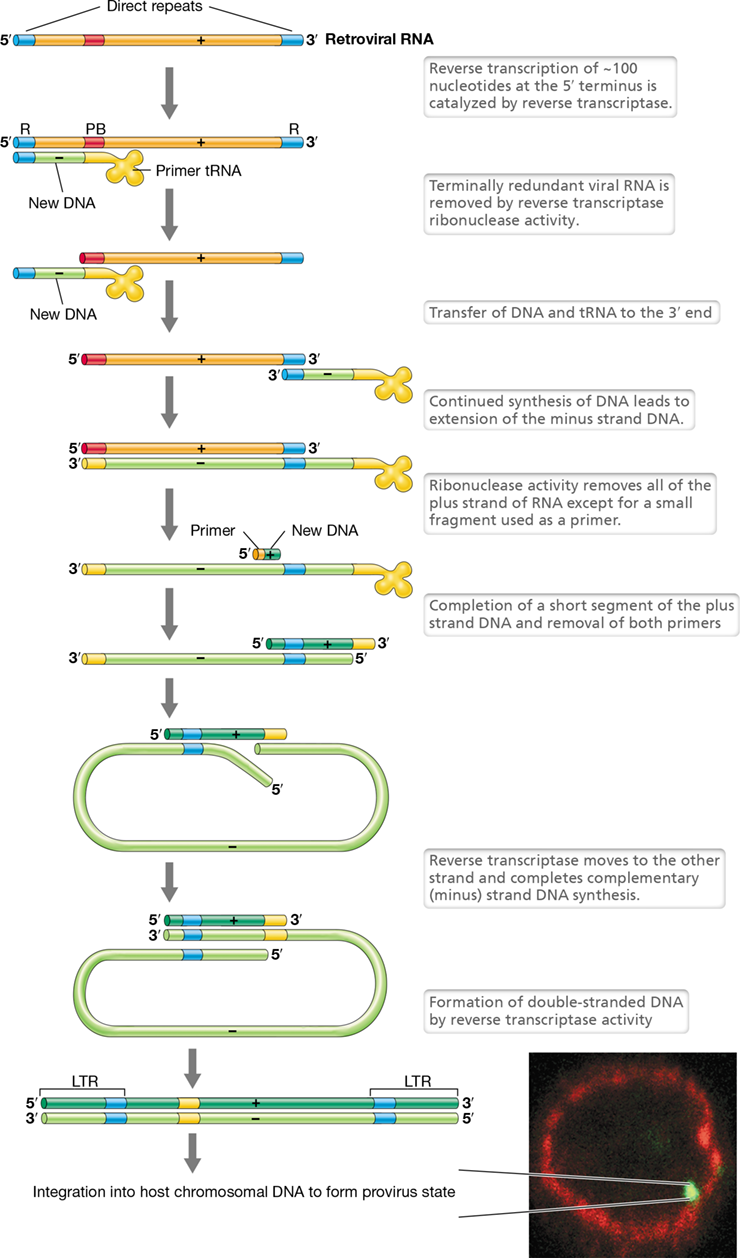
Retroviruses are animal viruses whose genomes consist of single-stranded RNA of the plus configuration but replicate through a double-stranded DNA intermediate (class VI). The process of copying the information found in RNA into DNA is called reverse transcription and is catalyzed by an enzyme called reverse transcriptase. Human immunodeficiency virus (HIV, the causative agent of AIDS) is a retrovirus. Finally, class VII viruses are those highly unusual viruses (human hepatitis B virus is an example) whose genomes consist of double-stranded DNA but which replicate through an RNA intermediate. As we will see later, these viruses also use reverse transcriptase (Section 11.11).
Hosts for Viruses of Each Baltimore Class
Only certain of the Baltimore classes of viruses are known to infect cells of a particular phylogenetic domain. For example, only two classes are known in Archaea and only four in Bacteria; only in animals do we find examples of all seven Baltimore classes of viruses (**Figure 11.3*a***). Examples of viruses from each of the Baltimore classes are drawn to scale in Figure 11.3b.
Figure 11.3 Viral hosts and viral diversity.

(a) Virus host preferences by cellular domain (left graph) and in different major groups of eukaryotes (right graph). RT, reverse transcription. Data adapted from Nasir, A., and G. Caetano-Anollés. 2015. Sci. Adv. 1: e1500527. (b) Drawings to scale of several viruses discussed in this chapter. The symbols + and − next to viral names indicate the complementarity of the single-stranded DNA or RNA genome of that virus. By convention, all viral mRNAs are designated as + in sequence complementarity.
Double-stranded DNA viruses (class I) are the primary viruses infecting prokaryotic cells, while single-stranded plus-sense RNA viruses (class IV) are the major viral predators of eukaryotic cells (Figure 11.3a). As far as is known, fungi are only infected by RNA viruses of classes III and IV, whereas the vast majority of class I viruses that infect eukaryotes replicate in animal hosts rather than plants. By contrast, plants serve as hosts to many more class II viruses than do animals, whereas virtually all class V viruses (viruses with a single-strand minus-sense genome) infect animals rather than plants. And finally, retroviruses (class VI) are known only from animal hosts, while class VII viruses, which like retroviruses depend on reverse transcriptase to replicate their genome, are much more common in plants than in animals (Figure 11.3a). Although the reasons that different genomic classes of virus show specific host preferences is unclear, the fact that Bacteria and Archaea are hosts for only a relatively small group of viruses suggests that some viral classes may have evolved later in the course of evolution when more complex eukaryotic hosts were available for infection. However, computational analyses (to be discussed in Chapter 13) suggest that this hypothesis may be incorrect, in that most viral groups, especially the RNA viruses, appear to be of ancient origin.
Viral Protein Synthesis
During the course of viral replication, once (+) mRNA is made (Figure 11.2), viral proteins can be synthesized. In all viruses, these proteins can be grouped into two broad categories: (1) proteins synthesized soon after infection, called early proteins, and (2) proteins synthesized later in the infection, called late proteins. Both the timing and amount of viral protein synthesis is highly regulated. Early proteins are typically enzymes and are therefore synthesized in relatively small amounts. These include not only nucleic acid polymerases but also proteins that function to shut down host cell transcription and translation. By contrast, late proteins are typically structural components of the virion and other proteins that are not needed until virion assembly begins, and these are made in much larger amounts (Section 5.5).
Virus infection upsets the regulatory mechanisms of the host because there is a marked overproduction of viral nucleic acid and protein in the infected cell. Eventually, when the proper proportions of viral genome copies and virion structural components have been synthesized, new virions are assembled—typically spontaneously—and exit the host cell by either lysing and killing it or by budding or secretion processes in which the host cell may remain alive.
Check Your Understanding
Distinguish between a positive-strand RNA virus and a negative-strand RNA virus.
Contrast mRNA production in the two classes of single-stranded RNA viruses.
What is unusual about genetic information flow in retroviruses?
11.2 Viral Taxonomy and Phylogeny
Because of the remarkable diversity and abundance of viruses, the Baltimore scheme and host specificity are the easiest way to classify viruses. However, these classifications do not consider relationships between viruses. For example, both rabies and influenza viruses are negative-strand RNA viruses (Baltimore class V), but they possess very different morphology, infection strategies, and genomes. While we will discuss the evolutionary timeline for viral appearance on Earth and their relationship to cells in Chapter 13, here we focus on taxonomic and phylogenetic relationships between viruses.
Viral Taxonomy
Like microorganisms, viruses can be characterized using a polyphasic approach that includes phenotypic, genotypic, and phylogenetic analyses, or taxonomy (Section 13.12). Within viral taxonomy there are orders, families, subfamilies, genera, and species. The International Committee on Taxonomy of Viruses or ICTV is composed of virologists that oversee the classification and naming of viruses (http://ictv.global). ICTV uses a combination of host range, structural morphology, replication cycle, nucleic acid sequence, and pathogenicity to classify viruses. ICTV puts the Baltimore class V rabies and influenza viruses mentioned above in the orders Mononegavirales and Articulavirales, respectively. Another example of the utility of viral taxonomy is the classification of archaeal viruses. While only two Baltimore classes of viruses have been described to infect Archaea (classes I and II; Figure 11.3a), genomic and morphological diversity of these viruses (Section 11.5) places them into 17 different families within unclassified orders.
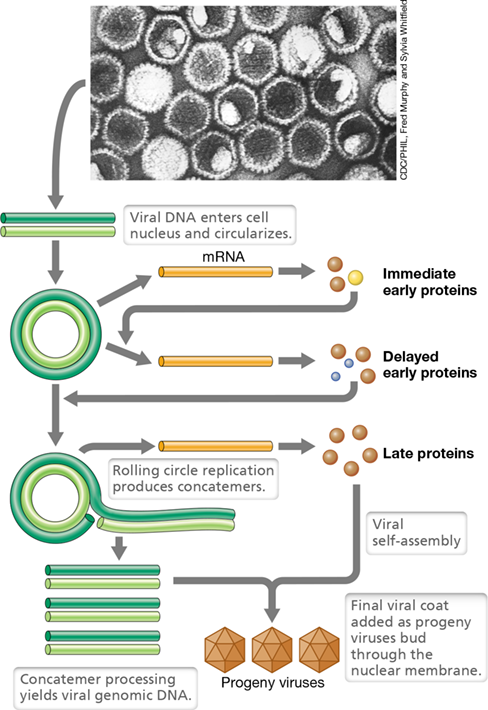
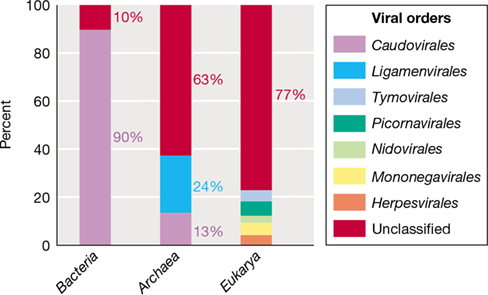
Figure 11.4 illustrates the distribution of seven major orders of cultured viruses among the three domains of life. While the number of bacteriophages isolated outnumber those of archaeal viruses by a factor of ∼20 (Figure 11.3a), 90% of bacteriophages belong to the Caudovirales order. Members of the Caudovirales are the tailed bacteriophages with double-stranded DNA genomes (Figure 11.3b and Section 11.4). In contrast, only 13% of cultured archaeal viruses belong to this order with over 60% of archaeal viruses belonging to unclassified orders (Figure 11.4). The high number of unclassified archaeal viruses stems from the diverse morphology displayed by those in culture (Section 11.5). Eukaryotic viral classification shares the same diversity issue, with most viruses comprising unclassified orders (Figure 11.4).
Figure 11.4 Taxonomic classification of viruses infecting the three domains of life.

The International Committee on Taxonomy of Viruses assignment of viruses into orders based on host. Data adapted from Mahmoudabadi, G., and Phillips, R. 2018. eLife 7: e31955.
Viral Phylogeny
Evolutionary relationships (phylogeny, Chapter 13) among viruses can be difficult to map because they do not possess the ribosomal RNA molecules used to make phylogenetic trees of cells (Section 1.15). Viruses also possess high rates of mutation, especially those with RNA genomes, compared to that of cells. To determine phylogenetic relationships between viruses, the sequences of conserved proteins have been compared along with some structural features. However, in only a few groups of viruses has it been possible to reliably trace phylogenies, and in these cases, trees have been assembled from sequences of a group of genes or proteins shared in common among the group. One such example is Mimivirus and its relatives, one of the larger known viruses (Figure 11.5; Chapter 5). Mimivirus capsids are multilayered and icosahedral. The virion is surrounded by spikes and is nearly 0.75 μm in diameter (Figure 11.5a), larger than some prokaryotic cells. Mimivirus contains a 1.2-megabase double-stranded DNA genome. Mimivirus infects the protozoan Acanthamoeba and belongs to a group of giant viruses with large genomes called nucleocytoplasmic large DNA viruses (NCLDV) (Figure 11.5b). The NCLDV comprise several virus families, including pox viruses (Section 11.6), iridoviruses, and certain plant viruses. These viruses share a set of highly homologous proteins, most of which function in DNA metabolism. A phylogenetic tree of these viruses constructed from DNA sequences encoding these proteins shows how they have diverged from a common ancestor (Figure 11.5b). It is thus possible to track the phylogeny of particular viral groups with some confidence, but to do so, one needs to start with a group that is already known to share a number of properties in common.
Figure 11.5 Phylogeny of nucleocytoplasmic large DNA viruses (NCLDV).

(a) Transmission electron micrograph of Mimivirus, a member of the NCLDV group. A virion is about 0.75 μm in diameter. (b) Phylogeny of major groups of NCLDV based on comparative sequences of several proteins of DNA metabolism. See Chapter 5 for additional coverage of large viruses.
It is clear that diversity in the viral world is enormous and that obtaining a detailed phylogenetic tree of all viruses will remain a challenge. The continual isolation of highly unusual new viruses makes this difficult task even more challenging. For example, only about 7% of the genes of Pandoravirus (Figure 11.1) have gene homologs in existing genomic databases. What this means is that over 90% of the genome of this giant virus will likely be new to biology—a striking example of what awaits discovery in the fascinating world of viruses.
Check Your Understanding
What type of viruses make up the Caudovirales order?
Lacking ribosomes, how can viruses be placed on the universal tree of life?
II DNA Viruses
Prokaryotic cells are infected primarily by double-stranded DNA viruses, whereas eukaryotic cells host viruses of all genomic variants. Some DNA viruses are tiny while others are huge, and some DNA animal viruses have novel replication steps or cause tumors in their hosts.
DNA viruses infect a wide variety of organisms, in particular, species of Bacteria and Archaea. In fact, the majority of viruses that infect prokaryotic cells are DNA viruses, mainly of the double-stranded variety (Figure 11.3). We examine several of these here along with some DNA viruses that infect eukaryotes, and we keep our focus on the genetic information flow that occurs during replication of DNA viruses of different genomic makeups.
11.3 Single-Stranded DNA Bacteriophages: ϕX174 and M13
11.3 Single-Stranded DNA Bacteriophages: ϕX174 and M13
11.3 Single-Stranded DNA Bacteriophages: ϕX174 and M13
In this section we discuss two single-stranded DNA bacteriophages, ϕX174 and M13. Many single-stranded DNA plant and animal viruses are also known, and because these share with bacterial viruses the fact that their genomes are of the plus complementarity (Baltimore class II, Figure 11.2a), many molecular events are similar. Hence, our focus here will be on the phages.
Bacteriophage ϕX174
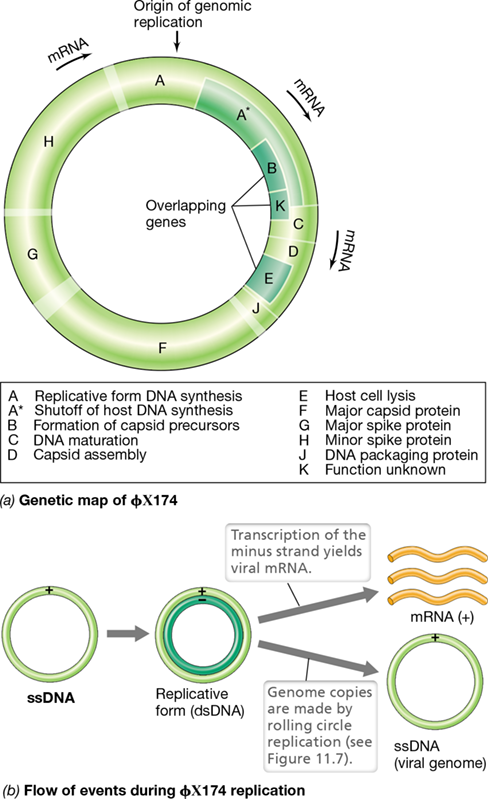
Bacteriophage ϕX174 contains a circular genome of 5386 nucleotides inside a tiny icosahedral virion, about 25 nm in diameter. Phage ϕX174 has only a few genes and shows the phenomenon of overlapping genes, a condition in which there is insufficient DNA to encode all viral-specific proteins; to overcome this problem, parts of the genome are transcribed in more than one reading frame. For example, in the ϕX174 genome, gene B resides within gene A, and gene K resides within both genes A and C (**Figure 11.6*a***). Genes D and E also overlap, gene E being contained completely within gene D. Also, the termination codon of gene D overlaps the initiation codon of gene J.
Figure 11.6 Bacteriophage ϕX174, a single-stranded DNA phage.


(a) Genetic map. Note regions of gene overlap. Protein Ais formed using only part of the coding sequence of gene A by reinitiation of translation. The key indicates the functions of the proteins encoded by each gene. Unlabeled parts of the chromosome are regions of noncoding DNA.(b)* Genetic information flow in ϕX174. Progeny single-stranded DNA is produced from the replicative form by rolling circle replication (see Figure 11.7).
Mastering Microbiology
Art Activity: Figure 11.7 Rolling circle replication in phage ϕX174
The distinct gene products from overlapping genes are made by reinitiating transcription in a different reading frame within a gene to yield a second (and distinct) transcript. In addition to overlapping genes, a small protein in ϕX174 called A, which functions to shut down host DNA synthesis, is synthesized by the reinitiation oftranslation(not transcription) within the mRNA for gene A. The A protein is read from the same mRNA reading frame as the A protein but has a different in-frame start codon and is thus a shorter protein.
As we saw earlier (Section 11.1), before a single-stranded DNA genome can be transcribed, a complementary strand of DNA must be synthesized, forming the double-stranded replicative form. This can then be used as a source of both (+) mRNA and genome copies. Upon infection of an Escherichia coli cell by ϕX174, the viral DNA is separated from the protein coat and the genome is converted into the replicative form by host enzymes. From this, several copies are made by semiconservative replication, and phage-specific transcripts are made by transcription of the negative strand of the replicative form (Figure 11.6b). The replicative form is also the starting point for making copies of the phage genome by a mechanism called rolling circle replication (Figure 11.7). This same mechanism is used by phage lambda, a widely used phage in molecular biology and biotechnology (Section 11.4).
Figure 11.7 Rolling circle replication in phage ϕX174.

Replication begins at the origin of the double-stranded replicative form with the cutting of the plus strand of DNA by gene A protein (both strands of DNA are shown in light green here for simplification). After one new progeny strand has been synthesized (one revolution of the circle), the gene A protein cleaves the new strand and ligates its two ends.
In the synthesis of the ϕX174 genome, the rolling circle facilitates the continuous production of positive strands from the replicative form. To do this, the positive strand of the replicative form is nicked and the 3′ end of the exposed DNA is used to prime synthesis of a new strand (Figure 11.7). Cutting of the plus strand is accomplished by the A protein (Figure 11.6a). Continued rotation of the circle leads to the synthesis of a linear ϕX174 genome. Note that rolling circle synthesis differs from semiconservative replication (Section 6.3) because only the negative strand serves as a template.
When the growing viral strand reaches unit length (5386 residues for ϕX174), the A protein cleaves it and then ligates the two ends of the newly synthesized single strand to give a single-stranded DNA circle. Ultimately, assembly of mature ϕX174 virions occurs and cell lysis follows. The E protein (Figure 11.6a) promotes cell lysis by inhibiting the activity of an enzyme in peptidoglycan synthesis (Section 8.5) in the host cell. Because of the resulting weakness in newly synthesized cell wall material, the host cell ruptures, releasing the phage virions.
Bacteriophage M13
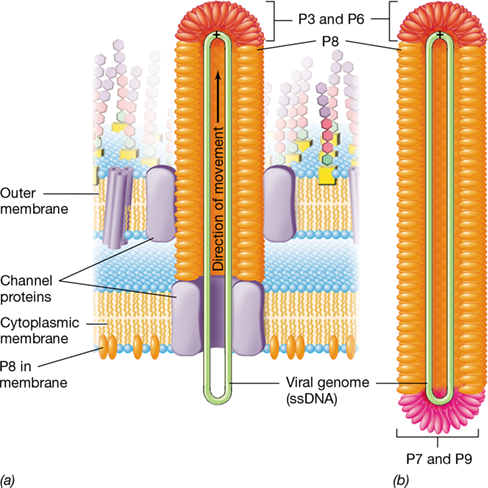
Bacteriophage M13 is a filamentous virus with helical symmetry; the virion is long and thin and attaches to the pilus of its host cell (Figure 5.14). Filamentous phages such as M13 have the unusual property of being released from the host cell without the cell undergoing lysis; infected cells continue to grow, and typical viral plaques (Section 5.3 and Figure 5.10) are not observed. To facilitate the nonlytic release, M13 DNA is covered with coat proteins as it exits across the cell envelope. Four minor coat proteins cover the tips of the virion while the major coat protein covers the sides (Figure 11.8). Thus, mature M13 virions do not accumulate in the cell as occurs with typical lytic bacteriophages. Instead, these filamentous bacteriophages cause chronic infections.
Figure 11.8 Release of phage M13.

The virions of phage M13 exit infected cells without lysis. (a) Budding. The virus DNA crosses the cell envelope through a channel constructed from virus-encoded proteins. As this occurs, the DNA is coated with phage proteins that have been embedded in the cytoplasmic membrane. (b) Complete virion. The two ends of the virion are covered with small numbers of the minor coat proteins P3 and P6 (front end) or P7 and P9 (rear end). P8, major coat proteins. Because bacteriophage M13 is a single-stranded DNA phage, it was widely used in the past as a tool for molecular cloning and DNA sequencing (Chapters 10 and 12).
Several features of phage M13 have made it useful as a cloning and DNA sequencing vehicle in the past. For example, many aspects of DNA replication in M13 are similar to those of ϕX174 and the genome is very small; this facilitates sequencing efforts. Second, a double-stranded form of genomic DNA essential for cloning purposes is produced naturally when M13 produces its replicative form. Third, as long as infected cells are kept growing, phage can be produced indefinitely, yielding a continuous source of the cloned DNA. These and other features of M13 made this phage a workhorse of the genetic engineering field for many years, although today M13 has been replaced for most genetic engineering tasks by a variety of even more convenient and useful tools, as we will see in Chapter 12.
We transition now from single-stranded DNA bacteriophages to double-stranded DNA bacteriophages, where, because of the nature of the genome, replication events resemble those in cells but have some unique features as well.
Check Your Understanding
Why is formation of the replicative form of ϕX174 necessary in order to make phage-specific mRNA?
In the ϕX174 genome, describe the difference between how the gene B and gene A* proteins are made.
How can M13 virions be released without killing the infected host cell?
11.4 Double-Stranded DNA Bacteriophages: T4, T7, and Lambda
11.4 Double-Stranded DNA Bacteriophages: T4, T7, and Lambda
11.4 Double-Stranded DNA Bacteriophages: T4, T7, and Lambda
The double-stranded DNA (dsDNA) (Baltimore class I, Figure 11.2a) bacteriophages are among the best studied of all viruses, and we have already discussed two important ones, T4 and lambda, in Chapter 5. Because of their importance in molecular biology, gene regulation, and genomics, we consider details regarding their replication and also that of bacteriophage T7.
Bacteriophage T4
Bacteriophage T4 is a large, tailed, double-stranded DNA virus of Escherichia coli that possesses complex morphology (90 nm×200 nm; Figure 5.7). As a virulent bacteriophage, T4 always kills its host cell following infection (Section 5.5). The T4 genome differs from many other bacteriophage genomes in that its DNA contains the modified cytosine base 5-hydroxymethylcytosine in place of cytosine (**Figure 11.9*a***); this modification is catalyzed by a phage-encoded enzyme and further glycosylated (a glucose molecule is added) to produce a viral nucleotide that cannot be degraded by the cell’s restriction modification systems (Sections 5.5 and 9.12).
Figure 11.9 Circular permutation and the unique DNA of bacteriophage T4.

(a) The unique base 5-hydroxymethylcytosine in the DNA of bacteriophage T4. Once this base is glucosylated, the T4 DNA is resistant to restriction enzyme attack. (b) Generation of virus-length T4 DNA molecules with permuted sequences by an endonuclease that cuts off constant lengths of DNA from a concatemer regardless of their sequence.
Because of its large, 170-kilobase-pair linear genome, bacteriophage T4 encodes its own DNA polymerase. Other proteins that function in viral DNA replication such as primases and helicases (Section 6.3) are also encoded by the T4 genome. In fact, T4 produces its own eight-protein DNA replisome complex (Section 6.4) to facilitate phage-specific genome synthesis. However, the T4 genome does not encode its own RNA polymerase to express these genes. Instead T4-specific proteins modify the specificity of the host RNA polymerase so that it recognizes only phage promoters; these modification proteins are encoded by T4 genes as part of the “early proteins” in the replication cycle (Figure 5.16).
To facilitate expression of only T4-encoded genes, a phage-encoded anti-sigma factor (Section 7.16) is produced that binds to the host RNA polymerase sigma factor and prevents it from recognizing promoters on host genes. This effectively switches the activity of host RNA polymerase from transcribing host-specific genes to transcribing T4-specific genes. Later in the infection process, other phage proteins modify the host RNA polymerase so that it now recognizes T4 promoters of genes encoding middle proteins. Finally, transcription of T4 late genes begins, and this requires a new T4-encoded sigma factor that directs host RNA polymerase to promoters for these genes only. At this point, viral assembly can begin.
Besides encoding an unusual nucleotide and its own replication machinery, the T4 genome has another unusual feature: In a population of T4 virions, although each copy of the genome contains the same set of genes, they are arranged in a different order. This is a phenomenon called circular permutation, which is a feature of many virus genomes. The term circular permutation is derived from the fact that DNA molecules that are circularly permuted appear to have been linearized by opening identical circular genomes at different locations. Circularly permuted genomes are also terminally redundant, meaning that some DNA sequences are duplicated on both ends of the DNA molecule as a result of the mechanism that generated them.
The T4 genome is first replicated as a unit and then several genomic units are recombined end to end to form a long DNA molecule called a concatemer (Figure 11.9b). When the T4 DNA is packaged into capsids, the concatemer is not cut at a specific sequence; instead linear segments of DNA just long enough to fill a phage head are generated. This is called headful packaging and is common among bacteriophages. However, because the T4 head holds slightly more than a genome length, the headful mechanism generates terminal repeats of about 3–6 kbp at each end of the DNA molecule (Figure 11.9b).
Bacteriophage T7
Bacteriophage T7 is a relatively small virulent DNA virus that infects Escherichia coli and a few related enteric bacteria. The virion has an icosahedral head and a very short tail (Figure 11.3b), and the T7 genome is a linear double-stranded DNA molecule of about 40 kilobase pairs.
When a T7 virion attaches to a host cell and the DNA is injected, early genes are quickly transcribed by host RNA polymerase and then translated. One of these early proteins inhibits the host restriction system, a mechanism for protecting the cell from foreign DNA (Sections 5.5 and 9.12). This occurs very rapidly, as the T7 anti-restriction protein is made and becomes active before the entire T7 genome has entered the cell. Other early proteins include a T7 RNA polymerase and proteins that inhibit host RNA polymerase activity. T7 RNA polymerase recognizes only T7 gene promoters distributed along the T7 genome. This transcriptional strategy differs from that of phage T4 because, as we saw, T4 uses the host RNA polymerase throughout its replication cycle but modifies the host polymerase to recognize only phage genes.
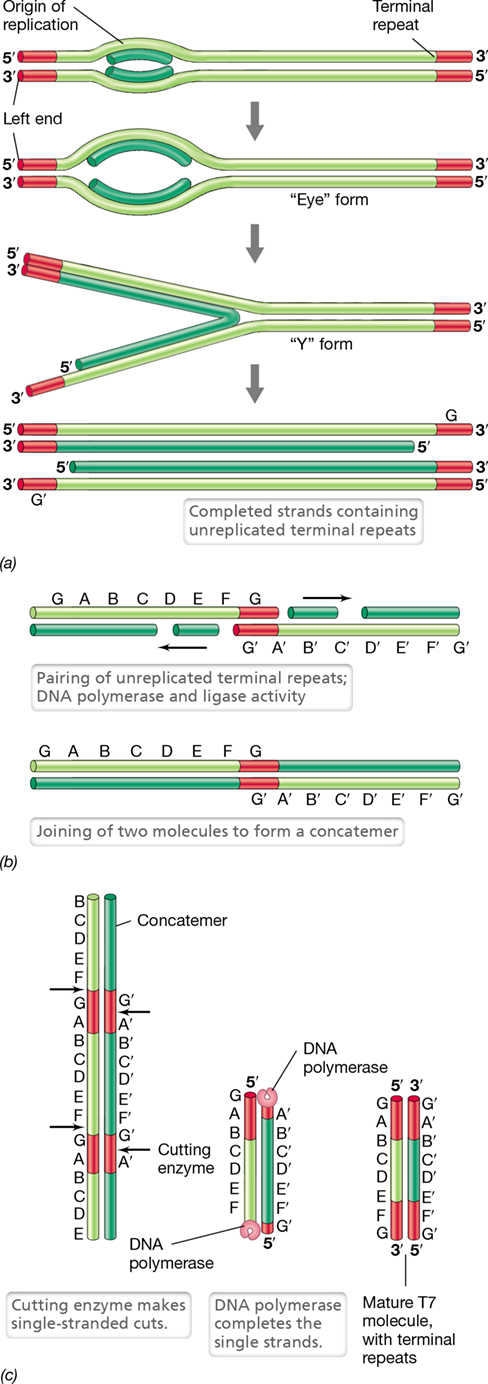
Genome replication in T7 begins at an origin of replication within the molecule and proceeds bidirectionally from this point (**Figure 11.10*a***). Phage T7 uses its own DNA polymerase, which is a composite protein including one polypeptide encoded by the phage and one by the host. As in phage T4, T7 DNA contains terminal repeats at both ends of the molecule and these are eventually used to form concatemers (Figure 11.10b). Continued replication and recombination leads to concatemers of considerable length, but ultimately a phage-encoded endonuclease cuts each concatemer at a specific site, resulting in the formation of linear DNA molecules with terminal repeats that are packaged into phage heads (Figure 11.10c). However, because T7 endonuclease cuts the concatemer at specific sequences, the DNA sequence in each T7 virion is identical. This differs from the situation in phage T4 discussed above, where DNA concatemers are processed using a “headful mechanism” that generates circularly permuted genomes.
Figure 11.10 Replication of the bacteriophage T7 genome.

(a) The linear, double-stranded DNA undergoes bidirectional replication, giving rise to intermediate “eye” and “Y” forms (for simplicity, both template strands are shown in light green and both newly synthesized strands in dark green). (b) Formation of concatemers by joining DNA molecules at their unreplicated terminal ends. (c) Production of mature viral DNA molecules from T7 concatemers by activity of the cutting enzyme, an endonuclease.
Mastering Microbiology
Art Activity: Figure 11.10 Replication of the bacteriophage T7 genome
Bacteriophage Lambda
Bacteriophage lambda, which also infects Escherichia coli, is a double-stranded DNA virus with a head and tail (**Figure 11.11*a***). Lambda is a temperate bacteriophage and thus has two possible pathways—lytic or lysogenic. During the lytic cycle, lambda kills its host cells in the same manner as T4 and T7. However, in lysogeny lambda differs from T4 and T7 by maintaining a stable relationship with its host. Inside its capsid, the 48.5-kilobase lambda genome is linear and possesses a single-stranded region 12 nucleotides long at the 5′ end of each DNA strand. These single-stranded “cohesive” ends are complementary in base sequence; when lambda DNA enters the host cell, they base-pair to form the cos site and cyclize (circularize) the genome (Figure 11.11b).
Figure 11.11 Bacteriophage lambda: virions, integration of viral DNA and rolling circle replication.

(a) Transmission electron micrograph of phage lambda virions. The head of each virion is about 65 nm in diameter and contains linear dsDNA. (b) Lambda DNA integrates at specific attachment (att) sites on both the host and phage genomes. Host genes near att include gal, galactose utilization; bio, biotin synthesis; and moa, molybdenum cofactor synthesis. Lambda integrase is required, and specific pairing of the complementary ends results in integration of lambda DNA. (c) During rolling circle replication, as one strand (dark green) rolls out, it is both replicated at its opposite end and serves as a template for synthesis of the complementary strand.
If lambda enters the lytic pathway, long, linear concatemers of genomic DNA are synthesized by the rolling circle replication mechanism described for ϕX174 (Section 11.3). The double-stranded concatemer is then cut into genome-sized lengths at the cos sites and the resulting genomes packaged into lambda phage heads (Figure 11.11c). Once the tail has been added and mature lambda virions have been assembled (Figure 11.11a), cell lysis occurs and the virions are released. In its role as a lytic phage, lambda can also package a few chromosomal genes from its lysed host in newly synthesized virions and then transfer these to a second host cell, a process called transduction. Transduction is an important means of horizontal gene transfer in nature and is also an important tool in bacterial genetics (Section 9.7).
Instead of the lytic pathway, if lambda takes the lysogenic route, its genome integrates into the E. coli chromosome. This requires a protein called lambda integrase, a phage-encoded enzyme that recognizes the phage and bacterial genome attachment sites (att in Figure 11.11b) and facilitates integration of the lambda genome. From this relatively stable state, certain events such as host DNA damage can initiate the lytic cycle once again. After such a trigger, a lambda excision protein excises the lambda genome from the host chromosome, transcription of lambda DNA begins, and lytic events unfold. With these two contrasting events—lysis and lysogeny—in mind, what controls the path that phage lambda will take?
Lysis or Lysogeny: Regulation of the Lambda Lifestyle
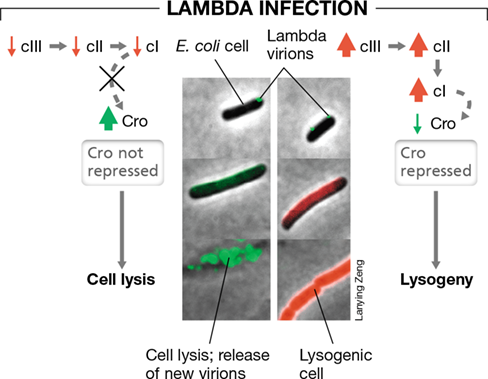
Whether lysis or lysogeny occurs in a lambda infection depends in large part on the levels of two key repressor proteins that can accumulate in the cell following infection: the lambda repressor, also called the cI protein, and a second repressor called Cro. In a nutshell, the first repressor that accumulates will control the outcome of the infection.
If genes encoding the cI protein are rapidly transcribed following infection and cI accumulates, it represses the transcription of all other lambda-encoded genes, including Cro. When this happens, the lambda genome integrates into the host’s genome and becomes a prophage (Figure 11.12). The host continues to grow, yielding more lysogens, until an event that triggers lysis is encountered. Cro, on the other hand, represses expression of a protein called cII whose function is to activate the synthesis of cI. Hence, following infection, if cI is present at insufficient levels to repress expression of phage-specific genes, Cro accumulates in the cell; if this happens, lambda travels the lytic pathway.
Figure 11.12 Regulation of lytic and lysogenic events in phage lambda.

The photomicrographs show time courses of cells of Escherichia coli following a course of lytic (left panel, green) or lysogenic (right panel, red) events, as controlled by various repressors. The colors originate from genetically engineered lambda phage that trigger the production of specific fluorescent proteins when either lytic genes (green) or lysogenic genes (red) are expressed. Lytic cells are killed whereas E. coli lysogens continue to grow and divide.
Control of these alternative lifestyles—lysis or lysogeny—of lambda has been likened to a “genetic switch,” where a defined series of events must occur for one pathway to be favored over the other. Although infection of an E. coli cell by a lambda virion typically results in the lytic cycle, as we have said, lytic events can be switched off if sufficient concentrations of cII are present to ensure adequate levels of cI (Figure 11.12). But how does this come about? Levels of protein cII are controlled by the relative activity of a protease in the cell that slowly degrades cII and by levels of yet another protein, cIII, whose function is to stabilize cII and protect it from protease attack. We thus have a cascade of regulatory events here: cIII controls cII, which in turn controls cI. But even this is not the end of the story. Several other proteins not described here also play a role in the lambda lytic/lysogenic “decision,” and hence the progress of a lambda infection is a highly intricate series of events.
Indeed, this tiny bacteriophage employs some of the most elaborate regulatory systems known in virology. One of the few lambda proteins expressed during lysogeny prevents the growing lysogen from entering dormancy during stressful conditions (Section 8.12). This regulatory mechanism helps ensure the continued spread of the lambda virus in a mixed population of Escherichia coli lysogens and nonlysogens.
Cells of both Bacteria and Archaea are prokaryotic, and one might predict that what is true for bacterial viruses is also true of archaeal viruses. We see next that this is definitely not the case.
Check Your Understanding
How does the mechanism used by bacteriophage T7 to evade the host cell’s restriction system differ from that of T4?
What commits lambda to the lytic versus the lysogenic pathway?
11.5 Viruses of *Archaea*
Many bacteriophages and archaeal viruses have been isolated and characterized thus far. For Bacteria, these include both DNA and RNA phages, some with single-stranded and others with double-stranded genomes. However, all characterized archaeal viruses have DNA genomes, and with rare exception, double-stranded DNA genomes (Figure 11.3a).
DNA Archaeal Viruses
Several DNA viruses have been discovered that infect species in the two main phyla of Archaea, the Euryarchaeota and the Crenarchaeota (Chapter 17). Most viruses that infect species of Euryarchaeota, including both methanogenic and halophilic Archaea, are of the “head and tail” type, resembling the structurally complex phages that infect enteric bacteria, such as phage T4 (Section 5.5; Section 11.4). Besides two special cases discussed later, all other characterized archaeal DNA viruses contain double-stranded and typically circular DNA genomes.
The most morphologically distinctive archaeal viruses infect hyperthermophilic Crenarchaeota. For example, the sulfur chemolithotroph Sulfolobus is host to several structurally unusual viruses. One such virus, called SSV, forms spindle-shaped virions that often cluster in rosettes (**Figure 11.13*a***). Such viruses are widespread in acidic hot springs worldwide. Virions of SSV contain a circular DNA genome of about 15 kilobase pairs. A second morphological type of Sulfolobus virus forms a rigid, helical rod-shaped structure (Figure 11.13b). Viruses in this class, nicknamed SIFV, contain linear DNA genomes about twice the size of that of SSV. Many variations on the spindle- and rod-shaped patterns have been seen in archaeal viral isolation studies, and a few species of Crenarchaeota are also infected by filamentous viruses.
Figure 11.13 Archaeal viruses.

Electron micrographs of viruses of Crenarchaeota (parts a, b, d, e), and a virus of a euryarchaeote (c). (a) Spindle-shaped virus SSV1 that infects Sulfolobus solfataricus (virions are 40×80 nm). (b) Filamentous virus SIFV that infects S. solfataricus (virions are 50×900−500 nm). (c) Spindle-shaped virus PAV1 that infects Pyrococcus abyssi (virions are 80×120 nm). (d) ATV, the virus that infects the hyperthermophile Acidianus convivator. When released from the cell the virions are lemon-shaped (left), but they proceed to grow appendages on both ends (right). ATV virions are about 100 nm in diameter. (e) Filamentous virus ACV of Aerophilum pernix. ACV virions are about 220 nm long.
A spindle-shaped virus that infects the hyperthermophile Acidianus displays a novel behavior. The virion, called ATV, contains a circular genome of about 68 kilobase pairs and is lemon-shaped when first released from the host cells. However, shortly after release from its lysed host cell, the virion produces long, thin tails, one at each end (Figure 11.13d). The tails are actually tubes, and as they form, the virion becomes thinner and its volume is reduced. Remarkably, this is the first example of virus development in the complete absence of host cell contact. It is thought that the extended tails of ATV help the virus in some way survive in its hot (85 °C), acidic (pH 1.5) environment or perhaps assist in attachment to a new host cell. This unusually shaped virus is also lysogenic, a property rarely seen in other archaeal viruses.
A spindle-shaped virus also infects the hyperthermophile Pyrococcus (Euryarchaeota). This virus, named PAV1, resembles SSV but is larger and contains a very short tail (Figure 11.13c). PAV1 has a small circular DNA genome and is released from host cells without cell lysis, probably by a budding mechanism similar to that of the Escherichia coli bacteriophage M13 (Section 11.3). Pyrococcus has a growth temperature optimum of 100 °C and thus PAV1 virions must be extremely heat-stable. Despite their similar morphologies, genomic comparisons of PAV1 and SSV-type viruses show little sequence similarity, indicating that the two types of viruses do not have common evolutionary roots.
Although double-stranded DNA genomes seem to predominate in the Archaea, two archaeal viruses have been described that possess single-stranded DNA genomes. Pleomorphic virus 1 (HRPV-1) infects the extreme halophile Halorubrum and is unusual not only because of its circular single-stranded DNA genome (of the + complementarity a property it shares with bacteriophage ϕX174, Figure 11.6), but also due to its enveloped nonuniform morphology. Thus, the virus is released by secretion versus the host cell lysis mechanism common to virulent bacteriophage. Over half of the 7048-nucleotide HRPV-1 genome encodes viral proteins that have also been found in the double-stranded DNA haloarchaeal virus His2. This highlights the challenges of classifying viruses based on genome type and the Baltimore scheme (Section 11.1) but also indicates the likely relationships that exist between archaeal viruses of quite different genomic structure. A second archaeal virus possessing a single-stranded DNA genome is the Aeropyrum coil-shaped virus (ACV) that infects the hyperthermophile Aeropyrum pernix (Figure 11.13e). A filamentous capsid surrounds this circular single-stranded 24.9-kilobase viral genome, which is double the size of the second largest known single-stranded DNA viral genome. Thus, much diversity obviously exists among archaeal viruses.
Genome Content of DNA Archaeal Viruses
While over 2000 bacteriophage genomes have been sequenced, only a little over 100 archaeal viral genome sequences are available in the databases. This disparity in number is due to the difficulty in culturing many archaeal hosts versus their bacterial counterparts. Metagenomic approaches (Section 10.7) are helping expand this number and identify viruses that are not in culture (see discussion of archaeal RNA viruses in next subsection). On average, archaeal virus genomes are smaller than bacteriophage genomes (65 versus 101 open reading frames). Comparative analysis of viral genomes suggests that the genome replication strategies of many archaeal viruses are more similar to those employed by eukaryotic viruses than those used by bacteriophages. Unlike many bacteriophages such as phage T7 (Section 11.4), archaeal viral genomes do not encode a virus-specific RNA polymerase. Thus, archaeal viruses are dependent on the host cell’s RNA polymerase for gene expression, and transcription factors predicted to commandeer the archaeal RNA polymerase have been annotated in many archaeal viral genomes.
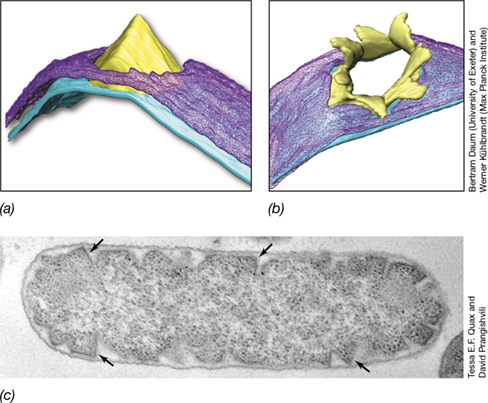
Besides the genes for transcription factors mentioned above and some characterized structural proteins, analyses indicate that up to 90% of the genes within many archaeal viral genomes encode proteins of unknown function unrelated to any other proteins in the databases. However, some have been characterized. For example, a unique method of viral release occurs with two archaeal viruses that infect Sulfolobus islandicus. Not only do the S. islandicus rod-shaped virus 2 (SIRV2) and turreted icosahedral virus (STIV) genomes encode structural proteins for their capsids, they also encode a protein called PVAP (protein forming virus-associated pyramid) that facilitates viral release from the cell. PVAP assembles into seven-fold pyramid structures that rupture the host cell’s S-layer (Section 2.5) and open outwards to create a portal for viruses to exit the host cell (**Figure 11.14a,*b***). Heterologous expression (Section 10.5 and Chapter 12) of PVAP in the bacterium Escherichia coli also resulted in the formation of pyramid structures that protruded into the periplasmic space (Figure 11.14c), thus indicating that a single archaeal viral protein alone is sufficient for producing these structures. The large number of unique archaeal viral proteins that remain to be characterized will undoubtedly lead to even more remarkable discoveries.
Figure 11.14 Virus-associated pyramids (VAPs).

Electron cryotomograph showing a Sulfolobus islandicus cell with SIRV2 VAP (a) closed and (b) open. The VAPs open to allow release of virions from the Sulfolobus host cell. (c) Transmission electron micrograph showing heterologous expression of the protein for archaeal VAP formation (PVAP) in Escherichia coli. The arrows point to pyramid structures formed in the periplasm. The E. coli cell is about 0.7 μm in diameter.
RNA Archaeal Viruses
Thus far, RNA viruses that can replicate in the laboratory on archaeal hosts are unknown, despite the fact that a variety of RNA viruses infect Bacteria and eukaryotes (Figure 11.3). Although no concrete examples of archaeal RNA viruses have emerged, environmental genomics have shown that they almost certainly exist. In some acidic hot springs of Yellowstone National Park (USA) that support large communities of Crenarchaeota, a large number of unusually shaped and structurally tough archaeal viruses have been discovered (Figure 11.15) and grown in the laboratory. Thus far, all of these have been DNA viruses. However, using the powerful tools of metagenomics (Section 10.7), researchers studying these hot springs have discovered viral RNAs whose RNA sequences bear no resemblance to those of any known RNA viruses that infect Bacteria. Because these springs are too hot for eukaryotes and the cell numbers of Bacteria are few, the unusual RNA is almost certainly from RNA archaeal viruses that are yet to be propagated in the laboratory.
Figure 11.15 An acidic Yellowstone hot spring and its archaeal viruses.

This spring is a boiling mudpot with a pH of about 2 and is loaded with hyperthermophilic and acidophilic Crenarchaeota (Section 17.10). Inset: Transmission electron micrograph of a mixture of archaeal viruses from the spring. Compare with Figure 11.13.
Sequence analyses of the hot spring viral RNA show that it originated from single-stranded plus-sense RNA viruses (Baltimore class IV, Figure 11.2b and Section 11.8). These viral genomes also encode an RNA replicase—a hallmark of RNA viruses—and are likely to replicate by way of polyprotein formation, a replication mechanism employed by some class IV viruses of eukaryotes, such as poliovirus (Section 11.8). Replication steps of the putative RNA archaeal viruses, including important molecular details such as the extent to which viral (rather than host) polymerases participate in the replication process, are unclear and await laboratory cultivation of the viruses. However, now that scientists know that such viruses almost certainly exist, they can be on the lookout for them in viral enrichment and isolation studies.
We move on now to consider animal viruses—a group in which there is no shortage of genomic diversity and fascinating replication schemes, many unique to biology.
Check Your Understanding
What type of genome is seen in most archaeal viruses?
Compared with other archaeal viruses, what are two unusual features of the virus that infects Acidianus?
Describe the unique mechanism some Sulfolobus viruses use to exit the host cell.
11.6 Uniquely Replicating DNA Animal Viruses
Unusual replication strategies are seen in two groups of double-stranded DNA (Baltimore class I, Figure 11.2a) animal viruses: pox viruses and adenoviruses. Pox viruses are unique because all replication events, including DNA replication, occur in the host cytoplasm instead of the nucleus, and adenoviruses are unique because the replication of their genome proceeds in a leading fashion on both DNA template strands.
Pox Viruses
Pox viruses have been important historically as well as medically. Smallpox virus was the first virus to be studied in any detail and was the first virus for which a vaccine was developed (over 200 years ago the British physician Edward Jenner was the first to protect people from infection by smallpox virus by exposing them to the similar but much less virulent cowpox virus). Pox viruses are among the largest of all viruses, the brick-shaped vaccinia virions measuring almost 400 nm in diameter (Figure 11.16). Because it closely resembles the smallpox virus but is not pathogenic, vaccinia is used as a smallpox vaccine today and as a safe laboratory model for smallpox virus molecular biology.
Figure 11.16 Smallpox virus.
Transmission electron micrograph of a negatively stained thin section of smallpox virus virions. The virions are approximately 350 nm (0.35 μm) long. The dumbbell-shaped structure inside the virion is the nucleocapsid, which contains the double-stranded DNA genome. All replication functions for pox virus occur in the host cytoplasm.
Mastering Microbiology
Art Activity: Figure 11.17b Adenoviruses and their genomic replication
The vaccinia virus genome consists of linear double-stranded DNA of about 190 kilobase pairs encoding about 250 genes. Following attachment, vaccinia virions are taken up into host cells and the nucleocapsids (Figure 11.16) are liberated in the cytoplasm; all replication events take place in the cytoplasm. Uncoating of the viral genome requires the activity of a viral protein that is synthesized after infection (the gene encoding this protein is transcribed by a viral RNA polymerase contained within the virion). In addition to this uncoating gene, a number of other viral genes are transcribed, including genes that encode a DNA polymerase that synthesizes copies of the viral genome. These are then incorporated into virions that accumulate in the cytoplasm, and the virions are released when the infected cell lyses.
Vaccinia virus has been genetically engineered to contain certain proteins from other viruses for use in recombinant vaccines (Section 12.8). A vaccine is a substance capable of eliciting an immune response in an animal that protects the animal from future infection with the same agent (Chapter 28). Vaccinia virus causes no serious health effects in humans but elicits a strong immune response. Therefore, as a carrier of proteins from pathogenic viruses, vaccinia virus is a relatively safe and effective tool for stimulating an immune response against these pathogens. Success has been obtained with vaccinia virus vaccines against the viruses that cause influenza, rabies, herpes simplex type 1, and hepatitis B. We discuss all of these viruses in upcoming sections.
Adenoviruses
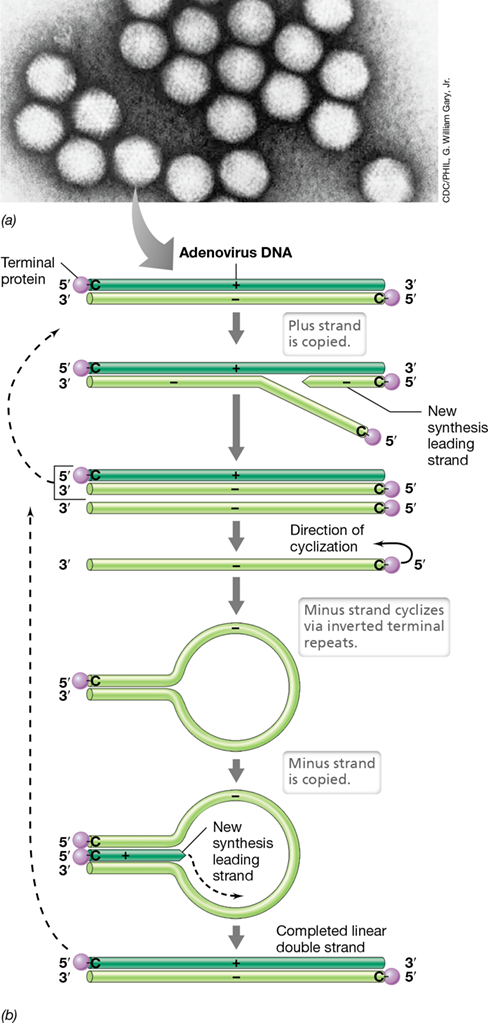
Adenoviruses are a group of small and naked icosahedral viruses (**Figure 11.17*a***) that contain linear double-stranded DNA genomes. Adenoviruses are of minor health importance, causing mild respiratory infections in humans, but they have unique stature in virology because of the mechanism by which they replicate their genomes. Attached to the 5′ end of adenoviral genomic DNA is a protein called the adenoviral terminal protein, and it is essential for replication of the adenoviral genome. The complementary DNA strands also have inverted terminal repeats that play a role in the replication process (Figure 11.17b).
Figure 11.17 Adenoviruses and their genomic replication.

(a) Transmission electron micrograph of adenoviral virions. Note the icosahedral structure. (b) Adenoviral genome replication. Because of loop formation (cyclization), there is no lagging strand; DNA synthesis is leading on both strands. A cytosine (C) is attached to the terminal protein. Adenoviruses are one of several classes of human viruses that cause upper respiratory infections such as the common cold (Section 31.7). Rhinoviruses (single-stranded plus-sense RNA viruses, Section 11.8) cause the vast majority of colds.
Following infection, the adenoviral nucleocapsid is released into the host cell nucleus, and transcription of the early genes proceeds by activity of the host RNA polymerase. Most early transcripts encode important replication proteins such as the terminal protein and a viral DNA polymerase. Replication of the adenoviral genome begins at either end of the DNA genome and the terminal protein facilitates this process because it contains a covalently bound cytosine that functions as a primer for DNA polymerase (Figure 11.17b). The products of this initial replication are a completed double-stranded viral genome and a single-stranded minus-sense DNA molecule. At this point, a unique replication event occurs. The single DNA strand cyclizes by means of its inverted terminal repeats, and a complementary (plus-sense) DNA strand is synthesized beginning from its 5′ end (Figure 11.17b). This mechanism is unique because double-stranded DNA is replicated without the formation of a lagging strand, as occurs in conventional semiconservative DNA replication (Section 6.3). Once sufficient copies of the adenoviral genome have formed and virion structural components accumulate in the host cell, mature adenoviral virions are assembled and released from the cell following lysis.
In addition to infectious diseases, some animal viruses cause cancer, and we consider these now.
Check Your Understanding
What is unusual about genome replication in pox viruses?
What is unusual about genome replication in adenoviruses?
Why is the adenovirus terminal protein essential for replicating its genome?
11.7 DNA Tumor Viruses
Besides catalyzing lytic events or becoming integrated into a genome in a latent state, some DNA animal viruses can induce the formation of tumors. These include viruses of the polyomavirus family and some herpesviruses, both of which contain double-stranded DNA genomes (Baltimore class I, Figure 11.2a).
Polyomavirus SV40
Polyomavirus SV40 is a naked icosahedral virus that can cause tumors in small mammals, such as hamsters and rats. Its circular genome consists of double-stranded DNA (**Figure 11.18*a***). The genome is too small (5.2 kb) to encode its own DNA polymerase, so host DNA polymerases are used and SV40 DNA is replicated in a bidirectional fashion from a single origin of replication. Because of the small genomes of polyomaviruses, the strategy of overlapping genes, typical of many small bacteriophages (Sections 11.3 and 11.8), is also employed here. Transcription of the viral genome occurs in the nucleus, and the mRNAs are exported to the cytoplasm for protein synthesis. Eventually SV40 virion assembly occurs (in the nucleus) and the cell is lysed to release the new virions.
Figure 11.18 Polyomaviruses and tumor induction.

(a) Transmission electron micrograph of relaxed (nonsupercoiled) circular DNA from a tumor virus. The contour length of each circle is about 1.5 μm. (b) Events in cell transformation by a polyomavirus such as SV40. Viral DNA becomes incorporated into the host genome. From there, viral genes encoding cell transformation events are transcribed and transported to the cytoplasm for translation.
When SV40 infects a host cell, one of two outcomes can occur, depending on the host cell. In permissive hosts, virus infection results in the usual formation of new virions and the lysis of the host cell. In nonpermissive hosts, lytic events do not occur; instead, the viral DNA becomes integrated into host DNA, genetically altering the cells in the process (Figure 11.18b). Such cells can show loss of growth inhibition and become malignant, a process called transformation (Figure 5.22). As in certain tumor-causing retroviruses (Section 11.11), expression of specific SV40 genes is required to convert the cell to the transformed state. These tumor-inducing proteins bind to and inactivate host cell proteins that control cell division, and in this way, they promote uncontrolled cell development.
Herpesviruses
Herpesviruses are a large group of double-stranded DNA viruses that cause a variety of human diseases, including fever blisters (cold sores), venereal herpes, chicken pox, shingles, and infectious mononucleosis. An important group of herpesviruses cause cancer. For example, Epstein–Barr virus causes Burkitt’s lymphoma, a tumor endemic in children of central Africa and New Guinea. A widespread herpesvirus is Cytomegalovirus (CMV), present in nearly three-quarters of all adults in the United States over 40 years of age. For healthy individuals, infection with CMV comes with no apparent symptoms or long-term health consequences. However, CMV can cause pneumonia, retinitis (an eye condition), and certain gastrointestinal disorders, as well as serious disease or even death in immune-compromised individuals.
Herpesviruses can remain latent in the body for long periods of time and become active under conditions of stress or when the immune system is compromised. Herpesvirus virions are enveloped and can have many distinct structural layers over the icosahedral nucleocapsid (Figure 11.19). Following viral attachment, the host cytoplasmic membrane fuses with the virus envelope, and this releases the nucleocapsid into the cell. The nucleocapsid is transported to the nucleus, where the viral DNA is uncoated and three classes of mRNA are produced: immediate early, delayed early, and late (Figure 11.19). Immediate early mRNA encodes certain regulatory proteins that stimulate the synthesis of the delayed early proteins. Among the key proteins synthesized during the delayed early stage is a viral-specific DNA polymerase and a DNA-binding protein, both of which are needed for viral DNA replication. As for other viruses, late proteins are primarily viral structural proteins.
Figure 11.19 Herpesvirus.

Flow of events in replication of herpes simplex virus starting from a transmission electron micrograph of herpes simplex virus (diameter of a single virion is about 150 nm). Although the viral genome is linear within the virion, it circularizes once inside the host.
Mastering Microbiology
Herpesvirus DNA replication takes place in the nucleus. After infection, the herpesvirus genome circularizes and replicates by a rolling circle mechanism (Section 11.3). Long concatemers are formed that become processed into virus-length genomic DNA during the assembly process (Figure 11.19). Viral nucleocapsids are assembled in the nucleus, and the viral envelope is added during budding through the nuclear membrane. Mature herpesvirus virions are subsequently released through the endoplasmic reticulum to the outside of the cell. The assembly of herpesvirus virions thus differs from that of other enveloped viruses, which typically receive their envelope from the cytoplasmic membrane during exit from the cell.
Although we have seen a number of unique genomes and replication schemes with DNA viruses, more is in store with RNA viruses, microbes that cause most common human infectious diseases of viral etiology.
Check Your Understanding
What genomic feature does SV40 share with bacteriophage ϕX174?
How can the outcome of an SV40 viral infection differ in permissive versus nonpermissive hosts?
III RNA Viruses
Viruses that carry an RNA genome cause some of the most notorious diseases of humans and display the most intricate of all viral replication schemes. Because most cellular RNA polymerases are unable to make RNA from an RNA template, RNA viruses carry special enzymes in their virions to accomplish this task.
RNA viruses make up Baltimore classes III–VI (Figure 11.2b) and infect both prokaryotic and eukaryotic cells. As in the foregoing sections that dealt with DNA viruses, we organize our coverage of RNA viruses here by genomic characteristics. We also consider Baltimore class VII viruses, which have a partially double-stranded DNA genome, in the final section of this part of the chapter.
11.8 Positive-Strand RNA Viruses
Many viruses contain single-stranded RNA genomes of the plus sense and are therefore positive-strand RNA viruses. In these viruses, the sequence of the genome and the mRNA are the same (both are of the + complementarity, Figure 11.2b). A number of positive-strand animal and bacterial viruses are known, so we restrict our coverage here to just a few well-studied cases. We begin with the tiny bacteriophage MS2.
Bacteriophage MS2
Bacteriophage MS2 is about 25 nm in diameter and has an icosahedral capsid. The virus infects cells of Escherichia coli by attaching to the cell’s pilus (**Figure 11.20*a***), a structure that normally functions in a form of horizontal gene exchange (conjugation) in bacteria (Section 9.8). How MS2 RNA actually gets inside the E. coli cell from the pilus is unknown, but once it has, MS2 replication events begin quickly; the genetic map and major activities of this virus are shown in Figure 11.20b and c.
Figure 11.20 A small RNA bacteriophage, MS2.
(a) Transmission electron micrograph of the pilus of a cell of Escherichia coli showing virions of phage MS2 attached. (b) Genetic map of MS2. Note how the lysis protein gene overlaps with both the coat protein and replicase genes. The numbers refer to the nucleotide positions on the RNA, the entire genome being 3569 nucleotides in length. (c) Flow of events during MS2 replication.
The MS2 genome is just 3.5 kb in size and encodes only four proteins, including the maturation protein, coat protein, lysis protein, and one subunit of RNA replicase, the enzyme that replicates the viral RNA (Section 11.1). Interestingly, MS2 RNA replicase is a composite protein, with three subunits encoded by the host genome and one subunit by the viral genome. The gene encoding the MS2 lysis protein overlaps that encoding the coat protein and replicase subunit (Figure 11.20b). We have seen this phenomenon of overlapping genes before (Sections 11.3 and 11.7) as a strategy for making small genomes encode more proteins.
Because the genome of phage MS2 is plus-sense RNA, it is also the mRNA and is translated directly upon entry into the cell by the host RNA polymerase. When RNA replicase is made, it begins synthesis of minus-sense RNA using plus strands as templates. As minus-sense RNA copies accumulate, more plus-sense RNA is made using the minus-sense strands as templates, and some of these are translated for continued synthesis of viral structural proteins.
Phage MS2 regulates synthesis of its proteins by controlling access of host ribosomes to translational start sites on its RNA. MS2 genomic RNA is folded into a complex secondary structure. Of the four AUG translational start sites (Section 6.9) on the MS2 RNA, the most accessible to the cell’s translation machinery is that for the coat protein and replicase. Hence, translation begins at these sites very early following infection. However, as coat protein molecules accumulate, they bind to the RNA around the AUG start site for the replicase protein, effectively turning off synthesis of replicase. Although the gene for the maturation protein is at the 5′ end of the RNA, the extensive folding of the RNA limits access to the maturation protein translational start site, and consequently, only a few copies are synthesized. In this way, all MS2 proteins are made in the relative amounts needed for virus assembly. Ultimately, spontaneous assembly of MS2 virions begins, and the virions are released from the cell as a result of cell lysis.
Poliovirus
Several positive-strand RNA animal viruses cause disease in humans and other animals. These include poliovirus, the rhinoviruses that cause many cases of the common cold, the coronaviruses that cause respiratory syndromes, including severe acute respiratory syndrome (SARS), and the hepatitis A virus. We focus here on poliovirus and coronaviruses, both of which have linear RNA genomes.
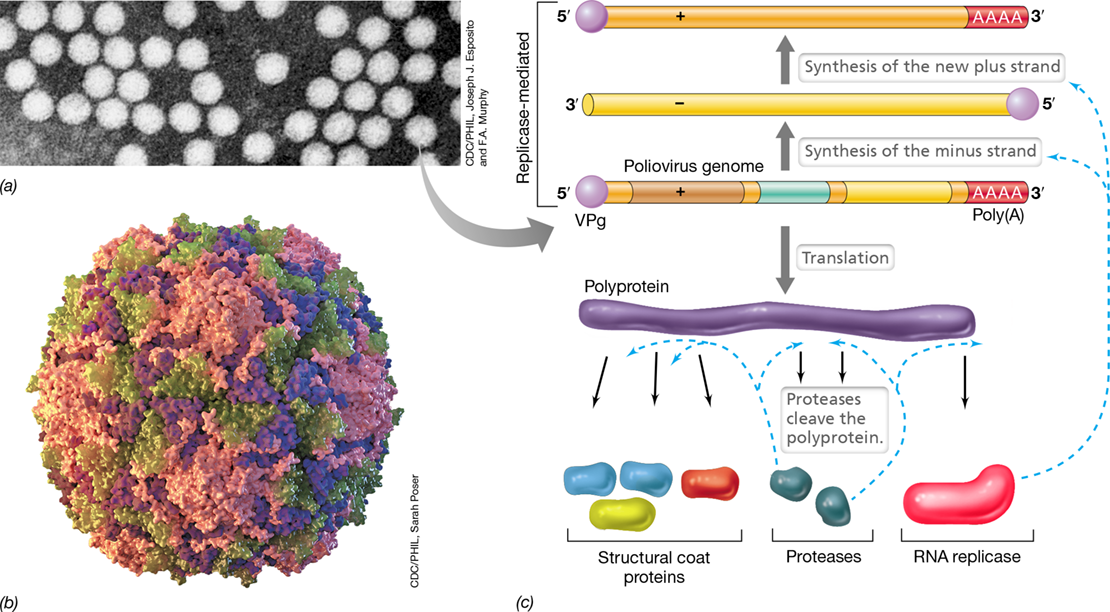
Poliovirus is one of the smallest of all viruses with a 30-nm icosahedral structure containing the minimum 60 morphological units (faces) per virion (**Figure 11.21a,*b***). At the 5′ terminus of the viral RNA is a protein, called the VPg protein, that is attached covalently to the genomic RNA, and at the 3′ terminus is a poly (A) tail (Figure 11.21c), a common feature of eukaryotic cell transcripts (Section 6.6). The poliovirus genome (about 7.4 kb) is also the mRNA, and the VPg protein facilitates binding of the RNA to host ribosomes. Translation yields a polyprotein, a single protein that self-cleaves into several smaller proteins including virion structural proteins. Other proteins generated from the polyprotein include the VPg protein, an RNA replicase responsible for synthesis of both minus-strand and plus-strand RNA, and a virus-encoded protease, which carries out the polyprotein cleavage (Figure 11.21c). This mechanism is called post-translational cleavage and is common in many animal viruses as well as animal cells.
Figure 11.21 Poliovirus.

(a) Transmission electron micrograph of poliovirus virions; a single virion is about 30 nm in diameter. (b) A computer model of a poliovirus virion. The various structural proteins are shown in distinct colors. (c) Genomic replication and formation of poliovirus proteins.
Poliovirus replication occurs in the host cell cytoplasm. To initiate infection, the poliovirus virion attaches to a specific receptor on the surface of a sensitive cell and enters the cell. Once inside the cell, the virion is uncoated, and the genomic RNA is attached to ribosomes and translated to yield the polyprotein. Replication of viral RNA by the poliovirus RNA replicase begins shortly after infection. Both the positive and negative strands that are made bind the VPg protein, which also functions as a primer for RNA synthesis following its uridylylation (reaction with uridylic acid; Figure 11.21c). Once poliovirus replication begins, host events are inhibited, and about 5 h after infection, cell lysis occurs with the release of new poliovirus virions.
Check Your Understanding
How can poliovirus RNA be synthesized in the cytoplasm whereas host RNA must be made in the nucleus?
How are protein synthesis and genomic replication similar or different in poliovirus and coronavirus?
11.9 Negative-Strand RNA Animal Viruses
A number of animal viruses have minus-sense RNA genomes (Baltimore class V, Figures 11.2b and 11.3). In contrast to the plus-strand viruses just considered, the genomes of these negative-strand RNA viruses are complementary in base sequence to the mRNA that is formed. We discuss here two important examples of negative-strand RNA viruses: rabies virus and influenza virus. There are no known negative-strand RNA bacteriophages or archaeal viruses.
Rabies Virus
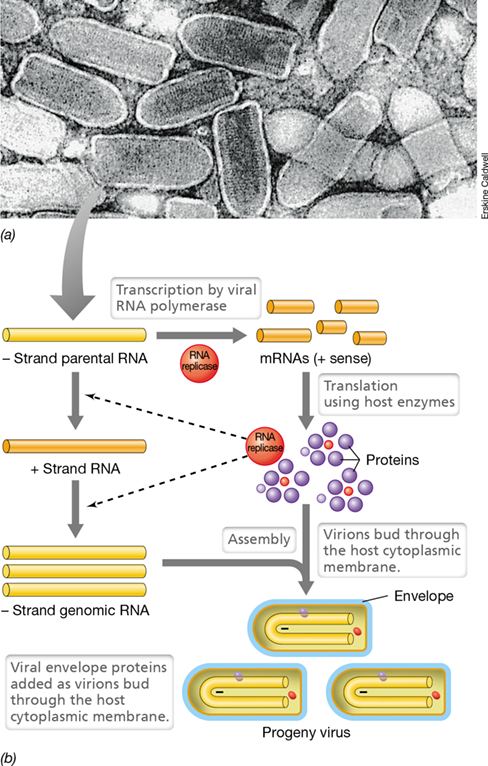
Rabies virus, which causes the fatal neuroinflammatory disease rabies (Section 32.1), is a rhabdovirus, a name that refers to the characteristic shape of the virion (rhabdos is Greek for rod). Rhabdoviruses are commonly bullet-shaped (**Figure 11.23*a***) and have an extensive and complex lipid envelope surrounding the helically symmetrical nucleocapsid. A rhabdovirus virion contains several enzymes that are essential for the infection process, including an RNA replicase. Unlike positive-strand viruses, a rhabdovirus genome cannot be directly translated but must first be transcribed by the replicase. This occurs in the cytoplasm and generates two classes of RNAs. The first is a series of mRNAs encoding each of the viral proteins, and the second is a complementary copy of the entire viral genome; the latter functions as a template for the synthesis of genomic RNA copies (Figure 11.23b).
Figure 11.23 Rhabdoviruses.

(a) Transmission electron micrograph of vesicular stomatitis virus virions. A virion is about 65 nm in diameter. (b) Flow of events during replication of a negative-strand RNA virus. Note the importance of the viral-encoded RNA replicase.
Mastering Microbiology
Art Activity: Figure 11.23b Negative-strand RNA virus: rhabdovirus
Assembly of a rhabdovirus virion is a highly orchestrated process. Two different coat proteins are made, nucleocapsid and envelope. The nucleocapsid is formed first by assembly of nucleocapsid protein molecules around the viral RNA genome. The envelope proteins are glycoproteins and they migrate to the cytoplasmic membrane where they are inserted into the membrane. Nucleocapsids then migrate to areas on the cytoplasmic membrane where these virus-specific glycoproteins are embedded and bud through them, becoming coated by the glycoprotein-enriched cytoplasmic membrane in the process. The final result is the release of new virions that can infect neighboring cells.
Influenza Virus
Another group of negative-strand RNA viruses contains the important human pathogen influenza virus. Influenza virus has been well studied over many years, beginning with early work during the 1918 influenza pandemic that killed millions of people worldwide (Sections 30.8 and 31.8). Influenza virus is an enveloped virus in which the viral genome is present in the virion in a number of separate pieces, a condition called a segmented genome. In the case of influenza A virus, a common strain, the genome is segmented into eight linear single-stranded molecules ranging in size from 890 to 2341 nucleotides and totaling 13.5 kb. The nucleocapsid of the virus is of helical symmetry, about 6–9 nm in diameter and about 60 nm long, and is embedded in an envelope that has a number of virus-specific proteins as well as lipid derived from the host cytoplasmic membrane. Because of the way influenza virus buds as it leaves the cell, virions do not have a uniform shape and instead are pleomorphic (**Figure 11.24*a***).
Figure 11.24 Influenza virus.

(a) Transmission electron micrograph of thin sections of human influenza virus virions. (b) Some of the major components of the influenza virus, including the segmented genome.
Several proteins on the outside of the influenza virion envelope interact with the host cell surface. One of these is hemagglutinin (Figure 11.24b). Hemagglutinin is highly immunogenic (capable of stimulating the immune system), and antibodies against it prevent the virus from infecting a cell. This is the mechanism by which immunity to influenza is brought about by immunization (Section 31.8). A second important influenza virus surface protein is the enzyme neuraminidase (Figure 11.24b). Neuraminidase breaks down sialic acid (a derivative of neuraminic acid) in the host cytoplasmic membrane. Neuraminidase functions primarily in virus assembly, destroying host membrane sialic acid that would otherwise block assembly or become incorporated into the virion. In addition to hemagglutinin and neuraminidase, influenza virions possess two other key enzymes. These include an RNA replicase, which converts the minus-strand genome into a plus strand, and an RNA endonuclease, which cuts the cap from host mRNAs (Section 6.6) and uses them to cap viral mRNAs so they can be translated by the host translational machinery.
After the influenza virion enters the cell, the nucleocapsid separates from the envelope and migrates to the nucleus. Uncoating activates the virus RNA replicase and transcription begins. Ten proteins are encoded by the eight segments of the influenza virus genome; the mRNAs transcribed from six segments each encode a single protein, while the other two segments encode two proteins each. Some of the viral proteins are needed for influenza virus RNA replication, whereas others are structural proteins of the virion. The overall pattern of genomic RNA synthesis resembles that of the rhabdoviruses (Figure 11.23b), with full-length positive-strand RNA used as a template for making negative-strand genomic RNA. The complete enveloped virion forms by budding, as for the rhabdoviruses.
The segmented genome of the influenza virus has important practical consequences. Influenza virus exhibits a phenomenon called antigenic shift in which segments of the RNA genome from two different strains of the virus infecting the same cell are reassorted. This generates hybrid influenza virions that express unique surface proteins unrecognized by the immune system. Antigenic shift is thought to trigger major outbreaks of influenza because immunity to the new forms of the virus is essentially absent from the population. We discuss antigenic shift, and a related phenomenon called antigenic drift, in Section 31.8.
Check Your Understanding
Why is it essential that negative-strand viruses carry an enzyme in their virions?
11.10 Double-Stranded RNA Viruses
Viruses with double-stranded RNA genomes (Baltimore class III, Figure 11.2b) infect animals, plants, fungi, and a few bacteria. Reoviruses are an important family of animal viruses, and we focus on them here.
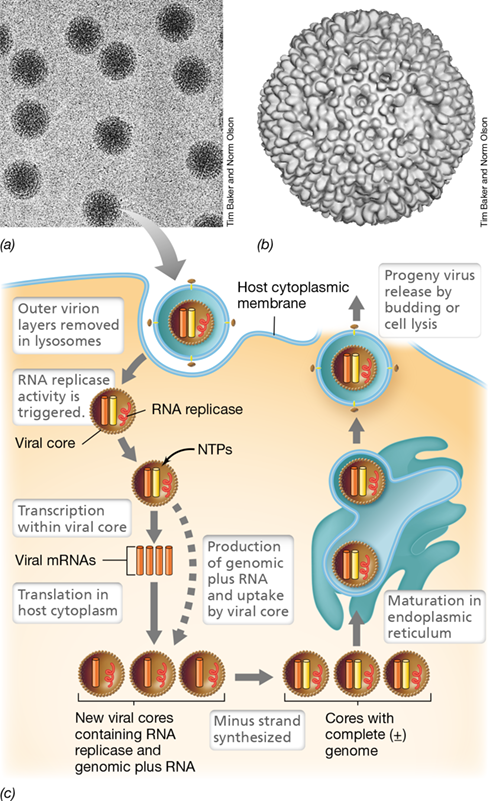
Rotavirus is a typical reovirus and is the most common cause of diarrhea in infants 6 to 24 months of age. Other reoviruses cause respiratory infections, and some infect plants. Reovirus virions consist of a nucleocapsid 60–80 nm in diameter, surrounded by a double shell of icosahedral symmetry (**Figure 11.25a,*b***). As we have seen with single-stranded RNA viruses, the virions of double-stranded RNA viruses must carry their own enzyme to synthesize their mRNA and replicate their RNA genomes. Like the influenza virus genome, the reoviral genome is segmented, in this case into 10–12 molecules of linear double-stranded RNA totaling 18 kb.
Figure 11.25 The reoviruses.

(a) Transmission electron micrograph showing reovirus virions (diameter, about 70 nm). (b) Three-dimensional computer reconstruction of a reovirus virion calculated from electron micrographs of frozen-hydrated virions. (c) The reovirus life cycle. All replication and transcription steps occur inside the nucleocapsids. NTPs, nucleotide triphosphates.
To initiate infection, a reovirus virion binds to a cellular receptor protein. The attached virus then enters the cell and is transported into lysosomes, where normally it would be destroyed (Section 2.15). However, only the outer coats of the virion are removed by the lysosome, revealing the nucleocapsid; the latter is released into the cytoplasm. This uncoating process activates the viral RNA replicase and initiates virus replication (Figure 11.25c).
Reovirus Replication
Reovirus replication occurs exclusively in the host cytoplasm but within the nucleocapsid itself (Figure 11.25c) because the host has enzymes that recognize double-stranded RNA as foreign and would destroy it. The plus strand of the reoviral genome is inactive as mRNA, and thus the first step in replication is the synthesis of plus-sense mRNA by the viral-encoded RNA replicase, using minus-strand RNA as a template; the nucleotide triphosphates necessary for RNA synthesis are supplied by the host (Figure 11.25c). The mRNAs are then capped with a methylated nucleotide (as is typical of eukaryotic mRNAs, Section 6.6) by viral enzymes and exported from the nucleocapsid into the cytoplasm and translated by host ribosomes.
Most RNAs in the reovirus genome encode a single protein, although in a few cases the protein formed is cleaved to yield the final products. However, one of the reovirus mRNAs encodes two proteins but the RNA does not have to be processed in order to translate both of these. Instead, a ribosome occasionally “misses” the start codon for the first gene in this mRNA and travels on to the start codon of the second gene to begin translation. When this occurs, the second protein, needed in small amounts, is made but the first protein is not. This “molecular mistake” can be viewed as a primitive form of translational control that ensures that viral proteins are made in their proper amounts.
As viral proteins are formed in the host cytoplasm, they aggregate to form new nucleocapsids, trapping copies of RNA replicase inside as they form (Figure 11.25c). Newly formed nucleocapsids then take up the correct complement of genomic (plus-strand) RNA fragments—probably by recognition of specific sequences on each fragment—and as each single-stranded RNA enters a newly formed nucleocapsid, a double-stranded form is produced from it by RNA replicase. Once genomic synthesis is complete, viral coat proteins are added in the host’s endoplasmic reticulum, and the mature reoviral virions are released by budding or cell lysis (Figure 11.25c).
Reoviruses and RNA Replication
In addition to the unusual genome structure of reoviruses that forces them to employ special mechanisms to protect their double-stranded RNA genomes from cleavage by host ribonucleases (Figure 11.25c), genomic replication in these viruses is also unique and differs in a fundamental way from that of cells and all other viruses.
Because the reovirus RNA genome is double-stranded, one might predict that reovirus replication would parallel that of organisms with double-stranded DNA genomes, but this is not the case. RNA replication in reoviruses is actually a conservative process rather than the well-known semiconservative process typical of cellular DNA replication and replication in viruses that contain double-stranded DNA genomes (Section 6.3 and Figure 11.2). This is because in the synthesis of reovirus mRNA only the minus strand is a template in the infecting nucleocapsids, whereas only the plus strand is a template in the synthesis of double-stranded genomic RNA from assimilated plus-strand RNA copies in progeny virions (Figure 11.25c). Hence, in addition to their unique double-stranded RNA genomes, reoviruses also display their unusual molecular biology by employing a unique genomic replication mechanism that is neither semiconservative nor rolling circle (Figure 11.7) in nature.
Check Your Understanding
How does reovirus genome replication resemble that of influenza virus, and how does it differ?
Why must reoviral replication events occur within the nucleocapsid?
11.11 Viruses That Use Reverse Transcriptase
Two different Baltimore classes of viruses use reverse transcriptase, an enzyme that can produce DNA from an RNA template, but they differ in the type of nucleic acid in their genomes; retroviruses have RNA genomes while hepadnaviruses have DNA genomes (Baltimore classes VI and VII, respectively, Figure 11.2). Besides their unique molecular properties, both classes of viruses include important human pathogens, including HIV (a retrovirus) and hepatitis B (a hepadnavirus).
Retroviruses and Integration into the Host Genome
Retroviruses have enveloped virions that contain two identical copies of the single-stranded plus complementarity RNA genome (Figure 11.2b and **Figure 11.26*a***). The prefix retro means “backward,” and the term retrovirus refers to the fact that these viruses transfer information from RNA to DNA (in contrast to genetic information flow in cells, which occurs from DNA to RNA). The virion of a retrovirus contains several enzymes, including reverse transcriptase and integrase, and also a specific viral tRNA. Enzymes for retrovirus replication must be carried in the virion because although the retroviral genome is of the plus sense, it is not used directly as mRNA. **Figure 11.26*b*** illustrates the general scheme for retroviral genome conversion to DNA, integration into the host chromosome, and production of new virions. To begin the process, the genome is converted to DNA by reverse transcriptase inside the nucleocapsid and then the DNA is released to the cytoplasm. The double-stranded DNA is then integrated into the host genome by integrase.
Figure 11.26 Retrovirus structure and replication.

(a) The virion carries two identical copies of the RNA genome (orange). (b) Top, an overview of the infection process. Replication steps: Reverse transcriptase, carried in the virion, makes single-stranded DNA from viral RNA and then double-stranded DNA that integrates into the host genome as a provirus. Transcription and translation of proviral genes leads to the production of new virions that are then released by budding.
Reverse transcriptase has three enzymatic activities: (1) reverse transcription (to synthesize DNA from an RNA template), (2) ribonuclease activity (to degrade the RNA strand of an RNA:DNA hybrid), and (3) DNA polymerase (to make double-stranded DNA from single-stranded DNA). A more detailed look at steps in reverse transcription is presented in Figure 11.27. Reverse transcriptase needs a primer for DNA synthesis and this is the function of the viral tRNA. Using this primer, nucleotides near the 5′ terminus of the RNA are reverse-transcribed into DNA. Once reverse transcription reaches the 5′ end of the RNA, the process stops. To copy the remaining RNA, a different mechanism comes into play. First, terminally redundant RNA sequences at the 5′ end of the molecule are removed by reverse transcriptase. This leads to the formation of a small, single-stranded DNA that is complementary to the RNA segment at the other end of the viral RNA. This short, single-stranded piece of DNA then hybridizes with the other end of the viral RNA molecule, where synthesis of DNA begins once again.
Figure 11.27 Formation of double-stranded DNA from retrovirus single-stranded genomic RNA.

The sequences labeled R on the RNA are direct repeats found at either end. The sequence labeled PB is where the primer (tRNA) binds. Note that DNA synthesis yields longer direct repeats on the DNA than were originally on the RNA. These are called long terminal repeats (LTRs). Inset: A special form of fluorescence microscopy allows visualization of HIV genome integration (green) into a chromosome region near the nuclear membrane (red) of the host cell, a CD4 lymphocyte. Image courtesy of Marina Lusic, University Hospital, Heidelberg, Germany.
Mastering Microbiology
Art Activity: Figure 11.27 Formation of double-stranded DNA from retrovirus single-stranded genomic RNA
Continued reverse transcription leads to the formation of a double-stranded DNA molecule with long terminal repeats, and these assist in integration of the retroviral DNA into the host chromosome by the integrase enzyme carried in the virion (Figure 11.27). For HIV, the chromosomal integration site is not random. Through the use of a special form of fluorescence microscopy, scientists have shown that HIV incorporates into chromosomal loci near the outer shell of the nucleus (photo inset, Figure 11.27). This location is likely favored due to the short life of the viral integrase. Thus the retroviral DNA must be integrated quickly into the host genome upon entry into the nucleus.
Once integrated, retroviral DNA becomes a permanent part of the host chromosome and is considered a provirus (Section 5.7); the genes may be expressed or they may remain in a latent state indefinitely. However, if induced, retroviral DNA is transcribed by a cellular RNA polymerase to form RNA transcripts (plus-sense mRNA) that can be either packaged into virions as genomic RNA or translated to yield retroviral proteins.
With retroviruses we thus see a replication scheme that is perhaps the most complex of all known viruses. Despite the complexity of retroviruses, molecular studies of them suggest an ancient origin and possible central importance in the transition from self-replicating RNA life forms to the DNA world of cellular organisms. It is the signature enzyme of retroviruses—reverse transcriptase, the only enzyme known that can make DNA from RNA—that brings retroviruses into this evolutionary limelight, and so it is possible that all cells owe their very existence to this class of viruses. We revisit this topic in Section 13.5.
Check Your Understanding
Why are protease inhibitors an effective treatment for human AIDS?
Contrast the genomes of HIV and hepatitis B virus.
How does the role of reverse transcriptase in the replication cycles of retroviruses and hepadnaviruses differ?
IV Subviral Agents
A few noncellular microbes are even more streamlined than viruses; some contain RNA but lack protein while others contain protein but lack nucleic acids of any type. Although biologically minimalist entities, these structures can trigger devastating diseases.
We conclude our genomic tour of the viral world by considering two subviral agents: the viroids and the prions. These are infectious agents that resemble viruses but lack either nucleic acid (prions) or protein (viroids) and are thus not viruses.
11.12 Viroids
Viroids are infectious RNA molecules that lack a protein component. Viroids are small, circular, single-stranded RNA molecules that are the smallest known pathogens. They range in size from 246 to 399 nucleotides and show a considerable degree of sequence homology to each other, suggesting that they have common evolutionary roots. Viroids cause a number of important plant diseases and can have a severe agricultural impact (Figure 11.29). A few well-studied viroids include coconut cadang-cadang viroid (246 nucleotides) and potato spindle tuber viroid (359 nucleotides). No viroids are known that infect animals or microorganisms.
Figure 11.29 Viroids and plant diseases.

Photograph of healthy tomato plant (left) and one infected with potato spindle tuber viroid (PSTV) (right). The host range of most viroids is quite restricted. However, PSTV infects tomatoes as well as potatoes, causing growth stunting, a flat top, and premature plant death.
Viroid Structure and Function
The extracellular form of a viroid is naked RNA; there is no protein capsid. Although the viroid RNA is a single-stranded, covalently closed circle, its extensive secondary structure forms a hairpin-shaped double-stranded molecule with closed ends (Figure 11.30). This makes the viroid sufficiently stable to exist outside the host cell and protects the viroid while inside the cell from cellular ribonucleases. Because it lacks a capsid, a viroid does not use a receptor to enter the host cell. Instead, the viroid enters a plant cell through a wound, as from insect or other mechanical damage. Once inside, viroids move from cell to cell via plasmodesmata, which are the thin strands of cytoplasm that link plant cells (Figure 11.31).
Figure 11.30 Viroid structure.

Viroids consist of single-stranded circular RNA that forms a seemingly double-stranded structure by intra-strand base pairing.
Figure 11.31 Viroid movement inside plants.

After entry into a plant cell, viroids (orange) replicate either in the nucleus or the chloroplast. Viroids can move between plant cells via the plasmodesmata (thin threads of cytoplasm that penetrate the cell walls and connect plant cells). On a larger scale, viroids can also move around the plant via the plant vascular system.
Viroid RNAs do not encode proteins and thus the viroid is totally dependent on its host for replication. Plants have several RNA polymerases, one of which has RNA replicase activity, and this is the enzyme that replicates the viroid. The replication mechanism itself resembles the rolling circle mechanism used for genome synthesis by some small viruses (Sections 11.3, 11.4, and 11.7). The result is a large RNA molecule containing many viroid units joined end to end. The viroid has ribozyme activity (catalytic RNA, Section 13.1) and uses it to self-cleave the large RNA molecule, releasing individual viroids.
Viroid Disease
Viroid-infected plants can be symptomless or show symptoms that range from mild to lethal, depending on the viroid (Figure 11.29). Most disease symptoms are growth related, and it is believed that viroids mimic or in some way interfere with plant small regulatory RNAs. In fact, viroids could themselves be derived from regulatory RNAs that have evolved away from carrying out beneficial roles in the cell to inducing destructive events. Viroids are known to yield small interfering RNAs (siRNAs) as a by-product of replication. It has been proposed that these siRNAs may function by way of an RNA-interference silencing pathway, similar to that of regulatory RNAs in Bacteria and Archaea (Section 7.12), to suppress the expression of plant genes that show some homology to viroid RNA, and in this way induce disease symptoms. This mechanism of regulation is similar to how some bacterial and archaeal regulatory small RNAs target mRNAs for degradation (Figure 7.28b).
Check Your Understanding
If viroids are circular molecules, why are they depicted as hairpins?
11.13 Prions
Prions represent the opposite extreme from that of viroids. Prions are infectious agents whose extracellular form consists entirely of protein. That is, a prion lacks both DNA and RNA. Prions cause several neurological diseases such as scrapie in sheep, bovine spongiform encephalopathy (BSE or “mad cow disease”) in cattle, chronic wasting disease in deer and elk, and kuru and variant Creutzfeldt–Jakob disease in humans by catalyzing protein conformational changes that lead to protein clumping and accumulation. No prion diseases of plants are known, although prions have been detected in yeast. Collectively, animal prion diseases are known as transmissible spongiform encephalopathies.
Prion Proteins and the Prion Infectious Cycle
If prions lack nucleic acid, how is prion protein encoded? The answer to this conundrum is that the host cell itself encodes the prion. The host contains a gene, Prnp (Prion protein), which encodes the native form of the prion, known as PrPC (Prion Protein Cellular). This is primarily found in the neurons of healthy animals, especially in the brain (**Figure 11.32*a***). The pathogenic form of the prion protein is designated PrPSc (prion protein Scrapie) because the first prion disease to be discovered was that of scrapie in sheep. PrPSc is identical in amino acid sequence to PrPC from the same animal species, but it has a different conformation. For example, native prion proteins are largely α-helical, whereas the pathogenic forms contain less α-helix and more β-sheet secondary structure. Prion proteins from different species of mammals are similar but not identical in amino acid sequence, and host range is linked in some way to protein sequence. For example, PrPSc from BSE-diseased cattle can infect humans, whereas PrPSc from scrapie-infected sheep apparently cannot.
Figure 11.32 Prions.

(a) Section through brain tissue of a human with variant Creutzfeldt–Jakob disease. Note the spongy nature of the tissue (clearings, arrow) where neural tissue has been lost. (b) Mechanism of prion misfolding. Neuronal cells produce the native form of the prion protein. The pathogenic form catalyzes the refolding of native prions into the pathogenic form. The pathogenic form is protease resistant, insoluble, and forms aggregates in neural cells. This eventually leads to destruction of neural tissues (see part a) and neurological symptoms. About 70% of those diagnosed with variant Creutzfeldt–Jakob disease die within a year.
When the PrPSc form enters a host cell that is expressing PrPC, it promotes the conversion of PrPC into the pathogenic form (Figure 11.32b). That is, the pathogenic prion “replicates” by converting preexisting native prions into the pathogenic form. As the pathogenic prions accumulate and aggregate, they form insoluble crystalline fibers referred to as amyloids in neural cells; this leads to disease symptoms including the destruction of brain and other nervous tissue (Figure 11.32a). PrPC functions in the cell as a cytoplasmic membrane glycoprotein, and it has been shown that membrane attachment of pathogenic prions is necessary before disease symptoms commence. Mutant versions of PrPSc that can no longer attach to nerve cell cytoplasmic membranes still aggregate but no longer cause disease.
Besides the transmissible spongiform encephalopathies, amyloids are also associated with debilitating human diseases such as Alzheimer’s, Huntington’s, Parkinson’s, and type 2 diabetes. Whether any or all of these are truly prion diseases is unclear. However, protein aggregation has been linked to these diseases, so the possibility remains that they are manifestations of prion infection.
Nonpathogenic Prions
Certain fungi have proteins that fit the prion definition of an inherited self-perpetuating change in protein structure, although these proteins do not cause noticeable disease. Instead they adapt the fungal cells to survive dynamic environmental conditions by conferring traits such as altered nutrient utilization, antibiotic resistance, and biofilm formation. In yeast, for example, the [URE3] prion is a protein that regulates the transcription of genes encoding certain nitrogen metabolism functions. The normal (soluble) form of this protein functions to repress genes encoding proteins that metabolize certain nitrogen sources. However, when the [URE3] prion accumulates, it forms insoluble aggregates and when this occurs, the normally repressed genes are derepressed and the nitrogen sources are metabolized.
Humans also have beneficial prions. MAVS is a human protein that is part of our innate immune system and has been shown to convert to a self-perpetuating prion-like form in cells that become infected with a virus. The aggregation of MAVS protein triggers the production of immune modulators called interferons (Section 26.10). These proteins trigger the recruitment of phagocytic cells such as macrophages (cells that ingest and destroy pathogens, foreign particles, and cell debris) and other immune factors to the viral-infected cell, resulting in its destruction. While the conversion of MAVS to a prion-like form ultimately kills the cell, it prevents the virus from hijacking the cell’s molecular machinery, which would allow the virus to replicate and lyse the host cell to infect adjacent cells.
The viral and subviral worlds have revealed an almost limitless variation on the molecular principles that govern information flow in cells. We turn our attention in the following chapter to how scientists have exploited some of these unusual molecular principles (along with the enormous repertoire of genetic tools available from cells) to create genetically modified or even totally synthetic organisms for the benefit of mankind.
Check Your Understanding
On what basis can prions be differentiated from all other infectious agents?
What is the difference between the native and pathogenic forms of the prion protein?
How does a prion differ from a viroid? How does a prion differ from a virus?
Chapter Review
Go to Mastering Microbiology for videos, animations, practice tests, and more.
I Viral Genomes and Classification
11.1 Viral genomes can be single-stranded or double-stranded DNA or RNA and vary from a few to hundreds of kilobases in size. Viral mRNA is always of the plus configuration by definition, but single-stranded genomes can be of the plus or minus configuration. Viruses with RNA genomes must either carry an RNA replicase in their virions or encode this enzyme in their genomes in order to synthesize RNA from an RNA template.
Q Describe the classes of viruses based on their genomic characteristics. For each class, describe how viral mRNA is made and how the viral genome is replicated.
11.2 Besides classification based on genomic characteristics, viruses can also be assigned to orders, families, genera, and species based on taxonomy. Taxonomic classifications are made on a combination of host range, structural morphology, replication cycle, nucleic acid sequence, and pathogenicity. Phylogenetic trees illustrating viral relationships can also be constructed using genes conserved within a group of viruses.
Q What trait associated with archaeal viruses is likely responsible for the high percentage of viruses assigned to unclassified orders?
II DNA Viruses
11.3 Single-stranded DNA viruses contain DNA of the plus configuration, and a double-stranded replicative form is necessary for transcription and genome replication. The genome of the virus ϕX174 is so small that some of its genes overlap, and the genome replicates by a rolling circle mechanism. Some related viruses, such as M13, have filamentous virions that are released from the host cell without lysis.
Q Describe how the genome of bacteriophage ϕX174 is transcribed and translated.
11.4 Bacteriophage T4 contains a double-stranded DNA genome that is both circularly permuted and terminally redundant. T4 encodes its own DNA polymerase and several other replication proteins, but it utilizes the host RNA polymerase to express early, middle, and late viral genes. Bacteriophage T7 contains a double-stranded DNA genome that encodes both early genes, transcribed by the host RNA polymerase, and late genes, transcribed by a virus-encoded RNA polymerase. Replication of the T7 genome employs T7 DNA polymerase and involves terminal repeats and concatemers. Bacteriophage lambda possesses a linear double-stranded DNA genome that circularizes at the cos sites once injected into its host. Depending on cellular conditions, lambda can either integrate into the host chromosome and be maintained as a prophage through lysogeny or enter the lytic cycle and be replicated by rolling circle replication.
Q Why can it be said that transcription of the bacteriophage T4 genome requires one enzyme, while the T7 genome requires two enzymes? How does lambda get incorporated into a host cell’s chromosome?
11.5 Several double-stranded DNA viruses infect cells of Archaea, most of which inhabit extreme environments. Many of these genomes are circular, in contrast to the linear double-stranded DNA genomes of bacteriophages. Although many head-and-tail viruses are known, other archaeal viruses have an unusual spindle-shaped morphology.
**Q List three unusual features of the archaeal virus that infects Acidianus that distinguish it from bacteriophage T7.**
11.6 Pox viruses are large, double-stranded DNA viruses that replicate entirely in the cytoplasm and are responsible for several human diseases, including smallpox. Adenoviruses are double-stranded DNA viruses whose genome replication employs protein primers and a mechanism that occurs without lagging-strand synthesis.
Q Of all the double-stranded DNA animal viruses, pox viruses stand out concerning one unique aspect of their DNA replication process. What is this unique aspect, and how can this be accomplished without special DNA replication enzymes being packaged in the virion?
11.7 Some double-stranded DNA viruses cause cancer in humans. SV40 is such a tumor virus and has a tiny genome containing overlapping genes. The activity of certain SV40 genes can trigger cell transformation (tumor induction). Some herpesviruses also cause cancer, but most cause various human infectious diseases. Herpesviruses can maintain themselves in a latent state in the host indefinitely, initiating viral replication periodically.
Q What is unusual about the envelope of the herpesvirus?
III RNA Viruses
11.8 In single-stranded plus-sense RNA viruses, the genome is also the mRNA, and a negative strand is synthesized to produce more mRNA and genome copies. The tiny bacteriophage MS2 contains only four genes, one of which encodes a subunit of its RNA replicase. In poliovirus, the viral RNA is translated directly, producing a polyprotein that is cleaved into several small viral proteins. Coronaviruses are large RNA viruses that resemble poliovirus in some but not all of their replication features.
Q What is the function of the VPg protein of poliovirus, and how can coronaviruses replicate without a VPg protein?
11.9 In negative-strand viruses, the virus RNA is not mRNA but must first be copied to form mRNA by RNA replicase present in the virion. The positive strand is the template for production of genome copies. Important pathogenic negative-strand viruses include rabies virus and influenza virus.
Q Rabies virus and poliovirus both have single-stranded RNA genomes, but only in poliovirus can the genome be translated directly. Explain.
11.10 Reoviruses contain segmented linear double-stranded RNA genomes. Like negative-strand RNA viruses, reoviruses contain an RNA-dependent RNA polymerase within the virion. Newly forming virions take up plus-strand RNA that is the template for the complementary strand produced in the new virions.
Q Compare the reovirus genome to those of influenza virus and bacteriophage MS2. Why do reovirus replication events have to happen in the nucleocapsid?
11.11 Some viruses employ reverse transcriptase, including retroviruses (HIV) and hepadnaviruses (hepatitis B). Retroviruses have single-stranded RNA genomes and use reverse transcriptase to make a DNA copy. Hepadnaviruses contain partially double-stranded DNA genomes and use reverse transcriptase to make a single strand of genomic DNA from a full-length complementary strand of RNA.
Q Why do both hepadnaviruses and retroviruses require reverse transcriptase when their genomes are double-stranded DNA and single-stranded RNA, respectively?
IV Subviral Agents
11.12 Viroids are circular single-stranded RNA molecules that do not encode proteins and are dependent on host-encoded enzymes for replication. Unlike viruses, viroid RNA is not enclosed within a capsid, and all known viroids are plant pathogens.
Q What are the similarities and differences between viruses and viroids?
11.13 Prions are made of protein but have no nucleic acid of any kind. Prions exist in two conformations, the native cellular form and the pathogenic form, which takes on a different protein structure. The pathogenic form “replicates” itself by converting native prion proteins, encoded by the host cell, into the pathogenic conformation.
Q What are the similarities and differences between prions and viruses?
Application Questions
Not all proteins are made from the RNA genome of bacteriophage MS2 in the same amounts. Can you explain why? One of the proteins functions very much like a repressor, but it functions at the translational level. Which protein is it and how does it function?
Replication of both strands of DNA in adenoviruses occurs in a continuous (leading) fashion. How can this happen without violating the rule that DNA synthesis always occurs in a 5′→3′ direction?
Imagine that you are a researcher at a pharmaceutical company charged with developing new drugs against human RNA viral pathogens. Describe at least two types of drugs you might pursue, what class of virus they would affect, and why you feel that the drugs would not harm the patient.
Reoviruses contain genomes that are unique in all of biology. Why? Why can’t reovirus replication occur in the host cytoplasm? Contrast reovirus genomic replication events with those of a cell. Why can it be said that reovirus genome replication is not semiconservative even though the reovirus genome consists of complementary strands?
Chapter Glossary
a genomic nucleic acid molecule formed from the recombination of several genomic units Hepadnavirus
a virus whose DNA genome replicates by way of an RNA intermediate Negative strand
a nucleic acid strand that has the opposite sense to (is complementary to) the mRNA Overlapping genes
two or more genes in which part or all of one gene is embedded in another gene Polyprotein
a large protein expressed from a single gene and subsequently cleaved to form several individual proteins Positive strand
a nucleic acid strand that has the same sense as the mRNA Prion
an infectious protein whose extracellular form lacks nucleic acid Replicative form
a double-stranded molecule that is an intermediate in the replication of viruses with single-stranded genomes Retrovirus
a virus whose RNA genome is replicated via a DNA intermediate Reverse transcriptase
the retroviral enzyme that produces DNA from an RNA template Reverse transcription
the process of copying genetic information found in RNA into DNA RNA replicase
an enzyme that can produce RNA from an RNA template Rolling circle replication
a mechanism, used by some plasmids and viruses, of replicating circular DNA, which starts by nicking and unrolling one strand. For a single-stranded genome, this is preceded by using the still-circular strand as a template for DNA synthesis; for a double-stranded genome, the unrolled strand is used as a template for DNA synthesis Viroid
an infectious RNA whose extracellular form lacks protein