14 Metabolic Diversity of Microorganisms
## Chapter 14 Metabolic Diversity of Microorganisms
III Respiratory Processes Defined by Electron Donor
IV Respiratory Processes Defined by Electron Acceptor
Ferreting Out the Peculiar Life of Iron Bacteria
Life requires the movement of electrons from one place to another, and microbes have found many different ways to accomplish this. Many iron-oxidizing bacteria are autotrophic chemolithotrophs that conserve energy by oxidizing Fe2+ and transporting electrons to an electron acceptor such as O2. Fe2+ is present in anoxic groundwater, and the “iron bacteria” grow where this ferrous-rich water reaches the oxic surface. However, aerobic iron bacteria face an unusual challenge because when reduced iron meets O2, it spontaneously oxidizes to form rust. To survive, iron bacteria must outcompete this abiotic process. Another challenge is that iron bacteria form iron oxide minerals as waste products, and as they grow, they risk becoming entombed within a metallic crust of their own making.
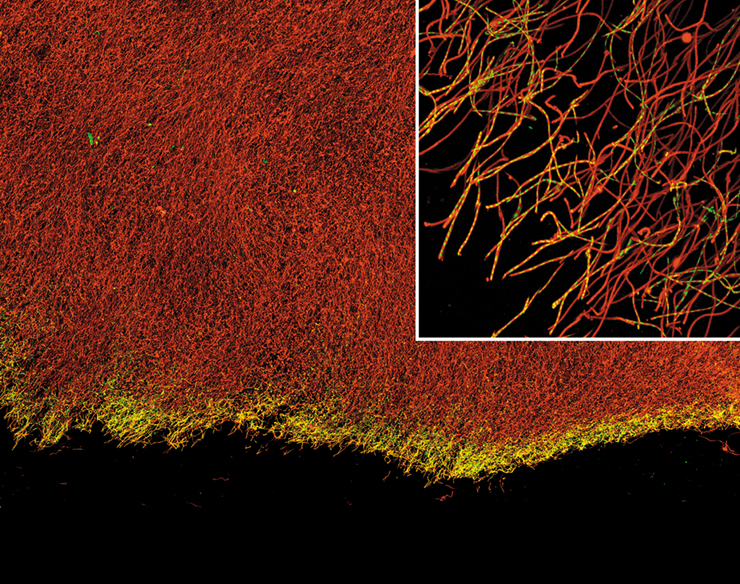
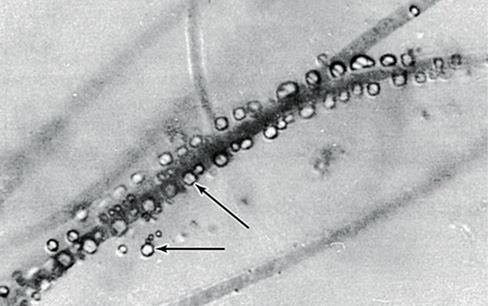
Iron bacteria have evolved elegant strategies for overcoming these obstacles. They have specialized mechanisms for donating electrons to extracellular electron acceptors, and this prevents iron oxides from forming within cells. In addition, they catalyze iron biomineralization, forming stalks and sheaths that are extruded away from the cell. The photos here show cells of the iron bacterium Leptothrix ochracea forming a microbial mat in a spring in Spruce Point, Maine (USA). In these fluorescence micrographs, the cells (about 5 μm long) are stained yellow and occur only at the tips of hollow iron oxide sheaths, stained red. The aerobic oxidation of iron provides only a very small amount of energy and because of this, iron bacteria leave long empty iron oxide sheaths as they grow. The cells sense and constantly move toward O2, meaning that sheath production functions almost as a form of motility that allows L. ochracea to position itself effectively in response to gradients of Fe2+ and O2.
Microbes are often defined by their metabolic properties, and to understand bacteria like L. ochracea, we must first understand their metabolism.
Source: Chan, C.S., et al. 2016. The architecture of iron microbial mats reflects the adaptation of chemolithotrophic iron oxidation in freshwater and marine environments. Front. Microbiol. 7: 796.
A major theme of microbiology is the great phylogenetic diversity of microbial life on Earth. We will explore this microbial diversity in the following four chapters. In this chapter we focus on the metabolic diversity of microorganisms, with special emphasis on the processes and mechanisms that underlie this diversity. Metabolism provides one framework we can use to make sense of microbial diversity. In the following chapters, we will see that microbial diversity can be understood in terms of metabolic diversity, ecological diversity, and phylogenetic diversity.
I Introduction to Metabolic Diversity
All cells conserve energy, obtain reducing power, and achieve redox balance by carrying out oxidation–reduction reactions. Metabolic pathways are modular and often reversible, facilitating the evolution of metabolic diversity through horizontal gene transfer.
While microbial metabolism is amazingly diverse, all microbes follow the same basic metabolic principles. These principles were introduced in Chapter 3, and we revisit them now and expand upon them to see how they apply to diverse metabolic types. In all forms of metabolism, energy is conserved and reducing power is obtained during catabolic reactions (Figure 3.1), and cells grow by coupling this energy to anabolic (biosynthetic) reactions such as CO2 fixation. We begin our review of metabolic diversity by reviewing the mechanisms of energy conservation and their relationship to redox reactions.
14.1 Foundational Principles of Metabolic Diversity: Energy and Redox
14.1 Foundational Principles of Metabolic Diversity: Energy and Redox
14.1 Foundational Principles of Metabolic Diversity: Energy and Redox
The diversity of metabolic types in the microbial world is tremendous, and these diverse organisms are essential to sustain our biosphere. In part, this diversity is driven by the modularity of metabolic reactions. Metabolic modularity facilitates the evolution of metabolic diversity by allowing the formation of new pathways from the horizontal transfer and modification of existing enzymes and pathways. For example, the components of electron transport allow for a tremendous diversity of respirations, most of which do not involve O2 and are thus anaerobic respirations (Figure 14.1). By understanding the principles that underlie metabolism, we can move beyond the memorization of pathways as unique entities and instead learn to predict the metabolic needs of any organism we encounter in the microbial world.
Figure 14.1 Diversity of respiration reactions.

The redox couples (right, yellow box) are arranged in order from most electronegative E0′ (top) to most electropositive E0′ (bottom), assuming neutral pH. An exergonic reaction results when an electron donor (left, green box) donates electrons to an electron acceptor from a more electropositive (lower) redox couple. See Figure 3.4 and Table 3.1 to compare how the energy yields of some of these anaerobic respirations vary. The E0′ of the Fe3+/Fe2+ couple at pH 2 is +0.77 V. NH3 is shown to indicate its relative E0′ value as a donor. Corg, organic carbon compounds (glucose, ethanol, etc.).
All organisms need to conserve energy by converting chemical or light energy into ATP. They also need a source of reducing power, a source of highly electronegative electrons for performing redox reactions. Finally, they need to achieve redox balance by regenerating oxidized electron carriers (such as NAD+) through the use of an external electron acceptor (in respiration), or by recycling electrons back onto a metabolic intermediate and excreting them as fermentation products (in fermentation).
Conservation of Energy
All cells conserve energy by coupling a flow of electrons to the synthesis of ATP. Electron flow in the cell consists of a series of redox reactions. Recall from Chapter 3 that redox reactions are reactions in which electrons are transferred from one substance (in one half reaction that donates electrons) to another substance (in a second half reaction that accepts electrons). The tendency of a substance to donate or accept electrons is defined by its reduction potential (E0′, Figure 14.1; Figure 3.4 and Table 3.1). The farther apart the two half reactions are in terms of the E0′ of their redox couples, the greater the amount of energy available (Figure 3.4).
Based on these simple principles governing redox reactions, a wide variety of organic or inorganic electron donors can be coupled to a wide variety of external electron acceptors (through respiration, Figure 14.1) or internal metabolic products (through fermentation), generating the tremendous diversity of metabolic options that exist in the microbial world. These reactions can support growth, provided sufficient energy is released for the production of ATP (−31.8 kJ/mol). Ultimately, the flow of electrons through redox reactions provides the energy for ATP synthesis by one of three mechanisms: substrate-level phosphorylation, oxidative phosphorylation, or photophosphorylation (Chapter 3). Chemotrophs conserve energy by the former two mechanisms while phototrophs use the latter. We will consider phototrophs in detail in Sections 14.3, 14.4, 14.5 and 14.6 and focus first on chemotrophic metabolism.
Chemotrophic Metabolism
Chemotrophic organisms can conserve energy through either fermentation or respiration. Fermentation does not require an external electron acceptor and ATP is generated primarily by substrate-level phosphorylation, with electron balance obtained by reducing metabolic intermediates that are then excreted as fermentation products (Section 3.7). Respiration, by contrast, requires an external electron acceptor, and ATP is generated by oxidative phosphorylation resulting from electron transport reactions that generate a proton motive force (pmf); the enzyme complex ATP synthase harnesses this force to make ATP (Section 3.9). Chemolithotrophs are chemotrophic organisms that use inorganic electron donors in respiration reactions (Table 14.1). Note in Table 14.1 how the energy yield of each reaction shown is determined by the ΔE0′ of its redox couples.
Table 14.1 Energy yields from the oxidation of various inorganic electron donorsa

aData are from Gf 0 values in Table 3.1 (or references therein) and Figure 3.4, and from bioenergetics calculations as described in Sections 3.2 and 3.3; E0′ values for Fe2+ are for pH 2, and others are for pH 7. At pH 7, the E0′ for the Fe3+/Fe2+ couple is about +0.2 V.
bExcept for phosphite, all reactions are shown coupled to O2 as electron acceptor. The only known phosphite oxidizer couples to SO4 2− as electron acceptor. H2 and most sulfur compounds can be oxidized anaerobically using one or more electron acceptors, and Fe2+ can be oxidized at neutral pH with NO3 − as electron acceptor. For other chemolithotrophic reactions of sulfur compounds, see Table 14.2.
Respiration can occur under both oxic and anoxic conditions. Anaerobic respirations are those whose electron acceptors are other than O2. From Figure 14.1 we can see that almost any half reaction can serve as an electron acceptor if coupled with a sufficiently electronegative electron donor. Because the O2/H2O couple is so electropositive, more energy is typically available from aerobic respiration than from anaerobic respiration. Therefore, for any given electron donor, aerobic organisms will always conserve more energy—and will therefore outcompete—anaerobic organisms. However, because O2 is such a good electron acceptor, and because it is poorly soluble in water, it is consumed rapidly in poorly mixed environments. Thus, anoxic habitats and anaerobic organisms are widespread in nature.
Reducing Power and Redox Balance
The conservation of energy on its own is not sufficient to support life. Cells also need a source of reducing power for biosynthesis and need to achieve redox balance by balancing electron flow within the cell. The reducing power required for biosynthetic reactions is typically in the form of low-potential electron carriers such as NAD(P)H (E0′=−0.32 V) or reduced ferredoxin (Fdred, E0′=−0.37 to −0.50 V). In the case of most chemoorganotrophs, this reducing power is readily generated during the oxidation of reduced organic molecules (Sections 3.6 and 3.7). The challenge for chemoorganotrophs is managing their redox balance; that is, they need a way to export electrons from the cell in order to regenerate the oxidized NAD(P)+ or Fdox they need for catabolic reactions. Chemoorganotrophs achieve redox balance by donating electrons to external electron acceptors (in respiration) or metabolic intermediates (in fermentation) that are reduced and excreted from the cell. In contrast, the problem for many chemolithotrophs and some phototrophs (Section 3.11) is a lack of reducing power. Their electron donors (such as S0, H2S, NH3, and the like) are unable to reduce the NAD(P)+ or Fdox they need for catabolic and biosynthetic reactions. Hence, these organisms must generate reducing power by coupling the endergonic reduction of these electron carriers to some other exergonic reaction.
Three mechanisms of energy coupling are known that can increase reducing power or achieve redox balance at the expense of energy. The first is by coupling a reaction directly to ATP hydrolysis; this requires a substantial energy expenditure by the cell and so is used primarily by organisms that perform highly exergonic catabolic reactions. The second is reverse electron transport (Section 3.11) wherein the endergonic reduction of NAD(P)+ or Fdox is driven by dissipation of the pmf. For example, during electron transport, an NADH:quinone oxidoreductase (such as Complex I, Figure 3.19) can donate electrons to an oxidized quinone, an exergonic reaction that contributes to the formation of the pmf. However, during reverse electron transport, this enzyme runs in the reverse direction and transfers electrons from a reduced quinone to NAD+, an endergonic reaction driven by the pmf.
The third, and most energy-efficient, mechanism of energy coupling is flavin-based electron bifurcation (Figure 14.2). In electron bifurcation, the endergonic reduction of a low-potential electron acceptor such as Fdox is driven by the exergonic reduction of a higher-potential electron acceptor such as NAD+ (Figure 14.2). Electron-bifurcating enzymes contain a flavin coenzyme. The flavin accepts two electrons at a time and donates one of these electrons in an exergonic reaction to a higher-potential electron acceptor (such as NAD+) in order to drive the endergonic reduction of a lower-potential electron acceptor (such as Fdox) by the other electron (Figure 14.2). For example, Figure 14.2 depicts a bifurcation reaction in which 2 NADH(2×2 e−, E0′=−0.32 V) are oxidized to reduce one low-potential Fdox (2×1 e−, E0′=−0.42 V) and one crotonyl-CoA (2×1 e−, E0′=−0.01 V). Some electron-bifurcating systems are even reversible, resulting in electron confurcation (see Sections 14.12 and 14.17).
Figure 14.2 The reaction scheme for flavin-based electron bifurcation.

Many obligate anaerobes require reducing power in the form of reduced ferredoxin (Fdred) or flavodoxin (Fldred) to perform difficult reactions (such as CO2 fixation), but lack electron donors sufficiently electronegative to reduce ferredoxin. In flavin-based electron bifurcation, two electrons from an electron donor (such as H2) are transferred to a flavin (FAD), and one electron is used to reduce a favorable electron acceptor (such as NAD+), making it possible to drive the second electron to an unfavorable electron acceptor (such as Fdox). The reaction can be reversed yielding electron confurcation, for example by organisms that need to achieve redox balance by driving the endergonic formation of H2 from NADH.
Flavin-based electron-bifurcating enzymes have two important functions. (1) They allow the cell to make a highly electronegative intermediate (such as Fdred) that can drive difficult endergonic reductions and can also be used to conserve energy through pmf formation (see Sections 14.12, 14.15, 14.16 and 14.17, and 14.19). (2) They can increase fermentative energy yields by allowing the cell to oxidize NADH to NAD+ through H2 production from Fdred (see Sections 14.17, 14.19). Many obligate anaerobes use flavin-based electron bifurcation reactions, particularly those having very low energy yields such as fermenters, sulfate reducers, acetogens, and methanogens. We will learn about these processes later in this chapter.
Assimilative and Dissimilative Processes
While chemoorganotrophs often get nutrients from organic materials, many chemotrophs and phototrophs can assimilate nutrients by reducing inorganic molecules such as N2, NO3 −, SO4 2−, and CO2. Assimilative processes are those processes used to assimilate inorganic nutrients into cell material. By contrast, dissimilative processes are those processes used to conserve energy.
Assimilative and dissimilative processes differ markedly. In assimilative metabolism, energy is consumed as nutrients are assimilated into cellular material, and so assimilatory reactions are performed only to acquire those nutrients needed to satisfy the biosynthetic needs of growing cells. Since assimilatory reactions contribute to biosynthesis, they require energy in the form of ATP and reducing power. By contrast, in dissimilative metabolism, energy is conserved, and this means that a large amount of the electron acceptor must be reduced and then excreted from the cell. Most microbes can perform a variety of assimilative reduction reactions, whereas dissimilative reduction reactions are the result of anaerobic respiration.
The most important assimilative process in the biosphere is CO2 fixation performed by autotrophic organisms. A diversity of pathways for fixing CO2 into cellular material exist, as we will see next, and by exploring how these pathways differ we will shed light on the evolutionary forces that have produced the vast metabolic diversity within the microbial world.
Check Your Understanding
In a coupled reaction, how can you tell the electron donor half reaction from the electron acceptor half reaction?
How does aerobic respiration differ from anaerobic respiration, and why does aerobic respiration repress anaerobic respiration?
Describe the major differences between assimilative and dissimilative processes.
14.2 Autotrophic Pathways
An autotroph is an organism that can assimilate CO2 into cell material. Many microbes are autotrophic, including virtually all phototrophs and chemolithotrophs. In all autotrophs, CO2 fixation supplies carbon for the biosynthesis of cellular materials, and in phototrophs, CO2 is also the ultimate electron acceptor for photosynthesis.
The evolutionary origin of CO2 fixation likely occurred very soon after the origin of life when Earth was still anoxic. As a result, most CO2 fixation pathways contain enzymes inhibited by O2. Pathways of CO2 fixation often share some enzymes with catabolic pathways, revealing the modularity of enzyme systems. Indeed, the presence of shared metabolites between pathways for catabolism and CO2 fixation create opportunities for some organisms to grow as mixotrophs, meaning that rather than growing as either a heterotroph or an autotroph, they exist on a continuum between heterotrophy and autotrophy depending on the opportunities available to them in their environments. We will also see, as is the case with the Calvin cycle, that enzymes that provide a strong evolutionary benefit are often shared between microorganisms by horizontal gene transfer.
The Calvin Cycle
The Calvin cycle (Section 3.12) is the most widespread and globally important pathway for CO2 fixation. The Calvin cycle is used by all oxygenic phototrophs (including cyanobacteria, algae, and plants), most purple bacteria, and most aerobic chemolithotrophic bacteria (Section 3.11). This wide distribution indicates that many microbes acquired Calvin cycle genes from horizontal gene transfer. The key enzyme of the Calvin cycle is the enzyme RuBisCO, which reduces CO2 to the level of glyceraldehyde 3-phosphate (Section 3.12). The Calvin cycle requires 12 NAD(P)H and 18 ATP to synthesize one molecule of fructose 6-phosphate from 6 CO2 (Figure 3.27).
Many bacteria that perform the Calvin cycle have carboxysomes (Figure 14.3), which are proteinaceous microcompartments and are the site of RuBisCO activity. RuBisCO has very low affinity for CO2 and is inhibited by O2. This inhibition occurs because O2 competes with CO2 for access to the RuBisCO active site, resulting in a waste of reducing power and decreasing autotrophic efficiency. Carboxysomes improve the efficiency of RuBisCO dramatically because they concentrate CO2 and exclude O2 at the site of RuBisCO activity. Even with carboxysomes, RuBisCO is a relatively inefficient and sluggish enzyme and so cells performing the Calvin cycle must contain very large amounts of RuBisCO protein. As a result, RuBisCO is thought to be the single most abundant protein on the Earth.
Figure 14.3 Crystalline Calvin cycle enzymes: Carboxysomes.

Electron micrograph of carboxysomes purified from the chemolithotrophic sulfur oxidizer Halothiobacillus neapolitanus. The structures are about 100 nm in diameter. Carboxysomes are present in a wide variety of obligately autotrophic aerobic Bacteria.
The evolutionary origins of the Calvin cycle remain unclear, but the low affinity for CO2 and high sensitivity to O2 of its key enzyme, RuBisCO, indicates that the cycle originated prior to the Great Oxidation Event (Section 13.2). Carboxysomes were unnecessary at that time because the Earth’s atmosphere had much higher levels of CO2 and lower levels of O2 than it has today. Indeed, carboxysomes are completely absent from chloroplasts and this infers that the cyanobacterial ancestor of chloroplasts possessed the Calvin cycle but lacked carboxysomes. Molecular sequence analysis indicates that carboxysomes originated in nonphototrophic bacteria and were acquired by cyanobacteria during horizontal gene transfer events well after the origin of chloroplasts. There is intense interest in discovering a way to bioengineer carboxysomes into the chloroplasts of plants because such a modification would increase photosynthetic efficiency and plant growth.
Although the Calvin cycle is the most widespread and important pathway of CO2 fixation in the biosphere, many autotrophic Bacteria and Archaea employ alternative pathways for fixing CO2.
The Reverse Citric Acid Cycle
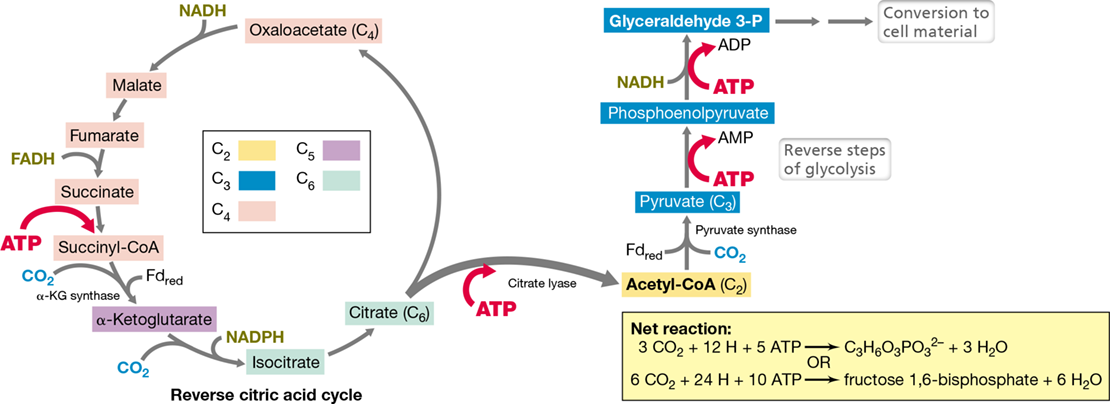
Not all phototrophic organisms rely on the Calvin cycle for CO2 fixation. The reverse citric acid cycle (also called the reductive tricarboxylic acid cycle or rTCA cycle) is a pathway of CO2 fixation used by green sulfur bacteria such as Chlorobium (Section 15.6). The rTCA cycle is also used by many anaerobic and microaerophilic chemolithotrophic Bacteria. In the rTCA cycle, CO2 is reduced by a reversal of steps in the citric acid cycle (Section 3.6; Figure 14.4). The rTCA cycle is more energy efficient than the Calvin cycle, requiring 24 H (that come from 4 NADH, 2 NADPH, 2 FADH, and 4 Fdred) but only 10 ATP to fix 6 CO2 into one molecule of fructose 1,6-bisphosphate. [Note that in the cycle (Figure 14.4), although only four ATP are shown, one ATP is converted into AMP instead of ADP and hence 5 ATP equivalents are consumed per glyceraldehyde 3-P formed. Two molecules of glyceraldehyde 3-P can be converted into fructose 1,6-bisphosphate by reversal of the aldolase step of glycolysis (Figure 3.11).]
Figure 14.4 Reverse citric acid cycle.

The reverse citric acid cycle is the mechanism of CO2 fixation in green sulfur bacteria and many microaerophilic and anaerobic chemolithotrophs. Fdred indicates carboxylation reactions requiring reduced ferredoxin. Starting from oxaloacetate, each turn of the cycle results in three molecules of CO2 being incorporated and pyruvate as the product. Pyruvate is a key metabolite that can feed directly into the glycolytic pathway, generating sugars and other important biosynthetic intermediates.
As the name implies, most of the reactions of the reverse citric acid cycle are catalyzed by reverse reactions of enzymes of the citric acid cycle. However, the cycle also requires the activity of several unique enzymes. These include in particular the enzymes α-ketoglutarate synthase and pyruvate synthase, which catalyze the reductive fixation of CO2 using electrons supplied by Fdred. In green sulfur bacteria, Fdred is produced in the light reactions of photosynthesis (see Figure 14.16), whereas chemolithotrophic bacteria must produce it from reverse electron transport or flavin-based electron bifurcation (Figure 14.2). The two ferredoxin-linked reductions are (1) the carboxylation of succinyl-CoA to α-ketoglutarate, and (2) the carboxylation of acetyl-CoA to pyruvate (Figure 14.4). The rTCA cycle also replaces the enzyme citrate synthase from the citric acid cycle (Figure 3.12) with the enzyme citrate lyase (an ATP-dependent enzyme that cleaves citrate into acetyl-CoA and oxaloacetate), and the enzyme succinate dehydrogenase from the citric acid cycle by the FADH-linked fumarate reductase (an enzyme that forms succinate from fumarate) in the reverse cycle (Figure 14.4).
Other Pathways of CO2 Fixation
In addition to the Calvin cycle and the rTCA cycle, at least four other pathways of CO2 fixation are known, and we consider them here only briefly. Green nonsulfur bacteria such as Chloroflexus (Section 15.7) grow autotrophically with either H2 or H2S as electron donor. However, neither the Calvin cycle nor the reverse citric acid cycle operates in this organism. Instead, two molecules of CO2 are reduced to glyoxylate by the 3-hydroxypropionate bi-cycle. This cycle is so named because hydroxypropionate, a three-carbon compound, is a key intermediate and because it couples two cycles to ultimately yield pyruvate, which can then be converted into glyceraldehyde 3-P as in the rTCA cycle (Figure 14.4).
Archaea appear to lack the Calvin and rTCA cycles as well as the 3-hydroxypropionate bi-cycle. Instead, many chemolithotrophic Archaea use either the 3-hydroxypropionate/4-hydroxybutyrate cycle or the dicarboxylate/4-hydroxybutyrate cycle. Each of these autotrophic cycles includes two connected pathways in which bicarbonate and/or CO2 is reduced to acetyl-CoA, and they are named for their key intermediates, 3-hydroxypropionate, 4-hydroxybutyrate, and C4 dicarboxylic acids.
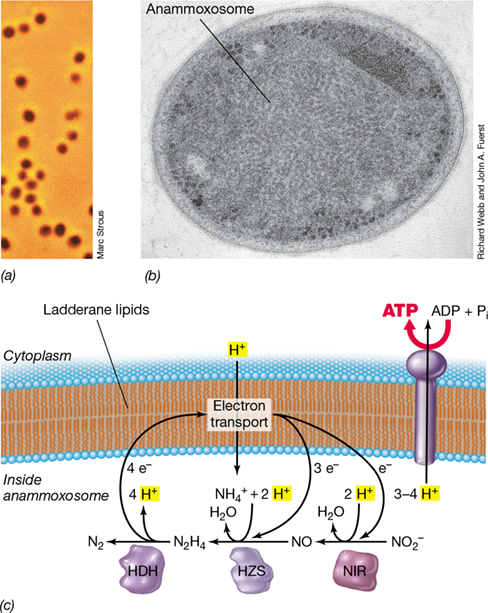
The final pathway of CO2 fixation is the reductive acetyl-CoA pathway (see Figure 14.34). This pathway is present in some obligately anaerobic species of Bacteria and Archaea, and because it is limited to anaerobes and found in both domains, it is thought to be of ancient origin. The reductive acetyl-CoA pathway is found in methanogens, many acetogens, and Planctomyces that carry out the anammox reaction (metabolisms to be described later in this chapter). The reductive acetyl-CoA pathway is the most efficient of all CO2 fixation pathways and requires only about 1 ATP, 3 Fdred, and 2 H2 per 3 molecules of CO2 fixed. It is also the only CO2 fixation pathway coupled directly to energy conservation. The reductive acetyl-CoA pathway was likely one of the earliest mechanisms of CO2 fixation to appear on Earth and we will consider it in detail in Section 14.14.
We now pick up where we left off in Chapter 3 and explore the process of photosynthesis and the amazing diversity it supports.
Check Your Understanding
What is the purpose of carboxysomes in cyanobacteria, and why are they not present in the chloroplasts of plants and algae?
Make a hypothesis to explain why the rTCA cycle is present in diverse unrelated species of bacteria.
What evidence supports the hypothesis that the reductive acetyl-CoA cycle was one of the first to appear on Earth?
II Phototrophy
Phototrophs, which can be anoxygenic or oxygenic, use pigments to transfer light energy to photosynthetic reaction centers. Here they excite an electron to become a strong electron donor for photophosphorylation.
Phototrophy—the use of light energy—is widespread in the microbial world. Phototrophic organisms appeared early in the history of life, they are tremendously diverse, and they have transformed the biosphere. Ultimately, the origin of cyanobacteria billions of years ago converted Earth from an anoxic to an oxic world and set the stage for an explosion of eukaryotic microbial diversity that eventually gave rise to plants and animals (Sections 1.5 and 13.2). In this section, we examine the properties and energy-conserving strategies of phototrophic microorganisms.
14.3 Photosynthesis and Chlorophylls
The most important biological process on Earth is photosynthesis, the use of light energy to drive biosynthesis. Phototrophs are organisms that convert light energy into chemical energy. Photosynthetic organisms are phototrophs that are also autotrophs. Hence, photosynthetic organisms harness light energy to drive the reduction of CO2 to organic compounds; this lifestyle is called photoautotrophy. It is important to note however that not all phototrophs are autotrophic. Some phototrophs use organic carbon as their carbon source; this lifestyle is called photoheterotrophy.
Phototrophy originated within the Bacteria, and a wide diversity of bacterial species can harvest energy from light. No less than seven different phototrophic systems have evolved within Bacteria and these systems are found in diverse phyla. These include the Acidobacteria (genus Chloracidobacterium), Chlorobi (green sulfur bacteria), Chloroflexi (green nonsulfur bacteria, also known as filamentous anoxygenic phototrophs), Cyanobacteria, Firmicutes (heliobacteria), Gemmatimonadetes (genus Gemmatimonas), and Proteobacteria (purple bacteria). Ultimately, phototrophy also evolved within the Eukarya as a result of the endosymbiotic origin of chloroplasts from cyanobacterial relatives (Section 13.4). These phototrophic systems all differ in characteristic ways, but they all reveal similar underlying principles that we will review in the sections that follow.
Photosynthesis supporting autotrophic growth is comprised of two distinct sets of reactions that operate in parallel: the light reactions and the light-independent reactions. The term “light reactions” is used to describe those reactions that convert light energy into chemical energy in the form of the proton motive force and ATP. By contrast, the light-independent reactions are those of CO2 fixation (Section 14.2). CO2 fixation requires both ATP and reducing power (such as NAD(P)H or Fdred); hence in addition to light, photosynthetic organisms need a source of electrons to fix CO2 into cellular carbon.
Water (H2O) is the electron donor for photosynthesis in cyanobacteria, algae, and green plants. When electrons are removed from H2O, molecular oxygen (O2) is produced as a waste product. Consequently, the term oxygenic photosynthesis describes phototrophic organisms that consume H2O and produce O2 as a waste product (Figure 14.5). However, phototrophic organisms can use many different electron donors other than H2O and these organisms do not produce O2 as a waste product. For example, in green and purple sulfur bacteria the donor could be a reduced sulfur compound such as hydrogen sulfide (H2S), or even molecular hydrogen (H2). The term anoxygenic photosynthesis describes photosynthetic organisms that do not produce O2 (Figure 14.5).
Figure 14.5 Patterns of photosynthesis and examples of each.
Energy and reducing power synthesis in (a) anoxygenic and (b) oxygenic phototrophs. Note that oxygenic phototrophs produce O2, while anoxygenic phototrophs do not. Photos: Left, photomicrograph of cells of a typical anoxygenic phototroph, the purple sulfur bacterium Chromatium; cells are about 5 μm in diameter. Note the sulfur globules inside the cells produced from the oxidation of H2S. Right, photomicrograph of cells of the cyanobacterium Halothece; cells are about 5 μm in diameter.
Photosynthesis requires light-sensitive pigments, the chlorophylls—present in plants, algae, and the cyanobacteria—and bacteriochlorophylls, present in anoxygenic phototrophs. Absorption of light energy by chlorophylls and bacteriochlorophylls begins the process of photosynthetic energy conversion, and the net result is chemical energy, ATP.
Chlorophyll and Bacteriochlorophyll
Chlorophyll and bacteriochlorophyll are tetrapyrroles that are related to the parent structure of the cytochromes. But unlike cytochromes, chlorophylls contain magnesium instead of iron at the center of the ring. Chlorophylls also contain specific substituents on the tetrapyrrole ring and a hydrophobic alcohol that helps anchor the chlorophyll into photosynthetic membranes. The structure of chlorophyll a, the principal chlorophyll of oxygenic phototrophs, is shown in **Figure 14.6*a***. Chlorophyll a is green because it absorbs red and blue light and transmits green light; its absorption spectrum shows strong absorbance near 680 nm and 430 nm (Figure 14.6b). Several structurally distinct chlorophylls are known, each distinguished by its unique absorption spectrum. Cyanobacteria contain chlorophyll a (a few species contain chlorophyll d in place of a), while their relatives the prochlorophytes contain chlorophylls a and b.
Figure 14.6 Structures and spectra of chlorophyll *a* and bacteriochlorophyll *a*.

(a) The two molecules are identical except for those portions contrasted in yellow and green. (b) Absorption spectrum (green curve) of cells of the green alga Chlamydomonas. The peaks at 680 and 430 nm are due to chlorophyll a, and the peak at 480 nm is due to carotenoids. Absorption spectrum (red curve) of cells of the phototrophic purple bacterium Rhodopseudomonas palustris. Peaks at 870, 805, 590, and 360 nm are due to bacteriochlorophyll a, and peaks at 525 and 475 nm are due to carotenoids.
Anoxygenic phototrophs produce one or more bacteriochlorophylls. Bacteriochlorophyll a (Figure 14.6 and Figure 14.7), present in most purple bacteria (Sections 15.4 and 15.5), absorbs maximally between 800 and 925 nm. This range of wavelengths results from the fact that different species of purple bacteria synthesize photocomplexes of slightly different protein structure, and this affects the absorption maxima of bacteriochlorophyll a. Other bacteriochlorophylls, whose distribution runs along phylogenetic lines, absorb in other regions of the visible and near-infrared spectrum (Figure 14.7). This diversity in photosynthetic pigments allows phototrophs, collectively, to absorb energy across a wide range of the electromagnetic spectrum. By employing different pigments with distinct absorption properties, different phototrophs can coexist in the same habitat, each absorbing wavelengths of light that others cannot. Thus, pigment diversity has ecological significance for the successful coexistence of different phototrophs in the same habitat.
Figure 14.7 Structure of all known bacteriochlorophylls (Bchl).

The different substituents present in the positions R1 to R7 in the structure at the right are listed. In vivo absorption maxima are the physiologically relevant absorption peaks. The spectrum of bacteriochlorophylls extracted from cells and dissolved in organic solvents is often quite different.
Reaction Centers and Antenna Pigments
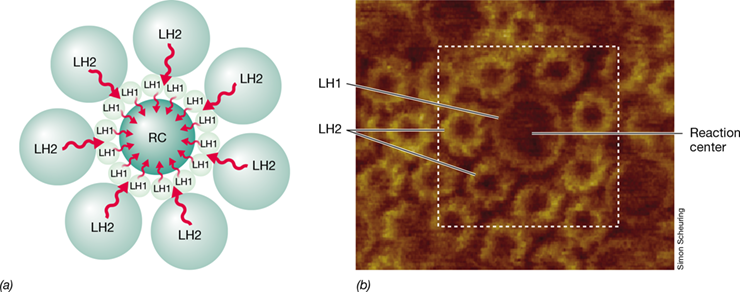
In oxygenic phototrophs and in purple anoxygenic phototrophs, chlorophyll/bacteriochlorophyll molecules do not exist freely in the cell but are attached to proteins and housed within membranes to form photocomplexes consisting of anywhere from 50 to 300 chlorophyll/bacteriochlorophyll molecules. A small number of these pigment molecules are present within photosynthetic reaction centers (Figure 14.8), the complex macromolecular structures that participate directly in the reactions that lead to energy conservation. Photosynthetic reaction centers are surrounded by larger numbers of light-harvesting chlorophylls/bacteriochlorophylls (shown as LH1 and LH2 in Figure 14.8). These so-called antenna pigments function to absorb light and funnel some of the energy to the reaction center (Figure 14.8). At the low light intensities that are often found in nature, this arrangement for concentrating energy allows reaction centers to receive light energy that would otherwise be missed.
Figure 14.8 Arrangement of light-harvesting chlorophylls/bacteriochlorophylls and reaction centers within a photosynthetic membrane.

(a) Light energy absorbed by light-harvesting (LH) molecules (light green) is transferred to the reaction centers (RC) where photosynthetic electron transport reactions begin. Pigment molecules are secured within the membrane by specific pigment-binding proteins. (b) Atomic force micrograph of photocomplexes of the purple bacterium Phaeospirillum molischianum. This organism has two types of light-harvesting complexes, LH1 and LH2. LH2 complexes transfer energy to LH1 complexes, and these transfer energy to the reaction center (see Figure 14.14b).
Photosynthetic Membranes, Chloroplasts, and Chlorosomes
The chlorophyll pigments and all the other components of the light-gathering apparatus exist within membranes in the cell. The location of these photosynthetic membranes differs between prokaryotic and eukaryotic phototrophs. In eukaryotic phototrophs, photosynthesis takes place in intracellular organelles, the chloroplasts, which contain sheetlike photosynthetic membrane systems called thylakoids (Figure 14.9); stacks of thylakoids within the chloroplast form grana. The thylakoids are arranged so that the chloroplast is divided into two regions, the matrix space that surrounds the thylakoids, called the stroma, and the inner space within the thylakoid array, called the lumen. This arrangement makes possible the generation of a light-driven proton motive force used to synthesize ATP (see Section 14.5).
Figure 14.9 The chloroplast.
Details of chloroplast structure, showing how the convolutions of the thylakoid membranes define an inner space called the stroma and form membrane stacks called grana. Inset: Photomicrograph of cells of the green alga Makinoella. Each of the four cells in a cluster contains several chloroplasts.
Chloroplasts are absent from prokaryotic phototrophs. In purple bacteria, the photosynthetic pigments are integrated into internal membrane systems that arise from invagination of the cytoplasmic membrane. Membrane vesicles called chromatophores or membrane stacks called lamellae are common membrane arrangements in purple bacteria (Figure 14.10). In cyanobacteria, photosynthetic pigments reside in lamellar membranes (Figure 14.10b) that are also called thylakoids because of their resemblance to the thylakoids in the chloroplasts of algae (Figure 14.9).
Figure 14.10 Membranes in anoxygenic phototrophs.

(a) Chromatophores. Transmission electron micrograph of a section through a cell of the purple bacterium Rhodobacter showing vesicular photosynthetic membranes. The vesicles are continuous with and arise by invagination of the cytoplasmic membrane. A cell is about 1 μm wide. (b) Lamellar membranes in the purple bacterium Ectothiorhodospira. A cell is about 1.5 μm wide. These membranes are also continuous with and arise from invagination of the cytoplasmic membrane, but instead of forming vesicles, they form membrane stacks.
The ultimate structure for capturing energy from low light intensities is the chlorosome (Figure 14.11). Chlorosomes are present in the green sulfur bacteria (Chlorobium, Section 15.6), green nonsulfur bacteria (Chloroflexus, Section 15.7), and phototrophic Acidobacteria (Chloracidobacterium, Section 15.8), all of which are anoxygenic phototrophs. Chlorosomes function as giant antenna systems, but unlike the antennae of purple bacteria or cyanobacteria, bacteriochlorophyll molecules in the chlorosome are not attached to proteins. Chlorosomes contain bacteriochlorophyll c, d, or e (Figure 14.7) arranged in dense arrays running along the long axis of the structure. Light energy absorbed by these antenna pigments is transferred to bacteriochlorophyll a in the reaction center in the cytoplasmic membrane through a small protein called the FMO protein (Figure 14.11).
Figure 14.11 The chlorosome of green sulfur and green nonsulfur bacteria.

(a) Transmission electron micrograph of a cross section of a cell of the green sulfur bacterium Chlorobaculum tepidum. Note the chlorosomes (arrows). (b) Model of chlorosome structure. The chlorosome (green) lies appressed to the inside surface of the cytoplasmic membrane. Antenna bacteriochlorophyll (Bchl) molecules are arranged in tubelike arrays inside the chlorosome, and energy is transferred from these to reaction center (RC) Bchl a in the cytoplasmic membrane through a protein called FMO. Base plate (BP) proteins function as connectors between the chlorosome and the cytoplasmic membrane.
Chlorosomes allow these phototrophs to grow at extremely low light intensities, and hence these phototrophs are typically found in the deepest waters of lakes, inland seas, and other anoxic aquatic habitats where light levels are too low to support other phototrophs. Green nonsulfur bacteria and Chloracidobacterium are major components of microbial mats, thick biofilms that form in hot springs and highly saline environments (Section 20.5). Microbial mats experience a steep light gradient, with light levels even a few millimeters into the mat approaching darkness. Green sulfur bacteria are common near the sediments of sulfidic aquatic systems and can even carry out photosynthesis in the darkness of the deep ocean by using infrared radiation emitted from hydrothermal vents. Hence, chlorosomes allow phototrophs that produce them to carry out photosynthesis with only the minimal light intensities available.
Check Your Understanding
What is the fundamental difference between an oxygenic and an anoxygenic phototroph?
What is the purpose of chlorophyll and bacteriochlorophyll molecules? In what ways do they resemble cytochromes and in what ways do they differ?
Why can phototrophic green bacteria grow at light intensities that will not support purple bacteria or cyanobacteria?
14.4 Carotenoids and Phycobilins
Although chlorophyll/bacteriochlorophyll is required for photosynthesis, phototrophic organisms contain other pigments as well. These pigments include, in particular, the carotenoids and phycobilins.
Carotenoids
The most widespread accessory pigments in phototrophs are the carotenoids. Carotenoids are hydrophobic pigments that are firmly embedded in the photosynthetic membrane. Figure 14.12 shows the structure of a common carotenoid, β-carotene. Carotenoids are typically yellow, red, brown, or green and absorb light in the blue region of the spectrum. Some of the major carotenoids of anoxygenic phototrophs are shown in Figure 14.13. Because they tend to mask the color of bacteriochlorophylls, carotenoids are responsible for the brilliant colors of red, purple, pink, green, yellow, or brown that are observed in different species of anoxygenic phototrophs (Figure 15.12).
Figure 14.12 Structure of β-carotene, a typical carotenoid.

The conjugated double-bond system is highlighted in orange.
Figure 14.13 Structures of some common carotenoids found in anoxygenic phototrophs.

Carotenes are hydrocarbon carotenoids, and xanthophylls are oxygenated carotenoids. Compare the structure of β-carotene shown in Figure 14.12 with how it is drawn here. For simplicity in the structures shown here, methyl (CH3) groups are designated by bond only.
Carotenoids are closely associated with chlorophyll or bacteriochlorophyll in photosynthetic complexes, and some of the energy absorbed by carotenoids can be transferred to the reaction center. However, carotenoids function primarily as photoprotective agents. Bright light can be harmful to cells because it can catalyze photooxidation reactions that can produce toxic forms of oxygen, such as singlet oxygen (1O2). Like superoxide and other forms of toxic oxygen (Section 4.16), singlet oxygen can spontaneously oxidize photocomplexes, rendering them nonfunctional. Carotenoids quench toxic oxygen species by absorbing much of this harmful light and in this way prevent these dangerous photooxidations. Because phototrophic organisms by their very nature must live in the light, the photoprotection conferred by carotenoids is clearly advantageous.
Phycobiliproteins and Phycobilisomes
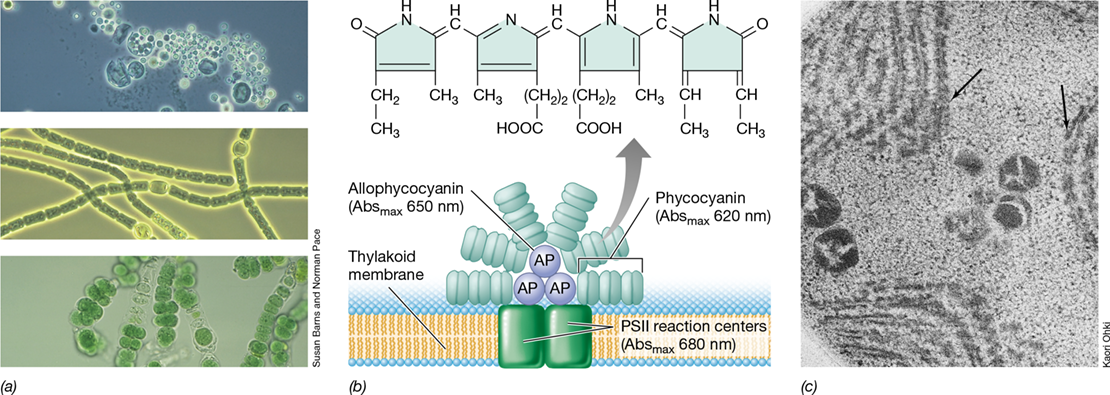
Cyanobacteria and the chloroplasts of red algae (which are descendants of cyanobacteria, Section 18.1) contain pigments called phycobiliproteins, which are the main light-harvesting systems of these phototrophs. Phycobiliproteins consist of red or blue-green linear tetrapyrroles, called bilins, bound to proteins, and give cyanobacteria and red algae their characteristic colors (Figure 14.14). The red phycobiliprotein, called phycoerythrin, absorbs most strongly at wavelengths around 550 nm, whereas the blue phycobiliprotein, phycocyanin (Figure 14.14b), absorbs most strongly at 620 nm. A third phycobiliprotein, called allophycocyanin (Figure 14.14b) absorbs at about 650 nm.
Figure 14.14 Phycobiliproteins and phycobilisomes.

(a) Light photomicrographs of cells of the cyanobacteria (top to bottom) Dermocarpa, Anabaena, and Fischerella, showing the typical blue-green color of cells due to phycobiliproteins. (b) Structure of phycocyanin (top) and a phycobilisome. Phycocyanin absorbs at higher energies (shorter wavelengths) than allophycocyanin. Chlorophyll a absorbs at longer wavelengths (lower energies) than allophycocyanin. Energy flow is thus phycocyanin→allophycocyanin→chlorophyll a of PSII (see Figure 14.16). (c) Electron micrograph of a thin section of the cyanobacterium Synechocystis. Note the darkly staining phycobilisomes (arrows) attached to the lamellar membranes.
Phycobiliproteins assemble into aggregates called phycobilisomes that attach to cyanobacterial thylakoids (Figure 14.14c). Phycobilisomes are arranged such that the allophycocyanin molecules are in direct contact with the photosynthetic membrane. Allophycocyanin is surrounded by phycocyanin or phycoerythrin (or both, depending on the organism). Phycocyanin and phycoerythrin absorb light of shorter wavelengths (higher energy) and transfer some energy to allophycocyanin, which is positioned closest to the reaction center chlorophyll and transfers energy to it (Figure 14.14b). Thus, in a fashion similar to how antenna bacteriochlorophyll systems function in anoxygenic phototrophs (Figure 14.8), energy transfer proceeds “downhill” from phycobilisomes to the reaction center. Phycobilisomes thus facilitate energy transfer to cyanobacterial reaction centers, allowing cyanobacteria to grow at lower light intensities than would otherwise be possible.
Check Your Understanding
In which phototrophs are carotenoids found? Phycobiliproteins?
How does the structure of a phycobilin compare with that of a chlorophyll?
Phycocyanin is blue-green. What color of light does it absorb?
14.5 Anoxygenic Photosynthesis
In the photosynthetic light reactions, electrons travel through an electron transport chain whose components are arranged in a photosynthetic membrane in order of their increasingly more electropositive reduction potential (E0′), similar to the electron transport chains used in respiration (Sections 3.9, 3.10 and 3.11). This electron transport generates a proton motive force that drives ATP synthesis (Section 3.11). Key parts of this process include photosynthetic reaction centers and photosynthetic membranes (Section 14.3).
Photosynthetic reaction centers are complex macromolecular structures localized within photosynthetic membranes. They are composed of multiple protein subunits and cofactors (including chlorophylls or bacteriochlorophylls), and they interact with both antenna pigments and components of the electron transport chain. Photosynthetic antenna pigments funnel light energy to the reaction center to excite a pair of chlorophylls or bacteriochlorophylls (referred to as the special pair), thereby generating highly electronegative electrons that can be donated to a subsequent electron transport chain. Several different types of reaction centers have been described in anoxygenic phototrophs, but all fall into one of two classes; they are either of a quinone type (Q-type) or iron–sulfur type (FeS-type) depending on the electron acceptor in the reaction center.
Photosynthetic Electron Flow in Purple Bacteria
Purple bacteria use a Q-type reaction center that contains three polypeptides, designated L, M, and H. These proteins along with a molecule of cytochrome c are firmly embedded in the photosynthetic membrane (Figure 14.10) and wind through the membrane several times (Figure 14.15). The L, M, and H polypeptides bind four molecules of bacteriochlorophyll a, two of which are the special pair and another two that transfer electrons within the reaction center, two molecules of bacteriopheophytin a (bacteriochlorophyll a minus its magnesium atom), two molecules of quinone (Section 3.8), and one carotenoid molecule (Figure 14.15).
Figure 14.15 Structure of the reaction center of a purple phototrophic bacterium.

(a) Arrangement of pigment molecules in the reaction center. The “special pair” of bacteriochlorophyll molecules (orange) overlap and occur at the top of the reaction center structural diagram; adjacent to and below the special pair are a pair of accessory bacteriochlorophylls (yellow near the top). Bacteriopheophytin molecules (blue) are arranged below the bacteriochlorophylls, and quinones (yellow at the bottom) are present at the bottom of the structural model. Compare the structure of these molecules in the reaction center to their role in electron transfer (see Figure 14.16). (b) Molecular model of the protein structure of the reaction center. The molecular structures (C in gray, O in red, N in blue) of pigments described in part a are depicted in context with ribbon diagrams of reaction center protein subunits: H (red), M (white), and L (blue). The reaction center pigment–protein complex is integrated into the lipid bilayer.
Photosynthetic light reactions begin when light energy absorbed by antenna pigments is transferred to the special pair of bacteriochlorophyll a molecules (Figure 14.15a, Figure 3.25). This excites the special pair, converting them from a relatively weak to a very strong electron donor (very electronegative E0′, Sections 3.3 and 3.11). Once this strong donor has been produced, the remaining steps in photosynthetic electron flow are highly reminiscent of those we have seen before in respiration (Sections 3.9 and 3.10); that is, electrons flow through a membrane from carriers of low E0′ to those of high E0′ and in the process generate a proton motive force that is used to synthesize ATP (Section 3.11).
Before excitation, the purple bacterial reaction center, which is called P870, has an E0′ of about +0.5 V; after excitation, it has a potential of about −1.0 V (Figure 14.16). An excited electron within P870 proceeds to reduce a molecule of bacteriochlorophyll a within the reaction center (Figures 14.15 and 14.16). This transition takes place incredibly fast, taking only about three-trillionths (3×10−12) of a second. Once reduced, bacteriochlorophyll a proceeds to reduce bacteriopheophytin a, and the latter reduces quinone molecules within the membrane (Figure 14.16). These transitions are also very fast, taking less than one-billionth of a second. From the quinone, electrons are transported through the membrane more slowly (on a millisecond scale) through a series of iron–sulfur proteins and cytochromes used to generate the proton motive force (Figure 14.16), eventually returning to the reaction center. This overall process is referred to as cyclic photophosphorylation because the electrons cycle in a closed loop, thereby generating the proton motive force needed to synthesize ATP (Section 3.11).
Figure 14.16 A comparison of electron flow in anoxygenic and oxygenic phototrophs.

Purple bacteria and green sulfur bacteria have Q-type and FeS-type reaction centers (RC), respectively, while oxygenic phototrophs have both types. The FeS-type reaction center of green sulfur bacteria produces highly electronegative electrons that can reduce ferredoxin (Fd), allowing for CO2 fixation through the reverse citric acid cycle (Figure 14.4). Q-type reaction centers cannot reduce Fd, which makes reverse electron transport necessary to generate the NADH needed to fix CO2 by the Calvin cycle (Figures 3.26 and 3.27). Cyclic electron flow occurs in purple bacteria but not in green sulfur bacteria. Electron flow in oxygenic phototrophs is noncyclic, resulting in the characteristic “Z-scheme” pattern of electron flow. If photosystem II is blocked, oxygenic phototrophs can perform cyclic photosynthesis at photosystem I (dashed line in the cyanobacteria panel, see also Figure 14.17). Bchl, bacteriochlorophyll; BPh, bacteriopheophytin; Chl, chlorophyll; Cyt, cytochrome; FeS, iron–sulfur protein; FNR, ferredoxin–NADP oxidoreductase; P870, P840, P680, and P700, reaction centers; PC, plastocyanin; Ph, pheophytin; PQ, plastoquinone; Q, quinone; QA, QB, intermediate quinones; MQ, menaquinone.
Mastering Microbiology
Art Activity: Figure 14.12a Electron flow in anoxygenic photosynthesis
Overview of Electron Flow in Anoxygenic Phototrophs
Thus far, we have focused on electron flow in purple bacteria. Although analogous membrane-associated reactions drive photophosphorylation in other phototrophs, there are significant differences in the details. Q-type reaction centers that contain quinone molecules are found in purple bacteria (Sections 15.4 and 15.5), green nonsulfur bacteria (Section 15.7), and Gemmatimonadetes (genus Gemmatimonas, Section 15.8), but FeS-type reaction centers that lack quinones and contain instead FeS clusters are found in the green sulfur bacteria (Section 15.6), Acidobacteria (genus Chloracidobacterium, Section 15.8), and heliobacteria (Section 15.8). The Q-type and FeS-type reaction centers have important structural differences. For example, whereas Q-type reaction centers contain bacteriochlorophyll, FeS-type reaction centers contain both bacteriochlorophyll and chlorophyll a (green sulfur bacteria) or a modified form of chlorophyll a (hydroxychlorophyll a in heliobacteria). The structural differences between Q-type and FeS-type reaction centers cause major differences in electron flow (Figure 14.16).
The bacteriochlorophyll molecules of FeS-type reaction centers have an excited state that is significantly more electronegative than Q-type reaction centers (Figure 14.16). In purple bacteria, the Q-type reaction center donates electrons directly to a quinone (Figure 14.16). However, these quinones (E0′ about 0 V) are insufficiently electronegative to provide the reducing power (E0′ of −0.32 V for NAD(P)H and −0.42 V for ferredoxin) needed for CO2 fixation and other biosynthetic reactions (Section 14.2). In contrast, FeS-type reaction centers donate electrons directly to low-potential FeS-proteins able to reduce ferredoxin. Green sulfur bacteria then use this Fdred when they fix CO2 using the rTCA cycle (Section 14.2 and Figure 14.4). Electrons from ferredoxin can also pass to ferredoxin–NAD+ oxidoreductase for the production of NAD(P)H needed for biosynthesis.
Phototrophs having a Q-type reaction center perform cyclic photophosphorylation providing cells with the ability to produce ATP as long as light is present, but these organisms still require a source of reducing power in order to perform the biosynthetic reactions required for growth. Since Q-type reaction centers produce electrons that are insufficiently electronegative to reduce NAD(P)+, these phototrophs require reverse electron transport (Section 14.1and Section 3.11) to produce the NAD(P)H they need to carry out CO2 fixation and other biosynthetic reactions.
It remains unclear whether electron transfer in green sulfur bacteria is cyclic or noncyclic. It has been proposed that the FeS-type reaction centers in these phototrophs can transfer electrons directly to menaquinone, thereby generating a proton motive force resulting in cyclic photophosphorylation as seen in purple bacteria (Figure 14.16). However, little evidence for cyclic photophosphorylation has been observed in green sulfur bacteria. Alternatively, these phototrophs may employ noncyclic electron flow whereby electrons from external electron donors, such as H2S, enter at the level of the menaquinone pool. These electrons would be transferred through the reaction center and then to ferredoxin where they would ultimately be channeled into biosynthetic reactions without the need for reverse electron transport (Figure 14.16). In heliobacteria, FeS-type phototrophs that are not autotrophic, cyclic photophosphorylation has also not been demonstrated. However, in this group of strictly photoheterotrophic bacteria, reducing power needs are less extensive than for photoautotrophs and thus it is more likely that cyclic flow occurs to generate sufficient ATP for biosynthesis.
Mastering Microbiology
Art Activity: Figure 14.14 Electron flow in oxygenic photosynthesis, the “Z” scheme
We move on now to consider photosynthesis in oxygenic phototrophs such as the cyanobacteria. These organisms contain both FeS-type and Q-type reaction centers that likely evolved from those of green and purple bacteria, respectively. These two photocomplexes function in parallel to both generate ATP and reducing power, the latter from H2O (Figure 14.16). We explore how this fascinating process occurs now.
Check Your Understanding
What parallels exist in the processes of photophosphorylation and oxidative phosphorylation?
Why is reverse electron transport needed by phototrophs having Q-type reaction centers and not needed by those having FeS-type reaction centers?
What is the difference between cyclic and noncyclic photophosphorylation?
14.6 Oxygenic Photosynthesis
In contrast to photosynthetic electron flow in anoxygenic phototrophs, which have either FeS-type or Q-type photosynthetic reaction centers, oxygenic phototrophs have both types of reaction centers. In oxygenic phototrophs, electrons flow through two distinct photosystems called photosystem I (PSI, or P700), which has an FeS-type reaction center, and photosystem II (PSII, or P680), which has a Q-type reaction center. PSI and PSII interact in the “Z scheme” of photosynthesis, so named because the pathway resembles the letter Z turned on its side (Figure 14.16). As in anoxygenic photosynthesis, the light reactions in oxygenic photosynthesis occur in photocomplexes embedded in specialized photosynthetic membranes. In eukaryotic cells, the membranes are in the chloroplast (Figure 14.9), whereas in cyanobacteria, the membranes are arranged in stacks within the cytoplasm (Figure 14.14c).
Electron Flow and ATP Synthesis in Oxygenic Photosynthesis
PSII performs the first—and most distinctive—step in oxygenic photosynthesis, the splitting of water into oxygen and electrons (Figure 14.17). Upon absorbing light energy, the P680 chlorophyll a molecule in PSII is excited to a very electronegative reduction potential that allows it to donate an electron to pheophytin a (chlorophyll a lacking its magnesium atom), a molecule with an E0′ of about −0.5 V. This creates a charge separation that causes P680 to become so strongly electropositive that it can accept electrons from H2O, an extremely weak electron donor. The oxidation of water by PSII occurs at the water-oxidizing complex and is catalyzed by a Mn4Ca cluster (Figure 14.17), which binds 2 molecules of H2O. P680 removes one electron from the Mn4Ca cluster of the water-oxidizing complex for each photon absorbed. In this way 4 electrons are sequentially removed from the 2 H2O molecules bound to the Mn4Ca cluster, resulting in the production of O2 and 4 H+. Each electron transferred to pheophytin then travels through a series of quinones (QA and QB) within the PSII photocomplex. Finally, two electrons from the PSII photocomplex are used to reduce plastoquinone (PQ) to PQH2, a step that allows for the generation of the proton motive force.
Figure 14.17 Electron transport in oxygenic photosynthesis.

Photosystem II (PSII) is activated by photons, causing H2O to be oxidized on the Mn4Ca cluster of the water-oxidizing complex. Electrons are transferred from PSII to the plastoquinone pool (PQ/PQH2). Protons are exchanged across the membrane when plastoquinone is oxidized by cytochrome b6f. Per two molecules of water oxidized to one O2, a total of 12 protons are released to the lumen to fuel ATP synthase. Electrons are then transferred to plas-tocyanin (PC), which carries them to photosystem I (PSI). Upon activation by light, PSI reduces ferredoxin (Fd), with sequential reduction of ferredoxin:NADP oxidoreductase (FNR), and then NADP+. The ATP and NADPH produced by the light reactions are used in CO2 fixation by the Calvin cycle (Section 3.12 and Section 14.2). Cyclic photophosphorylation occurs when FNR donates electrons to cytochrome b6f instead of to NADP+. During cyclic photophosphorylation, more ATP and less NADPH are produced than during noncyclic photophosphorylation.
The proton motive force is generated in oxygenic photosynthesis by electron transport through quinones and cytochromes of increasingly positive reduction potential. These electron transport reactions are similar to those encountered during our discussion of aerobic respiration (Section 3.9). Electrons from PQH2 are transferred through cytochrome b6f and through a copper-containing protein called plastocyanin before being donated to the PSI reaction center (Figures 14.16 and 14.17). The absorption of light by P700 of PSI allows it to accept electrons donated from plastocyanin. Electrons travel through several intermediates in PSI terminating with the reduction of NADP+ to NADPH (Figures 14.16 and 14.17). The electrons from NADPH are used in CO2 fixation (by the Calvin cycle, Section 3.12 and Section 14.2) and other biosynthetic reactions, thereby regenerating NADP+ and making CO2 the ultimate electron acceptor for oxygenic photosynthesis. Two protons are generated for each water molecule that is split by PSII and four protons are translocated across the membrane for every two electrons transferred through the electron transport chain, resulting in a total of 12 protons translocated for every molecule of O2 produced. This proton motive force is then used by ATP synthase to produce ATP.
Oxygenic photosynthesis results in noncyclic photophosphorylation because electrons do not cycle back to reduce the oxidized P680, but instead are used in the reduction of NADP+. However, when the cell requires less NADPH for biosynthesis, oxygenic phototrophs can perform cyclic photophosphorylation. This occurs when, instead of reducing NADP+, electrons from PSI are returned to the electron transport chain that connects PSII to PSI. When this happens, the recycled electrons can be used to generate a proton motive force that supports additional ATP synthesis (dashed line in the cyanobacteria panel of Figure 14.16; Figure 14.17).
Anoxygenic Photosynthesis in Oxygenic Phototrophs
Photosystems I and II normally function in tandem in oxygenic photosynthesis (Figures 14.16 and 14.17). However, if PSII activity is blocked, some oxygenic phototrophs can perform photosynthesis using only PSI. Under these conditions, cyclic photophosphorylation (Figure 14.16) occurs exclusively and reducing power for CO2 reduction comes from sources other than water. In effect, this is anoxygenic photosynthesis occurring in oxygenic phototrophs.
Many cyanobacteria can use H2S as an electron donor under these conditions, and many green algae can use H2. When H2S is used, it is oxidized to elemental sulfur (S0), and sulfur granules similar to those produced by green sulfur bacteria (Figure 14.5) are deposited outside the cyanobacterial cells. Figure 14.18 shows this in the filamentous cyanobacterium Oscillatoria limnetica. This organism lives in anoxic salt ponds where it oxidizes sulfide and carries out anoxygenic photosynthesis along with green and purple bacteria.
Figure 14.18 Oxidation of H2S by *Oscillatoria limnetica*.

Note the globules of S0 (arrows), the oxidation product of H2S, formed outside the cells. O. limnetica carries out oxygenic photosynthesis, but cells revert to the anoxygenic process in the presence of H2S.
From an evolutionary standpoint, the process of photophosphorylation in both oxygenic and anoxygenic phototrophs is one of many indications of their close relationship. Indeed, the structure of the Q-type reaction centers is homologous with PSII, and the structure of the FeS-type reaction centers is homologous with PSI. Because the evidence is strong that purple and green bacteria preceded cyanobacteria on Earth by perhaps as many as 0.5 billion years (Section 13.2), it is clear that anoxygenic photosynthesis was the first form of photosynthesis on Earth. The key evolutionary event that ultimately led to the evolution of oxygenic photosynthesis in cyanobacteria was to acquire both types of reaction centers by horizontal gene transfer. The ancestors of cyanobacteria were certainly anoxygenic phototrophs, but the presence of redundant reaction centers facilitated their evolutionary diversification, as often results from gene duplication events (Section 13.8). Ultimately, this process of functional diversification and experimentation led to the origin of oxygenic photosynthesis when chance events caused PSII to acquire the ability to use H2O as an electron donor. The evolutionary origin of oxygenic photosynthesis was a seminal event in Earth’s history since it not only oxygenated the planet (Figure 13.1) but also allowed photoautotrophs to tap an inexhaustible supply of electrons.
In Part III we turn our attention to chemotrophic organisms that conserve energy through the respiration of inorganic electron donors.
Check Your Understanding
Differentiate between cyclic and noncyclic electron flow in oxygenic photosynthesis.
What is the key role of light energy in the initial step of the photosynthetic light reactions?
What evidence is there that anoxygenic and oxygenic photosynthesis are related processes?
III: Respiratory Processes Defined by Electron Donor
III: Respiratory Processes Defined by Electron Donor
III Respiratory Processes Defined by Electron Donor
Chemolithotrophs use inorganic electron donors such as H2S, Fe2+, or NH3 in electron transport, generating ATP by oxidative phosphorylation. However, these electropositive donors force these organisms to generate NADH through endergonic reactions.
As we learned in Chapter 3 and reviewed in Section 14.1, energy is conserved in respiration by redox reactions that transfer electrons from an initial electron donor to a final electron acceptor. A tremendous diversity of respirations exist, and the microbes that carry them out are typically characterized by the nature of their electron donors and/or electron acceptors. In this part of the chapter, we focus on diversity from the standpoint of electron donors.
14.7 Oxidation of Sulfur Compounds
Many reduced sulfur compounds can be electron donors for the colorless sulfur bacteria, called colorless to distinguish them from the pigmented green and purple sulfur bacteria discussed earlier in this chapter (Figure 14.5 and Section 14.5). Historically, the concept of chemolithotrophy emerged in the late nineteenth century from studies of the sulfur bacteria by the Russian microbiologist Sergei Winogradsky (Section 1.13) and was a radically new idea at the time. However, as our understanding of metabolic diversity has improved, it has become clear that chemolithotrophy, and in particular sulfur chemolithotrophy, is a major metabolic lifestyle of many Bacteria and Archaea.
Energetics of Sulfur Oxidation
The most common sulfur compounds used as electron donors are hydrogen sulfide (H2S), elemental sulfur (S0), thiosulfate (S2O3 2−), and sulfite (SO3 2−) (Table 14.1, Table 14.2, and see Table 14.4). In most cases, the final oxidation product is sulfate (SO4 2−). Sulfide oxidation occurs in stages, with the first oxidation step yielding elemental sulfur, S0. Some sulfide-oxidizing bacteria, such as Beggiatoa, deposit this elemental sulfur inside the cell (**Figure 14.19*a***), where the sulfur exists as a potential energy (electron) reserve. When the supply of sulfide has been depleted, additional energy can then be conserved from the oxidation of sulfur to sulfate. When S0 is present externally, the organism must attach itself to the sulfur particle because elemental sulfur is rather insoluble (Figure 14.19b).
Table 14.2 Comparison of the energetics of oxidation of some common reduced sulfur compounds

a All reactions are balanced, both atomically and electrically. See Table 3.3 and Section 3.3 for details of calculations. For the reaction and energetics of the oxidation of sulfide to sulfur and sulfur to sulfate, see Table 14.1.
Figure 14.19 Sulfur bacteria.

(a) Internal sulfur granules in Beggiatoa (arrows). (b) Attachment of cells of the sulfur-oxidizing archaeon Sulfolobus acidocaldarius to a crystal of elemental sulfur. Cells were visualized by fluorescence microscopy after being stained with the dye acridine orange, which does not stain sulfur.
One product of the oxidation of reduced sulfur compounds is protons (Tables 14.1 and 14.2). Consequently, one result of sulfur chemolithotrophy is acidification of the environment. Because of this, many sulfur bacteria have evolved to be acid-tolerant or even acidophilic. Acidithiobacillus thiooxidans, for example, grows best at a pH between 2 and 3.
Biochemistry of Sulfur Oxidation: The Sox System
There are diverse pathways for conserving energy from the oxidation of sulfur compounds. One of the best characterized is the Sox (for sulfur oxidation) system (Figure 14.20), which has been detailed in the bacterium Paracoccus pantotrophus and is widespread among sulfur-oxidizing bacteria. The Sox system contains over 15 genes encoding various cytochromes and other proteins necessary for the oxidation of reduced sulfur compounds directly to sulfate. Elements of the Sox system are found in diverse sulfur chemolithotrophs and also in some phototrophic sulfur bacteria, organisms that oxidize sulfide to obtain reducing power for CO2 fixation rather than for energy conservation. The fact that this biochemical system is distributed among bacteria that oxidize sulfide for very different reasons is a good indication that the genes that encode Sox have been transferred between species by horizontal gene flow (Section 13.9 and Chapter 9).
Figure 14.20 Oxidation of reduced sulfur compounds by sulfur chemolithotrophs.

There are several different pathways for conserving energy through the oxidation of sulfide (H2S), thiosulfate (S2O3−), and elemental sulfur (S0). In the Sox system (sulfur oxidation), SoxXA attaches a reduced sulfur compound to the carrier protein SoxYZ. The protein SoxCD, sulfur dehydrogenase, catalyzes removal of 6 e− from the bound sulfur atom, and is a key enzyme for bacteria that use the complete Sox system for sulfur oxidation (such as Paracoccus pantotrophus). Sulfate (SO4 2−) is released by the action of SoxB. In contrast, bacteria that form sulfur granules, such as Beggiatoa (Figure 14.19a), lack SoxCD and instead oxidize sulfur compounds using the enzymes DsrAB, dissimilatory sulfite reductase, and APS reductase (see Section 14.12). In sulfur oxidation, these enzymes are run backwards to oxidize sulfur compounds. In certain sulfur oxidizers, APS reductase is replaced by sulfite reductase. Reactions of the Sox cycle take place in the periplasm and electrons enter the electron transport chain through the activity of a periplasmic c-type cytochrome (Cyt c), while reactions of Sox/Dsr systems take place instead in the cytoplasm and electrons can enter electron transport at either the level of flavoproteins (Fp) or c-type cytochromes.
There are four key proteins in the Sox system: SoxXA, SoxYZ, SoxB, and SoxCD. All of these proteins are present in the periplasm (Section 2.4). The pathway begins when the enzyme SoxXA forms a heterodisulfide bond between the sulfur compound to be oxidized (which can be HS−, S0, or S2O3 2−) and the carrier protein, SoxYZ (Figure 14.20). The sulfur compound remains bound to the carrier throughout the pathway, being ultimately released as sulfate through the activity of SoxB. The enzyme SoxCD (sulfur dehydrogenase) is the key enzyme that mediates the removal of 6 electrons from the sulfur compound bound to the carrier (Figure 14.20). Electrons from the Sox system are funneled into the electron transport chain (see later), while the protons generated in the periplasm are released to and acidify the external environment.
Other Aspects of Chemolithotrophic Sulfur Oxidation
Sulfur-oxidizing microbes that store sulfur granules within the cell (see Figure 14.19a) also use components of the Sox system but lack the key enzyme sulfur dehydrogenase (SoxCD). In the absence of SoxCD, a sulfur atom bound to SoxYZ is added to a growing sulfur granule in the periplasm (Figure 14.20). The sulfur in the granule can be reductively activated and transported to the cytoplasm where it is eventually oxidized to sulfite (SO3 2−) by the reverse activity of DsrAB, an enzyme homologous to the enzyme sulfite reductase found in sulfate-reducing bacteria (Section 14.12). The sulfite is then oxidized to sulfate plus two electrons through one of two different pathways. The most widespread system employs the reverse activity of the cytoplasmic enzyme sulfite reductase. This enzyme oxidizes sulfite and transfers the electrons to the electron transport chain. By contrast, some sulfur chemolithotrophs oxidize SO3 2− to SO4 2− via a reversal of the activity of the enzyme adenosine phosphosulfate (APS) reductase (an enzyme essential for the metabolism of sulfate-reducing bacteria, see Section 14.12). The oxidation of SO3 2− to SO4 2− yields an energy-rich phosphate bond by substrate-level phosphorylation when AMP is converted to ATP (Figure 14.20).
Electrons from the oxidation of reduced sulfur compounds eventually reach the electron transport chain, as shown in Figure 14.20. Though the exact details remain unclear, electrons likely enter at the flavoprotein or cytochrome c (E0′=+0.3 V) levels and are transported through the chain to O2, generating a proton motive force that triggers ATP synthase activity (Figure 14.20). Electrons for CO2 fixation come from reverse electron transport (Section 14.1), eventually yielding NADH, and autotrophy is driven by reactions of the Calvin cycle or some other autotrophic pathway (Section 14.2). Although the sulfur chemolithotrophs are primarily an aerobic group, some species can grow by anaerobic respiration using nitrate as an electron acceptor. The sulfur bacterium Thiobacillus denitrificans is a classic example, reducing nitrate to dinitrogen gas (the process of denitrification, Section 14.11).
We now consider iron oxidation, a process that at acidic pH poses the greatest energy challenge of all chemolithotrophic energy metabolisms.
Check Your Understanding
How many electrons are available from the oxidation of H2S if S0 or SO4 2− is the final product?
In terms of intermediates, how does the Sox system differ from other sulfide-oxidizing systems?
Why are many sulfur- and sulfide-oxidizing bacteria acidophilic?
14.8 Iron (Fe2+) Oxidation
The aerobic oxidation of ferrous iron (Fe2+) to ferric iron (Fe3+) supports growth of the chemolithotrophic “iron bacteria” (Section 15.14 and see MicrobiologyNow on page 424). At acidic pH, only a small amount of energy is available from this reaction (Table 14.1), and for this reason the iron bacteria must oxidize large amounts of iron in order to produce only tiny amounts of cell material. The ferric iron produced becomes hydrated to form insoluble ferric hydroxide (Fe3++3 H2O→Fe(OH)3+3 H+) and other iron precipitates in aquatic environments (Figure 14.21), and this drives down the pH. This inevitable chemical reaction probably explains why many iron-oxidizing bacteria have evolved to be strongly acidophilic.
Figure 14.21 Iron-oxidizing bacteria.
(a) Acid mine drainage, showing the confluence of a normal river and a creek draining a coal-mining area. At low pH values, Fe2+ does not oxidize spontaneously in air, but Acidithiobacillus ferrooxidans carries out the oxidation; insoluble Fe(OH)3 and complex ferric salts precipitate. (b) Cultures of A. ferrooxidans. Shown is a dilution series, with no growth in the tube on the left and increasing amounts of growth from left to right. Growth is evident from the production of Fe(OH)3.
Iron-Oxidizing Bacteria
The best-known iron bacteria, Acidithiobacillus ferrooxidans and Leptospirillum ferrooxidans, can both grow autotrophically using ferrous iron (Figure 14.21) as electron donor at pH values as low as 1; growth is optimal at pH 2–3. These bacteria are common in acid-polluted environments such as coal-mining runoff waters (Figure 14.21a). Ferroplasma, a species of Archaea, is an extremely acidophilic iron oxidizer and can grow at pH values below 0 (Section 17.3). We discuss the role of all of these organisms in acid mine pollution and mineral oxidation in Sections 21.6, 22.1, and 22.2.
At neutral pH, Fe2+ spontaneously oxidizes to Fe3+, so opportunities for the iron bacteria in neutral habitats are restricted to locations where Fe2+ is transitioning from anoxic to oxic conditions. For example, anoxic groundwater often contains dissolved Fe2+, and when it is released, as in iron-rich spring water, it becomes exposed to O2. At such interfaces, iron bacteria oxidize Fe2+ to Fe3+ before it oxidizes spontaneously. Gallionella ferruginea, Sphaerotilus natans, and Leptothrix discophora are examples of bacteria that live at these interfaces. They are typically seen mixed in with the characteristic ferric iron deposits they form (Figures 15.55 and 21.6).
Energy from Iron Oxidation
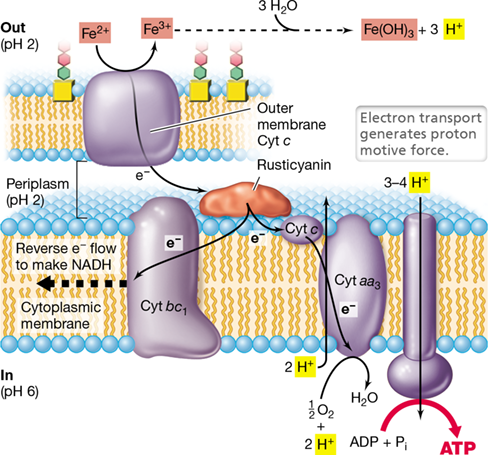
The bioenergetics of ferrous iron oxidation by Acidithiobacillus ferrooxidans and other acidophilic iron oxidizers are of considerable interest because of the very electropositive reduction potential of the Fe3+/Fe2+ couple at acidic pH (E0′=+0.77 V at pH 2). The respiratory chain of A. ferrooxidans contains cytochromes of the c and aa3 types and a periplasmic copper-containing protein called rusticyanin (Figure 14.22). There is also an iron-oxidizing protein located in the outer membrane of this gram-negative bacterium.
Figure 14.22 Electron flow during Fe2+ oxidation by the acidophile *Acidithiobacillus ferrooxidans*.

The periplasmic copper-containing protein rusticyanin receives electrons from Fe2+ oxidized by a c-type cytochrome located in the outer membrane. From here, electrons travel through a short electron transport chain, resulting in the reduction of O2 to H2O. Reducing power comes from reverse electron flow. Note the steep pH gradient across the cytoplasmic membrane.
Because the reduction potential of the Fe3+/Fe2+ couple is so high, steps in electron transport to oxygen (12O2/H2O, E0′=+0.82 V) are necessarily few. Iron oxidation begins in the outer membrane where the organism contacts either soluble Fe2+ or insoluble ferrous iron minerals such as FeS. Fe2+ is oxidized to Fe3+, a one-electron transition (Table 14.1), by an outer membrane cytochrome c that transfers electrons into the periplasm where rusticyanin (E0′=+0.68 V) is the electron acceptor. This thermodynamically slightly unfavorable reaction is thought to be pulled forward by the immediate consumption of Fe3+ by Fe(OH)3 formation (Figure 14.22). Rusticyanin then reduces a periplasmic cytochrome c that transfers electrons to cytochrome aa3, and it is the latter protein that reduces O2 to H2O; ATP is synthesized by ATPase in the usual fashion (Figure 14.22).
The nature of the proton motive force in A. ferrooxidans is of interest. In a highly acidic environment, a large gradient of protons already exists across the A. ferrooxidans cytoplasmic membrane (the periplasm is pH 1–2, whereas the cytoplasm is about pH 6, Figure 14.22). Although one might think that with this gradient A. ferrooxidans could make ATP at no energetic cost, this is not the case; the organism cannot make ATP from this preformed proton motive force in the absence of an electron donor. This is because H+ ions that enter the cytoplasm via ATPase must be consumed in order to maintain the internal pH within acceptable limits. Proton consumption occurs during the reduction of O2 in the electron transport chain and this reaction requires electrons; the latter come from the oxidation of Fe2+ to Fe3+ (Figure 14.22).
Autotrophy in A. ferrooxidans is supported by the Calvin cycle (Section 14.2), and because of the high potential of the electron donor, much energy must be consumed in reverse electron flow reactions to obtain the reducing power (NADH or NADPH) necessary to drive CO2 fixation (Section 14.1). NADH is formed by reduction of NAD+ by electrons obtained from Fe2+ that are forced backwards through cytochrome bc1 and the quinone pool at the expense of the proton motive force (Figure 14.22).
The relatively poor energetic yield from ferrous iron oxidation coupled with the large energetic demands of the Calvin cycle means that A. ferrooxidans must oxidize large amounts of Fe2+ to produce even a very small amount of cell material. Thus, in environments where acidophilic iron-oxidizing bacteria thrive, their presence is signaled not by the formation of high cell numbers but by the presence of the extensive ferric iron precipitates they have generated (Figure 14.21).We consider the ecology of iron bacteria in Chapters 21 and 22.
Ferrous Iron Oxidation under Anoxic Conditions
Ferrous iron can be oxidized under anoxic conditions by certain chemolithotrophs and phototrophic purple and green bacteria (Figure 14.23). In these cases, Fe2+ is used either as an electron donor in energy metabolism (chemolithotrophs) and/or as a reductant for CO2 fixation (phototrophs). An important point to consider here is that at neutral pH where these organisms thrive, the E0′ of the Fe3+/Fe2+ couple is significantly more electronegative than at acidic pH (+0.2 V versus +0.77 V, respectively, Figure 14.1). Hence, electrons from Fe2+ can reduce cytochrome c to initiate electron transport reactions. For chemolithotrophs, the electron acceptor is nitrate (NO3 −) with either nitrite (NO2 −) or dinitrogen gas (N2) being the final product of this anaerobic respiration. For Fe2+-oxidizing purple and green bacteria, either soluble Fe2+ or iron sulfide (FeS) can be used as electron donor. With FeS, both Fe2+ and S2− are oxidized, Fe2+ to Fe3+ (one electron) and HS− to SO4 2− (eight electrons).
Figure 14.23 Fe2+ oxidation by anoxygenic phototrophic bacteria.

(a) Oxidation in anoxic tube cultures. Left to right: Sterile medium, inoculated medium, a growing culture showing Fe(OH)3. (b) Phase-contrast photomicrograph of an Fe2+-oxidizing purple bacterium, strain L7. The bright refractile areas within cells are gas vesicles (Section 2.7). The granules outside the cells are iron precipitates. Strain L7 is phylogenetically related to the purple sulfur bacterium Chromatium.
We now consider chemolithotrophic metabolisms of nitrogen compounds, major components of the nitrogen cycle in nature (Section 21.3).
Check Your Understanding
Why is only a very small amount of energy available from the oxidation of Fe2+ to Fe3+ at acidic pH?
What is the function of rusticyanin and where is it found in the cell?
14.9 Nitrification
The reduced inorganic nitrogen compounds ammonia (NH3) and nitrite (NO2 −) are oxidized aerobically by the chemolithotrophic nitrifying bacteria in the process of nitrification (Section 15.10). Nitrifying bacteria are widely distributed in soils, water, wastewaters, and the oceans. Nitrification consists of two different sets of reactions; the first set of reactions catalyze oxidation of ammonia to nitrite, and the second set catalyze oxidation of nitrite to nitrate (NO3 −). Most nitrifying microbes are only able to catalyze one set of these reactions. For example, Bacteria such as Nitrosomonas and Archaea such as Nitrosopumilus oxidize NH3 only to NO2 −, and we call these organisms ammonia oxidizers. The full nitrification pathway is ultimately completed when other Bacteria such as Nitrobacter oxidize NO2 − to NO3 −, and we call these organisms nitrite oxidizers. As far as is known, only certain bacteria in the genus Nitrospira can catalyze both sets of reactions, oxidizing NH3 all the way to NO3 −.
Bioenergetics and Enzymology of Ammonia and Nitrite Oxidation
The bioenergetics of nitrification is based on the same principles that govern other chemolithotrophic reactions: Electrons from reduced inorganic substrates (in this case, reduced nitrogen compounds) enter an electron transport chain, and electron transport reactions establish a proton motive force that drives ATP synthesis. The complete oxidation of NH3 to NO3 − releases eight electrons, but the electron donors for the nitrifying bacteria are not particularly strong. For example, the E0′ of the NO2 −/NH3 couple (the first step in the oxidation of NH3) is +0.34 V, and the E0′ of the NO3 −/NO2 − couple is even more positive, about +0.43 V. By necessity, these reduction potentials force the nitrifying bacteria to donate electrons to rather high-potential electron acceptors, and this of course limits the amount of energy that can be conserved (Section 14.1).
Mastering Microbiology
Art Activity: Figure 14.32 Oxidation of NH3 and electron flow in ammonia-oxidizing bacteria
Several key enzymes participate in the oxidation of reduced nitrogen compounds. In ammonia-oxidizing bacteria such as Nitroso-monas, NH3 is oxidized by ammonia monooxygenase (AMO; monooxygenases are discussed in Section 14.23), producing hydroxylamine (NH2OH) and H2O (Figure 14.24). A second key enzyme, hydroxylamine oxidoreductase (HAO), then oxidizes NH2OH to NO2 −, removing four electrons in the process. AMO is an integral membrane protein, whereas HAO is periplasmic (Figure 14.24). In the reaction carried out by AMO, NH3+O2+2 H++2 e−→NH2OH+H2O
two electrons and two protons are needed to reduce one molecule of oxygen (O2) to H2O. These electrons originate from the oxidation of hydroxylamine and are supplied to AMO from HAO (Figure 14.24). Thus, for every four electrons generated from the oxidation of NH3 to NO2 −, only two actually reach the terminal oxidase that interacts with O2 to form H2O (Figure 14.24).
Figure 14.24 Oxidation of NH3 and electron flow in the cytoplasmic membrane of ammonia-oxidizing bacteria.

The reactants and the products of this reaction series are highlighted. The top surface of the membrane is the beginning of the periplasm. The cytochrome c (Cyt c) in the periplasm is a different form of Cyt c than that in the cytoplasmic membrane. AMO, ammonia monooxygenase; HAO, hydroxylamine oxidoreductase; Q, ubiquinone.
Mastering Microbiology
Art Activity: Figure 14.33 Oxidation of NO2 − to NO3 − by nitrifying bacteria
Nitrite-oxidizing bacteria such as Nitrobacter oxidize NO2 − to NO3 − by the enzyme nitrite oxidoreductase, with electrons traveling a very short electron transport chain (because of the high potential of the NO3 −/NO2 − couple) to the terminal oxidase (Figure 14.25). Cytochromes of the c and bd types are present in the electron transport chain of nitrite oxidizers, and the activity of the bd-type cytochrome generates a proton motive force (Figure 14.25). As is the case with the iron bacteria (Section 14.8), only small amounts of energy are available from nitrite oxidation. Hence, minimal amounts of cell material are obtained even though large amounts of nitrite may be oxidized.
Figure 14.25 Oxidation of NO2 − to NO3 − by nitrifying bacteria.

The reactants and products of this reaction series are highlighted to show the reaction clearly. NXR, nitrite oxidoreductase; Cyt, cytochrome.
Carbon Metabolism and Ecology of Nitrifying Bacteria
Like sulfur- and iron-oxidizing chemolithotrophs (Sections 14.7 and 14.8), aerobic nitrifying Bacteria employ the Calvin cycle for CO2 fixation. The ATP and reducing power requirements of the Calvin cycle place additional burdens on an energy-generating system that already has a relatively low yield; NAD(P)H to drive the Calvin cycle in nitrifiers is formed by reverse electron flow (Figures 14.24 and 14.25). The energetic constraints are particularly severe for nitrite oxidizers, and it is perhaps for this reason that most of these organisms have alternative energy-conserving mechanisms, being able to grow chemoorganotrophically on glucose and a few other organic substrates. By contrast, species of ammonia-oxidizing bacteria are either obligate chemolithotrophs and autotrophs, or are mixotrophic, both fixing CO2 and using organic compounds (Section 14.2). Autotrophy in ammonia-oxidizing Archaea is supported by the 3-hydroxypropionate/4-hydroxybutyrate cycle (Section 14.2).
Nitrifying microbes play key ecological roles in the nitrogen cycle, converting ammonia in soils into nitrate, a key plant nutrient. Nitrifiers are also important in sewage and wastewater treatment, removing toxic amines and ammonia and releasing less toxic nitrogen compounds (Sections 22.6 and 22.7). Nitrifiers play a similar role in the water column of lakes, where ammonia produced in the sediments from the decomposition of organic nitrogenous compounds is oxidized to nitrate, a more usable fixed nitrogen source for algae and cyanobacteria.
Although classical nitrifiers are obligate aerobes, we will see next that ammonia can be oxidized anaerobically as well, by a unique group of Bacteria.
Check Your Understanding
What are the substrates for the enzyme ammonia monooxygenase?
What is the difference between ammonia oxidation and nitrite oxidation, and in what types of organisms are these reactions found?
Why do both ammonia- and nitrite-oxidizing bacteria require reverse electron transport?
14.10 Anaerobic Ammonia Oxidation (Anammox)
In addition to the oxygen-dependent process of ammonia oxidation by organisms such as Nitrosomonas and Nitrosopumilus, NH3 can also be oxidized under anoxic conditions. This process is called anammox (for anaerobic ammonia oxidation) and is catalyzed by an unusual group of obligately anaerobic Bacteria.
Ammonia is oxidized in the anammox reaction using NO2 − as the electron acceptor to yield N2: NH4 ++NO2 −→N2+2 H2OΔG0′=−357kJ
A major anammox organism, Brocadia anammoxidans, is a species of the Planctomycetes phylum of Bacteria (Section 16.16). Planctomycetes are structurally unusual Bacteria because their cytoplasm can contain membrane-enclosed compartments of various types (Figure 14.26). In cells of B. anammoxidans, this compartment is the anammoxosome, and it is within this structure that the anammox reaction occurs (Figure 14.26c). In addition to Brocadia, several other genera of anammox bacteria are known, including Kuenenia, Anammoxoglobus, Jettenia, and Scalindua, all of which are related to Brocadia and also contain anammoxosomes. Like aerobic ammonia oxidizers, anammox bacteria are also autotrophs, but they do not fix CO2 using the pathways employed by aerobic ammonia oxidizers. Instead, anammox bacteria fix CO2 by way of the reductive acetyl-CoA pathway, an autotrophic pathway widespread among some obligately anaerobic autotrophic Bacteria and Archaea (see Section 14.14).
Figure 14.26 Anammox.

(a) Phase-contrast photomicrograph of cells of Brocadia anammoxidans. A single cell is about 1 μm in diameter. (b) Transmission electron micrograph of a Brocadia cell; note the membrane-enclosed compartments including the large fibrillar anammoxosome. (c) Reactions in the anammoxosome. NIR, nitrite reductase, HZS, hydrazine synthase; HDH, hydrazine dehydrogenase.
The Anammoxosome and Its Reactions
The anammoxosome is a unit membrane–enclosed structure (Figure 14.26b) and in this respect is technically an organelle in the eukaryotic sense of the term. Lipids that form the anammoxosome membrane are not the typical lipids of Bacteria but instead consist of fatty acids constructed of multiple cyclobutane (C4) rings bonded to glycerol by both ester and ether bonds. These ladderane lipids, as they are called, aggregate in the membrane to form an unusually dense membrane structure that prevents diffusion of substances from the anammoxosome into the cytoplasm.
The sturdy anammoxosome membrane is required to protect the cell from toxic intermediates produced during anammox reactions. These include, in particular, the compound hydrazine (N2H4), a very strong reductant. In the anammox reaction, NO2 − is first reduced to nitric oxide (NO) by nitrite reductase, and then NO reacts with ammonium (NH4 +) to yield N2H4 by activity of the enzyme hydrazine synthase (Figure 14.26c). (Note that while NH3 is the substrate for AMO in aerobic ammonia-oxidizing bacteria [Figure 14.24], NH4 + is the substrate for hydrazine synthase in the anaerobic anammox bacteria.) N2H4 is then oxidized to N2 plus electrons by the enzyme hydrazine dehydrogenase, and the electrons are funneled into the electron transport chain where they are used to reduce NO2 − and NO earlier in the pathway. In this way, anammox generates a cyclical series of electron transfer reactions in order to generate a proton motive force; ATP is formed from the latter by ATPases in the anammoxosome membrane (Figure 14.26c).
Reducing power for CO2 fixation by anammox bacteria is derived from reverse electron transport (Section 14.1), but because electron transfer reactions are cyclic, the electrons needed for reverse electron transport derive from an independent set of reactions that oxidize NO2 − to NO3 − by a nitrite oxidoreductase, a reaction also present in Nitrobacter (Figure 14.25). Interestingly, then, NO2 − serves two different purposes for anammox bacteria: The reduction of nitrite is required to generate ATP by chemiosmosis, whereas the oxidation of nitrite is required to generate reducing power for CO2 fixation.
Ecology of Anammox
In nature, the source of NO2 − for the anammox reaction is presumably aerobic ammonia-oxidizing Bacteria and Archaea. These organisms coexist with anammox bacteria in ammonia-rich habitats such as sewage and other wastewaters. The suspended particles that form in these habitats contain both oxic and anoxic zones in which ammonia oxidizers of different physiologies can coexist in close association. In mixed laboratory cultures, high levels of oxygen inhibit anammox and favor classic nitrification, and thus it is likely that in nature, the fraction of ammonia oxidation catalyzed by anammox bacteria is governed by the concentration of O2 in the habitat.
From an environmental standpoint, anammox is a very beneficial process in the treatment of wastewaters. The anoxic removal of ammonia and amines by the formation of N2 (Figure 14.26c) helps reduce the input of fixed nitrogen from wastewater treatment effluents that flow into rivers and streams, thereby maintaining higher water quality than would otherwise be possible. Also, marine anammox bacteria are likely responsible for the large amount of NH3 that is known to disappear during mineralization processes in anoxic marine sediments. At least some ammonia-rich freshwater lake sediments also support anammox, and thus it appears that anammox can occur in any anoxic environment in which NH3 and NO2 − coexist.
We move on to Part IV now where chemotrophic energy metabolisms will be defined not by the electron donor that is used, but by the electron acceptor that is used to receive electrons from a wide variety of donors.
Check Your Understanding
Name the electron donor and the electron acceptor in the anammox process.
What does electron transport for reducing power synthesis in chemolithotrophs (e.g., anammox bacteria, nitrifiers, and sulfide oxidizers) have in common with that of purple sulfur bacteria?
Compare autotrophy in anammox bacteria and purple sulfur bacteria. What characteristics do these processes share and how are they different?
IV: Respiratory Processes Defined by Electron Acceptor
IV: Respiratory Processes Defined by Electron Acceptor
IV Respiratory Processes Defined by Electron Acceptor
Anaerobic respiration occurs when an alternative electron acceptor is substituted for oxygen in respiration. Many forms of anaerobic respiration are possible using oxidized nitrogen and sulfur compounds, and some metals and organic compounds support the process as well.
We examined the process of aerobic respiration in Chapter 3 and reviewed the bioenergetics of respiration in Section 14.1. Here we consider the details of anaerobic respiration in its many variations found in the microbial world. Diverse compounds function as electron acceptors in anaerobic respirations (Figure 14.1), and each acceptor has its own redox properties and is typically linked to a specific group or groups of microbes. We begin with a common form of anaerobic respiration in which nitrate functions as electron acceptor.
14.11 Nitrate Reduction and Denitrification
Inorganic nitrogen compounds are some of the most common electron acceptors in anaerobic respiration. Table 14.3 summarizes the relevant forms of inorganic nitrogen with their oxidation states. One of the most common alternative electron acceptors for dissimilative purposes is nitrate (NO3 −), which can be reduced with two electrons to nitrite (NO2 −), or reduced further to nitric oxide (NO), nitrous oxide (N2O), or dinitrogen (N2). Because NO, N2O, and N2 are all gases, they can be lost from the environment, and their biological production is called denitrification (Figure 14.27).
Table 14.3 Oxidation states of key nitrogen compounds

Figure 14.27 Steps in the dissimilative reduction of nitrate.

Some organisms can carry out only the first step. All enzymes involved are derepressed by anoxic conditions. Also, some bacteria are known that can reduce NO3 − to NH4 + in dissimilative metabolism. Note that colors used here match those used in Figure 14.28.
Some nitrate reducers, for example Escherichia coli, are not true denitrifiers, but only carry out the first step (NO2 − to NO3 −) in the process (Figure 14.27and Figure 3.23). Moreover, some organisms can reduce NO2 − to ammonia (NH3) in a dissimilative process. But it is the production of gaseous products—denitrification—that is of greatest global significance because it consumes fixed nitrogen and produces some polluting gases.
Denitrifying Microorganisms and their Ecological Activities
Many denitrifying Bacteria are phylogenetically Proteobacteria and physiologically facultative aerobes. Pseudomonas species, for example, are typically strong denitrifiers. Aerobic respiration occurs when O2 is present, even if NO3 − is also present in the medium. Many denitrifying bacteria also reduce other electron acceptors anaerobically, such as Fe3+ and certain organic electron acceptors (Figure 14.1), and some denitrifiers can even ferment. Thus, denitrifying bacteria are metabolically diverse in terms of alternative energy-generating mechanisms. Some species of Archaea can grow anaerobically by nitrate reduction to nitrite, and several others can denitrify. Interestingly, at least one eukaryote has also been shown to be a denitrifier. The protist Globobulimina pseudospinescens, a shelled amoeba (a foraminiferan, Section 18.6), can denitrify and likely employs this form of metabolism in its habitat, anoxic marine sediments.
Denitrification is a process with significant ecological ramifications. For agricultural purposes, denitrification is a detrimental process, as it removes nitrate—often intentionally added by grain and other crop farmers as potassium nitrate fertilizer—from the soil. Gaseous products of denitrification other than N2 (N2O and NO) are also of significant environmental concern. N2O is a strong greenhouse gas (contributing to climate change) and can also be converted to NO by sunlight; NO reacts with and consumes ozone (O3) in the upper atmosphere to form NO2 −. When it rains, NO2 − returns to Earth as nitrous acid (HNO2) in acid rain. In contrast to these environmentally harmful processes, for a desirable process such as sewage treatment, denitrification (as well as anammox, see earlier) is beneficial. This is because denitrification removes fixed nitrogen, a major trigger of algal growth if a nitrate-rich sewage effluent is released into rivers or lakes (Sections 22.6 and 22.7).
Biochemistry of Dissimilative Nitrate Reduction
The electron transport pathway of denitrification is shown in Figure 14.28. The enzyme that catalyzes the first step of dissimilative nitrate reduction is nitrate reductase, a molybdenum-containing, membrane-integrated enzyme whose synthesis is repressed by O2. All subsequent enzymes of the pathway are coordinately regulated and thus also repressed by O2. In addition to anoxic conditions, nitrate must also be present before these enzymes are fully expressed.
Figure 14.28 Denitrification.

Scheme for electron transport in membranes of Pseudomonas stutzeri during denitrification. Nitrate and nitric oxide reductases are integral membrane proteins, whereas nitrite and nitrous oxide reductases are periplasmic enzymes. Fp, fla-voprotein; Q, ubiquinone; Cyt, cytochrome.
The biochemistry of dissimilative nitrate reduction has been studied in detail in E. coli, in which NO3 − is reduced only to NO2 − (Section 3.10) and Paracoccus denitrificans and Pseudomonas stutzeri, in which denitrification occurs. In P. denitrificans and P. stutzeri, nitrogen oxides are formed enzymatically from NO2 − by nitrite reductase, nitric oxide reductase, and nitrous oxide reductase (Figure 14.28). NO and N2O are gaseous intermediates that are free to escape from the cell, and N2O in particular is a major product of denitrification, though these intermediates are often reduced all the way to N2. During these electron transport reactions, a proton motive force is established (Figure 14.28), and ATPase couples this to the synthesis of ATP.
We move on from coverage of nitrogen compounds as electron acceptors in anaerobic respiration to consider sulfur compounds as electron acceptors in the next section. A diverse group of organisms will be in play here that generate a potentially toxic product but one that is central to the sulfur cycle. Following that, we round out this part of the chapter by considering a few key metals and organic compounds as electron acceptors.
Check Your Understanding
Under what condition would you expect the genes for denitrification to be repressed?
If the final product of denitrification is N2 then why is N2O released into the environment?
Where is dissimilative nitrate reductase found in the cell? What unusual metal does it contain?
14.12 Sulfate and Sulfur Reduction
Several inorganic sulfur compounds are important electron acceptors in anaerobic respiration, and Table 14.4 lists the oxidation states of the key compounds. Sulfate (SO4 2−), the most oxidized form of sulfur, is reduced by the sulfate-reducing bacteria, a highly diverse group of obligately anaerobic bacteria widely distributed in nature. The product of sulfate reduction is hydrogen sulfide, H2S (which occurs mostly as HS− at pH 7), an important and potentially toxic natural product that participates in many biogeochemical processes (Sections 21.4, 22.6, and 22.7). Sulfate-reducing species in the genus Desulfovibrio, in particular D. desulfuricans, have been widely studied, and the general properties of sulfate-reducing bacteria are discussed in Section 15.11.
Table 14.4 Sulfur compounds for sulfate reduction

As was the case with nitrate metabolism (Section 14.11), it is necessary to distinguish between assimilative and dissimilative sulfate metabolism. Most microbes can incorporate sulfate for biosynthetic purposes to make cysteine, methionine, and many other organosulfur compounds; this is assimilative sulfate metabolism. By contrast, the ability to use sulfate as an electron acceptor for energy conservation requires its large-scale reduction and is restricted to the sulfate-reducing bacteria. H2S is produced on a very large scale by these organisms and is excreted from the cell, free to be oxidized by air, used by other organisms, or combined with metals to form metal sulfides. Although a major electron donor to many microbes, an environmental concern of H2S production is the fact that sulfide is highly toxic to higher organisms such as fish and wildlife. When anoxic conditions develop in sulfate-containing waters, the production of H2S can cause die-offs of fish and other aquatic organisms and reduce water quality for recreational use or consumption.
Biochemistry and Energetics of Sulfate Reduction
As shown in Figure 14.1, SO4 2− is an energetically much less favorable electron acceptor than is O2 or NO3 −. Hydrogen (H2) is used as an electron donor by virtually all species of sulfate reducers, whereas the use of organic electron donors is more restricted. For example, lactate and pyruvate are widely used by sulfate-reducing bacteria found in freshwater anoxic environments while acetate and longer-chain fatty acids are widely used by marine sulfate-reducing bacteria. Many morphological and physiological types of sulfate-reducing bacteria are known, and with the exception of Archaeoglobus (Section 17.4), a genus of Archaea, all known sulfate reducers are Bacteria (Section 15.11).
The reduction of SO4 2− to H2S requires eight electrons and proceeds through a number of intermediate stages. The reduction of SO4 2− requires that it first be activated in a reaction requiring ATP. The enzyme ATP sulfurylase catalyzes the attachment of SO4 2− to a phosphate of ATP, forming adenosine phosphosulfate (APS) as shown in **Figure 14.29*a***. Activation raises the extremely electronegative E0′ of the SO4 2−/SO3 2− couple (−0.52 V) to near 0 V, making reduction of the sulfate moiety possible with electron donors such as NADH (−0.32 V). In dissimilative sulfate reduction, the SO4 2− in APS is reduced directly to sulfite (SO3 2−) by the enzyme APS reductase with the release of AMP. In assimilative reduction, by contrast, a second phosphate is added to APS to form phosphoadenosine phosphosulfate (PAPS) (Figure 14.29a), and only then is the SO4 2− reduced. However, in both cases the product of sulfate reduction is sulfite (SO3 2−), and once SO3 2− is formed, it is reduced to H2S by a sulfite reductase enzyme (Figure 14.29b).
Figure 14.29 Biochemistry of sulfate reduction: Activated sulfate.

(a) Two forms of active sulfate can be made, adenosine 5′-phosphosulfate (APS) and phosphoadenosine 5′-phosphosulfate (PAPS). Both are derivatives of adenosine diphosphate (ADP), with the second phosphate of ADP being replaced by SO4 2−. (b) Schemes of assimilative and dissimilative sulfate reduction.
Sulfate reduction offers only a very low free-energy yield and so organisms have developed a variety of mechanisms to optimize electron flow and energy conservation. During dissimilative sulfate reduction, electron transport reactions generate a proton motive force and this drives ATP synthesis by ATPase. Cytochrome c3, a periplasmic low-potential cytochrome (Figure 14.30), is a major intermediate in these reactions. Cytochrome c3 can accept electrons from several different periplasmic hydrogenase enzymes and can transfer these electrons to a number of different membrane-associated protein complexes. Hydrogenases are enzymes that perform redox reactions that produce or consume H2. Sulfate reducers that use organic substrates typically do so by oxidizing these substrates to produce H2 that then feeds into the electron transport chain by way of these periplasmic hydrogenases and cytochrome c3 (Figure 14.30).
Figure 14.30 Electron transport and energy conservation in sulfate-reducing bacteria.

H2 serves as primary electron donor for sulfate reduction, and growth on organic compounds such as lactate result in the production of H2 as an intermediate. The periplasmic cytochrome c3 transfers electrons from a system of periplasmic hydrogenases (H2ase) to membrane-bound proteins that mediate electron transfer reactions. Core components of electron transfer are shown in purple, and also shown are components required for growth exclusively on H2 (in green) and on lactate (in blue). The scheme is proposed to include three bifurcating enzymes. Electron bifurcation at the APS reductase complex results in the donation of electrons to APS. Electron bifurcation at the heterodisulfide reductase complex (Hdr) results in the donation of electrons to Dsr-C and the release of HS−. An electron confurcation at the CO-induced hydrogenase complex (Coo) contributes to redox balance during growth on lactate. Energy-converting hydrogenases (see Section 14.17) contribute to energy conservation in addition to quinone cycling.
Dissimilative sulfate reduction requires several enzymes that perform flavin-based electron bifurcation (Section 14.1). For example, the reduction of APS to SO3 2− requires the activity of the APS reductase complex (Figure 14.30). The APS reductase complex includes membrane-bound proteins that funnel electrons from a reduced quinone to a cytoplasmic enzyme that performs an electron confurcation reaction (Section 14.1) combining 2 e− from reduced ferredoxin (Fdred) and 2 e− from the quinone to reduce 2 APS to 2 SO3 2−. In addition, the dissimilative reduction of SO3 2− to HS− occurs in two steps. The first step is a two-electron transfer mediated by dissimilatory sulfite reductase (DsrAB); this reaction forms a heterodisulfide bond between S (from SO3 2−) and the protein DsrC (Figure 14.30). The heterodisulfide is reduced in a four-electron transfer mediated by an electron-bifurcating heterodisulfide reductase (Hdr, reaction 4 in Figure 14.30). This Hdr reaction is similar to a key reaction in methanogenesis (see Section 14.15 and Figure 14.38). These cells also balance electron flow using an energy-converting hydrogenase, a membrane-bound hydrogenase enzyme that uses the exergonic reduction of protons by Fdred to generate a pmf (Figure 14.30). This reaction has a standard free-energy change (ΔG0′) close to zero, but under natural conditions, the ΔG is typically exergonic. A net of about two ATP is produced for each SO4 2− reduced to HS− by H2, and the reaction is 4 H2+SO4 2−+H+→HS−+4 H2 OΔG0′=−152 kJ
Oxidation of organic compounds, such as lactate, is used to produce H2 through energy-converting hydrogenases or by electron confurcation (Figure 14.30). This H2 crosses the cytoplasmic membrane and is oxidized by the periplasmic hydrogenase to electrons—which are fed back into the system—and protons, which establish the proton motive force (Figure 14.30). When lactate or pyruvate is the electron donor, ATP is produced not only from pmf but also by substrate-level phosphorylation during the oxidation of pyruvate to acetate plus CO2 (reaction 9 in Figure 14.30). Many strains are ultimately unable to further oxidize acetate and so while some acetate is assimilated during biosynthetic reactions, the remainder is excreted from the cell.
Marine but not freshwater species of sulfate-reducing bacteria can couple sulfate reduction and the oxidation of acetate (and longer-chain fatty acids) to CO2: CH3COO−+SO4 2−+3 H+→2 CO2+H2S+2 H2OΔG0′=−57.5 kJ
The mechanism for acetate oxidation in most of these species is the reductive acetyl-CoA pathway, a series of reversible reactions used by many anaerobes for acetate synthesis or acetate oxidation (Section 14.14). A few sulfate-reducing bacteria can also grow autotrophically with H2. Under these conditions, the organisms use the reductive acetyl-CoA pathway for making acetate as a carbon source. Such species can be cultured in a completely organic-free medium containing only mineral salts, sulfate, CO2, and H2.
Special Metabolisms of Sulfate-Reducing Bacteria
Certain species of sulfate-reducing bacteria can catalyze reactions not characteristic of all species. These include sulfur reduction, disproportionation, and even phosphite oxidation. Most sulfate-reducing bacteria can also conserve energy from the reduction of elemental sulfur to sulfide (S0+2 H→H2S). The electrons for sulfur reduction come from H2 or any of a number of organic compounds. For example, Desulfuromonas acetoxidans can oxidize acetate or ethanol to CO2 coupled to the reduction of S0 to H2S.
Many microbes can reduce elemental sulfur but not sulfate in anaerobic respiration. These are the sulfur reducers, a diverse group of Bacteria and Archaea that often coexist with sulfate-reducing bacteria in anoxic, sulfur-rich habitats. Sulfur reducers lack the capacity to activate sulfate to APS (Figure 14.29), and presumably this is what excludes them from using SO4 2− as an electron acceptor.
Disproportionation is a process in which one molecule of a substance is oxidized while a second molecule is reduced, ultimately forming two different products. For example, Desulfovibrio sulfodismutans can disproportionate thiosulfate (S—SO3 2−) as follows: S—SO3 2−+H2O→H2S+SO4 2−ΔG0′=−21.9 kJ/reaction
Note that in this reaction, the right-hand sulfur atom of S—SO3 2− is oxidized (forming SO4 2−), while the left-hand atom is reduced (forming H2S). The free energy available from the oxidation of thiosulfate by D. sulfodismutans is insufficient to couple to substrate-level phosphorylation and so instead is coupled to a proton “pump” that uses the minimal energy available in the reaction to establish a proton motive force. Other partially reduced sulfur compounds such as sulfite (SO3 2−) and sulfur (S0) can also be disproportionated.
At least one sulfate-reducing bacterium can couple phosphite (HPO3 −) oxidation to SO4 2− reduction. This bacterium, Desulfotignum phosphitoxidans, is an autotroph and also a strict anaerobe. The chemolithotrophic reaction carried out by D. phosphitoxidans yields phosphate and sulfide: 4 HPO3−+SO4 2−+H+→4 HPO4 2−+HS−ΔG0′=−364 kJ
The natural sources of phosphite are likely to be organophosphorous compounds called phosphonates that are generated from the anoxic degradation of nucleic acids, phospholipids, and other cellular sources. Along with sulfur respiration, sulfur disproportionation (also a chemolithotrophic process), and H2 utilization, phosphite oxidation underscores the diversity of metabolic reactions carried out by sulfate-reducing bacteria.
Check Your Understanding
What mechanisms do sulfate reducers use to balance electron flow and conserve energy when growing on organic substrates such as lactate?
Why is cytochrome c3 so important for energy conservation in sulfate-reducing bacteria?
What difference in metabolism allows some sulfate reducers to oxidize organic carbon substrates all the way to CO2 while others stop at acetate?
14.13 Other Electron Acceptors
Bacteria can perform a wide diversity of anaerobic respirations (Figure 14.1) including the respiration of several metals, metalloids, and associated minerals. In addition, certain organic and halogenated compounds can also serve as important electron acceptors in anaerobic respiration. We consider these forms of anaerobic respiration here.
Metal and Metalloid Reduction
A variety of metals, metalloids (elements such as arsenic, As, whose properties are intermediate between those of a metal and a nonmetal), and associated minerals can be reduced through anaerobic respiration. Ferric iron (Fe3+) and manganic ion (Mn4+) are the most common such electron acceptors, but other possible electron acceptors include As5+, Co3+, Cr6+, Se6+, Tc7+, Te6+, U6+. The reduction potential of the Fe3+/Fe2+ couple is +0.2 V (at pH 7), and that of the Mn4+/Mn2+ couple is +0.8 V; thus, several electron donors can couple to Fe3+ and Mn4+ reduction. A major challenge for metal-reducing organisms is that oxidized minerals of Fe and Mn are often insoluble, making transport into the cell difficult if not impossible. Hence, cells performing metal respiration need a mechanism for exporting electrons outside of the cell to allow the reduction to occur on the outer cell surface.
A number of different systems have evolved in diverse bacteria for reducing extracellular insoluble substrates. One well-studied example is Geobacter sulfurreducens, a gram-negative bacterium that can couple the oxidation of H2, acetate, or other organic electron donors to the reduction of a number of different metals and minerals (Section 15.13). G. sulfurreducens can oxidize acetate using the citric acid cycle (Section 3.6), and electrons make their way to electron transport by way of NADH. The electrons pass through quinones and are used to generate a pmf before reducing c-type cytochromes in the periplasm. These periplasmic cytochromes transfer electrons to a transmembrane porin–cytochrome complex embedded in the outer membrane. Finally, the electrons are transferred to specialized type IV pili (Section 2.6) that are electrically conductive (Figure 14.31). These electrically conductive type IV pili are called nanowires because they function like electrical wires. The pili are enriched in aromatic amino acids that function to conduct electrons. Attached to the pili are multiheme c-type cytochromes that function as an adaptor to allow electrons to flow out of the pili and into an insoluble terminal electron acceptor, such as ferrihydrite (Fe2O3).
Figure 14.31 Iron-reducing bacteria reduce extracellular electron acceptors.

Iron oxides are insoluble and cannot be transported into the cell. The bacterium Geobacter makes electrically conductive type IV pili (nanowires) that transfer electrons out of the cell where they reduce oxidized iron minerals.
Other inorganic substances can function as electron acceptors for anaerobic respiration, including the metalloids selenium, tellurium, and arsenic, and various oxidized chlorine compounds (some of these are shown in Figure 14.1). For example, the sulfate-reducing bacterium Desulfotomaculum auripigmentum can reduce AsO4 3− to AsO3 3−, and sulfate to sulfide, precipitating the yellow mineral orpiment (As2S3) in the process (Figure 14.32). This process is an example of biomineralization, the formation of a mineral by bacterial activity. The presence of soluble forms of arsenic in groundwater is a major problem in many parts of the world and so microbes that convert arsenic from a soluble to an insoluble form provide a means of detoxifying contaminated habitats. Metal- and metalloid-reducing bacteria are of great importance in groundwater chemistry, and their metabolic activities are often used to remediate contaminated sites (Section 22.3).
Figure 14.32 Biomineralization during arsenate reduction by the sulfate-reducing bacterium *Desulfotomaculum auripigmentum*.

Left, appearance of culture bottle after inoculation. Right, following growth for 2 weeks and biomineralization of arsenic trisulfide, As2S3, also called orpiment. Center, synthetic sample of As2S3.
Organic and Halogenated Electron Acceptors
Certain organic compounds can be electron acceptors in anaerobic respirations. Of those listed in Figure 14.1, the compound that has been most extensively studied is fumarate, a citric acid cycle intermediate that is reduced to succinate; the reaction in this anaerobic respiration is the opposite of the reaction that takes place in the citric acid cycle aerobically (Figure 3.12). The role of fumarate as an electron acceptor for anaerobic respiration derives from the fact that the fumarate/succinate couple has a reduction potential near 0 V, which allows coupling of fumarate reduction to the oxidation of NADH, FADH, or H2. Many facultatively aerobic bacteria can grow anaerobically on fumarate as electron acceptor, including Escherichia coli.
Trimethylamine oxide (TMAO) and dimethyl sulfoxide (DMSO) (Figure 14.1) are also important organic electron acceptors. TMAO is a product of marine fish, and several bacteria can reduce it to trimethylamine (TMA), which has a strong odor and flavor (the odor of spoiled seafood is due primarily to TMA produced by bacterial action). Dimethyl sulfoxide (DMSO), which is reduced to dimethyl sulfide (DMS), is a common natural product and is found in both marine and freshwater environments. The reduction potentials of the TMAO/TMA and DMSO/DMS couples are about the same (near +0.15 V, Figure 14.1) and so the electron transport chains that terminate with TMAO or DMSO reductases typically contain cytochromes of the b type.
Several halogenated organic compounds function as electron acceptors in reductive dechlorination (also called dehalorespiration). For example, the sulfate-reducing bacterium Desulfomonile grows anaerobically with H2 or organic electron donors and chlorobenzoate as an electron acceptor that is reduced to benzoate and hydrochloric acid (HCl): C7H4O2Cl−+2 H→C7H5O2−+HCl
Several other anaerobic bacteria can reductively dechlorinate, and some specialize in the use of chlorinated compounds as electron acceptors for anaerobic respiration. For example, Dehalobacter and Dehalococcoides oxidize H2 and reduce tetrachloroethylene (TCE) to dichloroethylene and ethene, respectively. Dehalococcoides can also reduce polychlorinated biphenyls (PCBs). TCE and PCBs are widespread organic pollutants that seep into groundwater and freshwater environments from underground storage tanks, leaking transformers, and other human inputs. However, removal of the chlorine groups from these molecules greatly reduces their toxicity and hence reductive dechlorination is not only a form of anaerobic respiration but also an environmentally significant process of bioremediation (Sections 22.3, 22.4 and 22.5).
We now explore the fascinating metabolisms of species of Bacteria and Archaea that specialize in the transformations of compounds that contain only one carbon: the C1 metabolizers.
Check Your Understanding
With H2 as electron donor, why is reduction of Fe3+ a more favorable reaction than reduction of fumarate?
Explain two different processes of anaerobic respiration that can be used to remediate contaminated environments.
How can bacteria transfer electrons to an insoluble extracellular electron acceptor?
V One-Carbon (C1) Metabolism
Acetogenesis and methanogenesis are energy-conserving metabolisms that exploit exergonic reductions of CO2 to form acetate and methane, respectively. Methanotrophs oxidize CH4 while methylotrophs oxidize C1 compounds other than CH4.
Carbon dioxide (CO2) and methane (CH4) are abundant in many anoxic habitats, and a wide diversity of microbes have evolved metabolic pathways that conserve energy from either the reduction of CO2 or the oxidation of CH4. A number of the enzymatic reactions in one-carbon metabolisms are unique. In this part of the chapter, we consider the metabolism of organisms that either consume or produce C1 compounds, highlighting the major similarities and differences.
14.14 Acetogenesis
Two major groups of strictly anaerobic microbes use CO2 as an electron acceptor for energy conservation. One of these is the acetogens, and we discuss them here. The other group, the methanogens, are considered in the next section. Hydrogen (H2) is a major electron donor for both groups of organisms, and an overview of their energy metabolism, acetogenesis and methanogenesis, is shown in Figure 14.33. Both processes are linked to ion pumps, of either protons (H+) or sodium ions (Na+), as the mechanism of energy conservation, and these pumps fuel ATP synthases in the membrane. The pathway of acetogenesis also conserves energy in a substrate-level phosphorylation reaction.
Figure 14.33 The contrasting processes of methanogenesis and acetogenesis.

Note the difference in free energy released in the reactions.
Organisms and Pathway
Acetogens carry out the reaction 4 H2+H++2 HCO3 −→CH3COO−+4 H2OΔG0′=−105 kJ
In addition to H2, electron donors for acetogenesis include various C1 compounds such as methanol, several methoxylated aromatic compounds, sugars, organic and amino acids, alcohols, and certain nitrogen bases, depending on the organism. Many acetogens can also grow by fermentation or by the anaerobic respiration of nitrate (NO3 −) or thiosulfate (S2O3 2−). However, CO2 reduction is a major feature of the acetogenic lifestyle, with CO2 serving both as electron acceptor and as an intermediate through which carbon is assimilated into cellular material.
A major unifying thread among acetogens is the pathway of CO2 reduction. Acetogens reduce CO2 to acetate by the reductive acetyl-CoA pathway (also called the Wood–Ljungdahl pathway), the major pathway in obligate anaerobes for CO2 fixation and the production of acetate (Section 14.2). The reactions of the reductive acetyl-CoA pathway are reversible, and some microbes reverse this pathway to oxidize acetate.
Acetogens such as Acetobacterium woodii and Clostridium aceticum can grow either chemoorganotrophically by fermentation of sugars (reaction A) or chemolithotrophically and autotrophically through the reduction of CO2 to acetate with H2 as electron donor (reaction B). In either case, the sole organic product is acetate:
When growing on glucose, acetogens use glycolysis (Figure 3.11) to oxidize the glucose into two molecules of pyruvate and two molecules of NADH. The pyruvate is then further oxidized to produce two molecules of acetate: C.
The CO2 generated in reaction (C) is then used as a terminal electron acceptor in the reductive acetyl-CoA pathway. The NADH generated during glycolysis and pyruvate oxidation is used as an electron donor in CO2 reduction. Starting from pyruvate, then, the overall production of acetate can be written as 2 Pyruvate−+4 H→3 acetate−+H+
Most acetogenic bacteria that produce acetate in energy metabolism are gram-positive Bacteria, and many are species of the genera Clostridium or Acetobacterium. A few other gram-positive and many different gram-negative Bacteria and Archaea use the reductive acetyl-CoA pathway for autotrophic purposes, reducing CO2 to acetate as a source of cell carbon; these include autotrophic sulfate-reducing bacteria (Section 14.12), anammox bacteria (Section 14.10), and methanogens (Section 14.15). And finally, some microbes run the acetyl-CoA pathway in the reverse direction as a means of oxidizing acetate to CO2; these include acetate-utilizing methanogens and sulfate-reducing bacteria. The acetyl-CoA pathway is thus a metabolically highly versatile series of reactions.
The Reductive Acetyl-CoA Pathway and Energy Conservation in Acetogenesis
Unlike other autotrophic pathways (Section 14.2), the reductive acetyl-CoA pathway of CO2 fixation is not a cycle. Instead, it catalyzes the reduction of CO2 along two linear pathways, with one molecule of CO2 being reduced to the methyl group of acetate (the methyl branch of the pathway) and the other to the carbonyl group of acetate (the carbonyl branch of the pathway). These two C1 units are then combined to form acetyl-CoA (Figure 14.34).
Figure 14.34 Reactions of acetogenesis from H2 and CO2 in *Acetobacterium woodii*.

The reductive acetyl-CoA pathway is used to reduce CO2 to acetate. The pathway has a methyl branch (blue arrows) and a carbonyl branch (red arrows). Carbon monoxide from the carbonyl branch and a methyl group from the methyl branch are combined into acetyl-CoA by carbon monoxide dehydrogenase ➀, a key enzyme for the pathway. Note that the reductive acetyl-CoA pathway itself does not conserve energy. An electron-bifurcating hydrogenase ➁ (see also Figure 14.2) is used to reduce ferredoxin (Fdred), and energy is conserved at the Rnf complex ➂, which generates a Na+ motive force. THF, tetrahydrofolate; CoFeSP, Co/Fe-containing corrinoid iron–sulfur protein.
A key enzyme of the acetyl-CoA pathway is carbon monoxide (CO) dehydrogenase. CO dehydrogenase contains Ni, Zn, and Fe as cofactors and catalyzes the reaction CO2+H2→CO+H2O
The CO produced by CO dehydrogenase ends up as the carbonyl carbon of acetate (reaction 1 in Figure 14.34). The methyl group of acetate originates from the reduction of CO2 by a series of reactions in which the coenzyme tetrahydrofolate (THF) plays a major role (Figure 14.34). The methyl group is then transferred from tetrahydrofolate to a cobalt- and iron-containing corrinoid iron–sulfur protein (CoFeSP) coenzyme. In the final step of the pathway, the methyl group is combined with CO by the activity of both CO dehydrogenase and acetyl-CoA synthase to form acetyl-CoA. Conversion of acetyl-CoA to acetate by the combined activities of the enzymes phosphotransacetylase and acetate kinase is the last step in the pathway, generating one ATP by substrate-level phosphorylation (Figure 14.34; see Table 14.5). However, this ATP is consumed in the first step of the acetyl-CoA pathway. Considering this, how then do acetogens get their ATP?
Acetogens conserve energy by the generation of an ion motive force. In Acetobacterium woodii, the best-studied acetogen, the ion motive force is generated by activity of the Rnf complex; this enzyme uses reduced ferredoxin (Fdred) as electron donor and NAD+ as electron acceptor. The Rnf complex pumps one Na+ across the membrane for each electron exchanged, thereby generating a Na+ motive force, which can be used to make ATP using a Na+-dependent ATP synthase (Figure 14.34). Alternatively, other acetogens such as Clostridium ljungdahlii use a proton motive force instead of a sodium motive force. The Fdred required by the Rnf cluster is provided by a flavin-based electron bifurcation reaction (Figure 14.2) in which the oxidation of 2 H2 is coupled to the reduction of one Fdox and one NAD+ (Figure 14.34). This bifurcation reaction is critical for producing the reduced ferredoxin required to conserve energy during acetogenic growth on H2. Ultimately, only 0.3 ATP are produced for every 4 H2 and 2 CO2 that are consumed by the acetogens, making these organisms minimalists in terms of the energy that can be conserved from their metabolism.
As we have seen, acetogens start with a C1 or C6 compound and make a C2 product in acetate. We now examine methanogens, whose metabolism starts with a C1 or C2 (or rarely a C3) compound and makes a C1 product, natural gas (CH4).
Check Your Understanding
If acetogens conserve energy using the Rnf complex, then what is the purpose of the reductive CoA pathway?
What is electron bifurcation and what role does it play in acetogens?
14.15 Methanogenesis
The biological production of methane—methanogenesis—is catalyzed by a diverse assemblage of strictly anaerobic Archaea called the methanogens. These organisms are common in freshwater sediments and wetlands (Figure 14.35), the guts of many animals including humans, sewage sludge digesters (Section 22.8), and other bioreactors. The reduction of CO2 by H2 to form CH4 is a major pathway of methanogenesis and is a form of anaerobic respiration. We consider the basic properties, phylogeny, and taxonomy of the methanogens in Section 17.2. Here we focus on their bioenergetics and unique biochemistry.
Figure 14.35 Methanogenesis.

Methane is collected in a funnel from swamp sediments where it was produced by methanogens and then ignited in a demonstration experiment at Woods Hole, Massachusetts (USA).
Mastering Microbiology
Art Activity: Figure 14.47 Methanogenesis from CO2 plus H2
C1 Carriers in Methanogenesis
Methanogenesis from CO2 requires eight electrons, and these electrons are added two at a time. This leads to intermediary oxidation states of the carbon atom from +4 (CO2) to −4 (CH4). Several novel coenzymes participate in methanogenesis and can be divided into two classes: (1) those that carry the C1 unit along its path of enzymatic reduction (C1 carriers) and (2) those that donate electrons (redox coenzymes) (Figure 14.36). We consider the C1 carriers first.
Figure 14.36 Coenzymes of methanogenesis.

The atoms shaded in brown or yellow are the sites of oxidation–reduction reactions (brown in F420 and CoB) or the position to which the C1 moiety is attached during the reduction of CO2 to CH4 (dark yellow in methanofuran, methanopterin, and coenzyme M). The same colors used to highlight a particular coenzyme name (CoB is orange, for example) are also used in Figures 14.38 and 14.39 to follow the reactions in each figure. Coenzyme F430 participates in the terminal step of methanogenesis catalyzed by the enzyme methyl reductase, with the methyl group binding to Ni+ in F430 prior to its reduction to CH4.
The coenzyme methanofuran (MF) is required for the first step of methanogenesis. Methanofuran contains the five-membered furan ring and an amino nitrogen atom that binds CO2 (Figure 14.36a). Methanopterin (MP, Figure 14.36b, also called tetrahydromethanopterin) is a methanogenic coenzyme that resembles the vitamin folic acid and plays a role analogous to that of tetrahydrofolate (see Figure 14.34) by carrying the C1 unit in the intermediate steps of CO2 reduction to CH4. Coenzyme M (CoM) (Figure 14.36c) is required for the terminal step of methanogenesis, the reduction of the methyl group (−CH3) to CH4. Although not a C1 carrier, the nickel (Ni+)-containing tetrapyrrole coenzyme F430 (Figure 14.36d) also participates in the terminal step of methanogenesis as part of the methyl reductase enzyme complex (discussed later).
Redox Coenzymes
The coenzymes F420 and 7-mercaptoheptanoylthreonine phosphate (also called coenzyme B, CoB) are electron donors in methanogenesis. Coenzyme F420 (Figure 14.36e) is a flavin derivative, structurally resembling the flavin coenzyme FMN (Figure 3.15). F420 participates in methanogenesis as the electron donor in several steps of CO2 reduction (see Figure 14.38). Coenzyme F420 takes its name from the fact that its oxidized form absorbs light at 420 nm and fluoresces blue-green. Such fluorescence is useful for the microscopic identification of a methanogen (Figure 14.37). CoB is required for the terminal step of methanogenesis catalyzed by the methyl reductase enzyme complex. As shown in Figure 14.36f, the structure of CoB resembles the vitamin pantothenic acid, which is part of acetyl-CoA (Figure 3.8).
Figure 14.37 Fluorescence due to the methanogenic coenzyme F420.

(a) Autofluorescence in cells of the methanogen Methanosarcina barkeri due to the presence of the unique electron carrier F420. A single cell is about 1.7 μm in diameter. The organisms were made visible by excitation with blue light in a fluorescence microscope. (b) F420 fluorescence in cells of the methanogen Methanobacterium formicicum. A single cell is about 0.6 μm in diameter.
Methanogenesis from CO2+H2
Nearly all methanogens can use H2 as the source of electrons for the reduction of CO2 to CH4. Some can also use a few additional substrates including acetate, formate, methanol, and methylamines. Methanogens use the reductive acetyl-CoA pathway (Sections 14.2 and 14.14) to assimilate CO2 into cellular material. With respect to energy conservation, two major groups of methanogens are differentiated by the presence or absence of cytochromes. We will first consider methanogenesis from H2 and CO2 in methanogens that lack cytochromes (Figure 14.38):
CO2 is activated by a methanofuran-containing enzyme and reduced to the formyl (CHO) level. The immediate electron donor is ferredoxin (Fdred), a strong reductant with a reduction potential (E0′) of about −0.45 V.
The formyl group is transferred from methanofuran (MF) to an enzyme containing methanopterin (MP). It is subsequently dehydrated and reduced in two separate steps (total of 4 H) to the methylene and methyl levels. The 2 H required for each of these reductions are derived from H2 and transferred by F420.
The methyl group is transferred from MP to an enzyme containing CoM by the enzyme methanopterin:coenzyme M methyltransferase. This highly exergonic reaction results in energy conservation because it drives 2 Na+ across the membrane, resulting in the formation of a sodium motive force.
Methyl-S-CoM is reduced to CH4 by methyl-coenzyme M reductase. In this reaction, F430 and CoB are required. Coenzyme F430 removes the CH3 group from CH3-S-CoM, forming a Ni+−CH3 complex. This complex is reduced by CoB, generating CH4 and a heterodisulfide bond between CoM and CoB (CoM-S—S-CoB).
Free CoM and CoB are regenerated by the reduction of CoM-S—S-CoB (E0′=−0.14) catalyzed by heterodisulfide reductase through a flavin-based electron-bifurcation reaction (see red-shaded region in Figure 14.38; see also Figure 14.2) with H2 (−0.42 V) as the electron donor. The high reduction potential of CoM-S—S-CoB is used to drive electrons in the bifurcation reaction to reduce ferredoxin. The Fdred so generated is then used for CO2 reduction in the first step of the pathway.
Figure 14.38 Methanogenesis from CO2 plus H2.

The carbon atom reduced is highlighted in green, and the source of electrons is highlighted in brown. See Figure 14.36 for the structures of the coenzymes. Methanogens without cytochromes reduce the heterodisulfide bond in CoM-S-S-CoB using an electron bifurcation reaction (red box and reaction arrow) that also generates reduced ferredoxin (Fdred). Methanogens with cytochromes conserve more energy by using CoM-S-S-CoB as the electron acceptor in electron transport (blue box and arrow). MF, methanofuran; MP, methanopterin; CoM, coenzyme M; F420H2, reduced coenzyme F420; F430, coenzyme F430; Fd, ferredoxin; CoB, coenzyme B; M, methanophenazine.
Under standard conditions, the free energy of methanogenesis from H2+CO2 is −131 kJ/mol; however, the free energy available under natural conditions is likely in the range of −17 to −40 kJ/mol. In methanogens that lack cytochromes, a sodium motive force drives ATP formation by activity of a Na+-linked ATPase. The heterodisulfide reductase reaction produces a net of two H+ in the cytoplasm and these are exchanged for one Na+ by an H+: Na+ antiporter in the membrane (Figure 14.38). Hence, the production of one CH4 molecule from H2+CO2 yields a net of one Na+ ion pumped across the membrane, sufficient to form less than 0.5 ATP.
In methanogens containing cytochromes, H2 is oxidized at a membrane-bound hydrogenase containing b-type cytochromes. This enzyme passes electrons to a membrane-associated electron carrier called methanophenazine (M, highlighted in blue in Figure 14.38). Methanophenazine in turn passes electrons to a membrane-bound heterodisulfide reductase complex that reduces CoM-S—S-CoB (Figure 14.38). The net result of these electron transport reactions is the generation of a pmf. Methanogens with cytochromes have H+-linked ATPases in place of the Na+-fueled ATPases found in many other methanogens. Since Fdred is still required for CO2 reduction, some of the pmf drives Fdred formation using electrons from H2 (Figure 14.38). The net result is that methanogens with cytochromes conserve more energy than those without, forming >1ATP per molecule of CH4 formed from H2+CO2.
In methanogens we thus see at least two mechanisms for energy conservation that have different energy yields: (1) a sodium motive force (which can be converted to a proton motive force) linked to the methyl transferase reaction, and (2) a proton motive force linked to cytochrome-dependent electron transport reactions. In addition to these energetic differences, cytochrome-containing methanogens are more metabolically versatile than those that lack cytochromes, with many species able to metabolize acetate, methanol, and other methylated compounds, as we see next.
Methanogenesis from Methyl Compounds and Acetate
Cytochrome-containing methanogens, such as Methanosarcina, can form CH4 from acetate and certain methylated compounds, as well as from H2+CO2. Methyl compounds such as methanol are catabolized by donating methyl groups to an enzyme containing a corrinoid coenzyme to form CH3–corrinoid. Corrinoids are the parent structures of compounds such as vitamin B12 and contain a porphyrin-like ring with a central cobalt atom. The CH3–corrinoid complex then transfers the methyl group to CoM, yielding CH3-S-CoM (**Figure 14.39*a***). Energy is then conserved during electron transport reactions in the same manner as for growth on H2+CO2 (Figure 14.38). However, in the absence of H2, the reducing power for driving electron transport reactions comes from running the methanogenic pathway in reverse from CH3-S-CoM to produce CO2, one molecule of Fdred, and two molecules of F420H2. These reduced F420H2 molecules can donate electrons to the electron transport chain in place of H2 (Figure 14.39a). In addition, the Fdred can be oxidized by an energy-converting hydrogenase (see Section 14.12) to produce H2 (which can donate electrons to the electron transport chain along with F420H2) and contribute further to pmf formation. The net result for growth on methanol is that one molecule of methanol must be oxidized to CO2 for every three reduced to CH4.
Figure 14.39 Methanogenesis from methanol and acetate.

Both reactions funnel carbon into the methanogenic pathway (Figure 14.38) but at different points. (a) For growth on methanol (CH3OH), redox balance is achieved because one CH3OH is oxidized to CO2 for every 3 reduced to CH4. (b) Acetate is split into CH4 and CO2. Abbreviations and color-coding are as in Figures 14.36 and 14.38; CODH, carbon monoxide dehydrogenase.
When acetate is the substrate for methanogenesis by acetate-degrading methanogens (called acetoclastic methanogens), it is first activated to acetyl-CoA, which interacts with CO dehydrogenase from the acetyl-CoA pathway (Section 14.14 and Figure 14.34). The methyl group of acetate is then transferred to the corrinoid enzyme, ultimately forming CH3-MP (Figure 14.39b). Formation of CH4 from CH3-MP proceeds as in the H2+CO2 pathway (Figure 14.38), resulting in the formation of both a Na+ and a H+ motive force. Simultaneously, the CO group is oxidized to yield CO2 and Fdred (Figure 14.39b). The reduced ferredoxin can be oxidized by an energy-converting hydrogenase (see Figure 14.43) to produce H2 and contribute further to pmf formation.
Autotrophy
Autotrophy in methanogens is supported by the reductive acetyl-CoA pathway (Sections 14.2 and 14.14). As we have just seen, parts of this pathway are already integrated into the catabolism of methanol and acetate by methanogens that contain cytochromes (Figure 14.39). However, methanogens lack the tetrahydrofolate-driven series of reactions of the acetyl-CoA pathway that lead to the production of a methyl group (Figure 14.34). But this is not a problem because methanogens produce methyl groups during methanogenesis from H2 and CO2. In addition, methylotrophic methanogens derive the required methyl groups directly from their electron donors. Finally, acetoclastic methanogens have no need for CO2 fixation because acetyl-CoA, which is formed during acetate oxidation (Figure 14.39b), can be funneled directly into biosynthesis.
Check Your Understanding
Why is it that methanogens lacking cytochromes are unable to form CH4 from methyl compounds?
Explain why methanogens that contain cytochromes can conserve more energy than those that lack cytochromes.
Explain how a reaction that produces a free energy yield of less than −31.8 kJ/mol, and produces a net translocation of just a single Na+ ion, is enough to drive the formation of ATP.
14.16 Methanotrophy
Methane (CH4) and many other C1 organic compounds can be catabolized both aerobically and anaerobically. In this section, we consider the oxidation of these compounds by methylotrophs, organisms that use organic compounds that lack C—C bonds as electron donors and carbon sources. The oxidations of CH4 and methanol (CH3OH) have been the best-studied reactions, and we focus here on the oxidation of CH4 as an example of a methylotrophic lifestyle.
Aerobic Methane Oxidation
Not all methylotrophs can use methane. Methanotrophs are those methylotrophs that can use CH4, and methanotrophy has been well studied in aerobic methanotrophs such as the gram-negative bacterium Methylococcus capsulatus. The steps in CH4 oxidation to CO2 can be summarized as CH4→CH3OH→CH2O→HCOO−→CO2
The initial step in the aerobic oxidation of CH4 is catalyzed by the enzyme methane monooxygenase (MMO). Monooxygenases incorporate one oxygen atom from O2 into a carbon compound (see Section 14.23 and Figure 14.54a). M. capsulatus contains two MMOs, one cytoplasmic (soluble MMO, sMMO) and the other membrane-integrated (particulate MMO, pMMO). In the MMO reaction, an atom of oxygen is introduced into CH4, forming CH3OH, and the second atom of O is reduced to form H2O. CH3OH is oxidized to formaldehyde (CH2O) and then to CO2, yielding NADH that can be used to conserve energy in electron transport to O2. In addition, formaldehyde can be siphoned off to provide carbon for biosynthesis through one of two different pathways.
C1 Assimilation by Aerobic Methanotrophs
At least two distinct pathways exist for the incorporation of C1 units into cell material in methanotrophs. The serine pathway is outlined in **Figure 14.40*a***. In this pathway, acetyl-CoA is synthesized from one molecule of CH2O and one molecule of CO2. The serine pathway requires reducing power and energy in the form of two molecules each of NADH and ATP, respectively, for each acetyl-CoA synthesized. The serine pathway employs a number of enzymes of the citric acid cycle and one enzyme, serine transhydroxymethylase, unique to the pathway (Figure 14.40a).
Figure 14.40 The serine and ribulose monophosphate pathways for the assimilation of C1 units into cell material by methylotrophic bacteria.

(a) Serine pathway. The product, acetyl-CoA, is used as the starting point for making new cell material. The key enzyme of the pathway is serine transhydroxymethylase. (b) Ribulose monophosphate pathway. Three molecules of CH2O are required, with the product being glyceraldehyde 3-phosphate. The key enzyme of this pathway is hexulosephosphate synthase. The sugar rearrangements require enzymes of the pentose phosphate pathway (Figure 3.31).
An alternative pathway for C1 incorporation is the ribulose monophosphate pathway (Figure 14.40b). This pathway is more energy efficient than the serine pathway because all of the carbon for cell material is derived from CH2O. Because CH2O is at the same oxidation level as cell material, no reducing power is needed for its incorporation. Hence, all of the NADH from the oxidation of methane can be oxidized in the electron transport chain.
The ribulose monophosphate pathway consumes one molecule of ATP for each molecule of glyceraldehyde 3-phosphate (G-3-P) synthesized (Figure 14.40b); two G-3-Ps can then be converted into glucose by reversal of the glycolytic pathway (Figure 3.11). Enzymes unique to the ribulose monophosphate pathway are hexulosephosphate synthase, which condenses one molecule of formaldehyde with one molecule of ribulose 5-phosphate, and hexulose 6-P isomerase, which forms fructose derivatives that can be converted into glyceraldehyde 3-P (Figure 14.40b). The remaining enzymes of this pathway are enzymes of intermediary metabolism widely distributed in bacteria.
Anaerobic Oxidation of Methane (AOM)
The anaerobic oxidation of methane uses a variety of enzymes and cofactors common to other forms of C1 metabolism; however, anaerobic methanotrophy shows some novel features, as well. Methane can be oxidized anaerobically by a microbial association (called a consortium) of two organisms, a sulfate-reducing bacterium (SRB) plus a species of Archaea phylogenetically related to methanogens. These consortia thrive in anoxic marine sediments and are responsible for oxidizing more than 90% of the methane produced there. The components of the consortium coexist in spatially structured aggregates (Figure 14.41). The archaeal component, called ANME (anaerobic methanotroph), of which there are several different types, oxidizes CH4 as an electron donor. Electrons from methane oxidation are then transferred to the sulfate reducer, which uses them to reduce SO4 2− to H2S (Figure 14.41b).
Figure 14.41 Anaerobic oxidation of methane.

(a) Methane-oxidizing cell aggregates from marine sediments. The aggregates contain methane-oxidizing (methanotrophic) Archaea (red) surrounded by sulfate-reducing bacteria (green). Each cell type has been stained by a different FISH probe (Section 19.5). The aggregate is about 30 μm in diameter. (b) Mechanism for the cooperative degradation of CH4. Electrons are transferred from the archaeal methanotrophic (ANME) cell to the sulfate reducer by direct electron transfer through the “nanowire” connections. (c) Transmission electron micrograph of a section through a methane-oxidizing aggregate showing an ANME cell transferring electrons to a sulfate reducer.
ANME Archaea oxidize CH4 to CO2 by reversing the steps of methanogenesis (Figure 14.38). This process is endergonic but is made possible by the SRB partner organism, which consumes electrons from ANME, thereby making the oxidation CH4 to CO2 energetically favorable. Remarkably, electrons are transferred between the ANME and SRB partners by direct electron transfer (Figure 14.41b, c). Cells of ANME make electrically conductive multiheme cytochromes that span their outer cell layer and transfer electrons from the cytoplasmic membrane to the outside of the cell. The SRB have similar large electrically conductive cytochromes as well as pili that serve as electrically conductive “nanowires” (Section 14.13). These pili can be more than 1 μm in length and electrically connect the cells within aggregates to facilitate the two microbes’ metabolisms (Figure 14.41b, c; Section 23.3). Such direct electron transfer likely explains why some ANME can also use insoluble metals such as iron (Fe3+) and manganese oxides (Mn4+) as terminal electron acceptors (Section 14.13) in CH4 oxidation. Since these insoluble materials cannot enter the cell, the cell has to “reach out” to them in order to transfer electrons.
AOM is not limited to consortia of ANME and SRB. ANME Archaea include Methanoperedens nitroreducens, which uses nitrate (NO3 −) as a terminal electron acceptor for the anaerobic oxidation of CH4. This organism couples reverse methanogenesis to the reduction of NO3 − to NO2 −. M. nitroreducens can be found in consortia with denitrifying bacteria that then use NO2 − as an electron acceptor. These consortia are active in anoxic environments where CH4 and NO3 − coexist, such as certain freshwater sediments.
Intra-Aerobic Methanotrophy
The methanotrophic denitrifying bacterium Methylomirabilis oxyfera is an obligate anaerobe that catalyzes AOM linked to NO2 − as an electron acceptor. M. oxyfera can grow on CH4 in pure culture and has a highly unusual polygonal morphology (Figure 14.42). Analysis of the M. oxyfera genome reveals all of the genes required for the aerobic oxidation of CH4 to CO2. M. oxyfera is also able to reduce NO2 − to N2, and it has most of the genes required for denitrification although it lacks nitric oxide reductase and nitrous oxide reductase (Section 14.11 and Figure 14.28). This leads to the interesting question of why an anaerobic bacterium would use aerobic pathways for CH4 oxidation.
Figure 14.42 The cell morphology of *Methylomirabilis oxyfera*.

The denitrifying methanotroph M. oxyfera has a unique polygonal morphology as revealed in this transmission electron micrograph of cells from a microbial community grown in a bioreactor. A cell is about 0.5 μm in diameter.
Mastering Microbiology
Art Activity: Figure 14.54 The essentials of fermentation
The answer to this conundrum is that M. oxyfera reduces nitrite in a novel way. The organism reduces NO2 − to nitric oxide (NO) as a normal denitrifying bacterium would, but then M. oxyfera does something remarkable: The organism generates O2 through the reaction 2 NO→N2+O2 and proceeds to use this O2 as the electron acceptor for CH4 oxidation. Because M. oxyfera produces its own O2, its methanotrophic metabolism has been termed intra-aerobic methanotrophy. As it turns out, O2 is toxic for the anaerobic M. oxyfera. However, if the O2 is consumed (by its reduction to H2O using electrons from the oxidation of CH4) as soon as it is produced, O2 never accumulates, and the organism’s environment remains anoxic.
We transition to Part VI now and consider metabolisms in which ATP is not a product of electron transport reactions. These are the fermentations—metabolisms in which there is no lack of physiological diversity.
Check Your Understanding
When using CH4 as electron donor, why is Methylococcus capsulatus an obligate aerobe?
In which two ways does the ribulose monophosphate pathway save energy over reactions of the serine pathway?
What is unique about methanotrophy in Methylomirabilis oxyfera?
VI Fermentation
Fermentations are anaerobic metabolisms that do not require external electron acceptors. Fermenting microbes achieve redox balance and conserve energy by forming various fermentation products that are excreted from the cell.
Thus far we have considered phototrophy and many types of aerobic and anaerobic respiration; these processes are unified by the fact that they all conserve energy primarily by using an ion gradient (H+ or Na+) that fuels photophosphorylation or oxidative phosphorylation. Here we turn our focus to fermentation, in which energy conservation depends primarily upon substrate-level phosphorylation reactions.
14.17 Energetic and Redox Considerations
Fermentation is a form of chemotrophic metabolism defined by the lack of an external electron acceptor. In fermentation, cells conserve energy primarily by substrate-level phosphorylation and they achieve redox balance by donating electrons to metabolic intermediates excreted as fermentation products. Fermentative organisms face two major challenges. First, their reliance on substrate-level phosphorylation means that they typically conserve much less energy than respiratory organisms. Second, their lack of external electron acceptors makes it difficult to achieve redox balance.
A tremendous diversity of fermentation reactions are possible. Many fermentative organisms, such as Escherichia coli, perform diverse metabolic reactions and only ferment when external electron acceptors are absent and conditions are anoxic. However, there are many more organisms, such as most lactic acid bacteria and the clostridia, which have exclusively fermentative metabolisms. Over the next few sections, we will see that fermentative organisms have evolved diverse strategies for facing the challenges of energy scarcity and redox balance.
Energy-Rich Compounds and Substrate-Level Phosphorylation
Energy can be conserved by substrate-level phosphorylation from many different compounds. However, as we learned in Chapter 3, central to an understanding of substrate-level phosphorylation is the concept of energy-rich compounds (Figure 3.8). These are organic compounds that contain an energy-rich phosphate bond or a molecule of coenzyme A. The bond is “energy-rich” because its hydrolysis is highly exergonic. Table 14.5 lists some energy-rich compounds formed during metabolism; the hydrolysis of most of these yields sufficient free energy to be coupled to ATP synthesis (ΔG0′=−31.8 kJ/mol). If an organism can form one of these compounds during fermentative metabolism, it can make ATP by transferring the phosphate bond from the energy-rich compound to ADP to form ATP— in other words, substrate-level phosphorylation.
Table 14.5 Energy-rich compounds that can couple to substrate-level phosphorylationa

aData from Thauer, R.K., K. Jungermann, and K. Decker. 1977. Energy conservation in chemotrophic anaerobic bacteria. Bacteriol. Rev. 41: 100.
b The ΔG0′ values shown here are for “standard conditions,” which are not necessarily those of cells. Including heat loss, the energy costs of making an ATP are more like −60 kJ/mol than −31.8 kJ/mol, and the energy of hydrolysis of the energy-rich compounds shown here is thus likely higher. But for simplicity and comparative purposes, the values in this table will be taken as the actual energy released per reaction.
Many anaerobic bacteria produce acetate or other fatty acids as a major or minor fermentation product. The production of fatty acids offers the organism the opportunity to conserve energy by substrate-level phosphorylation. The key to this process is the production of a coenzyme-A derivative of the fatty acid, since these are energy-rich compounds (Table 14.5). For example, acetyl-CoA can be converted to acetyl phosphate and the phosphate group subsequently transferred to ADP, yielding ATP (see Figures 14.46 and 14.47). Fatty acid production is common in fermentations, and if the fatty acid is metabolized through a CoA intermediate, the potential for ATP synthesis by substrate-level phosphorylation is a possibility.
Achieving Redox Balance
In any fermentation there must be atomic and redox balance. That is, the total number of each type of atom and electrons in the reaction products must balance those in the reactants (the substrates). Redox balance is achieved by the excretion of fermentation products, reduced substances such as acids or alcohols that are made as products of the fermentation. However, many reactions are difficult to balance, and so fermentative organisms have evolved alternative mechanisms for achieving redox balance.
In many fermentations, redox balance is facilitated by the production of molecular hydrogen (H2). Fermentative organisms can produce H2 in several ways (Figure 14.43). First, H2 can be produced from the C1 fatty acid formate using the enzyme formate hydrogenlyase. This mechanism is the source of fermentative H2 production used by many facultative aerobes including most enteric bacteria such as E. coli. Second, H2 can be produced from proton reduction using the iron–sulfur protein ferredoxin (Fdred), in a reaction catalyzed by a hydrogenase enzyme. Many organisms perform this reaction using a membrane-bound energy-converting hydrogenase because this enzyme couples H2 production to proton pumping, thereby contributing to pmf formation and energy conservation (Figure 14.43).
Figure 14.43 Production of H2 to achieve redox balance during fermentation.

Several mechanisms exist for reducing protons to H2, thereby allowing the cell to achieve redox balance by exporting electrons. Energy-converting hydrogenases allow for coupling of H2 production with energy conservation by pumping an H+ across the membrane to generate a proton-motive force.
Finally, some organisms produce H2 using electron confurcation (Figure 14.43, see also Section 14.1). The advantage of electron confurcation is that it provides the ability to regenerate NAD+ by producing H2 from NADH at low energetic cost. Consider that ΔG0′=+17 kJ/mol for the confurcation reaction producing two H2, while ΔG0′=+20 kJ/mol for the direct coupling of NADH to H2 production; hence, confurcation allows the cell to produce two H2 for less energy than it takes to produce one H2 from the oxidation of NADH by reverse electron transport.
Fermentative organisms excrete H2 to achieve redox balance by reoxidizing electron carriers such as NAD(P)H and Fdred. The four mechanisms for H2 production (Figure 14.43) all have ΔG0′ very close to zero under standard conditions. However, fermentation can produce copious amounts of NADH and Fdred that would accumulate in the cell if these electron carriers were not reoxidized. Under actual physiological conditions, then, these H2 production reactions are usually slightly exergonic, and in the case of energy-converting hydrogenases (Figure 14.43) can even be used to conserve energy. Another factor to consider is that H2, excreted as a waste product by fermentative organisms, is a very powerful electron donor for respiratory organisms, and this reality typically keeps H2 levels in anoxic environments extremely low.
With H2 we thus see that the waste product of one organism becomes a resource for another. This metabolic coupling, in which one organism’s waste is another organism’s resource, is a theme that runs throughout the microbial world. With these bioenergetic and redox principles in mind, we explore the metabolic diversity of fermentations beginning with species that produce acidic fermentation products, common and widespread bacteria in most anoxic environments.
Check Your Understanding
Why is H2 produced during many types of fermentation?
Why is acetate formation in fermentation energetically beneficial to the cell?
What is an energy-converting hydrogenase, and what role does it play in achieving redox balance?
14.18 Lactic and Mixed-Acid Fermentations
Fermentations are categorized by either the substrate fermented or the products formed. Table 14.6 lists some major fermentations classified on the basis of the products formed, such as alcohol, lactic acid, or other reduced organic compounds. Other fermentations are classified by the substrate fermented rather than the fermentation product; for instance, the amino acid, purine/pyrimidine, or succinate fermentations. We begin with two very common fermentations of sugars in which lactic acid is the sole or major product.
Table 14.6 Common fermentations and their energetics and example organisms

Lactic Acid Fermentation
Lactic acid bacteria are gram-positive nonsporulating bacteria that produce lactic acid as a major or sole fermentation product from the fermentation of sugars (Section 16.6). Two fermentative patterns are observed. One, called homofermentative, yields a single fermentation product, lactic acid. The other, called heterofermentative, yields products in addition to lactate, mainly ethanol plus CO2.
Figure 14.44 summarizes pathways for the fermentation of glucose by homofermentative and heterofermentative lactic acid bacteria. The differences observed can be traced to the presence or absence of the enzyme aldolase, a key enzyme of glycolysis (Figure 3.11). Homofermentative lactic acid bacteria contain aldolase and produce two molecules of lactate from glucose by the glycolytic pathway (Figure 14.44a). Heterofermenters lack aldolase and thus cannot break down fructose bisphosphate to triose phosphate. Instead, they oxidize glucose 6-phosphate to 6-phosphogluconate and then decarboxylate this to ribulose 5-phosphate and on to xylulose 5-phosphate. The latter is then converted to triose phosphate and acetyl phosphate by the key enzyme phosphoketolase (Figure 14.44b). The early steps in catabolism by heterofermentative lactic acid bacteria are those of the pentose phosphate pathway (Figure 3.31).
Figure 14.44 The fermentation of glucose in *(a)* homofermentative and *(b)* heterofermentative lactic acid bacteria.

Note that no ATP is made in reactions leading to ethanol formation in heterofermentative organisms.
In heterofermenters, triose phosphate is converted to lactic acid with the production of ATP (Figure 14.44b). However, to achieve redox balance the acetyl phosphate produced is used as an electron acceptor and is reduced by NADH (generated during the production of pentose phosphate) to ethanol. This occurs without ATP synthesis because the energy-rich CoA bond is lost during ethanol formation. Because of this, heterofermenters produce only one ATP per glucose instead of the two ATP per glucose produced by homofermenters. In addition, because heterofermenters decarboxylate 6-phosphogluconate, they produce CO2 as a fermentation product; homofermenters do not produce CO2. Thus, an easy way to differentiate a homofermenter from a heterofermenter is to observe for the production of CO2 in laboratory cultures.
Entner–Doudoroff Pathway
The initial stages of glucose fermentation often rely on glycolysis (Figure 3.11) or a variant of the glycolytic pathway called the Entner–Doudoroff pathway (this variation is shown in Figure 14.44b). In the Entner–Doudoroff pathway, glucose 6-phosphate is oxidized to 6-phosphogluconic acid and NADPH; the 6-phosphogluconic acid is dehydrated and split into pyruvate and glyceraldehyde 3-phosphate (G-3-P), a key intermediate of the glycolytic pathway. G-3-P is then catabolized as in glycolysis, generating NADH and two ATP, and used as an electron acceptor to balance redox reactions as occurs in both lactic acid fermentations (Figure 14.44).
Because pyruvate is formed directly in the Entner–Doudoroff pathway and cannot yield ATP as can G-3-P (Figure 14.44), the Entner–Doudoroff pathway yields only half the ATP of the glycolytic pathway. Organisms using the Entner–Doudoroff pathway therefore share this physiological characteristic with heterofermentative lactic acid bacteria (Figure 14.44b). Zymomonas, an obligately fermentative gram-negative bacterium, and Pseudomonas, a strictly respiratory bacterium (Section 16.4), are major genera that employ the Entner–Doudoroff pathway for glucose catabolism, although a modified version of this pathway is present in some species of hyperthermophilic and extremely halophilic Archaea.
Mixed-Acid Fermentations
In mixed-acid fermentations (Table 14.6), characteristic of enteric bacteria such as Escherichia coli and relatives (Section 16.3), three different acids—acetic, lactic, and succinic—are formed from the fermentation of glucose or other sugars that can be converted into glucose. Ethanol, CO2, and H2 are also typically formed as fermentation products. Glycolysis (Figure 3.11) is the pathway used to degrade glucose by mixed-acid fermenters.
Some enteric bacteria produce acidic products in lower amounts than E. coli and balance redox in their fermentations by producing larger amounts of neutral products. One key neutral product is the four-carbon alcohol butanediol. In this variation of the mixed-acid fermentation, the main products observed are butanediol, ethanol, CO2, and H2 (Figure 14.45). In the mixed-acid fermentation of E. coli, equal amounts of CO2 and H2 are produced, whereas in a butanediol fermentation, considerably more CO2 than H2 is produced. This is because mixed-acid fermenters produce CO2 only from formic acid by means of the enzyme formate hydrogenlyase (Figure 14.45, see also Figure 14.43): HCOOH→H2+CO2
Figure 14.45 Butanediol production and mixed-acid fermentations.

Note how only one NADH but two molecules of pyruvate are used to make one butanediol. This leads to redox imbalance and the production of more ethanol by butanediol producers than by mixed-acid fermenters. A yellow highlight is used to show the path that two of the four pyruvate carbons take to end up in α-acetolactate.
By contrast, butanediol producers, such as Enterobacter aerogenes, produce CO2 and H2 from formic acid but also produce two additional molecules of CO2 during the formation of each molecule of butanediol. However, because butanediol production consumes only one-half of the NADH generated in glycolysis, more ethanol is produced by these organisms than by non–butanediol fermenters in order to achieve redox balance (Figure 14.45).
Lactic acid and enteric bacteria are facultative, meaning that they can grow either aerobically or anaerobically. By contrast, some fermenters are obligate anaerobes and we explore their fermentation patterns now.
Check Your Understanding
How can homo- and heterofermentative metabolism be differentiated in pure cultures of lactic acid bacteria?
Why does butanediol production lead to greater ethanol production than the mixed-acid fermentation of Escherichia coli?
How and why does E. coli produce H2 during fermentation?
14.19 Fermentations of Obligate Anaerobes
Many fermentative organisms are obligate anaerobes. These organisms grow under highly reducing conditions, often use enzymes that cannot tolerate O2, and often produce H2 as a fermentation product. Species of Clostridium (Section 16.8) and their relatives are the most well known of this group. Different species of clostridia ferment different groups of substrates, including sugars, amino acids, purines and pyrimidines, and a few other compounds. In all cases, ATP synthesis is linked primarily to substrate-level phosphorylation. However, we will see that many obligately anaerobic fermentative organisms can also conserve some energy through the generation of pmf. We begin with saccharolytic (sugar-fermenting) clostridia.
Sugar Fermentation by *Clostridium* Species
A number of clostridia ferment sugars, producing butyric acid and H2 as major fermentation products. A generalized scheme for the biochemical steps in the formation of butyric acid and neutral products from sugars is shown in Figure 14.46. In saccharolytic clostridia, such as the well-studied Clostridium pasteurianum, 1.5 glucose is converted to 3 pyruvate and 3 NADH via the glycolytic pathway, and each pyruvate is split to yield acetyl-CoA, CO2, and Fdred (the “phosphoroclastic reaction” in Figure 14.46). One acetyl-CoA gives rise to acetate and an ATP and the other two are combined and reduced to butyrate to form another ATP. One NADH is reoxidized during the formation of β-hydroxybutyryl-CoA and the other 2 NADH are oxidized in an electron bifurcation reaction (Figure 14.2) that couples the exergonic reduction of crotonyl-CoA to the endergonic reduction of Fdox. Redox balance is achieved ultimately by a cytoplasmic hydrogenase that uses Fdred to reduce protons to H2 (Figure 14.43).
Figure 14.46 The butyric acid and butanol/acetone fermentation.

All fermentation products from glucose are shown in bold (dashed lines indicate minor products). Note how the production of acetate and butyrate lead to additional ATP by substrate-level phosphorylation. By contrast, formation of butanol and acetone reduces the ATP yield because the butyryl-CoA to butyrate step is bypassed. The production of butyryl-CoA is an electron bifurcation reaction (Figure 14.2) in which 2 NADH donate electrons to crotonyl-CoA and Fdox. Fdred, reduced ferredoxin; Fdox, oxidized ferredoxin.
The actual fermentation products observed differ between species, and the ratio of these products is influenced by environmental conditions. Clostridium acetobutylicum provides a classic example of this pattern. During the early stages of the butyric fermentation, C. acetobutylicum produces mostly butyrate and a small amount of acetate, similar to that of C. pasteurianum. But the production of butyrate and acetate leads to acid production (Figure 14.46), and acidification of the environment eventually inhibits growth of C. acetobutylicum. Hence, as the pH of the medium drops, the organism shifts its fermentation products toward acetone and butanol, “neutral products” that do not lower the pH of the environment. However, if the pH of the medium is kept neutral by buffering, butyric acid production continues, and very little acetone and butanol will be formed because butyrate formation provides a higher yield of ATP (Figure 14.46).
Interestingly, the production of butanol is a consequence of the production of acetone. For each acetone produced, two NADH cannot be reoxidized in the reduction of crotonyl-CoA. To achieve redox balance, the cell then uses butyrate as an electron acceptor, producing butanol as a fermentation product (Figure 14.46). Although neutral product formation helps C. acetobutylicum keep its environment from becoming too acidic, there is an energetic price to pay.
Amino Acid Fermentation by Clostridia and the Stickland Reaction
Some clostridia ferment amino acids. These are the proteolytic clostridia, organisms that degrade proteins released from dead organisms. Some of these, such as the animal pathogen Clostridium tetani (tetanus), are strictly proteolytic, while other species are both saccharolytic and proteolytic.
Depending on the species, some proteolytic clostridia ferment individual amino acids, typically glutamate, glycine, alanine, cysteine, histidine, serine, or threonine. The biochemistry behind these fermentations is quite complex, but the metabolic strategy is simple. In virtually all cases, the amino acids are catabolized in such a way as to eventually yield a fatty acid–CoA derivative, typically acetyl (C2), butyryl (C4), or caproyl (C6). From these, ATP is produced by substrate-level phosphorylation (Table 14.5). Other typical products of amino acid fermentation include ammonia (NH3) and CO2.
Some clostridia ferment only an amino acid pair. In this situation one amino acid functions as the electron donor and is oxidized, whereas the other amino acid is the electron acceptor and is reduced. This coupled amino acid fermentation is called a Stickland reaction, named for the scientist who discovered it. For example, Clostridium sporogenes ferments glycine and alanine, and in this reaction, alanine is the electron donor and glycine is the electron acceptor (Figure 14.47). The products of the Stickland reaction are invariably NH3, CO2, and a carboxylic acid with one fewer carbons than the amino acid that was oxidized (Figure 14.47).
Figure 14.47 The Stickland reaction.

This example shows the co-catabolism of the amino acids alanine and glycine. The structures of key substrates, intermediates, and products are shown in brackets to allow the chemistry of the reaction to be followed. Note that in the reaction shown, alanine is the electron donor and glycine is the electron acceptor.
Many of the products of amino acid fermentation by clostridia are foul-smelling substances, and the odor that results from putrefaction is mainly a result of clostridial activity. In addition to fatty acids, other odoriferous compounds produced in proteolytic fermentations include hydrogen sulfide (H2S), methyl mercaptan (CH3SH, derived from sulfur-containing amino acids), cadaverine (from lysine), putrescine, and NH3. Purines and pyrimidines (Figure 3.34), released from the degradation of nucleic acids, lead to many of the same fermentation products and also yield ATP by substrate-level phosphorylation from the hydrolysis of fatty acid–CoA derivatives (Table 14.5) produced in their respective fermentative pathways.
Energy-Converting Hydrogenases
Pyrococcus furiosus is a hyperthermophilic species of Archaea that grows optimally at 100 °C (Section 17.4) by fermenting sugars and small peptides. It uses the early reactions of glycolysis (Figure 3.11) to convert glucose into two glyceraldehyde 3-phosphate, but then produces two molecules of 3-phosphoglycerate instead of two 1,3-bisphosphoglycerate, as normally occurs in glycolysis (Figure 14.48). In so doing, P. furiosus forgoes the ability to make ATP from the energy-rich phosphate bonds in 1,3-bisphosphoglycerate and instead forms Fdred. Additional ATP and Fdred are formed in the subsequent conversion of pyruvate to acetate (Figure 14.48). P. furiosus is then able to use an energy-converting hydrogenase (Figure 14.43) to reoxidize Fdred and achieve redox balance, but this hydrogenase also has the ability to translocate one Na+ across the membrane for each proton reduced. A Na+-linked ATPase then uses the sodium motive force to synthesize ATP (Figure 14.48).
Figure 14.48 Modified glycolysis and proton reduction in anaerobic respiration in the hyperthermophile *Pyrococcus furiosus*.

Hydrogen (H2) production is linked to H+ pumping by an energy-converting hydrogenase (see Figure 14.43) that receives electrons from reduced ferredoxin (Fdred). Compare this figure with classical glycolysis in Figure 3.11. G-3-P, glyceraldehyde 3-phosphate; 3-PGA, 3-phosphoglycerate; PEP, phosphoenolpyruvate.
Ultimately, P. furiosus obtains two-thirds (4 ATP) of its energy from substrate-level phosphorylation and one-third (2 ATP) from oxidative phosphorylation driven by proton respiration linked to a Na+ pump. By using this strategy, P. furiosus yields a net of 4 ATP per glucose (Figure 14.48), whereas lactic acid bacteria only obtain 2 ATP per glucose (Figure 14.44). Like P. furiosus, many obligately anaerobic fermentative organisms use energy-converting hydrogenases to achieve redox balance and thereby conserve some energy in the form of a proton or sodium motive force. Is this respiration or is it fermentation? Some metabolic reactions are difficult to classify, but the majority of energy is conserved in substrate-level phosphorylation, a trait that is a defining feature of fermentative metabolisms.
Is fermentation the “end of the line” in terms of the energy extracted from the initial substrate? Not quite, as we see in the final three sections of this part of the chapter.
Check Your Understanding
Why does Clostridium acetobutylicum need to produce butanol as a fermentation product when it is producing acetone?
What type of substrates are fermented by saccharolytic clostridia? By proteolytic clostridia?
What aspects of metabolism in Pyrococcus furiosus are unusual for a fermentative organism?
14.20 Secondary Fermentations
In addition to classifying fermentations by the substrate fermented or product(s) produced (Section 14.18 and Table 14.6), fermentations can be classified as primary or secondary. Primary fermentations are carried out by organisms that break down and ferment various polymers and monomers of carbohydrate, protein, and fat to reduced products including acids and alcohols as well as H2 and CO2. Secondary fermentations use these fermentation products as substrates for additional fermentation reactions whose products are mainly volatile fatty acids (such as butyrate, propionate, and acetate), H2, and CO2.
Propionic Acid Fermentation
The gram-positive bacterium Propionibacterium and some related bacteria produce propionic acid as a major fermentation product from either glucose or lactate. Lactate, a fermentation product of the lactic acid bacteria (Section 14.18), is probably the major substrate for propionic acid bacteria in nature, where these two groups live in close association. Propionibacterium is an important agent in the ripening of Emmental (Swiss) cheese, which gets its unique bitter and nutty taste from the propionic and acetic acids produced; the CO2 produced during the fermentation forms bubbles that leave the characteristic holes (eyes) in the cheese.
Figure 14.49 shows the reactions leading from lactate to propionate. When glucose is the starting substrate, it is first catabolized to pyruvate by the glycolytic pathway. Then pyruvate, produced either from glucose or from the oxidation of lactate, is converted to acetate plus CO2 or carboxylated to form methylmalonyl-CoA; the latter is converted into oxaloacetate and, eventually, propionyl-CoA (Figure 14.49). Propionyl-CoA reacts with succinate in a step catalyzed by the enzyme CoA transferase, producing succinyl-CoA and propionate. This results in a lost opportunity for ATP production from propionyl-CoA (Table 14.5) but avoids the energetic costs of having to activate succinate with ATP to form succinyl-CoA. The succinyl-CoA is then isomerized to methylmalonyl-CoA and the cycle is complete when propionate is formed and CO2 regenerated (Figure 14.49).
Figure 14.49 The propionic acid fermentation of *Propionibacterium*.

Products are shown in bold. The four NADH made from the oxidation of three lactate are reoxidized in the reduction of oxaloacetate and fumarate, and the CoA group from propionyl-CoA is exchanged with succinate during the formation of propionate.
NADH is oxidized in the steps between oxaloacetate and succinate. The reduction of fumarate to succinate (Figure 14.49) is linked to electron transport reactions (Section 14.13) and the formation of a proton motive force; this yields one ATP by oxidative phosphorylation. The propionate pathway also converts some pyruvate to acetate plus CO2, which allows for additional ATP to be made by substrate-level phosphorylation because this reaction proceeds through the energy-rich intermediate acetyl-CoA (Table 14.5 and Figure 14.49). Thus, in the propionate fermentation, both substrate-level and oxidative phosphorylation occur.
Propionate is also formed in the fermentation of succinate by the bacterium Propionigenium, but by a completely different mechanism than that described here for Propionibacterium. Propionigenium, to be considered in Section 14.21, is phylogenetically and ecologically unrelated to Propionibacterium, but aspects of its energy metabolism are of considerable interest from the standpoint of metabolic diversity and the energetic limits to life.
*Clostridium kluyveri* Fermentation
Clostridium kluyveri ferments two substrates: ethanol plus acetate. This is an unusual secondary fermentation in that ethanol is the electron donor and acetate is the electron acceptor, and butyrate, caproate, and H2 are the products. The overall reaction in the caproate/butyrate fermentation is shown in Table 14.6 and begins with 6 ethanol and 3 acetate as substrates. The oxidation of 6 ethanol to 6 acetyl-CoA results in the production of 12 NADH. Then, using the same reactions we saw in butyric acid fermentation (Figure 14.46), one acetyl-CoA is used to produce acetate and ATP by substrate-level phosphorylation. Next, the 3 acetate molecules are activated by adding CoA, giving a total of 8 acetyl-CoA. These molecules are used in the butyric acid pathway to accept electrons from the 12 NADH generated earlier, thereby producing 4 butyryl-CoA (as in Figure 14.46).
As in the butyric acid fermentation, the production of 4 butyryl-CoA is mediated by an electron bifurcation reaction (Figure 14.2) producing 4 Fdred, two of which give rise to H2. However, C. kluyveri, unlike C. pasteurianum, has a membrane-bound NAD+–ferredoxin oxidoreductase (RnfA–G) that uses the exergonic reduction of NAD+ by Fdred to pump protons across the membrane, thereby generating a pmf (Rnf is also found in acetogens, see Figure 14.34). The two remaining Fdred are used by the Rnf enzyme complex to produce 2 NADH and to pump 4 H+ across the membrane, enough to make one ATP by oxidative phosphorylation. Finally, we are left with 2 NADH, 1 acetyl-CoA, and 4 butyryl-CoA. The CoA are transferred from 3 of the butyryl-CoA to activate the 3 acetate at the beginning of the pathway, producing 3 butyrate. Lastly, the 2 remaining NADH are used to make caproate from the final butyryl-CoA and the acetate produced earlier in the pathway, yielding the final fermentation products: 3 butyrate, 1 caproate, and 2 H2.
We can see several themes at work in the fermentation of C. kluyveri. First, we see the modularity of metabolism in the way that C. kluyveri has adapted the butyric acid pathway to perform a butyrate-based fermentation that is distinct from that of C. pasteurianum. Second, we can see how metabolic intermediates are often recycled and reused within fermentative pathways to achieve redox balance. Third, we see that organisms can enhance their metabolic efficiency by using mechanisms such as electron bifurcation and using reduced ferredoxin as electron donor to generate a proton motive force with protons or NAD+ as electron acceptor.
In the final two sections of this part of the chapter, we will see more examples of microbes that employ innovative strategies for living at the thermodynamic edge of life.
Check Your Understanding
What aspects of energy conservation are shared by both Propionibacterium and Clostridium kluyveri?
What is meant by the terms primary and secondary fermentation?
How is the Rnf enzyme complex of Clostridium kluyveri similar to the energy-converting hydrogenase of Pyrococcus furiosus? How are they different?
14.21 Fermentations That Lack Substrate-Level Phosphorylation
14.21 Fermentations That Lack Substrate-Level Phosphorylation
14.21 Fermentations That Lack Substrate-Level Phosphorylation
Certain fermentations yield insufficient energy to synthesize ATP by substrate-level phosphorylation (that is, less than −31.8 kJ, Table 14.5), yet still support anaerobic growth without external electron acceptors. In these cases, catabolism of the compound is linked to ion pumps that establish an ion gradient across the cytoplasmic membrane. Examples of these include the fermentation of succinate by Propionigenium modestum and the fermentation of oxalate by Oxalobacter formigenes.
Propionigenium modestum
Propionigenium modestum was first isolated in anoxic enrichment cultures lacking electron acceptors and fed succinate as an electron donor. Propionigenium inhabits marine and freshwater sediments and can also be isolated from the human oral cavity. The organism is a gram-negative short rod and, phylogenetically, is a species of Fusobacteria. During studies of the physiology of P. modestum, it was shown to require sodium chloride (NaCl) for growth and to catabolize succinate under strictly anoxic conditions: Succinate2−+H2O→propionate−+HCO3−ΔG0′=−20.5 kJ
This is a disproportionation reaction (Section 14.12) in which part of the succinate molecule is reduced to propionate and the other part is oxidized to CO2 (**Figure 14.50*a***). Energy is conserved by an unusual enzyme that decarboxylates (S)-methylmalonyl-CoA and adds a proton to it to produce propionyl-CoA. The enzyme uses the energy released from this decarboxylation reaction to translocate two Na+ across the membrane. Energy is then conserved using a Na+-linked ATPase as we have seen for acetogens (Section 14.14), methanogens (Section 14.15), and Pyrococcus (Section 14.19).
Figure 14.50 The unique fermentations of succinate and oxalate.

(a) Succinate fermentation by Propionigenium modestum. Sodium export is linked to the energy released by succinate decarboxylation, and a sodium-translocating ATPase produces ATP. (b) Oxalate fermentation by Oxalobacter formigenes. Oxalate import and formate export by a formate–oxalate antiporter (Figure 2.6) consume cytoplasmic protons. ATP synthesis is linked to a proton-driven ATPase. All substrates and products of the fermentation are shown in black bold.
In a related decarboxylation reaction, the bacterium Malonomonas rubra decarboxylates the C3 dicarboxylic acid malonate, forming acetate plus CO2. As for Propionigenium, energy metabolism in Malonomonas is linked to Na+ and a sodium-driven ATPase. But the free energy available from malonate fermentation by Malonomonas (−17.4 kJ) is even less than that of succinate fermentation by P. modestum. Sporomusa, an endospore-forming bacterium and also an acetogen (Section 14.14), is also capable of fermenting malonate, as are a few other anaerobic Bacteria.
Oxalobacter formigenes
Oxalobacter formigenes is a bacterium present in the intestinal tract of animals, including humans. It performs a disproportionation reaction using the C2 dicarboxylic acid oxalate, reducing one part of the molecule to formate and oxidizing the other part to CO2 (Figure 14.50b). Oxalate degradation by O. formigenes is thought to be important in the human colon for preventing the accumulation of oxalate, a substance that can form calcium oxalate kidney stones. O. formigenes is a gram-negative strict anaerobe that carries out the following reaction: Oxalate2−+H2O→formate−+HCO3−ΔG0′=−26.7 kJ
As in the catabolism of succinate by P. modestum, insufficient energy is available from this reaction to drive ATP synthesis by substrate-level phosphorylation (Table 14.5). In addition, remarkably, the organism does not have a mechanism for pumping ions out of the cell. Energy is conserved because protons are consumed in the cytoplasm during the oxidation of oxalate and production of formate. Looked at another way, a divalent molecule (oxalate) enters the cell while a univalent molecule (formate) is excreted, and this contributes to the energy of the electrochemical gradient. Together, these forces allow for the formation of a proton motive force sufficient to drive ATP synthesis (Figure 14.50b).
What Can Be Learned from the Decarboxylations of Succinate and Oxalate?
The unique aspect of decarboxylation-type fermentations is that ATP is made without substrate-level phosphorylation or electron transport reactions. In both cases, ATP synthesis is driven by a disproportionation reaction in which a redox reaction is used to generate an ion gradient directly, either by consuming protons, as in Oxalobacter, or by fueling an ion pump, as in Propionigenium. We also see this trend reflected in energy-converting hydrogenases (as in some sulfate reducers, acetogens, methanogens, and fermenters) and in the Rnf enzyme complex (as in some acetogens and fermenters), which both couple a redox reaction directly to an ion-pumping mechanism. All of these mechanisms offer an important lesson in microbial bioenergetics: ATP synthesis from reactions that yield less than −31.8 kJ/mol is still possible if the reaction can be coupled to an ion pump.
At a minimum, then, an energy-conserving reaction must yield sufficient free energy to pump at least one ion. This energy requirement is estimated to be near −12 kJ. Reactions that release less free energy than this should not be able to drive ion pumps and should therefore not be potential energy-conserving reactions. However, as we will see in the next section, bacteria are known that push this theoretical limit even lower, and their energetics, consequently, are still incompletely understood. These are the syntrophs, bacteria living on the energetic margin of existence.
Check Your Understanding
Why does Propionigenium modestum require sodium for growth?
Of what benefit is the organism Oxalobacter to human health?
How can a fermentation that yields insufficient free energy to make an ATP still support growth?
14.22 Syntrophy
There are many examples in microbiology of syntrophy, a situation in which two different microbes cooperate to perform a metabolic reaction that neither can accomplish alone. Most syntrophic reactions are secondary fermentations in which organisms ferment the fermentation products of other anaerobes. We will see in Chapter 21 how syntrophy is often a key step in anaerobic food webs and in the anaerobic metabolisms that lead to the production of methane in nature. Here we consider the microbiology and energetic aspects of syntrophy.
Interspecies Electron Transfer
Many organic compounds can be degraded syntrophically, including even aromatic and aliphatic hydrocarbons. But the major compounds of interest in syntrophic environments are fatty acids and alcohols. The heart of syntrophic reactions is interspecies electron transfer, defined as a relationship between two organisms in which one species serves as the electron acceptor for another species that is the electron donor. This relationship can be direct (direct interspecies electron transfer, DIET), mediated by either direct contact between the cells or by direct electrical connections established by using nanowires (Section 14.13). Alternatively, the relationship can be mediated (mediated interspecies electron transfer, MIET) by metabolic products that are exchanged by diffusion between cells. H2 mediates many syntrophic partnerships since many fermentative organisms readily produce it and many respiratory organisms readily consume it. We will encounter examples of DIET in Chapter 23. Here we consider only how the exchange of metabolic products allows both syntrophic partners to thrive.
As an example of syntrophy, consider the fermentation of ethanol to acetate plus H2 by the syntroph Pelotomaculum coupled to the production of methane (Figure 14.51). The syntroph carries out a reaction whose standard free-energy change (ΔG0′) is positive. Hence, in pure culture, the organism will not grow. However, the H2 produced by Pelotomaculum can be used as an electron donor by a methanogen to produce methane, an exergonic reaction. When the two reactions are summed, the overall reaction is exergonic (Figure 14.51a), and when Pelotomaculum and a methanogen are cultured together (cocultured), both organisms grow.
Figure 14.51 Syntrophy: Interspecies H2 transfer.

Shown is the fermentation of ethanol to methane and acetate by syntrophic association of an ethanol-oxidizing syntroph and a H2-consuming partner (in this case, a methanogen). (a) Reactions involved. The two organisms share the energy released in the coupled reaction. (b) Overview of the syntrophic transfer of H2.
A second example of syntrophy is the oxidation of a fatty acid such as butyrate to acetate plus H2 by the fatty acid–oxidizing bacterium Syntrophomonas (**Figure 14.52*a***). Syntrophomonas oxidizes butyrate by reversing most steps in the butyric acid fermentation pathway (see Figure 14.46). However, the electron-bifurcating enzyme of butyric acid fermentation is replaced by an enzyme that oxidizes butyryl-CoA to produce NADH (Figure 14.52a). The two NADH produced in butyrate oxidation are oxidized in an electron bifurcation reaction to reduce Fdox and menaquinone (Q). The Fdred is then used by a cytoplasmic hydrogenase to produce one H2, and the QH2 is used in a membrane-associated endergonic reaction to reduce 2 H+ to H2. The free-energy change of this overall reaction is even more unfavorable than that of ethanol oxidation and in pure culture Syntrophomonas will obviously not grow on butyrate (Figure 14.52a). However, as with ethanol fermentation by Pelotomaculum, if the H2 produced by Syntrophomonas is consumed by a partner organism, Syntrophomonas will grow on butyrate in coculture with the H2-consuming partner. How does this occur?
Figure 14.52 Energetics of growth of *Syntrophomonas* in syntrophic culture and in pure culture.

(a) In syntrophic culture, growth requires a H2-consuming organism, such as a methanogen. H2 production is driven by reverse electron transport because the E0′ values of the NADH couple are more electropositive than that of 2 H+/H2. (b) In pure culture, energy conservation is linked to anaerobic respiration with crotonate reduction to butyrate. Inset: photomicrograph of cells of a fatty acid–degrading syntrophic bacterium (red) in association with a methanogen (green-yellow).
Energetics of H2 Transfer
In a syntrophic relationship, the removal of H2 by a partner organism shifts the equilibrium of the entire reaction and pulls it in the direction of product formation; this can greatly affect the energetics of the reaction. Recall from our consideration of the principles of free energy (Section 3.3) that the concentration of reactants and products in a reaction can have a major effect on the reaction’s energetics. Said another way, the ΔG0′ of a reaction calculated under standard conditions may well differ from the ΔG of the reaction calculated under actual conditions (Figure 3.7). At very low H2 concentration, the energetics of oxidizing butyrate (or ethanol, or other fatty acids) to acetate plus H2 become exergonic. For example, if a partner organism keeps the concentration of H2 extremely low, the oxidation of butyrate by Syntrophomonas yields −18 kJ/mol (Figure 14.52a).
While H2 transfer is characteristic of many syntrophic associations, particularly those associated with fermentation, it is also possible to have syntrophic associations in which only electrons are transferred. In direct electron transfer, such as the anoxic consumption of CH4 by the ANME–sulfate reducer consortium (Section 14.16 and Figure 14.41), we learned that some respiratory syntrophs can use electrically conductive proteins (large multiheme cytochromes and “nanowires”) to transfer electrons directly between cells separated by significant distances. In such syntrophic reactions, direct electron transfer does not depend on diffusion rates or H2 concentrations because the electron transfer results from a direct connection between two cells.
Ecology of Syntrophs
Ecologically, syntrophic bacteria are key links in the anoxic steps of the carbon cycle (Section 21.2). Syntrophs consume highly reduced fermentation products and release a key product, H2, for anaerobic respirations. Without syntrophs, a bottleneck would develop in anoxic environments in which electron acceptors (other than CO2) were limiting. By contrast, when conditions are oxic or alternative electron acceptors are abundant, syntrophic relationships are unnecessary. For example, if O2 or NO3 − is available as an electron acceptor, the energetics of the respiration of a fatty acid or an alcohol is so favorable that syntrophic relationships are unnecessary. Thus, syntrophy is characteristic of anoxic catabolisms in which primarily methanogenesis or acetogenesis are the terminal processes in the ecosystem. Methanogenesis is a major process in anoxic wastewater biodegradation, and microbiological studies of sludge granules that form in such systems have shown the close physical relationship that develops between H2 producer and H2 consumer in such habitats (Figure 14.52a inset).
The final part of this chapter, Part VII, considers the catabolism of a specific class of substrates: hydrocarbons. These electron donors, common in nature, can be degraded both aerobically and anaerobically, and once again, metabolic diversity is on display.
Check Your Understanding
Give an example of interspecies H2 transfer. How is it that both organisms benefit from this process?
Could Syntrophomonas grow in a syntrophic association with a butyric acid fermenter like Clostridium pasteurianum?
How does Syntrophomonas produce H2, and what makes this possible?
VII Hydrocarbon Metabolism
Hydrocarbons are organic compounds that are devoid of oxygen and can be catabolized both aerobically and anaerobically. Hydrocarbon catabolism requires oxygen, and anaerobically, a series of unique reactions solves this biochemical conundrum.
Hydrocarbons, molecules that contain only carbon and hydrogen atoms, are widely used by microbes as electron donors, and we wrap up our coverage of metabolic diversity with a consideration of this process. Unlike methane, hydrocarbons containing two or more carbons typically have to be oxygenated before they can be catabolized. We first consider the aerobic catabolism of aliphatic and aromatic hydrocarbons, where this oxygenation involves O2. We then proceed with a consideration of anoxic hydrocarbon metabolism, a situation where oxygenation of the hydrocarbon is still necessary, but where O2 obviously plays no role.
14.23 Aerobic Hydrocarbon Metabolism
We previously discussed the role of molecular oxygen (O2) as an electron acceptor in energy-generating reactions. By contrast, O2 also plays an important role as a reactant in the catabolism of hydrocarbons, and oxygenase enzymes are key players in the process.
Oxygenases and Aliphatic Hydrocarbon Oxidation
Oxygenases are enzymes that catalyze the incorporation of O2 into organic compounds and in some cases, inorganic compounds (Section 14.9). There are two classes of oxygenases: dioxygenases, which catalyze the incorporation of both atoms of O2 into the molecule, and monooxygenases, which catalyze the incorporation of only one of the two oxygen atoms of O2 into an organic compound with the second atom of O2 being reduced to H2O. For most monooxygenases, the required electron donor is NADH or NADPH.
In the initial oxidation step of a saturated aliphatic (straight-chain) hydrocarbon, one of the atoms of O2 is incorporated, typically at a terminal carbon atom. This reaction is catalyzed by a monooxygenase, and a typical reaction sequence is shown in **Figure 14.53*a***. The end product of the reaction sequence is a fatty acid of the same length as the original hydrocarbon. The fatty acid is then oxidized by beta-oxidation, a series of reactions in which two carbons of the fatty acid are split off at a time (Figure 14.53b). During beta-oxidation, NADH is formed and is oxidized in the electron transport chain for energy conservation purposes (Figure 3.19). A single round of beta-oxidation releases acetyl-CoA plus a new fatty acid that is two carbon atoms shorter than the original fatty acid. The process of beta-oxidation is then repeated, and another acetyl-CoA molecule is released. The acetyl-CoA formed by beta-oxidation is either oxidized through the citric acid cycle (Figure 3.12) or used to make new cell material. With the exception of how the hydrocarbon is oxygenated, much of the biochemistry of anoxic hydrocarbon catabolism is the same as that shown for aerobic catabolism (Figure 14.53), with beta-oxidation reactions being of prime importance in both cases.
Figure 14.53 Monooxygenase activity and beta-oxidation.

(a) Steps in the oxidation of an aliphatic hydrocarbon, the first of which is catalyzed by a monooxygenase. (b) Fatty acid oxidation by beta-oxidation leads to the successive formation of acetyl-CoA.
Aromatic Hydrocarbon Oxidation
Many aromatic (ringed) hydrocarbons can also be used as electron donors aerobically by microorganisms. The metabolism of these compounds, some of which contain multiple rings, such as naphthalene or biphenyls, typically has as its initial stage the formation of catechol or a structurally related compound via catalysis by oxygenase enzymes, as shown in Figure 14.54. Once catechol is formed it can be cleaved and further degraded into compounds that can enter the citric acid cycle, such as succinate, acetyl-CoA, and pyruvate.
Figure 14.54 Roles of oxygenases in catabolism of aromatic compounds.

Monooxygenases introduce one atom of oxygen from O2 into a substrate, whereas diooxygenases introduce both atoms of oxygen. (a) Hydroxylation of benzene to catechol by a monooxygenase in which NADH is an electron donor. (b) Cleavage of catechol to cis, cis-muconate by an intradiol ring-cleavage dioxygenase. (c) The activities of a ring-hydroxylating dioxygenase and an extradiol ring-cleavage dioxygenase in the degradation of toluene. The oxygen atoms that each enzyme introduces are distinguished by different colors. Compare aerobic toluene catabolism to anoxic toluene catabolism shown in Figure 14.55b.
Several steps in the aerobic catabolism of aromatic hydrocarbons require oxygenases. Figure 14.54a–c shows four different oxygenase-catalyzed reactions, one using a monooxygenase, two using a ring-cleaving dioxygenase, and one (toluene dioxygenase) using a ring-hydroxylating mechanism. As in aerobic aliphatic hydrocarbon catabolism (Figure 14.53), aromatic compounds, whether single- or multi-ringed, are typically oxidized completely to CO2, with electrons entering an electron transport chain or used to make new cell material.
Check Your Understanding
How do monooxygenases differ in function from dioxygenases?
What is the final product of catabolism of a hydrocarbon?
14.24 Anaerobic Hydrocarbon Metabolism
Although aerobic hydrocarbon oxidation is a major process in nature, anaerobic hydrocarbon oxidation linked to the reduction of nitrate, sulfate, or ferric iron as electron acceptors in anaerobic respirations (Sections 14.11, 14.12 and 14.13) is also possible. And, as for the aerobic process, both aliphatic and aromatic hydrocarbons can be degraded anaerobically.
Aliphatic Hydrocarbons
Aliphatic hydrocarbons are substrates for denitrifying and sulfate-reducing bacteria. Saturated aliphatic hydrocarbons as long as C20 support growth, although shorter-chain hydrocarbons are more soluble and therefore more readily catabolized. The mechanism of anoxic hydrocarbon degradation has been well studied for hexane (C6H14) metabolism in denitrifying bacteria, organisms that use NO3 − as an electron acceptor (Section 15.10). However, the mechanism appears to be the same for the anoxic catabolism of longer-chain hydrocarbons and for anoxic hydrocarbon oxidation linked to other electron acceptors, and so we focus on the hexane/nitrate system here.
In anoxic hexane metabolism, hexane is modified on carbon atom 2 by attachment of a molecule of fumarate, a C4 intermediate of the citric acid cycle (Figure 3.12), forming the compound 1-methylpentylsuccinate (**Figure 14.55*a***). The enzymatic addition of fumarate to hexane oxygenates the hydrocarbon without the need for O2 and allows the molecule to be further catabolized anaerobically. Following the addition of coenzyme A, a series of reactions occurs that includes beta-oxidation (Figure 14.53b) and regeneration of fumarate (Figure 14.55a). The electrons released during beta-oxidation generate a proton motive force and are consumed in nitrate or sulfate reduction (Sections 14.11 and 14.12, respectively).
Figure 14.55 Anoxic catabolism of two hydrocarbons.

(a) In anoxic catabolism of the aliphatic hydrocarbon hexane, the addition of fumarate provides the oxygen atoms necessary to form a fatty acid derivative that can be catabolized by beta-oxidation (see Figure 14.53b) to yield acetyl-CoA. Electrons (H) generated from hexane catabolism are used to reduce sulfate or nitrate in anaerobic respirations. (b) Fumarate addition during the anoxic catabolism of the aromatic hydrocarbon toluene forms benzylsuccinate.
Aromatic Hydrocarbons
Aromatic hydrocarbons can also be degraded anaerobically through anaerobic respirations. For anoxic catabolism of the aromatic hydrocarbon toluene, oxygen needs to be added and this also occurs by the addition of fumarate (Figure 14.55b). The reaction series eventually yields benzoyl-CoA, which is further degraded by ring reduction (see Figure 14.56). Benzene (C6H6) can also be catabolized anaerobically, likely by a similar mechanism. Multi-ringed aromatic hydrocarbons such as naphthalene (C10H8) can be degraded by certain sulfate-reducing and denitrifying bacteria. But in contrast to other hydrocarbons, the oxygenation of multi-ringed hydrocarbons occurs by the addition of CO2 to the ring to form a carboxylic acid derivative rather than by fumarate addition. However, this carboxylation reaction serves the same biochemical purpose as oxygenase reactions (Figures 14.53a and 14.54) or the addition of fumarate (Figure 14.55); an O atom becomes part of the hydrocarbon and facilitates its catabolism.
Many bacteria can catabolize certain aromatic hydrocarbons anaerobically, including even fermentative and phototrophic purple bacteria. However, except for toluene, only aromatic compounds that already contain an O atom are degraded, typically by a common mechanism. In contrast to aerobic catabolism that occurs by way of ring oxidation (Figure 14.54), anaerobic catabolism proceeds by ring reduction. The anaerobic degradation of aromatic hydrocarbons is often facilitated by their conversion into benzoate followed by aromatic ring cleavage and benzoate catabolism performed by the benzoyl-CoA pathway (Figure 14.56). Benzoate catabolism in this pathway begins by forming the coenzyme A derivative followed by ring cleavage to yield fatty or dicarboxylic acids (Figure 14.56) that can be further catabolized to intermediates of the citric acid cycle (Figure 3.12).
Figure 14.56 Anoxic degradation of benzoate by the benzoyl-CoA pathway.

This pathway operates in the purple phototrophic bacterium Rhodopseudomonas palustris and many other facultative bacteria, both phototrophic and chemotrophic. Note that all intermediates of the pathway are bound to coenzyme A. The acetate produced is further catabolized in the citric acid cycle.
As we reach the end of our survey of metabolic diversity, we need not keep in mind all of the metabolic details, for they are numerous and formidable, but instead, we need to see the “metabolic big picture” formed by the major themes of this chapter: the modularity of metabolism, and the key processes of phototrophy, chemolithotrophy, and autotrophy, respirations by electron donor or electron acceptor, C1 metabolisms, fermentations, and hydrocarbon metabolism. These overarching principles will guide us through the next four chapters where our focus will be the diversity of the microbes themselves rather than their metabolisms. Chapters 15, 16 and 17 in particular will cover many of the Bacteria and Archaea whose metabolisms we have described here, and we will witness there the many cases in which metabolic diversity and prokaryotic diversity are inextricably linked.
Check Your Understanding
What is the benzoyl-CoA pathway, and how might it participate in the anaerobic degradation of toluene?
How is hexane oxygenated during anoxic catabolism?
Chapter Review
Go to Mastering Microbiology for videos, animations, practice tests, and more.
I Introduction to Metabolic Diversity
14.1 All cells must conserve energy, have a source of reducing power, and achieve redox balance in their metabolism. Energy is conserved through substrate-level phosphorylation, oxidative phosphorylation, or photophosphorylation. Reducing power is supplied by electron donors or must be generated in endergonic reactions. Redox balance is achieved by using terminal electron acceptors (in respiration) or fermentation products (including H2) to export electrons from the cell.
Q What are reverse electron transport and electron bifurcation, and what role do they play in generating reducing power and achieving redox balance?
14.2 Autotrophy is supported in most phototrophic and chemolithotrophic bacteria by the Calvin cycle, in which the enzyme RuBisCO plays a key role. The reverse citric acid and hydroxypropionate cycles are autotrophic pathways in green sulfur and green nonsulfur bacteria, respectively.
Q What is a carboxysome, and what is its role in CO2 fixation? Plants do not have carboxysomes; how might this affect the efficiency with which they fix CO2?
II Phototrophy
14.3 In photosynthesis, ATP is generated from light and then consumed in the reduction of CO2. Two forms of photosynthesis are known: oxygenic, where O2 is produced (for example, in cyanobacteria), and anoxygenic, where it is not (for example, in purple and green bacteria). Photosynthetic reaction centers and photosynthetic pigments reside in membranes where the light reactions of photosynthesis are carried out.
Q What are the functions of light-harvesting and reaction center chlorophylls?
14.4 Accessory pigments including carotenoids and phycobilins absorb light and transfer the energy to reaction center chlorophyll, thus broadening the wavelengths of light usable in photosynthesis. Carotenoids also play an important photoprotective role in preventing photooxidative damage to cells.
Q What accessory pigments are present in phototrophs, and what are their functions?
14.5 In anoxygenic phototrophs, electron transport reactions in photosynthetic membranes are driven by the photosynthetic reaction center, forming a proton motive force and ATP.
Q Why is reverse electron transport necessary in purple sulfur bacteria but not in green sulfur bacteria?
14.6 In oxygenic photosynthesis, H2O donates electrons to drive CO2 fixation, and O2 is a by-product. There are two separate but interconnected photosystems in oxygenic phototrophs, PSI and PSII, whereas anoxygenic phototrophs contain a single photosystem.
**Q How does the reduction potential (E0′) of chlorophyll a in PSI and PSII differ? Why must the reduction potential of PSII chlorophyll a be so highly electropositive?**
III Respiratory Processes Defined by Electron Donor
14.7 Reduced sulfur compounds such as H2S, S2O3 2−, and S0 are electron donors for energy conservation in sulfur chemolithotrophs. Electrons from these substances enter electron transport chains, yielding a proton motive force. Sulfur chemolithotrophs are also autotrophs and fix CO2 by the Calvin cycle.
**Q Compare and contrast the utilization of H2S by a purple phototrophic bacterium and by a colorless sulfur bacterium such as Beggiatoa. What role does H2S play in the metabolism of each organism?**
14.8 Chemolithotrophic iron bacteria oxidize Fe2+ as an electron donor. Most iron bacteria grow at acidic pH and are often associated with acidic pollution from mineral and coal mining. A few chemolithotrophic and phototrophic bacteria can oxidize Fe2+ to Fe3+ anaerobically.
Q Why is it necessary for iron-oxidizing bacteria to have cytochromes in their outer membranes even though electron transport and energy conservation takes place at the cytoplasmic membrane?
14.9 The ammonia-oxidizing Bacteria and Archaea produce nitrite from ammonia, which is oxidized by nitrite-oxidizing Bacteria to nitrate.
Q What is the key enzyme that mediates ammonia oxidation, and how does this enzyme function in the cell?
14.10 Anoxic ammonia oxidation (anammox) consumes both ammonia and nitrite, forming N2. The anammox reaction occurs within a membrane-enclosed compartment called the anammoxosome.
Q Contrast classical nitrification with anammox in terms of oxygen requirements, organisms involved, and the need for monooxygenases.
IV Respiratory Processes Defined by Electron Acceptor
14.11 Nitrate is a common electron acceptor in anaerobic respiration. Nitrate reduction is catalyzed by the enzyme nitrate reductase, reducing NO3 − to NO2 −. Denitrification is an anaerobic respiratory process through which NO3 − is reduced to gaseous nitrogen compounds including NO, N2O, or N2.
Q Why is it that denitrifiers that reduce NO3 − to N2 can still release significant amounts of N2O?
14.12 Sulfate-reducing bacteria are obligately anaerobic bacteria that reduce SO4 2− to H2S in a process in which SO4 2− must first be activated to adenosine phosphosulfate (APS). Some sulfur-reducing organisms can reduce S0 but not SO4 2− to H2S.
**Q Why is the enzyme hydrogenase useful to Desulfovibrio even when it is not grown on H2 as electron donor?**
14.13 Iron-reducing bacteria can use nanowires to reduce insoluble electron acceptors on the outside of the cell. Many different electron-accepting respiratory reactions are possible including the reduction of various metals and metalloids, fumarate and other organic compounds, and various chlorinated compounds.
Q In what way might bacteria alter the toxicity of groundwater contaminated with arsenic, chromium, or even uranium?
V One-Carbon (C1) Metabolism
14.14 Acetogens are strict anaerobes that reduce CO2 to acetate, usually with H2 as electron donor. The mechanism of acetate formation is the reductive acetyl-CoA pathway, a pathway widely distributed in obligate anaerobes for either autotrophic purposes or acetate oxidation.
Q Describe the mechanism of energy conservation for acetogens growing on H2+CO2.
14.15 Methanogenesis is the production of CH4 from CO2+H2 or from acetate or methanol by strictly anaerobic methanogenic Archaea. Several unique coenzymes are required for methanogenesis, and energy conservation is linked to either a proton or sodium motive force.
Q Why is it that methanogens that lack cytochromes are unable to grow on methanol?
14.16 Methanotrophy is the use of CH4 as both carbon source and electron donor. Aerobic methanotrophs use the enzyme methane monooxygenase and oxidize CH4 to formaldehyde, which is then assimilated by the cell through one of two pathways. Methane can also be oxidized anaerobically by diverse organisms through syntrophic partnerships or by generating O2 as a catalyst for CH4 oxidation.
**Q Which pathway for C1 assimilation found in aerobic methanotrophs is most energetically efficient and why? In what ways does Methylomirabilis oxyfera resemble denitrifiers and aerobic methanotrophs, and in what ways does it differ?**
VI Fermentation
14.17 In the absence of external electron acceptors, organic compounds can be catabolized anaerobically only by fermentation. In fermentation, most ATP is typically formed by substrate-level phosphorylation. Redox balance is achieved by the excretion of fermentation products.
Q Define the term substrate-level phosphorylation: How does it differ from oxidative phosphorylation? What compound(s) do fermentative bacteria need to synthesize in order to make ATP by substrate-level phosphorylation?
14.18 The lactic acid fermentation is carried out by homofermentative species, where lactate is the sole product, and heterofermentative species, where lactate, ethanol, and CO2 are produced. The mixed-acid fermentation typical of enteric bacteria yields various acids plus neutral products (ethanol, butanediol), depending on the organism.
**Q What are the major fermentation products of Lactobacillus and Escherichia?**
14.19 Clostridia ferment sugars, amino acids, and other organic compounds, with butyric and acetic acids and H2 being major products. Butyrate production allows for additional ATP to be produced. Energy-converting hydrogenases allow some fermenters to generate a proton or sodium motive force by producing H2.
**Q When fermenting 1.5 glucose to butyrate and acetate, Clostridium pasteurianum produces 3 NADH during glycolysis. How does it achieve redox balance?**
14.20 The bacterium Propionibacterium produces propionate, acetate, and CO2 in a secondary fermentation of lactate. By contrast, Clostridium kluyveri produces butyrate and caproate by oxidizing ethanol and reducing acetate.
Q What is the difference between primary and secondary fermentation?
14.21 Energy conservation in Propionigenium, Oxalobacter, and Malonomonas is linked to decarboxylation reactions that pump Na+ or H+ across the membrane; ATPases use the energy in the ion gradient to form ATP. The reactions catalyzed by these organisms yield insufficient free energy to make ATP by substrate-level phosphorylation but sufficient energy to pump ions.
Q Give an example of a fermentation that does not employ substrate-level phosphorylation.
14.22 In syntrophy, two organisms cooperate to degrade a compound that neither can degrade alone. In this process, H2 produced by one organism is consumed by another. H2 consumption affects the energetics of the reaction carried out by the H2 producer, allowing it to make ATP where it otherwise could not.
Q When do syntrophic relationships not involve H2 transfer?
VII Hydrocarbon Metabolism
14.23 In addition to its role as a terminal electron acceptor, O2 can also be introduced directly into other substances. In aerobic metabolism, oxygenases introduce atoms of oxygen from O2 into hydrocarbons. Once oxygenated, aliphatic hydrocarbons can be further degraded by beta-oxidation and aromatic hydrocarbons by ring splitting and oxidation.
Q How do monooxygenases differ from dioxygenases in terms of the reactions they catalyze? Why are oxygenases necessary for the aerobic catabolism of hydrocarbons?
14.24 Hydrocarbons can be oxidized under anoxic conditions following addition of the dicarboxylic acid fumarate. Aromatic compounds are catabolized anaerobically by ring reduction and cleavage to form intermediates that can be catabolized in the citric acid cycle.
Q How do denitrifying and sulfate-reducing bacteria degrade hydrocarbons anaerobically and without oxygenases?
Application Questions
The growth rate of the phototrophic purple bacterium Rhodobacter, grown phototrophically with H2 as electron donor, is about twice as fast in a medium containing malate as in a medium containing CO2 as the only carbon source. Discuss the reasons why this is true and list the nutritional class in which we would place Rhodobacter when growing under each of the two different conditions.
A fatty acid such as butyrate cannot be fermented in pure culture, but in many anoxic habitats, fermentative organisms oxidize it readily. Why can butyrate be fermented in a community of organisms when this process cannot be performed by a pure culture?
Compare and contrast the methanogenic pathway for growth on H2+CO2 in methanogens that have cytochromes with those that lack cytochromes. Why do acetate-degrading (acetoclastic) methanogens not need to fix CO2?
Chapter Glossary
energy metabolism in which acetate is produced from either H2 plus CO2 or from organic compounds Anaerobic respiration
a form of respiration in which the terminal electron acceptor is not O2 Anammox
anoxic ammonia oxidation Anoxygenic photosynthesis
photosynthesis in which O2 is not produced Antenna pigments
light-harvesting chlorophylls or bacteriochlorophylls in photocomplexes that funnel energy to the reaction center Autotroph
an organism capable of biosynthesizing all cell material from CO2 as the sole carbon source Bacteriochlorophyll
the chlorophyll pigment of anoxygenic phototrophs Calvin cycle
the series of biosynthetic reactions by which most phototrophs and most aerobic chemolithotrophs convert CO2 into organic compounds Carboxysomes
a polyhedral cellular inclusion of crystalline ribulose biphosphate carboxylase (RuBisCO), the key enzyme of the Calvin cycle Carotenoid
a hydrophobic accessory pigment present along with chlorophyll in photosynthetic membranes Chlorophyll
a light-sensitive, Mg-containing porphyrin of phototrophic organisms that initiates the process of photophosphorylation Chlorosome
a cigar-shaped structure present in the periphery of cells of green sulfur and green nonsulfur bacteria and containing the antenna bacteriochlorophylls (c, d, or e) Conserve energy
to convert energy from chemical reactions or light into a form that can do work, such as ATP Denitrification
anaerobic respiration in which NO3 − or NO2 − is reduced to nitrogen gases, primarily N2 Electron bifurcation (confurcation)
an exergonic redox reaction including three simultaneous half reactions whereby 4 electrons (2×2 e−) from a donor are split between an exergonic reduction (2×1 e−) and an endergonic reduction (2×1 e−); the reverse reaction is a confurcation causing an endergonic reduction involving two e− that is driven by an exergonic reduction performed with the other two e− Energy-converting hydrogenase
a membrane-bound enzyme that uses reduced ferredoxin to reduce protons to H2 and couples this reaction to the generation of a proton or sodium motive force Fermentation
anaerobic catabolism in which an organic compound is both an electron donor and an electron acceptor and ATP is produced by substrate-level phosphorylation Heterofermentative
in reference to lactic acid bacteria, capable of making a mixture of fermentation products typically including lactate, ethanol, and CO2, from the fermentation of glucose Homofermentative
in reference to lactic acid bacteria, producing only lactic acid as a fermentation product Hydrogenase
an enzyme, widely distributed in anaerobic microorganisms, capable of oxidizing or evolving H2 Methanogen
a methane-producing species of the Archaea Methanogenesis
an organism capable of oxidizing methane (CH4) as an electron donor in energy metabolism Methylotroph
an organism capable of oxidizing organic compounds that do not contain carbon–carbon bonds; if able to oxidize CH4 also a methanotroph Mixotroph
an organism that conserves energy from the oxidation of inorganic compounds but requires organic compounds serve as a carbon source Nanowires
electrically conductive type IV pili used to exchange electrons with extracellular objects, either other cells or abiotic structures Nitrification
the microbial oxidation of ammonia to nitrate Oxygenase
an enzyme that catalyzes the incorporation of oxygen from O2 into organic or inorganic compounds Oxygenic photosynthesis
photosynthesis carried out by cyanobacteria and green plants in which O2 is evolved Photosynthesis
the series of reactions in which ATP is synthesized by light-driven reactions and CO2 is fixed into cell material Phototroph
an organism that uses light as its source of energy Phycobiliprotein
the antenna pigment complex in cyanobacteria that contains phycocyanin and allophycocyanin or phycoerythrin coupled to proteins Phycobilisome
a photosynthetic complex containing chlorophyll or bacteriochlorophyll and several other components; the initial electron transfer reactions of photosynthetic electron flow occur here Redox balance
having the same number of electrons in the reactants and the products of metabolism Reducing power
the ability to donate electrons in metabolic reactions Reductive acetyl-CoA pathway
a pathway of CO2 fixation used by obligate anaerobes such as acetogens and methanogens; the pathway reduces CO2 to acetate and can be reversed to oxidize acetate to CO2 Reductive dechlorination
an anaerobic respiration in which a chlorinated organic compound is used as an electron acceptor, usually with the release of Cl− Respiration
the process in which an electron donor is oxidized and electrons are passed through electron transport reactions to an external electron acceptor in order to generate a proton motive force and synthesize ATP by oxidative phosphorylation Reverse citric acid cycle
a pathway of CO2 fixation used in green sulfur bacteria as well as microaerophilic and anaerobic chemolithotrophs Reverse electron transport
a method of generating reducing power, usually in the form of NADH or Fdred, whereby the endergonic reduction of electron carriers is driven by dissipation of a proton motive force Ribulose monophosphate pathway
a reaction series in certain methylotrophs in which formaldehyde is assimilated into cell material using ribulose monophosphate as the C1 acceptor molecule RuBisCO
ribulose bisphosphate carboxylase, a key enzyme for CO2 fixation by the Calvin cycle Secondary fermentation
a fermentation in which the substrates are the fermentation products of other organisms Serine pathway
a reaction series in certain methylotrophs in which CH2O plus CO2 are assimilated into cell material by way of the amino acid serine Stickland reaction
a process whereby two or more microorganisms cooperate to degrade a substance neither can degrade alone Thylakoids
membrane stacks containing the photosynthetic pigments in cyanobacteria or in the chloroplast of eukaryotic phototrophs