15 Ecological Diversity of Bacteria
**II Ecological Diversity of Phototrophic *Bacteria***

**III Diversity of Bacteria Defined by Metabolic Traits**
**IV Morphologically and Ecologically Distinctive *Bacteria***
Cyanobacterial Diversity and Environmental Change
Cyanobacteria have had an enormous impact on the Earth for over 2 billion years: They caused Earth’s oxygenation and still produce much of the oxygen we breathe today. Cyanobacteria show tremendous ecological diversity, and they are an essential component of healthy ecosystems worldwide. However, the ecological traits that make cyanobacteria so successful also allow them to respond rapidly to environmental change. Human activities alter ecosystem function, and when these activities promote the unrestricted growth of bacteria, the results can be catastrophic.

Intensive agriculture often produces nitrogen and phosphorus runoff that causes ecosystem eutrophication (nutrient enrichment). Phosphorus availability limits cyanobacterial growth in freshwater, and excess phosphorus causes rapid growth of certain cyanobacteria, resulting in harmful blooms. These cyanobacterial blooms release toxins and smother aquatic plants, and subsequent bloom decomposition fuels massive growth of heterotrophic bacteria, causing anoxic conditions that kill aquatic animals. Lake Erie (USA) was once so highly eutrophic it was considered a dead lake, but environmental regulations restricted phosphorus inputs, restoring its health. However, eutrophication is a problem once again, now largely because of nitrogen runoff.
Microcystis is a genus of freshwater cyanobacteria. While many cyanobacteria can fix N2, Microcystis cannot, and hence nitrogen enrichment favors the growth of Microcystis. Recent blooms in Lake Erie (see natural-color NASA photo, September 2017) were dominated by Microcystis species that produce microcystin, a toxin that can kill animals (and humans). A massive Microcystis bloom left the city of Toledo, Ohio (USA) without drinking water in 2014, and a similar bloom in Wuxi, China, affected millions of people in 2007.
Elevated temperatures, CO2, and nutrient levels associated with climate change are predicted to make such blooms more frequent and more intense in the future. It is thus essential that we understand the ecological diversity of microbes such as cyanobacteria if we are to predict and manage their responses to environmental change.
Source: Steffen, M.M., et al. 2017. Ecophysiological examination of the Lake Erie Microcystis bloom in 2014: Linkages between biology and the water supply shutdown of Toledo, OH. Environ. Sci. Technol. 51: 6745.
I Ecological Diversity Among Microorganisms
Diversity in the microbial world can be described in many different ways. Ecological diversity emphasizes the characteristics of microbial groups in the context of their environments and the microbes they interact with.

Microbial diversity defies simple definitions. If we are to make sense of microbial diversity, we must first identify and measure how one organism differs from another. Microbes differ in many ways, but at a fundamental level we define these differences in terms of three aspects: metabolism, ecology, and phylogeny. Ultimately, these three threads weave together in the tapestry that we call microbial diversity, and only by examining each thread can we understand their connections. We explored microbial metabolic diversity in the previous chapter, and we will explore the phylogenetic diversity of microorganisms in the chapters that follow. Now we will consider and define microbial diversity in terms of ecological differences between microorganisms.
15.1 Making Sense of Microbial Diversity
Metabolic diversity, as explored in the last chapter, is defined in terms of the cellular processes that support growth. Ecological diversity, which we review in the present chapter, is defined in terms of microbial interactions between organisms and their environments. Finally, phylogenetic diversity, which we will review in the coming chapters, is defined by evolutionary relationships between organisms.
Ecological diversity is the component of microbial diversity that deals with how microbes interact with each other and with their environments. In the microbial world, ecological interactions are often defined by metabolic traits. For example, consider lactic acid bacteria (Section 14.18). These fermentative organisms achieve redox balance by excreting lactic acid as a fermentation product. The excretion of lactic acid also lowers the pH of the environment. Lactic acid bacteria grow well in these acidic conditions, and this acidity inhibits their neutrophilic competitors. Hence, lactic acid production allows these bacteria to interact with their environment and their competitors in a way that facilitates their own growth. In addition to transforming the environment, the products of metabolism often serve as microbial substrates for other microbes, leading to complex mutualistic interactions (Section 14.22). For example, the lactic acid just mentioned is still an energy-rich substance and is actively consumed by propionic acid bacteria, secondary fermenters that ferment lactate into propionate (Section 14.20). Hence, in the microbial world, ecology is interdependent with metabolism.
While it is useful to consider ecological diversity in terms of metabolism, it is incorrect to consider ecological diversity solely in terms of metabolism. Microbes that have similar metabolic traits can still differ substantially in their ecology. For example, organisms that have similar metabolic traits can differ with respect to their morphology, motility, growth rate, environmental tolerances, mutualisms and antagonisms, as well as their systems of gene regulation and development. All of these traits can have major impacts on microbial distribution and activity in the environment. These complex traits cannot be defined solely in terms of metabolism and are best understood in terms of ecological interactions.
Most fundamentally, phylogenetic diversity deals with the diversity of evolutionary lineages such as phyla, genera, and species. At its broadest, phylogenetic diversity encompasses the genetic and genomic diversity of evolutionary lineages and so can be defined on the basis of either genes or organisms (Section 13.11). Most commonly, though, phylogenetic diversity is defined on the basis of ribosomal RNA gene phylogeny, which is thought to reflect the phylogenetic history of the entire organism (Sections 13.3 and 13.12). Phylogenetic diversity is the overarching theme of our coverage of microbial diversity in Chapters 16, 17 and 18.
In many cases, metabolic and ecological characteristics align with phylogenetic groups (for example, with the organisms described in Sections 15.3, 15.4, 15.6, 15.7, and 15.17). Microbial ecological diversity, however, often does not correspond with phylogenetic diversity as defined by the 16S ribosomal RNA gene. We will see many examples in this chapter where metabolic and ecological traits are widely distributed among the Bacteria and Archaea (Figure 15.1), and at least three reasons can account for why metabolic and ecological traits are shared between divergent organisms. The first is gene loss, a situation where a trait present in the common ancestor of several lineages is subsequently lost in some lineages but retained in others that over evolutionary time became quite divergent. The second is convergent evolution, in which a trait has evolved independently in two or more lineages and is not encoded by homologous genes shared by these lineages. The third is horizontal gene transfer (Sections 9.5, 9.6, 9.7, 9.8, 9.9, 9.10 and 9.11 and 13.9, 13.10 and 13.11), a situation where genes that confer a particular trait are homologous and have been exchanged between distantly related lineages. Figure 15.1 emphasizes these points by showing how many common metabolisms are dispersed among distinct phylogenetic groups of microbes.
Figure 15.1 Major functional traits mapped across major phyla of *Bacteria* and *Archaea*.

The dendrogram shows relationships between microbial phyla as inferred by analysis of 16S ribosomal RNA gene sequences. Blue branches are used to denote phyla of Bacteria and red branches denote phyla of Archaea. Colored circles indicate phyla that contain at least one species with a functional trait indicated in the color key.
The examples of ecological diversity provided in this chapter are meant to be illustrative and not exhaustive, and we will consider other organisms with important ecological functions in Chapters 16, 17 and 18 and 20, 21, 22 and 23. In particular, we will explore the causes and consequences of ecological diversity when we consider the science of microbial ecology in Unit 5 (Chapters 19, 20, 21, 22 and 23) and bring microbial ecology into a human focus when we consider the human microbiome in Chapter 24.
Check Your Understanding
Describe a metabolic process that would have a major impact on an organism’s ecology.
What are three reasons that metabolic or ecological traits might not correspond with distinct phylogenetic groups as defined by 16S ribosomal RNA gene sequences?
Why is it useful to consider microbial diversity in terms of metabolic diversity, ecological diversity, and phylogenetic diversity?
II Ecological Diversity of Phototrophic *Bacteria*
Light is a powerful energy source and has been tapped by a wide variety of microorganisms—the phototrophs—whose metabolisms share many key attributes yet differ in major ways.
In this section we consider the ecological diversity of phototrophic microorganisms, those microbes that conserve energy from light. We will see that phototrophy is widespread within the domain Bacteria and that several distinct types of phototrophs can be defined on the basis of their metabolic and ecological traits.
15.2 Overview of Phototrophic *Bacteria*
The ability to conserve energy from light evolved early in the history of life, when the Earth was anoxic (Section 13.2). Photosynthesis originated within the Bacteria, and the first phototrophic organisms were anoxygenic phototrophs, organisms that do not generate O2 as a product of photosynthesis (Section 14.5). Instead of H2O, these early phototrophs likely used H2, ferrous iron (Fe2+), or H2S as the electron donor for photosynthesis. Anoxygenic photosynthesis is present in six bacterial phyla: the Proteobacteria, Chlorobi, Chloroflexi, Firmicutes, Acidobacteria, and Gemmatimonadetes. Oxygenic photosynthesis, by contrast, is known only within the Cyanobacteria (Figure 15.1). There is extensive metabolic diversity among the anoxygenic phototrophs, which are found in a wide range of habitats. It is clear that horizontal gene exchange has had a major impact on the evolution of photosynthesis and on the distribution of photosynthetic genes across the phylogenetic tree of Bacteria.

Phototrophic bacteria have several common features. All phototrophic bacteria use chlorophyll-like pigments and various accessory pigments to harvest energy from light and transfer this energy to a membrane-bound reaction center where it is used to drive electron transfer reactions that ultimately result in the production of ATP (Sections 14.3, and 14.4 and 14.5). There are two different types of photosynthetic reaction centers: FeS-type reaction centers, which are found in photosystem I of oxygenic phototrophs, and Q-type reaction centers, which are found in photosystem II of oxygenic phototrophs (Sections 14.3, 14.4, 14.5 and 14.6). Both types of reaction centers are present in Cyanobacteria (Section 14.6), whereas only one type or the other is present in anoxygenic phototrophs. In some cases, photosynthetic pigments are found in the cytoplasmic membrane, but often they are present in intracellular photosynthetic membrane systems that originate from invaginations of the cytoplasmic membrane. These internal membranes allow phototrophic bacteria to increase the amount of pigment they contain for better use of light of low intensities.
Many phototrophic bacteria couple light energy to carbon fixation through a variety of different mechanisms (Section 14.2), but not all phototrophs fix CO2; some instead either prefer or require organic sources of carbon to support growth. We will see that many of the characteristics of phototrophic bacteria, including their membrane systems and photosynthetic pigments, have evolved as a result of niche adaptation for the light environment.
As we begin our tour of ecological diversity, it can be seen that each of the sections in this and the next three chapters begins with a list of a few “Key Genera.” These names are simply meant to familiarize the student with some of the most common microbes in the microbial world and are often the names that appear if one travels beyond this text book to explore the primary scientific literature.
Check Your Understanding
What form of photosynthesis was most likely the first to appear on Earth?
What are the different types of photosynthetic reaction centers?
Phylogeny and Classification of *Cyanobacteria*
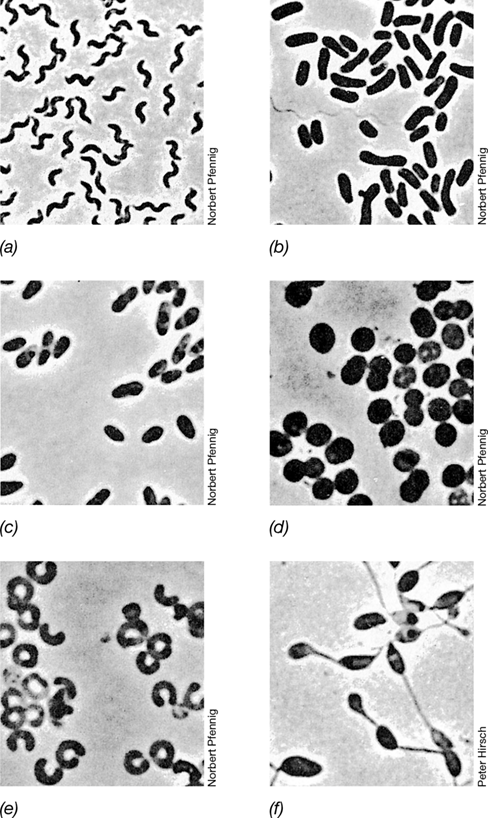
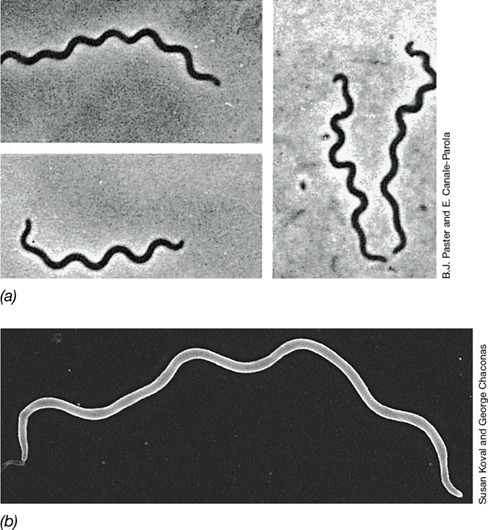
The morphological diversity of the Cyanobacteria is impressive (Figure 15.2). Both unicellular and filamentous forms exist, comprising considerable variation in morphology. Cyanobacterial cells range in size from 0.5 μm in diameter to cells as large as 100 μm in diameter. Cyanobacteria can be divided into five morphological groups: (1) Chroococcales are unicellular, dividing by binary fission (Figure 15.2a); (2) Pleurocapsales are unicellular, dividing by multiple fission (colonial) (Figure 15.2b); (3) Oscillatoriales are filamentous nonheterocystous forms (Figure 15.2c); (4) Nostocales are filamentous, divide along a single axis, and are capable of cellular differentiation (Figure 15.2d); and (5) Stigonematales are morphologically similar to Nostocales except that cells divide in multiple planes, forming branching filaments (Figure 15.2e). Finally, the prochlorophytes are a lineage of unique unicellular Cyanobacteria once thought to be distinct but now classified within the Chroococcales.
Figure 15.2 The five major morphological types of *Cyanobacteria*.

(a) Unicellular, Gloeothece; a single cell measures 5–6 μm in diameter; (b) colonial, Pleurocapsa; these structures are >50 μm in diameter and contain hundreds of cells; (c) filamentous, Lyngbya; a single cell measures about 10 μm wide; (d) filamentous heterocystous, Nodularia; a single cell measures about 10 μm wide; (e) filamentous branching, Fischerella; a cell is about 10 μm wide. See how morphological diversity relates to phylogenetic diversity in Figure 15.3.
Some of the major morphological classifications of Cyanobacteria correspond to coherent phylogenetic groups, but others do not (Figure 15.3). Species of Pleurocapsales form a coherent group within the cyanobacteria, indicating that reproduction by multiple fission arose only once in the evolutionary history of Cyanobacteria (Figure 15.3). Likewise, species of the Nostocales and Stigonematales share a common ancestor and form a coherent phylogenetic group, indicating a single origin of cellular differentiation within the Cyanobacteria (Figure 15.3). All Stigonematales share a single ancestor within the clade composed of Nostocales and Stigonematales, indicating that the capacity to form branching filaments arose only once within the lineage of Cyanobacteria capable of cellular differentiation (Figure 15.3). In contrast, unicellular and simple filamentous Cyanobacteria (Chroococcales and Oscillatoriales, respectively) are dispersed in the cyanobacterial phylogeny, and these morphological groups do not represent coherent evolutionary lineages (Figure 15.3).
Figure 15.3 Taxonomically informative traits mapped onto the phylogeny of *Cyanobacteria*.

The dendrogram depicts phylogenetic relationships inferred from analysis of conserved protein families in cyanobacterial genomes. Colored circles are used to indicate species traits as indicated by the key. Color shading is used to indicate taxonomic groupings. “Prochloro.” is used to indicate Prochlorococcus, which is a distinct group within the Chroococcales. Note that the Chroococcales and Oscillatoriales are not monophyletic in origin, meaning that their shared trait of nitrogen fixation has arisen independently on multiple occasions in the phylogeny.
Physiology and Photosynthetic Membranes
Cyanobacteria are oxygenic phototrophs and therefore have both FeS-type and Q-type photosystems. All species are able to fix CO2 by the Calvin cycle, many can fix N2, and most can synthesize their own vitamins (Sections 3.12 and 4.1). Cells harvest energy from light and fix CO2 during the day. During the night, cells generate energy by fermentation or aerobic respiration of carbon storage products such as glycogen. While CO2 is the predominant source of carbon for most species, some Cyanobacteria can assimilate simple organic compounds such as glucose and acetate if light is present, a process called photoheterotrophy. A few Cyanobacteria, mainly filamentous species, can also grow in the dark on glucose or sucrose, using the sugar as both carbon and energy source. Finally, when sulfide concentrations are high, some Cyanobacteria are able to switch from oxygenic photosynthesis to anoxygenic photosynthesis using hydrogen sulfide rather than water as electron donor for photosynthesis (Figure 14.18).
Cyanobacteria have specialized membrane systems called thylakoids that increase the ability of cells to harvest light energy (Figure 14.9). Photosynthesis takes place in the thylakoid membrane, a complex and multilayered photosynthetic membrane system containing photopigments and proteins that mediate photosynthesis (Sections 14.3 and 14.4). In most unicellular Cyanobacteria, the thylakoid membranes are arranged in regular concentric circles around the periphery of the cytoplasm (Figure 15.4). Cyanobacteria produce chlorophyll a, and most also have characteristic pigments called phycobilins (Figure 14.14), which function as accessory pigments in photosynthesis. One class of phycobilins, phycocyanins, are blue and, together with the green chlorophyll a, are responsible for the blue-green color of most cyanobacteria. Some Cyanobacteria produce phycoerythrin, a red phycobilin, and species producing phycoerythrin are red or brown. Photopigments are fluorescent and emit light when visualized using a fluorescence microscope; chlorophyll a, for example, fluoresces bright red (Figure 15.5). Prochlorophytes, such as Prochlorococcus and Prochloron, are unique among Cyanobacteria in that all members of this group contain chlorophyll a and b but do not contain phycobilins.
Figure 15.4 Thylakoids in *Cyanobacteria*.
Electron micrograph of a thin section of the cyanobacterium Synechococcus lividus. A cell is about 5 μm in diameter. Note thylakoid membranes running parallel to the cell wall.
Figure 15.5 Phycocyanin fluorescence in *Cyanobacteria*.

Fluorescence micrograph of Spirulina. Filaments consist of chains of helical cells with each cell approximately 5 μm wide.
Cellular Structures and Motility
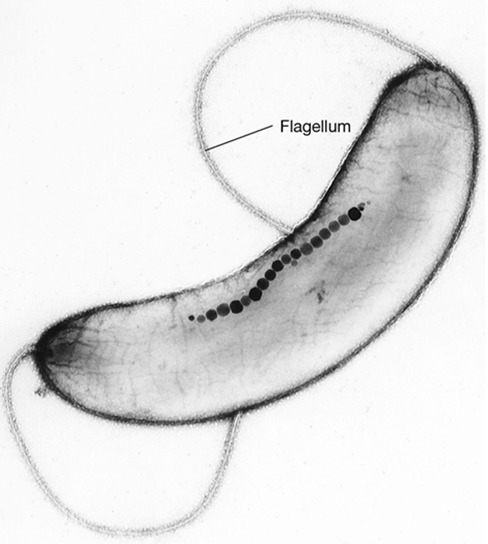
The cell wall of Cyanobacteria contains peptidoglycan and is structurally similar to that of other gram-negative bacteria. And, like many bacteria, Cyanobacteria possess several mechanisms for motility but do not contain flagella. Many cyanobacteria exhibit gliding motility (Section 2.10). Gliding occurs only when a cell or filament is in contact with a solid surface or with another cell or filament. In some Cyanobacteria, gliding is not a simple translational movement but is accompanied by rotations, reversals, and flexing of filaments (Figure 2.37b). Most gliding species exhibit directional movement toward light (phototaxis), and chemotaxis (Section 2.11) may occur as well. Synechococcus exhibits an unusual form of swimming motility that does not require flagella or any other extracellular organelle. The cell surface of Synechococcus has specialized proteins that provide direct thrust by way of a mechanism that has yet to be resolved. Gas vesicles (Section 2.7) are also found in a variety of aquatic Cyanobacteria and are important in positioning cells in the water column. The function of gas vesicles is to regulate cell buoyancy such that cells can remain in a position in the water column where light intensity is optimal for photosynthesis.
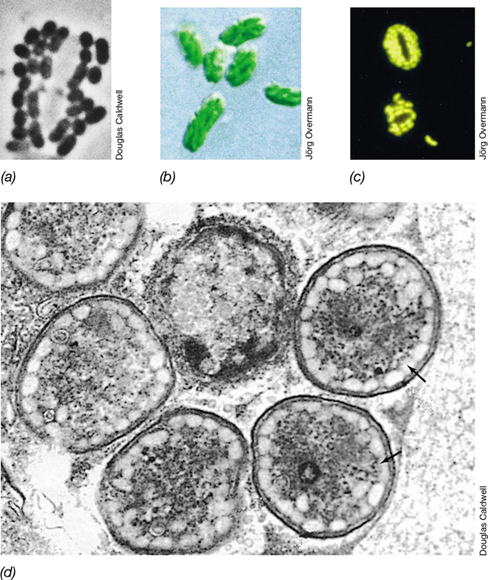
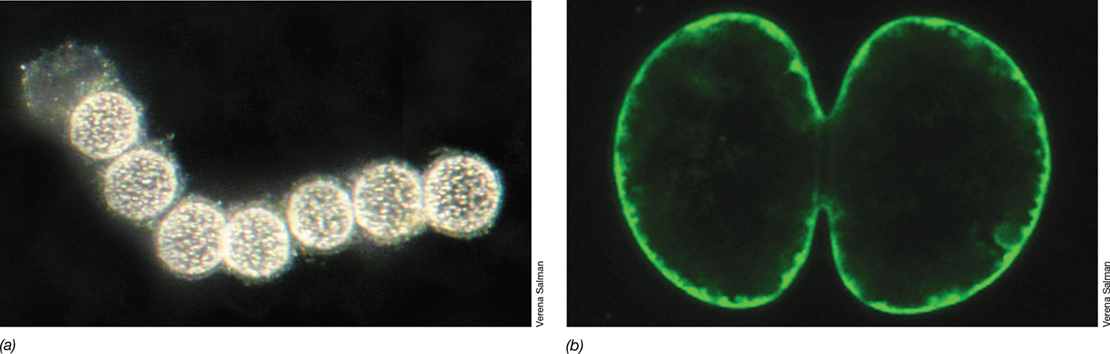
Cyanobacteria are able to form a variety of structures associated with energy storage, reproduction, and survival. Many Cyanobacteria produce extensive mucilaginous envelopes, or sheaths, that bind groups of cells or filaments together (Figure 15.2a). Some filamentous cyanobacteria form hormogonia (Figure 15.6), short, motile filaments that break off from longer filaments to facilitate dispersal in times of stress. Some species also form resting structures called akinetes (Figure 15.6c), which protect the organism during periods of darkness, desiccation, or cold. Akinetes are cells with thickened outer walls. When conditions improve, akinetes germinate by breaking down their outer wall and initiating growth of a new vegetative filament. Many Cyanobacteria also form a structure called cyanophycin. This structure is a copolymer of aspartic acid and arginine and is a nitrogen storage product; when nitrogen in the environment becomes deficient, cyanophycin is broken down and used as a cellular nitrogen source. Many species of the Nostocales and Stigonematales are also able to form heterocysts, as discussed next.
Figure 15.6 Structural differentiation in filamentous *Cyanobacteria*.

(a) Initial stage of hormogonium formation in Oscillatoria. Notice the empty spaces where the hormogonium is separating from the filament. (b) Hormogonium of a smaller Oscillatoria species. Notice that the cells at both ends are rounded. Cells are about 10 μm wide. Differential interference contrast microscopy. (c) Akinete (resting spore) of Anabaena in a phase-contrast micrograph, cells about 5 μm wide.
Heterocysts and Nitrogen Fixation
Many Cyanobacteria are capable of nitrogen fixation (Figure 15.3). The nitrogenase enzyme, however, is inhibited by oxygen, and thus nitrogen fixation cannot occur along with oxygenic photosynthesis (Section 3.12). Cyanobacteria have evolved several regulatory mechanisms for separating nitrogenase activity from photosynthesis (Section 8.9). For example, many unicellular Cyanobacteria, such as Cyanothece and Crocosphaera (**Figure 15.7*a***), fix nitrogen only at night when photosynthesis does not occur. In contrast, the filamentous cyanobacteria Trichodesmium (Figure 15.7b) fixes nitrogen only during the day through a mechanism that remains unclear but appears to require transient suppression of photosynthetic activity (and its O2 production) within filaments. Finally, many filamentous Cyanobacteria of the Nostocales and Stigonematales facilitate nitrogen fixation by forming specialized cells called heterocysts, either on the ends of filaments (**Figure 15.8*a***, b) or along the filament (Figure 15.8c, d).
Figure 15.7 Marine Cyanobacteria that fix N2.

(a) Unicellular Crocosphaera-like cells in the process of dividing; cells are approximately 5 μm diameter. (b) Colonial “tuft” of Trichodesmium. The tuft is composed of many attached, undifferentiated, unbranched filaments and has a diameter of approximately 100 μm. (c) A diatom containing the cyanobacterial symbiont Richelia (scale in micrometers). The Richelia symbiont is an unbranched filament with a terminal heterocyst; cells are about 5 μm wide.
Figure 15.8 Heterocysts.

Differentiation of heterocysts causes the loss of photopigments and inability to carry out photosynthesis. (a) Phase-contrast micrograph of Calothrix with terminal heterocysts. (b) Fluorescence micrograph of the same Calothrix filaments; cells are about 10 μm wide. (c) Phase-contrast micrograph of Fischerella. (d) Fluorescence micrograph of the same Fischerella filaments; cells are about 10 μm wide. See how heterocyst formation is regulated at the genetic level in the well-studied cyanobacterium Anabaena in Figure 8.21.
Heterocysts arise from differentiation of vegetative cells and are the sites of nitrogen fixation in heterocystous Cyanobacteria. Heterocysts are surrounded by a thickened cell wall that slows the diffusion of O2 into the cell and permits nitrogenase activity to occur in an anoxic environment. Heterocysts lack photosystem II, the oxygen-evolving photosystem that generates reducing power from H2O (Section 14.6), and thus do not fluoresce as strongly as vegetative cells (Figure 15.8). Without photosystem II, heterocysts are unable to fix CO2 and thus lack the necessary electron donor (pyruvate) for nitrogen fixation. However, heterocysts have intercellular connections with adjacent vegetative cells that allow for mutual exchange of materials between these cells. Fixed carbon is imported by the heterocyst from adjacent vegetative cells, and this is oxidized to yield electrons for nitrogen fixation. The products of photosynthesis move from vegetative cells to heterocysts, and fixed nitrogen moves from heterocysts to vegetative cells (Figure 8.21).
Ecology of *Cyanobacteria Cyanobacteria* are of central importance to the productivity of the oceans. Small unicellular *Cyanobacteria*, such as *Synechococcus* and *Prochlorococcus* (Section 20.11), are the most abundant phototrophs in the oceans. Together these organisms contribute 80% of marine photosynthesis and 35% of all photosynthetic activity on Earth.
Cyanobacterial nitrogen fixation represents the dominant input of new nitrogen into vast segments of Earth’s oceans, particularly in oligotrophic tropical and subtropical waters. Marine nitrogen fixation is dominated by two groups of Cyanobacteria, the unicellular species, such as Crocosphaera, and the filamentous Trichodesmium. Crocosphaera (Figure 15.7a) and relatives dominate nitrogen fixation in most of the Pacific Ocean and are widespread in tropical and subtropical habitats. Trichodesmium is the dominant nitrogen-fixer in the North Atlantic Ocean and parts of the Pacific where dissolved iron concentrations are elevated. Trichodesmium forms macroscopically visible tufts of filaments (Figure 15.7b) and relies on gas vesicles to remain suspended in the photic zone, where it is often observed in dense masses of cells called blooms. In addition, other marine nitrogen-fixers including species of Calothrix and Richelia form symbiotic associations with diatoms (Figure 15.7c); these symbiotic associations are often observed in tropical and subtropical oceans. Finally, heterocystous cyanobacteria such as Nodularia (Figure 15.2d) and Anabaena can sometimes dominate nitrogen fixation in cold waters of the Northern Hemisphere and are often observed in the Baltic Sea.
Cyanobacteria are also widely found in terrestrial and freshwater environments. In general, they are more tolerant of environmental extremes, particularly extremes of desiccation, than are eukaryotic algae. Cyanobacteria are often the dominant or sole oxygenic phototrophic organisms in hot springs, saline lakes, desert soils, and other extreme environments. In some of these environments, cyanobacterial mats of variable thickness may form (Figure 20.8). Freshwater lakes, especially those rich in inorganic nutrients, often develop blooms of Cyanobacteria, especially in late summer when temperatures are warmest (Figures 20.1 and 20.19b). A few Cyanobacteria are symbionts of liverworts, ferns, and cycads, and a number are phototrophic components of lichens, a symbiosis between a phototroph and a fungus (Section 23.1).
Several metabolic products of Cyanobacteria are of considerable practical importance. Some Cyanobacteria produce potent neurotoxins, and toxic blooms may form when massive accumulations of Cyanobacteria develop. Animals ingesting water containing these toxic products may be killed. Many Cyanobacteria are also responsible for the production of earthy odors and flavors in some freshwater, and if such waters are used as drinking water sources, aesthetic problems may arise. The major compound produced is geosmin, a substance also produced by many actinomycetes (Section 16.12).
Check Your Understanding
What are the differentiating properties of the five major morphological groups of Cyanobacteria?
Why are cyanobacteria such as Synechococcus and Prochlorococcus so important to the global oxygen budget?
Key Genera: *Chromatium, Thermochromatium, Ectothiorhodospira*
Purple sulfur bacteria are anoxygenic phototrophs that use hydrogen sulfide (H2S) as an electron donor for photosynthesis (Figure 14.5). Purple sulfur bacteria are a phylogenetically coherent group found within the order Chromatiales in the Gammaproteobacteria.
Purple sulfur bacteria are generally found in illuminated anoxic zones where H2S is present. Such habitats occur commonly in lakes, marine sediments, and “sulfur springs,” where H2S produced geochemically or biologically can support the growth of purple sulfur bacteria (Figure 15.9). Purple sulfur bacteria are also commonly found in microbial mats (Section 20.5) and in salt marsh sediments. The characteristic color of purple sulfur bacteria comes from their carotenoids, accessory pigments involved in light harvesting (Section 14.4). These bacteria use a Q-type photosystem (Figure 14.16), contain either bacteriochlorophyll a or b, and carry out CO2 fixation by the Calvin cycle (Section 3.12).
Figure 15.9 Anoxygenic phototrophs in aquatic environments.

Groundwater passing through sedimentary rock can introduce sulfur minerals into lakes and ponds. (a) Purple sulfur bacteria, such as Lamprocystis roseopersicina (shown here), grow on the sediment of a spring-fed pool and float to the surface (by virtue of their gas vesicles) when disturbed. The purple color is from the photopigments of the purple sulfur bacteria and the green color is from cells of the alga Spirogyra. (b) Sample of water from a depth of 7 m in Lake Mahoney, British Columbia; the major phototroph is the purple sulfur bacterium Amoebobacter purpureus. These phototrophs grow in a dense purple cloud at the chemocline, where sulfide-rich bottom water is exposed to light from the surface. (c) Phase-contrast photomicrograph of water from the chemocline of Lake Dagow, Germany, containing several purple and green sulfur bacteria. Abbreviations: Tr, Thiopedia rosea (purple sulfur bacteria); Ca, green sulfur bacteria partially disaggregated from “Chlorochromatium aggregatum” (a phototrophic consortium, see Section 15.6); Cn, Chloronema sp. (green nonsulfur bacteria, see Section 15.7); Pr, “Pelochromatium roseum” (a phototrophic consortium, Section 23.2); Plr, Planktothrix rubescens (cyanobacteria, Section 15.3). Gas vesicles (Section 2.7) within Tr, Cn, and Plr cells cause them to appear refractile. Each Cn cell is about 5 μm long.
During autotrophic growth of purple sulfur bacteria, H2S is oxidized to elemental sulfur (S0), which is deposited as sulfur granules (Figure 15.10). When sulfide is limiting, the sulfur is used as an electron donor for photosynthesis, resulting in the oxidation of S0 to sulfate (SO4 2−). Many purple sulfur bacteria can also use other reduced sulfur compounds as photosynthetic electron donors; for example, thiosulfate (S2O3 2−) is commonly used to grow laboratory cultures.
Figure 15.10 Bright-field and phase-contrast photomicrographs of purple sulfur bacteria.

(a) Chromatium okenii; cells are about 5 μm wide. Note the globules of elemental sulfur inside the cells. (b) Thiospirillum jenense, a very large, polarly flagellated spiral; cells are about 30 μm long. Note the sulfur globules. (c) Thiopedia rosea; cells are about 1.5 μm wide. (d) Phase-contrast micrograph of cells of Ectothiorhodospira mobilis; cells are about 0.8 μm wide. Note external sulfur globules (arrow).
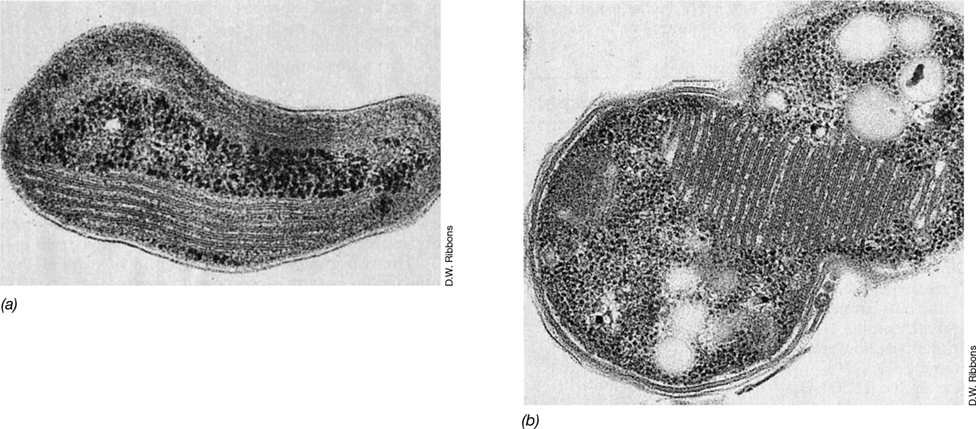
The purple sulfur bacteria form two families: the Chromatiaceae and the Ectothiorhodospiraceae. Species of the two families are readily distinguished by the location of sulfur granules and by their photosynthetic membranes. Chromatiaceae, including the genera Chromatium and Thiocapsa, store S0 granules inside their cells (in the periplasmic space) and have vesicular intracellular photosynthetic membrane systems (**Figure 15.11*b***). These organisms are common in stratified lakes containing sulfide and in the anoxic sediments of salt marshes. Ectothiorhodospiraceae, including the two main genera Ectothiorhodospira and Halorhodospira, oxidize H2S to S0 that is deposited outside the cell (Figure 15.10d) and have lamellar intracellular photosynthetic membrane systems (Figure 15.11a). These genera are also interesting because many species are extremely halophilic (salt loving) or alkaliphilic (alkalinity loving) and are among the most extreme in these characteristics of all known Bacteria. These organisms are typically found in saline lakes, soda lakes, and salterns, where abundant levels of SO4 2− support sulfate-reducing bacteria (Section 21.4 and Section 15.11), the organisms that produce H2S.
Figure 15.11 Membrane systems of phototrophic purple bacteria as revealed by transmission electron microscopy.

(a) Ectothiorhodospira mobilis, showing the photosynthetic membranes in flat sheets (lamellae). (b) Allochromatium vinosum, showing the membranes as individual, spherical vesicles.
Purple sulfur bacteria are often observed in high density in meromictic (permanently stratified) lakes. Meromictic lakes form layers because they have denser (usually saline) water on the bottom and less dense water (usually freshwater) nearer the surface. If sufficient sulfate is present to support sulfate reduction, sulfide is produced in the sediments and diffuses upward into the anoxic bottom waters. The presence of sulfide and light in the anoxic layers of the lake allows purple sulfur bacteria to form dense cell masses (Figure 15.9b, c), usually in association with green phototrophic bacteria.
Thermochromatium, a thermophilic purple sulfur bacterium, inhabits sulfidic hot springs where it forms thin microbial biofilms (Section 4.9 and Figure 4.17a). The only known species of Thermochromatium, T. tepidum, has unique absorption properties and has become a model organism for the study of energy transfer from light-harvesting (antenna) pigments to the photosynthetic reaction center (Section 14.5).
Check Your Understanding
What is the source of the purple color from which the purple sulfur bacteria get their name?
Where would you expect to find purple sulfur bacteria in nature?
15.5 Purple Nonsulfur Bacteria and Aerobic Anoxygenic Phototrophs
15.5 Purple Nonsulfur Bacteria and Aerobic Anoxygenic Phototrophs
Key Genera: *Rhodospirillum, Rhodoferax, Rhodopseudomonas, Rhodobacter*
The purple nonsulfur bacteria are the most metabolically versatile of all microbes. Despite their name, they are not always purple; these organisms synthesize an array of carotenoids (Section 14.4) that can lend them a variety of spectacular colors (Figure 15.12). Together, these pigments give purple bacteria their colors, usually purple, red, or orange. Purple nonsulfur bacteria are typically photoheterotrophs (a condition where light is the energy source and an organic compound is the carbon source), and species are able to use a wide range of carbon sources and electron donors for photosynthesis, including organic acids, amino acids, alcohols, sugars, and even aromatic compounds like benzoate or toluene. Like purple sulfur bacteria, purple nonsulfur bacteria use a Q-type photosystem and contain either bacteriochlorophyll a or b. The purple nonsulfur bacteria are phylogenetically and morphologically diverse (Figure 15.13) and reside within the Alphaproteobacteria (e.g., Rhodospirillum, Rhodobacter, Rhodopseudomonas) or Betaproteobacteria (e.g., Rubrivivax, Rhodoferax).
Figure 15.12 Photograph of liquid cultures of phototrophic purple bacteria showing the color of species with various carotenoid pigments.

All species contain bacteriochlorophyll a. The blue culture is a carotenoidless mutant strain of Rhodospirillum rubrum showing that bacteriochlorophyll a is actually blue. The bottle on the far right (Rhodobacter sphaeroides strain G) lacks one of the carotenoids of the wild type and thus is less red and more green.
Figure 15.13 Representatives of several genera of purple nonsulfur bacteria.

(a) Phaeospirillum fulvum; cells are about 3 μm long. (b) Rhodoblastus acidophilus; cells are about 4 μm long. (c) Rhodobacter sphaeroides; cells are about 1.5 μm wide. (d) Rhodopila globiformis; cells are about 1.6 μm wide. (e) Rhodocyclus purpureus; cells are about 0.7 μm in diameter. (f) Rhodomicrobium vannielii; cells are about 1.2 μm wide.
One interesting characteristic shared by all species of purple nonsulfur bacteria (including the aerobic species to be discussed in the next subsection) is the presence of a “photosynthetic gene cluster” (PGC). The PGC is a set of several connected operons whose genes encode the necessary proteins to carry out anoxygenic photosynthesis: pigment biosynthesis proteins, light-harvesting and reaction center proteins, and a variety of regulatory and other photosynthesis proteins (Section 14.5). Gene co-localization enables coordinated regulation of expression, but such co-localization also facilitates horizontal gene transfer. Genome sequence data indicates that horizontal transfer of PGC likely contributed to the evolution of phototrophy in lineages such as Gemmatimonadetes and Chloroflexi.
Purple nonsulfur bacteria can conserve energy through a variety of metabolic processes. For example, some species can grow photoautotrophically using H2, low levels of H2S, or even ferrous iron (Fe2+) as the electron donor for photosynthesis with CO2 fixation carried out by the Calvin cycle. Most species are also able to grow in darkness by using aerobic respiration of organic or even some inorganic compounds; synthesis of the photosynthetic machinery is typically repressed by O2. Finally, some species can also grow by fermentation or anaerobic respiration using a variety of electron donors and acceptors.
Enrichment and isolation of purple nonsulfur bacteria is easy using a mineral-salts medium supplemented with an organic acid (for example, acetate, malate, or succinate) as carbon source. Such media, inoculated with a mud, lake water, or sewage sample and incubated anaerobically in the light, invariably select for purple nonsulfur bacteria. Enrichment cultures can be made even more selective by omitting fixed nitrogen sources (for example, ammonia) or organic nitrogen sources (for example, yeast extract or peptone) from the medium and supplying a gaseous headspace of N2. Virtually all purple nonsulfur bacteria can fix N2 (Section 3.12) and will thrive under such conditions, rapidly outcompeting other bacteria.
Key Genera: *Roseobacter, Erythrobacter*
The aerobic anoxygenic phototrophs are obligatory aerobic heterotrophs that use light as a supplemental source of energy to support growth. Like purple nonsulfur bacteria, aerobic anoxygenic phototrophs are phylogenetically diverse and are Alphaproteobacteria or Betaproteobacteria. Their primary physiological difference from the purple nonsulfur bacteria is that aerobic anoxygenic phototrophs are strict heterotrophs and employ anoxygenic photosynthesis only under oxic conditions as a supplemental source of energy. Aerobic anoxygenic phototrophs contain bacteriochlorophyll a and a Q-type photosystem but are unable to fix CO2 and thus rely on organic forms of carbon as their carbon source. Carotenoids of various types lend colors of yellow, orange, or pink to cultures.
Aerobic anoxygenic phototrophs are only able to photosynthesize when grown on a day/night cycle. Under these conditions, bacteriochlorophyll a is made only in the dark and then used to conserve energy by photophosphorylation when the light returns. Aerobic anoxygenic phototrophs can account for as much as a quarter of the microbial community inhabiting coastal marine waters and 5% of gross photosynthesis in such systems (Section 20.11). Common genera found in coastal marine habitats include Roseobacter and Erythrobacter.
Check Your Understanding
What are some similarities between purple nonsulfur bacteria and aerobic anoxygenic phototrophs? What are the differences between these two groups?
Where would you expect to find aerobic anoxygenic phototrophs?
Key Genera: *Chlorobium, Chlorobaculum, Prosthecochloris, “Chlorochromatium”*
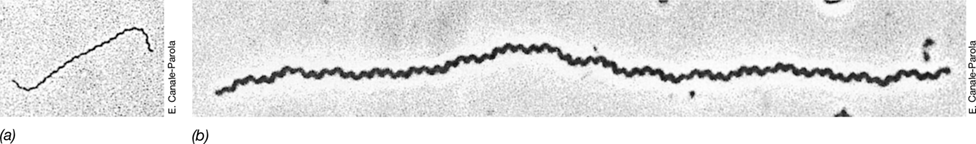
Green sulfur bacteria are a phylogenetically coherent group of anoxygenic phototrophs that forms the phylum Chlorobi. Green sulfur bacteria have little metabolic versatility, and they are typically nonmotile and strictly anaerobic anoxygenic phototrophic bacteria. The group is also morphologically restricted and includes primarily short to long rods (Figure 15.14).
Figure 15.14 Phototrophic green sulfur bacteria.

(a) Phase-contrast micrograph of Chlorobium limicola; cells are about 0.8 μm wide. Note the spherical sulfur granules deposited extracellularly. (b) Phase-contrast micrograph of Chlorobium clathratiforme, a bacterium forming a three-dimensional network; cells are about 0.8 μm wide. (c) Scanning electron micrograph of cells of Prosthecochloris aestuarii showing numerous prosthecae on each cell; cells are about 1.2 μm long. (d) Transmission electron micrograph of a P. aestuarii cell showing chlorosomes (arrow) within prosthecae.
Like purple sulfur bacteria, green sulfur bacteria oxidize hydrogen sulfide (H2S) as an electron donor for autotrophic growth, oxidizing it first to sulfur (S0) and then to sulfate (SO4 2−). But unlike most purple sulfur bacteria, the S0 produced by green sulfur bacteria is deposited only outside the cell (Figure 15.14a). Autotrophy is supported not by the reactions of the Calvin cycle, as in purple bacteria, but instead by a reversal of steps in the citric acid cycle (Section 14.2), a unique means of autotrophy in phototrophic bacteria.
Pigments and Ecology
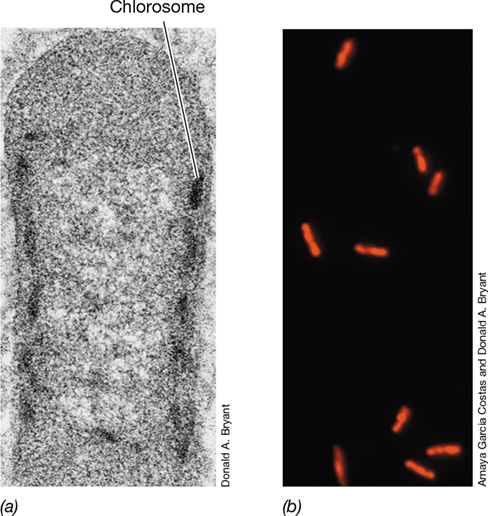
Green sulfur bacteria contain bacteriochlorophyll c, d, or e and house these pigments in unique structures called chlorosomes (Section 14.3 and Figure 15.15). A small amount of bacteriochlorophyll a is present in the reaction center and FMO protein, the latter of which connects the chlorosome to the cytoplasmic membrane (Figure 14.11b). Chlorosomes are oblong, bacteriochlorophyll-rich bodies bounded by a thin, nonunit membrane and attached to the cytoplasmic membrane in the periphery of the cell (Figure 15.15and Figure 14.11). Chlorosomes function to funnel energy into the photosystem, and this eventually leads to ATP synthesis. Unlike purple anoxygenic phototrophs, green sulfur bacteria use an FeS-type photosystem. Both green- and brown-colored species of green sulfur bacteria are known, the brown-colored species containing bacteriochlorophyll e and carotenoids that turn dense cell suspensions brown (Figure 15.16).
Figure 15.15 The thermophilic green sulfur bacterium *Chlorobaculum tepidum*.

Transmission electron micrograph. Note chlorosomes (arrow) in the cell periphery. A cell is about 0.7 μm wide.
Figure 15.16 Green and brown chlorobia.

Tube cultures of (a) Chlorobaculum tepidum and (b) Chlorobaculum phaeobacteroides. Cells of C. tepidum contain bacteriochlorophyll c and green carotenoids, and cells of C. phaeobacteroides contain bacteriochlorophyll e and isorenieratene, a brown carotenoid. The structures of bacteriochlorophylls c and e and of green bacteria carotenoids were shown in Figures 14.7 and 14.13, respectively.
Like purple sulfur bacteria (Section 15.4), green sulfur bacteria live in anoxic, sulfidic, illuminated aquatic environments. However, the chlorosome is a very efficient light-harvesting structure, which allows green sulfur bacteria to grow at light intensities much lower than those required by other phototrophs. Green sulfur bacteria also tend to have a greater tolerance of H2S than do other anoxygenic phototrophs. As a result, green sulfur bacteria are typically found at the greatest depths of all phototrophic microorganisms in lakes or microbial mats, where light intensities are low and H2S levels the highest. As an example, a species of green sulfur bacteria isolated from a deep-sea hydrothermal vent (Section 20.16) was found to be growing phototrophically on the weak glow of infrared radiation emitted from the geothermally heated rock.
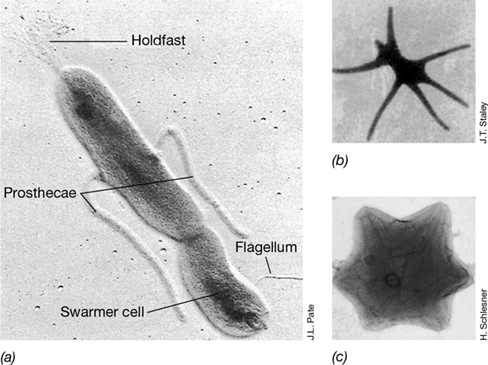
Although most green sulfur bacteria are freshwater species, some are marine and at least one species is thermophilic. Prosthecochloris is a marine species whose cells form extensions of the cytoplasm and cell wall called prosthecae (see Section 15.18 and Figure 15.53) that contain chlorosomes (Figure 15.14c, d). Some Prosthecochloris species inhabit saline lakes and appear to be the only green sulfur bacteria to be widespread outside of freshwater habitats. Chlorobaculum tepidum (Figure 15.15) is thermophilic and forms dense microbial mats in high-sulfide hot springs. C. tepidum also grows rapidly and is amenable to genetic manipulation by both conjugation and transformation (Chapter 9). In addition, C . tepidum can use thiosulfate as an electron donor for autotrophic CO2 fixation, a property not universal among green sulfur bacteria. Because of these features, C. tepidum has become the model organism for studying the molecular biology of green sulfur bacteria.
Green Sulfur Bacteria Consortia
Certain species of green sulfur bacteria form an intimate two-membered association, called a consortium, with a chemoorganotrophic bacterium. In the consortium, each organism benefits, and thus a variety of such consortia containing different phototrophic and chemotrophic components probably exist in nature. The phototrophic component, called the epibiont, is physically attached to the nonphototrophic central cell (Figure 15.17) and communicates with it in various ways (Section 23.2).
Figure 15.17 “*Chlorochromatium aggregatum*.”

Consortia of green sulfur bacteria and a chemoorganotroph (see also Figure 15.9 and Section 23.2). (a) In a phase-contrast micrograph, the nonphototrophic central organism is lighter in color than the pigmented phototrophic bacteria. (b) Green carotenoids lend their color to the phototrophs in a differential interference contrast micrograph. (c) A fluorescence micrograph shows the cells stained with a fluorescent probe specific for green sulfur bacteria. (d) Transmission electron micrograph of a cross section through a single consortium; note the chlorosomes (arrows) in the epibionts. The entire consortium is about 3 μm in diameter.
The name “Chlorochromatium aggregatum” (not a formal name because this is a mixed culture) has been used to describe a commonly observed green-colored consortium that is green because the epibionts are green sulfur bacteria that contain green-colored carotenoids (Figure 15.17b and Figure 15.9c). Evidence that the epibionts are indeed green sulfur bacteria comes from pigment analyses, the presence of chlorosomes (Figure 15.17d), and phylogenetic staining (Figure 15.17c). A structurally similar consortium called “Pelochromatium roseum” is brown because its epibionts produce brown-colored carotenoids (Figure 15.9c and Figures 23.3–23.5). We examine the symbiotic nature of the Chlorochromatium consortium in more detail in (Section 23.2).
Check Your Understanding
What evidence exists that the epibionts of green bacterial consortia are truly green sulfur bacteria?
Key Genera: *Chloroflexus, Heliothrix, Roseiflexus*
Green nonsulfur bacteria, which are also called filamentous anoxygenic phototrophs, are anoxygenic phototrophs of the phylum Chloroflexi. The latter contains several distinct lineages, one of which, the class Chloroflexi, contains green nonsulfur bacteria. The remainder of the phylum contains metabolically diverse organisms including both aerobic and anaerobic chemoorganotrophs as well as the Dehalococcoidetes, a group of dehalogenating bacteria that use halogenated organic compounds as electron acceptors in anaerobic respiration (Section 14.13). Analyses of 16S ribosomal RNA sequences from environmental samples (Sections 19.6 and 19.8) indicate that species of the phylum Chloroflexi are widespread and that most species in the phylum have yet to be cultivated in isolation; thus, the extent of the metabolic diversity of this phylum remains unclear.
All cultured representatives of the green nonsulfur bacteria are filamentous bacteria that are capable of gliding motility. Chloroflexus, one of the most studied of the green nonsulfur bacteria, forms thick microbial mats in neutral to alkaline hot springs along with thermophilic cyanobacteria (Figure 15.18; Figure 20.8b). Green nonsulfur bacteria grow best as photoheterotrophs using simple carbon sources as electron donors for photosynthesis. However, growth also occurs photoautotrophically using H2 or H2S as electron donors for photosynthesis. The 3-hydroxypropionate bi-cycle, a pathway of CO2 incorporation unique to only a few Bacteria and Archaea, supports autotrophic growth (Section 14.2). Most green nonsulfur bacteria also grow well in the dark by aerobic respiration of a wide variety of carbon sources.
Figure 15.18 Green nonsulfur bacteria.

(a) Phase-contrast micrograph of the anoxygenic phototroph Chloroflexus aurantiacus; cells are about 1 μm in diameter. (b) Phase-contrast micrograph of the large phototroph Oscillochloris; cells are about 5 μm wide. The brightly contrasting material on the top is a holdfast, used for attachment. (c) Phase-contrast micrograph of filaments of a Chloronema species; the cells are wavy filaments and about 2.5 μm in diameter. (d) Tube cultures of C. aurantiacus (right) and Roseiflexus (left). Roseiflexus is yellow because it lacks bacteriochlorophyll c and chlorosomes.
The photosynthetic features of the green nonsulfur bacteria form a “hybrid” between those of both green sulfur bacteria (Section 15.6) and purple phototrophic bacteria (Sections 15.4, 15.5). Green nonsulfur bacteria have reaction centers that contain bacteriochlorophyll a and chlorosomes that contain bacteriochlorophyll c and in this way are similar to green sulfur bacteria (Figure 15.15). However, in contrast to green sulfur bacteria, green nonsulfur bacteria contain a Q-type photosynthetic reaction center (Section 14.5 and Figure 14.16) and in this respect resemble purple bacteria.
Other *Chloroflexi*
In addition to Chloroflexus, other phototrophic green nonsulfur bacteria include the thermophile Heliothrix and the large-celled mesophiles Oscillochloris (Figure 15.18b) and Chloronema (Figure 15.18c). Oscillochloris and Chloronema form rather large cells, 2–5 μm wide and up to several hundred micrometers long (Figure 15.18c). Species of both genera inhabit freshwater lakes containing H2S. Roseiflexus and Heliothrix are similar to Chloroflexus in their filamentous morphology and thermophilic lifestyle but differ in a major photosynthetic property. Roseiflexus and Heliothrix lack bacteriochlorophyll c and chlorosomes and thus more closely resemble purple phototrophic bacteria (Sections 15.4, 15.5) than Chloroflexus. This can be seen in cultures of Roseiflexus that are yellow-orange instead of green from their extensive carotenoid pigments and lack of bacteriochlorophyll c (Figure 15.18d).
Thermomicrobium is a nonphototrophic genus of Chloroflexi and a strictly aerobic, gram-negative rod, growing optimally in complex media at 75 °C. Besides its phylogenetic properties, Thermomicrobium is also of interest because of its membrane lipids (Figure 15.19). Recall that the lipids of Bacteria and Eukarya contain fatty acids esterified to glycerol (Section 2.1). By contrast, the lipids of Thermomicrobium are formed on 1,2-dialcohols instead of glycerol and have neither ester nor ether linkages (Figure 15.19; Section 2.1). In addition, cells of Thermomicrobium contain only small amounts of peptidoglycan (Section 2.3), and the cell wall is composed primarily of protein.
Figure 15.19 The unusual lipids of *Thermomicrobium*.

(a) Membrane lipids from Thermomicrobium roseum contain long-chain diols like the one shown here (1,2-nonadecanediol). Note that unlike the lipids of other Bacteria or of Archaea, neither ester- nor ether-linked side chains are present. (b) To form a bilayer membrane, dialcohol molecules oppose each other at the methyl groups, and the —OH groups are the inner and outer hydrophilic surfaces. Small amounts of the diols have fatty acids esterified to the secondary —OH group (shown in red), whereas the primary —OH group (shown in green) can bond with a hydrophilic molecule like phosphate.
Check Your Understanding
In what ways do Chloroflexus and Roseiflexus resemble Chlorobium? Rhodobacter?
Key Genera: *Heliobacterium, Heliorestis*
Heliobacteria are a phylogenetically coherent group of phototrophic gram-positive Bacteria found within the phylum Firmicutes (Section 16.8). The heliobacteria are anoxygenic phototrophs that have an FeS-type photosystem (Section 14.5) and that produce a unique pigment, bacteriochlorophyll g (Figure 14.7). Heliobacteria grow photoheterotrophically using a narrow range of organic compounds including pyruvate, lactate, acetate, or butyrate, and the group contains five genera: Heliobacterium, Heliophilum, Heliorestis, Heliomonas, and Heliobacillus. All known heliobacteria form rod-shaped or filamentous cells (Figure 15.20), although Heliophilum is unusual because its cells form into bundles (Figure 15.20b) that are motile as a unit.
Figure 15.20 Cells and endospores of heliobacteria.

(a) Electron micrograph of Heliobacillus mobilis, a peritrichously flagellated species. (b) Heliophilum fasciatum cell bundles as observed by electron microscopy. (c) Phase-contrast micrograph of endospores from Heliobacterium gestii. Most heliobacteria cells are about 1–2 μm in diameter.
Heliobacteria are strict anaerobes, but in addition to phototrophic growth, they can grow chemotrophically in darkness by pyruvate fermentation (as can many clostridia, close relatives of the heliobacteria). Heliobacteria produce endospores, the highly resistant structures produced by certain gram-positive bacteria (Section 2.8). Like the endospores of Bacillus or Clostridium species, the endospores of heliobacteria (Figure 15.20c) contain elevated calcium (Ca2+) levels and the signature molecule of the endospore, dipicolinic acid. Heliobacteria reside in soil, especially paddy (rice) field soils, where their nitrogen fixation activities may benefit rice productivity. A large diversity of heliobacteria have also been found in highly alkaline environments, such as soda lakes and surrounding alkaline soils.
Key Genus: *Chloracidobacterium*
A novel group of anoxygenic phototrophs has been discovered growing in photosynthetic microbial mats of certain thermal springs in Yellowstone National Park (USA). Chloracidobacterium thermophilum is a thermophilic oxygen-tolerant anoxygenic phototroph of the phylum Acidobacteria (Section 16.21). Like green sulfur bacteria, C. thermophilum produces bacteriochlorophyll a and c, the latter in chlorosomes (Figure 15.21), and uses an FeS-type photosystem. However, unlike green sulfur bacteria, C. thermophilum can also grow aerobically, as is true for the aerobic anoxygenic phototrophs (Section 15.5). In terms of its carbon metabolism, C. thermophilum is a photoheterotroph that uses short-chain fatty acids as carbon sources, but unlike green sulfur or green nonsulfur bacteria, it is incapable of autotrophy.
Figure 15.21 Chlorosomes in *Chloracidobacterium thermophilum*, a phototrophic member of the phylum *Acidobacteria.*

(a) Electron micrograph of C. thermophilum showing chlorosomes. (b) Fluorescence photomicrograph of C. thermophilum. The red color is the fluorescence of bacteriochlorophyll c present in chlorosomes. A cell of C. thermophilum is about 0.8 μm wide.
Key Genus: *Gemmatimonas*
Another novel group of anoxygenic phototrophs has been discovered in a freshwater lake in the western Gobi desert (part of China and Mongolia). Gemmatimonas phototrophica is an aerobic facultative photoheterotroph of the phylum Gemmatimonadetes. It gains most of its energy through the aerobic respiration of organic compounds, whether in the light or in the dark. However, in the light, G. phototrophica uses photophosphorylation to supplement energy generated by aerobic respiration. G. phototrophica cannot grow as an obligate phototroph, it cannot fix CO2, and it cannot grow anaerobically. G. phototrophica contains a photosynthetic gene cluster that resembles those of aerobic anoxygenic phototrophs (Section 15.5), and it produces bacteriochlorophyll a and a Q-type reaction center, both of which are characteristic properties of purple bacteria (Section 14.5). It thus seems likely that G. phototrophica acquired its photosynthetic gene cluster and the ability to perform photophosphorylation as the result of an ancient horizontal gene transfer event.
Check Your Understanding
What types of anoxygenic phototrophs contain chlorosomes?
What kind of phototrophic bacteria make endospores?
III: Diversity of Bacteria Defined by Metabolic Traits
III: Diversity of Bacteria Defined by Metabolic Traits
III Diversity of *Bacteria* Defined by Metabolic Traits
Some microbial metabolisms, such as those where sulfur or iron are oxidized to yield energy, are so characteristic of a specific group of microbes that the metabolism becomes the defining characteristic of the group.
Certain types of microbial metabolism have major impacts on biogeochemical cycling of elements such as nitrogen and sulfur (see Chapter 21). Hence, it is useful to define the diversity of these organisms in terms of their metabolism. All cells must assimilate nitrogen, sulfur, and some metals for growth, and thus all organisms catalyze certain assimilatory reactions. The Bacteria and Archaea, however, are the only domains containing organisms that can conserve energy from the dissimilative metabolism of inorganic nitrogen, sulfur, and metallic compounds.
In this part of the chapter, we focus on the diversity of microorganisms defined by metabolic traits. We will see that considerable ecological and phylogenetic diversity often exists within any metabolic group and that ecological characteristics often govern the distribution and activity of species within each metabolic group (for example, see Section 15.12). We start our tour of the nitrogen cycle by considering those microbes that reduce atmospheric nitrogen: the nitrogen fixers.
Key Genera: *Mesorhizobium, Desulfovibrio, Azotobacter*
Diazotrophs are microorganisms that fix dinitrogen gas (N2) into NH3, which can be assimilated as a source of nitrogen for cells. [The term diazotroph is derived from a combination of the French azote, which means “nitrogen,” the Greek troph, which means “eating,” and di, which means two. Thus, a diazotroph is an organism that “eats two nitrogens” (N2)]. Nitrogen fixation is an assimilative process and requires ATP and the enzyme nitrogenase (Section 3.12). Diazotrophs typically fix N2 only when other forms of N are absent, and nitrogenase expression is inhibited when NH3 is available to cells. Nitrogenase is irreversibly inhibited by O2, and this is one cause of ecological diversification among diazotrophs; we will see that different organisms have evolved different solutions to protect nitrogenase from O2.
Nitrogen fixation is widespread among Bacteria and is also found in a few Archaea. It is thought to be an ancient process that was present in the last universal common ancestor (Section 13.1). The nifH gene encodes the dinitrogenase reductase component of nitrogenase (Section 3.12) and can be used as a measure of diazotroph diversity. More than 30,000 unique nifH gene sequences have been described, spanning nine bacterial phyla and one archaeal phylum (Figure 15.1). The phylogenetic distribution of nitrogenase in the tree of life has been influenced strongly by horizontal gene exchange. As a result, the phylogeny of nifH is largely inconsistent with the 16S ribosomal RNA gene phylogeny (Figure 15.22). We consider here the diversity of both symbiotic and free-living diazotrophic Bacteria.
Figure 15.22 Relationships among diazotrophic (nitrogen-fixing) bacteria as inferred from 16S ribosomal RNA gene sequences and NifH amino acid sequences.

Branches in each tree are colored to indicate phyla. The dashed lines indicate branches shared between the two trees. The incongruence between the two trees has resulted from multiple horizontal transfer events of the nifH gene. Red text denotes obligate anaerobes and underlined text indicates species that form symbioses with Eukarya.
Symbiotic Diazotrophs
Diazotrophs form several symbiotic relationships with plants, animals, and fungi. These relationships typically consist of a host that provides a hospitable environment, a source of carbon and energy, and a system for regulating O2 concentrations, and a microbial symbiont that provides a supply of fixed nitrogen to the host.
The symbiosis between rhizobia and leguminous plants is one of the best-characterized nitrogen-fixing symbiotic associations (Section 23.4). Root-nodule-forming bacteria are Alphaproteobacteria (e.g., Mesorhizobium, Bradyrhizobium, Sinorhizobium), Betaproteobacteria (e.g., Burkholderia), or Actinobacteria (e.g., Frankia). Other genera of symbiotic diazotrophs are found in association with shipworms (Teredinibacter), termite guts (Treponema) (Section 23.9), endomycorrhizal fungi (Glomeribacter) (Sections 18.12 and 23.5), and several fungi, algae, and plants (Cyanobacteria) (Sections 23.1 and 23.4). These different symbioses have evolved independently multiple times as a result of convergent evolution (Figure 15.22).
Free-Living Diazotrophs
Free-living diazotrophs need a mechanism for protecting nitrogenase from oxygen (Section 3.12 and Section 15.3). The simplest solution to this problem is to grow only in anoxic environments. The origin of nitrogen fixation predates the origin of oxygenic photosynthesis, which means the first nitrogen-fixing organisms were free-living anaerobes. Obligately anaerobic free-living diazotrophs are common in anoxic environments including marine and freshwater sediments and microbial mats. Obligately anaerobic free-living diazotrophs are found in the bacterial phyla Firmicutes (e.g., Clostridium), Chloroflexi (e.g., Oscillochloris), Chlorobi (e.g., Chlorobium), Spirochaetes (e.g., Spirochaeta), and Proteobacteria (e.g., Desulfovibrio, Chromatium, Rhodobacter) and in the archaeal phylum Euryarchaeota (e.g., Methanosarcina). Desulfovibrio species occur in anoxic salt marsh sediments dominated by Spartina grass, and their N2 fixation is an important nitrogen source to plants that live in this ecosystem.
Other simple mechanisms for protecting nitrogenase from oxygen include fixing N2 only at times when oxygen is absent or present in low concentration. For example, facultative aerobes will often fix N2 only while growing anaerobically (e.g., Klebsiella). Some aerobic nitrogen-fixers are microaerophiles; these organisms fix nitrogen only in environments where oxygen is present at low concentration (typically less than 2%). However, some organisms have evolved more complex mechanisms for protecting nitrogenase from oxygen and are able to grow in the presence of air.
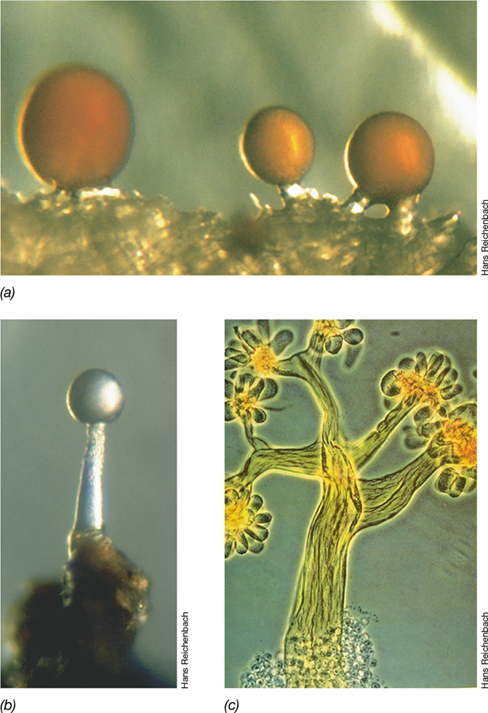
Obligately aerobic free-living diazotrophs include the Cyanobacteria (which have evolved a variety of mechanisms of protecting nitrogenase from oxygen, see Section 15.3) and several unicellular free-living chemoorganotrophic bacteria. Obligately aerobic free-living diazotrophs include Azotobacter, Azospirillum, and Beijerinckia. Azotobacter cells are large rods or cocci with diameters of 2–4 μm or more. When they are growing on N2 as a nitrogen source, extensive capsules or slime layers are typically produced (Figure 15.23 and Figures 2.16 and 3.29a, b). It is thought that the high respiratory rate characteristic of Azotobacter cells and the abundant capsular slime they produce help protect nitrogenase from O2. Azotobacter is able to grow on many different carbohydrates, alcohols, and organic acids, and metabolism is strictly oxidative.
Figure 15.23 Examples of slime production by free-living N2-fixing bacteria.

(a) Cells of Derxia gummosa encased in slime. Cells are about 1–1.2 μm wide. (b) Colonies of Beijerinckia species growing on a carbohydrate-containing medium. Note the raised, glistening appearance of the colonies due to abundant capsular slime.
Azotobacter can form resting structures called cysts (**Figure 15.24*b***). Like bacterial endospores, Azotobacter cysts show negligible endogenous respiration and are resistant to desiccation, mechanical disintegration, and ultraviolet and ionizing radiation. In contrast to endospores, however, cysts are not very heat resistant, and they are not completely dormant because they rapidly oxidize carbon sources if supplied.
Figure 15.24 *Azotobacter vinelandii.*

(a) Vegetative cells and (b) cysts visualized by phase-contrast microscopy. A cell measures about 2 μm in diameter and a cyst about 3 μm.
*Azotobacter* and Alternative Nitrogenases
We considered the important process of biological N2 fixation in Section 3.12 and discussed the central importance of the metals molybdenum (Mo) and iron (Fe) to the enzyme nitrogenase. The species Azotobacter chroococcum was the first nitrogen-fixing bacterium shown to grow on N2 in the absence of molybdenum. This is because either of two “alternative nitrogenases” are formed when Mo limitation prevents the MoFe nitrogenase from being synthesized. These nitrogenases are less efficient than the MoFe nitrogenase and contain either vanadium (V) or Fe in place of Mo. The three different types of nitrogenase (MoFe, VFe, and FeFe) are encoded by paralogous genes and likely arose as the result of gene duplications (Section 13.8). Subsequent investigations of other nitrogen-fixing bacteria have shown that these genetically distinct “backup” nitrogenases are widely distributed among nitrogen-fixing microbes, in particular in the Cyanobacteria and Archaea.
Check Your Understanding
What mechanisms do free-living diazotrophs use to protect nitrogenase from oxygen?
Where might you expect to find nitrogen-fixing bacteria?
15.10 Diversity of Nitrifiers and Denitrifiers
Microorganisms that grow by the anaerobic respiration of inorganic nitrogen (NO3 −, NO2 −) to the gaseous products NO, N2O, and N2 are called denitrifiers (Section 14.11). These organisms are typically facultative aerobes and chemoorganotrophs that use organic carbon as both carbon source and electron donor.
Microorganisms able to grow chemolithotrophically at the expense of reduced inorganic nitrogen compounds (NH3, NO2 −) are called nitrifiers (Figure 15.25; Section 14.11). These organisms are typically obligate aerobes that can also grow autotrophically; most species fix CO2 by the Calvin cycle. A few species have also been shown to grow mixotrophically by assimilating organic carbon in addition to CO2.
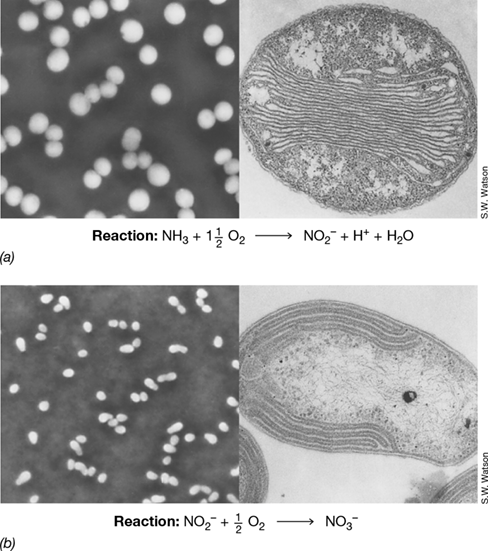
Figure 15.25 Nitrifying bacteria.

(a) Phase-contrast photomicrograph (left) and electron micrograph (right) of the ammonia-oxidizing bacterium Nitrosococcus oceani. A single cell is about 2 μm in diameter. (b) Phase-contrast photomicrograph (left) and electron micrograph (right) of the nitrite-oxidizing bacterium Nitrobacter winogradskyi. A cell is about 0.7 μm in diameter. Beneath each panel is the energy-conserving reaction that each organism catalyzes. The distinct internal membranes of each species are sites of key enzymes of nitrification.
Physiology of Nitrifying *Bacteria* and *Archaea*
Nitrification often results from the sequential activities of two physiological groups of organisms, the ammonia oxidizers (which oxidize NH3 to nitrite, NO2 −) (Figure 15.25a) and the nitrite oxidizers, the actual nitrate-producing microorganisms, which oxidize NO2 − to NO3 − (Figure 15.25b). Ammonia oxidizers typically have genus names beginning in Nitroso-, whereas genus names of nitrate producers begin with Nitro-. However, certain microbes within the genus Nitrospira are able to carry out both ammonia oxidation and nitrite oxidation and are therefore able to oxidize ammonia all the way to nitrate.
Many species of nitrifiers have internal membrane stacks (Figure 15.25) that closely resemble the photosynthetic membranes found in their close phylogenetic relatives, the purple phototrophic bacteria (Section 15.4) and the methane-oxidizing (methanotrophic) bacteria (Section 15.15). The membranes are the location of key enzymes in nitrification: ammonia monooxygenase, which oxidizes NH3 to hydroxylamine (NH2OH), and nitrite oxidoreductase, which oxidizes NO2 − to NO3 − (Section 14.9).
Enrichment cultures of nitrifying bacteria can be achieved using mineral salts media containing NH3 or NO2 − as electron donors and bicarbonate (HCO3 −) as the sole carbon source. Because these organisms produce very little ATP from their electron donors (Section 14.9), visible turbidity may not develop in cultures even after extensive nitrification has occurred. An easy means of monitoring growth is thus to assay for the production of NO2 − (with NH3 as electron donor) or NO3 − (with NO2 − as electron donor).
Key Genera: *Nitrosomonas, Nitrosospira, Nitrosopumilus*
Ammonia oxidizers are found in the Betaproteobacteria (e.g., Nitrosomonas, Nitrosospira, Nitrosolobus, Nitrosovibrio) and Gammaproteobacteria (Nitrosococcus), in the phylum Nitrospirae, and in the archaeal phylum Thaumarchaeota (Nitrosopumilus, Nitrosocaldus, Nitrosoarchaeum, Nitrososphaera).
Ammonia oxidizers are widespread in soil and water. Bacterial ammonia-oxidizers are present in highest numbers in habitats where NH3 is abundant, such as sites with extensive protein decomposition (ammonification), and also in sewage treatment facilities (Sections 22.6 and 22.7). Nitrifying bacteria develop especially well in lakes and streams that receive inputs of sewage or other wastewaters because these are frequently high in NH3. Nitrosomonas is often observed in the activated sludge present in aerobic wastewater treatment facilities. Bacterial ammonia-oxidizers are also common in soils (e.g., Nitrosospira, Nitrosovibrio) and in the oceans (e.g., Nitrosococcus).
Archaeal ammonia-oxidizers (Thaumarchaeota, Section 17.5) appear to be most common in habitats where NH3 is present in low concentration. These organisms are thought to be the dominant ammonia-oxidizers in the oceans where ammonia levels are very low (Sections 20.10 and 20.12). Archaeal ammonia-oxidizers are also common in soils, and in some soils they outnumber bacterial ammonia-oxidizers by several orders of magnitude. The availability of NH3 relative to NH4 + declines with pH (the substrate for the enzyme that oxidizes ammonia, ammonia monooxygenase, is NH3, not NH4 +) and thus acid soils (pH<6.5), which are common, may favor organisms able to grow at low NH3 concentration.
Key Genera: *Nitrospira, Nitrobacter*
Nitrite oxidizers are found in the classes Alpha- (Nitrobacter), Beta- (Nitrotoga), Gamma- (Nitrococcus), and Deltaproteobacteria (Nitrospina), as well as in the phylum Nitrospirae (genus Nitrospira) (Section 16.21).
Like nitrite-oxidizing Proteobacteria, Nitrospira oxidizes nitrite (NO2 −) to nitrate (NO3 −) and grows autotrophically (Figure 15.26). However, Nitrospira lacks the extensive internal membranes found in species of nitrifying Proteobacteria. Nevertheless, Nitrospira inhabits many of the same environments as nitrite-oxidizing Proteobacteria such as Nitrobacter, so it has been suggested that its capacity for NO2 − oxidation may have been acquired by horizontal gene flow from nitrifying Proteobacteria (or vice versa). As we know, this mechanism for acquiring physiological traits has been widely exploited in the bacterial world (Chapter 9 and Section 13.9). However, environmental surveys for the presence of nitrifying bacteria in nature have shown Nitrospira to be much more abundant than Nitrobacter; thus most of the NO2 − oxidized in natural environments is probably due to the activities of Nitrospira.
Figure 15.26 Nitrifying bacteria.

Nitrospira are a genus of nitrifying bacteria often found in biofilms and aggregates. The cell aggregates shown were obtained from activated sludge in a wastewater treatment facility. (a) Multiple cell aggregates (about 100 μm in diameter and >106 cells each) containing “Candidatus Nitrospira inopinata” (yellow), a species capable of completely oxidizing ammonia to nitrite. The biofilm was imaged by confocal laser microscopy using a phylogenetic stain specific for “Ca. N. inopinata” (Section 19.5). (b) A single aggregate of Nitrospira cells. Individual cells are curved (arrows) and group into tetrads, which then cluster together to form the larger aggregate. A single cell of Nitrospira is about 0.3×1–2 μm.
Key Genera: *Paracoccus, Pseudomonas*
Denitrifiers are capable of growth by the anaerobic respiration of NO3 − or NO2 − to the gaseous products NO, N2O, and N2 (Section 14.11). Nearly all denitrifiers are chemoorganotrophs that use organic carbon as both carbon source and electron donor. Exceptions include the denitrifying sulfur-oxidizers discussed in Section 15.12. Denitrifiers are typically facultative aerobes, and in nearly all cases they will grow preferentially as aerobes if O2 is present. Denitrifiers are of great importance in agricultural soils where they cause the loss of nitrogen fertilizers and the production of N2O, which is a dominant component of greenhouse gases produced by agricultural soils (Section 21.9).
Denitrifiers are phylogenetically and metabolically diverse and include two archaeal phyla (Section 17.10) and six bacterial phyla, including five classes of Proteobacteria (Figure 15.1). One of the best-characterized denitrifiers is Paracoccus denitrificans (Alphaproteobacteria). Denitrification of NO3 − to N2 requires several key enzymatic steps (Section 14.11), and the genes that encode these enzymes are present throughout the tree of life, indicating the strong influence of horizontal gene exchange. However, many nitrate reducers possess only part of the denitrification pathway and are thus unable to reduce NO3 − completely to N2, producing final products such as NO2 −, NO, or N2O.
Check Your Understanding
Under what conditions would you expect microorganisms to grow as a result of denitrification?
Which traits are shared among ammonia oxidizers and nitrite oxidizers?
15.11 Dissimilative Sulfur- and Sulfate-Reducers
Sulfur metabolism may have fueled the earliest forms of life on our planet (Section 13.1 and Figure 13.5), and the sulfur cycle (Section 21.4) continues to support an enormous diversity of microorganisms today. In this section, we consider the diversity of organisms capable of dissimilative sulfur metabolism; that is, organisms that conserve energy through the oxidation or reduction of sulfur compounds (Sections 14.7 and 14.12).
The remarkable diversity of Bacteria and Archaea capable of dissimilative sulfur metabolism is in part a function of the chemical diversity in which sulfur occurs in the biosphere. Sulfur has eight oxidation states that range from its most oxidized form, sulfate (SO4 2−, oxidation state of +6), to thiosulfate (S2O3 2−, oxidation state of +2), to elemental sulfur (S0, oxidation state of 0), and finally to hydrogen sulfide (H2S, oxidation state of −2), its most reduced form. In addition, sulfur compounds can take on diverse chemical forms including inorganic sulfur compounds, organosulfur compounds, and metal sulfides.
Key Genera: *Desulfovibrio, Desulfobacter*
Dissimilative sulfate-reducers gain energy by coupling the oxidation of H2 or organic compounds to the reduction of SO4 2− (anaerobic respiration, Section 14.12). The more than 30 known genera of sulfate reducers are found across five phyla of Bacteria and Archaea (Figure 15.27). Most sulfate reducers reside in the Deltaproteobacteria, though sulfate reducers are also found in the Firmicutes (e.g., Desulfotomaculum and Desulfosporosinus), Thermodesulfobacteria (e.g., Thermodesulfobacterium), and Nitrospirae (e.g., Thermodesulfovibrio). Sulfate reduction also occurs in Archaeoglobus, a genus of the archaeal phylum Euryarchaeota.
Figure 15.27 Dissimilative sulfate-reducers.

The dendrogram depicts phylogenetic relationships among some genera of sulfate reducers as inferred by analysis of their 16S ribosomal RNA gene sequences. Color shading is used to differentiate the five main phyla that contain genera of sulfate reducers. Colored circles indicate whether species are complete oxidizers, which are able to oxidize acetate to CO2, or incomplete oxidizers, which cannot oxidize acetate. The physiology of sulfate-reducing bacteria is considered in Section 14.12 and their role in the sulfur cycle in Section 21.4.
Physiology of Sulfate-Reducing Bacteria
Sulfate-reducing bacteria are morphologically and biochemically diverse. The biochemistry of sulfate reduction was discussed in Section 14.12, so here we consider some of the more general physiological properties of this group. Sulfate reducers are generally obligate anaerobes, and strict anoxic techniques must be used in their cultivation (**Figure 15.28*g***).
Figure 15.28 Representative sulfate-reducing and sulfur-reducing bacteria.

(a) Desulfovibrio desulfuricans; cell diameter about 0.7 μm. (b) Desulfonema limicola; cell diameter 3 μm. (c) Desulfobulbus propionicus; cell diameter about 1.2 μm. (d) Desulfobacter postgatei; cell diameter about 1.5 μm. (e) Desulfosarcina variabilis; cell diameter about 1.25 μm. (f) Desulfuromonas acetoxidans; cell diameter about 0.6 μm. (g) Enrichment culture of sulfate-reducing bacteria. Left, sterile medium; center, a positive enrichment showing black FeS; right, colonies of sulfate-reducing bacteria in a dilution tube (Section 19.2 and Figure 19.3b). Photos a–d and f are phase-contrast photomicrographs; part e is a differential interference contrast micrograph.
Sulfate reducers use H2 or organic compounds as electron donors for growth, and the range of organics used is broad. Lactate and pyruvate are almost universally used, and many species also oxidize short-chain alcohols (ethanol, propanol, and butanol) as electron donors. Some species, such as Desulfosarcina and Desulfonema, grow chemolithotrophically and autotrophically with H2 as an electron donor, SO4 2− as an electron acceptor, and CO2 as the sole carbon source. A few sulfate reducers can oxidize hydrocarbons as electron donors (Section 14.24).
There are two physiological types of dissimilative sulfate-reducers, the complete oxidizers, which can oxidize acetate and other fatty acids completely to CO2, and the incomplete oxidizers, which are unable to oxidize acetate to CO2. The latter group includes the best studied of the sulfate-reducing bacteria, Desulfovibrio (Figure 15.28a), along with Desulfomonas, Desulfotomaculum, and Desulfobulbus (Figure 15.28c). The acetate oxidizers include Desulfobacter (Figure 15.28d), Desulfococcus, Desulfosarcina (Figure 15.28e), and Desulfonema (Figure 15.28b), among many others. These bacteria specialize in the oxidation of fatty acids (in particular acetate) to CO2 and reducing SO4 2− to H2S. These two physiological groups are not phylogenetically coherent but instead are distributed widely across the phylogeny of sulfate-reducing bacteria (Figure 15.27).
Some sulfate-reducing bacteria can exploit alternative metabolic pathways. In addition to SO4 2− or S0, some sulfate reducers can also reduce nitrate and sulfonates (such as isethionate, HO—CH2—CH2—SO3 −). Sulfate-reducing bacteria can also ferment certain organic compounds such as pyruvate. Moreover, although generally obligate anaerobes, a few sulfate-reducing bacteria are quite O2-tolerant (primarily strains that coexist with O2- producing cyanobacteria in microbial mats). At least one species, Desulfovibrio oxyclinae, can actually grow with O2 as the electron acceptor under microaerophilic conditions.
Ecology of Sulfate-Reducing Bacteria
Sulfate reducers are widespread in aquatic and terrestrial environments that contain SO4 2− and become anoxic as a result of microbial decomposition. Sulfate reducers are abundant in marine sediments, and the H2S they generate is responsible for the pungent smell (like that of rotten eggs) often encountered in decaying vegetation near coastal ecosystems. Desulfotomaculum, phylogenetically a species of Firmicutes (gram-positive Bacteria), consists of endospore-forming rods found primarily in soil. Growth and reduction of SO4 2− by Desulfotomaculum in certain canned foods leads to a type of spoilage called sulfide stinker. Species of Thermodesulfobacterium, Thermodesulfovibrio, and Archaeoglobus (an archaeon) are all thermophilic and found in geothermally heated environments such as hot springs, hydrothermal vents, and oil reserves. The remaining genera of sulfate reducers are indigenous to anoxic marine and freshwater environments and can occasionally be isolated from the mammalian gut.
The enrichment of Desulfovibrio species is straightforward in an anoxic lactate–sulfate medium containing ferrous iron (Fe2+). A reducing agent, such as thioglycolate or ascorbate, is required to achieve a low reduction potential (E0′) in the medium. When sulfate-reducing bacteria grow, the H2S they form combines with the ferrous iron to form black, insoluble ferrous sulfide (Figure 15.28g). Purification can be accomplished by diluting the culture in molten agar tubes (Section 19.2 and Figure 19.3b). Upon solidification, individual cells of sulfate-reducing bacteria become distributed throughout the agar and grow to form black colonies (Figure 15.28g) that can be removed aseptically to yield pure cultures.
Key Genera: *Desulfuromonas, Wolinella*
Here we consider the dissimilative sulfur-reducers, microorganisms that are able to use the respiratory reduction of S0 to conserve energy. Dissimilative sulfur-reducing bacteria can reduce S0 and other oxidized forms of sulfur (such as SO3 2−) to H2S but are unable to reduce SO4 2−. There are more than 25 genera of dissimilative sulfur-reducers spread across five bacterial and archaeal phyla (Figure 15.1).
Most sulfur-reducing bacteria are Proteobacteria, primarily Deltaproteobacteria (e.g., Desulfuromonas, Pelobacter, Desulfurella, Geobacter), with some genera residing in the Epsilonproteobacteria (e.g., Wolinella and Sulfurospirillum) and Gammaproteobacteria (e.g., Shewanella and Pseudomonas mendocina). Other sulfur-reducing bacteria are species of Firmicutes (e.g., Desulfitobacterium and Ammonifex), Aquificae (e.g., Desulfurobacterium and Aquifex), Synergistetes (e.g., Dethiosulfovibrio), Deinococcus–Thermus (e.g., Thermus) or Deferribacteres (e.g., Geovibrio). The sulfur-reducing Archaea—of which there are many—are all genera of the phylum Crenarchaeota (e.g., Acidianus, Pyrodictium, and Thermodiscus).
Physiology and Ecology of Sulfur-Reducing Bacteria
The physiology of sulfur reducers is more diverse than that of sulfate reducers. Most sulfur reducers are obligate anaerobes, but facultatively aerobic species are also common. Sulfur reducers are often able to reduce electron acceptors such as nitrate, ferrous iron, or thiosulfate as alternatives to S0. Like sulfate reducers, the physiology of sulfur reducers is characterized by whether they completely oxidize acetate and other fatty acids to CO2. Species of Desulfuromonas (Figure 15.28f) are complete oxidizers that grow anaerobically by coupling the oxidation of acetate, succinate, ethanol, or propanol to the reduction of S0. In contrast, Sulfospirillum and Wolinella are incomplete oxidizers and cannot use acetate as an electron donor. Sulfospirillum can reduce S0 using either H2 or formate as electron donor.
Dissimilative sulfur-reducing bacteria reside in many of the same habitats as dissimilative sulfate-reducing bacteria and often form associations with bacteria that oxidize H2S to S0, such as green sulfur bacteria (Section 15.6). The S0 produced from H2S oxidation is then reduced back to H2S during metabolism of the sulfur reducer, completing an anoxic sulfur cycle (Section 21.4).
Check Your Understanding
What are the typical electron donors used by dissimilative sulfur-reducers?
What are the typical electron donors used by dissimilative sulfate-reducers?
What bacterial phyla are known to contain dissimilative sulfate-reducers?
Key Genera: *Thiobacillus, Achromatium, Beggiatoa*
Dissimilative sulfur-oxidizers are chemolithotrophs that oxidize reduced sulfur compounds such as H2S, S0, thiosulfate, or thiocyanate (SCN−) as electron donors in energy conservation, typically with O2 as electron acceptor. These organisms are common in environments such as marine sediments, sulfur springs, and hydrothermal systems where H2S produced by sulfate- or sulfur-reducing bacteria (Section 15.11), or abiotically by geothermal reactions, is released into oxygenated waters (Figure 15.29). The sulfur oxidizers are found in three phyla of Bacteria (Proteobacteria, Aquificae, Deinococcus–Thermus) and one of Archaea (Crenarchaeota) (Figure 15.1). Most sulfur-oxidizing bacteria are Beta- (Thiobacillus), Gamma- (Achromatium, Beggiatoa), or Epsilonproteobacteria (Thiovulum, Sulfurimonas).
Figure 15.29 Habitats of sulfur oxidizers.

(a) A sulfidic artesian spring in Florida (USA). The outside of the spring is coated with a mat of Thiothrix (see Figure 15.31b). The mat is about 1.5 m in diameter. (b) Hydrothermal chimneys at Cathedral Hill in the Guaymas Basin (Mexico), 2000-m depth. Sulfide-rich waters vent from the chimneys, which are covered by mats composed of orange, white, and yellow cells of Beggiatoa.
Physiological Diversity of Sulfur-Oxidizing Bacteria
The morphological and physiological diversity of sulfur oxidizers is significant. Cells can be less than 1 micrometer in diameter (e.g., Sulfurimonas denitrificans) or as large as 750 micrometers in diameter (e.g., Thiomargarita namibiensis). Most sulfur oxidizers are obligate aerobes; however, species of Thiomargarita and Sulfurimonas can also reduce NO3 − in denitrification (Section 14.11 and Section 15.10). Many species oxidize H2S to elemental sulfur (S0), which they deposit as either intracellular or extracellular granules for later use as an electron donor (Figure 14.19) if H2S becomes limiting.
Some sulfur chemolithotrophs are obligate chemolithotrophs, locked into a lifestyle of using inorganic instead of organic compounds as electron donors. When growing in this fashion, they are also autotrophs, converting CO2 into cell material by reactions of the Calvin cycle. Carboxysomes are often present in cells of obligate chemolithotrophs (**Figure 15.30*a***). These structures contain high levels of Calvin cycle enzymes and probably increase the rate at which these organisms fix CO2 (Section 14.2).
Figure 15.30 Nonfilamentous sulfur chemolithotrophs.

(a) Transmission electron micrograph of cells of the chemolithotrophic sulfur-oxidizer Halothiobacillus neapolitanus. A single cell is about 0.5 μm in diameter. Note the polyhedral bodies (carboxysomes) distributed throughout the cell (arrows) (Figure 14.3). (b) Achromatium. Cells photographed by differential interference contrast microscopy. The small globular structures near the periphery of the cells (arrow) are elemental sulfur, and the large granules are calcium carbonate. A single Achromatium cell is about 25 μm in diameter.
Other sulfur chemolithotrophs are facultative chemolithotrophs, facultative in the sense that they can grow either chemolithotrophically (and thus, also as autotrophs) or chemoorganotrophically. Most species of Beggiatoa can obtain energy from the oxidation of inorganic sulfur compounds but lack enzymes of the Calvin cycle. They thus require organic compounds as carbon sources. Organisms that use a mix of carbon and energy sources, for example those that simultaneously assimilate carbon from both CO2 and organic sources, are called mixotrophs.
*Thiobacillus* and *Achromatium*
The genus Thiobacillus and related genera include several gram-negative, rod-shaped Betaproteobacteria (Figure 15.30a) that are the best studied of the sulfur chemolithotrophs. The oxidation of H2S, S0, or thiosulfate by Thiobacillus generates sulfuric acid (H2SO4), and thus thiobacilli are often acidophilic. One highly acidophilic species, Acidithiobacillus ferrooxidans, can also grow chemolithotrophically by the oxidation of Fe2+ and is a major biological agent for the oxidation of this metal. Iron pyrite (FeS2) is a major natural source of ferrous iron as well as of sulfide. The oxidation of FeS2, especially in mining operations, can be both beneficial (because leaching of the ore releases the iron from the sulfide mineral) and ecologically disastrous (the environment can become acidic and contaminated with toxic metals such as aluminum, cadmium, and lead) (Sections 22.1 and 22.2).
Achromatium is a spherical sulfur-oxidizing chemolithotroph that is common in freshwater sediments of neutral pH containing H2S. Cells of Achromatium are large cocci that can have diameters of 10–100 μm (Figure 15.30b). Achromatium is a species of Gammaproteobacteria and is specifically related to purple sulfur bacteria, such as its phototrophic counterpart Chromatium (Section 15.4 and Figure 15.10a). Like Chromatium, cells of Achromatium store S0 internally (Figure 15.30b); the granules later disappear as S0 is oxidized to SO4 2−. Cells of Achromatium also store large granules of calcite (CaCO3) (Figure 15.30b), possibly as a carbon source (in the form of CO2) for autotrophic growth. The physiology of chemolithotrophic sulfur-oxidizers is discussed in Section 14.7.
Ecological Diversity and Strategies of Sulfide-Oxidizing Bacteria
Aerobic sulfide-oxidizers provide a case study that demonstrates the degree of ecological diversification that can occur among microbes that share the same basic metabolic features. The chemical oxidation of H2S to H2SO4 is spontaneous and rapid in the presence of O2. Hence, aerobic H2S-oxidizers have evolved diverse ecological strategies that allow them to metabolize two molecules that otherwise react with each other spontaneously. We consider here six different strategies used by aerobic sulfide-oxidizers to cope with the chemical instability of H2S in the presence of O2.
Thiothrix is a filamentous sulfur chemolithotroph that forms filaments that group together at their ends by way of a holdfast to form cell arrangements called rosettes (Figure 15.31). The ecological strategy of Thiothrix is to use its holdfast to position itself in high-flow environments downstream from a source of H2S. Such environments are common near sulfur springs and in creeks draining sulfidic salt marshes where abundant H2S is produced and carried away in waters rich with O2 (Figure 15.31a). Physiologically, Thiothrix is an obligately aerobic mixotroph, and in this and most other respects, it resembles Beggiatoa.
Figure 15.31 *Thiothrix.*

(a) Filaments of Thiothrix attached to plant material found in the outwash stream of a sulfidic cave in Frasassi, Italy. From the plant branch point, the longest branch is about 4 mm long. (b) Phase-contrast photomicrograph of a rosette of cells of Thiothrix isolated from the sulfide-containing artesian spring shown in Figure 15.29a. Note the internal sulfur globules produced from the oxidation of sulfide. Each filament is about 4 μm in diameter.
Beggiatoa are filamentous, gliding, sulfur-oxidizing bacteria that are usually large in both diameter and length, consisting of many short cells attached end to end (**Figure 15.32*a***). Filaments can flex and twist so that many filaments become intertwined to form a complex tuft. Beggiatoa is found primarily in microbial mats, sediments, sulfur springs, and hot springs. The ecological strategy of Beggiatoa is to use gliding motility to position itself at the point where H2S and O2 co-occur in an environment. For example, Beggiatoa in microbial mats can move vertically by as much as several centimeters per day in response to cyanobacterial O2 production, moving up to obtain O2 when photosynthesis ceases at night and down during the day when cyanobacterial O2 production at the mat surface causes H2S to be found deeper in the mat.
Figure 15.32 Filamentous sulfur-oxidizing bacteria.

(a) Phase-contrast photomicrograph of a Beggiatoa species isolated from a sewage treatment plant. Note the abundant elemental sulfur granules in some of the cells. (b) Cells of a large marine Thioploca species. Cells contain sulfur granules (yellow) and are about 40–50 μm wide.
The genus Thiomargarita contains some of the largest bacteria yet observed, with diameters that can be as much as 0.75 millimeter (Figure 15.33). Thiomargarita is nonmotile, and its ecological strategy is to separate in time the oxidation of H2S from the reduction of O2. To accomplish this, Thiomargarita contains a giant vacuole (Figure 15.33b) that it fills with high concentrations of nitrate (NO3 −). This vacuole can fill almost the entire volume of the cell. Cells live in sulfide-rich marine sediments that are mixed occasionally with O2-rich waters, such as that in salt marshes and in ocean upwelling zones. When buried in sediment, cells oxidize H2S to S0 anaerobically by reducing NO3 − stored in the vacuole to ammonium (NH4 +). They then store the S0 as intracellular granules (Figure 15.33a). When turbulent waters mix the cells into the water column where H2S is lacking, they switch to the aerobic oxidation of stored S0. The energy they gain from S0 oxidation is used to refill their vacuole with NO3 − from the water column so they will be able to survive the next period of anoxia.
Figure 15.33 The giant sulfide-oxidizing bacterium, *Thiomargarita.*

(a) Thiomargarita namibiensis recovered from the Namibian upwelling (off the Namibian coast, southwest Africa). Cells are about 100 μm in diameter. (b) Dividing cells of vacuole-containing sulfide-oxidizers recovered from the same location. The fluorescence micrograph shows ribosomes of Thiomargarita stained with a fluorescent nucleic acid probe. Ribosomes are found in the cytoplasm, which is present as a thin layer along the outer edge of the cells. The cytoplasm is squeezed between the cell wall and the large central vacuole, which appears dark in the image. Cells are about 50 μm wide.
Thioploca are large filamentous bacteria that use a strategy similar to that of Thiomargarita. Thioploca also have intracellular S0 granules and large vacuoles filled with NO3 − (Figure 15.32b). However, filaments of Thioploca are motile by gliding and they occur in large sheaths that can be filled with many parallel filaments (Figure 15.32b). Sheaths are arranged vertically in the sediments and filaments glide up and down in the sheaths, going down to anaerobically respire H2S using stored NO3 − as electron acceptor and going up to aerobically respire S0 and to refill their vacuoles with NO3 − (Figure 20.10).
Thiovulum are found in freshwater and marine habitats in which sulfide-rich muds interface with oxic zones (Figure 15.34). Thiovulum cells are fairly large (10–20 μm) cocci, and when motile, they swim at exceptionally high speed, perhaps the fastest of all known bacteria (∼0.6 mm/sec). The ecological strategy of Thiovulum is to actually control the flow of nutrients to cells. Thiovulum cells secrete a slime that links cells together in a veil-like structure that can be centimeters in diameter (Figure 15.34a). The veils, composed of many Thiovulum cells (Figure 15.34b), are formed over a source of H2S. Cells have long flagella that attach to the veil and to solid surfaces. Since the terminal end of the flagellum is attached and immobile, flagellar rotation causes cells to rotate along their flagellar axis (Figure 15.34c). The simultaneous unidirectional rotation of all the Thiovulum cells in the veil creates a flow of water through the veil, allowing the cells (Figure 15.34b, d) to generate and regulate the gradients of H2S and O2 they require to generate energy (Figure 15.34c).
Figure 15.34 The sulfide-oxidizing bacterium *Thiovulum.*

(a) Photo of a Thiovulum veil forming over marine sediment. The veil, composed of many cells within a polysaccharide matrix, appears like a spider web covering the sediment surface. Under the veil, H2S is released from the sediment. (b) Close-up photograph of Thiovulum cells (yellow dots) within their veil. (c) Thiovulum cells attach to the sediment with a long holdfast and begin to rotate. The collective rotation of the many cells within the veil cause O2-rich water above the veil to be pumped down across the veil. Hence, through collective action, Thiovulum cells manipulate their environment to optimize local concentrations of O2 and H2S. (d) Transmission electron micrograph of a section from a dividing cell of Thiovulum. Arrows point to sulfur (S0) globules. Single cells of Thiovulum are typically 10–20 μm in diameter.
The final ecological strategy of sulfur chemolithotrophs is for the organism to form a symbiotic association with a eukaryote. There are diverse symbiotic associations in which the host provides a mechanism for regulating H2S and O2 levels and the sulfide-oxidizing symbiont fixes CO2 and provides a source of carbon and energy to the host. The best example is the tube worm Riftia, which contains sulfide-oxidizing endosymbionts and lives at deep-sea hydrothermal vents (Section 23.11). A variety of other such symbiotic associations are present at hydrothermal vent ecosystems, including symbionts living in the gill tissue of the giant clam Calyptogena magnifica and on the surface of the yeti crab, which farms sulfide-oxidizing bacteria by waving its claws over sulfide-rich vent fluid. Symbioses involving invertebrates are also common in the sulfide-rich marine sediments of shallow coastal systems. For example, bivalves in the family Solemyidae burrow into sulfide-rich sediments and pump sulfide- and oxygen-rich water over gills that contain sulfide-oxidizing bacteria.
From these examples it should be clear how ecological diversity drives bacteria that carry out the same energy metabolism—in this case sulfide oxidation—to best exploit the different environments they inhabit. In each case, the goal of the organism is the same, to obtain the electron donor and acceptor it needs. But also in each case, the strategy to accomplish this is unique and the best fit to both the properties of the organism and the habitat it exploits.
Check Your Understanding
Describe the energy and carbon metabolism of Thiobacillus in terms of how ATP and new cell material are made.
What are some ecological strategies that sulfur oxidizers use to compete with the spontaneous (chemical) oxidation of H2S?
Key Genera: *Geobacter, Shewanella*
Dissimilative iron-reducers couple the reduction of oxidized metals or metalloids to cellular growth. These organisms need to overcome the fundamental obstacle of using an insoluble solid material as an electron acceptor in respiration (Section 14.13). A variety of microorganisms are able to enzymatically reduce metals as a consequence of either fermentation reactions or sulfur or sulfate reduction, but such organisms do not conserve energy from metal reduction. In contrast, dissimilative iron-reducers carry out metal respiration by coupling the oxidation of H2 or organic compounds to the reduction of ferric iron (Fe3+) (**Figure 15.35*a***) or manganese (Mn6+).
Figure 15.35 The dissimilative iron-reducing bacterium *Geobacter.*

(a) The uninoculated tube (left) contains an anoxic medium that includes acetate and ferrihydrite, a poorly magnetic iron oxide. Following growth of Geobacter (right tube) the ferrihydrite is reduced to magnetite, which is magnetic. (b) Transmission electron micrograph of Geobacter sulfurreducens showing flagella and pili (Figure 14.31). The cell is about 0.7×3.5 μm. (c) Transmission electron micrograph of G. sulfurreducens showing immunogold labeling of cytochrome OmcS on the pili (arrow).
Dissimilative iron-reducers are phylogenetically diverse (Figure 15.1). Iron-reducing bacterial genera are found in the Proteobacteria (Geobacter, Shewanella), Acidobacteria (Geothrix), Deferribacteres (Geovibrio), Deinococcus–Thermus (Thermus), Thermotogae (Thermotoga), and Firmicutes (Bacillus, Thiobacillus), while archaeal genera are found in the Crenarchaeota (Pyrobaculum). Iron respiration likely evolved early in the history of life, and its wide distribution may be due to its presence in the universal ancestor coupled with subsequent gene loss in some lineages and horizontal gene transfer to others.
Physiology
Dissimilative iron-reducers specialize in using insoluble external electron acceptors, and these organisms are typically extremely versatile at anaerobic respiration. Dissimilative iron-reducers are unusual in that they possess outer membrane cytochromes that facilitate electron transfer with insoluble minerals (Figure 14.31). Most species are able to use either iron oxides or manganese oxides as electron acceptors, and various species are also able to use nitrate, fumarate, and oxidized inorganic sulfur, cobalt, chromium, uranium, tellurium, selenium, arsenic, and humic compounds (Section 14.13). Most genera of iron-reducing bacteria are obligate anaerobes, but some, such as Shewanella and relatives, are facultative aerobes. Electron donors are typically organic compounds such as fatty acids, alcohols, sugars, and in certain cases, even aromatic compounds. Many species are also able to use H2 as an electron donor, but they are generally unable to grow autotrophically, requiring a source of organic carbon to support growth.
The family Geobacteraceae in the Deltaproteobacteria contains four genera of dissimilative iron-reducing bacteria (Geobacter, Desulfuromonas, Desulfuromusa, Pelobacter) that aptly demonstrate the physiological diversity of the obligately anaerobic metal reducers. Geobacter, Desulfuromonas, and Desulfuromusa can all use acetate as an electron donor as well as a diversity of other small organics, and they oxidize these substrates completely to CO2. These genera typically specialize in anaerobic respiration. Geobacter in particular can use a wide range of electron donors and acceptors. Geobacter produce pili (Figure 15.35b) that contain cytochromes (Figure 15.35c), and these pili facilitate electron transfer to the surface of iron oxide minerals. Pelobacter, in contrast, are primarily fermentative organisms having a more limited respiratory capacity. For example, Pelobacter carbinolicus can only use lactate as the electron donor and can only use ferric iron or S0 as the electron acceptors. Pelobacter species are unable to oxidize their carbon substrates completely to CO2.
Shewanella and its relatives Ferrimonas and Aeromonas in the Gammaproteobacteria are facultative aerobes and will grow aerobically when O2 is available. Shewanella are able to use a wide diversity of electron donors and acceptors in addition to ferric iron and manganese. However, like Pelobacter species, they are unable to oxidize their carbon substrates completely to CO2 and are unable to oxidize acetate as an electron donor for anaerobic respiration.
Ecology
Dissimilative iron-reducers are common in anoxic freshwater and marine sediments. These organisms are thought to play an important role in organic matter oxidation in many anoxic habitats. Dissimilative iron-reducers are also common in shallow aquifers as well as in the deep subsurface environment (Section 20.8). In addition, several thermophilic and hyperthermophilic iron-reducing species are known (e.g., Thermus, Thermotoga) and are often found in hot springs and other geothermally heated systems, including the deep subsurface.
Check Your Understanding
In what phylogenetic groups are Geobacter and Shewanella found?
Which genera of dissimilative iron-reducers contain facultative aerobes?
Key Genera: *Acidithiobacillus, Gallionella*
The ability to couple the oxidation of ferrous iron (Fe2+) to cell growth is widespread in the tree of life and thought to be a trait that evolved early in Earth’s history. Genera capable of using ferrous iron as an electron donor to support growth are spread across five bacterial and two archaeal phyla (Figure 15.1).
Aerobic iron-oxidizer diversity and distribution are influenced strongly by pH and O2. Ferrous iron oxidizes spontaneously to form insoluble precipitates in the presence of O2 at neutral to alkaline pH (pH>7) but is stable either under anoxic conditions or aerobically at acidic pH (pH<4). Iron oxidizers can be divided into four functional groups on the basis of their physiology: acidophilic aerobic iron-oxidizers, neutrophilic aerobic iron-oxidizers, anaerobic chemotrophic iron-oxidizers, and anaerobic phototrophic iron-oxidizers.
Acidophilic Aerobic Iron-Oxidizing Bacteria
The growth of iron-oxidizing bacteria is favored in iron-rich acidic environments where soluble ferrous iron is present. Aerobic iron-oxidizers are often abundant in acid mine drainage generated from abandoned coal or iron mines or from mine tailings (Sections 22.1 and 22.2). Acidophilic aerobic iron-oxidizers also inhabit iron-rich acidic springs in volcanic areas. In these environments, sulfur is often present along with ferrous iron, and many acidophilic aerobic iron-oxidizers are able to oxidize both elemental sulfur and ferrous iron. Species can be either autotrophic or heterotrophic, and commonly observed genera include Acidithiobacillus (Gammaproteobacteria), Leptospirillum (Nitrospirae), and Ferroplasma (Euryarchaeota). Other acidophilic aerobic iron-oxidizers can be found in the Actinobacteria and Firmicutes.
Neutrophilic Aerobic Iron-Oxidizing Bacteria
Neutrophilic aerobic iron-oxidizers are organisms adapted to a specialized niche (Section 14.8 and see Microbiology Now). This is because ferrous iron is relatively insoluble at neutral pH and its chemical oxidation is spontaneous and rapid in the presence of air. Furthermore, at neutral pH, iron oxidation at the cell surface causes the formation of an iron oxide crust that can effectively entomb growing cells. Neutrophilic aerobic iron-oxidizers therefore thrive where iron-rich anoxic waters are exposed to air. Such habitats are common near wetlands or soils where anoxic groundwater forms a spring, but iron oxidizers also inhabit the rhizosphere of wetland plants and certain submarine hydrothermal systems.
Few genera of neutrophilic aerobic iron-oxidizers have been described and they all belong to the Proteobacteria. Those species found in freshwater habitats belong to a set of closely related genera in the Betaprotobacteria, while species found in marine habitats belong to the Zetaproteobacteria. The metabolism of these organisms is fairly narrow. Species are typically microaerophiles and obligate chemolithotrophs, though in certain cases mixotrophy has been observed. The genera Leptothrix and Sphaerotilus are exceptions (Section 15.19). Leptothrix and Sphaerotilus are common in freshwater environments containing neutrophilic aerobic iron-oxidizers. They catalyze the oxidation of both iron and manganese but do not appear to conserve energy from these reactions, conserving energy instead from the oxidation of organic matter.
Characteristic species of neutrophilic aerobic iron-oxidizers are found in the genus Gallionella (freshwater) and the marine genus Mariprofundus (marine)(Section 21.6 and Figure 21.19). Species of Gallionella and Mariprofundus each form a twisted stalklike structure containing Fe(OH)3 from the oxidation of ferrous iron (Figure 15.36). The iron-encrusted stalk contains an organic matrix on which Fe(OH)3 accumulates as it is excreted from the cell surface. Stalk formation is presumably an adaptation that prevents cells from becoming entombed in an iron oxide crust.
Figure 15.36 The neutrophilic ferrous iron-oxidizer *Gallionella ferruginea*, from an iron seep near Ithaca, New York.

(a) Photomicrograph of two bean-shaped cells with stalks that combine to form one twisted mass. (b) Transmission electron micrograph of a thin section of a Gallionella cell with stalk. Cells are about 0.6 μm wide.
Gallionella is common in the waters draining bogs, iron springs, and other habitats where ferrous iron is present. Mariprofundus was first isolated from Lo‘ihi Seamount, a submarine volcano near Hawaii (Section 21.5 and Figure 21.18). Gallionella and Mariprofundus are both autotrophic chemolithotrophs containing enzymes of the Calvin cycle (Sections 3.12 and 14.2).
Anaerobic Iron-Oxidizing Bacteria
Anaerobic ferrous iron oxidation can be mediated by both chemotrophic and phototrophic bacteria. These groups are common in anoxic sediments and wetlands. Anoxic conditions promote the solubility of ferrous iron across a wide range of pH and so, unlike the aerobic iron-oxidizing bacteria, growth of anaerobic iron-oxidizers is not strictly limited to neutral pH. These groups contain organisms that are metabolically diverse and able to grow by using a variety of different electron donors and acceptors.
Phototrophic iron oxidation occurs in select species of purple nonsulfur bacteria of the Alphaproteobacteria (e.g., Rhodopseudomonas palustris), select species of purple sulfur bacteria of the Gammaproteobacteria (Figure 14.23), and select species of green sulfur bacteria found in the Chlorobi (Chlorobium ferrooxidans). In all cases ferrous iron is one of several compounds that these organisms can use as an electron donor in photosynthesis.
Anaerobic chemotrophic iron-oxidizers couple the oxidation of ferrous iron to nitrate reduction, producing either NO2 − or nitrogen gases (denitrification). These organisms are Alpha-, Beta-, Gamma-, or Deltaproteobacteria, and most are also able to use various organic electron donors in nitrate reduction; many can also grow aerobically. The bacterial genera Acidovorax, Aquabacterium, and Marinobacter all contain anaerobic iron-oxidizers. While most species are mixotrophs when growing with ferrous iron as electron donor, species such as Marinobacter aquaeolei and Thiobacillus denitrificans are able to grow autotrophically as iron-oxidizing chemolithotrophs.
Check Your Understanding
What habitat characteristics govern the diversity and distribution of iron oxidizers?
How do aerobic neutrophilic iron-oxidizers keep their cells from becoming entombed in a crust of iron?
15.15 Methanotrophs and Methylotrophs
Methylotrophs are organisms that grow using organic compounds lacking C—C bonds as electron donors in energy metabolism and as carbon sources. Methylotrophy occurs in the bacterial phyla Proteobacteria, Firmicutes, Actinobacteria, Bacteroidetes, Verrucomicrobia, and in the archaeal phylum Euryarchaeota (Figure 15.1). Methanotrophs are a subset of methylotrophs defined by their ability to use methane as a substrate for growth (Section 14.16).
Aerobic methylotrophs are common in soil and aquatic environments where O2 is present. Anaerobic methylotrophs are common in anoxic environments, particularly in marine sediments. Many anaerobic methylotrophs are methanogenic Archaea. In addition, a consortium of methanogenic Archaea and sulfate-reducing bacteria combine to oxidize methane from gas hydrates found in deep-sea sediments (Section 14.16 and Figure 14.41). We consider here only the aerobic methylotrophs.
Key Genera: *Hyphomicrobium, Methylobacterium*
Aerobic facultative methylotrophs are unable to use methane but can use many other methylated compounds. They are species of Alpha-, Beta-, and Gammaproteobacteria, Actinobacteria, and Firmicutes. Facultative methylotrophs are metabolically diverse and, in addition to methylated substrates, most species can grow aerobically using other organic compounds, such as organic acids, ethanol, and sugars. When growing as methylotrophs, most species can grow aerobically with methanol and some can also metabolize methylated amines, methylated sulfur compounds, and halomethanes. Most are obligate aerobes, though some species are capable of denitrification.
The genus Hyphomicrobium provides an example of the metabolic versatility of the aerobic facultative methylotrophs. Certain species of Hyphomicrobium can grow as aerobic methylotrophs using methanol, methylamine, or dimethyl sulfide. Species of Hyphomicrobium can also grow as anaerobic methylotrophs using methanol as an electron donor coupled to denitrification. Finally, Hyphomicrobium can grow aerobically on a range of C2 and C4 compounds.
Key Genera: *Methylomonas, Methylosinus*
Aerobic methanotrophs are methylotrophs that can use methane as an electron donor and typically can use it as a carbon source as well. Table 15.1 gives a taxonomic overview of the methanotrophs. Most methanotrophs are Proteobacteria and are classified into two major groups based on their internal cell structure, phylogeny, and carbon assimilation pathway. Type I methanotrophs assimilate one-carbon compounds via the ribulose monophosphate cycle and are Gammaproteobacteria. By contrast, type II methanotrophs assimilate C1 intermediates via the serine pathway and are Alphaproteobacteria (Table 15.1). We discussed the biochemical details of these pathways in Section 14.16. Most methanotrophs are metabolically specialized for aerobic growth on methane, though some can grow on either methane or methanol. Methanotrophs are typically obligate methylotrophs; however, the methanotrophic genus Methylocella contains species that can also grow on acetate or organic acids such as pyruvate and succinate.
Table 15.1 Some characteristics of methanotrophic *Bacteria*

aInternal membranes: type I, bundles of disk-shaped vesicles distributed throughout the organism; type II, paired membranes running along the periphery of the cell. See Figure 15.37.
cAcidophiles. For the properties of Verrucomicrobiaceae, see Section 16.17.
In addition to the proteobacterial methanotrophs described above, the phylum Verrucomicrobia contains the bacterium Methylacidiphilum. Genome analysis has shown that species of Methylacidiphilum lack key enzymes of both the ribulose monophosphate and serine pathways. Instead, Methylacidiphilum uses the Calvin cycle to assimilate carbon from CO2.
Physiology
Methanotrophs possess a key enzyme, methane monooxygenase, that catalyzes the incorporation of an atom of oxygen from O2 into CH4, forming methanol (CH3OH, Section 14.16). The requirement for O2 as a reactant in the initial oxygenation of CH4 explains why these methanotrophs are obligate aerobes. Methane monooxygenase is located in extensive internal membrane systems that are the site of methane oxidation. Membranes in type I methanotrophs are arranged as bundles of disk-shaped vesicles distributed throughout the cell (**Figure 15.37*b***). Type II species possess paired membranes running along the periphery of the cell (Figure 15.37a). Verrucomicrobial methanotrophs possess membrane vesicles. Methylotrophs unable to use methane lack these internal membrane arrays.
Figure 15.37 Methanotrophs.

(a) Electron micrograph of a cell of Methylosinus, illustrating a type II membrane system. Cells are about 0.6 μm in diameter. (b) Electron micrograph of a cell of Methylococcus capsulatus, illustrating a type I membrane system. Cells are about 1 μm in diameter. Compare with Figure 15.25.
Methanotrophs are virtually unique among bacteria in possessing relatively large amounts of sterols. Sterols are rigid planar molecules found in the cytoplasmic and other membranes of eukaryotes but are absent from most bacteria. Sterols may be an essential part of the complex internal membrane system for methane oxidation (see Figure 15.37). The only other group of bacteria in which sterols are widely distributed is the mycoplasmas, bacteria that lack cell walls and thus probably require a tougher cytoplasmic membrane (Section 16.9). Many methylotrophs contain various carotenoid pigments and high levels of cytochromes in their membranes, and these features often render colonies of aerobic methylotrophs pink.
Ecology
Aerobic methylotrophs are found in the open ocean, in soils, in association with plant roots and leaf surfaces, and at the oxic interface of many anoxic environments. Methanol is produced during the breakdown of plant pectin and this is likely an important substrate for methylotrophs in terrestrial ecosystems. In addition, soils contain aerobic methanotrophs that consume atmospheric methane and are an important biological sink for atmospheric methane. Aerobic methanotrophs are also common at the oxic interface of anoxic environments found in lakes, sediments, and wetlands where methanogens provide a constant source of methane. These methanotrophs play an important role in the global carbon cycle by oxidizing CH4 and converting it into cell material and CO2 before it reaches the atmosphere (CH4 is a strong greenhouse gas).
Methanotrophs also form a variety of symbioses with eukaryotic organisms. For example, some marine mussels live in the vicinity of hydrocarbon seeps on the seafloor, places where CH4 is released in substantial amounts. Methanotrophic symbionts reside within the animal’s gill tissue (Figure 15.38), which ensures effective gas exchange with seawater. Assimilated CH4 is distributed throughout the animal by the excretion of organic compounds by the methanotrophs. These methanotrophic symbioses are therefore conceptually similar to those that develop between sulfide-oxidizing chemolithotrophs and hydrothermal vent tube worms and giant clams (Section 23.11).
Figure 15.38 Methanotrophic symbionts of marine mussels.

(a) Electron micrograph of a thin section at low magnification of gill tissue from a marine mussel living near hydrocarbon seeps in the Gulf of Mexico. Note the symbiotic methanotrophs (arrows) in the tissues. (b) High-magnification view of gill tissue showing methanotrophs with type I membrane bundles (arrows). Cells of the methanotrophs are about 1 μm in diameter. Compare with Figure 15.37b.
Methylomirabilis oxyfera is a methanotroph isolated from anoxic waters in the Black Sea, and it was the first isolate obtained from the unique bacterial phylum NC-10. M. oxyfera is an obligate anaerobe; however, it uses the O2-dependent enzyme of aerobic methanotrophs (methane monooxygenase) to oxidize methane to CO2. M. oxyfera accomplishes this by reducing nitrite to nitric oxide (NO), which is then dismutated to N2 and O2 (2 NO→N2+O2). The O2 produced by this pathway is then consumed by methane monooxygenase during the oxidation of CH4 (Section 14.16). Like the methanotroph Methylacidiphilum, M. oxyfera assimilates C1 units as CO2, probably by the Calvin cycle.
Mastering Microbiology
Art Activity: Figure 15.40 Developmental cycle of the bacterial predator Bdellovibrio bacteriovorus
Check Your Understanding
What is the difference between a methanotroph and a methylotroph?
What is unique about the methanotroph Methylomirabilis?
IV: Morphologically and Ecologically Distinctive Bacteria
IV: Morphologically and Ecologically Distinctive Bacteria
IV Morphologically and Ecologically Distinctive *Bacteria*
**Some Bacteria are so morphologically distinct that they can be instantly recognized, even in natural samples. Other Bacteria display some unique behavioral property—such as predation or motility controlled by magnetic particles—that sets them apart from all other microbes in an ecological sense.**
Many microbes have distinctive lifestyles or morphological characteristics that differentiate them from other organisms. These traits are often associated with specific ecological properties. For example, some chemoorganotrophs obtain their carbon and energy by consuming other organisms. We begin this section by discussing microbial predators.
Key Genera: *Bdellovibrio, Myxococcus*
Some bacteria are predators that consume other bacteria. Known bacterial predators reside among several classes of Proteobacteria and in the Bacteroidetes, Chloroflexi, and Melainabacteria. Several different methods of predation have been observed. Some predators, such as Vampirococcus (phylogeny unknown), Micavibrio (Alphaproteobacteria), and the green algal predator Vampirovibrio (Melainabacteria) are epibiotic predators; they attach to the surface of their prey and acquire nutrients from its cytoplasm or periplasm. Other predators, such as Daptobacter (Epsilonproteobacteria), are cytoplasmic predators, as they invade their host cells and replicate in the cytoplasm, consuming their prey from the inside out. Bdellovibrio have a similar lifestyle as periplasmic predators; they invade and replicate within the periplasmic space of their prey cells. Finally, predators such as Lysobacter (Gammaproteobacteria) and Myxococcus (Deltaproteobacteria) are social predators. These gliding bacteria use swarming behavior to find prey, which they lyse and feed upon collectively. Bdellovibrio and Myxococcus are the most thoroughly described genera of bacterial predators.
Bdellovibrio
Bdellovibrio are small, highly motile and curved bacteria that prey on other bacteria, using the cytoplasmic constituents of their hosts as nutrients (bdello is a prefix meaning “leech”). After attachment of a Bdellovibrio cell to its prey, the predator penetrates the cell wall of the prey and replicates in the periplasmic space, eventually forming a spherical structure called a bdelloplast. Two stages of penetration are shown in electron micrographs in Figure 15.39 and diagrammatically in **Figure 15.40*b***. A wide variety of gram-negative prey bacteria can be attacked by Bdellovibrio, but gram-positive cells are not attacked.
Figure 15.39 Attack on a prey cell by *Bdellovibrio.*

Thin-section electron micrographs of Bdellovibrio attacking a cell of Delftia acidovorans. (a) Entry of the predator cell. (b) Bdellovibrio cell inside the host. The Bdellovibrio cell is enclosed in the bdelloplast and replicates in the periplasmic space. A Bdellovibrio cell measures about 0.3 μm in diameter.
Figure 15.40 Developmental cycle of the bacterial predator *Bdellovibrio bacteriovorus.*

(a) Electron micrograph of a cell of Bdellovibrio bacteriovorus; note the thick flagellum. A cell is 0.3 μm wide. (b) Events in predation. Following primary contact with a gram-negative bacterium, the highly motile Bdellovibrio cell attaches to and penetrates into the prey periplasmic space. Once inside the periplasmic space, Bdellovibrio cells elongate, and within 4 h progeny cells are released. The number of progeny cells released varies with the size of the prey; 5–6 bdellovibrios are released from Escherichia coli and 20–30 for a larger prey cell, such as Aquaspirillum.
Bdellovibrio is an obligate aerobe, obtaining its energy from the oxidation of amino acids and acetate. In addition, Bdellovibrio assimilates nucleotides, fatty acids, peptides, and even some intact proteins directly from its host without first hydrolyzing them. Prey-independent derivatives of Bdellovibrio can be isolated and grown on complex media, however, showing that predation is not an obligatory lifestyle.
Phylogenetically, bdellovibrios are Deltaproteobacteria, and they are widespread in aquatic habitats. Procedures for their isolation are similar to those used to isolate bacterial viruses (Section 5.3). Prey bacteria are spread on the surface of an agar plate, forming a lawn, and the surface is inoculated with a small amount of soil suspension that has been filtered through a membrane filter; the filter retains most bacteria but allows the small Bdellovibrio cells to pass. On incubation of the agar plate, plaques analogous to bacteriophage plaques (Figure 5.10b) are formed at locations where Bdellovibrio cells are multiplying. Pure cultures of Bdellovibrio can then be isolated from these plaques. Bdellovibrio are widely distributed, as cultures have been obtained from many soils and from sewage.
Myxobacteria
Myxobacteria exhibit the most complex behavioral patterns of all known bacteria. The life cycle of myxobacteria results in the formation of multicellular structures called fruiting bodies. The fruiting bodies are often strikingly colored and morphologically elaborate (Figure 15.41), and these can often be seen with a hand lens on moist pieces of decaying wood or plant material. The fruiting myxobacteria are classified on morphological grounds using characteristics of the vegetative cells, the myxospores, and fruiting body structure.
Figure 15.41 Fruiting bodies of three species of fruiting myxobacteria.

(a) Myxococcus fulvus (125 μm high). (b) Myxococcus stipitatus (170 μm high). (c) Chondromyces crocatus (560 μm high).
The life cycle of a typical myxobacterium is shown in Figure 15.42. The vegetative cells of the myxobacteria are simple, nonflagellated, gram-negative rods (Figure 15.42) that glide across surfaces and obtain their nutrients primarily by using extracellular enzymes to lyse other bacteria and use the released nutrients. A vegetative cell excretes slime, and as it moves across a solid surface, it leaves behind a slime trail (Figure 15.42). The vegetative cells form a swarm that exhibits self-organizing behavior, and this allows them to behave as a single coordinated entity in response to environmental cues.
Figure 15.42 Life cycle of *Myxococcus.*

Dormant myxospores germinate under favorable conditions to yield vegetative cells. Vegetative cells exhibit adventurous motility characterized by gliding (lower inset, Myxococcus fulvus cells, 0.8 μm diameter). When nutrients are limiting, cells aggregate into swarming colonies (upper inset, Myxococcus xanthus colony on agar, 10 mm diameter) exhibiting social motility mediated by type IV pili and twitching. Cells eventually aggregate to form fruiting bodies, within which some vegetative cells develop into myxospores.
Mastering Microbiology
Art Activity: Figure 15.42 Life cycle of Myxococcus xanthus
Upon nutrient exhaustion, vegetative cells of myxobacteria begin to migrate toward each other, aggregating together in mounds or heaps (Figure 15.43). Aggregation is likely mediated by chemotactic or quorum-sensing responses (Sections 7.6 and 7.7). As the cell masses become higher, they begin to differentiate into fruiting bodies (Figure 15.44) containing myxospores. Myxospores are specialized cells that are resistant to drying, ultraviolet radiation, and heat, but the degree of heat resistance is much less than that of the bacterial endospore (Section 2.8). Fruiting bodies can be simple, consisting of masses of myxospores embedded in slime, or complex, consisting of a stalk and heads (Figure 15.44). The fruiting body stalk is composed of slime within which a few cells are trapped. The majority of the cells migrate to the fruiting body head, where they undergo differentiation into myxospores (Figure 15.42).
Figure 15.43 Scanning electron micrographs of fruiting body formation in *Chondromyces crocatus.*

(a) Early stage, showing aggregation and mound formation. (b) Initial stage of stalk formation. Slime formation in the head has not yet begun and so the cells that compose the head are still visible. (c) Three stages in head formation. Note that the diameter of the stalk also increases. (d) Mature fruiting bodies. The entire fruiting structure is about 600 μm in height (compare with Figure 15.41c).
Figure 15.44 The myxobacterium *Stigmatella aurantiaca.*

Scanning electron micrograph of a fruiting body growing on a piece of wood. Note the individual cells visible in each fruiting body. Inset: Phase-contrast photomicrograph of a single fruiting body about 150 μm high. The color is due to the production of glucosylated carotenoid pigments.
Check Your Understanding
What environmental conditions trigger fruiting body formation in myxobacteria?
What are the different ways in which species of Myxococcus and Bdellovibrio kill their prey?
Key Genera: *Spirochaeta, Treponema, Cristispira, Leptospira, Borrelia*
Spirochetes are morphologically unique bacteria found only within the bacterial phylum Spirochaetes. Spirochetes are gram-negative, motile, tightly coiled Bacteria, typically slender and flexuous in shape (Figure 15.45). Spirochetes are widespread in aquatic sediments and in animals. Some cause diseases, including syphilis, an important human sexually transmitted disease. Spirochetes are classified into eight genera (Table 15.2) primarily on the basis of habitat, pathogenicity, phylogeny, and morphological and physiological characteristics.
Figure 15.45 Morphology of spirochetes.

Two spirochetes at the same magnification, showing the wide size range in the group. (a) Spirochaeta stenostrepta, by phase-contrast microscopy. A single cell is 0.25 μm in diameter. (b) Spirochaeta plicatilis. A single cell is 0.75 μm in diameter and can be up to 250 μm (0.25 mm) in length.
Table 15.2 Genera of spirochetes and their characteristics

Spirochetes have an unusual mode of motility conveyed by their unusual morphology. Spirochetes contain endoflagella, which resemble normal flagella but are found in the cell periplasm (Figure 15.46). The endoflagella are anchored at the cell poles and extend back along the length of the cell. Both the endoflagella and the protoplasmic cylinder are surrounded by a flexible membrane called the outer sheath (Figure 15.46b). Endoflagella rotate, as do typical bacterial flagella. However, when both endoflagella rotate in the same direction, the protoplasmic cylinder rotates in the opposite direction, placing torsion on the cell (Figure 15.46b). This torsion causes the spirochete cell to flex, resulting in a corkscrew-like motion that allows cells to burrow through viscous materials or tissues.
Figure 15.46 Motility in spirochetes.

(a) Electron micrograph of a negatively stained cell of Spirochaeta zuelzerae, showing the position of the endoflagellum; the cell is about 0.3 μm in diameter. (b) Diagram of a spirochete cell, showing the arrangement of the protoplasmic cylinder, endoflagella, and external sheath, and how rotation of the endoflagellum generates rotation of both the protoplasmic cylinder and the external sheath.
Spirochetes are often confused with spirilla. Spirilla are helically curved rod-shaped cells, usually motile by means of polar flagella (Figure 15.47). The word spirillum refers to a general cell shape that is widespread among Bacteria and Archaea. The number of helical turns in a single spirillum may vary from less than one complete turn (in which case the organism looks like a vibrio) to many turns. In addition, spirilla that divide terminally, such as the cyanobacterium Spirulina (Figure 15.5), can form long helical filaments that superficially resemble spirochetes. Spirilla, however, lack the outer sheath, endoflagella, and corkscrew-like motility of spirochetes. In addition, spirilla are typically fairly rigid cells while spirochetes are highly flexible and quite thin (< 0.5 μm).
Figure 15.47 Spirilla.

(a) Spirillum volutans, visualized by dark-field microscopy, showing flagellar bundles and volutin (polyphosphate) granules. Cells are about 1.5×25 μm. (b) Scanning electron micrograph of an intestinal spirillum. Note the polar flagellar tufts and the spiral structure of the cell surface. (c) Scanning electron micrograph of cells of Ancylobacter aquaticus. Cells are about 0.5 μm in diameter.
*Spirochaeta* and *Cristispira*
The genus Spirochaeta includes free-living, anaerobic, and facultatively aerobic spirochetes. These organisms, of which several species are known, are common in aquatic environments such as freshwater and sediments, and also in the oceans. Spirochaeta plicatilis (Figure 15.45b) is a large spirochete found in sulfidic freshwater and marine habitats. The 20 or so endoflagella inserted at each pole of S. plicatilis are arranged in a bundle that winds around the coiled protoplasmic cylinder. Another species, Spirochaeta stenostrepta (Figure 15.45a), is an obligate anaerobe commonly found in H2S-rich black muds. It ferments sugars to ethanol, acetate, lactate, CO2, and H2.
Cristispira (Figure 15.48) is a unique spirochete found in nature only in the crystalline style of certain molluscs, such as clams and oysters. The crystalline style is a flexible, semisolid rod seated in a sac and rotated against a hard surface of the digestive tract, thereby mixing and grinding the small particles of food taken in by the animal. Cristispira lives in both freshwater and marine molluscs, but not all species of molluscs possess this organism. Unfortunately, Cristispira has not been cultured, and so the physiological rationale for its restriction to this unique habitat is unknown.
Figure 15.48 *Cristispira.*

Electron micrograph of a thin section of a cell of Cristispira. This large spirochete is about 2 μm in diameter. Notice the numerous endoflagella.
*Treponema* and *Borrelia*
Anaerobic or microaerophilic host-associated spirochetes that are commensals or pathogens of humans and animals reside in the genus Treponema. T. pallidum, the causal agent of syphilis (Section 31.13), is the best-known species of Treponema. It differs in morphology from other spirochetes in that the Treponema cell is not helical but flat and wavy. The T. pallidum cell is remarkably thin, measuring only 0.2 μm in diameter. Because of this, dark-field microscopy has long been used to examine exudates from suspected syphilitic lesions (Figure 31.13).
Other species of Treponema are also often found as commensals in humans and other animals. For example, Treponema denticola is common in the human oral cavity and is associated with gum disease. It ferments amino acids such as cysteine and serine, forming acetate as the major fermentation acid, as well as CO2, NH3, and H2S. Spirochetes are also common in the rumen, the digestive forestomach of ruminant animals (Section 23.15). For instance, Treponema saccharophilum (**Figure 15.49*a***) is a large, pectinolytic spirochete found in the bovine rumen where it ferments pectin, starch, inulin, and other plant polysaccharides. Treponema primitia can be found in the hindgut of certain termites. In the termite gut, fermentation of cellulose causes production of H2 and CO2. T. primitia is an acetogen (Section 14.14) that grows on H2 plus CO2, forming acetate, which is an important component of the insect’s nutrition. Treponema azotonutricium is also found in the termite hindgut and is capable of nitrogen fixation (Section 3.12).
Figure 15.49 *Treponema* and *Borrelia*.

(a) Phase-contrast micrographs of Treponema saccharophilum, a large pectinolytic spirochete from the bovine rumen. A cell measures about 0.4 μm in diameter. Left, regularly coiled cells; right, irregularly coiled cells. (b) Scanning electron micrograph of a cell of Borrelia burgdorferi, the causative agent of Lyme disease.
The majority of species of Borrelia are animal or human pathogens. Borrelia burgdorferi (Figure 15.49b) is the causative agent of the tickborne Lyme disease, which infects humans and animals (Section 32.4). B. burgdorferi is also of interest because it is one of the few bacteria that has a linear (as opposed to a circular) chromosome (Sections 6.2 and 10.3). Other species of Borrelia are primarily of veterinary importance, causing diseases in cattle, sheep, horses, and birds. In most cases, the bacterium is transmitted to the animal host from the bite of a tick.
*Leptospira* and *Leptonema*
The genera Leptospira and Leptonema contain strictly aerobic spirochetes that oxidize long-chain fatty acids (for example, the C18 fatty acid oleic acid) as electron donors and carbon sources. With few exceptions, these are the only substrates utilized for growth. Leptospiras are thin, finely coiled, and usually bent at each end into a semicircular hook. At present, several species are recognized in this group, some free-living and many parasitic. Two major species of Leptospira are L. interrogans (parasitic) and L. biflexa (free-living). Strains of L. interrogans are parasitic for humans and animals. Rodents are the natural hosts of most leptospiras, although dogs and pigs are also important carriers of certain strains.
In humans the most common leptospiral syndrome is leptospirosis, a disorder in which the organism localizes in the kidneys and can cause renal failure or even death. Leptospiras ordinarily enter the body through the mucous membranes or through breaks in the skin during contact with an infected animal. After a transient multiplication in various parts of the body, the organism localizes in the kidneys and liver, causing nephritis and jaundice. Domestic animals such as dogs are vaccinated against leptospirosis with a killed virulent strain in the combined distemper–leptospira–hepatitis vaccine.
Check Your Understanding
What are the major differences between spirochetes and spirilla?
Name two diseases of humans caused by spirochetes.
Key Genera: *Hyphomicrobium, Caulobacter*
The growth of most bacteria is coupled to cell division by the well-known process of binary fission (Section 4.6 and Figure 4.8). In this section, we consider organisms that grow and divide in different ways, including budding and the formation of appendages. Budding and appendaged species often have life cycles that are distinct among bacteria.
Budding Division
As we learned in Section 4.10, budding bacteria divide as a result of unequal cell growth. Cell division in stalked and budding bacteria forms a totally new daughter cell, with the mother cell retaining its original identity (Figure 4.18). In contrast, binary fission produces two equivalent cells.
A fundamental difference between budding bacteria and bacteria that divide by binary fission is the formation of new cell wall material from a single point (polar growth) rather than throughout the whole cell (intercalary growth) as in binary fission (Chapter 8). Several genera not normally considered to be budding bacteria show polar growth without differentiation of cell size (Figure 4.18). An important consequence of polar growth is that internal structures, such as membrane complexes, are not partitioned in the cell division process and must be formed de novo. However, this has an advantage in that more complex internal structures can be formed in budding cells than in cells that divide by binary fission, since the latter cells would have to partition these structures between the two daughter cells. Not coincidentally, many budding bacteria, particularly phototrophic and chemolithotrophic species, contain extensive internal membrane systems.
Budding Bacteria: *Hyphomicrobium*
Two well-studied budding bacteria are closely related Alphaproteobacteria: Hyphomicrobium (Figure 15.50), which is chemoorganotrophic, and Rhodomicrobium, which is phototrophic. These organisms release buds from the ends of long, thin hyphae. The hypha is a direct cellular extension and contains cell wall, cytoplasmic membrane, and ribosomes, and can contain DNA.
Figure 15.50 Stages in the *Hyphomicrobium* cell cycle.

The single chromosome of Hyphomicrobium is circular.
Mastering Microbiology
Art Activity: Figure 15.53 Stages in the Hyphomicrobium cell cycle
Figure 15.50 shows the life cycle of Hyphomicrobium. The mother cell, which is often attached by its base to a solid substrate, forms a thin outgrowth that lengthens to become a hypha. At the end of the hypha, a bud forms. This bud enlarges, forms a flagellum, breaks loose from the mother cell, and swims away. Later, the daughter cell loses its flagellum and after a period of maturation forms a hypha and buds. More buds can also form at the hyphal tip of the mother cell, leading to arrays of cells connected by hyphae. In some cases, a bud begins to form directly from the mother cell without the intervening formation of a hypha, whereas in other cases a single cell forms hyphae from each end (Figure 15.51). Nucleoid replication events occur before the bud emerges, and then once a bud has formed, a copy of the chromosome moves down the hypha and into the bud. A cross-septum then forms, separating the still-developing bud from the hypha and mother cell (Figure 15.51).
Figure 15.51 Morphology of *Hyphomicrobium*.

(a) Phase-contrast micrograph of cells of Hyphomicrobium. Cells are about 0.7 μm wide. (b) Electron micrograph of a thin section of a single Hyphomicrobium cell. The hypha is about 0.2 μm wide.
Mastering Microbiology
Art Activity: Figure 15.56a Prosthecate bacteria
Physiologically, Hyphomicrobium is a methylotrophic bacterium (Section 14.16 and Section 15.15), and it is widespread in freshwater, marine, and terrestrial habitats. Preferred carbon sources are methanol (CH3OH), methylamine (CH3NH2), formaldehyde (CH2O), and formate (HCOO−). A fairly specific enrichment procedure for Hyphomicrobium is to use CH3OH as an electron donor with nitrate (NO3 −) as an electron acceptor in a dilute medium incubated under anoxic conditions. The only rapidly growing denitrifying bacterium known that uses CH3OH as an electron donor is Hyphomicrobium, and so this procedure can select this organism out of a wide variety of environments.
Prosthecate and Stalked *Bacteria*
A variety of bacteria are able to produce cytoplasmic extrusions including stalks (Figure 15.52), hyphae, and appendages (Table 15.3). Extrusions of these kinds, which are smaller in diameter than the mature cell and contain cytoplasm and a cell wall, are collectively called prosthecae (Figure 15.53). Prosthecae allow organisms to attach to particulate matter, plant material, or other microorganisms in aquatic habitats. In addition, prosthecae can be used to increase the ratio of surface area to cell volume. Recall that the high surface-to-volume ratio of prokaryotic cells in general confers an increased ability to take up nutrients and expel wastes (Section 1.3). The unusual morphology of appendaged bacteria (Figure 15.53) carries this theme to an extreme and may be an evolutionary adaptation to life in oligotrophic (nutrient-poor) waters where these organisms are most commonly found.
Figure 15.52 Stalked bacteria.

(a) A Caulobacter rosette. A single cell is about 0.5 μm wide. The five cells are attached by their stalks, which are also prosthecae. Two of the cells have divided, and the daughter cells have formed flagella. (b) Negatively stained preparation of a Caulobacter cell in division. (c) A thin section of Caulobacter showing that cytoplasm is present in the stalk. Parts b and c are transmission electron micrographs.
Table 15.3 Characteristics of major genera of stalked, appendaged (prosthecate), and budding *Bacteria*

Figure 15.53 Prosthecate bacteria.

(a) Electron micrograph of a shadow-cast preparation of Asticcacaulis biprosthecum, illustrating the location and arrangement of the prosthecae, the holdfast, and a swarmer cell. The swarmer cell breaks away from the mother cell and begins a new cell cycle. Cells are about 0.6 μm wide. (b) Negatively stained electron micrograph of a cell of Ancalomicrobium adetum. The prosthecae are bounded by the cell wall, contain cytoplasm, and are about 0.2 μm in diameter. (c) Electron micrograph of the star-shaped prosthecate bacterium Stella. Cells are about 0.8 μm in diameter.
Prosthecae may also function to reduce cell sinking. Because these organisms are aquatic and their metabolism is typically aerobic, prosthecae may keep cells from sinking into anoxic zones in their aquatic environments where they would be unable to respire. Some prosthecate bacteria produce gas vesicles (Section 2.7), which would also help prevent sinking.
Caulobacter
Two common stalked bacteria are Caulobacter (Figure 15.52) and Gallionella (Figure 15.36). The former is a chemoorganotroph that produces a cytoplasm-filled stalk, that is, a prostheca, while the latter is a chemolithotrophic iron-oxidizing bacterium whose stalk is composed of ferric hydroxide [Fe(OH)3] (Section 15.14). Caulobacter cells are often seen on surfaces in aquatic environments with the stalks of several cells attached to form rosettes (Figure 15.52a). At the end of the stalk is a structure called a holdfast by which the stalk anchors the cell to a surface.
The Caulobacter cell division cycle (Figure 15.54; Section 8.8 and Figure 8.20) is unique because cells undergo unequal binary fission. A stalked cell of Caulobacter divides by elongation of the cell followed by binary fission, and a single flagellum forms at the pole opposite the stalk. The flagellated cell so formed, called a swarmer, separates from the nonflagellated mother cell and eventually attaches to a new surface, forming a new stalk at the flagellated pole; the flagellum is then lost. Stalk formation is a necessary precursor of cell division and is coordinated with DNA synthesis (Figure 15.54). The cell division cycle in Caulobacter is thus more complex than simple binary fission or budding division because the stalked and swarmer cells are structurally different and the growth cycle must include both forms.
Figure 15.54 Growth of *Caulobacter*.

Stages in the Caulobacter cell cycle, beginning with a swarmer cell. Compare with Figure 8.20.
Mastering Microbiology
Art Activity: Figure 15.57 Growth of Caulobacter
Check Your Understanding
How does budding division differ from binary fission? How does binary fission differ from the division process in Caulobacter?
What advantage might a prosthecate organism have in a very nutrient-poor environment?
Key Genera: *Sphaerotilus, Leptothrix*
Bacteria in many phyla form sheaths made of polysaccharide or protein that encase one or many cells. Sheaths often function to bind cells together into long multicellular filaments (Sections 15.3 and 15.12). Sphaerotilus and Leptothrix are filamentous bacteria that grow within a sheath and have a unique life cycle. Under favorable conditions, the cells grow vegetatively, leading to the formation of long, cell-packed sheaths. Flagellated swarmer cells form within the sheath under unfavorable growth conditions, and the swarmer cells break out and are dispersed to new environments, leaving behind the empty sheath.
Sphaerotilus and Leptothrix are common in freshwater habitats that are rich in organic matter, such as wastewaters and polluted streams. Because they are typically found in flowing waters, they are also abundant in trickling filters and activated sludge digesters in sewage treatment plants (Section 22.6). In habitats in which reduced iron (Fe2+) or manganese (Mn2+) is present, the sheaths may become coated with ferric hydroxide [Fe(OH)3] or manganese oxides from the oxidation of these metals.
Leptothrix
The ability of Sphaerotilus and Leptothrix to precipitate iron oxides on their sheaths is well established, and when sheaths become iron encrusted, as occurs in iron-rich waters, they can frequently be seen microscopically (Figure 15.55). Iron precipitates form when ferrous iron (Fe2+), chelated to organic materials such as humic or tannic acids, is oxidized. These chemoorganotrophic bacteria use the organic materials as a carbon or energy source and, when no longer chelated, the ferrous iron becomes oxidized and precipitates on the sheath. Iron oxidation is fortuitous and though these organisms are closely related to dissimilative iron-oxidizers (Section 15.14), the organism does not gain energy from iron oxidation. In a similar way, Leptothrix can also oxidize manganese.
Figure 15.55 *Leptothrix* and iron precipitation.

Transmission electron micrograph of a thin section of Leptothrix growing in a ferromanganese film in a swamp in Ithaca, New York. A single cell measures about 0.9 μm in diameter. Note the protuberances of the cell envelope that contact the sheath (arrows).
Sphaerotilus
The Sphaerotilus filament is composed of a chain of rod-shaped cells enclosed in a closely fitting sheath. This thin, transparent structure is difficult to see when it is filled with cells, but when it is partially empty, the sheath can more easily be resolved (**Figure 15.56*a***). Individual cells are 1–2×3–8 μm in dimensions and stain gram-negatively. The cells within the sheath (Figure 15.56b) divide by binary fission, and the new cells synthesize new sheath material at the tips of the filaments. Eventually, motile swarmer cells are liberated from the sheaths (Figure 15.56c) and then migrate, attach to a solid surface, and begin to grow, with each swarmer being the forerunner of a new filament. The sheath, which is devoid of peptidoglycan, consists of protein and polysaccharide.
Figure 15.56 *Sphaerotilus natans*.

A single cell is about 2 μm wide. (a) Phase-contrast photomicrographs of material collected from a polluted stream. Active growth stage (above) and swarmer cells leaving the sheath. (b) Electron micrograph of a thin section through a filament, clearly showing the sheath. (c) Electron micrograph of a negatively stained swarmer cell. Notice the polar flagellar tuft.
Sphaerotilus species are nutritionally versatile and use simple organic compounds as carbon and energy sources; one species can grow mixotrophically with thiosulfate as electron donor. Befitting its habitat in flowing waters, Sphaerotilus is an obligate aerobe. Large masses (blooms) of Sphaerotilus often occur in the fall of the year in streams and brooks when leaf litter causes a temporary increase in the organic content of the water. In addition, its filaments are the main component of a microbial complex that wastewater engineers call “sewage fungus,” a filamentous slime found on the rocks in streams receiving sewage pollution. In activated sludge of sewage treatment plants (Section 22.6), Sphaerotilus is often responsible for a condition called bulking, where the tangled masses of Sphaerotilus filaments so increase the bulk of the sludge that it remains suspended and does not settle as it should. This has a negative effect on the oxidation of organic matter and the recycling of inorganic nutrients and leads to treatment plant discharges with high nitrogen and carbon loads.
Check Your Understanding
Describe how a sheathed bacterium such as Sphaerotilus grows.
List two metals that are oxidized by sheathed bacteria.
Key Genera: *Magnetospirillum*
In a magnetic field, magnetic bacteria demonstrate a dramatic directed movement called magnetotaxis. Within these cells are structures called magnetosomes, which consist of chains of magnetic particles made of magnetite (Fe3O4) or greigite (Fe3S4) (Section 2.7 and Figure 2.24). Magnetosomes are localized within invaginations of the cell membrane that are organized in a linear conformation by a protein scaffold. Magnetic bacteria orient along the north–south magnetic moment of a magnetic field, aligning parallel to the field lines in much the same manner as a compass needle. Magnetic bacteria are typically microaerophilic or anaerobic and are most often found near the oxic–anoxic interface in sediments or stratified lakes. The magnetosomes of aerobic species typically contain the mineral magnetite while those of anaerobes contain exclusively greigite.
Although the ecological role of bacterial magnets is unclear, the ability to orient in a magnetic field may be of selective advantage in maintaining these organisms in zones of low O2 concentration. Generally, the concentration of O2 decreases with depth through sediments or the water column of stratified lakes. Since Earth is spherical, its magnetic field lines have a strong vertical component in the Northern and Southern Hemispheres. Thus, bacteria that orient along these field lines can preferentially swim down and away from O2. The magnetosome functions like a compass needle to “point” the bacterium in the right direction; rotation of the flagellum, by contrast, is controlled by a chemotactic response to O2 (Section 2.11).
Magnetic bacteria display one of two magnetic polarities depending on the orientation of magnetosomes within the cell. Cells in the Northern Hemisphere have the north-seeking pole of their magnetosomes forward with respect to their flagella and thus move in a northward direction (which in the Northern Hemisphere is downward). Cells in the Southern Hemisphere have the opposite polarity and move southward.
Most of the magnetic bacteria that have been described are species of Alphaproteobacteria, but species have also been observed in the Gammaproteobacteria, the Deltaproteobacteria, and the Nitrospira group. One of the best-characterized species is Magnetospirillum magnetotacticum (Figure 15.57), which is a chemoorganotrophic microaerophile that can also grow anaerobically by reducing NO3 − or N2O. In contrast, the species Desulfovibrio magneticus is a sulfate reducer and an obligate anaerobe. In addition, magnetosomes have been observed in a few species of sulfur oxidizers and purple nonsulfur bacteria. Multicellular magnetotactic bacteria are also known. These are Deltaproteobacteria that form multicellular aggregates of 10–20 cells organized as a hollow sphere. While multicellular magnetotactic bacteria are obligate anaerobes, the basis of their metabolism has not yet been determined.
Figure 15.57 A magnetotactic spirillum.
Electron micrograph of a single cell of Magnetospirillum magnetotacticum; a cell measures 0.3× 2μm. The cell contains particles of magnetosomes made of Fe3O4 arranged in a chain.
We transition now from viewing Bacteria through the lens of their ecological diversity to consider some other important phyla in a phylogenetic context in Chapter 16. We then conclude our coverage of prokaryotic microbes with Chapter 17 dedicated to the Archaea.
Check Your Understanding
What benefit do magnetic bacteria accrue from having magnetosomes?
Would you expect to find greigite or magnetite in the magnetosomes of Desulfovibrio magneticus?
Chapter Review
Go to Mastering Microbiology for videos, animations, practice tests, and more.
I Ecological Diversity Among Microorganisms
15.1 Microbial diversity can be defined in several different ways. Metabolic diversity is defined in terms of the cellular processes that support growth. Ecological diversity is defined in terms of microbial interactions between organisms and their environments. Phylogenetic diversity is defined by evolutionary relationships between organisms.
Q What is convergent evolution and how is it different from horizontal gene transfer?
II Ecological Diversity of Phototrophic *Bacteria*
15.2 Anoxygenic phototrophs, which do not produce oxygen, were the first phototrophic organisms to evolve. The evolution of photosynthesis has been affected strongly by patterns of horizontal gene transfer.
15.3 Cyanobacteria is the only bacterial phylum that contains oxygenic phototrophs. All species of cyanobacteria can fix CO2 and many can fix N2, making these organisms important primary producers in many ecosystems.
**Q How are prochlorophytes, such as Prochlorococcus, different from other cyanobacteria?**
15.4 Purple sulfur bacteria are anoxygenic phototrophic Gammaproteobacteria. Purple sulfur bacteria use H2S and S0 as electron donors and fix CO2 by the Calvin cycle. These phototrophs have bacteriochlorophylls a or b and use a Q-type reaction center.
Q Compare and contrast the metabolism, morphology, and phylogeny of purple sulfur and purple nonsulfur bacteria.
15.5 Purple nonsulfur bacteria are anoxygenic phototrophic Alpha- and Betaproteobacteria. Purple nonsulfur bacteria are metabolically diverse, growing best as photoheterotrophs, and can also grow in darkness. These phototrophs have bacteriochlorophylls a or b and use type II photosystems with Q-type reaction centers. Aerobic anoxygenic phototrophs have type II photosystems but only possess bacteriochlorophyll a.
Q Compare and contrast the metabolism of purple nonsulfur bacteria and aerobic anoxygenic phototrophs.
15.6 Green sulfur bacteria are anoxygenic phototrophs of the phylum Chlorobi. Green sulfur bacteria use H2S or S0 as electron donors and fix CO2 by the reverse citric acid cycle. These phototrophs contain bacteriochlorophylls c, d, or e localized in chlorosomes and bacteriochlorophyll a localized in their FeS-type photosynthetic reaction centers.
Q Compare and contrast the metabolism, morphology, and phylogeny of green sulfur and purple sulfur bacteria.
15.7 Green nonsulfur bacteria, also known as filamentous anoxygenic phototrophs, are anoxygenic phototrophs of the phylum Chloroflexi and grow best as photoheterotrophs. These phototrophs contain bacteriochlorophyll c in chlorosomes and bacteriochlorophyll a in their Q-type photosynthetic reaction centers.
Q What traits do green nonsulfur bacteria share with green sulfur bacteria and purple sulfur bacteria?
15.8 Heliobacteria are anoxygenic phototrophic Firmicutes that grow as photoheterotrophs or in darkness as chemotrophs. Heliobacteria produce bacteriochlorophyll g and have FeS-type reaction centers. Chloracidobacterium thermophilum is an anoxygenic phototrophic acidobacterium that grows photoheterotrophically, possesses bacteriochlorophyll a and c as well as chlorosomes, and has an FeS-type reaction center.
**Q In what ways is Chloracidobacterium thermophilum similar to green sulfur bacteria, and in what ways is it different?**
III Diversity of *Bacteria* Defined by Metabolic Traits
15.9 Diazotrophs are bacteria that assimilate N2 through activity of the enzyme nitrogenase. Diazotrophs are metabolically and phylogenetically diverse and employ various adaptations to protect nitrogenase from oxygen inactivation.
Q What are some ways that diazotrophs protect nitrogenase from O2?
15.10 Nitrifying bacteria are aerobic chemolithotrophs that oxidize NH3 to NO2 − (genus prefix Nitroso-) or NO2 − to NO3 − (genus prefix Nitro-). Ammonia oxidizers are Proteobacteria or Thaumarchaeota, while nitrite oxidizers are Proteobacteria or Nitrospirae. Denitrifiers are metabolically and phylogenetically diverse facultative aerobes and chemoorganotrophs that reduce NO3 − to the gaseous products NO, N2O, and N2.
Q Compare and contrast the nitrogen metabolism of nitrifiers with that of denitrifiers.
15.11 Dissimilative sulfate-reducers are obligate anaerobes that grow by reducing SO4 2− with H2 or simple organic compounds as electron donors. Most sulfate reducers are Deltaproteobacteria. Dissimilative sulfur-reducers are metabolically and phylogenetically diverse organisms that grow by reducing S0 and other oxidized sulfur compounds (other than SO4 2−) as electron acceptors.
Q In what ways are sulfur-reducing bacteria different from sulfate-reducing bacteria, and in what ways are they similar? With respect to sulfate-reducing bacteria, what is the difference between complete and incomplete oxidizers?
15.12 Sulfur chemolithotrophs, most of which are species of Proteobacteria, oxidize H2S and other reduced sulfur compounds as electron donors with O2 or NO3 − as electron acceptors and use either CO2 or organic compounds as carbon sources. Sulfur chemolithotrophs use a variety of ecological strategies to conserve energy from H2S and O2, substances that otherwise react together spontaneously.
Q What are some ecological strategies that aerobic sulfide-oxidizers use to compete with the chemical oxidation of H2S by atmospheric O2?
15.13 Dissimilative iron-reducers reduce insoluble electron acceptors in anaerobic respirations. Most species can grow anaerobically by reducing ferric iron using H2 or simple organic compounds as electron donor. The best-characterized genera include Geobacter, which contains exclusively obligate anaerobes, and Shewanella, which contains facultative aerobes.
**Q In what ways are the dissimilative iron-reducing bacteria Shewanella and Geobacter similar, and in what ways are they different?**
15.14 Dissimilative iron-oxidizers conserve energy from the aerobic oxidation of ferrous iron. These microbes use several ecological strategies to cope with the chemical instability of ferrous iron in oxic habitats at neutral pH. Iron oxidizers are found in four physiological groups: aerobic acidophiles, aerobic neutrophiles, anaerobic chemotrophs, and anaerobic phototrophs.
Q Which group of dissimilative iron-oxidizers is the least diverse, and in what way is this related to oxygen and pH?
15.15 Methylotrophs grow on organic compounds that lack carbon–carbon bonds. Some methylotrophs are also methanotrophs, organisms able to catabolize methane. Most methanotrophs are Proteobacteria that contain extensive internal membranes and incorporate carbon by either the serine or ribulose monophosphate pathways.
Q What are the differences between type I and type II methanotrophs?
IV Morphologically and Ecologically Distinctive *Bacteria*
15.16 Bacterial predators such as Bdellovibrio and Myxococcus consume other microorganisms. Myxobacteria have a complex developmental cycle that involves the formation of fruiting bodies that contain myxospores.
**Q Compare and contrast the life cycle of Myxococcus with that of Bdellovibrio.**
15.17 The phylum Spirochaetes contains helically shaped bacteria that show a novel form of motility that allows them to “corkscrew” through viscous materials. These organisms are common in anoxic habitats and are the cause of many well-known human diseases, such as syphilis.
Q Contrast the motility of spirochetes with that of spirilla.
15.18 Prosthecate bacteria, such as Hyphomicrobium, Caulobacter, and Gallionella, are appendaged cells that form stalks or prosthecae used for attachment or nutrient absorption and are primarily aquatic. Some prosthecate bacteria, such as Hyphomicrobium, have a complex life cycle in which new cells form by budding from hyphae.
**Q Contrast the life cycle of Hyphomicrobium with that of Caulobacter.**
15.19 Sheathed bacteria are filamentous Proteobacteria in which individual cells form chains within an outer layer called the sheath. Sphaerotilus and Leptothrix are major genera of sheathed bacteria and can oxidize metals, such as Fe2+ and Mn2+.
**Q In what environment might you expect to find Leptothrix?**
15.20 Magnetosomes are specialized magnetic structures present in magnetotactic bacteria. Magnetosomes orient cells along the magnetic field lines of Earth, and this allows cells to use their normal chemotactic response to move vertically in a directed fashion in sediments or stratified aquatic systems.
Q In what way does a magnetosome contribute to the fitness of microaerophilic bacteria in sediments?
Application Questions
Describe a key physiological feature of the following Bacteria that would differentiate each from the others: Rhodobacter, Methylococcus, Azotobacter, Desulfovibrio, and Spirillum.
Describe the metabolism for each of the following Bacteria and state whether the organism is an aerobe or an anaerobe: Thiobacillus, Nitrosomonas, Methylomonas, Heliobacterium, Rhodospirillum, Myxococcus, and Gallionella.
Using an example from each of the morphologically diverse groups of Bacteria (Sections 15.16, 15.17, 15.18, 15.19 and 15.20), describe how you could distinguish them from each other using only microscopy. How do the habitats of your example organisms differ from each other? Could you find any of these organisms in or on the human body? Despite their ability to oxidize inorganic electron donors, why are Sphaerotilus and Leptothrix not considered chemolithotrophs?
Chapter Glossary
an organism that is an aerobic heterotroph that uses anoxygenic photosynthesis as a supplemental source of energy Carboxysome
a polyhedral cellular inclusion of crystalline ribulose bisphosphate carboxylase (RuBisCO), the key enzyme of the Calvin cycle Chemolithotroph
an organism able to oxidize inorganic compounds (such as H2, Fe2+, S0, or NH4 +) as energy sources (electron donors) Chlorosome
a cigar-shaped structure present in the periphery of cells of green sulfur and green nonsulfur bacteria and containing the antenna bacteriochlorophylls (c, d, or e) Consortium
a two- or more-membered association of bacteria, usually living in an intimate symbiotic fashion Convergent evolution
a circumstance where a trait or set of traits that are similar in form and/or function between two organisms are not inherited from a shared ancestor (that is, the traits are similar but not homologous) Cyanobacteria
prokaryotic oxygenic phototrophs containing chlorophyll a and in most cases phycobilins Denitrifier
an organism that carries out anaerobic respiration with NO3 − or NO2 −, reducing it to the gaseous products NO, N2O, and N2 Diazotroph
an organism that can assimilate N2 into biomass by activity of the enzyme nitrogenase Dissimilative sulfate-reducer
an anaerobic microorganism that conserves energy through the reduction of SO4 2− Dissimilative sulfur-oxidizer
a microorganism that gains energy for growth through oxidation of reduced sulfur compounds Dissimilative sulfur-reducer
an anaerobic or facultatively aerobic microorganism that conserves energy through the reduction of S0 but cannot reduce SO4 2− Ecological diversity
a concept for understanding microbial diversity that defines diversity in terms of differences in the interactions between organisms and their environments. Green nonsulfur bacteria
anoxygenic phototrophs containing chlorosomes, Q-type photosynthetic reaction centers, bacteriochlorophylls a and c as light-harvesting chlorophylls, and typically growing best as photoheterotrophs Green sulfur bacteria
anoxygenic phototrophs containing chlorosomes, FeS-type photosynthetic reaction centers, bacteriochlorophylls c, d, or e as antenna bacteriochlorophylls, and typically growing with H2S as an electron donor Heliobacteria
anoxygenic phototrophs containing bacteriochlorophyll g and FeS-type reaction centers Horizontal gene transfer
the unidirectional transfer of genes between cells through a process uncoupled from reproduction Metabolic diversity
a concept for understanding microbial diversity that defines diversity in terms of differences in the cellular processes that support microbial growth Methanotroph
an organism capable of oxidizing methane (CH4) as an electron donor in energy metabolism Methylotroph
an organism capable of oxidizing organic compounds that do not contain carbon–carbon bonds; if able to oxidize CH4, also a methanotroph Mixotroph
an organism that conserves energy from the oxidation of inorganic compounds but requires organic compounds as a carbon source Nitrifier
a chemolithotroph capable of carrying out the oxidation of NH3 or NO2 − Phycobilins
the pigments phycocyanin, allophycocyanin, and phycoerythrin that function as photosynthetic accessory pigments in cyanobacteria and are coupled to proteins to form phycobiliproteins Phylogenetic diversity
a concept for understanding microbial diversity that defines diversity in terms of differences in the evolutionary relationships between organisms Prochlorophyte
a bacterial oxygenic phototroph that contains chlorophylls a and b but lacks phycobilins Prosthecae
extrusions of cytoplasm, often forming distinct appendages, bounded by the cell wall Purple nonsulfur bacteria
a group of phototrophic bacteria that contain bacteriochlorophyll a or b and Q-type reaction centers, and that grow best as photoheterotrophs Purple sulfur bacteria
a group of phototrophic bacteria that contain bacteriochlorophylls a or b, have Q-type reaction centers, and can oxidize H2S as photosynthetic electron donor Spirilla (singular, spirillum)
a slender, tightly coiled, gram-negative bacterium of the phylum Spirochaetes characterized by possession of endoflagella used for motility