18 Diversity of Microbial Eukarya
**I Organelles and Phylogeny of Microbial *Eukarya***
Coccolithophores, Engineers of Global Climate
Coccolithophores are single-celled phototrophic eukaryotes. They live near the ocean surface worldwide, where they comprise a large portion of the planktonic community. Coccolithophores (photo inset, cell 6 μm in diameter) have armor plates made of calcium carbonate, formed from autotrophic CO2 fixation, and often develop immense blooms, dense clouds of cells so large they are visible from space (photo, Barents Sea, July 2016). Coccolithophores play an important role in global climate because their carbonate armor is heavy. It sinks to the ocean floor and is stabilized, eventually forming chalk, a type of sedimentary rock. Chalk deposits worldwide reveal the power of coccolithophores to remove carbon from the atmosphere and turn it into rock. Our ability to predict global climate depends, in part, on how much CO2 coccolithophores remove from the atmosphere.

The more than 200 species of coccolithophores vary tremendously in their distribution, ecology, and morphology (see Figure 18.15). No one knows for certain why they form armor plates, but we do know that forming these carbonate shells requires a very large amount of cellular energy. The shells may provide coccolithophores protection from predators or prevent bacterial or virus attachment. Alternatively, the shells might enhance photosynthetic efficiency by functioning as lenses, or in some way trap and concentrate CO2. They also might function to protect cells from ultraviolet radiation.
Since coccolithophores vary dramatically in shape, it seems likely that the costs and benefits of shell formation vary from species to species. Climate change is causing elevated CO2, higher temperatures, and greater ocean acidity. The former two conditions may enhance coccolithophore growth, whereas the latter should inhibit it. The fate of coccolithophores and their role in determining Earth’s future likely depends on their diversity and the costs and benefits of forming their carbonate armor in oceans gradually being transformed by climate change.
Source: Monteiro, F.M., et al. 2016. Why marine phytoplankton calcify. Sci. Adv. 2: e1501822.
I Organelles and Phylogeny of Microbial *Eukarya*
**Microbial Eukarya include protists, fungi, and algae. Primary symbioses caused these microbes to acquire mitochondria and red and green algae to also acquire chloroplasts. Many other protists acquired chloroplasts from the uptake of red or green algae in secondary symbioses.**
In this chapter, we consider the phylogeny and diversity of microbial eukaryotes. While plants and animals are the most conspicuous members of the Eukarya, microbial eukaryotes are far older and more phylogenetically diverse than are their macroscopic relatives. Eukarya exhibit tremendous morphological and ecological complexity but a limited range of metabolic diversity. Metabolically, eukaryotic organisms are almost exclusively chemoorganotrophic or phototrophic and most species are obligate aerobes. Nearly all Eukarya depend on their organelles—mitochondria and chloroplasts (Section 2.14)—for generating energy. The nutritional lifestyles of microbial eukaryotes are highly varied and include phagotrophy (obtaining energy by consuming other cells or detritus), saprotrophy (obtaining energy by absorbing organic matter from the environment), phototrophy (generating energy from light), and/or symbiosis (a persistent interaction with another organism that can be beneficial, commensal, or parasitic, Chapter 23).
Microbial eukaryotes can be defined by their morphology as protists, algae, or fungi. Protist is an informal term referring to any eukaryotic microorganism that is not a plant, animal, or fungus. Protists are found in many different phylogenetic lineages of Eukarya and are typically phagotrophic, though some are phototrophic. Algae include unicellular and multicellular eukaryotes that perform photosynthesis but are not plants. While some algal groups share an ancestor with plants, as we will see shortly, phototrophic microbes have evolved independently in multiple different lineages of Eukarya. Fungi are nonphototrophic eukaryotic microorganisms with rigid cell walls composed of chitin. The fungi form a single phylogenetic group, which includes yeasts, molds, and mushrooms. Fungi typically exhibit saprotrophic and/or symbiotic lifestyles and are often associated with (primarily aerobic) decomposition.
One thing that unites all Eukarya is that their ancestors had complex cell structure and true organelles. The structural complexity of the eukaryotic cell (Sections 2.13, 2.14 and 2.15) has allowed the domain Eukarya to evolve tremendous morphological diversity. As a result, complex cellular structures and multicellularity are common traits of eukaryotic microorganisms. We begin our consideration of microbial eukaryotes by examining the evolutionary history of the domain Eukarya and the endosymbiotic origins of organelles.
18.1 Endosymbioses and the Eukaryotic Cell
The exact evolutionary origin of the eukaryotic cell remains uncertain, but biologists agree that the eukaryotic cell is a genetic chimera containing genes from multiple sources. The last common ancestor of all eukaryotic cells was a single-celled microorganism closely related to Archaea. This microorganism had certain features common to all eukaryotic cells, including a nucleus, a cytoskeleton, spliceosomes, and a genome with spliceosomal introns (that is, those introns processed by spliceosomes), but it lacked energy-producing organelles—the mitochondria and chloroplasts. These were obtained by endosymbiosis (Section 13.4).
The acquisition of mitochondria was a foundational event in the origin of Eukarya. Mitochondria are derived from Bacteria and they contain bacterial DNA (Figure 18.1; Section 2.14). The acquisition of mitochondria and the establishment of eukaryotic cell structure set the stage for the evolution of diverse microbial lineages in addition to complex multicellular organisms such as plants and animals.
Figure 18.1 Organellar DNA.

Cells of the yeast Saccharomyces cerevisiae have been stained with the fluorescent dye DAPI that binds to DNA. Each mitochondrion has two to four circular chromosomes that stain blue with the dye.
Primary Endosymbioses
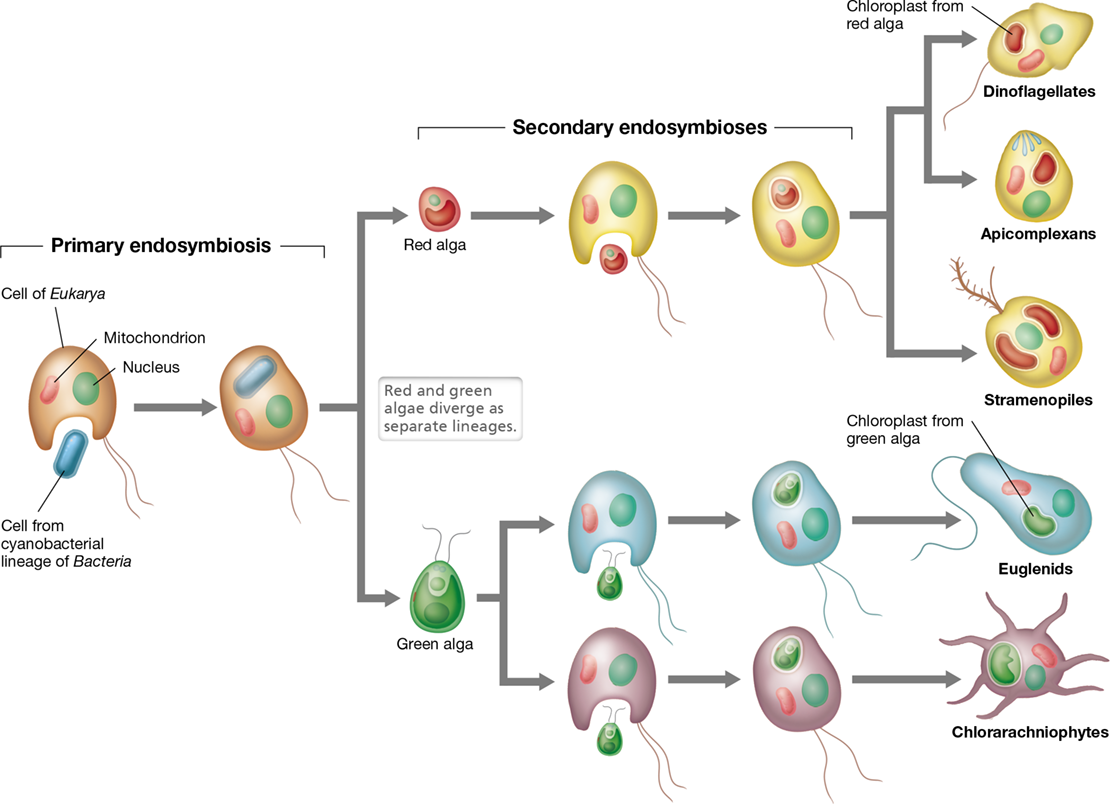
Endosymbiosis has played a major role in the origin and diversification of Eukarya. All Eukarya are descended from cells that contained respiratory mitochondria. Mitochondria evolved through endosymbiosis, in which the ancestor of Eukarya formed a symbiotic relationship with a bacterial cell capable of respiration (Sections 2.14 and 13.4). This bacterium may have initially invaded the cytoplasm of the eukaryotic cell as an intracellular parasite or it may have been acquired by phagocytosis, but for some reason these partners entered into a stable symbiotic relationship. Over time, the bacterial endosymbiont and its eukaryotic partner evolved mutual dependency until they eventually became a single cell, inseparable. This event is considered primary endosymbiosis because the bacterial symbiotic partner was acquired directly by the eukaryotic host cell.

All phototrophic eukaryotes—plants and algae—require chloroplasts to perform photosynthesis. Chloroplasts, like mitochondria, were acquired through primary endosymbiosis (Figure 18.2). In this case, a eukaryotic cell, which already contained mitochondria, acquired a phototrophic cyanobacterium as an endosymbiont (Section 13.4). As with the mitochondria, this cyanobacterial endosymbiont evolved mutual dependency with its eukaryotic host until they became inseparable. However, primary endosymbiosis is not the end of the story for the evolution of organelles.
Figure 18.2 Endosymbioses.

Following primary endosymbiotic association(s) leading to the mitochondrion, primary endosymbioses with phototrophic Bacteria led to the red and green algae. Secondary symbioses of green and red algae spread the property of photosynthesis to many independent lineages of protists.
Secondary Endosymbioses
Primary endosymbiosis with a cyanobacterium gave rise to the Archaeplastida, a lineage that includes green algae, red algae, and plants (Figure 18.2 and see Figure 18.3). However, following this primary endosymbiosis event, several different groups of protists acquired chloroplasts through secondary endosymbiosis. Secondary endosymbiosis occurs when a cell that already contains an endosymbiont is engulfed by a second host cell (Figure 18.2). For example, many protists became phototrophic by acquiring an algal endosymbiont that already contained a chloroplast. Over time, many features of the algal endosymbiont were lost but its chloroplasts remained, as did the ability to perform photosynthesis.
Secondary endosymbiosis has occurred multiple times within Eukarya, giving rise to a diversity of phototrophic eukaryotes (Figure 18.2). The chloroplasts of euglenids and chlorarachniophytes were acquired through secondary endosymbiosis with green algae, while those of the Alveolata (ciliates, apicomplexans, and dinoflagellates) and Stramenopiles were obtained through secondary endosymbioses with red algae (Figure 18.2 and see Figure 18.3). The ancestral red algal chloroplasts were apparently lost from some lineages, such as the ciliates, or became greatly reduced in size in others, such as the apicomplexans. In some other organisms, such as the dinoflagellates, the red algal chloroplast was replaced altogether with chloroplasts from different algae, including green algae. The legacy of endosymbiosis can be detected even if an endosymbiont is lost, because some endosymbiont genes are transferred to the host and become part of the host genome (Section 13.4); these genes are then revealed when the genome is sequenced.
These many examples of endosymbiotic events underscore the importance of endosymbiosis in the evolution and diversification of microbial eukaryotes. It is unlikely that primary endosymbiotic events occurred only twice in evolutionary history—after all, trial and error is the essence of evolution—and secondary endosymbioses are known to have occurred multiple times in several lineages (Figure 18.2). There are even examples of animals that engulf phototrophic protists to carry out photosynthesis for extended periods (Section 23.13). Indeed, it appears that endosymbioses are a common and ongoing occurrence in the eukaryotic world, conferring obvious metabolic advantages on the eukaryotic partner and perhaps other, not-so-obvious advantages, that remain to be discovered.
Check Your Understanding
Which was acquired first: chloroplasts or mitochondria?
Describe how euglenids came to acquire chloroplasts.
Distinguish between primary and secondary endosymbioses.
18.2 Phylogenetic Lineages of *Eukarya*
Based on conclusions drawn from molecular phylogenetic studies and supported by microfossil discoveries, it appears likely that a major phylogenetic radiation took place rather early in eukaryote evolution. This was likely triggered by the endosymbiotic acquisition of mitochondria, the foundational event in the evolution of the eukaryotic cell. All extant Eukarya contain mitochondria, structures homologous to mitochondria, or some genetic trace of these structures. What promoted this primary endosymbiotic event is unknown, but quite possibly it was the accumulation of O2 in the atmosphere from cyanobacterial photosynthesis (Figure 13.1). Mitochondria would have improved the ability of the early eukaryotic cell to generate energy, providing eukaryotes a considerable advantage over anaerobic competitors. Somewhat later, another primary endosymbiotic event occurred between eukaryotic cells and cyanobacteria, giving rise to the first algae. This event was followed by a series of secondary endosymbioses (Section 18.1 and Figure 18.2) that gave rise to the diversity of phototrophic eukaryotes we see today.
Eukaryotic Evolution: The Big Picture
Although phylogenies based on ribosomal RNA gene sequences (Chapter 13) confirm the three domains of life—Bacteria, Archaea, and Eukarya—our picture of eukaryotic evolution has changed dramatically with the incorporation of data from whole genome sequences. There are currently five supergroups of Eukarya (a supergroup is not an official taxon but rather a cluster of related phyla): Archaeplastida, the SAR clade, Excavates, Amoebozoa, and Opisthokonta (Figure 18.3). The Archaeplastida include the entire plant kingdom as well as all red algae, green algae, and glaucophytes; these phototrophic lineages resulted from the primary endosymbiosis of chloroplasts (Figure 18.2). The Stramenopiles, Alveolata, and Rhizaria include highly diverse protists including both heterotrophic and phototrophic species. These three groups share an ancestor and they are classified together into the SAR clade (an acronym for Stramenopiles, Alveolata, and Rhizaria). Where it occurs, phototrophy within these lineages was acquired as a result of secondary endosymbiosis (Figure 18.3 and Section 18.1). Excavates are diverse protists, many of which are anaerobic, and some of which have acquired phototrophy as a result of secondary endosymbiosis (Figures 18.2 and 18.3). The Amoebozoa include many forms of amoebae and slime molds, though cells with an amoeboid morphology occur in many other lineages of Eukarya as well. Finally, the Opisthokonta include the well-known kingdoms Fungi and Animalia.
Figure 18.3 Phylogenetic tree of *Eukarya*.

The tree diagram provides a schematic view of phylogenetic relationships for five of the best-characterized supergroups within the Eukarya. Colors are used to identify the supergroups in the tree. Dashed lines indicate both primary endosymbioses of mitochondria and chloroplasts, and the secondary endosymbioses of red and green algae. Protists are common in all eukaryotic lineages except the Opisthokonta (Fungi and Animalia) and the plants.
The phylogenetic tree shown in Figure 18.3 should not be considered the final word on eukaryotic evolution. For example, many phyla of Eukarya have been discovered that do not fall neatly within these five groups and their position in the tree of life remains uncertain. For example, the haptophytes (Section 18.7) are an unusual and globally important group of microbes that form their own separate branch deep within the eukaryotic phylogenetic tree (Figure 18.3). New genome sequences continue to be unraveled, new organisms continue to be discovered, and new aspects of eukaryotic biology continue to be revealed. With each new discovery, we shed more light on eukaryotic phylogeny, and hence our understanding of eukaryotic phylogeny will continue to improve as our knowledge grows.
Phylogenetic Insights on Endosymbiosis
As we have seen, endosymbiosis has clearly been an important aspect of eukaryotic evolution, and the acquisition of the mitochondrion by primitive Eukarya was central to the evolutionary success of this domain. However, some parasitic microbial eukaryotes such as Giardia and Microsporidia lack mitochondria. The Microsporidia were once thought to be ancient members of the Eukarya, descended from an ancestor of Eukarya that lacked mitochondria. We now know, however, that these amitochondriate eukaryotes are descended from eukaryotic ancestors that once had mitochondria (see the position of Microsporidia in Figure 18.24) but then lost them for some reason, perhaps while reverting to an anaerobic lifestyle such as in Giardia, Entamoeba, and several other anaerobic parasitic eukaryotes. However, the genomes of amitochondriate eukaryotes typically retain a few genes of mitochondrial origin, and these molecular leftovers are strong evidence that the organisms once had mitochondria.
The tree of Eukarya (Figure 18.3) also shows how secondary endosymbioses account for the origin of chloroplasts in some unicellular phototrophic eukaryotes. Following primary endosymbiosis of the cyanobacterial ancestor of chloroplasts by early mitochondrion-containing eukaryotes, these now phototrophic eukaryotes diverged into red and green algae. Then, in secondary endosymbioses, ancestors of the euglenids, kinetoplastids, and chlorarachniophytes engulfed green algae while ancestors of the Alveolata and Stramenopiles engulfed red algae (Section 18.1). These secondary endosymbioses account for the great phylogenetic diversity of phototrophic eukaryotes, which we explore later in this chapter.
With a feeling for the “big picture” of eukaryotic evolution, we now move on to consider microbial eukaryotes themselves. During our tour, a diverse array of nutritional lifestyles, cellular accessories, interesting habitats, and ecological strategies for survival will unfold.
Check Your Understanding
What are the five major supergroups within the Eukarya?
What supergroup was formed as a result of the primary endosymbiosis of chloroplasts?
What group of microbial eukaryotes is most closely related to animals?
II Protists
The term “protist” describes any microbial eukaryote that is not a plant, animal, or fungus. Protists are highly diverse in their phylogeny, morphology, and ecology, and their metabolisms range from chemoorganotrophic to phototrophic.
With the big picture of eukaryotic phylogeny as our guidepost, we begin with the protists, an enormously diverse group defined loosely by their morphology. Protists include both phototrophic and nonphototrophic microbial eukaryotes, and while most are unicellular, some are multicellular. Protists are widely distributed in nature, exhibit a wide range of morphologies, and show great phylogenetic diversity.
Key Genera: *Giardia, Trichomonas, Trypanosoma, Euglena*
The Excavates are a diverse group of protists including both chemoorganotrophs and phototrophs, with some species being anaerobic. Once considered as a coherent taxonomic group, the Excavates include a collection of distantly related organisms whose phylogeny remains controversial. We start with the diplomonads and parabasalids, flagellated protists that lack mitochondria and chloroplasts. These microbes live in anoxic habitats, such as animal intestines, either symbiotically or as parasites, and conserve energy from fermentation. Some diplomonads cause serious and common diseases in fish, domestic animals, and humans, and one parabasalid causes a major sexually transmitted disease of humans. Both groups share a relatively recent common ancestor before they diverged to form separate phylogenetic lineages (Figure 18.3).
Diplomonads
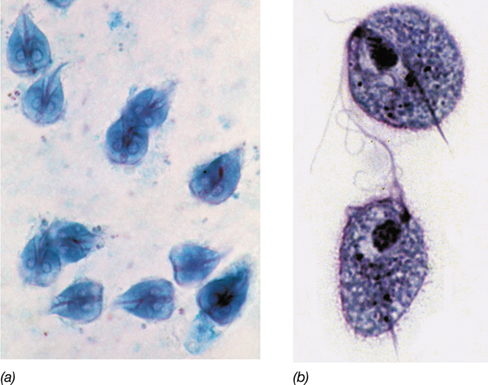
Diplomonads (**Figure 18.4*a***) characteristically contain two nuclei of equal size, and also contain mitosomes, much reduced mitochondria lacking electron transport proteins and enzymes of the citric acid cycle (Figure 3.12). The diplomonad Giardia has a relatively small genome for a eukaryote, about 12 megabase pairs (Mbp). The genome is also quite compact, contains few introns, and lacks genes for many metabolic pathways, including the citric acid cycle. These characteristics likely account for the organism’s parasitic and anaerobic lifestyle. Giardia intestinalis (Figure 18.4a), also known as Giardia lamblia, causes giardiasis, one of the most common waterborne diarrheal diseases in the United States.We examine human giardiasis in Section 34.4.
Figure 18.4 Diplomonads and parabasalids.

(a) Light photomicrograph of cells of Giardia intestinalis, a typical diplomonad. Note the dual nuclei. Cells are about 10 μm wide. (b) Light photomicrograph of cells of the parabasalid Trichomonas vaginalis. Cells are about 6 μm wide. The spearlike structure (axostyle) is used to attach the cell to urogenital tissues.
Parabasalids
Parabasalids contain a parabasal body that, among other functions, gives structural support to the cell’s Golgi complex. These anaerobic microbial eukaryotes lack mitochondria but contain hydrogenosomes. Hydrogenosomes are anaerobic organelles that resemble and evolved from mitochondria and produce ATP through fermentation reactions resulting in the production of H2 and acetate. Parabasalids live in the intestinal and urogenital tract of vertebrates and invertebrates as parasites or as commensal symbionts. The parabasalid Trichomonas vaginalis is motile by a tuft of flagella (Figure 18.4b) and causes a widespread sexually transmitted disease in humans (Section 34.4 and Figure 34.9).
The genomes of parabasalids are unique among eukaryotes in that most of them lack introns, the noncoding sequences characteristic of eukaryotic genes (Sections 6.6 and 10.4). In addition, the genome of T. vaginalis is surprisingly huge for a parasitic organism, about 160 Mbp, and includes genes acquired from bacteria by horizontal gene transfer. Much of the genome of T. vaginalis contains repetitive DNA sequences and transposable elements (Section 13.9), which has made genomic analyses difficult. But Trichomonas is still thought to contain nearly 60,000 genes, about twice that of the human genome and near the upper limit observed thus far for eukaryotic genomes.
Kinetoplastids
Kinetoplastids are a well-studied group of Excavates and are named for the presence of the kinetoplast, a mass of DNA present in their single, large mitochondrion. Kinetoplastids live primarily in aquatic habitats, where they feed on bacteria. Some species, however, are parasites of animals and cause serious diseases in humans and vertebrate animals. Cells of Trypanosoma, a genus of organisms that infect humans, are small, about 20 μm long, thin, and crescent-shaped. Trypanosomes have a single flagellum that originates in a basal body and folds back laterally across the cell where it is enclosed by a flap of cytoplasmic membrane (Figure 18.5). Both the flagellum and the membrane participate in propelling the organism, making effective movement possible even in viscous liquids, such as blood, where pathogenic trypanosomes are often found.
Figure 18.5 Trypanosomes.

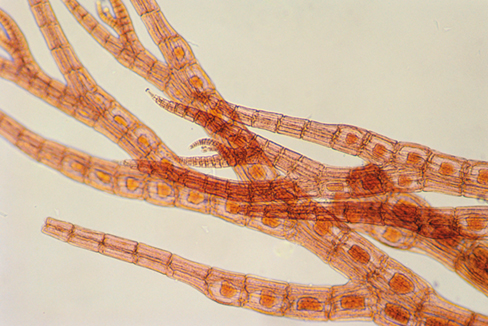
Photomicrograph of the flagellated kinetoplastids Trypanosoma brucei, the causative agent of African sleeping sickness. Blood smear preparation. The cells of T. brucei are about 3 μm wide.
Trypanosoma brucei (Figure 18.5) causes African sleeping sickness, a chronic and usually fatal human disease. The parasite lives and grows primarily in the bloodstream, but in the later stages of the disease it invades the central nervous system, causing an inflammation of the brain and spinal cord that is responsible for the characteristic neurological symptoms of the disease. The parasite is transmitted from host to host by the tsetse fly, Glossina spp., a bloodsucking fly found only in certain parts of Africa. After moving from the human to the fly in a blood meal, the parasite proliferates in the intestinal tract of the fly and invades the insect’s salivary glands and mouthparts, from which it is transferred to a new human host by a fly bite (Section 34.6).
Other kinetoplastids that are human parasites include Trypanosoma cruzi, the causative agent of Chagas disease, and Leishmania species, the causative agents of cutaneous and systemic leishmaniasis. Chagas disease is spread by the bite of a blood-feeding insect called the “kissing bug.” The disease is usually self-limiting, but it can become chronic and lead to a fatal infection. Leishmaniasis is a disease of tropical and subtropical regions transmitted to humans and other mammals by a bite from the sand fly. This potentially fatal disease can be localized to the skin surrounding a fly bite or can infect the spleen and liver and cause systemic infection.Both Chagas disease and leishmaniasis are covered in more detail in Section 34.6.
Euglenids
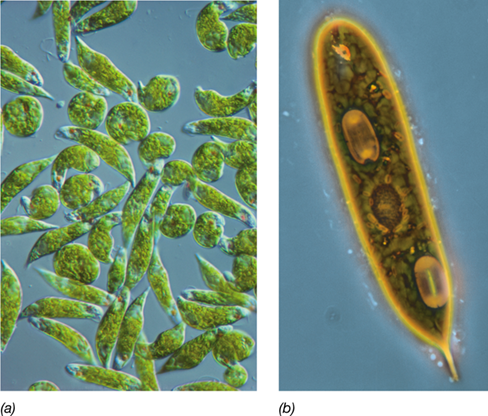
Another well-studied group of excavates are the euglenids (Figure 18.6). Unlike the kinetoplastids, these motile microbial eukaryotes are nonpathogenic and are both chemotrophic and phototrophic. Most euglenids contain two flagella, dorsal and ventral, and their active motility allows the organisms to access both illuminated and dark habitats in their environment to support their alternate nutritional lifestyles.
Figure 18.6 *Euglena*, a euglenid.

(a) This phototrophic protist, like other euglenids, is not pathogenic. A cell is about 15 μm wide. (b) High-magnification view.
Euglenids live exclusively in aquatic habitats, both freshwater and marine, and contain chloroplasts, which support phototrophic growth (Figure 18.6). In darkness, however, cells of Euglena, a typical euglenid, can lose their chloroplasts and exist as chemoorganotrophs. Many euglenids can also feed on bacterial cells via phagocytosis, a process of surrounding a particle with a portion of their flexible cytoplasmic membrane to engulf the particle and bring it into the cell. The engulfed particle remains in this structure (called the phagosome) where it is digested, and the released nutrients are made available to nourish the cell.
Check Your Understanding
Contrast the two nutritional options for Euglena.
What diseases are caused by Trypanosoma cruzi, Leishmania, and Giardia?
Key Genera: *Gonyaulax, Plasmodium, Paramecium*
The Alveolata as a group are characterized by their alveoli, cytoplasmic sacs located just under the cytoplasmic membrane. Although the function of alveoli is unknown, they may help the cell maintain osmotic balance by controlling water influx and efflux, and in the dinoflagellates, they may function as armor plates. Three phylogenetically distinct, although related, kinds of Alveolata are known: the ciliates, which use cilia for motility; the dinoflagellates, which are motile by means of a flagellum; and the apicomplexans, which are parasites of humans and other animals (Figure 18.3).
Ciliates
Ciliates possess cilia (Figure 18.7) at some stage of their life cycle. Cilia are structures that function in motility and may cover the cell or form tufts or rows, depending on the species (Section 2.15 and Figure 2.48a). Probably the best-known and most widely distributed ciliates are those of the genus Paramecium (Figure 18.7). Like many other ciliates, Paramecium use cilia both for motility and to obtain food by ingesting particulate materials such as bacterial cells through a distinctive funnel-shaped oral groove. Cilia that line the oral groove move material down the groove to the cell mouth, also called the gullet (Figure 18.7b). Once in the gullet, the material is enclosed in a vacuole by phagocytosis. Digestive enzymes secreted into the vacuole then break down the material as a source of nutrients.
Figure 18.7 *Paramecium*, a ciliated protist.

(a) Phase-contrast photomicrograph. (b) Scanning electron micrograph. Note the cilia in both micrographs. A single Paramecium cell is about 60 μm in diameter.
Ciliates are unique among protists in having two kinds of nuclei, micronuclei and macronuclei. Genes in the macronucleus regulate basic cellular functions, such as growth and feeding, whereas those of the micronucleus are involved in sexual reproduction, which occurs through a partial fusion of two Paramecium cells and exchange of micronuclei. The genome of Paramecium is huge, with macronuclear genes numbering about 40,000, nearly twice that of humans.
Many Paramecium species (as well as many other protists) are hosts for endosymbiotic Bacteria, Archaea, or eukaryotes, the latter usually green algae. These organisms may play a nutritional role, synthesizing vitamins or other growth factors used by the host cell. Several anaerobic ciliated protists also contain endosymbionts. For example, ciliated protists in the termite hindgut contain endosymbiotic methanogens (Archaea) that consume H2 plus CO2 to form methane (CH4). Ciliates themselves can also be symbiotic: Obligately anaerobic ciliates are present in the rumen, the forestomach of ruminant animals, and play an important role in the digestive and fermentative processes of the animal (Sections 23.15 and 23.16).
Some ciliates are animal parasites, although this lifestyle is less common in ciliates than in some other groups of protists. The species Balantidium coli (Figure 18.8), for example, is primarily an intestinal parasite of domestic animals but occasionally infects the intestinal tract of humans, producing dysentery-like symptoms. Cells of B. coli form cysts (Figure 18.8) that promote their transmission in infected food or water.
Figure 18.8 *Balantidium coli*, a ciliated protist that causes a dysentery-like disease in humans.
The dark blue–stained lobed structure in this B. coli cyst obtained from swine intestine is a dividing macronucleus. The cell is about 5 μm wide.
Dinoflagellates
Dinoflagellates are a diverse group of motile marine and freshwater phototrophic organisms (Figure 18.9) that acquired the capacity to photosynthesize through secondary endosymbioses (Figures 18.2 and 18.3). Eukaryotic flagella (Section 2.15 and Figure 2.48) encircle the cell and impart spinning movements that give dinoflagellates their name (dinos is Greek for “whirling”). Dinoflagellates have two flagella of different lengths and with different points of insertion into the cell, transverse and longitudinal. The transverse flagellum is attached laterally, whereas the longitudinal flagellum originates from the lateral groove of the cell and extends lengthwise (see Figure 18.10b). Some dinoflagellates are free-living, whereas others live a symbiotic existence with animals that form coral reefs, obtaining a sheltered and protected habitat in exchange for supplying phototrophically fixed carbon as a food source for the reef. A number of free-living species are capable of bioluminescence and emit light when disturbed at night. This bioluminescence results in a “sparkling” effect that can often be observed in coastal seas and bioluminescent bays.
Figure 18.9 The marine dinoflagellate *Ornithocercus magnificus* (an alveolate).

The cell proper is the globular central structure; the attached ornate structures are called lists. A cell is about 30 μm wide.
Several species of dinoflagellates are toxic and can cause harmful algal blooms (see MicrobiologyNow in the Chapter 15 opener for other toxic microbial blooms). For example, dense suspensions of Gonyaulax cells, called “red tides” (**Figure 18.10*a***) because of the red-colored pigments of this organism, can form in warm and typically polluted coastal waters. Such blooms are often associated with fish kills and poisoning in humans following consumption of mussels that have accumulated Gonyaulax cells (Figure 18.10b) through filter feeding. Toxicity results from a neurotoxin that can cause a condition called paralytic shellfish poisoning in humans and some marine animals, such as sea otters. Symptoms include numbness of the lips, dizziness, and difficulty breathing; in severe cases, death can result from respiratory failure. Pfiesteria is another toxic dinoflagellate. Toxic spores of Pfiesteria piscicida (Figure 18.10c) infect fish and eventually kill them by way of neurotoxins that affect movement and destroy skin. Lesions form on areas of the fish, allowing opportunistic bacterial pathogens to grow (Figure 18.10d). Symptoms of human toxemia from Pfiesteria poisoning include skin rashes and respiratory problems.
Figure 18.10 Toxic dinoflagellates (alveolates).

(a) Photograph of a “red tide” caused by massive growth of toxin-producing dinoflagellates such as Gonyaulax. The toxin is excreted into the water and also accumulates in shellfish that feed on the dinoflagellates. (b) Colorized scanning electron micrograph of a cell of the dinoflagellate Gonyaulax. The cell is about 30 μm wide. (c) Scanning electron micrograph of a toxic spore of Pfiesteria piscicida; the structure is about 12 μm wide. (d) A fish killed by P. piscicida; note the lesions of decaying flesh.
Apicomplexans
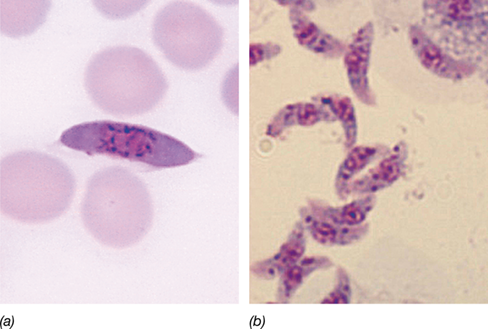
Apicomplexans are nonphototrophic obligate parasites that cause severe human diseases such as malaria (Plasmodium species, **Figure 18.11*a***), toxoplasmosis (Toxoplasma, Figure 18.11b), and coccidiosis (Eimeria). These organisms are characterized by nonmotile adult stages, and nutrients are taken up in soluble form across the cytoplasmic membrane as in bacteria and fungi.
Figure 18.11 Apicomplexans.

(a) A gametocyte of Plasmodium falciparum in a blood smear. The gametocyte is the stage in the malarial parasite life cycle that infects the mosquito vector. (b) Sporozoites of Toxoplasma gondii.
Apicomplexans produce structures called sporozoites (Figure 18.11b; Section 34.5 and Figure 34.12), which function in transmission of the parasite to a new host, and the name apicomplexan derives from the presence at one apex of the sporozoite of a complex of organelles that penetrate host cells. Apicomplexans also contain apicoplasts. These are degenerate chloroplasts that lack pigments and photosynthetic capacity but contain a few of their own genes. Apicoplasts catalyze fatty acid, isoprenoid, and heme biosyntheses, and export their products to the cytoplasm. It is hypothesized that apicoplasts are derived from red algal cells engulfed by apicomplexans in a secondary endosymbiosis (Figures 18.2 and 18.3). Over time, the chloroplast of the red algal cell degenerated to play a nonphototrophic role in the apicomplexan cell.
Both vertebrates and invertebrates can be hosts for apicomplexans. In some cases, an alternation of hosts takes place, with some stages of the life cycle linked to one host and some to another. Important apicomplexans are the coccidia, which are typically parasites of birds, and species of Plasmodium, which are malarial parasites (Figure 18.11a).We reserve detailed discussion of malaria—a disease that has killed more humans than any other disease—for Section 34.5.
Check Your Understanding
What health problem is associated with the organism Gonyaulax?
What are apicoplasts, which organisms have them, and what are their functions?
Key Genera: *Phytophthora, Nitzschia, Ochromonas, Macrocystis*
The Stramenopiles include both chemoorganotrophic and phototrophic microorganisms as well as macroorganisms. Members of this group bear flagella with many short, hairlike extensions (Figure 18.2), and this morphological feature gives the group its name (from Latin stramen for “straw” and pilus for “hair”). The diatoms, oomycetes, golden algae, and brown algae are the major groups of Stramenopiles (Figure 18.3).
Diatoms
Diatoms include over 200 genera of unicellular, phototrophic, microbial eukaryotes, and are major components of the planktonic (suspended) phytoplankton microbial community in marine and freshwaters. Diatoms characteristically produce an exoskeleton made of silica to which protein and polysaccharide are added. The exoskeleton, which protects the cell against predation, exhibits widely different shapes in different species and can be highly ornate (Figure 18.12). This external structure, called a frustule, often remains after the cell dies and the organic materials have disappeared. Diatom frustules typically show morphological symmetry, including pinnate symmetry (having similar parts arranged on opposite sides of an axis, as in the common diatom Nitzschia, Figure 18.12b), and radial symmetry, as in the marine diatoms Thalassiosira and Asterolampra (Figure 18.12c, d). Because the diatom frustules, which are composed mainly of silica, are resistant to decay, these structures can remain intact for long periods and often sink and remain in the sediments for millions of years. Diatom frustules constitute some of the best-known unicellular eukaryotic fossils, and from dating of frustule samples, it has been shown that diatoms first appeared on Earth relatively recently, about 200 million years ago.
Figure 18.12 Diatom frustules.

(a) Dark-field photomicrograph of a collage of frustules from different diatom species showing various forms of symmetry. (b–d) Scanning electron micrographs of diatom frustules showing pinnate (part b) or radial (parts c, d ) symmetry. Diatoms vary considerably in size from very small species about 5 μm wide to larger species up to 200 μm wide.
Oomycetes
The oomycetes, also called water molds, were previously grouped with fungi based on their filamentous growth and the presence of coenocytic (that is, multinucleate) hyphae, morphological traits characteristic of fungi (Section 18.9). Phylogenetically, however, the oomycetes are distant from fungi and are closely related to other Stramenopiles (Figure 18.3). Oomycetes differ from fungi in other fundamental ways, as well. For example, the cell walls of oomycetes are typically made of cellulose instead of the chitin cell walls of fungi, and the water molds have flagellated cells, which are lacking in all but a few fungi. Nonetheless, oomycetes are ecologically similar to fungi in that they grow as a mass of hyphae decomposing dead plant and animal material in aquatic habitats.
Oomycetes have had a major impact on human society, as many species are plant pathogens (phytopathogens). The oomycete Phytophthora infestans, which causes late blight disease of potatoes, contributed to massive famines in Ireland in the mid-nineteenth century. The famines led to the death of a million Irish and triggered great waves of Irish immigration to North America. Other major phytopathogens include Pythium, a common pathogen of greenhouse seedlings, and Albugo, which causes “white rusts” on several agricultural crops.
Golden Algae and Brown Algae
Along with the diatoms, golden and brown algae form major lineages of Stramenopiles. Golden algae, also called chrysophytes, are primarily unicellular marine and freshwater phototrophs. Some species are chemoorganotrophs and feed either by phagocytosis or by transporting soluble organic compounds across the cytoplasmic membrane. Some golden algae, such as Dinobryon (Figure 18.13a), found in freshwater, are colonial. However, most golden algae are unicellular and motile by the activity of two flagella of unequal length.
Figure 18.13 Golden and brown algae.

(a) Dinobryon, a golden alga (family Chrysophyceae) that forms branched colonies. (b) Macrocystis, a marine kelp belonging to the brown algae (family Phaeophyceae). (c) Ochromonas, a unicellular chrysophyte. The golden or brown color of the chloroplasts of these algae is due to the pigment fucoxanthin.
Golden algae are so named because of their golden-brown color (Figure 18.13a, c). This is due to chloroplast pigments dominated by the brown-colored carotenoid fucoxanthin. The major chlorophyll pigment in golden algae is chlorophyll c rather than chlorophyll a, and they lack the phycobiliproteins present in red algal chloroplasts (Section 18.15). Cells of the unicellular golden alga Ochromonas, the best-studied genus of this group, have only one or two chloroplasts (Figure 18.13c).
Brown algae are primarily marine and are multicellular and typically macroscopic. No unicellular brown algae are known. The kelps, such as the giant kelp Macrocystis (Figure 18.13b), which can grow up to 50 m in length, are perhaps the most widespread of brown algae. Fucus, another common seaweed of intertidal regions, can grow up to 2 m. As their name implies, brown algae are brown or green-brown in color depending on how much of the carotenoid pigment fucoxanthin they produce. Most marine “seaweeds” are brown algae and their rapid growth, especially in cold marine waters, can cause nuisance odor problems when they wash ashore and decay.
Check Your Understanding
What structure of diatoms accounts for their excellent fossil record?
In what ways do oomycetes differ from and resemble fungi?
Which chlorophyll pigment is found in golden and brown algae?
Chlorarachniophytes
Chlorarachniophytes are freshwater and marine amoeba-like phototrophs that develop a flagellum for dispersal. The group’s acquisition of green algal chloroplasts is a prime example of a secondary endosymbiosis (Figure 18.2) and shows how extensively this process has molded several phylogenetically distinct lineages of microbial eukaryotes (Figure 18.3).
Chloroplasts typically have two membranes (Section 2.14; Figures 2.46 and 14.9), derived from the inner and outer membranes of Cyanobacteria, which have a gram-negative cell envelope (Sections 2.3 and 2.4). Chloroplasts in chlorarachniophytes, however, have four membranes. In addition, they have nucleomorphs tucked in between the two sets of chloroplast membranes. Nucleomorphs are remnant nuclei left over from when the algal endosymbionts were acquired during secondary endosymbiosis (Section 18.1). Both the nucleomorphs and the extra membranes were derived from this algal endosymbiont (Figure 18.2).
It is a testament to endosymbiosis to consider that chlorarachniophytes have merged a minimum of five different genomes: the host genome, the host mitochondrial genome, the algal endosymbiont genome (which has become the nucleomorph), the algal endosymbiont mitochondrial genome, and the algal endosymbiont chloroplast genome! Most nonessential or duplicate genes have been deleted over time, and many genes from the mitochondria and chloroplasts have been transferred to the host genome. As a result, the majority of genes in a chlorarachniophyte are present in the nucleus of the host cell. The nucleomorph itself is greatly reduced in size relative to its ancestor and over time may be lost completely from the chloroplast.
Foraminiferans
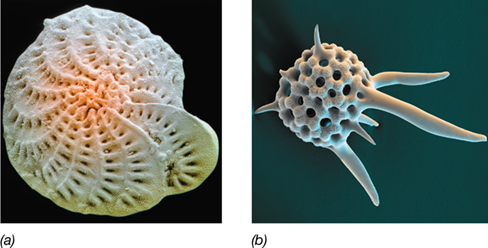
In contrast to chlorarachniophytes, foraminiferans are exclusively marine microbes and form shell-like structures called tests, which have distinctive characteristics and are often quite ornate (**Figure 18.14*a***). Tests are typically made of organic materials reinforced with calcium carbonate. The test is not firmly attached to the cell, and the amoeba-like cell may extend partway out of the test during feeding. However, because of the weight of the test, the cell usually sinks to the bottom of the water column, and it is thought that the organisms feed on dissolved organic matter and particulate deposits, primarily bacteria, other protists, and the remains of dead organisms near the sediments. Foraminiferan cells can also host a variety of algae that form endosymbiotic relationships with the protist and supply it with organic carbon, probably in exchange for inorganic nutrients derived from the breakdown of dead organisms. Phototrophs are found primarily in planktonic foraminifera that remain suspended in the water column to provide their endosymbionts with sufficient sunlight.
Figure 18.14 Foraminifera and radiolaria.

(a) A foraminiferan. Note the ornate and multilobed test. The test is about 1 mm wide. (b) A spiked radiolarian of the Nassellaria group. A test is about 150 μm wide. Both a and b are colorized scanning electron micrographs.
Foraminiferan tests (Figure 18.14a) are relatively resistant to decay and are readily fossilized. These buried and preserved tests are quite useful to geologists. Because particular taxa of foraminifera are typically associated with particular strata in the geological record, fossilized foraminiferan tests in samples obtained from exploratory wells are used by oil industry paleontologists as a means to date and assess the petroleum potential of a given drill site.
Radiolarians
Radiolarians are chemoorganotrophic and mostly planktonic marine eukaryotes that reside in the upper 100 m of the ocean where they consume bacteria and particulate organic matter. Some species associate with algae that take on a symbiotic (but not endosymbiotic) role and supply nutrients to the radiolarian.
The name “radiolarian” comes from the radial symmetry of their tests, transparent or translucent mineral skeletons made of silica in one fused piece (Figure 18.14b). Along with the accumulation of lipid droplets and large cytoplasmic vacuoles, the needle-like pseudopodia of radiolarians probably help keep the organisms from sinking in their mainly open ocean (planktonic) habitats. However, when cells eventually die, their tests settle to the ocean floor and can build up over time into thick layers of slowly decaying cell material.
Check Your Understanding
What structure distinguishes rhizaria from all other protists?
In what way are foraminifera similar to diatoms? In what ways are they different?
How has endosymbiosis shaped the genomes of chlorarachniophytes?
KEY GENUS: *Emiliania*
Haptophytes are a group of unicellular and free-living phototrophic protists found in aquatic environments. Members of this group are widespread and abundant in the oceans, where they have a major impact on the biosphere. All haptophytes share several structural features including the presence of two flagella, two chloroplasts (derived from secondary endosymbiosis of red algae, Figures 18.2 and 18.3), and a unique structure called a haptonema. While all haptophytes are phototrophic, some are mixotrophic, obtaining energy both from photosynthesis and from the ingestion of smaller organisms or detritus. The haptonema, for which the group is named, is used when feeding, but it can also facilitate attachment and the avoidance of predators.
Coccolithophores
Haptophytes, at some point in their life cycle, typically possess rigid polysaccharide plates on their outer surface. The morphology of these plates can vary dramatically between species. One major group of haptophytes, the coccolithophores, are distinguished by their ability to form intricate mineralized plates made of calcite, a form of calcium carbonate.
Coccolithophores (Figure 18.15) are one of the most abundant and widely distributed unicellular phototrophic eukaryotes in the oceans. One of the most widespread and well characterized of the coccolithophores is Emiliania huxleyi. Coccolithophores are found in the photic zone of the ocean from tropical to subpolar latitudes. They can contribute 1 to 10% of the primary productivity in these systems under normal conditions, but they can also form massive blooms in which they account for more than 40% of photosynthetic activity. These blooms can span hundreds of thousands of square kilometers and are visible from space (see MicrobiologyNow in the chapter opener).
Figure 18.15 Haptophytes.

Scanning electron micrographs showing a collection of diploid cells of different species of coccolithophores. These phototrophic microbes have an exoskeleton composed of interlocking coccoliths made from calcium carbonate. Emiliania huxleyi, center top, is about 10 μm in diameter.
Coccolithophores have an exoskeleton composed of coccoliths, which are plates made from calcium carbonate (Figure 18.15). The plates could have several purposes. For example, they might be used to enhance photosynthesis by attenuating or focusing sunlight into the cell. Alternatively, the plates may protect the cell from grazing or from attack by bacteria or viruses.
Whatever their purpose, the production of coccolithophore plates has a major impact on Earth’s carbon cycle and, by extension, its climate. This is because the plates make coccoliths heavy, and the cells eventually sink, exporting carbon to the seafloor in the form of calcite. In this way, coccolithophores are a “biological pump” that removes CO2 from the atmosphere and deposits it on the seafloor where it will eventually form sedimentary rocks (Section 21.7 and Figure 21.20c). Indeed, coccoliths are a main component of the chalk deposits found in many regions of the world such as those of the famous White Cliffs of Dover in England. These deposits represent millions of years of carbon deposited on the seafloor, and they are a dramatic illustration of the power of microbes to influence the biosphere.
Life Cycle
Haptophytes have a life cycle that alternates between haploid and diploid cells; only haploid cells contain a haptonema. In the case of E. huxleyi (Figure 18.15; Figure 21.20a), diploid cells are observed most commonly and are responsible for the formation of blooms. Diploid cells are nonmotile, reproduce by binary fission to form diploid progeny, and form structurally complex exoskeletons composed of interlocking coccolith scales (Figure 18.15). In response to an environmental trigger, the nature of which remains poorly characterized, diploid cells undergo meiosis resulting in haploid progeny. The haploid cells are motile and do not form interlocking calcified exoskeletons. These noncalcified haploid cells divide by binary fission until some environmental trigger causes them to fuse with another haploid cell, returning the organism to the diploid state.
The two life stages of the coccolithophores are thus so morphologically different that they can easily be mistaken for different species. In nature, however, it is likely that the exoskeleton-containing (diploid) form predominates because this is the form seen in the massive blooms of coccolithophores in oceans worldwide.
Check Your Understanding
In what ways are coccolithophores similar to diatoms? How do they differ?
In what way do coccolithophores influence the global climate?
Key Genera: *Amoeba, Entamoeba, Physarum, Dictyostelium*
The Amoebozoa are a large group of terrestrial and aquatic protists that use lobe-shaped pseudopodia for movement and feeding, in contrast to the threadlike pseudopodia of rhizaria. The major groups of Amoebozoa are the gymnamoebas, the entamoebas, and the plasmodial and cellular slime molds. Phylogenetically, the Amoebozoa diverged from a lineage that eventually led to the fungi and animals (Figure 18.3).
Gymnamoebas and Entamoebas
The gymnamoebas are free-living protists that inhabit aquatic and soil environments. They use pseudopodia to move by a process called amoeboid movement (Figure 18.16) and feed by phagocytosis on bacteria, other protists, and particulate organic materials. Amoeboid movement results from streaming of the cytoplasm as it flows forward at the less contracted and viscous cell tip, taking the path of least resistance. Cytoplasmic streaming is facilitated by microfilaments (Section 2.15), which exist in a thin layer just beneath the cytoplasmic membrane. Amoeba (Figure 18.16) is a common organism in pond waters, with species varying in size from 15 μm in diameter (clearly microscopic) to over 750 μm (visible with the naked eye).
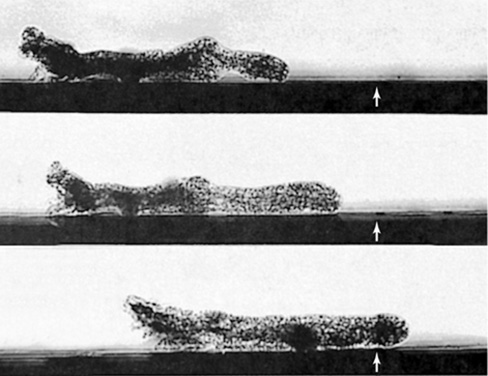
Figure 18.16 Time-lapse view of the amoebozoan *Amoeba proteus*.

The time interval from top to bottom is about 6 sec. The arrows point to a fixed spot on the surface. A single cell is about 80 μm wide.
In contrast to gymnamoebas, the entamoebas are parasites of vertebrates and invertebrates. Their usual habitat is the oral cavity or intestinal tract of animals. The anaerobe Entamoeba histolytica is pathogenic in humans and causes amebic dysentery, an ulceration of the intestinal tract that results in bloody diarrhea. This parasite forms cysts that are transmitted from person to person by fecal contamination of water, food, and eating utensils. In Section 34.00 we discuss the etiology and pathogenesis of amebic dysentery, an important cause of death from intestinal parasites in humans.
Slime Molds
The slime molds were previously grouped with fungi since they undergo similar life cycles and produce fruiting bodies with spores for dispersal. As protists, however, slime molds are motile and can move across a solid surface fairly quickly (see Figures 18.17–18.19). Slime molds are divided into two groups: plasmodial slime molds (also called acellular slime molds), whose vegetative forms are masses of protoplasm of indefinite size and shape called plasmodia (Figure 18.17), and cellular slime molds, whose vegetative forms are single amoebae. Slime molds live primarily on decaying plant matter, such as leaf litter, logs, and soil, where they consume other microorganisms, especially bacteria. Slime molds can maintain themselves in a vegetative state for long periods but eventually form differentiated spore-like structures that can remain dormant and then germinate later to once again generate the active amoeboid state.
Figure 18.17 Slime mold.

The plasmodial slime mold Physarum growing on an agar surface. The plasmodium is about 5 cm long and 3.5 cm wide.
Plasmodial slime molds, such as Physarum, exist in the vegetative phase as an expanding single mass of protoplasm called the plasmodium that contains many diploid nuclei (Figure 18.17). The plasmodium is actively motile by amoeboid movement, and from this phase, a sporangium containing haploid spores can be produced. When conditions are favorable the spores germinate to yield haploid flagellated swarm cells. The fusion of two swarm cells then regenerates a diploid plasmodium.
In contrast to their plasmodial relatives, cellular slime molds are individual haploid cells and form diploids only under certain conditions. The well-studied cellular slime mold Dictyostelium discoideum undergoes an asexual life cycle in which vegetative cells aggregate, migrate as a cell mass, and eventually produce fruiting bodies in which cells differentiate and form spores (Figures 18.18 and 18.19). When cells of Dictyostelium are starved, they aggregate and form a pseudoplasmodium; in this stage cells lose their individuality, but do not fuse. Aggregation is triggered by the production of cyclic adenosine monophosphate (cAMP). The first cells of Dictyostelium that produce this compound attract neighboring cells and eventually aggregate into motile masses of cells called slugs. Fruiting body formation begins when the slug becomes stationary and vertically oriented. The emerging structure differentiates into a stalk and a head. The stalk cells form cellulose, which provides the rigidity of the stalk, and the head cells differentiate into spores. Eventually, spores are released and dispersed, with each spore forming a new amoeba (Figures 18.18 and 18.19).
Figure 18.18 Photomicrographs of various stages in the life cycle of the cellular slime mold *Dictyostelium discoideum*.

(a) Amoebae in pre-aggregation stage. (b) Aggregating amoebae. Amoebae are about 300 μm in diameter. (c) Low-power view of aggregating amoebae. (d) Migrating pseudoplasmodia (slugs) moving on an agar surface and leaving trails of slime behind. (e, f) Early stages of fruiting body. (g) Mature fruiting bodies. Figure 18.19 shows the sizes of these structures. Dictyostelium has long served as a model for development in multicellular organisms, and its genome of 12,500 genes is about half that of the human genome.
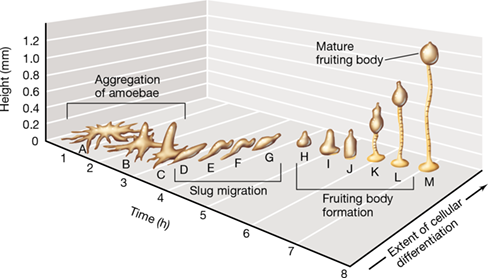
Figure 18.19 Stages in fruiting body formation in the cellular slime mold *Dictyostelium discoideum*.

(A–C) Aggregation of amoebae. (D–G) Migration of the slug formed from aggregated amoebae. (H–L) Culmination of migration and formation of the fruiting body. (M) Mature fruiting body composed of stalk and head. Cells from the rear of the slug form the head and become spores. Dictyostelium also undergoes sexual reproduction (not shown) when two amoebae fuse to form a macrocyst; the fused nuclei in the macrocyst return to the haploid stage when meiosis forms new vegetative amoebae.
Mastering Microbiology
Art Activity: Figure 18.18 Stages in fruiting body formation in cellular slime mold Dictyostelium discoideum
In addition to this asexual process, Dictyostelium can produce sexual spores. These form when two amoebae in an aggregate fuse to form a single giant amoeba. A thick cellulose wall develops around this cell to form a structure called the macrocyst, and this can remain dormant for long periods. Eventually, the diploid nucleus undergoes meiosis to form haploid nuclei that become integrated into new amoebae that can once again initiate the asexual cycle.
In Part III, we consider the fungi, a huge group of microbial eukaryotes whose service to the ecosystem by decomposing dead organic material cannot be understated.
Check Your Understanding
How can amoebozoans be distinguished from rhizaria?
Compare and contrast the lifestyles of gymnamoebas and entamoebas.
Describe the major steps in the life cycle of Dictyostelium discoideum.
III Fungi
Fungi are nonphototrophic eukaryotic microbes that form rigid cell walls made of chitin. Fungi are major agents of decomposition in soil and other habitats, and some form symbiotic relationships with plants.
The Fungi are a large, diverse, and widespread group of mostly nonmotile organisms that includes such well-known groups as the molds, mushrooms, and yeasts. Approximately 100,000 fungal species have been described, and as many as 1.5 million species could exist. Fungi form a phylogenetic cluster distinct from protists and are the microbial group most closely related to animals (Figure 18.3).
Most fungal species are microscopic and terrestrial. They inhabit soil or dead plant matter and play crucial roles in the decomposition of organic matter. A large number of fungal species are plant pathogens, and a few cause diseases of animals, including humans (Chapter 34). Certain species of fungi also establish symbiotic associations with plants, facilitating the plant’s acquisition of minerals from soil, and many fungi benefit humans through fermentation and the synthesis of antibiotics.
18.9 Fungal Physiology, Structure, and Symbioses
In this section we describe some general features of fungi, including their physiology, cell structure, and the symbiotic associations they develop with plants and animals. In the following section, we examine fungal reproduction and phylogeny.
Nutrition, Physiology, and Ecology
Fungi are chemoorganotrophs—typically displaying simple nutritional requirements—and most are aerobic. Fungi nourish themselves by secreting extracellular enzymes that digest polymeric materials, such as polysaccharides or proteins, into monomers that are assimilated as sources of carbon and energy. As decomposers, fungi digest dead animal and plant materials. As parasites of plants or animals, fungi use the same mode of nutrition but take up nutrients from the living cells of the plants and animals they invade rather than from dead organic materials.
A major ecological activity of fungi, especially the basidiomycetes, is the decomposition of wood, paper, cloth, and other products derived from these natural sources. Lignin, a complex polymer in which the building blocks are phenolic compounds, is an important constituent of woody plants, and in association with cellulose, it confers rigidity on them. Lignin is decomposed in nature almost exclusively through the activities of certain basidiomycetes called wood-rotting fungi. Two types of wood rot are known: brown rot, in which the cellulose is attacked preferentially while the lignin is left unmetabolized, and white rot, in which both cellulose and lignin are decomposed. The white rot fungi are of major ecological importance because they play such a key role in decomposing woody materials in forests.
Fungal Morphology, Spores, and Cell Walls
Most fungi are multicellular, forming a network of filaments called hyphae (singular, hypha) from which asexual spores are produced (Figure 18.20). Hyphae are tubular cell walls that surround the cytoplasmic membrane. Fungal hyphae are often septate, with cross-walls dividing each hypha into separate cells. In some cases, however, the vegetative cell of a fungal hypha contains more than one nucleus, and hundreds of nuclei can form as a result of repeated nuclear divisions without the formation of cross-walls, a condition called coenocytic. Each hyphal filament grows mainly at the tip by extension of the terminal cell (Figure 18.20).
Figure 18.20 Fungal structure and growth.

(a) Scanning electron micrograph of asexual spores of a typical mold, Penicillium chrysogenum. The conidiophore is shown in green and the conidia are yellow. (b) Diagram of a mold life cycle. The conidia can be dispersed by either wind or animals and are about 2 μm wide.
Mastering Microbiology
Art Activity: Figure 18.19 Fungal structure and growth
Hyphae typically grow together across and above a surface to form a compact, macroscopically visible tuft called a mycelium (**Figure 18.21*a***). From the mycelium, aerial hyphae reach up into the air above the surface, and spores called conidia are formed on their tips (Figure 18.21b). Conidia are asexual spores and they are often pigmented black, green, red, yellow, or brown (Figure 18.21). Conidia give the mycelium a dusty appearance (Figure 18.21a) and function to disperse the fungus to new habitats. Some fungi form macroscopic reproductive structures called fruiting bodies (mushrooms or puffballs, for example), in which millions of spores are produced that can be dispersed by wind, water, or animals (Figure 18.22). In contrast to mycelial fungi, some fungi grow as single cells; these are the yeasts.
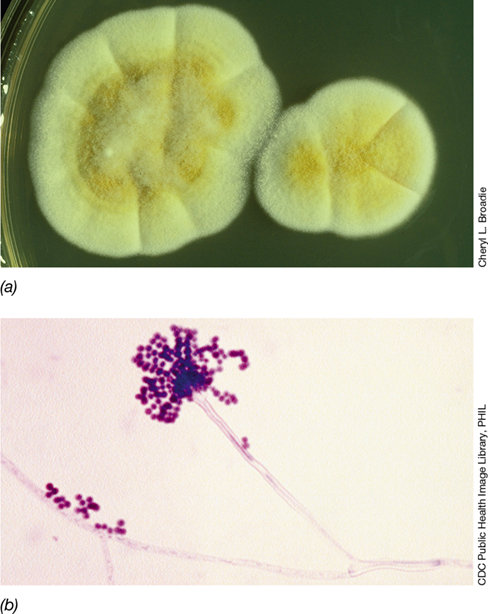
Figure 18.21 Hyphal fungi (molds).

(a) Colonies of an Aspergillus species (ascomycete), growing on an agar plate. Note the masses of filamentous cells (mycelia) and asexual spores that give the colonies a dusty, matted appearance. (b) Conidiophore and conidia of Aspergillus fumigatus (see Figure 18.20b). The conidiophore is about 300 μm long and the conidia here are about 3 μm wide. These cells were stained to improve contrast. Besides being a common saprophyte, Aspergillus can be pathogenic, causing serious pulmonary and occasional systemic infections in humans and some domestic animals. Cancer patients and others with weakened immune systems are particularly susceptible to aspergillosis.
Figure 18.22 Mushroom life cycle.

Mushrooms typically develop underground and then emerge on the surface rather suddenly (usually overnight), triggered by an influx of moisture. Photos of stages in formation of a common lawn mushroom (see also Section 18.14).
Most fungal cell walls consist of chitin, a polymer of N-acetylglucosamine. Chitin is arranged in the walls in microfibrillar bundles, as is cellulose in plant cell walls, to form a thick, tough wall structure. Other polysaccharides such as mannans and galactosans, or even cellulose itself, replace or supplement chitin in some fungal cell walls. Fungal cell walls are typically 80–90% polysaccharide, with only small amounts of proteins, lipids, polyphosphates, and inorganic ions making up the wall-cementing matrix.
Symbioses and Pathogenesis
Most plants are dependent on certain fungi to facilitate their uptake of minerals from soil. These fungi form symbiotic associations with the plant roots called mycorrhizae (the word means, literally, “fungus roots”). Mycorrhizal fungi establish close physical contact with the roots and help the plant obtain phosphate, as well as other minerals, and water from the soil. In return, the fungi obtain nutrients such as sugars from the plant root (Figure 23.22). There are two kinds of mycorrhizal associations. One, ectomycorrhizae, typically forms between basidiomycetes (Section 18.14) and the roots of woody plants, while the second, endomycorrhizae, forms between glomeromycete fungi (Section 18.12) and many nonwoody plants. Some fungi also form associations with cyanobacteria or green algae. These are the lichens, the colorful and crusty growths often seen on the surfaces of trees and rocks. We explore the biology of lichens and mycorrhizae in more detail in Sections 23.1 and 23.5, respectively.
Fungi can invade and cause disease in plants and animals. Fungal plant pathogens cause widespread crop and plant damage worldwide, and fruit and grain crops in particular suffer significant yearly losses as a result of fungal infection. Human fungal diseases, called mycoses, range from relatively minor and easily cured conditions, such as athlete’s foot and jock itch, to serious, life-threatening systemic mycoses, such as histoplasmosis.We consider some major fungal diseases of humans in Chapter 34.
Check Your Understanding
What are conidia? How does a conidium differ from a hypha? From a mycelium?
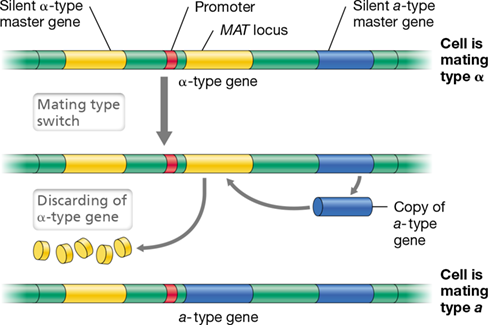
18.10 Fungal Reproduction and Phylogeny
Fungi reproduce by asexual means in one of three ways: (1) by the growth and spread of hyphal filaments; (2) by the asexual production of spores (conidia; Figures 18.20 and 18.21); or (3) by simple cell division, as in budding yeasts (Figure 18.23). Most fungi also form sexual spores, typically as part of an elaborate life cycle. Some fungi, such as the well-known mold Penicillium (the source of the antibiotic penicillin), were long thought to lack a sexual stage and reproduce only by way of conidia. But it has now been shown that the life cycle of Penicillium includes a sexual stage.
Figure 18.23 The common baker’s and brewer’s yeast *Saccharomyces cerevisiae* (*Ascomycota*).

In this colorized scanning electron micrograph, note the budding division and scars from previous buds. A single cell is about 6 μm in diameter.
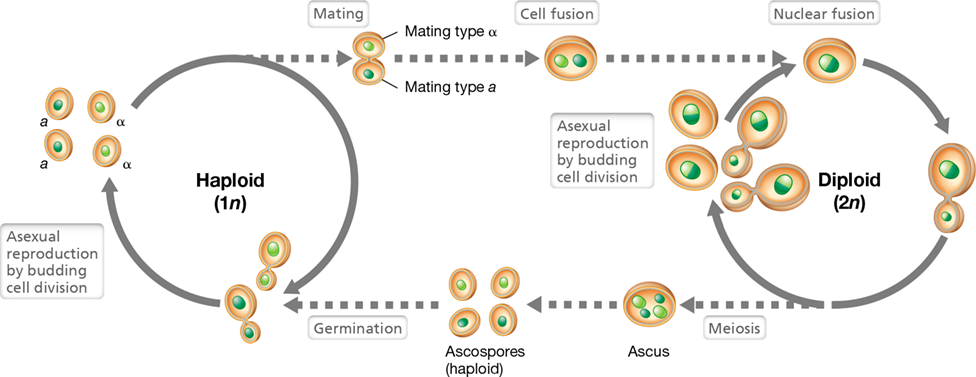
Sexual Spores of Fungi
Some fungi produce spores as a result of sexual reproduction. The spores develop from the fusion of either unicellular gametes or specialized hyphae called gametangia. Alternatively, sexual spores can originate from the fusion of two haploid cells to yield a diploid cell; this then undergoes meiosis and mitosis to yield individual haploid spores.
Depending on the group, different types of sexual spores are produced. Spores formed within an enclosed sac (ascus) are called ascospores. Many yeasts produce ascospores, and we consider sporulation in the common baker’s yeast Saccharomyces cerevisiae in Section 18.13. Sexual spores produced on the ends of a club-shaped structure (basidium) are basidiospores (Figure 18.22 and see Figure 18.31c). Zygospores, produced by zygomycetous fungi such as the common bread mold Rhizopus (Section 18.12), are macroscopically visible structures that result from the fusion of hyphae and genetic exchange. Eventually the zygospore matures and produces asexual spores that are dispersed by air and germinate to form new fungal mycelia. Chytrid fungi produce sexual spores, motile by eukaryotic flagella, called zoospores.
Sexual spores of fungi are typically resistant to drying, heating, freezing, and some chemical agents. However, neither sexual nor asexual spores of fungi are as resistant to heat as bacterial endospores (Section 2.8). Both asexual and sexual spores of fungi can germinate and develop into a new hypha.
The Phylogeny of *Fungi*
Fungi share an ancestor with animals and are more closely related to animals than any other eukaryotic group (Figure 18.3). The last common ancestor of all fungi likely existed sometime between 450 million and 1.5 billion years ago. One of the earliest fungal lineages is thought to be the Chytridiomycota, an unusual group of motile fungi that produce zoospores. Thus, the lack of flagella in most fungi indicates that motility is a characteristic that has been lost at various times in different fungal lineages.
Some of the major groups of fungi are shown in the evolutionary tree in Figure 18.24. The phylogeny shown in this figure includes several distinct fungal groups: the Microsporidia, Chytridiomycota, Mucoromycota, Glomeromycota, Ascomycota, and Basidiomycota (these groups are considered in detail in the next four sections). The vast majority of described fungal species belong to the Ascomycota and Basidiomycota. The Ascomycota are a large and diverse group of fungi that includes yeasts, such as Saccharomyces (Figures 18.23 and 18.24), and molds, such as Aspergillus (Figure 18.21). The Basidiomycota include fungi that form mushrooms (Figures 18.22, 18.24, and see Figure 18.31), as well as many important plant pathogens such as the rusts and smuts. While a tremendous diversity of fungal species have already been cultured and described, phylogenetic analyses of fungal DNA sequences recovered from environmental samples (Sections 19.6 and 19.8) indicate that more than 90% of fungal species remain uncultured. Clearly, we have much to learn about the biology and phylogeny of the fungi.
Figure 18.24 Phylogeny of fungi.

This phylogenetic tree depicts the relationships among the major groups (phyla) of fungi. A typical genus is listed for each group and depicted in the tree.
Check Your Understanding
Why is the mold Penicillium economically important?
What are the major differences between ascospores and conidia?
To what major group of macroorganisms are fungi most closely related?
Key Genera: *Allomyces, Batrachochytrium, Encephalitozoon*
The Microsporidia and Chytridiomycota are ancient phylogenetic groups of parasitic or saprophytic fungi (Figure 18.24). Microsporidia are obligate parasites of a wide variety of animal hosts including humans, whereas the Chytridiomycota are primarily aquatic fungi whose species are either parasites or saprophytes.
*Microsporidia Microsporidia* are tiny (2–5 μm) unicellular parasites of animals and protists. Based on 18S ribosomal RNA gene sequencing and their lack of mitochondria, *Microsporidia* were once thought to form a very early-branching lineage of *Eukarya*. However, composite gene and protein sequencing has shown the microsporidians are fungi closely related to the *Chytridiomycota* (Figure 18.24). There remains debate as to whether the *Microsporidia* should be included as one of the most deeply divergent lineages within the fungi or whether they should instead be classified as a distinct lineage closely related to fungi.
Microsporidia have adapted to a parasitic lifestyle. They exist as spores when outside of the host. When near a host cell they extend a helical polar tubule that penetrates the host cytoplasmic membrane. The spore then injects its sporoplasm into the host cell. The sporoplasm replicates within the cytoplasm of the host cell, forming new spores as it completes its life cycle. Eventually, the cell membrane of the host is disrupted and the spores are released into the surrounding environment, free to infect new cells.
Like most obligate parasites, microsporidia have undergone significant genome reduction, losing many features that would allow them to live outside of a host cell. The microsporidium Encephalitozoon (Figure 18.24 and **Figure 18.25*a***), for example, lacks not only mitochondria but also a Golgi complex (another key eukaryotic cell structure, Section 2.15 and Figure 2.42). Moreover, Encephalitozoon cells contain a very small genome of only 2.9 Mbp and only about 2000 genes (this is 1.5 Mbp and 2600 genes smaller than that of the bacterium Escherichia coli). The Encephalitozoon genome lacks genes for major metabolic pathways, such as the citric acid cycle (Section 3.6), meaning that this pathogen must depend on its host for even the most basic of metabolites and metabolic processes.
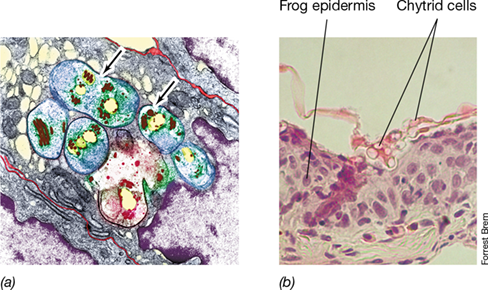
Figure 18.25 *Microsporidia* and *Chytridiomycota*.

(a) Colorized transmission electron micrograph of thin sections of cells of the microsporidium Encephalitozoon intestinalis (arrows) growing in human intestinal cells. (b) Cells of the chytrid Batrachochytrium dendrobatidis stained pink growing on the surface of frog epidermis.
In humans, Encephalitozoon causes chronic debilitating diseases of the intestine, lung, eye, muscle, and some internal organs but is uncommon among healthy adults with normal immune systems. However, microsporidial diseases have appeared with increasing frequency in immune-compromised individuals, such as those with HIV/AIDS or those on long-term administration of immune-suppressing drugs, such as those who have had organ transplants.
*Chytridiomycota Chytridiomycota*, or *chytrids*, are one of the earliest diverging lineage of fungi (Figure 18.24), and their name refers to the structure of the fruiting body, which contains their *zoospores* (Section 18.10). These spores are unusual among fungal spores in being flagellated and motile and are ideal for dispersal of these organisms in the aquatic environments, mostly freshwater and moist soils, where they are commonly found.
Many species of chytrids are known and some exist as single cells, whereas others form colonies with hyphae. They include both free-living forms that degrade organic material, such as Allomyces, and parasites of animals, plants, and protists. The chytrid Batrachochytrium dendrobatidis causes chytridiomycosis of frogs (Figure 18.25b), a condition in which the organism infects the frog’s epidermal layers, leading to a loss of ions across the membrane and osmotic imbalance. Chytrids have been implicated in the massive die-off of frogs and some other amphibians worldwide, probably in response to increases in global temperatures that have stimulated chytrid proliferation and to increased animal susceptibility due to habitat loss and aquatic pollution.
Unresolved aspects of the phylogeny of chytrids suggest that this group is not monophyletic. That is, some organisms currently classified as chytrids may actually be more closely related to species of other fungal groups, such as the Mucoromycota, to which we turn next. As is true for the protists, much about the evolution of the chytrids and other groups of fungi remains to be learned.
Check Your Understanding
What animal group has been most affected by chytrids?
What are some features of Microsporidia that distinguish them from chytrids?
Key Genera: *Rhizopus, Glomus*
We consider two groups of fungi here, the Mucoromycota, known primarily for their role in food spoilage, and the Glomeromycota, important fungi in certain mycorrhizal associations. Mucoromycota are commonly found in soil and on decaying plant material, whereas Glomeromycota form symbiotic relationships with plant roots. All of these fungi are coenocytic (multinucleate), and while Glomeromycota have an exclusively asexual life cycle, Mucoromycota often have a life cycle that includes sexual spores called zygospores (Section 18.10).
Mucoromycota
The common black bread mold Rhizopus nigricans (Figure 18.26) is a widespread zygomycete (zygomycetes are fungi that produce zygospores). This organism undergoes a complex life cycle that includes both asexual and sexual reproduction. In the asexual phase the mycelia form sporangia within which haploid spores are produced. Once released, these haploid spores disperse and eventually germinate, giving rise to vegetative mycelia. In the sexual phase, mycelial gametangia of different mating types (analogous to male and female, see Section 18.13) fuse to yield a cell with two nuclei. This diploid cell eventually develops into a zygospore, which can remain dormant and resist dryness and other unfavorable conditions. When conditions are favorable, the zygospore can germinate, forming either a diploid mycelium or a sporangium that produces haploid spores for dispersal.
Figure 18.26 *Mucoromycota*.

(a) Moldy bread from growth of the zygomycete Rhizopus nigricans. (b) Stained mycelium of Rhizopus showing the black aerial sporangia containing asexual spores.
Most species of Rhizopus and related zygomycetes are harmless saprophytes whose airborne spores land and form spreading colonies on stale bread (Figure 18.26a) and various moist surfaces in the home or on walls and crevices in buildings where moisture is trapped. However, some species are human pathogens. If inhaled in sufficient amounts, spores of pathogenic Rhizopus species can cause serious infections of the lungs, sinuses, eyes, nose, and mouth leading to swelling of facial features, asthma-like symptoms, and even fatal systemic fungal infections if the initial infection is not promptly treated.
Glomeromycota
The Glomeromycota are a relatively small and unique group of obligately symbiotic fungi in which all known species form associations with plants called endomycorrhizae (Section 18.9). As many as 80% or more of land plant species form these associations in which the fungal hyphae enter the plant cell and aid the plant’s acquisition of phosphate from the soil in return for fixed carbon from the plant. Most of these Glomeromycota are arbuscular mycorrhizae, fungi that form structures called arbuscules that penetrate cells of their plant host and specialize in nutrient exchange (Section 23.5). As plant symbionts, glomeromycetes are thought to have played a pivotal role in the ability of early vascular plants to colonize land.
As far as is known, glomeromycetes reproduce only asexually and are mostly coenocytic in their hyphal morphology. Asexual spores of Glomus (Figure 18.24), a major genus of endomycorrhizae, are collected from the roots of cultivated plants and used as an agricultural inoculant to promote vigorous symbiotic associations between plant and fungus. This natural approach to plant fertilization is a widespread practice in small, sustainable farming operations and has been shown to increase both the growth and nutrient content of tomato, pepper, squash, bean, and several other small fruit and vegetable plants.
Check Your Understanding
Contrast the habitats of Mucoromycota and Glomeromycota.
How does the fungus Glomus aid the acquisition of nutrients by plants?
Key Genera: *Saccharomyces, Candida, Aspergillus*
The Ascomycota, also called the ascomycetes, are the largest and most diverse group of fungi and they range from single-celled species, such as the baker’s yeast Saccharomyces (Figure 18.27 and Figure 18.23), to species that grow as filaments, such as the common mold Aspergillus (Figure 18.21). Ascomycetes are found in aquatic and terrestrial environments and take their name from the production of asci (singular, ascus), cells in which two haploid nuclei from different mating types fuse to form a diploid nucleus that eventually undergoes meiosis to form haploid ascospores. In addition to ascospores, ascomycetes reproduce asexually by the production of conidia that form at the tips of specialized hyphae called conidiophores (Figure 18.21). Both saprophytic and pathogenic yeasts, such as Candida albicans, are common in nature. We focus here on the yeast Saccharomyces as a model ascomycete.
Figure 18.27 Growth by budding division in *Saccharomyces cerevisiae*.

A time-lapse series of phase-contrast micrographs shows the budding division process starting from a single cell. Note the pronounced nucleus. A single cell of S. cerevisiae is about 6 μm in diameter.
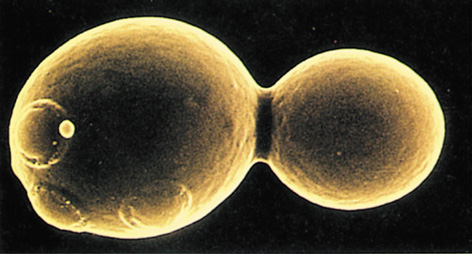
*Saccharomyces cerevisiae*
The cells of Saccharomyces and other single-celled ascomycetes are spherical, oval, or cylindrical, and cell division typically takes place by budding. In the budding process, a new cell forms as a small outgrowth of the old cell; the bud gradually enlarges and then separates from the parent cell (Figures 18.23 and 18.27).
Yeast cells are typically much larger than bacterial cells and can be distinguished from bacteria microscopically by their larger size and by the obvious presence of internal cell structures, such as the nucleus or cytoplasmic vacuoles (Figure 18.27). Yeasts flourish in sugar-rich habitats such as fruits, flowers, and the bark of trees. Yeasts are typically facultative aerobes, growing aerobically as well as by fermentation. Several yeasts live symbiotically with animals, especially insects, and a few species are pathogenic for animals including humans (Section 34.1 and Figure 34.1a,c). The most important commercial yeasts are the baker’s and brewer’s yeasts, which are species of Saccharomyces. The yeast S. cerevisiae has been studied as a model eukaryote for many years and was the first eukaryote to have its genome completely sequenced (Section 10.4).
Mastering Microbiology
Art Activity: Figure 18.27 Life cycle of a typical ascomycete yeast, Saccharomyces cerevisiae
Mating Types and Sexual Reproduction in *Saccharomyces*
The yeast Saccharomyces can reproduce by sexual means in which two cells fuse. Within the fused cell, called a zygote, meiosis occurs and ascospores are eventually formed. The life cycle of S. cerevisiae is depicted in Figure 18.28. S. cerevisiae can grow vegetatively (nonsexually) as either haploid or diploid cells. S. cerevisiae forms two different types of haploid cells called mating types designated α (alpha) and a (encoded by genes α and a); these are analogous to male and female gametes. The genes α and a regulate the production of the peptide hormones α factor and a factor, respectively, which are excreted by yeast cells during mating. The hormones bind to cells of the opposite mating type and bring about changes in their cell surfaces that enable the cells to fuse; once mating has occurred, the nuclei fuse, forming a diploid zygote (Figure 18.29). The zygote undergoes vegetative growth by budding, but under starvation conditions it undergoes meiosis and generates ascospores (Figure 18.28).
Figure 18.28 Life cycle of a typical ascomycete yeast, *Saccharomyces cerevisiae*.

Vegetative growth can occur for long periods as haploid cells or as diploid cells before life cycle events (dashed lines) generate the alternate genetic form.
Figure 18.29 Electron micrographs of mating in the ascomycete yeast *Hansenula wingei*.

(a) Two cells have fused at the point of contact. (b) Late stage of mating. The nuclei of the two cells have fused, and a diploid bud has formed at a right angle to the mating cells. This bud becomes the progenitor of a diploid cell line. A cell of Hansenula is about 10 μm in diameter.
Haploid strains of S. cerevisiae are genetically predisposed to be either a or α but are able to switch their mating type. This switch occurs when the active mating-type gene is replaced with one of two otherwise “silent” genes, as shown in Figure 18.30. There is a single location on one of the S. cerevisiae chromosomes called the MAT (for mating type) locus, at which either gene a or gene α can be inserted. At this locus, the MAT promoter controls transcription of whichever gene is present. If gene a is at that locus, then the cell is mating-type a, whereas if gene α is at that locus, the cell is mating-type α. Elsewhere in the yeast genome are copies of genes a and α that are not expressed, and these are the source of the inserted gene. In the switch (Figure 18.30), the appropriate gene, a or α, is copied from its silent site and inserted into the MAT location, replacing the gene already present. The old mating-type gene is excised and discarded, and the new gene is inserted.
Figure 18.30 The cassette mechanism that switches an ascomycete yeast from mating type α to *a*.

The cassette inserted at the MAT locus determines the mating type. The process shown is reversible, so type a can also revert to type α.
Whichever gene is inserted in the MAT locus is the one that will govern the mating type of the strain. It is thus possible for cells from a pure culture of S. cerevisiae derived from a single cell to mate, following a mating-type switch in one or more cells in the culture.
Check Your Understanding
Explain how a single haploid cell of Saccharomyces can eventually yield a diploid cell.
Figure 18.31 Mushrooms.

(a) Amanita, a highly poisonous mushroom. (b) Gills on the underside of the mushroom fruiting body contain the spore-bearing basidia. (c) Light micrograph of basidia and basidiospores from the mushroom Coprinus.
Mushroom Development
During most of its existence, a mushroom fungus lives as a simple haploid mycelium, growing vegetatively in soil, leaf litter, or decaying logs. It is the sexual reproductive phase of basidiomycetes that produces the visible mushroom structure (Figures 18.22 and 18.31). In this process, mycelia of different mating types fuse, and the faster growth of the dikaryotic (two nuclei per cell) mycelium formed from that fusion overgrows and crowds out the parental haploid mycelia. Then, when environmental conditions are favorable, usually following periods of wet and cool weather, the dikaryotic mycelium develops rapidly into the fruiting body.
The mushroom fruiting body, called a basidiocarp, begins as a mycelium that differentiates into a small button-shaped structure underground that then expands into the full-grown basidiocarp that we see aboveground, the mushroom (Figures 18.22 and 18.31a, b). The dikaryotic basidia are borne on the underside of the basidiocarp on flat plates called gills, which are attached to the cap of the mushroom (Figure 18.31b, c). The basidia then undergo a fusion of the two nuclei, forming basidia with diploid nuclei. The two rounds of meiotic division generate four haploid nuclei in the basidia, and each of the nuclei becomes a basidiospore. The genetically distinct basidiospores can then be dispersed by wind to new habitats to begin the cycle again, germinating under favorable conditions and growing as haploid mycelia (Figure 18.22).
Pathogenic Basidiomycetes
The smuts and rusts are plant pathogenic basidiomycetes. Smuts are pathogens of cereal grains and other crops, and the genus Ustilago contains several species whose hosts include corn, sugarcane, wheat, and several other grains. The fungus targets the reproductive system of the plant, and in corn, smut causes the developing kernels to form massive tumorlike kernels that give the cob a swollen and burned appearance.
Rust fungi attack rapidly growing plant tissues such as emerging shoots, leaves, and fruits. Rust fungi of the genus Puccinia can infect any of a wide variety of plants whereas other rust fungi are more host-restricted. Some common rusts include stem rust of wheat and white pine blister rust, a disease of several pine species that can trigger branch dieback and even death of the entire tree. Pine trees weakened by rust fungi are typically more susceptible to insect attack such as from infestations of the pine bark beetle, a highly destructive pest that has destroyed major stands of various pine species in the western United States and central Europe in recent years. Hence, even if the rust fungus itself is not lethal, the damage it does to the overall vigor of the tree can set the stage for lethal attack by a secondary biological agent or even by strong winds.
We now move on to the final part of this chapter where we will explore the world of algae, phototrophic eukaryotes widespread in nature.
Check Your Understanding
What are some distinguishing features of basidiospores and of zoospores (Section 18.10)?
IV Archaeplastida
**The Archaeplastida are a diverse phototrophic lineage that includes plants as well as several groups of algae. Species of red and green algae can be microscopic or macroscopic and are found worldwide, primarily in aquatic habitats.**
We conclude our tour of eukaryotic microbial diversity with the kingdom Archaeplastida, a large group that includes both red and green algae as well as land plants. As we have previously discussed, only the red and green algae originated from primary endosymbiotic events, whereas other protists containing chloroplasts were the result of secondary endosymbioses (Figures 18.2 and 18.3). Here we focus on the red and green algae, a large and diverse group of eukaryotic organisms that contain chlorophylls and carry out oxygenic photosynthesis (Section 14.6).
Key Genera: *Polysiphonia, Cyanidium, Galdiera*
The red algae, also called rhodophytes, mainly inhabit the marine environment, but a few species are found in freshwater and terrestrial habitats. Both unicellular and multicellular species are known, and some of the latter are macroscopic.
Major Properties of Red Algae
Red algae are phototrophic and contain chlorophyll a; their chloroplasts lack chlorophyll b but contain phycobiliproteins, the major light-harvesting pigments of the cyanobacteria (Section 14.4). The reddish color of many red algae (Figure 18.32) results from phycoerythrin, an accessory pigment that masks the green color of chlorophyll. This pigment is present along with phycocyanin and allophycocyanin in structures called phycobilisomes, the light-harvesting (antenna) components of cyanobacteria. At greater depths in aquatic habitats, where less light penetrates, cells compensate by producing more phycoerythrin and are a darker red, whereas shallow-dwelling species often have less phycoerythrin and can be green in color (see Figure 18.33).
Figure 18.32 *Polysiphonia*, a filamentous marine red alga.

Light micrograph. Polysiphonia grows attached to the surfaces of marine plants. Cells are about 150 μm wide.
Most species of red algae inhabit marine environments and are multicellular and lack flagella. Some are “seaweeds” and are the source of agar, the solidifying agent used in bacteriological culture media, and carrageenans, thickening and stabilizing agents used in the food industry. Other species of red algae, such as the genus Porphyra, are harvested, dried, and used as a wrap in sushi. Different species of red algae are filamentous, leafy, or, if they deposit calcium carbonate, coralline (coral-like) in morphology. Coralline red algae play an important role in the development of coral reefs and help strengthen reefs against wave damage (Section 23.13).
Polysiphonia (Figure 18.32) is a genus of filamentous and branched red algae found worldwide in marine environments. The organism is found primarily near shore where the cells grow attached to rocks, other algae, and man-made surfaces such as jetties, concrete abutments, retaining walls, and docks. Nearly 200 species of this genus are recognized, and the organism undergoes a complex life cycle in which an alternation of generations occurs. In this cycle, haploid male and female gametes released from a diploid multicellular organism develop into haploid male and female multicellular organisms. The male alga then releases haploid “sperm” cells that fuse with a specialized reproductive structure on the female alga to yield a diploid zygote; the latter develops a multicellular form and, following meiosis, releases male and female gametes to complete the life cycle.
*Cyanidium, Galdieria*, and Relatives
In addition to multicellular red algae like Polysiphonia (Figure 18.32), unicellular species are also known. One such group, members of the Cyanidiales that include the genera Cyanidium, Cyanidioschyzon, and Galdieria (Figure 18.33), live in hot, acidic and metal-rich hot springs at temperatures from 30 to 60 °C and acidic pH (0.5 to 4.0); under these extreme conditions, no other phototrophic microorganisms (including anoxygenic phototrophs) can exist. The unicellular red algae are unusual in other ways as well. For example, cells of Cyanidioschyzon merolae are unusually small (1–2 μm in diameter) for eukaryotes, and the genome of this species, approximately 16.5 Mbp, is one of the smallest genomes known for a phototrophic eukaryote.
Figure 18.33 *Galdieria*, a unicellular red alga.

(a) Galdieria inhabits acidic and sulfidic hot springs where it often grows attached to mineral debris as shown here (arrow). (b) Cells of Galdieria are about 25 μm in diameter and are more blue-green than red in color because the phototroph contains mainly phycocyanin rather than phycoerythrin as its phycobilin (Section 14.4).
Molecular analyses of the Galdieria genome have revealed the surprising result that this eukaryotic phototroph contains at least 75 genes acquired by horizontal transfer from various prokaryotic sources. Some of the key genes transferred encode protections against salt stress and metal toxicity and the construction of a toughened cytoplasmic membrane to withstand the heat and acidity of the Galdieria habitat (Figure 18.33a). The ability of Galdieria to grow in darkness—an unusual feature for an alga—has also been linked to horizontally transferred genes, in particular to genes encoding transport systems for various organic compounds. Although these genetic transfers are unusual in the sense that donor and recipient belong to different phylogenetic domains, they once again underscore the importance of horizontal gene transfer in molding microbial genomes (Section 13.9).
Check Your Understanding
What physiological properties would be necessary for Galdieria to live in its habitat?
Key Genera: *Chlamydomonas, Volvox*
The green algae, also called chlorophytes, have chloroplasts containing chlorophylls a and b, which give them their characteristic green color, but they lack phycobiliproteins and so do not develop the red or blue-green colors of red algae (Figures 18.32 and 18.33). In the composition of their photosynthetic pigments, green algae are similar to plants and are closely related to plants phylogenetically. There are two main groups of green algae, the chlorophytes, examples of which are the microscopic Chlamydomonas and Dunaliella (**Figure 18.34*a***), and the charophyceans such as Chara (Figure 18.34b), macroscopic organisms that often resemble land plants and are actually most closely related to land plants.
Figure 18.34 Green algae.

(a) A single-celled, flagellated green alga, Dunaliella. A cell is about 5 μm wide. (b) The plantlike green alga Chara. (c) Micrasterias. This single multilobed cell is about 100 μm wide. (d) Scenedesmus, showing packets of four cells each. (e) Spirogyra, a filamentous alga with cells about 20 μm wide. Note the green spiral-shaped chloroplasts. (f) Volvox carteri colony with eight daughter colonies. (g) The petroleum-producing green alga Botryococcus braunii. Note the excreted oil droplets surrounding the cell.
Most green algae inhabit freshwater while others are found in moist soil or growing in snow, to which they impart a pink color (Figure 4.23). Other green algae live with fungi in a symbiotic relationship to form lichens (Section 23.1). The morphology of chlorophytes ranges from unicellular (Figure 18.34a, c), to filamentous (with individual cells arranged end to end, Figure 18.34e), to colonial (as aggregates of cells, Figure 18.34f). Even multicellular species exist, an example of which is the seaweed Ulva. Most green algae have a complex life cycle, with both sexual and asexual reproductive stages.
Very Small Green Algae and Colonial Green Algae
One of the smallest eukaryotes known is the green alga Ostreococcus tauri, a common unicellular species of marine phytoplankton (Section 20.11 and Figure 20.27b). Cells of O. tauri have a diameter of approximately 2 μm, and the organism contains the smallest genome of any known phototrophic eukaryote, approximately 12.6 Mbp. Ostreococcus has thus provided a model organism for research into the evolution of genome reduction and specialization in eukaryotes.
At the colonial level of organization in green algae is Volvox (Figure 18.34f). This alga forms colonies composed of several hundred flagellated cells, some of which are motile and primarily carry out photosynthesis, while others specialize in reproduction. Cells in a Volvox colony are interconnected by thin strands of cytoplasm that allow the entire colony to swim in a coordinated fashion. Volvox has been a long-term model for research on the genetic mechanisms controlling multicellularity and the distribution of functions among cells in multicellular organisms.
Some colonial green algae have potential as sources of biofuels. For example, the colonial green alga Botryococcus braunii excretes long-chain (C30−C36) hydrocarbons that have the consistency of crude oil (Figure 18.34g). About 30% of the B. braunii cell dry weight consists of this petroleum, and there has been heightened interest in using this and other oil-producing algae as renewable sources of petroleum. Evidence from biomarker studies has shown that some known petroleum reserves originated from green algae such as B. braunii that settled in lake beds in ancient times. Hence, if the scale-up challenges for commercial algal petroleum production could ever be met, it is possible that some fraction of the world’s oil supply could someday come from photosynthesis by green algae.
Endolithic Phototrophs
Some green algae grow inside rocks. These endolithic (endo means “inside”) phototrophs inhabit porous rocks, such as those containing quartz, and are typically found in layers near the rock surface (**Figure 18.35*a***). Endolithic phototrophic communities are most common in dry environments such as deserts or cold, dry environments such as Antarctica. For example, in the McMurdo Dry Valleys of Antarctica, where temperatures and humidity are extremely low (Figure 4.22d, e), life within a rock has its advantages. Rocks in these harsh environments are heated by the sun, and water from snowmelt can be absorbed and retained for relatively long periods, supplying moisture needed for growth. Moreover, water absorbed by a porous rock makes the rock more transparent, thus funneling more light to the algal layers.
Figure 18.35 Endolithic phototrophs.

(a) Photograph of a limestone rock from the McMurdo Dry Valleys region of Antarctica broken open to show the layer of endolithic green algae. (b) Light micrograph of cells of the green alga Trebouxia, a widespread endolithic alga in Antarctica.
A wide variety of phototrophs can form endolithic communities, including cyanobacteria and various green algae (Figure 18.35b). In addition to being free-living phototrophs, green algae and cyanobacteria coexist with fungi in endolithic lichen communities (Section 23.1 for discussion of the lichen symbiosis). Metabolism and growth of these internal rock microbial communities slowly weathers the rock, allowing gaps to develop where water can enter, freeze and thaw, and eventually crack the rock, producing new habitats for microbial colonization. The decomposing rock also forms a crude soil that can support development of plant and animal communities in environments where conditions (temperature, moisture, and so on) permit.
Check Your Understanding
What phototrophic properties link green algae and plants?
What is unusual about the green algae Ostreococcus, Volvox, and Botryococcus?
Chapter Review
Go to Mastering Microbiology for videos, animations, practice tests, and more.
I Organelles and Phylogeny of Microbial *Eukarya*
18.1 Key metabolic organelles of eukaryotes are the chloroplast, which functions in photosynthesis, and the mitochondrion, which functions in respiration. These organelles were originally Bacteria that established permanent residence inside other cells (endosymbiosis).
Q Distinguish between a primary and a secondary endosymbiosis. Which groups of protists are derived from which form of endosymbiosis?
18.2 The modern phylogeny of eukaryotes suggests a major radiation of eukaryotic diversity emerging at some time following symbiotic events that led to the mitochondrion. The latter provided eukaryotic cells with a chemotrophic energy-generating powerhouse that is present today in nearly all eukaryotic cells.
**Q What are the five major supergroups within the Eukarya?**
II Protists
18.3 Diplomonads such as Giardia are unicellular, flagellated, nonphototrophic protists. Parabasalids such as Trichomonas are human pathogens and contain huge genomes that lack introns. Euglenids and kinetoplastids are unicellular, flagellated protists. Some are phototrophic. This group includes some important human pathogens, such as Trypanosoma, and some well-studied nonpathogens, such as Euglena.
Q What are hydrogenosomes and what is their function?
18.4 Three groups make up the Alveolata: ciliates, dinoflagellates, and apicomplexans. Most ciliates and dinoflagellates are free-living organisms, whereas apicomplexans are obligate parasites of animals.
Q What organism causes “red tides,” and why is this organism toxic?
18.5 Stramenopiles are protists that bear a flagellum with fine, hairlike extensions. They include oomycetes, diatoms, and brown and golden algae.
Q In terms of their photosynthetic pigments, how are brown and golden algae similar?
18.6 The Rhizaria include diverse protists such as the phototrophic chlorarachniophytes and foraminiferans, as well as radiolarians, which are chemoorganotrophs.
**Q What morphological trait unites the Rhizaria and distinguishes them from other protists?**
18.7 Haptophytes include the coccolithophores, unicellular phototrophic microbes that form intricate calcite exoskeletons. Coccolithophores such as Emiliania huxleyi are abundant in the oceans and important to global climate because they remove carbon from the atmosphere and deposit it as calcite in deep ocean sediments.
**Q Describe the life cycle of E. huxleyi and explain why this life cycle can make it difficult to identify species of coccolithophores in the environment.**
18.8 Amoebozoa are protists that use pseudopodia for movement and feeding. Within Amoebozoa are gymnamoebas, entamoebas, and slime molds. Plasmodial slime molds form masses of motile protoplasm, whereas cellular slime molds are individual cells that aggregate to form fruiting bodies from which spores are released.
**Q Explain the major differences between the slime molds Dictyostelium and Physarum.**
III Fungi
18.9 Fungi include the molds, mushrooms, and yeasts. Other than phylogeny, fungi primarily differ from protists by their rigid chitin-containing cell wall, production of spores, and the fact that most fungal species are nonmotile.
Q What are the major differences between mushrooms, molds, and yeasts?
18.10 A variety of sexual spores are produced by fungi, including ascospores, basidiospores, and zygospores. From a phylogenetic standpoint, fungi are the closest relatives of animals.
Q List the different types of sexual spores of fungi. Are conidia sexual or asexual spores?
18.11 Chytridiomycota and Microsporidia are basal to all other known fungal groups in the fungal phylogeny. Some chytrids are amphibian pathogens, while microsporidians are parasites of a variety of animals.
Q In what way do chytrids and microsporidians differ from other fungi?
18.12 Mucoromycota form coenocytic hyphae and undergo both asexual and sexual reproduction, and the common bread mold Rhizopus is a good example. Glomeromycota are fungi that form endomycorrhizal associations with plants.
Q What is the major feature of the ecology of glomeromycetes?
18.13 The Ascomycota are a large and diverse group of mostly saprophytic fungi. Some, such as Candida albicans, can be pathogenic in humans. There are two mating types in the yeast Saccharomyces cerevisiae, and yeast cells can convert from one type to the other by a genetic switch mechanism.
Q How is the mating type of a yeast cell determined?
18.14 Basidiomycota include the mushrooms, puffballs, smuts, and rusts. Basidiomycetes undergo both vegetative (asexual) reproduction as haploid mycelia and sexual reproduction via fusion of mating types and formation of haploid basidiospores.
**Q What morphological feature unites the Basidiomycota, and where is this feature found?**
IV Archaeplastida
18.15 Red algae are mostly marine and range from unicellular to multicellular. Their reddish color is due to the pigment phycoerythrin, a key cyanobacterial pigment, present in their chloroplast.
Q In what kinds of habitats would one likely find red algae?
18.16 Green algae are common in aquatic environments and can be unicellular, filamentous, colonial, or multicellular. A unicellular green alga, Ostreococcus, has the smallest genome known for a phototrophic eukaryote, while the green alga Volvox is a model colonial phototroph.
Q What traits link green algae and plants? What traits link green algae and red algae?
Application Questions
Explain the differences between mitochondria, chloroplasts, hydrogenosomes, and mitosomes. Explain the evolutionary process that gave rise to hydrogenosomes and mitosomes. Explain why the chloroplasts of red and green algae have two membranes, while those of many other photosynthetic protists have four membranes.
Considering all of the groups of microbes covered in this chapter, which of them seem most similar to prokaryotic cells in cell structure? Discuss at least two lines of evidence that show that these microbes are not prokaryotic cells.
Chapter Glossary
an informal term for phototrophic eukaryotes other than plants; algae are polyphyletic, being found in diverse groups of Eukarya as a result of secondary endosymbiosis Chitin
a polymer of N-acetylglucosamine commonly found in the cell walls of fungi Ciliate
any protist characterized in part by rapid motility driven by numerous short appendages called cilia Coenocytic
the presence of multiple nuclei in fungal hyphae without septa Colonial
the growth form of certain protists and green algae in which several cells live together and cooperate for feeding, motility, or reproduction; an early form of multicellularity Conidia
nonphototrophic eukaryotic microorganisms with rigid cell walls made of chitin Hydrogenosomes
organelles that produce ATP through fermentation reactions resulting in the production of H2 and acetate Mushroom
the aboveground fruiting body, or basidiocarp, of basidiomycete fungi Nucleomorphs
remnant nuclei found in certain algae; derived from an ancestral algal endosymbiont and associated with chloroplasts acquired by secondary endosymbiosis Phagocytosis
a mechanism for ingesting particulate material in which a portion of the cytoplasmic membrane surrounds the particle and brings it into the cell; for phagocytes of the immune system, the process of engulfing and killing foreign particles and cells Primary endosymbiosis
the endosymbiotic acquisition by a eukaryotic host cell of a respiring bacterium or a phototrophic cyanobacterium, followed by evolution of the endosymbiont into mitochondria or chloroplasts Protist
an informal term used to describe eukaryotic microorganisms that are not plants, animals, or fungi; can be heterotrophic or phototrophic, unicellular or multicellular Secondary endosymbiosis
the endosymbiotic acquisition by a mitochondrion-containing eukaryotic cell of a red or green algal cell, which itself contains a chloroplast derived from primary endosymbiosis Slime mold
a nonphototrophic protist that lacks cell walls and that aggregates to form fruiting structures (cellular slime molds) or masses of protoplasm (acellular slime molds) Yeast
UNIT 5 Microbial Ecology and Environmental Microbiology
Unit 5 Microbial Ecology and Environmental Microbiology
Unit 5 Microbial Ecology and Environmental Microbiology
Chapter 19 Taking the Measure of Microbial Systems
Chapter 23 Microbial Symbioses with Microbes, Plants, and Animals