Living on Fumes
Antarctica is one of the harshest environments on Earth, experiencing extremely cold temperatures, high levels of ultraviolet radiation, and limited availability of carbon, nitrogen, and water. Even so, rocky desert Antarctic soils (photo) support microbial communities whose diversity rivals that of temperate soils. Cyanobacteria fix enough carbon dioxide (CO2) to support heterotrophic communities in many soils of Antarctica. However, the virtual absence of cyanobacteria in some other Antarctic soils raises the question of what energy source (other than light) sustains these microbial communities.

Researchers combined metagenomics with biochemical measurements to probe this mystery. The first hint was the discovery of Calvin cycle genes for CO2 fixation in Actinobacteria that composed about half of the total soil microbiota (the Calvin cycle is a major autotrophic pathway). Since these Actinobacteria are not phototrophic, CO2 fixation—which requires ATP—had to be supported by energy sources other than sunlight. The answer again emerged from metagenomics by identifying genes in the Actinobacteria encoding aerobic respiration of molecular hydrogen (H2) and carbon monoxide (CO) and then showing that these genes were actively transcribed at below-freezing temperatures. Additionally, comparison of these gene sequences to those of cultured close relatives suggested these enzymes could scavenge H2 and CO at the parts per billion concentrations typically found in Earth’s lower atmosphere. This was subsequently confirmed by direct measurements of activity where it was shown that atmospheric concentrations of H2 stimulated rates of CO2 fixation sufficient to support about 107 bacteria per gram of Antarctic soil.
Because low concentrations of H2 and CO are constantly available, they are dependable energy sources for the slow-growing soil microbial communities in Antarctica. Whereas most ecosystems are sustained by solar or geologically derived energy, it appears that bacteria in some Antarctic surface soils are supported by atmospheric trace gases. In other words, these microbes are “living on fumes”!
Source: Mukan Ji, Chris Greening, Inka Vanwonterghem, et al. 2017. Atmospheric trace gases support primary production in Antarctic desert surface soil. Nature 552: 400.
Microorganisms do not live alone in nature but instead interact with other organisms and with their environment. In so doing, microorganisms carry out many essential activities that support all life on Earth. In this chapter we explore some of the major habitats of microorganisms; these include soil, freshwater, and the oceans. In addition to these, microbes have also established more specific, and often very intimate, associations with plants and animals. We examine a few examples of such microbial partnerships and symbioses in Chapters 23 and 24.
I Microbial Ecology
Microbial ecology borrows many concepts from classical ecology but differs in the enormous diversity of microbial habitats and metabolisms it must consider—habitats and metabolisms that collectively drive the major nutrient cycles in nature and provide life support to all macroorganisms.
We begin with a broad overview of the science of microbial ecology, including ways that organisms interact with each other and their environments and the difference between species diversity and species abundance. These basic ecological concepts pervade this and the next three chapters.
20.1 General Ecological Concepts
The distribution of microorganisms in nature resembles that of macroorganisms in the sense that a given species resides in certain places but not others; that is, everything is not everywhere. Also, environments differ in their abilities to support diverse microbial populations, from the highly diverse microbial world of undisturbed fertile soil to the rather restricted world of some highly extreme environments.
Ecosystems and Habitats
An ecosystem is a dynamic complex of plant, animal, and microbial communities and their abiotic surroundings, all of which interact as a functional unit. An ecosystem contains many different habitats, parts of the ecosystem best suited to one or a few populations. Although microorganisms are present in any habitat containing plants and animals, many microbial habitats are unsuitable for plants and animals. For example, microorganisms are ubiquitous on Earth’s surface and even deep within it; they inhabit boiling hot springs and solid ice, acidic environments near pH 0, saturated brines, environments contaminated with radionuclides and heavy metals, and the interior of porous rocks that contain only traces of water. Therefore, some ecosystems are mostly or even exclusively microbial.

Collectively, microorganisms show great metabolic diversity and are the primary catalysts of nutrient cycles in nature (Chapter 21). The types of microbial activities possible in an ecosystem are a function of the species present, their population sizes, and the physiological state of the microorganisms in each habitat. By contrast, the rates of microbial activities in an ecosystem are controlled by the nutrients and growth conditions that prevail. Depending on several factors, microbial activities in an ecosystem can have minimal or profound impacts and can diminish or enhance the activities of both the microorganisms themselves and the macroorganisms that may coexist with them.
Species Diversity in Microbial Habitats
A group of microorganisms of the same species that reside in the same place at the same time constitutes a microbial population and may be descendants of a single cell. A microbial population differs from a microbial community. A community consists of populations living in association with other populations. The microbial species that reside in a certain habitat are those best able to grow with the nutrients and conditions that prevail there, and the number of different populations present is a measure of the community’s complexity.

The diversity of microbial species in a community can be expressed in two ways: richness and abundance (Figure 20.1). Species richness is an expression of the total number of different species present. Identifying cells is, of course, basic to determining microbial species richness, but this need not require their isolation and culture. Species richness may also be expressed in molecular terms by the diversity of phylotypes (for example, the diversity of ribosomal RNA genes, Section 19.6) observed in a given community. Species abundance, by contrast, is the proportion of each species in the community (compare Figure 20.1b and c). Species richness and abundance can change quickly over a short time as shown by the change in abundance of cyanobacteria in a lake receiving nutrient-rich agricultural runoff (Figure 20.1a). One goal of microbial ecology is to understand what factors control species richness and abundance in microbial communities along with the community’s associated activities and the abiotic environment. Once all of these factors are known, microbial ecologists can model the ecosystem by perturbing it in some way and observing whether predicted changes match experimental results.
Figure 20.1 Microbial species diversity: Richness versus abundance.

(a) Collecting samples from Lake Taihu, China, following a bloom of the cyanobacterium Microcystis. (b) High species richness in St. John’s River, Florida, shown by microscopy of planktonic microorganisms including cyanobacteria, diatoms, green algae, flagellates, and bacteria. (c) Shift of St. John’s River community to low richness but high abundance following a bloom of the cyanobacterium Microcystis.
The microbial species richness and abundance of a community are functions of the conditions that prevail and the kinds and amounts of nutrients available in the habitat. Table 20.1 lists common nutrients and conditions relevant to microbial growth. In some microbial habitats, such as undisturbed organic-rich soils, high species richness is common (see Figure 20.14), with most species present at only moderate abundance. Nutrients in such a habitat are of many different types, and this helps select for high species richness. In other habitats, such as some extreme environments, species richness is often very low and abundance of one or a few species very high. This is because the physical and chemical conditions in the environment exclude all but a handful of species, and key nutrients are present at such high levels (Figure 20.1a) that the highly adapted species can grow to high cell densities. Bacteria that catalyze acid mine runoff from the oxidation of iron are an example of this. These organisms thrive in highly acidic, iron-rich but organic-poor waters, where the acidic conditions and the dearth of organic carbon limit species richness. However, the elevated levels of ferrous iron (Fe2+) present, which is oxidized to Fe3+ in energy-yielding reactions (Section 14.8), fuel high species abundance. We examine the activities of acidophilic iron-oxidizing microorganisms in Sections 21.6, 21.1, and 22.2. Although the microbial diversity of extreme environments may at first seem mundane, extreme environments have been well-studied by microbial ecologists because the limited community complexity makes it easier to dissect the ecological interactions of the microbes that inhabit them.
Table 20.1 Resources and conditions that govern microbial growth in nature

Check Your Understanding
What is the difference between species richness and species abundance?
What are the characteristics of a microbial population?
How does a microbial population differ from a microbial community?
20.2 Ecosystem Service: Biogeochemistry and Nutrient Cycles
20.2 Ecosystem Service: Biogeochemistry and Nutrient Cycles
20.2 Ecosystem Service: Biogeochemistry and Nutrient Cycles
In any ecosystem whose resources and growth conditions are suitable, microorganisms will grow to form populations. Metabolically similar microbial populations that exploit the same resources in a similar way are called guilds. A habitat that is shared by a guild and supplies the resources and conditions the cells require for growth is called a niche. Sets of guilds form microbial communities (Figure 20.2). Microbial communities interact with macroorganisms and abiotic factors in the ecosystem in a way that defines the workings of that ecosystem.
Figure 20.2 Populations, guilds, and communities.

Microbial communities consist of populations of cells of different species. A freshwater lake ecosystem would likely have the communities shown here. The reduction of NO3 −, Fe3+, SO4 2−, S0, and CO2 are examples of anaerobic respirations. The region of greatest activity for each of the different respiratory processes would differ with depth in the sediment. As more energetically favorable electron acceptors are depleted by microbial activity near the surface, less favorable reactions occur deeper in the sediment. All guild metabolisms shown are described in Chapter 14.
Energy Inputs to the Ecosystem
Energy enters ecosystems as sunlight, organic carbon, and reduced inorganic substances. Light is used by phototrophs to make ATP and synthesize new organic matter (Figure 20.2). In addition to carbon (C), new organic matter contains nitrogen (N), sulfur (S), phosphorus (P), iron (Fe), and the other elements of life (Section 3.1). This newly synthesized organic material along with organic matter that enters the ecosystem from the outside (called allochthonous organic matter) fuels the catabolic activities of chemoorganotrophic organisms. These activities oxidize the organic matter to CO2 by respiration or ferment it to various reduced substances. If chemolithotrophs are present and metabolically active in the ecosystem, they can conserve energy from the oxidation of inorganic electron donors, such as H2, Fe2+, S0, or NH3 (Chapters 14 and 15), and contribute new organic matter through their autotrophic activities.
Biogeochemical Cycling
Microorganisms play an essential role in cycling elements, in particular C, N, S, and Fe, between their different chemical forms. The study of these transformations is part of biogeochemistry, an interdisciplinary science that includes biology, geology, and chemistry. Figure 20.2 shows how the activities of different guilds of microorganisms influence the chemistry of one environment, a lake ecosystem. The sequence of changing chemistry with increasing depth in the sediments corresponds to the layers of different microbial guilds. The location of each guild in the ecosystem is primarily determined by the availability of electron donors and acceptors, both of which tend to decrease with increasing depth in the sediments.

A biogeochemical cycle defines the transformations of an element that are catalyzed by either biological or chemical means (or both). Many different microorganisms participate in biogeochemical cycling reactions, and in many cases, microorganisms are the only biological agents capable of regenerating forms of the elements needed by other organisms, particularly plants. Thus, biogeochemical cycles are often also nutrient cycles, reactions that generate important nutrients for other organisms.
Most biogeochemical cycles proceed by oxidation–reduction reactions as the element moves through the ecosystem and are often tightly coupled, with transformations in one cycle affecting one or more other cycles. For example, hydrogen sulfide (H2S) is oxidized by phototrophic and chemolithotrophic microbes to sulfur (S0) and sulfate (SO4 2−), the latter being a key nutrient for plants. Phototrophs and chemolithotrophs are also autotrophs, and thus affect the carbon cycle by producing new organic carbon from CO2. However, SO4 2− can be reduced to H2S by the sulfate-reducing bacteria, organisms that consume organic carbon, and this reduction closes the biogeochemical sulfur cycle while regenerating CO2. The cycling of nitrogen is also a microbial process and is key to the regeneration of forms of nitrogen usable by plants and other organisms. The nitrogen cycle is driven by both chemolithotrophic and chemoorganotrophic bacteria, organisms that produce and consume organic carbon, respectively. We considered the microbiology of biogeochemical cycles and their coupled nature in Chapters 14 and 15 and will revisit this theme in more detail in Chapter 21.
Check Your Understanding
How does a microbial guild differ from a microbial community?
What is a biogeochemical cycle? Give an example based on sulfur. Why are biogeochemical cycles also called nutrient cycles?
II The Microbial Environment
Being so small, microbes directly experience only a tiny portion of their environment. Microbial interactions, often as part of attached or layered communities, are based on consumption, generation, and diffusion of nutrients and the products of metabolism.
Microorganisms define the limits of life throughout aquatic and terrestrial environments on our planet. Specific conditions required by a particular organism or group of organisms may be subject to rapid change as a result of inputs to and outputs from their habitat or as a result of microbial activities or physical disturbances. Thus, within one environment there can be multiple habitats, some relatively stable and others changing rapidly over time and space.
20.3 Environments and Microenvironments
Besides living in the common habitats of soil and water, microorganisms thrive in extreme environments and also reside on and within the cells of other organisms. The intimate associations developed between microorganisms and other organisms will be explored in Chapters 23 and 24. Here we focus on terrestrial and aquatic microbial habitats.
The Microorganism, Niches, and the Microenvironment
The habitat in which a microbial community resides is governed by physicochemical conditions that are determined in part by the metabolic activities of the community. For example, the organic material used by one species may have been a metabolic by-product of a second species. Another example is oxygen (O2), which can become limiting if biological consumption exceeds the rate at which it is supplied.
Because microbes are very small, they directly experience only a tiny local environment; this small space is called their microenvironment. For example, for a typical 3-μm rod-shaped bacterium, a distance of 3 mm is equivalent to that which a human would experience over a distance of 2 km! As a consequence of the smallness of microorganisms, the variable metabolic activities of nearby microbes, and the changes in physicochemical conditions over short intervals of time and distance, several microenvironments can exist within a given microbial habitat. The conditions supporting growth within a microenvironment correspond to the general requirements for growth we considered in Chapters 3, 4, and 8.
Ecological theory states that for every organism there exists at least one niche, the realized niche (also called the prime niche), where it will be most successful. The organism dominates the realized niche but may also inhabit other niches; in other niches it is less ecologically successful than in its realized niche, but it may still be able to compete. The full range of environmental conditions under which an organism can exist is called its fundamental niche (we considered the realized and fundamental niche in the context of enrichment culture and isolation in Sections 19.1, 19.2 19.3). The word “niche” should not be confused with the word “microenvironment” because the microenvironment describes conditions at a specific location and can change rapidly. In other words, the general conditions that describe a specific niche may be transient at many places in a microenvironment.
Another important consequence of microbes being so small is that diffusion often determines the availability of resources. Consider, for example, the distribution of an important microbial nutrient such as O2 in a soil particle. Microsensors (Section 19.9) can be used to measure oxygen concentrations throughout small soil particles. As shown in the data from an actual microsensor experiment (Figure 20.3), soil particles are not homogeneous in terms of their O2 content but instead contain many adjacent microenvironments. The outer layer of the soil particle may be fully oxic (21% O2) while the center, only a very short distance away (in human terms, but of course a great distance from a microbial standpoint), may be anoxic (O2-free). The microorganisms near the outer edges consume all of the O2 before it can diffuse to the center of the particle. Thus, anaerobic organisms could thrive near the center of the particle, microaerophiles (aerobes that require very low oxygen levels) farther out, and obligately aerobic organisms in the outermost region of the particle. Facultatively aerobic bacteria (organisms that can grow either aerobically or anaerobically, Section 4.16) could be distributed throughout the particle. Nutrient transfer is particularly important in thick assemblages of cells, such as biofilms and microbial mats, and we explore this in Section 20.4.
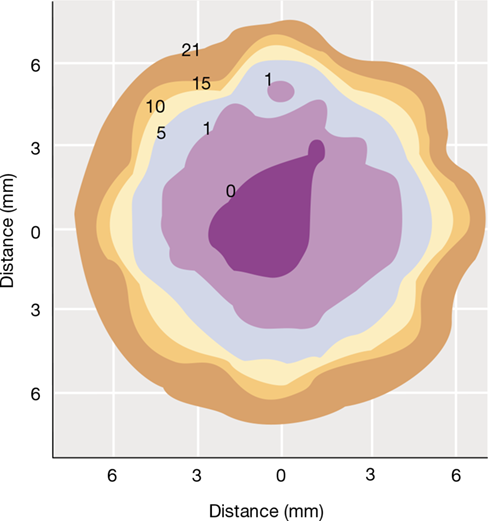
Figure 20.3 Oxygen microenvironments.

Contour map of O2 concentrations in a small soil particle as determined by a microsensor (Section 19.9). The axes show the dimensions of the particle. The numbers on the contours are percentages of O2 concentration (air is 21% O2). Each zone can be considered a different microenvironment.
Physicochemical conditions in a microenvironment are subject to rapid change in both time and space. For example, the O2 concentrations shown in the soil particle in Figure 20.3 represent “instantaneous” values. Measurements taken in the same particle following a period of intense microbial respiration or disturbance due to wind, rain, or disruption by soil animals could differ dramatically from those shown. During such events certain populations may temporarily dominate the activities in the soil particle and grow to high numbers, while others remain dormant or nearly so. However, if the microenvironments shown in Figure 20.3 are eventually reestablished, the various microbial activities characteristic of different regions of the soil particle will eventually return as well.
Nutrient Levels and Growth Rates
Resources (Table 20.1) typically enter an ecosystem intermittently. A large pulse of nutrients—for example, an input of leaf litter or the carcass of a dead animal—may be followed by a period of nutrient deprivation. Because of this, microorganisms in nature often face a “feast-or-famine” existence. It is thus common for them to produce storage polymers as reserve materials when resources are abundant and draw upon these reserves in periods of starvation. Examples of storage materials are poly-β-hydroxyalkanoates, polysaccharides, and polyphosphate (Section 2.7).

Extended periods of exponential microbial growth in nature are typically rare, except in certain extreme environments—for example, a hot spring—where conditions and resources may stay optimal indefinitely. Instead, microorganisms typically grow in spurts, linked closely to the availability and types of resources and ideal growth conditions. Because all relevant physicochemical conditions in nature are rarely optimal for microbial growth at the same time, growth rates of microorganisms in nature are usually well below the maximum growth rates recorded in the laboratory. For instance, the generation time of Escherichia coli in the intestinal tract of a healthy adult eating at regular intervals is about 12 h (two doublings per day), whereas in pure culture it can grow much faster, with a minimum generation time of about 20 min under optimal conditions. In addition, research-based estimates indicate that most cultured soil bacteria typically grow in nature at less than 1% of the maximal growth rate measured in the laboratory.
These slower growth rates in nature than in laboratory culture reflect the facts that (1) resources and growth conditions (Table 20.1) are frequently suboptimal; (2) the distribution of nutrients throughout the microbial habitat is not uniform; and (3) except in rare instances, microorganisms in nature grow in mixed populations rather than pure culture. An organism that grows rapidly in pure culture may grow much slower in a natural environment where it must compete with other organisms that may be better suited to the resources and growth conditions available.
Microbial Competition and Cooperation
Competition among microorganisms for resources in a habitat may be intense, with the outcome dependent on several factors, including rates of nutrient uptake, inherent metabolic rates, and ultimately, growth rates. A typical habitat contains a mixture of different species (Figures 20.1 and 20.2), with the density of each population dependent on how closely its local environment resembles its realized niche.
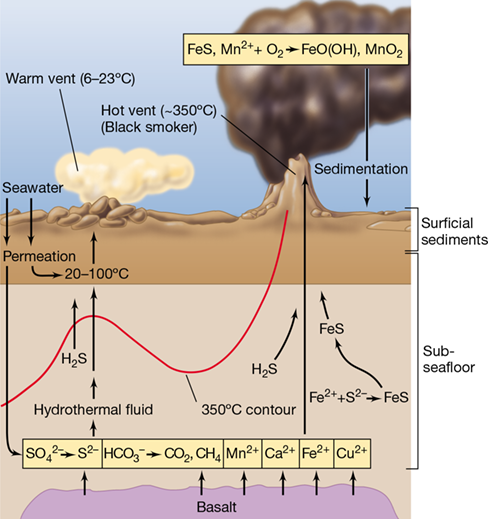
Some microbes work together to carry out transformations that neither can accomplish alone—a process called syntrophy—and these microbial partnerships are particularly important for anoxic carbon cycling (Section 14.22 and Section 21.2). Metabolic cooperation can also be seen in the activities of organisms that carry out complementary metabolisms. For example, we have previously considered metabolic transformations that are carried out by two distinct groups of organisms, such as those of the nitrifying Bacteria and Archaea (Sections 14.9, 15.10, and 17.5). Together, these nitrifiers oxidize ammonia (NH3) to nitrate (NO3 −). Because nitrite (NO2 −), the final product of most ammonia-oxidizing nitrifiers (Section 15.10 and Section 21.3), is the substrate for the nitrite-oxidizing bacteria, the two groups of organisms often live in nature in tight association within their habitats (Figure 19.13a).
Winogradsky discovered nitrification in 1890 (Section 1.13), but for over one hundred years no organism capable of complete ammonia oxidation was ever discovered. However, some Nitrospira species (Bacteria) can oxidize both ammonia and nitrite (Section 15.10), a process that has been coined comammox. With the use of molecular tools (Chapter 19) to survey various habitats for comammox bacteria, related organisms have been identified in wetlands, riverbeds, aquifers, lake sediments, and wastewater treatment systems but not in the oceans. In some of these environments, comammox bacteria seem to outnumber classical nitrifying bacteria (species that catalyze only one step in nitrification), whereas in other environments, comammox organisms may be part of the rare biosphere (Section 19.6), low in abundance yet nevertheless ecologically important microbes.
Check Your Understanding
What characteristics define the realized niche of a particular microorganism?
Why can many different physiological groups of organisms live in a single habitat?
20.4 Surfaces and Biofilms
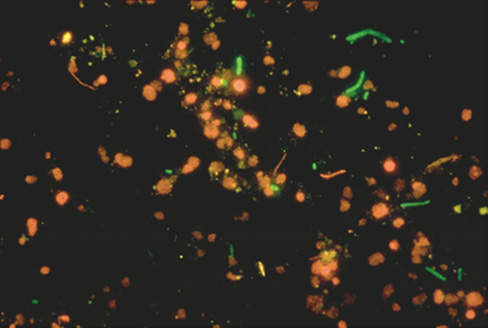
Surfaces are important microbial habitats, typically offering microbes greater access to nutrients and protection from predation and physicochemical disturbances. Attachment to a surface also offers cells a means to remain in a favorable habitat, modify the habitat by their own activities, and not be washed away. In addition, flow across a colonized surface increases the flux of nutrients to the surface, providing more resources than are available to planktonic cells (cells that live a floating existence) in the same environment. A surface may be abiotic, such as a particle of organic matter, or provided by another organism. For example, plant root surfaces become heavily colonized by soil bacteria thriving on organic compounds excreted by the plant, as revealed when fluorescent stains such as acridine orange (Section 19.4) are used on natural samples (**Figure 20.4*a***).
Figure 20.4 Microbial biofilms on surfaces.

(a) Fluorescence photomicrograph of a natural microbial community living on plant roots in soil and stained with acridine orange. Note microcolony development. (b) Bacterial microcolonies developing on a microscope slide that was immersed in a river. The bright particles are mineral matter. The short, rod-shaped cells are about 3 μm long. (c) Confocal scanning laser microscopy through a natural biofilm (top view) on a leaf surface. The color of the cells indicates their depth in the biofilm: red, surface; green, 9-μm depth; blue, 18-μm depth. (d) A cross-sectional view of an experimental biofilm composed of cells of Pseudomonas aeruginosa. The yellow layer (about 15 μm in depth) contains cells and is stained by a reaction showing activity of the enzyme alkaline phosphatase.
Virtually any natural or artificial surface exposed to microorganisms will be colonized. For example, microscope slides have been used as experimental surfaces to which organisms can attach and grow. A slide can be immersed in a microbial habitat, left for a period of time, and then retrieved and examined microscopically (Figure 20.4b). Clusters of a few cells that develop from a single colonizing cell—called microcolonies—form readily on such surfaces, much as they do on natural surfaces (Figure 20.4a). Periodic microscopic examination of immersed slides has been used to measure growth rates of attached organisms in nature.
Surface colonization may be sparse, consisting only of microcolonies not visible to the naked eye, or may consist of so many cells that microbial accumulation becomes visible as, for example, in a stagnant toilet bowl. Surface growth can be particularly problematic in a hospital setting where microbial colonization of indwelling devices such as catheters and intravenous lines can cause serious infection (Figure 4.17b). In a few extreme environments that lack small animal grazers (for example, hot springs), microbial accumulation on a surface can be layered and several centimeters thick. Called microbial mats (Section 20.5), such accumulations often contain highly complex yet very stable assemblages of phototrophic, chemolithotrophic, autotrophic, and heterotrophic microbes.
Layered communities can even form where light penetrates the near-surface interior of minerals, such as calcium sulfate (gypsum) crusts in solar salterns. Despite the extremely high salt content of these crystals, they are readily colonized by a community of halophilic phototrophs and heterotrophs (Figure 20.5). The phototrophic components in the gypsum crusts include cyanobacteria (Section 15.3; orange- and green-colored layers in Figure 20.5) and purple sulfur bacteria (Section 15.4, red layer in Figure 20.5). Purple bacteria grow in the deepest layer of the crust because of their need for H2S (produced by sulfate-reducing bacteria deeper in the crust) and ability to grow at low light intensities.
Figure 20.5 Endolithic microbial biofilm.

Section through a submerged calcium sulfate (gypsum) crust that formed in a Guerrero Negro (Baja California, Mexico) crystallizer pond showing layered phototrophic microbial communities. The orange layer just under the crust surface contains the unicellular cyanobacterium Halothece, the green layer a mixture of unicellular and filamentous cyanobacteria, and the red layer purple sulfur bacteria of the genera Ectothiorhodospira and Halochromatium. Various chemotrophic bacteria are present in each of the layers as well, including sulfate-reducing bacteria, the source of the H2S that feeds the purple sulfur bacteria.
Biofilms
As bacterial cells grow on surfaces, they commonly form biofilms—assemblages of bacterial cells attached to a surface and enclosed in an adhesive matrix that is the product of excretion by cells and cell death (Figure 20.4c, d). The matrix is typically a mixture of polysaccharides, proteins, and nucleic acids that bind the cells together. Biofilms trap nutrients and help prevent the detachment of cells on dynamic surfaces, such as in flowing systems (Figure 20.6). We examined the basic properties of biofilms in Section 4.9 and genetic and regulatory features of biofilm formation in Section 8.10, and so here we focus on their ecological and medical consequences.
Figure 20.6 Phototrophic biofilms in rivers and streams.

(a) Phototrophic biofilms colonizing the rocky bottom of a stream flowing from the Rhone Glaciers, Switzerland (water is flowing toward the viewer). The striped patterns emerge from microbial growth along different flow paths of the glacial outflow. (b) Cyanobacteria attached to the river rocky substratum form “tower-like” clusters with apical oxygen bubbles forming from high photosynthesis rates within the clusters. The clusters are about 0.5–1 cm high.
Biofilms typically contain multiple layers of cells embedded in the porous matrix material, and the cells in each layer can be examined by confocal scanning laser microscopy (Section 1.9; Figure 20.4c). Biofilms may contain one or two species or, more commonly, many species of bacteria. The biofilms that form on tooth and soft surfaces of the mouth, for example, contain between 100 and 200 different phylotypes (Section 19.6), including species of both Bacteria and Archaea; in total, the human mouth is a habitat for at least 700 phylotypes (Sections 24.3 and 25.2). Biofilms are thus functional and growing microbial communities and not just cells trapped in a sticky matrix.
Wherever submerged surfaces are present in natural environments, biofilm growth is almost always more extensive and diverse than the planktonic growth in the liquid above the surface. Biofilms differ from planktonic communities in supporting critical transport and transfer processes, which generally control growth in biofilm environments. For example, if consumption of O2 by populations near the surface exceeds diffusion of O2 into deeper regions of the biofilm, the deeper regions will become anoxic, opening up new niches for colonization by obligate anaerobes or facultative aerobes. This is analogous to the depletion of O2 in the interior of a soil particle that was depicted in Figure 20.3. Hence, different metabolisms can be active in different regions of a biofilm and this broadens the diversity of microbes that can exist there.
Why Bacteria Form Biofilms
At least three reasons have been proposed for the formation of biofilms. First, biofilms are a means of microbial self-defense that increase survival. Biofilms resist physical forces that could otherwise remove cells only weakly attached to a surface. Biofilms also resist phagocytosis by protozoa and cells of the immune system and retard the penetration of toxic molecules such as antibiotics. These advantages improve the chances for survival of cells in the biofilm. Second, biofilm formation allows cells to remain in a favorable niche. Biofilms attached to nutrient-rich surfaces, such as animal tissues, or to surfaces in flowing systems (Figure 20.6) fix bacterial cells in locations where nutrients may be more abundant or are constantly replenished. Third, biofilms form because they allow bacterial cells to live in close association with each other. This facilitates cell-to-cell communication, offers more opportunities for nutrient and genetic exchange, and in general increases chances for survival.
The fact that biofilms form on virtually any surface capable of supporting bacterial growth suggests that biofilms are the “default” growth mode for bacteria in natural habitats. Planktonic growth, so common in shaken or otherwise mixed laboratory cultures, may thus be an atypical growth mode and the norm only for those bacteria adapted to life at extremely low nutrient concentrations (oligotrophs, Sections 20.9, 20.10, and 20.12). For example, the rocky and pebbly substratum of rivers and streams is an important biofilm habitat, as any hiker attempting to cross over the slippery rocks of a mountain stream is aware. This type of biofilm is in part nourished by phototrophic algae and cyanobacteria that rapidly colonize newly exposed surfaces (Figure 20.6a).
Although biofilms are known to provide some resistance to grazing, they are not immune to grazing and typically attract high densities of micrograzers, including heterotrophic flagellates, amoebae, and ciliates. These specialized biofilm micrograzers, together with small metazoans called meiofauna (rotifers, nematodes, micro-crustaceans, oligochaetes, and flies), feed on components of the biofilm. In doing so, they incorporate a significant amount of the carbon fixed by biofilm autotrophs and also remove planktonic cells and organic carbon from the water column. Thus, by existing at the interface between an inert substratum and surface water, river and stream biofilms provide significant ecosystem services by coupling benthic and water column microbial communities and nourishing higher trophic levels in aquatic systems.
Biofilms on Microplastics
In addition to natural surfaces, plastics provide an artificial surface for microbial colonization, and biofilms that develop on plastics can have both beneficial and harmful aspects. Each year humans add millions of metric tons of plastic to the oceans alone. Biodegradation and weathering processes degrade this plastic into small particles and fibers less than 5 mm in diameter called microplastics (MPs, Figure 20.7), and the surfaces of MPs get colonized by microbes. On the one hand, the formation of biofilms on these surfaces signals the initial stages of biodegradation and can be considered a good thing. But on the other hand, more than 200 marine organisms are known to ingest microplastics, including zooplankton, fish, seabirds, marine mammals, and benthic invertebrates. Depending on the animal, MPs can cause a variety of health issues including neurotoxicity, DNA damage, reproductive defects, and decreased filtration rates in filter feeders such as mussels. Unfortunately, biofilm-coated (as opposed to clean) MP may actually escalate this environmental problem.
Figure 20.7 Biofilm formation on a microplastics particle.

Microbial colonization and biofilm formation (arrows) on a polystyrene microplastics particle following immersion for three months in a bog in Germany. This microplastic particle is 3–4 mm in diameter and about 0.1 mm thick.
Surface colonization and biodegradation of plastic by microorganisms is suspected to increase harmful environmental effects by altering the surface chemistry, density, and sinking rates of MPs. In addition, since microbially colonized MPs (Figure 20.7) can be greatly enriched in hydrophobic organic contaminants relative to surrounding seawater, they are also a conduit for the delivery of these adsorbed contaminants to filter-feeding biota. Thus, apart from the possibility that filter feeders may preferentially ingest biofilm-encrusted MPs because of their higher nutritional quality, most studies point to only adverse impacts of MPs on aquatic ecosystems.
Problem Biofilms for Humans
In addition to aquatic systems, biofilms have significant implications in human medicine and commerce. In the body, bacterial cells within a biofilm are protected from attack by the immune system, and antibiotics and other antimicrobial agents often fail to penetrate the biofilm. One of the most clinically and industrially relevant properties of biofilm microbial communities is their inherent tolerance of antibiotics and other antimicrobial chemicals. A given species growing in a biofilm can be up to 1000 times more tolerant of an antimicrobial substance than planktonic cells of the same species. Reasons for this greater tolerance include slower growth rates in biofilms, reduced penetration of antimicrobial substances through the extracellular matrix, and the expression of genes that increase tolerance to stress. This tolerance of antimicrobial substances may explain why biofilms are responsible for many untreatable or difficult-to-treat chronic infections and are also hard to eradicate in industrial systems, such as wastewater plants, where surface growth (fouling) by microbes may impair important processes.
As we saw in earlier chapters, the bacterium Pseudomonas aeruginosa forms tenacious biofilms (Sections 4.9 and 8.10 and Figure 4.16), and biofilms of this organism form in the lungs of humans suffering from the genetic disease cystic fibrosis. Once in the biofilm state, P. aeruginosa is difficult to treat with antibiotics and the biofilm helps the organism persist. Burkholderia (previously Pseudomonas) cepacia is also a threat to those with cystic fibrosis (Section 16.2). Besides cystic fibrosis, biofilms have been implicated in several medical and dental conditions, including periodontal disease, chronic wounds, kidney stones, tuberculosis, Legionnaires’ disease, and Staphylococcus infections (Figure 4.17b). Medical implants are ideal surfaces for biofilm development. These include both short-term devices, such as urinary catheters, as well as long-term implants, such as artificial joints. It is estimated that 10 million people a year in the United States experience biofilm infections from implants or intrusive medical procedures. Biofilms also explain why routine oral hygiene is so important for maintaining dental health. Dental plaque is a typical biofilm and contains acid-producing bacteria responsible for dental caries (Sections 24.3 and 25.2 and Figures 25.7, 25.8).
Biofilms affect commerce because they can slow the flow of water, oil, or other liquids through pipelines and can accelerate corrosion of the pipes themselves (Figure 1.17). Biofilms also initiate the degradation of submerged objects, such as structural components of offshore oil platforms, boats, and shoreline installations. The safety of drinking water may be compromised by biofilms that develop in water distribution pipes. These biofilms mostly contain harmless microbes, but if pathogens successfully colonize a biofilm, water purification practices (Chapter 22) may fail to kill them. Periodic releases of pathogenic cells can then lead to outbreaks of disease. For example, it is thought that Vibrio cholerae, the causative agent of cholera (Section 33.3), may be propagated in this manner.
Biofilm control is big business, and thus far, only a limited number of tools exist to fight biofilms. Collectively, industries commit huge financial resources to treating pipes and other surfaces to keep them free of biofilms. New antimicrobial agents that can penetrate biofilms, as well as drugs that eliminate biofilm formation by interfering with intercellular communication, are being developed, but at this point, human battles with biofilms continue at a brisk pace.
Check Your Understanding
Why might a biofilm be a good habitat for bacterial cells living in a flowing system?
Give an example of a medically relevant biofilm that forms in virtually all healthy humans.
How is it possible for both aerobes and obligate anaerobes to coexist in the same biofilm?
20.5 Microbial Mats
Microbial mats are among the most visibly conspicuous of microbial communities and can be thought of as extremely thick biofilms. Supported by phototrophic or chemolithotrophic bacteria, these layered microbial communities can be several centimeters thick (**Figure 20.8a,*b***). The layers are composed of species of different microbial guilds whose activities are governed by light availability and other resources (Table 20.1). The combination of microbial metabolism and nutrient transport controlled by diffusion results in steep concentration gradients of different microbial nutrients and metabolites, creating unique niches at different depth intervals in the mats. The most abundant and versatile phototrophic mat builders are filamentous cyanobacteria, which are oxygenic phototrophs and many of which tolerate extreme environmental conditions. For example, some species of cyanobacteria grow in waters as hot as 73 °C or as cold as 0 °C, and others tolerate salinities in excess of 12% and pH values as high as 10.
Figure 20.8 Microbial mats.

(a) Mat specimen collected from the bottom of a hypersaline pond at Guerrero Negro, Baja California (Mexico). Most of the bottom of this shallow pond is covered with mats built by the major primary producer, the filamentous cyanobacterium Microcoleus chthonoplastes. (b) Microbial mat core from an alkaline Yellowstone National Park (USA) hot spring. The upper (green) layer contains mainly cyanobacteria, while the reddish layers contain anoxygenic phototrophic purple bacteria. (c) Daylight oxygen (O2), H2S, and pH profiles through a hot spring mat core such as that shown in part b.
Cyanobacterial Mats
Cyanobacterial mats (Figure 20.8a, b) are complete microbial ecosystems, containing large numbers of primary producers (cyanobacteria and other phototrophic bacteria) that use light energy to synthesize new organic material from CO2. These along with populations of consumers in the mat community mediate all key nutrient cycles.
Microbial mats have existed for over 3.5 billion years (stromatolites, Section 13.2 and Figure 13.6) but are found today only in aquatic environments where environmental stresses such as high temperatures or high salt concentrations restrict grazing by small animals and insects. Well-studied microbial mats are found in hypersaline solar evaporation basins; such basins have either formed naturally, such as Solar Lake (Sinai, Egypt), or have been constructed for the recovery of sea salt (Figure 20.8a). Layered phototrophic communities also form within the salt crusts of these evaporation basins (Figure 20.5). Because microbial mats are restricted to extreme environments, most are found in remote locations and many are not readily accessible to study. In contrast, however, the cyanobacterial mats that colonize the outflow channels of hot springs in Yellowstone National Park (USA), Iceland, and many other thermal regions in the world are easily accessible and have been widely studied (Figure 20.8b, c).
The chemical and biological structure of a microbial mat can change dramatically during a 24-h period (called a diel cycle) as a consequence of light to dark transitions. Using microsensors (Section 19.9) it is possible to measure pH, H2S, and O2 repeatedly over a diel cycle in zones in the mat separated vertically by only a few micrometers. During the day, there is intense oxygen production in the cyanobacterial surface layer of microbial mats and active sulfate reduction throughout the lower regions. Near the zone where O2 and H2S begin to mix, intense metabolic activity by phototrophic and chemolithotrophic sulfur bacteria may consume these substrates rapidly over very short vertical distances. Detecting the rate of these changes reveals the zones of greatest microbial activity (Figure 20.8c). These gradients disappear at night when photosynthesis stops, and the entire mat turns anoxic as H2S accumulates. Some mat organisms rely on motility to follow the shifting chemical gradients. For example, sulfur-oxidizing filamentous phototrophic bacteria such as Chloroflexus and Roseiflexus (Section 15.7) track the up-and-down movement of the O2–H2S interface on a diel basis.
Iron-Rich Microbial Mats
Microbial mats of oxidized iron also form in metal-rich, low-pH environments, such as occurs with the mining of metals (**Figure 20.9*a***; see acid mine drainage, Section 22.2) and in iron-rich hot springs, such as that of Echinus Geyser in Yellowstone National Park (Figure 20.9b). As Fe2+-rich water is exposed to air, iron-oxidizing Bacteria and Archaea generate a thick mat of oxidized iron. Metagenomic analyses of the Echinus Geyser mat revealed a novel phylum of microaerophilic Archaea, provisionally named the Marsarchaeota (see Figure 20.16) after the resemblance of their red iron-oxide-rich habitat to the color of Mars. Enrichment cultures of Marsarchaeota show large cocci that can oxidize both organic carbon and ferrous iron. Their heterotrophic capabilities suggest that they receive organic carbon fixed by chemolithotrophs coexisting in these iron-rich mat communities. An example of such a chemolithotroph is Metallosphaera, a relative of the sulfur- and iron-oxidizing Sulfolobus, a genus of Archaea (Section 17.10). We will see in Chapter 22 that acidophilic iron-oxidizing microbes in mining operations can cause considerable environmental damage to both flora and fauna when the acidity and toxic heavy metals their activities release enter aquatic systems.
Figure 20.9 Iron-oxidizer microbial mats.

(a) A mat of iron-oxidizing bacteria attached to rocks in the Rio Tinto, Spain. As ferrous (Fe2+)-rich water from metal mining activities flows over and through the biofilm, the organisms oxidize Fe2+ to Fe3+. (b) An iron-rich microbial mat in the acidic (pH 3–4) and hot (70–80 °C) outflow of Echinus Geyser (Norris Geyser Basin, Yellowstone National Park, Wyoming, USA). The terraces in the outflow channel are formed by surface growth of ferrous-iron-oxidizing Sulfolobales (Metallosphaera yellowstonensis) (Section 17.10) that form insoluble ferric oxides. The mat also contains significant populations of Geoarchaeota (a heterotrophic candidate archaeal phylum) and the phylum Marsarchaeota. Photo courtesy of William Inskeep, Research Permit YELL-2016-SCI-05068.
Chemolithotrophic Microbial Mats
Like iron-rich mats (Figure 20.9), which typically lack phototrophic components and are thus supported by the activities of iron-oxidizing chemolithotrophs, some microbial mats develop from the activities of sulfur-oxidizing chemolithotrophs. The most common of these are mats composed of filamentous sulfur bacteria such as Beggiatoa and Thioploca species that grow on marine sediment surfaces at the interface between O2 supplied from oxic overlying waters and H2S produced by sulfate-reducing bacteria residing in the anoxic sediments. In these dark habitats, photosynthesis cannot occur and so the bacteria oxidize H2S to yield energy and support autotrophy (Sections 14.7 and 15.12).
Chemolithotrophic mats composed of sulfur-oxidizing Thioploca species on sediments of the Chilean and Peruvian continental shelf are thought to be the most extensive microbial mats of any type on Earth (Figure 20.10). The bottom waters near the sediments in this region of the Pacific Ocean are oxygen-depleted (an oxygen minimum zone, Section 20.10) but contain significant levels (about 25 μm) of nitrate (NO3 −). Thioploca (Figure 15.32b) can oxidize sulfide anaerobically with nitrate as electron acceptor and has evolved a remarkable strategy to bridge these spatially separated resources—nitrate in the overlying water and sulfide in the sediments. Thioploca cells contain large internal vacuoles that can concentrate nitrate from the surrounding water to levels as high as 0.5 M. To do this, they migrate upward from the sediments by gliding motility (Section 2.10) and charge their vacuoles with NO3 − (Figure 20.10a, b). They then return into the anoxic sediment (gliding at speeds of 3–5 mm per hour) to use their stored NO3 − as an electron acceptor for H2S oxidation.
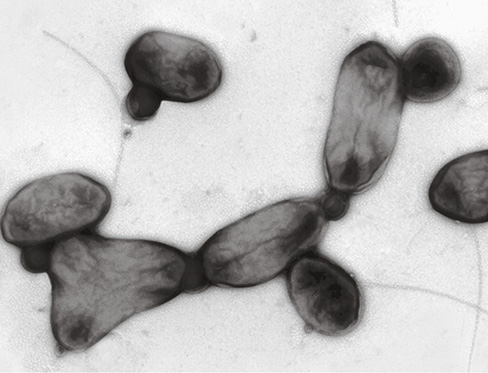
Figure 20.10 *Thioploca* mats.

(a, c) Filaments of the large sulfur-oxidizing chemolithotroph Thioploca extend into the water above the sediment (87-m depth) in the Bay of Concepción off the Chilean coast. (b) Thioploca form bundles of 10 to 20 filaments (trichomes) held together by a gelatinous sheath, each bundle approximately 1.5 mm in diameter and 10–15 cm in length. Two species of Thioploca commonly inhabit the same bundle: T. chileae, about 20 μm in diameter, and T. araucae, about 40 μm in diameter. Individual trichomes glide independently within the sheaths and can extend up to 3 cm into the water.
The physical and biological structures of both biofilms and microbial mats are determined by metabolic interactions among the microbes within them and the diffusion of nutrients. Thus, as biofilms form on a surface they become increasingly more complex, and in so doing generate new niches for organisms of differing physiologies. This diversity reaches a maximum in mature microbial mats (Figure 20.8a, b), as molecular community sampling (Section 19.6) has shown these structures to be among the most complex microbial communities yet discovered.
Check Your Understanding
What is a microbial mat, and what major nutrient changes occur in phototrophic mats during a diel cycle?
How would motile aerobic bacteria in a microbial mat respond to changing O2 concentrations over a diel cycle?
What is the strategy used by Thioploca to oxidize sulfide in anoxic regions below the surface of marine sediments?
III Terrestrial Environments
Terrestrial environments range from wet to dry, warm to cold, alkaline to acidic, oxic to anoxic, and nutrient-rich to nutrient-poor. These characteristics control the diversity and activity of microorganisms that inhabit the terrestrial environments found on Earth.
Extensive microbial habitats exist in two terrestrial environments on Earth: surface soils and the deep subsurface. We cover these microbial habitats in the following three sections, the first two on soils and the third one on the subsurface. We consider the basic characteristics of each habitat, explore their microbial diversity, and discuss reasons why the microbes that are present in each habitat may be well suited to their environment.
20.6 Soils: General Properties
The word soil refers to the loose outer material of Earth’s surface, a layer distinct from the bedrock that lies underneath (Figure 20.11). Soil develops over long periods through complex interactions among the parent geological materials (rock, sand, glacial drift materials, and so on), the topography, climate, and the presence and activities of living organisms.
Figure 20.11 Soil.

(a) Profile of a mature fertile soil. The soil horizons are zones defined by soil scientists, and the graph to the left shows approximate levels of organic carbon in different soil horizons. (b) Photo of a soil profile, showing O, A, and B horizons. This soil from Carbondale, Illinois (USA), is rich in clay and very compact. Such soils are not as well drained as those rich in sand. Note the clear color delineation between the organic-rich A horizon and the less-organic-rich B horizon.
Mastering Microbiology
Soils can be divided into two broad groups: Mineral soils are derived from the weathering of rock and other inorganic materials, and organic soils are derived from sedimentation in bogs and marshes. Most soils are a mixture of these two basic types. Although mineral soils, which are the primary focus of this section, predominate in most terrestrial environments, there is increasing interest in the role that organic soils play in carbon storage. A detailed understanding of carbon storage (sinks) and sources (such as release of CO2) is of great relevance to the science of climate change. The carbon cycle is a major focus of Chapter 21.
Soil Composition
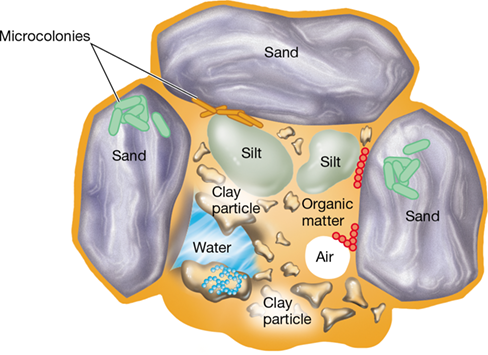
Vegetated soils have at least four components. These include (1) inorganic mineral matter, typically 40% or so of the soil volume; (2) organic matter, usually about 5%; (3) air and water, roughly 50%; and (4) microorganisms and macroorganisms, about 5%. Particles of various sizes are present in soil. Soil scientists classify soil particles on the basis of size: Those in the range of 0.1–2 mm in diameter are called sand, those between 0.002 and 0.1 mm silt, and those less than 0.002 mm clay. Different textural classes of soil are then given names such as “sandy clay” or “silty clay” based on the percentages of sand, silt, and clay they contain. A soil in which no one particle size dominates is called a loam.
Soils are categorized based on their depth in a soil profile, and the absolute depths vary from soil to soil. In a typical mineral soil the O horizon (“O” for “organic”) (Figure 20.11a) is the uppermost layer and is dominated by decomposing organic matter; because of this, vast numbers (as high as 1010/g) of microbial cells are often present here. The O horizon segues into the “A” horizon, the soil layer commonly referred to as “topsoil,” which is also organic-rich and extends anywhere from a few centimeters to about 50 cm from the surface, depending on a soil’s structural characteristics. The “B” horizon, called “subsoil,” can extend over a meter below the A horizon and is rich in iron- and aluminum-containing clay minerals and thus red/brown in color (Figure 20.11b). Plant roots, especially tree roots, often extend into the B horizon. The “C” horizon, also called “parent material,” transitions into underlying bedrock and can extend for several meters from the surface.
The organic content of a soil is typically inversely related to depth, with the organic-rich O–A horizons transitioning into the more organic carbon–depleted B and nearly organic carbon–free C horizons (Figure 20.11a). Organic matter content of a soil varies with the type, but in the soil shown in Figure 20.11b (a soil that the United States Department of Agriculture would classify as an “alfisol”), values of organic carbon as high as 5% can be present in the O–A horizons; values drop to less than 1% in the B horizon and less than 0.25% beneath this (Figure 20.11a). The nitrogen and phosphorus content of soils follows a similar pattern to that of carbon, reflecting the fact that the bulk of these key nutrients are tied up in decaying plants and animals and in microbes, both dead and alive.
Soil Formation
Physical, chemical, and biological processes all contribute to the formation of soil. An examination of almost any exposed rock reveals the presence of algae, lichens, or mosses. These organisms are phototrophic and produce organic matter, which supports the growth of chemoorganotrophic bacteria and fungi. More complex chemoorganotrophic communities composed of Bacteria, Archaea, and eukaryotes then develop as the extent of the earlier colonizing organisms increases. Carbon dioxide produced during respiration becomes dissolved in water to form carbonic acid (H2CO3), which slowly dissolves the rock, especially rocks containing limestone (CaCO3). In addition, many chemoorganotrophs excrete organic acids, which also promote the dissolution of rock into smaller particles.
Freezing, thawing, and other physical processes assist in soil formation by forming cracks in the rocks. As the particles generated combine with organic matter, a crude soil forms in these crevices, providing sites needed for pioneering plants to become established. The plant roots penetrate farther into the crevices, further fragmenting the rock; the excretions of the roots promote development in the rhizosphere (the soil that surrounds plant roots and receives plant secretions) of high microbial cell abundance (Figure 20.4a). When the plants die, their remains are added to the soil and become nutrients for more extensive microbial development. Minerals are rendered soluble, and as water percolates, it carries some of these substances deeper into the soil.
As weathering proceeds, the soil increases in depth and becomes able to support the development of larger plants and small trees. Soil animals such as earthworms colonize the soil and play an important role in keeping the upper layers of the soil mixed and aerated. Eventually, the movement of materials downward results in the soil profile (Figure 20.11). The rate of development of a typical soil profile depends on climatic and other factors, but it can take hundreds to thousands of years from bare rock to fertile soil.
Water Availability: Vegetated and Dryland Soils as Microbial Habitats
The limiting nutrients in soils are often inorganic, such as phosphorus and nitrogen compounds, key components of several classes of macromolecules. Another major factor affecting microbial activity in soil is the availability of water, and we have previously emphasized the importance of water for microbial growth (Section 4.15).
Water is a highly variable component of soil, and a soil’s water content depends on soil composition, rainfall, drainage, and plant cover. Water is held in the soil in two ways—by adsorption onto surfaces or as free water in thin sheets or films between soil particles (Figure 20.12). The water present in soils has materials dissolved in it, and the mixture is called the soil solution. In well-drained soils, air penetrates readily, and the oxygen concentration of the soil solution can be high, similar to that of the soil surface. In waterlogged soils, however, the only oxygen present is that dissolved in water, and this can be rapidly consumed by the resident microbiota. Such soils then become anoxic, and, as described for freshwater environments (Section 20.9), show profound changes in their biological activities. There is also water in the larger channels in soil, where bulk flow is important for rapid transport of microorganisms and their substrates and products.
Figure 20.12 A soil microbial habitat.

Very few microorganisms are free in the soil solution; most of them reside in microcolonies attached to the soil particles. Note the relative size differences among sand, clay, and silt particles.
Arid Soils
The greatest microbial activity in soils is in the organic-rich surface layers in and around the rhizosphere (Figure 20.4a). However, some soils are so dry that plant coverage is greatly limited, and only special microbial communities can thrive. These are arid soils, and approximately 35% of Earth’s landmass is permanently or seasonally arid. Aridity can be defined by the aridity index, expressed as the ratio of precipitation to potential evapotranspiration (P/PET). Evapotranspiration is the sum of water loss through evaporation and plant transpiration. A region is deemed arid if there is a P/PET of less than 1; that is, water entering through precipitation (and fog and dew) is less than that lost through evapotranspiration.
Arid soils are among the most extreme environments on Earth, with temperature highs in excess of 60 °C and lows of −24 °C, high insolation (exposure to solar rays), and low water activity (Section 4.15). Although arid regions are typically nearly devoid of leafy plants, they sustain important microbial communities that assemble in and stabilize soil near the surface and reside within and on the surfaces of rocks (Section 19.8 and Figure 19.27). The dominant microorganisms present in these carbon-limited environments are cyanobacteria, with lesser numbers of green algae, fungi, heterotrophic bacteria, lichens, and mosses.
Dryland microbial habitats include biological soil crusts (BSCs) (Figure 20.13), ventral surfaces of translucent stones (hypolithic colonists), exposed rock surfaces (epilithic colonists), and the interior pore spaces, cracks, and fissures of rocks or minerals (endolithic colonists; Figure 18.35 and Figure 20.5). The soil crusts are dominated by the filamentous cyanobacterium Microcoleus (Figure 20.13b, c; Figure 19.27), whereas coccoid Chroococcidiopsis species are the predominant endolithic population. The rock colonists play an important role in weathering and soil formation; here we primarily consider the BSC communities.
Figure 20.13 Biological soil crust (BSC).

(a) BSC on the Colorado Plateau shown adjacent to lighter, disturbed soils. (b, c) Scanning electron micrographs of filamentous cyanobacteria (Microcoleus species) that bind sand grains together with their sheath material (Figure 19.27). The sand grains in part b are about 100 μm in diameter and the filaments in part c about 5 μm in diameter.
The BSC functions in soil stabilization of desert ecosystems. Stabilization is critical because of the very slow rate of desert soil formation (<1 cm per 1000 years). Here, the Microcoleus and fungi provide soil cohesion, which is further stabilized aboveground by lichens and mosses when present. Importantly, this microbial network functions to virtually eliminate soil erosion from wind and water. The BSCs are major determinants of water infiltration and influence local hydrological cycles and water availability to vegetation. Remarkably, when moisture and temperature conditions are optimal, the photosynthetic rates of BSC are comparable to those of vascular plant leaves. In soil crusts dominated by Microcoleus, nitrogen-fixing bacteria (Sections 8.9, 3.12, and 15.3) provide most of the nitrogen, and much of the fixed nitrogen is released immediately and made available to Microcoleus and other soil biota (Section 19.8). To respond to variability in moisture conditions, desert soil microbial communities have higher relative abundances of genes encoding osmoregulation and dormancy functions; this reflects their adaptation to the extremes in water availability they experience, from rarely very wet to usually dry or even arid.
The disruption of BSCs is a major contributor to desertification, a process exacerbated by climate change and human activities. Dust storms resulting from BSC destruction reduce soil fertility, and when heavy dust is deposited on nearby snowfields it accelerates melt and evapotranspiration rates, thereby reducing freshwater inputs to rivers. Once compromised, soil crusts have recovery times varying from 15 to 50 years. Given the expansive terrestrial presence of BSCs, their importance to human and ecosystem function, and the projected increase in aridity associated with climate change (Section 21.9), a better understanding of BSC formation and the rehabilitation of compromised BSCs is important for a healthy planet Earth. In this connection, important new understanding of the metabolic interactions among the microbes that sustain these critical communities is emerging from the new research area of metabolomics (Section 19.8, subsection Utility of Metabolomics: An Environmental Case Study of Biocrusts). As we saw in this section, biocrusts contain both producer and consumer microbes, and the exchange of nutrients between these two groups is extensive (Figure 19.28), bordering on what could be called a symbiosis.
With this brief background on soils, we next consider soil microbial diversity. Although some microbial eukaryotes reside in soils, our focus will be on prokaryotic cells because these are present in soils in highest numbers and greatest diversity.
Check Your Understanding
What four components make up a typical vegetated soil?
Which region of soil is the most microbially active? What are important sources and types of carbon in soil?
Why is the cyanobacterium Microcoleus important in arid soils?
20.7 Prokaryotic Diversity in Soils
As we saw in Figure 20.3, even a single soil particle can contain many different microenvironments, and as a consequence, soils typically support the growth of several physiological types of microbes. To examine soil particles directly for microbes, fluorescence microscopes are often used, the organisms in the soil having been previously stained with a fluorescent dye. To visualize a specific microorganism or groups of related microorganisms in a soil particle, staining methods with fluorescent gene probes (including FISH), are used (Section 19.5). Microorganisms can also be observed on soil surfaces directly by scanning electron microscopy (Figure 20.13b, c).
Mastering Microbiology
Art Activity: Figure 20.12 Soil bacterial and archaeal diversity
We learned in Chapter 19 that sequence analyses of 16S ribosomal RNA (rRNA) genes obtained from the environment are commonly used as a measure of bacterial and archaeal diversity (Section 19.6). As yet, no natural communities have been so thoroughly characterized by these techniques that all resident species have been identified. However, within limits, the method is widely considered to be a valid measure of microbial diversity and avoids the more serious problems of enrichment bias that plague culture-dependent diversity studies (Section 19.1).
Here and in later sections of this chapter we present “phylogenetic snapshots” of major microbial habitats, with the goal of emphasizing trends, patterns, and ecological rationales rather than comprehensive and exhaustive surveys. And importantly, it should be kept in mind when viewing these phylogenetic summaries that the proportions of each group shown in the diagrams reflect the diversity of that group (expressed as a fraction of the whole) and not necessarily the abundance of the group. Indeed, some environments contain highly diverse groups that are few in number, and vice versa.
Soil Bacterial and Archaeal Diversity
Molecular community sampling of a typical vegetated surface soil has shown typically thousands of different species of Bacteria and Archaea in a single gram of soil, likely reflecting the numerous microenvironments (Figure 20.3) present there. A “species” is defined here as a 16S rRNA gene sequence obtained from a microbial community that differs from all other sequences by more than 3% (Section 13.12). Such an environmental sequence is called a phylotype (also commonly called an operational taxonomic unit, or OTU).
Besides very large species numbers, soil microbial diversity studies have also showed that diversity varies with soil type and geographical location. For example, analysis of an Alaska forest soil, an Oklahoma prairie soil, and a Minnesota farm soil (all sites in the USA) revealed approximately 5000, 3700, and 2000 different phylotypes, respectively. The Alaska and Minnesota soils showed similar distributions at the phylum level of taxonomy (for example, Proteobacteria, Acidobacteria, Bacteroidetes, Actinobacteria, Verrucomicrobia, and Planctomycetes) but shared only about 20% of their species in common. This indicates that although the proportions of the dominant phyla in different soils are relatively constant, the actual species present within a phylum may vary considerably between different soils. In addition, lower bacterial diversity was observed in the farm soil than the Alaska soil, an indication that modern intensive agricultural practices that rely heavily on fertilization, low plant diversity, and the chemical suppression of unwanted plants and animals, reduce bacterial diversity.
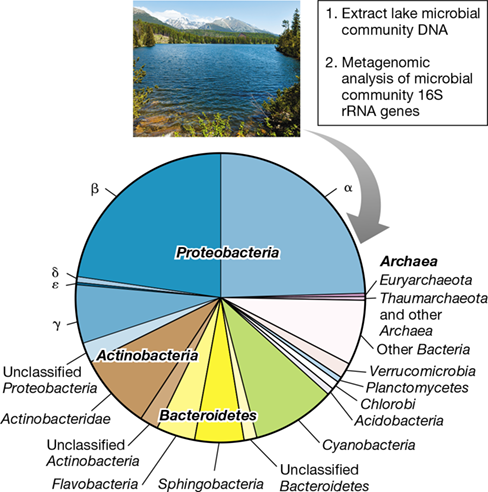
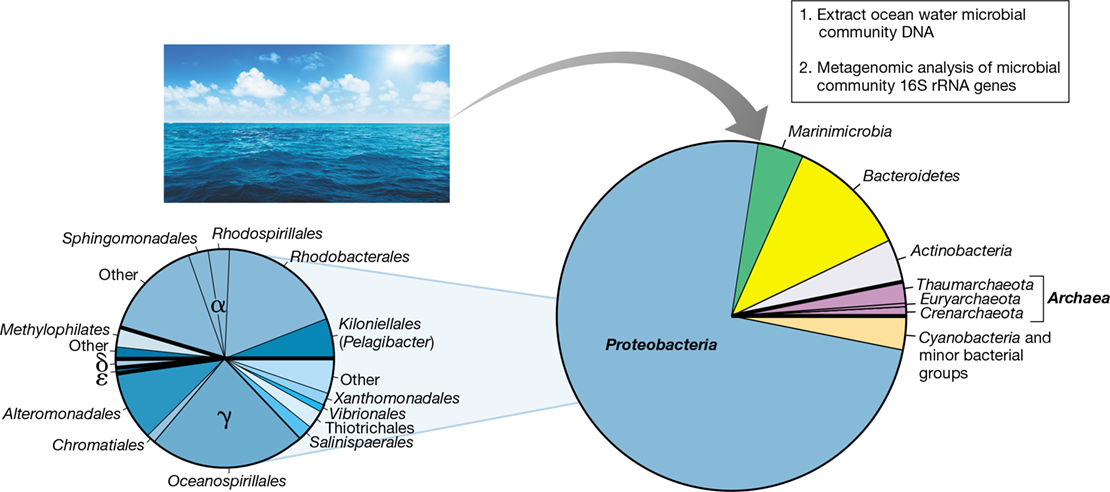
Figure 20.14 shows a diversity snapshot of prokaryotic soil communities based on pooled 16S rRNA sequence data taken from several soils. As can be seen, Proteobacteria (Chapters 15 and 16) make up nearly half of the total phylotypes recovered, with all major subgroups except for Epsilonproteobacteria well represented. The dominance of Proteobacteria in soils is not surprising because the enormous phylogenetic diversity shown by this group is reflected in their enormous metabolic diversity. Phototrophy, chemolithotrophy, and a broad range of chemoorganotrophic metabolisms—both aerobic and anaerobic—are hallmarks of the Proteobacteria (Chapters 14, 15 16). Many pseudomonads, for example, are genetically equipped to use any of over 100 different organic compounds as carbon and energy sources, a logical metabolic strategy for thriving in soils where the timing and nature of organic inputs can be highly variable. Likewise, many species of Rhizobiales can fix N2, others specialize in the use of C1 compounds, and some are even chemolithotrophic or phototrophic, all metabolisms that can be supported in surface soils. In short, the diversity and abundance of Proteobacteria in soils is no surprise.
Figure 20.14 Soil bacterial and archaeal diversity.

The results are pooled analyses from several studies of the 16S rRNA gene content of soil environments. Many of these groups are covered in Chapters 15 and 16 (Bacteria) or 17 (Archaea). For Proteobacteria and Bacteroidetes, major subgroups are indicated. Note high species richness as indicated by the large proportion of the total community composed of unclassified and minor bacterial groups. Also note the relatively low proportion of the total prokaryotic soil diversity represented by Archaea. Data assembled and analyzed by Nicolas Pinel.
Acidobacteria (Section 16.21) and Bacteroidetes (Sections 16.13 and 16.14) are also diverse groups in soil (Figure 20.14), whereas Actinobacteria and Firmicutes are less so. Although relatively few species of Acidobacteria have been cultured, many that have are acid-tolerant and mostly aerobic chemoorganotrophs that collectively use a variety of organic compounds, including plant-derived aromatic compounds, sugars, and polymers common in soils, such as cellulose, xylan, and chitin. Metagenomic analyses (Sections 10.7 and 19.8) are broadening our understanding of Acidobacteria metabolic diversity. Such studies suggest that in addition to a known capacity of some Acidobacteria to grow by anaerobic respiration with ferric iron (Fe3+) as electron acceptor (Section 14.13), other species can reduce nitrite, nitric oxide, sulfate, and sulfite (Sections 14.10, 14.11 and 14.12). Such capacities should serve these organisms well in soils transitioning from oxic to anoxic conditions. Similar to the Acidobacteria, the major groups of soil Bacteroidetes (Sphingobacteria and Flavobacteria) are chemoorganotrophs specialized in the use of complex soil polysaccharides (Section 16.14). Their abilities to hydrolyze these polymers—which are common in soil—and use the monomers generated as electron donors in energy metabolism makes these groups ideally suited to a soil habitat.
In the case of soils, diversity also reflects abundance. Proteobacteria are the most abundant bacteria in most soils followed by Acidobacteria and Actinobacteria. Bacteroidetes, although a highly diverse group in most soils, is not very abundant, averaging about the same number of cells per gram of soil as the Chloroflexi, a group much less diverse than the Bacteroidetes (Figure 20.14).
In contrast to Bacteria, the diversity of Archaea in soil is minimal, with relatively few sequences that group with only two major phyla—the Euryarchaeota and the Thaumarchaeota—commonly observed. But low diversity does not necessarily mean low cell numbers; an archaeal assemblage comprising a small slice of overall diversity can have major biogeochemical importance. For example, species of Thaumarcheota, comprised solely of ammonia-oxidizing Archaea (Section 17.5), are a small part of overall soil diversity (Figure 20.14) but are present in relatively high abundance, greatly outnumbering the ammonia-oxidizing Bacteria in the Nitrosomonadales (Betaproteobacteria). Indeed, Thaumarchaeota, with their characteristic ability to oxidize ammonia present in trace amounts, are now thought to be responsible for the majority of nitrification in most soils (Section 14.9). Nevertheless, when comparing entire domains, the abundance of Bacteria in most soils greatly exceeds that of Archaea by several orders of magnitude.
Polluted Soils
A similar study to that shown in Figure 20.14 but performed on hydrocarbon-polluted soil showed that the general taxonomic makeup of polluted and unpolluted soils is similar: Proteobacteria comprise the largest fraction in both soil types, followed by significant representation of Acidobacteria, Bacteroidetes, Actinobacteria, and Firmicutes. However, there was a significant shift in fractional representation of these taxa in the two soils. Polluted soils are enriched in Actinobacteria, Gammaproteobacteria, and Euryarchaeota but diminished in Bacteroidetes, Acidobacteria, and unclassified Bacteria relative to unpolluted soils. Hydrocarbon-polluted soils contained a single dominant Bacteroidetes phylotype, whereas unpolluted soils contained several phylotypes of Bacteroidetes (Figure 20.14). Notably, Thaumarchaeota are absent from all surveys of hydrocarbon-polluted soils, suggesting that hydrocarbon pollutants suppress ammonia-oxidizing Thaumarchaeota.
Although the functional significance of the observed diversity of microbial communities in polluted versus unpolluted soils is unknown, the shifts observed signal that the two soils will likely differ in their capacity to process carbon and nitrogen and to carry out other important nutrient cycling events. However, despite this lack of a functional connection, different 16S rRNA gene surveys of soils agree on two things: (1) undisturbed, unpolluted soils support very high prokaryotic diversity, and (2) soil perturbations trigger measurable shifts in community composition toward species that are more competitive in the disturbed soil and are accompanied by an overall reduction in diversity.
Climate and Disturbance Effects on Soil Microbes
The importance of understanding the diversity of microbes and microbial processes in soils has been highlighted by the concern about the impact climate change will have on soil systems, including the microbial response to higher temperatures, rainfall patterns, and the rapid changes taking place in land use. From what is known today of microbial and metabolic diversity, climate change will almost certainly impact biogeochemical cycles, and such functional understanding is essential for predicting whether soils will be a sink or a source of atmospheric CO2.
Since cultured diversity from soil is still but a fraction of the diversity that is known to be there, metagenomic analyses combined with 16S rRNA surveys are relied upon to provide insight into the effects of climate and land use changes. For example, a recent metagenomic study of the consequences of deforestation in the Brazilian Amazon revealed distinct microbial community shifts between pasturelands and adjacent rain forests. Acidobacteria (Figure 20.14) were associated exclusively with pasture soils. In contrast, the candidate division Rokubacteria was almost exclusively associated with rain forest soils and displayed distinct metabolic properties, using carbon sources ranging from methane to complex hydrocarbons.
This and other examples of shifts in microbial diversity due to land use changes illustrate the fact that microbial diversity in any environment is a dynamic entity and will change—sometimes dramatically—in response to environmental changes in conditions and resources. Since large-scale changes in microbial activities typically follow large-scale shifts in diversity, future soil problems associated with climate change and land disturbances could be significant. And, since humans rely on soil one way or the other for the majority of their food, it is easy to see why studying the effect on microbes of climate and land changes is an urgent and important problem that will require more intensive cultivation and metagenomic surveys.
Check Your Understanding
How might a microbial group having relatively minor representation in soil diversity still have major biogeochemical importance?
Which three phyla of Bacteria dominate the bacterial diversity of soils? Which phylum of Archaea dominates and how does it obtain energy?
Why should humans be concerned if a climate-induced major shift in soil microbial diversity occurred?
20.8 The Terrestrial Subsurface
Surface soils and subsurface terrestrial environments provide quite different habitats for microorganisms. Microbes in vegetated soils benefit from relatively high levels of nutrients because of their association with growing plants, periodic large inputs of nutrients from dead plants and animals, and inputs of moisture from rain events. By contrast, microbes living below the C horizon (Figure 20.11) have few if any of these advantages. Because of nutrient limitations, microbial diversity and abundance in the deepest terrestrial environments are typically extremely low.
Sampling deep subsurface or marine sediments is expensive and difficult, especially in extracting samples from great depths without contamination from upper subsurface zones. Nevertheless, a number of studies of subsurface soils, deep confined aquifers, and deep marine sediments have been done. Many have focused on the microbial diversity of easily accessible groundwater systems that are within a few hundred meters of the surface. More limited surveys, often requiring more specialized drilling equipment, have examined deep marine sediments and deep confined aquifers.
From research made possible by the development of improved drilling and aseptic sampling technology (Figure 20.15), it is now known that microbial life extends to at least 3000 meters (3 kilometers or 1.9 miles) below the Earth’s surface. Results show that the microbial diversity of relatively shallow subsurface areas is similar to that of surface soils; Proteobacteria, Actinobacteria, Bacteroidetes, and Firmicutes generally predominate (Figure 20.14), but their abundances decrease with depth, likely a result of decreasing organic carbon availability (Figure 20.11a). Less highly represented groups, such as Acidobacteria, Verrucomicrobia, and unclassified lineages, are also commonly encountered. Many unclassified Bacteria and Archaea affiliate with novel phyla and are extremely small and likely grow as symbionts of other microbes (see Figure 20.17). The only group that occasionally shows increases in relative abundance with depth are the Firmicutes, in particular, the Clostridiales. These are anaerobic bacteria capable of forming endospores (Sections 2.8 and 16.8) and thus could be expected to survive long periods of dormancy.
Figure 20.15 Sampling the deep subsurface.

(a) Drilling to 600 m in Allendale, South Carolina (USA), for the U.S. Department of Energy (DOE) Deep Subsurface Microbiology Program. Subsurface microbiology is both expensive and challenging from the standpoint of obtaining uncontaminated samples from deep underground. (b) Sampling hot (55 °C) fissure water from a depth of 3000 m in the Tau Tona South African gold mine.
*Bacteria* in the Deep Subsurface
Research on the deepest microbial biosphere has been facilitated by drilling of deep marine sediments and mining and drilling operations that expose aquifer waters in fractured granitic rock at great depths. For example, samples collected from a nearly 3-km-deep gold-mining operation in South Africa (Figure 20.15b) revealed phylotypes of species of Bacteria and Archaea. A common geochemical feature of deep granitic aquifers is the presence of significant sulfate at depths greater than 1000 meters. The possibility that sulfate respiration could contribute to a subsurface biosphere was confirmed by analysis of DNA extracted from fissure water deep in this mine, showing that a H2-oxidizing, sulfate-reducing bacterium (Sections 14.12 and 15.11) was virtually the only bacterium present. Genome analysis of the organism, as yet uncultured but given the provisional name Desulforudis audaxviator, indicated that it should be moderately thermophilic (the aquifer temperature at this depth was approximately 50 °C) and should be capable of autotrophic growth using H2 as the electron donor for anaerobic respiration and CO2 fixation. In addition, the organism contained genes encoding nitrogen fixation (Section 3.12), meaning that it should be able to live in an anoxic environment on a diet of a few minerals, CO2, SO4 2−, N2, and H2—a lean diet indeed!
D. audaxviator should be well suited to long-term isolation in the deep subsurface, as would other autotrophic and nitrogen-fixing bacteria that could use H2 as electron donor. Possible subsurface sources of H2 include the radiolysis of water by uranium, thorium, and other radioactive elements, and geochemical processes such as the release of H2 from the oxidation of iron silicate minerals in aquifers. H2 can satisfy the electron donor needs of bacteria that carry out many different anaerobic respirations, including sulfate reduction, acetogenesis, ferric iron reduction, and methanogenesis (Chapter 14), and examples of all these physiologies have been identified from metagenomic analyses of subsurface materials. Hence, Bacteria supported by these physiologies undoubtedly inhabit the subsurface microbial ecosystem along with D. audaxviator.
*Archaea* in the Deep Subsurface
As 16S rRNA sequencing is increasingly complemented by environmental genomics (Sections 19.6 and 19.8), additional novel organisms and metabolisms almost certainly await discovery. Notably, although generally in lower abundance than Bacteria, many novel Archaea inhabit the terrestrial subsurface and marine sediments (Section 20.15). As one example, an archaeon affiliated with marine anaerobic methane oxidizers (Section 14.16) is associated with sulfate-dependent methane oxidation in some deep aquifer communities. In addition to archaeal species from phyla having cultivated representatives (Euryarchaeota, Crenarchaeota, Thaumarchaeota, Chapter 17), several phyla so far identified only through PCR-based and metagenomic surveys include the Bathyarchaeota, the Asgard group, and several others (Figure 20.16; Section 17.8). These surveys have also revealed a remarkable diversity of extremely small Archaea 0.15–1.2 μm in diameter (Figure 20.17) that contain small genomes (∼500–1000 genes) and inhabit the subsurface and other nutrient-depleted environments (see Chapter 1 Explore the Microbial World, “Tiny Cells”). These minute Archaea form a single deep evolutionary divergence within the Archaea that encompasses multiple phyla called DPANN (a “superphylum,” Figure 20.16). The first described member of DPANN was the hyperthermophile Nanoarchaeum equitans, a small parasitic species of limited metabolic capacity that grows in obligate physical association with a host archaeon, Ignicoccus (Section 17.6). However, besides the parasitic lifestyle, metagenomics suggest that some DPANN species may be autotrophs and thus less metabolically dependent than Nanoarchaeum.
Figure 20.16 Diversity and abundance of subsurface *Archaea*.

Phylogeny of archaeal clades found predominantly in the terrestrial and marine deep subsurfaces (this is a consensus tree based on 16S rRNA gene sequences and other conserved marker genes). The novel phyla previously unknown until studies of the deep subsurface are highlighted in light blue. Members of the Asgard superphylum are the closest known relatives of eukaryotic cells (Section 13.4).
Figure 20.17 Small *Archaea*.
Electron micrographic section of a cell of a small species of Archaea inhabiting acid mine drainage (Section 22.2). The small Archaea found in this acidic environment is a member of the DPANN superphylum (Figure 20.16). The archaeal cell is approximately 0.4 μm in diameter.
At least two major surprises have emerged from metagenomic studies of the Bathyarchaeota. First was the discovery of genomes from subsurface Bathyarchaeota (Figure 20.16) that contained genes homologous to those encoding methanogenesis and the acetyl-CoA pathway (the mechanisms of energy conservation and autotrophy, respectively, in methanogenic Archaea, Sections 14.14 and 14.15). Previously, methanogenesis was thought to be restricted to a single phylum of Archaea, the Euryarchaeota. Second, genomic analyses indicate that some Bathyarchaeota likely oxidize methane and others may oxidize longer-chain hydrocarbons, all under anoxic conditions (Section 14.24). Metagenomics also tells us that some Bathyarchaeota are heterotrophs, hydrolyzing peptides as carbon and energy sources or degrading polymers such as lignin, a legacy of ancient buried plants.
Other unusual Archaea, such as the Asgard group (Section 17.8; Figure 20.16), have emerged from deep marine subsurface metagenomic analyses. Species in this group appear to be heterotrophs and play a role in carbon degradation in anoxic sediments. Some seem genetically equipped to degrade methane—a large reservoir of which exists in the oceanic subsurface—and other short-chain hydrocarbons, such as butane. As discussed in Section 17.8, there is mounting evidence that the eukaryotic domain of cellular life emerged from within the Archaea, specifically the Asgard superphylum. This phylogenetic connection is supported by criteria other than just ribosomal RNA gene sequences, including the fact that Asgard genomes encode several eukaryotic-like proteins, such as actin (Section 2.15). Thus, analysis of the deep subsurface biosphere has not only expanded the known phylogenetic diversity of Archaea (Figure 20.16) but has revealed new information about the origin of the eukaryotic cell.
Growth Rates and the Future of Subsurface Microbiology
Bacterial numbers in uncontaminated groundwater vary by several orders of magnitude (102–108 per ml), primarily as a function of dissolved organic carbon content. Measured and estimated generation times for deep subsurface bacteria also vary by orders of magnitude, from days to centuries, as determined by the physicochemical environment, the physiology of the resident populations, and nutrient availability. Microbes in the deep subsurface likely originate from sedimentary accumulation of near-surface populations. Increasing nutrient limitation with depth of burial then results in selective death of starving populations, and remaining populations are sustained by organic carbon and other nutrients released from dead cells. Only in scattered regions of higher organic carbon content, such as in ancient coal deposits (Figure 20.41), can higher cell numbers be supported.
The many unanswered questions in subsurface microbiology have encouraged the establishment of permanent science laboratories at great depths in the earth. For example, the Sanford Underground Research Facility in Lead, South Dakota (USA) (2400 m deep), is supported by government and private agencies for research in physics, geology, and microbiology. The International Ocean Discovery Program has probed for microbial populations at great depths below the seafloor. Results thus far have shown Archaea and Bacteria at depths greater than 2000 m below the seafloor (Section 20.15) and in rocks more than 100 million years old. Although this may sound ancient, such ages are actually relatively young compared with viable bacteria that have been recovered from salt crystals dated to nearly a half billion years old. Obviously, microbial cells can remain viable for enormously long periods of time.
We leave soils and the subsurface now to explore the microbial diversity of aquatic environments in the final part of this chapter. Like terrestrial environments, aquatic environments vary considerably. On the one hand, aquatic environments can be nutrient-rich and highly dynamic ecosystems that support large and diverse microbial populations and a wealth of plants and animals. But on the other hand, some aquatic environments provide the same nutrient limitation problems for microbes as is characteristic of the deep subsurface.
Check Your Understanding
What are possible sources of biologically available energy in the terrestrial subsurface?
What is the major environmental factor that controls the abundance and type of cells in the deep subsurface?
How have metagenomic analyses of the Bathyarchaeota and the Asgard group brought new metabolic and evolutionary concepts to the forefront?
IV Aquatic Environments
Marine and freshwater environments host a tremendous diversity of microorganisms and viruses that infect them. Variations in light intensity, pressure, and nutrient availability, along with the effects of human pollution, greatly influence the activities, abundance, and diversity of aquatic microbes.
Freshwater and marine environments differ in many ways including salinity, average temperature, depth, and nutrient content, but both provide many excellent habitats for microorganisms. In this part of the chapter we focus first on freshwater microbial habitats. We then consider two marine environments: (1) coastal and open ocean waters, and (2) the deep sea. Much new information is emerging about marine microorganisms from studies using the molecular tools of microbial ecology, especially genetic stains, microbial community sampling, and metagenomics (Chapter 19).
20.9 Freshwaters
Freshwater environments vary significantly in the resources and conditions (Table 20.1) available for microbial growth because some lakes and streams are isolated and nearly pristine while others are highly polluted from agricultural, industrial, or residential runoff. Both oxygen-producing and oxygen-consuming organisms are present in aquatic environments, and the balance between photosynthesis and respiration (Figure 20.2) controls the natural cycles of oxygen, carbon, and other nutrients (nitrogen, phosphorus, metals).
Among microorganisms, oxygenic phototrophs include algae and cyanobacteria, both of which are primary producers. These organisms can be either planktonic (floating) and distributed throughout the water columns of lakes or benthic, meaning they are attached to the bottom or sides of a lake or stream. The activity and diversity of chemoorganotrophic aquatic microbial communities depend to a major extent on primary production, in particular its rates and temporal and spatial distributions. If primary production rates are very high, the resultant excessive organic matter production can lead to bottom-water O2 depletion from respiration and the development of anoxic conditions. This in turn activates anaerobic metabolisms such as anaerobic respirations and fermentations (Chapter 14).
Oxygen Relationships in Freshwater Lakes
The biological and nutrient structure of lakes is greatly influenced by seasonal changes in physical gradients of temperature and salinity. In many lakes in temperate climates, the water column becomes separated into layers of differing physical and chemical characteristics that constitute a stratified water column. During the summer, warmer and less dense surface layers, called the epilimnion, are separated from the colder and denser bottom layers (the hypolimnion). The thermocline is the transition zone from epilimnion to hypolimnion (Figure 20.18).
Figure 20.18 Development of anoxic conditions in waters of a temperate lake due to summer stratification.

The colder bottom waters are more dense and contain H2S from bacterial sulfate reduction. The thermocline is the zone of rapid temperature transition. As surface waters cool in the fall and early winter, they reach the temperature and density of hypolimnetic waters and sink, displacing bottom waters and effecting lake turnover. Data are from a small freshwater lake in northern Wisconsin (USA).
In the late fall and early winter, lake surface waters become colder and thus more dense than the bottom layers. This, combined with wind-driven mixing, causes the cooled surface water to sink and the lake to “turn over,” mixing surface and bottom waters and their nutrients. The separation of a relatively well-mixed surface layer from a relatively static bottom layer limits the transfer of nutrients between layers until fall turnover once again mixes the water column.
During periods of stratification, transfer between surface and bottom waters is controlled not by mixing but by the much slower process of diffusion. As a result, bottom waters can experience seasonal periods of either low or no dissolved O2. Whether a lake actually becomes O2-depleted depends on several factors, including the amount of organic matter present and the degree of mixing of the water column. Organic matter that is not consumed in surface layers sinks to the depths and is decomposed by anaerobes (Figure 20.2). Lakes may contain high levels of dissolved organic matter because inorganic nutrients that run off the surrounding land can trigger algal and cyanobacterial blooms; these organisms typically excrete various organic compounds and also release complex organic compounds when they die and decay. The combination of water body stratification during early summer, high organic loading, and limited O2 transfer results in O2 depletion of the bottom waters (Figure 20.18), making them unsuitable for aerobic organisms such as plants and animals.
The annual turnover cycle allows the bottom waters of a lake to pass from oxic to anoxic and back to oxic. Microbial activity and community composition is altered with these changes in oxygen content, but other factors that accompany fall turnover of the water column, especially changes in temperature and nutrient levels, govern microbial diversity and activity as well. If organic matter is sparse, as it is in unpolluted lakes or in the open ocean, there may be insufficient substrate available for chemoorganotrophs to consume all the oxygen. The microorganisms that dominate such environments are typically oligotrophs, organisms adapted to growth under very dilute conditions (Section 20.12). Alternatively, where currents are strong or there is turbulence because of wind mixing, the water column may be well mixed, and consequently oxygen may be transferred to the deeper layers.
Oxygen levels in rivers and streams are also of interest, especially those that receive inputs of organic matter from urban, agricultural, or industrial pollution. Rivers and streams are typically flowing systems and thus rarely become strongly stratified, but even in a river well mixed by water flow and turbulence, large organic inputs can lead to a marked oxygen deficit from bacterial respiration (**Figure 20.19*a***). As the water moves away from a point source input, for example, from an input of sewage, organic matter is gradually consumed, and the oxygen content returns to previous levels. As in lakes, nutrient inputs to rivers and streams from sewage or other pollutants can trigger massive blooms of cyanobacteria and algae (Figure 20.1) and aquatic plants (Figure 20.19b), thereby diminishing overall water quality and growth conditions for aquatic animals.
Figure 20.19 Effect of the input of organic-rich wastewaters into aquatic systems.

(a) In a river, bacterial numbers increase and O2 levels decrease with a spike of organic matter. The rise in algae and cyanobacteria is a response to inorganic nutrients, especially PO4 3−. (b) Photo of a eutrophic (nutrient-rich) lake, Lake Mendota, Madison, Wisconsin (USA), showing algae, cyanobacteria, and aquatic plants that bloom in response to nutrients from agricultural runoff. (See also Figure 20.1).
Biochemical Oxygen Demand
The microbial oxygen-consuming capacity of a body of water is called its biochemical oxygen demand (BOD). The BOD of water, such as that from a temperate lake, is determined by taking a sample, aerating it well to saturate the water with dissolved O2, placing it in a sealed bottle, incubating it in the dark (to prevent photosynthetic oxygen production) for 5 days at 20 °C, and determining the residual oxygen in the water at the end of incubation. A BOD determination gives a measure of the amount of organic material in the water that can be oxidized by the microbes present in the water. As a lake or river recovers from an input of organic matter or from excessive primary production, the initially high BOD becomes lower and is accompanied by a corresponding increase in dissolved oxygen in the ecosystem (Figure 20.19a).
In freshwaters, the oxygen and carbon cycles are linked, with the levels of organic carbon and oxygen being inversely related. In other words, high BOD leads to low dissolved oxygen levels. Anoxic aquatic environments, which are typically rich in organic material, are the end result of respiratory processes that remove dissolved oxygen from the ecosystem, leaving the remaining organic material to be mineralized by organisms employing the anaerobic energy metabolisms previously discussed (Chapter 14). It is also important to recognize the importance of storms, floods, and droughts in determining delivery, transport, and cycling of organic matter and inorganic nutrients in freshwater systems, including streams, rivers, lakes, and reservoirs. These less predictable but nonetheless important changes also affect microbial productivity, diversity, distribution, and interactions.
A Phylogenetic Snapshot of Freshwater Prokaryotic Diversity
The importance of Bacteria and Archaea in lakes, streams, and rivers to the production, regeneration, and mobilization of nutrients is well recognized, and, as for soil (Section 20.7), 16S ribosomal RNA gene sequencing has been used as a culture-independent method to identify and quantify phylotypes in aquatic microbial habitats. Since most molecular studies of freshwater systems have explored the diversity in lakes, we focus on lakes here.
Figure 20.20 shows the major prokaryotic groups that inhabit lake surface samples (the epilimnion, Figure 20.18). Five major bacterial phyla are routinely observed: Proteobacteria, Actinobacteria, Bacteroidetes, Cyanobacteria, and Verrucomicrobia. Euryarchaeota, Crenarchaeota, and Thaumarchaeota are the major Archaea present but collectively account for only a small fraction of the total prokaryotic abundance and diversity.
Figure 20.20 Freshwater lake bacterial and archaeal diversity.

Distribution of 16S ribosomal RNA gene sequences by phylum determined from analysis of a collective dataset of 16S genes collected from the epilimnion of several freshwater lakes. Data assembled and analyzed by Nicolas Pinel.
The phylum-level composition of Bacteria in lakes shares features in common with both surface soils and the oceans, environments where Proteobacteria and Bacteroidetes also comprise the greater part of diversity (Figure 20.14 and see Figure 20.31). However, compared with lakes and soils, where Alpha- and Betaproteobacteria show roughly equivalent diversity, we will see in Section 20.12 that in the oceans, Alpha- and Gammaproteobacteria predominate and Betaproteobacteria are only minor components. Betaproteobacteria tend to be fast-growing species that respond quickly to periodic pulses of organic nutrients (for example, leaf litter) that enter lake systems. By contrast, Alphaproteobacteria tend to dominate under the more nutrient-limiting conditions of the open ocean.
In addition to leading in the diversity category, Proteobacteria are also the most abundant bacteria in most lake systems, with Actinobacteria and Bacteroidetes species being significant but in lesser abundance. Interestingly, Acidobacteria, major components of both diversity and abundance in soils (Figure 20.14), are but a tiny fraction of bacterial diversity and abundance in freshwater lakes.
With the exception of the Acidobacteria, it is likely that a similar rationale exists for why the major bacterial phyla found in soil (Figure 20.14) are also found in freshwater: metabolic diversity. Proteobacteria excel in this realm and Bacteroidetes are well known for their degradation of various biopolymers and aromatic compounds, such as humic materials. Actinobacteria are chemoorganotrophs that can also break down polymers, in particular nucleic acids and proteins. In addition, metagenomic analyses (Section 19.8) have shown that at least some Actinobacteria contain genes that encode a bacteriorhodopsin-like protein, similar to that which converts light energy into ATP in extremely halophilic Archaea (Section 17.1); thus, these Actinobacteria may be phototrophic. Moreover, many anoxygenic phototrophs are Proteobacteria (Sections 15.4 and 15.5), and their competitive advantage in freshwaters would also rely on photosynthesis. A rationale for cyanobacteria in overall lake diversity (Figure 20.20) hardly needs mentioning, as oxygenic photosynthesis requires nothing but a few inorganic nutrients, CO2, and plenty of sunshine. Cyanobacterial abundance can also be quite high in freshwater lakes, especially if the lake receives inputs of key inorganic nutrients such as phosphorus and nitrogen; these nutrients often trigger massive cyanobacterial blooms (Figures 20.1 and 20.19b).
Collectively, the high prokaryotic diversity observed in freshwater lakes (Figure 20.20) reflects the dynamic character of these habitats. Lakes typically receive seasonally variable inputs of endogenous and exogenous nutrients, a pattern that sustains a phylogenetically and metabolically complex community of Bacteria and a few groups of Archaea.
Check Your Understanding
What is a primary producer? In a freshwater lake, would primary producers such as cyanobacteria more likely reside in the epilimnion or the hypolimnion, and why?
Will addition of organic matter to a water sample increase or decrease its BOD?
What factor(s) might account for the prokaryotic diversity of freshwater lakes?
20.10 Oxygen Relationships in the Marine Environment
20.10 Oxygen Relationships in the Marine Environment
20.10 Oxygen Relationships in the Marine Environment
With the exception of oxygen, nutrients in the open ocean (called the pelagic zone) are typically at very low levels compared with lakes and other freshwaters. This is especially true of key inorganic nutrients for phototrophic organisms, such as nitrogen, phosphorus, and iron. In addition, water temperatures in the oceans are cooler and more constant seasonally than those of most freshwater lakes. The activity of marine phototrophs is limited by these factors, and thus total microbial cell numbers are typically about 10-fold lower in the oceans than in freshwater environments (∼106/ml versus 107/ml, respectively). These are average numbers, and studies of marine prokaryotic diversity are just beginning to reveal recurrent temporal patterns of diversity and abundance.
Tracking Seasonal Changes in Marine Microbial Composition
The Bermuda Atlantic Time-Series Study has a history of continuous biogeochemical monitoring of ocean waters since the mid-1950s, and since the dawn of the molecular ecology era, it has also incorporated diversity studies of microbial population structure. These studies have revealed three major microbial communities in ocean waters that follow a seasonal pattern: (1) the community corresponding to the spring surface water bloom, consisting of small eukaryotic algae, marine Actinobacteria, and two groups of Alphaproteobacteria; (2) the summertime community in the upper water column associated with water column stratification, consisting of Pelagibacter, Puniceispirillum, and two groups of Gammaproteobacteria; and (3) the more stable deep-water community, consisting of Nitrosopumilus, representatives of the SAR11 group with which the genus Pelagibacter (see Figure 20.30) affiliates, a group of Deltaproteobacteria, and species of two additional groups related to the Chloroflexi and Fibrobacter. Except for Nitrosopumilus (domain Archaea), all other deep-water microbes are Bacteria (Chapter 16). In Section 20.11, we will return to a more specific breakdown of prokaryotic cell diversity in the open oceans.
Most pelagic Bacteria and Archaea are very small cells, a characteristic often seen in organisms that inhabit nutrient-poor (oligotrophic) environments. Smallness is an adaptive feature for nutrient-limited microorganisms in that it requires less energy for cellular maintenance and yields a larger surface-to-volume ratio in the cell (Section 1.3). As a consequence of their oligotrophic lifestyle, the genomes of pelagic microbes typically encode very high-affinity transport systems for acquiring the nutrients they need.
Microbial Activities in Marine Environments and Oxygen Minimum Zones
In pelagic waters there is a lower return of nutrients from the bottom waters to upper waters than in freshwater lakes because the waters do not completely turn over seasonally; thus, average primary productivity in the pelagic photic zone is less than in a lake. However, because the oceans are so large, the collective carbon dioxide sequestration and oxygen production from oceanic photosynthesis are major factors in Earth’s carbon balance. Terrestrial runoff, retention of nutrients, and upwelling of nutrient-rich waters combine to support higher populations of phototrophic microorganisms in near-shore waters than in pelagic waters (Figure 20.21); these more productive near-shore waters in turn support higher densities of chemoorganotrophic bacteria and aquatic animals, such as fish and shellfish.
Figure 20.21 Distribution of chlorophyll in the western North Atlantic Ocean as recorded by satellite.

The eastern coastline of the United States from the Carolinas to northern Maine is shown in dotted outline. Areas rich in phototrophic plankton are shown in red (chlorophyll concentrations of >1 mg/m3); blue and purple areas have lower chlorophyll concentrations (<0.01 mg/m3). Note the high primary productivity of coastal areas and the Great Lakes.
In shallow marine waters such as marine bays and inlets, eutrophication resulting from nutrient inputs can lead to the waters becoming intermittently anoxic from the removal of O2 by respiration and the production of H2S by sulfate-reducing bacteria. This can lead to oxygen minimum zones (OMZs), regions of oxygen-depleted waters at intermediate depths, typically in waters between 100 and 1000 m, that extend over wide expanses of the open and coastal ocean (Figure 20.22). These oxygen-depleted regions arise when the respiratory demand for oxygen exceeds oxygen availability, and they are associated with nutrient-rich, highly productive regions. In this way they are similar to the depletion of oxygen caused by agricultural runoff in coastal zones (Figure 20.22). However, OMZs predate human activity and originate naturally in regions of high surface production and little mixing with oxygen-rich water.
Figure 20.22 Diversity of marine systems and associated microbial metabolic processes.

Decreasing electron acceptor availability with depth into the sediment or with increasing distance into an oxygen minimum zone is indicated by red wedges. Sulfate becomes limiting only at greater depths in marine sediments.The indicated metabolic diversity is described in Chapter 14.
The oxygen saturation values of the largest of the OMZs in the eastern Pacific off the coast of Peru are less than 10% of that at the surface, but in some other pelagic waters the values actually reach zero. Because of this, OMZs have been recognized as significant sinks for the loss of fixed nitrogen through denitrification (Section 14.11) and anammox (Section 14.10), and as a significant source of nitrous oxide (N2O), a potent greenhouse gas (Section 21.9; Figure 20.22). Ongoing studies of OMZs have shown that these regions are expanding, and that their recent expansion is almost certainly associated with global warming. As the oceans absorb more heat, warming of the surface waters increases stratification of near-surface waters and reduces oxygen transfer through mixing to deeper regions. Expansion of the OMZs will favor anaerobic microbial processes at the expense of the aerobic processes that sustain critical oceanic food webs. Ultimately, these changes are expected to impact commercial fisheries.
Oxygen-Depleted Marine Dead Zones and the Deepwater Horizon Catastrophe
An extensive region (6000–8000 square miles) of seasonal oxygen depletion in the Gulf of Mexico is associated with high loads of nitrogen and phosphorus carried in by the Mississippi River from agricultural runoff in the Mississippi Valley. This input triggers excessive oxygen consumption from chemoorganotrophs and the formation of hypoxic (oxygen-depleted) water. This region, called the Gulf of Mexico Dead Zone, contributes to the loss and impairment of fish and benthic sea life that sustain major seafood industries in this region. In addition to the chronic degradation of the Gulf of Mexico ecology through agricultural runoff, increased offshore oil drilling also poses significant environmental risk. A major catastrophe for the Gulf of Mexico was the April 2010 explosion and sinking of the Deepwater Horizon offshore drilling platform; failure to control well pressure resulted in the rupture of the wellhead at a depth of 1.5 km and the release of over 4 million barrels of oil before the well was capped three months later (Figure 20.23).
Figure 20.23 The Deepwater Horizon oil spill in the Gulf of Mexico.

(a) Inferno resulting from the wellhead blowout. (b) NASA Terra satellite image taken on May 24, 2010, of the Gulf of Mexico near New Orleans, Louisiana. A large plume of oil was released at about 1500-m depth, some of which reached the surface where sunlight reflects off of the oil slick (arrows).
This largest marine oil spill ever was unique in that most of the oil was released as a plume at great depths in the water column. Typically, marine oil spills contaminate surface waters, resulting in rapid volatilization and loss to the atmosphere of low-molecular-weight oil components such as naphthalene, ethylbenzene, toluene, and xylene. By contrast, the Deepwater Horizon spill released both low-molecular-weight components and natural gas (methane, ethane, propane) deep into the water column. These components comprised about 35% of the hydrocarbon plume that extended across many miles of the Gulf from the surface to depths greater than 800 m (Figure 20.23b) and offered an opportunity for microbiologists to measure the effect of a large input of oil to marine waters.
The microbial response to the Gulf spill was tracked over several months using both culture-based and molecular methods, including 16S ribosomal RNA gene and metagenomic sequencing, and GeoChip microarray analyses (Sections 19.6, 19.7 and 19.8). Hydrocarbon-degrading bacteria were already enriched in Gulf waters as a consequence of natural seepage of petroleum and natural gas from sediments in coastal regions rich in petroleum reserves (Figure 20.24). These were hydrocarbon-degrading Gammaproteobacteria species related to genera in the Oceanospirillales group, and of the genera Colwellia and Cycloclasticus. Microbial diversity and abundance measurements showed that the initial response to the oil spill in May and June of 2010 was a bloom of these very species, probably because of their ability to catabolize gaseous hydrocarbons released from natural seeps (Figure 20.24). This was confirmed with enrichment cultures (Section 19.1) that grew rapidly when ethane or propane was the substrate. Colwellia species also contributed to the degradation of a variety of other hydrocarbons, as indicated by their growth in crude-oil enrichment cultures lacking natural gas and by stable isotope probing experiments (SIP, Section 19.10) that showed incorporation of 13C benzene. Other oil-degrading Bacteria were detected in the Gulf spill as well. In studies of oil consumption on beach sand, a succession was observed in the microbial community during the course of oil consumption as lighter fractions were consumed and more refractory components remained for specialist organisms to consume.
Figure 20.24 Marine petroleum seep.

(a) Methane and CO2 are produced by microbial degradation of marine subsurface oil in the Coal Oil Point seep field and migrate through overlying geological strata to be released 21 m underwater in the Santa Barbara Channel off the California Coast. The bubbles also contain small amounts of additional hydrocarbons and H2S from sulfate reduction. Continuous release of methane and trace hydrocarbons to the water maintain populations of hydrocarbon-degrading microbes capable of responding to accidental petroleum releases. (b) Approximately half the released methane dissolves into the water and becomes available to the microbial community.
Although there remains uncertainty about the fate of all the hydrocarbons released during the Deepwater Horizon spill, it appears that the early stimulation of hydrocarbon-degrading bacteria by the more soluble, low-molecular-weight gaseous components in the oil reduced the environmental impact of this immense oil spill. Nevertheless, billions of dollars in losses to commercial fisheries and other human activities in the Gulf of Mexico and significant losses of wildlife and wildlife habitat occurred because of the spill.
Check Your Understanding
Why are marine prokaryotic cells typically quite small?
What is an oxygen minimum zone and why is expansion of these zones a problem for marine and global ecology?
What did the Gulf oil spill tell us about how mixed hydrocarbons are degraded in nature?
20.11 Major Marine Phototrophs
The oceans contain large numbers of phototrophic microbes, including both prokaryotic and eukaryotic oxygenic phototrophs as well as significant numbers of a special group of purple (anoxygenic) phototrophs. We consider these organisms here as a prelude to exploring the marine prokaryotic world in general in Section 20.12.
Primary Productivity: *Prochlorococcus*
Much of the primary productivity in the open oceans, even at significant depths, comes from photosynthesis by prochlorophytes, tiny bacterial phototrophs that are phylogenetically cyanobacteria (Section 15.3). However, unlike most cyanobacteria, prochlorophytes do not contain phycobilins, although they do contain chlorophylls a and b. The organism Prochlorococcus is a particularly important primary producer in the marine environment (Figure 20.25). Because Prochlorococcus lacks phycobilins, which are the accessory pigments of the cyanobacteria (Section 14.4), dense suspensions of Prochlorococcus cells are olive green (as are green algae) rather than the blue-green color of cyanobacteria (compare Figures 20.1c and 20.25).
Figure 20.25 *Prochlorococcus*, the most abundant oxygenic phototroph in the oceans.

A bottle of Prochlorococcus showing the olive-green color of the cells containing chlorophylls a and b. Inset: FISH-stained cells (Section 19.5) of Prochlorococcus in a marine water sample.
Prochlorococcus accounts for up to half of the photosynthetic biomass and primary production in the tropical and subtropical regions of the world’s oceans, reaching cell densities of 105/ml. A number of strains of Prochlorococcus have now been identified in culture, and each inhabits its own depth range in pelagic waters. The different Prochlorococcus strains are considered distinct ecotypes, genetic variants of a species that differ physiologically and therefore occupy slightly different niches. For example, different Prochlorococcus ecotypes photosynthesize at different light intensities (high-light versus low-light ecotypes) and use different inorganic and organic nitrogen and phosphorus sources. Prochlorococcus is thus distributed in both surface waters and deeper waters to depths of 200 m, and when an oxygen minimum zone (Section 20.10) is present, Prochlorococcus extends into the upper regions of this zone (Figure 20.22). This is near the bottom of the photic zone where light intensities are very low (see Figure 20.29).
Genome sequences of about a dozen cultured Prochlorococcus strains revealed that although each contains about 2000 genes, only about 1100 genes (the core genome, Section 13.10) are shared by all strains. Each presumptive ecotype contains approximately 200 unique genes, which likely have adaptive significance for growth in the realized niche of that ecotype. This was illustrated in Chapter 19 where we compared the genome of a single cultured Prochlorococcus ecotype to metagenome sequences obtained from pelagic waters (Section 19.8 and Figure 19.24).
Single-cell genomics (Section 10.11) of natural Prochlorococcus populations have refined our understanding of genetic diversity and the relationship between environmental conditions and the fitness of individual genotypes. These analyses revealed a high degree of fine-scale genetic diversity within the high-light (HL) and low-light (LL) ecotypes. Each ecotype is actually composed of hundreds of subpopulations, each of which is united by a common set of core genes encoding functions that control the interaction of the organism with its environment (e.g., transport functions, oxidative stress responses, and cell surface structure) (**Figure 20.26*b***). However, each subpopulation also contains a small set of genes termed “flexible” genes (ranging from 4 to 14 genes per characterized genome) that vary within that subpopulation. The flexible genes are packaged as “cassettes” localized to highly variable regions (genomic islands) within the Prochlorococcus genome (Sections 13.10, 19.8, and Figure 19.24). Variation in flexible gene content contributes to exceptionally high microdiversity within natural populations of Prochlorococcus, pointing to tremendous versatility in their adaptive response to new niche opportunities. The importance of genotypic variability in sustaining large numbers of this major marine phototroph is shown by seasonal shifts in different genotypes in response to changing light intensity, nutrient availability, and predator populations (Figure 20.26a).
Figure 20.26 Seasonal variation of *Prochlorococcus* ecotypes in marine surface waters.

(a) Single-cell genome sequencing (Section 10.11) has shown that the abundance of different genotypes, presumptive ecotypes (represented by cell color and surface features), correlate with seasonal changes in temperature, light, nutrients, and predators (grazers and bacteriophages). (b) Adaptations to different physical conditions (light and temperature) partition at higher taxonomic levels than do adaptations to different nutrient conditions (e.g., nitrogen availability) and resistance to specific predators, which show closer genetic relationships among ecotypes (see also Figure 19.24).
Mastering Microbiology
Art Activity: Figure 20.23 Seasonal variation of Prochlorococcus ecotypes in marine surface waters
Other Pelagic Oxygenic Phototrophs
In tropical and subtropical oceans, the planktonic filamentous marine cyanobacterium Trichodesmium (**Figure 20.27*a***) is a widespread and occasionally abundant phototroph. Cells of Trichodesmium form colonies of filaments called puffs. Each puff can contain many hundreds of individual filaments, each filament composed of 20–200 cells. In the Caribbean Sea surface waters, colonies of Trichodesmium can approach 100/m3. Trichodesmium is a nitrogen-fixing cyanobacterium, and the production of fixed nitrogen by this organism is thought to be an important link in the marine nitrogen cycle. Trichodesmium contains phycobilins, absent from prochlorophytes, and thus differs from these organisms in its absorption properties (Section 14.4).
Figure 20.27 *Trichodesmium* and *Ostreococcus*.

(a) Light photomicrograph of a puff of cells of the nitrogen-fixing cyanobacterium Trichodesmium. The filaments in the puff are chains of cells, each of which is about 6 μm in diameter. (b) Transmission electron micrograph of a cell of Ostreococcus, a small green alga found primarily in marine coastal waters. The arrow points to the chloroplast. An Ostreococcus cell is about 0.7 μm in diameter.
Very small eukaryotic phototrophs also inhabit coastal and pelagic waters, and some of these are among the smallest eukaryotic cells known. Three common genera—Bathycoccus, Micromonas, and Ostreococcus—are so small that they contain only one mitochondrion and one chloroplast per cell (a human cell can have hundreds or even thousands of mitochondria). These genera belong to the family Prasinophyceae, a group of green algae that diverged early from other lineages of green algae (Section 18.16). Cells of Ostreococcus are cocci that measure only about 0.7 μm in diameter (Figure 20.27b), which is even smaller than a cell of Escherichia coli.
Although cells of the eukaryotic Ostreococcus and the prokaryotic Prochlorococcus are of roughly the same dimensions and they are both oxygenic phototrophs, their genomes are distinct. The genome of Ostreococcus is 12.6 Mbp (distributed over 20 chromosomes), which is more than seven times the size of the Prochlorococcus genome. Even though this is large relative to cyanobacteria, the Ostreococcus genome is unusual for that of a eukaryote in being very gene dense, containing about 8000 genes, and is thought to be near the minimum genome size of a free-living phototrophic eukaryote. By comparison, the genome of a common plant, Japanese rice (Oryza sativa subsp. japonica), is 420 Mbp and contains about 50,000 genes.
In many marine waters, other small eukaryotic cells are present at about 104/ml. Although many of these are Ostreococcus or relatives, some are chemoorganotrophs and some are phototrophs unrelated to Ostreococcus that incorporate small amounts of organic matter to supplement their primarily phototrophic lifestyle.
Aerobic Anoxygenic Phototrophs
Besides oxygenic phototrophs, anoxygenic phototrophs are also present in coastal and pelagic marine waters. Like purple anoxygenic phototrophs, these organisms contain bacteriochlorophyll a (Sections 14.3, 14.5, 15.4, and 15.5). However, unlike classical purple bacteria that carry out photosynthesis only under anoxic conditions, these anoxygenic phototrophs carry out photosynthetic light reactions only under oxic conditions.
Aerobic anoxygenic phototrophs include bacteria such as Erythrobacter, Roseobacter, and Citromicrobium (Figure 20.28), all genera of Alphaproteobacteria (Section 16.1). Aerobic anoxygenic phototrophs synthesize ATP by photophosphorylation when oxygen is present (which is all of the time in oxic pelagic waters), but they are unable to grow autotrophically and thus rely on organic carbon for their carbon sources (a nutritional condition called photoheterotrophy). These organisms thus use the ATP produced by photophosphorylation to supplement their otherwise chemoorganotrophic metabolism.
Figure 20.28 Aerobic anoxygenic phototrophic bacteria.
Transmission electron micrograph of negatively stained cells of Citromicrobium. Cells of this marine, aerobic anoxygenic phototroph produce bacteriochlorophyll a only under oxic conditions and divide by both budding and binary fission, yielding morphologically unusual and irregular-shaped cells.
Surveys have shown that a great diversity of aerobic anoxygenic phototrophs exist in marine waters, especially near-shore waters. Oligotrophic and highly oxic freshwater lakes are also habitats for these interesting phototrophic bacteria. The physiology of aerobic anoxygenic phototrophs is thus ideal for their illuminated and highly oxic habitats.
Check Your Understanding
How does Ostreococcus differ from Prochlorococcus? What do they have in common?
How does the organism Prochlorococcus contribute to both the carbon and oxygen cycles in the oceans?
How does Roseobacter differ from Prochlorococcus?
20.12 Pelagic *Bacteria* and *Archaea*
Despite vanishingly low nutrient levels, significant numbers of Bacteria and Archaea live a planktonic existence in pelagic marine waters. Of these, one species in particular has garnered significant attention, a bacterium named Pelagibacter.
Distribution and Activity of *Archaea* and *Bacteria* in Pelagic Waters
The abundance of prokaryotic cells in the open oceans decreases with depth. In pelagic waters, the photic zone extends to a depth of about 200 m, and within this region cell numbers are the highest. In surface waters, cell numbers average about 106/ml. Below 1000 m, however, total cell numbers fall to between 103 and 105/ml. The distribution of Bacteria and Archaea with depth has been tracked in pelagic waters using fluorescence in situ hybridization (FISH) technology (Section 19.5). Species of Bacteria tend to predominate in waters above 1000 m, although cells of Bacteria and Archaea are found in near-equal abundance in deeper waters (Figure 20.29). Deep-water Archaea are almost exclusively species of Thaumarchaeota (Section 17.5), and many, perhaps even most, are ammonia-oxidizing chemolithotrophs (Sections 14.9 and 17.5) that play an important role in coupling the marine carbon and nitrogen cycles (Chapter 21). Extrapolating to a global basis using the data in Figure 20.29, it is estimated that 1.3×1028 and 3.1×1028 cells of Archaea and Bacteria, respectively, exist in the world’s oceans. This means that the oceans contain the largest microbial biomass on Earth’s surface.
Figure 20.29 Distribution of *Archaea* and *Bacteria* in North Pacific Ocean water.

(a) Percentage of Archaea and Bacteria with depth. (b) Approximate absolute numbers of Archaea and Bacteria per milliliter with depth in the open ocean.
Pelagic Bacteria and Archaea are ecologically important because they consume dissolved organic carbon in the oceans, one of the largest pools of usable organic carbon on Earth. These small and free-living planktonic microbes consume about half the total oceanic organic carbon produced from photosynthesis and are responsible for about half of all marine respiration and nutrient regeneration. Planktonic marine microbes thus return organic matter to the marine food web that would otherwise be lost because of the inability of larger marine organisms to take up such diluted organic nutrients. This so-called “secondary production” (primary production being that from photosynthesis) is balanced by cell losses from grazing protists and from virus attack (see Figures 20.32, 20.33, and 20.34). This leads to a near-steady state in which the abundance of prokaryotic cells in the open oceans remains roughly constant over time. But importantly, secondary production allows some of the dissolved organic carbon in seawater to reach larger organisms, including fish, because microbes are eventually passed up the food web by the feeding activities of larger organisms.
*Pelagibacter*: The Most Abundant Bacterium
Small planktonic chemoorganotrophic bacteria inhabit pelagic marine waters in numbers of 105–106 cells/ml. The most abundant of these are members of the “SAR11 group” within the Alphaproteobacteria (Section 16.1) that includes the genus Pelagibacter. Environmental metagenomic studies (Sections 10.7 and 19.8) and cell counts done using FISH (Section 19.5) have revealed a great abundance of SAR11 group organisms in pelagic waters. The total oceanic population of this group is estimated to be about 2.4×1028 cells—about three-quarters of all Bacteria—making it the most successful microbial group, as reflected by abundance, on the planet. Pelagibacter is an oligotroph, an organism that grows best at very low concentrations of nutrients and grows in laboratory culture only up to the densities it is found in nature.
What makes Pelagibacter so successful in the open oceans? In part, its success is related to small size. Cells of Pelagibacter are small rods with a diameter of only 0.2–0.5 μm and a volume of just 0.01 μm3, making cells difficult to resolve with the light microscope but not the electron microscope (Figure 20.30). The resulting high surface-to-volume ratio (Section 1.3 and Figure 1.7) facilitates nutrient transport, increasing substrate concentration and processing rates within the cell. Proteomic analyses (Section 10.9) of Pelagibacter have also revealed a high abundance of periplasmic binding proteins (Section 2.2) for soluble nutrients such as phosphate, amino acids, and sugars. Another adaptive feature of Pelagibacter is its fairly small genome (1.3 Mbp). Consistent with the proteome analysis, the genome encodes an unusually high number of ABC-type transport systems—transporters that have an extremely high affinity for their substrates (Section 2.2)—and other enzymes useful for an oligotrophic lifestyle. The Pelagibacter genome is also highly “streamlined,” with intergenic spacings averaging only 3 base pairs; such a highly compact genome reduces the cost of replication.
Figure 20.30 *Pelagibacter*, the most abundant bacterium in the ocean.

Electron micrograph taken by electron tomography, a technique that introduces a three-dimensional effect. A single cell of Pelagibacter is about 0.2 μm in diameter.
Rhodopsins in Marine Microbes
The Pelagibacter genome also contains genes encoding a form of the visual pigment rhodopsin that can convert light energy into ATP. In Section 17.1 we discussed the now well-studied molecule bacteriorhodopsin, a light-activated protein complex present in the extreme halophile Halobacterium (Archaea). Bacteriorhodopsin functions in ATP synthesis as a simple light-driven proton pump; protons pumped across the membrane by this system fuel ATPase in the membrane, generating ATP (Figure 17.4). The form of rhodopsin in Pelagibacter and other pelagic Bacteria is structurally similar to bacteriorhodopsin and has been called proteorhodopsin (“proteo” referring to Proteobacteria). Although proteorhodopsin was first discovered in species of Proteobacteria, it is actually fairly widely distributed in Bacteria; the system is found in many Gamma- and Alphaproteobacteria, Bacteroidetes, and Actinobacteria (all covered in Chapter 16), and also in nonhalophilic species of Archaea, such as species of the marine Euryarchaeota that inhabit the photic zone. The different variants of proteorhodopsins in marine microbes have absorption properties that reflect changing spectral properties of light at increasing depths in the water column, with near-surface variants absorbing green light and those at greater depths absorbing blue light.
Experiments have shown that proteorhodopsin-containing marine bacteria survive starvation better in the light than in the dark. This indicates that energy-starved cells use light-mediated ATP production to compensate for energy unavailable from carbon respiration when organic carbon levels are low. Proteorhodopsins are thought to exist in ∼80% of bacteria in some marine waters and are thus a widespread strategy to supplement the energy metabolism of marine microbes such that they need not rely solely on scarce organic carbon for their energy needs. Metagenomics has shown that a variant of proteorhodopsin called heliorhodopsin is widely distributed among marine Archaea, Bacteria, and Eukarya, many of which were previously unsuspected to sense light. However, unlike the energy-generating proteorhodopsins, heliorhodopsins appear to function only as a photosensor, most likely assisting in cellular positioning at an optimal location in a light field that would benefit light-mediated energy-generating systems, such as rhodopsins or chlorophylls.
A Phylogenetic Snapshot of Marine Bacterial and Archaeal Diversity
Several studies have documented the diversity of planktonic marine Bacteria and Archaea from sequences of 16S ribosomal RNA genes obtained from seawater. The existence of abundant alphaproteobacterial populations with which Pelagibacter is affiliated was first revealed by such methods. Mesophilic Archaea subsequently shown to be related to ammonia-oxidizing Nitrosopumilus maritimus (Section 17.5) were also first observed using this approach.
Major bacterial groups now recognized as abundant in the ocean include Alpha- and Gammaproteobacteria, Cyanobacteria, Bacteroidetes, and to a lesser extent Marinimicrobia and Actinobacteria; Firmicutes are only minor components. Long-term studies of oceanic bacterial and archaeal population dynamics using molecular methods (Section 19.6) have provided an unprecedented view of microbial diversity in marine systems. A six-year seasonal sampling study at a coastal site (near Plymouth, England) is one such example (Figure 20.31). Notably, Cyanobacteria, which include the ecologically important and relatively abundant phototroph Prochlorococcus, comprise only about 1% of diversity in these coastal waters because of competition from eukaryotic phytoplankton (including diatoms and green algae). In contrast, Cyanobacteria—and Prochlorococcus in particular—dominate the nutrient-limited open ocean.
Figure 20.31 Coastal ocean bacterial and archaeal diversity.

The results are pooled analyses of nearly one million 16S ribosomal RNA sequences from seasonal sampling near the coast of Plymouth (United Kingdom) over a six-year period. Many of these groups are covered in Chapters 15 and 16 (Bacteria) or 17 (Archaea). For Proteobacteria, major subgroups are indicated. Note the high proportion of Alphaproteobacteria and Gammaproteobacteria sequences. Data assembled and analyzed by Nicolas Pinel. Compare the prokaryotic diversity of seawater with that of freshwater shown in Figure 20.20.
Proteobacteria, in particular the alpha and gamma classes, clearly dominate the microbial diversity of the oceans. Many marine Alpha- and Gammaproteobacteria are chemoorganotrophs that metabolize dimethylsulfoniopropionate (DMSP; Table 4.6), including the alphaproteobacterium Ruegeria pomeroyi and species of the gammaproteobacterial genus Marinomonas. DMSP is a soluble organic-sulfur compound produced by marine phytoplankton (microscopic eukaryotic phototrophs) that functions primarily as a compatible solute to control osmotic tension in the cell (Section 4.15). However, much of this DMSP gets excreted, and bacteria degrade it, forming the volatile compound dimethyl sulfide (DMS). This gas has a significant effect on Earth’s climate. As the single most important natural source of sulfur released to the atmosphere, DMS becomes photooxidized to sulfuric acid in the atmosphere, and this promotes water condensation and the formation of clouds. The latter deflect sunlight back into space and help cool our planet (Sections 21.4 and 21.9).
R. pomeroyi is a nonphototrophic member of the Roseobacter lineage within the Rhodobacterales (Section 16.1), an assemblage that also includes many aerobic purple bacteria (Section 15.5) and composes up to a third of the bacterioplankton in coastal waters and a few percent in open ocean surface waters. R. pomeroyi and relatives are often the dominant taxa to develop during blooms of DMSP-producing phytoplankton, pointing to the importance of this bacterium in generating DMS and influencing atmospheric chemistry.
As is the case for soil microbial diversity (Figure 20.14), a large group of Bacteroidetes species and unclassified and minor bacterial groups are also present in seawater. A major group of marine Gammaproteobacteria is the yet-to-be-cultured “SAR86 group.” Although accounting for approximately 10% of the total prokaryotic community in the open oceans’ surface layer, this group is only poorly represented in higher-nutrient coastal waters (Figure 20.31). The candidate bacterial phylum Marinimicrobia is widely distributed in the ocean, including coastal and pelagic surface waters, oxygen minimum zones (Figure 20.22), and depths greater than 6000 meters. Metagenomic analysis of this phylum has revealed tremendous flexibility in the use of electron acceptors other than oxygen to support anaerobic respirations, including sulfoxides, nitrous oxide, polysulfides, and nitrate, and some species can oxidize thiosulfate as an electron donor. With the exception of the cyanobacteria, most marine Bacteria are likely chemoorganotrophs adapted to extremely low nutrient availability in the open ocean, with some augmenting energy conservation through proteorhodopsin or aerobic anoxygenic phototrophy (Section 20.11).
Archaea in pelagic waters comprise a rather restricted diversity of Euryarchaeota, Crenarchaeota, and Thaumarchaeota, most of which have not yet been brought into laboratory culture. The isolation of Nitrosopumilus maritimus, affiliated with the Thaumarchaeota, revealed that, although not highly diverse (Figure 20.31), ammonia-oxidizing chemolithotrophic Archaea are also among the most abundant marine microbes, comprising 20–40% of all marine prokaryotic cells beneath the photic zone. Although species of Euryarchaeota can also be abundant in the upper ocean, comprising 4–20% of the prokaryotic community in surface waters, none have thus far been cultured. However, based on metagenomic analyses, most encode proteorhodopsin, suggesting they grow primarily as photoheterotrophs.
“Dilution culture” and single cell isolation methods employing very dilute culture media have successfully brought some pelagic microbes into culture (Sections 19.2 and 19.3). It appears that most of these organisms have evolved to grow only at very low nutrient concentrations and very low cell densities (105−106/ml), so it is either difficult or impossible to generate large amounts of cell material in the laboratory. This limitation renders many of the common tools for measuring cell growth (turbidity or microscopic counts) useless on samples that are not first concentrated. Nevertheless, there have been notable successes with dilution culturing of marine bacteria, and the aforementioned bacterium Pelagibacter is perhaps the best example of this (Figure 19.6).
Check Your Understanding
What is proteorhodopsin and why is it so named? Why might proteorhodopsin make a bacterium such as Pelagibacter more competitive in its habitat? How might heliorhodopsin be helpful to a phototrophic microbe?
What important role does the bacterium Ruegeria pomeroyi play in the health of our planet?
Why are dilute culture media used for isolating pelagic microbes?
20.13 Pelagic Marine Viruses
Viruses are found everywhere on and in Earth where cellular life is present (including on and in plants and animals) and are present in some environments in enormous numbers. The number of bacterial and archaeal cells on Earth is far greater than the total number of eukaryotic cells; estimates of total prokaryotic cell numbers are on the order of 1029. However, the number of viruses is even greater than this, an estimated 1030. Thus, one might expect that, despite their small size, viruses would play a major ecological role in nature. Soil, freshwater, Earth’s deep subsurface, and microbial mats, among many other habitats, are teeming with microbes and the viruses that feed on them. We focus here on seawater to give a feel for the numbers involved and how viruses interact with their prokaryotic hosts.
Bacteriophages and Archaeal Viruses in Seawater
There are about 106 prokaryotic cells/ml of seawater and approximately ten times as many viruses. Although viruses account for most of the total microbes present in seawater in terms of numbers, because of their very small sizes they constitute only about 5% of the total microbial biomass. However, using special fluorescence microscopy techniques, viruses can be visualized in seawater (Figure 20.32). It has been estimated that at least 5% and as many as 50% of the Bacteria in seawater are killed by bacteriophages each day, and most of the others are eaten by protozoa. For example, Syn5 is a bacteriophage that attacks and lyses Synechococcus species (Figure 20.33), who, along with their relative Prochlorococcus, are the major primary producers of the ocean and account for over 30% of the CO2 fixed globally (Section 20.11).
Figure 20.32 Viruses in seawater.
A water sample collected on a 0.02-μm (20-nm) pore size filter is stained with SYBR Green and viewed by epifluorescence microscopy. The tiny green dots are viruses while the larger, brighter dots are prokaryotic cells about 0.5 μm in diameter. Viruses are typically 10 times more abundant than the total of prokaryotic cells in seawater. Inset transmission electron micrographs show various marine bacterial viruses (scale bars, 100 nm in all images).
Jed Fuhrman, Jennifer R. Brum, Ryan O. Schenck, N. Solonenko, and Matt Sullivan
Figure 20.33 Cyanophage infection of *Synechococcus*.

Phase contrast–electron cryotomography sections of cells of Synechococcus in various stages of infection with phage Syn5. (a) Early. (b) Intermediate. (c) Late. Arrows point to phage particles.
The two primary mechanisms of bacterial mortality, grazing by protists and lysis by bacteriophages, contribute about equally to cell death but differ greatly in their impact on marine food webs. Grazing moves carbon up through the food chain when bacteria are consumed by protozoa, which are then consumed by zooplankton, which in turn sustain higher trophic levels up to fish and larger organisms. In contrast, the cytoplasm released as dissolved organic matter (DOM) by viral attack on Prochlorococcus (Figure 20.33) and other marine microbes—estimated to be as much as 3 gigatons of carbon per year—decreases the transfer of carbon to higher trophic levels. Although providing a significant amount of organic matter for other microbes, this released carbon amounts to about 25% of primary production that could otherwise be used by grazing populations.
Most described marine bacteriophages are head-and-tail bacteriophages containing double-stranded DNA genomes (Baltimore class I, Figure 11.2); by contrast, described RNA-containing phages and non-tailed doubled-stranded DNA viruses are relatively rare. However, electron microscopic observations of marine water samples have revealed that non-tailed viruses appear to dominate in the global ocean, even though this group is almost completely missing from marine virus isolates and sequence collections. This mystery was at least partially solved by the discovery of an entirely new lineage of non-tailed double-stranded DNA bacteriophage called the Autolykiviridae (the group is named for Autolykos of Greek mythology, a shape-changing thief notable for being difficult to catch). The virion of the virus has a more-or-less icosahedral morphology (Figure 20.34). The Autolykiviridae were recovered from plaque assays (Section 5.3 and Figure 5.10) using marine Vibrio cells as host. When genome sequences of isolated Autolykiviridae were used to screen marine metagenomic and genomic libraries for related viruses, sequences homologous to the gene encoding the capsid protein of the Autolykiviridae were identified in 13 bacterial and archaeal phyla, including two of the most abundant marine groups, the Alphaproteobacteria and the Thaumarchaeota.
Figure 20.34 *Autolykiviridae*.

Thin-section electron micrograph of lysed marine Vibrio cells containing virions of Autolykiviridae, a family of non-tailed dsDNA viruses. The viruses contain a lipid-bilayer membrane with an outer wall surrounding the virus core. A single virion is approximately 30 nm in diameter. Arrows point to individual virions.
The discovery of this previously unrecognized virus family of very broad host range (infecting both Bacteria and Archaea) has filled a major gap in our understanding of environmental virology. It is likely that this approach to discovery—combining classic methods of bacteriophage isolation in laboratory culture and comparative metagenomic sequence analysis—will continue to expand our knowledge of the marine virosphere. For example, recognition that bacteriophages infecting Pelagibacter species (Figure 20.30) are widespread in the marine environment was also dependent on first describing the bacteriophage infecting laboratory cultures of the host bacterium.
Marine Viral Diversity
Although laboratory characterization remains the definitive approach to virus description, additional understanding of viral diversity is being provided by viral metagenomics. Since viruses lack universal marker genes that can be used for targeted surveys (such as rRNA genes in cellular microbes), metagenomic approaches are used to conduct diversity surveys. Viral metagenomics combines virus particle purification and sequencing of DNA isolated from the virus particles. The viral metagenome is the sum total of all the virus genes in a particular environment. When combined with next-generation sequencing technologies, this approach has shown that a large part of genetic diversity on Earth resides with bacteriophages.
Several viral metagenomic studies have been undertaken, and they invariably show that immense viral diversity exists on Earth. For example, approximately 75% of the gene sequences found in viral metagenomic studies show no similarity to any other genes known, either viral or cellular. By comparison, surveys of bacterial metagenomes typically reveal approximately 10% unknown genes. However, viral metagenomics alone is generally not sufficient to identify the host(s) infected by a specific virus. Methods in development to improve host assignment include matching of CRISPR spacers (antiviral defenses of Bacteria and Archaea, Section 9.12) to environmental viral sequences and identifying patterns of co-occurrence between viral populations and putative host bacteria. Thus, most viruses still await discovery and most viral genes have unknown functions. This makes the study of the virosphere and viral diversity one of the most exciting areas of microbiology today. A rough estimate thus far of marine viral diversity indicates that at least 200,000 different kinds of viruses exist in the world’s oceans, harboring an enormous number of novel genes.
Viruses as Agents of Genetic Transfer in the Marine Environment
As we have seen, lysogenic bacteriophages can integrate into the genomes of their bacterial hosts (Section 5.6), and when they do, they can confer new properties on the cell. Moreover, some lytic phages facilitate the transfer of bacterial genes from one cell to another through the process of transduction, a major means of horizontal gene transfer in which a virus carries host genes between cells, for example by picking up host DNA and becoming a nonlytic transducing particle (Section 9.7). As agents of transduction, bacteriophages are thought to have a major influence on bacterial evolution by conferring new metabolic or other beneficial properties on recipient cells that allow them to successfully colonize new habitats.
A good example of bacteriophage gene transfer in marine systems are the “cyanophages,” phages that can transfer certain photosynthesis genes among strains of Synechococcus and Prochlorococcus. When these phages are released from their lysed host cells (Figure 20.33), some of them incorporate host genes that encode photosystem (PS) II, one of the key components of oxygenic photosynthesis (Section 14.6). When such a phage infects a new host cell, it provides the cell with genes encoding a modified PSII. It is hypothesized that a more diverse complement of PSII proteins improves both cell and phage fitness by allowing the host cell to better adapt to changing environmental conditions—for example, changes in light intensity or light quality—and in the process, producing more cyanophage.
When host-derived genes were first found in temperate (nonlytic) bacteriophages, they were called lysogenic conversion factors. However, recognition that hostlike genes are widely distributed in both temperate and lytic viruses required the introduction of the much broader term auxiliary metabolic genes (AMGs). During lytic infection, expression of AMGs alters host metabolism toward pathways that maximize production of new viral particles. For example, in the cyanobacteria–cyanophage system just described, the AMGs encode proteins that supplement photosynthetic electron transport and redirect energy from carbon fixation to the pentose phosphate pathway (Section 3.13) for increased production of ribose 5-phosphate, a nucleotide precursor needed to support virus replication. Similarly, cyanophages isolated from marine regions of low phosphate are more likely to have AMGs involved in phosphorus acquisition, such as genes for phosphate-binding protein (pstS) and alkaline phosphatase (phoA); this ensures that the host will have sufficient phosphate to produce cyanophage DNA.
Marine viruses infecting eukaryotes also encode AMGs. For example, an AMG of a virus infecting the calcareous phytoplankton Emiliania huxleyi (Figure 21.20a and Section 18.7) alters host lipid biosynthesis pathways to produce lipids needed for viral particle assembly. Although viral metagenomics has shown that one way or the other, the most common AMGs affect nucleotide biosynthesis, viral AMGs can influence many different pathways of microbial metabolism, most often to the benefit of the virus and not the host.
Survival Strategies of Viruses in Nature
When hosts are plentiful in nature, it is thought that bacteriophages adopt the lytic lifestyle and thus large numbers of host cells are killed. By contrast, when host numbers are low, it may be difficult for viruses to find a new host cell, and under such circumstances, lysogeny would be favored if the virus is lysogenic (Section 5.6). Under these conditions, the virus would survive as a prophage until host numbers increased and a lytic lifestyle could once again be supported. This hypothesis is consistent with the observation that in the depths of the ocean where bacterial numbers are lower than in surface waters, around half the bacteria examined have been found to contain one or more lysogenic viruses. As far as is known, no single-stranded DNA viruses and no RNA viruses can enter a lysogenic state, and so how these viruses might survive periods of low host numbers is unknown.
Check Your Understanding
What type of bacteriophages are most common in the oceans, based on what evidence?
How can bacteriophages affect bacterial evolution?
What does the viral metagenome suggest about our understanding of viral diversity?
What is an AMG, and how do AMGs benefit marine microbes?
20.14 The Deep Sea
Light penetrates no farther than about 250 m in pelagic waters; as mentioned, this illuminated region is called the photic zone (Figure 20.29). Beneath the photic zone, down to a depth of about 1000 m, there is still considerable biological activity. However, water at depths greater than 1000 m is, by comparison, much less biologically active and is known as the deep sea. Greater than 75% of all ocean water is deep-sea water, lying primarily at depths between 1000 and 6000 m. The deepest waters in the oceans lie below 10,000 m. However, because depressions this deep are very rare, the waters in them make up only a very small proportion of all pelagic waters.
The biology of the deepest ocean regions, the abyssal zone (below 4000 m) and the hadal zone (below 6000 m), has stirred the imagination for centuries. These regions remain among the least explored of Earth’s biosphere because their study requires very specialized equipment. Only a few remotely operated vehicles (ROVs), autonomous underwater vehicles, and human-occupied submersibles can dive to the deepest marine waters, which exceed depths of 10,000 m; pressures at such depths are around 16,000 pounds per square inch (110,000 kilopascals). Among the newest generation of human-occupied submersibles is the Limiting Factor (Figure 20.35). This two-person vessel is capable of diving to 11,000 meters, allowing humans to directly visit and explore all regions of the ocean floor, including the very deepest—the Challenger Deep, which lies within the Mariana Trench (Pacific Ocean, depth of 10,900 m).
Figure 20.35 The deep-diving submersible *Limiting Factor*.

(a) Deployment of the vessel. (b) Beginning a dive. This two-person vessel is capable of reaching depths of 11,000 meters with an operational dive time of 16 hours.
Conditions in the Deep Sea
Organisms that inhabit the deep sea face three major environmental extremes: (1) low temperature, (2) high pressure, and (3) low nutrient levels. In addition, deep-sea waters are completely dark such that photosynthesis relying on sunlight is impossible. Thus, microbes that inhabit the deep sea must be chemotrophic and able to grow under high pressure and oligotrophic conditions in the cold.
Below depths of about 1000 m, ocean water temperatures stay constant at 2–3 °C. We discussed the responses of microorganisms to changes in temperature in Sections 4.11, 4.12 and 4.13. As would be expected, many bacteria isolated from marine waters below 1000 m are psychrophilic (cold-loving) or at least psychrotolerant. However, since mesophilic microorganisms from warmer and more productive surface waters are constantly sinking through the water column, a variety of mesophiles can also be isolated from water collected at great depths. Deep-sea microbes must also be able to withstand the enormous hydrostatic pressures associated with great depths. Pressure increases by 1 atm for every 10 m of depth in a water column. Thus, an organism growing at a depth of 5000 m must be able to withstand pressures of 500 atm. We will see that microorganisms in general are remarkably tolerant of high hydrostatic pressures; many species can withstand pressures of 500 atm, and some species can withstand far more than this. Moreover, from studies of deep-sea microbial diversity performed thus far, some unusual microbes call this extreme environment home.
Piezotolerant and Piezophilic *Bacteria* and *Archaea*
Different physiological responses to pressure are observed in different deep-sea microorganisms. Some organisms simply tolerate high hydrostatic pressure, but do not grow optimally under such pressure; these organisms are piezotolerant (Figure 20.36). By contrast, others actually grow best under elevated hydrostatic pressure; these are called piezophiles. Organisms isolated from surface waters down to about 3000 m are typically piezotolerant. In piezotolerant organisms, higher metabolic rates are observed at 1 atm than at 300 atm, although growth rates at the two pressures may be similar (Figure 20.36). However, piezotolerant isolates typically do not grow at pressures greater than about 500 atm (Figure 20.36).
Figure 20.36 Growth of piezotolerant, piezophilic, and extremely piezophilic bacteria.

Compare the slower growth rate of the extreme piezophile (right ordinate) with the growth rate of the piezotolerant and piezophilic bacteria (left ordinate) and note the inability of the extreme piezophile to grow at low pressures.
Mastering Microbiology
Art Activity: Figure 20.30 Growth of piezotolerant, piezophilic, and extremely piezophilic bacteria
By contrast, cultures derived from samples taken at greater depths, 4000–6000 m, are typically piezophilic, growing optimally at pressures of around 300–400 atm. However, although piezophiles grow best under high pressure, they can still grow at 1 atm (Figure 20.36). In even deeper waters (for example, 10,000 m), extreme piezophiles (also called hyperpiezophiles) are present. These organisms require very high pressure for growth (Figure 20.37). For example, the extreme piezophile Moritella MT-5 isolated from the Mariana Trench (Pacific Ocean, >10,000-m depth) (Figure 20.38) grows optimally at a pressure of 700–800 atm (Figure 20.37) and grows nearly as well at 1035 atm, the pressure it experiences in its natural habitat. The current pressure champion, Colwellia marinimaniae, was isolated from the Challenger Deep. This obligately hyperpiezophilic psychrophile is a member of the Gammaproteobacteria and grows at pressures between 80 and 140 MPa, with an optimum growth pressure of 120 MPa and optimum growth temperature of 6 °C.
Figure 20.37 Pressure and temperature optima for cultured bacterial and archaeal piezophiles.

Pressure is in pascals (Pa), the SI unit for pressure. One megapascal (MPa) corresponds to approximately 10 atmospheres. Note that different species of the same genus (for example, Moritella) can have vastly different pressure optima. Data assembled by Doug Bartlett.
Figure 20.38 Sampling the deep sea.
The unmanned Japanese submersible Kaiko collecting a sediment sample on the seafloor of the Mariana Trench off the Philippines at a depth of 10,897 m. The tubes of sediment are used for enrichment and isolation of piezophilic bacteria, such as the extreme piezophile (Moritella) isolated from this seafloor.
Molecular Effects of High Pressure
High pressure affects cellular physiology and biochemistry in many ways. In general, pressure decreases the ability of the subunits of multi-subunit proteins to interact. Thus, large protein complexes in extreme piezophiles must interact in such a way as to minimize pressure-related effects. Protein synthesis, DNA synthesis, and nutrient transport are sensitive to high pressure. Piezophilic bacteria grown under high pressure have a higher proportion of unsaturated fatty acids in their cytoplasmic membranes than when grown at 1 atm. Unsaturated fatty acids allow membranes to remain functional and keep from gelling at high pressures or at low temperatures. The rather slow growth rates of extreme piezophiles compared with other marine bacteria (Figure 20.36) are likely due to the combined effects of pressure and low temperature; low temperature slows down the reaction rates of enzymes, directly affecting cell growth (Section 4.11).
Studies of gene expression and adaptive features contributing to growth at high pressure require special pressurized incubation devices (Figure 20.39). These studies have shown that when a gram-negative piezophile is grown under high pressure, a specific outer membrane protein called OmpH (outer membrane protein H) is present that is absent from cells grown at 1 atm. OmpH is a type of porin. Porins are proteins that form channels through which molecules diffuse into the periplasm (Section 2.4 and Figure 2.12c). Presumably, the porin made by cells grown at 1 atm cannot function properly at high pressure and thus a different porin must be synthesized. Interestingly, pressure controls transcription of ompH, the gene encoding OmpH. In characterized gram-negative piezophiles, a pressure-sensitive membrane protein complex is present that monitors pressure and triggers transcription of ompH only when conditions of high pressure warrant it. Transcriptomic analyses (Section 10.8) indicate that even relatively modest changes in hydrostatic pressure alter the expression of a large number of genes in piezophiles, so it is likely that many other pressure-monitoring proteins exist in these organisms.
Figure 20.39 Pressure cells for growing piezophiles under elevated pressure.

(a) Photo of several pressure cells incubating in a cold room (4 °C). (b) Schematic design of a pressure cell. These vessels are designed to maintain pressures of 1000 atm. Illustration based on drawing by Doug Bartlett.
There remains much to explore in the abyssal and hadal zones of the ocean. As we discussed in Chapter 19, recent advances in metagenomics and high-throughput sequencing have revolutionized studies of deep-sea microbes and their adaptive strategies, such as alterations in membrane and protein structure that allow growth under such extreme conditions. Diversity studies have shown that both free-living and particle-attached archaeal and bacterial piezophiles are abundant, and that most have not yet been cultured. For example, Bacteria related to the phyla Marinimicrobia (Figure 20.31) and Gemmatimonadetes are enriched at these great depths. Since no member of the Marinimicrobia has been isolated and the few Gemmatimonadetes available are from soils or wastewater, a more complete understanding of the unique properties of these hadal Bacteria awaits their future cultivation. Ammonia-oxidizing Archaea related to Nitrosopumilus (Thaumarchaeota, Section 17.5), an organism common in the upper marine water column, are also among the most abundant free-living Archaea in hadal waters, highlighting their global dominance in both marine and terrestrial environments. Thus, presence of these novel Archaea and Bacteria at great depths points to a unique hadal microbiology that we are only now beginning to more fully explore.
Check Your Understanding
How does pressure change with depth in a water column?
What molecular adaptations are found in piezophiles that allow them to grow optimally under high pressure?
20.15 Deep-Sea Sediments
In addition to deep-sea waters, another vast and mostly unexplored microbial biosphere exists below the seafloor in the deep-sea sediments. Drilling expeditions to explore far below the ocean seafloor have revealed both archaeal and bacterial populations in sediment cores taken at depths greater than 2000 m (Figure 20.40). Most studies thus far have focused on relatively organic-rich sediments along continental margins. Here, cell numbers typically decrease from about 109 cells/g in surface sediment to about 106 cells/g at depths as great as 1000 m below the seafloor. In these coastal sediments, sulfate-reducing bacteria and other anaerobes deplete sulfate and other electron acceptors within a few meters of the sediment–water interface (Figure 20.22). This depletion of electron acceptors and organic matter with increasing depth constrains energy available to the deep subsurface microbial communities, accounting for the major decrease in cell numbers with depth (Figure 21.29).
Figure 20.40 Drilling deep-sea sediments.

(a) Deep-sea drilling vessel the JOIDES Resolution. Inset: Red dot indicates the location of sediment sampling in the Peru Basin. (b) Sediment cores recovered from the Peru Basin at 4800-m depth. Cores were split lengthwise to allow subsampling for molecular characterization. See Section 20.5 and Figure 20.10 for discussion of sulfide-oxidizing microbial mats that develop on the sediment surface off the Chilean and Peruvian coasts.
Cell Numbers in Deep-Sea Sediments
The better-studied continental margins and shelf sediments are not representative of most of the ocean floor, about 90% of which is at greater than 2000-m depth and associated with marine waters of low productivity and therefore of much lower organic matter content (Figure 20.22). In the absence of significant transport of organic material to the sediment surface, sulfate and other electron acceptors may permeate all the way through the sediments to the underlying bedrock. However, because of the dearth of organic matter, cell numbers in these sediments are several orders of magnitude lower than in organic-rich sediments, ranging from about 106 cells/gram at the surface to fewer than 103 cells/gram at depths of a few hundred meters (Figure 20.41). Cell counts in all sediments generally decline significantly with depth, reflecting lower organic carbon availability and catabolic quality in older deep sediment material. Despite these low numbers, because deep-sea sediments are so vast, it has been estimated that a total of ∼5.4×1029 prokaryotic cells exist in deep-sea sediments (∼4×1015 g), a number similar to the sum total in all the oceans.
Figure 20.41 Microbial cell numbers in deep-sea sediments.

Depth-related cell abundance from the analysis of one coastal sediment (filled circles) and the global average for all sampled ocean sediments (dashed regression line) based on cell count data.
Figure 20.41 also shows that cell numbers can deviate from the global regression as a function of spotty distributions of resources with depth at any particular drill site. In the example shown in Figure 20.41, cell numbers in coastal plain sediments are elevated near methane (cold) seeps (Figure 20.22), a region associated with active anaerobic methane oxidation (Section 14.16), and in coal bed deposits (Figures 20.22 and 20.41), where, in addition to coal, associated organic materials boost electron donor availability. Other than these nutrient “oases,” nutrient limitation in deep sea sediments greatly restricts the size of indigenous microbial communities.
A Phylogenetic Snapshot of Marine Sediment Prokaryotic Diversity
Marine sediment communities have been explored only to a limited extent because of the difficulty and expense of obtaining uncontaminated drilling cores from great depths (Figure 20.40). However, analyses of 16S ribosomal RNA gene sequences obtained by using PCR methods (Section 19.6) on deep core samples have clearly established that novel Archaea make up a large fraction of the archaeal diversity and that bacterial diversity in both deep and shallow marine sediments is dominated by Proteobacteria (Figure 20.42). This abundance of Proteobacteria is also true of all the other habitats explored by culture-independent techniques (Figures 20.14, 20.20, 20.31, and see Figure 20.47).
Figure 20.42 Marine sediment bacterial and archaeal diversity.

The results are pooled analyses of 13,360 16S ribosomal RNA gene sequences from several studies of shallow and deep marine sediments. Many of the groups indicated are covered in Chapters 15 and 16 (Bacteria) or 17 (Archaea). For Proteobacteria, major subgroups are indicated. Note the high proportion of archaeal sequences and of Gamma-, Delta-, and Epsilonproteobacteria. Data assembled and analyzed by Nicolas Pinel. Compare the prokaryotic diversity of marine sediments with that of open ocean water shown in Figure 20.31.
Within marine sediment Proteobacteria, phylotypes associated with sulfate-reducing bacteria such as the Desulfobacterales are quite common (Figure 20.42), which is consistent with the fact that sulfate reduction (Sections 14.12 and 15.11) is the major form of anaerobic respiration in marine sediments. Bacteroidetes and the unclassified/minor groups are also well represented in shallow marine sediments. Although abundant in marine waters, cyanobacteria compose just a tiny proportion of the total cell population in the permanently dark and anoxic sediments and probably represent cells that have reached the sediments after attaching to a particle or dead animal that eventually sank.
In addition to a variety of Bacteria, novel phyla of Archaea unrelated to cultured representatives are widespread in the deep subsurface. Genome sequences of uncultured Bathyarchaeota recovered from both terrestrial and marine sediments (Figure 20.16) revealed a physiological capacity to degrade and assimilate protein and a capacity for some species to degrade carbohydrates. Other Bathyarchaeota appear to carry out methanogenesis and are autotrophs, likely existing on the small amounts of H2 produced from geochemical reactions (Section 20.8) in deep sediments.
Energy Limitation and Microbial Life Below the Seafloor
How microbes in the deepest marine sediments survive in their nutrient-depleted environment remains unclear, but it is likely that they employ many of the strategies we have seen in marine pelagic microbes including small cell size (Figure 20.17) and small, compact genomes. Sequencing of 16S ribosomal RNA genes selectively amplified by PCR (Section 19.6) using DNA extracted from deep drilling cores, as well as from more limited metagenomic surveys, has identified relatively few sequences related to the classical sulfate-reducing bacteria (Section 15.11) or methanogenic and known methane-oxidizing Archaea (Sections 14.16, 15.15, and 17.2), organisms that by contrast are quite common in surface sediments. Thus, from a nutritional standpoint, how do buried cells thrive under conditions of extreme electron donor limitation?
In the marine environment, easily degraded organic material is removed by microbial respiration in the water column and anaerobic respiration in surficial sediments (the sediments at and near the sediment surface), leaving behind a dilute pool of less readily degradable organic material that slowly trickles down into deeper sediments. Microbes that inhabit the deep sediments presumably utilize this low-quality organic material along with organic carbon released from dead cells as electron donors in energy metabolism. However, since representative organisms have yet to be isolated from deep sediments, our understanding of the supporting energy-conserving metabolisms is sketchy and can only be inferred from a few partial genome sequences assembled from metagenomes (Sections 10.7 and 19.8) or obtained from single cells isolated directly from sediments, using single-cell genomic technology (Section 10.11). Various known and yet-to-be-discovered forms of anaerobic respiration and fermentation are the most likely candidates for deep-sea sediment metabolisms, perhaps along with other metabolic options not obvious from the conditions known to exist in deep marine sediments. Also, with growth rates of deep sediment microbes likely to be near the slowest of any on Earth, tiny amounts of organic material may be all that these cell numbers need to survive in small populations for extended periods.
Since sediment microorganisms largely control the fate of carbon in this vast subsurface reservoir of organic matter, the discovery of novel Archaea (Figure 20.16) living at the thermodynamic edge of life has given microbial ecologists a new perspective on carbon cycling in marine sediments. However, the extent to which abiotic processes may also contribute energy sources to these buried microbes is unclear but could be significant. For example, the higher temperatures found at greater depths may promote the alteration of organic material, releasing methane and other hydrocarbons, H2, acetate, and CO2 that may diffuse upwards to nourish microbes inhabiting the deep sediment biosphere. There is clearly much more to learn about the microbial ecology of Earth’s deep subsurface, both in terrestrial and marine sediment contexts.
Check Your Understanding
Give two reasons why sulfate-reducing bacteria are common in shallow marine sediments.
How and why do the numbers of bacterial and archaeal cells vary with depth in marine deep sediments?
What alternative sources of energy are suggested to nourish microorganisms in extremely deep marine sediments?
20.16 Hydrothermal Vents
Although we have thus far described the deep sea and deep-sea sediments as a remote, low-temperature, high-pressure, oligotrophic environment suitable only for slow-growing piezotolerant and piezophilic microorganisms (Section 20.14), there are some amazing exceptions. Thriving animal and microbial communities cluster in and around deep-sea hot springs throughout the world. These hot springs are located at depths from less than 1000 m to greater than 4000 m in regions of the seafloor where volcanic magma and hot rock have caused the floor to split open at crustal spreading centers (Figure 20.22 and Figure 20.43), or where iron and magnesium minerals associated with ancient rocks react with seawater and generate heat. Seawater seeping into these dynamic cracking regions of the crust reacts with hot rock, resulting in hot springs saturated with inorganic chemicals and dissolved gases. Collectively, these types of underwater hot springs are called hydrothermal vents. We discuss several remarkable symbiotic associations between hydrothermal vent–associated animals and microorganisms in Chapter 23. Here we consider the vent environment as a habitat for free-living microbes.
Figure 20.43 Hydrothermal vents.

Schematic showing geological formations and major inorganic chemicals and minerals that are emitted from warm vents and black smokers. In warm vents, the hot hydrothermal fluid is cooled by cold 2–3 °C seawater permeating the sediments. In black smokers, hot hydrothermal fluid near 350 °C reaches the seafloor directly. The term “surficial” is a geological term meaning surface, in this case, the top layers of sediment.
Types of Vents
Volcanic hydrothermal systems are typically either warm (∼5 to >50 °C), diffuse vents or very hot vents that emit hydrothermal fluids at 270 to >400 °C. The gently flowing, warm, diffuse fluids are emitted from cracks in the seafloor and the exterior walls of hydrothermal chimneys. The fluids originate from the mixing of cold seawater with hot hydrothermal fluids in subsurface regions of the sediments. Hot vents, called black smokers, form upright sulfide edifices called chimneys that can be less than 1 m to over 30 m in height. Chimneys form when acidic hydrothermal fluids rich in dissolved metals and magmatic gases are suddenly mixed with cold, oxygenated seawater. The rapid mixing causes fine-grained metal sulfide minerals such as pyrite and sphalerite to precipitate out, forming dark, buoyant plumes that rise above the seafloor (Figure 20.44).
Figure 20.44 A hydrothermal vent black smoker emitting sulfide- and mineral-rich water at temperatures of 350°C.

The walls of the black smoker chimneys display a steep temperature gradient and contain several types of Bacteria and Archaea.
A quite different type of hydrothermal vent environment is the “Lost City” formation located in the mid-Atlantic Ocean. Lost City is formed from the exposure of minerals associated with ocean crust 1–2 million years old that was once deep beneath the seafloor. Geological faults in these slow-spreading systems exposed magnesium and iron-rich rocks called peridotites at the seafloor. Chemical reactions of seawater and newly exposed peridotite are highly exothermic, generating heat and also driving the pH up to as high as pH 11. Extremely high levels of H2, CH4, and other low-molecular-weight hydrocarbons are also present in the hot (200 °C) hydrothermal fluids. In contrast to the acidic volcanic black smoker systems (Figure 20.44), which are relatively transient, mixing of these alkaline fluids with seawater results in the formation of calcium carbonate (limestone) chimneys (Figure 20.45) that can reach up to 60 m in height and be active for 100,000 years or more.
Figure 20.45 Massive carbonate chimney formation at Lost City peridotite-hosted vent system.

Microbial colonization of freshly exposed mineral surfaces was studied by placing sterile mineral fragments in the green-topped device positioned over an actively venting area of the chimney. The diameter of the cylindrical collection device is approximately 10 cm. In contrast to the highly sulfidic vents (Figure 20.44), Lost City hydrothermal fluids are alkaline and lack sulfide but are rich in H2 and CH4.
*Bacteria* and *Archaea* in Hydrothermal Vents
Chemolithotrophic metabolisms (Table 14.1) dominate hydrothermal vent microbial ecosystems (Figure 20.22). Sulfidic vents support sulfur-oxidizing bacteria, whereas vents that emit other inorganic electron donors support nitrifying, hydrogen-oxidizing, iron- and manganese-oxidizing, or methylotrophic bacteria, the latter presumably growing on the CH4 and carbon monoxide (CO) emitted from the vents. Table 20.2 summarizes the inorganic electron donors and electron acceptors that are thought to play a role in chemolithotrophic metabolisms at hydrothermal vents.
Table 20.2 Chemolithotrophic *Bacteria* and *Archaea* present near deep-sea hydrothermal ventsa

aSee Chapter 14 for detailed discussions of these metabolisms and Chapters 15, 16 and 17 for further coverage of each group of organisms.
Although microbes cannot survive in the superheated hydrothermal fluids of black smokers, thermophilic and hyperthermophilic organisms do thrive in the gradients that form as the superheated water mixes with cold seawater. For example, within the walls of smoker chimneys can be found hyperthermophiles such as Methanopyrus, a species of Archaea that oxidizes H2 and makes CH4 (Section 17.2). In fact, phylogenetic FISH staining (Section 19.5) has detected cells of both Bacteria and Archaea in smoker chimney walls (Figure 20.46). The most thermophilic of all known sulfur-reducing microbes, species of Pyrolobus and Pyrodictium (Chapter 17), were isolated from black smoker chimney walls. In contrast to the significant microbial diversity in volcanic vent chimney walls, the carbonate chimney walls of the Lost City vents are primarily populated by methanogens of the genus Methanosarcina and are nourished by the H2-rich fluids that permeate the porous, CO2-rich chimney walls.
Figure 20.46 Phylogenetic FISH staining of sulfidic black smoker chimney material.

Taken from the Snake Pit vent field in the Mid-Atlantic Ridge, depth of 3500 m. A green fluorescing dye was conjugated to a probe that reacts with the 16S rRNA of all Bacteria and a red dye to a 16S rRNA probe for Archaea. The hydrothermal fluid going through the center of this chimney was at 300 °C.
When smokers plug up from mineral debris, hyperthermophiles presumably drift away to colonize active smokers and somehow become integrated into the growing chimney wall. Surprisingly, although they require very high temperatures for growth, hyperthermophiles are quite tolerant of cold temperatures and oxygen. Thus, transport of cells from one vent site to another in cold oxic seawater apparently is not a problem.
A Phylogenetic Snapshot of Hydrothermal Vent Prokaryotic Diversity
Using a variety of molecular methods (Section 19.6, 19.7 and 19.8), studies of the prokaryotic diversity present near volcanic hydrothermal vents have revealed an enormous diversity of Bacteria. These 16S rRNA gene sequence surveys include waters near both warm and hot vents. Hydrothermal vent microbial communities are dominated by Proteobacteria, in particular Epsilonproteobacteria (Section 16.5; Figure 20.47). Alpha-, Delta-, and Gammaproteobacteria are also abundant, whereas Betaproteobacteria are much less so.
Figure 20.47 Hydrothermal vent bacterial and archaeal diversity.

The results are pooled analyses from several studies of the 16S rRNA gene content of warm and hot hydrothermal vents. Many of these groups are covered in Chapters 15 and 16 (Bacteria) or 17 (Archaea). For Proteobacteria, major subgroups are indicated. Note the high proportion of Archaea and of Epsilonproteobacteria. The physiology of many of these organisms is summarized in Table 20.2. Most of the chemolithotrophs listed in Table 20.2 grow with CO2 as carbon source (autotrophic growth). Data assembled and analyzed by Nicolas Pinel.
Many Epsilon- and Gammaproteobacteria oxidize sulfide and sulfur as electron donors with either O2 or nitrate (NO3 −) as electron acceptors (Chapter 16). As shown in the detailed diagram of Proteobacteria in Figure 20.47, vent Epsilonproteobacteria phylotypes most closely match those of chemolithotrophic sulfur bacteria such as Sulfurimonas, Arcobacter, Sulfurovum, and Sulfurospirillum. These bacteria oxidize reduced sulfur compounds as electron donors (Sections 14.7 and 15.12), and such a physiology is consistent with their presence near vent fluids charged with sulfur and sulfide. In addition, most Deltaproteobacteria specialize in anaerobic metabolisms using oxidized sulfur compounds as electron acceptors.
In contrast to Bacteria, the diversity of volcanic hydrothermal vent Archaea is more limited. Estimates of the number of unique phylotypes indicate that the diversity of Bacteria near hydrothermal vents is about 10 times that of Archaea. However, Archaea prevail in samples recovered from the walls of hot vent chimneys (Figure 20.46). Most of the Archaea detected near hydrothermal vents are either methanogens (Section 17.2) or species of marine Crenarchaeota and Euryarchaeota (Chapter 17). With the exception of the ammonia-oxidizing Nitrosopumilus (Thaumarchaeota, Section 17.5), organisms in these groups remain uncultured and their physiologies poorly understood.
As we close this chapter dedicated to microbial ecosystems, it should be clear that any environment capable of supporting microbial life will be colonized with one or more microorganisms. The microbes present in each environment are typically a reflection of those whose metabolisms are most compatible with the conditions and resources available. But whether an ecosystem is nutrient robust or nearly devoid of nutrients, microbial communities will develop, and populations within a community will rise or fall in response to the dynamics of their environment.
Check Your Understanding
How does a warm hydrothermal vent differ from a black smoker, both chemically and physically?
Why is 350 °C water emitted from a black smoker not boiling?
Which phylum of Bacteria and which subgroups of this phylum dominate hydrothermal vent ecosystems, and why?
Chapter Review
Go to Mastering Microbiology for videos, animations, practice tests, and more.
I Microbial Ecology
20.1 Ecosystems consist of organisms, their environments, and all of the interactions among the organisms and environments. The organisms are members of populations and communities and are adapted to habitats. Species richness and abundance are aspects of species diversity in a community and an ecosystem.
Q What types of environmental disturbance would likely reduce species richness in an aquatic habitat? What types of environments would support high species diversity?
20.2 Microbial communities consist of guilds of metabolically similar organisms. Microorganisms play major roles in energy transformations and biogeochemical processes that result in the recycling of elements essential to living systems.
Q In what forms does potential energy enter a microbial ecosystem? Which energy classes of microorganisms can exploit each?
II The Microbial Environment
20.3 The niche for a microorganism consists of the specific assortment of biotic and abiotic factors within a microenvironment in which that microorganism can be competitive. Microorganisms in nature often live a feast-or-famine existence such that only the best-adapted species reach high population densities in a given niche. Cooperation among microorganisms is also important in many microbial interrelationships.
Q Why is it possible to isolate both obligately anaerobic and obligately aerobic bacteria from the same soil sample?
20.4 When surfaces are available, bacteria grow in attached masses of cells called biofilms. Biofilm formation confers several protective advantages on cells. Biofilms can have significant medical and economic effects on humans when unwanted biofilms develop on inert as well as living surfaces.
Q The surface of a rock in a flowing stream will often contain a biofilm. What specific advantages could be conferred on bacteria growing in a biofilm compared with growth within the flowing stream?
20.5 Microbial mats can be phototrophic or chemolithotrophic. Phototrophic cyanobacterial mats are thick biofilms consisting of microbial cells and trapped particulate materials and are widespread in hypersaline or thermal waters. Mats of iron-oxidizing microorganisms are common in low-pH and metals-rich waters of geothermal springs and mining waste waters. Sulfur-oxidizing chemolithotrophs form extensive mat communities on marine sediment surfaces below the photic zone at the oxygen–sulfide interface.
Q What is the importance of gliding motility in the formation of marine chemolithotrophic mat systems?
III Terrestrial Environments
20.6 Soils are complex microbial habitats consisting of sand, silt, and clay and contain numerous microenvironments and niches. Microorganisms are present in the soil primarily attached to soil particles or plant roots. The most important factors influencing microbial activity in soil are the availability of water and nutrients. However, in very arid soils microorganisms play important roles in stabilizing soil structure.
Q In what soil horizon are microbial numbers and activities the highest, and why?
20.7 Proteobacteria, Acidobacteria, and Bacteroidetes are the most diverse groups of Bacteria in soil because their metabolisms fit best with the available resources. Among Archaea, only Euryarchaetoa and Thaumarchaeota are commonly present in soils. Polluted soils contain different proportions of the same major groups of Bacteria but are also enriched in Actinobacteria and nearly devoid of Thaumarchaeota. Changes in soil microbial diversity due to pollution and climate change threaten global food supplies.
**Q Based on their metabolism, why are Thaumarchaeota well suited to inhabit soils?**
20.8 The deep subsurface is a significant microbial habitat that sustains chemolithotrophic populations whose resource demands are few and inorganic, such as CO2, SO4 2−, N2, and H2. Novel phyla of small-celled Archaea are present in the deep subsurface of both terrestrial and marine sediment environments. The discovery of the Asgard superphylum has provided new understanding of the origin of eukaryotes.
Q Studies of the deep subsurface have identified populations of chemoorganotrophs. What types and sources of nutrients are available to those organisms?
IV Aquatic Environments
20.9 In freshwater aquatic ecosystems, phototrophic microorganisms are the main primary producers. Most of the organic matter produced is consumed by bacteria, which can lead to depletion of oxygen in the environment. The BOD of a body of water indicates its relative content of organic matter that can be biologically oxidized.
Q How and in what way does an input of organic matter, such as sewage, affect the oxygen content of a river or stream?
20.10 Pelagic marine waters are more nutrient deficient than most freshwaters, yet substantial numbers of microbes inhabit the oceans. However, in some highly productive and expansive oceanic regions, oxygen can be drawn down to low or undetectable levels at depths between 100 and 1000 m; oxygen-depleted waters at these depths are called oxygen minimum zones (OMZs). Bacteria specialized in the biodegradation of petroleum are enriched in regions of natural seepage of petroleum and natural gas.
Q How is climate change related to the ongoing increase in size of OMZs?
20.11 The major oxygenic microbial phototrophs in the open oceans are the cyanobacterium Prochlorococcus and the alga Ostreococcus. Marine anoxygenic phototrophs include Roseobacter and its relatives, the aerobic phototrophic purple bacteria. Unlike other anoxygenic phototrophs, aerobic purple bacteria carry out photosynthesis only under aerobic conditions and thus are well suited to open ocean redox conditions.
**Q How has single-cell genomics contributed to new understanding of factors controlling the diversity of Prochlorococcus in the ocean?**
20.12 Species of Bacteria tend to predominate in marine surface waters, whereas in deeper waters Archaea and Bacteria make up similar shares of the microbial community. Most pelagic Bacteria are specialized to grow at very low nutrient concentrations, and many found in the photic zone use light to make ATP by rhodopsin-driven proton pumps. Thaumarchaeota and Euryarchaeota are the major Archaea inhabiting the oceans.
**Q Many pelagic Bacteria can use light energy but are not considered “phototrophs” in the same sense as cyanobacteria or purple bacteria. Explain.**
20.13 Marine viruses far outnumber total cell numbers in the ocean, greatly impact marine food webs, and confer new properties to their hosts as gene transfer agents. Phage-encoded auxiliary metabolic genes typically function to subvert host metabolism for increased virus production.
Q Describe two types of viral auxiliary metabolic genes and how they benefit the virus.
20.14 The deep sea is a cold, dark habitat where hydrostatic pressure is high and nutrient levels are low. Piezophiles grow best under pressure but do not require pressure, whereas extreme piezophiles require high pressure, typically several hundred atmospheres, for growth. Ammonia-oxidizing Archaea are among the most abundant free-living microbes in the deepest regions of the ocean.
Q What property is shared by piezotolerant, piezophilic, and extremely piezophilic microorganisms?
20.15 Deep-sea sediments show decreasing levels of nutrients with depth, and thus the microbes that reside there experience constant near-starvation conditions. Although not yet cultured, novel phyla of Archaea inhabit deep marine sediments (as well as the terrestrial deep subsurface), and these organisms likely survive by scavenging the trace levels of both organic matter and electron acceptors present there.
Q What sources of organic matter in deep-sea sediments could be used to support growth?
20.16 Hydrothermal vents are deep-sea hot springs where either volcanic activity or unusual chemistry generates fluids containing large amounts of inorganic electron donors (such as sulfide, ammonia, and hydrogen) that can be used by various chemolithotrophic bacteria.
Q Would you expect to find the same types of microorganisms associated with black smoker and carbonate vent systems? Explain.
Application Questions
Imagine a sewage plant that is releasing sewage containing high levels of ammonia and phosphate and very low levels of organic carbon. Which types of microbial blooms might be triggered by this sewage? How would the graphs of oxygen near and beyond the plant’s release point differ from the graph shown in Figure 20.19a?
Keeping in mind that the open-ocean waters are highly oxic, predict the possible metabolic lifestyles of open-ocean Archaea and Bacteria. How might rhodopsin-like pigments be more abundant in one group of organisms than in the other?
Warming associated with climate change has been suggested to result in reduced transfer of oxygen to deeper waters in the ocean. How might such warming also result in reduced nutrient availability to planktonic species in marine surface waters?
Chapter Glossary
virus-encoded genes that alter host metabolism toward pathways that maximize production of new virus particles Biochemical oxygen demand (BOD)
the relative amount of dissolved oxygen consumed by microorganisms for complete oxidation of bioavailable organic and inorganic material in a water sample Biofilm
colonies of microbial cells encased in a porous organic matrix and attached to a surface Biogeochemistry
the study of biologically mediated chemical transformations in the environment Community
two or more cell populations coexisting in a certain area at a given time Ecosystem
a dynamic complex of organisms and their physical environment interacting as a functional unit Epilimnion
the warmer and less dense surface waters of a stratified lake Extreme piezophile
an organism requiring several hundred atmospheres of pressure for growth Guild
metabolically similar microbial populations that exploit the same resources in a similar way Habitat
a part of the ecosystem best suited to one or a few populations Hydrothermal vent
a deep-sea hot spring emitting warm (∼20 °C) to superheated (>300 °C) water Hypolimnion
the colder, denser, and often anoxic bottom waters of a stratified lake Microbial mat
a thick, layered, diverse community nourished either by light in a hypersaline or an extremely hot aquatic environment, in which cyanobacteria are essential; or by chemolithotrophs growing on the surface of sulfide-rich marine sediments Microenvironment
a micrometer-scale space surrounding a microbial cell or group of cells Niche
in ecological theory, the biotic and abiotic characteristics of the microenvironment that contribute to an organism’s competitive success Oligotroph
an organism that grows only or grows best at very low levels of nutrients Oxygen minimum zone (OMZ)
an oxygen-depleted region of intermediate depth in the marine water column Piezophile
an organism that grows best under a hydrostatic pressure greater than 1 atm Piezotolerant
able to grow under elevated hydrostatic pressures but growing best at 1 atm Population
a group of organisms of the same species in the same place at the same time Primary producer
an organism that synthesizes new organic material from CO2 and obtains energy from light or from oxidation of inorganic compounds Prochlorophyte
a bacterial oxygenic phototroph that contains chlorophylls a and b but lacks phycobiliproteins Proteorhodopsin
a light-sensitive protein present in some pelagic Bacteria that fuels a proton pump that yields ATP Rhizosphere
the region immediately adjacent to plant roots Species abundance
the proportion of each species in a community Species richness
the total number of different species present in a community Stratified water column
a body of water separated into layers having distinct physical and chemical characteristics