23 Microbial Symbioses with Microbes, Plants, and Animals
23 Microbial Symbioses with Microbes, Plants, and Animals

## Chapter 23 Microbial Symbioses with Microbes, Plants, and Animals
Coral Fluorescence Provides the Guiding Light for Their Symbiotic Algae
Although many symbiotic microorganisms are transferred directly between generations of their host (vertical transfer), others are acquired from the environment (horizontal transfer) and must find their way to their host. Chemical signaling is one attraction mechanism. However, in at least one symbiosis, light from the green fluorescent protein (GFP) controls the process.
GFP is widely distributed among marine invertebrates, including jellyfish, anemones, and corals. Although often used in protein tagging experiments to measure gene expression and localize specific proteins in the cell, GFP and other proteins that fluoresce in different colors have natural functions that remain unclear. Some possibilities include shifting the color spectrum of bioluminescent organisms for prey attraction or avoidance or functioning in light sensory systems. However, GFP is now known to guide the symbiosis between stony corals (cnidarians) and Symbiodinium, the algal symbiont that nourishes the coral.
A significant fraction of Symbiodinium acquisition is by horizontal transfer, mostly during the coral larval and juvenile stages. How does the alga find the coral? The answer lies in the phototaxis of Symbiodinium toward light emitted from the coral’s GFP (photo). Researchers demonstrated the importance of phototaxis by painting GFP on traps placed in laboratory aquaria containing Symbiodinium and in natural reef communities. Symbiodinium cells migrated to the painted traps, but not to unpainted traps. The GFP light emission directing Symbiodinium phototaxis matched the spectrum optimal for its photosynthesis. Thus, the coral is simply exploiting the natural capacity of Symbiodinium to use phototaxis for positioning optimally in the marine light gradient. Phototaxis may also be important for coral reacquisition of their symbionts following a bleaching event. As coral bleaching becomes a more frequent occurrence with climate change, it is essential to better understand mechanisms of repair. Without recovery from bleaching, the major marine ecosystems supported by corals could soon be lost.
Source: Aihara, Y., Maruyama, S., Baird, A. H., Iguchi, A., Takahashi, S., and Minagawa, J. 2019. Green fluorescence from cnidarian hosts attracts symbiotic algae. Proc. Natl. Acad. Sci. (USA) 116: 2118.
Many microbial species form intimate and often mutually beneficial associations, called symbioses, with other microorganisms, plants, or animals. In a mutualism, both symbiotic partners benefit; in parasitism, one symbiotic partner benefits and the other is harmed; in commensalism, one symbiotic partner benefits and the other is unaffected. Symbioses typically develop through prolonged association and coevolution of the partners, evolution that proceeds jointly in a pair of intimately associated species owing to the effects each has on the other. In this chapter we explore some well-studied examples of these microbial symbioses as a prelude to our coverage in the following chapter of microbial symbioses with humans—symbioses that are not only extensive, but in many instances, absolutely essential for our well-being.
I Symbioses Between Microorganisms
Microorganisms can form intimate associations of two or more species to use resources more efficiently. Common associations form between phototrophic and chemotrophic partners, such as in lichens and some aquatic bacteria. Other associations exploit conductive surface structures to electrically couple two species for the oxidation of organic carbon.
Many microbial species form mutualisms with other microbial species. Direct microscopic observations of natural samples show that many microbes are not solitary entities but are associated with other microbes on surfaces or as suspended aggregates of cells. In most cases, the advantages conferred by an association are unknown. Because microbial ecologists have recognized that communities of interacting microbial populations—not individual organisms—control environmental processes, research to discover the nature of strictly microbial symbioses has increased. In Part I we present three microbial mutualisms where the advantages to both partners are clear.
23.1 Lichens
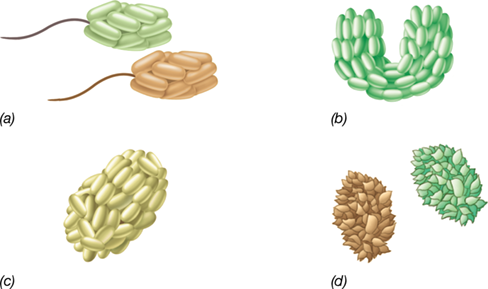
Lichens are readily visible, leafy or encrusting microbial symbioses often found growing on bare rocks, tree trunks, house roofs, man-made surfaces, and bare soils—surfaces where other organisms typically do not grow (Figure 23.1). The lichen symbiosis originates from an ancient mutualistic association between two dominant microorganisms: a fungus, usually an ascomycete (Section 18.13), and either an alga or a cyanobacterium. This association is one of the earliest forms of terrestrial plant life and can be traced back 400 million years to the Devonian period. The alga or cyanobacterium is the phototrophic partner and produces organic matter that feeds the fungus. The fungus is a chemotroph and provides a firm anchor within which the phototrophic partner can grow. The fungus also releases lichen acids that promote the dissolution and chelation of inorganic nutrients from the rock or other surface that are needed by the phototroph. The fungus facilitates the uptake of water and sequesters some for the phototroph, a characteristic that enables lichens to colonize almost all terrestrial environments, including deserts and other arid environments. Cells of the phototroph (also called the photobiont) are embedded in defined layers or clumps among cells of the fungus, together forming the thallus (Figure 23.2), and are protected from erosion by rain or wind.
Figure 23.1 Lichens.

Characteristic forms of lichens associated with (a) living and (b) dead tree branches, and (c) coating the surface of a large rock. Lichens are generally categorized by morphology and nearly all can be classified into one of the three types shown here, (a) foliose, (b) fruticose, and (c) crustose.
Figure 23.2 Lichen structure.

(a) Magnified image of a lichen thallus. The distance between the two arrows is approximately 6 mm. (b) Photomicrograph of a cross section through lichen thallus. The algal layer is positioned within the lichen structure near the top so as to receive the most sunlight. The fungal partners form the structural layers (cortex) positioned above and below the algal layer.
Lichen Components
Lichen photobionts are mostly green algae (Section 18.16) but are sometimes species of brown algae (Section 18.5). When cyanobacteria partner with lichens, they are often nitrogen-fixing species, organisms such as Anabaena or Nostoc (Sections 3.12 and 15.3). In a few lichens, two phototrophic species are present within a single thallus, and molecular studies have revealed that additional low-abundance species of photobionts can also be co-residents. Variation in the lichen physical structure is primarily determined by the fungus (Figure 23.1), and more than 18,000 species of fungi are able to form lichen associations with a variety of photobionts. Their habitat range spans tropical to polar climatic zones, and coastal to high-altitude habitats. Lichens typically grow quite slowly, with growth rates varying from 1 mm or less per year to over 3 cm per year, depending on the organisms composing the symbiosis and the temperature and amount of rainfall and sunlight received.
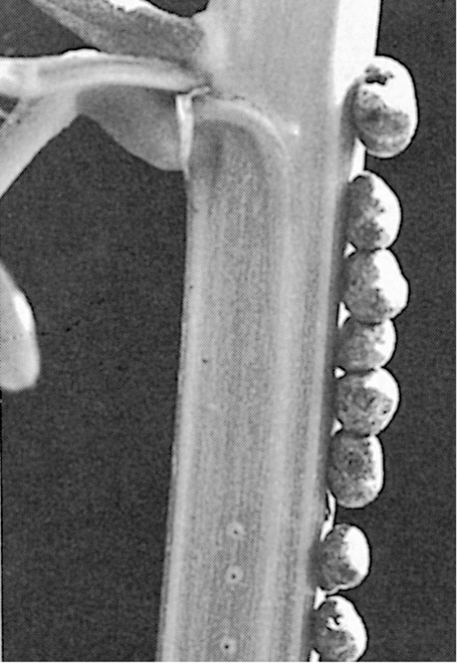
Diversity among the photobionts is lower than that of the fungi, and thus many different kinds of lichens can have the same phototrophic partner. The partner photobiont is primarily determined by lichen reproduction, which may be either asexual or sexual depending on the fungal species. During asexual reproduction, clonally derived fungal propagules co-disperse with the photobiont, preserving the specific symbiotic association. In contrast, fungal spores released by sexual reproduction lack the photobiont and require independent acquisition of a new symbiotic partner after dispersal. Such transmission provides an opportunity for the fungus to form new symbiotic associations that may have locally adaptive features. In contrast, lichens that disperse by propagules have less adaptive flexibility.
Molecular Studies of Lichens
The conventional view that lichens are simple two-partner assemblages has been challenged by culture-independent molecular studies (Section 19.6) showing that in addition to multiple photobionts, lichens may have more than one fungus and also host bacteria that may benefit the association. The cortex of many lichens, formed by fungal structural tissue bounding the phototroph layer (Figure 23.2), can also contain a yeast (basidiomycete, Section 18.14) in addition to the ascomycete. Similarly, a common feature of many lichens is association with Alphaproteobacteria of the order Rhizobiales—known N2-fixing bacteria—suggesting that in addition to cyanobacteria, other organisms may contribute fixed nitrogen to the symbiosis.
Metagenomic and metabolomic studies (Sections 10.7, 10.10, and 19.8) have identified other metabolic benefits of the bacterial association, including the production of vitamins and protection from toxic compounds. A striking example of the latter is the increased abundance of genes encoding arsenic resistance detected in lichens inhabiting environments contaminated with arsenic. These examples demonstrate how culture-independent methods can reveal exciting new information about even well-studied symbioses.
Check Your Understanding
What two types of microbes form a partnership in the lichen symbiosis? What are the benefits to both partners?

Besides organic compounds, of what benefit to the fungus is a mutualism with Anabaena?
What might be advantages of having bacteria present in the lichen symbiosis?
23.2 “Chlorochromatium aggregatum”
Microbial mutualisms called consortia form in freshwater environments. A commonly observed consortium develops between nonmotile green sulfur bacteria (phototrophs that are colored either green or brown) and certain motile, nonphototrophic bacteria. These consortia are found worldwide in stratified sulfidic freshwater lakes and can account for up to 90% of the green sulfur bacteria present and nearly 70% of the bacterial biomass in these lakes. The basis of the mutualism of these consortia is in the phototrophic production of organic matter by the green sulfur bacterium and the motility and organic matter consumption of the chemotrophic partner organism. Each consortium has been given a genus and species name, but since these names do not denote true species (because they are not a single organism), the names are enclosed in quotation marks. We examined the general biology of these consortia in Section 15.6.
Nature of the Consortium
The morphology of a green sulfur bacterial consortium depends on its species composition. The consortium generally consists of 13–69 green sulfur bacteria, called epibionts, surrounding and attached to a central, colorless, flagellated, rod-shaped bacterium (Figure 23.3). Several distinct motile phototrophic consortia have been recognized based on the color, morphology, and presence or absence of gas vesicles (Section 2.7) of the epibionts. For example, in “Chlorochromatium aggregatum” the central bacterium is surrounded by rod-shaped green bacteria, whereas in “Pelochromatium roseum” the epibiont is brown. The consortium “Chlorochromatium glebulum” is bent and includes green epibionts that contain gas vesicles (Figure 23.3).
Figure 23.3 Drawings of some motile phototrophic green bacterial consortia found in freshwater lakes.

Green epibionts: (a) “Chlorochromatium aggregatum,” (b) “C. glebulum,” (c) “C. magnum,” (d) “C. lunatum.” Brown epibionts: (a) “Pelochromatium roseum,” (d) “P. selenoides.” The epibionts are 0.5–0.6 μm in diameter. Adapted from Overmann, J., and H. van Gemerden. 2000. FEMS Microbiol. Rev. 24: 591.
Green sulfur bacteria are obligately anaerobic phototrophs that form a distinct phylum (Chlorobi, Section 15.6). The green and brown species differ in the types of bacteriochlorophyll and carotenoids they contain. Both green and brown species are found in stratified lakes where light penetrates to depths at which the water contains hydrogen sulfide (H2S), the primary electron donor for photosynthetic CO2 fixation by the phototroph. In stratified lakes, the motile consortia reposition rapidly to remain in regions where conditions are most favorable for photosynthesis in the constantly changing gradients of light, oxygen, and sulfide that occur throughout the course of a day. Water samples collected from depths where these conditions are most favorable are enriched in this morphologically conspicuous consortium (Figure 23.4). The consortia show dark aversion (scotophobotaxis, Section 2.12) and positive chemotaxis toward sulfide.
Figure 23.4 Phase-contrast micrograph of “*Pelochromatium roseum*” from Lake Dagow (Brandenburg, Germany).
The preparation was compressed between a coverslip and microscope slide to reveal the central rod-shaped bacterium (arrow). A single consortium is about 3.5 μm in diameter. Used with permission from J. Overmann and H. van Gemerden. 2000. FEMS Microbiol. Rev. 24: 591.
Some free-living green sulfur bacteria, such as Pelodictyon (Chlorobium) phaeoclathratiforme, produce gas vesicles that regulate buoyancy and vertical position in the water column. However, because they are so small, the time they require for repositioning in the water column is from one to several days, which is not fast enough for tracking the more rapidly changing gradients. By contrast, motile consortia move up and down in the water column fast enough to follow the gradients of light and sulfide as they change on a diel basis.
Although green bacterial consortia were discovered almost a century ago, only with the advent of molecular techniques and newer culture methods has it become possible to study certain aspects of these remarkable associations. Sequencing of 16S ribosomal RNA (rRNA) genes revealed a significant geographic distribution of epibionts in lakes of Europe and the United States, many of which were genetically distinct. Epibionts in neighboring lakes have identical 16S rRNA gene sequences, whereas the sequences of morphologically similar epibionts in widely separated lakes differ. Phylogenetic analysis indicates that mechanisms of cell–cell recognition between epibionts and their central bacterium arose through coevolution of each specific partnership.
Phylogeny and Metabolism of a Consortium
The epibiont of “Chlorochromatium aggregatum” has been isolated and grown in pure culture. Although this green sulfur bacterium, named Chlorobium chlorochromatii, can be grown in pure culture, no naturally free-living variant has been observed, supporting the view that in nature a symbiotic lifestyle is obligate for epibionts. The central bacterium of “Chlorochromatium aggregatum” is a species of Betaproteobacteria (Section 16.2). This bacterium requires α-ketoglutarate (an intermediate of the citric acid cycle, Figure 3.12) for growth, and this is presumably supplied to it by the epibiont. However, the central cell only assimilates fixed carbon in the presence of light and sulfide—conditions in which the epibionts are active and can transfer nutrients to the central bacterium. Genomic analysis of the central bacterium of one consortium revealed massive gene loss, indicating that this organism is unable to grow independently of the green sulfur bacterium.
Recent studies comparing the transcriptome and proteome (Sections 10.8 and 10.9) of C. chlorochromatii growing alone or in association with the central rod bacterium have identified some features specifically related to the symbiosis. Approximately 50 C. chlorochromatii proteins are unique to the symbiotic state. Most of approximately 350 differentially regulated genes are repressed when the organism is symbiotically associated, whereas only 19 genes are more highly expressed. Many of the more highly expressed genes encode proteins of amino acid metabolism and nitrogen regulation. These include the enzyme glutamate synthase (Section 3.14) and a transport protein for branched amino acids, suggesting that in addition to α-ketoglutarate, the metabolic coupling between the epibiont and central bacterium involves the exchange of amino acids as well.
Scanning electron microscopy of the consortium (Figure 23.5) has revealed that tubular extensions of the central bacterium’s periplasm (Section 2.4) cover much of its surface and appear to fuse with the periplasm of the epibiont. If the two bacterial partners actually share a common periplasmic space, this would facilitate the transfer of nutrients from phototroph to chemotroph and vice versa (however, whether the chemotroph reciprocates and transfers nutrients to the phototroph is unknown). The fact that the central bacterium is unable to grow without its phototrophic partner (while the phototroph can be grown in pure culture), and that organic compounds are only assimilated by the chemotroph in the light, is strong evidence that nutrients flow from the phototroph to feed the chemotroph and that the chemotroph is obligatorily dependent on its phototrophic partner.
Figure 23.5 Scanning electron micrographs of “*Chlorochromatium aggregatum*.”

(a) Chlorobium chlorochromatii epibionts tightly clustered around a flagellated central bacterium. (b) The central bacterium exhibits numerous protrusions of its outer membrane that make intimate contact with the epibionts, possibly fusing the periplasms of the two gram-negative organisms. Cells of the epibiont are about 0.6 μm in diameter. Used with permission from G. Wanner et al. 2008. J. Bacteriol. 190: 3721.
Check Your Understanding
What is the evidence that “Chlorochromatium aggregatum” is a stable product of evolution?
What advantage does motility offer a phototrophic consortium?
How might nutrients be shuttled between phototroph and chemotroph in the consortium?
23.3 Methanotrophic Consortia: Direct Interspecies Electron Transfer
23.3 Methanotrophic Consortia: Direct Interspecies Electron Transfer
23.3 Methanotrophic Consortia: Direct Interspecies Electron Transfer
Methanotrophic consortia that couple the activities of two anaerobic microbes effectively oxidize methane to CO2 in anoxic marine sediments. The biochemistry of this process was discussed in Section 14.16.
Anaerobic Oxidation of Methane and Short-Chain Alkanes
It has long been known that methane is oxidized in anoxic marine sediments. Subsequent culture-independent studies showed that specific methane-oxidizing (methanotrophic) Archaea form intimate associations with sulfate-reducing bacteria (**Figure 23.6*a***) to do this, and working together, the organisms carry out the following reaction: CH4+SO4 2−+H+→CO2+HS−+2 H2OΔG0′=−18 kJ
Figure 23.6 Anaerobic methane-oxidizing consortia.

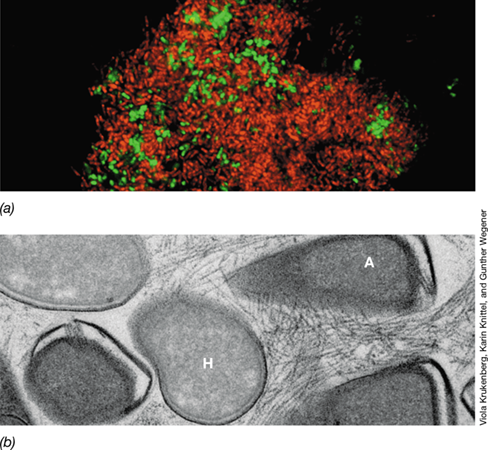
(a) A methanotrophic consortia enriched from geothermally heated sediments (Guaymas Basin, Gulf of California, Mexico) by cultivation with methane as sole electron donor and stained by CARD-FISH (Section 19.5) using probes selective for the ANME methanotroph (red fluorescence) and its sulfate-reducing bacterial partner (green fluorescence). (b) Electron micrograph of a thin section through the consortia, showing the electrically conductive “nanowires” produced by the sulfate reducer (H), connecting it electrically to cytochrome-rich proteins on the surface of the ANME methanotroph (A).
The methanotrophic Archaea comprise three different groups of anaerobic methane (ANME)-oxidizing Euryarchaeota. Members of the ANME-1 cluster belong to the Methanomicrobia, ANME-2 the order Methanosarcinales, and ANME-3 to Methanococcoides species (Section 17.2).
All ANME Archaea employ part of the metabolic pathway used by methanogens to generate methane, but ANME run the pathway in the opposite direction. The first step of anaerobic oxidation is activation of methane by methyl-coenzyme M reductase (MCR), producing coenzyme M–bound methyl groups that are further oxidized by the C1 pathway (Section 14.15). Some species of ANME belonging to the Methanomicrobia can completely oxidize short-chain alkanes (ethane, propane, and butane) using variants of MCR that activate the alkanes for further oxidation in a similar fashion to how MCR activates methane in ANME species. For example, ethane-oxidizing enrichment cultures produce ethyl-coenzyme M (ethyl-CoM) as an initial intermediate. Despite this variation in substrates, short-chain alkane oxidizers depend on syntrophic interactions with a sulfate reducer or other appropriate electron-accepting partner bacteria.
Sulfate-Reducing Bacteria as Partner Organisms in Anoxic Hydrocarbon Oxidations
A variety of sulfate-reducing Bacteria are syntrophic partners, including the Deltaproteobacteria Desulfosarcina and Desulfococcus species, and species of the family Desulfobulbaceae (Section 15.11). These bacteria accept electrons from hydrocarbon-oxidizing methanogens and dispose of them by reducing sulfate to sulfide. The transfer of electrons occurs by direct electron transfer between the two species, using electrically conductive appendages similar to those described for metal-reducing bacteria (Sections 14.13, 15.13, and 21.5 and Figures 14.31 and 21.14). The conductive structures (also called “nanowires”) bridge the intercellular space between the organisms (Figure 23.6b).
Anaerobic methane oxidation may not depend on syntrophic association in all cases. For example, enrichment cultures dominated by certain ANME Archaea established from freshwater samples were shown to couple methane oxidation to the reduction of nitrate to nitrite, rather than to the reduction of sulfate. In addition, some evidence suggests that redox-active metals such as ferric iron may also function as electron acceptors to support anoxic methanotrophy. However, in both of these cases the presence of a second organism (the electron consumer) to form a syntrophic association cannot be ruled out, as pure cultures of ANME organisms have not yet been obtained.
Direct Interspecies Electron Transfer (DIET)
Direct interspecies electron transfer (abbreviated DIET) of the type that drives the ANME–sulfate reducer association is much more common than was earlier suspected. For example, apart from the direct reduction of insoluble metals by Geobacter species (Section 21.5), species of Geobacter also form partnerships with methanogens. When paired in an association with G. metallireducens, the normally acetate-utilizing (acetoclastic) methanogen Methanosaeta harundinacea reduces carbon dioxide to methane using electrons delivered by G. metallireducens from acetate oxidation. In this partnership, Geobacter benefits because it uses the acetate as its carbon and energy source. And, because abiotic electron acceptors are limiting or absent, Geobacter uses the methanogen as its electron acceptor. The methanogen benefits because instead of splitting acetate to form methane, it reduces CO2 to CH4 instead, a more energetically favorable reaction (Section 14.15). As we learn more about organisms participating in DIET, greater metabolic variety will almost surely be revealed.
Check Your Understanding
What observations suggest that direct interspecies electron transfer may be more widely distributed among Bacteria and Archaea than now recognized?
Which of the two partner organisms, the ANME or the sulfate-reducing bacterium, may have the capacity to live independently? How might this influence the specificity of interaction?
II Plants as Microbial Habitats
**Symbiotic associations between microorganisms and plants can be mutualistic, where the microbe and the plant exchange nutrients, or parasitic, where the plant produces nutrients only for the bacterium. Prime examples of the former exist in the Rhizobium–root nodule symbiosis and of the latter in crown gall disease.**
Plants interact closely with microbes through their roots and leaf surfaces and even more intimately within their vascular tissue and cells. Most mutualisms between plants and microorganisms increase nutrient availability to the plants or defend them against pathogens. We consider three examples in the next three sections: (1) a mutualism where the nature of the symbiosis is understood in exquisite detail (root nodules), (2) a mutualism in which plants expand and interconnect their root system through association with a fungus (mycorrhizae), and (3) a symbiosis that is harmful to the plant (crown gall disease).
23.4 The Legume–Root Nodule Symbiosis
A plant–bacterial mutualism of great importance to humans is that of leguminous plants and nitrogen-fixing bacteria. Legumes are flowering plants that bear their seeds in pods and include such agriculturally important members as soybeans, clover, alfalfa, beans, and peas. These crops are key commodities for the food and agricultural industries, and the ability of legumes to grow without added nitrogen saves farmers millions of dollars in fertilizer costs yearly and reduces the polluting effects of fertilizer runoff.
The partners in a symbiosis are called symbionts, and most nitrogen-fixing bacterial symbionts of plants are collectively called rhizobia, derived from the name of a major genus, Rhizobium. Species of rhizobia are Alpha- or Betaproteobacteria (Figure 23.7; Sections 16.1 and 16.2) that can grow freely in soil or infect leguminous plants and establish a symbiotic relationship. The same bacterial genus (or even species) of legume can contain both rhizobial and nonrhizobial strains of nitrogen fixers. Infection of legume roots by rhizobia leads to the formation of root nodules (Figure 23.8) in which the bacteria convert N2 to NH3 (Sections 3.12 and 15.9). Nitrogen fixation in root nodules accounts for a fourth of the N2 fixed annually on Earth and is of enormous agricultural importance, as it increases the fixed nitrogen content of soil. Nodulated legumes can grow well on unfertilized bare soils that are nitrogen deficient, while other plants grow only poorly on them (Figure 23.9).
Figure 23.7 Phylogeny of rhizobial (genus names in boldface) and related genera inferred from analysis of 16S rRNA gene sequences.

More than 70 species of rhizobia are found in 12 genera of Alpha- and Betaproteobacteria.
Figure 23.8 Soybean root nodules.

The nodules developed from infection by Bradyrhizobium japonicum. The main stem of this soybean plant is about 0.5 cm in diameter.
Figure 23.9 Effect of nodulation on plant growth.

A field of unnodulated (left) and nodulated (right) soybean plants growing in nitrogen-poor soil. The yellow color is typical of chlorosis, the result of nitrogen starvation.
Mastering Microbiology
Art Activity: Figure 23.10 Steps in the formation of a root nodule in a legume infected by Rhizobium
Leghemoglobin and Cross-Inoculation Groups
In the absence of its bacterial symbiont, a legume cannot fix N2. Rhizobia, on the other hand, can fix N2 when grown in pure culture under microaerophilic conditions (a low-oxygen environment is necessary because the key nitrogen-fixing enzyme nitrogenase is inactivated by high levels of O2, Section 3.12). In the nodule, O2 levels are precisely controlled by the O2-binding protein leghemoglobin. Production of this iron-containing protein in healthy N2-fixing nodules (Figure 23.10) is induced through the interaction of the plant and bacterial partners. Leghemoglobin functions as an “oxygen buffer,” cycling between the oxidized (Fe3+) and reduced (Fe2+) forms of iron to supply sufficient O2 for bacterial respiration while keeping unbound O2 within the nodule low. The ratio of leghemoglobin-bound O2 to free O2 in the root nodule is thus maintained on the order of 10,000:1.
Figure 23.10 Root nodule structure.

Sections of root nodules from the legume Coronilla varia, showing the reddish pigment leghemoglobin.
There is a marked specificity between the species of legume and rhizobium that can establish a symbiosis. A particular rhizobial species is able to infect certain species of legumes but not others. A group of related legumes that can be infected by a particular rhizobial species is called a cross-inoculation group. Each group consists of all the legume species that will develop nodules when inoculated with rhizobia obtained from any other legume of the group (Table 23.1). If legumes are inoculated with the correct rhizobial strain, leghemoglobin-rich, N2-fixing nodules develop on their roots (Figures 23.8–23.10).
Table 23.1 Major cross-inoculation groups of leguminous plants

aSeveral varieties (biovars) of Rhizobium leguminosarum exist, each capable of nodulating a different legume.
Steps in Root Nodule Formation
How root nodules form is well understood, and the steps are as follows (Figure 23.11):
Recognition of the correct partner by both plant and bacterium and attachment of the bacterium to the root hairs
Secretion of oligosaccharide signaling molecules (Nod factors) by the bacterium
Movement of bacteria to the main root by way of the infection thread
Formation of modified bacterial cells (bacteroids) within the plant cells, development of the N2-fixing state, and continued plant and bacterial cell division forming the mature root nodule
Figure 23.11 Steps in the formation of a root nodule in a legume infected by *Rhizobium*.

Formation of the bacteroid state is a prerequisite for nitrogen fixation. The time course of nodulation events from infection to effective nodule is about 1 month for soybeans. See Figure 23.16 for physiological activities in the nodule.
Attachment and Infection
The roots of leguminous plants secrete organic compounds that stimulate the growth of a diverse rhizosphere microbial community. If rhizobia of the correct cross-inoculation group are in the soil, they will form large populations and eventually attach to the root hairs that extend from the roots of the plant (Figure 23.11). An adhesion protein called rhicadhesin is present on the cell surfaces of rhizobia. Other substances, such as carbohydrate-containing proteins called lectins and specific receptors in the plant cytoplasmic membrane, also play roles in plant–bacterium attachment.
After attaching, a rhizobial cell penetrates into the root hair, which curls in response to substances (Nod factors) secreted by the bacterium. The bacterium then induces the plant to form a cellulosic tube, called the infection thread (**Figure 23.12*a***), which spreads down the root hair. Root cells adjacent to the root hairs subsequently become infected by rhizobia, and plant cells divide. Continued plant cell division forms the tumorlike nodule (Figure 23.12a) consisting of plant cells filled with bacteroids (discussed below and shown in Figure 23.12b, c). A different mechanism of infection is used by some rhizobia adapted to aquatic or semiaquatic tropical legumes (see Figure 23.17). These rhizobia enter the plant at the loose cellular junctions of roots emerging perpendicular from an established root (lateral roots), or on a stem in or just above the water. Following entry into the plant, some of the rhizobia develop infection threads, whereas others do not.
Figure 23.12 The infection thread and formation of root nodules.

(a) Light micrograph of an early-stage nodule from a legume (Lotus japonicus) infected with a rhizobium strain containing the lacZ gene. The nodule was sectioned and bacterial distribution (blue) determined using an activity stain (X-gal) that turns blue when cleaved by the enzyme β-galactosidase (Section 12.2 and Figure 12.10). An infection thread, consisting of a cellulosic tube through which bacteria move to root cells, is clearly visible extending from the surface to the interior. (b) Transmission electron micrograph through a soybean nodule infected with Bradyrhizobium japonicum, showing bacteroid-filled plant cells. The plant cell is approximately 50 μm long. (c) Higher-magnification micrograph showing individual symbiosomes, each filled with several bacteroids. The clear areas in each bacteroid are the storage polymer poly-β-hydroxybutyrate (Section 2.7). Bacteroids are about 2 μm long.
Bacteroids
The rhizobia multiply rapidly within the plant cells and become transformed into swollen, misshapen, and branched cells called bacteroids. A microcolony of bacteroids becomes surrounded by portions of the plant cytoplasmic membrane to form a structure called the symbiosome (Figure 23.12c), and only after the symbiosome forms does N2 fixation begin. Nitrogen-fixing nodules can be detected experimentally by the reduction of acetylene to ethylene (Section 19.9). When the plant dies, the nodule deteriorates, releasing bacteroids into the soil. Although bacteroids are incapable of division, a small number of dormant rhizobial cells are always present in the nodule. These now proliferate, using some of the products of the deteriorating nodule as nutrients. The bacteria can then initiate infection the next growing season or maintain a free-living existence in the soil.
Nodule Formation: *nod* Genes, Nod Proteins, and Nod Factors
Rhizobial genes that direct the steps in nodulation of a legume are called nod genes. It is thought that the ability to form nodules has independently emerged multiple times through the horizontal transfer of such genes as nod and nif that are located on plasmids or transferable regions of chromosomal DNA. In Rhizobium leguminosarum biovar viciae, which nodulates peas, ten nod genes have been identified. The nodABC genes encode proteins that produce oligosaccharides called Nod factors; these induce root hair curling and trigger cell division in the pea plant, eventually leading to formation of the nodule (see Figure 23.16 for a description of root nodule biochemistry).
Nod factors are lipochitin oligosaccharides to which various substituents are bonded (Figure 23.13) that function as primary rhizobial signaling molecules triggering legumes to develop new plant organs: root nodules that host the bacteria as nitrogen-fixing bacteroids (Figure 23.12). Resolving the details of the signaling pathway triggered by Nod binding to cell surface receptors (NFR1 and NFR2) and leading to the induction of organogenesis (nodule formation) is an active area of research (Figure 23.14). Interestingly, many elements of the signaling pathway leading to nodulation are also used by the mycorrhizal fungi for infection of plant roots (Figure 23.14 and Section 23.5).
Figure 23.13 Nod and Myc factors.

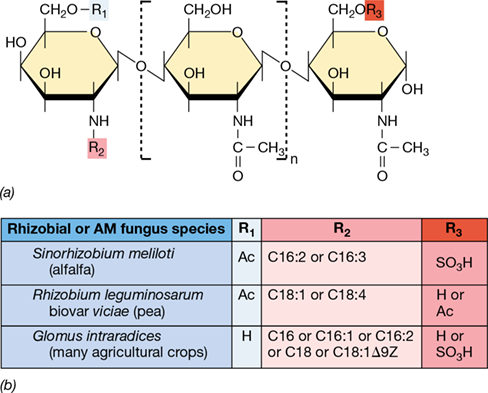
(a) General structure of Nod factors produced by two rhizobial species and the Myc factor produced by an arbuscular mycorrhizal (AM) fungus (Section 23.5). The central hexose unit can repeat up to three times for different Nod factors and repeat either two or three times for the different Myc factors. (b) Table of the structural differences (R1, R2, R3) that define the precise signaling factors of each species. C16:1, C16:2, and C16:3, palmitic acid with either one, two, or three double bonds, respectively; C18:1, oleic acid with one double bond; C18:1Δ9Z, the trans isomer of oleic acid with one double bond at the 9th C–C bond; C18:4, oleic acid with four double bonds; Ac, acetyl.
Figure 23.14 Nod and Myc signaling pathways in root nodule and mycorrhizal arbuscule formation.

Nod factor (NF) signaling involves at least three membrane-associated receptors (NFR1, NFR5, and SYMRK) that together initiate nodulation via protein phosphorylation. NFR1 and SYMRK have active kinase domains (blue), whereas NFR5 kinase is inactive. The direct binding of NF to a complex of NFR1 and NFR5 at the plant cell cytoplasmic membrane initiates signal transduction by activation of the NFR1 kinase. The resulting phosphorylation of the NFR cytoplasmic domains triggers events leading to formation of the infection thread. Signal transduction to the SYMRK by the NFR1–NFR5–Nod factor complex (or by an unidentified receptor for the Myc factor) is part of a conserved symbiosis program in which induction of calcium signaling in the plant cell nucleoplasm triggers gene expression changes and production of plant growth hormones (cytokinins) required for nodule or arbuscule formation. See Section 23.5 for a discussion of mycorrhizae.
Which plants a given rhizobial species can infect is in part determined by the structure of the Nod factor it produces. Besides the nodABC genes, which are universal and whose products synthesize the Nod backbone, each cross-inoculation group contains nod genes that encode proteins that chemically modify the Nod factor backbone to form its species-specific molecule (Figure 23.13). In R. leguminosarum biovar viciae, nodD encodes the regulatory protein NodD, which controls transcription of other nod genes. After interacting with inducer molecules, NodD promotes transcription and is thus a positive regulatory protein (Section 7.3). NodD inducers are plant flavonoids, organic molecules that are widely secreted by plants. Some flavonoids that are structurally very closely related to nodD inducers in R. leguminosarum biovar viciae inhibit nod gene expression in other rhizobial species (Figure 23.15). This indicates that part of the specificity observed between plant and bacterium in the rhizobia–legume symbioses is controlled by the chemistry of the flavonoids secreted by each species of legume.
Figure 23.15 Plant flavonoids and nodulation.

Structures of flavonoid molecules that are (a) an inducer of nod gene expression and (b) an inhibitor of nod gene expression in Rhizobium leguminosarum biovar viciae. Note the similarities in the structures of the two molecules. The common name of the structure shown in part a is luteolin, and it is a flavone derivative. The structure in part b is called genistein, an isoflavone derivative.
Mastering Microbiology
Art Activity: Figure 23.13 Nod and Myc signalling pathways in root nodule formation and mycorrhizal arbuscule formation
Biochemistry of Root Nodules
As discussed in Section 3.12, N2 fixation is catalyzed by nitrogenase. Nitrogenase from bacteroids shows the same biochemical properties as the enzyme from free-living N2-fixing bacteria, including O2 sensitivity and the ability to reduce acetylene as well as N2. Bacteroids are dependent on the plant for the electron donor for N2 fixation. The major organic compounds transported across the symbiosome membrane (Figure 23.12c) and into the bacteroid proper are citric acid cycle intermediates, in particular, the C4 organic acids succinate, malate, and fumarate (Figure 3.12; Figure 23.16). These are used as electron donors for ATP production and, following conversion to pyruvate, as the ultimate source of electrons for the reduction of N2.
Figure 23.16 The root nodule bacteroid.

Schematic of major metabolic reactions and nutrient exchanges in the bacteroid. The symbiosome is a collection of bacteroids surrounded by a membrane originating from the plant (see Figure 23.12c).
The product of N2 fixation is ammonia (NH3), and the plant assimilates most of this NH3 by forming organic nitrogen compounds. The NH3-assimilating enzyme glutamine synthetase is present in high levels in the plant cell cytoplasm and can convert glutamate and NH3 into glutamine (Section 3.14). Glutamine and a few other organic nitrogen compounds transport bacterially fixed nitrogen throughout the plant.
Stem-Nodulating Rhizobia
Although most leguminous plants form N2-fixing nodules on their roots, a few legume species bear nodules on their stems. Stem-nodulated leguminous plants are widespread in tropical regions where soils are often nitrogen deficient because of leaching and intense biological activity. The best-studied system is the tropical aquatic legume Sesbania, which is nodulated by the bacterium Azorhizobium caulinodans (Figure 23.17). Stem nodules typically form in the submerged portion of the stems or just above the water level. The general sequence of events by which stem nodules form in Sesbania resembles that of root nodules: attachment, formation of an infection thread, and bacteroid formation.
Figure 23.17 Stem nodules formed by stem-nodulating *Azorhizobium*.

The right side of this stem of the tropical legume Sesbania rostrata was inoculated with Azorhizobium caulinodans, but the left side was not.
Some stem-nodulating rhizobia produce bacteriochlorophyll a and thus have the potential to carry out anoxygenic photosynthesis (Section 14.5) in their illuminated habitat. Bacteriochlorophyll-containing rhizobia, called “photosynthetic Bradyrhizobium,” are widespread in nature, particularly in association with tropical legumes. In these species, light energy converted to chemical energy (ATP) in photosynthesis probably supplies part of the energy source needed by the bacterium to support N2 fixation.
Mastering Microbiology
Art Activity: Figure 23.15 The root nodule bacterioid
Nonlegume N2-Fixing Symbioses: *Azolla*–*Anabaena* and *Alnus*–*Frankia*
Various nonleguminous plants form N2-fixing symbioses with bacteria other than rhizobia. For example, the water fern Azolla harbors within small pores of its fronds a species of heterocystous N2-fixing cyanobacteria (Section 15.3) called Anabaena azollae (Figure 23.18). Azolla has been used for centuries to enrich Asian rice paddies with fixed nitrogen. Before planting rice, the farmer allows the surface of the rice paddy to become densely covered with Azolla. As the rice plants grow, they eventually crowd out the Azolla, causing its death and the release of its nitrogen, which is assimilated by the rice plants. By repeating this process each growing season, rice farmers can obtain high yields of rice without applying nitrogenous fertilizers.
Figure 23.18 *Azolla–Anabaena* symbiosis.

(a) Intact association showing a single plant of Azolla pinnata. The diameter of the plant is approximately 1 cm. (b) Cyanobacterial symbiont Anabaena azollae as observed in crushed leaves of A. pinnata. Single cells of A. azollae are about 5 μm wide. Vegetative cells are oblong; the spherical heterocysts (lighter color, arrows) are differentiated for nitrogen fixation.
The alder tree (genus Alnus) has N2-fixing root nodules (**Figure 23.19*a***) that harbor filamentous, N2-fixing actinomycetes of the genus Frankia. When assayed in cell extracts the nitrogenase of Frankia is sensitive to O2, but cells of Frankia fix N2 at full oxygen tensions. This is because Frankia protects its nitrogenase from O2 by localizing the enzyme in terminal swellings on the cells called vesicles (Figure 23.19b). The vesicles contain thick walls that retard O2 diffusion, thus maintaining the O2 tension within vesicles at levels compatible with nitrogenase activity. In this regard, Frankia vesicles resemble the heterocysts produced by some filamentous cyanobacteria as localized sites of N2 fixation (Sections 8.9 and 15.3).
Figure 23.19 *Frankia* nodules and *Frankia* cells.

(a) Root nodules of the common alder Alnus glutinosa. (b) Frankia culture purified from nodules of Comptonia peregrina. Note vesicles (arrows) on the tips of hyphal filaments.
Alder is a characteristic pioneer tree able to colonize nutrient-poor soils, probably because of its ability to enter into a symbiotic N2-fixing relationship with Frankia. A number of other small or bushy, woody plants are nodulated by Frankia. As is the case for the rhizobial symbionts of leguminous plants, a single strain of Frankia can form nodules on several different species of plants.
Check Your Understanding
What is a bacteroid, and what occurs within it? What is the function of leghemoglobin?
What are the major similarities and differences between rhizobia and Frankia?
23.5 Mycorrhizae
Mycorrhizae are mutualisms between plant roots and fungi in which nutrients are transferred in both directions. The fungus transfers inorganic nutrients—in particular, phosphorus and nitrogen—from the soil to the plant, and the plant in turn transfers primarily carbohydrates to the fungus. These mutualisms are harnessed in agricultural applications. From fungal spores produced in culture or from root scrapings of infected plants, soil inoculants are produced that enhance plant growth.
Classes of Mycorrhizae
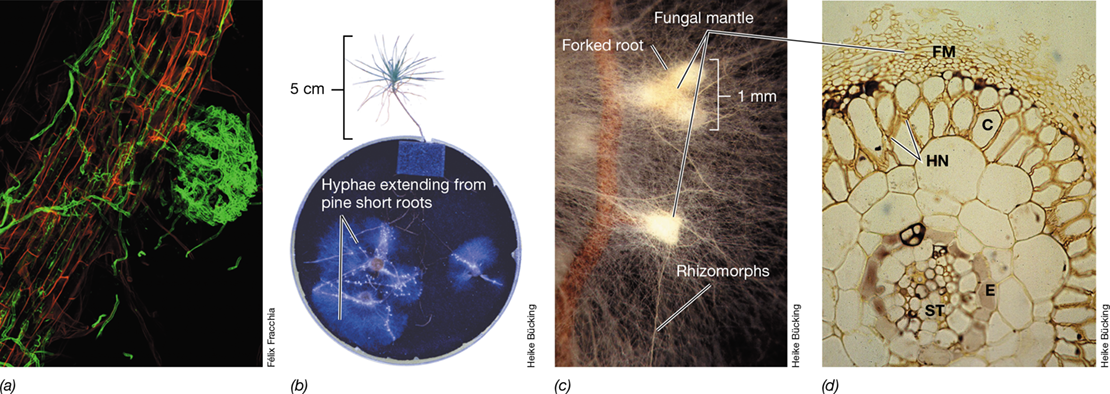
There are two classes of mycorrhizae. In ectomycorrhizae, fungal cells form an extensive sheath (fungal mantle) around the outside of the root with only a slight penetration into the root cellular structure (Figure 23.20). As rootlets emerge, they are rapidly colonized by the fungi (Figure 23.20a). In endomycorrhizae, a part of the fungus becomes deeply embedded within cells comprising the root tissue. Ectomycorrhizae are found mainly on the roots of forest trees, especially conifers, beeches, and oaks, and are most highly developed in boreal and temperate forests. In such forests, almost every root of every tree is mycorrhizal. The root system of a mycorrhizal tree such as a pine (genus Pinus) is composed of both long and short roots (Figure 23.20b, c). The short roots, which are characteristically dichotomously branched in Pinus (Figure 23.20b, c), show typical fungal colonization, and long roots are also frequently colonized.
Figure 23.20 Ectomycorrhizal colonization of pine and beech tree roots.

(a) Confocal image of an ectomycorrhizal fungus (stained green) aggregating on an emerging tree rootlet (stained red) and then “running” on the surface as it begins to further colonize the root. (b) Colonization of pine (Pinus sylvestris) roots with the ectomycorrhizal fungus Suillus bovinus. (c) Ectomycorrhizal root tips of pine roots enclosed by a mantle of ectomycorrhizal fungi (white) and associated hyphae extending into the soil matrix. The ectomycorrhizal fungus Suillus bovinus also forms rhizomorphs, hyphal aggregations that are involved in the long-distance transport from the soil to the mycorrhizal root. (d) Cross section of a thin beech root showing the fungal mantle (FM) and Hartig net (HN) within the root cortex (C). The Hartig net is the location of nutrient exchange between the plant and fungus. Also shown are root vascular tissue (ST) and endodermis (E).
Ectomycorrhizal hyphae extending from the fungal mantle and penetrating between the epidermal and cortical cells form a network called the Hartig net (Figure 23.20d) where nutrient exchange between the fungus and the host plant occurs that benefit the plant (see Figure 23.23). Most mycorrhizal fungi do not catabolize cellulose and other leaf litter polymers. Instead, they catabolize simple carbohydrates and typically have one or more vitamin requirements. They obtain their carbon from root secretions and obtain inorganic minerals from the soil. Mycorrhizal fungi are rarely found in nature except in association with roots, and most are probably obligate symbionts.
Despite the close symbiotic association between fungus and root, a single species of tree can form multiple mycorrhizal associations. One pine species can associate with over 40 species of fungi. This relative lack of host specificity allows ectomycorrhizal mycelia to interconnect trees, providing linkages for transfer of carbon and other nutrients between trees of the same or different species. Nutrient transfer from well-illuminated overstory plants to shaded trees is thought to help equalize resource availability, subsidizing young trees and increasing biodiversity by promoting the coexistence of different species.
Arbuscular Mycorrhizae
Although ectomycorrhizal fungi play a significant role in the ecology of forests, there is a greater diversity of endomycorrhizae. Most are arbuscular mycorrhizae (AM) that comprise a phylogenetically distinct fungal division, the Glomeromycota (Section 18.12), of which all or most species are obligate plant mutualists (the word arbuscular means “little tree”). AM colonize 70–90% of all terrestrial plants, including most grassland species and many crop species. The association between plants and the Glomeromycota is thought to be the ancestral type of mycorrhizae, established about 450 million years ago and an important evolutionary step in the successful invasion of dry land by terrestrial plants.
AM fungi produce lipochitin oligosaccharide signaling factors (Myc factors) closely related to the Nod factors in the rhizobium–legume symbiosis (Section 23.4), and Myc factors initiate formation of the mycorrhizal state (Figures 23.13 and 23.14). Root colonization by an AM fungus begins with germination of a soilborne spore, producing a branched germination mycelium that recognizes the host plant through reciprocal chemical signaling. Spore germination and mycelial branching is induced by strigolactones (**Figure 23.21a,*b***), plant hormones released by the roots that also play a key role in plant development. When a plant is nutrient-limited, this hormone represses aboveground plant growth (suppressing formation of secondary shoots) and stimulates the growth of the root system, enhancing the production of lateral roots and root hairs. These developmental changes help the plant secure nutrients before using them later for aboveground growth.
Figure 23.21 Arbuscular mycorrhizal root colonization.

(a) A spore (S) recognizes the presence of a nearby root by sensing strigolactones released by the plant root. (b) The structure of strigol, one type of strigolactone. Strigolactones stimulate spore germination and mycelial branching. Myc factor (see Figure 23.13) produced by the growing fungal mycelium then initiates the infection process. (c) The mycelium forms an attachment structure called the hyphopodium (HP) and then enters the inner cortex region of the root by penetrating epidermal cells and cells of the outer cortex. Arbuscules (dichotomously branched invaginations, A) are formed by mycelia spreading either intercellularly (left) or intracellularly (right). (d) Image of arbuscules in the root cells of Medicago truncatula (barrel clover) colonized by the fungus Rhizophagus irregularis engineered to express the green fluorescent protein.
Mastering Microbiology
Art Activity: Figure 23.20c Arbuscular mycorrhizal root colonization
The Myc factor produced by the AM fungal mycelium signals the plant to initiate the developmental process (Figure 23.14). The fungus then forms a contact structure called the hyphopodium with root epidermal cells (Figure 23.21c). Penetrating hyphae extend into the plant from each hyphopodium, usually forging an intracellular path through epidermal and outer cortical cell layers of the root before forming dichotomously branched or coiled hyphal structures called arbuscules within plant inner cortex cells, near vascular tissues (Figure 23.21c, d). However, the arbuscular hyphae remain separated from plant protoplasm by an extensive plant cytoplasmic membrane that forms a region called the apoplast (Figure 23.22), and this structure functions to increase the surface area of contact between plant and fungus. Inorganic nitrogen and phosphorus are then “mined” from the soil by the fungi, converted to arginine and polyphosphate, and translocated through the hyphae to the plant (Figure 23.22).
Figure 23.22 Pathways of N, P, and C exchange between plant and arbuscular mycorrhizal fungi.

Inorganic nitrogen (NH4 + and NO3 −) and phosphorus (Pi) mined from the soil by the extraradical (soil-associated) mycelia are translocated to the plant as arginine and polyphosphate (poly-P) through the mycelial network and delivered to the plant at the intraradical (plant cell–associated) mycelium. Ammonia and phosphate are regenerated in the intraradical mycelium for transfer to the plant cell. In exchange for the N and P, the plant provides organic carbon to the fungus.
Myc factors are very similar to the rhizobial Nod factors and only relatively minor modifications of the chitin backbone structure confer specificity (Figure 23.13). It is likely that the basic signaling and developmental systems used in the legume–root nodule symbiosis (Section 23.4), which arose about 60 million years ago, first evolved in the much more ancient AM fungi–plant symbiosis and were recruited and adapted for the legume–root nodule symbiosis (Figure 23.14).
Although the arbuscular mycorrhizae are a much more ancient and widely distributed microorganism–plant symbiosis, understanding of their signaling and developmental program has been slower to develop because AM fungi cannot be grown in pure culture. AM fungi are obligately biotropic (meaning that they obtain their nutrients only from living cells of their symbiotic partner), and unlike rhizobia, they have no supporting genetic system that can be exploited to unravel the developmental steps leading to the plant–fungus association.
Benefits for the Plant
The beneficial effect of the mycorrhizal fungus on the plant is best observed in poor soils where plants that are mycorrhizal thrive, but nonmycorrhizal ones do not. For example, if trees planted in prairie soils, which ordinarily lack a suitable fungal inoculum, are artificially inoculated at the time of planting, they grow much more rapidly than uninoculated trees (Figure 23.23). The mycorrhizal plant can absorb nutrients from its environment more efficiently (Figure 23.22) and thus has a competitive advantage. This improved nutrient absorption is due to the greater surface area provided by the fungal mycelium (see Figure 23.23 inset). In the pine seedling shown in Figure 23.20b and c, the ectomycorrhizal fungal mycelium makes up the overwhelming part of the absorptive capacity of the plant root system. The mycorrhizal plant is better able to function physiologically and compete successfully in a species-rich plant community, and the fungus benefits from a steady supply of organic nutrients.
Figure 23.23 Effect of mycorrhizal fungi on plant growth.

Six-month-old seedlings of Monterey pine (Pinus radiata) growing in pots containing prairie soil: left, nonmycorrhizal; right, mycorrhizal. Inset: A Pinus seedling showing the extensive fanlike development of its fungal partner. Also see Figure 23.20b, c.
In addition to helping plants absorb nutrients, mycorrhizae also play a significant role in supporting plant diversity. Field experiments clearly show a positive correlation between the abundance and diversity of mycorrhizae in a soil and the extent of the plant diversity that develops in it. However, although most mycorrhizae are true mutualistic symbioses, there are also parasitic mycorrhizae. In these less common mycorrhizal symbioses, either the plant parasitizes the fungus or, in some cases, the fungus parasitizes the plant.
There is much more to learn about fungal–plant symbioses, and such discoveries will be important not only in agriculture but also in facilitating the restoration of damaged ecosystems.
Check Your Understanding
How do endomycorrhizae differ from ectomycorrhizae?
What features of mycorrhizal fungi might have assisted in colonization of dry land by plants?
23.6 *Agrobacterium* and Crown Gall Disease
Some microorganisms develop parasitic symbioses with plants. The genus Agrobacterium, a relative of the root nodule bacterium Rhizobium (Section 23.4), is such an organism, causing the formation of tumorlike growths on diverse plants. The two species of Agrobacterium most widely studied are Agrobacterium tumefaciens (also called Rhizobium radiobacter), which causes crown gall disease, and Agrobacterium rhizogenes (also called Rhizobium rhizogenes), which causes hairy root disease.
The Ti Plasmid
Although wounded plants often form a benign accumulation of tissue called a callus, the growth in crown gall disease (Figure 23.24) is different in that it is uncontrolled growth, resembling an animal tumor. A. tumefaciens cells induce tumor formation only if they contain a large plasmid called the Ti plasmid (Ti for tumor inducing). In A. rhizogenes, a similar plasmid called the Ri plasmid is necessary for induction of hairy root disease. Following infection, a part of the Ti plasmid called the transferred DNA (T-DNA) is integrated into the plant’s genome. T-DNA carries the genes for tumor formation and also for the synthesis of a number of modified amino acids called opines. Octopine [N2-(1,3-dicarboxyethyl)-L-arginine] and nopaline [N2-(1,3-dicarboxypropyl)-L-arginine] are two common opines. Opines are produced by plant cells transformed by T-DNA and are a source of carbon and nitrogen, and sometimes phosphate, for the parasitic A. tumefaciens cells. These nutrients are the benefits for the bacterial symbiont.
Figure 23.24 Crown gall.

Photograph of a crown gall tumor (arrow) on a tobacco plant caused by Agrobacterium tumefaciens. The disease usually does not kill the plant but may weaken it and make it more susceptible to drought and diseases.
Recognition and T-DNA Transfer
To initiate the tumorous state, A. tumefaciens cells attach to a wound site on the plant. Following attachment, the synthesis of cellulose microfibrils by the bacteria helps anchor them to the wound site, and bacterial aggregates form on the plant cell surface. This sets the stage for plasmid transfer from bacterium to plant. The general structure of the Ti plasmid is shown in Figure 23.25. Only the T-DNA is actually transferred to the plant. The T-DNA contains genes that induce tumorigenesis. The vir genes on the Ti plasmid encode proteins that are essential for T-DNA transfer. Transcription of vir genes is induced by metabolites synthesized by wounded plant tissues such as the phenolic compounds acetosyringone and ferulate. The transmissibility genes on the Ti plasmid (Figure 23.25) allow the plasmid to be transferred by conjugation (Sections 9.8 and 9.9) from one bacterial cell to another.
Figure 23.25 Structure of the Ti plasmid of *Agrobacterium tumefaciens*.

T-DNA is the region transferred to the plant. Arrows indicate the direction of transcription of each gene. The entire Ti plasmid is about 200 kilobase pairs (kbp) of DNA and the T-DNA is about 20 kbp.
The vir genes are the key to T-DNA transfer. The virA gene encodes a protein kinase (VirA) that interacts with inducer molecules and then phosphorylates the product of the virG gene (Figure 23.26). VirG is activated by phosphorylation and functions to activate other vir genes. The product of the virD gene (VirD) has endonuclease activity and nicks DNA in the Ti plasmid in a region adjacent to the T-DNA. The product of the virE gene is a DNA-binding protein that binds the single strand of T-DNA in the plant cell to protect it from destruction by nucleases. The virB operon encodes 11 different proteins that form a type IV secretion system (Section 6.13) for single-strand T-DNA and protein transfer between bacterium and plant (Figure 23.26) and thus resembles bacterial conjugation. Laboratory studies of A. tumefaciens have shown that it can transfer T-DNA into many types of eukaryotic cells, including fungi, algae, protists, and even human cell lines.
Figure 23.26 Mechanism of transfer of T-DNA to the plant cell by *Agrobacterium tumefaciens.*

(a) VirA activates VirG by phosphorylation, and VirG activates transcription of other vir genes. (b) VirD is an endonuclease that nicks the Ti plasmid, exposing the T-DNA. (c) VirB functions as a conjugation bridge between the A. tumefaciens cell and the plant cell, and VirE is a single-strand binding protein that assists in T-DNA transfer. Plant DNA polymerase produces the complementary strand to the transferred single strand of T-DNA.
Mastering Microbiology
Art Activity: Figure 23.25 Mechanism of transfer of T-DNA to plant cells by Agrobacterium tumefaciens
Once inside the plant cell, T-DNA becomes inserted into the genome of the plant. Tumorigenesis (onc) genes on the Ti plasmid (Figure 23.25) encode enzymes for plant hormone production and at least one key enzyme of opine biosynthesis. Expression of these genes leads to tumor formation and opine production. The Ri plasmid responsible for hairy root disease also contains onc genes. However, in this case the genes confer increased auxin responsiveness on the plant, and this promotes overproduction of root tissue and the symptoms of the disease. The Ri plasmid also encodes several opine biosynthetic enzymes.
Genetic Engineering with the Ti Plasmid
From the standpoint of microbiology and plant pathology, crown gall disease and hairy root disease both require intimate interactions that lead to genetic exchange from bacterium to plant. In other words, tumor induction in these diseases is the result of a natural plant-transformation system. Because of this, recent interest in the Ti–crown gall system has shifted away from the disease itself toward applications of this natural genetic exchange process in plant genetic engineering and biotechnology.
Several modified Ti plasmids that lack disease genes but that can still transfer DNA to plants have been developed by genetic engineering. These have been used for the construction of genetically modified (transgenic) plants. Many transgenic plants have been constructed thus far, including crop plants carrying genes for resistance to herbicides, insect attack, and drought. We discuss the use of the Ti plasmid as a vector in plant biotechnology in Section 12.7.
Now we transition away from plants and look at some remarkable symbioses between bacteria and insects.
Check Your Understanding
How do the vir genes differ from T-DNA in the Ti plasmid?
How has an understanding of crown gall disease benefited plant agriculture?
III Insects as Microbial Habitats
Mutualisms between microorganisms and insects are common, contributing nutritional advantages or protection from pathogens and predators. Bacterial symbionts can reside within specialized insect cells, associate with specialized surface structures, or colonize the insect gut.
Insects are the most abundant class of animals living today, with over 1 million species known. As many as 20% of all insects are thought to support symbiotic microbes in a mutually beneficial way. The symbioses contribute to the insects’ ecological success by providing them either nutritional advantages or protection from parasites. Some symbionts are found on insects’ outer surfaces or in their digestive tracts. Endosymbionts are intracellular bacteria and are typically localized at specialized organs within the insect.
23.7 Heritable Symbionts of Insects
How symbionts are transferred from one generation to the next determines how a mutualism functions and how stable it is. Microbial symbionts can either be acquired by a host from an environmental reservoir (horizontal transmission) or be transferred directly from the parent to the next generation (heritable or vertical transmission). The mode of symbiont transmission is related to the specificity and persistence of an association. In general, less specificity is associated with horizontal transmission. In this section we focus only on mutualisms in which the microbial symbiont has no free-living form; that is, the symbionts are transmitted in a vertical fashion.
Types of Heritable Symbionts
All known heritable microbial symbionts of insects lack a free-living replicative stage. Thus, they are obligate symbionts. However, although these bacteria require the host for replication, not all hosts are dependent upon the symbiont. Relative to host dependence, heritable symbionts are either primary or secondary symbionts. Primary symbionts are required for host reproduction. They are restricted to a specialized region called the bacteriome present in several insect groups; within the bacteriome the bacterial cells reside in specialized cells called bacteriocytes. Secondary symbionts are not required for host reproduction. Unlike primary symbionts, secondary symbionts are not always present in every individual of a species and are not restricted to particular host tissues.
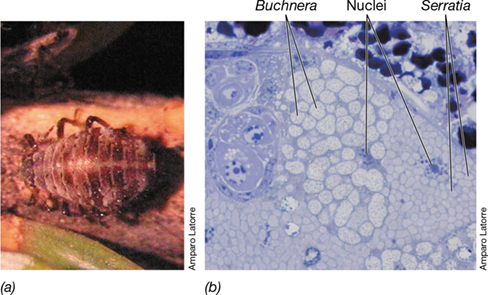
Secondary symbionts are broadly distributed among insect groups. Like pathogens, they invade different cell types and may live extracellularly within the insect’s hemolymph (the fluid bathing the body cavity). In insects with bacteriomes, such as the cedar aphid (**Figure 23.27*a***), secondary symbionts can invade the bacteriocytes, co-residing with or sometimes displacing the primary symbionts (Figure 23.27b). However, in order to persist in the insect host, the secondary symbiont must confer some advantage, such as a nutritional advantage or protection from environmental stresses such as heat. For example, whiteflies infected with the bacterium Rickettsia (Section 16.1) produce offspring at about twice the rate of uninfected flies, and more offspring survive to adulthood. Secondary symbionts may also provide protection against invasion by pathogens or predators. A Spiroplasma species (Section 16.9), which was first observed in Drosophila neotestacea in the 1980s, provides protection against a parasitic nematode worm. In most instances the basis for increased fitness or protection is unknown, but in one case a toxin encoded by a lysogenic bacteriophage (Section 5.6) carried by the symbiont is known to confer protection on the insect from infection by a parasitic wasp.
Figure 23.27 Primary and secondary symbionts of an aphid.

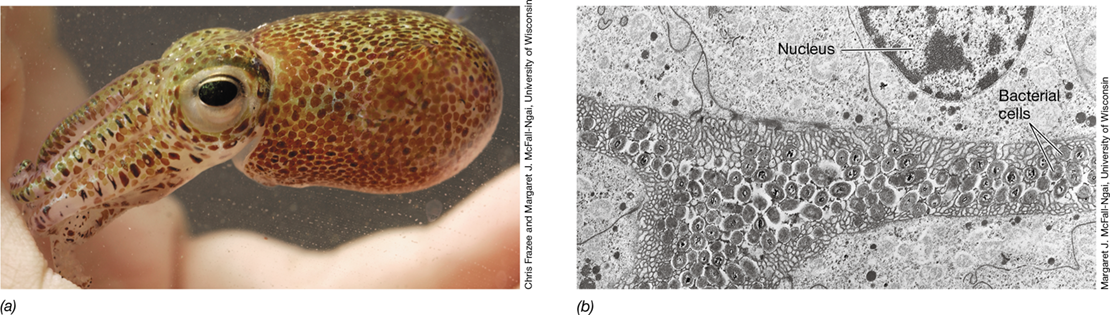
(a) The cedar aphid Cinara cedri, a model organism for studies of symbioses. (b) Transmission electron micrograph of the bacteriome of C. cedri showing two bacteriocytes. Packed within each bacteriocyte are cells of Buchnera aphidicola (the primary symbiont) or Serratia symbiotica, the smaller, secondary symbiont. The nucleus of each bacteriocyte is identified. The bacteriocyte containing Buchnera cells is about 40 μm wide.
There are heritable parasitic symbionts that manipulate the host’s reproductive system, increasing the frequency of female progeny (sex-ratio skewing, Figure 16.27). Because most heritable symbionts are transmitted maternally, the suppression of male progeny serves to expand the number of infected individuals and increase the rate of spread through an insect population. Since symbiont-conferred functions can spread rapidly within a population, acquisition of symbiont-encoded traits provides a mechanism for much more rapid adaptation than is possible through mutations in insect genes. Rickettsia infection of the whitefly population provides one example of how rapidly symbiont-conferred traits can spread through a population. Only 1% of whiteflies tested in the southwestern United States were infected with Rickettsia in 2000. In 2006, 97% of flies were infected.
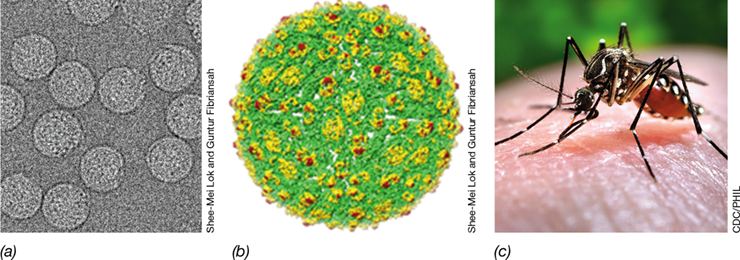
An important applied benefit of improved basic understanding of insect symbionts is the increased use of symbionts in insect pest management and the control of vectorborne diseases, such as malaria and filariasis in humans (Sections 34.5 and 34.7). For example, symbiotic Wolbachia, which are reproduction manipulators, are widely distributed among insect species (possibly infecting as many as 60–70% of all insect species). The sperm of Wolbachia-infected males can sterilize uninfected females. Although the mechanism for sterilization is not fully understood, the phenomenon is being tested as a means to suppress mosquito-borne human viral disease transmission (see Explore the Microbial World, “Combating Mosquito-Borne Viral Diseases with an Insect Symbiont”).
Nutritional Significance of Obligate Intracellular Symbionts of Insects
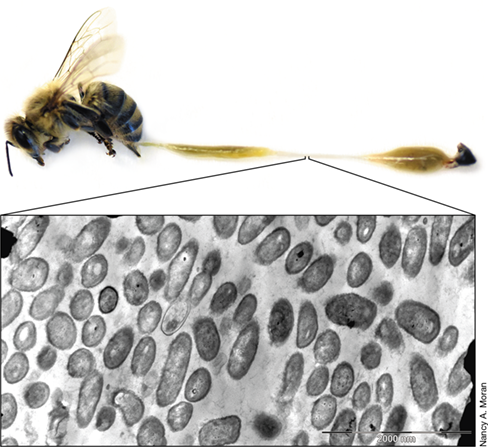
The association of bacteria and insects has allowed many insects to use food resources that are rich in some nutrients, but poor in others. To achieve adequate nutrition, some insects exploit the metabolic potential of their symbionts. For instance, aphids feed on the carbohydrate-rich but otherwise nutrient-poor sap of phloem vessels in plants. Early on it was suspected that obligate symbionts might benefit the insect by providing nutrients not provided by their primary diet, and this is now known to be true.
Molecular analyses have shown that most families of aphids harbor the bacterium Buchnera in their bacteriomes. The role of Buchnera in host nutrition was first indicated by experiments using defined diets to examine the nutrient requirements of aphids. Compared with infected controls, symbiont-free aphids required a diet containing all amino acids that are either lacking or rare in phloem sap. Subsequent genomic studies documented the presence in Buchnera of genes encoding the biosynthesis of nine amino acids missing from the sap. There are also examples of synergy between host and symbiont where the synthesis of certain amino acids becomes a joint venture. For example, Buchnera lacks the enzyme needed for the last step in leucine biosynthesis, but the necessary gene is present in the aphid’s genome. Presumably, this enzyme is made by the aphid and participates in the leucine biosynthetic pathway along with the bacterial enzymes.
A secondary symbiont can also contribute to a joint venture. For example, the Buchnera symbiont of the cedar aphid is unable to supply tryptophan to the aphid. Two genes in the tryptophan biosynthetic pathway are present in Buchnera, but the remaining genes for the pathway are located on the chromosome of a secondary endosymbiont (Figure 23.27). Thus, different parts of a required metabolic pathway can be encoded by different endosymbionts present in the same insect. The fungus-cultivating ants provide yet another example of a complex symbiosis that has formed between an insect and multiple microorganisms (as we review in the next section).
Mealybugs (Planococcus citri) present one of the most unusual examples of a partnership between two symbionts infecting the same insect. Mealybugs have two stable bacterial symbionts, “Candidatus Tremblaya princeps” (a Betaproteobacterium) and “Candidatus Moranella endobia” (a Gammaproteobacterium). (The term “Candidatus” means that these organisms are not yet in pure culture, Section 13.12, but we refer to them in the usual style below.) These symbionts cooperate in providing their host with essential amino acids missing in its diet, as is true for the symbionts of many sap-feeding insects. However, Moranella actually lives inside of Tremblaya, the only known example of a bacterium-within-a-bacterium symbiosis. The highly reduced Tremblaya genome has lost all genes for tRNA synthetases, an essential function supplied either by the host or by the Moranella residing within the cytoplasm of Tremblaya.
Genome Reduction and Gene Transfer Events
Common features of primary insect symbionts are extreme genome reduction (Table 10.1), high adenine plus thymine content DNA, and accelerated rates of mutation. Genomes of most insect symbionts fall within a range from 0.12 to 0.80 megabase pairs (Mbp) and 16.5 to 33% G + C (Table 23.2). The 0.14-Mbp (140 kilobase-pair) genome of Tremblaya princeps is among the smallest genomes known for any cell. In contrast, the genomes of related free-living bacteria range from 2 to 8 Mbp with a base composition closer to 50% G + C. Two common types of spontaneous mutation, cytosine deamination and the oxidation of guanosine, if not repaired, change a GC pair to an AT pair (Section 9.4). Symbionts with reduced genomes have fewer DNA repair enzymes (Section 9.4), and this likely facilitates a shift over time to genomes of lower G + C content.
Table 23.2 Genome features of some endosymbiotic *Bacteria* of animalsa

aAll listed symbionts are obligately associated with their hosts, with the exception of the symbiont of Riftia, which also has a free-living stage. For a comparison with the genomes of free-living Bacteria, see Table 10.1.
bThe free-living sulfur-oxidizing bacterium Thiomicrospira crunogena has a genome significantly smaller (2.4 Mb) than this symbiont.
The streamlined genomes of insect symbionts have lost genes from most functional categories (Chapter 10) and tend to retain only genes required for host fitness and essential molecular processes, such as translation, replication, and transcription. Genome reduction implies that the symbionts are reliant on the host for many functions no longer encoded in the symbiont genome (Section 10.3). For example, in many cases genes needed for the biosynthesis of cell wall components are missing, including lipid A and peptidoglycan, suggesting that the host supplies these functions or that the structures are not required to form stable cells within the bacteriocyte.
There is an interesting genomic contrast between primary symbionts and typical disease-causing bacteria (pathogens). While primary symbionts tend to lose genes encoding proteins required in catabolic pathways, pathogenic bacteria typically retain these, but lose genes for anabolic pathways. This reflects their differing relationships with their hosts; the insect symbiont provides the host with essential biosynthetic nutrients while the pathogen obtains important biosynthetic nutrients from the host.
Because genome sequences for a large number of insects and their symbionts are now appearing, microbiologists can begin to evaluate the frequency of gene transfer between them. Horizontal gene transfer is the movement of genetic information across normal mating barriers (Chapter 9 and 13). Although early research demonstrated that DNA of Wolbachia has been transferred to the nuclear genomes of their insect and nematode hosts, inspection of other insect mutualisms for which both host and symbiont genome sequences are available (e.g., aphid and body louse) indicate that DNA transfer is very rare. Even though transfer of genes between insect symbiont and host genomes appears to be very rare, as we will see in the next section, there can be extensive movement of genetic information between symbionts that associate with a specific insect host.
Check Your Understanding
What factors stabilize the presence of a secondary insect symbiont?
What are the consequences of symbiont genome reduction?
How could it be determined if a symbiont and its host have experienced a long period of coevolution?
23.8 Defensive Symbioses
In addition to symbioses that provide nutritional benefits, other symbioses between insects and microbes provide the insect with a defense weapon. A widespread defensive strategy used by insects to deter pathogens and predators is the production of toxic and antimicrobial chemicals. The chemical deterrent is sometimes produced by the insect. For example, a toxic mix of hydroquinone and hydrogen peroxide is prepared in a specialized abdominal chamber of the bombardier beetle before being squirted at an approaching predator. However, in many more instances, the defensive chemical is the product of microorganisms symbiotically associated with the insect.
The Rove Beetle
An example of this defensive strategy can be found in the Paederus beetle (commonly called the rove beetle), which deters predators using the chemical pederin (Figure 23.28) synthesized by an endosymbiotic Pseudomonas species. This cytotoxic chemical (inhibiting mitosis in eukaryotes) accumulates in the insect’s hemolymph and is deposited in its eggs, effectively deterring arthropod predation on the eggs. The beetle is better known in the tropics for causing severe dermatitis and blistering of the human skin if the insect is inadvertently crushed when brushed away, releasing the pederin-containing hemolymph.
Figure 23.28 Defensive chemical of insects.

(a) The rove beetle. (b) The insect’s bacterial symbiont produces large amounts of pederin, a highly cytotoxic defensive chemical that accumulates in the hemolymph of the beetle.
Many such defensive symbioses are now recognized, protecting the insect from predation, parasitism, and microbial pathogens. Defensive chemicals are made by a wide variety of bacterial symbionts, including members of the Proteobacteria, Firmicutes, and Actinobacteria. Some are intracellular, others reside within the gut, and others are associated with specialized anatomical structures on the insect exoskeleton. Comparative metagenomic analyses (Sections 10.7 and 19.8) of symbiont gene content has shown that the array of defensive chemicals deployed by individual symbionts is primarily determined by horizontal gene transfer. Pathways for chemical synthesis commonly reside on plasmids and other mobile genetic elements (Sections 6.2 and 9.11), such that even closely related symbionts can express very different defensive chemicals. Specific associations between insects and defensive symbionts are maintained by a variety of transmission mechanisms, including strictly vertical inheritance from the mother through the germ line and horizontal recruitment from the environment in each generation.
The Leafcutter Ants
The attine ants, more generally called leafcutter ants, are an example of an elaborate symbiotic association between multiple microbial species and insect. These ants have established an obligate mutualism with a fungus they cultivate, using small leaf fragments to feed the fungus. A close symbiotic relationship between ant and fungus was first indicated by the observation that one specific fungus was cultivated by each ant lineage. In addition to this close mutualistic relationship, the symbioses include four other microbial symbionts: a small fungus that is parasitic on the cultivated fungus, nitrogen-fixing bacteria (Sections 3.12 and 15.9) associated with the cultivated fungus, an actinobacterium that antagonizes the parasitic fungus, and a black yeast that interferes with the actinobacterium.
The fungus is vertically transmitted between ant generations by colony-founding queens. The queen collects a pellet of fungus and, after mating, uses the fungus pellet to establish a new nest containing the cultivated fungus (Figure 23.29). Nitrogen-fixing Klebsiella and Pantoea species associated with the fungus enrich the nutritional quality of the garden by adding new nitrogen to the nitrogen-poor leaf growth substrate. The cultivated fungus is susceptible to a parasitic fungus of the genus Escovopsis, and to repel this parasite, the ant has formed another symbiotic association with an actinobacterium (genus Pseudonocardia) that appears as a “waxy bloom” growing on the cuticle of the ant (Figure 23.29b). These bacterial cells, housed in specialized cuticular modifications on the ant’s body, secrete highly selective antimicrobial agents that inhibit the growth of Escovopsis but not the cultivated fungus.
Figure 23.29 Leafcutter attine ants.

(a) Queen and worker ants cultivating fungi. (b) Mutualism with the symbiont Pseudonocardia covers much of the exoskeleton of workers (white areas). The antifungal agent produced by the symbiont kills parasitic fungi but not the cultured fungi.
The Pseudonocardia cells likely receive nourishment from the ant in glandular secretions through pores localized in regions of cuticular modification. Comparative genomic sequencing has revealed good congruence between the phylogenies of the ants, fungal cultivars, Escovopsis, and Pseudonocardia, pointing to very specific interactions among microbes and ants in this complex symbiosis. The fourth and final microorganism identified in this symbiosis is a yeast that grows in the same cuticular regions colonized by the bacterium Pseudonocardia. This black-pigmented yeast interferes with chemical protection of the cultivated fungi by stealing nutrients from Pseudonocardia, thereby reducing its ability to suppress Escovopsis growth. The attine ant–microbial symbiosis is thus a complex maze of interactions between a macroorganism—the ant—and two groups of microbes, fungi and bacteria.
The Gut Microbiome of Bees
Bees and other animal pollinators provide vital ecosystem services. Thus, the loss of nearly 60% of honeybee colonies in the United States since 1947 is a major concern. Several factors are thought to contribute to this decline including fragmentation of habitats, pesticides, pollution, land use changes, and climate change. The decline of bee populations has also drawn attention to the importance of the bee gut microbiome for combating bee pathogens and environmental stressors.
The gut communities of bumblebees and honeybees (Figure 23.30) are surprisingly simple and consist of a core community of only five dominant and culturable bacterial species in adult worker bees. These include two gram-negative Proteobacteria (Snodgrassella alvi and Gilliamella apicola), two Lactobacillus species, and a Bifidobacterium species. S. alvi and G. apicola pack the lumen of the honeybee gut (Figure 23.30). Occasionally, up to four other species are present. Although different strains of each species in the core microbial community of the honeybee gut are closely related in 16S rRNA gene sequence, their genomes have diverged significantly through coevolution with bees over millions of years. As a result, strains have become host-specific. For example, S. alvi isolated from honeybees will not colonize bumblebees, and vice versa.
Figure 23.30 Scanning electron micrograph of honeybee gut microbiome.
Five dominant bacterial species pack the lumen of the honeybee midgut as shown in the thin sectioned transmission electron micrograph.
Metabolic interdependencies exist among microbes and the bee. For instance, S. alvi oxidizes the fermentation products of G. apicola, and G. apicola has genes for the degradation of pectin, a polysaccharide in the cell wall of pollen grains. Because bees do not produce pectinase and pollen is a key component of their diet, bee gut microbiota are essential for utilization of this important food source. Some bee gut microbes also carry genes for the utilization of uncommon sugars such as arabinose and raffinose that are indigestible and potentially toxic to the bee. This finely tuned microbiome plays a defensive role for the insect in that it inhibits invasive microorganisms through direct competition for space and resources and by stimulating the insect to produce antimicrobial peptides that target pathogenic bacteria.
Check Your Understanding
How does the leafcutter ant prevent its cultivated fungus from being parasitized by another fungus?
What is unusual about the diversity of the honeybee gut microbiome?
How does the rove beetle use its Pseudomonas symbiont for protection?
23.9 Termites
Microorganisms are primarily responsible for the degradation of wood and cellulose in nature. However, the activities of free-living microbial species have been exploited by certain groups of insects that have established microbial symbioses in order to digest lignocellulosic materials. Like the rumen of many herbivorous animals (see Section 23.15), the insect gut provides a protective niche for microbial symbionts, and in return, the insect gains access to nutrients derived from an otherwise indigestible carbon source. Termites are among the most abundant representatives of this type of symbiotic alliance.
Termite Natural History and Biochemistry
Microbial symbionts in termites decompose the greater part of cellulose and hemicellulose in the plant material termites ingest. In contrast to the heritable insect symbionts discussed in Section 23.7, most termites do not harbor intracellular bacteria. Instead, the symbiotic bacteria are present in digestive organs (guts) as in the case of mammals. Termite diets include lignocellulosic plant materials (either intact or at various stages of decay), dung, and soil organic matter (humus). About two-thirds of the terrestrial environment supports one or more termite species, with the greatest representation in tropical and subtropical regions, where termites may constitute as much as 10% of all animal biomass and 95% of soil insect biomass.
Termites are categorized as higher or lower based on their phylogeny, and this classification correlates with different symbiotic strategies. The posterior alimentary tract of higher termites (family Termitidae, comprising about three-fourths of termite species) contains a dense and diverse community of mostly anaerobic bacteria, including cellulolytic species. In contrast, the lower termites harbor diverse populations of both anaerobic bacteria and cellulolytic protists. Bacteria of lower termites participate little or not at all in cellulose digestion; only the protists phagocytize and degrade the wood particles ingested by the termites.
The termite gut consists of a foregut (including the crop and muscular gizzard), a tubular midgut (site of secretion of digestive enzymes and absorption of soluble nutrients), and a relatively large hindgut of about 1 microliter volume (Figure 23.31). In lower termites, the hindgut consists primarily of a single chamber, the paunch (Figure 23.31a). The hindgut of higher termites is more complex, being divided into several compartments (Figure 23.31b). For both higher and lower termites, the hindgut harbors a dense and diverse microbial community and is a major site of nutrient absorption. Acetate and other organic acids are produced during microbial fermentation of carbohydrate in the hindgut, and these products are primary carbon and energy sources for the termite (Figure 23.31c). High O2 consumption by bacteria near the gut wall keeps the interior of the hindgut anoxic. However, microsensor measurements (Section 19.9) have shown that O2 can penetrate up to 200 μm into the gut before it is completely removed by microbial respiration (Figure 23.31c). Thus, this tiny gut compartment offers distinct microbial niches with respect to O2 and can support diverse microbial activities.
Figure 23.31 Termite gut anatomy and function.

Gut architecture of lower (a) and higher (b) termites, showing the foregut, midgut, and differing complexity of the hindgut compartments. (c) Photo of workers, gut architecture, and biochemical activities of the lower termite Coptotermes formosanus. Microbial fermentation products, particularly acetate, are assimilated by the termite. Hydrogen produced by fermentation is consumed primarily by CO2-reducing acetogens, with a smaller amount going to hydrogenotrophic methanogens. Acetogenesis and methanogenesis are discussed in Sections 14.14 and 14.15 respectively. Syntrophic associations between protists and methanogens are discussed in Section 21.2 (Figure 21.8).
Bacterial Diversity and Lignocellulose Digestion in Higher Termites
The microbial gut communities differ significantly in termites of different genera. Analysis of 16S rRNA gene sequences from hindgut contents of two genera of wood-eating higher termites (Nasutitermes and Microcerotermes) and one genus of a wood-eating lower termite (Reticulitermes) revealed a high diversity of microbial species from 12 phyla of Bacteria, but few Archaea (Figure 23.32). Spirochetes of the genus Treponema (Section 15.17) dominated, with a lesser contribution from thus far uncultured organisms distantly related to the phylum Fibrobacteres (Section 16.22), a group also present in the rumen (Figure 23.47).
Figure 23.32 Microbial composition of termite hindgut inferred from 16S rRNA sequences.

The results are pooled analyses of 5075 sequences from amplified or metagenomic sequencing studies of three genera of wood-feeding higher termites, Nasutitermes, Reticulitermes, and Microcerotermes. The data provide a picture primarily of diversity, not relative abundance. Data assembled and analyzed by Nicolas Pinel.
Metagenomic analysis (Sections 10.7 and 19.8) of the Nasutitermes hindgut microbial community has revealed bacterial genes encoding glycosyl hydrolases that hydrolyze cellulose and hemicelluloses. These metagenomic data clearly implicate spirochetes and Fibrobacteres in the digestion of lignocellulose, although the corresponding cellulolytic bacteria have not yet been isolated from the higher termites. The abundance of Bacteroidetes varies greatly between different species of higher termites, depending on their dietary preferences. In particular, Bacteroidetes are enriched in higher termites that digest wood with the help of a lignin-degrading basidiomycete fungus (Termitomyces spp.). As do the leafcutter ants (see Section 23.8), these termites cultivate fungi that they feed lignocellulosic plant litter. The termites feed on both the fungus and the partially digested wood. The Bacteroidetes in these termites encode many glycosyl hydrolases that assist in the breakdown of plant and fungal cell wall material. In addition to the cellulolytic activities of Fibrobacteres and Spirochaetes in termites specializing in direct consumption of wood, other cellulolytic species include Ruminococcaceae, the latter comprising a significant fraction of Firmicutes diversity in the termite gut (Figure 23.32). At every molting of an individual termite, gut symbionts are lost, yet there is good conservation of the gut community within each termite species. Stable horizontal transmission of gut symbionts likely occurs as a result of the intimate social behavior and close contact characteristic of termites.
Acetogenesis and Nitrogen Fixation in the Termite Gut
Genes encoding enzymes of the acetyl-CoA pathway (Section 14.14) are highly represented in the spirochetes of the Nasutitermes hindgut, consistent with their function as the major CO2-reducing acetogens. The termite gut microbial communities have long been recognized as important to host nitrogen metabolism, providing new fixed nitrogen (Sections 3.12 and 15.9) and helping to conserve nitrogen by recycling excretory nitrogen back to the insect for biosynthesis. Consistent with this, metagenomic analyses have revealed that many bacteria, including Fibrobacteres and certain spirochetes, contain genes encoding nitrogenase, the enzyme required to fix N2.
From a simple energetic viewpoint, methanogenesis from H2 and CO2 is more favorable than acetogenesis from the same substrates (−34 kJ/mol of H2 versus −26 kJ/mol of H2, respectively), and thus methanogens should have a competitive advantage in all habitats in which the two processes compete (Sections 14.14–14.15). However, in termites they do not. There are at least two reasons for this. First, unlike methanogens, acetogens are able to use other substrates such as sugars or methyl groups from lignocellulose degradation as electron donors for energy metabolism. Second, termite acetogens (which seem to consist mostly of spirochetes) can for some reason better colonize the H2-rich termite gut center, whereas methanogens are largely restricted to the gut wall. On the gut wall, methanogens are located downstream of the H2 gradient and thus receive only a fraction of the H2 flux. In addition, the wall likely contains higher O2 tensions, which may negatively affect the physiology of methanogens. So, despite the fact that termites are methanogenic, producing up to 150 teragrams of CH4 per year on a global basis (1 teragram=1012 grams), carbon and electron flow in the termite gut favor acetogenesis in this interesting anoxic microbial habitat.
Check Your Understanding
How are anoxic conditions maintained in the termite hindgut?
Why does reductive acetogenesis predominate over methanogenesis in many termites?
Which group of morphologically unusual bacteria, absent from molecular surveys of prokaryotic cells in the rumen, seem to dominate activities in the termite hindgut?
IV Other Invertebrates as Microbial Habitats
Microbial symbionts of invertebrates perform different behavioral and nutritional functions. Some symbionts provide a light source to their host while others absorb light or consume inorganic chemicals to feed the animal. In some cases the invertebrate uses the symbiont as an offensive weapon, producing toxic compounds to kill its prey.
Thus far in this chapter we have discussed how certain macroorganisms that live in terrestrial environments provide habitats for microbial symbionts. We now consider some additional symbioses occurring in both terrestrial and aquatic environments. We begin in the marine environment as microbial symbioses with marine animals, especially with invertebrates, are common. By finding habitats in marine invertebrates, microorganisms establish a safe residence in a nutritionally rich environment.
23.10 Bioluminescent Symbionts and the Squid Symbiosis
23.10 Bioluminescent Symbionts and the Squid Symbiosis
23.10 Bioluminescent Symbionts and the Squid Symbiosis
Some bacteria can emit light, and this capacity has been exploited by several marine invertebrates. The light organ of the Hawaiian bobtail squid has become an important model system for such symbioses, especially for resolving the complex communication between microorganism and animal required for establishing a specific and stable symbiosis.
Key Bioluminescent Genera and Biochemistry of Bioluminescence
Several species of bacteria among the genera Vibrio, Aliivibrio, Photobacterium, Shewanella, and Photorhabdus (all Gammaproteobacteria, Section 16.4) can emit light, a process called bioluminescence (Figure 23.33). Most bioluminescent bacteria inhabit the marine environment, and some species colonize specialized light organs of certain marine fishes and squids, producing light that the animal uses for signaling, avoiding predators, and attracting prey (Figure 23.33c–f ). When living symbiotically in light organs of fish and squids, or saprophytically, for example on the skin of a dead fish, or parasitically in the body of a crustacean, luminous bacteria can be recognized by the light they produce.
Figure 23.33 Bioluminescent bacteria and their role as light organ symbionts in the flashlight fish.

(a) Two Petri plates of luminous bacteria photographed by their own light. Note the different colors. Left, Aliivibrio fischeri strain MJ-1, blue light, and right, strain Y-1, green light. (b) Colonies of Photobacterium phosphoreum photographed by their own light (Figure 1.3). (c) The flashlight fish Photoblepharon palpebratus; the bright area is the light organ containing bioluminescent bacteria. (d) Same fish photographed by its own light. (e) Underwater photograph taken at night of P. palpebratus. (f) Electron micrograph of a thin section through the light-emitting organ of P. palpebratus showing the dense array of bioluminescent bacteria (arrows).
Mechanism and Ecology of Bioluminescence
Although Photobacterium, Aliivibrio, and Vibrio isolates are facultative aerobes, they are bioluminescent only when O2 is present. Luminescence in bacteria requires the genes luxCDABE (Section 7.7) and is catalyzed by the enzyme luciferase, which uses O2, a long-chain aliphatic aldehyde (RCHO) such as tetradecanal, and reduced flavin mononucleotide (FMNH2) as substrates: FMNH2+O2+RCHO→LuciferaseFMN+RCOOH+H2O+light
The light-generating system constitutes a metabolic route for shunting electrons from FMNH2 to O2 directly, without employing other electron carriers such as quinones and cytochromes.
Luminescence in many luminous bacteria only occurs at high population density. The enzyme luciferase and other proteins of the bacterial luminescence system exhibit a population density–responsive induction, called autoinduction, in which transcription of the luxCDABE genes is controlled by a regulatory protein, LuxR, and an inducer molecule, acyl homoserine lactone (AHL, Section 7.7 and Figure 7.18). During growth, cells produce AHL, which can rapidly cross the cytoplasmic membrane in either direction, diffusing in and out of cells. Under conditions in which a high local population density of cells of a given species is attained, as in a test tube, a colony on a plate, or in the light organ of a fish or squid, AHL accumulates. Only when it reaches a certain concentration in the cell is AHL bound by LuxR, forming a complex that activates transcription of luxCDABE; cells then become luminous (Figure 23.33b, Figure 1.3).
This gene regulatory mechanism is called quorum sensing because of the population density–dependent nature of the phenomenon (Section 7.7). The strategy of quorum-sensing induction of luminescence ensures that luminescence develops only when population densities are high enough to allow the light produced to be visible to animals. The bacterial light can then attract animals to feed on the luminous material, thereby bringing the bacteria into the animal’s nutrient-rich gut for further growth. Alternatively, the luminous material may function as a light source in symbiotic light organ associations (Figure 23.33c, d). Quorum sensing is a form of regulation that has been detected in many different bacteria, controlling activities such as the production of extracellular enzymes and expression of virulence factors for which a high population density is beneficial if the bacteria are to have a biological effect.
The Squid–*Aliivibrio* System as a Model Symbiosis
The Hawaiian bobtail squid, Euprymna scolopes, is a small marine invertebrate (**Figure 23.34*a***) that harbors a large population of the bioluminescent bacterium Aliivibrio fischeri (Figure 23.33a) in a light organ located on its ventral side (Figure 23.34b and see Figure 23.35). Squid and bacterium are partners in a mutualism. The bacteria emit light that resembles moonlight penetrating marine waters, and this is thought to camouflage the squid from predators that strike from beneath. Several other species of Euprymna inhabit marine waters near Japan and Australia and in the Mediterranean, and these contain Aliivibrio symbionts as well.
Figure 23.34 Squid–*Aliivibrio* symbiosis.

(a) An adult Hawaiian bobtail squid, Euprymna scolopes, is about 4 cm long. (b) Thin-sectioned transmission electron micrograph through the E. scolopes light organ shows a dense population of bioluminescent Aliivibrio fischeri cells.
Many features of the E. scolopes–A. fischeri symbiosis have made it an important model for studies of animal–bacterial symbioses. These include the facts that the animals can be maintained in the laboratory and that only a single bacterial species is in the symbiosis in contrast to the large number of species in symbioses such as those of the termite (Figure 23.32) or the mammalian large intestine (Chapter 24). In addition, the symbiosis is not an essential one; both the squid and its bacterial partner can survive apart from each other in the laboratory. This allows juvenile squid to be grown without bacterial symbionts and then experimentally colonized. Experiments can be done to study specificity in the symbiosis, the number of bacterial cells needed to initiate an infection, the capacity of genetically defined mutants of A. fischeri to initiate infection of the squid, and many other aspects of the relationship. Moreover, because the genome of A. fischeri has been sequenced, microbial genomics may be employed to help interpret experimental results.
Establishing the Squid–*Aliivibrio* Symbiosis
Juvenile squid just hatched from eggs do not contain cells of A. fischeri. Thus, transmission of bacterial cells to juvenile squid is a horizontal (environmental) rather than a vertical (parent to offspring) event. Almost immediately after juveniles emerge from eggs, cells of A. fischeri in surrounding seawater begin to colonize them, entering through ciliated ducts that end in the immature light organ. Amazingly, the light organ becomes colonized specifically with A. fischeri and not with any of the many other species of gram-negative bacteria present in the seawater. Even if large numbers of other species of bioluminescent bacteria are offered to juvenile squid along with low numbers of A. fischeri, only A. fischeri establishes residence in the light organ. This implies that the animal recognizes and accepts A. fischeri cells and excludes those of other species.
Initial colonization of the squid light organ by Aliivibrio has three basic stages: (1) first contact, exclusion of unwanted microbes, and symbiont enrichment and aggregation, (2) chemotactic migration of the symbionts to the light organ colonization pore, and (3) migration of the symbionts into the crypts of the light organ interior, followed by loss of flagella and cell division (Figure 23.35). Contact of the squid with any bacterial cells triggers recognition in a very general way. Upon contact with peptidoglycan (a component of the cell wall of Bacteria, Section 2.3), the ciliated surface of the young squid’s immature light organ secretes an acidic mucus, trapping the bacteria on its developing light organ. The mucus is the first layer of specificity in the symbiosis, as it makes gram-negative but not gram-positive bacteria aggregate (Figure 23.35a).
Figure 23.35 Squid light organ symbiont colonization.

(a) Aliivibrio fischeri (green fluorescence) aggregates on the mucus-covered (blue) immature light organ surface of Euprymna scolopes (aggregates are approximately 100 μm long). (b, c) Chemotactic migration of A. fischeri cells (red fluorescence) along the ciliated surface (green) of the light organ, following a gradient of sugars released from the colonization pores. (d) A. fischeri cells migrate from the pore through the light organ along the direction of the white arrows to colonize the crypts (yellow arrows). Host nuclei are stained blue and A. fischeri cells fluoresce red or green (each cell aggregate is approximately 40 μm wide). (e) Following colonization of the light organs, the symbionts trigger the loss of the ciliated lobe-like appendages that are no longer needed for collection of A. fischeri cells from seawater. A. fischeri cells were genetically modified to express the green fluorescent protein or red fluorescent protein (Section 19.4).
Within the aggregates, A. fischeri outcompetes the other gram-negative bacteria to form a monoculture within 2 h of a juvenile’s hatching from an egg. Specific colonization is aided by the chemical environment generated by squid, including production of antimicrobial peptides and the gas nitric oxide (NO, see below), which helps the aggregating A. fischeri compete against attempted colonization by other gram-negative bacteria and facilitates their migration into the light organ (Figure 23.35b). The aggregated cells respond chemotactically to a gradient of specific sugars released from colonization pores (Figure 23.35c), migrating up this chemical gradient and into the pores. From there the cells continue their migration into the crypts of the light organ (Figure 23.35d). At this point, another critical selection process takes place. Only a single Aliivibrio cell enters and grows in each crypt of the light organ; when inside, it sheds its flagella and divides to form a dense population (Figures 23.35d and 23.34b). These initial colonization events trigger developmental events in the squid that lead to maturation of the light organ (Figure 23.35e). Each light organ in a mature E. scolopes contains between 108 and 109 A. fischeri cells.
Colonization by A. fischeri in the squid is assisted by nitric oxide (NO). Nitric oxide is a well-known defense response of animal cells to attack by bacterial pathogens; the gas is a strong oxidant and causes sufficient oxidative damage to kill bacterial cells ( Section 26.7). Nitric oxide produced by the squid is incorporated into the aggregates in the mucus and is present in the light organ itself. As A. fischeri colonizes the light organ, NO levels diminish rapidly. It appears that cells of A. fischeri can tolerate exposure to NO and actually consume it through the activity of NO-inactivating enzymes. The inability of other gram-negative bacteria in the aggregates to detoxify NO helps explain the sudden enrichment of A. fischeri in the ducts even before the actual colonization of the light organ. Then, after establishment, continued production of NO in the light organ prevents colonization by other bacterial species.
Propagating the Symbiosis
The squid matures into an adult in about 2 months and then lives a strictly nocturnal existence in which it feeds mostly on small crustaceans. During the day, the animal buries itself and remains quiescent in the sand. Each morning at dawn the squid nearly empties its light organ of A. fischeri cells and begins to grow a new population of the bacterium; by midafternoon, the structure contains the dense populations of A. fischeri cells required for the production of visible light. The actual emission of light requires a certain density of cells and is controlled by quorum sensing (Section 7.7). This daily growth and expulsion of bacterial cells is thought to be a mechanism for seeding the environment with cells of the bacterial symbionts; this increases the chances that the next generation of juvenile squid will be colonized and the symbiosis maintained.
A. fischeri clearly benefits from the squid symbiosis. The bacterium grows much faster in the light organ than in the open ocean, presumably because it is supplied with nutrients by the squid and is in a protected environment where it can avoid predation. A. fischeri is not a particularly abundant marine bacterium. Thus, the bacterium benefits by having an alternative habitat to seawater in which rapid growth and dense populations are possible. Daily expulsion of A. fischeri cells from the light organ increases the bacterium’s numbers in the microbial community. Hence, the symbiotic relationship of the bacterium with the squid helps maintain larger populations of A. fischeri than if the organism were strictly free-living. Because the competitive success of a microbial species is to some degree a function of population size (Section 20.1), this boost in cell numbers may confer an important ecological advantage on A. fischeri in its marine habitat.
Check Your Understanding
What substrates and enzyme are required for an organism such as Aliivibrio to emit visible light?
What is quorum sensing, and how does it control bioluminescence?
What features of the squid–Aliivibrio symbiosis make it an ideal model for studying animal–bacterial symbioses?
23.11 Marine Invertebrates at Hydrothermal Vents and Cold Seeps
23.11 Marine Invertebrates at Hydrothermal Vents and Cold Seeps
23.11 Marine Invertebrates at Hydrothermal Vents and Cold Seeps
Diverse invertebrate communities develop near undersea hot springs called hydrothermal vents. We covered the geochemistry and microbiology of hydrothermal vents and cold seeps of natural gas in Sections 20.16 and 21.1. Here we focus on hydrothermal vent invertebrates and their microbial symbionts.
Macroinvertebrates, including tube worms over 2 m in length and large clams and mussels, are present near these vents (Figure 23.36). Photosynthesis cannot support these invertebrate communities because they exist below the photic zone. However, hydrothermal fluids contain large amounts of reduced inorganic materials, including H2S, Mn2+, H2, and CO (carbon monoxide), and some vents contain high levels of ammonium (NH4 +) instead of H2S; all of these are good electron donors for chemolithotrophs—Bacteria and Archaea that use inorganic compounds as electron donors and fix CO2 as their carbon source (Chapter 14). Thus, hydrothermal vent invertebrates thrive in permanent darkness because they receive nourishment through a symbiotic association with these autotrophic bacteria.
Figure 23.36 Invertebrates living near deep-sea thermal vents.

(a) Riftia (tube worms, phylum Annelida), showing the sheath (white) and plume (red) of the worm bodies. (b) Mussel bed in vicinity of a warm vent. Note yellow deposition of elemental sulfur from the oxidation of H2S emitted from the vents.
Tube Worms, Mussels, and Giant Clams
Hydrothermal vent–associated animals either feed directly on cells of free-living chemolithotrophs or have formed tight symbiotic associations with them. Mutualistic chemolithotrophs are either tightly attached to the animal surface (that is, as epibionts, Section 23.2 and Figures 23.5 and 23.29b) or actually live within the animal tissues, supplying organic compounds to the animals in exchange for a safe residence and ready access to the electron donors needed for their energy metabolism. For example, the 2-m-long tube worms (Figure 23.36a) lack a mouth, gut, and anus, but contain an organ consisting primarily of spongy tissue called the trophosome. This structure, which constitutes half the worm’s weight, is filled with sulfur granules and large populations of spherical sulfur-oxidizing bacteria (Figure 23.37). Bacterial cells taken from trophosome tissue show activity of enzymes of the Calvin cycle, a major pathway for autotrophy (Section 3.12), but interestingly, they also contain enzymes of the reverse citric acid cycle, a second autotrophic pathway (Section 14.2). In addition, they contain the sulfur-oxidizing enzymes necessary to obtain energy from reduced sulfur compounds (Sections 14.7 and 15.12). The tube worms are thus nourished by organic compounds produced from CO2 and secreted by the sulfur chemolithotrophs.
Figure 23.37 Chemolithotrophic sulfur-oxidizing bacteria associated with the trophosome tissue of tube worms from hydrothermal vents.

(a) Scanning electron micrograph of trophosome tissue showing spherical chemolithotrophic sulfur-oxidizing bacteria. Cells are 3–5 μm in diameter. (b) Transmission electron micrograph of bacteria in sectioned trophosome tissue. The cells are frequently enclosed in pairs by an outer membrane of unknown origin. Reprinted with permission from Science 213: 340–342 (1981), © AAAS.
Along with tube worms, giant clams and mussels (Figure 23.36b) are also common near hydrothermal vents, and sulfur-oxidizing bacterial symbionts have been found in the gill tissues of these animals. Phylogenetic analyses have shown that each individual animal harbors one or more different strains of bacterial symbiont and that a variety of species of bacterial symbionts inhabit different species of vent animal. With the exception of the bacterial symbiont of Riftia (tube worms), which also has a free-living stage (Table 23.2), none of the bacterial symbionts of hydrothermal vent animals have yet been obtained in laboratory culture, even though they are fairly closely related to free-living sulfur chemolithotrophs (Sections 14.7 and 15.12).
The red plume of the tube worm (Figure 23.36a) is rich in blood vessels and is used to trap and transport inorganic substrates to the bacterial symbionts. Tube worms contain unusual hemoglobins that bind H2S and O2; these are then transported to the trophosome where they are released to the bacterial symbionts. The CO2 content of tube-worm blood is also high, about 25 mM, and presumably this is released in the trophosome as a carbon source for the symbionts. In addition, stable isotope analyses (Section 19.10) of elemental sulfur from the trophosome have shown that its 34S/32S composition is the same as that of the sulfide emitted from the vent. This ratio is distinct from that of seawater sulfate and is solid evidence that geothermal sulfide is actually entering the worm in large amounts.
Other Invertebrate Symbionts
Other marine invertebrates have coevolved bacterial symbioses that supply their nutrition as well (Table 23.3). For example, methanotrophic (CH4-consuming) symbionts are present in giant clams that live near cold seeps of natural gas at relatively shallow depths in the Gulf of Mexico. Although not autotrophs (CH4 is an organic compound), the methanotrophs do provide nutrition to the clams; the methanotrophs use CH4 as their electron donor and carbon source and secrete organic carbon to the clams.
Table 23.3 Marine animals with chemolithotrophic or methanotrophic endosymbiotic *Bacteria*

aWhale and wood falls are sunken whale carcasses and wood, respectively.
Molecular hydrogen (H2) is used as an electron donor by the mussel Bathymodiolus puteoserpentis, the most abundant macrofauna in vent fields associated with the peridotite-hosted vent systems of the Mid-Atlantic Ridge (Section 20.16 and Figure 20.45). These systems release high levels of H2 and CH4, with H2 concentrations as high as 19 mM. This mussel was previously shown to live in a dual symbiosis with methane-oxidizing bacteria and chemolithotrophic sulfur-oxidizing bacteria localized to the gill tissue. Remarkably, the sulfur-oxidizing symbiont of B. puteoserpentis also has the capacity to use H2 as an electron donor, making this mussel one of the most metabolically versatile of vent macrofauna.
Genomics and Hydrothermal Vent Symbioses
Genome sequencing is revealing additional features of the metabolic interaction and coevolution of marine invertebrates and their bacterial symbionts. The genome sequence of the gill endosymbiont of the giant vent clam Calyptogena magnifica provides direct evidence for autotrophy by way of the Calvin cycle because the genome encodes the key enzymes of the cycle, ribulose bisphosphate carboxylase and phosphoribulokinase (Section 3.12). This genome also contains genes encoding key sulfur-oxidation processes and genes encoding the biosynthesis of most vitamins and cofactors and all 20 amino acids needed to support the host. However, because few substrate-specific transporters are encoded by the symbiont genome—proteins necessary to secrete organic compounds to the host—it is likely that the clam actually digests symbiont cells for nutrition, as do mussels.
Like the obligate symbionts of insects, most symbionts of marine invertebrates have small genomes (Table 23.2), indicating reduced function and an obligate association with their host. The bacterial symbiont of the giant tube worm Riftia pachyptila is an exception, having a genome larger than some free-living sulfur-oxidizing chemolithotrophs (Table 23.2). The R. pachyptila symbiont is acquired by uninfected juvenile animals from the environment (horizontal transmission), and its larger genome is likely important for survival in the free-living state. For more on the tube worm–microbe symbiosis, see Explore the Microbial World “Pattern Recognition Receptors of Hydrothermal Vent Tube Worms Facilitate Endosymbiosis” in Chapter 26.
Check Your Understanding
What are the similarities of the obligate symbioses of insects and hydrothermal vent invertebrates?
Why might some marine invertebrates contain large genomes and others small genomes?
23.12 Entomopathogenic Nematodes
Two families of nematodes—the Heterorhabditidae and Steinernematidae—are obligate pathogens of insects and together constitute a group of entomopathogenic (insect-killing) invertebrates that have a wide range of insect hosts. The basis for their insect lethality is a specific association between the nematode and its bacterial symbionts, which produce a variety of insecticidal compounds. However, unlike the insects making defensive use of symbiont products that we explored in Section 23.8, the nematode uses the compounds produced by its bacterial symbiont to kill insect prey.
Specificity of Entomopathogenic Nematodes for Their Symbionts and Insect Hosts
Species of the gram-negative bacteria Photorhabdus and Xenorhabdus are the primary bacterial symbionts of the entomopathogenic nematodes. Comparative 16S rRNA sequence analysis (Section 13.11) has shown a high specificity of association between bacterial and nematode species. Species of Photorhabdus specifically associate with Heterorhabditidae, whereas Xenorhabdus species selectively associate with Steinernematidae. Each Steinernema species is thought to associate specifically with only one Xenorhabdus species. However, a single Xenorhabdus species may be associated with more than one nematode species. In turn, the nematodes comprising these two families can be separated into several phylogenetic groups that correspond to the phylogenetic relationships of their insect hosts. Together these data point to a history of coevolution between nematodes and their bacterial symbionts that has resulted in the various entomopathogenic nematodes having different insect host specificities.
The Nematode Life Cycle and Lethality
All entomopathogenic nematodes have a similar life cycle in which only one stage, the nonfeeding third-stage infective juvenile, survives outside the host (Figure 23.38). The long association between nematode and bacterial symbiont has also resulted in modification of the anterior part of the infective juvenile nematode’s intestine. This region forms a discrete chamber in the infective juvenile called the bacterial receptacle (Figure 23.38b, c). This vessel becomes filled with bacteria by growth of the symbiont during maturation of the infective juvenile, protecting the symbiont for the next infective cycle.
Figure 23.38 Nematode–*Xenorhabdus* life cycle.

(a) In the presence of nutrients released in the infected insect, the nematodes develop through four juvenile stages (J1–J4), molting between stages, to egg-laying adults. When nutrients become limiting, the nematodes develop into an alternative nonfeeding J3 stage (known as the infective juvenile) that closes off most of its intestine and forms a receptacle in the gut where symbiotic bacteria localize. The infective J3 stage nematodes are then released into the environment to infect other insects. (b) Xenorhabdus localized to the bacterial receptacle of the infective juvenile. (c) Bacteria-filled receptacle imaged by differential interference contrast microscopy. Xenorhabdus colonizing the nematodes shown in a and b were genetically engineered to express the green fluorescent protein (Sections 8.1 and 19.4) for imaging by fluorescence microscopy. Nematodes are approximately 50 μm in diameter.
When the infective juvenile encounters an appropriate insect host, it invades the insect’s hemocoel (the body cavity containing the bloodlike hemolymph of the insect’s circulatory system) through a natural opening such as the mouth or anus. The bacteria are then released from the bacterial receptacle into the hemolymph and grow rapidly. They are able to thwart the insect’s natural immune system in part by producing a variety of hemolysins, toxins, and digestive enzymes (e.g., proteases, lipases, chitinases) that promote the release of nutrients from host tissues. The bacterial symbionts also produce antibiotics to inhibit competitive colonization by other microbes. Both the multiplying bacteria and digested host tissues nourish the multiplying nematodes, and this results in a slow insect death, taking anywhere from 1 to 22 days. As the nutrients in the host become depleted, the adult nematodes produce new infective nonfeeding juveniles that are adapted to withstand the outside environment (Figure 23.38a), and the life cycle repeats.
Practical Applications of Entomopathogenic Nematodes
Entomopathogenic nematodes have been extensively studied as biocontrol agents, providing an alternative to broad-spectrum chemical pesticides in the control of insect pests. In addition to alleviating possible human health effects of chemical pesticides, biologically based insect control strategies can be highly specific, targeting the pest species but not other insects. This specificity helps maintain the natural insect biodiversity in environments that require pest species management, most commonly in agricultural settings. Apart from their application in pest management, this symbiosis has also provided a powerful model system for developing a detailed understanding of the evolution, physiology, and genetics of beneficial host–microbe interactions.
Check Your Understanding
What evidence suggest that the nematodes and their bacterial symbionts have coevolved?
What prevents other bacteria from colonizing the dead insect and competing with the nematode and Xenorhabdus for nutrients?
23.13 Reef-Building Corals
Coral reef ecosystems are the products of mutualistic associations between microscopic phototrophs and simple marine animals. The extensive ecosystems associated with the worldwide distribution of these mutualisms support thousands of species of fish and marine invertebrates.
Phototrophic Symbioses with Animals
Some animals establish mutualistic associations with algae or cyanobacteria (Table 23.4). The animals in most of these associations are in phyla that display very simple body plans, for example, the Porifera (sponges) and Cnidaria (corals, sea anemones, and hydroids). These mutualistic animal–bacterial associations live in clear tropical waters where nutrients for the animals are scarce, and the animal body typically has a large surface area relative to its volume and is thus well suited for capturing light.
Table 23.4 Symbioses between animals and phototrophic symbionts

aCyanobacteria are Bacteria; all others are eukaryotic phototrophs.
There are only a few instances of algae forming associations with more complex animals, such as those in the phyla Platyhelminthes (flatworms), Mollusca (snails and clams), and Urochordata (sea squirts). In these cases, either the animal has a suitable surface-to-volume ratio or has evolved specific light-gathering surfaces. The unicellular phototrophic symbionts are phylogenetically diverse and include cyanobacteria (Section 15.3), and red and green algae, diatoms, and dinoflagellates (Chapter 18). The most common symbionts are green algae (Chlorella, associating with sponges and freshwater hydras), cyanobacteria (associating with marine sponges), and species of the dinoflagellate genus Symbiodinium (Section 18.4; Figure 23.39). We focus here on the symbiotic association between the dinoflagellate Symbiodinium and the stony coral cnidarians (see also MicrobiologyNow on page 744).
Figure 23.39 *Symbiodinium* symbiont of marine invertebrates.

(a) Thin-section micrograph of Symbiodinium in the mantle tissue of a giant clam. (b) Symbiodinium cells recovered from a soft coral. (c) Transmission electron micrograph of a Symbiodinium cell within a vacuole of a cell of the stony coral Ctenactis echinata. The Symbiodinium cell is about 10 μm in diameter.
The Staghorn Coral Symbiosis
The staghorn coral is representative of the mutualism between the stony corals (phylum Cnidaria) and Symbiodinium. Stony corals are among the most spectacular and ecologically significant animal–phototroph associations. Each coral is a colony of genetically identical polyps (Figure 23.40) that together build the calcium carbonate structure. The Symbiodinium symbiont is distributed throughout the coral skeleton in the coral tissue (Figure 23.40c). Together the corals and dinoflagellates form the trophic and structural foundation of the coral reef ecosystem.
Figure 23.40 Staghorn coral.

(a) Photograph of a large, branched colony of reef-building staghorn coral (11 cm at its widest). (b) Close-up of a coral branch. Each polyp (arrows) is around half a millimeter in size. The brown bands are caused by the photosynthetic symbionts living within the coral tissue. (c) Fluorescence microscopy of a single polyp on a branch of the colony reveals symbionts (red autofluorescence from chlorophyll) living inside the polyp’s tentacles, and in bands along its sides.
The stony corals possess a simple two-tissue-layer body plan (ectoderm and gastroderm) and harbor their dinoflagellate symbiont intracellularly in membrane-bound vesicles called symbiosomes within cells of the inner (gastrodermal) tissue layer (Figure 23.39c). The coral symbiosomes are analogous to the bacteroid-filled vesicles that develop in plant cells of the legume root nodules (Section 23.4). The coral skeleton is an extremely efficient light-gathering structure that greatly enhances light harvesting by Symbiodinium cells to maintain the symbiosis.
Transmission, Specificity, and Benefits of the *Symbiodinium*–Coral Association
Reef-building corals reproduce sexually by releasing gametes into the seawater. A male and a female gamete fuse to form a free-swimming larva that later settles on a surface, where it may initiate a new coral colony. Algal symbionts can be present in the egg before it is released from the parent (vertical transmission), or acquired from the environment by juvenile corals (horizontal transmission). A developing coral that ingests dinoflagellates digests all of them except the particular strain of Symbiodinium of its mutualism. After establishing an association, the coral controls the growth of Symbiodinium via chemical signaling and, following each cell division, each Symbiodinium daughter cell is allocated to a new symbiosome.
Both partners in the stony coral–dinoflagellate mutualism have evolved adaptations for nutritional exchange. The dinoflagellates donate most of their photosynthetically fixed carbon (in the form of small molecules such as sugars, glycerol, and amino acids) to the cnidarian in exchange for inorganic nitrogen, phosphorus, and inorganic carbon from the host. Moreover, in addition to providing protection and inorganic nutrients, the calcium carbonate skeleton of corals is one of the most efficient collectors of solar radiation in nature, amplifying the incident light field for the symbionts by as much as fivefold; this benefits the symbiont in carrying out photosynthesis under a light-absorbing water column. This mutualism between animal and phototroph has allowed coral reefs to thrive in large expanses of nutrient-poor ocean waters.
Coral Bleaching—The Risk of Harboring a Phototrophic Symbiont in a Changing World
Many of the extensive coral reef systems in the oceans are threatened with extinction, primarily as a consequence of human activities. Ongoing loss of these beautiful and productive ecosystems is thought to be the result of elevated atmospheric CO2; namely, increased sea surface temperature, rising sea levels, and ocean acidification (Sections 21.7 and 21.9). Coastal development also threatens reef systems, contributing to pollution from sewage discharge, eutrophication from nutrient runoff, and overfishing. These environmental changes are contributing to high mortality through disease, loss of coral structure from reduced calcification caused by acidification, and bleaching. Healthy corals harbor millions of cells of Symbiodinium per square centimeter of tissue. Coral bleaching is the loss of color from host tissues caused by the lysis or expulsion of these pigmented symbionts, revealing the underlying white limestone skeleton (Figure 23.41).
Figure 23.41 Coral bleaching.

(a) Two colonies of the brain coral Colpophyllia natans. The coral on the left is a healthy brown color, whereas the coral on the right is fully bleached. (b) A large colony of partially bleached mountainous star coral (Orbicella faveolata).
Coral reefs live close to their optimum temperature, and it is the synergistic effect of increased sea surface temperature and irradiance that causes massive bleaching. Elevated temperature and high irradiance impair the photosynthetic apparatus of the dinoflagellates, resulting in the production of reactive oxygen species (for example, singlet oxygen and superoxide, Section 4.16) that cause damage to both host and symbiont. Bleaching is thought to be caused by a protective immune response of the host that destroys compromised symbionts. Increases in sea surface temperatures as small as 0.5–1.5 °C above the local maximum, if sustained for as little as several weeks, can induce rapid coral bleaching. Such thermal stress has resulted in bleaching of huge expanses of coral reefs, providing additional and highly visible evidence that climate change is upon us and that microbially based ecosystems are responding.
Although coral reefs are clearly threatened, there is some uncertainty in projecting their future because of a lack in basic knowledge of the heat stress capacity of individual coral–symbiont mutualisms (Figure 23.42). For example, thermal tolerance is in part conferred by the species or strain of Symbiodinium, and following a bleaching event, the mutualism can shift to a more thermally tolerant symbiont (Figure 23.42). Molecular studies have indicated that there are over 150 different Symbiodinium phylotypes, each possibly representing a distinct species with different stress tolerance. When a coral is bleached, a more heat-tolerant symbiont can replace the bleached species, likely by symbiont swapping. In this process, the species shift results from the more robust growth of a more heat-tolerant variant already associated with the coral but in very low numbers; the variant then thrives following loss of the original symbiont and the bleaching event. Because the type of symbiont influences the ability of the coral to adapt to stresses associated with climate change, a more complete understanding of this adaptive response to thermal stress, and the limits of the response, is essential for predicting the future health of corals, their symbionts, and the spectacular and ecologically important reefs that they build.
Figure 23.42 Differential stress tolerance of a coral species associated with different *Symbiodinium* phylotypes.

Pocillopora corals symbiotically associated with Symbiodinium type C1b-c were much more sensitive to a thermal stress event than the same coral species associated with Symbiodinium type D1. The more tolerant Symbiodinium type D1–Pocillopora association suffered low mortality. The response also suggested additional genetic variation within each Symbiodinium type, since the two mutualisms displayed a range of sensitivity to increasing thermal stress.
In the final part of this chapter, we explore the symbionts of the mammalian gut of herbivorous animals as a prelude to our coverage of the human microbiome in Chapter 24.
Check Your Understanding
Where do Symbiodinium cells localize within the coral skeleton and within individual coral cells?
Describe two functions of the calcium carbonate skeleton of the stony corals.
Besides thermal stress, list some environmental factors contributing to coral bleaching.
V Mammalian Gut Systems as Microbial Habitats
The guts of herbivorous animals are modified in such a way as to support the growth of dense populations of anaerobic microorganisms that specialize in the digestion of plant fibers. Consumption of the products of fiber digestion and the microbes themselves allows the animal to live on a plant diet rich in carbohydrates but poor in protein.
The evolution of animals has been shaped in part by a long history of symbiotic associations with microorganisms and includes vertebrates as well as the invertebrate systems we just explored. We end this chapter by considering microbial symbioses with herbivorous animals. Microbes inhabit all sites on mammalian bodies, but the greatest diversity and density of microbes are found in the mammalian gut, and so we center our discussion there.
23.14 Alternative Mammalian Gut Systems
Some mammals are herbivores, consuming only plant materials, whereas others are carnivores, eating primarily the flesh of other animals. Omnivores eat both plants and animals. As Figure 23.43 indicates, closely related mammals have evolved adaptations for differing diets. Notice that mammals of different lineages independently evolved the herbivorous lifestyle, mostly during the Jurassic period, an era in Earth’s history of roughly 60 million years beginning about 200 million years ago.
Figure 23.43 Phylogenetic tree showing multiple origins of herbivory among mammals.

Some of the herbivores listed are foregut fermenters, while others are hindgut fermenters (see Figure 23.44). Instead of animal flesh, some mammalian carnivores eat only insects (the insectivores, such as bats), or fish (the piscivores, such as the river otter).
The massive evolutionary radiation of mammals during the Jurassic led to the evolution of several feeding strategies. Most mammalian species evolved gut structures that foster mutualistic associations with microorganisms. As anatomical differences evolved, microbial fermentation remained important or essential in mammalian digestion. Monogastric mammals, such as humans, have a single compartment, the stomach, positioned before the intestine. Such animals may get a substantial part of their energy requirement from microbial fermentation of otherwise indigestible foods in the lower intestine, while herbivores are totally dependent on such fermentations.
Plant Substrates
Microbial associations with various herbivorous animals led to the capacity to catabolize plant fiber, the structural component of plant cell walls. Fiber is composed primarily of insoluble polysaccharides, of which cellulose is the most abundant component. Mammals—and indeed virtually all animals—lack the enzymes necessary to digest cellulose and certain other plant polysaccharides. However, many microbes have genes encoding the hydrolases and polysaccharide lyases required to decompose these polysaccharides.
As the most abundant organic compound on Earth and one composed exclusively of glucose, cellulose offers a rich source of carbon and energy for animals that can digest it. The two primary traits that evolved to support herbivory are (1) an enlarged anoxic fermentation chamber for holding ingested plant material and (2) an extended retention time—the time that ingested material remains in the gut. A longer retention time allows for a longer association of microorganisms with the ingested material and thus a more complete degradation of the plant polymers.
Digestion Systems of Herbivores
Two digestive patterns have evolved in herbivorous mammals, foregut and hindgut. In herbivores with a foregut fermentation, the microbial fermentation chamber precedes the small intestine. This gut architecture originated independently in ruminants—such as cattle and sheep—colobine monkeys, sloths, and macropod marsupials (Figure 23.44). These all share the common feature that ingested nutrients are degraded by the gut microbiota before reaching the acidic stomach and small intestine. We examine the digestive processes of ruminants, as examples of foregut fermenters, in the next section.
Figure 23.44 Variations on vertebrate gut architecture.

All vertebrates have a small intestine but vary in other gut structures. Most host absorption of dietary nutrients occurs in the small intestine, whereas microbial fermentation can occur in the forestomach, cecum, or large intestine (colon). Foregut fermentation is found in four major clades of mammals and one avian species (the hoatzin). Hindgut fermentation, either in the cecum or large intestine/colon, is common to many clades of mammals (including humans), birds, and reptiles. Compare with Figure 23.43.
Horses and rabbits are herbivorous mammals, but they are not foregut fermenters. Instead, these animals are hindgut fermenters. They have only one stomach but use an organ called the cecum, a digestive organ located between the small and large intestines, as their fermentation vessel (Figure 23.44). The cecum contains fiber- and cellulose-digesting (cellulolytic) microorganisms. Mammals, such as the rabbit, that rely primarily on microbial breakdown of plant fiber in the cecum are called cecal fermenters. In other hindgut fermenters, both the cecum and colon are major sites of fiber breakdown by microorganisms.
Anatomical differences among monogastric mammals, foregut fermenters, and hindgut fermenters are summarized in Figure 23.44. Nutritionally, foregut fermenters have an advantage over hindgut fermenters in that the cellulolytic microbial community of the foregut eventually passes through an acidic stomach. As this occurs, most microbial cells are killed by the acidity and become a protein source for the animal. By contrast, in animals such as horses and rabbits, the remains of the cellulolytic community pass out of the animal in the feces because of its position posterior to the acidic stomach. For this reason, many hindgut fermenters, for example, rabbits, commonly eat their own feces (a phenomenon called coprophagy) in order to recover protein from the microbial cells.
Check Your Understanding
How do animals with foregut and hindgut fermentation differ in recovery of nutrients from plants?
How does retention time affect microbial digestion of food in a gut compartment?
23.15 The Rumen and Rumen Activities
Earth’s dominant herbivores are the ruminants (Figure 23.44), mammals that possess a special foregut digestive organ, the rumen, within which cellulose and other plant polysaccharides are digested by microorganisms. Some of the most important domesticated animals—cows, sheep, and goats—are ruminants. Camels, buffalo, deer, reindeer, caribou, and elk are also ruminants. Because the human food economy depends to a great extent on ruminant animals, rumen microbiology is of considerable economic significance and importance.
Rumen Anatomy
Unique features of the rumen as a site of cellulose digestion are its relatively large size (capable of holding 100–150 liters in a cow, 6 liters in a sheep) and its position in the gastrointestinal system before the acidic stomach. The rumen’s warm and constant temperature (39 °C), narrow pH range (5.5–7, depending on when the animal was last fed), and anoxic environment are also important factors in overall rumen function. **Figure 23.45*a*** shows the relationship of the rumen to other parts of the ruminant digestive system. The digestive processes and microbiology of the rumen have been well studied, in part because it is possible to remove samples for analysis by way of a sampling port, called a fistula, implanted into the rumen of a cow (Figure 23.45b) or a sheep.
Figure 23.45 The rumen.

(a) Schematic diagram of the rumen and gastrointestinal system of a cow. Food travels from the esophagus into the reticulo-rumen, consisting of the reticulum and rumen. The reticulum collects smaller digesta particles and moves them into the omasum. Larger particles remain in the rumen and are regurgitated as cud (partially digested fiber). Cud is chewed until food particles are small enough to pass from the reticulum into the omasum, abomasum, and intestines, in that order. The abomasum is an acidic vessel, analogous to the stomach of monogastric animals like pigs and humans. (b) Photo of a fistulated Holstein cow. The fistula, shown unplugged, is a sampling port that allows access to the rumen.
After a cow swallows its food, the food enters the first chamber of the four-compartment stomach, the reticulum. Partially digested plant materials flow freely between the rumen and reticulum, sometimes referred to together as the reticulo-rumen. The main function of the reticulum is to collect smaller food particles and move them to the omasum. Larger food particles (called cud) are regurgitated, chewed, mixed with saliva containing bicarbonate, and returned to the reticulo-rumen where they are digested by rumen bacteria. Solids may remain in the rumen for more than a day during digestion. Eventually, small and more thoroughly digested food particles are passed to the omasum and from there to the abomasum, an organ similar to a true, acidic stomach. In the abomasum, chemical digestive processes begin that continue in the small and large intestine.
Microbial Fermentation in the Rumen
Food remains in the rumen for 20–50 h depending on the feeding schedule and other factors. During this relatively long retention time, cellulolytic microorganisms hydrolyze cellulose, which frees glucose. The glucose then undergoes bacterial fermentation with the production of volatile fatty acids (VFAs), primarily acetic, propionic, and butyric acids, and the gases carbon dioxide (CO2) and methane (CH4) (Figure 23.46). The VFAs pass through the rumen wall into the bloodstream and are oxidized by the animal as its main source of energy. The gaseous fermentation products CO2 and CH4 are released by eructation (belching).
Figure 23.46 Metabolic sequences and biochemical reactions in the rumen.

The major pathways are solid lines; dashed lines indicate minor pathways. Approximate steady-state rumen levels of volatile fatty acids (VFAs) are acetate, 60 mM; propionate, 20 mM; butyrate, 10 mM.
The rumen contains enormous numbers of bacteria (1010–1011 cells/g of rumen contents). Most of the bacteria adhere tightly to food particles. These particles proceed through the gastrointestinal tract of the animal where they undergo further digestive processes similar to those of nonruminant animals. Bacterial cells that digested plant fiber in the rumen are themselves digested in the acidic abomasum. Because bacteria living in the rumen biosynthesize amino acids and vitamins, the digested bacterial cells are a major source of protein and vitamins for the animal.
Check Your Understanding
What physical and chemical conditions prevail in the rumen?
What are VFAs, and of what value are they to the ruminant?
23.16 Rumen Microbes and Their Dynamic Relationships
23.16 Rumen Microbes and Their Dynamic Relationships
23.16 Rumen Microbes and Their Dynamic Relationships
Enormous numbers of prokaryotic and eukaryotic cells are present in the rumen, even greater than those present in fertile soils (Section 20.7). These include Bacteria and Archaea as well as various fungi and nonphototrophic protists. Together these microbes constitute a highly integrated and efficient microbial community whose dynamics vary according to the types and amounts of fermentable material consumed.
Rumen *Bacteria* and *Archaea*
Anaerobic Bacteria and Archaea dominate in the rumen because it is a strictly anoxic compartment, and some anaerobic microbial eukaryotes are also present. Cellulose is converted to fatty acids, CO2, and CH4 (Figure 23.46) in a multistep microbial food chain, with several different anaerobes participating in the process. Analysis of 16S rRNA gene sequences from rumen samples indicate that the typical rumen contains 300–400 bacterial species (Section 13.12; Figure 23.47), more than ten times higher than culture-dependent diversity estimates. Molecular surveys also show that species of Firmicutes and Bacteroidetes dominate bacterial diversity in the rumen, while methanogens make up virtually the entire archaeal population (Figure 23.47).
Figure 23.47 Ruminal microbial community inferred from 16S rRNA gene sequences.

Pooled analyses of 14,817 sequences from several studies of ruminant animals, including cow, sheep, goat, and deer. The data reflect prokaryotic diversity and not necessarily relative abundance. Data assembled and analyzed by Nicolas Pinel.
Several rumen anaerobes have been cultured and their physiology characterized (Table 23.5). Various rumen bacteria hydrolyze cellulose to sugars and ferment the sugars to VFAs. Fibrobacter succinogenes and Ruminococcus albus are the two most abundant cellulolytic rumen anaerobes. Although both organisms produce cellulases, Fibrobacter, a gram-negative bacterium, produces enzymes localized to its outer membrane, whereas Ruminococcus, a gram-positive bacterium (and therefore lacking an outer membrane), produces a cellulose-degrading protein complex bound to its cell wall. However, in both cases, cells of Fibrobacter and Ruminococcus need to physically bind to cellulose particles in order to degrade them. The gram-positive bacterium Butyrivibrio also participates in the digestion of plant cell wall polymers, specializing in the digestion of hemicellulose components such as xylan.
Table 23.5 Characteristics of some rumen *Bacteria* and *Archaea*

aExcept for the methanogens, which are Archaea, all organisms listed are Bacteria.
bThese species also degrade xylan, a major plant cell wall polysaccharide.
dAlso ferments amino acids, producing NH3. Several other rumen bacteria ferment amino acids as well, including Peptostreptococcus anaerobius and Clostridium sticklandii.
Rumen Protists and Fungi
In addition to huge populations of Bacteria and Archaea, the rumen has characteristic populations of ciliated protists (Chapter 18) present at a density of about 106 cells/ml. Many of these protists are obligate anaerobes, a physiology that is rare among eukaryotes. Although these protists are not essential for rumen fermentation, they contribute to the overall process. In fact, some protists are able to hydrolyze cellulose and starch and ferment glucose with the production of the same VFAs formed by cellulose-fermenting bacteria (Figure 23.46 and Table 23.5). Rumen protists also consume rumen bacteria and smaller rumen protists and are likely to play a role in controlling bacterial densities in the rumen. An interesting commensal interaction has been observed between rumen protists that produce VFAs and H2 as products and methanogenic bacteria that consume the H2, producing CH4. Because their cells autofluoresce (Section 14.15 and Figure 14.37), methanogens are easily observed in rumen fluid bound to the surface of H2-producing protists. A similar situation exists in the hindgut of termites, but in this case, methanogens actually live inside anaerobic cellulolytic protozoa (Figure 21.8c, d).
Anaerobic fungi also inhabit the rumen and play a role in its digestive processes. Rumen fungi are typically species that alternate between a flagellated and a thallus form, and studies with pure cultures have shown that they can ferment cellulose to VFAs. Neocallimastix, for example, is an obligately anaerobic fungus that ferments glucose to formate, acetate, lactate, ethanol, CO2, and H2. Although a eukaryote, this fungus lacks mitochondria and cytochromes and thus lives an obligately fermentative existence. However, Neocallimastix cells contain a redox organelle called the hydrogenosome; this mitochondrial analog evolves H2 and has otherwise been found only in certain anaerobic protists (Section 18.3).
Rumen fungi play an important role in the degradation of polysaccharides other than cellulose, including a partial solubilization of lignin (the strengthening agent in the cell walls of woody plants), hemicellulose (a derivative of cellulose that contains pentoses and other sugars), and pectin.
Dynamics of the Rumen Microbial Community
If a ruminant is gradually switched from cellulose to a diet high in starch (grain, for instance), the starch-digesting Gammaproteobacteria Ruminobacter amylophilus and Succinomonas amylolytica grow to high numbers in the rumen. On a low-starch diet these organisms are typically minor constituents. If an animal is fed legume hay, which is high in pectin, a complex polysaccharide containing both hexose and pentose sugars, then the pectin-digesting gram-positive bacterium Lachnospira multipara (Table 23.5) becomes an abundant member of the rumen microbial community. Some of the fermentation products of these rumen bacteria are used as energy sources by secondary fermenters in the rumen. For example, succinate is fermented to propionate plus CO2 (Figure 23.46) by the bacterium Schwartzia, and lactate is fermented to acetate and other fatty acids by the gram-positive bacteria Selenomonas and Megasphaera (Table 23.5). Hydrogen (H2) produced in the rumen by fermentative processes never accumulates because it is quickly consumed by methanogens for the reduction of CO2 to CH4. This H2 removal facilitates greater fermentative activity because H2 accumulation negatively affects the energetics of fermentative reactions that produce H2 (Section 14.22). Although acetate, a methanogenic substrate, is produced in large amounts in the rumen, it is absorbed by the ruminant (Figure 23.46) and thus unavailable for conversion to methane.
Dangerous and Protective Changes in the Rumen Microbial Community
Significant changes in the microbial composition of the rumen can cause illness or even death of the animal. For example, if a cow is changed abruptly from forage to a grain diet, the gram-positive bacterium Streptococcus bovis grows rapidly in the rumen. The normal level of S. bovis, about 107 cells/g, is an insignificant fraction of total rumen bacterial numbers. But if large amounts of grain are fed abruptly, numbers of S. bovis can quickly rise to over 1010 cells/g and dominate the rumen microbial community. This occurs because grasses contain mainly cellulose, which does not support growth of S. bovis, while grain contains high levels of starch, on which S. bovis grows rapidly.
Because S. bovis is a lactic acid bacterium (Sections 14.18 and 16.6), large populations are capable of producing large amounts of its fermentation product, lactic acid. Lactic acid is a much stronger acid than the VFAs produced during normal rumen function (Figure 23.46). Lactate production thus acidifies the rumen below its lower functional limit of about pH 5.5, thereby disrupting the activities of normal rumen bacteria. Rumen acidification, a condition called acidosis, causes inflammation of the rumen epithelium, and severe acidosis can cause hemorrhaging in the rumen, acidification of the blood, and death of the animal. Despite this dangerous condition typically associated with abrupt feeding changes, ruminants such as cattle can be fed a diet exclusively of grain. However, to avoid acidosis, the animals must be switched from forage to grain gradually over a period of many days. The slow introduction of starch selects for VFA-producing starch-degrading bacteria (Table 23.5) instead of S. bovis. Under these conditions, normal rumen functions continue, and the animal remains healthy.
The overgrowth of S. bovis is a potent example of how a single microbial species can have a deleterious effect on animal health. In contrast, there is at least one well-studied example of how a single bacterial species can enhance the health of ruminant animals. This occurs in animals fed the tropical legume Leucaena leucocephala. This plant has very high nutritional value but contains an amino acid–like compound called mimosine that is converted to the toxic compounds 3-hydroxy-4(1H)-pyridone (3,4-DHP) and 2,3-dihydroxypyridine (2,3-DHP) by rumen microorganisms (Figure 23.48). The observation that ruminants in Hawaii, but not Australia, could feed on Leucaena without toxic effect led investigators to hypothesize that further metabolism of DHP by bacteria present in Hawaiian ruminants alleviated DHP toxicity. This was subsequently confirmed by the isolation of the bacterium Synergistes jonesii, an anaerobe related to Deferribacter (Section 16.22) but not closely related to any other rumen bacteria. Inoculation of Australian ruminants with cells of S. jonesii conferred resistance to mimosine by-products, allowing the animals to feed on Leucaena without ill effect.
Figure 23.48 Conversion of mimosine to toxic pyridine and pyridone metabolites by ruminal microorganisms.

Mimosine is converted to toxic 3,4-DHP by normal ruminal microbiota. Synergistes jonesii converts 3,4-DHP to nontoxic metabolites through a 2,3-DHP intermediate, preventing buildup of toxic metabolites of mimosine.
The success of this single-organism modification of the rumen microbial community has encouraged further studies of this sort, including genetic engineering of bacteria to improve their ability to utilize available nutrients or to detoxify toxic substances. A notable success has been inoculation of the rumen of sheep with genetically engineered cells of Butyrivibrio fibrisolvens (Table 23.5) containing a gene encoding the enzyme fluoroacetate dehalogenase. This successfully prevented fluoroacetate poisoning of sheep fed plants containing high levels of this highly toxic citric acid cycle inhibitor.
With this extensive background on the symbiotic microbial communities associated with various nonhuman animals, we now move on to Chapter 24 where the microbial world that populates and affects the health of human beings will be center stage and considered in some detail.
Check Your Understanding
What groups of Archaea are present in the rumen, and how do they contribute to ruminant nutrition?
Fungi and protists are eukaryotic microbes. How can they survive in the anoxic rumen?
Give an example of how changes in the type of fiber given to a ruminant can result in a transition in its rumen microbes.
Why is the metabolism of Streptococcus bovis of special concern for ruminant nutrition, and under what conditions is this bacterium a threat to the animal’s health?
Explore the Microbial World Combating Mosquito-Borne Viral Diseases with an Insect Symbiont
Explore the Microbial World Combating Mosquito-Borne Viral Diseases with an Insect Symbiont
Combating Mosquito-Borne Viral Diseases with an Insect Symbiont
The global reemergence of mosquito-borne viral disease is correlated with rapid human population growth and urbanization, particularly in the tropics. Dengue (Section 32.5) is now the world’s most common mosquito-borne viral disease. In addition, major outbreaks of Zika virus (**Figure 1a,*b***) have occurred in the South Pacific and the Americas, with infection in the Americas associated with congenital abnormalities. Urbanization has created greater opportunities for mosquito reproduction and transmission, and subtropical cities are the preferred habitat of the primary disease vector, Aedes aegypti (Figure 1c), which lives and breeds in building structures, urban waste, and pools of standing water.
Figure 1 Zika virus and its mosquito vector.

(a) Transmission electron microscopic image of Zika viruses. Each virion is approximately 40 nm in diameter. (b) Zika virus structural model. (c) The Aedes aegypti mosquito, the principal vector for transmission of Zika fever, yellow fever, and dengue fever.
With the tremendous resurgence of mosquito-borne disease, conventional mosquito control methods—eliminating breeding sites or applying insecticides—are no longer effective. In response to what is becoming a global crisis, novel technologies based on the rearing and releasing of large numbers of modified mosquitos into wild populations have been developed and tested in the field. The basic concept is to introduce mosquitoes modified in such a way that their mating with the native population yields nonviable offspring, thus reducing the size of the vector population. Some attempts have been made using sterilized males or genetically modified males that carry lethal genes. However, the most successful and cost-effective approach to date, and one that does not require genetic engineering or repeated releases of altered males, is the use of Wolbachia, a bacterium that naturally infects 40–60% of all insects.
Wolbachia is an insect endosymbiont that uses a mechanism called cytoplasmic incompatibility to promote its spread through insect populations. Infected females can mate successfully with both infected and uninfected males, which enables the rapid spread of Wolbachia throughout a naive population. However, when infected males mate with uninfected females, the progeny are not viable, thus effectively sterilizing those females. Cytoplasmic incompatibility appears to be conferred by two incompatibility factors (cifA and cifB) encoded by a bacteriophage integrated into the Wolbachia genome (a prophage, Section 5.6). Infection with Wolbachia also inhibits virus replication in the insect. In fact, mosquito transmission of dengue, Zika, and chikungunya viruses ( Section 32.5) is greatly reduced when A. aegypti mosquitoes are infected with Wolbachia.
The potential for effective pathogen control by Wolbachia was shown by the release of a large number of Wolbachia-infected male Culex quinquefasciatus mosquitoes in locations in Myanmar. This insect is the vector of the filarial nematode that causes elephantiasis ( Section 34.7), and the release effectively eliminated the local mosquito population. Later release studies in Australia showed that the introduction of Wolbachia-infected mosquitoes into an uninfected population can achieve an infection rate of over 90% for years, thus requiring far fewer mosquitoes and releases than other control methods.
The success of these early studies was the genesis for the World Mosquito Program. This program is conducting large-scale release studies of Wolbachia-infected A. aegypti in Brazil, Colombia, and Indonesia that cover more than 2 million inhabitants in each study. These studies will yield the epidemiological evidence (disease tracking, Chapter 30) essential to fully document the efficacy of Wolbachia control of vectorborne diseases. However, as for all new technologies that affect natural systems, caution about unintended consequences is also essential. For example, whiteflies infected with the bacterium Hamiltonella (a symbiont related to enteric bacteria) are more likely than uninfected flies to transmit tomato yellow leaf curl virus to plants.
Chapter Review
Go to Mastering Microbiology for videos, animations, practice tests, and more.
I Symbioses Between Microorganisms
23.1 Lichens are a mutualistic association between one or two species of fungi and an oxygenic phototroph, either an alga or a cyanobacterium.
Q Describe some similarities and differences between the lichen and coral symbioses (Section 23.13).
23.2 The consortium “Chlorochromatium aggregatum” is a mutualism between a phototrophic green sulfur bacterium and a motile heterotrophic bacterium. Mutual benefit is based on the phototroph supplying organic matter to the heterotroph in exchange for motility that permits rapid repositioning in stratified lakes to obtain optimal light and nutrients.
**Q What mechanisms do the consortia use to orient at the appropriate depth in the water column? What have genomic studies revealed about the nature of the “Chlorochromatium” mutualism?**
23.3 Anaerobic microorganisms capable of direct interspecies electron transfer use cytochrome-rich surface structures to form direct electrical connections between cells of different species, such as between a methane-oxidizing archaeon and a sulfate-reducing bacterium. Electrons originating from the oxidation of a hydrocarbon compound (such as methane or ethane) by one species are directly transferred to the electrically connected second organism, which then uses those electrons to reduce an electron acceptor such as sulfate.
Q Assuming that methane was the only available energy source for a sulfate-reducer–ANME consortium, why can it be said that neither partner could exist without the other?
II Plants as Microbial Habitats
23.4 One of the most agriculturally important plant–microbial symbioses is that between legumes and nitrogen-fixing bacteria. The bacteria induce the formation of root nodules within which nitrogen fixation occurs. The plant provides the energy needed by the root nodule bacteria, and the bacteria provide fixed nitrogen for the plant.
**Q Describe the steps in the development of root nodules on a leguminous plant. What is the nature of the recognition between plant and bacterium, and how do Nod factors help control this? How does this compare with recognition in the Agrobacterium–plant system (Section 23.6)?**
23.5 Mycorrhizae are mutualistic associations between fungi and the roots of plants that allow the plant to extend its root system via intimate interaction with an extensive network of fungal mycelia. Both ectomycorrhizae and endomycorrhizae are known. The mycelial network provides the plant with essential inorganic nutrients, and the plant, in turn, supplies organic compounds to the fungus.
Q How do mycorrhizae improve the growth of trees? In what way(s) are the root nodule and mycorrhizal symbioses similar? In what major way do they differ?
23.6 The crown gall bacterium Agrobacterium enters into a unique relationship with plants. Part of the Ti plasmid in the bacterium can be transferred into the genome of the plant, initiating crown gall disease. The Ti plasmid has also been used for the genetic engineering of crop plants.
**Q Compare and contrast the production of a plant tumor by Agrobacterium tumefaciens and a root nodule by a Rhizobium species. In what ways are these structures similar? In what ways are they different? Of what importance are plasmids to the development of both structures?**
III Insects as Microbial Habitats
23.7 A large proportion of insects have established obligate mutualisms with bacteria, the basis of the mutualism often being bacterial biosynthesis of nutrients such as amino acids that are absent from the food the insect feeds on. Long-established obligate mutualisms are marked by extreme genome reduction of the symbiont, with retention of only those genes essential for the mutualism.
Q How is it possible for aphids to feed only on the carbohydrate-rich but otherwise nutrient-poor sap of phloem vessels in plants? Why do symbionts that are transmitted horizontally show less genome reduction, as opposed to the significant genome reduction observed in heritable symbionts?
23.8 The symbionts of many insects produce defensive chemicals that are toxic to specific pathogens and predators of the insect. The biosynthetic capacity to make defensive chemicals often resides on mobile genetic elements such as plasmids and transposons, allowing insects and their protective symbionts to adapt to changing predator threats by horizontal gene transfer.
Q How would horizontal versus vertical transmission of symbionts influence transfer of genes for defensive chemicals between different strains or species of symbionts?
23.9 Termites associate symbiotically with bacteria and protists capable of digesting plant cell walls. The unique termite gut configuration and the hindgut microbial community composed largely of cellulolytic bacteria and protists and acetogenic bacteria result in high levels of acetate, the primary source of carbon and energy for the termite.
Q How do the microbial communities of higher and lower termite guts differ in composition and degradation of cellulose?
IV Other Invertebrates as Microbial Habitats
23.10 Several genera of primarily marine microorganisms have the capacity to generate light through a biochemical process known as bioluminescence, sometimes forming symbiotic associations with marine animals. A light-emitting organ on the underside of the Hawaiian bobtail squid provides a habitat for bioluminescent cells of the bacterium Aliivibrio fischeri. From the mutualism in the light organ, the squid gains protection from predators while the bacterium benefits from a habitat in which it grows quickly and contributes cells to its free-living population.
**Q How is the correct bacterial symbiont selected in the squid–Aliivibrio symbiosis?**
23.11 Most invertebrates living on the seafloor near regions receiving hydrothermal fluids have established obligate mutualisms with chemolithotrophic bacteria. These mutualisms are nutritional, allowing the invertebrates to thrive in an environment enriched in reduced inorganic materials, such as H2S, that are abundant in vent fluids. The invertebrates provide the symbionts an ideal nutritional environment in exchange for organic nutrients.
Q Why is the genome of the tube-worm symbiont thought to be so much larger than the genomes of insect symbionts?
23.12 Entomopathogenic nematodes have established symbiotic associations with species of Photorhabdus and Xenorhabdus bacteria. Following invasion of an insect by the nematode, bacterial symbionts are released into the insect’s hemolymph and multiply rapidly by thwarting the insect’s immune system, killing the insect by the release of toxins and digestive enzymes. When nutrients are depleted, the nematodes then transition to a nonfeeding juvenile form that harbors the symbiont in a specialized receptacle and goes on to infect another insect.
Q Why are entomopathogenic nematodes so attractive for the biocontrol of insect pest species?
23.13 The mutualism between the dinoflagellate Symbiodinium and the stony corals produces the extensive worldwide coral reef ecosystems that sustain a tremendous diversity of marine life. Coral bleaching caused by climate change threatens these ecosystems.
**Q How does the body plan of corals influence their ability to symbiotically associate with Symbiodinium?**
V Mammalian Gut Systems as Microbial Habitats
23.14 Microbial fermentation is important for digestion in all mammals. Several microbial mutualisms have evolved in different mammals that allow for the digestion of different types of food. Herbivores derive almost all of their carbon and energy from plant fiber.
Q From a gastrointestinal standpoint, how do a sheep and a rabbit differ and how are they similar?
23.15 The rumen, the foregut digestive organ of ruminant animals, specializes in cellulose digestion, which is carried out by various microbes. The position of the rumen at the beginning of the animal’s digestive tract allows for protein recovery when rumen microbes are killed in the animal’s acidic stomach.
Q How can the rumen be studied and sampled in a living animal?
23.16 Bacteria, protists, and fungi in the rumen produce volatile fatty acids that provide energy for the ruminant while Archaea convert CO2 and H2 into CH4 that is released to the atmosphere. Rumen microorganisms synthesize vitamins and amino acids and are also a major source of protein—all used by the ruminant.
Q Give an example of two rumen microbes and describe how they contribute to herbivore nutrition.
Application Questions
Choose one invertebrate discussed in this chapter and compare its microbial symbionts with those in the rumen. Why is the microbial community of these two animals considerably different? In your comparison, consider major aspects of the biology of the host (body temperature, organ systems need to resist predators, and the like), its habitat, and its food source(s), as well as the biology of the symbionts, including their habitat, metabolism, and genome characteristics.
Imagine that you have discovered a new animal that consumes only grass in its diet. You suspect it to be a ruminant and have available a specimen for anatomical inspection. If this animal is a ruminant, describe the position and basic components of the digestive tract you would expect to find and any key microorganisms and substances you might look for. What metabolic types of microorganisms or specific genes would you predict would be present?
Why would you be very surprised to find the exact same microbial symbionts inhabiting a lichen and the rumen of a cow? Consider both the physical and chemical conditions of the habitat and the requirement for specific microbes to be present in order for the symbiosis to be successful.
Chapter Glossary
a branched or coiled hyphal structure within cells of the inner cortex of plants with a mycorrhizal infection Autoinduction
the induction of transcription of a set of genes, such as those responsible for bioluminescence, that occurs only when population density is great enough Bacteriocyte
a specialized insect cell in which bacterial symbionts reside Bacteriome
a specialized region in several insect groups that contains insect bacteriocyte cells packed with bacterial symbionts Bacteroid
a misshapen rhizobial cell inside a leguminous plant root nodule; can fix N2 Bioluminescence
the enzymatic production of visible light by living organisms Coevolution
evolution that proceeds jointly in a pair of intimately associated species owing to the effects each has on the other Commensalism
a symbiosis in which one symbiotic partner benefits and the other is unaffected Consortium
a two- or more-membered association of bacteria, usually living in an intimate symbiotic fashion Infection thread
in the formation of root nodules, a cellulosic tube through which Rhizobium cells can travel to reach and infect root cells Leghemoglobin
an O2-binding protein found in root nodules Lichen
one or two species of fungi and an alga (or cyanobacterium) living in symbiotic association Mutualism
a symbiosis in which both partners benefit Myc factors
lipochitin oligosaccharides produced by mycorrhizal fungi to initiate symbiosis with a plant Mycorrhizae
a symbiotic association between a fungus and the roots of a plant in which nutrients are transferred in both directions Nod factors
lipochitin oligosaccharides produced by root nodule bacteria that help initiate the plant–bacterial symbiosis Parasitism
a symbiosis in which one symbiotic partner benefits and the other is harmed Root nodule
a tumorlike growth on plant roots that contains symbiotic nitrogen-fixing bacteria Rumen
the major fermentation vessel in the multichambered gut of ruminant animals, where most cellulose digestion occurs Symbiosis
an intimate relationship between organisms, often developed through prolonged association and coevolution Ti plasmid
a conjugative plasmid in the bacterium Agrobacterium tumefaciens that can transfer genes into plants Volatile fatty acids (VFAs)
also called short-chain fatty acids (SCFAs), these are the major fatty acids (acetate, propionate, and butyrate) produced during fermentation in the rumen
UNIT 6 Microbe–Human Interactions and the Immune System
Unit 6 Microbe–Human Interactions and the Immune System
Unit 6 Microbe-Human Interactions and the Immune System
Chapter 26 Innate Immunity: Broadly Specific Host Defenses
Chapter 27 Adaptive Immunity: Highly Specific Host Defenses
Chapter 28 Immune Disorders and Antimicrobial Therapy