27 Adaptive Immunity: Highly Specific Host Defenses
27 Adaptive Immunity: Highly Specific Host Defenses
## Chapter 27 Adaptive Immunity: Highly Specific Host Defenses
III The Major Histocompatibility Complex (MHC)
Controlling HIV through “Public” T Cell Receptors on CD4 T Cells
Control of human immunodeficiency virus (HIV) proliferation in infected individuals typically requires surveillance of viral activity and highly active anti-retroviral therapy (HAART; see Section 31.15). HAART requires administration of combination anti-retroviral drugs to reduce the viral load and prevent HIV mutation into drug-resistant strains. However, in a small fraction (<1%) of cases, HIV-positive individuals exhibit natural resistance to HIV and maintain an undetectable viral load even in the absence of HAART. These rare individuals, termed “HIV controllers,” maintain high numbers of CD4 T cells that produce a robust immune response against HIV.
As discussed in this chapter, CD4 T cells (see scanning electron micrograph) are “helper” T cells that secrete cytokines to coordinate the adaptive immune response. This is initiated through the interaction of T cell receptors (TCRs), found on the surface of T cells, with antigens presented on major histocompatibility complex (MHC) proteins expressed on the surface of antigen-presenting cells. MHC proteins are encoded by human leukocyte antigen (HLA) genes, and because of a phenomenon called polymorphism, many different HLA alleles occur in the human population.
Research has shown that HIV controllers produce CD4 T cells with high-affinity “public” TCRs that recognize a highly conserved HIV capsid epitope called Gag293 when presented by a broader than usual array of distinct HLA class II molecules. By generating an especially efficient “immunological synapse”—the highly organized interaction that occurs when a TCR engages antigen presented by MHC proteins—the public TCRs conferred unusual cytotoxic activity on the CD4 T cells, allowing them to selectively destroy HIV-infected host cells. This property appears to be a key factor in suppressing HIV replication in HIV controllers and suggests that high-affinity public TCRs are linked to spontaneous control of HIV infection, a finding that ultimately may prove to be of enormous clinical significance.
Source: Galperin, M., et al. 2018. CD4+ T cell-mediated HLA class II cross-restriction in HIV controllers. Sci. Immunol. 3, eaat0687 doi:10.1126/sciimmunol.aat0687.
In the previous chapter we discussed the key features of innate immunity and how this system protects against infection and disease from a broad range of pathogens. Here, we build on this foundation with a focus on the powerful and highly specific immune mechanisms of adaptive immunity, mechanisms that depend on both cellular and molecular components and that complement the innate response.
I Principles of Adaptive Immunity
Adaptive immune mechanisms are highly specific and exhibit immune memory. B and T lymphocytes are the primary effector cells activated in response to immunogen exposure.
Innate and adaptive immunity can be viewed as two arms of the immune response that work together to protect the host from attack by foreign substances. Whereas innate immunity is characterized by broadly targeted responses triggered by common structural features found on microorganisms, adaptive immunity is directed toward specific molecular components of the microbes (their antigens). Thus, it is the individual antigenic properties of different pathogens that orchestrate the adaptive immune response.
We begin this part of the chapter by considering the major characteristics of the adaptive immune response and explore how adaptive immune responses are tailored to address invasions from various pathogens, including viruses, bacteria, and fungi.
27.1 Specificity, Memory, Selection Processes, and Tolerance
27.1 Specificity, Memory, Selection Processes, and Tolerance
27.1 Specificity, Memory, Selection Processes, and Tolerance
Adaptive immunity is primarily a function of B and T lymphocytes. B lymphocytes (B cells) specialize in the production of antibodies that interact with and protect against extracellular antigens, thus conferring antibody-mediated (humoral) immunity on the host. T lymphocytes (T cells) express antigen-specific receptors called T cell receptors (TCRs) on their surfaces that defend against intracellular pathogens, such as viruses and certain bacteria, thus conferring cell-mediated (cellular) immunity on the host. The combination of antibody-mediated and cell-mediated defenses comprises adaptive immunity, a system that is characterized by three major features: specificity, memory, and tolerance. None of these features is characteristic of the innate immune response (Chapter 26).
Immune Specificity and Memory
Overall, the immune response is highly specific, but the innate and the adaptive systems differ in their degree of specificity. Innate immunity is activated upon recognition of features common to a broad diversity of microorganisms, such as the peptidoglycan of all gram-positive bacteria or the lipopolysaccharide (LPS) of all gram-negative bacteria. By contrast, adaptive immune mechanisms are directed against pathogen-specific antigens, such as a specific protein associated with a single strain of a particular pathogen.

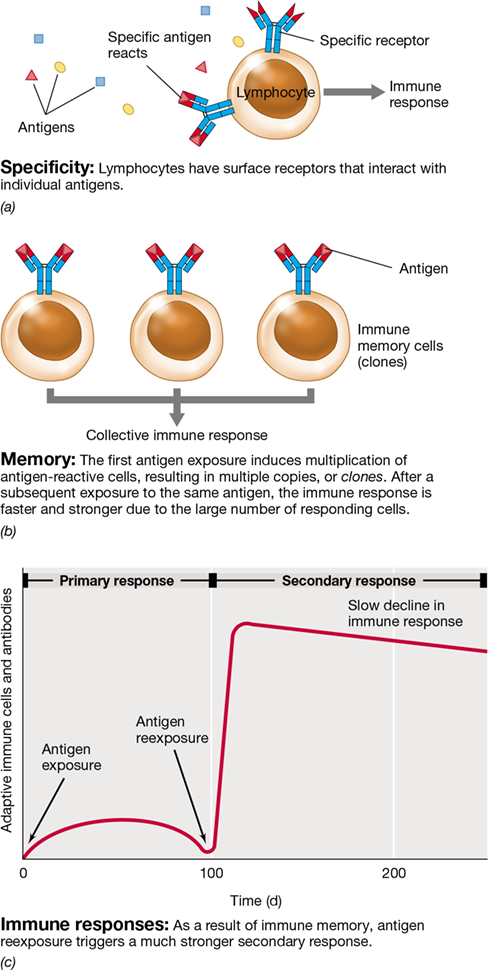
Each B cell or T cell expresses protein receptors that exhibit specificity for a single type of antigen. Just as T cells have antigen-binding TCRs on their surfaces, the antigen-binding proteins of B cells are membrane-bound immunoglobulins called B cell receptors (BCRs). The specificity of the antigen–BCR or antigen–TCR interaction is dependent on the capacity of the lymphocyte cell receptor to interact with a particular antigen but not with other antigens (**Figure 27.1*a***). A pathogen that invades the body introduces multiple antigens that are specific to that pathogen, and each of these antigens will be recognized by distinct lymphocytes bearing receptors that specifically bind them.
Figure 27.1 Specificity and memory in the adaptive immune response.

Key features of antibody-mediated and cell-mediated immunity are (a) specificity and (b) memory. (c) The primary response induces both immune cells and antibodies. The antigens given at day 0 and day 100 must be identical to induce a secondary response. The secondary response may generate more than 10-fold increases in immune cells and antibody concentrations.
In addition to specificity, the adaptive immune system exhibits memory (immune memory) (Figure 27.1b). Lymphocytes must encounter antigen to proliferate in the body and, in the case of B cells, stimulate production of detectable and effective antigen-activated antibodies. The first exposure to an antigen generates a primary immune response in which antigen recognition by specific B or T lymphocytes leads to B and T cell activation, proliferation, and differentiation, thereby resulting in large numbers of antigen-specific B and T cell clones. Some of these clones differentiate into long-lived memory cells, cells which persist in the body for years and confer long-term antigen-specific immunity. Subsequent exposure to the same antigen activates these clones and generates a faster and stronger secondary immune response that peaks within a few days (Figure 27.1c). This capacity to respond more quickly and vigorously to subsequent exposures to the eliciting antigen provides the host with immediate resistance to previously encountered pathogens, a topic we develop in more detail in Section 27.3. Immune memory is one of the most important outcomes of an immune response because vaccinating humans or other animals with killed or weakened pathogens (or their products) is a major and relatively inexpensive means of conferring immunity to dangerous pathogens; this in turn promotes long-term public health (see Section 27.2).
T Cell Selection and Tolerance
The adaptive immune system must be able to discriminate between harmless host antigens (“self”) and potentially dangerous foreign antigens (“nonself”) and function to destroy only the latter. T cells undergo immune selection for potential antigen-reactive cells and selection against those cells that react strongly with self antigens. Selection against self-reactive cells results in the development of another key characteristic of the adaptive immune response: tolerance. Tolerance is a key component of the immune response and ensures that adaptive immunity is directed against agents that pose genuine threats to the host and not against the host’s own proteins. The failure to develop tolerance may result in dangerous reactions to self antigens, a condition called autoimmunity (Section 28.1).
T lymphocyte precursors leave the bone marrow via the bloodstream and enter the thymus (Figure 27.2); the thymus and bone marrow are the primary lymphoid organs (Section 26.4). During the process of T cell maturation in the thymus, immature T cells undergo a two-step selection process to (1) select potential antigen-reactive cells (positive selection) and (2) eliminate cells that react strongly with self antigens (negative selection). Positive selection requires the interaction of immature T cells in the thymus with peptide antigens that are actually of self origin. As we discussed in Chapter 26, T cells interact only with antigens that are presented on major histocompatibility complex proteins (MHCs) expressed on the surface of host cells. Using their TCRs, some immature T cells bind to MHCs that present self peptides on the thymic epithelial tissue. The T cells that do not bind MHC–peptide complexes will be of no use in the immune response and are therefore permanently eliminated via apoptosis (programmed cell death). By contrast, those T cells that bind thymic MHC proteins receive survival signals and therefore remain viable. Positive selection retains T cells that recognize self-peptide:self-MHC and deletes T cells that do not recognize self-peptide:self-MHC and would therefore be unable to recognize MHC–peptide outside the thymus.
Figure 27.2 T cell selection and clonal deletion.

T cells undergo positive selection for functionality and negative selection for dangerously strong recognition of self antigens in the thymus.
Mastering Microbiology
Art Activity: Figure 27.2 T cell selection and clonal deletion
The second aspect of T cell maturation is negative selection. In this process, the positively selected T cells continue to interact with thymic MHC–peptide. Precursor T cells that recognize self-peptide:self-MHC too strongly could potentially mount a harmful immune response against host tissues, and therefore these cells are eliminated by apoptosis. The removal of self-reactive precursor T cells through the processes of positive and negative selection is called clonal deletion, and this important mechanism prevents the propagation of precursor T cells that could potentially cause autoimmune complications. Precursor T cells having TCRs that react less strongly with self-peptide:self-MHC survive negative selection and continue further differentiation (Figure 27.2).
This two-stage T cell selection in the thymus ensures the generation of self-tolerant T cells capable of reacting strongly to foreign antigens only. Precursor T cells that are either useless (do not bind) or harmful (bind too tightly) are deleted in the thymus. Approximately 95–98% of all immature T cells do not survive the thymic selection process. The remaining selected T cells are destined to interact very strongly with nonself antigens. They are not destroyed in the thymus because their weak binding interactions with thymic self antigens signal them to proliferate. The selected and growing T cells leave the thymus and migrate to the spleen, mucosa-associated lymphoid tissue (MALT), and lymph nodes—the secondary lymphoid organs or tissues—where they can engage foreign antigens presented by B cells and other antigen-presenting cells (APCs).
B Cell Selection and Tolerance
To respond to the nearly infinite variety of environmental antigens, the immune system must have the capacity to generate an essentially limitless variety of antigen-specific lymphocytes. To this end, and as we shall see in Section 27.4, the body produces an enormous diversity of B cells, each expressing on its surface BCRs that are specific for a single antigen. B cell selection occurs in the bone marrow—the primary lymphoid organ responsible for B cell maturation in mammals—when each immature B cell is exposed to self antigen. As with T cells, the development of tolerance in B cells is necessary because antibodies produced by self-reactive B cells (autoantibodies) may cause autoimmunity and damage to host tissues (Section 28.1). Therefore, B cells must undergo their own clonal deletion process. Immature B cells whose BCRs bind to self-cell-surface antigens undergo apoptosis and are removed from the B cell repertoire. B cells whose BCRs do not react with self antigens complete development and migrate to peripheral tissues.
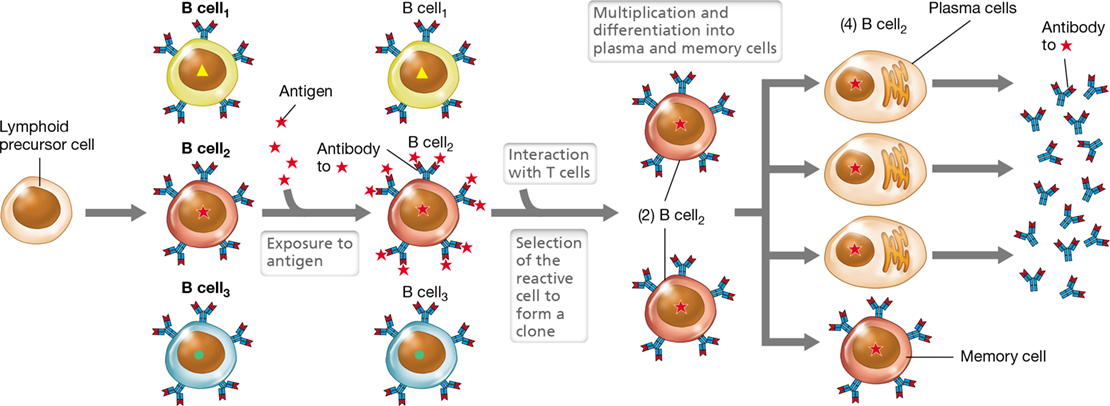
In the periphery (secondary lymphoid tissues), any B cell that recognizes (binds to) its specific foreign antigen becomes activated and can then proliferate and differentiate in a process called clonal expansion (Figure 27.3). This produces a pool of B cells, each of which expresses identical antigen-specific receptors. B cells that have not interacted with antigen are not activated and, therefore, do not proliferate. B cell activation and clonal expansion may occur from encountering either T-independent or T-dependent antigens. T-independent antigens are typically common microbial components composed of repeating subunits, such as lipopolysaccharides and bacterial capsules. Cross-linking of BCRs by T-independent antigens activates the B cell and triggers clonal expansion in the absence of T cell help. Although relatively rapid, immune responses to T-independent antigens are comparatively weak, producing antibody-secreting plasma cells but no long-lived memory cells.
Figure 27.3 B cell clonal selection and expansion.

Among the repertoire of individual B cells (represented by B cell1, B cell2, and B cell3), only B cell2 specifically interacts with the introduced antigen (shown as a star). The antigen drives selection and, following interaction with a Th cell, proliferation of B cell2 to form a clone of antigen-specific B cells. Clonal copies of the original antigen-reactive B cell have the same antigen-specific surface antibody. Continued exposure to antigen results in continued expansion of the clone. Clones differentiate further into many antibody-producing plasma cells and comparatively few long-lived memory B cells. Compare this figure with Figure 27.6.
B cells that interact with T-dependent antigens, which are typically pathogen-specific peptides, must engage a corresponding T-helper (Th) cell (Section 27.8) to become activated and generate a clone of antigen-specific B cells. Upon activation, the newly produced B cells differentiate primarily into antibody-producing plasma cells but also a smaller population of antigen-specific memory cells (Figure 27.3). It is the production of memory cells that confers long-term immunity to the eliciting antigen. Upon subsequent exposure to the same antigen, memory B cells do not require T cell help for activation and, therefore, are able to rapidly undergo clonal expansion and trigger an adaptive response. We explore these concepts further in Section 27.3.
In addition to clonal deletion, clonal anergy (clonal unresponsiveness) also plays a role in final selection of the B cell pool. Some immature B cells are reactive to self antigens but are not activated by them. This is because in addition to antigen recognition, B cell activation requires a second signal from a T-helper cell. The nature of this second signal is a positive interaction between proteins on the surface of B cells and Th cells that triggers the release of cytokines, especially IL-4 (interleukin-4), from the Th cell that activates the B cell. If no second signal is generated because the available Th cells have been rendered tolerant to the antigen in the thymus, the B cell remains unresponsive. Using a similar mechanism, a second signal is also required to activate T cells that are interacting with antigens presented on MHCs (Section 27.7). The requirement for a second activation signal is the key to establishing and maintaining clonal anergy in potentially dangerous self-reactive B and T lymphocytes.
With their key properties of specificity, memory, and tolerance, lymphocytes stand ready to deploy the adaptive immune response. The selection and control processes just described ensure that this response will be directed only against foreign antigens. We now examine the nature of antigens in more detail and consider the basic classes of adaptive immunity.
Check Your Understanding
Distinguish between immune specificity, memory, and tolerance.
Distinguish between positive and negative T cell selection. How do positive selection and negative selection contribute to the development of tolerance in T cells?
Distinguish between clonal deletion and clonal anergy in B cells.
27.2 Immunogens and Classes of Immunity
The adaptive immune response can be induced by a vast range of foreign antigens. As we have discussed, antigens are substances that interact with antibodies or TCRs. Most, but not all, foreign antigens are immunogens, substances that induce an immune response. Here we examine the features of effective immunogens, define the features of antigens that promote interactions with antibodies and TCRs, and discuss the classes of immunity.
Immunogens and Antigen Binding
Immunogens share several intrinsic properties that enable them to induce an adaptive immune response. First, molecular size is an important property of immunogenicity; for a molecule to be immunogenic, it must be sufficiently large. Certain low-molecular-weight compounds called haptens—such as the disaccharide lactose—do not induce immune responses themselves. However, antibodies can still bind haptens, and they may induce an immune response if attached to a larger carrier molecule. Because antibodies can bind them, haptens are considered antigens, but they are not themselves immunogens. Proteins and complex carbohydrates are effective immunogens, whereas nucleic acids, simple polysaccharides with repeating subunits, and lipids are typically not. Thus, sufficient molecular complexity is another key property of immunogenicity. Finally, unlike soluble molecules, large, insoluble macromolecules are usually excellent immunogens because phagocytes readily engulf and process them. Thus, appropriate physical form is another intrinsic property of immunogenicity.
Extrinsic factors, such as the immunogen dose and the route of administration, also influence immunogenicity. The administered dose of an immunogen can be important for an effective immune response, and in general, doses of 10 μg to 1 g are effective in most mammals. Immunogens administered parenterally (outside of the gastrointestinal tract), usually by injection, are normally more effective than those given topically or orally. When administered by oral or topical routes, antigens may be significantly degraded before coming in contact with immune cells and, as a result, may exhibit diminished or no immunogenicity.
As discussed in Chapter 26, an antibody or TCR does not interact with an immunogen as a whole but only with a distinct portion of the molecule called an epitope (also called an antigenic determinant) (Figure 27.4). Epitopes may include sugars, short peptides of four to six amino acids, and other organic molecules that are components of a larger immunogen. Antibodies are able to recognize epitopes on proteins or polysaccharides in their native conformations. By contrast, TCRs recognize epitopes only after the antigens have been partially degraded or processed into peptides by antigen-presenting cells or target cells. T cells recognize peptides shorter than about 20 amino acids presented to them in association with MHC molecules on the surface of host cells.
Figure 27.4 Antigens and epitopes for antibodies.

Antigens may contain several different epitopes, each capable of reacting with a different antibody (Ab). In some cases, epitopes may be formed from nonlinear parts of a folded polypeptide, as is the case for epitope 1, recognized here by Ab1. The folding brings two distant portions of the polypeptide together to make a single epitope.
Antibodies and TCRs can discriminate between closely related epitopes. However, specificity is not absolute, and an individual antibody or TCR may interact to some extent with several different but structurally similar epitopes. The antigen that induces the production of an antibody and, in turn, interacts with that antibody is called the homologous antigen, whereas any noninducing antigens that interact with the antibody are called heterologous antigens. An interaction between an antibody or a TCR and a heterologous antigen is called a cross-reaction.
The engagement of BCRs and TCRs on lymphocytes with naturally encountered immunogens induces an adaptive immune response that often leads to long-term immunity. But as we now discuss, immunity may also be conferred either by purposely introducing an eliciting antigen (vaccination) or through passive transfer mechanisms, administered either naturally or artificially.
Classes of Adaptive Immunity: Active and Passive
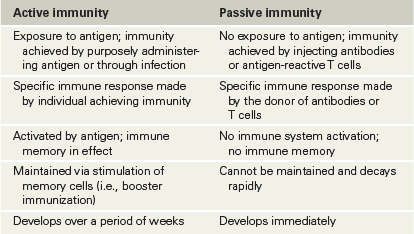
There are two broad classes of adaptive immunity. Active adaptive immunity may develop following a natural or intentional exposure to an immunogen. By contrast, passive adaptive immunity is generated from the transfer of antibodies or immune cells from an immune individual to a nonimmune one. Active and passive immunity are illustrated in Figure 27.5 and contrasted in Table 27.1.
Figure 27.5 Active and passive immunity.

Photos, left to right: (1) Childhood measles showing typical systemic measles rash. Natural active immunity requires infection with a pathogen to activate the adaptive immune response. (2) Vaccination by nasal inhalation of antigen. Artificial active immunity occurs from exposure to particular antigens in a vaccine. (3) A 1934 United States government poster promoting breastfeeding. Natural passive immunity occurs when immunity is transferred from one individual to another by natural means, such as the transfer of maternal antibodies in breast milk. (4) Timber rattlesnakes produce highly toxic venom. An antivenom consisting of purified antibodies to timber rattlesnake venom can be prepared in horses and artificial passive immunity conferred on a snakebite victim by injecting the victim with the antivenom.
Table 27.1 A comparison of active and passive immunity


Active and passive immunity can be further classified as being either natural or artificial. Natural active immunity is the outcome of exposure to antigens through infection and is typically mediated by both B and T cells. By contrast, natural passive immunity occurs when a nonimmune person acquires immune cells or antibodies through natural transfer of cells or antibodies from an immune person, such as from a mother to her fetus before birth or from a mother to a newborn through breastfeeding (Figure 27.5).
Artificial active immunity is conferred by vaccination (immunization) and is a major weapon for the prevention and treatment of many infectious diseases. The introduction of antigen into the host triggers B and T cell activation, resulting in antibody production in a primary immune response and, more importantly, production of a population of immune memory cells. A second dose, often called a “booster,” of the same antigen results in a secondary immune response in which existing memory cells are quickly activated, producing much higher levels of antibodies and a larger population of immune memory cells (Figure 27.1c). Active immunity may persist throughout life as a result of immune memory.
Artificial passive immunity is conferred when an individual receives antibodies from an immune individual through injection of an antiserum. In passively immunized individuals, transferred antibodies gradually disappear from the body, no immune memory is conferred, and a later exposure to the antigen does not elicit a secondary response. Artificial passive immunity is often used as postexposure therapy for acute infectious diseases, such as tetanus or rabies, for individuals not already protected against such diseases by previous vaccination. This approach is also used for treating individuals following bites from venomous animals (antivenom, Figure 27.5). Antisera containing a concentrated suspension of antibodies specific to the venom (toxin) are generated in large immunized animals, such as horses, or obtained from humans following a natural or artificial active immune response against the antigen and stored for use when needed to treat a human or other animal from a potentially fatal toxemia.
Check Your Understanding
Identify the intrinsic and extrinsic properties of an immunogen.
Describe an epitope recognized by an antibody and compare it to an epitope recognized by a TCR.
Give an example for each: natural and artificial active immunity and natural and artificial passive immunity.
II Antibodies
In response to the presence of immunogens, B cells produce antibodies (immunoglobulins) to specifically bind, neutralize, and—with the help of other immune cells—eliminate foreign substances from the body.
Antibodies provide antigen-specific immunity that protects against pathogens and dangerous soluble proteins, such as toxins. Here we look at the production, structural diversity, and antigen-binding function of antibodies. We also consider the organization and recombination of genes that encode antibodies, which underlie the nearly unlimited potential for the adaptive immune response to react to foreign molecules.
27.3 Antibody Production and Structural Diversity
An antibody, or immunoglobulin (Ig), is a soluble protein made by a B lymphocyte or a plasma cell (Figure 26.5) in response to antigen exposure. Each antibody binds to a specific antigen. Antibodies help control the spread of infections by recognizing and binding antigens from pathogens and their products in extracellular environments, such as blood and mucus secretions, and in so doing facilitate the removal of these foreign substances from the body by a variety of mechanisms.
B Cells, Antibodies and Their Activities, and Memory
Each B cell has about 100,000 immunoglobulins of identical antigen specificity that function as B cell receptors (BCRs) on the cell’s surface. To make antibodies, a B cell must first bind antigen by way of its BCR (Figure 27.6). The BCR–antigen interaction causes the B cell to ingest the bound antigen by endocytosis. The B cell then digests the antigen and generates from it a suite of pathogen-derived antigenic peptides that are affixed to proteins of the class II major histocompatibility complex (MHC II) (Section 27.5) and displayed (presented) on the surface of the B cell. The B cell now functions as an antigen-presenting cell and presents the MHC II–peptide complex to a T-helper (Th) cell, which in turn stimulates the B cell to proliferate and differentiate into antibody-secreting plasma cells and memory B cells (Figure 27.6). We consider MHCs and T cells in more detail later in this chapter.
Figure 27.6 B cell–T cell interaction and antibody-mediated immunity.

B cells interact with antigen and Th cells to produce antibodies. B cells initially function as antigen-presenting cells. First, their antigen-specific Ig receptor traps antigen. Following endocytosis, antigens are processed into peptide fragments, which are bound by MHC II and transported to the B cell surface. The MHC II–peptide complex is bound by the TCR on the Th cell, causing the Th cell to secrete IL-4 and IL-5. These cytokines activate the B cell to produce clones that differentiate into many antibody-producing plasma cells and a smaller number of memory cells. The latter are long-lived and can differentiate into antibody-producing plasma cells upon secondary exposure to the same antigen.
Th cells do not interact directly with pathogens but instead stimulate, or “help,” other immune cells, such as B cells and macrophages, to become activated to carry out an immune response. In this case, the cell being “helped” is the antigen-presenting B cell on which the Th cell recognized the MHC II–peptide complex. The recognition of the MHC II–peptide complex on the B cell surface by a cognate Th cell (the Th cell with a TCR specific for that peptide) stimulates the Th cell to produce cytokines and express cell surface receptors that, in turn, stimulate antigen-specific B cells to grow and divide. This results in the expansion of B cell clones having specificity to the original antigen. Activated B cell clones also differentiate into plasma cell clones, each with the ability to produce and secrete large amounts of antibodies of identical antigenic specificity (Figure 27.6). This primary antibody response (Figure 27.1c) is detectable within about five days after antigen exposure, and serum levels of antibodies reach their peak within a few weeks.
Some of the activated B cell clones remain in circulation in the immune system as long-lived memory B cells (Figure 27.6). Subsequent exposure to the same antigen, for example by reinfection with the same pathogen, stimulates the antigen-reactive memory B cells, producing a secondary antibody response characterized by more efficient production of higher quantities of high-affinity antibodies (Figures 27.1c and 27.6). Recall that the secondary response, conferred by immune memory, is the basis for vaccination (Section 27.2).
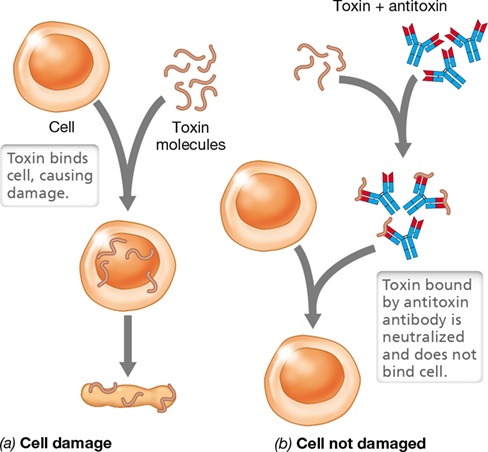
Antibodies released from plasma cells interact with antigen, which is often located directly on a pathogen. Antibody binding may have multiple effects on a pathogen, but most antibody interactions do not directly kill pathogens; instead they block interactions between pathogens (or their products) and host cells. For example, antibodies present on mucous membranes may specifically interact with influenza virus antigens able to bind to host cells, thereby blocking attachment of the influenza virus to host cells on the mucosal surface. In addition, circulating antibodies in blood and lymph serum can neutralize toxins by binding them and thereby prevent their deleterious effects on host cells (Figure 27.7). In other cases, antibodies coat pathogens by binding to their surface antigens, a process called opsonization, thereby marking them for destruction by receptor-mediated phagocytosis (Section 26.9). Phagocytes have antibody receptors called Fc receptors (FcR) that bind to any antibody attached to an antigen, resulting in enhanced phagocytosis of the antibody-coated pathogens (Figure 26.15a).
Figure 27.7 Neutralization of an exotoxin by an antitoxin antibody.

(a) Toxin results in cell destruction. (b) Antitoxin prevents cell destruction. Examples of exotoxins include botulism and tetanus toxins and are discussed in Section 25.6.
Immunoglobulin G Structure and Function
There are five classes of immunoglobulins—IgG, IgM, IgA, IgD, and IgE—distinguished from one another by their different physical, chemical, and immunological properties. Based on these distinctions, each antibody class has a defined distribution and general function (Table 27.2). IgG is the most common circulating antibody, comprising up to 80% of serum immunoglobulins, and it can activate complement through the classical pathway (Section 26.9). IgG is composed of four polypeptide chains that are interconnected by disulfide (S—S) bonds (Figure 27.8). In each IgG molecule, two identical short chains of about 25 kilodaltons (kDa) (called light chains) are paired with two identical longer chains of about 50 kDa (called heavy chains), yielding a Y-shaped, symmetrical molecule. Each heavy chain interacts with a light chain to form a functional antigen-binding site. Therefore, an IgG antibody molecule is bivalent because it has two identical sites that bind two identical antigenic determinants.
Table 27.2 Properties of human immunoglobulins

aAll immunoglobulins may have either λ or κ light-chain types, but not both.
Figure 27.8 Immunoglobulin G structure.

(a) IgG consists of two heavy chains (50,000 molecular weight) and two light chains (25,000 molecular weight), with a total molecular weight of 150,000. One heavy and one light chain interact to form an antigen-binding unit. The variable domains of the heavy (VH) and light chains (VL) bind antigen. The constant domains (CH1, CH2, CH3, CL) are identical in all IgG proteins. The chains are covalently joined with disulfide bonds, shown here as connecting black lines. The Fab (f ragment, antigen-binding) region contains the antigen-binding site. The Fc (f ragment, crystalline) stem of the antibody binds receptor molecules on phagocytes to facilitate phagocytosis of opsonized pathogens. (b) Space-filling model of an IgG molecule. The heavy chains are red and dark blue. The light chains are green and light blue.
Heavy and light chains consist of variable and constant protein domains. A heavy-chain variable domain is connected to three heavy-chain constant domains (Figure 27.8a). The variable domain (VH) and the first constant domain (CH1) compose part of the Fab (fragment, antigen-binding) portion of the immunoglobulin, so named because it contains the antigen-binding site. The two constant domains located most distal to the variable domain (CH2 and CH3) compose the Fc (fragment, crystalline) region of the antibody, named for its tendency to crystallize in solution. As previously mentioned, it is the Fc region of the antibody that binds FcR molecules on the surface of phagocytes to facilitate phagocytosis. The light chains of the antibody consist of one variable (VL) and one constant (CL) domain each and contribute to the two Fab regions only (Figure 27.8a).
Because the variable domains of a given antibody bind a specific antigenic determinant, their amino acid sequences vary in each different antibody. The variable domain of a light chain (VL) interacts with the variable domain of a heavy chain (VH) to bind antigen. By contrast, the constant domains of each antibody are identical in amino acid sequence for all Ig molecules of a given class.
Other Immunoglobulin Classes and Their Functions
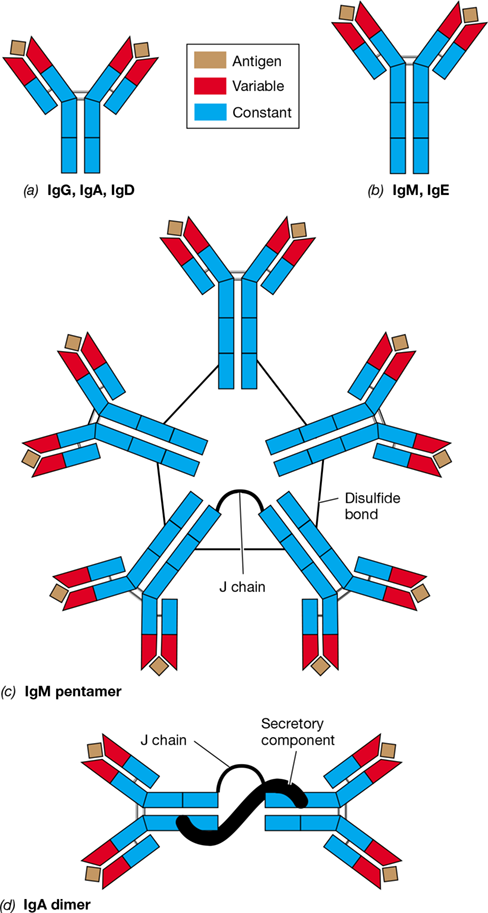
Immunoglobulins of the other classes differ from IgG in structure and function. Because the amino acid sequence of heavy-chain constant domains determines antibody class, the heavy chain called gamma (γ) defines the IgG class. Likewise, alpha (α) defines IgA; mu (μ) defines IgM; delta (δ) defines IgD; and epsilon (ε) defines IgE (Table 27.2). The heavy chains of IgM and IgE contain four constant domains, whereas the heavy chains of IgG, IgA, and IgD contain only three (**Figure 27.9*a, b***).
Figure 27.9 Immunoglobulin classes.

All classes of Igs have VH and VL that bind antigen. (a) IgG, IgA, and IgD have three constant domains. (b) The heavy chains of IgM and IgE have a fourth constant domain. (c) IgM is found in serum as a pentameric protein consisting of five IgM proteins covalently linked to one another via disulfide bonds and a joining (J) chain protein. Because it is a bivalent pentamer, IgM can bind up to ten antigens, as shown. (d) Secretory IgA is often found in body secretions as a dimer consisting of two IgA proteins covalently linked to one another by a J chain protein. The secretory component aids in transport of IgA across mucosal membranes.
The monomeric and pentameric structures of IgM are shown in Figure 27.9b and c, respectively. Circulating IgM forms an aggregate of five or six immunoglobulin molecules attached by at least one J (joining) peptide (Figure 27.9c). Pentameric IgM is the first class of antibody produced in a typical immune response to a bacterial infection, but each individual antigen-binding site of an IgM pentamer generally has low affinity (binding strength) for antigen. However, the avidity (combined antigen-binding strength of multiple epitopes) of IgM antibodies is enhanced because each IgM pentamer has ten binding sites available for interaction with antigen (Table 27.2 and Figure 27.9c). The ten antigen-binding sites make IgM especially effective at agglutinating (clumping) antigens, thereby increasing phagocytic efficiency. Like IgG, IgM is a strong activator of complement through the classical pathway. Whereas up to 10% of serum antibodies are pentameric IgM, IgM monomers do not circulate in the blood but rather are one of the classes of antibodies that function as BCRs.
Dimers of IgA are present in high numbers in secreted body fluids, including saliva, tears, breast milk colostrum, and mucosal secretions from the gastrointestinal, respiratory, and genitourinary tracts. The mucosal surfaces in an average adult, which total about 400 m2 (skin has about 6 m2), contain MALT and produce large amounts (about 10 g) of secretory IgA every day. By comparison, an individual produces only about 5 g of serum IgG per day. Whereas serum IgA typically occurs in monomeric form (Table 27.2), secretory IgA consists of two IgA molecules covalently linked by a J chain peptide and complexed with a protein called the secretory component that aids in transport of IgA across membranes (Figure 27.9d). The high concentration of secretory IgA in breast milk colostrum likely plays a key role in preventing gastrointestinal disease in newborns.
Although IgE is present in small amounts in serum, most IgE is bound to cells. For example, through its Fc region IgE binds eosinophils, arming these granulocytes to target parasitic pathogens such as schistosomes and other helminths (Section 34.7). IgE also binds to tissue mast cells, where subsequent binding of antigen to antigen-binding sites of IgE causes degranulation of the mast cells. The ensuing release of chemical mediators, such as histamine and serotonin, triggers immediate-type hypersensitivities (a type of allergic response; Section 28.1). Like IgM, IgE has a fourth heavy-chain constant domain (Figure 27.9b), and this allows for binding of IgE to eosinophils and mast cells, the key step for activating the protective and allergic reactions, respectively, associated with these cell types.
IgD (Figure 27.9a), present in serum in low concentrations, has no known unique immune protective function. However, IgD, like IgM, is abundant on the surfaces of mature B cells, especially memory B cells, where it functions as a BCR.
Antigen Exposure, Immune Memory, and the Primary and Secondary Responses
We have seen that immune memory is a major outcome of the adaptive immune response (Figure 27.1c). Starting with a B cell, an antibody-mediated immune response begins with antigen exposure and culminates with the production and secretion of antigen-specific antibodies, and the type and route of antigen exposure influences the class of the antibodies produced. Blood and lymph, as well as the spleen and MALT (Chapter 26), are key sites for engagement of antigens by immune components. Bloodborne pathogens/antigens are captured in the spleen, where IgM, IgG, and serum IgA antibodies are produced. Antigen introduced subcutaneously, intradermally, topically, or intraperitoneally is carried through the lymphatic system to the nearest lymph nodes, again stimulating production of IgM, IgG, and serum IgA. Antigen introduced to mucosal surfaces is delivered to the nearest MALT. For example, antigen delivered orally is delivered to the MALT in the intestinal tract, stimulating production of secretory IgA in the gut.
Following initial BCR–antigen contact, each B cell stimulated by T-dependent antigen multiplies and differentiates to form antibody-secreting plasma cells and memory B cells (Figure 27.6). Plasma cells in this primary antibody response are relatively short-lived (less than one week) and secrete large amounts of mostly IgM antibody (Figure 27.10). After a latent period of several days, antibody appears in the blood and a gradual increase in antibody titer (antibody quantity) occurs. As antigen disappears, the primary antibody response slowly diminishes.
Figure 27.10 Primary and secondary antibody responses in serum.

The antigen exposures at day 0 and day 100 must be to identical antigen to induce a secondary response. The secondary response, also called a booster response, may be more than 10-fold greater than the primary response. Note the class switch from IgM production in the primary response to IgG production in the secondary response.
Mastering Microbiology
Art Activity: Figure 27.10 Primary and secondary antibody responses in serum
Memory B cells generated by the initial exposure to antigen may circulate in the host for years. A second exposure to the same antigen stimulates memory B cells to rapidly differentiate into plasma cells and produce antibody without input from helper T cells. The second and each subsequent exposure to antigen causes the antibody titer to rise rapidly to a level often 10–100 times greater than the titer following the first exposure (Figure 27.10). This rise in antibody titer is called the secondary antibody response. The secondary response is a function of immune memory and typically results in a switch from the production of IgM to another antibody class, a transition called class switching (Figure 27.10). This switch is induced by a cytokine signal produced in response to the specific type of pathogen. The signal prompts a recombination event that switches the heavy-chain constant region, resulting in the production of the antibody class that can most efficiently address the threat. For example, in serum, the most common antibody class switch is from IgM to IgG, whereas in mucosal tissues, switches to IgA predominate. It is important to note that class switching does not cause a change in antigen specificity; the new class of antibody generated remains specific to the original antigen.
Serum antibody titer slowly decreases over time, but subsequent exposure(s) to the same antigen can trigger another secondary response. This results in the maintenance of high levels of memory cells and circulating antibody specific for the antigen, providing long-term active immunity against individual infectious diseases.
Check Your Understanding
Compare and contrast B cell activation from T-dependent and T-independent antigens (Section 27.1).
Differentiate among antibody classes using structural characteristics, distribution patterns, and functional roles.
Explain the rationale for periodic revaccination in children and adults.
27.4 Antigen Binding and the Genetics of Antibody Diversity
27.4 Antigen Binding and the Genetics of Antibody Diversity
27.4 Antigen Binding and the Genetics of Antibody Diversity
We have seen that antibodies consist of four polypeptides, two heavy (H) chains and two light (L) chains (Figure 27.8), and each chain is composed of constant (C) and variable (V) domains. The V domains of one H and one L chain interact to form the antigen-binding site. Here we examine the structural features of the V domains that define the antigen-binding site and then explore the genetics behind the enormous diversity of antibodies that, collectively, B cells are capable of producing.
Variable Domains and Antigen–Antibody Interaction
The V domains of an H and an L chain bind antigen strongly but not covalently. To accommodate all possible antigens that a host might encounter in a lifetime, the synthesis of billions of different antibodies, each with its own unique antigen-binding site, is necessary. The V domains define these unique antigen-binding sites.
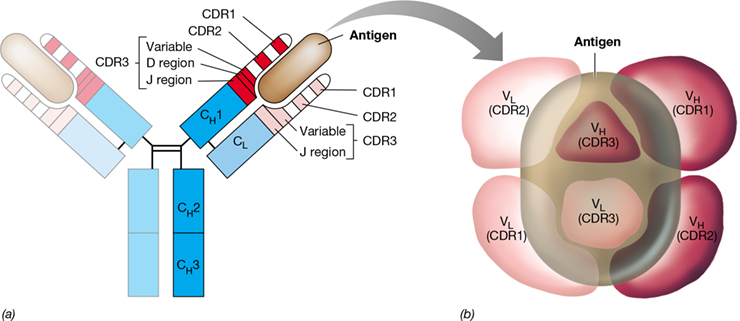
Amino acid sequences differ in the V domains of Igs that bind different antigens. Amino acid variability is especially apparent in several complementarity-determining regions (CDRs). Three CDRs in each of the V domains provide most of the molecular contacts with antigen (Figure 27.11). The amino acid sequences of CDR1 and CDR2 differ in minor ways between different Igs, while CDR3s differ dramatically in sequence from one another. Three distinct gene segments encode CDR3 of the H chain: the carboxy-terminal portion of the V domain, followed by a short “diversity” (D) segment of about three amino acids, and a longer “joining” (J) segment about 13–15 amino acids long. The light-chain CDR3 is similar except that it lacks the D segment. The heavy- and light-chain CDRs, six in total, confer antigen specificity on the antibody molecule (Figure 27.11).
Figure 27.11 Antigen binding by immunoglobulin light and heavy chains.

(a) An Ig is shown schematically, with bound antigen (light brown). The V domains on the H and L chains are shown in red (heavy chain) and pink (light chain) with their antigen-binding complementarity-determining regions (CDR1, CDR2, and CDR3). CH1, CH2, and CH3 (dark blue) are constant domains in the H chain, and CL (light blue) is the constant domain in the L chain. (b) CDRs from both H (red) and L (pink) chains, shown from above, are conformed to make a single antigen-binding site on the Ig. The highly variable CDR3s from both H and L chains cooperate at the center of the site. Antigen (light brown) may contact all CDRs. The shape of the site may be a shallow groove or a deep pocket, depending on the antibody–antigen pair involved.
The Ig three-dimensional structure was shown in Figure 27.8b. Each antigen–antibody interaction requires the specific binding of an antigenic epitope with the CDR domains of the H and L chains. A single antigen-binding site of an antibody molecule measures about 2×3 nm, creating a space large enough to accommodate an epitope of 10 to 15 amino acids. Antigen binding is ultimately a function of the Ig folding pattern of the H and L polypeptide chains. The Ig folds of the V region bring all six CDRs together, resulting in the formation of a unique and specific antigen-binding site (Figure 27.11).
Because antigen diversity is enormous, the immune system must be capable of producing antibodies to recognize a virtually unlimited variety of antigens. In order to accomplish this from a relatively small number of immunoglobulin genes, several unusual genetic mechanisms come into play. Table 27.3 summarizes these diversity-generating mechanisms, and we explore each below.
Table 27.3 Generation of antigen-binding receptor diversity in B cells and T cells

Organization of Immunoglobulin Genes
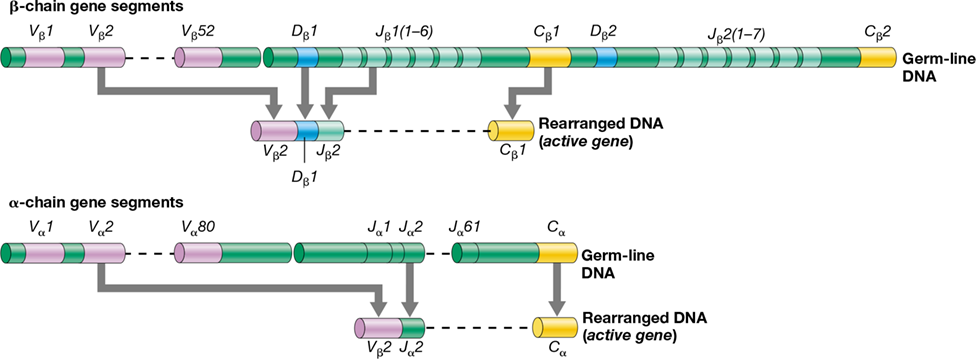
The gene encoding each immunoglobulin H or L chain is constructed from several gene segments. In each B cell, Ig gene segments undergo a series of random, somatic rearrangements characterized by genetic recombination followed by deletion of intervening sequences. These events produce a single, functional antibody gene derived from a relatively small pool of Ig gene segments. As a B cell matures, V, D, and J gene segments are enzymatically recombined to form a single Ig heavy-chain gene (**Figure 27.12*a***). The single V gene segment encodes CDR1 and CDR2, whereas CDR3 is encoded by a mosaic of the 3’ end of the V region, followed by the D and J gene segments.
Figure 27.12 Immunoglobulin gene rearrangement in human B cells.

Ig genes are arranged in tandem on three different chromosomes. (a) The H chain gene complex on human chromosome 14. The filled boxes represent Ig coding gene segments. The dashed lines indicate intervening sequences and are not shown to scale. (b) The κ light-chain complex on human chromosome 2. The λ light-chain genes are in a similar complex on human chromosome 22. (c) Assembly of one-half of an antibody molecule.
In each B cell, only one productive Ig-encoding rearrangement each occurs in the heavy- and light-chain genes. Called allelic exclusion, this mechanism causes each B cell to express Ig genes from only one of the two inherited parental alleles, thus ensuring that all Igs from a given clone of B cells have identical antigen specificity. Finally, separate C gene segments encode the class-defining constant domains of Igs. Therefore, four different gene segments—V, D, J, and C—recombine to form one functional heavy-chain gene (Figure 27.12a). Light chains, which lack D segments, are encoded in a similar way by recombination of light-chain V, J, and C segments (Figure 27.12b). The gene segments required for all Igs exist in all nucleated cells of the body but undergo recombination only in developing B lymphocytes.
V, D, J, and C gene segments are separated by noncoding DNA sequences typical of gene arrangements in eukaryotes (Section 6.6). Genetic recombination occurs in each B cell during its development. One each of the V, D, and J segments is recombined at random to form a functional heavy-chain gene, while randomly recombined V and J segments form a complete light-chain gene. The active gene, still containing an intervening sequence between the VDJ or VJ gene segments and the C gene segments, is transcribed, and the resulting primary RNA transcript is spliced to yield the final messenger RNA (mRNA). The mRNA is then translated to make the heavy and light chains of the Ig molecule.
VDJ Recombination and Somatic Hypermutation
If one considers all of the random reassortments possible in genes encoding immunoglobulin heavy and light chains and from these calculates the number of unique molecules that could be encoded, the total diversity is astonishing. In humans, for example, based on the numbers of kappa (κ) light-chain gene segments, there are about 40 V×5 J possible rearrangements, or 200 possible κ light chains. For the alternative lambda (λ) light chain, the type produced instead of κ by some B cells in every individual, there are about 30 V×4 J=120 possible combinations. About 10,500 possible heavy chains can be formed by the rearrangement of about 65 V×27 D×6 J genes. Assuming that each heavy chain and light chain has an equal chance to be expressed, there are 10,500×200=2,100,000 possible immunoglobulins with κ light chains and 10,500×120=1,260,000 possible immunoglobulins with λ chains. Therefore, at least 3,360,000 possible antibodies can be expressed based on random reassortment alone!
On top of this impressive diversity, additional diversity is generated in the CDR3 regions of both heavy and light chains by several unique mechanisms. First, the DNA-joining mechanism that constructs V-D or D-J gene segments in the heavy chain or the V-J gene segments in the light chain is an imprecise process. The final nucleotide sequence at these regions occasionally varies by a few nucleotides from the original genomic sequence, and when this occurs, it will alter the amino acid sequence in this region. Even more diversity is generated at V-D and D-J coding joints in the heavy-chain genes and at V-J coding joints in light-chain genes by either random (N) or template-specific (P) nucleotide additions. Because these coding joints are contained within the sequences that encode CDR3 on both heavy and light chains, N and P diversity at V-domain coding joints changes or adds amino acids in the CDR3 of both heavy and light chains.
VDJ recombination events are antigen-independent and occur as B cells develop in the bone marrow. In mature B cells that have migrated to lymphoid tissues, immunoglobulin diversity is further expanded upon antigen exposure by a process called somatic hypermutation—the mutation of B cell Ig genes at much higher rates than those observed in other genes. Somatic hypermutation of Ig genes is typically evident after a second exposure to an immunizing antigen and occurs only in the V regions of rearranged heavy- and light-chain genes, creating B cells with slightly altered Ig cell surface receptors. These mutated B cells compete for available antigen, and B cells whose receptors have a higher affinity for antigen than the original B cell receptor are selected. This process, called affinity maturation, is one of the factors responsible for a dramatically stronger secondary immune response (Figures 27.1c and 27.10). Affinity maturation also adds more diversity to the pool of antibody specificity in the adaptive immune response, thus making the potential antibody repertoire essentially infinite.
Check Your Understanding
Draw a complete Ig molecule and identify antigen-binding sites on the antibody.
Describe antigen binding to the CDR1, 2, and 3 regions of the heavy-chain and light-chain variable domains.
Describe the recombination events that produce a mature heavy-chain gene and other somatic events that further enhance antibody diversity.
III The Major Histocompatibility Complex (MHC)
The major histocompatibility gene complex (MHC) encodes cell surface proteins that present antigens to T cells, providing an important link between innate and adaptive immune mechanisms.
The major histocompatibility complex (MHC) is a series of genes found in all vertebrates that encodes a group of proteins important in antigen presentation. The MHC proteins in humans are called human leukocyte antigens (HLAs) and were first identified as the major antigens responsible for immune-mediated tissue transplant rejection. We now know that MHC proteins function primarily as antigen-presenting molecules, binding pathogen-derived peptides and displaying these peptides for interaction with T cell receptors.
27.5 MHC Proteins and Their Functions
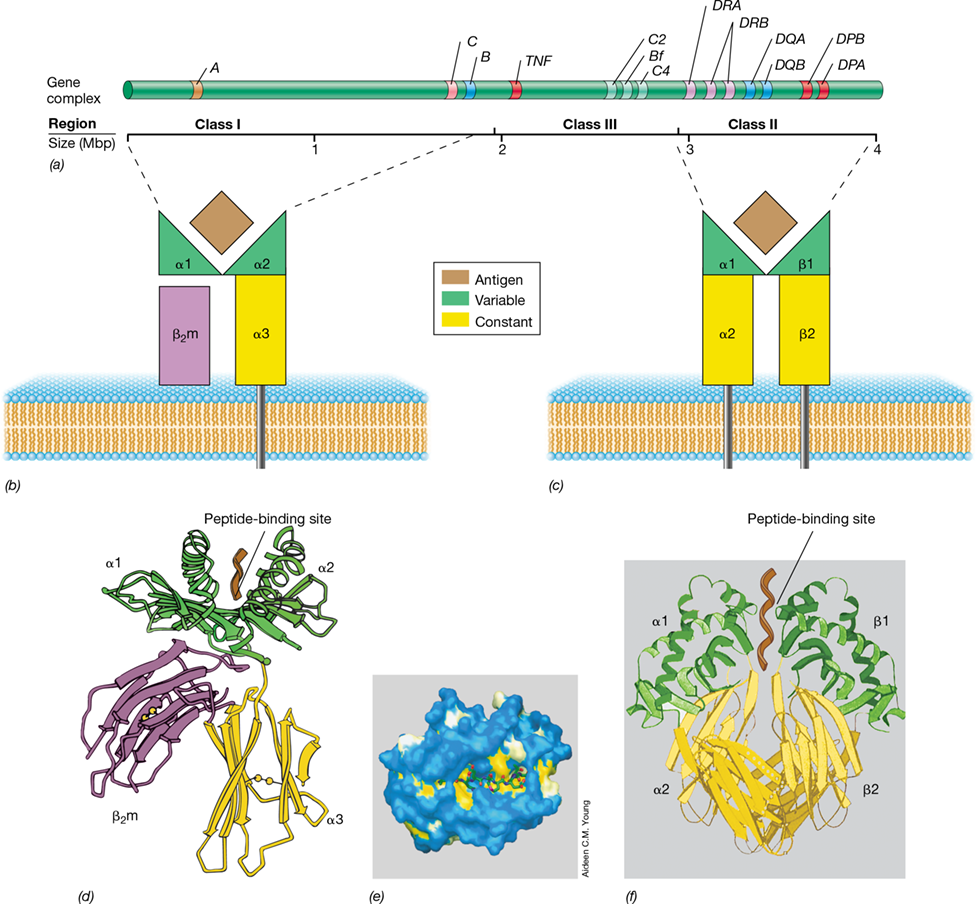
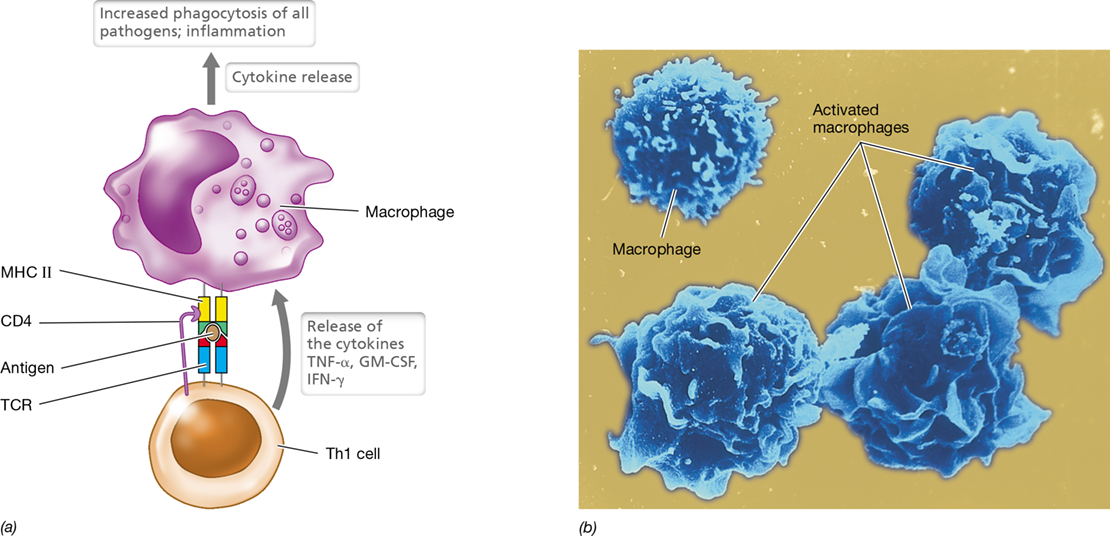
The MHC proteins consist of two protein classes encoded by about 4 megabase pairs (Mbp) of DNA (**Figure 27.13*a***). MHC class I proteins are expressed on the surfaces of all nucleated cells and function to present peptide antigens to T-cytotoxic (Tc) cells. If a peptide antigen presented by MHC class I is recognized by the TCR of a Tc cell, the antigen-containing cell is quickly destroyed (Section 27.8). MHC class II proteins are found only on the surface of B lymphocytes, macrophages, and dendritic cells—the antigen-presenting cells (APCs; Section 26.4). Through their MHC class II proteins, APCs present antigens to T-helper (Th) cells, stimulating cytokine production that leads to antibody-mediated immunity or inflammatory responses (Section 27.8).
Figure 27.13 Human leukocyte antigen (HLA) genes and MHC proteins.

(a) The HLA complex, located on human chromosome 6, contains more than 4 megabase pairs (Mbp). Class II genes DPA and DPB encode class II proteins DPα and DPβ; DQA and DQB encode DQα and DQβ; DRA and two DRB loci encode DRα and DRβ proteins. The class I MHC proteins HLA-A, HLA-C, and HLA-B are encoded by genes A, C, and B. The class II and class I loci are highly polymorphic and encode peptide-binding proteins. Class III MHC genes encode proteins associated with immune-related functions, such as complement proteins C4 and C2 and the cytokine TNF (tumor necrosis factor). (b) Schematic of MHC class I protein. The α1 and α2 domains interact to form the peptide antigen–binding site. (c) Schematic of MHC class II protein. The α1 and β1 domains combine to form the peptide antigen–binding site. (d) MHC class I protein structure. Beta-2 microglobulin (β2m) binds noncovalently to the α chain. The antigen peptide (brown) is bound cooperatively by the α1 and α2 domains. (e) An MHC I protein with a bound peptide, as seen from above. A peptide of nine amino acids is shown as a molecular backbone structure embedded in a space-filling model of a mouse MHC I protein. (f) Structure of MHC class II protein dimer. The antigen peptide (brown) is shown embedded in the peptide-binding site of the MHC II proteins.
Class I and Class II MHC Proteins
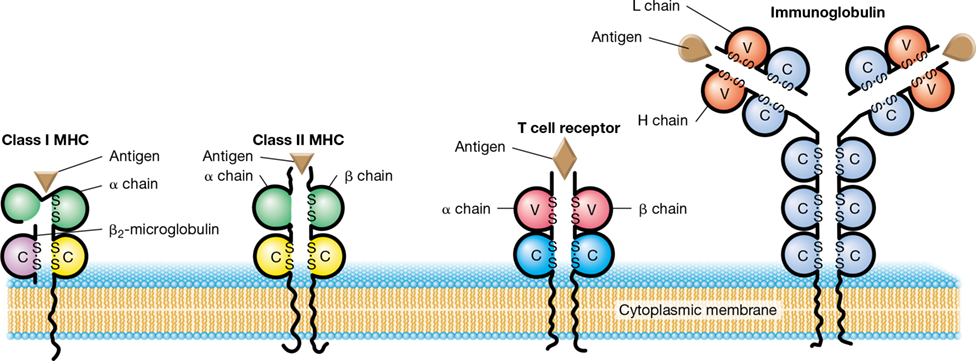
Class I MHC proteins consist of the membrane-embedded alpha (α) chain, encoded by a gene located in the MHC gene region, and the beta-2 microglobulin (β2m), a smaller, noncovalently associated protein encoded by a gene not included in the MHC gene cluster (Figure 27.13b and d). The α chain folds to form a peptide-binding groove that accommodates peptides of 8 to 11 amino acids (Figure 27.13d and e). The peptides in infected cells are derived from endogenous (intracellular) antigens, for example, from viral proteins produced inside the cell. Following translocation to the cell surface, the MHC–virus-peptide complexes are recognized by TCRs on activated Tc cells, and the Tc cells then kill the infected target cells.
Class II MHC proteins consist of two membrane-integrated polypeptides, α and β, expressed primarily by APCs. Together, one α and one β polypeptide form a functional heterodimer (Figure 27.13c, f ). The α1 and β1 domains of the MHC class II protein interact to form a binding site for TCR–peptide similar to the MHC class I binding site for TCR–peptide. However, the peptide-binding groove in the class II MHC is able to bind and display peptides that may be significantly longer than 8–11 amino acids. Peptides presented by class II MHCs are typically 10 to 20 amino acids long and are proteolytic fragments derived from exogenous (extracellular) antigens from pathogens that have been phagocytosed and digested by APCs. The APCs use the MHC class II–peptide complex to interact with TCRs on Th cells, leading to Th activation and an adaptive immune response (Section 27.8).
Antigen Processing and Presentation to T Cells
MHC proteins are expressed on cell surfaces only when they are complexed with a peptide, either a self peptide or a foreign peptide. In essence, then, MHC–peptide complexes reveal to the immune system the protein composition of the cell and, in this capacity, function to alert the immune system when a cell contains foreign antigens. T cells, through their TCRs, scan cell surfaces to identify any cells expressing foreign antigens as an indication of infection. When the latter are encountered, the TCR interacts with the foreign antigen presented on the MHC I protein, and this interaction targets the cell for destruction. T cells do not destroy cells presenting MHC–self peptide complexes because self-reactive T cells that escape clonal deletion in the thymus typically become anergic upon recognition of self antigens (Section 27.7).
Mastering Microbiology
Activities: Microbiology Animation: Antigen Processing and Presentation: Steps
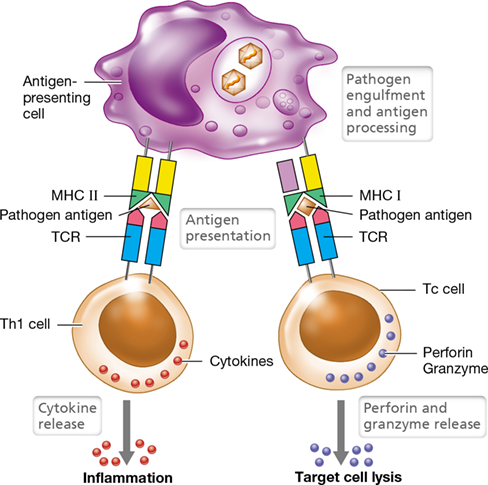
Two distinct antigen-processing schemes are at work to initiate adaptive immune responses: one for MHC I antigen presentation and another for MHC II antigen presentation. MHC I proteins present peptide from viruses or other intracellular pathogens; such infected cells are called target cells (**Figure 27.14*a***). Proteins derived from infecting viruses, for example, are taken up and digested in the cytoplasm in a structure called the proteasome. Peptides about ten amino acids long are transported into the endoplasmic reticulum (ER) through a pore called the transporter for antigen processing (TAP). Once the foreign peptides have entered the ER they are bound by MHC I, and the MHC I–peptide complex moves to the cell surface where it integrates into the cell membrane. When the TCR on the surface of a Tc cell interacts with both the foreign peptide (recognized as “nonself”) and the MHC I protein (recognized as “self”) on the surface of the target cell, the Tc cell releases perforin and granzyme, cytotoxic proteins that kill the virus-infected target cell.
Figure 27.14 Antigen presentation by MHC I and MHC II proteins.

(a) 1 Protein antigens, such as virus components manufactured within the cell, are degraded by the proteasome in the cytoplasm. The peptide fragments are transported into the endoplasmic reticulum (ER) through a pore formed by the TAP proteins. 2 MHC I proteins in the ER are stabilized by chaperonins until peptide fragments are bound. 3 When peptide fragments are bound by MHC I, the complex is transported to the cell surface. 4 The MHC I– peptide complex interacts with T cell receptors (TCRs) on the surface of Tc cells. 5 The CD8 coreceptor on the Tc cell engages MHC I, resulting in a stronger complex. The Tc cell is activated by the binding events, causing it to release cytokines and cytolytic toxins and kill the target cell. (b) 1 External foreign proteins are imported into the cell and digested into peptide fragments in phagolysosomes. 2 MHC II proteins in the ER are assembled with Ii, a blocking protein that prevents MHC II from binding with peptides in the ER. 3 The MHC II–Ii assembly is transported to the lysosome, where it remains until the lysosome fuses with the phagosome, forming a phagolysosome where Ii is degraded, 4 freeing the MHC II protein to bind the foreign peptide fragments. 5 The MHC II–peptide complex is transported to the cell surface, where it interacts with TCRs and 6 the CD4 coreceptor on Th cells. The Th cells then release cytokines that interact with other cells to promote an immune response. Note that the APC in part b may be either a B cell, which ingests antigen by endocytosis (shown), or a macrophage or dendritic cell, which engulf antigens through phagocytosis.
MHC II proteins are expressed on APCs, where they function to present peptide antigens from engulfed pathogens (Figure 27.14b). Similar to MHC I, MHC II proteins are assembled in the endoplasmic reticulum. In the ER, the MHC II peptide-binding groove is blocked by a protein called invariant chain (Ii) until the MHC II complex is situated in a lysosome (Figure 27.14b). After ingestion of a pathogen or a pathogen product by an APC, the phagosome containing the foreign antigen fuses with a lysosome to form a phagolysosome (Section 26.7). Within the phagolysosome, enzymes digest both the foreign antigens and the Ii peptide but not MHC II. The foreign peptides are then loaded onto the newly freed MHC II peptide-binding groove, and the complex is inserted into the cell surface for presentation to T-helper cells. The latter, through their TCRs, recognize the MHC II–peptide complex and secrete cytokines that activate APCs, such as macrophages for enhanced phagocytosis or B cells for antibody production.
In addition to the TCR, each T cell expresses a unique cell surface protein that functions as a coreceptor. Tc cells express the CD8 coreceptor, and Th cells express the CD4 coreceptor (Figure 27.14). When the TCR binds to the peptide–MHC complex, the coreceptor on the T cell also binds to the MHC protein on the antigen-presenting cell, strengthening the molecular interactions between the cells and enhancing activation of the T cell. CD4 binds only to MHC II, strengthening Th cell interaction with APCs that express MHC II protein. Likewise, CD8 binds only to MHC I, enhancing the binding of Tc cells to MHC I–bearing target cells. In clinical medicine, the CD4 and CD8 proteins are used as T cell markers to differentiate Th (CD4) cells from Tc (CD8) cells in diagnostic tests, for example, in assessing the course of disease in an AIDS patient (Section 31.15 and Figure 31.48).
We now consider the fact that human MHC proteins allow for some sequence variation from individual to individual and how this variation affects the immune response.
Check Your Understanding
Identify the cells that display MHC class I and MHC class II proteins on their surfaces.
Compare the MHC I and MHC II protein structures and peptide-binding sites. How do they differ? How are they similar?
Define the sequence of events for processing and presenting antigens from both intracellular (endogenous) and extracellular (exogenous) pathogens.
27.6 MHC Polymorphism, Polygeny, and Peptide Binding
27.6 MHC Polymorphism, Polygeny, and Peptide Binding
27.6 MHC Polymorphism, Polygeny, and Peptide Binding
Although MHC class I and class II proteins theoretically can bind all possible antigen peptides for presentation to T cells, MHC proteins in different individuals of the same species are not identical. Different individuals typically have subtle differences in the amino acid sequence of homologous MHC proteins. These genetically encoded MHC variants, called polymorphisms, are the major immunological barriers for successful tissue transplantation from one individual to another.
Mastering Microbiology
Art Activity: Figure 27.14 Antigen presentation by MHC I and MHC II proteins
Polymorphism, Polygeny, and the Immune Barrier to Tissue Transplantation
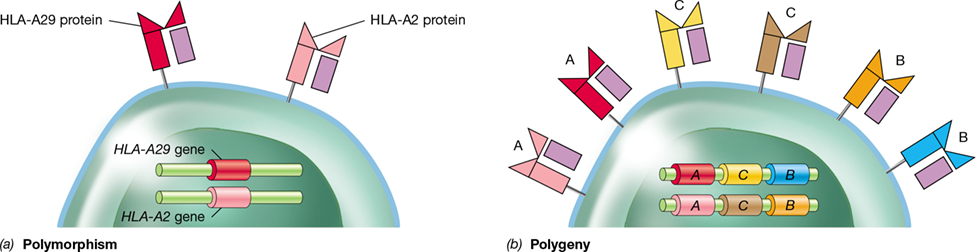
Polymorphism is the occurrence, within a population, of multiple alleles (alternate forms of a gene) at a specific locus (the location of the gene on the chromosome). For example, the MHC class I locus HLA-A (Figure 27.13a) has over 2000 known alleles, each of which encodes a distinct HLA-A protein that occurs within the human population. The genome of each person, however, contains only two of the HLA-A alleles; one allele is of paternal origin and one is of maternal origin. The two allelic protein products are expressed equally (**Figure 27.15*a***).
Figure 27.15 Polymorphism and polygeny in MHC genes and proteins.

(a) Polymorphism in HLA-A loci results in equal expression of proteins encoded by both alleles. There are over 2000 HLA-A alleles in the human population, but only two (one at each locus) are found in each individual. HLA-B and HLA-C exhibit similar levels of polymorphism. (b) Polygeny in MHC results in duplicated polymorphic HLA-A, HLA-B, and HLA-C genes that potentially encode three pairs of different MHC proteins. The colors represent alternate alleles of each gene and their respective protein.
Additional MHC class I protein diversity is a result of polygeny, in which evolutionary gene duplication events have led to the occurrence of two additional genetically, structurally, and functionally related loci, HLA-B and HLA-C. These gene loci are also polymorphic, having more than 2600 and 1500 allelic variants, respectively. Thus, an individual typically displays six structurally distinct proteins derived from the three polymorphic class I loci (three products derived from maternal origin and three products derived from paternal origin) (Figure 27.15b). Likewise, highly polymorphic and equally expressed alleles encode MHC class II proteins at the HLA-DR, HLA-DP, and HLA-DQ alpha- and beta-chain loci (Figure 27.13a).
As a result of polymorphism and polygeny, most individuals have unique MHC profiles. Only closely related family members are likely to have all of the same MHC genes and proteins, and this property can be used to prove (or disprove) paternity and has also been used to trace ancestral lineages and pathways of early human migration (Figure 27.16). These highly polymorphic variations in MHC proteins are major barriers to successful tissue transplants because the MHC proteins on the donor tissue (graft) are recognized as foreign by the recipient’s immune system. An immune response directed against the graft MHC proteins thus causes rejection and death of the graft tissue. However, matching MHC alleles between donors and recipients minimizes tissue graft rejection. Control of tissue rejection may also be accomplished by administering drugs that suppress the immune system.
Figure 27.16 Polymorphic *HLA* genes and tracing ancestral lineages.
The image shows what a Neanderthal man might have looked like. By analyzing the distribution of specific HLA alleles within different populations, researchers are increasingly able to reconstruct migration and interbreeding patterns of ancient humans and other hominids, including as far back as the Neanderthal era. The intermixing of HLA genes between disparate ancient populations increased genetic diversity and likely enhanced survival by helping to strengthen and shape the modern immune response.
Peptide Antigen Binding
Most of the allelic variations in MHC proteins occur as amino acid changes concentrated in the peptide-binding groove (Figure 27.13d–f ), and each polymorphic variation of the MHC protein binds a different set of peptide antigens. The peptides bound by a single MHC protein share a common structural pattern—a peptide motif—and each different MHC protein binds a different motif. For example, for a certain MHC class I protein, the bound peptides contain eight amino acids with a phenylalanine (F) at position 5 and a leucine (L) at position 8 (Figure 6.27 for the structures of amino acids). All other positions in the peptide can be occupied by any amino acid (designated by an X). Thus, all peptides sharing the sequence X-X-X-X-F-X-X-L would bind to that MHC protein. Another MHC class I protein encoded by a different MHC allele binds a peptide motif of nine amino acids with a tyrosine (Y) at position 2 and an isoleucine (I) at position 9 (X-Y-X-X-X-X-X-X-I).
The invariant amino acids in each motif are called anchor residues—they bind directly and specifically within an individual MHC peptide-binding groove. Thus, an individual MHC protein can bind and present many different peptide antigens if the peptides contain the same anchor residues. Because each MHC protein binds a different motif with different anchor residues, the six possible MHC I proteins encoded in an individual’s genome bind six different motifs. In this way, each individual can present a large number of different peptide antigens using the limited number of MHC I molecules available. MHC II proteins bind peptides in a similar manner.
Because of polymorphisms and polygeny within the human species, at least a few peptide antigens from virtually any pathogen will display a motif that can be bound and presented by the MHC proteins. This system of generating antigen-binding diversity is therefore quite different from the genetic mechanisms used to synthesize Igs (Section 27.4) and TCRs (Section 27.7), in which each receptor interacts specifically with only a single antigen or antigenic determinant.
Check Your Understanding
Define polymorphism and polygeny as they apply to MHC genes.
How does a single MHC protein present many different peptides to T cells?
IV T Cells and Their Receptors
There are many different classes of T cells, and each class carries out specific functions. Using their T cell receptors, T cells of various types interact with antigens presented on MHCs to initiate and implement an adaptive immune response to pathogen invasion.
Adaptive immunity is ultimately initiated by the recognition of foreign peptides on infected cells by T lymphocytes. The infected cells that are first recognized by T cells may include the same phagocytes that participate in the innate immune response (Chapter 26). Antigen presentation activates naive (uncommitted) T lymphocytes to differentiate into effector T cells that carry out cell-mediated immunity. In the absence of antigen-activated T cells, there is little antigen-specific immunity and no immune memory.
27.7 T Cell Receptors: Proteins, Genes, and Diversity
27.7 T Cell Receptors: Proteins, Genes, and Diversity
27.7 T Cell Receptors: Proteins, Genes, and Diversity
APCs ingest bacteria, viruses, and other antigenic material by phagocytosis (in the case of macrophages and dendritic cells) or through internalization of a molecular antigen bound to a BCR. Phagosomal or cytoplasmic antigens are then digested, complexed with MHC proteins, and moved to the cell surface for antigen presentation to T cells. The TCRs of T cells can recognize (bind) antigens only when the peptides are complexed with MHC proteins on host cell surfaces. For example, a macrophage or dendritic cell that has encountered a virus will display MHC I and MHC II proteins embedded with viral peptides (Figure 27.17). These viral peptide–MHC complexes are the sites for T cell activation and interaction.
Figure 27.17 T cell–mediated immunity.
Antigen-presenting cells, such as the phagocytes in innate immunity, ingest, degrade, and process antigens. They then present antigens to T cells that secrete protein cytokines that activate the adaptive immune response. T-helper 1 (Th1) cells produce cytokines that activate other cells and induce inflammation. T-cytotoxic (Tc) cells produce perforin and granzyme, proteins that destroy nearby target cells.
Mastering Microbiology
Each T cell expresses a TCR that is specific for a single peptide–MHC complex. Antigen-specific T cells are found closely associated with APCs in the spleen, lymph nodes, and MALT. T cells constantly sample surrounding APCs for peptide–MHC complexes. Peptide–MHC complexes that interact with the TCR signal the T cell to grow and divide, producing antigen-reactive clones that coordinate cell-mediated killing, induce inflammation, and activate antibody-producing B cells. Here we examine how the TCR interacts with antigens presented on an APC or an infected target cell.
TCR Structure and Diversity
The TCR is a membrane-spanning protein that extends from the T cell surface into the extracellular environment. Similar to B cells with their BCRs, each T cell has thousands of copies of its specific TCR on its surface. A functional TCR consists of an α and a β polypeptide chain, and like Igs, each TCR chain has a variable (V) domain and a constant (C) domain (Figure 27.18). The Vα and Vβ domains interact cooperatively to form an antigen-binding site that contains complementarity-determining region (CDR1, CDR2, and CDR3) segments. However, unlike Igs, which can bind antigens in a native conformation, TCRs recognize only peptides bound to self MHC; other antigens, such as complex polysaccharides, are not recognized by TCRs.
Figure 27.18 The TCR:MHC I–peptide complex.

(a) A three-dimensional structure showing the orientation of TCR, peptide (brown), and MHC. This structure was derived from data deposited in the Protein Data Bank. (b) A diagram of the TCR:MHC–peptide structure. Note that the peptide is bound by both MHC and TCR proteins and has a distinct surface structure that interacts with each (a motif interacts with the MHC and an epitope with the TCR, as discussed in the text).
The three-dimensional structure of the TCR bound to MHC I–peptide is shown in Figure 27.18a. Both TCR and MHC proteins bind directly to peptide antigen. The MHC protein binds one face of the peptide—the MHC motif—and the TCR binds the other peptide face—the T cell epitope (Figure 27.18b). The CDR regions of the TCR bind directly to the MHC–peptide complex, and each CDR has a specific binding function. The CDR3 regions of the TCR α and β chains bind the antigen epitope; the CDR1 and CDR2 regions of the TCR α and β chains bind mainly to the MHC protein.
The adaptive immune response can generate TCR diversity sufficient to bind nearly every possible peptide antigen generated by APCs or target cells. T cells generate receptor diversity in ways similar to the generation of Ig diversity in B cells, and Table 27.3 compares the receptor diversity–generating mechanisms for each cell type. Analogous to the H and L chains of Igs, the TCR α and β chains are encoded by distinct constant- and variable-domain gene segments. TCR V-region gene segments are grouped together sequentially; those encoding the α chain include 80 V and 61 J segments, whereas those encoding the β chain include 52 V, 2 D, and 13 J gene segments (Figure 27.19). As we have seen for Igs (Section 27.4), antigen-binding diversity in TCRs is generated by somatic recombination, imprecise V-J (α-chain) or V-D-J (β-chain) joining, and random reassortment. Additional TCR diversity is generated because the D region of the β chain can be transcribed in all three reading frames (Section 6.9 and Figure 6.34), leading to production of three separate transcripts from each D-region gene segment.
Figure 27.19 Organization of the human TCR α- and β-chain genes.

The α-chain gene segments are located on chromosome 14, and the β-chain gene segments are on chromosome 6. Compare this figure with Figure 27.12.
Similar to the assembly of Ig H and L chains, individual α and β chains are produced by each T cell at random and assembled to form the α:β heterodimer. The somatic hypermutation mechanisms responsible for increased receptor diversity in Ig genes do not operate in T cells, and thus additional TCR diversity from these events is not possible. Despite this, potential TCR diversity is still enormous; an estimated 1015 different TCRs can be generated! It is important to note that not all TCR interactions with MHC–peptide result in activation of the T cell. As we now consider, many T cells that escape deletion in the thymus never receive the full set of activation signals necessary to become effector cells.
T Cell Activation and Anergy
Like B cells, T cells require a second signal for activation in addition to interactions with MHC–peptide. As we described in Section 27.1, T cells that react strongly with self antigens are deleted in the thymus to prevent autoimmunity. However, many self antigens are not expressed in the thymus, meaning that many T cell clones responsive to nonthymus self antigens avoid deletion. Yet, because of the requirement of a second signal for activation, these self-reactive T cells remain unresponsive and become anergic after they leave the thymus.
T cells that migrate from the thymus to secondary lymphoid organs and have not yet encountered specific antigen are naive and must be activated to become competent effector cells. For example, the first requirement for activation of a naive Tc cell is TCR binding of an MHC–peptide complex presented by an APC (**Figure 27.20*a***). The next step requires the interaction of two more proteins: B7, found on the APC, and CD28, expressed on the surface of the Tc cell. The binding of B7 to CD28 is the second signal necessary to activate the Tc cell and make it an effector cell; without the B7–CD28 interaction, the naive Tc cell is not activated (Figure 27.20). Once a T cell is activated, only the first signal (TCR binding to MHC–peptide) is required to induce an effector T cell. Thus, the activated (effector) Tc cell in Figure 27.20b can kill any target cell presenting the recognized peptide antigen on its MHC I, even those cells that do not display B7.
Figure 27.20 T cell activation.

(a) A naive Tc cell interacts through its TCR with an MHC–peptide presented by an APC, the first required activation signal. In addition, CD28 on the Tc cell interacts with B7 on the APC, the second signal required to activate the naive T cell. (b) The activated Tc cell can kill any target cell bearing the same MHC–peptide. (c) A naive Tc cell interacts through its TCR with an MHC–peptide presented by any cell. Conditions for the first signal are met, but the second signal is not generated because only APCs display the B7 protein. (d) In the absence of the second signal, the Tc cell becomes permanently unresponsive, or anergized.
The requirement for a second activation signal has major implications for establishing and maintaining clonal anergy. A naive Tc cell that interacts with a self antigen on a cell that is not an APC will receive only an MHC–peptide signal because non-APCs do not express B7. In the absence of the B7–CD28 interaction, a T cell that engages MHC–peptide is permanently anergized and can never be activated (Figure 27.20c and d), thus providing protection against harmful autoimmune reactions.
Structural Similarities of Antigen-Binding Proteins
As we have seen, antigen-binding proteins of the adaptive immune response have common structural features. Ig, TCR, and MHC protein complexes all consist of two nonidentical polypeptides: MHC and TCR proteins are composed of α and β polypeptide chains, and Igs have separate heavy and light chains (Section 27.4). Therefore, these protein complexes are believed to have arisen from duplication and selection of genes encoding primordial antigen receptors. Because of their shared structural, evolutionary, and functional features, genes encoding Ig, TCR, and MHC proteins are part of an extended gene family called the immunoglobulin gene superfamily, and a comparison of Ig superfamily proteins is shown in Figure 27.21.
Figure 27.21 Immunoglobulin gene superfamily proteins.

Constant (C) domains have homologous amino acid sequences and higher-order structures. The Ig-like C domains in each protein chain indicate evolutionary relationships that identify the proteins as members of the Ig gene superfamily. The variable (V) domains of Igs and TCRs are also Ig-like, but the peptide-binding domains of MHC class I and class II proteins (green) are not because their structures vary considerably from the basic features of the Ig domain.
Mastering Microbiology
Art Activity: Figure 27.21 Immunoglobulin gene superfamily proteins
The constant (C) domain of each protein in the superfamily has a highly conserved amino acid sequence consisting of about 100 amino acids with an intrachain disulfide bond spanning 50–70 amino acids. C domains provide structural integrity for the antigen-binding molecules, anchor the antigen-binding V domains to the cytoplasmic membrane, and give each protein its characteristic shape. C domains can also provide recognition sites for accessory molecules. For example, C domains of most IgG and all IgM proteins are bound by the C1q component of complement, a critical first step in initiating the classical complement activation sequence (Section 26.9). Likewise, MHC I C domains bind to the accessory CD8 protein on Tc cells, and homologous MHC II C domains bind CD4 on Th cells. As we have discussed, such interactions are critical steps for T cell activation and initiation of adaptive immunity (Section 27.5).
The variable (V) domains of TCR and Ig molecules are about the same length as the C domains, but the structures of V domains can vary considerably from one another and from C domains. As we have seen, Ig and TCR V domains interact specifically with a nearly limitless variety of antigens, whereas the V domains of MHC proteins, which have likely evolved independently of Ig and TCR V domains, interact with foreign peptides that share a common motif (Section 27.6).
Although all T cells produce and express antigen-binding TCRs, T cells are not all functionally equivalent. As we will see next, there are several different subsets of T cells, each with their own specific functions in the adaptive immune response.
Check Your Understanding
Identify diversity-generating mechanisms unique to TCRs as compared to diversity-generating mechanisms in Igs.
What does it mean when a T cell becomes “anergized,” and how does this process relate to the prevention of autoimmunity?
Describe and compare the structural features of Ig gene superfamily constant and variable domains.
27.8 T Cell Subsets and Their Functions
Antigen-reactive T cells consist of multiple T cell subsets having different functional properties. As we have discussed, T-cytotoxic (Tc) cells, also called cytotoxic T lymphocytes (CTLs), are CD8 T cells (Section 27.5) that recognize the peptide–MHC I complex on an infected cell. By contrast, T-helper (Th) cells, which can differentiate into several more specialized Th cell subsets, are CD4 T cells that interact with peptide–MHC II complexes on the surface of APCs and function to activate macrophages and stimulate antibody-mediated immunity.
T-Cytotoxic Cells
When an effector Tc cell (a CTL) recognizes a foreign peptide on an infected cell, it kills the peptide-bearing target cell. For example, a viral peptide embedded in MHC I, displayed on a virus-infected cell, marks the cell for interaction and killing by a CTL whose TCRs recognize the viral antigen. Contact between a CTL and the target cell is required to initiate killing of the infected cell (**Figure 27.22*a***). The point of initial contact is between the TCR and the peptide–MHC I complex. The CD8 protein on the CTL then binds the MHC I protein, strengthening the interaction and establishing an immunological synapse between the two cells. In addition to engaging and destroying virus-infected host cells, CTLs also have the ability to identify and kill cancer cells that present abnormal peptides (Figure 27.22b).
Figure 27.22 T-cytotoxic cells.

(a) When the TCR on a cytotoxic T lymphocyte (CTL) binds MHC I–peptide complexes on any cell, degranulation of the CTL releases molecules of perforin and granzyme, cytotoxins that perforate the target cell and cause apoptosis, respectively. (b) Three CTLs containing granules of perforin and granzyme (red) surround a cancer cell (blue, center). The CTL on the top left is in the process of flattening its membrane surface (green) against the cancer cell to form the immunological synapse.
Upon contact with the target cell, granules in the CTL migrate to the contact site, where the contents of the granules—cytolytic proteins called perforin and granzyme (Section 27.5)—are released into the synapse (Figure 27.22). Perforin molecules enter the membrane of the target cell and combine to form a transmembrane pore through which granzyme molecules then enter the target cell. Granzymes are cytotoxins that induce apoptosis (programmed cell death), characterized by organized killing and degradation of the target cell from within. The released perforin and granzyme molecules affect only the target cell because degranulation within the synapse is confined to the contact surface between the CTL and the target cell bearing peptide–MHC I. Cells lacking the peptide recognized by the CTL do not make contact and are not killed.
Different Classes of T-Helper Cells
Interactions with APCs drive naive CD4 Th cells to differentiate into one of several Th subsets, each producing unique combinations of cytokines that direct the immune response (Table 27.4). Dendritic cells and, to a lesser extent, macrophages (Section 26.4) play a central role as APCs in the activation of Th cells. A naive Th cell that recognizes peptide–MHC II through its TCR and also receives the costimulatory B7–CD28 second signal (Section 27.7) is further stimulated by the release of cytokines from the APC. This causes the Th cell to proliferate and differentiate into long-lived memory cells and effector Th cells that enter the circulation and migrate to sites of infection. The population of effector Th cells is comprised of different Th cell subsets, which may include Th1, Th2, and Th17 cells.
Table 27.4 T-helper cell subsets

Th1 cells produce IL-2, a cytokine that promotes growth and activation of other T cells, and also activate macrophages through the cytokines interferon gamma (IFN-γ), tumor necrosis factor alpha (TNF-α), and granulocyte–monocyte colony-stimulating factor (GM-CSF) (**Figure 27.23*a***). Th1-activated macrophages take up and kill foreign cells more efficiently than nonactivated (resting) macrophages (Figure 27.23b), and they may also kill tumor cells because activated macrophages recognize as nonself the tumor-specific antigens expressed by cancer cells.
Figure 27.23 Th1 cells and macrophage activation.

(a) Th1 cells are activated by antigens presented on macrophages in the context of MHC II protein. Activated Th1 cells produce cytokines that activate macrophages, leading to increased phagocyte activity and inflammation. (b) Activated macrophages are generally larger than resting macrophages and have a ruffled surface, often with cytoplasmic extensions that “feel” for pathogens. In addition to carrying out more aggressive phagocytosis, activated macrophages express genes that encode bactericidal enzymes found in lysosomes, such as proteases and enzymes that produce reactive oxygen species, all of which are designed to quickly kill ingested microorganisms inside the phagolysosome (Figure 26.11).
Mastering Microbiology
Art Activity: Figure 27.23 Th1 cells and macrophage activation
Activated macrophages also produce several cytokines and chemokines that function as proinflammatory mediators and chemoattractants, respectively. Table 27.5 summarizes the source and activities of these and other important immune cytokines and chemokines. In the case of organ or tissue transplants, Th1-activated macrophages can actually harm the host if the host’s Th1 cells recognize nonself MHC proteins on the transplant and trigger macrophage activation and transplant destruction (Section 27.6).
Table 27.5 Major immune cytokines and chemokines

aIL, interleukin; bIFN, interferon; cGM-CSF, granulocyte–monocyte colony-stimulating factor; dTNF, tumor necrosis factor; eMCP, macrophage chemoattractant protein.
Th2 cells play a pivotal role in B cell activation and antibody production. As we have discussed (Section 27.3), B cells make antibodies, and differentiated B cells are coated with antibodies (BCRs) that function as antigen receptors. When antigen binds the BCR (Figure 27.6), the antibody-bound antigen is taken into the B cell by endocytosis and degraded. Peptides from the degraded antigen are then loaded into the B cell’s MHC II protein for presentation to a Th2 cell. The Th2 cell responds by producing IL-4 and IL-5, cytokines that activate the B cell (Table 27.5) and cause it to proliferate and differentiate into plasma cells that produce and secrete antibodies specific to the presented antigen (Section 27.3).
Th17 cells are important in the first stages of the immune response. Naive Th cells differentiate into Th17 cells primarily through the activity of dendritic cells. When dendritic cells encounter pathogens, they present antigen and secrete IL-6 and transforming growth factor-β (TGF-β), cytokines that catalyze differentiation of naive Th cells to Th17 cells (**Figure 27.24*a***). Th17 cells then produce IL-17, a cytokine that activates other tissue cells to produce cytokines and chemokines that attract proinflammatory neutrophils to the site of infection (Table 27.5). Thus, the function of Th17 cells is to produce IL-17, starting a cascade that draws neutrophils to infection sites. By recruiting neutrophils, Th17 cells amplify innate immunity triggered by interaction of a pathogen with the dendritic cell.
Figure 27.24 Th17 and Treg cells.

(a) Th17 cells interact with pathogen-stimulated dendritic cells to draw neutrophils to the site of pathogen invasion, leading to inflammation and pathogen control. PRR, pattern recognition receptor. (b) Treg cells interact with nonactivated dendritic cells and respond by making immunosuppressive cytokines that control reactions to self antigens.
Mastering Microbiology
We conclude our consideration of T cells with a description of the T regulatory (Treg) cell, a Th cell subset that plays a critical role in controlling the immune response. Undifferentiated Th cells remain naive unless they are stimulated to mature and differentiate by certain cytokines, as is the case for IL-6 stimulation to produce Th17 cells. However, in the absence of a pathogen, Th cells can still interact with dendritic cells through MHC II–peptide–TCR (Figure 27.24b). In this case, the peptide is usually a self peptide, and an immune response to it could trigger autoimmunity. However, because the dendritic cells did not interact with a pathogen, they cannot produce IL-6 to promote Th17 differentiation. Instead, the absence of IL-6 pushes differentiation to Treg cells that make IL-10 and TGF-β, two cytokines that suppress immunity and inflammation. In the presence of self antigens and in the absence of IL-6, Treg cells shut down the immune response and inhibit inflammation. This is important for controlling immune responses to self antigen and preventing autoimmunity. In this regard, Treg cells play a key role in the maintenance of a healthy gut by interacting with short-chain fatty acids produced by the normal colonic microbiota to suppress host inflammatory responses against these “friendly” bacteria (Section 24.9 and Figure 24.23).
Check Your Understanding
Describe the mechanism used by Tc cells to recognize infected host cells.
Describe the effector system (the cell-killing mechanism) used by Tc cells.
Compare and contrast the roles and activities of the different Th cells. What cytokines do they produce, and which effector cells do these cytokines act upon?
Chapter Review
Go to Mastering Microbiology for videos, animations, practice tests, and more.
I Principles of Adaptive Immunity
27.1 The adaptive immune response is characterized by specificity for the antigen, the ability to respond more vigorously when reexposed to the same antigen (memory), and the acquired inability to interact with self antigens (tolerance). Tolerance in lymphocytes is acquired through a selection process. Immature T cells that do not interact with MHC–peptide (positive selection) or that react strongly with self antigens (negative selection) are eliminated by clonal deletion in the thymus. T cells that survive positive and negative selection leave the thymus and can participate in the immune response. Lymphocyte reactivity to self antigens is controlled through clonal deletion and anergy.
Q Why is it necessary that all three defining characteristics of the adaptive immune response (specificity, memory, and tolerance) be in place to maintain a functional and effective adaptive immune system?
27.2 Immunogens are foreign macromolecules that induce an immune response. Immunogens initiate an immune response when introduced into a host. Antigens are molecules recognized (bound) by antibodies or TCRs. Adaptive immunity develops naturally and actively through immune responses to infections, or naturally and passively through antibody transfer from the placenta or breast milk. Artificial passive immunity occurs when antibodies or immune cells are transferred from an immune individual to a nonimmune individual. Immunization induces artificial active immunity and is widely used to prevent infectious diseases.
Q What properties are required for a vaccine to induce an immune response? What type of immunity results from vaccination, and what role does a “booster” play in this process?
II Antibodies
27.3 Antibody production is initiated when an antigen contacts an antigen-specific B cell. Whereas B cell activation from T-independent antigens requires no T cell help, B cells that bind T-dependent antigens must process the antigen and present it to an antigen-specific Th cell. The Th cell becomes activated, producing cytokines that signal the B cell to clonally expand and differentiate to produce soluble, antigen-specific antibodies. Each antibody (immunoglobulin) protein consists of two heavy and two light chains. The antigen-binding site is formed by the interaction of the variable regions of one heavy and one light chain. Each antibody class has different structural characteristics, expression patterns, and functional roles. Clones of activated B and T cells can live for years as memory cells and can rapidly expand and differentiate to produce high titers of antibodies after reexposure to antigen.
Q Describe the structural and functional differences among the five major classes of antibodies. What cellular and molecular interactions occur to initiate production of antibodies?
27.4 The antigen-binding site of an immunoglobulin (Ig) is composed of the V (variable) domains of one heavy chain and one light chain. Each V region contains three complementarity-determining regions, or CDRs, that are folded together to form the antigen-binding site. Ig diversity is generated by several mechanisms. Somatic recombination of gene segments allows shuffling of the various Ig gene segments. Random reassortment of the heavy- and light-chain genes, imprecise joining of VDJ and VJ gene segments, and hypermutation mechanisms contribute to nearly unlimited Ig diversity.
Q Which Ig chains are used to construct a complete antigen-binding site? Which domains? Which CDRs? Calculate the total number of germ-line-encoded VH and VL domains that can be constructed from the available Ig genes.
III The Major Histocompatibility Complex (MHC)
27.5 T cells, with their TCRs, bind peptide antigens presented by MHC proteins on infected cells or APCs. Class I MHC proteins are expressed on all nucleated cells and present endogenous antigenic peptides to TCRs on Tc cells. Class II MHC proteins are expressed only on APCs. They function to present exogenously derived peptide antigens to TCRs on Th cells. These interactions activate T cells to kill antigen-bearing cells or to promote inflammation or antibody production.
Q Describe the basic structure of class I and class II major histocompatibility complex (MHC) proteins. In what functional ways do they differ?
27.6 Class I and class II MHC genes are highly polymorphic, and the many allelic variations challenge successful tissue transplantation. Different alleles of MHC class I and class II genes encode proteins that bind and present different peptide subsets, each characterized by a specific structural motif.
Q For the MHC class I polymorphisms, how many different MHC proteins are expressed in an individual? How many by the entire human population?
IV T Cells and Their Receptors
27.7 T cell receptors bind to peptide antigens presented by MHC proteins. The CDR3 regions of both the α chain and the β chain bind to the antigen epitope; the CDR1 and CDR2 regions bind to the MHC protein. VDJ gene segments encode the β-chain V domain of TCRs, and VJ gene segments encode the α-chain V domain. TCR diversity, generated by a variety of mechanisms, is nearly unlimited. The immunoglobulin gene superfamily encodes proteins that are evolutionarily, structurally, and functionally related to immunoglobulins. The antigen- binding Igs, TCRs, and MHC proteins are members of this superfamily.
Q What diversity-generating mechanisms function to produce the nearly unlimited variety of antigen-specific TCRs? What structural and functional features are common to proteins classified within the immunoglobulin gene superfamily?
27.8 T-cytotoxic (Tc) cells recognize antigens on virus-infected host cells and tumor cells through antigen-specific TCRs, and recognition triggers killing via perforin and granzyme. T-helper (Th) cells differentiate into several subsets. Through the action of cytokines, Th1 inflammatory cells activate macrophage effector cells, whereas Th2 cells activate B cells. Th17 cells are activated by pathogen-activated dendritic cells and secrete IL-17 to recruit neutrophils to the site of infection. Treg cells produce cytokines that suppress adaptive immunity.
Q What mechanism do Tc cells use to identify and destroy infected cells in the body? How do Th cells differ from Tc cells, and how do the different subsets of Th cells differ from each other?
Application Questions
Antibodies of the IgA class are probably more prevalent than those of the IgG class. Explain why this might be the case. What benefits might this have for the host?
Although genetic recombination events are important for generating significant diversity in the antigen-binding site of Igs, post-recombination somatic events may be even more important in achieving overall Ig diversity. Do you agree or disagree with this statement? Explain.
Polymorphism implies that each different MHC protein binds a different peptide motif. However, for the MHC class I proteins, only 6 peptide motifs can be recognized in an individual, whereas over 6000 motifs can be recognized by the entire human population. What advantage does recognition of multiple motifs have for the individual? What potential advantage does recognition of the extremely large number of motifs have for the population? Can everyone process and present the same antigens?
Chapter Glossary
a molecule capable of interacting with specific components of the immune system and that often functions as an immunogen to elicit an adaptive immune response B cell receptor (BCR)
a cell surface immunoglobulin that acts as an antigen receptor on a B cell CD4 coreceptor
a protein found on Th cells that interacts with MHC II on an antigen-presenting cell CD8 coreceptor
a protein found exclusively on Tc cells that interacts with MHC I on a target cell Clonal anergy
the inability to produce an immune response to specific antigens due to the neutralization of effector cells Clonal deletion
for T cell selection in the thymus or B cell selection in the bone marrow, the elimination by apoptosis of useless or self-reactive precursor lymphocytes Clonal expansion
following its activation, the proliferation and differentiation of a naive T or B lymphocyte into a clone of effector cells Clone
lymphocytes of identical antigen specificity that have arisen from a single, activated progenitor cell Complementarity-determining region (CDR)
a varying amino acid sequence within the variable domains of immunoglobulins or T cell receptors where contacts with antigen are made Epitope
the portion of an antigen that reacts with a specific antibody or T cell receptor Human leukocyte antigen (HLA)
an antigen-presenting protein encoded by a major histocompatibility complex gene in humans Immune memory (memory)
the ability to rapidly produce large quantities of specific immune cells or antibodies after subsequent exposure to a previously encountered antigen Immunogen
a molecule capable of eliciting an adaptive immune response Immunoglobulin gene superfamily
a family of genes that are evolutionarily, structurally, and functionally related to immunoglobulins Memory cell
a long-lived B or T lymphocyte responsive to a specific antigen MHC class I protein
an antigen-presenting molecule found on all nucleated vertebrate cells MHC class II protein
an antigen-presenting molecule found on macrophages, B cells, and dendritic cells Motif
in antigen presentation, a specific amino acid sequence found in all peptides that bind to a given MHC protein Negative selection
in T cell selection, the deletion of T cells that interact strongly with self antigens in the thymus (see also clonal deletion) Polygeny
the occurrence of two or more genetically, structurally, and functionally related gene loci due to an evolutionary gene duplication event Polymorphism
in a population, the occurrence of multiple alleles for a gene locus at a higher frequency than can be explained by recent random mutations Positive selection
in T cell selection, the growth and development of T cells that interact with self MHC–peptide in the thymus Primary immune response
the production of antibodies or immune T cells on first exposure to antigen; the antibodies are mostly of the IgM class Secondary immune response
the enhanced production of antibodies or immune T cells on second and subsequent exposures to antigen; the antibodies are mostly of the IgG class Somatic hypermutation
the mutation of immunoglobulin genes at rates higher than those observed in other genes Specificity
the ability of cells of the adaptive immune response to interact with particular antigens T cell receptor (TCR)
an antigen-specific receptor protein on the surface of T cells T-cytotoxic (Tc) cell
a lymphocyte that interacts with MHC I–peptide complexes through its T cell receptor and produces cytotoxins that kill the interacting target cell T-helper (Th) cell
a lymphocyte that interacts with MHC II–peptide complexes through its T cell receptor and produces cytokines that act on other cells. Th subsets include Th1 cells, which activate macrophages; Th2 cells, which activate B cells; Th17 cells, which activate neutrophils; and Treg cells, which suppress adaptive immunity Tolerance
the acquired inability to produce an immune response to particular antigens Vaccination (immunization)
the inoculation of a host with inactive or weakened pathogens or pathogen products to stimulate protective active immunity