28 Immune Disorders and Antimicrobial Therapy
## Chapter 28 Immune Disorders and Antimicrobial Therapy
I Disorders and Deficiencies of the Immune System
Preventing Autoimmunity with . . . Parasitic Worms?
Some immune disorders are a result of autoimmunity, a harmful condition in which an immune response is directed against “self” antigens. For example, type 1 diabetes (T1D) is an autoimmune disorder in which immune cells attack and destroy insulin-secreting β cells in the pancreas, resulting in an inability to metabolize sugars and control blood glucose levels. With no cure, type 1 diabetics must treat their condition by strict diet control and regular injection (or subcutaneous infusion) of insulin.
The prevalence of T1D has risen markedly in recent years; currently, nearly 80,000 children around the world are diagnosed annually. Interestingly, most of these cases occur in developed nations associated with elevated socioeconomic conditions and modern sanitation. The “hygiene hypothesis” has been offered as an explanation for the rising prevalence of autoimmune diseases. The hypothesis asserts that the overly sanitized, primarily indoor upbringing typical for those in developed nations precipitates the onset of allergies and autoimmunity due to reduced early exposure to parasites and other microbes that traditionally associate with humans and shape immune development.
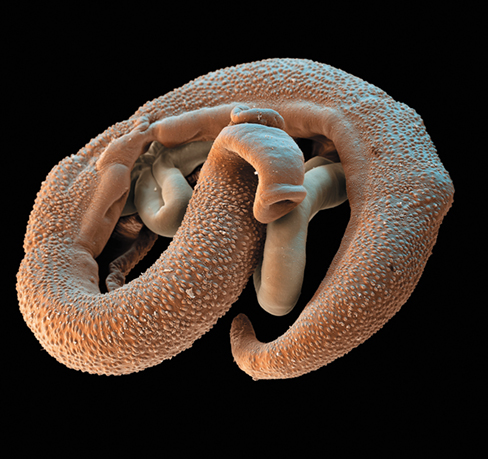
In this connection, researchers have reported that infection with parasitic helminths such as Schistosoma mansoni (photo), the causative agent of schistosomiasis, may improve and even prevent T1D. Recent studies have shown that Schistosoma infection can suppress Th1 cell activation and instead trigger the propagation of regulatory T (Treg) cells, thereby inhibiting inflammation and controlling the immune response. A similar helminth-mediated immune dampening has been demonstrated for other autoimmune disorders, such as the treatment of Crohn’s disease by introducing whipworm (Trichuris suis) infection.
These studies suggest that short-term parasitic worm infection (cured using antihelminthic drugs) may bring about long-term autoimmune protection. The next step is to determine what components of the worms actually cause the immunoregulatory effect so that it might become possible to administer only those purified products and avoid the intentional helminth infection.
Source: Tang, C.-l., et al. 2019. Helminths protect against type 1 diabetes: Effects and mechanisms. Parasitol. Res. 118: 1087. doi:10.1007/s00436-019-06247-4.
We discussed the key features of the innate and adaptive immune responses in Chapters 26 and 27, respectively, but of course the immune system does not always effectively halt microbial infection and disease progression. In fact, the immune response itself can be detrimental if self antigens are targeted. Such events cause immune disorders and immunodeficiency syndromes, which we explore in the first part of this chapter. We then discuss strategies to both prevent and treat diseases by either optimizing the immune response through vaccinations and immunotherapy or administering antimicrobial drugs to kill or inhibit established pathogens. We conclude the chapter with a discussion of the important implications of antimicrobial drug resistance.
Mastering Microbiology
Art Activity: Figure 27.24 Immediate hypersensitivity
I: Disorders and Deficiencies of the Immune System
I: Disorders and Deficiencies of the Immune System
I Disorders and Deficiencies of the Immune System
In some cases, the immune system fails to provide an adequate or beneficial response, and this may result in hypersensitivity, autoimmunity, superantigen-induced overstimulation, or immunodeficiency. All of these immune disorders are harmful, and some can even be fatal.
In some cases, reactions of the adaptive immune response can damage the host. For example, hypersensitivity is an immune response that results in host damage and, in some cases, even host death. Hypersensitivities are grouped according to the antigens and the mechanisms that produce disease. Likewise, exposure to superantigens, proteins produced by certain bacteria and viruses that initiate massive inflammatory responses, also causes severe immune reactions that result in host damage. We consider both of these along with immunodeficiency, a condition in which a host’s immune response is either absent or insufficient to effectively fight infections.
28.1 Allergy, Hypersensitivity, and Autoimmunity
There are two major types of hypersensitivity: antibody-mediated and cell-mediated. Antibody-mediated, or immediate hypersensitivity, is more commonly called allergy. Cell-mediated hypersensitivities also cause allergy-like syndromes but, because of the typically delayed onset of symptoms, are termed delayed-type hypersensitivity (DTH). As discussed in MicrobiologyNow on type 1 diabetes in the opening of this chapter, autoimmunity is a harmful immune reaction directed against self antigens. Hypersensitivities are categorized as type I, II, III, or IV based on immune effectors, antigens, and symptoms (Table 28.1), and we consider these first.
Table 28.1 Hypersensitivitya
aAutoimmune diseases may be caused by type II, type III, or type IV reactions.
Immediate Hypersensitivity
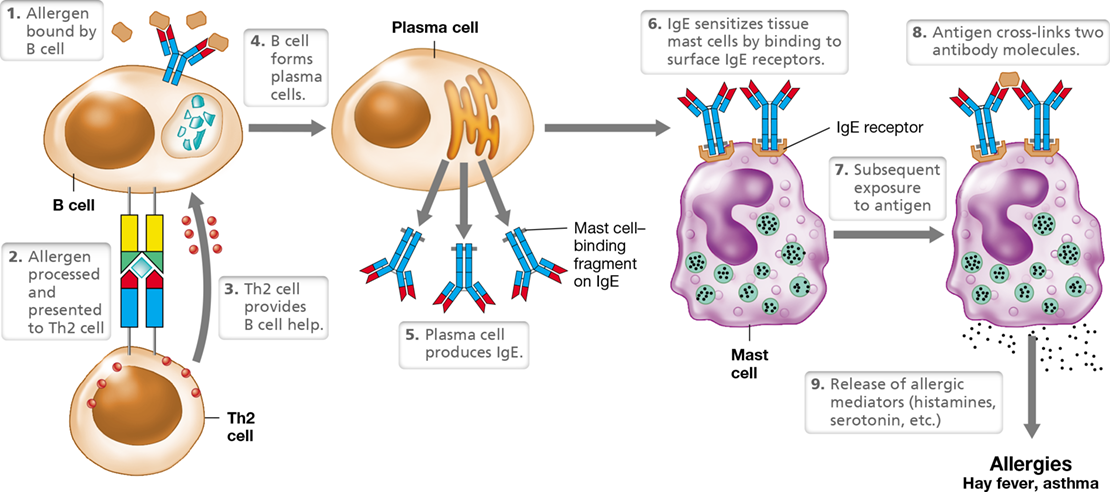
Immediate (type I) hypersensitivity occurs because mast cells coated with IgE release substances that either increase or decrease blood pressure or heart rate (vasoactive products) (Figure 28.1). Immediate hypersensitivity reactions occur within minutes after exposure to an allergen, the antigen that caused the type I hypersensitivity. Depending on the individual and the allergen, immediate hypersensitivity reactions can be mild allergic reactions or can cause a life-threatening reaction called anaphylaxis.
Figure 28.1 Immediate hypersensitivity.

Certain antigens, such as pollens, stimulate IgE production. IgE binds to mast cells by means of a high-affinity surface receptor and arms the mast cell. Antigen cross-links surface IgE, causing release of soluble mediators, including histamine. These mediators produce symptoms ranging from mild allergic symptoms to life-threatening anaphylaxis.
Nearly 25% of the population suffers from immediate hypersensitivity allergies to pollens, molds, animal dander, certain foods (strawberries, nuts, and shellfish), insect venoms, dust mites, and other agents. Most allergens enter the body at the surface of mucous membranes, especially those lining the lungs and the gut. Initial exposure to allergens stimulates Th2 cells to produce cytokines that induce B cells to make IgE antibodies. The allergen-specific IgE antibodies bind to IgE receptors on mast cells (Figure 28.1). Mast cells are nonmotile granulocytes (Section 26.4) associated with connective tissue adjacent to capillaries throughout the body. With any subsequent exposure to the immunizing allergen, the mast-cell-bound IgE molecules bind the antigen. Cross-linking of IgEs by an antigen triggers degranulation—the release of soluble allergic mediators from the mast cells. These mediators cause allergic symptoms within minutes of antigen exposure. After initial sensitization by an allergen, the allergic individual responds to each subsequent reexposure to the allergen.
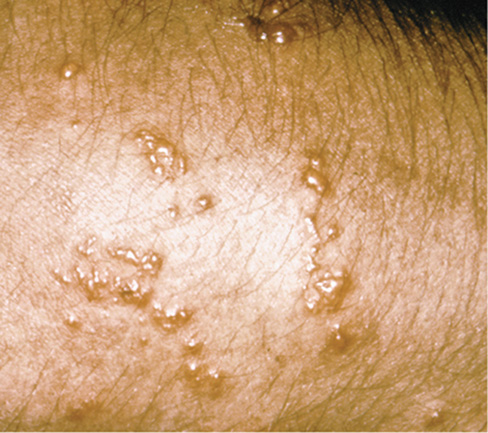
The principal chemical mediators released from mast cells are histamine and serotonin, modified amino acids that cause rapid dilation of blood vessels and contraction of smooth muscle, initiating symptoms ranging from mild local discomfort to systemic anaphylactic shock. Local symptoms typically include mucus production, rash, sneezing, itchiness, watery eyes, and hives (Figure 28.2). Symptoms of anaphylactic shock may include vasodilation (causing a sharp drop in blood pressure) and asthma due to smooth muscle constriction in the lungs.
Figure 28.2 Hives due to immediate hypersensitivity.

The raised, red areas are typical symptoms after contact with allergens that cause immediate hypersensitivity.
Severe anaphylaxis is treated immediately with the hormone epinephrine to counter smooth muscle contraction, promote breathing, and increase blood pressure. Less serious allergic symptoms may be treated with antihistamines, which are drugs that neutralize histamine, or with anti-inflammatory steroids. Finally, immunization with increasing doses of the allergen (beginning with a tiny amount) can be used to shift antibody production from IgE to IgG and IgA. The IgG and IgA then interact with the allergen during future exposures and thereby block antigen binding to IgE on sensitized mast cells. This process, called desensitization, inhibits IgE production and arrests allergic symptoms.
Delayed-Type Hypersensitivity
Delayed-type (type IV) hypersensitivity (DTH) is cell-mediated hypersensitivity characterized by tissue damage due to inflammation initiated by Th1 cells (Table 28.1). DTH symptoms appear several hours after secondary exposure to the eliciting antigen, with a maximal response usually occurring in 24 to 48 hours. Typical DTH antigens include chemicals that are not normally immunogens but become so when they covalently bind to skin proteins, creating new antigens and eliciting a DTH response. Hypersensitivity to these newly created antigens is known as contact dermatitis and results in, for example, skin reactions to poison ivy (Figure 28.3), jewelry, cosmetics, latex, and other chemicals that react with host tissues. Several hours after a second or subsequent exposure to the antigen, the skin feels itchy at the site of contact. Erythema (reddening) and edema (swelling) appear, often with localized tissue destruction in the form of blistering (Figure 28.3) and reach a maximum in several days. The delayed onset and the progress of the inflammatory response are the hallmarks of the DTH reaction. As discussed below, certain self antigens may also elicit DTH responses, resulting in autoimmune disease.
Figure 28.3 Delayed-type hypersensitivity.
Poison ivy blisters on an arm. The raised rash appears 24–48 hours after exposure to plants of the genus Rhus as a result of macrophage activation by Th1 cells sensitized to Rhus antigens.
Another example of delayed-type hypersensitivity is the development of protective immunity to the bacterium that causes tuberculosis, Mycobacterium tuberculosis (Section 31.4). The German physician-turned-microbiologist Robert Koch discovered this cellular immune response during his classic studies on tuberculosis over a century ago (Section 1.12). When antigens derived from the bacterium were injected subcutaneously into a person previously infected with M. tuberculosis, Koch observed that a skin reaction called the tuberculin reaction developed within 24–48 hours (Figure 28.4). We now know that during this DTH, local Th1 cells stimulated by the introduced M. tuberculosis antigens release cytokines that attract and activate large numbers of macrophages, which in turn produce a Characteristic local inflammation, including induration (hardening), edema, erythema, pain, and heating of the skin. The activated macrophages then ingest and destroy the invading antigen. Today, the DTH-based tuberculin skin test is used to test for a current or previous infection with M. tuberculosis or previous immunization with the tuberculosis vaccine (Section 29.5).
Figure 28.4 The tuberculin test and macrophage activation.

This tuberculin test shows a positive reaction. Macrophages activated by antigen-specific Th1 cells cause the localized, delayed-type reaction to a tuberculosis antigen, tuberculin, at the site of injection. The raised area of inflammation on the forearm is about 1.5 cm in diameter.
A number of other infectious diseases due to intracellular pathogens elicit DTH reactions. These include bacterial diseases such as leprosy, brucellosis, and psittacosis; viral diseases such as mumps; and fungal diseases such as coccidioidomycosis, histoplasmosis, and blastomycosis (many of these are covered in Chapters 31, 32, 33 and 34). Visible, antigen-specific skin responses similar to the tuberculin reaction occur after injection of antigens derived from the pathogens, indicating pathogen exposure and Th1-mediated immunity.
Autoimmune Conditions
As lymphocytes develop, T and B cells that react with self antigens are normally eliminated (Section 27.1). Autoimmune diseases result when these cells are instead activated to produce immune reactions against self proteins (Table 28.2). For example, Th1-mediated DTH can cause autoimmune responses directed against self antigens, as in the case of the Th1-mediated response to brain-derived antigens in allergic encephalitis. In type 1 diabetes mellitus (see MicrobiologyNow in the chapter opener), Th1 cells mount an immune response directed to antigens on insulin-producing β cells in the pancreas. The destruction of these cells leads to insulin insufficiency and an inability to control blood sugar levels without an insulin supplement.
Table 28.2 Some autoimmune diseases of humans

In addition to these examples, some autoimmune diseases are caused by autoantibodies, antibodies that bind self antigens, many of which are organ-specific. For example, in Hashimoto’s disease (hypothyroidism), autoantibodies are made against thyroglobulin, a protein product of the thyroid gland that assists in the synthesis of thyroid hormones. In this disease, antibodies to thyroglobulin bind complement proteins (Section 26.9), leading to local inflammation and the destruction of host cells, the hallmarks of a type II hypersensitivity disease (Table 28.2).
Systemic lupus erythematosus (SLE) is an autoimmune disease caused by a type III hypersensitivity. This disease and others like it are caused by autoantibodies directed against soluble, circulating self antigens. In SLE, the antigens include nucleoproteins and even DNA itself. Autoantibodies that bind to soluble proteins produce insoluble immune complexes, and disease symptoms result when these circulating antigen–antibody complexes deposit in different body tissues, such as the kidney, lungs, and spleen. In the tissues, the antibodies bind complement, resulting in inflammation and local, often severe, tissue damage. Type III hypersensitivities are also called immune complex disorders (Table 28.1).
Treating Autoimmunity
Organ-specific autoimmune diseases are sometimes more easily treated than diseases that affect multiple organs. For example, the product of organ function, such as thyroxine in autoimmune hypothyroidism or insulin in type 1 diabetes, can often be supplied in pure form from another source, such as genetically modified bacteria (Section 12.6 and Table 12.1). SLE, rheumatoid arthritis, and other autoimmune diseases that affect multiple organs and sites can often be controlled only by general immunosuppressive therapy, such as the use of steroid drugs. Unfortunately, the immunosuppression associated with these treatments significantly increases the chance of developing infections from opportunistic pathogens.
As an alternative to traditional immunosuppressive treatments, monoclonal antibodies (a single antibody type, Section 29.5) have emerged as a promising therapeutic strategy to combat autoimmune diseases. For example, adalimumab (marketed as the drug Humira®) is a monoclonal antibody that neutralizes tumor necrosis factor alpha (TNF-α), an inflammatory cytokine linked to several autoimmune diseases including rheumatoid arthritis, Crohn’s disease, and psoriasis. Similarly, the monoclonal antibody belimumab (marketed as the drug Benlysta®) targets B-cell activating factor, a cytokine that stimulates B cell maturation but is overexpressed in SLE patients, causing the persistence of B cells that produce autoantibodies. Other immunotherapeutic strategies to control autoimmunity focus on limiting the proliferation of proinflammatory Th17 cells by stimulating the production of regulatory T (Treg) cells; the latter function as “brakes” of the immune system to limit harmful inflammatory responses (Section 27.8). We explore immunotherapeutic treatments further in Section 28.4.
Heredity often influences the incidence, type, and severity of autoimmune diseases. Many autoimmune diseases correlate strongly with the presence of certain major histocompatibility proteins (Section 27.6). Studies of autoimmune diseases in mice support a genetic link, but the precise conditions necessary for developing autoimmunity may also depend on other factors, such as prior infections, gender, age, and health status. Women, for example, are about ten times more likely to develop SLE than are men. It is thus likely that the balance between a normal immune response and autoimmunity is tipped by a combination of factors in which genetic predisposition plays a central role.
Check Your Understanding
Discriminate between immediate hypersensitivity and delayed-type hypersensitivity with respect to antigens and immune effectors.
Provide examples and mechanisms for an antibody-mediated autoimmune disease directed against a specific organ and one that produces circulating immune complexes.
28.2 Superantigens and Immunodeficiency
Extremes in the adaptive immune response, whether overactive or deficient, can have devastating effects on the host. The exotoxins (Sections 25.6 and 25.7) called superantigens damage host cells indirectly by subverting the immune system such that T cells and their cytokine products destroy host tissues through an exaggerated immune response. By contrast, some diseases—either genetic or infectious—cause immunodeficiency, resulting in increased susceptibility of the host to infectious diseases. We now consider these opposing extremes in the adaptive immune response.
Superantigens
Superantigens are proteins that, upon exposure to the immune system, activate many more T cells than normal and are therefore capable of eliciting an unusually strong immune response. A variety of viruses and bacteria produce superantigens. For example, streptococci and staphylococci, especially certain strains of Streptococcus pyogenes and Staphylococcus aureus, produce several different extremely potent superantigens in the condition known as toxic shock syndrome (Figure 28.5)(Sections 31.2 and 31.9).
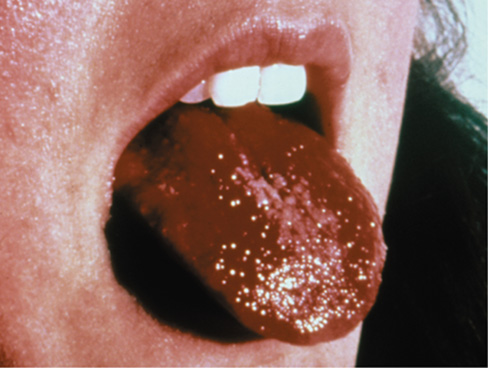
Figure 28.5 Toxic shock syndrome.

This individual has a swollen and bumpy tongue called “strawberry tongue,” a symptom of toxic shock syndrome caused by a Staphylococcus aureus superantigen.
Superantigen interaction with TCRs differs from conventional antigen–TCR binding. Ordinarily, foreign antigens presented by MHC proteins, such as those on the surface of an antigen-presenting cell, bind to a TCR at a defined antigen-binding site (Figure 27.18). By contrast, superantigens bind to sites on TCR and MHC proteins that are outside the antigen-specific binding site (Figure 28.6). A superantigen binds to all TCRs that share a common structure, and many different TCRs share the same structure outside the antigen-binding site. Whereas less than 0.01% of all available T cells interact with a conventional foreign antigen in a typical immune response, some superantigens can bind up to 25% of all T cells in the body. These interactions mimic conventional antigen presentation and activate large numbers of T cells to grow and divide. As in normal immune responses, the activated T cells produce cytokines that stimulate phagocytes and other immune cells. However, the unusually large cytokine production by the enhanced population of superantigen-activated T cells triggers a widespread cell-mediated response characterized by systemic inflammatory reactions. The resulting fever, diarrhea, vomiting, mucus production, and even systemic shock may be fatal in extreme cases. The clinical symptoms of superantigen shock are indistinguishable from those of septic shock, a condition in which a bacterial infection has spread throughout the body (Section 26.8).
Figure 28.6 Superantigens.


Superantigens bind to conserved regions of both the MHC and TCR proteins at positions outside the normal binding site. Superantigens interact with large numbers of T cells, causing large-scale T cell activation, cytokine release, and systemic inflammation.
One of the most common superantigen diseases is Staphylococcus aureus food poisoning, characterized by fever, vomiting, and diarrhea, and caused by one of several superantigen staphylococcal enterotoxins (Section 33.8). S. aureus also produces the superantigen responsible for toxic shock syndrome (Figure 28.5), and Streptococcus pyogenes produces erythrogenic toxin, the superantigen responsible for scarlet fever (Section 31.2).
Immunodeficiency
Active adaptive immunity is critical for infectious disease resistance. We know this because of problems caused by genetic defects and diseases that affect the adaptive immune system. For example, animals that cannot produce antibodies because of genetic defects in their B cells acquire serious infections from extracellular pathogens, especially bacteria. In addition, animals with genetic defects that prevent development of T cells suffer from recurrent infections with viruses and other intracellular pathogens.
Severe combined immune deficiency syndrome (SCID) is a genetic disorder that prevents proper formation of B and T cells. Individuals with SCID essentially have no effective adaptive immunity. A lack of proper T cell function results in a deficiency of cell-mediated immunity and indirectly causes a loss of antibody-mediated immunity because B cell activation against most antigens requires functional Th cells. Unless SCID patients receive supportive therapy, such as a bone marrow transplant and antibiotic treatments, SCID eventually causes death from multiple recurrent infections. The transplantation of compatible bone marrow tissue provides the afflicted individual with hematopoietic stem cells (Sections 26.3 and 26.4) free of the genetic defects that cause SCID.
Gene therapy has also shown promise as a curative treatment option for some forms of SCID. In this procedure, defective genes in hematopoietic stem cells are replaced through a transduction process using viral vectors (Section 9.7). As with bone marrow transplantation, successful gene therapy restores function of the adaptive immune system. However, using viral vectors—even genetically engineered vectors—can be risky because of secondary effects that the viruses may have, such as activating host genes that initiate tumor growth. Because of this, powerful new tools for genome editing (Section 12.13) that do not require viral vectors are being developed, and thus safe genetic therapies for SCID may be on the horizon.
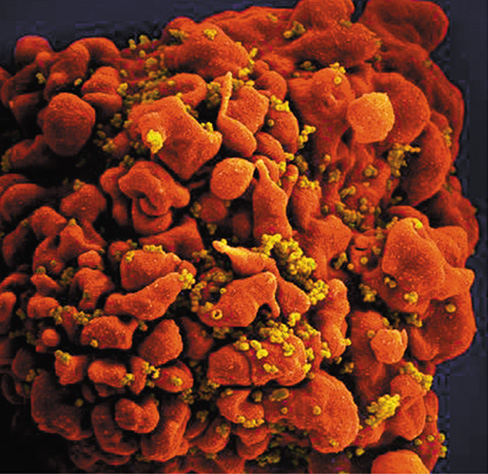
In some cases, immunodeficiency is not the result of a genetic disorder but rather is caused by microbial infection. The best-studied example of this is the loss of the adaptive immune response due to acquired immunodeficiency syndrome (AIDS). Human immunodeficiency virus (HIV) infects host cells that express the CD4 cell surface protein. The virus initially infects macrophages but later attacks and replicates primarily in Th cells (Figure 28.7). Once infected, Th cells cease dividing and eventually die. Therefore, in untreated cases, HIV infection causes a gradual depletion of Th cells, resulting in a lack of effective immunity and the eventual onset of AIDS (Section 31.15 and Figure 31.45).
Figure 28.7 HIV infection of a Th cell.

Colorized scanning electron micrograph of HIV virions (yellow) attacking a Th cell (red).
In most AIDS patients, the actual cause of death is not directly attributed to HIV but rather to secondary microbial infections caused by opportunistic pathogens (Section 25.4). Many of these infections are caused by fungal, bacterial, and viral pathogens that only rarely cause serious disease symptoms in individuals that have a fully functional immune system. The deficiency of adaptive immunity in AIDS patients allows these opportunistic pathogens to colonize and invade body tissues, eventually leading to death. We discuss the pathogenicity, symptoms, treatment, and other aspects of HIV/AIDS in more detail in Chapter 31.
Check Your Understanding
Describe the binding site for superantigens on T cells and antigen-presenting cells. How does this correlate to the activation of a much larger population of T cells than normal upon exposure to superantigens?
Compare and contrast the immunodeficiency observed in SCID patients to that of AIDS patients. What cell types are affected by each condition?
In the absence of treatment, what is the prognosis for individuals afflicted with SCID and AIDS?
II Vaccines and Immunotherapy
Vaccines are indispensable preventive tools to train the immune system to recognize and attack specific pathogens upon exposure. Immunotherapy is a treatment option that uses therapeutic agents to either activate or suppress immune responses to specific diseases, including cancer.
As we have seen, malfunctions of the immune system can either contribute to disease signs and symptoms (hypersensitivity and autoimmunity) or result in a failure to contain them (immunodeficiency). Here we move on to discuss ways in which attributes of the immune system can be harnessed to develop tools and strategies to both prevent and treat diseases.
28.3 Vaccination Against Infectious Diseases
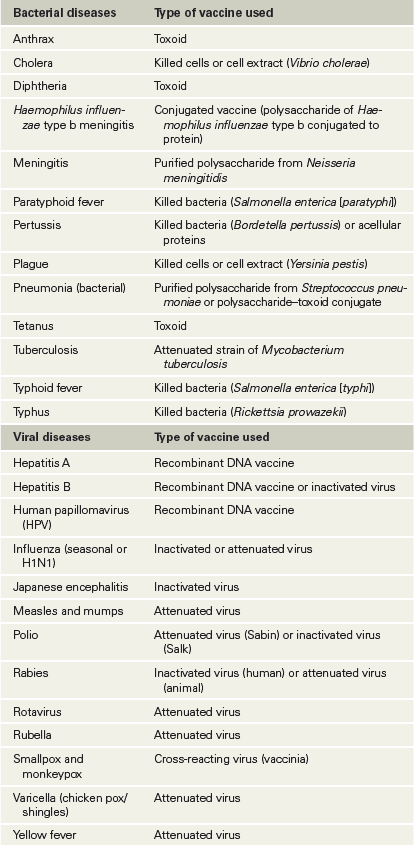
A major means of disease prevention is vaccination (immunization), in which deliberate exposure to an antigen triggers an adaptive immune response intended to protect an individual against future infection by a pathogen. The immunogen used to induce this artificial active immunity is called a vaccine. A summary of major diseases for which vaccines are available for human use is given in Table 28.3.
Table 28.3 Vaccines for infectious diseases in humans

The Nature of Vaccines
Infants are immune to many common infectious diseases during the first several months of life because they acquire natural passive immunity from maternal antibodies transferred across the placenta and in breast milk (Section 24.8). However, it is recommended that infants be immunized against key infectious diseases as soon as possible so that their own active immunity can replace the temporary maternal passive immunity (Figure 28.8). As discussed in Section 27.3, a single exposure to antigen does not lead to a high antibody titer. After an initial immunization, a series of “booster” immunizations are given to produce a secondary response and a high antibody titer.
Figure 28.8 Immunization recommendations for infants and children in the United States.

The U.S. Centers for Disease Control and Prevention website (http://www.cdc.gov) has recommendations for timing and dose of immunizations for all age groups and for special populations, such as international travelers, women of childbearing age, and those with immunodeficiency or chronic disease.
It is well established that the introduction of effective vaccines into a population has reduced the incidence of formerly epidemic childhood diseases such as measles and mumps (Section 31.6) and has eliminated smallpox altogether (Section 30.5). However, lifelong immunity is rarely achieved by vaccination because the immune cells and antibodies generated by immunization gradually disappear from the body. In the absence of environmental antigenic stimulation, the length of effective immunity varies considerably with different vaccines. For example, the tetanus toxoid vaccine provides effective immunity for ten years, but immunity induced by inactivated influenza virus vaccine generally disappears within a year or two (the reasons for this short-term immunity in the case of influenza are considered in Section 31.8).
To minimize the risk of infection or other adverse reactions, pathogens or pathogen products used in vaccines must be made harmless. To achieve this, many vaccines consist of pathogens killed either by heat treatment or by chemical agents. For example, formaldehyde is used to inactivate polio virions for preparation of the Salk polio vaccine. In addition, many exotoxins can be chemically modified to create a toxoid, a molecule that retains its antigenicity but is no longer toxic. Toxoid vaccines, such as the vaccine for Clostridium tetani (the causative agent of tetanus, Section 32.9), safely induce long-term protective immunity against the exotoxin.
Immunization with intact pathogens is usually more effective than immunization with dead or inactivated material. It is often possible to isolate attenuated strains of pathogens—those that have lost their virulence but still retain the immunizing antigens (Figure 25.10). However, because attenuated strains of pathogens are still viable, some individuals may inadvertently acquire active disease from the vaccination. Nevertheless, attenuated vaccines are widely used because they tend to provide long-lasting immunity and a strong secondary booster response. By contrast, killed virus vaccines generally provide short-lived immune responses with less long-term memory.
Most bacterial vaccines are composed of key cellular antigens in an inactivated form, such as the toxoids that protect against tetanus and diphtheria. Inactivated bacterial vaccines induce antibody-mediated protection without exposing recipients to the risk of infection. However, variability in primary and secondary responses with each vaccine and individual make periodic reimmunization necessary to establish and maintain immunity.
Synthetic and Genetically Engineered Vaccines
An alternative approach to vaccine development is to deploy the tools of genetic engineering (Chapter 12) to produce synthetic peptides. To make such a vaccine, a genetic engineer can synthesize a peptide that corresponds to an antigen from an infectious agent. For example, the toxin from the foot-and-mouth disease virus, an important animal pathogen, must be modified from its native form to render it harmless for use as a vaccine. The toxin contains a peptide of 20 amino acids that is an important antigenic determinant in the protein, but the peptide is too small to be an effective vaccine by itself. However, genetic engineers attached the small peptide to a larger, innocuous protein that acts as a carrier molecule, creating a conjugate vaccine against foot-and-mouth disease virus. This strategy has great promise for creating vaccines for a number of other pathogens as well, especially because the complete genomic sequences of many pathogens are now known, and this provides the insight necessary to pinpoint the most likely antigenic molecules.
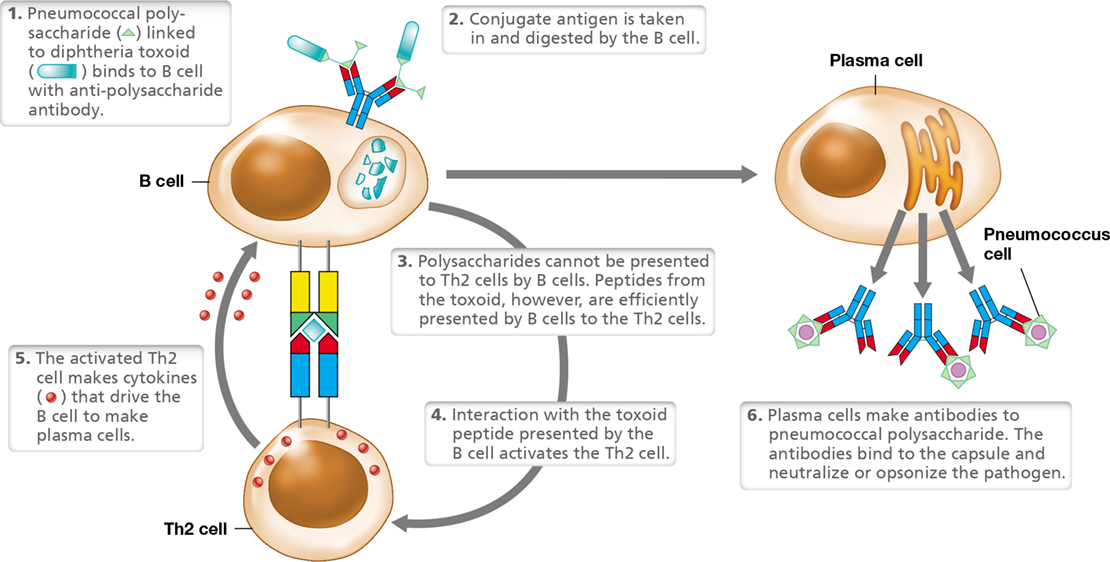
Two widely available conjugate vaccines couple extracted bacterial polysaccharides, which are poorly immunogenic by themselves, to a protein toxoid, thereby inducing a more robust immune response with better immune memory than injection of the polysaccharide antigen alone. The first of these is a pneumococcal vaccine that pairs diphtheria toxoid to polysaccharide capsule antigens derived from pathogenic strains of Streptococcus pneumoniae, the causative agent of bacterial pneumonia (Section 31.2) (Figure 28.9). The second is the Haemophilus influenzae type b (Hib) vaccine, which couples Hib polysaccharide to tetanus toxoid. The conjugation of polysaccharide antigens to protein toxoids efficiently activates Th2 cells, resulting in a strong immune response that confers immune memory.
Figure 28.9 Conjugate vaccines.

Conjugate vaccines, such as Streptococcus pneumoniae (pneumococcus) polysaccharide covalently linked to diphtheria toxoid (shown) or Haemophilus influenzae type B (Hib) polysaccharide coupled to tetanus toxoid provide effective immunity to polysaccharide antigens, which are poor immunogens in the absence of protein carriers.
Genomic information is particularly useful for making viral vaccines. Genes that encode antigens from virtually any virus can be cloned into the vaccinia virus genome and expressed (Section 12.8). Inoculation with the antigen-producing vaccinia virus can induce immunity to the product of the cloned gene. Such a preparation is called a recombinant-vector vaccine, an example of which is the recombinant vaccinia–rabies vaccine used in animals. A second immunization strategy uses proteins made from cloned DNA as immunogens. After a pathogen gene is cloned and expressed in a suitable microbial host, the pathogen protein is harvested and used to produce a recombinant-antigen vaccine. For example, the current vaccine for hepatitis B virus (Section 31.11) is a major hepatitis B surface protein antigen (HbsAg) expressed in genetically modified yeast cells. Similarly, a vaccine effective against human papillomavirus (HPV, Section 31.14) is also a recombinant-antigen vaccine made in yeast cells.
Nucleic Acid Vaccines and Plant-Based Vaccines
A novel vaccine delivery method is seen with the DNA vaccines, vaccines in which the antigen(s) of interest are not injected into the animal but are actually synthesized by the animal. DNA vaccines are bacterial plasmids that contain cloned DNA that encodes the antigen of interest. Typically, the vaccine is injected intramuscularly into a host animal. Once host cells take up the plasmid, the DNA is transcribed and translated to produce immunogenic proteins, triggering a conventional immune response of activated T cells and antibodies directed to the protein encoded by the cloned DNA. A major advantage of DNA vaccines is that, because only a single pathogen gene is cloned into the plasmid and administered to the host, there is no chance of an infection, as there might be with an attenuated vaccine.
A second nucleic acid–based vaccine methodology that has shown promise as an alternative to conventional methods is mRNA vaccines. In this approach, RNA that encodes an antigen of interest and has been engineered to resemble a fully processed, mature mRNA suitable for translation in eukaryotic cells (Section 6.6) is administered to the animal, and production of the protein by host cells produces an adaptive immune response to the antigen. As with DNA vaccines, there is no risk of infection from mRNA vaccines, but in addition, the short half-life of mRNA allows for greater control of protein expression and immunogenicity. Early challenges associated with instability and inefficient in vivo delivery of mRNA vaccines have largely been overcome, and therefore this vaccine platform may be of great importance for future disease control and prevention.
Lastly, a novel variation of the recombinant-antigen vaccine described above uses fast-growing plants as the transgenic host to produce plant-based vaccines. In this method, the pathogen-derived target gene is transferred to the plant using the plant pathogen Agrobacterium tumefaciens as a bacterial vector (Sections 12.7 and 23.6). Following plant infection by A. tumefaciens, recombinant proteins are extracted from the plants and purified to produce virus-like particles (VLPs), lipid bilayer constructs embedded with pathogen-specific protein antigens. VLPs produce a strong immune response, but because they lack the genetic material of the native pathogen, they cannot replicate or cause infection. Promising VLP vaccines are in development for a wide variety of viral pathogens, including rotavirus, norovirus, West Nile virus, hepatitis B virus, and pandemic influenza.
Check Your Understanding
Compare and contrast live attenuated vaccines, inactivated vaccines, and toxoids. Which of these has the greatest potential to cause active disease in the recipient? Which typically provides the longest-lasting immunity?
Identify the advantages of alternative immunization strategies as compared to traditional immunization procedures.
28.4 Immunotherapy
As we have seen, the immune system can be manipulated to provide protection against future pathogen exposures through vaccination strategies. In addition to this, immunotherapy is a burgeoning field in clinical medicine that seeks to harness cells and other components of the immune system to fight or prevent diseases, including various forms of cancer. Diagnostic tests for cancer typically begin with either a tissue biopsy for solid tumors or a blood sample for hematological cancers or circulating cancer cells. Key immunotherapeutic approaches currently in development to combat cancer include anticancer vaccination, the use of checkpoint inhibitors to stimulate the antitumor immune response, and adoptive T cell transfer, an encouraging approach that optimizes T cell activity against a variety of cancers (Figure 28.10).
Figure 28.10 Overview of cancer immunotherapy.

Anticancer vaccines can potentially be designed from properties of extracted tumor cells or the patient’s dendritic cells. Checkpoint blockade uses monoclonal antibodies to neutralize immunosuppressive components, such as PD-1/PD-L1 and CTLA-4, in the tumor microenvironment. CARs are genetically engineered receptors that reprogram cytotoxic T cells to recognize and be directly activated by tumor-specific antigens. CAR, chimeric antigen receptor; mAb, monoclonal antibody; TIL, tumor-infiltrating lymphocyte.
Anticancer Vaccines
Attempts to develop anticancer vaccines have historically been ineffective in practice, but due to recent advances in molecular immunology, interest is once again growing in this area. Although a universal vaccine to protect against all cancer is unlikely to be realized considering the many different types of cancer that exist, immunotherapeutic vaccines are currently in development for certain cancers, including melanoma, pancreatic cancer, and glioblastoma, an aggressive brain cancer with an otherwise poor prognosis.
Anticancer vaccines can be either prophylactic (preventive) or therapeutic. Rather than preventing cancer directly, prophylactic vaccines target oncogenic pathogens, such as human papillomavirus, the causative agent of most cervical and anal cancers, and hepatitis B virus, which can cause liver cancer. Therapeutic anticancer vaccines are designed to stimulate the body’s natural immune response to cancer cells. Advances in genomics, including digital innovations that allow for detailed data analyses, have led to personalized cancer vaccines. By mapping specific oncogenic mutations in extracted tumor cells, a patient’s dendritic cells can be sensitized to new antigens (called “neoantigens”) that appear on the transformed tumor cells, resulting in a potent anticancer response mediated by activated T cells (Figure 28.10).
Checkpoint Inhibitors
The tumor microenvironment has been shown to be immunosuppressive, which is an obstacle to cancer treatment. This is largely due to the production and activity of immune checkpoint proteins, including programmed cell death protein-1 (PD-1) and cytotoxic T-lymphocyte-associated protein-4 (CTLA-4), which function to suppress the adaptive immune response. PD-1 and CTLA-4 inhibit T cell activation by regulating the TCR costimulatory protein CD28 and competing with CD28 for protein B7 on APCs, respectively (Figure 27.20). Normally, these functions prevent uncontrolled inflammation and autoimmune responses, but cancer cells exploit these checkpoint proteins to suppress immunity and evade anticancer effector cells.
The goal of checkpoint inhibitor therapy (also called checkpoint blockade), therefore, is to prevent neutralization of cytotoxic T cells and stimulate their tumoricidal effector activity. The most successful checkpoint inhibitors are monoclonal antibody (mAb)-based drugs (Section 29.5), such as pembrolizumab (marketed as Keytruda®), that target PD-1 on T cells (or its ligand PD-L1 on cancer cells) in solid tumors and block inhibition of the antitumor immune response (Figure 28.10). Checkpoint inhibitors are often combined with steroids or other immunity-modulating drugs to boost their efficiency, but the safety and efficacy of these combined treatment strategies are still being assessed in clinical trials. Recall from Section 28.1 that mAbs are also used to treat autoimmune disorders, and therefore it is important to note that, depending on their activity, these drugs may be used to either suppress (to combat autoimmunity) or stimulate (to treat cancer) the immune response.
Adoptive T Cell Transfer
Adoptive T cell transfer (ACT) is a novel type of cancer immunotherapy that focuses on harnessing the tumor-targeting abilities of T cells. Two main approaches have been developed for ACT therapy: tumor-infiltrating lymphocytes (TILs) and chimeric antigen receptor (CAR) T cells (Figure 28.10).
TILs are effector T cells having natural anticancer activity that are extracted from tumors, propagated in the laboratory and then infused back into the patient to amplify the immune response against the cancer cells. TIL therapy has proven especially effective in treating melanoma—a dangerous and often metastasizing (spreading) form of cancer—and clinical trials using TILs against this and other forms of cancer, including ovarian, renal cell, and nasopharyngeal cancers, are under way.
CAR T cell therapy is a form of ACT that employs genetically engineered T cells to treat various cancers, especially B cell malignancies such as leukemia and lymphoma. CARs are engineered receptors that are an amalgam of components from TCRs, costimulatory molecules such as CD28, and the antigen-binding domains of antibodies (Figure 27.11) specific to tumor neoantigens (Figure 28.10). The synthesis of CAR T cells is complex and, like TIL therapy, requires extraction of the patient’s T cells for laboratory manipulation. The manipulation includes introducing a CAR-containing viral vector and clonal expansion of the transgenic T cells before infusion back into the patient. Essentially, CAR T cells are customized to recognize tumor-specific antigens and kill cancer cells even in the absence of MHC antigen presentation(Section 27.5). The latter is possible because the antigen-binding domain of CARs is of immunoglobulin origin rather than TCR origin.
Although treatments such as checkpoint inhibitors and CAR T cells are undeniably powerful immunotherapeutic tools, these technologies are also accompanied by several unresolved challenges, including the requirement to overcome immunosuppressive tumor microenvironments and side effects such as neurologic toxicity and excessive proinflammatory cytokine release. In addition, reactions to the same therapies can vary widely in different individuals, a response that could be the result of person-to-person differences in our gut microbiota (Chapter 24), a consideration we turn to now (Figure 28.11).
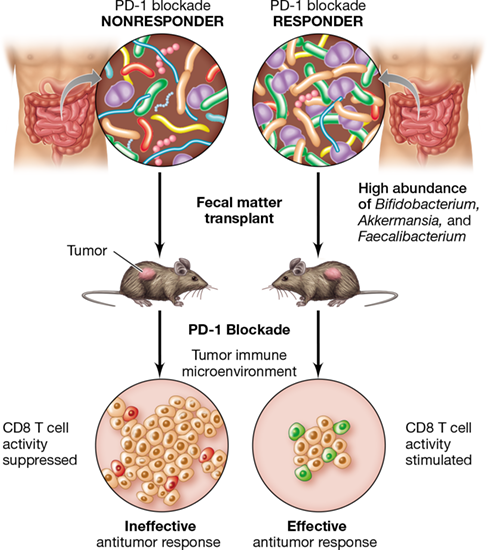
Figure 28.11 Immunotherapy and the gut microbiome.

Checkpoint blockade responders tend to have high abundance of Bifidobacterium, Akkermansia, and Faecalibacterium spp. in their gut microbiota, whereas nonresponders do not. A fecal transplant from responders into germ-free mice expressing tumors results in increased efficacy of checkpoint inhibition and tumor reduction. Fecal transplants from nonresponders confers no antitumor effect on mice.
Mastering Microbiology
Art Activity: Figure 28.27 Mechanisms of action of major antibacterial agents
Immunotherapeutic Efficacy and the Gut Microbiome
The composition of each person’s gut microbiota plays a significant role in the efficacy of a given immunotherapeutic treatment. For example, studies in mice have shown that administering Bifidobacterium as an oral probiotic produces an antitumor effect, and the use of PD-1/PD-L1 checkpoint inhibitors in addition to the probiotic can eliminate tumor growth entirely. Subsequent studies revealed that the abundance of the bacterium Bifidobacterium is particularly high in feces of melanoma patients that respond favorably to PD-1/PD-L1 blockade, and transplanting samples of the feces into germ-free mice (fecal transplant, Sections 24.9 and 24.11) improved the anticancer activity of checkpoint inhibitors and CD8 T cells in the mice (Figure 28.11). Using data from mouse studies as proof-of-concept, clinical trials are now under way to determine whether fecal transplants from patients who respond favorably to PD-1 checkpoint inhibitor therapy into those who do not respond to the treatment can ameliorate the effect of checkpoint blockade in nonresponders.
The mechanism responsible for the beneficial effect of the transplants has been linked to the release of immune-stimulating cytokines by gut-associated lymphoid tissue upon interaction with the beneficial bacteria. The goal now is to streamline the procedure and increase its reproducibility by determining as precisely as possible which microbes are conferring the beneficial effect and formulating a probiotic suspension of just those organisms. In addition to Bifidobacterium, research has focused on species of Bacteroides, Faecalibacterium, and Akkermansia (Figure 28.11) and species of the family Ruminococcaceae. Knowledge of the major contributors will minimize potential problems with quality control, as well as the risk of inadvertent transfer of pathogens or immunosuppressive strains in fecal transplants, both of which could undermine the benefits of the treatment.
Check Your Understanding
How do checkpoint inhibitors stimulate the immune response in the tumor microenvironment? How do chimeric antigen receptors (CARs) differ from T cell receptors (TCRs)?
Explain how the composition of the gut microbiome may influence the efficacy of cancer immunotherapy. What evidence do we have for this phenomenon?
III Drug Treatments for Infectious Diseases
Antimicrobial drugs kill or inhibit microorganisms by a variety of mechanisms and are of immense importance in clinical medicine. However, antimicrobial drug resistance is an escalating threat and a major impediment to the treatment of certain infections.
28.5 Antibacterial Drugs
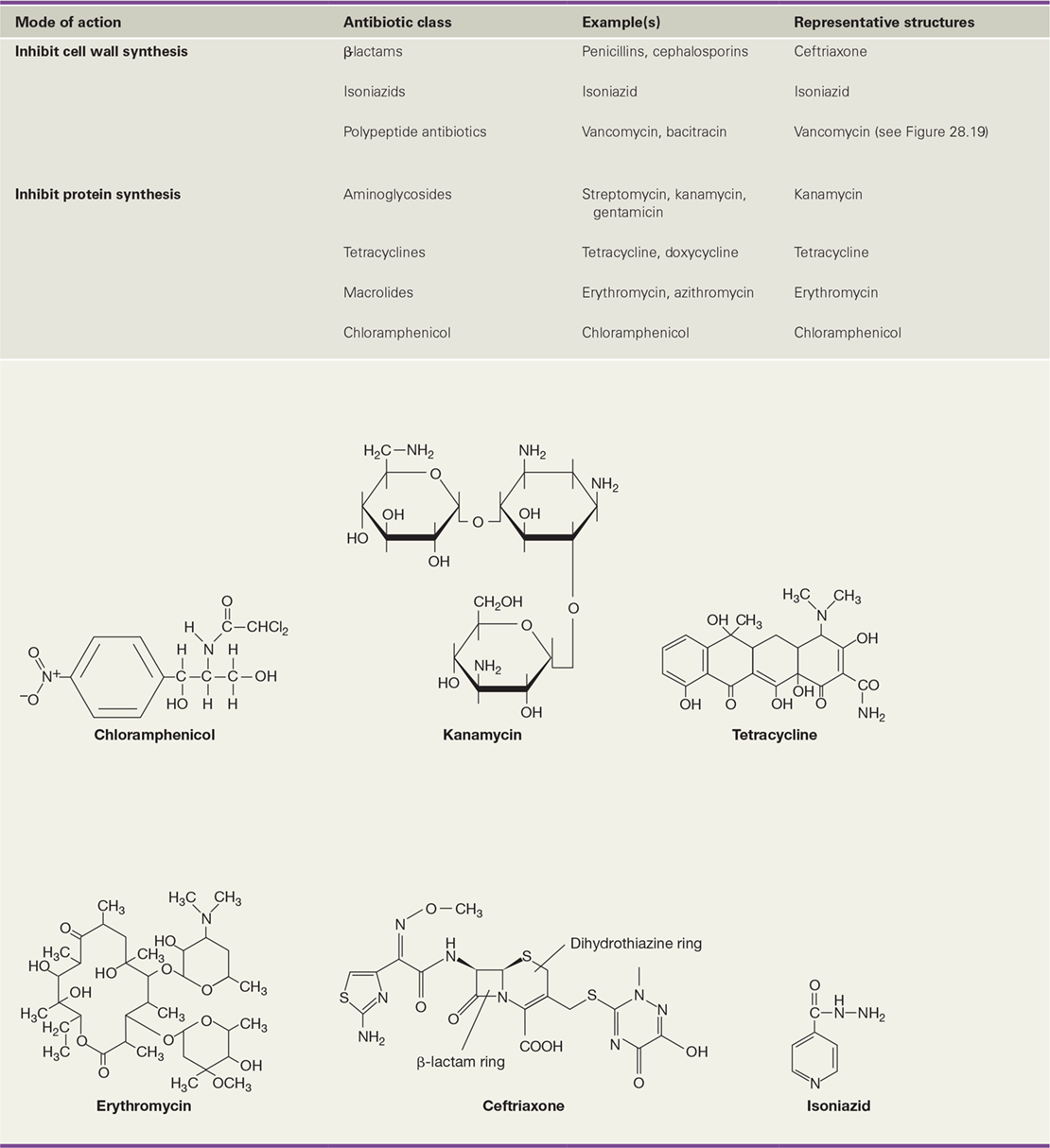
In cases where immunization is either not an option or fails to prevent disease, antimicrobial drug therapy is the primary weapon against infections. Antimicrobial drugs either kill or inhibit the growth of microorganisms in the host (in vivo) and are classified based on their molecular structure, mechanism of action (Figure 28.12), and spectrum of antimicrobial activity. Antibiotics are antimicrobial agents that are produced naturally by certain microorganisms, typically species of Bacteria or fungi. In addition to natural antibiotics, many other drugs are produced synthetically. Regardless of their origin, it is desirable that antimicrobial drugs exhibit selective toxicity—they inhibit or kill pathogens without adversely affecting the host. Although thousands of antibiotics are known, fewer than 1% are clinically useful, often because of problems with host toxicity or lack of uptake by host cells.
Figure 28.12 Mechanisms of action of major antibacterial agents.

Agents are classified according to their target structures in the bacterial cell. THF, tetrahydrofolate; DHF, dihydrofolate; PABA, p-aminobenzoic acid.
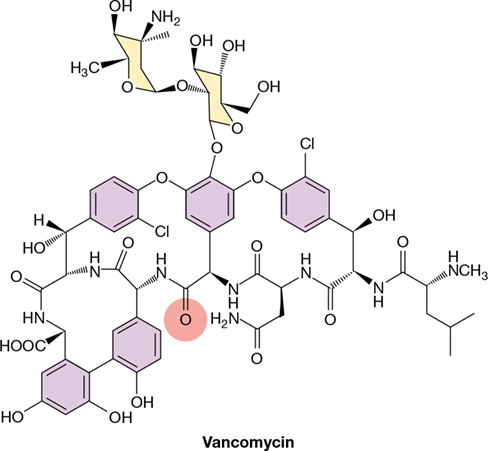
The susceptibility of individual microbes to different antimicrobial agents varies significantly. For example, gram-positive Bacteria are often sensitive to natural penicillin, whereas gram-negative Bacteria are generally resistant; thus, natural penicillin exhibits a relatively narrow spectrum of activity. By contrast, antibiotics that exhibit broad-spectrum activity, such as tetracycline, are generally effective against both groups. Although broad-spectrum antibiotics often find wider medical use than narrow-spectrum antibiotics, antibiotics with a limited spectrum of activity may be quite useful against certain pathogens, especially those that fail to respond to other antibiotics. A good example is vancomycin, a narrow-spectrum antibiotic that is an effective bactericidal agent for penicillin-resistant, gram-positive Bacteria, including certain enterococci, staphylococci, and clostridia.
Important targets of antibiotics in Bacteria include the cell wall, ribosomes, enzymes that facilitate nucleic acid synthesis or catalyze metabolic processes, and the cytoplasmic membrane (Figure 28.12). We provided an overview of antibiotic targets in Section 8.11 and now consider the mechanisms of drug activity in further detail.
The Bacterial Cell Wall as a Drug Target
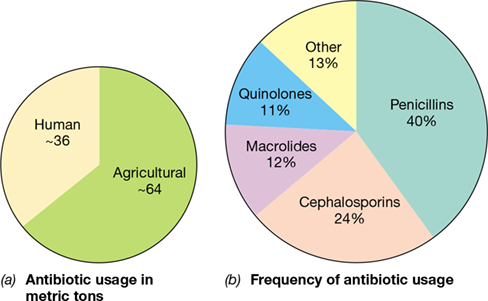
Worldwide, an estimated 100,000 metric tons of antibacterial drugs are manufactured and used annually (**Figure 28.13*a***), and the vast majority of these target the bacterial cell wall. β-lactam antibiotics, which include penicillins and cephalosporins, inhibit cell wall synthesis and account for nearly two-thirds of all antibiotics produced and used worldwide (Figure 28.13b). These antibiotics share a characteristic structural component, the β-lactam ring (Figure 28.14), and historically have been some of the most effective weapons against many different types of bacterial infections.
Figure 28.13 Annual worldwide production and use of antibiotics.

(a) An estimated 100,000 metric tons of antimicrobial agents are manufactured worldwide per year, and nearly two-thirds of this amount is used in an agricultural context. (b) By far, the β-lactam antibiotics (penicillins and cephalosporins) constitute the most important and widely used class of antibiotic. “Other” includes trimethoprim combinations, tetracyclines, chloramphenicol, aminoglycosides, and all other antimicrobial drug classes. Data are from the Center for Disease Dynamics, Economics, and Policy, Washington, DC.
Figure 28.14 Structure of selected penicillins.

The red arrow (top panel) indicates the site within the β-lactam ring cleaved by most β-lactamases, enzymes that destroy penicillin and other β-lactam antibiotics. Although the N-acyl group (blue shading) varies among different penicillin drugs, all penicillins have a common nucleus consisting of a β-lactam ring and a thiazolidine ring. Whereas penicillin G must be injected, most acid-stable penicillins can be administered orally (carbenicillin is an exception).
The first β-lactam antibiotic ever characterized was penicillin G, isolated from the fungus Penicillium chrysogenum in 1929 by Alexander Fleming. This new drug was dramatically effective for controlling staphylococcal and pneumococcal infections and was more effective for treating streptococcal infections than the synthetic sulfa drugs, discussed later in this section. Penicillin and other β-lactam antibiotics interfere with an important feature of bacterial cell wall synthesis called transpeptidation, the reaction catalyzed by transpeptidases that results in the cross-linking of two glycan-linked peptide chains (Sections 2.3 and 8.5). Because peptidoglycan synthesis mechanisms are unique to Bacteria, the β-lactam antibiotics are highly selective and nontoxic to host cells.
Penicillin G is active primarily against gram-positive Bacteria because gram-negative Bacteria are impermeable to the antibiotic. However, chemical modification of the N-acyl group of penicillin G produces semisynthetic penicillins, such as ampicillin and carbenicillin, which have broader activity and are effective against certain gram-negative Bacteria (Figure 28.14). The structural differences in these semisynthetic penicillins allow them to be transported across the gram-negative outer membrane (Section 2.4), where they inhibit cell wall synthesis. Penicillin G is also sensitive to β-lactamases, enzymes that destroy β-lactam antibiotics and are produced by many penicillin-resistant Bacteria (see Section 28.7). Oxacillin and methicillin are widely used β-lactamase-resistant semisynthetic penicillins (Figure 28.14).
Cephalosporins, produced by species of the fungus Cephalosporium, differ structurally from the penicillins. They retain the β-lactam ring but have a six-member dihydrothiazine ring joined to it instead of the five-member thiazolidine ring of penicillins (Table 28.4 and Figure 28.14). The cephalosporins have the same mode of action as the penicillins; they bind irreversibly to transpeptidases and prevent the cross-linking of peptidoglycan. Clinically important cephalosporins are semisynthetic antibiotics with a broader spectrum of activity than the penicillins. In addition, cephalosporins are typically more resistant to β-lactamases. For example, ceftriaxone (Table 28.4) is highly resistant to β-lactamases and has replaced penicillin for treatment of gonorrhea because many Neisseria gonorrhoeae strains have become resistant to penicillin (see Section 28.7).
Table 28.4 Selected antibacterial compounds


Some antimicrobial drugs mimic growth factors, organic nutrients obtained from an organism’s environment that are required by the organism for growth and survival (Section 3.1). A growth factor analog is a synthetic compound that chemically resembles a growth factor, but whose subtle structural differences prevent the analog from functioning as a growth factor in the cell, thereby disrupting cell metabolism. Isoniazid (Table 28.4) is an important growth factor analog with a very narrow spectrum of activity. Effective against mycobacteria only, isoniazid is an analog of nicotinamide, a vitamin required for mycolic acid synthesis, which is, in turn, required to construct the mycobacterial cell wall. Isoniazid is one of the most effective drugs for treatment of tuberculosis (Section 31.4), but isoniazid resistance in strains of Mycobacterium tuberculosis is increasing.
Protein Synthesis as a Drug Target
Some antibiotics inhibit bacterial pathogens by disrupting protein synthesis (translation), often through interactions with the ribosome that may include binding to ribosomal RNA (rRNA) (Figure 28.12). Most of these drugs target bacterial (70S) ribosomes only and, therefore, have no effect on the structurally distinct, cytoplasmic ribosomes of eukaryotic cells. However, because mitochondria also contain 70S ribosomes (Section 13.4), many antibiotics that inhibit protein synthesis in Bacteria also potentially inhibit mitochondrial protein synthesis in eukaryotic cells. Nevertheless, these drugs are still medically useful because 70S ribosomes in organelles are affected only at higher concentrations than are used for antimicrobial therapy.
The aminoglycosides are antibiotics that inhibit translation by targeting the 30S (small) subunit of the ribosome. Aminoglycosides contain amino sugars linked by glycosidic bonds, and clinically useful examples include streptomycin (produced by the bacterium Streptomyces griseus), kanamycin (Table 28.4), neomycin, and gentamicin. These broad-spectrum antibiotics are useful for treating infections caused by gram-negative Bacteria, but because of host toxicity effects that include kidney and hearing damage, aminoglycoside use has decreased since the development of the semisynthetic penicillins and tetracyclines. With the exception of gentamicin, which is routinely used to combat Pseudomonas infections, and neomycin, which is used in topical antibiotic ointments, aminoglycosides are generally administered only when other antibiotics fail.
Like the aminoglycoside antibiotics, tetracyclines interfere with the function of the 30S subunit of the ribosome. Tetracyclines are broad-spectrum antibiotics produced by several species of Streptomyces, and they inhibit most clinically relevant gram-positive and gram-negative Bacteria. The basic structure of tetracycline consists of a naphthacene ring system (Table 28.4), and side-chain substitutions (either natural or synthetic) to the various rings form new tetracycline analogs. Physicians are cautioned against the administration of tetracyclines to pregnant women and to young children because these antibiotics bind calcium in bones and teeth, weakening them and causing permanent staining of the latter (Figure 28.15). Tetracyclines are widely used in veterinary medicine, though, and in some countries, they are used routinely in healthy animals as nutritional supplements to suppress the growth of pathogens in poultry and swine, a practice that strongly selects for antibiotic resistance (Section 8.11).
Figure 28.15 Staining of teeth from the use of tetracycline.

Tetracycline binds calcium in developing bones and teeth, weakening them and causing permanent staining of tooth enamel. Therefore, tetracycline should not be administered to pregnant women or children unless absolutely necessary.
Macrolide antibiotics inhibit translation by targeting the 50S (large) subunit of the bacterial ribosome. Their basic structure contains a lactone ring bonded to sugars (as in erythromycin in Table 28.4), and variations in these constituents result in a diversity of macrolide antibiotics. Macrolides account for about 12% of global antibiotic production (Figure 28.13) and include, for example, erythromycin (produced by Streptomyces erythreus), clarithromycin, and azithromycin. The partial inhibition of protein synthesis by erythromycin, in particular, leads to preferential translation of some proteins and restricts translation of others, resulting in an imbalance in the proteome and potentially disrupting metabolic functions at all levels. Often used clinically in patients allergic to penicillin or other β-lactam antibiotics, erythromycin is particularly useful for treating legionellosis (Section 33.4).
Nucleic Acid Synthesis as a Drug Target
The quinolones are synthetic antibacterial compounds that disrupt bacterial metabolism by interfering with DNA gyrase, thus preventing the supercoiling and packaging of DNA in the bacterial cell (Section 6.1). Because DNA gyrase is found in all Bacteria, quinolones are effective for treating both gram-positive and gram-negative bacterial infections. Fluoroquinolones, such as ciprofloxacin (Table 28.4), are fluorinated derivatives of quinolones that are routinely used to treat urinary tract infections and have been widely used in the beef and poultry industries for prevention and treatment of respiratory diseases in animals. Ciprofloxacin is also the drug of choice for treating anthrax because some strains of Bacillus anthracis, the causative agent of anthrax (Section 32.8), are resistant to penicillin. The fluoroquinolone moxifloxacin is effective for treatment of tuberculosis; in combination with other anti-tuberculosis drugs, moxifloxacin may significantly reduce treatment time, a major problem with isoniazid-based treatments. However, in some individuals the use of fluoroquinolones has been accompanied by serious side effects, including tendon and organ damage and neurotoxicity, although why these occur is not well understood.
Some antibiotics disrupt transcription by inhibiting RNA synthesis (Figure 28.12). For example, rifamycins, such as rifampin (Table 28.4), inhibit RNA synthesis by binding to the β-subunit of RNA polymerase in Bacteria. Rifampin, often used in concert with isoniazid for treatment of tuberculosis, has the odd side effect of causing body secretions, including tears, urine, and sweat, to turn reddish-orange (Figure 28.16). Actinomycin inhibits RNA synthesis by combining with DNA and blocking RNA elongation. This agent binds most strongly to DNA at guanine–cytosine base pairs, fitting into the major groove in the double strand where RNA is synthesized.
Figure 28.16 Reddish-orange urine from rifampin usage.
The antibiotic rifampin is often administered to tuberculosis patients. However, a side effect of rifampin usage is that it turns urine and other body fluids reddish-orange.
Other Antibacterial Drug Targets
Like isoniazids, sulfa drugs (also called sulfonamides) are synthetic growth factor analogs. However, instead of affecting the cell wall, sulfa drugs block a key biosynthetic pathway in Bacteria. Sulfanilamide (Table 28.4), the simplest sulfa drug, is an analog of p-aminobenzoic acid (PABA), which is a component of the vitamin folic acid, a nucleic acid precursor. By mimicking PABA, sulfanilamide blocks the synthesis of folic acid, thereby inhibiting nucleic acid synthesis. Sulfanilamide is selectively toxic because bacteria synthesize their own folic acid, unlike humans and most animals, which obtain folic acid from their diet. Antimicrobial therapy with sulfamethoxazole (a sulfa drug) plus trimethoprim, a related folic acid synthesis competitor, is highly effective because the drug combination blocks two sequential steps in the folic acid synthesis pathway (Figure 28.12); resistance to this drug combination therefore requires two mutations in genes of the same pathway, a relatively rare event. However, resistance to sulfonamides is increasing as many formerly susceptible pathogens develop the ability to import folic acid from their environment.
Some antibiotics have atypical structures or targets. For example, the antibiotic daptomycin (Table 28.4) is produced by a species of Streptomyces and is a cyclic lipopeptide with a unique mode of action. Used mainly to treat infections by gram-positive Bacteria, including pathogenic streptococci and staphylococci, daptomycin binds specifically to bacterial cytoplasmic membranes, forms a pore, and induces rapid depolarization of the membrane. The depolarized cell quickly loses its ability to synthesize macromolecules, such as nucleic acids and proteins, resulting in cell death. However, mutations leading to alterations in cytoplasmic membrane structure may lead to resistance.
Platensimycin (Table 28.4), produced by Streptomyces platensis, is an unusual antibiotic that inhibits fatty acid and lipid biosynthesis. Platensimycin is effective against a broad range of gram-positive Bacteria, including nearly untreatable infections caused by methicillin-resistant Staphylococcus aureus (MRSA) and vancomycin-resistant enterococci. Platensimycin has a unique mode of action, shows no host toxicity, and there is no known potential for development of resistance by pathogens. We discuss the discovery of platensimycin in Section 28.7.
Check Your Understanding
Explain the concept of selective toxicity in terms of antimicrobial therapy.
How does the activity of each antibiotic class lead to control of the affected pathogens?
What are the sources of aminoglycosides, tetracyclines, macrolides, daptomycin, and platensimycin?
28.6 Antimicrobial Drugs That Target Nonbacterial Pathogens
28.6 Antimicrobial Drugs That Target Nonbacterial Pathogens
28.6 Antimicrobial Drugs That Target Nonbacterial Pathogens
Many pathogens are nonbacterial—such as viruses or eukaryotes—and are often difficult to treat. Antiviral drugs often adversely affect the host as well as the pathogen because viruses depend on host cell biosynthetic machinery for their replication (Chapter 5). As a result, selective toxicity for viruses is achieved only with agents that preferentially affect unique viral replication pathways or the assembly of viral components. In spite of these limitations, a number of drugs are more toxic for viruses than for the host, and a few agents specifically target individual viruses. Fungi, protozoa, and helminths (worms) pose special problems for the development of effective drugs because these pathogens are eukaryotic, and therefore their cellular biology is similar to that of humans.
Antiviral Drugs
The most successful and commonly used agents for antiviral chemotherapy are the nucleoside analogs. These drugs are nucleoside reverse transcriptase inhibitors (NRTIs), which work by inhibiting elongation of the viral nucleic acid chain by a nucleic acid polymerase. The first NRTI to be widely used was zidovudine, also called azidothymidine (AZT) (Table 28.5; Figure 31.49a), which effectively blocks reverse transcription and production of complementary DNA (cDNA) in HIV and other retroviruses (Section 31.15). It is structurally similar to thymidine but is a dideoxy derivative, lacking the 3′-hydroxyl group. Another widely used nucleoside analog, acyclovir (Table 28.5), resembles guanosine and has been successfully used to control the symptoms of herpesvirus infections (Section 31.14).
Table 28.5 Antiviral compounds

Some antiviral agents target the key enzyme of retroviruses, reverse transcriptase. Nevirapine(Figure 31.49b), a nonnucleoside reverse transcriptase inhibitor (NNRTI), binds directly to reverse transcriptase and inhibits reverse transcription (Table 28.5). Phosphonoformic acid, an analog of inorganic pyrophosphate, inhibits normal internucleotide linkages, preventing synthesis of viral nucleic acids. Because their action may affect normal host cell nucleic acid synthesis, both NRTIs and NNRTIs often induce some host toxicity.
Other anti-retroviral drugs include protease inhibitors and enfuvirtide, a fusion inhibitor (Table 28.5). Protease inhibitors disrupt viral replication by binding the active site of HIV protease (see Figure 28.20a), preventing this enzyme from processing large viral polyproteins into several individual viral components (Section 11.8) including reverse transcriptase and integrase. Enfuvirtide (marketed as Fuzeon®) is a 36-amino-acid synthetic peptide that binds to the gp41 membrane protein of HIV; this stops the conformational changes necessary for the fusion of HIV with T lymphocyte membranes, thus preventing infection of host immune cells by HIV (Figure 31.44).
A single category of drugs limits influenza infection: The neuraminidase inhibitors include oseltamivir (marketed as Tamiflu®), zanamivir (marketed as Relenza®), and peramivir (marketed as Rapivab®). These drugs block the active site of neuraminidase (Figure 11.24) in influenza A and B viruses, inhibiting virus release from infected cells (Table 28.5). Oseltamivir is used for both treatment and prophylaxis of influenza, but its use has, in some cases, been linked to adverse side effects that include behavioral abnormalities, especially in children. Zanamivir and peramivir are suitable for children, but because zanamivir is an inhaled drug, it is not recommended for those with asthma or other respiratory conditions.
Recall from Chapter 26 that virus-infected cells release small cytokine proteins called interferons that trigger a defensive response in neighboring host cells (Section 26.10). If produced and administered properly, interferons may have potential use as prescribed antimicrobial agents. The clinical utility of interferons depends on whether they can be delivered to specific areas in the host to stimulate the production of antiviral proteins in uninfected host cells. Alternatively, appropriate interferon stimulators, such as viral nucleotides, nonvirulent viruses, or even synthetic nucleotides, if given to host cells prior to infection, may induce natural production of interferon.
Drugs that Target Eukaryotic Pathogens
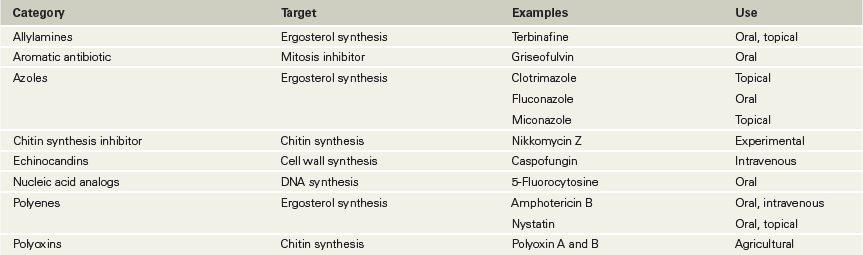
Fungi cause a number of serious diseases (Sections 34.1 and 34.2), and their treatment is complicated by the fact that many antifungal agents target metabolic pathways that are shared by fungi and their host cells, thus making the drugs toxic. As a result, many antifungal drugs can be used only for topical (surface) applications (Table 28.6).
Table 28.6 Antifungal agents

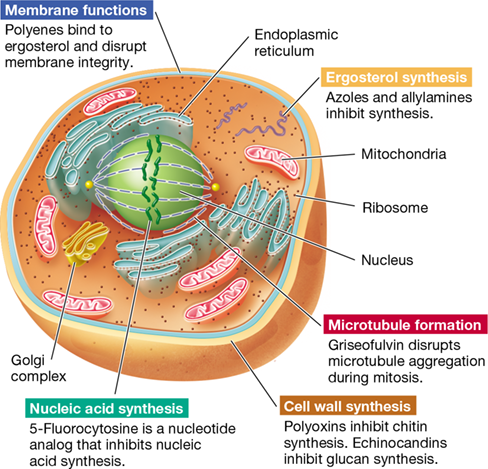
A major group of antifungal compounds are ergosterol inhibitors, which work either by interacting directly with ergosterol or inhibiting its synthesis (Table 28.6). Ergosterol is present in fungal cytoplasmic membranes in place of the cholesterol found in animal cell cytoplasmic membranes. Important ergosterol inhibitors include the polyene antibiotics, which are produced by species of Streptomyces bacteria. Polyenes bind specifically to ergosterol, causing membrane permeability and cell death (Figure 28.17). By contrast, azoles and allylamines are broad-spectrum antifungal drugs that work by selectively inhibiting ergosterol biosynthesis. Treatment with these drugs causes membrane damage and alteration of critical membrane transport activities.
Figure 28.17 Targets of some antifungal agents.

Traditional antibacterial agents are generally ineffective because fungi are eukaryotic cells. The cell wall and ergosterol (present in the cytoplasmic membrane) of fungal cells make good selective targets because they are unique structures not present in vertebrate host cells.
Echinocandins are cell wall inhibitors that block the activity of 1,3-β-d-glucan synthase, the enzyme that forms β-glucan polymers in the fungal cell wall (Figure 28.17 and Table 28.6). Because mammalian cells do not have 1,3-β-d-glucan synthase (or cell walls), the activity of these agents selectively kills fungal cells. Echinocandins are often used to treat Candida yeast infections, as well as some fungal pathogens that are resistant to azoles and other agents.
Fungal cell walls also contain chitin, a polymer of N-acetylglucosamine found only in fungi and insects. Several polyoxins inhibit cell wall synthesis by interfering with chitin biosynthesis (Figure 28.17). Although not used clinically, polyoxins are widely used as agricultural fungicides. Other antifungal drugs inhibit folate biosynthesis, interfere with DNA topology during replication, or, in the case of drugs such as griseofulvin, disrupt microtubule aggregation during mitosis (Figure 28.17). Moreover, the nucleic acid analog 5-fluorocytosine (flucytosine) is an effective nucleic acid synthesis inhibitor in fungi.
Historically, protozoan-caused diseases, especially malaria, have been treated with quinine or, more recently, quinine derivatives, such as chloroquine and mefloquine. However, extensive resistance to these drugs among the Plasmodium spp. that cause malaria (Section 34.5) has led to the development of artemisinin-based alternatives. Artemisinin is produced in low amounts by the Chinese wormwood plant (Artemisia), which has traditionally been used to control the cyclic fever associated with malaria. A more recent discovery has shown artemisinin to also be an effective treatment for certain helminth diseases, such as schistosomiasis (Section 34.7). Using the techniques of synthetic biology, a genetically engineered yeast strain is now being used to produce artemisinin, making the drug more widely available (Section 12.11 and Figure 12.37).
The drug of choice to combat many parasitic protozoans is metronidazole (or the related drug tinidazole), which targets anaerobic pathogens and is therefore also useful against infections caused by clostridia. Infections with Giardia intestinalis (giardiasis), Trichomonas vaginalis (trichomoniasis), and especially Entamoeba histolytica (amebic dysentery) have all been treated successfully with these drugs (Sections 34.3 and 34.4). The related protist Cryptosporidium parvum (cryptosporidiosis) is often treated with nitazoxanide, a drug that has also been effective against helminths. However, perhaps the most widely used antihelminthic drugs are praziquantel, for treating schistosomiasis and tapeworm infections, and mebendazole (and similar drugs), for treating a variety of helminth infections, including tapeworms, pinworms, hookworms, trichinosis, and ascariasis (Section 34.7).
Mastering Microbiology
Art Activity: Figure 28.32 Targets of some antifungal agents
We now tackle the topic of drug resistance, a consequence of the widespread use of antimicrobial agents in clinical and veterinary medicine and animal husbandry.
Check Your Understanding
What steps in the viral maturation process are inhibited by nucleoside analogs? By protease inhibitors? By interferons?
Why are there fewer clinically effective antifungal and antiparasitic agents than antibacterial agents?
28.7 Antimicrobial Drug Resistance and New Treatment Strategies
28.7 Antimicrobial Drug Resistance and New Treatment Strategies
28.7 Antimicrobial Drug Resistance and New Treatment Strategies
Antimicrobial drug resistance is the acquired ability of a microorganism to resist the effects of an antimicrobial agent to which it was formerly susceptible. As we have discussed, many microorganisms produce antibiotics, and genes encoding antibiotic resistance are present in virtually all of them. Horizontal gene transfer (Chapters 9 and 13) between and among microorganisms disseminates antimicrobial drug resistance.
Antimicrobial Drug Resistance
Common mechanisms of bacterial resistance to antibiotics were discussed in the context of microbial growth in Section 8.11 and depicted in Figure 8.28, and some examples are listed in Table 28.7. Antibiotic resistance can be genetically encoded on the bacterial chromosome, but more often, antibiotic-resistant bacteria isolated from patients contain drug-resistance genes located on horizontally transmitted R (resistance) plasmids(Section 6.2). Enzymes encoded by genes on R plasmids confer resistance by any of three classes of mechanisms: modifying and inactivating the drug, preventing uptake of the drug, or actively pumping the antibiotic out of the cell, a process called efflux (Table 28.7).
Table 28.7 Bacterial resistance to antibiotics

The widespread use of antibiotics (Figure 28.13a) provides favorable conditions for the spread of R plasmids because they carry genes that confer an immediate selective advantage. The ubiquity of resistance genes limits the long-term use of any single antibiotic as an effective antimicrobial agent. A classic example is the development of multidrug resistance in the bacterium Neisseria gonorrhoeae. Penicillin, widely used to treat gonorrhea into the 1980s, became ineffective and was replaced by ciprofloxacin, which, in turn, lost much of its efficacy after just 10 years of use (Figure 28.18). This prompted a switch in treatment to the β-lactamase-resistant ceftriaxone, and more recently, a combination of ceftriaxone and azithromycin. Such combinations of two unrelated antimicrobial agents often reduce resistance because it is less likely that a mutant strain resistant to one antibiotic will also be resistant to the second antibiotic. However, certain R plasmids confer multiple drug resistance and can thwart multiple antibiotic therapy as a clinical strategy.
Figure 28.18 The emergence of multidrug resistance in *Neisseria gonorrhoeae*.

Resistance to penicillin developed in the 1980s to the point that it could no longer be recommended for treatment of gonorrhea, at which point ciprofloxacin, a quinolone, became the drug of choice. By the early 2000s, resistance to ciprofloxacin prompted a change to ceftriaxone. The current gonorrhea treatment recommendation is a combination of ceftriaxone plus azithromycin.
Overuse of antibiotics also accelerates resistance. In addition to their traditional use as a treatment for infections, antibiotics are widely used as supplements to farm animal feeds both as growth-promoting substances and as prophylactic additives to prevent the occurrence of disease. For example, broad-spectrum antibiotics widely used clinically, such as the fluoroquinolone ciprofloxacin, were used extensively as a feed additive, especially by the poultry industry. In addition, although the trend is improving, antibiotics are used in clinical practice far more often than necessary, and this problem is compounded by patient noncompliance—many patients stop taking antibiotics as soon as they feel better. Antibiotic resistance can be minimized if drugs are used only for treatment of susceptible pathogens and are given in sufficiently high doses and for sufficient lengths of time to eradicate the pathogen before resistant mutants can propagate.
To prevent the further emergence of multidrug-resistant pathogens, the U.S. Centers for Disease Control and Prevention (CDC) publishes guidelines stressing the importance of vaccination, rapidly and accurately diagnosing and treating infections, using antimicrobial agents prudently, and preventing pathogen transmission.
New Drugs and New Treatment Strategies
Conservative, appropriate administration of antibiotics can prolong or even resurrect the clinical usefulness of available drugs, but the long-term solution to antimicrobial drug resistance requires continuous development of new drugs through design or discovery. Developing new analogs of existing antimicrobial compounds is usually more cost effective than discovering new drugs, and analogs may actually have greater antimicrobial activity than the parent compound. For example, using natural penicillin as the starting compound, systematic chemical substitution of the N-acyl group can generate hundreds of penicillin derivatives, many with broad-spectrum activity (Figure 28.14). Using this basic strategy, semisynthetic analogs of β-lactam antibiotics, tetracycline, and vancomycin (Figure 28.19) have been synthesized.
Figure 28.19 Vancomycin.

Many pathogenic strains now show intermediate drug resistance to the parent structure of vancomycin, but chemically substituting the carbonyl oxygen at the position shown in red with a methylene (=CH2) group restores much of the lost activity. Like penicillin, vancomycin prevents cross-linking of peptidoglycan and is most effective against gram-positive pathogens.
Novel antimicrobial compounds are more difficult to identify than analogs of existing drugs because new antimicrobial compounds must work at unique sites in metabolism or be structurally dissimilar to existing compounds to avoid resistance mechanisms. Computer-based methods accelerate novel compound design by maximizing molecular binding in a virtual environment. A dramatic success in computer-directed drug design was the development of saquinavir, a protease inhibitor that impedes the multiplication of HIV in infected individuals by binding the active site of the HIV protease enzyme (Figure 28.20). As an analog of the HIV precursor protein, saquinavir (marketed as Invirase®) displaces the authentic protease substrate and inhibits virus maturation. Several other computer-designed protease inhibitors, including indinavir (marketed as Crixivan®) (Figure 28.20b), are currently in use for the treatment of HIV/AIDS (Section 31.15).
Figure 28.20 Computer-generated anti-HIV drugs.

(a) The HIV protease homodimer. Individual polypeptide chains are shown in green and blue. A peptide (yellow) is bound in the active site. HIV protease cleaves an HIV precursor protein, a necessary step in virus maturation. Blocking of the protease site by the bound peptide inhibits precursor processing and HIV maturation. This structure is derived from coordinates in the Protein Data Bank. (b) These anti-HIV drugs are peptide analogs called protease inhibitors that were designed by computer to block the active site of HIV protease. The areas highlighted in orange show the regions analogous to peptide bonds in proteins.
Another strategy to overcome antimicrobial resistance is to select for antibiotics that interact with unexploited targets. Platensimycin is the first antimicrobial drug targeted to disrupt bacterial lipid biosynthesis (Table 28.4 and Figure 28.12). Platensimycin is especially active against gram-positive pathogens, including drug-resistant staphylococci and enterococci. To select an agent for a defined target, in this case an enzyme in the bacterial lipid synthesis pathway, scientists used antisense RNA (Section 7.12) to limit the amount of accessible mRNA required to produce a key lipid biosynthesis enzyme in Staphylococcus aureus. This reduced fatty acid synthesis in the cells and increased the sensitivity of the crippled S. aureus strain to antibiotics that inhibit fatty acid synthesis. After screening thousands of natural products from potential antibiotic producers, scientists isolated platensimycin from the soil bacterium Streptomyces platensis, a species of the filamentous Actinobacteria(Section 16.12). This strategy identifies target-specific antibiotics present in low concentrations and is applicable to virtually any target for which the gene sequence (and, hence, the corresponding antisense RNA sequence) is known.
The efficacy of some antibiotics can be preserved if they are administered with compounds that thwart antibiotic resistance mechanisms. For example, several β-lactam antibiotics can be combined with β-lactamase inhibitors to preserve antibiotic activity in β-lactam-resistant microbes. For instance, amoxicillin (similar to ampicillin, Figure 28.14) can be mixed with the β-lactamase inhibitor clavulanic acid to produce the combination drug Augmentin®. The inhibitor binds β-lactamase (produced by the resistant pathogen) irreversibly. This prevents amoxicillin from being degraded and maintains the efficacy of the antibiotic.
Drug combination therapy has also revolutionized treatment of HIV infections. Currently, a combination therapy consisting of nucleoside analogs and a protease inhibitor is recommended. This drug treatment protocol is termed HAART, for highly active anti-retroviral therapy (Section 31.15). As with antibacterial combination regimens, HAART is designed to target two independent viral functions: nucleoside analogs target virus replication, and protease inhibitors target virus maturation. Because the probability of a virus developing resistance to multiple drugs is much less than the probability of it becoming resistant to a single drug, HAART therapy has been a successful strategy in controlling HIV infections.
Check Your Understanding
Identify the basic mechanisms of antibiotic resistance in bacteria and describe what practices encourage their dissemination.
What does vancomycin have in common with penicillin? How can native vancomycin be improved?
Explain the advantages and disadvantages of developing new drugs based on existing drug analogs. What other methods exist for developing new drugs?
Chapter Review
Go to Mastering Microbiology for videos, animations, practice tests, and more.
I Disorders and Deficiencies of the Immune System
28.1 Hypersensitivity is the induction by foreign antigens of antibody-mediated or cell-mediated immune responses that damage host tissue. In autoimmunity, the immune response is directed against self antigens. Damage to host tissue is caused by the inflammation produced by immune mechanisms.
Q How do immediate and delayed-type hypersensitivities differ from one another in terms of immune effectors, target tissues, antigens, and clinical outcome?
28.2 Superantigens are components of certain bacterial and viral pathogens that bind outside the antigen-specific binding site of TCRs and, therefore, activate large numbers of T cells. Superantigen-activated T cells may produce diseases characterized by systemic inflammatory reactions. Immunodeficiency is the inability to generate a proper immune response, resulting in recurrent, uncontrollable infections by opportunistic pathogens. Some of the most severe immunodeficiency syndromes are SCID, caused by a genetic disorder, and AIDS, caused by HIV infection.
Q How do superantigens differ from conventional antigens in terms of initial T cell activation and clinical outcome? How does immunodeficiency resulting from SCID differ from that caused by HIV infection and AIDS?
II Vaccines and Immunotherapy
28.3 Immunization induces artificial active immunity and is widely used to prevent infectious diseases. Vaccines are either attenuated or inactivated pathogens or pathogen products or are genetically engineered antigens. The latter eliminate exposure to pathogens and, in some cases, even to protein antigen. Application of this strategy is providing safer vaccines targeted to individual pathogen antigens.
Q List the immunizations recommended for children in the United States. For which of these have you been immunized? List the diseases for which you may have acquired immunity naturally.
28.4 A growing number of diseases, including a variety of cancers and autoimmune disorders, are being treated by immunotherapy, a clinical strategy that optimizes the adaptive immune response to a disease using bioengineered immune components. The identification of tumor-specific neoantigens in cancer cells can allow for production of customized monoclonal antibodies (mAbs), chimeric antigen receptor (CAR) T cells, and anticancer vaccines. The efficacy of immunotherapeutic treatments varies between individuals, and this may be linked to the composition of their gut microbiota.
Q What is a “neoantigen”? How are they identified, and how can they be used to develop mAbs, CAR T cells, and anticancer vaccines?
III Drug Treatments for Infectious Diseases
28.5 Antibiotics are chemically diverse antimicrobial compounds produced by microorganisms. Each antibiotic works by inhibiting a specific cellular process in the target microorganisms. The β-lactam antibiotics, including penicillins and cephalosporins, target bacterial cell wall synthesis and are the most important class of clinical antibiotics. The aminoglycosides, macrolides, and tetracycline antibiotics selectively interfere with protein synthesis in Bacteria. The quinolones are an important class of synthetic antibacterial drugs that inhibit DNA synthesis. Daptomycin and platensimycin are structurally novel antibiotics that target cytoplasmic membrane functions and lipid biosynthesis, respectively.
Q What are the common sources for natural antibiotics? How do these antimicrobial drugs differ from growth factor analogs, such as the sulfa drugs? Why are β-lactam antibiotics generally more effective against gram-positive bacteria than against gram-negative bacteria?
28.6 Antiviral agents selectively target virus-specific enzymes and processes. Useful agents include analogs and compounds that inhibit nucleic acid polymerases and viral genome replication. Protease inhibitors interfere with viral maturation steps. Because fungi are Eukarya, antifungal agents exhibiting selective toxicity are difficult to find. Nevertheless, some effective antifungal agents are available, and they primarily target fungi-specific structures and biosynthetic processes.
Q Why is host toxicity a common problem with antiviral and antifungal drugs? Identify the targets that allow for selective toxicity of antifungal agents.
28.7 The use of antimicrobial drugs inevitably leads to resistance in the targeted microorganisms. The development of resistance can be accelerated by the indiscriminate use of antimicrobial drugs, and many pathogens have developed resistance. New antimicrobial compounds must continually be discovered and developed to deal with drug-resistant pathogens and to enhance our ability to treat infectious diseases. Computer-based modeling and other novel strategies are helping to address this challenge.
Q What practices contribute to the spread of antibiotic resistance? Explain how antisense RNA strategies can extend traditional methods of natural product selection for antibiotic discovery.
Application Questions
What problems would arise if a person had a hereditary deficiency that resulted in an inability to present antigens to Tc cells? What would the problems be if the person had a deficiency in presenting antigen to Th1 cells? To Th2 cells? To all T cells? What molecules might be deficient in each situation? Could a person having any one of these deficiencies survive in a normal environment? Explain for each.
Viruses and fungi present special problems for drug therapy. Explain the issues inherent in drug treatment of both groups and explain whether you agree with the preceding statement. Give specific examples and suggest at least one group of agents that might target both types of infectious agents.
Describe three important reasons why semisynthetic penicillins were first developed. Which clinical challenges do each of these reasons address? What key part of the penicillin molecule must be retained for any semisynthetic penicillin to be active?
Chapter Glossary
an antigen that elicits an immediate hypersensitivity (allergic) reaction upon exposure Aminoglycoside
an antibiotic, such as streptomycin, containing amino sugars linked by glycosidic bonds Antibiotic
a chemical substance produced by a microorganism that kills or inhibits the growth of another microorganism Antimicrobial drug resistance
the acquired ability of a microorganism to resist the effects of an antimicrobial agent to which it was formerly susceptible Autoantibody
an antibody that reacts to self antigens Autoimmunity
a harmful immune reaction directed against self antigens β-lactam antibiotic
penicillin, cephalosporin, or a related antibiotic that contains the four-membered heterocyclic β-lactam ring Delayed-type hypersensitivity (DTH)
an inflammatory allergic response mediated by Th1 lymphocytes Fusion inhibitor
a synthetic polypeptide that binds to viral glycoproteins, inhibiting fusion of viral and host cell membranes Growth factor analog
a chemical agent that has a similar structure to and blocks the uptake or utilization of a growth factor Hypersensitivity
an immune response leading to damage to host tissues Immediate hypersensitivity
an allergic response mediated by vasoactive products released from IgE-sensitized mast cells Immunotherapy
the treatment of a disease, especially cancer, using targeted therapeutic agents that either enhance or suppress the immune response Macrolide
erythromycin or a related antibiotic that contains a lactone ring bonded to sugars Nonnucleoside reverse transcriptase inhibitor (NNRTI)
a nonnucleoside compound that inhibits the action of retroviral reverse transcriptase by binding directly to the catalytic site Nucleoside reverse transcriptase inhibitor (NRTI)
a nucleoside analog compound that inhibits the action of retroviral reverse transcriptase by competing with nucleosides Protease inhibitor
a compound that inhibits the action of viral protease by binding directly to the catalytic site, preventing viral protein processing Quinolone
a synthetic antibacterial compound that inhibits DNA gyrase and prevents supercoiling of bacterial DNA Selective toxicity
the ability of a compound to inhibit or kill a pathogen without adversely affecting the host Sulfa drugs
synthetic growth factor analogs that inhibit folic acid biosynthesis in Bacteria Superantigen
a pathogen product capable of eliciting an inappropriately strong inflammatory immune response by stimulating greater than normal numbers of T cells Tetracycline
an antibiotic characterized by the four-member naphthacene ring structure Vaccination (immunization)
the inoculation of a host with inactive or weakened pathogens or pathogen products to stimulate protective active immunity Vaccine
an inactivated or attenuated pathogen, or a harmless pathogen product, used to induce artificial active immunity
Unit 7 Infectious Diseases
Chapter 31 Person-to-Person Bacterial and Viral Diseases
Chapter 32 Vectorborne and Soilborne Bacterial and Viral Diseases
Chapter 33 Waterborne and Foodborne Bacterial and Viral Diseases
Chapter 34 Eukaryotic Pathogens: Fungi, Protozoa, and Helminths