The Central Visual System
The Segregation of Input by Eye and by Ganglion Cell Type
Innervation of Other Cortical Layers from Layer IVC
BOX 10.2 BRAIN FOOD: Cortical Organization Revealed by Optical and Calcium Imaging
BOX 10.3 PATH OF DISCOVERY: Finding Faces in the Brain, by Nancy Kanwisher
BOX 10.4 OF SPECIAL INTEREST: The Magic of Seeing in 3D
Although our visual system provides us with a unified picture of the world around us, this picture has multiple facets. Objects we see have shape and color. They have a position in space, and sometimes they move. For us to see each of these properties, neurons somewhere in the visual system must be sensitive to them. Moreover, because we have two eyes, we actually have two visual images in our head, and somehow they must be merged.
In Chapter 9, we saw that in many ways the eye acts like a camera. But starting with the retina, the rest of the visual system is far more elaborate, far more interesting, and capable of doing far more than any camera. For example, we saw that the retina does not simply pass along information about the patterns of light and dark that fall on it. Rather, the retina extracts information about differences in brightness and color. There are roughly 100 million photoreceptors in the retina, but only 1 million axons leave the eye carrying information to the rest of the brain. What we perceive about the world around us, therefore, depends on what information is extracted by the output cells of the retina and how this information is analyzed and interpreted by the rest of the central nervous system (CNS). An example is color. There is no such thing as color in the physical world; there is simply a spectrum of visible wavelengths of light that are reflected by objects around us. Based on the information extracted by the three types of cone photoreceptors, however, our brain synthesizes a rainbow of colors and fills our world with it.
In this chapter, we explore how the information extracted by the retina is analyzed by the central visual system. The pathway serving conscious visual perception includes the lateral geniculate nucleus (LGN) of the thalamus and the primary visual cortex, also called area 17, V1, or the striate cortex. We will see that the information funneled through this geniculocortical pathway is processed in parallel by neurons specialized for the analysis of different stimulus attributes. The striate cortex then feeds this information to more than two dozen extrastriate cortical areas in the occipital, temporal, and parietal lobes, and many of these appear to be specialized for different types of analysis.
Much of what we know about the central visual system was first worked out in the domestic cat and then extended to the rhesus monkey, Macaca mulatta. The macaque monkey, as it is also called, relies heavily on vision for survival in its habitat, as do we humans. In fact, tests of the performance of this primate’s visual system show that in virtually all respects, it rivals that of humans. Thus, although most of this chapter concerns the organization of the macaque visual system most neuroscientists agree that it approximates very closely the situation in our own brain.
Visual neuroscience cannot yet explain every aspect of visual perception (Figure 10.1). However, significant progress has been made in answering a basic question: How do neurons represent the different facets of the visual world? By examining those stimuli that make different neurons in the visual cortex respond, and how these response properties arise, we begin to see how the brain portrays the visual world around us.

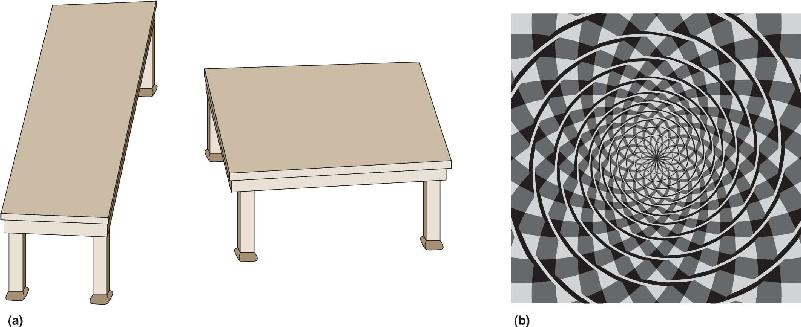
FIGURE 10.1 Perceptual illusions. (a) The two tabletops are of identical dimensions and are imaged on similarly sized patches of retina. To prove this to yourself, compare the vertical extent of the left table with the horizontal size of the right table. Because of the brain’s 3D interpretation of the 2D image, the perceived sizes are quite different. (b) This is an illusory spiral. Try tracing it with your finger. (Source: Part a adapted from R. Shepard, 1990, p. 48; part b adapted from J. Fraser, 1908.)
The neural pathway that leaves the eye, beginning with the optic nerve, is often referred to as the retinofugal projection. The suffix -fugal is from the Latin word meaning “to flee” and is commonly used in neuroanatomy to describe a pathway that is directed away from a structure. Thus, a centrifugal projection goes away from the center, a corticofugal projection goes away from the cortex, and the retinofugal projection goes away from the retina.
We begin our tour of the central visual system by looking at how the retinofugal projection courses from each eye to the brain stem on each side, and how the task of analyzing the visual world initially is divided among, and organized within, certain structures of the brain stem. Then, we focus on the major arm of the retinofugal projection that mediates conscious visual perception.
The ganglion cell axons “fleeing” the retina pass through three structures before they form synapses in the brain stem. The components of this retinofugal projection are, in order, the optic nerve, the optic chiasm, and the optic tract (Figure 10.2). The optic nerves exit the left and right eyes at the optic disks, travel through the fatty tissue behind the eyes in their bony orbits, then pass through holes in the floor of the skull. The optic nerves from both eyes combine to form the optic chiasm (named for the X shape of the Greek letter chi), which lies at the base of the brain, just anterior to where the pituitary gland dangles down.

FIGURE 10.2 The retinofugal projection. This view of the base of the brain shows the optic nerves, optic chiasm, and optic tracts.
At the optic chiasm, the axons originating in the nasal retinas cross from one side to the other. The crossing of a fiber bundle from one side of the brain to the other is called a decussation. Because only the axons originating in the nasal retinas cross, we say that a partial decussation of the retinofugal projection occurs at the optic chiasm. Following the partial decussation at the optic chiasm, the axons of the retinofugal projections form the optic tracts, which run just under the pia along the lateral surfaces of the diencephalon.
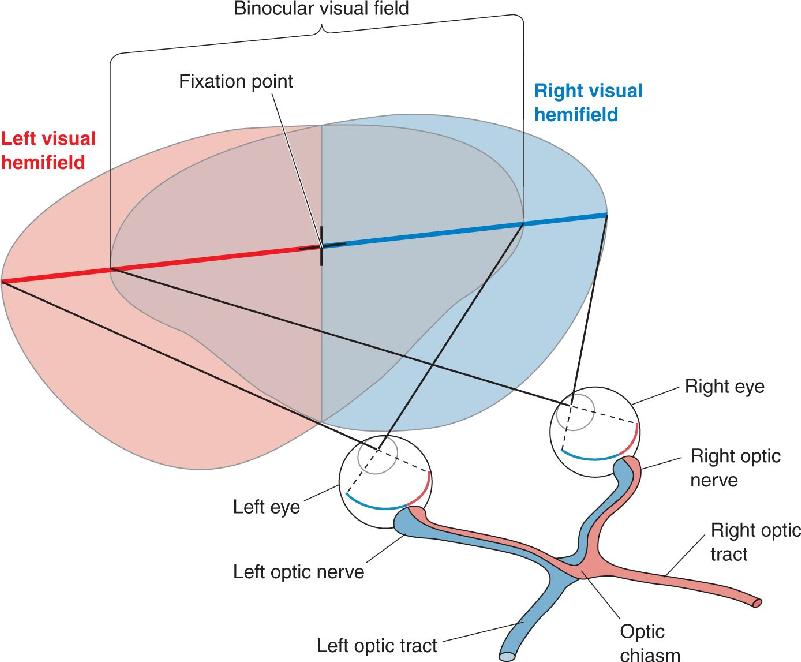
To understand the significance of the partial decussation of the retinofugal projection at the optic chiasm, let’s review the concept of the visual field introduced in Chapter 9. The full visual field is the entire region of space (measured in degrees of visual angle) that can be seen with both eyes looking straight ahead. Fix your gaze on a point straight ahead. Now imagine a vertical line passing through the fixation point, dividing the visual field into left and right halves. By definition, objects appearing to the left of the midline are in the left visual hemifield, and objects appearing to the right of the midline are in the right visual hemifield (Figure 10.3).

FIGURE 10.3 Right and left visual hemifields. Ganglion cells in both retinas that are responsive to visual stimuli in the right visual hemifield project axons into the left optic tract. Similarly, ganglion cells “viewing” the left visual hemifield project into the right optic tract. Description
By looking straight ahead with both eyes open and then alternately closing one eye and then the other, you will see that the central portion of both visual hemifields is viewed by both retinas. This region of space is therefore called the binocular visual field. Notice that objects in the binocular region of the left visual hemifield will be imaged on the nasal retina of the left eye and on the temporal retina of the right eye. Because the fibers from the nasal portion of the left retina cross to the right side at the optic chiasm, all the information about the left visual hemifield is directed to the right side of the brain. Remember this rule of thumb: Optic nerve fibers cross in the optic chiasm, such that the left visual hemifield is “viewed” by the right hemisphere and the right visual hemifield is “viewed” by the left hemisphere. You may recall from Chapter 7 that there is also a decussation in the descending pyramidal tract such that one side of the brain controls movement of the opposite side of the body. For reasons we do not understand, decussations are common in the sensory and motor systems.
A small number of optic tract axons peel off to form synaptic connections with cells in the hypothalamus, and another 10% or so continue past the thalamus to innervate the midbrain. But most of them innervate the lateral geniculate nucleus (LGN) of the dorsal thalamus. The neurons in the LGN give rise to axons that project to the primary visual cortex. This projection from the LGN to the cortex is called the optic radiation. Lesions anywhere in the retinofugal projection from the eye to the LGN to the visual cortex in humans cause blindness in part or all of the visual field. Therefore, we know that it is this pathway that mediates conscious visual perception (Figure 10.4).

FIGURE 10.4 The visual pathway that mediates conscious visual perception. (a) A side view of the brain with the retinogeniculocortical pathway shown inside (blue). (b) A horizontal section through the brain exposing the same pathway. Description
From our knowledge of how the visual world is represented in the retinofugal projection, we can predict the types of perceptual deficits that would result from its destruction at different levels, as might occur from a traumatic injury to the head, a tumor, or an interruption of the blood supply. As shown in Figure 10.5, while a transection of the left optic nerve would render a person blind in the left eye only, a transection of the left optic tract would lead to blindness in the right visual field as viewed through either eye. A midline transection of the optic chiasm would affect only the fibers that cross the midline. Because these fibers originate in the nasal portions of both retinas, blindness would result in the regions of the visual field viewed by the nasal retinas—that is, the peripheral visual fields on both sides (Box 10.1). Because unique deficits result from lesions at different sites, neuroophthalmologists and neurologists can locate sites of damage by assessing visual field deficits.

FIGURE 10.5 Visual field deficits from lesions in the retinofugal projection. (a) If the optic nerve on the left side is cut, vision will be lost completely in the left eye. Note that the resultant blindness is only in the monocular portion of the left hemifield because the right eye still sees most of the left visual field. (b) If the optic tract on the left side is cut, vision will be lost in the right visual field of each eye. (c) If the optic chiasm is split down the middle, only the crossing fibers will be damaged, and peripheral vision will be lost in both eyes. Description
Many of you are familiar with the famous story of David and Goliath, which appears in the Hebrew scriptures (Old Testament). The armies of the Philistines and the Israelites were gathered for battle when Goliath, a Philistine, came forth and challenged the Israelites to settle the dispute by sending out their best man to face him in a fight to the death. Goliath, it seems, was a man of great proportions, measuring more than “six cubits” in height. If you consider that a cubit is the distance from the elbow to the tip of the middle finger, about 20 inches, this guy was more than 10 feet tall! Goliath was armed to the teeth with body armor, a javelin, and a sword. To face this giant, the Israelites sent David, a young and diminutive shepherd, armed only with a sling and five smooth stones. Here’s how the action is described in the Revised Standard Version of the Bible (1 Samuel 17:48):
When the Philistine arose and came and drew near to meet David, David ran quickly toward the battle line to meet the Philistine. And David put his hand in his bag and took out a stone, and slung it, and struck the Philistine on his forehead; the stone sank into his forehead, and he fell on his face to the ground.
Now why, you might ask, are we giving a theology lesson in a neuroscience textbook? The answer is that our understanding of the visual pathway offers an explanation, in addition to divine intervention, for why Goliath was at a disadvantage in this battle. Body size is regulated by the secretion of growth hormone from the anterior lobe of the pituitary gland. In some cases, the anterior lobe becomes hypertrophied (swollen) and produces excessive amounts of the hormone, resulting in body growth to unusually large proportions. Such individuals are called pituitary giants and can be well over 8 feet tall.
Pituitary hypertrophy also disrupts normal vision. Recall that the optic nerve fibers from the nasal retinas cross in the optic chiasm, which butts up against the stalk of the pituitary. Any enlargement of the pituitary compresses these crossing fibers and results in a loss of peripheral vision called bitemporal hemianopia, or tunnel vision. (See if you can figure out why this is true from what you know about the visual pathway.) We can speculate that David was able to draw close and smite Goliath because the pituitary giant had completely lost sight of him.
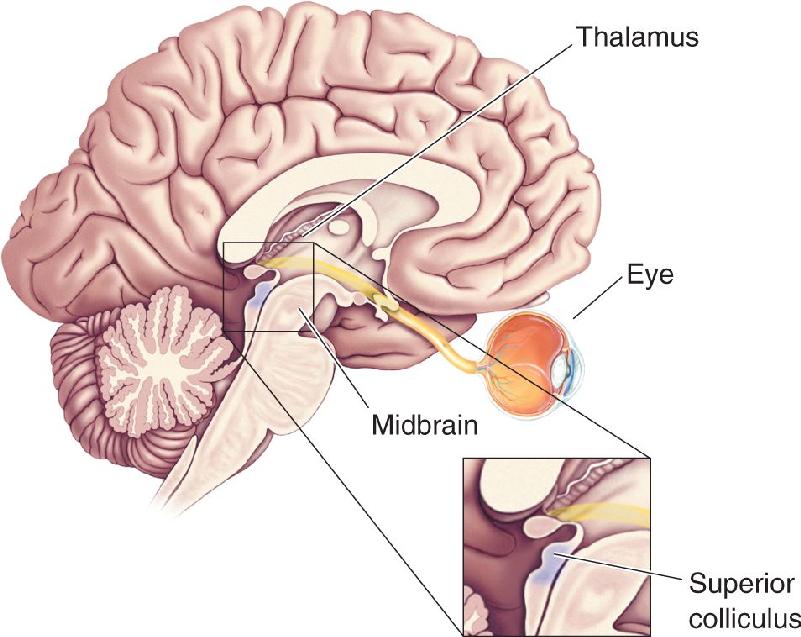
Nonthalamic Targets of the Optic Tract. As we have said, some retinal ganglion cells send axons to innervate structures other than the LGN. Direct projections to part of the hypothalamus play an important role in synchronizing a variety of biological rhythms, including sleep and wakefulness, with the daily dark–light cycle (see Chapter 19). Direct projections to part of the midbrain, called the pretectum, control the size of the pupil and certain types of eye movement. And about 10% of the ganglion cells in the retina project to a part of the midbrain tectum called the superior colliculus (Latin for “little hill”) (Figure 10.6).

FIGURE 10.6 The superior colliculus. Located in the tectum of the midbrain, the superior colliculus is involved in generating saccadic eye movements, the quick jumps in eye position used to scan across a page while reading. Description
While 10% may not sound like much of a projection, bear in mind that in primates, this is about 100,000 neurons, which is equivalent to the total number of retinal ganglion cells in a cat! In fact, the tectum of the midbrain is the major target of the retinofugal projection in all nonmammalian vertebrates (fish, amphibians, birds, and reptiles). In these vertebrate groups, the superior colliculus is called the optic tectum. This is why the projection from the retina to the superior colliculus is often called the retinotectal projection, even in mammals.
In the superior colliculus, a patch of neurons activated by a point of light, via indirect connections with motor neurons in the brain stem, commands eye and head movements to bring the image of this point in space onto the fovea. This branch of the retinofugal projection is thereby involved in orienting the eyes in response to new stimuli in the visual periphery. We saw in Chapter 9 that only the fovea has a dense concentration of cones sufficient for high-acuity vision. Therefore, it is critical that eye movements move our fovea to objects in our environment that might be threatening or of interest. We will return to the superior colliculus when we discuss motor systems in Chapter 14.
The right and left lateral geniculate nuclei, located in the dorsal thalamus, are the major targets of the two optic tracts. Viewed in cross section, each LGN appears to be arranged in six distinct layers of cells (Figure 10.7). By convention, the layers are numbered 1 through 6, starting with the most ventral layer, layer 1. In three dimensions, the layers of the LGN are arranged like a stack of six pancakes, one on top of the other. The pancakes do not lie flat, however; they are bent around the optic tract like a knee joint. This shape explains the name geniculate, from the Latin geniculatus, meaning “like a little knee.”

FIGURE 10.7 The LGN of the macaque monkey. The tissue has been stained to show cell bodies, which appear as purple dots. Notice particularly the six principal layers and the larger size of the cells in the two ventral layers (layers 1 and 2). (Source: Adapted from Hubel, 1988, p. 65.)
The LGN is the gateway to the visual cortex and, therefore, to conscious visual perception. Let’s explore the structure and function of this thalamic nucleus.
The Segregation of Input by Eye and by Ganglion Cell Type
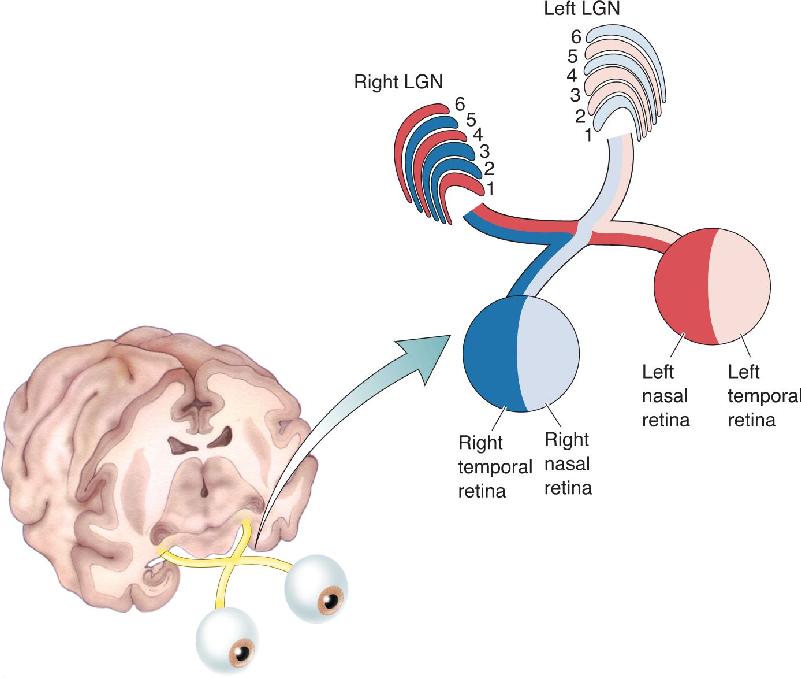
LGN neurons receive synaptic input from the retinal ganglion cells, and most geniculate neurons project an axon to the primary visual cortex via the optic radiation. The segregation of LGN neurons into layers suggests that different types of retinal information are being kept separate at this synaptic relay, and indeed this is the case: Axons arising from M-type, P-type, and nonM–nonP ganglion cells in the two retinas synapse on cells in different LGN layers.
Recall from our rule of thumb that the right LGN receives information about the left visual field. The left visual field is viewed by both the nasal left retina and the temporal right retina. At the LGN, input from the two eyes is kept separate. In the right LGN, the right eye (ipsilateral) axons synapse on LGN cells in layers 2, 3, and 5. The left eye (contralateral) axons synapse on cells in layers 1, 4, and 6 (Figure 10.8).

FIGURE 10.8 Retinal inputs to the LGN layers. The retinal axons project such that the LGN is excited by light in the contralateral visual field presented to either eye.
A closer look at the LGN in Figure 10.7 reveals that the two ventral layers, 1 and 2, contain larger neurons, and the four more dorsal layers, 3 through 6, contain smaller cells. The ventral layers are therefore called magnocellular LGN layers, and the dorsal layers are called parvocellular LGN layers. Recall from Chapter 9 that ganglion cells in the retina may also be classified into magnocellular and parvocellular groups. As it turns out, P-type ganglion cells in the retina project exclusively to the parvocellular LGN, and M-type ganglion cells in the retina project entirely to the magnocellular LGN.
In addition to the neurons in the six principal layers of the LGN, it was later discovered that there are numerous tiny neurons that lie just ventral to each layer. Cells in these koniocellular LGN layers (konio is from the Greek for “dust”), sometimes referred to as layers K1–K6, receive input from the nonM–nonP types of retinal ganglion cells and also project to the visual cortex. For the most part, each koniocellular layer gets input from the same eye as the overlying M or P layer. For example, layer K1 receives input from the contralateral eye just as layer 1 neurons do. In Chapter 9, we saw that in the retina, M-type, P-type, and nonM–nonP ganglion cells respond differently to light and color. In the LGN, the different information derived from the three categories of retinal ganglion cells from the two eyes remains largely segregated.
The anatomical organization of the LGN supports the idea that the retina gives rise to streams of information that are processed in parallel. This organization is summarized in Figure 10.9.

FIGURE 10.9 The organization of the LGN. (a) Ganglion cell inputs to the different LGN layers. (b) A thin koniocellular layer (shown in pink) is ventral to each of the six principal layers. Description
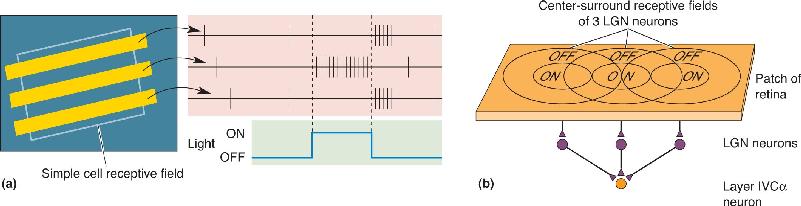
In Figure 9.25, we saw how the receptive field of a retinal ganglion cell can be mapped out by recording from the neuron while spots of light are shone on the retina. Similarly, by inserting a microelectrode into the LGN, it is possible to study the action potential discharges of a geniculate neuron in response to visual stimuli and map its receptive field. The surprising conclusion of such studies is that the visual receptive fields of LGN neurons are almost identical to those of the ganglion cells that feed them. For example, magnocellular LGN neurons have relatively large center-surround receptive fields, respond to stimulation of their receptive field centers with a transient burst of action potentials, and are insensitive to differences in wavelength. All in all, they are just like M-type ganglion cells. Likewise, parvocellular LGN cells, like P-type retinal ganglion cells, have relatively small center-surround receptive fields and respond to stimulation of their receptive field centers with a sustained increase in the frequency of action potentials; many of them exhibit color opponency. Receptive fields of cells in the koniocellular layers are center-surround and have either light/dark or color opponency. Within all layers of the LGN, the neurons are activated by only one eye (i.e., they are monocular) and ON-center and OFF-center cells are intermixed.
What makes the similarity of LGN and ganglion cell receptive fields so surprising is that the retina is not the main source of synaptic input to the LGN. In addition to the retina, the LGN receives inputs from other parts of the thalamus and the brain stem. The major input, constituting about 80% of the excitatory synapses, comes from primary visual cortex. Thus, one might reasonably expect that this corticofugal feedback pathway would significantly alter the qualities of the visual responses recorded in the LGN. So far, however, a role for this massive input has not been clearly identified. One hypothesis is that “top–down” modulation from the visual cortex to the LGN gates subsequent “bottom-up” input from the LGN back to the cortex. For example, if we want to selectively pay attention to a portion of our visual field, we might be able to suppress inputs coming from outside the attended area. We’ll have more to say about this in our discussion of attention in Chapter 21.
The LGN also receives synaptic inputs from neurons in the brain stem whose activity is related to alertness and attentiveness (see Chapters 15 and 19). Have you ever “seen” a flash of light when you are startled in a dark room? This perceived flash might be a result of the direct activation of LGN neurons by this pathway. Usually, however, this input does not directly evoke action potentials in LGN neurons. But it can powerfully modulate the magnitude of LGN responses to visual stimuli. (Recall modulation from Chapters 5 and 6.) Thus, the LGN is more than a simple relay from the retina to the cortex; it is the first site in the ascending visual pathway where what we see is influenced by how we feel.
The LGN has a single major synaptic target: the primary visual cortex. Recall from Chapter 7 that the cortex may be divided into a number of distinct areas based on their connections and cytoarchitecture. The primary visual cortex is Brodmann’s area 17 and is located in the occipital lobe of the primate brain. Much of area 17 lies on the medial surface of the hemisphere, surrounding the calcarine fissure (Figure 10.10). Other terms used interchangeably to describe the primary visual cortex are V1 and the striate cortex. (The term striate refers to the fact that area V1 has an unusually dense stripe of myelinated axons running parallel to the surface that appears white in unstained sections.)

FIGURE 10.10 The primary visual cortex. Top views are lateral; bottom views are medial.
We have seen that the axons of different types of retinal ganglion cells synapse on anatomically segregated neurons in the LGN. In the following sections, we look at the anatomy of the striate cortex and trace the connections different LGN cells make with cortical neurons. Later, we’ll explore how this information is analyzed by cortical neurons. As we did in the LGN, in the striate cortex we’ll see a close correlation between structure and function.
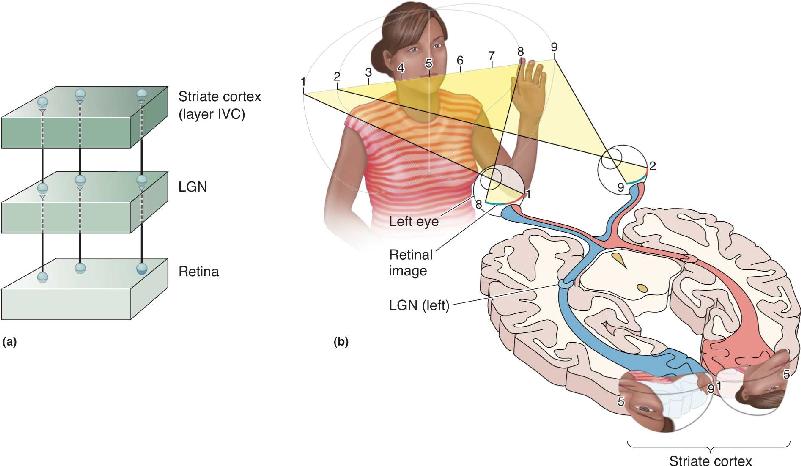
The projection starting in the retina and extending to the LGN and V1 illustrates a general organizational feature of the central visual system called retinotopy. Retinotopy is an organization whereby neighboring cells in the retina feed information to neighboring places in their target structures—in this case, the LGN and striate cortex. In this way, the two-dimensional surface of the retina is mapped onto the two-dimensional surface of the subsequent structures (Figure 10.11a).

FIGURE 10.11 The retinotopic map in the striate cortex. (a) Neighboring locations on the retina project to neighboring locations in the LGN. This retinotopic representation is preserved in the LGN projection to V1. (b) The lower portion of V1 represents the top half of visual space, and the upper portion of V1 represents the bottom half of visual space. Notice also that the map is distorted, with more tissue devoted to analysis of the central visual field. Similar maps are found in the superior colliculus, LGN, and other visual cortical areas. Description
There are three important points to remember about retinotopy. First, the mapping of the visual field onto a retinotopically organized structure is often distorted because visual space is not sampled uniformly by the cells in the retina. Recall from Chapter 9 that there are many more ganglion cells with receptive fields in or near the fovea than in the periphery. Corresponding to this, the representation of the visual field is distorted in the striate cortex: The central few degrees of the visual field are overrepresented, or magnified, in the retinotopic map (Figure 10.11b). In other words, there are many more neurons in the striate cortex that receive input from the central retina than from the peripheral retina.
The second point to remember is that a discrete point of light can activate many cells in the retina, and often many more cells in the target structure, due to the overlap of receptive fields. The image of a point of light on the retina actually activates a large population of cortical neurons; every neuron that contains that point in its receptive field is potentially activated. Thus, when the retina is stimulated by a point of light, the activity in the striate cortex is a broad distribution with a peak at the corresponding retinotopic location.
Finally, don’t be misled by the word “map.” There are no pictures in the primary visual cortex for a little person in our brain to look at. While it’s true that the arrangement of connections establishes a mapping between the retina and V1, perception is based on the brain’s interpretation of distributed patterns of activity, not literal snapshots of the world. (We discuss visual perception later in this chapter.)
The neocortex in general, and the striate cortex in particular, have neuronal cell bodies arranged into about a half-dozen layers. These layers can be seen clearly in a Nissl stain of the cortex, which, as described in Chapter 2, leaves a deposit of dye (usually blue or violet) in the soma of each neuron. Starting at the white matter (containing the cortical input and output fibers), the cell layers are named by Roman numerals VI, V, IV, III, and II. Layer I, just under the pia mater, is largely devoid of neurons and consists almost entirely of axons and dendrites of cells in other layers (Figure 10.12). The full thickness of the striate cortex from white matter to pia is about 2 mm, the height of the lowercase letter m.

FIGURE 10.12 The cytoarchitecture of the striate cortex. The tissue has been Nissl stained to show cell bodies, which appear as purple dots. (Source: Adapted from Hubel, 1988, p. 97.) Description
As Figure 10.12 shows, describing the lamination of the striate cortex as a six-layer scheme is somewhat misleading. There are actually at least nine distinct layers of neurons. To maintain Brodmann’s convention that the neocortex has six layers, however, neuroanatomists combine three sublayers into layer IV, labeled IVA, IVB, and IVC. Layer IVC is further divided into two tiers called IVCα and IVCβ. The anatomical segregation of neurons into layers suggests that there is a division of labor in the cortex, similar to what we saw in the LGN. We can learn a lot about how the cortex handles visual information by examining the structure and connections of its different layers.
The Cells of Different Layers. Many different neuronal shapes have been identified in striate cortex, but here we focus on two principal types, defined by the appearance of their dendritic trees (Figure 10.13). Spiny stellate cells are small neurons with spine-covered dendrites that radiate out from the cell body (recall dendritic spines from Chapter 2). They are seen primarily in the two tiers of layer IVC. Outside layer IVC are many pyramidal cells. These neurons are also covered with spines and are characterized by a single thick apical dendrite that branches as it ascends toward the pia mater and by multiple basal dendrites that extend horizontally. In Figure 10.13, the axon is the single neurite descending from the soma of each pyramidal cell.

FIGURE 10.13 The dendritic morphology of some cells in the striate cortex. Pyramidal cells are found in layers III, IVB, V, and VI, and spiny stellate cells are found in layer IVC. Description
Notice that a pyramidal cell in one layer may have dendrites extending into other layers. For the most part, only pyramidal cells send axons out of the striate cortex to form connections with other parts of the brain. The axons of stellate cells, which are indistinguishable from dendrites in Figure 10.13, generally make local connections only within the cortex. An exception to this rule is spiny stellate neurons in layer IVB that project to area V5, which we will discuss shortly.
In addition to the spiny neurons, inhibitory neurons, which lack spines, are sprinkled in all cortical layers as well. These neurons form only local connections.
The distinct lamination of the striate cortex is reminiscent of the layers we saw in the LGN. In the LGN, every layer receives retinal afferents and sends efferents to the visual cortex. In the visual cortex, the situation is different; only a subset of the layers receives input from the LGN or sends output to a different cortical or subcortical area.
Axons from the LGN terminate in several different cortical layers, with the largest number going to layer IVC. We’ve seen that the output of the LGN is divided into streams of information, for example, from the magnocellular and parvocellular layers serving the right and left eyes. These streams remain anatomically segregated in layer IVC.
Magnocellular LGN neurons project primarily to layer IVCα, and parvocellular LGN neurons project to layer IVCβ. Imagine that the two tiers of layer IVC are pancakes, stacked one (α) on top of the other (β). Because the input from the LGN to the cortex is arranged topographically, we see that layer IVC contains two overlapping retinotopic maps, one from the magnocellular LGN (IVCα) and the other from the parvocellular LGN (IVCβ). Koniocellular LGN axons follow a different path, making synapses primarily in layers II and III.
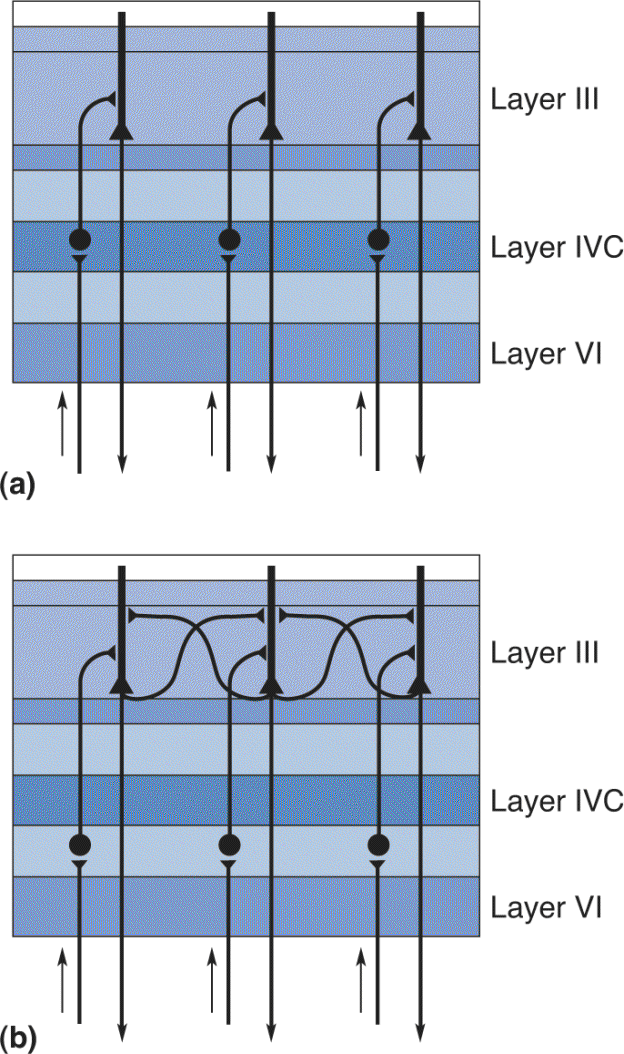
Innervation of Other Cortical Layers from Layer IVC. Most intracortical connections extend perpendicular to the cortical surface along radial lines that run across the layers, from white matter to layer I. This pattern of radial connections maintains the retinotopic organization established in layer IV. Therefore, a cell in layer VI, for example, receives information from the same part of the retina as does a cell above it in layer IV (Figure 10.14a). However, the axons of some layer III pyramidal cells extend collateral branches that make horizontal connections within layer III (Figure 10.14b). Radial and horizontal connections play different roles in the analysis of the visual world, as we’ll see later in the chapter.

FIGURE 10.14 Patterns of intracortical connections. (a) Radial connections. (b) Horizontal connections. Description
Leaving layer IV, there continues to be considerable anatomical segregation of the magnocellular and parvocellular processing streams. Layer IVCα, which receives magnocellular LGN input, projects mainly to cells in layer IVB. Layer IVCβ, which receives parvocellular LGN input, projects mainly to layer III. In layers III and IVB, an axon may form synapses with the dendrites of pyramidal cells of all layers.
Ocular Dominance Columns. How are the left eye and right eye LGN inputs arranged when they reach the striate cortex? Do they randomly intermix, or are they kept segregated? The answer was provided by a ground-breaking experiment performed in the early 1970s at Harvard Medical School by neuroscientists David Hubel and Torsten Wiesel. They injected a radioactive amino acid into one eye of a monkey (Figure 10.15). This amino acid was incorporated into proteins by the ganglion cells, and the proteins were transported down the ganglion cell axons into the LGN (recall anterograde transport from Chapter 2). Here, the radioactive proteins spilled out of the ganglion cell axon terminals and were taken up by nearby LGN neurons. But not all LGN cells took up the radioactive material; only those cells that were postsynaptic to the inputs from the injected eye incorporated the labeled protein. These cells then transported the radioactive proteins to their axon terminals in layer IVC of striate cortex. The location of the radioactive axon terminals was visualized by first placing a piece of film over thin sections of striate cortex and later developing the film like a photograph, a process called autoradiography (introduced in Chapter 6). The resulting collection of silver grains on the film marked the location of the radioactive LGN inputs.

FIGURE 10.15 Transneuronal autoradiography. Radioactive proline is ① injected into one eye, where it is ② taken up by retinal ganglion cells and incorporated into proteins that are ③ transported down the axons to the LGN. Some radioactivity spills out of the retinal terminals and is ④ taken up by LGN neurons that then ⑤ transport it to the striate cortex. The location of radioactivity can be determined using autoradiography. Description
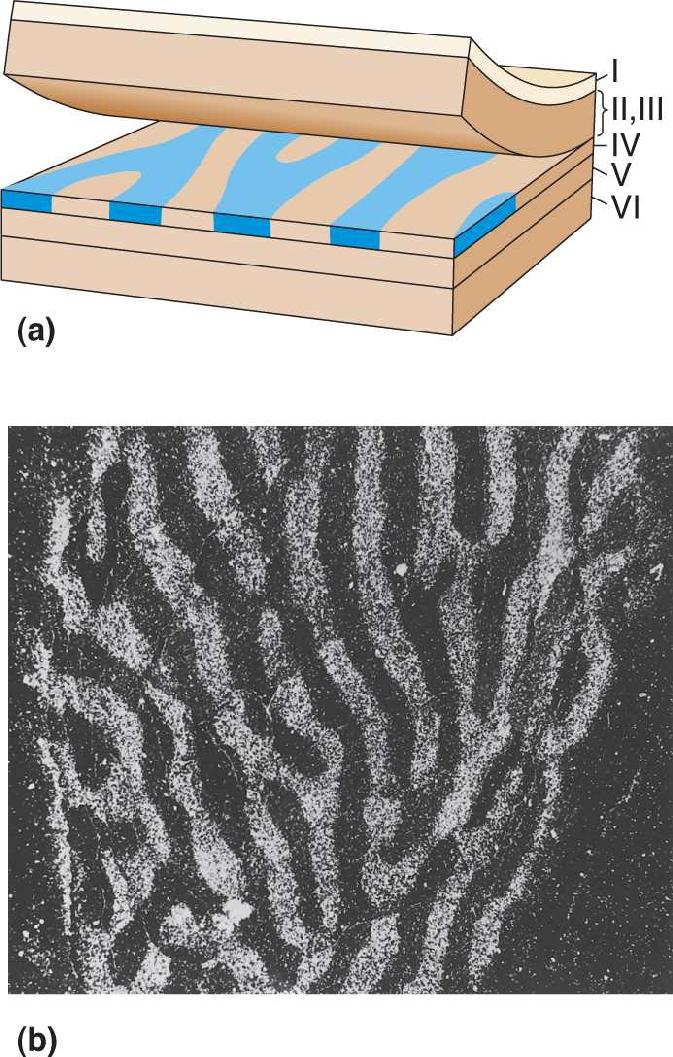
In sections cut perpendicular to the cortical surface, Hubel and Wiesel observed that the distribution of axon terminals relaying information from the injected eye was not continuous in layer IVC, but rather was split up into a series of equally spaced patches, each about 0.5 mm wide (Figure 10.16a). In later experiments, the cortex was sectioned tangentially, parallel to layer IV. This revealed that the left eye and right eye inputs to layer IV are laid out as a series of alternating bands, like the stripes of a zebra (Figure 10.16b). Rather than randomly mixing, neurons connected to the left and right eyes are as distinct in layer IV as they are in the LGN.

FIGURE 10.16 Ocular dominance columns in layer IV of the striate cortex. (a) The organization of ocular dominance columns in layer IV of macaque monkey striate cortex. The distribution of LGN axons serving one eye is shaded blue. In cross section (looking at layer IV from the side), these eye-specific zones appear as patches, each about 0.5 mm wide, in layer IV. Peeled-back layers reveal that the ocular dominance columns in layer IV look like zebra stripes. (b) An autoradiograph of a histological section of layer IV viewed from above. Two weeks prior to the experiment, one eye of this monkey was injected with radioactive proline. In the autoradiograph, the radioactive LGN terminals appear bright on a dark background. (Source: LeVay et al., 1980.) Description
Layer IVC stellate cells project axons radially up mainly to layers IVB and III where, for the first time, information from the left eye and right eye begins to mix (Figure 10.17). Whereas all layer IVC neurons receive input from only one eye, most neurons in layers II, III, V, and VI receive some amount of input from each eye. For example, a neuron above a left eye patch of neurons in layer IVC receives input from both left eye and right eye neurons in layer IVC, but more of the projections come from the left eye. It is said that the input to the neuron is “dominated” by the left eye. In Figure 10.17, the red and blue patches of cells in layer III are dominated by the right and left eye, respectively; the purple patches of neurons receive roughly equal input from the two eyes. Because of the alternating patches of left eye and right eye input reaching layer IV and the overall radial projections, neurons outside layer IV are organized into alternating bands dominated by the left and right eye. The bands of cells extending through the thickness of the striate cortex are called ocular dominance columns.

FIGURE 10.17 The mixing of information from the two eyes. Axons project from layer IVC to more superficial layers. Most layer III neurons receive binocular input from both left and right eyes. There are layer III neurons with responses dominated by the right eye (red), left eye (blue), or roughly equally responsive to input from the two eyes (purple). Because of the radial connectivity in striate cortex, neurons in layers above and below layer IV are dominated by the same eye. Ocular dominance columns (between vertical dotted lines) contain neurons with input dominated by one eye, and the columns alternate between left and right eye dominance. Description
Striate Cortex Outputs. As previously mentioned, the pyramidal cells send axons out of the striate cortex into the white matter. The pyramidal cells in different layers innervate different structures. Layer II, III, and IVB pyramidal cells send their axons to other cortical areas. Layer V pyramidal cells send axons all the way down to the superior colliculus and pons. Layer VI pyramidal cells give rise to the massive axonal projection back to the LGN (Figure 10.18). Pyramidal cell axons in all layers also branch and form local connections in the cortex.

FIGURE 10.18 Outputs from the striate cortex. Description
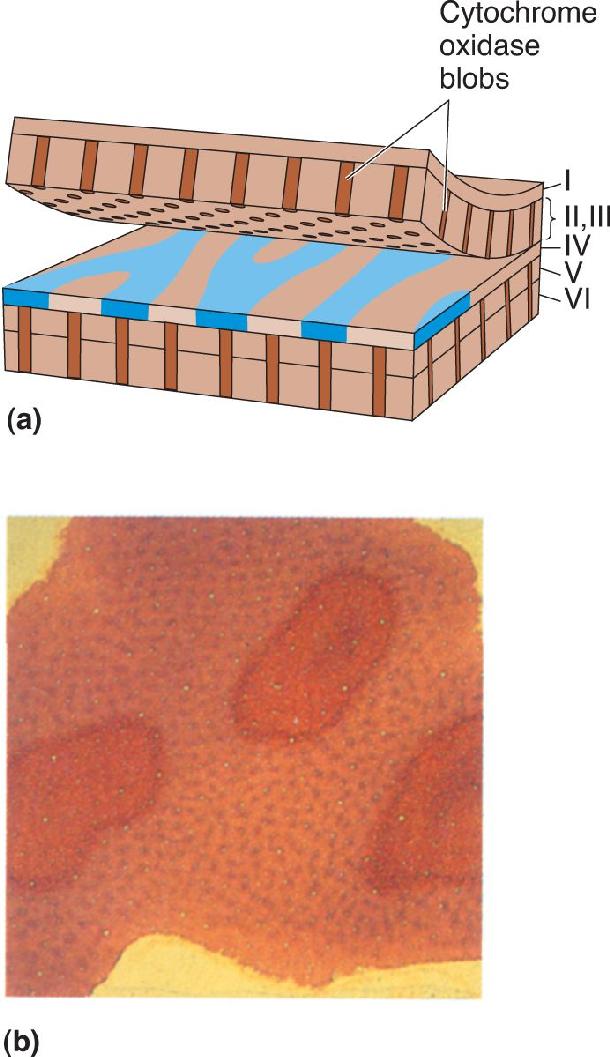
As we have seen, layers II and III play a key role in visual processing, providing most of the information that leaves V1 for other cortical areas. Anatomical studies suggest that the V1 output comes from two distinct populations of neurons in the superficial layers. When striate cortex tissue is stained to reveal the presence of cytochrome oxidase, a mitochondrial enzyme used for cell metabolism, the stain is not uniformly distributed in layers II and III. Rather, the cytochrome oxidase staining in cross sections of striate cortex appears as a colonnade, a series of pillars at regular intervals, running the full thickness of layers II and III and also in layers V and VI (Figure 10.19a). When the cortex is sliced tangentially through layer III, these pillars appear like the spots of a leopard (Figure 10.19b). These pillars of cytochrome oxidase–rich neurons have come to be called blobs. The blobs are in rows, each blob centered on an ocular dominance stripe in layer IV. Between the blobs are “interblob” regions. The blobs receive direct LGN input from the koniocellular layers, as well as parvocellular and magnocellular input from layer IVC of striate cortex.

FIGURE 10.19 Cytochrome oxidase blobs. (a) The organization of cytochrome oxidase blobs in macaque monkey striate cortex. The tissue that stains darkly for cytochrome oxidase looks like radial pillars in layers II, III, V, and VI. A cross section tangential to the surface shows the discrete patches that gave blobs their name (underside of layer III). (b) A photograph of a histological section of layer III, stained for cytochrome oxidase and viewed from above. The dark spots are cytochrome oxidase blobs. (Source: Courtesy of Dr. S.H.C. Hendry.) Description
Beginning in the early 1960s, Hubel and Wiesel were the first to systematically explore the physiology of the striate cortex with microelectrodes. They were students of Stephen Kuffler, who was then at Johns Hopkins University and later moved with them to Harvard. They extended Kuffler’s innovative methods of receptive field mapping to the central visual pathways. After showing that LGN neurons behave much like retinal ganglion cells, they turned their attention to the striate cortex, initially in cats and later in monkeys. (Here we focus on the monkey cortex.) The work that continues today on the physiology of the striate cortex is built on the solid foundation provided by Hubel and Wiesel’s pioneering studies. Their contributions to our understanding of the cerebral cortex were recognized with the Nobel Prize in 1981.
By and large, the receptive fields of neurons in layer IVC are similar to the magnocellular and parvocellular LGN neurons providing their input. This means they are generally small monocular center-surround receptive fields. In layer IVCα the neurons are insensitive to the wavelength of light, whereas in layer IVCβ the neurons exhibit center-surround color opponency. Outside layer IVC (and somewhat within), new receptive field characteristics, not observed in the retina or LGN, are present. We will explore these in some depth because they provide clues about the role V1 plays in visual processing and perception.
Binocularity. There is a direct correspondence between the arrangement of connections in V1 and the responses of the neurons to light in the two eyes. Each neuron in layers IVCα and IVCβ receives afferents from a layer of the LGN representing the left or right eye. Physiological recordings confirm that these neurons are monocular, responding to light only in one of the eyes. We have already seen that the axons leaving layer IVC diverge and innervate more superficial cortical layers, mixing the inputs from the two eyes (see Figure 10.17). Microelectrode recordings confirm this anatomical fact; most neurons in layers superficial to IVC are binocular, responding to light in either eye. The ocular dominance columns demonstrated with autoradiography are reflected in the responses of V1 neurons. Above the centers of ocular dominance patches in layer IVC, the layer II and III neurons are more strongly driven by the eye represented in layer IVC (i.e., their response is dominated by one eye even though they are binocular). In areas where there is more equal mixing of left eye and right eye projections from layer IVC, the superficial layer neurons respond about the same to light in either eye.
We say that the neurons have binocular receptive fields, meaning that they actually have two receptive fields, one in the ipsilateral eye and one in the contralateral eye. Retinotopy is preserved because the two receptive fields of a binocular neuron are precisely placed on the retinas such that they are “looking” at the same point in in the contralateral visual field. The construction of binocular receptive fields is essential in binocular animals, such as humans. Without them, we would probably be unable to use the inputs from both eyes to form a single image of the world around us and perform fine motor tasks that require stereoscopic vision, such as threading a needle.
Orientation Selectivity. Most of the receptive fields in the retina, LGN, and layer IVC are circular and give their greatest response to a spot of light matched in size to the receptive field center. Outside layer IVC, we encounter cells that no longer follow this pattern. While small spots can elicit a response from many cortical neurons, it is usually possible to produce a much greater response with other stimuli. Rather by accident, Hubel and Wiesel found that many neurons in V1 respond best to an elongated bar of light moving across their receptive fields. But the orientation of the bar is critical. The greatest response is given to a bar with a particular orientation; bars perpendicular to the optimal orientation generally elicit much weaker responses (Figure 10.20). Neurons having this type of response are said to exhibit orientation selectivity. Most of the V1 neurons outside layer IVC (and some within) are orientation selective. The optimal orientation for a neuron can be any angle around the clock.

FIGURE 10.20 Orientation selectivity. (a) The responses of an orientation-selective neuron are monitored as visual stimuli are presented in its receptive field. The visual stimulus is a bar of light. (b) Light bars of various orientations (left) elicit very different responses (right). The optimal orientation for this neuron is 45° counterclockwise from vertical. Description
If V1 neurons can have any optimal orientation, you might wonder whether the orientation selectivity of nearby neurons is related. From the earliest work of Hubel and Wiesel, the answer to this question was an emphatic yes. As a microelectrode is advanced radially (perpendicular to the surface) from one layer to the next, the preferred orientation remains the same for all the selective neurons encountered from layer II down through layer VI. Hubel and Wiesel called such a radial column of cells an orientation column.
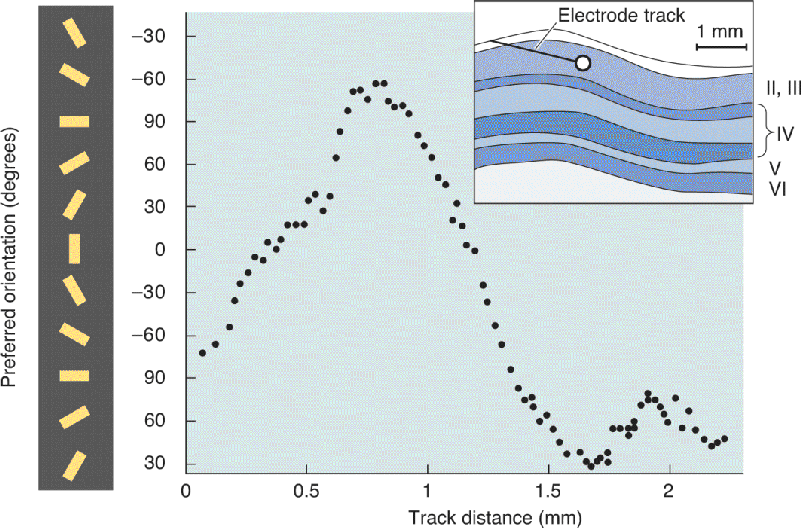
As an electrode passes tangentially (parallel to the surface) through the cortex in a single layer, the preferred orientation progressively shifts. We now know, from the use of a technique called optical imaging, that there is a mosaic-like pattern of optimal orientations in the striate cortex (Box 10.2). If an electrode is passed at certain angles through this mosaic, the preferred orientation rotates like the sweep of the minute hand of a clock, from the top of the hour to 10 past to 20 past, and so on (Figure 10.21). If the electrode is moved at other angles, more sudden shifts in preferred orientation occur. Hubel and Wiesel found that a complete 180° shift in preferred orientation required a traverse of about 1 mm, on average, within layer III.

FIGURE 10.21 Systematic variation of orientation preferences across the striate cortex. As an electrode is advanced tangentially across layers II and III of striate cortex, the orientation preference of the neurons encountered is recorded and plotted. In the recording shown, the preferred orientation of the neuron first encountered was near –70°, and as the electrode moved 0.7 mm, the preferred orientations rotated clockwise. As the electrode moved another millimeter, the preferred orientations rotated counterclockwise. (Source: Adapted from Hubel and Wiesel, 1968.) Description
Cortical Organization Revealed by Optical and Calcium Imaging
Most of what we know about the response properties of neurons in the visual system, and every other system in the brain, has been learned from intracellular and extracellular recordings with microelectrodes. These recordings give precise information about the activity of one or a few cells. However, unless one inserts thousands of electrodes, it is not possible to observe patterns of activity across large populations of neurons.
A view of neural coding at a scale much larger than individual neurons is provided by optical imaging of brain activity. In one version of optical recording, a voltage-sensitive dye is applied to the surface of the brain. The molecules in the dye bind to cell membranes, and an array of photodetectors or a video camera records changes in the optical properties that are proportional to variations in membrane potential. A second way to optically study cortical activity is to image intrinsic signals. When neurons are active, blood volume and oxygenation change to a degree correlated with neural activity. Blood flow and oxygenation influence the reflection of light from brain tissue, and reflectance changes can be used to indirectly assess neural activity. Light is projected onto the brain, and a video camera records the reflected light. Thus, when intrinsic signals are used to study brain activity, membrane potentials for action potentials are not directly measured.
Figure A is a photograph showing the vasculature in a portion of primary visual cortex. Figure B shows ocular dominance columns in the same patch of striate cortex obtained by optically imaging areas in which blood flow changes occurred during visual stimulation. This figure is actually a subtraction of two images—one made when only the right eye was visually stimulated, minus another when only the left eye was stimulated. Consequently, the dark bands represent cells dominated by the left eye, and the light bands represent cells dominated by the right eye. Figure C is a color-coded representation of preferred orientation in the same patch of striate cortex. Four different optical images were recorded while bars of light at four different orientations were swept across the visual field. Each location in the figure is colored according to the orientation that produced the greatest response at each location on the brain (blue = horizontal; red = 45°; yellow = vertical; green = 135°). Consistent with earlier results obtained with electrodes (see Figure 10.21), in some regions, the orientation changes progressively along a straight line. However, the optical recording technique reveals that cortical organization based on orientation is much more complex than an idealized pattern of parallel “columns.”

Figure A Vasculature on the surface of primary visual cortex. (Source: Ts’o et al., 1990, Fig. 1A.)

Figure B Intrinsic signal imaging map of ocular dominance columns. (Source: Ts’o et al., 1990, Fig. 1B.)

Figure C Intrinsic signal imaging map of preferred orientations. (Source: Ts’o et al., 1990, Fig. 1C.)
Another technique, in vivo two-photon calcium imaging, lets us see the activity of thousands of neurons with single-cell resolution. When a neuron fires action potentials, voltage-sensitive calcium channels open, and calcium concentration in the soma increases. These concentration changes can be measured by introducing a calcium-sensitive fluorescent dye into the neurons; the amount of fluorescent light emitted from the neuron is correlated with the amount of calcium in the cell body and, thus, the firing rate of the neuron. To see neural activity at fine spatial and temporal scales, two-photon microscopy is used. The top of Figure D shows a map of orientation preference obtained with intrinsic signal optical imaging from cat visual cortex. The bottom of Figure D shows the orientation preferences of individual neurons based on two-photon calcium imaging. Orientation columns are seen in the clumping of cells with the same color; the results confirm that the optical images result from highly consistent cell-to-cell preferences for orientation. Cells with progressively different orientation preferences are organized in a “pinwheel” fashion, confirmation at the level of single cells of the optical results.

Figure D A map of preferred orientation based on intrinsic signal optical imaging (top). Two-photo calcium imaging shows the orientation preference of individual neurons (bottom). (Source: Adapted from Ohki and Reid, 2006, Fig. 1.)
The analysis of stimulus orientation appears to be one of the most important functions of the striate cortex. Orientation-selective neurons are thus thought to be specialized for the analysis of object shape.
Direction Selectivity. Many V1 receptive fields also exhibit direction selectivity; they respond when a bar of light at the optimal orientation moves perpendicular to the orientation in one direction but not in the opposite direction. Direction-selective cells in V1 are a subset of the cells that are orientation selective. Figure 10.22 shows how a direction-selective cell responds to a moving stimulus. Notice in this example that the cell responds to an elongated stimulus swept rightward across the receptive field, but much less with leftward movement. Sensitivity to the direction of stimulus motion is a hallmark of neurons receiving input from the magnocellular layers of the LGN. Direction-selective neurons are thought to be specialized for the analysis of object motion.

FIGURE 10.22 Direction selectivity. With a bar stimulus at the optimal orientation, the neuron responds strongly when the bar is swept to the right but weakly when it is swept to the left.
Simple and Complex Receptive Fields. Neurons in the LGN have antagonistic center-surround receptive fields, and this organization accounts for the responses of these neurons to visual stimuli. For example, a small spot in the center of the receptive field may yield a much stronger response than a larger spot also covering the antagonistic surround. What do we know about the inputs to V1 neurons that might account for binocularity, orientation selectivity, and direction selectivity in their receptive fields? Binocularity is easy; we have seen that binocular neurons receive afferents from both eyes. The mechanisms underlying orientation and direction selectivity have proven more difficult to elucidate.
Many orientation-selective neurons have a receptive field elongated along a particular axis, with an ON-center or OFF-center region flanked on one or both sides by an antagonistic surround (Figure 10.23a). This linear arrangement of ON and OFF areas is analogous to the concentric antagonistic areas seen in retinal and LGN receptive fields. One gets the impression that the cortical neurons receive a converging input from LGN cells with receptive fields that are aligned along one axis (Figure 10.23b). Hubel and Wiesel called neurons of this type simple cells. The segregation of ON and OFF regions is a defining property of simple cells, and it is because of this receptive field structure that they are orientation selective.

FIGURE 10.23 A simple cell receptive field. (a) The response of a simple cell to optimally oriented bars of light at different locations in the receptive field. Notice that the response can be ON or OFF depending on where the bar lies in the receptive field. For this neuron, the middle location gives an ON response and the two flanking locations give OFF responses. (b) A simple cell receptive field might be built from convergent inputs of three LGN neurons with aligned center-surround receptive fields. Description
Other orientation-selective neurons in V1 do not have distinct ON and OFF regions and are therefore not considered simple cells. Hubel and Wiesel called most of these complex cells, because their receptive fields appeared to be more complex than those of simple cells. Complex cells give ON and OFF responses to stimuli throughout the receptive field (Figure 10.24). Hubel and Wiesel proposed that complex cells are constructed from the input of several like-oriented simple cells. However, this remains a matter of debate.

FIGURE 10.24 A complex cell receptive field. Like a simple cell, a complex cell responds best to a bar of light at a particular orientation. However, responses occur to both light ON and light OFF, regardless of position in the receptive field. Description
Simple and complex cells are typically binocular and sensitive to stimulus orientation. Different neurons show a range of sensitivities to color and direction of motion.
Blob Receptive Fields. We have seen repeatedly in the visual system that when two nearby structures label differently with some anatomical technique, there is good reason to suspect the neurons in the structures are functionally different. For example, we have seen how the distinctive layers of the LGN segregate different types of input. Similarly, the lamination of striate cortex correlates with differences in the receptive fields of the neurons. The presence of the distinct cytochrome oxidase blobs outside layer IV of striate cortex immediately raises the question of whether the neurons in the blobs respond differently from interblob neurons. The answer is controversial. The neurons in the interblob areas have some or all of the properties we discussed above: binocularity, orientation selectivity, and direction selectivity. They include both simple cells and complex cells; some are wavelength sensitive and some are not. The blobs receive input directly from the koniocellular layers of the LGN and magnocellular and parvocellular input via layer IVC. Early studies reported that blob cells, unlike interblob cells, are generally wavelength sensitive and monocular, and they lack orientation and direction selectivity. In other words, they resemble the koniocellular and parvocellular input from the LGN. The receptive fields of some blob neurons were found to be circular. Some have the color-opponent center-surround organization observed in the parvocellular and koniocellular layers of the LGN. Other blob cell receptive fields have red–green or blue–yellow color opponency in the center of their receptive fields, with no surround regions at all. Still other cells have both a color-opponent center and a color-opponent surround; they are called double-opponent cells. More recent studies of V1 have quantified the selectivity of blob and interblob cells and somewhat surprisingly found that, overall, neurons in blobs and interblobs are similar, showing selectivity for both orientation and color.
What should we conclude about the physiological properties of the blob neurons? Despite the distinct cytochrome oxidase labeling, at present there is no simple way to distinguish the receptive field properties of blob cells from neighboring interblob cells. Corresponding to the greater cytochrome oxidase activity in blobs, the firing rates of blob cells are on average higher than those of interblob cells. We can only speculate that future research may discover some receptive field difference that better correlates with the anatomical and firing rate distinctions. It is generally believed that neurons sensitive to wavelength are important for the analysis of object color, but we don’t know if we would be colorblind without functional cytochrome oxidase blobs.
We have seen that neurons in area V1 are clearly not all the same. Anatomical stains show that neurons in different layers, and even within a layer, exhibit a variety of shapes and neurite configurations. Coming into V1 are distinct projections from magnocellular, parvocellular, and koniocellular layers of the LGN. Within V1, cells are selective for different orientations, directions of motion, and colors. Some cells are monocular, while others are binocular. A big question is the extent to which this smorgasbord of neurons is organized into functional pathways that perform unique functions or modules that work cooperatively.
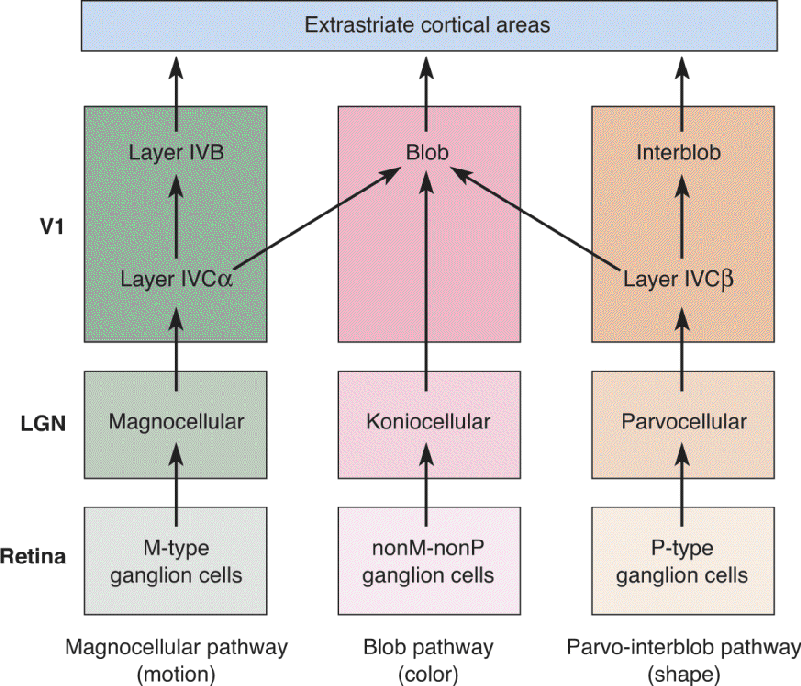
Parallel Pathways. Because there is great interest in how the brain makes sense of our complex visual world, possible systems by which visual analysis might take place have received considerable research attention. An influential model is based on the idea that there are three pathways within V1 that perform different functions in parallel. These can be called the magnocellular pathway, the parvo-interblob pathway, and the blob pathway (Figure 10.25). The magnocellular pathway begins with M-type ganglion cells of the retina. These cells send axons to the magnocellular layers of the LGN. These layers project to layer IVCα of striate cortex, which in turn projects to layer IVB. Because many of these cortical neurons are direction selective, the magnocellular pathway might be involved in the analysis of object motion and the guidance of motor actions.

FIGURE 10.25 A hypothetical model of parallel pathways in primary visual cortex. Based on receptive field properties and the pattern of innervation from LGN afferents, it has been suggested that there are three distinct pathways in striate cortex. Below each pathway, a functional role is suggested. Further research demonstrated mixing of magno, parvo, and koniocellular LGN signals and overlap in receptive field properties, raising questions about the distinctions, particularly between the blob and interblob pathways. Description
The parvo-interblob pathway originates with P-type ganglion cells of the retina, which project to the parvocellular layers of the LGN. The parvocellular LGN sends axons to layer IVCα of striate cortex, which projects to layer II and III interblob regions. Neurons in this pathway have small orientation-selective receptive fields, so perhaps they are involved in the analysis of fine object shape.
Finally, the blob pathway receives input from the subset of ganglion cells that are neither M-type cells nor P-type cells. These nonM–nonP cells project to the koniocellular layers of the LGN. The koniocellular LGN projects directly to the cytochrome oxidase blobs in layers II and III. Many neurons in the blobs are color selective, so they might be involved in the analysis of object color.
The description earlier is the simple version of the story; the reality is more complicated. Research has shown that the three proposed pathways do not keep magnocellular, parvocellular, and koniocellular signals separate; instead they mix. Also, receptive field properties such as orientation and color tuning are found across the proposed pathways. Thus, it is not the case that magnocellular, parvo-interblob, and blob neurons are strictly segregated and have entirely unique receptive field properties. At present, it appears that striate cortex input reflects the magnocellular, parvocellular, koniocellular segregation also seen in the LGN, but striate cortex output has a different form of parallel processing. For example, layer IVB contains many direction-selective neurons, its output appears to be dominated by magnocellular LGN input, and it projects to cortical areas thought to be involved in motion perception. Collectively, these observations are consistent with the notion that this is an output pathway particularly involved in navigation and the analysis of motion. The case for a distinction between form and color pathways is less compelling. We will see later that beyond the striate cortex, there appear to be two major pathways handling different types of visual information, one extending toward the parietal lobe that deals with motion and the other involved with color and form extending toward the temporal lobe.
Cortical Modules. Receptive fields in primary visual cortex range from a fraction of a degree to several degrees across, and nearby cells have receptive fields that overlap a great deal. For these reasons, even a small spot of light will activate thousands of V1 neurons. Hubel and Wiesel showed that the image of a point in the visual field falls within the receptive fields of neurons in a 2 × 2 mm chunk of macaque striate cortex. Such a block of cortex also contains two complete sets of ocular dominance columns, 16 blobs, and a complete sampling (twice over) of all 180° of possible orientations. Thus, Hubel and Wiesel argued that a 2 × 2 mm chunk of striate cortex is both necessary and sufficient to analyze the image of a point in space: necessary because its removal would leave a blind spot for this point in the visual field, and sufficient because it contains all the neural machinery required to analyze the form and color of objects viewed through either eye. Such a unit of brain tissue has come to be called a cortical module. Because of the finite size of receptive fields and some scatter in their locations, a cortical module processes information about a small patch of the visual field.
The striate cortex is constructed from perhaps a thousand cortical modules; one is shown in Figure 10.26. We can think of a visual scene being simultaneously processed by these modules, each “looking” at a portion of the scene. Just remember that the modules are an idealization. Optical images of V1 activity reveal that the regions of the striate cortex responding to different eyes and orientations are not nearly as regular as the “ice cube model” in Figure 10.26 suggests.

FIGURE 10.26 A cortical module. Each cortical module contains ocular dominance columns, orientation columns, and cytochrome oxidase blobs to fully analyze a portion of the visual field. The idealized cube shown here differs from the actual arrangement, which is not as regular or orderly. Description
The striate cortex is called V1, for “visual area one,” because it is the first cortical area to receive information from the LGN. Beyond V1 lie another two dozen distinct extrastriate areas of cortex that have unique receptive field properties. The contributions to vision of these extrastriate areas are still being vigorously debated. However, it appears that there are two large-scale cortical streams of visual processing, one stretching dorsally from the striate cortex toward the parietal lobe and the other projecting ventrally toward the temporal lobe (Figure 10.27).

FIGURE 10.27 Beyond the striate cortex in the macaque monkey brain. (a) Dorsal and ventral visual processing streams. (b) Extrastriate visual areas. (c) The flow of information in the dorsal and ventral streams. Description
The dorsal stream appears to serve the analysis of visual motion and the visual control of action. The ventral stream is thought to be involved in the perception of the visual world and the recognition of objects. These processing streams have primarily been studied in the macaque monkey brain, where recordings from single neurons can be made. However, functional magnetic resonance imaging (fMRI) research has identified areas in the human brain that have properties analogous to brain areas in the macaque. The locations of some of the human visual areas are shown in Figure 10.28.

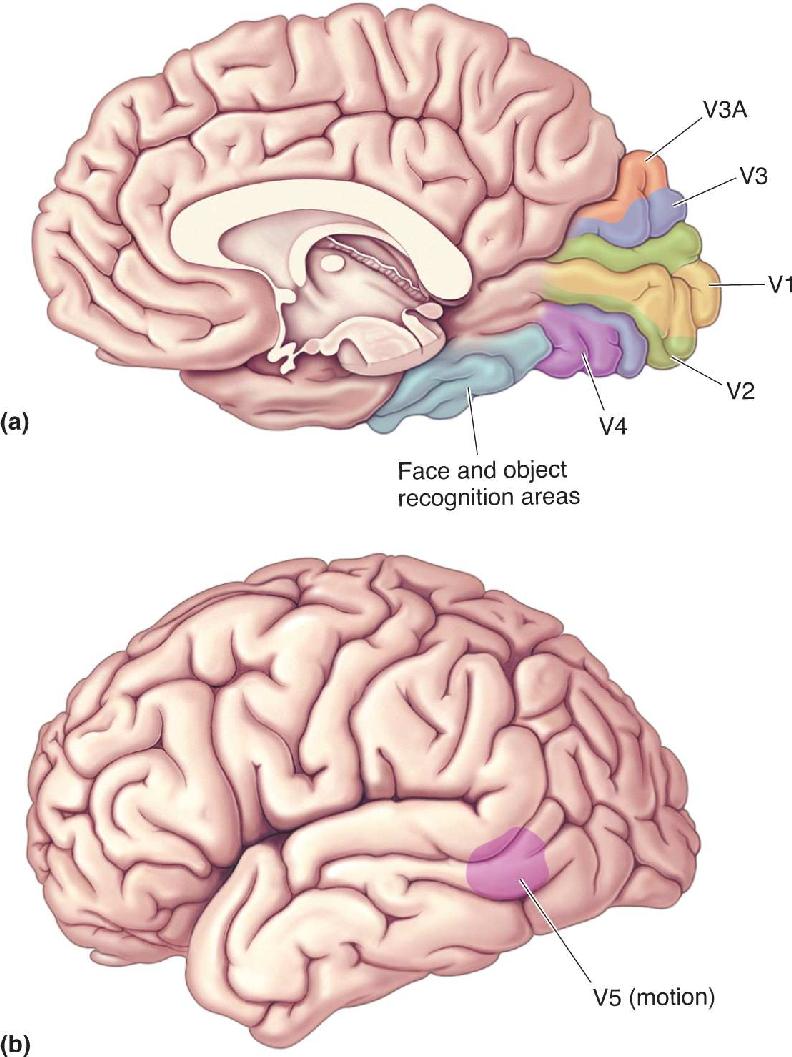
FIGURE 10.28 Visual areas in the human brain. (a) Compared to monkeys, human visual areas are shifted more to the medial wall of the occipital lobe, and many are buried in sulci. Early visual areas including V1, V2, V3, V3A, and V4 are retinotopically organized. Higher temporal lobe areas involved in face and object recognition are not retinotopic. (b) A variety of areas responsive to visual motion are found on the lateral aspect of the brain. The most studied of these is area V5, also known as MT. (Source: Zeki, 2003, Fig. 2.) Description
The properties of dorsal stream neurons are most similar to those of magnocellular neurons in V1, and ventral stream neurons have properties more like features of parvo-interblob and blob cells in V1. However, each extrastriate stream receives some amount of input from all the pathways in the primary visual cortex.
The cortical areas composing the dorsal stream are not arranged in a strict serial hierarchy, but there does appear to be a progression of areas in which more complex or specialized visual representations develop. Projections from V1 extend to areas designated V2 and V3, but we will skip farther ahead in the dorsal stream.
Area MT. In an area known as V5 or MT (because of its location in the middle temporal lobe in some monkeys), strong evidence indicates that specialized processing of object motion takes place. The location of area MT in the human brain is shown in Figure 10.28b. Area MT receives retinotopically organized input from a number of other cortical areas, such as V2 and V3, and it also is directly innervated by cells in layer IVB of the striate cortex. Recall that in layer IVB, the cells have relatively large receptive fields, transient responses to light, and direction selectivity. Neurons in area MT have large receptive fields that respond to stimulus movement in a narrow range of directions. Area MT is most notable for the fact that almost all the cells are direction selective, unlike areas earlier in the dorsal stream or anywhere in the ventral stream.
The neurons in MT also respond to types of motion, such as drifting spots of light, that are not good stimuli for cells in other areas—it appears that the motion of the objects is more important than their structure. Perhaps you have seen illusory motion in paintings or optical illusions; MT has been shown to be activated by some of these images too, suggesting that its neurons tell us what motion we perceive, not necessarily what motion is present. Further specialization for motion processing is evident in the organization of MT. This cortical area is arranged into direction-of-motion columns analogous to the orientation columns in V1. Presumably, the perception of movement at any point in space depends on a comparison of the activity across columns spanning a full 360° range of preferred directions.
William Newsome and his colleagues at Stanford University have shown that weak electrical stimulation in area MT of the macaque monkey appears to alter the direction in which small dots of light are perceived to move. For example, if electrical stimulation is applied to cells in a direction column preferring rightward movement, the monkey makes behavioral decisions suggesting that it has perceived motion in that direction. The artificial motion signal from electrical stimulation in MT appears to combine with visual motion input. The fact that the monkey behaviorally reports a perceived direction of motion based on the combination suggests that MT activity plays an important role in motion perception.
Dorsal Areas and Motion Processing. Beyond area MT, in the parietal lobe, are areas with additional types of specialized movement sensitivity. For example, in an area known as medial superior temporal (MST), there are cells selective for linear motion (as in MT), radial motion (either inward or outward from a central point), and circular motion (either clockwise or counterclockwise). We do not know how the visual system makes use of neurons with complex motion-sensitive properties in MST or of the “simpler” direction-selective cells in V1, MT, and other areas. However, three roles have been proposed:
- Navigation: As we move through our environment, objects stream past our eyes, and the direction and speed of objects in our peripheral vision provide valuable information that can be used for navigation.
- Directing eye movements: Our ability to sense and analyze motion must also be used when we follow objects with our eyes and when we quickly move our eyes to objects in our peripheral vision that catch our attention.
- Motion perception: We live in a world filled with motion, and survival sometimes depends on our interpretation of moving objects.
Striking evidence that cortical areas in the vicinity of MT and MST are critical for motion perception in humans comes from extremely rare cases in which brain lesions selectively disrupt the perception of motion. The clearest case was reported in 1983 by Josef Zihl and his colleagues at the Max Planck Institute for Psychiatry in Munich, Germany. Zihl studied a woman who experienced a stroke at the age of 43 years, bilaterally damaging portions of extrastriate visual cortex known to be particularly responsive to motion (Figure 10.28b). Although some ill effects of the stroke were evident, such as difficulty naming objects, neuropsychological testing showed the patient to be generally normal and to have relatively normal vision, except for one serious deficit: She appeared to be incapable of visually perceiving motion. Before you decide that not seeing motion would be a minor impairment, imagine what it would be like to see the world in snapshots. Zihl’s patient complained that when she poured coffee into a cup, it appeared at one moment to be frozen at the bottom of the cup and then suddenly it had overfilled the cup and covered the table. More ominously, she had trouble crossing the street—one moment she would perceive cars to be in the distance, and the next moment they would be right next to her. Clearly, this loss of motion perception had profound ramifications for the woman’s lifestyle. The implication of this case is that motion perception may be based on specialized mechanisms located beyond the striate cortex in the dorsal stream.
In parallel with the dorsal stream, a progression of areas from V1, V2, and V3 running ventrally toward the temporal lobes appears specialized for the analysis of visual attributes other than motion.
Area V4. One of the most-studied areas in the ventral stream is area V4 (see Figures 10.27b and 10.28a for the location of V4 in monkey and human brains). V4 receives input from the blob and interblob regions of the striate cortex via a relay in V2. Neurons in area V4 have larger receptive fields than cells in the striate cortex, and many of the cells are both orientation selective and color selective. Although there is a good deal of ongoing research into the function of V4, this area appears to be important for both shape perception and color perception. If this area is damaged in monkeys, perceptual deficits involving both shape and color result.
A rare clinical syndrome in humans known as achromatopsia is characterized by a partial or complete loss of color vision despite the presence of normal functional cones in the retina. People with this condition describe their world as drab, consisting of only shades of gray. Imagine how unappetizing a gray banana would look! Because achromatopsia is associated with cortical damage in the occipital and temporal lobes, without damage to V1, the LGN, or the retina, the syndrome suggests that there is specialized color processing in the ventral stream. Consistent with the coexistence of color-sensitive and shape-sensitive cells in the ventral stream, achromatopsia is usually accompanied by deficits in form perception. Some researchers have proposed that V4 is a particularly critical area for color and form perception, but the lesions associated with achromatopsia are generally not limited to V4, and severe visual deficits appear to require damage to other cortical areas in addition to V4.
Area IT. Beyond V4 in the ventral stream are cortical areas that contain neurons with complicated spatial receptive fields. A major output of V4 is an area in the inferior temporal lobe known as area IT (see Figure 10.27b and the recognition areas in Figure 10.28a). One reason this area is of particular interest is that it appears to be the farthest extent of visual processing in the ventral stream. A wide variety of colors and abstract shapes have been found to be good stimuli for cells in IT. As we will see in Chapter 24, output from area IT is sent to temporal lobe structures involved in learning and memory; IT itself may be important for both visual perception and visual memory. Recognizing an object clearly involves a combination or comparison of incoming sensory information with stored information.
One of the most intriguing findings concerning IT, as first noted by Charles Gross and his coworkers then at Princeton University, is that a small percentage of IT neurons in monkeys respond strongly to surprisingly complex objects such as pictures of faces. These cells may also respond to stimuli other than faces, but faces produce a particularly vigorous response, and some faces are more effective stimuli than others.
Human brain studies using fMRI appear to be consistent with the findings in monkeys. Nancy Kanwisher and her colleagues at MIT discovered that there is an area in the human brain that is more responsive to faces than to other stimuli (Box 10.3). This area is located on the fusiform gyrus and has come to be called the fusiform face area (Figure 10.29a). Might this area play a special role in the ability to recognize faces, which are of great behavioral significance to humans? The finding of face-selective cells and the fusiform face area has sparked much interest, in part because of a syndrome called prosopagnosia—difficulty recognizing faces even though vision is otherwise normal. This rare syndrome usually results from a stroke and is associated with damage to extrastriate visual cortex, perhaps including the fusiform face area.

FIGURE 10.29 Human brain activity elicited by pictures of faces. Using fMRI, brain activity was recorded first in response to faces and second in response to non-face stimuli. (a) In the horizontal brain section to the right, the red and yellow area on the left side and the symmetrical red area on the right side, known as the fusiform face area, showed significantly greater responses to faces. (b) Modified techniques in more recent studies have revealed multiple face-selective areas including occipital face area (OFA), anterior face patch 2 (AFP2), and fusiform face area (FFA). (Source: Part a courtesy of Drs. I. Gauthier, J.C. Gore, and M. Tarr; part b courtesy of Weiner and Grill-Spector, 2012.) Description
During my first year in graduate school in 1981, the first functional images of human visual cortex appeared on the cover of Science. Captivated by positron emission tomography (PET), a remarkable technology that enabled us for the first time to peer directly into the workings of the normal human brain, I wrote a research proposal using this method to investigate human vision and sent it to all of the PET labs in the world (I think there were five at the time). But it took a decade of pounding on the doors before I gained access to a PET scanner and got to do my first experiment.
Functional MRI (fMRI) was just starting to catch on then, and a few years later, in 1995, I was given the breathtaking privilege of my own weekly slot on the fMRI scanner at Massachusetts General Hospital. Collaborating with an undergraduate, Josh McDermott, and post-doc, Marvin Chun, I spent some of the happiest moments of my life lying inside the scanner bore, biting on a bite bar, and watching Marvin and Josh (upside down) through the mirror over my forehead, as they operated the scanner from out in the console room. What astonishing good fortune to get to use this amazing machine to explore the largely uncharted territory of human visual cortex!
We started out by trying to find brain regions engaged in the perception of object shape. Although we found some intriguing effects, they were weak. As I did not have a grant to pay for scan time, I knew that my scanning privileges would not continue unless I hit a home run—and quickly.
The extensive behavioral literature on normal and brain-damaged individuals strongly suggested that a special part of the brain might exist for face perception. We decided to look for it. Leslie Ungerleider and Jim Haxby and their colleagues at the National Institutes of Health (NIH) had already shown strong activations on the bottom of the temporal lobes when people look at faces. What they had not investigated was whether this response was specific to faces, or whether the same region might also be engaged during perception of other complex visual stimuli. This specificity question connected directly to one of the most long-standing and fierce debates in the histories of both cognitive science and neuroscience: To what extent are the mind and brain composed of special-purpose mechanisms, each processing a specific kind of information?
We figured that if a special-purpose part of the brain existed that was selectively involved in face perception, it should produce a stronger response when people look at faces than when they look at objects. To get enough face images, Marvin, Josh, and I went to Harvard’s “freshman face capture,” where the entire incoming class lined up for ID photos; we asked if we could use their ID photos in our experiments. We then scanned subjects while they looked at these face photos and at photos of common objects.
To our delight, we found that in almost everyone the image showed a nice clear blob on the lateral side of the fusiform gyrus, primarily in the right hemisphere, where the statistics told us that the response was higher when people were looking at faces than when they were looking at objects. However, it was not in exactly the same place in each subject. To deal with this anatomical variability, plus to make our statistical analyses bulletproof, we split the data for each subject in half, using half the data to find the region with our faces-versus-objects contrast, and the other half to quantify the response in that region. This “region of interest” method had already been used successfully by people studying lower level visual areas, and it was not a big leap to extend it to higher-level cortical areas.
Of course, demonstrating that a region of the brain is selectively responsive to faces requires much more than just showing that it responds more strongly to faces than to objects. Over the next few years, we (and others, notably Greg McCarthy and Aina Puce at Yale) tested the face specificity hypothesis against numerous alternative hypotheses. The fusiform face area (FFA) obliged, passing each new test (Figure A).

Figure A Nancy’s FFA, fusiform face area; OFA, occipital face area. (Source: Courtesy of N. Kanwisher.)
Other labs have used different methods to make breathtaking discoveries that have greatly extended our understanding of the FFA. After finding face-selective patches in monkeys with fMRI, Doris Tsao and her colleagues at Harvard went on to report that the vast majority of the cells in this region respond nearly exclusively to faces. (Not even I had thought the faces patches were that selective!) David Pitcher and his colleagues at University College London briefly disrupted a face-selective region just behind the FFA with transcranial magnetic stimulation, showing that this region is necessary for face perception (but not for perception of objects or bodies). And Yoichi Sugita at the Japan Science and Technology Agency reported that monkeys reared for 2 years without ever seeing a face show adult-like face discrimination abilities in the very first behavioral testing session, suggesting that experience with faces may not be necessary to wire the face-processing system.
Our original decision to work on faces was a pragmatic one (we needed a quick result), and it worked out well for us. But I am just as proud of the completely unanticipated discoveries we made later, such as the scene-selective parahippocampal place area (PPA) working with Russell Epstein, and the body-selective extrastriate body area (EBA) working with Paul Downing. Most astonishing to me was Rebecca Saxe’s discovery of a brain region that is selective for thinking about another person’s thoughts. (My only role was to tell her this experiment would never work!)
These findings show that the human mind and brain contain at least a few very specialized components, each of which is dedicated to solving a very specific computational problem. These discoveries open up a vast landscape of new questions. What computations go on in each of these regions? How are those computations implemented in neural circuits? What other specialized brain regions exist? How do these specialized regions develop? Why do some mental processes get their own private piece of real estate in the brain, while others do not? Tackling these questions will be a challenge and a thrill.
More recent experiments have revealed that there are actually about half a dozen patches of cortex within and near IT that are particularly sensitive to faces, and neurons in each patch have different degrees of sensitivity to face identity (Mary versus Sue) and other attributes such as the side of the head viewed (left, right, front, back) (Figure 10.29b). The implication is that multiple visual areas including portions of area IT may compose a system of areas specialized for facial recognition. In other human brain imaging studies, groups of patchy brain regions have been found that are reportedly involved in representations of color and biological objects.
Visual perception—the task of identifying and assigning meaning to objects in space—obviously requires the concerted action of many cortical neurons. But which neurons in which cortical areas determine what we perceive? How is the simultaneous activity of widely separated cortical neurons integrated, and where does this integration take place? Neuroscience research is just beginning to tackle these challenging questions. However, sometimes basic observations about receptive fields can give us insight into how we perceive (Box 10.4).
You have probably seen books or posters showing patterns of dots or splotches of color that supposedly contain pictures in 3D, if you contort your eyes just the right way. But how is it possible to see three dimensions on a two-dimensional piece of paper? The answer is based on the fact that our two eyes always see slightly different images of the world because of the distance between them in the head. The closer objects are to the head, the greater the difference in the two images. You can easily demonstrate this to yourself by holding a finger up in front of your eyes and alternately viewing it, at different distances, with the left or right eye closed.
Long before anything was known about binocular neurons in visual cortex, stereograms were a popular form of recreation. Two photographs were taken with lenses separated by a distance roughly the same as that of human eyes. By looking at the left photograph with the left eye and the right photograph with the right eye (by relaxing the eye muscles or with a stereoscope), the brain combines the images and interprets the different views as cues for distance, providing a three-dimensional percept (Figure A).

Figure A A nineteenth-century stereogram. (Source: Horibuchi, 1994, p. 38.)
In 1960, Bela Julesz, working at the Bell Telephone Laboratories, invented random-dot stereograms (Figure B). These paired images of random dots are, in principle, the same as the nineteenth-century stereograms. The big difference is that the object hidden in the images cannot be seen with normal binocular viewing. To see the 3D object, you must direct your left and right eyes to the left and right images. The principle in constructing the stereo images is to create a background of randomly spaced dots, and wherever an area should be closer or farther away in the fused image, the dots shown to one eye are horizontally shifted relative to those in the other eye. Imagine looking at a white index card covered with random black dots while you hold it in front of a large piece of white paper covered with similar dots. By alternately closing your eyes, the dots on the index card will shift horizontally more than those on the more distant piece of paper. The pair of stereo images captures this difference in viewpoint and erases any other indication that there is a square in front, such as the edge of the index card. Random-dot stereograms shocked many scientists, because in 1960, it was commonly thought that depth was perceived only after the objects in each eye were separately recognized.

Figure B A random-dot stereogram and the perception that results from binocularly fusing the images. (Source: Julesz, 1971, p. 21.)
In the 1970s, Christopher Tyler at the Smith–Kettlewell Eye Research Institute created autostereograms. An autostereogram is a single image that, when properly viewed, gives the perception of objects in 3D (Figure C). The colorful, and sometimes frustrating, autostereograms you see in books are based on an old illusion called the wallpaper effect. If you look at wallpaper that contains a repeating pattern, you can cross (or diverge) your eyes and view one piece of the pattern with one eye and the next cycle of the pattern with the other eye. The effect makes the wallpaper appear to be closer (or farther away). In an autostereogram, the wallpaper effect is combined with random-dot stereograms. To see the 3D skull in Figure C, you need to relax your eye muscles so that the left eye looks at the left dot on top and the right eye the right dot. You will know you are getting close when you see three dots at the top of the image. Relax and keep looking, and the picture will become visible.

Figure C An autostereogram. (Source: Horibuchi, 1994, p. 54.)
One of the fascinating things about stereograms is that you often must look at them for tens of seconds or even minutes, while your eyes become “properly” misaligned and your visual cortex “figures out” the correspondence between the left and right eye views. We do not know what is going on in the brain during this period, but presumably it involves the activation of binocular neurons in the visual cortex.
Comparing the receptive field properties of neurons at different points in the visual system might provide insight about the basis of perception. The receptive fields of photoreceptors are simply small patches on the retina, whereas those of retinal ganglion cells have a center-surround structure. The ganglion cells are sensitive to variables such as contrast and the wavelength of light. In the striate cortex, we encounter simple and complex receptive fields that have several new properties, including orientation selectivity and binocularity. We have seen that in extrastriate cortical areas, cells are selectively responsive to more complex shapes, object motion, and even faces. It appears that the visual system consists of a hierarchy of areas in which receptive fields become increasingly larger and more complex, moving away from V1 (Figure 10.30). Perhaps our perception of specific objects is based on the excitation of a small number of specialized neurons in some ultimate perceptual area that has not yet been identified. Is a person’s recognition of his or her grandmother based on the responses of 5 or 10 cells with receptive field properties so highly refined that the cells respond only to one person? The closest approximation to this is the face-selective neurons in area IT. However, even these fascinating cells do not respond to only one face.

FIGURE 10.30 A hierarchy of receptive fields. As one progresses from the retina to extrastriate visual cortex, receptive fields become larger and are selective for more complex shapes. At present, it seems unlikely that perception is based on undiscovered neurons highly selective for every object a person recognizes, such as one’s grandmother. Description
While it is by no means settled, there are several arguments against the idea that perception is based on extremely selective receptive fields such as those of hypothetical “grandmother cells.” First, recordings have been made from most parts of the monkey brain, but there is no evidence that a portion of cortex has different cells tuned to each of the millions of objects that we all recognize. Second, such great selectivity appears to be counter to the general principle of broad tuning that exists throughout the nervous system. Photoreceptors respond to a range of wavelengths, simple cells respond to many orientations, cells in MT respond to motion in a range of directions, and face cells usually respond to multiple faces. Moreover, cells that are selective for one property—orientation, color, or whatever—are always sensitive also to other properties. For example, we can emphasize the orientation selectivity of V1 neurons and the way in which this might relate to the perception of form, neglecting the fact that the same cells might selectively respond to size, direction of motion, and so on. Finally, it might be too “risky” for the nervous system to rely on extreme selectivity. A blow to the head might kill all five grandmother cells, and in an instant, one would lose the ability to recognize her. We will have more to say about the robustness of recognition when we explore learning and memory in Chapters 24 and 25.
If we do not rely on “grandmother cells,” how does perception work? One alternative hypothesis is formulated around the observation that parallel processing is used throughout the visual system (and other brain systems). We encountered parallel processing in Chapter 9 when we discussed ON and OFF and M and P ganglion cells. In this chapter, we saw three parallel channels coming into V1. Extending away from V1 are dorsal and ventral streams of processing, and different areas in these two streams are biased, or specialized, for various stimulus properties. Perhaps the brain uses a “division of labor” principle for perception. Within a given cortical area, many broadly tuned cells may serve to represent features of objects. At a bigger scale, a group of cortical areas may contribute to perception, some dealing more with color or form, others more with motion. In other words, perception may be more like the sound produced by an orchestra of visual areas, each with different roles, than by a single musician.
In this chapter, we have outlined the organization of the sensory pathway from the eye to the thalamus to the cortex. We saw that vision actually involves the perception of numerous different properties of objects—including color, form, and movement—and these properties are processed in parallel by different cells of the visual system. This processing of information evidently requires a strict segregation of inputs at the thalamus, some limited convergence of information in the striate cortex, and finally a massive divergence of information as it is passed on to higher cortical areas. The distributed nature of the cortical processing of visual information is underscored when you consider that the output of a million ganglion cells can recruit the activity of well over a billion cortical neurons throughout the occipital, parietal, and temporal lobes! Somehow, this widespread cortical activity is combined to form a single, seamless perception of the visual world.
Heed the lessons learned from the visual system. As we shall see in later chapters, the basic principles of organization in this system—parallel processing, topographic mappings of sensory surfaces, synaptic relays in the dorsal thalamus, cortical modules, and multiple cortical representations—are also features of the sensory systems devoted to hearing and touch.
1. Following a bicycle accident, you are disturbed to find that you cannot see anything in your left visual field. Where has the retinofugal pathway been damaged?
2. What is the source of most of the input to the left LGN?
3. A worm has eaten part of one lateral geniculate nucleus. You can no longer perceive motion in the right visual field of your right eye. What layer(s) of which LGN have most likely been damaged?
4. List the chain of connections that link a cone in the retina to a blob cell in the striate cortex. Is there more than one path by which cones connect to the blob cell?
5. What is meant by the statement that there is a map of the visual world in the striate cortex?
6. What is parallel processing in the visual system? Give two examples.
7. If a child is born cross-eyed and the condition is not corrected before the age of 10 years, binocular depth perception will be lost forever. This is explained by a modification in the circuitry of the visual system. From your knowledge of the central visual system, where do you think the circuitry has been modified?
8. What layers of the striate cortex send efferents to other visual cortex areas?
9. What new receptive field properties are found in the striate cortex and other cortical areas that are not seen in the retina or LGN?
10. What sort of experiment might you perform to investigate the relationship between visual perception and neural activity in the visual cortex?
De Haan EHF, Cowey A. 2011. On the usefulness of “what” and “where” pathways in vision. Trends in Cognitive Science 15:460–466.
Gegenfurtner KR. 2003. Cortical mechanisms of colour vision. Nature Reviews Neuroscience 4: 563–572.
Grill-Spector K, Malach R. 2004. The human visual cortex. Annual Reviews of Neuroscience 27: 649–677.
Hendry SHC, Reid RC. 2000. The koniocellular pathway in primate vision. Annual Reviews of Neuroscience 23:127–153.
Kreiman G. 2007. Single unit approaches to human vision and memory. Current Opinion in Neurobiology 17:471–475.
Milner AD, Goodale MA. 2008. Two visual systems reviewed. Neuropsychologia 46: 774–785.
Nasso JJ, Callaway EM. 2009. Parallel processing strategies of the primate visual system. Nature Reviews Neuroscience 10:360–372.
Sherman SM. 2012. Thalamocortical interactions. Current Opinion in Neurobiology 22: 575–579.
Tsao DY, Moeller S, Freiwald W. 2008. Comparing face patch systems in macaques and humans. Proceedings of the National Academy of Science 49:19514–19519.
Zeki S. 2003. Improbable areas in the visual brain. Trends in Neuroscience 26:23–26.
Additional figures