The Somatic Sensory System
BOX 12.1 OF SPECIAL INTEREST: Herpes, Shingles, and Dermatomes
BOX 12.3 PATH OF DISCOVERY: Cortical Barrels, by Thomas Woolsey
BOX 12.4 OF SPECIAL INTEREST: The Misery of Life Without Pain
Nociceptors and the Transduction of Painful Stimuli
BOX 12.6 OF SPECIAL INTEREST: Pain and the Placebo Effect
The somatic sensory system brings us some of life’s most enjoyable experiences, as well as some of the most aggravating. Somatic sensation enables our body to feel, to ache, to sense hot or chill, and to know what its parts are doing. It is sensitive to many kinds of stimuli: the pressure of objects against the skin, the position of joints and muscles, distension of the bladder, and the temperature of the limbs and of the brain itself. It is the origin of itch. When stimuli become so strong that they may be damaging, somatic sensation is also responsible for the feeling that is most offensive, but vitally important—pain.
The somatic sensory system is different from other sensory systems in two interesting ways. First, its receptors are distributed throughout the body rather than being concentrated at small, specialized locations. Second, because it responds to many different kinds of stimuli, we can think of it as a group of at least four senses rather than a single one: the senses of touch, temperature, pain, and body position. In fact, those four can in turn be subdivided into many more. The somatic sensory system is really a catch-all name, a collective category for all the sensations that are not seeing, hearing, tasting, smelling, and the vestibular sense of balance. The familiar idea that we have only five senses is obviously too simple.
If something touches your finger, you can accurately gauge the place, pressure, sharpness, texture, and duration of the touch. If it is a pinprick, there is no mistaking it for a hammer. If the touch moves from your hand to your wrist, and up your arm to your shoulder, you can track its speed and position. Assuming you are not looking, this information is described entirely by the activity of the sensory nerves in your limb. A single sensory receptor can encode stimulus features such as intensity, duration, position, and sometimes direction. But a single stimulus usually activates many receptors. The CNS interprets the activity of the vast receptor array and uses it to generate coherent perceptions.
In this chapter, we divide our discussion of somatic sensation into two main parts: the sense of touch and the sense of pain. As we shall see, these different categories depend on different receptors, different axonal pathways, and different regions of the brain. We’ll also describe sensations of itch, and how we sense changes in temperature. The sense of body position, also called proprioception, is discussed in Chapter 13, where we will explore how this type of somatic sensory information is used to control muscle reflexes.
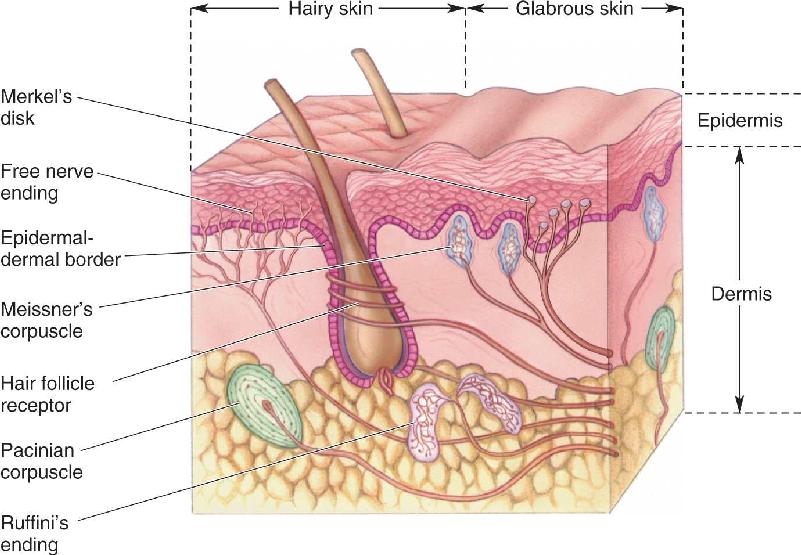
The sensation of touch begins at the skin (Figure 12.1). The two major types of skin are called hairy and glabrous (hairless), as exemplified by the backs and palms of your hands. Skin has an outer layer, the epidermis, and an inner layer, the dermis. Skin performs an essential protective function, and it prevents the evaporation of body fluids into the dry environment we live in. But skin also provides our most direct contact with the world; indeed, skin is the largest sensory organ we have. Imagine the beach without the squish of sand between your toes, or consider watching a kiss instead of experiencing it yourself. Skin is sensitive enough that a raised dot measuring only 0.006 mm high and 0.04 mm wide can be felt when stroked by a fingertip. In comparison, a Braille dot is 167 times higher.

FIGURE 12.1 Somatic sensory receptors in the skin. Hairy skin and glabrous skin have a variety of sensory receptors within the dermal and epidermal layers. Each receptor has an axon and, except for free nerve endings, all of them have associated non-neural tissues. Description
In this section, we will see how a touch of the skin is transduced into neural signals, how these signals make their way to the brain, and how the brain makes sense of them.
Most of the sensory receptors in the somatic sensory system are mechanoreceptors, which are sensitive to physical distortion such as bending or stretching. Present throughout the body, mechanoreceptors monitor skin contact, pressure in the heart and blood vessels, stretching of the digestive organs and urinary bladder, and force against the teeth. At the heart of all mechanoreceptors are unmyelinated axon branches that are sensitive to stretching, bending, pressure, or vibration.
The mechanoreceptors of the skin are shown in Figure 12.1. Most of them are named after the nineteenth century German and Italian histologists who discovered them. The largest and best-studied receptor is the Pacinian corpuscle, which lies deep in the dermis and can be as long as 2 mm and almost 1 mm in diameter. Each human hand has about 2500 Pacinian corpuscles, with the highest densities in the fingers. Ruffini’s endings, found in both hairy and glabrous skin, are slightly smaller than Pacinian corpuscles. Meissner’s corpuscles are about one-tenth the size of Pacinian corpuscles and are located in the ridges of glabrous skin (the raised parts of your fingerprints, for example). Located within the epidermis, Merkel’s disks each consist of a nerve terminal and a flattened, non-neural epithelial cell (the Merkel cell). In Krause end bulbs, which lie in the border regions of dry skin and mucous membrane (around the lips and genitals, for example), the nerve terminals look like knotted balls of string.
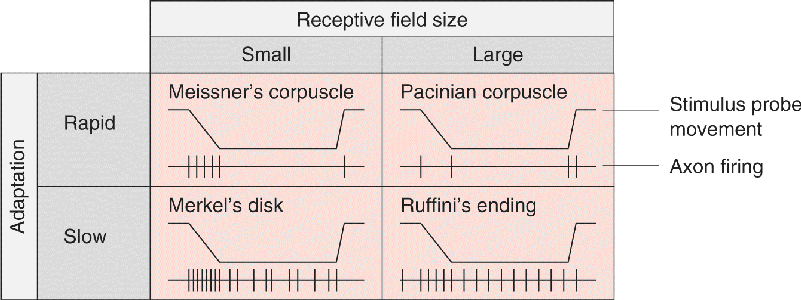
Skin can be vibrated, pressed, pricked, and stroked, and its hairs can be bent or pulled. These are quite different kinds of mechanical energy, yet we can feel them all and easily tell them apart. Accordingly, we have mechanoreceptors that vary in their preferred stimulus frequencies, pressures, and receptive field sizes. Swedish neuroscientist Åke Vallbo and his colleagues developed methods to record from single sensory axons in the human arm, so that they could simultaneously measure the sensitivity of mechanoreceptors in the hand and evaluate the perceptions produced by various mechanical stimuli (Figure 12.2a). When the stimulus probe was touched to the surface of the skin and moved around, the receptive field of a single mechanoreceptor could be mapped. Meissner’s corpuscles and Merkel’s disks had small receptive fields, only a few millimeters wide, while Pacinian corpuscles and Ruffini’s endings had large receptive fields that could cover an entire finger or half the palm (Figure 12.2b).

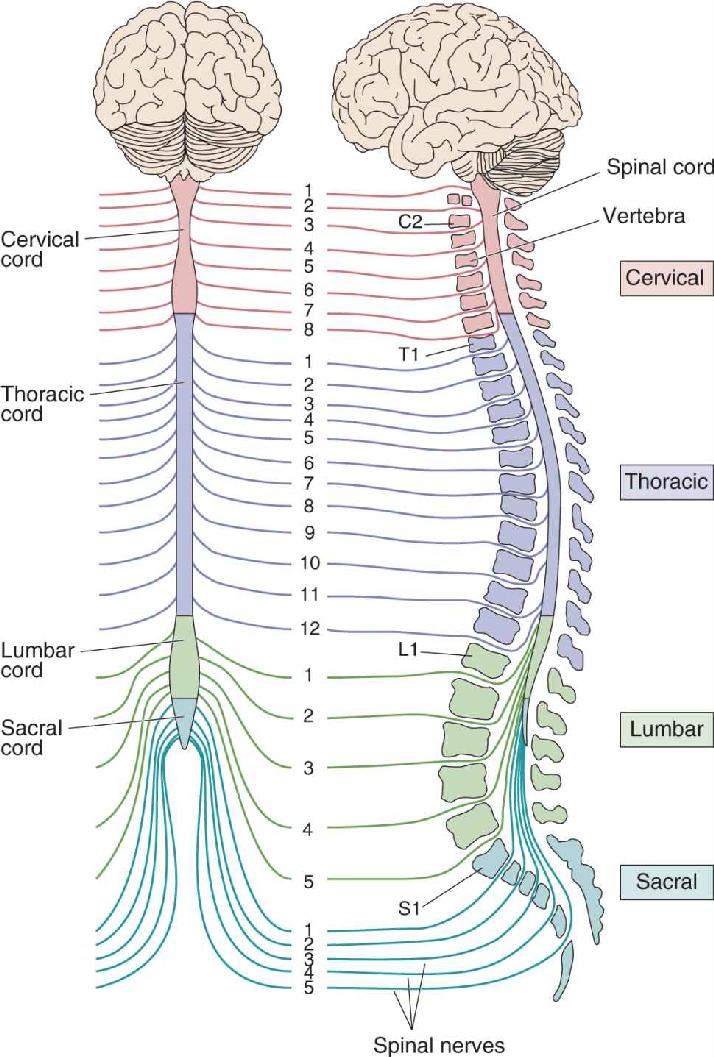
FIGURE 12.2 Testing the receptive fields of human sensory receptors. (a) By introducing a microelectrode into the median nerve of the arm, it is possible to record the action potentials from a single sensory axon and map its receptive field on the hand with a fine stimulus probe. (b) Results show that receptive fields are either relatively small, as in Meissner’s corpuscles, or large, as in Pacinian corpuscles. (Source: Adapted from Vallbo and Johansson, 1984.) Description
Mechanoreceptors also vary in the persistence of their responses to long-lasting stimuli. If a stimulus probe is suddenly pressed against the skin within the receptive field, some mechanoreceptors, such as Meissner’s and Pacinian corpuscles, tend to respond quickly at first but then stop firing even though the stimulus continues; these receptors are said to be rapidly adapting. Other receptors, such as Merkel’s disks and Ruffini’s endings, are slowly adapting, and generate a more sustained response during a long stimulus. Figure 12.3 summarizes the receptive field size and adaptation rate for four mechanoreceptors of the skin.

FIGURE 12.3 Variations of receptive field size and adaptation rate for four somatic sensory skin receptors. (Source: Adapted from Vallbo and Johansson, 1984.) Description
Hairs do more than adorn our head and keep a dog warm in winter. Many hairs are part of a sensitive receptor system. To demonstrate this, brush just a single hair on the back of your arm with the tip of a pencil; it feels like an annoying mosquito. For some animals, hair is a major sensory system. Imagine a rat slinking confidently through dark passageways and alleys. The rat navigates in part by waving its facial vibrissae (whiskers) to sense the local environment and derive information about the texture, distance, and shape of nearby objects.
Hairs grow from follicles embedded in the skin; each follicle is richly innervated by free nerve endings—the terminations of single axons—that either wrap around the follicle or run parallel to it (see Figure 12.1). There are several types of hair follicles, including some with erectile muscles (essential for mediating the strange sensation we call goose bumps), and the details of their innervation differ. In all cases, the bending of the hair causes a deformation of the follicle and surrounding skin tissues. This, in turn, stretches, bends, or flattens the nearby nerve endings, which then increase or decrease their action potential firing frequency. The mechanoreceptors of hair follicles may be either slowly adapting or rapidly adapting.
The different mechanical sensitivities of mechanoreceptors mediate different sensations. Pacinian corpuscles are most sensitive to vibrations of about 200–300 Hz, while Meissner’s corpuscles respond best around 50 Hz (Figure 12.4). Place your hand against a speaker while playing your favorite music loudly; you “feel” the music largely with your Pacinian corpuscles. If you stroke your fingertips across the coarse screen covering the speaker, each point of skin will hit the bumps at frequencies about optimal to activate Meissner’s corpuscles. You feel this as a sensation of rough texture. Stimulation at frequencies from about 1 to 10 Hz can also activate Meissner’s corpuscles, yielding a “fluttering” feeling.

FIGURE 12.4 Frequency sensitivity of two rapidly adapting skin mechanoreceptors. Pacinian corpuscles are most sensitive to high-frequency stimuli, and Meissner’s corpuscles are more sensitive to low-frequency stimuli. The skin was indented with a pressure probe, at various frequencies, while recording from the nerve. The amplitude of the stimulus was increased until it generated action potentials; threshold was measured as the amount of skin indentation in micrometers (μm). (Source: Adapted from Schmidt, 1978.) Description
Vibration and the Pacinian Corpuscle. The selectivity of a mechanoreceptive axon depends primarily on the structure of its special ending. For example, the Pacinian corpuscle has a football-shaped capsule with 20–70 concentric layers of connective tissue, arranged like the layers of an onion, with an axon terminal in the middle (see Figure 12.1). When the capsule is compressed, energy is transferred to the nerve terminal, its membrane is deformed, and mechanosensitive channels open. Current flowing through the channels generates a receptor potential, which is depolarizing (Figure 12.5a). If the depolarization is large enough, the axon will fire an action potential. But the capsule layers are slick, with viscous fluid between them. If the stimulus pressure is maintained, the layers slip past one another and transfer the stimulus energy in such a way that the axon terminal is no longer deformed, and the receptor potential dissipates. When pressure is released, the events reverse themselves; the terminal depolarizes again and may fire another action potential.

FIGURE 12.5 Adaptation in the Pacinian corpuscle. A single Pacinian corpuscle was isolated and stimulated by a probe that indented it briefly. The receptor potential was recorded from a nearby portion of the axon. (a) In the intact corpuscle, a large receptor potential was generated at the onset and offset of the stimulus; during maintained indentation, the receptor potential disappeared. (b) The onionlike encapsulation was dissected away, leaving a bare axon ending. When indented by the probe, a receptor potential was again generated, showing the capsule is not necessary for mechanoreception. But while the normal corpuscle responded only to the onset or offset of a long indentation, the stripped version gave a much more prolonged response; its adaptation rate was slowed. Apparently it is the capsule that makes the corpuscle insensitive to low-frequency stimuli. Description
In the 1960s, Werner Loewenstein and his colleagues, working at Columbia University, stripped away the capsule from single corpuscles and found that the naked nerve terminal became much less sensitive to vibrating stimuli and much more sensitive to steady pressure (Figure 12.5b). Clearly, it is the layered capsule (and not some property of the nerve ending itself) that makes the Pacinian corpuscle exquisitely sensitive to vibrating, high-frequency stimuli and almost unresponsive to steady pressure (see Figure 12.4). In order to communicate information about rapid vibrations to the central nervous system (CNS) in a timely way, the Pacinian corpuscles have some of the largest and fastest axons that originate in the skin.
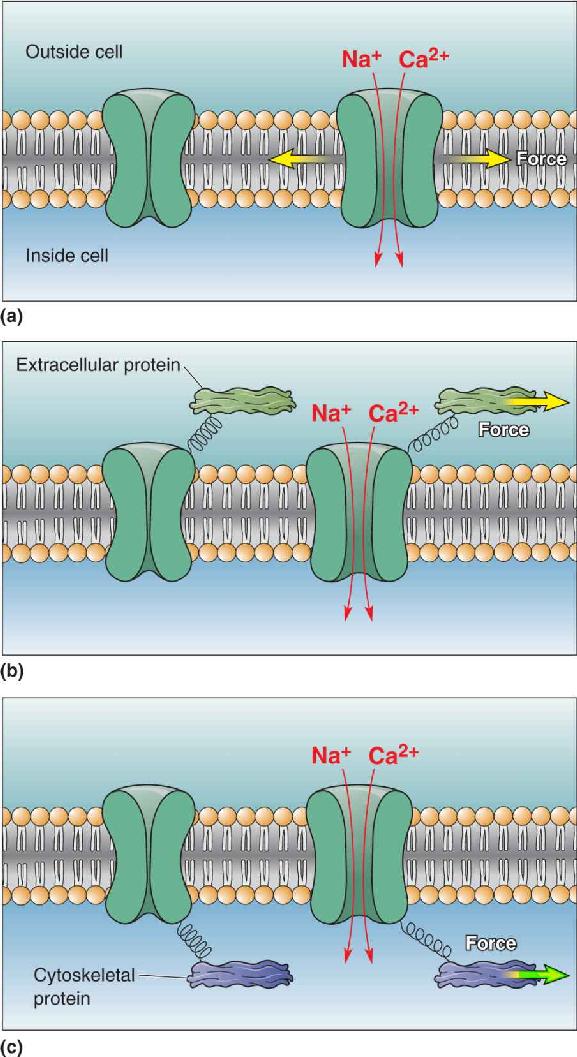
Mechanosensitive Ion Channels. The mechanoreceptors of the skin all have unmyelinated axon terminals, and the membranes of these axons have mechanosensitive ion channels that convert mechanical force into a change of ionic current. Forces applied to these channels alter their gating and either enhance or decrease channel opening. Force can be applied to a channel by the membrane itself when it is stretched or bent, or force may be applied through connections between the channels and extracellular proteins or intracellular cytoskeletal components (e.g., actin, microtubules) (Figure 12.6). Alternatively, mechanical stimuli may somehow trigger the release of second messengers (e.g., DAG, IP3) that secondarily regulate ion channels.

FIGURE 12.6 Mechanosensitive ion channels. (a) Some membrane ion channels are sensitive to stretching of the lipid membrane; tension in the membrane directly induces the channel to open and allow cations to flow. (b) Other ion channels open when force is applied to extracellular structures linked to the channels by peptides. (c) Mechanically sensitive channels may also be linked to intracellular proteins, especially those of the cytoskeleton; deformation of the cell and stress on its cytoskeleton generate forces that regulate channel gating. Description
A variety of ion channel types have been implicated in mechanosensation, but the specific types of channels in most of the somatic sensory receptors are still unidentified. Recent work on Merkel’s disks, which are sensitive to gentle pressure on the skin, suggests how complex some touch receptors are (see Figure 12.1). The epithelium-like Merkel cells make synapses onto nerve terminals, and it seems that both the Merkel cell and the axon terminal are mechanically sensitive. The Merkel cell has a mechanosensitive channel called Piezo2 that opens in response to pressure and depolarizes the cell. Depolarization triggers synaptic release of an unknown transmitter from the cell, which in turn excites the nearby nerve ending. Surprisingly, the nerve ending is also mechanically sensitive because of a second (unknown) ion channel in its own membranes. Thus, the actions of at least two different mechanosensitive channels and a synapse cooperate to activate Merkel’s disks and their associated axon.
Two-Point Discrimination. Our ability to discriminate the detailed features of a stimulus varies tremendously across the body. A simple measure of spatial resolution is the two-point discrimination test. You can do this yourself with a paper clip bent into the shape of a U. Start with the ends about an inch apart, and touch them to the tip of a finger; you should have no problem telling that there are two separate points touching your finger. Then bend the wire to bring the points closer together, and touch them to your fingertip again. Repeat, and see how close the points have to be before they feel like a single point. (This test is best done with two people, one testing and the other being tested without looking.) Now try it on the back of your hand, on your lips, on your leg, and any other place that interests you. Compare your results with those shown in Figure 12.7.

FIGURE 12.7 Two-point discrimination on the body surface. The pairs of dots show the minimum distance necessary to differentiate between two points touching the body simultaneously. Notice the sensitivity of the fingertips compared to the rest of the body. All measurements are shown at their actual scale. Description
Two-point discrimination varies at least twentyfold across the body. Fingertips have the highest resolution. The dots of Braille are 1 mm high and 2.5 mm apart; up to six dots make a letter. An experienced Braille reader can move an index finger across a page of raised dots and read about 600 letters per minute, which is roughly as fast as someone reading aloud. The Braille reader scans with fingertips because touch is most sensitive when the skin and the stimuli move across each other, compared to simply pressing one against the other. Practice also improves performance through a type of learning, and experienced Braille readers are particularly adept at discriminating patterns of small dots.
Several reasons explain why the fingertip is so much better than, say, the elbow for Braille reading: (1) There is a much higher density of mechanoreceptors in the skin of the fingertip than on other parts of the body, (2) the fingertips are enriched in receptor types that have small receptive fields (e.g., Merkel’s disks), (3) there is more brain tissue (and thus more raw computing power) devoted to the sensory information of each square millimeter of fingertip than elsewhere, and (4) there may be special neural mechanisms devoted to high-resolution discriminations.
The skin is richly innervated by axons that course through the vast network of peripheral nerves on their way to the CNS (Figure 12.8). Axons bringing information from the somatic sensory receptors to the spinal cord or brain stem are the primary afferent axons of the somatic sensory system. The primary afferent axons enter the spinal cord through the dorsal roots; their cell bodies lie in the dorsal root ganglia (Figure 12.9).


FIGURE 12.9 The structure of a segment of the spinal cord and its roots. Description
Primary afferent axons have widely varying diameters, and their size correlates with the type of sensory receptor to which they are attached. Unfortunately, the terminology approaches absurdity here because the different sizes of axons are designated by two sets of names, using Arabic and Greek letters and Roman numerals. As shown in Figure 12.10, in order of decreasing size, axons from skin sensory receptors are usually designated Aα, Aβ, Aδ, and C; axons of similar size, but innervating the muscles and tendons, are called groups I, II, III, and IV. Group C (or IV) axons are, by definition, unmyelinated axons, while all the rest are myelinated.

FIGURE 12.10 Various sizes of primary afferent axons. The axons are drawn to scale, but they are shown 2000 times larger than life size. The diameter of an axon is correlated with its conduction velocity and with the type of sensory receptor to which it is connected. Description
An interesting and simple point is hidden in the many axon names. Recall that the diameter of an axon, together with its myelin, determines its speed of action potential conduction. The smallest axons, the so-called C fibers, have no myelin and are less than about 1 μm in diameter. C fibers mediate pain, temperature sensation, and itch, and they are the slowest of axons, conducting at about 0.5–1 m/sec. To see how slow this is, take a big step, count to two, and then take another step. That’s about how fast the action potentials travel along C fibers. On the other hand, touch sensations, mediated by the cutaneous mechanoreceptors, are conveyed by the relatively large Aβ axons, which can conduct at up to 75 m/sec. For comparison, consider that an exceptional major league baseball pitcher can throw a fastball up to 100 miles per hour, which is about 45 m/sec.
Most peripheral nerves communicate with the CNS via the spinal cord, which is encased in the bony vertebral column.
Segmental Organization of the Spinal Cord. The arrangement of paired dorsal and ventral roots shown in Figure 12.9 is repeated 30 times down the length of the human spinal cord. Each spinal nerve, consisting of dorsal root and ventral root axons, passes through a notch between the vertebrae (the “back bones”) of the spinal column. There are as many spinal nerves as there are notches between vertebrae. As shown in Figure 12.11, the 30 spinal segments are divided into four groups, and each segment is named after the vertebra adjacent to where the nerves originate: cervical (C) 1–8, thoracic (T) 1–12, lumbar (L) 1–5, and sacral (S) 1–5.

FIGURE 12.11 Segmental organization of the spinal cord. The spinal cord is divided into cervical, thoracic, lumbar, and sacral divisions (left). The cross-sectional view (right) shows the spinal cord within the vertebral column. Spinal nerves are named for the level of the spinal cord from which they exit and are numbered in order from rostral to caudal. Description
The segmental organization of spinal nerves and the sensory innervation of the skin are related. The area of skin innervated by the right and left dorsal roots of a single spinal segment is called a dermatome; thus, there is a one-to-one correspondence between dermatomes and spinal segments. When mapped, the dermatomes delineate a set of bands on the body surface, as shown in Figure 12.12. The organization of the dermatomes is best revealed when one bends over to stand on both hands and feet (Figure 12.13). This organization presumably reflects our distant quadrupedal ancestry.

FIGURE 12.12 Dermatomes. These illustrations show the mapping of the approximate boundaries of the dermatomes on the body. Description

When a dorsal root is cut, the corresponding dermatome on that side of the body does not lose all sensation. The residual somatic sensation is explained by the fact that the adjacent dorsal roots innervate overlapping areas. To lose all sensation in one dermatome, therefore, three adjacent dorsal roots must be cut. However, the skin innervated by the axons of one dorsal root is plainly revealed by a condition called shingles, in which all the neurons of a single dorsal root ganglion become infected with a virus (Box 12.1).
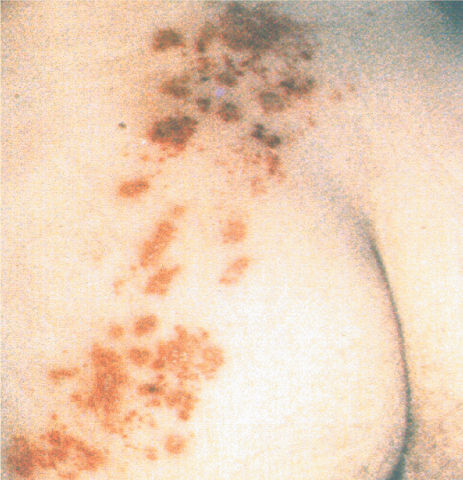
Many of us were infected by the varicella zoster virus, a type of herpes virus commonly known as chickenpox, when we were children. After a week or so covered with red, itchy spots on our skin, we usually recovered. Out of sight is not out of body, however. The virus remains in our primary sensory neurons, dormant but viable. Most people never notice it again, but in some cases, the virus revives decades later, wreaking havoc with the somatic sensory system. The result is shingles, a condition that can be agonizingly painful for periods of months or even years. The reactivated virus increases the excitability of the sensory neurons, leading to very low thresholds of firing as well as spontaneous activity. The pain is a constant burning, sometimes a stabbing sensation, and the skin is exquisitely sensitive to any stimulus. People with shingles often shun clothes because of their hypersensitivity. The skin itself becomes inflamed and blistered, then scaly—hence, the name (Figure A). Several useful treatments are available that often shorten the outbreak, relieve the pain, and prevent long-term complications.
Figure A Skin lesions caused by shingles, confined to the L4 dermatome on the left side.
Fortunately, the varicella zoster virus usually reactivates only in the neurons of one dorsal root ganglion. This means that the symptoms are restricted to the skin innervated by the axons of the affected dorsal root. In effect, the virus performs an anatomical labeling experiment for us by clearly marking the skin territory of one dermatome. Almost any dermatome may be involved, although the thoracic and facial areas are most common. Observations of many shingles patients and their infected areas were actually useful in mapping the dermatomes (see Figure 12.12).
Neuroscientists have learned how to use herpes viruses and other types of viruses to their advantage. Viruses are valuable research tools because they can be used to introduce new genes into neurons.
Notice in Figure 12.11 that the spinal cord in the adult ends at about the level of the third lumbar vertebra. The bundles of spinal nerves streaming down within the lumbar and sacral vertebral column are called the cauda equina (Latin for “horse’s tail”). The cauda equina courses down the spinal column within a sack of dura filled with cerebrospinal fluid (CSF). In a method called lumbar puncture (also called a spinal tap), used to collect CSF for medical diagnostic tests, a needle is inserted into this CSF-filled cistern at the midline. If the needle is inserted a little off center, however, a nerve can be touched. Not surprisingly, this causes a sensation of sharp pain in the dermatome supplied by that nerve.
Sensory Organization of the Spinal Cord. The basic anatomy of the spinal cord was introduced in Chapter 7. The spinal cord is composed of an inner core of gray matter, surrounded by a thick covering of white matter tracts that are often called columns. Each half of the spinal gray matter is divided into a dorsal horn, an intermediate zone, and a ventral horn (Figure 12.14). The neurons that receive sensory input from primary afferents are called second-order sensory neurons. Most of the second-order sensory neurons of the spinal cord lie within the dorsal horns.

FIGURE 12.14 The trajectory of the touch-sensitive Aβ axons in the spinal cord.
The large, myelinated Aβ axons conveying information about a touch to the skin enter the dorsal horn and branch. One branch synapses in the deep part of the dorsal horn on second-order sensory neurons. These connections can initiate or modify a variety of rapid and unconscious reflexes. The other branch of the Aβ primary afferent axon ascends straight to the brain. This ascending input is responsible for perception, enabling us to form complex judgments about the stimuli touching the skin.
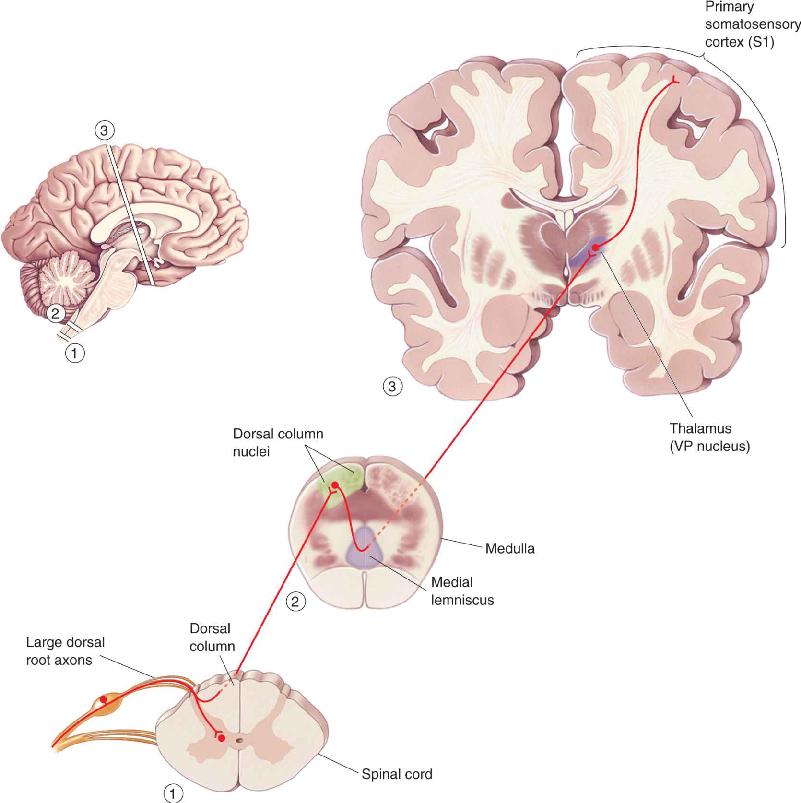
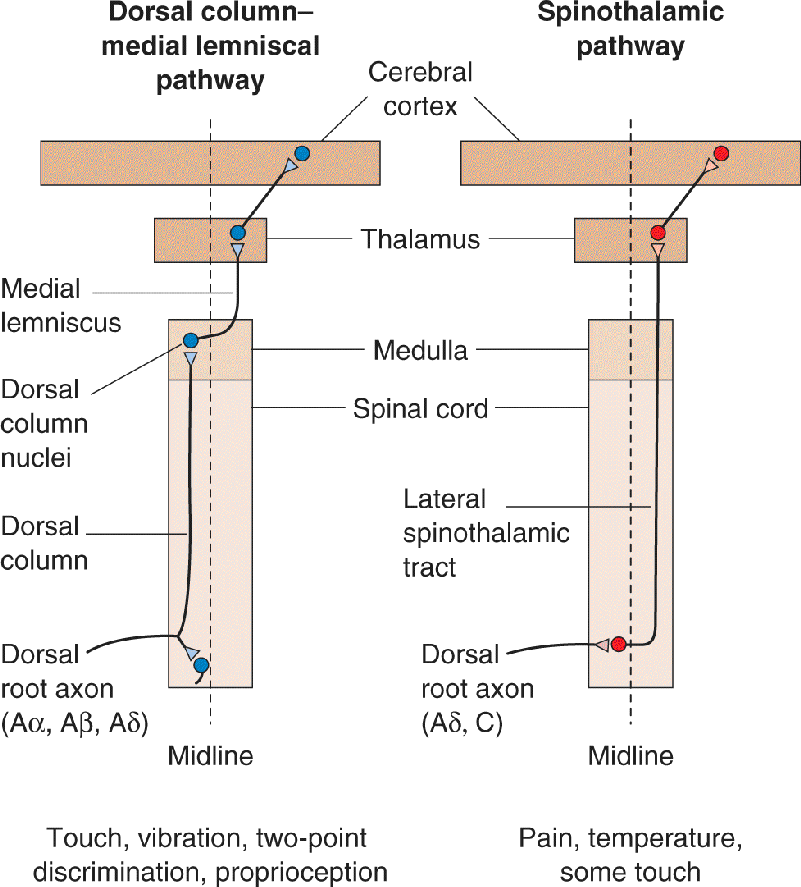
Information about touch or vibration of the skin takes a path to the brain that is entirely distinct from that taken by information about pain and temperature. The pathway serving touch is called the dorsal column–medial lemniscal pathway, for reasons we will see in a moment. The organization of this pathway is summarized in Figure 12.15.

FIGURE 12.15 The dorsal column–medial lemniscal pathway. This is the major route by which touch and proprioceptive information ascend to the cerebral cortex. Description
The ascending branch of the large sensory axons (Aβ) enters the ipsilateral dorsal column of the spinal cord, the white matter tract medial to the dorsal horn (see Figure 12.14). The dorsal columns carry information about tactile sensation (and limb position) toward the brain. They are composed of primary sensory axons, as well as second-order axons from neurons in the spinal gray matter. The axons of the dorsal column terminate in the dorsal column nuclei, which lie at the junction of the spinal cord and medulla. Consider that some of the longest axons in your body originate in the skin of your big toe and terminate in the dorsal column nuclei at the base of your head! This is a fast, direct path that brings information from the skin to the brain without an intervening synapse.
At this point in the pathway, information is still represented ipsilaterally; that is, touch information from the right side of the body is represented in the activity of cells in the right dorsal column nuclei. However, axons from cells of the dorsal column nuclei arch toward the ventral and medial medulla, and decussate. From this point onward, the somatic sensory system of one side of the brain is concerned with sensations originating from the other side of the body.
The axons of the dorsal column nuclei ascend within a conspicuous white matter tract called the medial lemniscus. The medial lemniscus rises through the medulla, pons, and midbrain, and its axons synapse upon neurons of the ventral posterior (VP) nucleus of the thalamus. Remember that almost no sensory information goes directly into the neocortex without first synapsing in the thalamus (olfaction is the exception). Thalamic neurons of the VP nucleus then project to specific regions of primary somatosensory cortex, or S1.
It is tempting to assume that sensory information is simply transferred, unchanged, through nuclei in the brain stem and thalamus on its way to the cortex, with the actual processing taking place only in the cortex. In fact, this assumption is demonstrated by the term relay nuclei, which is often used to describe specific sensory nuclei of the thalamus such as the VP nucleus. Physiological studies prove otherwise, however. In both dorsal column and thalamic nuclei, considerable transformation of information takes place. As a general rule, information is altered every time it passes through a set of synapses in the brain. In particular, inhibitory interactions between adjacent sets of inputs in the dorsal column–medial lemniscal pathway enhance the responses to tactile stimuli (Box 12.2). As we will see later, some synapses in these nuclei can also change their strength, depending on their recent activity. Neurons of both the thalamus and the dorsal column nuclei are also controlled by input from the cerebral cortex. Accordingly, the output of the cortex can influence the input of the cortex!
Information is usually transformed as it is passed from one neuron to the next in a sensory pathway. One common transformation is the amplification of differences in the activity of neighboring neurons, also known as contrast enhancement. We already saw this in retinal ganglion cell receptive fields (see Chapter 9). If all the photoreceptors providing input to a ganglion cell are evenly illuminated, the cell hardly notices. However, if there is a contrast border—a difference in illumination—within the cell’s receptive field, the cell’s response is strongly modulated. Contrast enhancement is a general feature of information processing in sensory pathways, including the somatic sensory system. One general mechanism underlying contrast enhancement is lateral inhibition, whereby neighboring cells inhibit one another. Let’s see how this works, using a simple model.
Consider the situation in Figure A. Dorsal root ganglion neurons lettered a through g relay information via excitatory synapses to dorsal column nucleus neurons A through G. All of the neurons fire with baseline rates of 5 spikes/sec, even in the absence of stimulation. Consider what happens when a stimulus is applied to the receptive field of just one sensory neuron, cell d in Figure A. The firing rate of cell d increases to 10 spikes/sec. Let’s assume the output of the dorsal column nucleus cells is simply the presynaptic input multiplied by a synaptic gain factor of 1. If the input activity of cell d is 10, the output activity of cell D is also 10. This simple relay does nothing to enhance the difference between the more active neuron, d, and the other neurons. The contrast in activity between neuron D and its neighbors C and E, for example, is 10 versus 5 spikes/sec.

Now consider the situation in Figure B, where inhibitory interneurons have been added that project laterally to inhibit each cell’s neighbor. The synaptic gain of the inhibitory synapses (black triangles) is –1, and we’ve adjusted the gain of the excitatory synapses, as shown in the figure. Calculate the activity of each cell by multiplying the input to each synapse by its synaptic gain, and then summing the effect of all the synapses on the cell. If you perform this calculation when the stimulus is again applied to cell d, you will see that there is significant contrast enhancement: The difference between the activity in cell d and its neighbors has been greatly amplified in the output of cell D. The contrast in activity between neuron D and its neighbors C and E is now 20 versus 0 spikes/sec.

Thus far, we have described only the part of the somatic sensory system that enters the spinal cord. If this were the whole story, your face and the top of your head would be numb. Somatic sensation of the face is supplied mostly by the large trigeminal nerves (cranial nerve V), which enter the brain at the pons (see Chapter 7). (The word is from the Latin tria, “three,” and geminus, “twin.”) There are twin trigeminal nerves, one on each side, and each breaks up into three peripheral nerves that innervate the face, mouth areas, the outer two-thirds of the tongue, and the dura mater covering the brain. Additional sensation from the skin around the ears, nasal areas, and pharynx is provided by other cranial nerves: the facial (VII), glossopharyngeal (IX), and vagus (X).
The sensory connections of the trigeminal nerve are analogous to those of the dorsal roots. The large-diameter sensory axons of the trigeminal nerve carry tactile information from skin mechanoreceptors. They synapse onto second-order neurons in the ipsilateral trigeminal nucleus, which is analogous to a dorsal column nucleus (Figure 12.16). The axons from the trigeminal nucleus decussate and project into the medial part of the VP nucleus of the thalamus. From here, information is relayed to the somatosensory cortex.

FIGURE 12.16 The trigeminal nerve pathway. Description
As with all other sensory systems, the most complex levels of somatosensory processing occur in the cerebral cortex. Most of the cortex concerned with the somatic sensory system is located in the parietal lobe (Figure 12.17). Brodmann’s area 3b, now regarded as the primary somatosensory cortex (S1), is easy to find in humans because it lies on the postcentral gyrus (right behind the central sulcus). (See also Figure 7.28 showing Brodmann’s areas.) Other cortical areas that also process somatic sensory information flank S1. These include areas 3a, 1, and 2 on the postcentral gyrus, and areas 5 and 7 on the adjacent posterior parietal cortex (see Figure 12.17).

FIGURE 12.17 Somatic sensory areas of the cortex. All of the illustrated areas lie in the parietal lobe. The lower drawing shows that the postcentral gyrus contains S1, area 3b. Description
Area 3b is the primary somatic sensory cortex because (1) it receives dense inputs from the VP nucleus of the thalamus; (2) its neurons are very responsive to somatosensory stimuli (but not to other sensory stimuli); (3) lesions here impair somatic sensation; and (4) when electrically stimulated, it evokes somatic sensory experiences. Area 3a also receives a dense input from the thalamus; however, this region is concerned with the sense of body position rather than touch.
Areas 1 and 2 receive dense inputs from area 3b. The projection from 3b to area 1 sends mainly texture information, while the projection to area 2 emphasizes size and shape. Small lesions in area 1 or 2 produce predictable deficiencies in discrimination of texture, size, and shape.
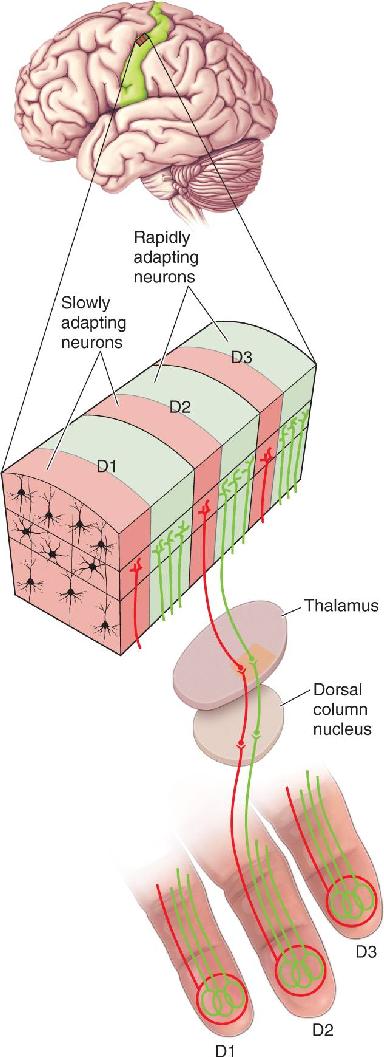
Somatic sensory cortex, like other areas of neocortex, is a layered structure. As is the case for visual and auditory cortex, the thalamic inputs to S1 terminate mainly in layer IV. The neurons of layer IV project, in turn, to cells in the other layers. Another important similarity with other regions of cortex is that S1 neurons with similar inputs and responses are stacked vertically into columns that extend across the cortical layers (Figure 12.18). In fact, the concept of the cortical column, so beautifully elaborated by Hubel and Wiesel in visual cortex, was actually first described in somatic sensory cortex by Johns Hopkins University scientist Vernon Mountcastle.

FIGURE 12.18 Columnar organization of S1’s area 3b. Each finger (D1–D3) is represented by an adjacent area of cortex. Within the area of each finger representation are alternating columns of cells with rapidly adapting (green) and slowly adapting (red) sensory responses. (Source: Adapted from Kaas et al., 1981, Fig. 8.) Description
Cortical Somatotopy. Electrical stimulation of the S1 surface can cause somatic sensations localized to a specific part of the body. Systematically, moving the stimulator around S1 will cause the sensation to move across the body. American-Canadian neurosurgeon Wilder Penfield, working at McGill University from the 1930s through the 1950s, actually used this method to map the cortex of neurosurgical patients. (It is interesting to note that these brain operations can be performed in awake patients, with only local anesthesia of the scalp, because the brain tissue itself lacks the receptors of somatic sensation.) Another way to map the somatosensory cortex is to record the activity of a single neuron and determine the site of its somatosensory receptive field on the body. The receptive fields of many S1 neurons produce an orderly map of the body on the cortex. The mapping of the body’s surface sensations onto a structure in the brain is called somatotopy. We have seen previously that the brain has maps of other sensory surfaces, such as the light-sensitive retina in the eye (retinotopy) and the sound frequency-sensitive cochlea in the inner ear (tonotopy).
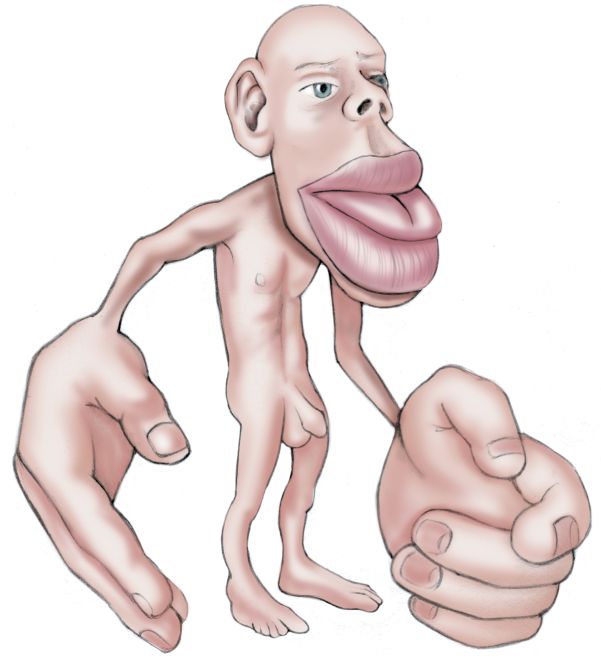
Somatotopic maps generated by electrical stimulation and neuronal recording methods are similar. The maps roughly resemble a body with its legs and feet at the top of the postcentral gyrus and its head at the opposite, lower end of the gyrus (Figure 12.19). A somatotopic map is sometimes called a homunculus (from the Latin diminutive of “man”; the little man in the brain).

FIGURE 12.19 A somatotopic map of the body surface onto primary somatosensory cortex. This map is a cross section through the postcentral gyrus (shown at top). Neurons in each area are most responsive to the parts of the body illustrated above them. (Source: Adapted from Penfield and Rasmussen, 1952, and from Kell et al., 2005, Fig. 3.) Description
Several things are obvious about the somatotopic map in S1. First, the map is not always continuous but can be broken up. Notice in Figure 12.19 that the representation of the hand separates that of the face and the head. Interestingly, Penfield’s original maps suggested that the male genitals were mapped onto the most distant and hidden part of S1, somewhere below the toes. However, a recent study using functional magnetic resonance imaging demonstrated that the penis is actually represented in a less surprising place on the map: in an area between the abdomen and the legs. Unfortunately, neither Penfield nor contemporary researchers have spent much time mapping the somatosensory maps of the female body and its unique features (what some have called the “hermunculus”).
Another obvious feature of the somatotopic map is that it is not scaled like the human body. Instead, it looks like a caricature (Figure 12.20): The mouth, tongue, and fingers are incongruously large, while the trunk, arms, and legs are tiny. The relative size of cortex devoted to each body part is correlated with the density of sensory input received from that part. Size on the map is also related to the importance of the sensory input from that part of the body; information from your index finger is more useful than that from your elbow. The importance of touch information from our hands and fingers is obvious, but why throw so much cortical computing power at the mouth? Two likely reasons are that tactile sensations are important in the production of speech and that your lips and tongue (feeling, as well as tasting) are the last line of defense when deciding if a morsel is delicious, nutritious food, or something that could choke you, break your tooth, or bite back. As we will see in a moment, the importance of an input, and the size of its representation in cortex, are also reflections of how often it is used.

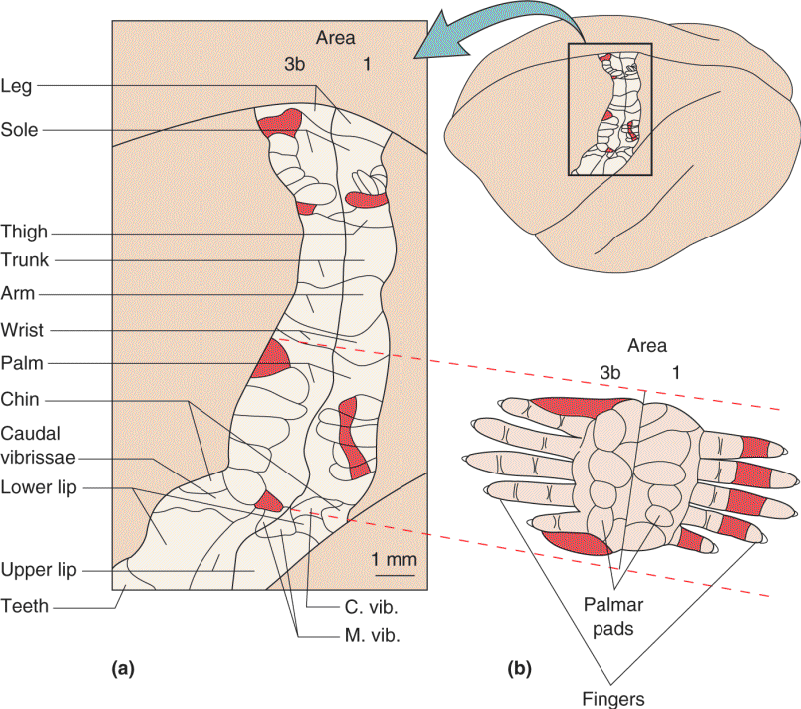
The importance of a body part can vary greatly in different species. For example, the large facial vibrissae (whiskers) of rodents receive a huge share of the territory in S1, while the digits of the paws receive relatively little (Figure 12.21). Remarkably, the sensory signals from each vibrissa follicle go to one clearly defined cluster of S1 neurons; such clusters are called barrels. The somatotopic map of rodent vibrissae is easily seen in thin sections of S1; the five rows of cortical barrels precisely match the five rows of facial vibrissae (Box 12.3). Studies of the “barrel cortex” in rats and mice have revealed much about the functions of sensory cortex.

FIGURE 12.21 A somatotopic map of the facial vibrissae on mouse cerebral cortex. (a) The positions of the major vibrissae on the face (dots). (b) A somatotopic map within S1 of the mouse brain. (c) Barrel cortex within S1. The cortex has been thinly sectioned parallel to the surface and Nissl-stained. The inset shows the pattern of barrels, laid out in five rows; compare with the five rows of vibrissae in the photograph in part a. (Source: Adapted from Woolsey and Van der Loos, 1970.) Description
In the mid-1960s, I completed a physiological study of the organization of touch, hearing, and vision in the mouse brain for an undergraduate research project at the University of Wisconsin. At Wisconsin, histology was done routinely on all such brains. After my first year at medical school, I came back to look at the sections. There was something odd about cortical layer IV just where I had recorded responses to moving the whiskers: The cell bodies were distributed unevenly. This was not new. Several different authors, in nearly forgotten papers written over 50 years earlier, showed this pattern of neurons; but that was before recording was possible, so no one knew the function of the cortex.
Cortex is usually studied in sections cut perpendicular to the brain surface. It occurred to me that cutting the brain parallel to its surface, which had only been done rarely, might give a view of the entire layer IV. The late H. Van der Loos, who taught neuroanatomy, gave me a place to work at Johns Hopkins during an elective period. I prepared specimens in a way so that I could accurately position them for cutting (I knew where I usually got responses to stimulating the face) and cut thicker sections than customary. About 10:00 on a bright late spring morning, after struggling to mount the first sections on slides, I took them down a corridor to the dark student histology lab, where I had a microscope. That first look showed a stunning pattern of cells in layer IV that obviously mimicked the whiskers. There was no doubt about what I had seen; I immediately showed the slides to Van der Loos, who was the second person in the world to know that whiskers are stamped in the mouse brain. We named the cell groups barrels. Later, the hypotheses that each barrel is associated with a single whisker and that each one forms part of a functional cortical column were proven.
Somatotopy in the cerebral cortex is not limited to a single map. Just as the visual system builds multiple retinotopic maps, the somatic sensory system has several maps of the body. Figure 12.22 shows the detailed somatotopy of S1 in an owl monkey. Carefully compare the maps in areas 3b and 1; they map the same parts of the body, literally in parallel along adjacent strips of cortex. The two somatotopic maps are not identical, but mirror images, as an enlargement of the hand regions makes clear (Figure 12.22b).

FIGURE 12.22 Multiple somatotopic maps. Recordings were made from areas 3b and 1 of an owl monkey. (a) Results show that each area has its own somatotopic map. (b) Detailed examination of the hand area shows that the two maps are mirror images. Shaded regions represent the dorsal surfaces of the hands and feet, unshaded regions the ventral surfaces. (Source: Adapted from Kaas et al., 1981.) Description
Cortical Map Plasticity. What happens to the somatotopic map in cortex when an input, such as the finger, is removed? Does the “finger area” of cortex simply go unused? Does it atrophy? Or is this tissue taken over by inputs from other sources? The answers to these questions could have important implications for the recovery of function after peripheral nerve injury. In the 1980s, neuroscientist Michael Merzenich and his associates at the University of California at San Francisco began a series of experiments to test the possibilities.
Some key experiments are summarized in Figure 12.23. First, the regions of S1 sensitive to stimulation of the hand in an adult owl monkey were carefully mapped with microelectrodes. Then, one finger (digit 3) on the hand was surgically removed. Several months later, the cortex was again mapped. The answer? The cortex originally devoted to the amputated digit now responded to stimulation of the adjacent digits (Figure 12.23c). There clearly had been a major rearrangement of the circuitry underlying cortical somatotopy.

FIGURE 12.23 Somototopic map plasticity. (a, b) The fingers of the hand of an owl monkey are mapped onto the surface of S1 cortex. (c) If digit 3 is removed, over time, the cortex reorganizes so that the representations of digits 2 and 4 expand. (d) If digits 2 and 3 are selectively stimulated, their cortical representations also expand. Description
In the amputation experiment, the cause of this map rearrangement was the absence of input from the missing digit. What happens when the input activity from a digit is increased? To answer this question, monkeys were trained to use selected digits to perform a task for which they received a food reward. After several weeks of this training, microelectrode mapping experiments showed that the representation of the stimulated digits had expanded in comparison with the adjacent, unstimulated ones (Figure 12.23d). These experiments reveal that cortical maps are dynamic and adjust depending on the amount of sensory experience. Subsequent experiments in other areas of cortex (visual, auditory, motor) have shown that this type of map plasticity is widespread in the brain.
The findings of map plasticity in animals have led to a search for similar changes in the human brain. One interesting example comes from studies of amputees. A common experience among amputees is the perception of sensations coming from the missing limb when other body parts are touched. These “phantom limb” sensations are usually evoked by the stimulation of skin regions whose somatotopic representations border those of the missing limb; for example, feeling can be evoked in a phantom arm by stimulating the face. Functional brain imaging reveals that the cortical regions originally devoted to the missing limb are now activated by stimulating the face. While this plasticity may be adaptive in the sense that the cortex does not go unused, the mismatch between sensory stimulation and perception in amputees shows that it can lead to confusion on how signals from S1 should be interpreted.
While having more cortex devoted to a body part may not necessarily be beneficial to amputees, it apparently is to musicians. Violinists and other string instrument players must continually finger the strings with their left hand; the other hand, holding the bow, receives considerably less stimulation of individual fingers. Functional imaging of S1 shows that the amount of cortex devoted to the fingers of the left hand is greatly enlarged in string musicians. It is likely that this is an exaggerated version of a continuous remapping process that goes on in everyone’s brain as each person’s life experiences vary.
The mechanisms of these types of map plasticity are not understood. However, as we shall see in Chapter 25, they may be related to processes involved in learning and memory.
The Posterior Parietal Cortex. As we have seen, the segregation of different types of information is a general rule for the sensory systems, and the somatic sensory system is no exception. However, information of different sensory types cannot remain separate forever. When we feel for a key in our pocket, we do not ordinarily sense it as a list of traits: a particular size and shape, textured and smooth edges, hard and smooth flat surfaces, a certain weight. Instead, without thinking much about it, we simply confirm with our fingers “key,” as opposed to “coin” or “wad of old chewing gum.” Separate aspects of a stimulus come effortlessly together as a meaningful object. We have a very poor understanding of how this occurs biologically within any sensory system, much less between sensory systems. After all, many objects have a distinct look, sound, feel, and smell, and the melding of these sensations is necessary for the complete mental image of something like your pet cat.
What we do know is that the character of neuronal receptive fields tends to change as information passes through the cortex and receptive fields enlarge. For example, neurons below the cortex and in cortical areas 3a and 3b are not sensitive to the direction of stimulus movement across the skin, but cells in areas 1 and 2 are. The stimuli that neurons prefer become increasingly complex. Certain cortical areas seem to be sites where simple, segregated streams of sensory information converge to generate particularly complex neural representations. When we discussed the visual system, we saw this in the complex receptive fields of area IT. The posterior parietal cortex is also such an area. Its neurons have large receptive fields with stimulus preferences that are a challenge to characterize because they are so elaborate. Moreover, the area is concerned not only with somatic sensation but also with visual stimuli, movement planning, and even a person’s state of attentiveness.
Damage to posterior parietal areas can yield some bizarre neurological disorders. Among these is agnosia, the inability to recognize objects even though simple sensory skills seem to be normal. People with astereognosia cannot recognize common objects by feeling them (e.g., a key), although their sense of touch is otherwise normal and they may have no trouble recognizing the object by sight or sound. Deficits are often limited to the side contralateral to the damage.
Parietal cortical lesions may also cause a neglect syndrome, in which a part of the body or a part of the world (the entire visual field left of the center of gaze, for example) is ignored or suppressed, and its very existence is denied (Figure 12.24). Neurologist Oliver Sacks described such a patient in his essay, “The Man Who Fell Out of Bed.” After suffering a stroke that presumably damaged his cortex, the man insisted that someone was playing a macabre joke on him by hiding an amputated leg under his blanket. When he tried to remove the leg from his bed, he and the leg ended up on the floor. Of course, the leg in question was his own, still attached, but he was unable to recognize it as part of his body. A neglect syndrome patient may ignore the food on one half of his plate, or attempt to dress only one side of his body. Neglect syndromes are most common following damage to the right hemisphere, and, fortunately, they usually improve or disappear with time.

FIGURE 12.24 Symptoms of a neglect syndrome. A patient who had had a stroke in the right posterior parietal cortex was asked to copy the model drawing but was unable to reproduce many of the features on the left side of the model. (Source: Springer and Deutsch, 1989, p. 193.)
In general, the posterior parietal cortex seems to be essential for the perception and interpretation of spatial relationships, accurate body image, and the learning of tasks involving coordination of the body in space. These functions involve a complex integration of somatosensory information with that from other sensory systems, particularly the visual system.
In addition to the mechanosensitive touch receptors we have described so far, somatic sensation depends strongly on nociceptors, the free, branching, unmyelinated nerve endings that signal that body tissue is being damaged or is at risk of being damaged. (The word is from the Latin nocere, “to hurt.”) The information from nociceptors takes a path to the brain that is largely distinct from the path taken by mechanoreceptors; consequently, the subjective experience elicited by activation of these two pathways is different. Selective activation of nociceptors can lead to the conscious experience of pain. Nociception, and pain, are vital to life (Box 12.4).
Pain teaches us to avoid harmful situations. It elicits withdrawal reflexes from noxious stimuli. It exhorts us to rest an injured part of our body so it can heal. Pain is vital. The most convincing arguments for the functional benefits of pain are the very rare people with a condition called congenital insensitivity to pain. They go through life in constant danger of destroying themselves because they do not realize the harm they are doing. They often die young.
A Canadian woman, for example, was born with an indifference to painful stimuli, had no other sensory deficits, and was quite intelligent. Despite early training to avoid damaging situations, she developed progressive degeneration of her joints and spinal vertebrae, leading to skeletal deformation, degeneration, infection, and, finally, death at the age of 28. Apparently, low levels of nociceptive activity are important during everyday tasks to tell us when a particular movement or prolonged posture is putting too much strain on our body. Even during sleep, nociception may be the prod that makes us toss and turn enough to prevent bedsores or skeletal strain.
People with congenital insensitivity to pain reveal that pain is a separate sensation and not simply an excess of the other sensations. Such people usually have a normal ability to perceive other somatic sensory stimuli. The causes of the disorder can include the failure of peripheral nociceptors to develop, altered synaptic transmission in the pain-mediating pathways of the CNS, and genetic mutations. A study of several afflicted families in Pakistan revealed mutations in a gene called SCN9A, which codes for a unique type of voltage-gated sodium channel expressed only in nociceptive neurons. The mutation leads to nonfunctional sodium channels, a selective absence of action potentials in nociceptors, and profound insensitivity to pain. Family members with the mutations sustain continual cuts, bruises, bitten lips and tongues, and broken bones.
It is important to realize, however, that nociception and pain are not always the same thing. Pain is the feeling, or the perception, of irritating, sore, stinging, aching, throbbing, miserable, or unbearable sensations arising from a part of the body. Nociception is the sensory process that provides signals that trigger pain. While nociceptors may fire away wildly and continually, pain may come and go. The opposite may also happen. Pain may be agonizing, even without activity in nociceptors. More than any other sensory system, the cognitive qualities of nociception can be controlled from within, by the brain itself.
Nociceptors and the Transduction of Painful Stimuli
Nociceptors are activated by stimuli that have the potential to cause tissue damage. Tissue damage can result from strong mechanical stimulation, extremes in temperature, oxygen deprivation, and exposure to certain chemicals, among other causes. The membranes of nociceptors contain ion channels that are activated by these types of stimuli.
Consider as an example the events that accompany your stepping on a thumbtack (recall Chapter 3). The simple stretching or bending of the nociceptor membrane activates mechanically gated ion channels that cause the cell to depolarize and generate action potentials. In addition, damaged cells at the site of injury can release a number of substances that cause ion channels on nociceptor membranes to open. Examples of released substances are proteases (enzymes that digest proteins), adenosine triphosphate (ATP), and K+. Proteases can break down an abundant extracellular peptide called kininogen to form another peptide called bradykinin. Bradykinin binds to specific receptor molecules that activate ionic conductances in some nociceptors. Similarly, ATP causes nociceptors to depolarize by binding directly to ATP-gated ion channels. And, as we learned in Chapter 3, the elevation of extracellular [K+] directly depolarizes neuronal membranes.
Now consider leaning against a hot stove. Heat above 43 °C causes tissues to burn, and heat-sensitive ion channels in nociceptor membranes open at this temperature. Of course, we also have nonpainful sensations of warmth when the skin is heated from 37 to 43 °C. These sensations depend on non-nociceptive thermoreceptors and their CNS connections, which we will discuss in a later section. But for now, note that the sensations of warmth and scalding are mediated by separate neural mechanisms.
Imagine you are a middle-aged runner on the last mile of a marathon. When your tissue oxygen levels do not meet the oxygen demand, your cells use anaerobic metabolism to generate ATP. A consequence of anaerobic metabolism is the release of lactic acid. The buildup of lactic acid leads to an excess of H+ in the extracellular fluid, and these ions activate H+-gated ion channels on nociceptors. This mechanism causes the excruciating dull ache associated with very hard exercise.
A bee stings you. Your skin and connective tissue contain mast cells, a component of your immune system. Mast cells can be activated by exposure to foreign substances (e.g., bee venom), causing them to release histamine. Histamine binds to specific cell surface receptors on nociceptors and causes membrane depolarization. Histamine also causes blood capillaries to become leaky, which leads to swelling and redness at the site of an injury. Creams containing drugs that block histamine receptors (antihistamines) can be helpful to reduce the pain and the swelling.
Types of Nociceptors. The transduction of painful stimuli occurs in the free nerve endings of unmyelinated C fibers and lightly myelinated Aδ fibers. The majority of nociceptors respond to mechanical, thermal, and chemical stimuli, and are therefore called polymodal nociceptors. However, like the mechanoreceptors of touch, many nociceptors show selectivity in their responses to different stimuli. Thus, there are also mechanical nociceptors, showing selective responses to strong pressure; thermal nociceptors, showing selective responses to burning heat or extreme cold (Box 12.5); and chemical nociceptors, showing selective responses to histamine and other chemicals.
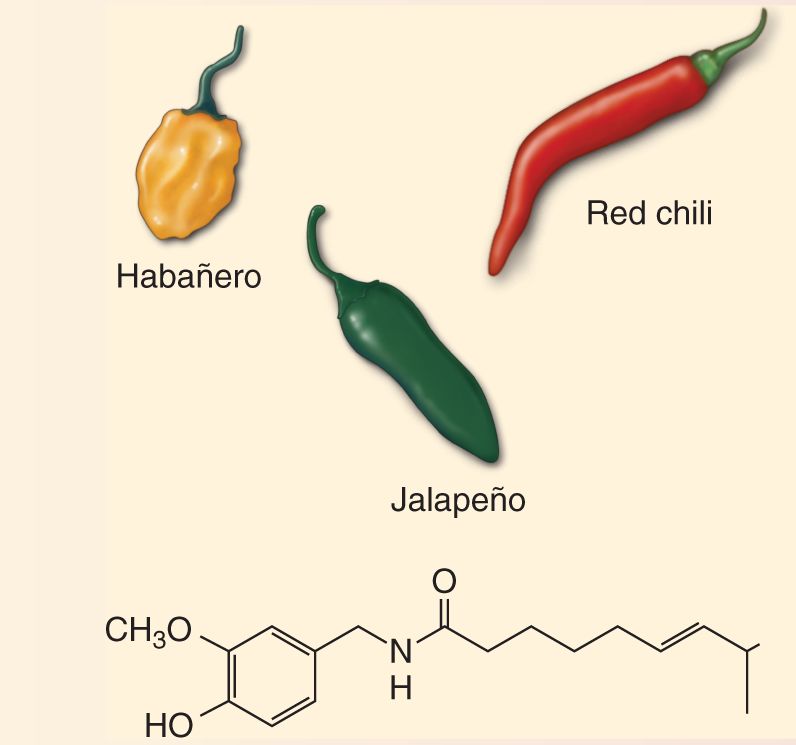
If you like spicy food, you should know that the active ingredient in a wide variety of hot peppers is capsaicin (Figure A). These peppers are “hot” because the capsaicin activates the thermal nociceptors that also signal painful elevations in temperature (above about 43 °C). Indeed, it was the peculiar fact that these nociceptive neurons are selectively activated by capsaicin that led to the discovery of the transduction mechanism for the sensation of hot. David Julius at the University of California, San Francisco, found that in some dorsal root ganglion cells, capsaicin activates a particular ion channel, called TRPV1, which is also activated by elevations in temperature greater than 43 °C. This ion channel causes the neuron to fire by admitting Ca2+ and Na+ and depolarizing it. TRPV1 is a member of a very large family of related TRP channels, originally identified in photoreceptors of the fruit fly Drosophila (TRP stands for transient receptor potential). Different TRP channels contribute to many different types of sensory transduction in organisms from yeast to humans.

Figure A Peppers containing capsaicin and the molecule’s chemical structure.
Why would a temperature-gated ion channel also be sensitive to hot peppers? Capsaicin appears to mimic the effect of endogenous chemicals released by tissue damage. These chemicals (and capsaicin) cause the TRPV1 channel to open at lower temperatures, explaining the heightened sensitivity of injured skin to increases in temperature. Indeed, inflammation-induced thermal hyperalgesia is absent in mice engineered to lack the TRPV1 channel. While all mammals normally express the TRPV1 channel, birds do not, thus explaining how birds can consume the spiciest of chili peppers. This fact also explains how birdseed laced with capsaicin can be enjoyed by birds without interference from raiding squirrels.
In addition to its protection of birdseed and widespread culinary use, capsaicin has a seemingly paradoxical clinical application. Applied in large quantities, it can cause analgesia, the absence of pain. The capsaicin desensitizes pain fibers and depletes the peptide substance P from their nerve terminals. Capsaicin ointments, sprays, and patches are useful treatments for pain associated with arthritis, strains, psoriasis, shingles, and other conditions (see Box 12.1).
Nociceptors are present in most body tissues, including skin, bone, muscle, most internal organs, blood vessels, and the heart. They are notably absent in the brain itself, except for the meninges.
Hyperalgesia and Inflammation. Nociceptors normally respond only when stimuli are strong enough to damage tissue. But we all know that skin, joints, or muscles that have already been damaged or inflamed are unusually sensitive. A light, sympathetic mother’s touch to a burned area of her child’s skin may elicit howls due to unbearable pain. This phenomenon is known as hyperalgesia, and it is the most familiar example of our body’s ability to control its own pain. Hyperalgesia can be a reduced threshold for pain, an increased intensity of painful stimuli, or even spontaneous pain. Primary hyperalgesia occurs within the area of damaged tissue, but tissues surrounding a damaged area may become supersensitive as well, by the process of secondary hyperalgesia.
Many different mechanisms appear to be involved in hyperalgesia, some in and around the peripheral receptors and others within the CNS. As noted earlier, when skin is damaged, a variety of substances, sometimes called the inflammatory soup, are released. The soup includes certain neurotransmitters (glutamate, serotonin, adenosine, ATP), peptides (substance P, bradykinin), lipids (prostaglandins, endocannabinoids), proteases, neurotrophins, cytokines, and chemokines, ions such as K+ and H+, and other substances (Figure 12.25). Together, they can trigger inflammation, which is a natural response of the body’s tissues as they attempt to eliminate injury and stimulate the healing process. The cardinal signs of inflammation in skin are pain, heat, redness, and swelling. A number of these chemicals can also modulate the excitability of nociceptors, making them more sensitive to thermal or mechanical stimuli (see Box 12.5).

FIGURE 12.25 Peripheral chemical mediators of pain and hyperalgesia. Description
Bradykinin was discussed earlier as one of the chemicals that directly depolarizes nociceptors. In addition to this effect, bradykinin stimulates long-lasting intracellular changes that make heat-activated ion channels more sensitive. Prostaglandins are chemicals generated by the enzymatic breakdown of lipid membrane. While prostaglandins do not elicit overt pain, they do increase greatly the sensitivity of nociceptors to other stimuli. Aspirin and other nonsteroidal anti-inflammatory drugs are a useful treatment for hyperalgesia because they inhibit the enzymes required for prostaglandin synthesis.
Substance P is a peptide synthesized by the nociceptors themselves. Activation of one branch of a nociceptor axon can lead to the secretion of substance P by the other branches of that axon in the neighboring skin. Substance P causes vasodilation (swelling of the blood capillaries) and the release of histamine from mast cells. Sensitization of other nociceptors around the site of injury by substance P is one cause of secondary hyperalgesia.
CNS mechanisms also contribute to secondary hyperalgesia. Following injury, the activation of mechanoreceptive Aβ axons by light touch can evoke pain. Thus, another mechanism of hyperalgesia involves cross-talk between the touch and pain pathways in the spinal cord.
Itch is defined as a disagreeable sensation that induces a desire or a reflex to scratch. Itch and the scratching it evokes can serve as a natural defense against parasites and plant toxins on the skin and scalp. Itch is usually a brief and minor annoyance. It can also become a chronic and seriously debilitating condition. Chronic itch may be caused by a wide variety of skin conditions such as allergic reactions, infections, infestations, and psoriasis; it can also be triggered by non-skin disorders such as cancers, iron deficiency, hyperthyroidism, liver disease, stress, and psychiatric conditions. Imagine that your worst itch has spread across most of your body and persists through every waking minute. The need to scratch can be unrelenting and irresistible. Chronic itch can be as dreadful as chronic pain, and it is notoriously difficult to treat with current drugs and therapies.
Itch has always been a hard sensation to categorize. Although pain and itch are unmistakably distinct, they also have many similarities. Both are mediated by thin sensory axons, although the axons carrying pain signals seem to be different from those triggering itch. Both can be triggered by various types of stimuli, including chemicals and touch. Some of the drugs and compounds that regulate pain can also trigger itch, and some signaling molecules transduce both sensations. Pain and itch also interact. For example, pain can suppress itch; this is why we sometimes aggressively scratch an itchy patch of skin.
Some types of itch are triggered by specific molecules and neural circuits. The very smallest C fibers (conduction velocity of 0.5 m/sec or less) are selectively responsive to histamine, the natural itch-producing substance that is released by mast cells in the skin during inflammation (see Figure 12.25). Histamine mediates itch by binding to histamine receptors, which then activate TRPV1 channels; surprisingly, these are the same types of TRPV1 channels that are stimulated by capsaicin and high temperature (see Box 12.5). Antihistamines—drugs that antagonize histamine receptors—can suppress this kind of itch. Not all itch is mediated by histamine, however. Itch can also be triggered by a wide variety of endogenous and exogenous substances, and thin itch-mediating axons seem to express a large number of other itch-producing types of receptors, signaling molecules, and membrane channels.
Many things about itch remain mysterious. It is not clear whether there are different types of itch-producing axons. The central circuits involved in itch are also poorly understood. One fascinating study implicated certain peptide neurotransmitters in specific itch-producing pathways in the spinal cord. If specific signaling molecules and receptors mediating itch can be identified and understood, it may be possible to develop effective and selective drugs to treat chronic itch without affecting pain and other somatic sensory processes.
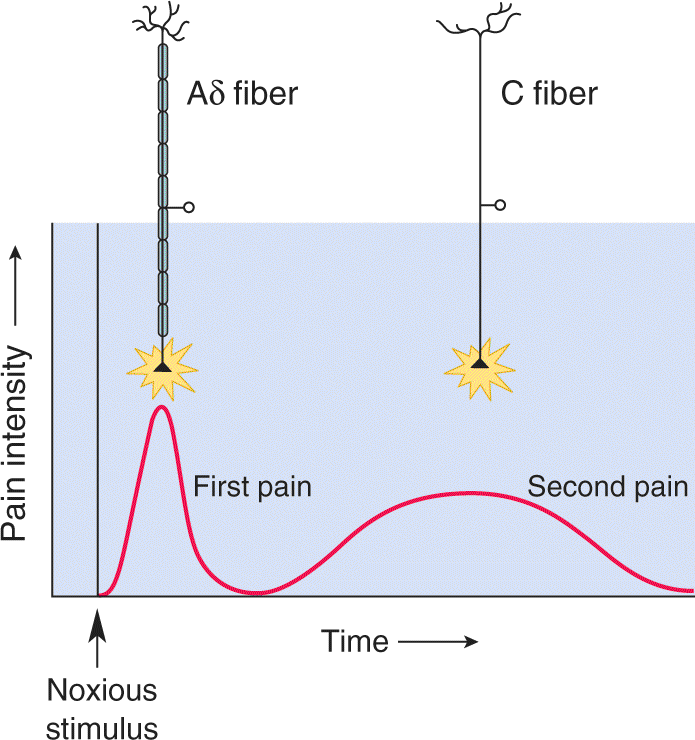
Aδ and C fibers bring information to the CNS at different rates because of differences in their action potential conduction velocities. Accordingly, the activation of skin nociceptors produces two distinct perceptions of pain: a fast, sharp, first pain followed by a duller, longer lasting second pain. First pain is caused by the activation of Aδ fibers; second pain is caused by the activation of C fibers (Figure 12.26).

FIGURE 12.26 First and second pain. The first pain sensation registered by noxious stimulation is mediated by fast Aδ axons. The second, longer lasting pain sensation is mediated by slow C fibers. Description
Like the Aβ mechanosensory fibers, the small-diameter fibers have their cell bodies in the segmental dorsal root ganglia, and they enter the dorsal horn of the spinal cord. The fibers branch immediately, travel a short distance up and down the spinal cord in a region called the zone of Lissauer, and then synapse on cells in the outer part of the dorsal horn in a region known as the substantia gelatinosa (Figure 12.27).

FIGURE 12.27 Spinal connections of nociceptive axons. Description
The neurotransmitter of the pain afferents is glutamate; however, as mentioned previously, these neurons also contain the peptide substance P (Figure 12.28). Substance P is contained within storage granules in the axon terminals (see Chapter 5) and can be released by high-frequency trains of action potentials. Recent experiments have shown that synaptic transmission mediated by substance P is required to experience moderate to intense pain.

FIGURE 12.28 Immunocytochemical localization of substance P in the spinal cord. Arrows point to strong concentrations of substance P in the substantia gelatinosa. (Source: Mantyh et al., 1997.)
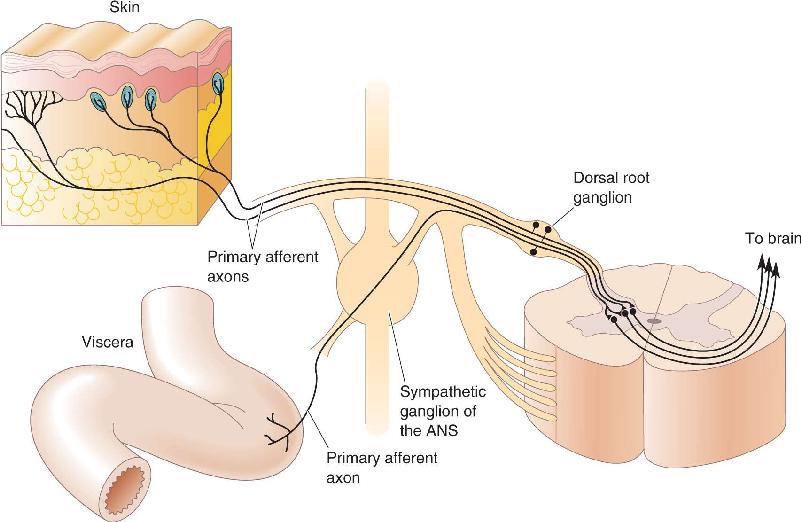
It is interesting to note that nociceptor axons from the viscera enter the spinal cord by the same route as the cutaneous nociceptors. Within the spinal cord, there is substantial mixing of information from these two sources of input (Figure 12.29). This cross-talk gives rise to the phenomenon of referred pain, where visceral nociceptor activation is perceived as a cutaneous sensation. The classic example of referred pain is angina, occurring when the heart fails to receive sufficient oxygen. Patients often localize the pain of angina to the upper chest wall and the left arm. Another common example is the pain associated with appendicitis, which in its early stages is referred to the abdominal wall around the navel.

FIGURE 12.29 The convergence of nociceptor input from the viscera and the skin. Description
Let’s briefly highlight the differences we’ve encountered between the touch and pain pathways. First, they differ with respect to their nerve endings in the skin. The touch pathway is characterized by specialized structures in the skin; the pain pathway has only free nerve endings. Second, they differ with respect to the diameter of their axons. The touch pathway is swift, using fat, myelinated Aβ fibers; the pain pathway is slow, using thin, lightly myelinated Aδ fibers and unmyelinated C fibers. Third, they differ with respect to their connections in the spinal cord. Branches of the Aβ axons terminate in the deep dorsal horn; the Aδ and C fibers branch, run within the zone of Lissauer, and terminate within the substantia gelatinosa. As we will now see, the two pathways also differ substantially in the way they transmit information to the brain.
The Spinothalamic Pain Pathway. Information about pain (as well as temperature) in the body is conveyed from the spinal cord to the brain via the spinothalamic pathway. Unlike the dorsal column–medial lemniscal pathway, the axons of the second-order neurons immediately decussate and ascend through the spinothalamic tract running along the ventral surface of the spinal cord (compare Figures 12.14 and 12.27). As the name implies, the spinothalamic fibers project up the spinal cord and through the medulla, pons, and midbrain without synapsing, until they reach the thalamus (Figure 12.30). As the spinothalamic axons journey through the brain stem, they eventually come to lie alongside the medial lemniscus, but the two groups of axons remain distinct from each other.

FIGURE 12.30 The spinothalamic pathway. This is the major route by which pain and temperature information ascend to the cerebral cortex. Description
Figure 12.31 summarizes the different ascending pathways for touch and pain information. Notice that information about touch ascends ipsilaterally, while information about pain (and temperature) ascends contralaterally. This organization can lead to a curious, but predictable, group of deficits when the nervous system is impaired. For example, if half of the spinal cord is damaged, certain deficits of mechanosensitivity occur on the same side as the spinal cord damage: insensitivity to light touch, the vibrations of a tuning fork on the skin, the position of a limb. On the other hand, deficits in pain and temperature sensitivity will show up on the side of the body opposite the cord damage. Other signs, such as motor deficiency and the exact map of sensory deficits, give additional clues about the site of spinal cord damage. For example, movements will be impaired on the ipsilateral side. The constellation of sensory and motor signs following damage to one side of the spinal cord is called Brown–Séquard syndrome.

FIGURE 12.31 An overview of the two major ascending pathways of somatic sensation. Description
The Trigeminal Pain Pathway. Pain (and temperature) information from the face and head takes a path to the thalamus that is analogous to the spinal path. The small-diameter fibers in the trigeminal nerve synapse first on second-order sensory neurons in the spinal trigeminal nucleus of the brain stem. The axons of these cells cross and ascend to the thalamus in the trigeminal lemniscus.
In addition to the spinothalamic and trigeminothalamic pathways, other closely related pain (and temperature) pathways send axons into a variety of structures at all levels of the brain stem, before they reach the thalamus. Some of these pathways are particularly important in generating sensations of slow, burning, agonizing pain, while others trigger a more general state of behavioral arousal and alertness.
The Thalamus and Cortex. The spinothalamic tract and trigeminal lemniscal axons synapse over a wider region of the thalamus than those of the medial lemniscus. Some of the axons terminate in the VP nucleus, just as the medial lemniscal axons do, but the touch and pain systems still remain segregated there by occupying separate regions of the nucleus. Other spinothalamic axons end in the small intralaminar nuclei of the thalamus (Figure 12.32). From the thalamus, pain and temperature information is projected to various areas of the cerebral cortex. As in the thalamus, this pathway covers a much wider territory than the cortical connections of the dorsal column–medial lemniscal pathway.

FIGURE 12.32 Somatic sensory nuclei of the thalamus. In addition to the VP nucleus, the intralaminar nuclei relay nociceptive information to a large expanse of the cerebral cortex.
The perception of pain is highly variable. Depending on the concurrent level of nonpainful sensory input and the behavioral context, the same level of nociceptor activity can produce more pain or less pain. Understanding this modulation of pain is of great importance because it may offer new strategies for the treatment of chronic pain, a condition that afflicts up to 20% of the adult population.
Afferent Regulation. We’ve already seen that light touch can evoke pain via the mechanisms of hyperalgesia. However, pain evoked by activity in nociceptors can also be reduced by simultaneous activity in low-threshold mechanoreceptors (Aβ fibers). Presumably, this is why it feels good to rub the skin around your shin when you bruise it. This may also explain an electrical treatment for some kinds of chronic, intractable pain. Wires are taped to the skin surface, and pain is suppressed when the patient simply turns on an electrical stimulator designed to activate large-diameter sensory axons.
In the 1960s, Ronald Melzack and Patrick Wall, then working at MIT, proposed a hypothesis to explain these phenomena. Their gate theory of pain suggests that certain neurons of the dorsal horns, which project an axon up the spinothalamic tract, are excited by both large-diameter sensory axons and unmyelinated pain axons. The projection neuron is also inhibited by an interneuron, and the interneuron is both excited by the large sensory axon and inhibited by the pain axon (Figure 12.33). By this arrangement, activity in the pain axon alone maximally excites the projection neuron, allowing nociceptive signals to rise to the brain. However, if the large mechanoreceptive axon fires concurrently, it activates the interneuron and suppresses nociceptive signals.

FIGURE 12.33 Melzack’s and Wall’s gate theory of pain. The relay of nociceptive signals by the projection neuron is gated by the activity of an inhibitory interneuron. Activity in the non-nociceptive mechanoreceptor can suppress, or close the “gate” on, nociceptive signals before they can proceed to the spinothalamic tract. The + signs indicate excitatory synapses and the – signs indicate inhibitory synapses. Description
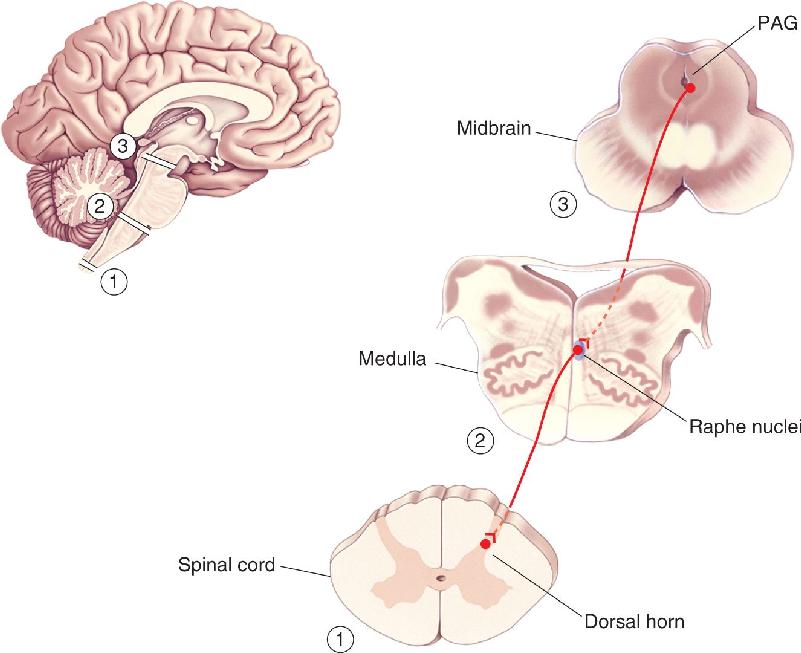
Descending Regulation. Stories abound of soldiers, athletes, and torture victims who sustained horrible injuries but apparently felt no pain. Strong emotion, stress, or stoic determination can powerfully suppress feelings of pain. Several brain regions have been implicated in pain suppression (Figure 12.34). One is a zone of neurons in the midbrain called the periventricular and periaqueductal gray matter (PAG). Electrical stimulation of the PAG can cause a profound analgesia that has sometimes been exploited clinically.

FIGURE 12.34 Descending pain-control pathways. A variety of brain structures, many of which are affected by behavioral state, can influence activity within the periaqueductal gray matter (PAG) of the midbrain. The PAG can influence the raphe nuclei of the medulla, which in turn can modulate the flow of nociceptive information through the dorsal horns of the spinal cord. Description
The PAG normally receives input from several brain structures, many of them appropriate for transmitting signals related to emotional status. PAG neurons send descending axons into various midline regions of the medulla, particularly to the raphe nuclei (which use the neurotransmitter serotonin). These medullary neurons in turn project axons down to the dorsal horns of the spinal cord, where they can effectively depress the activity of nociceptive neurons.
The Endogenous Opioids. Opium was probably known to the ancient Sumerians around 4000 BCE. Their pictograph for the poppy roughly translates to “joy plant.” By the seventeenth century, the therapeutic value of opium was undisputed. Opium, its active narcotic ingredients, and their analogues—including morphine, codeine, and heroin—are used and abused widely today, in most cultures of the world. These drugs, and others with similar actions, are called opioids, and they produce profound analgesia when taken systemically (see Chapter 6). They can also produce mood changes, drowsiness, mental clouding, nausea, vomiting, and constipation. The 1970s brought the stunning discoveries that opioids act by binding tightly and specifically to several types of opioid receptors in the brain, and that the brain itself manufactures endogenous morphine-like substances, collectively called endorphins (see Box 6.1). Endorphins are relatively small proteins, or peptides.
Endorphins and their receptors are distributed widely in the CNS, but they are particularly concentrated in areas that process or modulate nociceptive information. Small injections of morphine or endorphins into the PAG, the raphe nuclei, or the dorsal horn can produce analgesia. Because this effect is prevented by administering the specific blocker of opioid receptors, naloxone, the injected drugs must have acted by binding to opioid receptors in those areas. Naloxone can also block the analgesic effects induced by electrically stimulating these areas. At the cellular level, endorphins exert multiple effects that include suppressing the release of glutamate from presynaptic terminals and inhibiting neurons by hyperpolarizing their postsynaptic membranes. In general, extensive systems of endorphin-containing neurons in the spinal cord and brain stem prevent the passage of nociceptive signals through the dorsal horn and into higher levels of the brain where the perception of pain is generated (Box 12.6).
To test the efficacy of a new drug, clinical trials are often conducted in which one group of subjects receives the drug and the other receives an inert substance. Both groups of subjects may believe they have been given the drug. Surprisingly, the inert substance is often reported to have the effect that patients are told to expect from the drug. The term placebo is used to describe such substances (the word is from the Latin meaning, “I shall please”), and the phenomenon is called the placebo effect.
Placebos can be highly effective analgesics. A majority of patients suffering from postoperative pain have reported getting relief from an injection of sterile saline! Does this mean that these patients have only imagined their pain? Not at all. The opioid receptor antagonist naloxone can block the analgesic effect of the placebo, just as it antagonizes the effects of morphine, a true analgesic. Apparently, the belief that the treatment will work can be enough to cause activation of the endogenous pain-relief systems of the brain. The placebo effect is a likely explanation for the success of other treatments for pain, such as acupuncture, hypnosis, and, for children, a mother’s loving kiss.
As is the case for the sense of touch and pain, nonpainful temperature sensations originate from receptors in the skin (and elsewhere), and they depend on the neocortex for their conscious appreciation. Here, we briefly describe how this system is organized.
Because the rate of chemical reactions depends on temperature, the functioning of all cells is sensitive to temperature. However, thermoreceptors are neurons that are exquisitely sensitive to temperature because of specific membrane mechanisms. For example, we can perceive changes in our average skin temperature of as little as 0.01 °C. Temperature-sensitive neurons clustered in the hypothalamus and the spinal cord are important in the physiological responses that maintain stable body temperature, but it is the thermoreceptors in the skin that apparently contribute to our perception of temperature.
Temperature sensitivity is not spread uniformly across the skin. With a small cold or warm probe, you can map your skin’s sensitivity to temperature changes. Some spots about 1 mm wide are especially sensitive to either hot or cold but not both. The fact that the locations of hot and cold sensitivity are different demonstrates that separate receptors encode them. Also, small areas of skin between the hot and cold spots are relatively insensitive to temperature.
The sensitivity of a sensory neuron to a change in temperature depends on the type of ion channels the neuron expresses. Discovery of the channels responsive to painful increases in temperature above 43 °C (see Box 12.5) led researchers to wonder if other, closely related channels might be tuned to sense other temperature ranges. Just as the active ingredient in hot peppers was used to identify the “hot” receptor protein, called TRPV1, the active ingredient in mint was used to identify a “cold” receptor. Menthol, which produces a sensation of cold, was found to stimulate a receptor, called TRPM8, which is also activated by nonpainful decreases in temperature below 25 °C.
We now understand that there are six distinct TRP channels in thermoreceptors that confer different temperature sensitivities (Figure 12.35). As a rule, each thermoreceptive neuron appears to express only a single type of channel, thus explaining how different regions of skin can show distinctly different sensitivities to temperature. An exception appears to be some cold receptors that also express TRPV1 and are therefore also sensitive to increases in temperature above 43 °C. If such heat is applied to wide areas of skin, it is usually painful, but if the heat is restricted to small regions of skin innervated by a cold receptor, it can produce a paradoxical feeling of cold. This effect emphasizes an important point: The CNS does not know what kind of stimulus (in this case, heat) caused the receptor to fire, but it continues to interpret all activity from its cold receptor as a response to cold.

FIGURE 12.35 Thermoreceptor TRP channels tuned to detect different temperatures. (a) The arrangement of the known thermosensitive TRP channel protein molecules in the neuronal membrane. TRPM8 and TRPVI are responsive to menthol and capsaicin, respectively. (b) This graph plots the activation of the various TRP channels as a function of temperature. (Source: Adapted from Patapoutian, et al., 2003, Fig. 3.) Description
As with mechanoreceptors, the responses of thermoreceptors adapt during long-duration stimuli. Figure 12.36 shows that a sudden drop in skin temperature causes a cold receptor to fire strongly, while it silences a warm receptor. After a few seconds at 32 °C, however, the cold receptor slows its firing (but still fires faster than it did at 38 °C), while the warm receptor speeds up slightly. Notice that a return to the original warm skin temperature causes opposite responses—transient silence of the cold receptor and a burst of activity in the warm receptor—before both return to their steady, adapted rates. Thus, the differences between the response rates of warm and cold receptors are greatest during, and shortly after, temperature changes. Our perceptions of temperature often reflect these skin receptor responses.

FIGURE 12.36 Adaptations of thermoreceptors. The responses of cold and warm receptors to a step reduction in skin temperature are shown. Both receptors are most responsive to sudden changes in temperature, but they adapt over several seconds. Description
Try a simple experiment. Fill two buckets with tap water, one cold and one hot (but not painfully hot). Then plunge your right hand for one minute into each of them in turn. Notice the striking sensations of hot and cold that occur with each change, but also notice how transient the sensations are. With thermoreception, as with most other sensory systems, it is the sudden change in the quality of a stimulus that generates the most intense neural and perceptual responses.
At this point, you may be relieved to learn that the organization of the temperature pathway is virtually identical to that of the pain pathway already described. Cold receptors are coupled to Aδ and C fibers, while warm receptors are coupled only to C fibers. As we learned previously, small-diameter axons synapse within the substantia gelatinosa of the dorsal horn. The axons of second-order neurons immediately decussate and ascend in the contralateral spinothalamic tract (see Figure 12.30). Thus, if the spinal cord is transected on one side, there will be a loss of temperature sensitivity (as well as pain) on the opposite side of the body, specifically in those regions of skin innervated by spinal segments below the cut.
This concludes our discussion of the sensory systems. Although each one has evolved to be the brain’s interface with a different form of environmental energy, the systems are strikingly similar in organization and function. Different types of somatic sensory information are necessarily kept separate in the spinal nerves because each axon is connected to only one type of sensory receptor ending. Segregation of sensory types continues within the spinal cord and is largely maintained all the way to the cerebral cortex. In this way, the somatic sensory system repeats a theme common throughout the nervous system: Several flows of related, but distinct, information are passed in parallel through a series of neural structures. The mixing of these streams occurs along the way, but only judiciously, until higher levels of processing are reached in the cerebral cortex. We saw other examples of parallel processing of sensory information in the chemical senses, vision, and audition.
Exactly how the parallel streams of sensory data are melded into perception, images, ideas, and memories remains the Holy Grail of neuroscience. Thus, the perception of any handled object involves the seamless coordination of all facets of somatic sensory information. The bird in hand is rounded, warm, soft, and light in weight; its heartbeat flutters against your fingertips; its claws scratch; and its textured wings brush against your palm. Somehow, your brain knows it’s a bird, even without looking or listening, and would never mistake it for a toad. In the chapters that follow, we describe how the brain begins to use sensory information to plan and coordinate movement.
1. Imagine rubbing your fingertips across a pane of smooth glass and then across a brick. What kinds of skin receptors help you distinguish the two surfaces? As far as your somatic sensory system is concerned, what is different about the two surfaces?
2. What purpose is served by the encapsulations around some sensory nerve endings in the skin?
3. If someone tossed you a hot potato and you caught it, which information would reach your CNS first: the news that the potato was hot or that it was relatively smooth? Why?
4. At what levels of the nervous system are all types of somatic sensory information represented on the contralateral side: the spinal cord, the medulla, the pons, the midbrain, the thalamus, and the cortex?
5. What lobe of the cortex contains the main somatic sensory areas? Where are these areas relative to the main visual and auditory areas?
6. Where within the body can pain be modulated, and what causes its modulation?
7. Where in the CNS does information about touch, shape, temperature, and pain converge?
8. Imagine this experiment: Fill two buckets with water, one relatively cold and one hot. Fill a third bucket with water of an intermediate, lukewarm temperature. Put your left hand into the hot water, your right hand into the cold, and wait one minute. Now quickly plunge both hands into the lukewarm water. Try to predict what sensations of temperature you will feel in each hand. Will they feel the same? Why?
Abraira VE, Ginty DD. 2013. The sensory neurons of touch. Neuron 79:618–639.
Braz J, Solorzano C, Wang X, Basbaum AI. 2014. Transmitting pain and itch messages: a contemporary view of the spinal cord circuits that generate gate control. Neuron 82:522–536.
Di Noto PM, Newman L, Wall S, Einstein G. 2013. The hermunculus: what is known about the representation of the female body in the brain? Cerebral Cortex 23:1005–1013.
Eijkelkamp N, Quick K, Wood JN. 2013. Transient receptor potential channels and mechanosensation. Annual Review of Neuroscience 36:519–546.
Fain GL. 2003. Sensory Transduction. Sunderland, MA: Sinauer.
Hsiao S. 2008. Central mechanisms of tactile shape perception. Current Opinion in Neurobiology 18:418–424.
McGlone F, Wessberg J, Olausson H. 2014. Discriminative and affective touch: sensing and feeling. Neuron 82:737–755.
Vallbo Å. 1995. Single-afferent neurons and somatic sensation in humans. In The Cognitive Neurosciences, ed. Gazzaniga M. Cambridge, MA: MIT Press, pp. 237–251.
Additional figures