Introduction
Chapter 20 Integrating Cells into Tissues Antibody staining of mouse small intestinal villi, fingerlike projections of cell layers (see Figures 20-13 through 20-15). NPC1L1, a protein located primarily on the plasma membrane involved with cholesterol metabolism, is green. Villin, which binds to actin bundles in microvilli (very small membrane projections on the apical surfaces of absorptive cells that
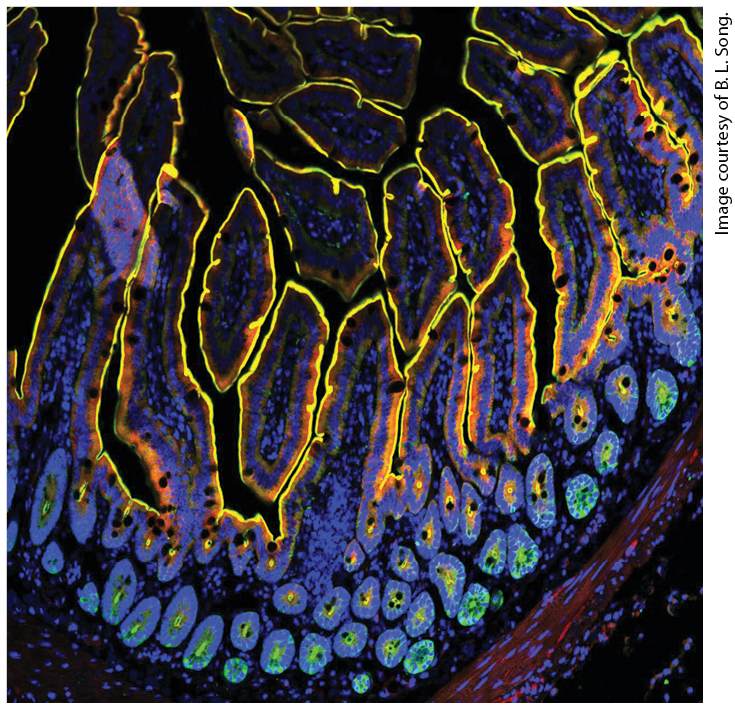
take up digested nutrients), is red. Nuclei (DNA) are blue. The image shows all colors merged together (colocalization of green and red appears yellow). The yellow lines show how the apical surfaces of the cells face the lumen of the small intestine (black) from which nutrients are absorbed.
20.1 Cell-Cell and Cell–Extracellular Matrix Adhesion: An Overview
20.2 Cell-Cell and Cell–Extracellular Matrix Junctions and Their Adhesion Molecules
20.4 The Extracellular Matrix II: Connective Tissue
20.5 Adhesive Interactions in Motile and Nonmotile Cells
20.6 Plant Tissues In the development of complex multicellular organisms such as plants and animals, progenitor cells differentiate into distinct types that have characteristic compositions, structures, and functions. Cells of a given type often aggregate into a tissue to cooperatively perform a common function — muscle contracts, neural tissue conducts electric impulses, xylem tissue in plants transports water. Different tissues can be organized into an organ, again to perform one or more specific functions. For instance, the muscles, valves, and blood vessels of a heart work together to pump blood. The coordinated functioning of many types of cells and tissues permits the organism to move, metabolize, reproduce, and carry out other essential activities. Indeed, the complex and diverse morphologies of plants and animals are examples of the whole being greater than the sum of the individual parts. Vertebrates have many hundreds of different cell types, including leukocytes (white blood cells) and erythrocytes (red blood cells), photoreceptors in the retina, fat-storing adipocytes, fibroblasts in
connective tissue, and the hundreds of different subtypes of neurons in the human brain. A large-scale international effort, the Human Cell Atlas Project, is using single-cell RNA sequencing and other methods to generate a comprehensive catalogue of all human cell types and subtypes. Multicellular animals — also called metazoans — usually comprise differentiated cells organized into distinct tissues, although there are some less complex exceptions (e.g., sponges). Even rather simple animals exhibit distinctive tissue organization. The adult form of the roundworm Caenorhabditis elegans contains a mere 959 cells, yet these cells fall into 12 different general cell types and many distinct subtypes. Despite their diverse forms and functions, animal cells typically can be classified as components of just five main classes of tissue: epithelial tissue, connective tissue, muscular tissue, neural tissue, and blood. Various cell types are arranged in precise patterns of staggering complexity to generate tissues and organs. The physiological costs of such complexity include increased requirements for information, material, energy, and time during the development of an individual organism. Although these costs of generating and maintaining complex tissues and organs are high, they confer the ability to thrive in varied and variable environments — a major evolutionary advantage. One of the defining characteristics of animals with complex tissues and organs (most metazoans) is that the external and internal surfaces of most of their tissues and organs — and indeed, the exterior of the entire organism — are built from tightly packed sheet-like layers of cells known as epithelia. The formation of an epithelium and its subsequent
remodeling into more complex collections of epithelial and nonepithelial tissues is a hallmark of the development of most metazoans. Sheets of tightly attached epithelial cells act as regulatable, selectively permeable barriers, which permit the generation of chemically and functionally distinct compartments in an organism, such as the stomach and bloodstream. As a result, distinct and sometimes opposite functions (e.g., digestion and synthesis) can efficiently proceed simultaneously within an organism. Such compartmentalization also permits more sophisticated regulation of diverse biological functions. In many ways, the roles of complex tissues and organs in an organism are analogous to those of organelles and membranes in individual cells. The assembly of distinct tissues and their organization into organs are determined by molecular interactions at the cellular level (Figure 20-1). These interactions would not be possible without the temporally, spatially, and functionally regulated expression of a wide array of adhesion molecules. Cell-surface molecules engaged in adhesion are called adhesion receptors. Cells in tissues can adhere directly to one another (cell-cell adhesion) through specialized membrane proteins called celladhesion molecules (CAMs) or cell-cell adhesion receptors, which often cluster on the plasma membrane, sometimes into specialized structures called cell junctions. In the fruit fly Drosophila melanogaster, at least 500 genes (∼4 percent of the total) are estimated to be involved in cell adhesion, and in mammals there are over 1000 such genes. Cells in animal tissues also adhere indirectly (cell-matrix adhesion) through the binding of cell-matrix adhesion receptors in the plasma membrane to components of the surrounding extracellular matrix (ECM), a complex
interdigitating meshwork of proteins and polysaccharides secreted by cells into the spaces between them. Some classes of adhesion receptors (e.g., integrins) can mediate both cell-cell adhesion as well as cell-matrix adhesion.
FIGURE 20-1 Overview of major cell-cell and cell-matrix adhesive interactions. Schematic cutaway drawing of a typical epithelial tissue, such as in the inner surface of the intestines. The apical (upper) surface of each cell is packed with fingerlike microvilli ( 1 ) that project into the intestinal lumen; the basal (lower) surface ( 2 ) rests on extracellular matrix (ECM). The ECM ( 3 ) associated with epithelial cells is usually organized into various interconnected layers — such as the basal lamina ( 3a ), connecting fibers (not shown), and connective tissue ( 3b ) — in which large, interdigitating ECM macromolecules bind to one another and to the cells ( 3 ). Cell-adhesion molecules (CAMs, also called cell-cell adhesion receptors) bind to CAMs on other cells, mediating cell-cell adhesion ( 4 ), and cell-matrix adhesion receptors bind to various components of the ECM, mediating cell-matrix adhesion ( 5 ). Both types of adhesion receptors are usually integral membrane proteins whose cytosolic domains often bind to multiple intracellular adapter proteins. These adapters, directly or indirectly, link the CAM to the cytoskeleton (actin or intermediate filaments) and to intracellular signaling pathways (see Figure 20-8). As a
consequence, information can be transferred by adhesion receptors and the macromolecules to which they bind from the cell exterior to the intracellular environment and vice versa. In some cases, a complex aggregate of adhesion receptors, adapters, and associated proteins is assembled. Specific, localized aggregates of adhesion receptors form various types of cell junctions, which play important roles in holding tissues together and facilitating communication between cells and their environment. Tight junctions ( 6 ), lying just under the apical surface, prevent the diffusion of many substances through the extracellular spaces between the cells. Through connexon channels, gap junctions ( 7 ) allow the movement of small molecules and ions between the cytosols of adjacent cells. The remaining three types of junctions, adherens junctions ( 8 and 4 ), desmosomes ( 9 ), hemidesmosomes ( 10 and 5 ), and focal adhesions (also called focal contacts; 11 ) link the cytoskeleton of a cell to other cells or to the ECM. See V. Vasioukhin and E. Fuchs, 2001, Curr. Opin. Cell Biol. 13:76–84. Description The illustration on the left shows two ovals, the top one is labeled tight junction and shows a vertical line of cells close together with pieces coming off to the right. The oval below is labeled gap junction and shows two yellow vertical lines with two orange tubes connecting them at the center. The top tube is closed and the bottom one is open and is labeled connexon. In the center of the illustration is a schematic of a sample of the intestinal wall. At the top is a wavy line labeled apical surface, representing the villi. Between two villi are examples of the chain of cells going down to the basal lamia at the bottom of the diagram. The ovals from the left side are indicated as enlargements of two areas on the left example. The right example has a green oval half way batten villi and lamia and labeled adherens junction, and a blue oval further down labeled desmosome. The lamia shows root-like lines. At the right of the illustration, an oval is at the top, labeled cell-cell adhesions. It shows chains of cells labeled, from left to right, Adapters, cell adhesion molecules (C A M's), then actin. The lower oval is labeled cellmatrix adhesions. It shows adapters attached to actin at the top, attached below to adhesion receptors, which are then lowered into a blue area labeled E C M. Cell-cell and cell-matrix adhesions not only allow cells to aggregate into distinct tissues, but also provide a means for the bidirectional transfer of
information between the exterior and the interior of cells. As we will see, both types of adhesions are intrinsically associated with the cytoskeleton and cellular signaling pathways. As a result, a cell’s surroundings influence its shape and functional properties (“outside-in” effects); likewise, cellular shape and function influence a cell’s surroundings (“inside-out” effects). Thus connectivity and communication are intimately related properties of cells in tissues. Information transfer is important to many biological processes, including cell survival, proliferation, differentiation, and migration. Therefore, it is not surprising that defects that interfere with adhesive interactions and the associated flow of information can cause or contribute to diseases, including a wide variety of neuromuscular and skeletal disorders, heart and vascular defects, clotting abnormalities, cancer, and others. In this chapter, we examine various types of adhesion molecules found on the surfaces of cells and in the surrounding extracellular matrix. Interactions between these molecules allow the organization of cells into tissues and have profound effects on tissue development, function, and pathology. Many adhesion molecules are members of families or superfamilies of related proteins. While each type of adhesion molecule performs a distinct role, we will focus on the common features shared by members of some of these families to illustrate the general principles underlying their structures and functions. Because of the particularly wellunderstood nature of the adhesion molecules in tissues that form tight epithelia, as well as their very early evolutionary development, we will initially focus on epithelial tissues, such as the walls of the intestinal tract and the skin. Epithelial cells are normally nonmotile (immobile), also
called sessile; however, during development, wound healing, and in certain pathological states (e.g., cancer), epithelial cells can transform into motile cells. Changes in the expression and function of adhesion molecules play a key role in this transformation, as they do in normal biological processes involving cell movement, such as the crawling of white blood cells into sites of infection. We therefore follow the discussion of epithelial tissues with a discussion of adhesion in nonepithelial, developing, and motile tissues. The evolutionary lineages of plants and animals diverged before multicellular organisms arose (see Figure 1-1). Thus multicellularity and the molecular means for assembling tissues and organs presumably arose independently in animal and plant lineages. Not surprisingly, then, animals and plants exhibit many differences in the organization and development of tissues. For this reason, we first consider the organization of tissues in animals and then deal separately with plants.
Cell-Adhesion Molecules Bind to One Another and to Intracellular Proteins
20.1 Cell-Cell and Cell–Extracellular Matrix Adhesion: An Overview There are many different types of cells in the body that dynamically interact with each other in a myriad of ways. These interactions, achieved via adhesion molecules, must be precisely and carefully controlled in time and space and in their physical characteristics (e.g., strength of adhesion) to correctly determine the structures and functions of tissues in a complex organism. It is not surprising, therefore, that cell-cell and cell-matrix adhesion molecules exhibit diverse structures, or that their expression levels vary in different cells and tissues. As a consequence, they mediate the very specific and distinctive cell-cell and cell-matrix interactions that hold tissues together as well as permit essential communication between cells and their environment. We begin this overview with a brief orientation to some of the various types of adhesion molecules present on cells and within the extracellular matrix, their major functions in organisms, and their evolutionary origin. In subsequent sections, we will examine in detail the unique structures and properties of various participants in cell-cell and cell-matrix interactions. Cell-Adhesion Molecules Bind to One Another and to Intracellular Proteins Cell-cell adhesion is mediated through membrane proteins called cell-adhesion molecules (CAMs, also called cell-cell adhesion receptors). Many CAMs fall into one of four major families: the cadherins, the immunoglobulin (Ig) superfamily, the integrins, and sugar-binding proteins called lectins, one subset of which are called selectins. As the schematic structures in Figure 20-2 illustrate, CAMs often comprise multiple, distinct domains, many of which can be found in more than one kind of protein. The functions of these domains vary. Some confer the ability to bind specifically to their partner CAMs on neighboring cells or even to CAMs on the same cell. Some of these domains are present in multiple copies and contribute to the length of the CAMs and thus help define the distance between the plasma membranes of cells bound together by the CAMs. Other membrane proteins, whose structures do not belong to any of the major classes of CAMs in Figure 20-2, are also adhesion receptors and participate in cell-cell and cell-matrix adhesion in various tissues. One example is a set of adhesion G protein–coupled receptors (GPCRs, see Chapter 15). As we will see later, integrins can function both as cell-cell and cell-matrix adhesion receptors. Figure 20-2 depicts an integrin serving as a cell-matrix adhesion receptor that binds to ECM components. Some Ig-superfamily CAMs can play this dual role as well.
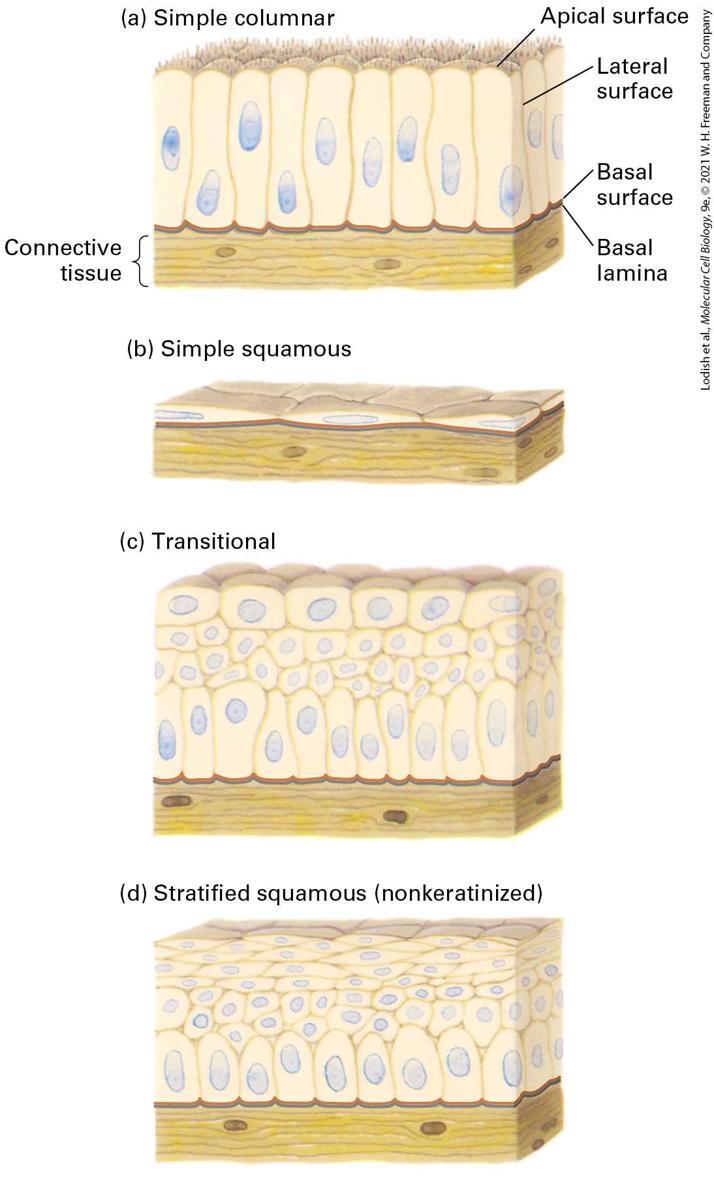
FIGURE 20-2 Major families of cell-adhesion molecules (CAMs) and adhesion receptors. Adhesion receptors can mediate binding, either by homophilic interactions (binding to other molecules of the same type; left) or heterophilic interactions (binding to different types of molecules; right). E-cadherins on one cell (light blue, cell 1) commonly mediate cell-cell adhesion by forming homophilic cross-bridges with identical E-cadherins on adjacent cells (dark blue, cell 2). E-cadherins also can bind to neighboring E-cadherin molecules on the same cells (see Figures 20-3 and 20-14). Members of the immunoglobulin (Ig) superfamily of adhesion receptors form homophilic linkages (as shown here for NCAM) or heterophilic linkages (to other types of CAMs, not shown). Heterodimeric integrins (e.g., αv and β3 chains) can function as cell-matrix adhesion receptors that bind to very large, multi-adhesive extracellular proteins such as fibronectin, only a small part of which is shown here. Selectins, shown as dimers, contain a carbohydrate-binding lectin domain that recognizes specialized sugar structures on glycoproteins (as shown here) or glycolipids on adjacent cells (heterophilic interactions). Note that CAMs such as E-cadherins, often form higher order oligomers within the plane of the plasma membrane. Many adhesion molecules contain multiple, distinct domains, some of which are found in more than one kind of CAM. The cytoplasmic domains of adhesion receptors are often associated with adapter proteins that link them to the cytoskeleton or to signaling pathways. See R. O. Hynes, 1999, Trends Cell Biol. 15(12):M33–M37; R. O. Hynes, 2002, Cell 110:673–687; and J. Brasch et al., 2012, Trends Cell Biol. 22:299–310. Description The illustration on the left shows two types of homophilic interactions. The diagram has a blue bar at the top labeled cell 1 and a blue bar at the bottom labeled cell 2. These interactions are connected between these cells. One is labeled Cadherins (E-Cadherin) and is represented as a set of 5 light blue beads hanging down and attached to a set of 5 darker blue beads. The connections between the light blue beads are labeled as calcium-binding sites. Next to this at the right is an interaction labeled Ig-superfamily C A M's (N C A M). The chain of bead-like structures here are two orange ovals followed by 10 green circles with the middle 4 joining each other, then two orange ovals at the bottom. None are labeled. On the heterophilic side, the first diagram is labeled integrins (alpha V Beta 3) A blue chain of beads labeled alpha hangs straight down. To the right, the beta chain hangs down at an angle to join with the alpha chain and join to a light orange chain that is moving sideways and is labeled fibronectin. The bottom cell is not shown here. The second type is labeled selectins (P-selectin). Two pink chains hang down parallel to each other and join to black branched lines labeled sugars. The sugars go to cell 2, which has the label glycoprotein. Through their extracellular domains, CAMs mediate adhesive interactions between cells of the same type (homotypic adhesion) or between cells of different types (heterotypic adhesion). A CAM on one cell can directly bind to the same kind of CAM on an adjacent cell (homophilic binding) or to a different class of CAM (heterophilic binding) (see Figure 20-2). CAMs can be broadly distributed along the regions of plasma membranes that contact other cells or clustered in discrete patches or spots that are sometimes called cell junctions. Cell-cell adhesions can be tight and long lasting or relatively weak and transient. For example, the associations between neurons in the spinal cord or the metabolic cells in the liver exhibit tight adhesion. In contrast, immune-system cells in the blood often exhibit only brief, weak interactions that allow them to roll along and pass through a blood vessel wall on their way to fight an infection within a tissue.
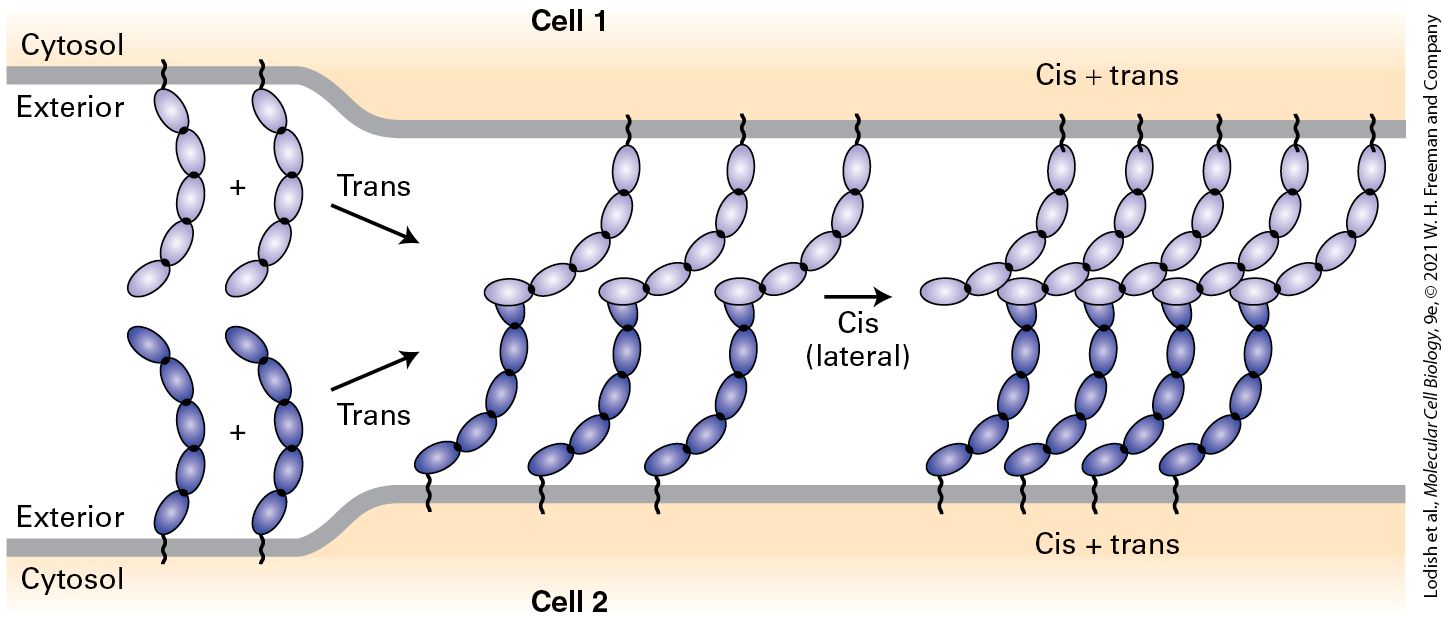
The cytosolic domains of adhesion receptors recruit sets of multifunctional adapter proteins (see Figure 20-1). These adapters act as linkers that directly or indirectly connect adhesion receptors to elements of the cytoskeleton (see Chapters 17 and 18); they can also recruit intracellular molecules that function in signaling pathways (see Chapters 15 and 16) to modify cellular behavior, including gene expression, and the activities of a variety of intracellular proteins including the adhesion receptors themselves. In many cases, a complex aggregate of adhesion receptors, adapter proteins, and other associated proteins is assembled at the inner surface of the plasma membrane. These complexes facilitate two-way, outside-in and inside-out, communication between cells and their surroundings. The formation of many cell-cell adhesions entails two types of molecular interactions, called trans and cis binding interactions (Figure 20-3). Trans interactions are also called intercellular or adhesive interactions, and cis interactions are also called lateral (in the same cell) interactions. In trans interactions, CAMs on one cell bind to the CAMs on an adjacent cell. In cis interactions, monomeric CAMs on one cell bind to one or more CAMs in the same cell’s plasma membrane. For some adhesion molecules the trans interactions between adjacent cells can increase the probability of stable cis interaction formation and thus determine the order in which these interactions occur (trans prior to cis is shown in Figure 20-3). It appears that trans and cis interactions are mutually reinforcing. Furthermore, the association of intracellular molecules with the cytosolic domains of CAMs can dramatically influence the intermolecular interactions of CAMs by promoting their clustering together (e.g., cis interactions) or by altering their conformation in a way that increases the affinity of trans interactions. Just like Velcro, CAMs can generate very tight adhesion when many weak interactions are combined, and this is especially the case when CAMs are concentrated in small, well-defined areas such as cell junctions.
FIGURE 20-3 Model for the generation of cell-cell adhesions. Trans interactions between CAMs on adjacent cells can hold the plasma membranes closely together and permit the subsequent formation of cis (lateral) interactions that collect the CAMs into clusters. The parts of the molecules that participate in these trans and cis interactions vary among the different CAMs. Multiple trans interactions concentrated in space by cis interactions generate a strong, Velcro-like adhesion between the cells. The models shown here are based on CAMs called classical cadherins (described in greater detail later in this chapter). See M. S. Steinberg and P. M. McNutt, 1999, Curr. Opin. Cell Biol. 11:554–560; and J. Brasch et al., 2012, Trends Cell Biol. 22:299–310. Description The illustration shows the surfaces of two neighboring cells. Cell 1, at the top of the diagram, has several chains of cadherins embedded in the surface. Cell 2 also contains these chains. The tips of the chains are interacting with the tips of chains embedded in the same
The Extracellular Matrix Participates in Adhesion, Signaling, and Other Functions
molecule laterally; this interaction is labeled cis. The chains of one cell, on encountering the cadherin chains of another cell, form trans interactions, the tips of the chains interacting with the cis chains on the other cell. Adhesive interactions between cells and the consequences of those interactions on cell structure and function vary considerably, depending on the tissue and the particular CAMs participating. For example, some CAMs require calcium ions to form effective adhesions while others do not. Among the many variables that determine the nature of adhesion between two cells are: the binding affinity of the interacting molecules (thermodynamic properties); the overall “on” and “off” rates of association and dissociation for each interacting molecule (kinetic properties); the spatial distribution or density of adhesion molecules (ensemble properties); the active versus inactive states of CAMs with respect to adhesion (biochemical properties); and external forces such as stretching and pulling, for example, in muscle or the laminar and turbulent flow of cells and surrounding fluids in the circulatory system (mechanical properties). The Extracellular Matrix Participates in Adhesion, Signaling, and Other Functions The ECM is a complex combination of proteins and polysaccharides that is secreted and assembled by cells into a network in which the components bind to one another. The ECM is often involved in holding cells and tissues together. The composition, physical properties, and functions of the ECM are carefully controlled and can vary depending on the tissue type, its location, its physiological state, and chemical modifications of its components, including changes that can occur as a consequence of pathology. These modifications include enzymatic phosphorylation, sulfation and desulfation, cross-linking, cleavage by proteases and glycosidases, and oxidation, as well as nonenzymatic addition of glucose (glycation). The ECM is usually sensed by cells as a consequence of binding to cell-matrix adhesion receptors on their plasma membranes, which then instruct the cells to behave appropriately in response to their environments or modulate the structure and function of the ECM based on the state of the cells. Different cells can bind to the same patch of ECM via their adhesion receptors and thus be indirectly bound together. ECM components include proteoglycans, a unique type of glycoprotein (a protein with covalently attached sugars); collagens and other proteins that often form fibers; soluble multi-adhesive matrix proteins; and others (Table 20-1). A common feature of many ECM proteins is that they comprise repeating structural domains (sometimes called repeats) that form very large proteins. Within one molecule, these repeats can be either very similar (homologous) to other repeats in the protein, distinct from other repeats, or both. The presence of nonidentical repeats that can exhibit distinct binding properties contributes to the multiligand and multifunctional characteristics of many adhesive proteins. For example, multi-adhesive matrix proteins, such as fibronectin and laminin, are long, flexible molecules that contain multiple repeats. They are responsible for binding various types of collagen, other matrix proteins, polysaccharides, and extracellular signaling molecules as well as cell-matrix adhesion receptors. These proteins are important organizers of the extracellular matrix. Through their interactions with adhesion receptors, they also regulate cell-matrix adhesion — and thus cell shape and behavior.
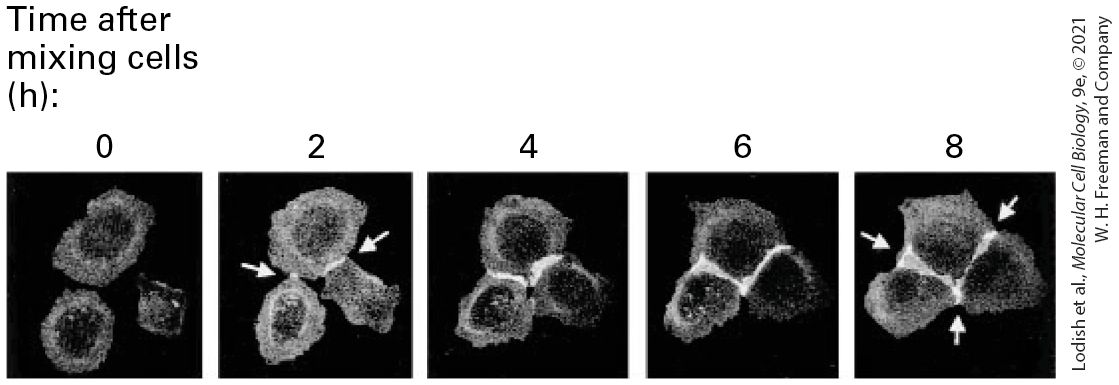
TABLE 20-1 • Extracellular Matrix Proteins Proteoglycans Collagens Description The illustration shows a purple stack of flattened ovals with three tails at the top, labeled Perlecan. Description The illustration shows two types of collagens. A blue line with an oval head on the right is labeled sheet forming; for example, type 4. An enlarged section shows a braid of 3 lines, with the lines stretched at the center. A blue rod-shaped structure is labeled fibrillar collagens; for example, types 1, 2, and 3. An enlarged section shows a braid of three lines evenly braided. Cells contribute to the assembly of the ECM not only by secreting its components, but also by participating directly in the assembly of those components into complex structures containing large fibrils and amorphous macromolecules. Once assembled, the ECM often is not static, but rather highly dynamic in that its chemical, physical, and biological properties can be altered quantitatively or qualitatively as a consequence of cells secreting enzymes, such as proteases, and other molecules into the extracellular space. These alterations in the ECM, which are usually referred to as remodeling, can involve covalent chemical modifications (including chemical cross-linking of ECM molecules), partial or essentially complete proteolytic cleavage of ECM
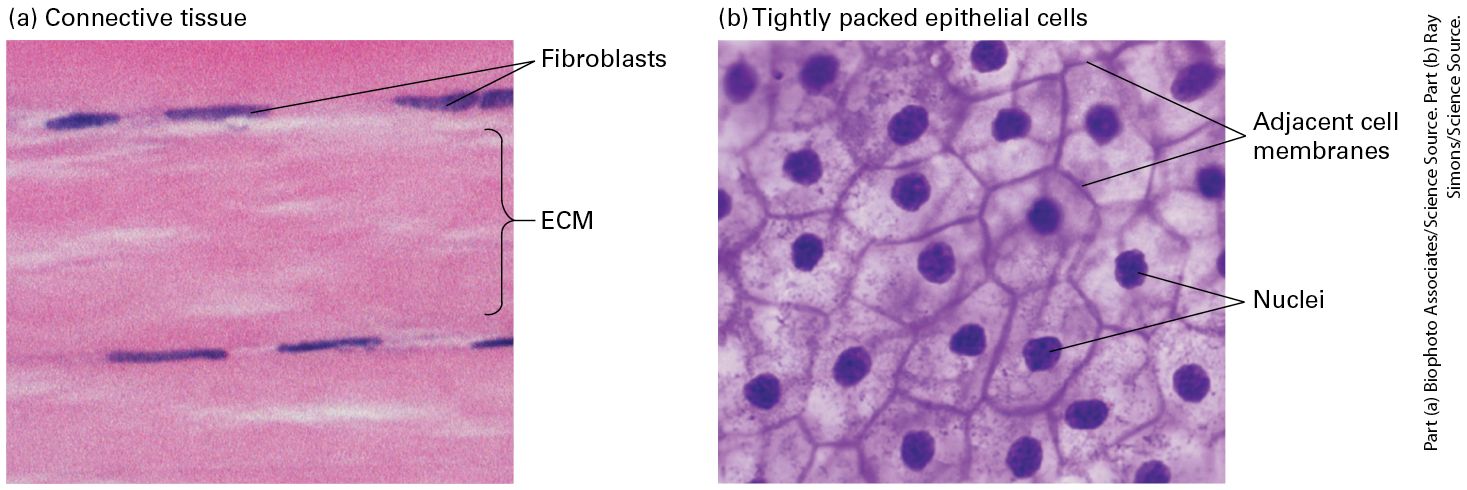
components, and addition of newly synthesized ECM molecules. An example we have already encountered, and will discuss in more detail in Section 20.4, is the role of ECM components in triggering activation of TGF-β by releasing it from its inhibitor (see Figure 16-22). The relative volumes occupied by cells and their surrounding matrix vary greatly among different animal tissues. Some connective tissue, for instance, is mostly matrix with relatively few cells (Figure 20-4a), whereas many other tissues, such as epithelia, are composed of very densely packed cells with relatively little matrix (Figure 20-4b). The density of packing of the molecules within the ECM itself can also vary greatly.
FIGURE 20-4 Variation in the relative density of cells and ECM in different tissues. (a) Dense connective tissue contains mostly extracellular matrix consisting of tightly packed ECM fibers (pink) interspersed with rows of relatively sparse fibroblasts, the cells that synthesized this ECM (purple). (b) Squamous epithelium viewed from the top, showing epithelial cells tightly packed into a quilt-like pattern with the plasma membranes of adjacent cells close to one another and little ECM between the cells (see also Figure 20-10b). Description The micrograph labeled (a) shows two rows of three narrow oval blue shapes labeled fibroblasts. The pink area between the fibroblasts is labeled E C M. The micrograph labeled (b) shows several irregular shaped cells with adjacent cell membranes. The blue dots in the center of the cells are labeled nuclei. H. V. Wilson’s classic studies of adhesion in marine sponge cells showed conclusively that one primary function of the ECM is to literally hold tissue together. Figures 20-5a and 20-5b, which re-create Wilson’s classic work, show that when sponges are mechanically dissociated and individual cells from two sponge species are mixed, the cells of one species will adhere to one another, but not to cells from the other species. This specificity is due in part to species-specific adhesive proteins in the ECM that bind to the cells via adhesion receptors. These adhesive proteins can be purified and used to coat colored beads, which, when mixed, aggregate with one another with a specificity similar to that of intact sponge cells (Figure 20-5c, d).
EXPERIMENTAL FIGURE 20-5 Mechanically separated marine sponges reassemble through species-specific homotypic cell adhesion. (a) Two intact sponges, Microciona prolifera (orange) and Halichondria panicea (yellow), growing in the wild. (b) After mechanical disruption and mixing of the individual cells from the two sponge species, their individual cells were allowed to reassociate for about 30 minutes with gentle stirring. The cells aggregated with species-specific homotypic adhesion, forming clumps of M. prolifera cells (orange) and H. panicea cells (yellow). In parts (c) and (d) red or green fluorescently labeled beads were coated with the proteoglycan aggregation factor (AF) from the ECM of either M. prolifera (MAF) or H. panicea (HAF). (c) When beads of both colors were coated with MAF, they all aggregated together, forming yellow aggregates (combination of red and green). (d) MAF (red) and HAF (green) coated beads do not readily form mixed aggregates, but rather assemble into distinct clumps held together by homotypic adhesion. [Parts (a) and (b) republished with permission from Springer, from X. Fernández-Busquets and M. M. Burger, 2003, “Circular Proteoglycans from Sponges: First Members of the Spongican Family,” Cell. Mol. Life Sci. 60(1):88–112; permission conveyed through Copyright Clearance Center, Inc. Parts (c) and (d) from J. Jarchow and M. M. Burger, 1998, “Species-Specific Association of the Cell-Aggregation Molecule Mediates Recognition in Marine Sponges,” Cell Adhes. Commun. 6(5):405–414; © Taylor and Francis, www.tandfonline.com.] Description The image labeled (a) shows sponges in their natural habitat. The micrograph labeled (b) shows agglomerated sponge cells after dispersion. The fluorescence micrographs labeled (c) and (d) show clumps of cells after the treatment with M A F - MA F and M A F - H A F, respectively. The micrograph labeled (c) shows more orange spots with a few green spots interspersed. The micrograph labeled (d) shows the orange and green spots clumped together away from each other. The ECM plays a multitude of other roles in addition to facilitating cell adhesion (Table 20-2). Different combinations of components tailor the ECM for specific purposes at different anatomic sites, for example, strength in a tendon, strength and rigidity in teeth and bones, cushioning in cartilage, and transparency in the vitreous humor in the eyeball. The composition of the ECM also provides positional and signaling information for cells, letting a cell know where it is and what it should do. ECM remodeling can modulate the interactions of a cell with its environment. Furthermore, the ECM serves as a reservoir for many extracellular signaling molecules that control cell growth and differentiation. In addition, it provides a lattice through or on which cells either can move or are prevented from moving, particularly in the early stages of tissue assembly. Morphogenesis — the stage of embryonic development in which tissues, organs, and body parts are formed by cell movements and rearrangements — is critically dependent on cell-matrix adhesion as well as cell-cell adhesion. For example, cell-matrix interactions are required for branching morphogenesis (formation of
branching structures) to form blood vessels, the air sacs in the lung, mammary and salivary glands, and other structures (Figure 20-6).
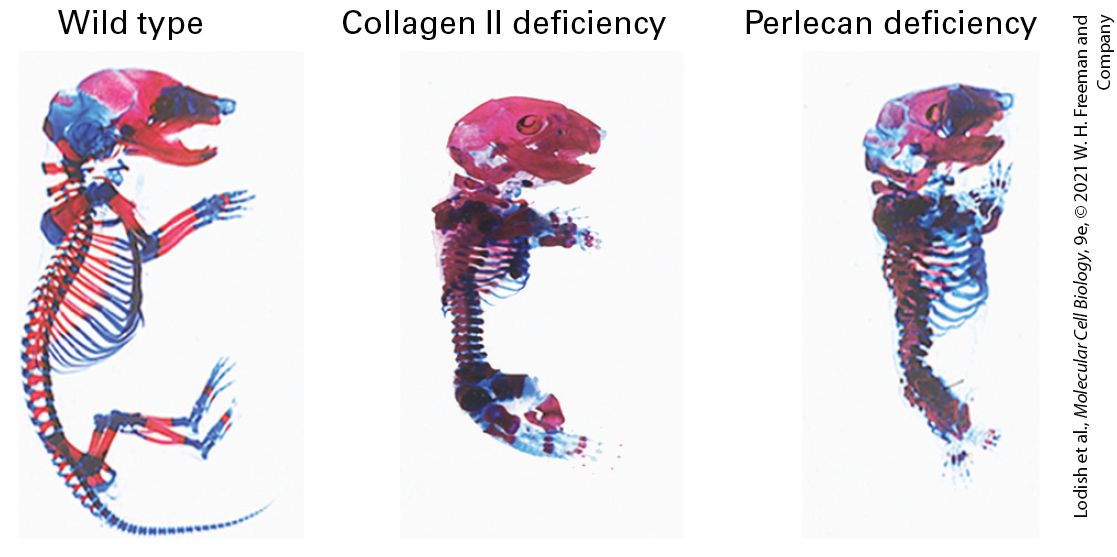
TABLE 20-2 • Functions of the Extracellular Matrix 1. Anchoring and surrounding cells to maintain solid-tissue three-dimensional architecture and define tissue boundaries 2. Determining the biomechanical properties (stiffness/elasticity, porosity, shape) of the extracellular environment 3. Controlling cellular polarity, survival, proliferation, differentiation, and fate (e.g., asymmetric division of stem cells; see Chapter 22), and thus embryonic and neonatal development and adult function and responses to the environment and to disease 4. Inhibiting or facilitating cell migration (e.g., serving as either a barrier to movement or, conversely, as a “track” along which cells — or portions of cells — can move) 5. Binding to and acting as a reservoir of growth factors; in some cases, the ECM (a) helps generate an extracellular concentration gradient of the growth factor, (b) serves as a co-receptor for the growth factor, or (c) aids in proper binding of the growth factor to its receptor (ECM component and growth factor jointly serve as a receptor’s combined ligand) 6. Activating cell surface signaling receptors EXPERIMENTAL FIGURE 20-6 Antibodies to fibronectin block branching morphogenesis in developing mouse tissues. Immature salivary glands were isolated from mouse embryos and allowed to undergo branching morphogenesis in vitro for 10 hours in the absence (a) or presence (b) of an antibody that binds to and blocks the activity of the ECM molecule fibronectin. Anti-fibronectin antibody (Anti-FN) treatment blocked branch formation (arrowheads). Inhibition of fibronectin’s adhesion receptor (an integrin) also blocks branch formation (not shown). Scale bar, 100 μm. [Republished with permission of Nature, from T. Sakai, M. Larsen, and K. M. Yamada, 2003, “Fibronectin Requirement in Branching Morphogenesis,” Nature 423(6942):876–881; permission conveyed through Copyright Clearance Center, Inc.] Description The micrograph labeled (a) shows a control mouse salivary gland. The gland features branched lobes with pits in-between. The pits are marked with arrows. The micrograph labeled (b) shows a mouse salivary gland treated with an antibody. The gland is smooth without branching. Disruptions in cell-matrix and cell-cell interactions can have devastating consequences for the development of tissues. Figure 20-7 shows the dramatic changes in the skeletal system of embryonic mice when the genes for either of two key ECM molecules, collagen II and perlecan, are inactivated. Disruptions in adhesion and ECM functions are also characteristic of various pathologies, including cardiovascular, musculoskeletal, kidney, skin,
eye, and bone diseases as well as metastatic cancer, in which cancer cells leave their normal locations and spread throughout the body. EXPERIMENTAL FIGURE 20-7 Inactivating the genes for some ECM proteins results in defective skeletal development in mice. These photographs show skeletons of normal (left), collagen II–deficient (center), and perlecan-deficient (right) murine embryos that were isolated and stained to visualize the cartilage (blue) and bone (red). Absence of these key ECM components leads to forms of short stature (e.g., dwarfism), with many skeletal elements shortened and disfigured. [Republished with permission from John Wiley & Sons, Inc., from E. Gustafsson et al., 2003, “Role of Collagen Type II and Perlecan in Skeletal Development,” Ann. NY Acad. Sci. 995:140–150; permission conveyed through Copyright Clearance Center, Inc.] Description The illustration on the left shows a wild type mouse skeleton stained to reveal areas of bone and cartilage. The illustration in the center shows a mouse skeleton with collagen 2 deficiency. The skeleton is stained to reveal areas of bone and cartilage. The skeleton is smaller than the wild type mouse skeleton and has shortened forelegs and almost no back legs. The illustration on the right shows a mouse skeleton with a perlecan deficiency. The skeleton is stained to reveal areas of bone and cartilage. The skeleton is smaller than the wild type mouse skeleton and has less bone, more cartilage, and small stubs. Although many CAMs and adhesion receptors were initially identified and characterized because of their adhesive properties, they also play major roles in signaling, using many of the pathways discussed in Chapters 15 and 16. Figure 20-8 illustrates how integrin adhesion receptors physically and functionally interact, via adapters and signaling molecules, with a broad array of intracellular signaling pathways to influence cell survival, gene transcription, cytoskeletal organization, cell motility, and cell proliferation. Conversely, changes in the activities of signaling pathways inside cells can influence the structures of CAMs and adhesion receptors — for example, by altering adapter binding to the cytosolic portions of the CAMs — and so modulate their ability to interact with other cells and with the ECM. Thus outside-in and inside-out signaling involve numerous interconnected pathways.
The Evolution of Multifaceted Adhesion Molecules Enabled the Evolution of Diverse Animal Tissues
FIGURE 20-8 Integrin adhesion receptor–mediated signaling pathways control diverse cell functions. Binding of integrins to their ligands induces conformational changes in their cytoplasmic domains, directly or indirectly altering their interactions with cytoplasmic proteins (outside-in signaling). These cytoplasmic proteins include adapter proteins (e.g., talins, kindlins, paxillin, vinculin) and signaling kinases [Src-family kinases, focal adhesion kinase (FAK), integrin-linked kinase (ILK)] that transmit signals via diverse signaling pathways, thereby influencing cell proliferation, cell survival, cytoskeletal organization, cell migration, and gene transcription. Components of several signaling pathways, some of which are associated directly with the plasma membrane, are shown in green boxes. Many of the components of the pathways shown here are shared with other cell-surface-activated signaling pathways (e.g., receptor tyrosine kinases shown on the right) and are discussed in Chapters 15 and 16. In turn, intracellular signaling pathways can, via adapter proteins, modify the ability of integrins to bind to their extracellular ligands (inside-out signaling). See W. Guo and F. G. Giancotti, 2004, Nat. Rev. Mol. Cell Biol. 5:816–826; and R. O. Hynes, 2002, Cell 110:673–687. Description The illustration shows a plasma membrane as a gray bar with exterior labeled above it and cytosol below it. Above the membrane is the label E C M with a blue and pink chain of beads labeled integrin (adhesion receptor). The two chains are joined at the top with a ligand. The integrin moves through the membrane to a rectangle with the label: various adaptors and signaling kinases. Using downward arrows, a series of these adaptors are represented as green rectangles with labels inside. The first column of rectangles, at left, is labeled, top to bottom: P I 3 K, P I (3, 4, 5) P subscript 3, and A k t slash P K B. The center set of green rectangles is labeled: R a c slash R h o slash C d c 42 (small G T Pases), N F-k B, J U N. The right-hand set is labeled: G R B 2, S o s, R a s, R a f, M E K, E R K slash M A P K. Below all of these is a box labeled cellular responses to adhesion receptor signaling. The following responses are listed in the box: Cell proliferation (cycle), cell survival, cytoskeletal organization, cell migration, gene transcription. At the right end of the plasma membrane is a receptor Tyrosine Kinase. It is a blue-colored four connected circles above the membrane with two triangle structures labeled bound ligands and blue irregular structures below the membrane.
Cell-Adhesion Molecules Mediate Mechanotransduction
The Evolution of Multifaceted Adhesion Molecules Enabled the Evolution of Diverse Animal Tissues Cell-cell and cell-matrix adhesions are responsible for the formation, composition, architecture, and function of animal tissues. Not surprisingly, some adhesion molecules are evolutionarily ancient and are among the most highly conserved proteins in multicellular organisms. Some sponges, the most primitive multicellular organisms, express certain CAMs and multi-adhesive ECM molecules whose structures are strikingly similar to those of the corresponding human proteins. The evolution of metazoans has depended on the evolution of diverse adhesion molecules with novel properties and functions whose levels of expression differ in different types of cells. Some CAMs and adhesion receptors (e.g., cadherins, integrins, and Ig-superfamily CAMs such as L1CAM) and some ECM components (e.g., type IV collagen, laminin, nidogen/entactin, and perlecan-like proteoglycans) are highly conserved because they play crucial roles in many different organisms, whereas other adhesion molecules are less well conserved. Fruit flies, for example, do not have certain types of collagen or the ECM protein fibronectin, which play important roles in mammals. The diversity of adhesion molecules arises in large part from two phenomena that can generate the numerous closely related proteins, called isoforms, that constitute a protein family. In some cases, the different members of a protein family are encoded by multiple genes that arose from a common ancestor by gene duplication and divergent evolution (see the human β-like globin gene cluster in Chapter 7). In other cases, a single gene produces an RNA transcript that can undergo alternative splicing to yield multiple mRNAs, each encoding a distinct protein isoform (see Figure 7-3 and Section 9.2). Both phenomena contribute to the diversity of some protein families, such as the cadherins. Particular isoforms of an adhesive protein are often expressed in some cell types and tissues, but not others. Cell-Adhesion Molecules Mediate Mechanotransduction Mechanotransduction is the reciprocal interconversion of a mechanical force — or stimulus — and biochemical processes. These interconversions underlie a variety of biological activities, such as signaling, regulated gene expression, cell proliferation, cell migration, and interactions among cells and between cells and the ECM. Evolution has produced a wide variety of mechanosensors that respond to a mechanical stimulus by changing their shape and activity. Examples include at least one G protein–coupled receptor that appears to respond directly to extracellular shear stress — a force parallel to the surface of the cell, such as that resulting from the flow of blood in a blood vessel — and a variety of ion channel mechanosensors whose opening is sensitive to mechanical stretch of the plasma membrane. These include certain transient receptor potential (TRP) channels as well as Piezo nonselective cation channels (see Chapter 23), which are found in neurons and nonneuronal cells and that respond to a variety of mechanical stimuli, including touch, airway stretch, blood pressure, and shear and osmotic forces.
Mechanotransduction in the context of cell-cell and cell-matrix interactions usually involves a cell-surface adhesion receptor that transmits mechanical force or biochemical information across the plasma membrane and one or more intracellular or extracellular mechanosensors that respond to the mechanical stimulus by changing shape and activity. For example, tension applied across the length of a multidomain mechanosensor protein, such as the ECM protein fibronectin or the integrin adapter protein talin, can separate otherwise tightly packed domains, or even literally pull apart (disassemble) individual domains. As a consequence, one or more binding sites that were otherwise inaccessible (cryptic) are exposed. Figure 20-9 shows examples of domains that are unfolded by mechanical force, exposing cryptic binding sites. The newly accessible binding sites can then recruit binding partners — in some cases after phosphorylation — and alter cellular or extracellular functions. For example, the stretching of fibronectins by integrins induces domain separation and consequently their assembly into fibrils, which is an early step in the assembly of collagen and other molecules into ECM. The mechanical forces in mechanotransduction can be forces generated within a cell, such as myosin-driven movement of actin filaments (Chapter 17), or outside a cell, such as blood flow, movement of adjacent cells, or contraction or expansion of ECM. As we will see later in this chapter, cell-cell and cell-matrix adhesion sites play particularly important roles in mechanotransduction.
FIGURE 20-9 Models of domains in mechanosensor proteins responding to mechanical forces. (a) Hypothetical model of the partial unfolding of a fibronectin type III domain in the ECM molecule fibronectin when that protein is subjected to mechanical force. Mechanical force generated within the cell by actin movement and mechanotransduced via multiple integrin adhesion receptors bound to the extracellular dimeric fibronectin can partially unfold the fibronectin. The unfolding is thought to expose a putative, previously hidden binding site on fibronectin (blue segment) that has the potential to form β sheets with other fibronectin molecules, recruiting them to form fibronectin fibrils, and helping assemble the ECM. (b) Hypothetical model of the partial unfolding of a domain (the R1 five-helix bundle) in the intracellular integrin adapter protein talin when it is subjected to mechanical stretching force. This force is generated by actin, which can bind to and pull on the C-terminus of talin while talin’s N-terminus is bound to the cytoplasmic tail of integrin’s β subunit. The unfolding is thought to expose this domain’s otherwise cryptic α-helical vinculin binding site (blue). Vinculin, an actin-binding protein, can then bind to the integrin-talin complex via the exposed site and in turn bind to actin, thus promoting the assembly of multiple actin fibers. The assembly of actin fibers indirectly linked by adapters to integrins strengthens integrin-mediated adhesion and helps to build focal adhesions (see Figures 20-14e and 20-40). [Part (a) Data from E. P. Gee et al., 2013, J. Biol. Chem. 288:21329–21340; and M. A. Schumacher et al., 2013, J. Biol. Chem. 288:33738– 33744. Part (b) Data from M. Yao et al., 2014, Sci. Rep. 4:4610; and E. Papagrigoriou et al., 2004, EMBO J. 23:2942–2951.] Description The illustration labeled (a) titled Fibronectin type 3 domain shows gray ribbons with one green, one red, and one blue area highlighted. The blue area is labeled inaccessible binding site. A sideward arrow from the ends of the domain is labeled mechanical force (stretching) and goes to a diagram where the red and blue parts are stretched into a single ribbon off to the right and the green highlight is still in the same place. The stretched blue ribbon is labeled accessible binding site. The illustration labeled (b) titled Talin five-helix bundle domain
shows gray ribbons with one blue area highlighted. The blue area is labeled inaccessible binding site. Then the sideward arrow with the stretching shows a blue ribbon off to the right of the bundle and labeled accessible binding site. KEY CONCEPTS OF SECTION 20.1 Cell-Cell and Cell–Extracellular Matrix Adhesion: An Overview Cell-cell and cell–extracellular matrix (ECM) interactions are critical for assembling cells into tissues, controlling cell shape and function, and determining the developmental fate of cells and tissues. Diseases may result from abnormalities in the structures or expression of adhesion molecules. Cell-cell adhesion molecules (CAMs, also called cell-cell adhesion receptors) mediate direct intercellular cell-cell adhesions (homotypic and heterotypic), and cell-matrix receptors mediate cell attachment to the ECM (see Figure 20-1). These interactions bind cells into tissues and facilitate communication between cells and their environments. The cytosolic domains of cell adhesion receptors bind adapter proteins that mediate interaction with cytoskeletal fibers and intracellular signaling proteins. The major families of CAMs are the cadherins, lectins, Ig-superfamily CAMs, and integrins (see Figure 20-2). Members of the integrin and IgCAM superfamilies can also function as cell-matrix adhesion receptors. Tight cell-cell adhesions entail both trans (adhesive or intercellular) interactions of like (homophilic) or different (heterophilic) CAMs and cis (lateral) oligomerization of CAMs (see Figure 20-3). The combination of trans and cis interactions produces a Velcro-like adhesion between cells. The ECM is a dynamic, complex meshwork of proteins and polysaccharides that contributes to the structure and function of a tissue (see Table 20-2). The major classes of ECM molecules are proteoglycans, collagens, and multi-adhesive matrix proteins, such as fibronectin and laminin. CAMs and adhesion receptors, together with their cytoplasmic adapter proteins, play major roles in “outside-in” and “inside-out” signaling, facilitating critically important communication between cells and their surroundings. The evolution of adhesion molecules with specialized structures and functions permits cells to assemble into diverse classes of tissues with varying functions. Mechanotransduction, the interconversion of a mechanical stimulus or force and biochemical processes, is mediated by adhesion receptors and mechanosensors. Adhesion receptors themselves can also be mechanosensors. Mechanotransduction permits cells to respond to mechanical forces from their environments and to exert mechanical forces on their surroundings.
Epithelial Cells Have Distinct Apical, Lateral, and Basal Surfaces
20.2 Cell-Cell and Cell–Extracellular Matrix Junctions and Their Adhesion Molecules Cells in epithelial and in nonepithelial tissues use many, but not all, of the same cell-cell and cell-matrix adhesion molecules. We begin our detailed discussion of adhesion with epithelia because of their relatively simple organization, as well as their fundamental role in evolution and development. In this section, we focus on regions of the cell surface that contain clusters of adhesion molecules in discrete patches or spots, called anchoring junctions, tight junctions, and gap junctions. Anchoring and tight junctions play critical roles in mediating cell-cell and cell-matrix adhesion, and all three types of junctions mediate intercellular or cell-matrix communication. Epithelial Cells Have Distinct Apical, Lateral, and Basal Surfaces Cells that form epithelial tissues are said to be polarized because their plasma membranes are organized into discrete regions. Typically, the distinct surfaces of a polarized epithelial cell are called the apical (top), lateral (side), and basal (base or bottom) surfaces (Figure 20-10; see also Figure 20-1). The apical, lateral, and basal surfaces of epithelial cells can exhibit distinctive characteristics. The area of the apical surface is often greatly expanded by the formation of microvilli — small, fingerlike projections (see Chapter 22). A complex set of evolutionarily conserved molecules, called polarity regulators, is required to generate the functional and structural asymmetry of epithelial and other cells (see Chapter 22). Adhesion molecules play essential roles in generating and maintaining these distinct surfaces.
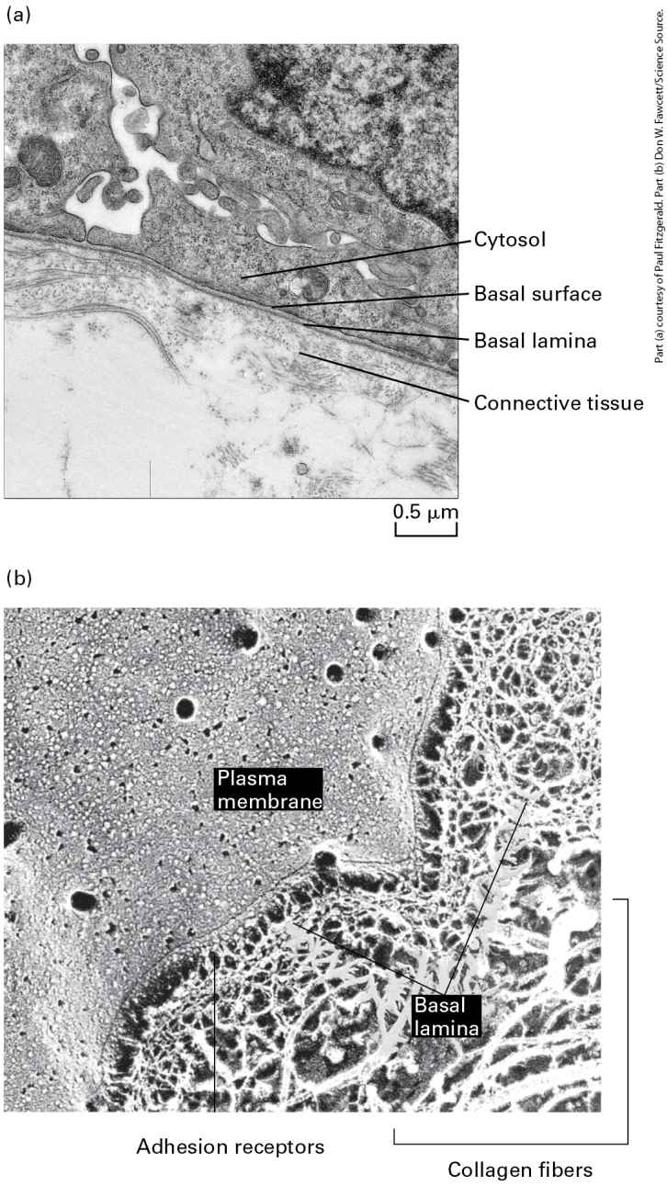
FIGURE 20-10 Principal types of epithelia. (a) Simple columnar epithelia consist of elongated cells, including mucus-secreting cells (in the lining of the stomach and cervical tract) and absorptive cells (in the lining of the small intestine). The thin protrusions at the apical surface are microvilli (see Figure 20-11). (b) Simple squamous epithelia, composed of thin cells, line the blood vessels (endothelial cells/endothelium) and many body cavities. (c) Transitional epithelia, composed of several layers of cells with different shapes, line certain cavities subject to expansion and contraction (e.g., the urinary bladder). (d) Stratified squamous (nonkeratinized) epithelia line surfaces such as the mouth and vagina; these linings resist abrasion and generally do not participate in the absorption or secretion of
Three Types of Junctions Mediate Many Cell-Cell and Cell-Matrix Interactions
materials into or out of the cavity. The basal lamina, a thin fibrous network of collagen and other ECM components, supports all epithelia and connects them to the underlying connective tissue. Description The illustration labeled (a) titled simple columnar shows tall thick rounded rectangular cells grouped above connective tissue, which is represented as a brown area of cells at the bottom. The top is labeled apical surface; the side of the rectangular cells has the label: lateral surface. At the bottom of the rectangles, the label reads basal surface, and the top of the connective tissue is labeled basal lamina. The illustration labeled (b) titled simple squamous shows the same connective tissue at the bottom, but the rectangles now look like a flat layer on top. The illustration labeled (c) titled transitional shows four layers of cells, and they are different sizes, thick on top, small ones in the middle, and taller ones at the bottom above the connective tissue. The illustration labeled (d) titled stratified squamous (non-keratinized) shows the cells flattened in the top layers. Epithelia in different body locations have characteristic morphologies and functions (see
Figure 20-10; see also Figure 1-4). Stratified (multilayered) epithelia commonly serve as barriers and protective surfaces (e.g., the skin), whereas simple (single-layered) epithelia often selectively move ions and small molecules from one side of the epithelium to the other. For instance, the simple columnar epithelium lining the stomach secretes hydrochloric acid into the lumen; a similar epithelium lining the small intestine transports products of digestion from the lumen of the intestine across the cells into the blood (see Figure 11-30). In simple columnar epithelia, adhesive interactions between the lateral surfaces hold the cells together in a two-dimensional sheet, whereas those at the basal surface connect the cells to a specialized underlying extracellular matrix called the basal lamina. Often the basal and lateral surfaces are similar in composition and are collectively called the basolateral surface. The basolateral surfaces of most simple epithelia are usually on the side of the cell closest to the blood vessels, whereas the apical surface is not in stable, direct contact with other cells or the ECM. In animals with closed circulatory systems, blood flows through vessels whose inner lining is composed of flattened epithelial cells called endothelial cells. In general, epithelial cells are sessile, in that adhesion molecules firmly and stably attach them to one another and their associated ECM. One especially important mechanism that generates strong, stable adhesions is the concentration of subsets of these molecules into clusters called cell junctions. Three Types of Junctions Mediate Many CellCell and Cell-Matrix Interactions
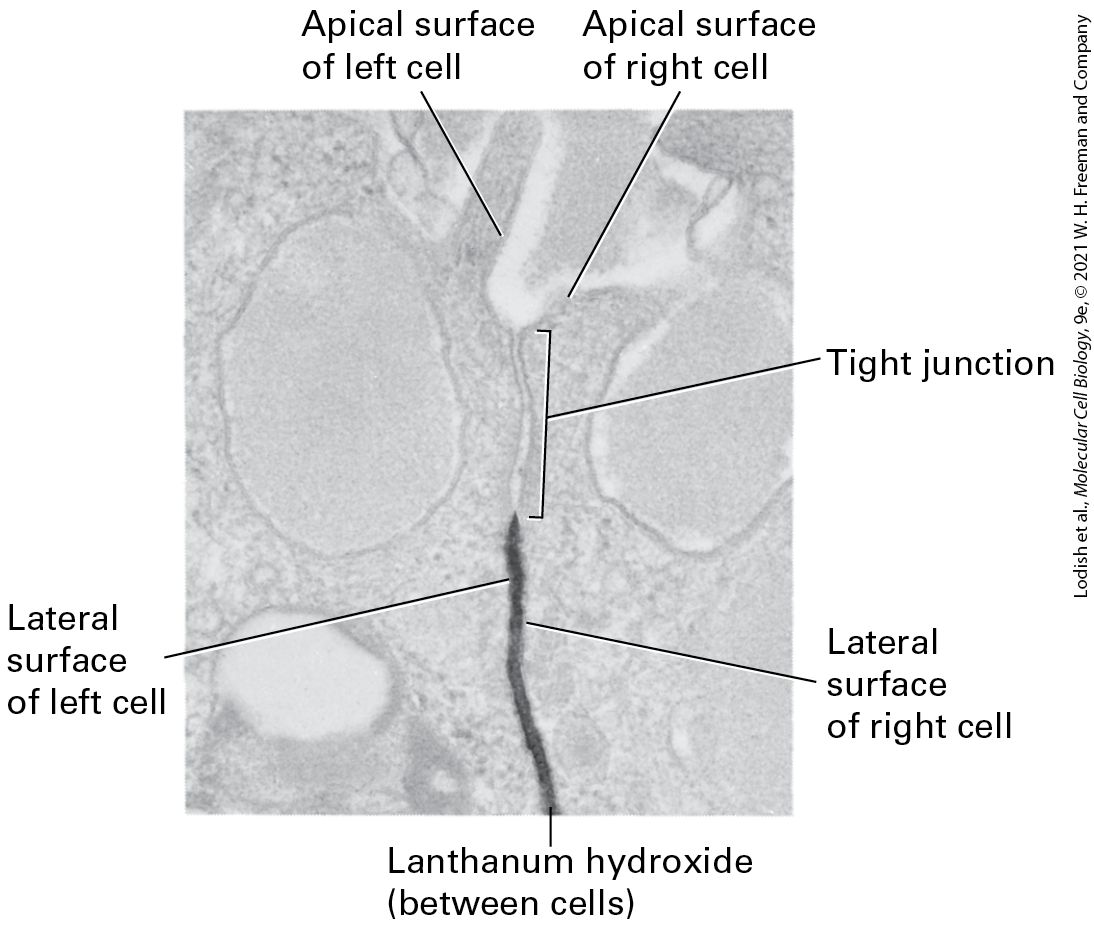
All epithelial cells in a sheet are connected to one another and to the ECM by specialized junctions. Although hundreds of individual dispersed adhesion molecule–mediated interactions are sufficient to cause cells to adhere, the clustered groups of adhesion molecules at cell junctions play special roles in imparting strength and rigidity to a tissue, transmitting information between the extracellular and the intracellular space, controlling the passage of ions and molecules across cell layers, and serving as conduits for the movement of ions and molecules from the cytoplasm of one cell to that of its immediate neighbor. Particularly important to epithelia is the formation of junctions that help form tight seals between the cells, allowing the epithelial sheet to serve as a barrier to the flow of molecules from one side of the sheet to the other. Three major classes of animal-cell junctions are prominent features of simple columnar epithelia (Figure 20-11 and Table 20-3): anchoring junctions, tight junctions, and gap junctions. Anchoring junctions and tight junctions perform the key task of holding the tissue together. As we shall see, tight junctions also control the flow of solutes through the extracellular spaces between the cells forming an epithelial sheet. Tight junctions are found primarily in epithelial cells, whereas anchoring junctions can be seen in both epithelial and nonepithelial cells. Anchoring junctions and tight junctions in epithelia are organized into three parts: (1) adhesive proteins (CAMs, adhesion receptors) in the plasma membrane that connect one cell to another cell on the lateral surfaces or to the ECM on the basal surfaces; (2) adapter proteins, which connect the CAMs or adhesion receptors to cytoskeletal filaments and signaling molecules; and (3) the cytoskeletal filaments themselves.
FIGURE 20-11 Principal types of cell junctions connecting the columnar epithelial cells lining the small intestine. (a) Schematic cutaway drawing of intestinal epithelial cells. The basal surface of the cells rests on a basal lamina, and the apical surface is packed with microvilli that project into the intestinal lumen. Tight junctions, lying just under the microvilli, prevent the diffusion of many substances between the intestinal lumen and internal body fluids (such as the blood) via the extracellular space between cells. Gap junctions allow the movement of small molecules and ions between the cytosols of adjacent cells. The remaining three types of junctions — adherens junctions, desmosomes, and hemidesmosomes — are critical to cell-cell and cell-matrix adhesion and signaling. (b) Electron micrograph of a thin section of epithelial cells in the rat intestine, showing the relative locations of the different junctions. [Part (b) M. G. Farquhar and G. E. Palade, 1963, J. Cell Biol. 17(2):375–412, Fig. 1; https://doi.org/10.1083/jcb.17.2.375.] Description The illustration labeled (a) shows a cube-shaped section of epithelial cells with microvilli as tan tubes on the top. A cutaway shows how various junctions are repeated and connected. The different types of junctions are labeled. At the top, below the microvilli is a tight junction with three layers of skinny ovals. Below this is a long red tube labeled adherens junction, with brown dots inside labeled actin and myosin filaments. Further down is an orange oval-shaped labeled gap junction. Next is a blue circle labeled desmosome. Connected to this, inside the cube are blue lines labeled intermediate filaments. At the bottom, but above the connective tissue layer is a pink oval labeled hemidesmosome. The micrograph labeled (b) shows the same area as that of the illustration. The tight junction, gap junction, adherens junction, and desmosome are labeled.
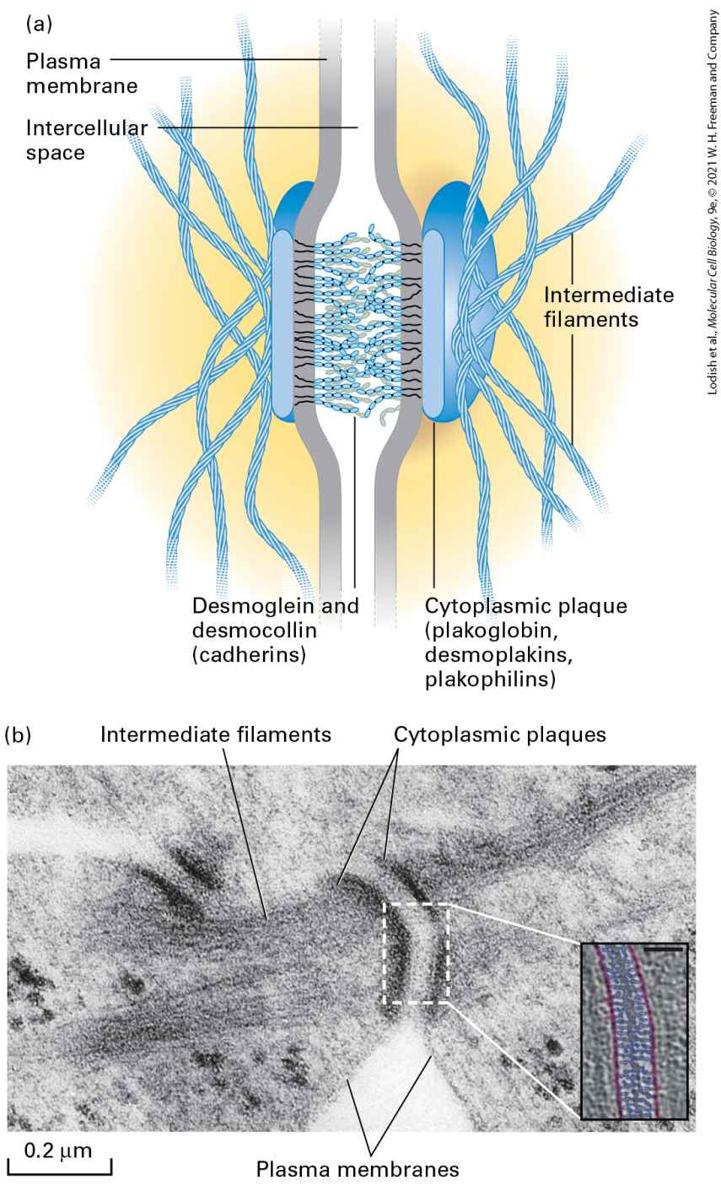
TABLE 20-3 • Cell Junctions Junction Adhesion Type Principal CAMs or Adhesion Receptors Cytoskeletal Attachment Intracellular Adapters Function
Anchoring junctions 1. Adherens junctions Cell-cell Cadherins Actin filaments Catenins, vinculin Shape, tension, signaling, force transmission 2. Desmosomes Cell-cell Desmosomal cadherins Intermediate filaments Plakoglobin, plakophilins, desmoplakins Strength, durability, signaling 3. Hemidesmosomes Cellmatrix Integrin (α6β4) Intermediate filaments Plectin, dystonin/BPAG1 Shape, rigidity, signaling 4. Focal contacts, fibrillar, and 3-D adhesions Cellmatrix Integrins Actin filaments Talin, kindlin, paxillin, vinculin Shape, signaling, force transmission, cell movement Tight junctions Cell-cell Occludin, claudins, JAMs Actin filaments
cingulin Controlling solute flow, cell polarity, signaling Gap junctions Cell-cell Connexins, innexins, pannexins Via adapters to other junctions
Communication, small-molecule transport between cells Plasmodesmata (plants only) Cell-cell Undefined Actin filaments
Communication, molecule transport between cells Gap junctions permit the rapid diffusion of small, water-soluble molecules between the cytoplasms of adjacent cells. Along with anchoring and tight junctions, gap junctions help a cell communicate with its environment. However, they are structurally very different from anchoring junctions and tight junctions and do not play a key role in strengthening cell-cell and cell-matrix adhesions. Found in both epithelial and nonepithelial cells, gap junctions resemble the distinct cell junctions in plants called plasmodesmata, which we discuss in Section 20.6.
Cadherins Mediate Cell-Cell Adhesions in Adherens Junctions and Desmosomes
Four types of anchoring junctions are present in cells. Two participate in cell-cell adhesion and two participate in cell-matrix adhesion. Adherens junctions connect the lateral membranes of adjacent epithelial cells and are usually located near the apical surface, just below the tight junctions (see Figure 20-11). A circumferential belt of actin and myosin filaments in a complex with the adherens junctions functions as a tension cable that can internally brace the cell and thereby control its shape. Epithelial and some other types of cells, such as smooth muscle and heart cells, are also bound tightly together by desmosomes, snap-like points of contact sometimes called spot desmosomes. Hemidesmosomes, found mainly on the basal surface of epithelial cells, and focal contacts, also called focal adhesions, anchor an epithelium to components of the underlying ECM, much like nails holding down a carpet. Adherens junctions, desmosomes, and focal adhesions are found in many different types of cells; hemidesmosomes appear to be restricted to epithelial cells. Bundles of intermediate filaments running parallel to the cell surface or through the cell connect desmosomes and hemidesmosomes, imparting shape and rigidity to the cell, as do actin filaments that connect the cytoskeleton with focal contacts and adherens junctions. The close interaction between these junctions and the cytoskeleton helps transmit shear forces from one region of a cell layer to the epithelium as a whole, providing strength and rigidity to the entire epithelial cell layer. Desmosomes and hemidesmosomes are especially important in maintaining the integrity of skin epithelia. As a consequence, mutations that interfere with hemidesmosomal anchoring in the skin can lead to a condition in which the epithelium becomes detached from its underlying matrix and extracellular fluid accumulates at the basolateral surface, forcing the skin to balloon outward, forming a blister. Cadherins Mediate Cell-Cell Adhesions in Adherens Junctions and Desmosomes The primary CAMs in adherens junctions and desmosomes belong to the cadherin family of glycoproteins that can contain oligosaccharides that are either N-linked (see Chapters 13 and 14) or O-linked (described in Figure 20-31 in Section 20.4). In vertebrates, this protein superfamily of more than a hundred members can be grouped into at least six subfamilies, including classical cadherins, clustered protocadherins, and desmosomal cadherins, which we will describe below. The diversity of cadherins arises from the presence of multiple
cadherin genes and alternative RNA splicing. It is not surprising that there are many different types of cadherins in vertebrates; many different types of cells in the widely diverse tissues of these animals use cadherins to mediate adhesion and communication, the detailed requirements for which may differ for different types of cells and tissues. Members of the cadherin superfamily can also control cell morphology, such as the assembly and tight packing of microvilli on the apical surfaces of some epithelial cells (see Figures 20-10a and 20-11a). The brain expresses the largest number of different cadherins, including protocadherins, presumably owing to the necessity of forming many specific cell-cell contacts to help establish its complex wiring pattern. Invertebrates, however, are able to function with fewer than 20 cadherins. Classical Cadherins The classical cadherins include E-, N-, and P-cadherins, named for the type of tissue in which they were initially identified (epithelial, neural, and placental, respectively). E- and N-cadherins are the most widely expressed, particularly during early differentiation. Sheets of polarized epithelial cells, such as those that line the small intestine or kidney tubules, contain abundant E-cadherin along their lateral surfaces. Although E-cadherin is concentrated in adherens junctions, it is present throughout the lateral surfaces, where it is thought to link adjacent cell membranes. The results of experiments with L cells, a line of cultured mouse fibroblasts, demonstrated that E-cadherins preferentially mediate homophilic interactions. L cells express no cadherins and adhere poorly to each other and to other types of cells. When the E-cadherin gene was introduced into L cells, the cells were found to adhere preferentially to other cells expressing E-cadherin (Figure 20-12). These engineered cadherin-expressing L cells formed epitheliumlike aggregates with one another and with epithelial cells isolated from lungs. Although most E-cadherins exhibit primarily homophilic binding, some mediate heterophilic interactions.
EXPERIMENTAL FIGURE 20-12 E-cadherin mediates -dependent adhesion of L cells. Under standard cell-culture conditions, in the presence of calcium in the extracellular fluid, L cells do not aggregate into sheets (left). Introduction of a gene that causes the expression of E-cadherin in these cells results in their aggregation into epithelium-like clumps in the presence of calcium (center), but not in its absence (right). Bar, 60 μm. [© C. L. Adams et al., 1998, J. Cell Biol. 142(4):1105–1119, Fig. 1E; https://doi.org/10.1083/jcb.142.4.1105.] The adhesiveness of cadherins depends on the presence of extracellular ; it is this property (calcium adhering) that gave rise to their name. For example, the adhesion of L cells expressing E-cadherin is prevented when the cells are bathed in a solution that is low in (see Figure 20-12). Some adhesion molecules require some minimal amount of in the extracellular fluid to function properly, whereas others, such as IgCAMs, are independent. The role of E-cadherin in adhesion can also be demonstrated by experiments with cultured epithelial cells called Madin-Darby canine kidney (MDCK) cells (see Figure 4-3). A green fluorescent protein–labeled form of E-cadherin has been used in these cells to show that clusters of E-cadherin mediate the initial attachment of the cells and the subsequent zippering of the cells into sheets (Figure 20-13). In this experimental system, the addition of an antibody that binds to E-cadherin, preventing its homophilic interactions, blocks the - dependent attachment of MDCK cells to one another and the subsequent formation of intercellular adherens junctions.
EXPERIMENTAL FIGURE 20-13 E-cadherin mediates adhesive connections in cultured MDCK epithelial cells. An E-cadherin gene fused to green fluorescent protein (GFP) was introduced into cultured MDCK cells. The cells were then mixed together in a calcium-containing medium, and the distribution of fluorescent E-cadherin was visualized over time (shown in hours). Clusters of E-cadherin mediate the initial attachment and subsequent zippering up of the epithelial cells and the formation of junctions (bicellular junctions are where two cells join and appear as lines; tricellular junctions are the sites of intersection of three cells). [© C. L. Adams et al., 1998, J. Cell Biol. 142(4):1105–1119, Fig. 2B; https://doi.org/10.1083/jcb.142.4.1105.] Description The label at the top of the photos reads, Time after mixing cells in hours (0, 2, 4, 6, 8 hours). The photo at 0 hours shows three dim and separate cells. In 2 hours, the cells are brighter, a connection is forming, and arrows point to the developing connection. In 4 hours, the connections are bigger. In 6 hours, the connections are bigger and longer. In 8 hours, the connections have extended three ways between the three cells. Each classical cadherin molecule contains a single transmembrane domain, a relatively short C-terminal cytosolic domain, and five extracellular cadherin domains (called EC1–EC5) (see
Figure 20-2). The extracellular domains are necessary for binding and cadherin-mediated cell-cell adhesion. Classical cadherin–mediated adhesion entails both cis lateral (on the same cell) clustering and trans adhesive (intercellular) molecular interactions (see Figures 20-3 and 20-14a–c). The binding of three ions at each of the sites located between the cadherin repeats (see Figures 20-2 and 20-14a) stabilizes the elongated and curved structure of the extracellular domain. As we shall see shortly, the curved structure of cadherin’s extracellular domain is necessary for the proper molecular complementarity that stabilizes cis and trans binding between cadherin molecules. The cis and trans interactions of cadherins, together with their interactions with cytoplasmic adapter and cytoskeletal molecules, permit the zippering up of cadherins into adhesive arrays. Binding of the EC1 domain of one cadherin molecule to the EC1 domain of another on the adjacent cell is responsible for trans binding (Figure 20-14a; see also Figure 20-3). Although the dissociation constant for EC1–EC1 homophilic binding
measured using isolated domains in solution is on the order of (relatively weak, or low-affinity, binding), the multiple low-affinity interactions in arrays of intact cadherin molecules on adjacent cells sum to produce a very tight intercellular adhesion.
FIGURE 20-14 Trans (intercellular) and cis (on the same cell) interactions of classical cadherins in typical adherens junctions and of protocadherins. (a) The exoplasmic cadherin domains [EC1-EC5, see ovals in (b)] of Ecadherins at adherens junctions on adjacent cells are clustered by homophilic cis and trans interactions (see Figure 203). The -dependent elongated and curved structure of cadherin’s extracellular domains is necessary for stable cis and trans interactions (examples highlighted by dashed circles). (b) EC1-EC2 cis interaction. Cis interactions are defined by the binding of an EC1 domain of one cadherin to an EC2 domain of an adjacent cadherin on the same cell.
In (b) and (c) the structure of each extracellular cadherin domain is represented using a ribbon diagram and highlighted by an oval. (c) EC1-EC1 trans interaction: Two views rotated by 90° of the trans binding of an EC1 domain of one cadherin to an EC1 domain of a cadherin on the adjacent cell. Only the EC1 and a portion of the EC2 domains are shown. Note that a small segment of polypeptide at the N-terminus of each of the two EC1 domains [highlighted in yellow (cell 1) and blue (cell 2)] swings out and replaces the equivalent segment from its binding partner (strand swap, dashed oval on the right). The strand swap places the side chain of a tryptophan residue on each of the segments into a binding pocket on the adjacent EC1 domain — an interaction that substantially stabilizes the trans binding. (d) The clustered protocadherins at neuron-to-neuron interfaces form a lattice of cis and trans interactions with their exoplasmic cadherin domains (ovals, EC1-EC6) that differs from that in classical cadherins (part a). Different protocadherin isoforms are represented by different colors (lighter colors expressed by Cell 1, darker colors expressed by Cell 2). The dimeric, homo- (same color) and heterophilic (different colors) cis interactions form between the EC6 domain of one molecule and EC5-EC6 domains of its partner (cis dashed circle). The dimeric, homophilic trans interactions form between the EC1-EC4 domains of one molecule with the EC1-EC4 domains of the same isoform (same color) on the adjacent cell. (e) The cytosolic domains of the classical E-cadherins bind directly or indirectly to multiple adapter proteins (e.g., β-catenin), which both connect the junctions to actin filaments (F-actin) of the cytoskeleton and participate in intracellular signaling pathways. Somewhat different sets of adapter proteins are illustrated in the two cells to emphasize the variety of adapters in adherens junctions. Some of these adapters, such as vinculin and ZO-1, can interact with several different CAMs. See V. Vasioukhin and E. Fuchs, 2001, Curr. Opin. Cell Biol. 13:76–84; and J. Brasch, et al., 2012, Trends Cell Biol. 22:299–310. [Parts (a–c) Data from O. J. Harrison et al., 2011, Structure 19: 244–256, PDB ID 3q2w. Part (d) Data from J. Brasch et al., 2019, Nature 569: 280–283, PDB ID 6e6b.] Description The illustration labeled (a) shows cell 1 at left and cell 2 at right, each with 5 rows of blue ovals moving toward the center between the cells. At the joint, where two of cell one ovals join, is the labeled cis, and a white arrow points to illustration (b). Also in illustration (a), where an oval from cell 1 joins an oval from cell 2 is labeled trans and a white arrow is pointing to illustration (c). The illustration labeled (b) shows a three-dimensional ribbon of the inside of each oval. Each oval is labeled from the cell 1 side E C 1 through E C 5. The E C 1 from the top row joins to the E C 1 from the next row down. The illustration labeled (c) shows the E C 1 from cell 1 joining to the E C 1 from cell 2, using three-dimensional ribbon diagrams. A small area of ribbon at the center is highlighted with yellow (cell 1) and blue (cell 2) colors showing how they interact. Next to the first ribbon diagram is another one from a 90-degree turn of the same ribbon diagram. The illustration labeled (d) titled protocadherin shows a schematic with cell 1 membrane at left and cell 2 membrane at right. From each cell, 3 sets of 6 ovals in chains extend toward the other cell. In this diagram, each pair of chains cross each other near the cell membrane, then the top chain from cell 1 pair joins four ovals to the bottom chain from cell 2. The illustration labeled (e) titled Ecadherin shows oval chains between the cell membranes of cell 1 at left and cell 2 at right. In the cytosol of cell 1, the ends of 4 chains have a protein attached, labeled from top to bottom: beta-catenin, alpha-catenin, F-actin, Z O- 1. In the cytosol of cell 2, four different proteins are diagramed and labeled, from top to bottom: V A S P, vinculin, alpha-actinin, and p120-catenin. Determination of the structures of the extracellular domains of cadherins, together with analyses of the structures and binding properties of many mutants of the key binding domains,
have provided a clear picture of the cis and trans interactions that underlie classical cadherin– mediated cell adhesion. The key features of cadherin cis and trans binding interactions are (1) the calcium-dependent curvature of the five extracellular cadherin domains that permits proper relative orientations of the EC1 and EC2 domains (see Figures 20-2 and 20-14a); (2) for cis interactions, the binding of one side of an EC1 domain to a complementary surface on the EC2 domain of an adjacent molecule on the same membrane (see Figures 20-2 and 20-14a and b); and (3) for trans interactions, the binding of a different surface of the EC1 domain to an EC1 domain from a cadherin molecule on the adjacent cell (Figure 20-14c). The trans EC1–EC1 binding is stabilized when a small segment of the protein at the N-terminus of each of the two EC1 domains swings out and replaces the equivalent segment from its binding partner (strand swap; see Figure 20-14c). Another group of CAMs, called clustered protocadherins, provides an excellent example of how evolution can employ common molecular building blocks — in this case, cadherin domains — for distinct structural and functional purposes. Clustered protocadherins play a role in establishing the complex wiring diagram of neurons in mammalian brains. Multiple protocadherin genes encode 52 different clustered protocadherin proteins (isoforms) in humans (58 in mice). The random combinations of 10–15 different isoforms that are expressed in any individual neuron provide a bar code that helps a neuron distinguish itself from other neurons and thus helps generate neuronal wiring diagrams. The domain architecture of these protocadherins with six extracellular cadherin domains (EC1–EC6) differs from that of classical cadherins with five domains, and the organization of the cis and trans binding (see
Figure 20-14d) differs markedly from that of classical cadherins (see Figure 20-14a–c). In clustered protocadherins, cis interactions are mediated by the EC6 domain of one molecule binding to the EC5 and EC6 domains of an adjacent molecule (Figure 20-14d). These cis interactions can be both homophilic and heterophilic, allowing the same or different isoforms to associate with each other into cis dimers [different colors represent different isoforms in part (d)]. In contrast, trans interactions are mediated by strictly homophilic, antiparallel binding of the EC1-EC4-domains (see Figure 20-14d). Trans binding only occurs when the sequences of all four of these EC domains perfectly match those of a corresponding protocadherin isoform on the adjacent cell. The lattices of intermixed protocadherin isoforms that form an interface between the membranes of neurons are responsible for their influence on neuronal wiring.
The cytosolic domains of CAMs have profound effects on the binding properties and functions of these adhesive molecules. For example, the C-terminal cytosolic domain of classical cadherins is linked to the actin cytoskeleton by adapter proteins (Figure 20-14e). These linkages are essential for strong adhesion. Indeed, cadherins together with their adaptor proteins in adherens junctions participate in mechanotransduction; a moderate increase in the mechanical tension at the junctions generated either intracellularly by actin and myosin, or by extracellular forces such as those transferred from adjacent cells or shear force due to blood flow, induces the formation of larger clusters of cadherins and stronger intercellular adhesion. An important mechanosensor is the adapter protein α-catenin, which is attached to the cytoplasmic domain of cadherin via the protein β-catenin and also binds directly to F-actin (see Figure 20-14e). When the cadherin/β-catenin/α-catenin/F-actin complex is subjected to mechanical stress, the C-terminal actin-binding domain of α-catenin undergoes a conformational change to bind more tightly to the actin. In addition, a tension-induced conformational change in α-catenin’s adhesion modulation domain uncovers otherwise cryptic binding sites for additional proteins, including the actin-binding adaptor vinculin. As a consequence, additional F-actin filaments can be recruited (see Figure 20-14e) and these further enlarge and strengthen the adherens junction. We will see vinculin playing a similar role in integrin-mediated adhesion later in this chapter. Disruption of the interactions between classical cadherins and α-catenin or β-catenin dramatically reduces cadherin-mediated cell-cell adhesion. This disruption occurs spontaneously in tumor cells, which sometimes fail to express α-catenin, and can be induced experimentally by depleting the cytosolic pool of accessible β-catenin. The cytosolic domains of cadherins also interact with other adaptor molecules including p120-catenin, which can influence the stability of the cadherin. Interestingly, β-catenin plays a dual role: it not only mediates cytoskeletal attachment, but also serves as a signaling molecule, translocating to the nucleus and altering gene transcription in the Wnt signaling pathway (see Figure 16-26). Classical cadherins play a critical role during tissue differentiation. Each classical cadherin has a characteristic tissue distribution. In the course of differentiation, the amounts or types of cell-surface cadherins change, affecting many aspects of cell-cell adhesion, cell migration, and cell division. For instance, the normal reorganization of tissues during morphogenesis is often accompanied by the conversion of nonmotile epithelial cells into motile cells, called mesenchymal cells, that are precursors for other tissues. This epithelial-to-mesenchymal
transition (EMT) is associated with a reduction in the expression of E-cadherin (Figure 2015a, b). The EMT is also associated with pathology, as in the conversion of epithelial cells into malignant carcinoma cells. For example, certain ductal breast tumors and hereditary diffuse gastric cancer (Figure 20-15c) characteristically involve a loss of E-cadherin activity. It is well known that animal cell-cell contact can inhibit cell proliferation. During tissue development, once dividing epithelial cells have formed a well-defined, tightly bound epithelium, they have no need for further cell division unless they are damaged or receive a signal to undergo the EMT. It is now clear that one mechanism used to inhibit proliferation of epithelial cells in epithelia is E-cadherin- and catenin-mediated regulation of the Hippo pathway that controls cell proliferation (see Chapter 21).
EXPERIMENTAL FIGURE 20-15 E-cadherin activity is lost during the epithelial-to-mesenchymal transition and during cancer progression. A protein called Snail that suppresses the expression of E-cadherin is associated with the epithelial-to-mesenchymal transition (EMT). (a) Normal epithelial MDCK cells grown in culture. (b) Expression of the snail gene in MDCK cells causes them to undergo an EMT. (c) Distribution of E-cadherin detected by immunohistochemical staining (dark brown) in a thin section of tissue from a patient with hereditary diffuse gastric cancer. E-Cadherin is seen at the intercellular borders of normal stomach gastric gland epithelial cells; no E-cadherin is seen at the borders of underlying invasive carcinoma cells.
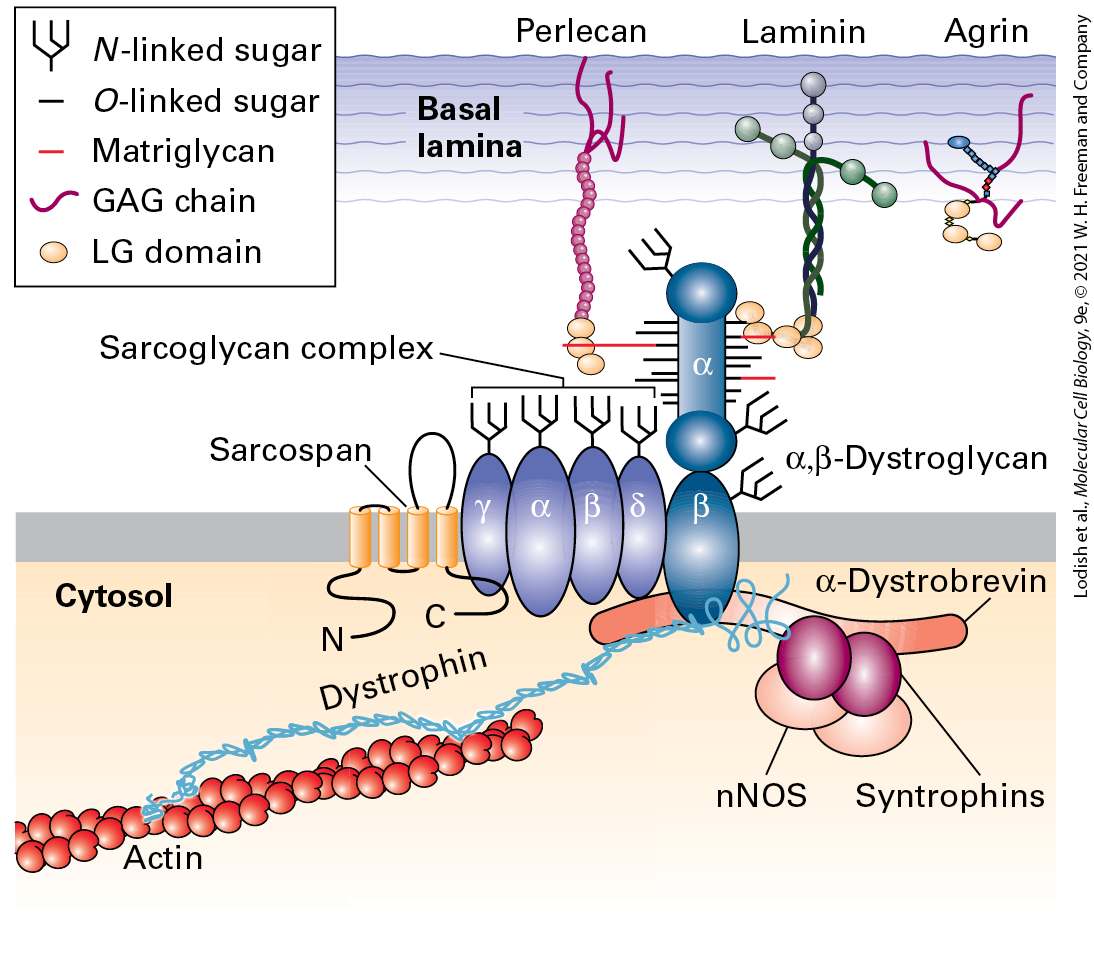
[Parts (a) and (b) republished with permission from Elsevier, from A. Martinez Arias, 2001, “Epithelial Mesenchymal Interactions in Cancer and Development,” Cell 105(4):425–431; permission conveyed through Copyright Clearance Center, Inc. Part (c) republished with permission from John Wiley & Sons, Inc., from F. Carneiro et al., 2004, “Model of the Early Development of Diffuse Gastric Cancer in E-Cadherin Mutation Carriers and Its Implications for Patient Screening,” J. Pathol. 203(2):681–687; permission conveyed through Copyright Clearance Center, Inc.] The firm epithelial cell-cell adhesions mediated by cadherins in adherens junctions permit the formation of a second class of intercellular junctions in epithelia — tight junctions, to which we will turn shortly. Infection with rhinoviruses (RV) is the most frequent cause of the common cold, and infection with virulent class C rhinoviruses (RV-C) can cause more severe illnesses, including exacerbation of asthma. To enter cells and replicate, RV-C must bind to cell-surface receptors. A cadherin family member called CDHR3, which is highly expressed in epithelial cells in the human airway, is a receptor for RV-C. Pathogens such as RV-C often evolve to co-opt proteins that have normal functions in their target (host) tissues. Genetic studies have shown that a naturally occurring mutation in humans that changes a cysteine to tyrosine (C → Y) in the EC5 domain of CDHR3 is associated with increased wheezing illnesses and hospitalizations for childhood asthma. In cultured cells, this C → Y mutation increases the cell-surface expression of CDHR3 and the binding and replication of RV-C. Treatments that disrupt the RV-C/cadherin (CDHR3) interaction have the potential to prevent or treat respiratory diseases caused by RVC. Desmosomal Cadherins Desmosomes (Figure 20-16) contain two specialized cadherins, desmoglein and desmocollin, whose cytosolic domains are distinct from those in the classical cadherins. The cytosolic domains of desmosomal cadherins bind to adapter proteins such as plakoglobin (similar in structure to β-catenin) and plakophilins, and these bind to a member of the plakin family of adapters, called desmoplakin. These adapters form the thick cytoplasmic plaques that are characteristic of desmosomes. The desmoplakins directly mediate plaque binding to intermediate filaments.
FIGURE 20-16 Desmosomes. (a) Model of a desmosome between epithelial cells with attachments to the sides of intermediate filaments. The key CAMs in desmosomes are the desmosomal cadherins desmoglein and desmocollin. Adapter proteins bound to the cytoplasmic domains of these cadherins include plakoglobin, desmoplakins, and plakophilins. See B. M. Gumbiner, 1993, Neuron 11:551–564; L. A. Staehelin and B. E. Hull, 1978, Sci. Am. 238:140; and D. R. Garrod, 1993, Curr. Opin. Cell Biol. 5:30–40. (b) Electron micrograph of a thin section of a desmosome connecting two cultured differentiated human keratinocytes. Bundles of intermediate filaments radiate from the two darkly staining cytoplasmic plaques that line the inner surface of the adjacent plasma membranes. Inset: Electron
Integrins Mediate Cell-Matrix Adhesions, Including Those in Epithelial-Cell Hemidesmosomes
microscopic tomograph of a desmosome linking two human epidermal cells (plasma membranes, pink; desmosomal cadherins, blue; bar, 35 nm). [Part (b) republished with permission from Nature, from A. Al-Amoudi et al., 2007, “The Molecular Architecture of Cadherins in Native Epidermal Desmosomes,” Nature 450(7171): 832–837; permission conveyed through Copyright Clearance Center, Inc.] Description The illustration labeled (a) shows a schematic of a desmosome in blue. Two cell membranes are with the blue connections of rows of blue ovals between them in the intercellular space. In the cytosol of each cell, intermediate filaments are labeled. The blue ovals are labeled desmoglein and desmocollin (cadherins). Inside the cell membrane on each side of the ovals is a large blue elongated oval labeled cytoplasmic plaque (plakoglobin, desmoplakins, plakophilins). The micrograph labeled (d) shows a desmosome with labels intermediate filaments, cytoplasmic plaques, and plasma membranes. The cadherin desmoglein was identified through studies of an unusual but revealing skin disease called pemphigus vulgaris, an autoimmune disease. Patients with autoimmune disorders synthesize self-attacking (or “auto”) antibodies that bind to a normal body protein. In pemphigus vulgaris, the auto-antibodies disrupt adhesion between epithelial cells, causing blisters of the skin and mucous membranes. The predominant auto-antibodies in patients were shown to be specific for desmoglein; indeed, the addition of such antibodies to normal skin induces the formation of blisters and disruption of cell adhesion. Integrins Mediate Cell-Matrix Adhesions, Including Those in Epithelial-Cell Hemidesmosomes To be stably anchored to solid tissues and organs, simple columnar epithelial sheets must be firmly attached via their basal surfaces to the underlying ECM (basal lamina). This attachment occurs via adhesion receptors called integrins (see Figure 20-2), which are located both within and outside of anchoring junctions called hemidesmosomes (see Figure 20-11a). Hemidesmosomes comprise integral membrane proteins linked via cytoplasmic adapter proteins (e.g., plakins) to keratin-based intermediate filaments. The principal ECM adhesion receptor in epithelial hemidesmosomes is integrin α6β4.
Integrins function as cell-cell and cell-ECM adhesion receptors in a wide variety of epithelial and nonepithelial cells, mediating many cell-matrix and cell-cell interactions (Table 20-4). In vertebrates, at least 24 integrin heterodimers, composed of 18 types of α subunits and 8 types of β subunits in various αβ heterodimeric combinations, are known. A single type of β chain can interact with any one of several different types of α chains, forming distinct integrins that bind different ligands. This phenomenon of combinatorial diversity allows a relatively small number of components to serve a large number of distinct functions. Although most cells express several distinct integrins that bind the same or different ligands, many integrins are expressed predominantly in certain types of cells. Not only do many integrins bind more than one ligand, but there are ligands that can bind to any one of several different integrins.
TABLE 20-4 • Selected Vertebrate Integrins Subunit Composition Primary Cellular Distribution Ligands α1β1 Many types Mainly collagens α2β1 Many types Mainly collagens; also laminins α3β1 Many types Laminins α4β1 Hematopoietic cells Fibronectin; VCAM-1 α5β1 Fibroblasts Fibronectin α6β1 Many types Laminins αLβ2 T lymphocytes
Icam-1, Icam-2
αMβ2 Monocytes Serum proteins (e.g., C3b, fibrinogen, factor X); ICAM-1 αIIbβ3 Platelets Serum proteins (e.g., fibrinogen, von Willebrand factor, vitronectin); fibronectin α6β4 Epithelial cells Laminin NOTE: The integrins are grouped into subfamilies having a common β subunit. Ligands shown in red are CAMs; all others are ECM or serum proteins. Some subunits can have multiple spliced isoforms with different cytosolic domains. SOURCE: Data from R. O. Hynes, 1992, Cell 69(1):11.
All integrins appear to have evolved from two ancient general subgroups: those that bind proteins containing the tripeptide sequence Arg-Gly-Asp, usually called the RGD motif (fibronectin is one such protein), and those that bind laminin. Several integrin α subunits contain a distinctive, inserted domain, the I-domain, which can mediate binding of certain integrins to various collagens in the ECM. Some integrins with I-domains are expressed exclusively on leukocytes (white blood cells) and red and white blood cell precursor (hematopoietic) cells. I-domains also recognize CAMs on other cells, including members of the Ig superfamily (e.g., ICAMs, VCAMs), and thus participate in cell-cell adhesion. Integrins typically exhibit low affinities for their ligands, with dissociation constants between and . However, the multiple weak interactions generated by the binding of hundreds or thousands of integrin molecules to their ligands on cells or in the ECM allow a cell to remain firmly anchored to its ligand-expressing target. Parts of both the α subunit and the β subunit of an integrin molecule contribute to the primary extracellular ligand-binding site (see Figure 20-2). Ligand binding to integrins also requires the simultaneous binding of divalent cations. Like that of other adhesion molecules, the cytosolic region of integrins interacts with adapter proteins, which in turn bind to the cytoskeleton and to intracellular signaling molecules (see Figure 20-8). Most integrins are linked via adapters to the actin cytoskeleton, including two of the integrins that connect the basal surface of epithelial cells to the basal lamina via the ECM molecule laminin. Some integrins, however, interact with intermediate filaments. The cytosolic domain of the β4 chain in the α6β4 integrin in hemidesmosomes (see Figure 20-1), which is much longer than the cytosolic domains of other integrin β chains, binds to specialized adapter proteins, which in turn interact with keratin-based intermediate filaments (see Table 20-4). Other integrins (e.g., α3β1) are the adhesion receptors in the focal contacts linking the epithelial basal lamina with the actin cytoskeleton (see Figure 20-1). As we will see, the diversity of integrins and their ECM ligands allows integrins to participate in a wide array of key biological processes, including the inflammatory response and the migration of cells to their correct locations during morphogenesis. The importance of integrins in diverse processes is highlighted by the defects exhibited by knockout mice engineered to have mutations in various integrin subunit genes. These defects include major abnormalities in development, blood vessel formation, leukocyte function, inflammation, bone remodeling, and
Tight Junctions Seal Off Body Cavities and Restrict Diffusion of Membrane Components
blood clotting. Despite their differences, all these processes depend on integrin-mediated interactions between the cytoskeleton and either the ECM or CAMs on other cells. In addition to their adhesion function, integrins can mediate outside-in and inside-out signaling (see Figure 20-8). The engagement of integrins by their extracellular ligands can, through adapter proteins bound to the integrin’s cytosolic region, influence the cytoskeleton and intracellular signaling pathways (outside-in signaling). Conversely, intracellular signaling pathways can alter the structure of integrins and consequently their abilities to adhere to their extracellular ligands and mediate cell-cell and cell-matrix interactions (inside-out signaling). We will see a detailed example of integrin-mediated signaling later in this chapter. Integrinmediated signaling pathways influence processes as diverse as cell survival, cell proliferation, and programmed cell death (see Chapter 21). Tight Junctions Seal Off Body Cavities and Restrict Diffusion of Membrane Components For polarized epithelial cells to function as barriers and mediators of selective transport, extracellular fluids surrounding their apical and basolateral membranes must be kept separate. Tight junctions between adjacent epithelial cells are usually located in a band surrounding the cell just below the apical surface (Figure 20-17; see also Figure 20-11). These specialized junctions form a barrier that seals off body cavities such as the intestinal lumen and separates the blood from the cerebral spinal fluid of the central nervous system (i.e., the blood-brain barrier).
FIGURE 20-17 Tight junctions. (a) Freeze-fracture preparation of tight-junction zone between two intestinal epithelial cells. The fracture plane passes through the plasma membrane of one of the two adjacent cells (see also
Figure 20-11). A honeycomb-like network of ridges and grooves below the microvilli constitutes the tight-junction zone. (b) Schematic drawing shows how a tight junction might be formed by the linkage of rows of protein particles in adjacent cells. In the inset micrograph of an ultrathin sectional view of a tight junction, the adjacent cells can be seen in close contact where the rows of proteins interact. See L. A. Staehelin and B. E. Hull, 1978, Sci. Am. 238:140. [Part (b) republished with permission from Nature, from S. Tsukita, M. Furuse, and M. Itoh, 2001, “Multifunctional Strands in Tight Junctions,” Nat. Rev. Mol. Cell Biol. 2(4):285–293; permission conveyed through Copyright
Clearance Center, Inc.] Description The micrograph labeled (a) shows gray rows of cells labeled tight junction, which is below an area labeled villi. The illustration labeled (b) shows the microvilli at the top, and the below is the tight junction labeled below. The tight junction has sections labeled intercellular spaces and linkage of protein particles in adjacent cells. Also labeled are rows of protein particles that stick out from the linkages. Next to this is a small micrograph of the linkage-intercellular space chain. Tight junctions prevent the diffusion of macromolecules and, to varying degrees, small watersoluble molecules and ions across an epithelium via the spaces between cells. They also help establish and maintain the polarity of epithelial cells by preventing the diffusion of membrane proteins and glycolipids between the apical and the basolateral regions of the plasma membrane, ensuring that these regions contain different membrane components. Indeed, the lipid compositions of the apical and basolateral regions of the plasma membrane’s exoplasmic leaflet (see Chapter 10) are distinct. Essentially all cell surface glycolipids are restricted to the exoplasmic face of the apical membrane, as are all proteins linked to the membrane by a glycosylphosphatidylinositol (GPI) anchor (see Figure 10-19). In contrast, the apical and basolateral regions of the plasma membrane’s cytosolic leaflet have uniform membrane composition in epithelial cells; their lipids and proteins can apparently diffuse laterally from one region of the membrane to the other. Tight junctions are composed of thin bands of plasma-membrane proteins that completely encircle the cell and are in contact with similar thin bands on adjacent cells. When thin sections of the tight junctions in cells are viewed in an electron microscope, the lateral surfaces of adjacent cells appear to touch each other at intervals and even to fuse in the zone just below the apical surface (see Figure 20-11b). In freeze-fracture preparations, tight junctions appear as an interlocking network of ridges and grooves in the plasma membrane (Figure 20-17a). Very high magnification reveals that rows of protein particles 3–4 nm in diameter form the ridges seen in freeze-fracture micrographs of tight junctions. In the model shown in Figure 20-17b, the tight junction is formed by a double row of these particles, one row donated by each cell. Treatment of an epithelium with the protease trypsin destroys the tight junctions, supporting the proposal that proteins are essential structural components of these junctions.
There are a number of integral membrane proteins found in tight junctions; two of the best studied are occludin and claudin (from the Latin claudere, “to close”). Each of these proteins has four membrane-spanning α helices (Figure 20-18) and are representative of a group of such cell-surface proteins called tetraspanins. The mammalian claudin gene family encodes as many as 27 homologous proteins that exhibit distinct tissue-specific patterns of expression and, as described below, exhibit distinct properties. A group of junction adhesion molecules (JAMs) have also been found to contribute to homophilic adhesion and other functions of tight junctions. JAMs and another junctional protein, the coxsackievirus and adenovirus receptor (CAR), contain a single transmembrane α helix and belong to the Ig superfamily of CAMs. The extracellular domains of rows of occludin, claudin, and JAMs in the plasma membrane of one cell apparently form extremely tight links with similar rows of the same proteins in an adjacent cell, creating a tight seal. -dependent cadherin-mediated adhesion also plays an important role in tight-junction formation, stability, and function.
FIGURE 20-18 Proteins that compose tight junctions. (a) Immunofluorescence localization of occludin (green) and tricellulin (red) in mouse intestinal epithelium. Note that tricellulin is predominantly concentrated in tricellular junctions. (b) The junction adhesion molecule (JAM) has a single transmembrane domain and an extracellular region with two immunoglobulin domains, whereas occludin and claudins contain four transmembrane helices. The transmembrane helices of claudin-15 form a four-helix bundle, and the extracellular loops contain a five-stranded β sheet (seen edgewise in this view) and α helices. These extracellular secondary structures participate in cis interactions that form a single-file row of claudins, which in turn interact via other cis contacts with an adjacent row of claudins in the same membrane of each cell. There are also trans interactions between the rows of claudins in adjacent cells (see also Figure 20-20). [Part (a) J. Ikenouchi et al., 2005, J. Cell Biol. 171(6):939–945, Fig. 3A; https://doi.org/10.1083/jcb.200510043. Part (b) structure of claudin-15 data from H. Suzuki et al., 2014, Science 344:304–307, PDB ID 4p79.] Description
The micrograph labeled (a) shows the fluorescence staining of occludin with a net of green threads attaching to green bead-like areas and the staining of tricellulin with a dim net of threads and enlargements. The illustration labeled (b) shows the schematic of J A M looking like a spring shape moving to a red square in the cell membrane. The N terminus and C terminus are labeled. The schematic of occludin looks like a pair of headphones above the cell membrane with long coiled wires leading into the cytosol. The claudin schematic looks like a pair of headphones with short wires into the cytosol. This schematic includes a three-dimensional ribbon diagram with four highlighted ribbons represents the areas that are in the cell membrane. At the intersection of three cells connected to one another by tight junctions (see Figure 20-13 and Figure 20-18a), two additional transmembrane proteins are incorporated into the tight junctions: tricellulin, which has four membrane-spanning helices; and angulins, which have a single transmembrane helix and one extracellular immunoglobulin domain and appear to be required for the assembly of tricellulin where the cells intersect. As with tight junctions, there appear to be tension-sensitive proteins that localize at tricellular vertices in at least some adherens junctions. As is the case for adherens junctions and desmosomes, cytosolic adapter proteins and their connections to the cytoskeleton are critical components of tight junctions and can mediate signaling. For example, when one specific claudin, claudin-6, is engaged in intercellular tight junctions, its cytoplasmic domain can initiate a kinase signaling cascade that results in the activation of nuclear receptors (e.g., the retinoic acid receptor γ and the estrogen receptor α) to control cell behavior (see Chapter 8). Also, the long C-terminal cytosolic segment of occludin binds to PDZ domains in some large, multidomain adapter proteins. PDZ domains are about 80 to 90 amino acids long and are found in various cytosolic proteins; they mediate binding to other cytosolic proteins or to the C-termini of particular plasma-membrane proteins. Cytosolic proteins containing a PDZ domain often have more than one of them. In the human genome, there are more than 250 PDZ domains in hundreds of proteins. Proteins with multiple PDZ domains can serve as scaffolds on which to assemble proteins into larger functional complexes. Several multiple-PDZ-domain–containing adapter proteins are associated with tight junctions, including the zonula occludens (ZO) proteins ZO-1, ZO-2, and ZO-3, which not only interact with occludin, claudin, and other adapter and signaling proteins but also mediate association with actin fibers. These interactions appear to stabilize the linkage between occludin and claudin molecules that is essential for maintaining the integrity of tight junctions. ZO proteins, which may form phase-separated biomolecular condensates at tight junctions, can also function as adapters in adherens junctions (see Figure 20-14) and gap junctions.
The role of tight junctions as permeability barriers has been demonstrated in many experiments. For example, in one simple experiment, lanthanum hydroxide (an electron-dense colloid of high molecular weight) is injected into the pancreatic blood vessel of an experimental animal; a few minutes later, the pancreatic epithelial acinar cells are fixed and prepared for microscopy. As shown in Figure 20-19, the lanthanum hydroxide diffuses from the blood into the space that separates the lateral surfaces of adjacent acinar cells, but it cannot penetrate past the tight junction. EXPERIMENTAL FIGURE 20-19 Tight junctions prevent passage of large molecules through extracellular spaces between epithelial cells. Tight junctions in the pancreas are impermeable to the large water-soluble colloid lanthanum hydroxide (dark stain) administered from the basolateral side of the epithelium. [From D. S. Friend and N. B. Gilula, 1972, J. Cell Biol. 53(3):758–776.] Description The micrograph has the following labels: at the top, the apical surface of the left cell and the right cell are labeled. Below them, a narrow area is labeled tight junction. Just below the tight junction is a black thick line labeled lanthanum hydroxide (between cells). The lateral surface of the left cell and right cell are labeled next to the black thick line.
As a consequence of tight junctions, many nutrients cannot move across an epithelium between cells; instead, their transport is achieved in large part through the transcellular pathway via specific membrane-bound transport proteins (Figure 20-20a; see also Figure 11-30). The barrier to diffusion provided by tight junctions, however, is not absolute; some can exhibit size- and ion-selective permeability, depending on the type of claudins incorporated into the tight junction. For some, but not all, claudins the intercellular trans association of their closely packed extracellular domains results in the formation of paracellular channels (pores) (Figure 20-20b). Thus certain small molecules and ions can move from one side of the epithelium to the other through the paracellular pathway (see Figure 20-20). The importance of selective permeability is highlighted by the evolutionary conservation of the molecules that establish it and the diseases that arise when it is disrupted. For example, murine embryos cannot develop properly if selective permeability is disrupted because proper fluid balance on the two sides of epithelia cannot be maintained. Similarly, the kidneys depend on proper tight-junction permeability to establish the ion gradients necessary for normal regulation of body fluids and waste removal. Owing at least in part to the varying properties of the different types of claudin molecules located in different tight junctions, the permeability of the tight junctions to ions, small molecules, and water varies enormously among different epithelial tissues.
FIGURE 20-20 Transcellular and paracellular pathways of transepithelial transport. (a) Transcellular transport requires the cellular uptake of molecules on one side of the cell and subsequent release on the opposite side by mechanisms for crossing membranes (see Chapter 11). In paracellular transport molecules move extracellularly through channels in the tight junctions, whose permeability to small molecules and ions depends on the composition of the junctional components (mainly claudins) and the physiological state of the epithelial cells. A cross-sectional plane through the tight junction linking the right-most two cells is indicated by dashed lines. Part (b) shows a perpendicular view of this plane (90 ° rotation). (b) A computational model of claudin-15 in a tight junction. No other tight-junctional proteins are included in this model. This perpendicular view of the cross-sectional plane illustrated in part (a) shows the transmembrane helices of claudin-15 molecules spanning the tight junctional membranes. In each of the two cells, there are adjacent, single-file rows of claudins (orange and blue) that form trans intercellular contacts with the rows of claudins in the adjacent cell. Extracellular channels that are oriented perpendicular to this cross-
sectional plane (outlined by gray circles) are formed by the extracellular domains of claudin-15. These channels permit the paracellular movement of small molecules and ions across the epithelium. There are negatively charged side chains of aspartic acid residues that line the channels and appear to contribute to the cation specificity of claudin15-dependent channels. [Part (a) Data from S. Tsukita et al., 2001, Nat. Rev. Mol. Cell Biol. 2:285. Part (b) Information from P. Samanta et al., 2018, J. Gen. Physiol. 150:949–968.] Description The illustration labeled (a) shows a schematic of three pathways between cells. 4 rectangular cells are with nuclei. Between the first two cells is a pathway labeled paracellular pathway. Between the second and third cells is a transcellular pathway, and between the third and fourth cells is an extracellular channel. The illustration labeled (b) shows a three-dimensional ribbon diagram of the extracellular channel. Cell 1 is at the left and the cytosol of cell 1 is labeled. The cell membranes of cells 1 and 2 are wide and have the ribbons inside them. Between cell 1 and cell 2, the connections between the ribbons are displayed. The area inside the membrane is also labeled claudins in the tight junction. The permeability of tight junctions can be altered by intracellular signaling pathways, especially G protein and cyclic AMP–coupled pathways (see Chapter 15). The regulation of tight-junction permeability is often studied by measuring ion flux (electrical resistance, called transepithelial resistance) or the movement of radioactive or fluorescent molecules across monolayers of MDCK or other epithelial cells. The importance of paracellular transport is apparent in several human diseases. In hereditary hypomagnesemia, a defect in the claudin16 gene prevents the normal paracellular flow of magnesium in the kidney. This defect results in an abnormally low blood level of magnesium, which can lead to convulsions. Furthermore, a mutation in the claudin14 gene causes hereditary deafness, apparently by altering transport around hair-cell epithelia in the cochlea of the inner ear. Some pathogens have evolved to exploit the molecules in tight junctions. Some use junctional proteins as co-receptors to attach to cells prior to infecting them (e.g., hepatitis C virus uses claudin-1 and occludin, together with two other co-receptors, to enter liver cells). Others break down the tight-junction barrier and cross epithelia via paracellular movement, and still others produce toxins that alter barrier function. For example, toxins produced by Vibrio cholerae, the enteric bacterium that causes cholera, alter the permeability barrier of the intestinal epithelium
Gap Junctions Composed of Connexins Allow Small Molecules to Pass Directly Between the Cytosols of Adjacent Cells
by altering the composition or activity of tight junctions. Vibrio cholerae also releases a protease that disrupts tight junctions by degrading the extracellular domain of occludin. Other bacterial toxins can affect the ion-pumping activity of membrane transport proteins in intestinal epithelial cells. Toxin-induced changes in tight-junction permeability (increased paracellular transport) and in protein-mediated ion pumping (increased transcellular transport) can result in massive losses of internal body ions and water into the gastrointestinal tract, which in turn leads to diarrhea and potentially lethal dehydration (see Chapter 11). It is clear that tight junctions are important for maintaining tissue integrity and function. Thus these junctions should and can respond to dynamic changes in epithelia, for example, when they are subjected to either external forces (such as stretching of the bladder by filling with urine) or internal forces (actin-myosin-induced changes in tension). It is clear that when there are changes in the mechanical forces at the tight junction there are coordinated changes in the composition of junctional adaptor proteins and paracellular permeability. The molecular mechanisms underlying mechanotransduction at tight junctions are not as well understood as those at adherens junctions, with some studies suggesting that ZO-1 may serve as a mechanosensor. Gap Junctions Composed of Connexins Allow Small Molecules to Pass Directly Between the Cytosols of Adjacent Cells Early electron micrographs of tissues revealed sites of cell-cell contact with a characteristic intercellular gap (Figure 20-21a). This feature, which was found in virtually all animal cells that contact other cells, prompted early morphologists to call these regions gap junctions. In retrospect, the most important feature of these junctions is not the 2–4 nm gap itself, but a well-defined set of cylindrical particles that cross the gap and compose pores connecting the cytosols of adjacent cells (Figure 20-21b, c). As we will see in Section 20.6, plant cells also assemble pores that connect the cytosols of adjacent cells, but those channels, called plasmodesmata, differ considerably in structure from gap junctions.
FIGURE 20-21 Gap junctions. (a) In this thin section through a gap junction connecting two mouse liver cells, the two plasma membranes are closely associated for a distance of several hundred nanometers, separated by a gap of 2–3 nm. (b) Numerous roughly hexagonal particles are visible in this perpendicular view of the cytosolic face of a region of plasma membrane enriched in gap junctions. Each hexagonal particle aligns with a similar particle on an adjacent cell, forming a channel connecting the two cells. (c) Schematic model of a gap junction connecting two plasma membranes. Both membranes contain connexon hemichannels, cylinders of six dumbbell-shaped connexin molecules. Two connexons join in the gap between the cells to form a gap-junction channel, 1.4–2.0 nm in diameter, that connects the cytosols of the two cells. (d) Structure of recombinant human Cx26 gap junction as determined by x-ray crystallography (3.5-Å resolution). Left: Space-filling model of a side view of the complete structure of two attached connexons oriented as in part (c). Each of the six connexins that comprise a connexon has four transmembrane helices and is shown in a distinct color. The structures of the loops connecting the transmembrane helices are not well defined and not shown. Right: View from the cytosol perpendicular to the membrane bilayers, looking down on the connexon with its central pore. The diameter of the pore’s channel is ∼14 Å, and it is lined by many polar/charged amino acids. See S. Nakagawa, S. Maeda, and T. Tsukihara, 2010, Curr. Opin. Struct. Biol. 20(4):423–430. [Part (a) courtesy of Daniel Goodenough. Part (b) D. L. Caspar et al., 1977, 74:605–628, Fig. 2b; https://doi.org/10:1083/jcb.74.2.605. Part (c) Data from L. A. Staehelin and B. E. Hull, 1978, Sci. Am. 238:140. Part (d) Data from S. Maeda et al., 2009, Nature 458:597–602, PDB ID 2zw3.] Description
The micrograph labeled (a) shows the gap junction as two long lines running vertically close together but not twisted. The micrograph labeled (b) shows many ends of tubes gathered. The illustration labeled (c) shows a schematic of a gap junction. The two cell membranes are vertical yellow lines with a pair of orange tubes connecting them at the center. The top tube looks closed and is labeled connexon hemichannel. The bottom tube looks open and is labeled gap junction channel. The area between the membranes is labeled intercellular area. A white arrow from the bottom tube points to illustration (d). The illustration labeled (d) shows two threedimensional space-filling models of the gap junction, one side view, and one top view. In the side view, the cytosol and membrane of cell 1 and cell 2 are labeled at the top, with the intercellular area labeled in the middle. At the bottom are width measurements for each part of the gap junction. From left to right: the area in cell 1 cytosol is 19 angstroms, the area inside the membrane is 38 angstroms, the area of the intercellular area is 40 angstroms, cell 2 membrane, 38 angstroms, and cell 2 cytosol, 19 angstroms. In many animal tissues, anywhere from a few to thousands of gap-junction particles cluster together in patches (e.g., along the lateral surfaces of epithelial cells; see Figure 20-11). When the plasma membrane is purified and then sheared into small fragments, some pieces mainly containing patches of gap junctions are generated. Owing to their relatively high protein content, these fragments have a higher density than the bulk of the plasma membrane and can be purified by equilibrium density-gradient centrifugation (see Figure 4-37). When these preparations are viewed perpendicular to the membrane, the gap junctions appear as arrays of hexagonal particles that enclose water-filled channels (see Figure 20-21b). The effective pore size of gap junctions can be measured by injecting a cell with a fluorescent dye covalently linked to membrane bilayer–impermeable molecules of various sizes and observing with a fluorescence microscope whether the dye passes into neighboring cells. Gap junctions between mammalian cells permit the passage of molecules as large as 1.2 nm in diameter. In insects, these junctions are permeable to molecules as large as 2 nm in diameter. Generally speaking, molecules smaller than 1200 Da pass freely and those larger than 2000 Da do not pass; the passage of intermediate-sized molecules is variable and limited. Thus ions, many low-molecular-weight precursors of cellular macromolecules, products of intermediary metabolism, and small intracellular signaling molecules can pass from cell to cell through gap junctions. In neural tissue, some neurons are connected by gap junctions through which ions pass rapidly, thereby allowing very rapid transmission of electrical signals. Impulse transmission through these connections, called electrical synapses, is almost a thousand times as rapid as at chemical synapses (see Chapter 23). Gap junctions are also present in many non-neural tissues,
where they help to integrate the electrical and metabolic activities of many cells. In the heart, for instance, gap junctions rapidly pass ionic signals among cardiac muscle cells, which are tightly bound together via desmosomes. Thus gap junctions contribute to the electrically stimulated coordinate contraction of cardiac muscle cells during a heartbeat. As discussed in
Chapter 15, some extracellular hormonal signals induce the production or release of small intracellular signaling molecules called second messengers (e.g., cyclic AMP, , and ) that regulate cellular metabolism. Because many second messengers can be transferred between cells through gap junctions, hormonal stimulation of one cell has the potential of triggering a coordinated response by that cell as well as many of its neighbors. Such gapjunction-mediated signaling plays an important role, for example, in the secretion of digestive enzymes by the pancreas and in the coordinated muscular contractile waves (peristalsis) in the intestine. Another vivid example of gap-junction-mediated transport is the phenomenon of metabolic coupling, or metabolic cooperation, in which a cell transfers nutrients or intermediary metabolites to a neighboring cell that is itself unable to synthesize them. Gap junctions play critical roles in the development of egg precursors (oocytes) in the ovaries by mediating the movement of both metabolites and signaling molecules, such as cyclic GMP, between an oocyte and its surrounding granulosa cells, as well as between neighboring granulosa cells. A current model of the structure of the gap junction is shown in Figure 20-21c–d. Vertebrate gap junctions are composed of connexins, a family of structurally related transmembrane proteins with molecular weights between 26,000 and 60,000. Each vertebrate hexagonal particle consists of 12 noncovalently associated connexin molecules: six form a cylindrical hemichannel, called a connexon, in one plasma membrane that is joined to a connexon in the adjacent cell membrane, forming a continuous aqueous channel (diameter ∼14 Å) between the cells. Each individual connexin molecule has four membrane-spanning α helices with a topology similar to that of claudin (see Figure 20-18), resulting in 24 transmembrane α helices in each connexon hemichannel. A completely different family of proteins, the innexins, forms the gap junctions in invertebrates. A third family of innexin-like proteins, called pannexins, has been found in both vertebrates and invertebrates. Pannexins form hexamer hemichannels (pannexons) whose opening can be regulated by changes in membrane potential or mechanical stress. When open, pannexons permit direct exchange of small molecules (such as ATP) and ions between the
intracellular and extracellular spaces. Pannexons are thought to play key roles in release of ATP from cells into the extracellular space. Extracellular ATP (as well as ADP and AMP) can function as an intercellular messenger or transmitter by binding to and activating the cellsurface purinergic receptors P1, P2X, or P2Y on target cells. There are 21 different connexin genes in humans, and different sets of connexins are expressed in different cell types. The existence of this diversity, together with the generation of mutant mice with inactivating mutations in connexin genes, has highlighted the importance of connexins in a wide variety of cellular systems. Some cells express a single connexin that forms homotypic connexons. Most cells, however, express at least two connexins; these different proteins can assemble into heteromeric connexons, which in turn form heterotypic gap-junction channels. Diversity in channel composition leads to differences in channel permeability. For example, channels made from a 43-kDa connexin isoform, Cx43 — the most ubiquitously expressed connexin — are more than a hundred times as permeable to ADP and ATP as those made from Cx32 (32 kDa). The permeability of gap junctions is regulated by post-translational modification of connexins (e.g., phosphorylation) and is sensitive to changes in environmental conditions such as intracellular pH and concentration, membrane potential, and the intercellular potential between adjacent interconnected cells (“voltage gating”). The N-termini of connexins appear to be especially important in the gating mechanism. Thus, as is the case for many ion channels (see Chapter 11), the channel in some gap junctions can be either opened or closed. One example of the physiological regulation of gap junctions occurs during mammalian childbirth. The smooth muscle cells in the mammalian uterus must contract strongly and synchronously during labor to expel the fetus. To facilitate this coordinated activity, immediately before and during labor there is an approximately five- to tenfold increase in the amount of the major connexin in these cells, Cx43, and an increase in the number and size of gap junctions, which is reversed rapidly postpartum (following childbirth). The assembly of connexins, their trafficking within cells, and the formation of functional gap junctions apparently depend on N-cadherin and its associated adapter proteins (e.g., α- and β-catenins, ZO-1, and ZO-2) as well as desmosomal proteins (plakoglobin, desmoplakin, and plakophilin-2). PDZ domains in ZO-1 and ZO-2 bind to the C-terminus of Cx43 and mediate its interaction with catenins and N-cadherin. The relevance of these relationships is
Tunneling Nanotubes Can Mediate Metabolic Coupling and Transfer Organelles Between Animal Cells
particularly evident in the heart, which depends on gap junctions for rapid coordinated electrical coupling and on adjacent adherens junctions and desmosomes for mechanical coupling between cardiomyocytes to achieve the intercellular integration of electrical activity and movement required for normal cardiac function. It is noteworthy that ZO-1 serves as an adapter for adherens (see Figure 20-14), tight, and gap junctions, suggesting that this and other adapters can help integrate the formation and functions of these diverse junctions. Mutations in connexin genes cause at least eight human diseases, including neurosensory deafness (Cx26 and Cx31), cataracts or heart malformations (Cx43, Cx46, and Cx50), and the X-linked form of Charcot-Marie-Tooth disease (Cx32), which is marked by progressive degeneration of peripheral nerves. Tunneling Nanotubes Can Mediate Metabolic Coupling and Transfer Organelles Between Animal Cells Tunneling nanotubes can serve as an alternative to gap junctions for moving molecules between cells. Tunneling nanotubes are tubelike projections of the plasma membrane that form a continuous channel connecting the cytosols of animal cells (Figure 20-22) and can transfer chemical and electrical signals between cells in a manner analogous to structures in plants called plasmodesmata (described in Section 20.6). Tunneling nanotubes are typically unbranched, straight tubes and can have a wide variety of diameters (50–300 nm) and lengths (extending between cells from , they can be longer than several cell diameters). All tunneling nanotubes have actin filaments passing through the central channel, and in some types of cells they also contain microtubules. Remarkably, functional mitochondria can travel between cells by passing through tunneling nanotubes in cell culture (see Figure 20-22) and in vivo, thereby rescuing receiving cells that have mitochondrial defects or deficiencies. Thus the concept of metabolic coupling described for gap junctions can be extended to include the movement of small molecules and organelles through tunneling nanotubes. Pathogens may also use tunneling nanotubes to spread between cells.
EXPERIMENTAL FIGURE 20-22 Microscopic visualization of a tunneling nanotube and mitochondria in cultured human cells. (a) Cultured human retinal pigment epithelial cells (ARPE-19 cell line) were incubated with a fluorescent dye (JC-1) that specifically stains mitochondria and then examined by a combination of conventional bright-field microscopy (see Chapter 4) to visualize the cells and fluorescence microscopy to visualize mitochondria (green intracellular fluorescence). A typical tunneling nanotube can be seen connecting cells 1 and 2. Inset (b) shows a higher magnification of the bright-field-only image with two bulges in the tunneling nanotube highlighted by dashed circles. Inset (c) shows a higher magnification of the same region of the combination image indicating two likely mitochondria within the tunneling nanotube at the positions of those bulges. [D. Wittig et al., 2012, “Multi-Level Communication of Human Retinal Pigment Epithelial Cells via Tunneling Nanotubes.” PLoS One 7(3):e33195; https://doi.org/10.1371/journal.pone.0033195.] KEY CONCEPTS OF SECTION 20.2 Cell-Cell and Cell–Extracellular Matrix Junctions and Their Adhesion Molecules Epithelial cells have distinct apical, basal, and lateral surfaces. Microvilli projecting from the apical surfaces of many epithelial cells considerably expand the cells’ surface areas. Three major classes of cell junctions — anchoring junctions, tight junctions, and gap junctions — assemble epithelial cells into sheets and mediate communication between them (see Figures 20-1 and 20-11). Anchoring junctions can be further subdivided into adherens junctions, focal contacts, desmosomes, and hemidesmosomes.
Adherens junctions and desmosomes are cadherin-containing anchoring junctions that bind the membranes of adjacent cells, giving strength and rigidity to the entire tissue. Cadherins are cell-adhesion molecules (CAMs) responsible for -dependent interactions among cells in epithelial and other tissues. They promote strong cell-cell adhesion by mediating both lateral (cis) and adhesive intercellular (trans) interactions. Adapter proteins that bind to the cytosolic domain of cadherins and other cell-cell and cell matrix adhesion receptors mediate the association of cytoskeletal and signaling molecules with the plasma membrane (see Figures 20-8 and 20-14). Strong cell-cell adhesion depends on the linkage of the interacting adhesion receptors to the cytoskeleton. Hemidesmosomes are integrin-containing anchoring junctions that attach cells to elements of the underlying extracellular matrix. Integrins are a large family of αβ heterodimeric cell-surface proteins that mediate both cell-cell and cell-matrix adhesions and inside-out and outside-in signaling in numerous tissues. Tight junctions block the diffusion of proteins and some lipids in the plane of the plasma membrane, contributing to the polarity of epithelial cells. They also limit and regulate the extracellular (paracellular) flow of water and solutes from one side of the epithelium to the other (see Figure 20-20). Two key integral membrane proteins found in tight junctions are occludin and claudin. Gap junctions are constructed of multiple copies of connexin proteins, which are assembled into a transmembrane channel that connects the cytosols of two adjacent cells (see Figure 20-21). Small molecules and ions can pass through gap junctions, permitting metabolic and electrical coupling of adjacent cells. Tunneling nanotubes are tubelike projections of plasma membranes that form a continuous channel connecting the cytosols of nearby animal cells (see Figure 20-22).
20.3 The Extracellular Matrix I: The Basal Lamina
20.3 The Extracellular Matrix I: The Basal Lamina In animals, the extracellular matrix (ECM) has multiple functions (see
Table 20-2). The ECM helps organize cells into tissues and coordinates their cellular functions by activating intracellular signaling pathways that control cell growth, proliferation, and gene expression. The ECM can directly influence cell and tissue structure and function. In addition, it can serve as a repository for inactive or inaccessible signaling molecules (e.g., growth factors) that are released to function when the ECM is disassembled or remodeled by hydrolyases, such as proteases. Indeed, hydrolyzed fragments of ECM macromolecules can themselves have independent biological activity. The ensemble of proteins that compose the ECM itself and associated proteins that covalently modify (e.g., chemically cross-link, phosphorylate, cleave), bind to, or otherwise regulate the composition and structure of the ECM is called the matrisome. Proteomic (Chapter 3) and genomic analyses suggest that there are approximately 1030 and 1110 genes that encode the human and mouse matrisomes, respectively. Dysfunction of matrisome components can cause a wide variety of diseases that affect many different tissues and organs. It is noteworthy that there are ECM components, as well as extracellular domains of plasma-membrane proteins, that are phosphorylated on serine, threonine, or tyrosine side chains. Kinases that are present in the luminal compartments of the secretory pathway and
The Basal Lamina Provides a Foundation for Assembly of Cells into Tissues
some that are apparently secreted into the extracellular space catalyze these phosphorylations. Many functions of the ECM and, indeed, some features of the assembly of the ECM require transmembrane adhesion receptors, including the integrins, that bind directly to ECM components and that also interact, through adapter proteins, with the cytoskeleton. Adhesion receptors bind to three types of molecules abundant in the ECM of all tissues (see Table 20-1): Proteoglycans, a group of glycoproteins that cushion cells and bind a wide variety of extracellular and cell-surface molecules Collagen fibers, which provide structural integrity, mechanical strength, and resilience Soluble multi-adhesive matrix proteins, such as laminin and fibronectin, which bind to and cross-link adhesion receptors and other ECM components We begin our description of the structures and functions of these major ECM components in the context of the basal lamina: the specialized sheet of ECM that plays a particularly important role in determining the overall architecture and function of epithelial tissues. In the following section, we discuss the ECM molecules commonly found in nonepithelial tissues, including connective tissue. The Basal Lamina Provides a Foundation for Assembly of Cells into
Tissues In animals, most organized groups of cells in epithelial and nonepithelial tissues are underlain or surrounded by the basal lamina, a sheet-like meshwork of ECM components usually no more than 60–120 nm thick (Figure 20-23). The basal lamina is structured differently in different tissues. In columnar and other epithelia such as intestinal lining and skin, it is a foundation on which only one surface of the cells rests. In other tissues, such as muscle or fat, the basal lamina surrounds each cell. Basal laminae play important roles in regeneration after tissue damage and in embryonic development. For instance, the basal lamina helps four- and eight-celled embryos adhere together in a ball (see Figures 22-2 and 22-3). In the development of the nervous system, neurons migrate along ECM pathways that contain basal laminal components. In higher animals, two distinct basal laminae are employed to form a tight barrier that limits diffusion of molecules between the blood and the brain (blood-brain barrier), and in the kidney, a specialized basal lamina serves as a selectively permeable blood filter. In muscle, the basal lamina helps protect the cell membranes from damage during contraction and relaxation. Thus the basal lamina is important for organizing cells into tissues and distinct compartments, repairing tissues, forming permeability barriers, and guiding migrating cells during development. It is therefore not surprising that its components have been highly conserved throughout evolution.
FIGURE 20-23 A basal lamina separates epithelial cells and some other cells from connective tissue. (a) Transmission electron micrograph of a thin section of cells (top) and underlying connective tissue (bottom). The electron-dense layer of the basal lamina can be seen to follow the undulations of the basal surfaces of the cells. (b) Electron micrograph of a quick-freeze deep-etch preparation of skeletal muscle, showing the plasma membrane, basal lamina, and surrounding connective-tissue collagen fibers. In this preparation, the basal lamina is revealed as a meshwork of filamentous proteins that associates with the plasma membrane and the thicker collagen fibers of the connective tissue. Description In the micrograph labeled (a) the bottom area is light in color and labeled connective tissue. At the top of the light area is a tube-like line going across it and labeled basal lamina. A dark line on top of this is labeled basal surface, and the area above this is labeled cytosol. The micrograph labeled (b) shows a micrograph of bone tissue. There is a thin area that separates the plasma membrane and basal lamina and moves in a crooked line from bottom left to top right of the micrograph. This area is labeled adhesion receptors. An area below and to the right of the basal lamina is labeled collagen fibers. Most of the ECM components in the basal lamina are synthesized by the cells that rest on it. Four ubiquitous protein components, each of which comprises multiple distinct repeating domains, are found in basal laminae (Figure 20-24): Type IV collagen, trimeric molecules with both rodlike and globular domains that form a two-dimensional network Laminins, a family of multi-adhesive, cross-shaped proteins that form a fibrous two-dimensional network with type IV collagen and that also bind to integrins and other adhesion receptors
Perlecan, a large proteoglycan whose protein contains multiple copies of seven different structural domains (total of 48 domains) and which binds to and cross-links many ECM components and cellsurface molecules Nidogen (also called entactin), a rodlike molecule that cross-links type IV collagen, perlecan, and laminin, which helps incorporate other components into the ECM and also stabilizes basal laminae
FIGURE 20-24 Major protein components of the basal lamina. Type IV collagen and laminin each form two-dimensional networks (see Figures 20-25 and 20-27), which are cross-linked by nidogen/entactin, perlecan, and laminin molecules, and which interact via laminins with the plasma membranes of adjacent cells. Description The basal lamina is composed of a layer of meshed collagen 4 and a layer composed of a mixture of laminin, which itself is composed of perlecan and laminin. Beneath the laminin layer, the plasma membrane is located. In this layer are small short strings of yellow circles, which are in a legend outside the diagram as nidogen slash entactin. Other ECM molecules, such as members of the evolutionarily ancient family of glycoproteins called fibulins, are incorporated into various basal
Laminin, a Multi-Adhesive Matrix Protein, Helps Cross-Link Components of the Basal Lamina
laminae, depending on the tissue and the particular functional requirements of the basal lamina. As depicted in Figure 20-1, one side of the basal lamina is linked to cells by adhesion receptors, including integrins in hemidesmosomes, which bind to laminin in the basal lamina. The other side of the basal lamina is anchored to the adjacent connective tissue by a layer of collagen fibers embedded in a proteoglycan-rich matrix. In stratified squamous epithelia (e.g., skin; see Figure 20-10d), this linkage between the dermis and epidermis is mediated by anchoring fibrils of type VII collagen. Loss-offunction mutations in the gene encoding type VII collagen results in the rare and debilitating blistering disease called dystrophic epidermolysis bullosa. Together, the basal lamina and the anchoring collagen fibrils form the structure called the basement membrane. Laminin, a Multi-Adhesive Matrix Protein, Helps Cross-Link Components of the Basal Lamina Laminin, the principal multi-adhesive matrix protein in basal laminae, is a heterotrimeric protein comprising α, β, and γ chains. At least 16 laminin isoforms in vertebrates are assembled from 5 α, 3 β, and 3 γ chains, with each chain numbered to reflect the chain composition: laminin-111 (α1β1γ1) or laminin-511 (α5β1γ1). Each laminin isoform exhibits a distinctive pattern of tissue- and developmental stage–specific expression. As shown in Figure 20-25, many laminins are large, cross-shaped proteins
(molecular weight of about 820,000), although some are Y- or rod-shaped. Globular domains at the N-terminus of each subunit bind to one another and thus mediate the self-assembly of laminins into mesh-like networks. Five globular LG domains at the C-terminus of the laminin α subunit mediate -dependent binding to cell-surface laminin receptors, including certain integrins (see Table 20-4) as well as sulfated glycolipids, syndecan, and dystroglycan, which will be described further in Section 20.4. Some of these interactions are via negatively charged carbohydrates on the receptors. LG domains are found in a wide variety of other proteins and can mediate binding to steroids and proteins as well as carbohydrates. Laminin is the principal basal laminal ligand of integrins.
FIGURE 20-25 Laminin is a heterotrimeric multi-adhesive matrix protein found in all basal laminae. (a) Schematic model of cross-shaped laminin molecule showing the general shape, location of globular domains, and coiled-coil region in which laminin’s three chains are covalently linked by several disulfide bonds. Different regions of laminin bind to adhesion receptors and various matrix components (indicated by arrows and brackets). Right: Laminins assemble into a lattice via interactions between their N-terminal globular domains. See G. R. Martin and R. Timpl, 1987, Annu. Rev. Cell Biol. 3:57–85; M. Durbeej, 2010, Cell Tissue Res. 339:259–268; and S. Meinen et al., 2007, J. Cell Biol. 176:979–993. (b) Electron micrographs of an intact laminin molecule, showing its characteristic cross shape (left), and the carbohydrate-binding LG domains near the C-terminus (right).
Sheet-Forming Type IV Collagen Is a Major Structural Component of the Basal Lamina
[Left: Republished with permission from Elsevier, from J. Engel et al., 1981, “Shapes, Domain Organizations and Flexibility of Laminin and Fibronectin, Two Multifunctional Proteins of the Extracellular Matrix,” J. Mol. Biol. 150(1):97–120; permission conveyed through Copyright Clearance Center, Inc. Right: Republished with permission from Elsevier, from R. Timpl et al., 2000, “Structure and Function of Laminin LG Modules,” Matrix Biol. 19(4):309–317; permission conveyed through the Copyright Clearance Center, Inc.] Description The illustration labeled (a) shows laminin with three chains, alpha, beta, and gamma. The alpha chain is at the top of a cross-shaped diagram and is labeled: laminins (selfassembly) integrins. The beta chain goes off to the left and is labeled: laminins (selfassembly) collagens. The gamma chain goes off to the right and is also labeled laminins (self-assembly) collagens, but adds a nidogen near the centerline of the cross shape. At the bottom of the cross shape are 5 pink circles labeled: LG domains, bind cellular receptors (integrins, syndecan, sulfated glycolipids, dystroglycan. A white arrow goes from the end of the gamma chain to a diagram that shows the cross shape binding to several cross shapes. Sheet-Forming Type IV Collagen Is a Major Structural Component of the Basal Lamina Type IV collagen is, together with laminin, a principal structural component of all basal laminae and can bind to adhesion receptors, including some integrins. Collagen IV is one of at least 28 types of collagen in humans that participate in the formation of distinct ECMs in various tissues (Table 20-5). There are also at least 20 additional collagen-
like proteins (such as host defense collagens) in the human proteome. Although collagen isoforms differ in certain structural features and in their tissue distribution, all collagens are trimeric proteins made from three polypeptides, each encoded by one of at least 43 genes in humans, usually called collagen α chains. The three α chains in a collagen molecule can be identical (forming a homotrimer) or different (forming a heterotrimer). All or parts of the three-stranded collagen molecule can twist together into a special triple helix called a collagenous triple helix. When there is more than one triple-helical segment, these segments are joined by nonhelical regions of the protein, as we will see shortly for type IV collagen. Within a helical segment, each of the three α chains twists into a left-handed helix, and the three chains then wrap around one another to form a right-handed triple helix (Figure 20-26).
TABLE 20-5 • Selected Collagens Type Molecule Composition Structural Features Representative Tissues Fibrillar Collagens I 300-nm-long fibrils Skin, tendon, bone, ligaments, dentin, interstitial tissues II 300-nm-long fibrils Cartilage, vitreous humor III 300-nm-long fibrils; often Skin, muscle, blood vessels
with type I V 390-nm-long fibrils with globular N-terminal extension; often with type I Cornea, teeth, bone, placenta, skin, smooth muscle Fibril-Associated Collagens VI Lateral association with type I; periodic globular domains Most interstitial tissues IX Lateral association with type II; N-terminal globular domain; bound GAG Cartilage, vitreous humor Sheet-Forming and Anchoring Collagens IV Twodimensional network All basal laminae VII Long fibrils Below basal lamina of the skin
XV Core protein of chondroitin sulfate proteoglycan Widespread; near basal lamina in muscle Transmembrane Collagens
Integral membrane protein Hemidesmosomes in skin Host Defense Collagens Collectins Oligomers of triple helix; lectin domains Blood, alveolar space C1q Oligomers of triple helix Blood (complement) Class A scavenger receptors Homotrimeric membrane proteins Macrophages SOURCE: Data from K. Kühn, 1987, in R. Mayne and R. E. Burgeson, eds., Structure and Function of Collagen Types, Academic Press, p. 2; M. Van Der Rest and R. Garrone, 1991, FASEB J. 5:2814. The collagen triple helix can form because of an unusual abundance of three amino acids in the α chains: glycine, proline, and a modified form of
proline called hydroxyproline (see Figure 2-15). They make up the characteristic repeating sequence motif Gly-X-Y, where X and Y can be any amino acid but are often proline in position X and hydroxyproline in position Y, and less often lysine and hydroxylysine. Glycine is essential because its small side chain, a hydrogen atom, is the only one that can fit into the crowded center of the three-stranded helix (see Figure 20-26b). Hydrogen bonds help hold the three chains together. Although the rigid peptidyl-proline and peptidyl-hydroxyproline linkages are not compatible with formation of a classic single-stranded α helix, they stabilize the distinctive collagenous triple helix. The hydroxyl group in hydroxyproline in the Y position helps hold its ring in a conformation that stabilizes the three-stranded helix.
FIGURE 20-26 The collagen triple helix. (a) Left: Side view of the crystal structure of a polypeptide fragment whose sequence is based on repeating sets of three amino acids, GlyX-Y, characteristic of collagen α chains. Center: Each chain is twisted into a left-handed helix, and three chains wrap around one another to form a right-handed triple helix. Right: A schematic model illustrates the triple-helical nature of the structure and shows the lefthanded twist of the individual collagen α chains (red line). (b) View down the axis of the triple helix. The proton side chains of the glycine residues (orange) point into the very narrow space between the polypeptide chains in the center of the triple helix. In collagen mutations in which other amino acids replace glycine, the proton in glycine is replaced by larger groups that disrupt the packing of the chains and destabilize the triple-helical structure. [Data from R. Z. Kramer et al., 2001, J. Mol. Biol. 311:131, PDB ID 1bkv.]
There are several distinct cell-surface receptors for collagen IV and other types of collagen (other collagens are discussed in the next section). These cell-surface receptors include certain integrins, discoidin domain receptors 1 and 2 (which are tyrosine kinase receptors), glycoprotein VI (on platelets), leukocyte-associated Ig-like receptor-1, members of the mannose receptor family, and a modified form of the protein CD44. They can play critical roles in helping to assemble the ECM and in integrating cellular activity with the ECM. The unique properties of each collagen isoform are due mainly to differences in (1) the number and lengths of the collagenous triple-helical segments; (2) the segments that flank or interrupt the triple-helical segments and that fold into other kinds of three-dimensional structures; and (3) covalent modification of the α chains (e.g., hydroxylation, glycosylation, oxidation, cross-linking). For example, the chains in type IV collagen are designated IVα chains. Mammals express six homologous IVα chains, which assemble into three different heterotrimeric type IV collagens with distinct properties. All subtypes of type IV collagen, however, form a 400-nm-long triple helix (Figure 20-27) that is interrupted about 24 times by nonhelical segments and flanked by a large globular domain at the C-terminus of the chain and a smaller globular domain at the N-terminus. The nonhelical regions introduce flexibility into the molecule. Through both lateral associations and interactions entailing the globular N- and C-termini, type IV collagen molecules assemble into a branching and covalently cross-linked, irregular twodimensional fibrous network that forms a lattice on which, together with the laminin lattice, the basal lamina is built (see Figures 20-24 and 20-27).
Multiple, unusual sulfilimine or thioether bonds between hydroxylysine (or lysine) and methionine residues covalently cross-link some adjacent C-terminal domains of type IV collagen molecules, and thus contribute to the stability of the network.
FIGURE 20-27 Structure and assembly of type IV collagen. (a) Schematic representation of type IV collagen. This 400-nm-long molecule has a small noncollagenous globular domain at the N-terminus and a large globular domain at the C-terminus. The collagenous triple helix is interrupted by nonhelical segments that introduce flexible kinks into the molecule. Lateral interactions between triple-helical segments, as well as head-to-head and tail-to-tail interactions between the globular domains, form dimers, tetramers, and higher order complexes, yielding a sheet-like network. See A. Boutaud et al., 2000, J. Biol. Chem. 275:30716. (b) Electron micrograph of type IV collagen network formed in vitro. The lacy appearance results from the flexibility of the molecule, the side-to-side binding between triple-helical segments (white arrows), and the interactions between C-terminal globular domains (yellow arrows). [Part (b) P. D. Yurchenco and G. C. Ruben, 1987, J. Cell Biol. 105(6):2559–2568, Fig. 1c; https://doi.org/10.1083/jcb.105.6.2559.] Description The illustration labeled (a) shows a collagen triple helix depicted schematically with a blue line. A region of the non-helical protein is above this. The N-terminal and C- terminal globular domains are highlighted as circles at each end. Downward arrows indicate that association can occur by the association of 2 C-terminal head groups (two blue lines joined), forming dimers, or association of four of the non-helical domains, forming tetramers (four blue lines joined at the center). The association of different collagen structures leads to a network structure. In the kidney, a double basal lamina called the glomerular basement membrane separates the epithelium that lines the urinary space from the endothelium that lines the surrounding blood-filled capillaries. Defects in this structure, which is responsible for ultrafiltration of the blood and initial urine formation, can lead to renal failure. For instance, mutations that alter the C-terminal globular domain of certain IVα chains are associated with progressive renal failure as well as sensorineural hearing
Perlecan, a Proteoglycan, Cross-Links Components of the Basal Lamina and Cell-Surface Receptors
loss and ocular abnormalities, a condition known as Alport’s syndrome. In Goodpasture’s syndrome, a relatively rare autoimmune disease, antibodies bind to the α3 chains of type IV collagen found in the glomerular basement membrane and lungs. This binding sets off an immune response that causes cellular damage, resulting in progressive renal failure and pulmonary hemorrhage. Perlecan, a Proteoglycan, Cross-Links Components of the Basal Lamina and Cell-Surface Receptors Perlecan, the major secreted proteoglycan in basal laminae, consists of a large multidomain core protein (∼470 kDa) to which polysaccharides are covalently attached. The core protein is made up of multiple repeats of five distinct domains, including laminin-like LG domains (3 copies), EGFlike domains (12 copies), and Ig domains (22 copies). The many globular repeats give it the appearance of an approximately 200-nm-long string of pearls when visualized by electron microscopy; hence the name perlecan. Perlecan contains three types of covalent polysaccharide chains: N-linked chains (see Chapter 14), O-linked chains, and glycosaminoglycans (GAGs) (O-linked sugars and GAGs are discussed further in Section 20.4). GAGs are long, linear polymers of repeating disaccharides. Glycoproteins containing covalently attached GAG chains are called proteoglycans. Both the protein and the GAG components of perlecan contribute to its ability to incorporate into and define the structure and function of basal laminae. Because its multiple domains and its polysaccharide chains have distinct
binding properties, perlecan binds to dozens of other molecules, including other ECM components (e.g., laminin, nidogen/entactin), cell-surface receptors, and polypeptide growth factors. Simultaneous binding to these molecules results in perlecan-mediated cross-linking. Perlecan can be found in basal laminae and in non–basal laminal ECM. The adhesion receptor dystroglycan can bind perlecan directly, via perlecan’s LG domains, and indirectly, via its binding to laminin. In humans, mutations in the perlecan gene can lead either to dwarfism or to muscle abnormalities, apparently due to dysfunction of the neuromuscular junction that controls muscle firing. KEY CONCEPTS OF SECTION 20.3 The Extracellular Matrix I: The Basal Lamina The matrisome is the ensemble of proteins that compose the ECM itself and associated proteins that covalently modify (e.g., chemically cross-link, phosphorylate, cleave) the ECM. The basal lamina, a thin meshwork of ECM molecules, separates most epithelia and other organized groups of cells from adjacent connective tissue. Together, the basal lamina and the immediately adjacent collagen network form a structure called the basement membrane. Four ECM proteins are found in all basal laminae (see Figure 20-24): laminin (a multiadhesive matrix protein), type IV collagen, perlecan (a proteoglycan), and nidogen/entactin. Adhesion receptors such as integrin anchor cells to the basal lamina, which in turn is connected to other ECM components (see Figure 20-1). Laminin in the basal lamina is the principal ligand of α6β4 integrin (see Table 20-4). Laminin and other multi-adhesive matrix proteins are multidomain molecules that bind multiple adhesion receptors and ECM components. The large, flexible molecules of type IV collagen interact end to end and laterally to form a mesh-like scaffold to which other ECM components and adhesion receptors can bind (see Figures 20-24 and 20-27).
Type IV collagen is a member of the collagen family of proteins, which is distinguished by the presence of repeating tripeptide sequences of Gly-X-Y that give rise to the collagen triple-helical structure (see Figure 20-26). Different collagens are distinguished by the length and chemical modifications of their α chains and by the presence or absence of segments that interrupt or flank their triple-helical regions. Perlecan, a large, multidomain, secreted proteoglycan that is present primarily in basal laminae, binds many ECM components and adhesion receptors. Proteoglycans consist of membrane-associated or secreted core proteins covalently linked to one or more specialized polysaccharide chains called glycosaminoglycans (GAGs).
20.4 The Extracellular Matrix II: Connective Tissue
20.4 The Extracellular Matrix II: Connective Tissue Connective tissue, such as tendon and cartilage, differs from other solid tissues in that, rather than being filled primarily with cells, most of its volume is made up of extracellular matrix packed with insoluble protein fibers (see Figure 20-4). ECM in connective tissue has several key components, some of which are found in other types of tissues as well: Collagens, trimeric molecules that are often bundled together into fibers (fibrillar collagens) Glycosaminoglycans (GAGs), specialized linear polysaccharide chains of specific repeating disaccharides that can be highly hydrated and confer diverse binding and physical properties (e.g., resistance to compression) Proteoglycans, glycoproteins containing one or more covalently bound GAG chains Multi-adhesive proteins, large multidomain proteins often comprising many copies (“repeats”) of a few distinctive domains that bind to and cross-link a variety of adhesion receptors and ECM components Elastin, a protein that forms the amorphous core of elastic fibers Collagen is the most abundant fibrous protein in connective tissue. Rubberlike elastin fibers, which can be stretched and relaxed, are also
Fibrillar Collagens Are the Major Fibrous Proteins in the ECM of Connective Tissues
present in deformable sites (e.g., skin, tendons, heart). The fibronectins, a family of multi-adhesive matrix proteins, form their own distinct fibrils in the ECM of most connective tissues. Although several types of cells are found in connective tissues, the various ECM components are produced largely by cells called fibroblasts. In this section, we explore the structure and function of the various ECM components in connective tissue, and we see how the ECM is degraded and remodeled by a variety of specialized proteases. Fibrillar Collagens Are the Major Fibrous Proteins in the ECM of Connective Tissues About 80–90 percent of the collagen in the body consists of fibrillar collagens (types I, II, and III), located primarily in connective tissues (see
Table 20-5). Because of its abundance in tendon-rich tissue such as rat tail, type I collagen is easy to isolate and was the first collagen to be characterized. Its fundamental structural unit is a long (300 nm), thin (1.5 nm diameter) triple helix (see Figure 20-26) consisting of two α1(I) chains and one α2(I) chain, each 1050 amino acids in length. The triple-stranded molecules pack tightly together and wrap around one another, forming microfibrils that associate into higher order polymers called collagen fibrils, which in turn often aggregate into larger bundles called collagen fibers (Figure 20-28).
FIGURE 20-28 Biosynthesis of fibrillar collagens. Step 1 : Procollagen α chains are synthesized on ribosomes associated with the endoplasmic reticulum (rough ER), and in the ER, asparagine-linked oligosaccharides are added to the C-terminal propeptide. Step 2 : Propeptides associate to form trimers and are covalently linked by disulfide bonds, and selected residues in the Gly-X-Y triplet repeats are covalently modified [certain prolines and lysines are hydroxylated; galactose or galactose-glucose (hexagons) are attached to some hydroxylysines; prolines are cis → trans isomerized]. Step 3 : The modifications facilitate zipper-like formation and stabilization of triple helices, and binding by the ER chaperone protein Hsp47, which may stabilize the helices or prevent premature aggregation of the trimers, or both. Steps 4 and 5 : The folded procollagens are packaged into large transport vesicles with the help of specialized membrane proteins in the ER (not shown) and then transported to and through the Golgi complex, where some lateral association into small bundles takes place. The chains are then secreted (step 6 ), the N- and C-terminal propeptides are removed (step 7 ), and the trimers assemble into fibrils and are covalently cross-linked (step 8 ). The 67 nm staggering of the trimers gives the fibrils a striated appearance in electron micrographs (inset). Step 9 : The fibrils can assemble into larger
Fibrillar Collagen Is Secreted and Assembled into Fibrils Outside the Cell
and larger bundles, some of which form the tendons that attach muscle to bone. See A. V. Persikov and B. Brodsky, 2002, Proc. Nat’l. Acad. Sci. USA 99:1101–1103. [Electron micrograph republished with permission from John Wiley & Sons, Inc., from J. Gross, 1953, “Evaluation of Structural and Chemical Changes in Connective Tissue,” Ann. NY Acad. Sci. 56(4):674–683; permission conveyed through Copyright Clearance Center, Inc.] Description In this schematic, the top left shows a cell inside the cytosol. Inside the cell, steps 1-5 a blue circle represents the ribosome associated with the rough endoplasmic reticulum. At step 3, a braid of 3 blue lines is with circles at each end and a green circle labeled H s p 47 attached toward the right end. The Golgi complex is at step 5 represented as light blue lines with blue circles on each end. Step 6 is the packages from the Golgi that enter the extracellular space and is arranged in rows to build collagen molecules. The molecules make fibrils, which look like blue wires that are gathered into blue cables in the last step to the right of the cell diagram. A micrograph is added at the bottom left that shows the cables of collagen as light gray lines. Classes of collagen that are less abundant, but nevertheless important, include fibril-associated collagens, which link the fibrillar collagens to one another or to other ECM components; sheet-forming and anchoring collagens, which form two-dimensional networks in basal laminae (type IV) and connect the basal lamina in skin to the underlying connective tissue (type VII); transmembrane collagens, which function as adhesion receptors; and host defense collagens, which help the body recognize and eliminate pathogens. Interestingly, several collagens (e.g., types IX, XVIII, and XV) are also proteoglycans with covalently attached GAGs (see Table 20-5).
Fibrillar Collagen Is Secreted and Assembled into Fibrils Outside the Cell Fibrillar collagens are secreted proteins, produced primarily by fibroblasts in the ECM. Collagen biosynthesis and secretion follow the normal pathway for a secreted protein, described in detail in Chapters 13 and 14. The collagen α chains are synthesized as longer precursors, called pro-α chains, by ribosomes attached to the endoplasmic reticulum (ER). The pro-α chains undergo a series of covalent modifications and fold into triple-helical procollagen molecules before their transport from the ER to the Golgi complex and subsequent release from cells (see Figure 20-28). Specialized membrane proteins in the ER allow the packaging of the very large collagen molecules into large transport vesicles for transfer to the Golgi complex. After the secretion of procollagen from the cell, extracellular peptidases remove the N-terminal and C-terminal propeptides. The resulting molecules, which consist almost entirely of a triple-stranded helix because of long stretches of the characteristic collagen repeating sequence motif Gly-X-Y, associate laterally to generate fibrils with a diameter of 50–200 nm. In fibrils, adjacent collagen molecules are displaced from one another by 67 nm, about one-quarter of their length. This staggered array produces a striated effect that can be seen in both light and electron microscopic images of collagen fibrils (see Figure 20-28, inset). The unique properties of the fibrillar collagens are mainly due to the formation of fibrils.
Short segments at either end of the fibrillar collagen α chains that are not composed of the repeating sequence motif Gly-X-Y, and thus are not triple-helical, are of particular importance in the formation of collagen fibrils. Lysine and hydroxylysine side chains in these segments are covalently modified by extracellular lysyl oxidases to form aldehydes in place of the amine group at the end of the side chain. These reactive aldehyde groups form covalent cross-links with lysine, hydroxylysine, and histidine residues in adjacent molecules. The cross-links stabilize the sideby-side packing of collagen molecules and generate a very strong fibril. The removal of the terminal propeptides and covalent cross-linking take place in the extracellular space to prevent the potentially catastrophic assembly of large fibrils within the cell. The post-translational modifications of pro-α chains are crucial for the formation of mature collagen molecules and their assembly into fibrils. Defects in these modifications have serious consequences, which ancient mariners frequently experienced. For example, ascorbic acid (vitamin C) is an essential cofactor for the hydroxylases responsible for adding hydroxyl groups to proline and lysine residues in pro-α chains. In cells deprived of ascorbate, as in the disease scurvy, the pro-α chains are not hydroxylated sufficiently to form stable triple-helical procollagen at normal body temperature, and the procollagen that forms cannot assemble into normal fibrils. Without the structural support of collagen, blood vessels, tendons, and skin become fragile. Fresh fruit in the diet can supply sufficient vitamin C to support the formation of normal collagen. Historically, British sailors were provided with limes to prevent scurvy,
Type I and II Collagens Associate with Nonfibrillar Collagens to Form Diverse Structures
leading to their being called “limeys.” Mutations in lysyl hydroxylase genes can also cause connective-tissue defects. Type I and II Collagens Associate with Nonfibrillar Collagens to Form Diverse Structures Collagens differ in the structures of the fibers they form and in how these fibers are organized into networks. Of the predominant types of collagen found in connective tissues, type I collagen forms long fibers, whereas networks of type II collagen are more mesh-like. In tendons, for instance, the long type I collagen fibers connect muscles to bones and must withstand enormous forces. Because type I collagen fibers have great tensile strength, tendons usually can be stretched without being broken. Indeed, gram for gram, type I collagen is stronger than steel. Two quantitatively minor fibrillar collagens, type V and type XI, co-assemble into fibers with type I collagen, thereby regulating the structures and properties of the fibers. Incorporation of type V collagen, for example, results in smaller-diameter fibers. Type I collagen fibrils are also used as the reinforcing rods in the construction of bone. Bones and teeth are hard and strong because they contain large amounts of dahllite, a crystalline calcium- and phosphatecontaining mineral. Most bones are about 70 percent mineral and 30 percent protein, the vast majority of which is type I collagen. Bones form
when certain cells (chondrocytes and osteoblasts) secrete collagen fibrils that are then mineralized by deposition of small dahllite crystals. In many connective tissues, particularly skeletal muscle, proteoglycans and a fibril-associated collagen called type VI collagen are noncovalently bound to the sides of type I fibrils and may bind the fibrils together to form thicker collagen fibers (Figure 20-29a). Type VI collagen is unusual in that the molecule consists of a relatively short triple helix with globular domains at both ends. The lateral association of two type VI monomers generates an antiparallel dimer. The end-to-end association of these dimers through their globular domains forms type VI microfibrils. These microfibrils have a beads-on-a-string appearance, with about 60-nm-long triple-helical regions separated by 40-nm-long globular domains.
FIGURE 20-29 Interactions of fibrillar collagens with fibril-associated collagens. (a) In tendons, type I fibrils are all oriented in the direction of the stress applied to the tendon. Proteoglycans and type VI collagen bind noncovalently to type I fibrils, coating the surface. The microfibrils of type VI collagen, which contain globular and triple-helical segments, bind to type I fibrils and link them together into thicker fibers. See R. R. Bruns et al., 1986, J. Cell Biol. 103:393. (b) In cartilage, type IX collagen molecules are covalently bound at regular intervals along type II fibrils. A chondroitin sulfate chain, covalently linked to the α2(IX) chain at the flexible kink, projects outward from the fibril, as does the globular N-terminal region. See L. M. Shaw and B. R. Olsen, 1991, Trends Biochem. Sci. 16:191. Description
The illustration labeled (a) shows two stacks of blue disks representing the type 1 collagen fibrils with orange short tubes going along between the stacks and labeled Type 6 collagen. Short black line segments are in the stacks and labeled proteoglycan. The illustration labeled (b) shows one stack of blue disks labeled Type 2 collagen fibril. On the front of this stake are two strings of orange cylinders with black lines between labeled kink. A red thread comes from between two orange cylinders and is labeled chondroitin sulfate. The orange cylinders are labeled Type 9 collagen. The fibrils of type II collagen, the major collagen in cartilage, are smaller in diameter than type I fibrils and are oriented randomly in a viscous proteoglycan matrix. The rigid collagen fibrils impart strength to the matrix and allow it to resist large deformations. Type II fibrils are crosslinked to matrix proteoglycans by type IX collagen, another fibrilassociated collagen. Type IX collagen and several related types have two or three triple-helical segments connected by flexible kinks and a globular N-terminal segment (Figure 20-29b). The globular N-terminal segment of type IX collagen extends from the type II fibril at the end of one of its helical segments, as does a chondroitin sulfate GAG chain (chondroitin sulfate is described below) that is sometimes linked to one of the type IX chains. These protruding nonhelical structures are thought to anchor the type II fibril to proteoglycans and other components of the matrix. The interrupted triple-helical structure of type IX and related collagens prevents them from assembling into fibrils, although they can associate with fibrils formed from other collagen types and form covalent crosslinks to them.
Proteoglycans and Their Constituent GAGs Play Diverse Roles in the ECM
Mutations affecting type I collagen and its associated proteins cause a variety of human diseases. Certain mutations in the genes encoding the type I collagen α1(I) or α2(I) chains lead to osteogenesis imperfecta, or brittle-bone disease. Because every third position in a collagen α chain must be a glycine for the triple helix to form (see Figure 20-26), mutations of glycine to almost any other amino acid are deleterious, resulting in poorly formed and unstable helices. A defect in only one of the three α chains in a collagen molecule can disrupt the whole molecule’s triplehelical structure and function. A mutation in a single copy (allele) of either the α1(I) gene or the α2(I) gene, both located on autosomes, can cause this disorder. Thus it normally shows autosomal dominant inheritance (see Chapter 6). Absence or malfunctioning of fibril-associated collagen in muscle tissue due to mutations in the type VI collagen genes cause dominant or recessive congenital muscular dystrophies with generalized muscle weakness, respiratory insufficiency, muscle wasting, and muscle-related joint abnormalities. Skin abnormalities have also been reported with type VI collagen disease. Proteoglycans and Their Constituent GAGs Play Diverse Roles in the ECM As we saw with perlecan in the basal lamina, proteoglycans play an important role in cell-matrix adhesion. Proteoglycans are a subset of
secreted or cell-surface glycoproteins containing covalently linked, specialized polysaccharide chains called glycosaminoglycans (GAGs). GAGs are long linear polymers of specific repeating disaccharides. Usually one sugar is either a uronic acid (D-glucuronic acid or L-iduronic acid) or D-galactose; the other sugar is N-acetylglucosamine or N-acetylgalactosamine (Figure 20-30). One or both of the sugars contain at least one anionic group (carboxylate or sulfate). Thus each GAG chain bears many negative charges. GAGs are classified into several major types based on the nature of the repeating disaccharide unit: heparan sulfate, chondroitin sulfate, dermatan sulfate, keratan sulfate, and hyaluronan (see
Figure 20-30). A hypersulfated form of heparan sulfate called heparin, produced mostly by mast cells, plays a key role in allergic reactions. It is also used medically as an anticlotting drug because of its ability to activate a natural clotting inhibitor called antithrombin III.
FIGURE 20-30 The repeating disaccharides of glycosaminoglycans (GAGs). Each of the four classes of GAGs is formed by polymerization of monomeric units into repeats of a particular disaccharide and subsequent modifications, including addition of sulfate groups and inversion (epimerization) of the carboxyl group on carbon 5 of D-glucuronic acid to yield L-iduronic acid. The squiggly lines represent covalent bonds that are oriented either above (D-glucuronic acid) or below (L-iduronic acid) the ring. Heparin is generated by hypersulfation of heparan sulfate, whereas hyaluronan is not sulfated. Description The illustration labeled (a) shows the chemical structure of hyaluronan, with twentyfive thousand dimers composed of D-glucuronic acid and N-acetyl-D-glucosamine. The illustration labeled (b) shows the chemical structure of chondroitin sulfate, with 250 dimers composed of D-glucuronic acid and N-acetyl D-galactosamine or L- iduronic acid and N-acetyl-D-galactosamine. The illustration labeled (c) shows the chemical structure of heparin sulfate, with two hundred dimers composed of D-glucuronic or L-iduronic acid and N-acetyl- or N- sulfo-D-glucosamine. The illustration labeled (d) shows the chemical structure of keratan sulfate, with 20 to 40 dimers composed of D-galactose and N-acetyl-d-glucosamine. With the exception of hyaluronan, which is discussed below, all the major GAGs occur naturally as components of proteoglycans. Like other secreted and transmembrane glycoproteins, proteoglycan core proteins are synthesized in the endoplasmic reticulum, and the GAG chains are assembled on and covalently attached to these cores in the Golgi complex. To generate heparan or chondroitin sulfate chains, a three-sugar linker is first attached to the hydroxyl side chains of certain serine residues in a
core protein (Figure 20-31a); thus these GAGs are O-linked oligosaccharides, several examples of which are shown in Figure 20-31.
FIGURE 20-31 Hydroxyl O-linked oligosaccharides. (a) Synthesis of a GAG, in this case chondroitin sulfate, is initiated by transfer of a xylose (Xyl) residue to a serine residue in the core protein, most likely in the Golgi complex, followed by sequential addition of two galactose (Gal) residues. Glucuronic acid (GlcUA) and N-acetylgalactosamine (GalNAc) residues are then added sequentially to these linking sugars and some of the GalNAc monomers are sulfated, forming the chondroitin sulfate chain. Heparan sulfate chains are connected to core proteins by the same three-sugar linker. Keratan sulfate GAGs are covalently attached to proteins via N-linked rather than O-linked connections. (b) Mucinlike O-linked chains are covalently bound to glycoproteins via an N-acetylgalactosamine (GalNAc). (c) In the integral membrane protein α-dystroglycan on cell surfaces, a complex polysaccharide is attached to hydroxyl side chains via an O-mannose (O-Man). A polymer of the GlcUA-Xyl disaccharide called matriglycan is attached to a phosphorylated O-linked mannose via an oligosaccharide containing N-acetylglucosamine (GlcNAc), ribitol 5-phosphate (Ribito5P), Xyl, and GlcUA. Matriglycan provides a binding site for ECM molecules, such as laminin and perlecan (described in detail in Section 20.5). O-linked oligosaccharides are attached to the hydroxyl side chains of specific serine and theonine residues in a variety of secreted proteins and the exoplasmic (extracellular) domains of some membrane proteins. For example, mucin-like O-linked chains are covalently bound to glycoproteins via an N-acetylgalactosamine (GalNAc) monosaccharide to which are covalently attached a variety of other sugars, often including sialic acid (SA; Figure 20-31b). The high concentration of long O-linked chains on mucin glycoproteins on the apical plasma membranes of epithelial cells apparently contributes to the bending of the membranes and the formation of microvilli (see Figures 20-11, 17-1, and 17-2). The only O-linked glycoproteins in baker’s yeast are mannose-linked (O-Man). In other eukaryotes (not in nematode worms or plants), certain specialized O-linked sugars are bound to proteins via mannose monosaccharides.
Some proteins, including classical cadherins and protocadherins (Section 20.2), have relatively short O-mannose linked oligosaccharides. Others have longer oligosaccharides attached via the O-mannose, such as that attached to the cell-surface protein dystroglycan (Figure 20-31c; described in detail in Section 20.5). In contrast to protein-linked heparan or chondroitin sulfate chains, the linkers for the addition of keratan sulfate chains are oligosaccharide chains attached to asparagine residues; such N-linked oligosaccharides are present in many glycoproteins (see Chapters 13 and 14), although only a subset carry GAG chains. All GAG chains are elongated by the alternating addition of sugar monomers to form the disaccharide repeats characteristic of a particular GAG; the chains are often modified subsequently by the covalent linkage of small molecules such as sulfate. The mechanisms responsible for determining which proteins are modified with GAGs, the sequence of disaccharides to be added, the sites to be sulfated, and the lengths of the GAG chains are unknown. The ratio of polysaccharide to protein in all proteoglycans is much higher than that in most other glycoproteins. Function of GAG Chain Modifications Similar to the sequence of amino acids in proteins, the arrangement of the sugar residues in GAG chains and the modification of specific sugars in those chains can determine their function and that of the proteoglycans that contain them. For example, groupings of certain modified sugars in the GAG chains of heparin sulfate proteoglycans can control the binding
of growth factors to certain receptors or the activities of proteins in the blood-clotting cascade. In the past, the chemical and structural complexity of proteoglycans posed a daunting barrier to analyzing and understanding their structures and their many diverse functions. In recent years, investigators employing classic and state-of-the-art biochemical techniques, mass spectrometry, and genetics have begun to elucidate the detailed structures and functions of these ubiquitous ECM molecules. The results of ongoing studies suggest that sets of sugar-residue sequences containing common modifications, rather than single unique sequences, are responsible for specifying distinct GAG functions. A case in point is a set of five-residue (pentasaccharide) sequences found in a subset of heparin GAGs that controls the activity of antithrombin III (ATIII), an inhibitor of the key blood-clotting protease thrombin. When these pentasaccharide sequences in heparin are sulfated at two specific positions (Figure 20-32), heparin can activate ATIII, thereby inhibiting clot formation. Several other sulfates can be present in the active pentasaccharide in various combinations, but they are not essential for the anticlotting activity of heparin. The rationale for generating sets of similar active sequences rather than a single, unique sequence is not well understood.
FIGURE 20-32 The pentasaccharide GAG sequence that regulates the activity of antithrombin III (ATIII). Sets of modified five-residue sequences in the much longer GAG called heparin with the composition shown here bind to ATIII and activate it, thereby inhibiting blood clotting. The sulfate groups in red type are essential for this heparin function; the modifications in blue type may be present but are not essential. Other sets of modified GAG sequences are thought to regulate the activity of other target proteins. Diversity of Proteoglycans The proteoglycans constitute a remarkably diverse group of molecules that are abundant in the ECM of all animal tissues and are also expressed on the cell surface. For example, of the five major classes of heparan sulfate proteoglycans, three are located in the ECM (perlecan, agrin, and type XVIII collagen) and two are cell-surface proteins. The latter include integral membrane proteins (syndecans) and GPI-anchored proteins (glypicans); the GAG chains in both types of cell-surface proteoglycans extend into the extracellular space. The sequences and lengths of proteoglycan core proteins vary considerably, and the number of attached GAG chains ranges from just a few to more than 100. Moreover, a core protein is often linked to two different types of GAG chains, generating a
hybrid proteoglycan. The basal laminal proteoglycan perlecan is primarily a heparan sulfate proteoglycan with three to four GAG chains, although it can sometimes have a bound chondroitin sulfate chain. Additional diversity in proteoglycans occurs because the numbers, compositions, and sequences of the GAG chains attached to otherwise identical core proteins can differ considerably. Laboratory generation and analysis of mutants with defects in proteoglycan production in Drosophila, C. elegans, and mice have clearly shown that proteoglycans play critical roles in development; for example, as participants in various signaling pathways (see Chapter 16 for examples in the TGF-β and Wnt pathways). Syndecans are cell-surface proteoglycans expressed by epithelial and nonepithelial cells that bind to collagens and multi-adhesive matrix proteins such as fibronectin, anchoring cells to the ECM. Like that of many integral membrane proteins, the cytosolic domain of syndecan interacts with the actin cytoskeleton and in some cases with intracellular regulatory molecules. In addition, cell-surface proteoglycans such as syndecan bind many protein growth factors and other external signaling molecules, thereby helping to regulate cellular metabolism and function. For instance, syndecans in the hypothalamic region of the brain modulate feeding behavior in response to food deprivation. They do so by participating in the binding to cell-surface receptors of antisatiety peptides that help control feeding behavior. In the fed state, the syndecan extracellular domain decorated with heparan sulfate GAG chains is released from the cell surface by proteolysis, thus suppressing the activity of the antisatiety peptides and feeding behavior. In mice engineered to overexpress the syndecan-1 gene in the hypothalamic region of the brain
Hyaluronan Resists Compression, Facilitates Cell Migration, and Gives Cartilage Its Gel-Like Properties
and other tissues, normal control of feeding by antisatiety peptides is disrupted, and the animals overeat and become obese. Hyaluronan Resists Compression, Facilitates Cell Migration, and Gives Cartilage Its Gel-Like Properties Hyaluronan, also called hyaluronic acid (HA) or hyaluronate, is a nonsulfated GAG (see Figure 20-30a) made by a plasma-membrane-bound enzyme called HA synthase and is secreted directly into the extracellular space as it is synthesized. (A similar approach is used by plant cells to make the ECM component cellulose.) Hyaluronan is a major component of the ECM that surrounds migrating and proliferating cells, particularly in embryonic tissues. In addition, it forms the backbone of complex proteoglycan aggregates found in many ECMs, particularly cartilage. Because of its remarkable physical properties, hyaluronan imparts stiffness and resilience as well as a lubricating quality to many types of connective tissue such as joints. Hyaluronan molecules range in length from a few disaccharide repeats to about 25,000. The typical hyaluronan in joints such as the elbow has 10,000 repeats for a total mass of Da and a length of 10 μm (about the diameter of a small cell). Individual segments of a hyaluronan molecule fold into a rodlike conformation because of the β glycosidic linkages between the sugars and extensive intrachain hydrogen bonding. Mutual repulsion between negatively charged carboxylate groups that
protrude outward at regular intervals also contributes to these locally rigid structures. Overall, however, hyaluronan is not a long, rigid rod like fibrillar collagen; rather, it is very flexible in solution, bending and twisting into many conformations, forming a random coil. Because of the large number of anionic residues on its surface, the typical hyaluronan molecule binds a large amount of water and behaves as if it were a large hydrated sphere with a diameter of about 500 nm. As the concentration of hyaluronan increases, the long chains begin to entangle, forming a viscous gel. Even at low concentrations, hyaluronan forms a hydrated gel; when placed in a confining space, such as that between two cells, the long hyaluronan molecules tend to push outward. This outward pushing creates a swelling, or turgor pressure, within the extracellular space. In addition, the binding of cations by carboxylate groups on the surface of hyaluronan increases the concentration of ions and thus the osmotic pressure in the gel. As a result, large amounts of water are taken up, contributing to the turgor pressure. These swelling forces give connective tissues their ability to resist compression forces, in contrast to collagen fibers, which are best able to resist stretching forces. Other highly charged GAG chains are similarly hydrated. Hyaluronan is bound to the surface of many migrating cells by a number of adhesion receptors, such as the receptor called CD44, which contains hyaluronan-binding domains, each with a similar three-dimensional conformation. Because of its loose, hydrated, porous nature, the hyaluronan coating bound to cells appears to keep them apart from one another, giving them the freedom to move about and proliferate. The
cessation of cell movement and the initiation of cell-cell attachments are frequently correlated with a decrease in hyaluronan, a decrease in hyaluronan receptors, and an increase in the extracellular enzyme hyaluronidase, which degrades hyaluronan in the matrix. These alterations of hyaluronan are particularly important during the many cell migrations that facilitate differentiation and in the release of a mammalian egg cell from its surrounding cells after ovulation. The predominant proteoglycan in cartilage, called aggrecan, assembles with hyaluronan into very large aggregates, illustrative of the complex structures that proteoglycans sometimes form. The backbone of this proteoglycan aggregate is a long molecule of hyaluronan to which multiple aggrecan molecules are bound tightly but noncovalently (Figure 20-33). A single hyaluronan-aggrecan aggregate, one of the largest macromolecular complexes known, can be more than 4 μm long and have a volume larger than that of a bacterial cell. These aggregates give cartilage its unique gel-like properties and its resistance to deformation, essential for distributing the load in weight-bearing joints.
FIGURE 20-33 Structure of proteoglycan aggregate from cartilage. (a) Electron micrograph of an aggrecan aggregate from fetal bovine epiphyseal cartilage (artificially colored). Aggrecan core proteins are bound at ∼40-nm intervals to a molecule of hyaluronan. (b) Schematic representation of an aggrecan monomer bound to hyaluronan. In aggrecan, both keratan sulfate and chondroitin sulfate chains are attached to the core protein. The N-terminal domain of the core protein binds noncovalently to a hyaluronan molecule. Binding is facilitated by a link protein, which binds to both the hyaluronan molecule and the aggrecan core protein. Each aggrecan core protein has 127 Ser-Gly sequences at which GAG chains can be added. The molecular weight of an aggrecan monomer averages . The entire aggregate, which may contain upward of 100 aggrecan monomers, has a molecular weight in excess of and is about as large as the bacterium Escherichia coli. [Part (a) republished with permission from Elsevier, from J. A. Buckwalter and L. Rosenberg, 1983, “Structural Changes During Development in Bovine Fetal Epiphyseal Cartilage,” Collagen Rel. Res. 3(6):489–504; permission conveyed through Copyright Clearance Center, Inc.] Description The micrograph labeled (a) shows a horizontal squiggly yellow line labeled hyaluronan molecule. From this molecule, all along both sides and around the end are threads that start green and end red. The green part is labeled keratan sulfate and the red part is labeled chondroitin sulfate. The illustration labeled (b) takes one of the green and red threads and makes a schematic diagram of it using lines to represent the parts. There is a yellow tube at the top to represent the hyaluronan molecule. A gray circle attached to it below is labeled link protein. There is a central line attached to the hyaluronan. One end of the central line near hyaluronan is labeled N-terminal hyaluronan-binding domain and the other end is labeled aggrecan core protein. The green and red lines are attached along the sides with linking sugars. The aggrecan core protein (∼250,000 MW) has one N-terminal globular domain that binds with high affinity to a specific decasaccharide sequence within hyaluronan. This specific sequence is generated by covalent
Fibronectins Connect Cells and ECM, Influencing Cell Shape, Differentiation, and Movement
modification of some of the repeating disaccharides in the hyaluronan chain. The interaction between aggrecan and hyaluronan is facilitated by a link protein that binds to both the aggrecan core protein and hyaluronan (Figure 20-33b). Aggrecan and the link protein have in common a link domain, about 100 amino acids long, that is found in numerous ECM and cell-surface hyaluronan-binding proteins in both cartilaginous and noncartilaginous tissues. These proteins almost certainly arose in the course of evolution from a single ancestral gene that encoded just this domain. Fibronectins Connect Cells and ECM, Influencing Cell Shape, Differentiation, and Movement Many different cell types synthesize fibronectin, an abundant multiadhesive matrix protein found in all vertebrates. The discovery that fibronectin functions as an adhesion molecule stemmed from observations that it is present on the surfaces of normal fibroblasts, which adhere tightly to petri dishes in laboratory experiments, but is absent from the surfaces of tumorigenic (i.e., cancerous) cells, which adhere weakly. The 20 or so isoforms of fibronectin are generated by alternative splicing of the RNA transcript produced from a single gene (see Figure 5-28). Fibronectins are essential for the migration and differentiation of many cell types in embryogenesis. These proteins are also important for wound healing because they promote blood clotting and facilitate the migration of macrophages and other immune-system cells into the affected area.
Fibronectins help attach cells to the ECM by binding to other ECM components, particularly fibrillar collagens and heparan sulfate proteoglycans, and to adhesion receptors such as integrins (see Figure 202). Through their interactions with adhesion receptors, fibronectins influence the shape and movement of cells and the organization of the cytoskeleton. Conversely, by regulating their receptor-mediated attachments to fibronectin and other ECM components, cells can sculpt the immediate ECM environment to suit their needs. Fibronectins are dimers of two similar polypeptides linked at their C-termini by two disulfide bonds; each chain is about 60–70 nm long and 2– 3 nm thick. Partial digestion of fibronectin with small amounts of proteases and analysis of the fragments showed that each chain comprises several functional regions with different ligand-binding specificities (Figure 20-34a). Each region, in turn, contains multiple copies of certain domain-encoding sequences that can be classified into one of three types. These domains are designated fibronectin type I, II, and III repeats, on the basis of similarities in amino acid sequence, although the sequences of any two repeats of a given type are not identical. These linked repeats give the molecule the appearance of beads on a string. The combination of the different repeats composing the regions confers on fibronectin its ability to bind multiple ligands.
FIGURE 20-34 Organization of fibronectin and its binding to integrin. (a) Scale model of fibronectin is shown docked by two type III repeats to the extracellular domains of integrin. Only one of the two similar chains, which are linked by disulfide bonds near their C-termini, in the dimeric fibronectin molecule is shown. Each chain contains about 2446 amino acids and is composed of three types of repeating amino acid sequences (type I, II, or III repeats) or domains. The EIIIA, EIIIB — both type III repeats — and IIICS domain are variably spliced into the structure at locations indicated by arrows. Circulating fibronectin lacks EIIIA, EIIIB, or both. At least five different sequences may be present in the IIICS region as a result of alternative splicing (see Figure 5-28). Each chain contains several multi-repeat-containing regions, some of which contain specific binding sites for heparan sulfate, fibrin (a major constituent of blood clots), collagen, and cell-surface integrins. The integrin-binding domain is also known as the cell-binding domain. Structures of fibronectin’s domains were determined from fragments of the molecule. (b) A highresolution structure shows that the RGD sequence motif extends outward in a loop from its compact type III domain on the same side of fibronectin as the synergy region, which also contributes to high-affinity binding to integrins. [Data from D. J. Leahy, I. Aukhil, and H. P. Erickson, 1996, Cell 84:155, PDB ID 1fnf.] Description The illustration labeled (a) shows fibronectin which comprises type 1 repeats at the N-terminal end, where fibrin and heparan sulfate can bind, followed by type 2 repeats, where collagen can bind, followed by type three repeats. In the long region composed of type three repeats, there are several cell-binding regions. E 3 B, R G D, where integrin binds, and E 3 A. This section is followed by further binding sites for heparan sulfate and three C S. At the c-terminal, there are further type 1 repeats followed by disulfide bonds to other fibronectin fibers.
One of the type III repeats in the cell-binding region of fibronectin mediates binding to certain integrins. The results of studies with synthetic peptides corresponding to parts of this repeat identified the tripeptide sequence Arg-Gly-Asp, called the RGD motif, as the minimal sequence within this repeat required for recognition by those integrins. In one study, heptapeptides with and without the RGD motif were tested for their ability to mediate the adhesion of rat kidney cells to a culture dish. The results showed that heptapeptides containing the RGD motif mimicked intact fibronectin’s ability to stimulate integrin-mediated adhesion, whereas variant heptapeptides lacking this sequence were ineffective (Figure 2035).
EXPERIMENTAL FIGURE 20-35 A specific tripeptide sequence (RGD) in the cellbinding region of fibronectin is required for cell adhesion. The cell-binding region of fibronectin contains an integrin-binding hexapeptide sequence, GRGDSP in the single-letter amino acid code. Together with an additional C-terminal cysteine (C) residue, this heptapeptide and several variants were synthesized chemically. Different concentrations of each synthetic peptide were added to polystyrene dishes that had the protein immunoglobulin G (IgG) firmly attached to their surfaces; the peptides were then chemically cross-linked to the IgG. Subsequently, cultured normal rat kidney cells were added to the dishes and incubated for 30 minutes to allow adhesion. After the unbound cells were washed away, the relative amounts of cells that had adhered firmly were determined by staining the bound cells with a dye and measuring the intensity of the staining with a
spectrophotometer. The results shown here indicate that cell adhesion increased above the background level with increasing peptide concentration for those peptides containing the RGD motif, but not for the variants lacking this sequence (modification underlined). [Data from M. D. Pierschbacher and E. Ruoslahti, 1984, Proc. Nat’l. Acad. Sci. USA 81:5985.] Description In the graph, the vertical axis plots the relative amounts of bound cells by stain intensity ranging from 0 to 1.4 in increments of 0.2. The horizontal axis plots peptide concentration in nanomoles per milliliter ranging from 0 to ten thousand in increments of 9, 90, and 900. Curves for four proteins containing R G D sequences are sigmoidal. Curves of proteins without R G D sequence show no cell binding. A three-dimensional model of fibronectin binding to integrin, based on partial structures of both fibronectin and integrin, has been assembled. In a high-resolution structure of the integrin-binding fibronectin type III repeat and its neighboring type III domain, the RGD motif is at the apex of a loop that protrudes outward from the molecule, in a position facilitating binding to integrins (Figure 20-34b). Although the RGD motif is required for binding to several different integrins, its affinity for integrins is substantially less than that of intact fibronectin or of the entire cellbinding region in fibronectin. Thus structural features near the RGD motif in fibronectins (e.g., parts of adjacent repeats, such as the synergy region; see Figure 20-34b) and in other RGD-containing proteins must enhance their binding to certain integrins. Moreover, the simple, soluble dimeric forms of fibronectin produced by the liver or by fibroblasts are initially in a nonfunctional conformation that binds poorly to integrins because the RGD motif is not readily accessible. The adsorption of fibronectin onto a
collagen matrix or basal lamina — or, experimentally, to a plastic tissue culture dish — results in a conformational change that enhances the ability of fibronectin to bind to cells. Possibly, this conformational change increases the accessibility of the RGD motif for integrin binding. Microscopy and other experimental approaches (e.g., biochemical-binding experiments) have demonstrated the role of integrins in cross-linking fibronectin and other ECM components to the cytoskeleton. For example, the colocalization of cytoskeletal actin filaments and integrins within cells can be visualized by fluorescence microscopy (Figure 20-36a). The binding of cell-surface integrins to fibronectin in the ECM induces the actin cytoskeleton–dependent movement of some integrin molecules in the plane of the plasma membrane. The ensuing mechanical tension due to the relative movement of different integrins bound to a single fibronectin dimer stretches the fibronectin (see Figure 20-9), a mechanosensor, and promotes self-association of fibronectins into multimeric fibrils.
EXPERIMENTAL FIGURE 20-36 Integrins mediate linkage between fibronectin in the ECM and the cytoskeleton. (a) Immunofluorescent micrograph of a fixed cultured fibroblast showing colocalization of the α5β1 integrin (green) and actin-containing stress fibers (red). The cell was incubated with two types of monoclonal antibodies: an integrin-
specific antibody linked to a green-fluorescing dye and an actin-specific antibody linked to a red-fluorescing dye. Stress fibers are long bundles of actin microfilaments that radiate inward from points where the cell contacts a substratum. At the distal ends of these fibers, near the plasma membrane, the coincidence of actin and fibronectin-binding integrin produces a yellow fluorescence. (b) Electron micrograph of the junction of fibronectin and actin fibers in a cultured fibroblast. Individual actin-containing 7 nm microfilaments, components of a stress fiber, end at the obliquely sectioned cell membrane. The microfilaments appear aligned with and in close proximity to the thicker, densely stained fibronectin fibrils on the outside of the cell. [Part (a) J. L. Duband et al., 1988, J. Cell Biol. 107:1385–1396, cover; https://doi.org/10.1083/jcb.107.4.1385. Part (b) republished with permission from Elsevier, from I. I. Singer, 1979, “The Fibronexus: A Transmembrane Association of Fibronectin- Containing Fibers and Bundles of 5 nm Microfilaments in Hamster and Human Fibroblasts,” Cell 16(3):675–685; permission conveyed through Copyright Clearance Center, Inc.] Description The micrograph labeled (a) shows bright red streaks coming from upper left to lower right, with tiny green spots along the ends. The micrograph labeled (b) shows long gray streaks at the left labeled fibronectin fibrils. A light area above these is labeled cell exterior. Thick gray areas to the right are labeled plasma membrane, and tiny thin gray lines streaking left to right and on the right side of the micrograph are labeled actin- containing microfilaments. The area below these is labeled cell interior. The force needed to unfold and expose functional self-association sites in fibronectin is much less than that needed to disrupt fibronectin-integrin binding. Thus fibronectin molecules remain bound to integrin while cell- generated mechanical forces induce fibril formation. In effect, the integrins, through adapter proteins, transmit the intracellular forces generated by the actin cytoskeleton to extracellular fibronectin (inside-out signaling via mechanotransduction). Gradually, the initially formed
Elastic Fibers Permit Many Tissues to Undergo Repeated Stretching and Recoiling
fibronectin fibrils mature into highly stable matrix components by covalent cross-linking. In some electron micrographs, exterior fibronectin fibrils appear to be aligned in a seemingly continuous line with bundles of actin fibers within the cell (Figure 20-36b). These observations and the results from other studies provided the first example of a molecularly well-defined adhesion receptor forming a bridge between the intracellular cytoskeleton and the ECM components — a phenomenon now known to be widespread. Elastic Fibers Permit Many Tissues to Undergo Repeated Stretching and Recoiling Elastic fibers are found in the ECM of a wide variety of tissues that are subject to mechanical strain or deformation, such as the lungs, which expand and contract during breathing (Figure 20-37a); the blood vessels, through which blood pulses due to the heartbeat; and the skin and many other tissues that stretch and contract. Elastic fibers permit the rubberlike reversible elastic stretching and recoiling of these tissues.
FIGURE 20-37 Elastic and collagen fibers in connective tissue. (a) Light-microscopic image of loose connective tissue from the lung. Collagen fibers (bundles of collagen fibrils) are stained pink; elastic fibers and nuclei of cells are stained purple. (b) Longitudinal and (c) cross-sectional electron microscopic images of elastic fibers and collagen fibrils (coll) in the skin of a mouse. The elastic fibers have a solid core of elastin (e) integrated into and surrounded by a bundle of microfibrils (mf). Scale bars, 0.25 μm. [Parts (b) and (c) republished with permission from Elsevier, from J. Choi et al., 2009, “Analysis of Dermal Elastic Fibers in the Absence of Fibulin-5 Reveals Potential Roles for Fibulin-5 in Elastic Fiber Assembly,” Matrix Biol. 28(4):211–220; permission conveyed through Copyright Clearance Center, Inc.] Description The micrograph labeled (a) titled connective tissue shows a pink web of fibers in and around cells with dark nuclei. The collagen fibers and elastic fibers are labeled. The micrograph labeled (b) shows a lower darkest area labeled collagen fibrils, a layer of
dark and white splotches above it labeled elastic fiber, and an area at the top looking like many black circles clustered along and labeled collagen fibrils. The micrograph labeled (c) shows the cross-sectional view of the micrograph (b), which looks like a dark spot in the center surrounded by the black circles from the collagen fibrils. The major component of an elastic fiber, which can be several hundred to several thousand nanometers in diameter, is an insoluble, amorphous core composed of the protein elastin. Elastin consists of aggregates of monomeric tropoelastin molecules that are covalently cross-linked via a lysyl oxidase–mediated process similar to that seen in collagen. Repetitive proline- and glycine-enriched hydrophobic sequence motifs contribute to the ability of tropoelastins to self-associate, extend under stress, and recoil efficiently after stretching. The elastin core is surrounded by a collection of 10–12-nm-diameter microfibrils (Figure 20-37b) made up of the proteins fibrillin, fibulin, and associated proteins such as LTBPs (latent TGF-β binding proteins). The microfibrils serve as scaffolds for the assembly of the elastic fiber’s core. Elastin-free microfibrils are found in the eye, where they transmit muscular force to reshape the lens for focusing and may provide structural support to the cornea. Similar to other components of the ECM, microfibrils participate in cell signaling. In the secretory pathway, LTBPs bind the inactive form of the secreted signaling protein transforming growth factor β (TGF-β; a cytokine, see Figure 16-22) prior to their co-secretion and incorporation into microfibrils. Biomechanical stress mediated by cell-surface integrins binding to and pulling on the LTBP/TGF-β complex or proteolytic cleavage are thought to be the direct cause of active TGF-β release from
the ECM and subsequent signaling (see Figure 16-22). Similar integrinmediated activation of the LTBP/TGF-β complex bound to other ECM proteins provides active TGF-β for regulation of a wide variety of biological processes. A variety of diseases, many involving skeletal and cardiovascular abnormalities, are consequences of mutations in the genes encoding the structural proteins of elastic fibers or the proteins that contribute to their proper assembly. For example, mutations in the fibrillin-1 gene cause Marfan syndrome, whose varied symptoms can include bone overgrowth, loose joints, abnormally long extremities and face, and cardiovascular defects due to weakness in the walls of the aorta and other blood vessels. There has been considerable speculation that President Abraham Lincoln’s unusually tall, elongated body may have been a consequence of Marfan syndrome. In mammals, most tropoelastin synthesis occurs immediately before and after birth during the late fetal and neonatal periods. Thus most of the body’s elastin needs to be very durable, lasting an entire lifetime. The extraordinary stability of elastin has been measured in a variety of ways. Pulse-chase experiments (see Chapter 3) using radioactive amino acid administration can be used to measure the life span of elastin in animals. In humans, two other methods employed to study the longevity of elastin have revealed that the mean lifetime of an elastin molecule in human lungs is about 70 years! The first method takes advantage of a naturally occurring phenomenon: the slow, natural rate of conversion of L-aspartic
Metalloproteases Remodel and Degrade the Extracellular Matrix
acid — incorporated into proteins during their synthesis — to D-aspartic acid. Thus the age of a long-lived protein can be estimated using chemical analysis to determine what fraction of its L-aspartic acid has been converted over time to the D isomer, together with knowledge of the age of the tissue from which it was isolated. The second method is a variation on the classic pulse-chase experiments used in the laboratory. As a consequence of nuclear weapons testing in the 1950s and 1960s, was introduced into the atmosphere and hence the food chain. This environmental has been used as the radioactive pulse in what is essentially a pulse-chase experiment to determine the stability of proteins of interest. Metalloproteases Remodel and Degrade the Extracellular Matrix Many key physiological processes, including morphogenesis during development, control of cellular proliferation and motility, response to injury, and even survival, require not only the production of ECM, but also its remodeling or degradation. Because of its enormous importance as a key element in the extracellular environment of multicellular organisms, remodeling and degradation of the ECM must be carefully controlled. Degradation of the ECM is often mediated by zinc-dependent ECM metalloproteases. Given the wide array of ECM components, it is not surprising that there are many such metalloproteases with varying substrate specificities and sites of expression. In many cases, their names incorporate the names of their substrates, as for the metalloproteases
called collagenases, gelatinases, elastases, and aggrecanases. Some are secreted into the extracellular fluid, and others are closely associated with the plasma membranes of cells, either tightly bound in a noncovalent association with the membrane or as integral membrane proteins. Many are initially synthesized as inactive precursors that must be specifically activated to function. ECM metalloproteases are divided into three major subgroups based on the enzymes’ structures: matrix metalloproteases (MMPs) (of which there are 23 in humans), a disintegrin and metalloproteinases (ADAMs), and ADAMs with thrombospondin motifs (ADAMTSs). These proteases can degrade ECM components as well as non-ECM components such as adhesion receptors. Indeed, a key function of ADAMs is cleaving extracellular domains from integral membrane proteins. In Chapter 16 we learned how ADAMs play key roles in signaling by Notch and EGF receptors. One mechanism used to control the activities of these proteases is the production of protein inhibitors called TIMPs (tissue inhibitors of metalloproteinases) and RECK (reversion-inducing–cysteine-rich protein with kazal motifs). Some of these inhibitors have their own cell-surface receptors and functions independent of their ability to inhibit metalloproteinases. ECM-degrading proteases are associated with a variety of diseases, the best known of which is metastatic (spreading) cancer (see Chapter 25). KEY CONCEPTS OF SECTION 20.4 The Extracellular Matrix II: Connective Tissue
Connective tissue, such as tendon and cartilage, differs from other solid tissues in that most of its volume is made up of extracellular matrix (ECM) rather than cells (see
Figure 20-4). The synthesis of fibrillar collagen (e.g., types I, II, and III) begins inside the cell with the chemical modification of newly made α chains and their assembly into triplehelical procollagen within the endoplasmic reticulum. After secretion, procollagen molecules are cleaved, associate laterally, and are covalently cross-linked into bundles called fibrils, which can form larger assemblies called fibers (see Figure 2028). The various collagens are distinguished by the ability of their helical and nonhelical regions to associate into fibrils, to form sheets, or to cross-link other collagen types (see Table 20-5). Proteoglycans consist of membrane-associated or secreted core proteins covalently linked to one or more glycosaminoglycan (GAG) chains, which are linear polymers of disaccharides that are often modified by sulfation. Cell-surface proteoglycans such as the syndecans facilitate cell-matrix interactions and help present certain external signaling molecules to their cell-surface receptors. Hyaluronan, a highly hydrated GAG, is a major component of the ECM of migrating and proliferating cells. Certain adhesion receptors bind hyaluronan to cells. Large proteoglycan aggregates containing a central hyaluronan molecule noncovalently bound to the core proteins of proteoglycan molecules (e.g., aggrecan) contribute to the ability of the matrix to resist compression forces (see Figure 20-33). Fibronectins are abundant multi-adhesive matrix proteins that play a key role in migration and cellular differentiation. They contain binding sites for integrins and ECM components (collagens, proteoglycans) and thus can attach cells to the ECM (see Figure 20-34). The tripeptide RGD motif Arg-Gly-Asp, found in fibronectins and some other matrix proteins, is recognized by several integrins. Elastic fibers permit repeated stretching and recoiling of tissues because of their highly elastic core of cross-linked, amorphous elastin, which is surrounded by a network of microfibrils that help assemble the fibers and regulate signaling mediated by TGF-β. The remodeling or degradation of ECM is mediated by a large number of secreted and cell-membrane-associated zinc metalloproteases that fall into several families (MMPs, ADAMs, ADAMTSs) and whose activities are regulated by protein inhibitors (TIMPs and RECK).
Integrins Mediate Adhesion and Relay Signals Between Cells and Their Three-Dimensional Environment
20.5 Adhesive Interactions in Motile and Nonmotile Cells After adhesive interactions in epithelia form during differentiation, they are often very stable and can last throughout the life span of the cells or until the epithelium undergoes further differentiation. Although such longlasting nonmotile adhesion also exists in nonepithelial tissues, some nonepithelial cells must be able to crawl across or through a layer of ECM or other cells. Moreover, during development or wound healing and in certain pathological states (e.g., cancer), epithelial cells can transform into more motile cells (the epithelial-to-mesenchymal transition). Changes in expression of adhesion molecules play a key role in this transformation, as they do in other biological processes involving cell movement, such as the crawling of white blood cells into tissue sites of infection. In this section, we describe various cell-surface structures that mediate transient adhesive interactions that are especially adapted for the movement of cells as well as those that mediate long-lasting adhesion. The intracellular mechanisms used to generate the mechanical forces that propel cells and modify their shapes are covered in Chapters 17 and 18. Integrins Mediate Adhesion and Relay Signals Between Cells and Their Three-Dimensional Environment
As already discussed, integrins can connect epithelial cells to the basal lamina and, through adapter proteins, to intermediate filaments of the cytoskeleton (see Figure 20-1). In both nonepithelial and epithelial cells, integrins form a bridge between the ECM and the cytoskeleton. In epithelial and nonepithelial cells, integrins in the plasma membrane are also clustered with other molecules in various focal contacts (focal adhesions) and focal contact–like adhesive structures called focal complexes, 3-D adhesions, and fibrillar adhesions, as well as in circular adhesions called podosomes. These structures are multiprotein complexes that mediate (1) cell adhesion to the ECM — for example, via integrin binding to fibronectin (see Figure 20-36), collagen or laminin, (2) integrin association with the actin cytoskeleton, (3) adhesion-dependent outside-in and inside-out signaling (see Figure 20-8), and (4) mechanosensory coupling between cells and their environments. These complexes are readily observed by fluorescence microscopy with the use of antibodies that recognize integrins or other molecules clustered with them (Figure 20-38).
EXPERIMENTAL FIGURE 20-38 Integrins cluster into adhesive structures with various morphologies in nonepithelial cells. Immunofluorescence methods were used to detect integrin-containing adhesive structures (green) on cultured cells: (a) focal adhesions and (b) 3-D adhesions on the surfaces of human fibroblasts. Cells were grown (a) directly on the flat surface of a culture dish or (b) on a three-dimensional matrix of ECM components. The shape, distribution, and composition of the integrin-based adhesions formed by cells vary depending on the cells’ environment. [Part (a) republished with permission from Nature, from B. Geiger et al., 2001, “Transmembrane Crosstalk Between the Extracellular Matrix and the Cytoskeleton,” Nat. Rev. Mol. Cell Biol. 2(11):793–805; permission conveyed through Copyright Clearance Center, Inc. Part (b) Kenneth Yamada and Edna Cukierman.] Description The micrograph labeled (a) is titled focal adhesion. The cell is fluorescing at points where integrins are expressed in bright green spots that look like a butterfly pattern. Points of fluorescence are visible at points along the cell membrane where the cell is in contact with a surface. The micrograph labeled (b) is titled three-dimensional adhesion and shows green fluorescence in three elongated clumps leaning from top left to bottom right. Fluorescence image shows focal adhesion over the surface of the cells.
Integrin-containing adhesive structures are dynamic due to ongoing import, export, or covalent modification of their components, and each contains dozens of intracellular adapter and associated proteins. These adhesive structures may in some cases assemble due to the formation of phase-separated biomolecular condensates (see Chapter 3) in which each component has the potential of binding directly to many other components. The hundreds of such proteins identified to date have the potential to engage in many hundreds of distinct protein-protein interactions that may be subject to regulation. For example, binding sites generated by phosphorylation of integrin and its associated proteins, as well as by generation of phosphorylated derivatives of phosphatidylinositol in the adjacent membrane, recruit additional proteins into and can also cause release of some proteins from these multiprotein complexes. A tightly controlled choreography of internal signals, contributions of other signaling pathways such as those involving receptor tyrosine kinases (see Figure 20-8), and external signals (such as the composition and rigidity of the ECM) regulates these complexes. Together, they help define the precise composition and activity of the integrin multiprotein complex and the consequent influence that it has on cellular structure and activity (outside-in effect) as well as the influence of the cellular actin cytoskeleton on the ECM (inside-out effect). The adhesive structures can be stable and substantially contribute to the architecture of a tissue. They can also be transient. For example, the dynamic formation and dissolution of adhesive structures can result in cells grabbing onto the ECM for traction, exerting force to move relative to the adhesion site, and then disassembling to release the cell, permitting net movement as cells crawl through adjacent tissue.
Although found in many nonepithelial cells, integrin-containing adhesive structures have been studied most frequently in fibroblasts grown in cell culture on flat glass or plastic surfaces called substrata. These conditions only poorly approximate the three-dimensional ECM environment that normally surrounds such cells in vivo. When fibroblasts are cultured in three-dimensional ECMs derived from cells or tissues, they form adhesions to the three-dimensional ECM substratum, called 3-D adhesions. These structures differ somewhat in composition, shape, distribution, and activity from the focal or fibrillar adhesions seen in cells growing on the flat substrata typically used in cell culture experiments (see Figure 20-38). Cultured fibroblasts in three-dimensional ECMs have anchoring junctions that display greater adhesion and mobility, increased rates of cell proliferation, and spindle-shaped morphologies more like those of fibroblasts in tissues than do cells cultured on hard, flat surfaces. These and other observations indicate that the topological, compositional, and mechanical properties of the ECM all play a role in controlling the shape and activity of a cell, due at least in part to integrin-mediated bridging of the ECM to intracellular molecules. Tissue-specific differences in these ECM characteristics probably contribute to the tissuespecific properties of cells. The importance of the three-dimensional environment of cells has been highlighted by cell-culture studies of the morphogenesis, functioning, and stability of specialized milk-producing mammary epithelial cells and their cancerous transformed counterparts. For example, the three-dimensional ECM-dependent outside-in signaling mediated by integrins influences the epidermal growth factor–tyrosine kinase receptor signaling system, and
Regulation of Integrin-Mediated Adhesion and Signaling Controls Cell Function and Movement
vice versa. The three- dimensional ECM also permits the mammary epithelial cells to generate in vivo–like circular epithelial structures, called acini, which secrete the major protein constituents of milk. The use of such three-dimensional ECM cell culture systems permits more realistic comparisons of the responses of normal and cancer cells to potential chemotherapeutic agents. Analogous systems employing both natural and synthetic three-dimensional ECMs are being developed to provide more in vivo–like conditions to study other complex tissues and organs, such as the liver. In the presence of the appropriate ECM, many types of 3-D, organ-like tissues called organoids, similar to those in intact organisms, can now be generated from self-organizing, pluripotent or adult stem cells (see Chapters 4 and 22). Regulation of Integrin-Mediated Adhesion and Signaling Controls Cell Function and Movement Cells can exquisitely control the strength of integrin-mediated cell-matrix interactions by regulating integrin’s expression levels, ligand-binding activities, or both. Such regulation is critical to the role of these interactions in cell migration and other functions involving cell movement. Integrin Binding
Integrin binding to ECM and neighboring cells can influence the developmental fate of cells, determining the type of cell into which a progenitor cell will differentiate (see Chapter 22). Integrins participate in cells sensing the mechanical characteristics of their environments, such as the rigidity of the ECM, and responding appropriately. Most integrins can exist in at least three conformations. In the first, inactive state (bent closed conformation, Figure 20-39a, left), the closely apposed extracellular domains of the αβ heterodimer are bent over, placing the ligand-binding site — located at the interface of the propeller (α chain) and βA (β chain) domains — near the plasma membrane and distant from the ECM. In the bent closed conformation, there is only low-affinity ligand binding and the transmembrane domains and cytoplasmic C-terminal tails of the two subunits are closely bound together. In the second, partially active state, the transmembrane and cytoplasmic domains of the α and β chains at least partially separate and the extracellular domains partially straighten into an extended closed conformation in which the ligand-binding site is projected farther away from the cell surface and assumes an intermediate affinity for ligands, such as a variety of ECM proteins (Figure 20-39a, center). In the third, fully active, extended open conformation, tension on the protein fully extends the extracellular domain of the β chain and confers high affinity ligand binding that can be as much as 4000-fold tighter than binding to the bent closed conformation (Figure 20-39a, right). The changes in the conformations of integrin’s cytoplasmic tails can influence the binding of intracellular adaptor proteins. For example, two of integrin’s cytoplasmic adaptor proteins, talin and kindlin, can bind to short sequence motifs in the cytoplasmic tail of
the β chain when it is separated from the tail of the α chain in the extended closed and open forms, but not when these sites are blocked in the bent closed form (see Figure 20-39a). EXPERIMENTAL FIGURE 20-39 The three conformations of integrins. (a) Most integrins can be in one of three interconvertible conformations: inactive, low affinity for extracellular ligands (bent closed, left); partially active, intermediate affinity (extended closed, center); and fully active, high affinity (extended open, right). Tension across the integrin due to pulling by either its extracellular ligand or intracellular binding partners drives the integrin into the extended open conformation. The extracellular ligand binding site is at the interface of the propeller domain on the α chain and the βA domain on the β chain. The cytoplasmic domain of the β chain has binding sites for intracellular adaptor
proteins, e.g., talin and kindlin (green and yellow ovals, respectively). These sites are inaccessible (cryptic) in the bent closed form, but accessible in the extended forms due to separation of the transmembrane and cytoplasmic domains of the α and β chains (indicated by double-headed arrow). (b) Single inactive (bent closed) integrin αIIbβ3 molecules (left) were incorporated into phospholipid nanodiscs (small bilayers in which the extracellular and cytoplasmic domains of the integrin are exposed to a buffer), and the integrin-binding and activating head domain of the adapter protein talin was added to some of these preparations (right). Multiple electron microscopic images of individual nanodiscs were collected and averaged. Phospholipid nanodiscs are indicated by dashed white circles, and the heights of the integrin extracellular regions that extend above the nanodiscs are indicated by brackets. Binding of talin’s head domain to the cytoplasmic domain of the β chain traps the integrins in the extended closed conformation. [Part (a) Information from J. Wang et al., 2019, Nat. Commun. 10:5481. Part (b) F. Ye et al., 2010, J. Cell Biol. 188(1):157–173, Fig. 7; https://doi.org/10:1083/jcb.200908045.] Description The illustration labeled (a) shows three schematics. The first one, at left, is labeled inactive low affinity (bent closed). There is a cell membrane with a ligand represented as a blue thin line hanging from it and the integrin is bent down and not connected to the ligand. In the center schematic, the integrin has straightened upward and the blue side has connected to the membrane while the pink side top looks like an open circle starting to surround the ligand. This is labeled partially active, intermediate affinity extended closed. The last diagram shows the pink side has closed around the ligand and stretched straight. A dotted line is labeled tension along that pink line. The label read fully active, high affinity (extended open). The micrographs labeled (b) show the closed integrin and labeled inactive integrin, bent closed. A dotted line outlines the membrane nanodisc. Another micrograph shows the open integrin connected to the nanodisc labeled partially active integrin, extended closed. One helpful way to think about these three forms of integrins is that there is an equilibrium distribution of integrin molecules in all three conformations (they can readily interconvert). When the integrin is bound
to neither extracellular ligands (e.g., ECM) nor intracellular partners (e.g., talin), the large majority of unbound integrin molecules is in the bent, inactive conformation. Binding of extracellular ligands to the extended conformations traps more of the integrins in the extended conformations than in the absence of the ligands (shifts the equilibrium distribution) and thus facilitates the binding of intracellular adaptor proteins such as talin or kindlin to the now accessible sites in the β chain’s cytoplasmic domain. Similarly, binding of intracellular adaptors to the extended conformations increases the fraction of integrin molecules in extended conformations (again, a shift in the equilibrium distribution) and facilitates extracellular ligand binding to the intermediate and high-affinity ligand-binding sites. Thus binding of ligands to integrins on either side of the membrane can increase binding on the other side. The structural features of integrin’s α and β subunits that permit them to adopt the bent and extended conformations have been preserved through metazoan evolution, indicating their importance for integrin function. Indeed, these three conformations provide an attractive explanation for the ability of integrins to mediate outside-in and inside-out signaling. The binding to the integrin’s extracellular ligand-binding site of certain ECM molecules or CAMs on other cells would shift the distribution of conformations and thus result in more integrins in the extended closed or extended open forms with at least partially separated cytoplasmic tails. Intracellular adapter proteins could recognize the separation of the tails and, as a result, bind to the tails. Integrin-binding-dependent changes in these adapters can then alter the cytoskeleton and activate or inhibit intracellular signaling pathways. Conversely, changes in the metabolic or
signaling state of the cells could cause intracellular adapters to bind to or dissociate from the cytoplasmic tails of the integrins and thus alter the distribution of integrin conformations. Such binding can force the tails either to separate or to associate (see Figure 20-39a). As a consequence, a greater fraction of the integrins would be either in the bent closed (inactive), extended closed (partially active), or extended open (fully active) conformations, thereby altering their interactions with the ECM or with other cells. Indeed, in vitro EM studies of purified integrins reconstituted individually into lipid bilayer nanodiscs show that binding of the head domain of the adapter/mechanosensor protein talin (described in detail below; see Figure 20-40) to the cytoplasmic tail of integrin’s β chain is sufficient to partially activate integrin, inducing a straightening of the bent closed conformation into the extended closed form (see Figure 2039b). We can see an example of integrins mediating intracellular-extracellular communication in the initial formation of focal adhesions in Figure 20-40. Although there are many intracellular proteins involved, this figure focuses on talin, actin, and vinculin. Figure 20-40a shows a model of an active conformation of talin and Figures 20-40b and c show some of the key steps associated with integrin/talin-mediated inside-out signaling. In the absence of stimulation, most of the talin in the cytoplasm is condensed into an inactive, autoinhibited, compact structure in which interdomain binding sterically blocks talin binding to some of its many partners, including integrins and actin (see Figure 20-40b, lower left).
FIGURE 20-40 Talin- and integrin-mediated intracellular-extracellular communication. (a) Top: A hypothetical model of an elongated, active conformation of a talin monomer
based on structures of fragments of the intact protein with secondary structures illustrated as tubes (α helices) and ribbons with arrowheads (β sheets). The N-terminal “head” domain (four globular subdomains) is connected by an apparently unstructured domain to a rod domain that comprises 13 α-helical bundles (4 or 5 helices per bundle), 9 of which contain either 1 or 2 helices (red, total of 11) that are cryptic binding sites for the protein vinculin. Some bundles also contain cryptic actin-binding sites. These helices can only bind vinculin when exposed by unfolding (see Figure 20-9b). There is a single α helix at the extreme C-terminus that is a dimerization domain. Bottom: A cartoon model. Head subdomains = gray ovals; rod domain bundles = colored capsules; and dimerization domain = narrow rectangle. (b) Intracellular signaling induces the translocation of inactive talin in the cytoplasm to the plasma membrane and its unfolding/activation (steps 1 and 2 ). The binding of activated talin to integrin’s β chain cytoplasmic domain stabilizes integrin’s extended closed conformation, promoting binding to the ECM (step 3 ). The unfurled talin homodimerizes and binds several proteins, including actin filaments that bind via talin’s C-terminal dimerization domain (shown) and several other sites (not shown) (step 4 ). The actin filaments exert force (step 5 ), generating tension across the ECM/integrin/talin complex. This tension converts the integrin into the fully active, extended open form (step 6 ). (c) Subsequent steps illustrated for the region outlined in the light gray dashed box in part (b) are shown. Additional force unfolds one or more of talin’s helical bundles (step 7 ), exposing cryptic binding site(s) (red helix) that recruit vinculin and other proteins, including more actin (steps 8 and 9 ), resulting in growth of the focal adhesion. [Part (a) left information from B. T. Goult, J. Yan, and M. A. Schwartz, 2018, J. Cell Biol. 217(11):3776–3784; and M. Yao et al., 2014, Sci. Rep. 4:4610. Parts (b) and (c) information from J. Wang et al., 2019, Nat. Commun. 10:5481; D. Dedden et al., 2019, Cell 179(1):120– 131; S. Chakraborty et al., 2019, Biochemistry 58(47):4677–4695; Y. A. Kadry and D. A. Calderwood, 2020, Biochim. Biophys. Acta, Biomembr. 1862(5):183206.] Description The illustration labeled (a) shows a horizontal three-dimensional ribbon model of talin. At left, a gray group of ribbons labeled talin (extended conformation), connected to a center set of red and white ribbons labeled alpha-helical bundles, which are in turn connected to bundles labeled cryptic vinculin binding alpha helices (red). Below this is the same structure drawn in schematic and labeled, from left to right, talin "head" (F E R M domain), unstructured (an area of thin gray lines), rod domain (13 alpha-helical
bundles, and dimerization. The illustration labeled (b) shows a schematic that starts with membranes as gray bars with ligands, and an inactive integrin, with an inactive talin looking like a tangled web of rectangular beads. Next, to the right, the integrin is extended and attaches to the partially active integrin. Then the pink part of the integrin separates from the blue part and dimerization is labeled at the other end of the integrin. Next, the integrin is fully active, and now two parts are with the talin, each one stretched down with tension to meet together at an actin protein represented as red circles. In the illustration, labeled (c) the helical part of the talin bends and reveals vinculin-binding areas represented as light blue ovals, which now attach to the actin. Intracellular signaling can induce the translocation of talin to the inner leaflet of the plasma membrane, for example, by generating polyphosphoinositides to which globular subdomains in talin’s head domain can bind and by small GTP-binding proteins (Figure 20-40b, steps 1 and 2 ). In steps 2 and 3 , the compact autoinhibited rod domain unfolds into the active conformation (see Figure 20-40a) and a portion of the head domain binds to the cytoplasmic domain of integrin’s β chain. Talin binds to integrin’s β chain cytoplasmic domain when the transmembrane and cytoplasmic domains of integrin’s α and β chains at least partly separate in the extended closed conformation. In this conformation integrin binds to the ECM with intermediate affinity (see
Figure 20-39). The order of these events is not well established. The C-terminal dimerization domain in the unfurled, extended, and active talin mediates homodimerization, and actin filaments can bind to several sites in the rod domains in the dimeric talin, including the C-terminal dimerization domain (step 4 , also see Figure 20-40a). Other binding partners can also bind to the unfurled, active talin. Additional signaling steps mediated by the partially active, extended closed integrin (e.g.,
activation of kinases, such as focal adhesion kinase and scr-like kinases) induce formin-mediated actin filament formation and myosin engagement with actin filaments to exert force on the talin (step 5 , also see Chapter 17). As a consequence, tension develops in the ECM-bound integrin/talin complex, converting the integrin into the fully active, extended open form (step 6 ). Additional force applied to the integrin/talin complex, exerted either from the cytoplasm (e.g., actin filaments and myosin) or the ECM, can pull apart the α-helical bundles in talin (Figure 20-40c, step 7 ), exposing cryptic vinculin binding sites (red helix). The greater the force, the more α-helical bundles in talin are opened and the more vinculin, actin, and other proteins are recruited to the integrin/talin complex. Vinculin binding to exposed sites on talin can then recruit other proteins, such as actin filaments (steps 8 and 9 ). The unfolded bundles can readily refold when the force is reduced. Strikingly, the more rigid the structure of the ECM, the greater the tension across the integrin/talin complex and the larger and stronger the adhesion. During steps 3 – 9 , integrins begin to cluster and the focal adhesion can continue to grow and form a mature focal adhesion that firmly attaches the cell to the ECM. Alternatively, the multiprotein adhesion complex can dissociate with inactivation of the integrins and other components, in part due to recruitment of inhibitory proteins. Studies suggest that the efficient activation of integrins in intact cells may also require the participation of another class of adapter proteins called kindlins, which bind to a distinct site on the cytoplasmic tail of integrin’s β
chain (see Figure 20-39a). Kindlin plays a key role in the integrin- and microfibril-mediated activation of TGF-β (inside-out signaling involving elastic fibers and their microfibril-associated protein LTBP, described earlier) and other pathways of integrin-mediated signaling, such as recruitment of branched actin networks in lamellipodia (see Chapter 17). Platelet function, discussed in more detail below, provides a good example of how cell-matrix interactions are modulated by control of integrinbinding activity. Platelets are cell fragments that circulate in the blood and clump together with ECM molecules to form a blood clot. In its basal state, the αIIbβ3 integrin present on the plasma membranes of platelets cannot bind tightly to its protein ligands (including fibrinogen and fibronectin), all of which participate in the formation of a blood clot, because it is in the inactive (bent closed) conformation. During clot formation, platelets are activated by binding to ECM proteins such as collagen and a large protein called von Willabrand factor that, through binding to receptors, generate intracellular signals. Platelets may also be activated by ADP or the clotting enzyme thrombin. These signals induce changes in signaling pathways within the platelet that result in an activating conformational change in the platelet’s αIIbβ3 integrin. As a consequence, this integrin can bind tightly to extracellular clotting proteins and participate in clot formation. People with genetic defects in the β3 integrin subunit are prone to excessive bleeding, attesting to the role of the αIIbβ3 integrin in the formation of blood clots (see Table 20-4). Integrin Expression
Connections Between the ECM and Cytoskeleton Are Defective in Muscular Dystrophy
The attachment of cells to ECM components can also be modulated by altering the number of integrin molecules exposed on the cell surface. The α4β1 integrin, which is found on many hematopoietic cells, offers an example of this regulatory mechanism. For these hematopoietic cells to proliferate and differentiate, they must be attached to fibronectin synthesized by supportive (stromal) cells in the bone marrow. The α4β1 integrin on hematopoietic cells binds to a Glu-Ile-Leu-Asp-Val (EILDV) sequence in fibronectin in the ECM, thereby anchoring the cells to the matrix. This integrin also binds to a sequence in a CAM called vascular CAM-1 (VCAM-1), which is present on stromal cells of the bone marrow. Thus hematopoietic cells directly contact the stromal cells as well as the ECM. Late in their differentiation, hematopoietic cells decrease their expression of this integrin; the resulting reduction in the number of α4β1 integrin molecules on the cell surface is thought to allow mature blood cells to detach from the ECM and stromal cells in the bone marrow and enter the circulation. Connections Between the ECM and Cytoskeleton Are Defective in Muscular Dystrophy The importance of the adhesion receptor–mediated linkage between ECM components and the cytoskeleton is highlighted by a set of hereditary muscle-wasting diseases, collectively called muscular dystrophies. Duchenne muscular dystrophy (DMD), the most common
type, is a sex-linked disorder, affecting 1 in 3300 boys, that results in cardiac or respiratory failure, usually in the late teens or early twenties. The first clue to understanding the molecular basis of this disease came from the discovery that people with DMD carry mutations in the gene encoding a protein named dystrophin. This very large protein was found to be a cytosolic adapter protein that contains a long central rodlike region with 24 repeats of a three-α-helix bundle, flanked by an N-terminal Factin-binding domain and C-terminal domains that bind to several plasma membrane and cytoplasmic proteins (Figure 20-41). Several of the central repeats also bind to F-actin and dystrophin links the actin filaments to an adhesion receptor called dystroglycan (see Figure 20-41). It is noteworthy that when dystrophin is subjected to tension some of its α-helical repeats unfold and the protein substantially lengthens, a feature somewhat reminiscent of the force-induced unfolding of talin (see Figure 20-40). However, a role for dystrophin in mechanotransduction has not been established.
FIGURE 20-41 The dystrophin glycoprotein complex (DGC) in skeletal muscle cells. This schematic model shows that the DGC comprises three subcomplexes: the α, β-Dystroglycan subcomplex; the integral membrane protein sarcospan and the sarcoglycan complex; and the cytosolic adapter subcomplex comprising dystrophin, other adapter proteins, and signaling molecules. Through its O-linked matriglycan sugars (see Figure 2031c), α-dystroglycan binds to LG-domain-containing components of the basal lamina, such as laminin and the GAG-containing proteoglycans perlecan and agrin. Dystrophin — the protein that is defective in DMD — links β-dystroglycan to the actin cytoskeleton, and α-dystrobrevin links dystrophin to the sarcoglycan complex and sarcospan. Neuronal nitric oxide synthase (nNOS), which produces the gaseous signaling molecule nitric oxide, binds to dystrophin via the adaptors called syntrophins. See S. J. Winder, 2001, Trends Biochem. Sci. 26:118–124; D. E. Michele and K. P. Campbell, 2003, J. Biol. Chem. 278(18):15457– 15460; T. Yoshida-Moriguchi and K. P. Campbell, 2015, Glycobiology 25(7):702–713; and
Y. M. Kobayashi and K. P. Campbell, 2012, Chapter 66, “Skeletal Muscle DystrophinGlycoprotein Complex and Muscular Dystrophy,” in J. A. Hill and E. N. Olson, eds., Muscle, vol. 2, Elsevier/Academic Press. Description The illustration shows agrin, laminin, and perlecan in the basal lamina. These can interact with the O-linked sugars of the alpha-beta-dystroglycan complex. This complex is attached to the sarcoglycan complex in the cell membrane, which is also connected to dystrophin and various other cytosolic proteins such as syntrophins, nitric oxide synthase, and alpha dystrobrevin. Dystrophin is connected to actin. Dystroglycan is a heterodimeric (α and β subunits) cell-surface glycoprotein (see Figure 20-41). As noted in Section 20.4, about three of the more than 20 O-linked oligosaccharide chains in dystroglycan’s α subunit are directly linked to the hydroxyl group via a mannose sugar that is phosphorylated (see Figure 20-31c). A phosphorylated hexasaccharide connects the phosphorylated mannose to a GAG-like polymer of xylose– glucuronic acid disaccharides called matriglycan. One key enzyme in the synthesis of the matriglycan oligosaccharide that is located in the Golgi complex is called LARGE. The O-linked matriglycan binds to various components of the basal lamina, including the multi-adhesive matrix protein laminin (see Figures 20-25 and 20-41) and the proteoglycans perlecan and agrin. The neurexins, a family of cell-surface adhesion molecules expressed by neurons, also bind matriglycan. All of these bind to matriglycan via LG domains (see
Dystroglycan’s transmembrane β subunit associates with a group of integral membrane proteins called the sarcoglycan complex and sarcospan, while its cytosolic domain binds directly to dystrophin and directly or indirectly to other adapter and signaling proteins (see Figure 20-41). Sarcospan is a tetraspanin, as are claudin and occludin in tight junctions (see Figure 20-18b). The resulting large, heteromeric assemblage, the dystrophin glycoprotein complex (DGC), links the ECM to the actin cytoskeleton. Binding to laminin in the basal lamina leads to a cluster of DGCs on the cell surface. The DGC is associated with signaling pathways within muscle and other types of cells. For instance, the signaling enzyme neuronal nitric oxide synthase (nNOS) is associated through syntrophin with the DGC in skeletal muscle. The rise in intracellular during muscle contraction activates nNOS to produce nitric oxide (NO), a signaling molecule that diffuses into smooth muscle cells surrounding nearby blood vessels. NO promotes smooth muscle relaxation, leading to a local rise in the flow of blood supplying nutrients and oxygen to the active skeletal muscle. Heart (cardiac) muscle contraction may be influenced by similar NOS-syntrophin interactions. Mutations in the genes encoding dystrophin, other DGC components, laminin, or the multiple enzymes that mediate the addition of matriglycan to dystroglycan can all disrupt the DGC-mediated link between the exterior and the interior of muscle cells and cause muscular dystrophies. In addition, dystroglycan mutations have been shown to greatly reduce the clustering of acetylcholine receptors on muscle cells at the neuromuscular junctions, which is also dependent on the basal lamina proteins laminin and agrin. These and possibly other effects of DGC defects apparently lead
IgCAMs Mediate Cell-Cell Adhesion in Neural and Other Tissues
to a cumulative loss of the mechanical stability of muscle cells as they undergo contraction and relaxation, resulting in deterioration of the cells and muscular dystrophy. Dystroglycan is an elegant and medically relevant example of the intricate networks of connectivity in cell biology. While originally discovered in the context of studying muscular dystrophy, it was later shown to be expressed in nonmuscle cells and, through its binding to laminin, to play a key role in the assembly and stability of at least some basement membranes. The extent of glycosylation of dystroglycan, such as the length of the matriglycan repeats, varies in different tissues, suggesting that alterations in its glycosylation regulate function. Dystroglycan is essential for normal development, including the proper functioning of some neuronal synapses (see Chapter 23). Additional studies led to its identification as a cell-surface receptor for the virus that causes the frequently fatal human disease Lassa fever and other related viruses, all of which bind via matriglycan. Furthermore, dystroglycan is the receptor on specialized cells in the nervous system — Schwann cells — to which binds the pathogenic bacterium Mycobacterium leprae, the organism that causes leprosy. IgCAMs Mediate Cell-Cell Adhesion in Neural and Other Tissues Numerous transmembrane proteins characterized by the presence of multiple immunoglobulin domains in their extracellular regions constitute
the immunoglobulin (Ig) superfamily of CAMs, or IgCAMs (e.g., see NCAM in Figure 20-2). The Ig domain is a common protein domain, containing 70–110 residues. It was first identified in antibodies, the antigen-binding immunoglobulins (see Chapter 24), but has a much older evolutionary origin in CAMs. The human, Drosophila, and C. elegans genomes include about 765, 150, and 64 genes, respectively, that encode proteins containing Ig domains. Immunoglobulin domains are found in a wide variety of cell-surface proteins, including the T-cell receptors produced by lymphocytes and many proteins that take part in adhesive interactions. Among the IgCAMs are neural CAMs; intercellular CAMs (ICAMs), which function in the movement of leukocytes into tissues; and junction adhesion molecules (JAMs), which are present in tight junctions (see Figure 20-18b). As their name implies, neural CAMs are of particular importance in neural tissues. One type, the NCAMs, primarily mediate homophilic interactions. First expressed during morphogenesis, NCAMs play an important role in the differentiation of muscle cells, glial cells, and neurons. Their role in cell adhesion has been directly demonstrated by the inhibition of adhesion with anti-NCAM antibodies. Numerous NCAM isoforms, encoded by a single gene, are generated by alternative mRNA splicing and by differences in glycosylation. Other neural CAMs (e.g., L1-CAM) are encoded by different genes. In humans, mutations in different parts of the L1-CAM gene cause various neuropathologies (e.g., mental retardation, congenital hydrocephalus, and spasticity).
Leukocyte Movement into Tissues Is Orchestrated by a Precisely Timed Sequence of Adhesive Interactions
An NCAM comprises an extracellular region with five Ig domains and two fibronectin type III domains, a single membrane-spanning segment, and a cytosolic segment that interacts with the cytoskeleton (see Figure 20-2). In contrast, the extracellular region of L1-CAM has six Ig domains and four fibronectin type III domains. As with cadherins, cis (on the same cell) interactions and trans (intercellular) interactions probably play key roles in IgCAM-mediated adhesion (see Figure 20-3); however, adhesion mediated by IgCAMs is independent. Leukocyte Movement into Tissues Is Orchestrated by a Precisely Timed Sequence of Adhesive Interactions In adult organisms, several types of white blood cells (leukocytes) participate in defense against infection caused by bacteria and viruses and respond to tissue damage due to trauma or inflammation. To fight infection and clear away damaged tissue, these cells must move rapidly from the blood, where they circulate as unattached, relatively quiescent cells, into the underlying tissue at sites of infection, inflammation, or damage. We know a great deal about the movement into tissue, termed extravasation, of four types of leukocytes: neutrophils, which release several antibacterial proteins; monocytes, the precursors of macrophages, which can engulf and destroy foreign particles; and T and B lymphocytes, the antigen-recognizing cells of the immune system (see Chapter 24).
Extravasation requires the successive formation and breakage of cell-cell contacts between leukocytes in the blood and endothelial cells lining the vessels. Some of these contacts are mediated by selectins, a family of CAMs that mediate leukocyte–vascular endothelium interactions. Endothelial cells express P- and E-selectins on their blood-facing surfaces, activated platelets express P-selectin, and leukocytes express L-selectin. All selectins contain a -dependent lectin domain, which is located at the distal end of the extracellular region of the molecule and recognizes particular sugars in glycoproteins or glycolipids (see Figure 20-2). For example, the primary ligand for P- and E-selectins is an oligosaccharide called the sialyl Lewis-x antigen, a part of longer oligosaccharides present in abundance on leukocyte glycoproteins and glycolipids.
Figure 20-42 illustrates the basic sequence of cell-cell interactions leading to the extravasation of leukocytes. Various inflammatory signals released in areas of infection or inflammation first cause activation of the vascular endothelium. P-selectin exposed on the surfaces of activated endothelial cells mediates the weak adhesion of passing leukocytes. Because of the force of the blood flow and the rapid “on” and “off” rates of P-selectin binding to its ligands, these bound leukocytes are slowed, but not stopped, and literally roll along the surface of the endothelium. Among the signals that promote activation of the endothelium are chemokines, a group of small, secreted proteins (8–12 kDa) produced by a wide variety of cells, including endothelial cells and leukocytes.
FIGURE 20-42 Endothelium-leukocyte interactions: activation, binding, rolling, and extravasation. Step 1 : In the absence of inflammation or infection, leukocytes and endothelial cells lining blood vessels are in a resting state and not interacting. Step 2 : Inflammatory signals released only in areas of inflammation, infection, or both activate resting endothelial cells, resulting in the movement of vesicle-sequestered selectins to the cell surface. The exposed selectins mediate weak binding of leukocytes by interacting with carbohydrate ligands on leukocytes. Blood flow forces the loosely bound leukocytes to roll along the endothelial surface of the blood vessel (curved arrow). Activation of the endothelium also causes synthesis of platelet-activating factor (PAF) and ICAM-1, both expressed on the endothelial cell surface. PAF and other, usually secreted, activators, including chemokines, then induce changes in the shapes of the leukocytes and activation of leukocyte integrins such as αLβ2, which is expressed by T lymphocytes (step 3 ). The subsequent tight binding between activated integrins on leukocytes and CAMs on the endothelium (e.g., ICAM-2 and ICAM-1) results in firm adhesion (step 4 ) and subsequent movement (extravasation) into the underlying tissue (step 5 ). See R. O. Hynes and A. D. Lander, 1992, Cell 68:303. Description The steps involved in the process are as follows. Step 1. Leukocyte in resting state (represented as a blue circle with a nucleus). The leukocyte has alpha L beta 2 integrin on its surface, two yellow selectin ligands, and a P A F receptor. Endothelial cells below it contain I C A M - 2 receptors on the cell surface and contain vesicles containing P-selectin. Step 2. Endothelial activation by the release of P selectin and leukocyte attachment via the selectin ligand and I C A M - 2. Step 3. Leukocyte activation, by docking of P A F on the endothelial cell with the P A F receptor. Step 4.
Firm adhesion via integrin slash I C A M binding. Step 5. Extravasation, where the leukocyte squeezes between endothelial cells as it moves from the blood into the tissue. For tight adhesion to occur between activated endothelial cells and leukocytes, β2-containing integrins on the surfaces of the leukocytes must be activated indirectly by chemokines or by other local activation signals such as platelet-activating factor (PAF). Platelet-activating factor is unusual in that it is a phospholipid rather than a protein; it is exposed on the surfaces of activated endothelial cells at the same time that P-selectin is exposed. The binding of PAF or other activators to their G protein– coupled receptors on leukocytes leads to activation of the leukocyte integrins (see Figure 20-39). These activated integrins then bind to distinct IgCAMs on the surfaces of endothelial cells. These IgCAMs include ICAM-2, which is expressed constitutively, and ICAM-1, whose synthesis is induced by activation. ICAM-1 does not usually contribute substantially to leukocyte adhesion to endothelial cells immediately after activation, but rather participates at later times in cases of chronic inflammation. The tight adhesion mediated by these -independent integrin-ICAM interactions leads to the cessation of rolling and to the spreading of leukocytes on the surface of the endothelium; soon the adhered cells move between adjacent endothelial cells and into the underlying tissue. The extravasation step itself (also called transmigration or diapedesis; step 5 in Figure 20-42) requires the dissociation of otherwise stable adhesive interactions between endothelial cells that are primarily mediated by the CAM VE-cadherin. There is general agreement that the leukocyte interactions with endothelial cells mediated by CAMs initiate outside-in signaling in the endothelial cells that involves phosphorylation, activation
of small GTPases, and an increase in cytosolic calcium concentration. These signals weaken or disrupt VE-cadherin-mediated inter-endothelial- cell adherens junctions and increase actin-myosin contraction, which pulls the endothelial cells apart, thus permitting the paracellular, amoeboid movement of the leukocyte between adjacent endothelial cells that is responsible for most extravasation. The selective adhesion of leukocytes to the endothelium near sites of infection or inflammation thus depends on the sequential appearance and activation of several different CAMs on the surfaces of the interacting cells. Different types of leukocytes express different integrins, though all contain the β2 subunit. Nonetheless, all leukocytes move into tissues by the general mechanism depicted in Figure 20-42. Many of the CAMs used to direct leukocyte adhesion are shared among different types of leukocytes and target tissues, yet often only a particular type of leukocyte is directed to a particular tissue. How is this specificity achieved? A three-step model has been proposed to account for the celltype specificity of such leukocyte-endothelium interactions. First, endothelial activation promotes initial relatively weak, transient, and reversible binding (e.g., the interaction of selectins and their carbohydrate ligands). Without additional local activation signals, the leukocyte will quickly move on. Second, cells in the immediate vicinity of the site of infection or inflammation release or express chemical signals such as chemokines and PAFs that activate only special subsets of the transiently attached leukocytes, depending on the types of chemokine receptors those leukocytes express. Third, additional activation-dependent CAMs (e.g.,
integrins) engage their binding partners, leading to strong, sustained adhesion. Only if the proper combination of CAMs, binding partners, and activation signals are engaged together with the appropriate timing at a specific site will a given leukocyte adhere strongly. Such combinatorial diversity and cross talk allows a small set of CAMs to serve diverse functions throughout the body — a good example of biological efficiency. Leukocyte-adhesion deficiency is caused by a genetic defect in the synthesis of the integrin β2 subunit. People with this disorder are susceptible to repeated bacterial infections because their leukocytes cannot extravasate properly and thus cannot effectively fight infection within a tissue. Some pathogenic viruses have evolved mechanisms to exploit cell-surface proteins that participate in the normal response to inflammation. For example, many of the RNA viruses that cause the common cold (rhinoviruses) bind to and enter cells through ICAM-1, and chemokine receptors can be important entry sites for human immunodeficiency virus (HIV), the cause of AIDS. Integrins appear to participate in the binding or internalization of a wide variety of viruses, including reoviruses (which cause fever and gastroenteritis, especially in infants), adenoviruses (which cause conjunctivitis and acute respiratory disease), and foot-and-mouth disease virus (which causes fever in cattle and pigs). KEY CONCEPTS OF SECTION 20.5 Adhesive Interactions in Motile and Nonmotile Cells
Many cells have integrin-containing clusters of proteins (e.g., focal adhesions, 3-D adhesions, podosomes) that physically and functionally connect cells to the ECM and facilitate inside-out and outside-in signaling. Via interaction with integrins, the three-dimensional structure of the ECM surrounding a cell can profoundly influence the behavior of the cell. Integrins exist in at least three conformations (bent closed, extended closed, extended open) that differ in their affinity for ligands and in their interactions with cytosolic adapter proteins and extracellular ligands (see Figure 20-39). Switching between these conformations depends, in part, on the application of mechanical force and allows regulation of integrin activity, which is important for control of cell adhesion and movements. Talin, one of many cytoplasmic adaptors that interact with integrins, can function as a mechanosensor. Talin can mediate integrin’s interactions with the cytoskeleton and thus its switching between conformational and functional states. Dystroglycan, an adhesion receptor, forms a large complex with dystrophin, other adapter proteins, and signaling molecules (see Figure 20-41). This complex links the actin cytoskeleton to the surrounding ECM, providing mechanical stability to muscle. Mutations in various components of this complex cause different types of muscular dystrophy. Neural cell-adhesion molecules, which belong to the immunoglobulin (Ig) family of CAMs, mediate - independent cell-cell adhesion in neural and other tissues. The combinatorial and sequential interaction of several types of CAMs (e.g., selectins, integrins, and ICAMs) is critical for the specific adhesion of different types of leukocytes to endothelial cells in response to local signals induced by infection or inflammation (see Figure 20-42).
20.6 Plant Tissues
20.6 Plant Tissues We turn now to the assembly of plant cells into tissues. The overall structural organization of plants is generally simpler than that of animals. For instance, plants have only four broad types of cells, which in mature plants form four basic classes of tissue: dermal tissue interacts with the environment, vascular tissue transports water and dissolved substances such as sugars and ions, space-filling ground tissue constitutes the major sites of metabolism, and sporogenous tissue forms the reproductive organs. Plant tissues are organized into just four main organ systems: stems have support and transport functions, roots provide anchorage and absorb and store nutrients, leaves are the sites of photosynthesis, and flowers enclose the reproductive structures. Thus at the cell, tissue, and organ levels, plants are generally less complex than most animals. Moreover, unlike animals, plants do not replace or repair old or damaged cells or tissues; they simply grow new organs. Indeed, the developmental fate of any given plant cell is primarily based on its position in the organism rather than on its lineage, whereas both are important in animals (see Chapter 22). In both plants and animals, a cell’s direct communication with its neighbors is important. Most importantly for this chapter, and in contrast with animals, few cells in plants contact one another directly through molecules incorporated into their plasma membranes. Instead, plant cells are typically surrounded by a rigid cell wall that contacts the cell walls of adjacent cells (Figure 20-43a). Also in contrast with animal
cells, a plant cell rarely changes its position in the organism relative to other cells. These features of plants and their organization have determined the distinctive molecular mechanisms by which plant cells are incorporated into tissues and communicate with one another.
FIGURE 20-43 Structure of the plant cell wall. (a) Overview of the organization of a typical plant cell, in which the organelle-filled cell with its plasma membrane is surrounded by a well-defined extracellular matrix called the cell wall. (b) Schematic representation of the cell wall of an onion. Cellulose (highly ordered glucose polymers) and hemicellulose (amorphous polysaccharide) are arranged into at least three layers in a matrix of the polysaccharide pectin and glycoproteins. The sizes of the polymers and their separations are drawn to scale. To simplify the diagram, most of the hemicellulose cross-links and other matrix constituents (e.g., glycoproteins such as extensin, complex organic polymers called lignins) are not shown. (c) Quick-freeze deep-etch electron micrograph of the cell wall of a garden pea in which some of the pectin molecules were removed by chemical treatment. The abundant thicker fibers are cellulose microfibrils, and the thinner fibers are hemicellulose cross-links (red arrowheads). [Part (c) republished with permission from Oxford University Press, from T. Fujino et al., 2000, “Characterization of Cross-Links Between Cellulose Microfibrils, and Their Occurrence During Elongation Growth in Pea Epicotyl,” Plant Cell Physiol. 2000, 41(4):486–494; permission conveyed through Copyright Clearance Center, Inc.]
The Plant Cell Wall, a Plant’s ECM, Is a Laminate of Cellulose Fibrils in a Matrix of Polysaccharides and Glycoproteins
Description The illustration labeled (a) shows a typical plant cell drawing with labels nucleus, vacuole, Golgi, chloroplast, cell wall, and plasmodesmata. The illustration labeled (b) shows a white arrow from the cell wall area of illustration (a) to the three-dimensional ribbon model. A dotted line cube is filled with cellulose microfibril as orange ribbons above the plasma membrane. The dotted line is labeled primary wall. Pectin is represented in green lines and hemicellulose is represented in black lines. The electron micrograph labeled (c) shows a plant cell wall with cellulose and red arrows mark the hemicellulose areas. The Plant Cell Wall, a Plant’s ECM, Is a Laminate of Cellulose Fibrils in a Matrix of Polysaccharides and Glycoproteins The plant cell wall, an extracellular matrix that is mainly composed of polysaccharides and is about 0.2 μm thick, completely coats the outside of the plant cell’s plasma membrane. This structure serves some of the same functions as the ECM produced by animal cells, even though the two structures are composed of entirely different macromolecules and have a different organization. About 1000 genes in the plant Arabidopsis, a small flowering plant (also called “thale cress”; see Chapters 1 and 7), are devoted to the synthesis and functioning of its cell wall, including approximately 414 glycosyltransferase genes, which encode enzymes that transfer specific sugar residues to proteins or polysaccharides, and more than 316 glycosyl hydrolase genes, which encode enzymes that degrade
sugar containing polymers. Similar to animal ECMs, the plant cell wall organizes cells into tissues, signals a plant cell to grow and divide, and controls the shapes of plant organs (morphogenesis). It is a dynamic structure that plays important roles in controlling the differentiation of plant cells during embryogenesis and growth, and it provides a barrier to protect against pathogen infection. Just as the ECM helps define the shapes of animal cells, the cell wall defines the shapes of plant cells. When the cell wall is digested away from plant cells by hydrolytic enzymes, spherical cells enclosed by a plasma membrane are left. Because a major function of the plant cell wall is to withstand the turgor pressure of the cell (between 14.5 and 435 pounds per square inch; as much as three orders of magnitude greater than in animal cells and about 13 times greater than in an automobile tire; see Chapter 11), the cell wall is built for lateral strength. It is arranged into layers of cellulose microfibrils: bundles of 18–36 parallel chains of extensively hydrogenbonded, long (as much as 4 μm or greater), linear polymers of glucose in β glycosidic linkages. The cellulose microfibrils are embedded in a matrix composed of pectin, a negatively charged polymer of D-galacturonic acid and other monosaccharides, and hemicellulose, a short, highly branched polymer of several five- and six-carbon monosaccharides. The mechanical strength of the cell wall depends on cross-linking of the microfibrils by hemicellulose chains (Figure 20-43b, c). The layers of microfibrils prevent the cell wall from stretching laterally. Cellulose microfibrils are synthesized on the exoplasmic face of the plasma membrane from UDPglucose and ADP-glucose formed in the cytosol. The polymerizing enzyme, called cellulose synthase, moves within the plane of the plasma
membrane along tracks of intracellular microtubules as cellulose is formed, providing a distinctive mechanism for intracellular-extracellular communication and ensuring that the cellulose microfibrils are oriented properly to permit cell wall, and thus whole-cell, growth. Unlike cellulose, pectin and hemicellulose are synthesized in the Golgi complex and transported to the cell surface, where they form an interlinked network that helps bind the walls of adjacent cells to one another and cushions them. When purified, pectin binds water and forms a gel in the presence of and borate ions — hence the use of pectins in many processed foods. As much as 15 percent of the cell wall may be composed of extensin, a glycoprotein that contains abundant hydroxyproline and serine. Most of the hydroxyproline residues are linked to short chains of arabinose (a five-carbon monosaccharide), and the serine residues are linked to galactose. Carbohydrate accounts for about 65 percent of extensin by weight, and its protein backbone forms an extended rodlike helix with the hydroxyl or O-linked carbohydrates protruding outward. Lignin — a complex, insoluble polymer of phenolic residues — associates with cellulose and is a strengthening material. Like cartilage proteoglycans, lignin resists compression forces. The cell wall is a selective filter whose permeability is controlled largely by pectins. Whereas water and ions diffuse freely across and through cell walls, the diffusion of large molecules, including proteins larger than 20 kDa, is limited. This limitation may explain why many plant hormones are small, water-soluble molecules, which can diffuse across the cell wall and interact with receptors in the plasma membrane of plant cells.
Loosening of the Cell Wall Permits Plant Cell Growth
Loosening of the Cell Wall Permits Plant Cell Growth Because the cell wall surrounding a plant cell prevents it from expanding, the wall’s structure must be loosened when the cell grows. The amount, type, and direction of plant-cell growth are regulated by small-molecule hormones called auxins. The auxin-induced weakening of the cell wall permits the expansion of the intracellular vacuole (see Figure 20-42a) by uptake of water, leading to elongation of the cell. We can grasp the magnitude of this phenomenon by considering that, if all cells in a redwood tree were reduced to the size of a typical liver cell, the tree would have a maximum height of only 1 meter, about a hundredfold less than normal. The cell wall undergoes its greatest changes at the meristem in a root or shoot tip. Meristems are where cells divide and grow, as described in
Chapter 22. Young meristematic cells are connected by thin, flexible primary cell walls, which can be loosened and stretched to allow subsequent cell elongation. Their major components are cellulose, pectin, and polysaccharides called xyloglucans. After cell elongation ceases, the cell wall is generally thickened, either by the secretion of additional macromolecules into the primary wall or, more usually, by the formation of a secondary cell wall composed of several layers. Cellulose, lignin, and polysaccharides called xylans and glucomannans are the major components in secondary cell walls. In mature tissues such as the xylem — the tubes that conduct salts and water from the roots through the stems
Plasmodesmata Directly Connect the Cytosols of Adjacent Cells
to the leaves — most of the cell eventually degenerates, leaving only the cell wall. The unique properties of wood and of plant fibers such as cotton are due to the molecular properties of the cell walls in the tissues of origin. Plasmodesmata Directly Connect the Cytosols of Adjacent Cells The presence of a cell wall separating cells in plants imposes barriers to cell-cell communication not faced by animals. One distinctive mechanism used by plant cells to communicate directly is specialized cell junctions called plasmodesmata, which are large tubes that extend through the cell wall (Figure 20-44) and were discovered in 1885. Like gap junctions, plasmodesmata can function as channels that directly connect the cytosol of a cell to that of an adjacent cell without a membrane barrier. Plasmodesmata appear to play an especially important role in protection from pathogens and in regulating the development of plant cells and tissues, as is suggested by their ability to mediate intracellular movement of transcription factors and ribonuclear protein complexes.
FIGURE 20-44 Plasmodesmata. (a) Schematic model of plasmodesmata, showing the desmotubule, an extension of the endoplasmic reticulum (ER), and the annulus, a plasmamembrane-lined channel filled with cytosol that interconnects the cytosols of adjacent cells. The regulated deposition of a glucose polymer called callose in the extracellular spaces in the cell wall adjacent to the entrances of the channels has the potential to block intercellular transport through the plasmodesmata, apparently by forcing the closing of the channels by narrowing the annulus. (b) Electron micrographs of thin sections of a sugarcane leaf (brackets indicate individual plasmodesmata). Left: Longitudinal view, showing the ER and desmotubule running through each annulus. Right: Perpendicular cross-sectional views of plasmodesmata, each with its annulus, desmotubule, and cytoplasmic sleeve. (c) Cells from the plant Arabidopsis were grown in culture, frozen, and, while cold, fixed with glutaraldehyde and stained with osmium tetroxide and uranyl acetate. Multiple views of one plasmodesma were visualized by tilting the sample in an electron microscope and a threedimensional tomographic model of the structure was computed. Shown is the electron micrograph of one view (black and white) with the colored model of its plasmodesma; plasmamembrane = yellow; ER and desmotubule membrane = blue; and spoke-like tethers connecting these membranes = red. The gap between the two membranes is the cytoplasmic sleeve.. [Part (b) republished with permission from Springer, from K. Robinson-Beers and R. F. Evert, 1991, “Fine Structure of Plasmodesmata in Mature Leaves of Sugarcane,” Planta 184(3):307–318; permission conveyed through Copyright Clearance Center, Inc. Part (c) republished with permission from Springer Nature, from W. J. Nicolas et al., 2017, “Architecture and Permeability of Post-Cytokinesis Plasmodesmata Lacking Cytoplasmic Sleeves.” Nature Plants 3:17082; permission conveyed through Copyright Clearance Center, Inc.] Description The illustration labeled (a) shows a plasma membrane and rough endoplasmic reticulum on both sides of the cell membrane, having moved through an annule. The E R is pink with red dots and the cell wall looks like Swiss cheese, with the holes being desmata. A desmotubule is labeled coming through one of the desmata. The fluorescence micrograph labeled (b) shows two views of the desmata. The first view shows cell 1 and cell 2 with a nanotube between two cells containing two endoplasmic reticulums. The second view shows little black circles with black dots in the centers.
The dots are labeled desmotubules and the circles are labeled cytoplasmic sleeve. Together, both parts are labeled annulus. The electron micrograph labeled (c) shows the plasma membranes of cell 1 and cell 2 highlighted in bright yellow, a bright blue line between them labeled desmotubule with E R in it moving from cell 1 to cell 2. Threespoke like tethers are labeled attached to the desmotubule and the plasma membrane. Although plasmodesmata and gap junctions appear to resemble each other functionally with respect to forming channels for direct diffusion of molecules from one cell’s cytosol to that of another, their structures differ dramatically in two significant ways. First, in animal-cell gap junctions, the membranes of the connected cells are distinct, and the cytosols are connected by a protein-lined tube of connexon hemichannels (see Figure 20-21). In plant plasmodesmata (see Figure 20-44), the plasma membranes of the adjacent plant cells merge to form a continuous single membrane that lines the wall of the channel, called the annulus, that connects the two cells. The diameter of the annulus varies from about 30–60 nm; its length can vary and may be greater than 1 μm. The lipid and protein composition of the specialized plasma membrane lining the annulus differs from that of the rest of the plasma membrane that surrounds the cell. Second, passing though the center of the plasmodesmata’s annulus is a narrow tube, called a desmotubule, that is an extension of endoplasmic reticulum (ER) from each of the connected cells. Between the desmotubule and the plasmodesmata’s specialized plasma membrane is a gap whose size is regulated and varies between 2–10 nm. This gap is called the cytoplasmic sleeve, through which molecules diffuse from the cytosol of one cell to that of the other. Filamentous actin runs throughout the length of the cytoplasmic sleeve. The close apposition of the plasma membrane and
ER/desmotubule’s membrane in plasmodesmata is an example of a specialized membrane contact site (MCS), such as that described for mitochondria and ER in Chapter 12. A set of transmembrane proteins in the desmotubule called MCTPs can span the gap of the cytoplasmic sleeve and bind to anionic phospholipids on the plasma membrane, thus tethering these two membranes. The MCTP proteins appear to compose spoke-like projections observed by electron microscopy that tether the two membranes (see Figure 20-44c), help determine the size and properties of the cytoplasmic sleeve, and thus control the movement of molecules through plasmodesmata. In animal cells, tunneling nanotubes resemble plasmodesmata in that they are membrane-lined channels that permit molecules to flow between cells (see Section 20.2). However, unlike plasmodesmata, there is no evidence for ER passing through tunneling nanotubes. There are simple plasmodesmata (with a single pore, like those in Figure 20-44) and complex plasmodesmata that branch into multiple channels. The density of plasmodesmata varies depending on the plant and cell type, and even the smallest meristematic cells have more than a thousand connections with their neighbors. Proteomic analysis suggests that there are more than 115 different proteins that are concentrated in plasmodesmata and play important roles in determining their structures and functions. These include the MCTPs, actin-binding proteins (e.g., formin and the adaptor protein NET1A), a variety of receptor-like proteins (e.g., kinases), and enzymes.
The Molecules That Plants Depend on for Adhesion and Mechanotransduction Differ from Those in Animals
Many types of molecules travel from cell to cell through plasmodesmata. Molecules smaller than about 1000 Da, including a variety of metabolic and signaling compounds (ions, sugars, amino acids), can generally diffuse freely through the cytoplasmic sleeve, whose size and properties are highly regulated. In some circumstances, the channel is clamped shut; in others, it is dilated sufficiently to permit the passage of molecules larger than 10,000 Da, including some transcription factors, nucleic acid/protein complexes, metabolic products, and even plant viruses. Some of these require special chaperones to facilitate transport. Soluble molecules pass through the cytosolic sleeve, whereas membrane-bound molecules or certain proteins within the ER lumen may pass from cell to cell via the desmotubule. The deposition and breakdown of a glucose polymer called callose in the extracellular spaces adjacent to the entrances of the channels (see Figure 20-44a) is thought to regulate the closing and opening of the channels, respectively. Specialized kinases may also phosphorylate plasmodesmal components to regulate their activities (e.g., opening of the channels). Among the factors that affect the permeability of plasmodesmata is the cytosolic concentration: an increase in cytosolic reversibly inhibits movement of molecules through these structures. The Molecules That Plants Depend on for Adhesion and Mechanotransduction Differ from Those in Animals
As noted earlier in this section, there are dramatic differences in the organization of cells into tissues in plants and animals. These differences have determined the distinctive molecular mechanisms by which plant cells are incorporated into tissues and communicate with one another, including their cell-cell and cell-matrix interactions. Plant Adhesion Molecules Systematic analyses of the Arabidopsis genome and biochemical analyses of other plant species have provided no evidence for the existence of plant homologs of most animal CAMs, adhesion receptors, and ECM components; plants use other adhesive molecules for their cell-cell and cell-matrix interactions. Currently, our understanding of cell-matrix adhesion in plants is less advanced than our understanding of cell-matrix adhesion in animals. Because plant cells are surrounded by thick cell walls, their assembly into tissues is primarily determined by cell-matrix interactions. Most of the polysaccharides in the cell wall (cellulose, hemicellulose, pectins, glycoproteins) can cross-link to one another. The adhesive properties of pectins are thought to be especially important for determining both the physical characteristics of cell walls and the interactions of the walls with the plasma membranes of the underlying cells. Disruption of the gene encoding glucuronyltransferase 1, a key enzyme in pectin biosynthesis, dramatically interferes with the synthesis of specialized pectins that help hold the cells in meristems tightly together. As a consequence, the mutation prevents normal cell adhesion and differentiation into
photosynthetic cells. In vitro binding assays, combined with in vivo studies and analyses of plant mutants, have identified other macromolecules in the ECM that are important for specialized types of adhesion. For example, a cysteine-rich protein called stigma/stylar cysteine-rich adhesin (SCA) and a specialized pectin that binds to SCA mediate normal adhesion of pollen, which contains sperm cells, to the stigma or style in the female reproductive organ of the Easter lily. A number of plasma membrane proteins exhibit the potential to bind to cell wall polysaccharides, particularly pectins, and thus mediate cellmatrix adhesion. These include receptor-like proteins, such as WallAssociated Kinases (WAKs), GPI-anchored proteins, Hydroxyproline-Rich Glycoproteins (HRGPs), and Glycosyl Inositol Phosphoceramides (GIPCs). For example, five WAKs and WAK-like proteins are expressed in the plasma membrane of Arabidopsis cells. These transmembrane proteins have a cytoplasmic serine/threonine kinase domain, and their extracellular regions contain multiple epidermal growth factor (EGF) repeats, frequently found in animal cell-surface receptors. Some WAKs have an extracellular pectin-binding domain that can recognize and bind fulllength pectin and pectin degradation fragments. Such binding has been proposed to help cells monitor and respond to the status of the cell wall during normal growth and in the context of cell-wall damage (wounding) or infection by pathogens. Thus some WAKS in plant cells appear to be analogous to adhesion receptors in animal cells, binding and sensing the ECM and mediating outside-in signaling. Mechanosensation in Plants
As with animal cells, plant cells are subject to a variety of mechanical forces (stresses and strains), both internal (e.g., turgor pressure, cytoskeletal forces) and external (e.g., wind), which they sense (mechanosensing) and to which they respond (e.g., alterations in cytoskeletal organization, calcium flux, gene expression, wood formation, cell wall stiffness, and extent and direction of growth). Possible mechanosensors in the cell wall include components whose structures respond to changes in mechanical forces and thus change the properties of the wall and its interactions with the plasma membrane. Potential mechanosensors in the cell’s plasma membrane are those components that bind to the cell wall (e.g., receptor-like kinases such as pectin-binding WAKs) and mechanosensitive ion channels. There is evidence that suggests inside-out and outside-in communication via adhesive molecules contribute to plant responses to mechanical forces. One well-known example of macroscopic plant mechanosensation is the Venus flytrap (Dionaea muscipula), first described by the colonial governor of North Carolina, Arthur Dobbs, in 1759. Charles Darwin studied the Venus flytrap extensively, writing a book in 1875 in which he called the plant “one of the most wonderful in the world” and once noting that he cared more for one of them “than the origin of all the species in the world.” The Venus flytrap detects the presence of an insect on its leaf when an insect hits one of its sensory hairs, which are around 200 μm in diameter at the base and 2 mm long. The sensory hair is a multicellular organ that, when bent by prey, puts shear stress on a cluster of cells at its base, which opens mechanosensitive calcium channels. Calcium entry
triggers an action potential in the leaf (see Chapter 23) that leads to closing the trap and hormonally induced secretion of digestive enzymes that disassemble the prey into basic nutrients over a one-to-two-week period. We are all familiar with one aspect of cell-wall–mediated adhesion in plants: the softening of fruits, such as tomatoes and strawberries, during ripening. The ripening of fruits is a complex process that influences color, aroma, flavor, nutrient content, and texture (including softening and water content). Ripening of seed-bearing fruit is advantageous because it encourages fruit-eating animals to eat and disperse the seeds or breaks down the uneaten fruit’s structure (softening and decay) to release the seeds in situ. However, excessive softening can interfere with successful storage and transport and can reduce shelf life. Expression of a group of cell-wall–degrading enzymes, including pectate lyase (PL), and their remodeling of cell walls by degrading pectin plays a key role in determining the rate and extent of softening. Experiments using both strawberries and tomatoes have suggested that inhibiting the expression of pectate lyase using either CRISPR gene editing or RNA interference (see
Chapter 6) can substantially delay strawberry and tomato softening and may not block other desirable consequences of ripening (changes in color, flavor, etc.). In the future, targeted genetic modifications of cell-wall metabolism may improve the quality of available fruits while reducing the costs of production and distribution. KEY CONCEPTS OF SECTION 20.6
Plant Tissues The organization of cells into tissues in plants is fundamentally different from the assembly of animal tissues, primarily because each plant cell is surrounded by a relatively rigid cell wall. The plant cell wall comprises layers of cellulose microfibrils embedded within a matrix of hemicellulose, pectin, extensin, and other less abundant molecules. Cellulose, a large, linear glucose polymer, assembles spontaneously into microfibrils stabilized by hydrogen bonding. The cell wall defines the shapes of plant cells and restricts their elongation. Auxininduced loosening of the cell wall permits cell elongation. Adjacent plant cells can communicate through plasmodesmata, junctions formed by tubes of specialized plasma membrane and ER that connect the cytosols of adjacent cells, and by allowing molecules to pass between cells (see Figure 20-44). Plants do not produce homologs of the common adhesion molecules found in animals. The molecules mediating cell-matrix adhesion in plants are not as well-defined as those in animals, although both cell wall and plasma membrane molecules implicated in plant cell adhesion have been identified and are under investigation. As in the case of animal cells, mechanotransduction in plants plays an important role in maintaining tissue integrity and directing differentiation and development.
Key Terms
End of Chapter Visit Achieve to access study tools and to learn more about the content in this chapter. Perspectives for the Future Analyze the Data Chapter References Additional study tools, including videos, animations, and quizzes Key Terms adapter proteins adherens junction adhesion receptor anchoring junction apical basal basal lamina basolateral cadherin cell-adhesion molecule (CAM) or cell-cell adhesion receptor cell junction cell-matrix adhesion receptor cell wall cellulose
cis (lateral) interactions classical cadherin clustered protocadherin collagen connective tissue connexin desmosomal cadherin desmosome elastin epithelium epithelial-to-mesenchymal transition extracellular matrix (ECM) fibrillar collagen fibroblast fibronectin focal adhesion/focal contact gap junction glycosaminoglycan (GAG) heterophilic binding heterotypic adhesion homophilic binding homotypic adhesion hyaluronan inside-out signaling integrin isoform laminin lateral
Review the Concepts
matriglycan matrix metalloproteases (MMPs) mechanosensor mechanotransduction multi-adhesive matrix protein N-linked oligosaccharide O-linked oligosaccharide outside-in signaling paracellular pathway PDZ domain perlecan plasmodesmata proteoglycan RGD motif selectin tight junction talin trans (intercellular) interactions transcellular pathway Review the Concepts 1. Describe the two phenomena that give rise to the diversity of adhesion molecules, such as cadherins. What additional phenomenon gives rise to the diversity of integrins? 2. Cadherins are known to mediate homophilic interactions between cells. What is a homophilic interaction, and how can it
be demonstrated experimentally for E-cadherins? What component of the extracellular environment is required for the homophilic interactions mediated by cadherins, and how can this requirement be demonstrated? 3. Together with their role in connecting the lateral membranes of adjacent epithelial cells, adherens junctions play a role in controlling cell shape. What associated intracellular structure and proteins are involved in this role? 4. What is the normal function of tight junctions? What can happen to tissues when tight junctions do not function properly? 5. Gap junctions between cardiac muscle cells and gap junctions between uterine smooth-muscle cells form connections that provide for rapid communication. What is this phenomenon called? How is communication among uterine smooth-muscle cells up-regulated for parturition (childbirth)? 6. What is collagen, and how is it synthesized? How do we know that collagen is required for tissue integrity? 7. Explain how changes in integrin structure mediate outside-in and inside-out signaling. 8. Compare the functions and properties of each of three types of macromolecules that are abundant in the ECM of all tissues. 9. Many proteoglycans have signaling roles. Regulation of feeding behavior by syndecans in the hypothalamic region of the brain is one example. How is this regulation accomplished? 10. You have synthesized an oligopeptide containing an RGD motif surrounded by other amino acids. What is the effect of this peptide when added to a fibroblast cell culture grown on a layer
of fibronectin adsorbed to the tissue culture dish? Why does this happen? 11. Describe the major activity and possible localization of the three major subgroups of proteins that remodel or degrade the ECM in physiological or pathological tissue remodeling. Identify a pathological condition in which these proteins play a key role. 12. Blood clotting is a crucial function for mammalian survival. How do the multi-adhesive properties of fibronectin lead to the recruitment of platelets to blood clots? 13. How do changes in molecular connections between the ECM and the cytoskeleton give rise to Duchenne muscular dystrophy? 14. To fight infection, leukocytes move rapidly from the blood into sites of infection in the tissues. What is this process called? How are adhesion molecules involved in this process? 15. The structure of a plant cell wall needs to loosen to accommodate cell growth. What signaling molecule controls this process? 16. Compare plasmodesmata in plant cells with gap junctions and tunneling nanotubes in animal cells.