3 Microbial Metabolism
III Catabolism: Electron Transport and Metabolic Diversity
Life Begins with Metabolism
Metabolism is the foundation upon which life is formed. Microorganisms have evolved tremendous metabolic diversity, but all microbes in their vast diversity have a common set of requirements. Life requires liquid water, a source of energy to do work, a source of electrons to perform biochemical reactions, and nutrients required to build macromolecules. Once we understand these requirements, we can begin to make predictions about how life behaves in any environment, whether a caustic hypersaline pond, inside a rock from the cold deserts of Antarctica, in a hydrothermal vent chimney from the ocean’s depths, or even on another world. In short, the search for life begins with an understanding of metabolism.

Water is common in our solar system. There is water on Mars and on Earth’s moon, on asteroids and comets, and there is evidence for water on several other planets and moons. Liquid water, however, is harder to find. One place that has liquid water is the moon Enceladus, which orbits Saturn. Enceladus has an ice-covered ocean with cracks in its surface through which erupt geysers that spew materials into space (see photo). The Cassini spacecraft photographed and flew through these plumes, detecting silicate minerals that form only in the presence of liquid water.
The gravity from Saturn squeezes Enceladus, resulting in volcanic activity and liquid water beneath its icy crust. These plumes also contain diverse organic and inorganic compounds and the gases H2, CO2, and methane (CH4). Taken as a whole, these molecules are sufficient to provide free energy, a source of electrons, and nutrients required to support life. Indeed, microbes could likely survive at the hydrothermal vents thought to exist on Enceladus. We now know that all of the fundamental requirements for life are present on other worlds, and the search for extraterrestrial life is a search for microbial life.
Source: Waite, J.H., et al. 2017. Cassini finds molecular hydrogen in the Enceladus plume: Evidence for hydrothermal processes. Science 356: 155.
Metabolism is the series of biochemical reactions needed to sustain life. Metabolism includes catabolism—reactions used to obtain energy and break down complex molecules—and anabolism—reactions used to synthesize cellular material (Figure 3.1). Microorganisms have evolved tremendous metabolic diversity through which they influence all aspects of our biosphere. In this chapter, we will learn the unifying concepts of metabolism that underlie all living systems. In particular, we will examine the manner in which cells obtain energy and how they use this energy to synthesize the building blocks of macromolecules and cells (Figure 3.1a). We will also see that metabolism relies on the directed movement of electrons from an electron donor to an electron acceptor (Figure 3.1b). We will revisit these principles of metabolism in Chapter 6, where we explore the biosynthesis of macromolecules—the nucleic acids and proteins—and in Chapter 14, where the enormous metabolic diversity of the microbial world unfolds.
Figure 3.1 Metabolic coupling with respect to energy conservation and electron flow.

(a) Catabolism employs exergonic reactions to drive the synthesis of ATP. Anabolism employs endergonic reactions, which consume ATP, to drive the biosynthesis of cellular material. Some energy would be lost as heat and cannot be conserved in the formation of ATP (not shown). (b) Cells require reducing power, in the form of a reduced electron donor, as a source of electrons (e−) needed to carry out anabolic and catabolic reactions. Inputs to metabolism are labeled in {green} and outputs of metabolism are labeled in {yellow}.
I Fundamentals of Metabolism
Bioenergetics describes the transformation of energy during metabolism characterized by electron flow within the cell. Metabolism uses the energy currency of ATP to couple the exergonic reactions of catabolism to the endergonic reactions of anabolism.
According to the first law of thermodynamics, energy is neither created nor destroyed. Hence, in order to grow, cells must conserve energy by converting energy available from their surroundings into a form that can do work. Cells accomplish this by generating energy-rich compounds such as adenosine triphosphate (ATP)—a molecule capable of storing energy and releasing it to fuel cellular processes (Figure 3.1).
3.1 Defining the Requirements for Life
All living cells share certain fundamental metabolic requirements. All cells require water in which to perform metabolic reactions, as well as sources of carbon and other nutrients with which to synthesize cellular materials. All cells also require free energy—the energy available to do work—and reducing power—a source of electrons (e−) that can be used to both generate free energy and perform certain biosynthetic reactions (Figure 3.1). We will review the nutrient requirements of cells in Chapter 4. Here we will learn to categorize and understand all forms of metabolism in terms of free energy and reducing power.
Free Energy
All chemical reactions can be described in terms of their relationship to energy. Some chemical reactions release energy as they proceed, while others require an input of energy to proceed. Consider the reaction for the aerobic respiration of glucose: C6H12O6+6 O2⇌6 CO2+6 H2O
In this reaction the reactants are on the left (C6H12O6+6 O2) and the products are on the right (6 CO2+6 H2O). The change in free energy during a reaction is expressed as ΔG0′, where the symbol Δ is read as “change in.” The “0” and “prime” in ΔG0′ indicate that the free-energy value is for standard conditions: pH 7 (approximate cytoplasmic conditions), 25∘C, 1 atmosphere of pressure, and all reactants and products at molar concentrations. The term ΔG0′ is expressed in units of kilojoules/mole (abbreviated as kJ/mol), a unit of heat energy. If the ΔG0′ for a reaction is negative in arithmetic sign, then the reaction will proceed with the release of free energy and such reactions are exergonic. However, if ΔG0′ for the reaction is positive, the reaction requires energy in order to proceed and such reactions are endergonic. Thus, exergonic reactions release free energy whereas endergonic reactions require free energy. For example, the aerobic respiration of glucose provides a free-energy change of ΔG0′=−2895 kJ/mol of glucose. Hence, this reaction is exergonic and it provides a substantial amount of free energy that can be used by the cell to do work. We will learn how to calculate the free-energy change of a reaction in Section 3.3.
Catabolic pathways are exergonic processes in which cells generate free energy by transforming reactants into products (Figure 3.1a). Free energy is energy available to do work. The free energy produced in catabolism is conserved by synthesizing energy-rich molecules such as ATP (Figure 3.1a). The formation of ATP requires at least ΔG0′=−31.8 kJ/mol. Hence, the aerobic respiration of a mole of glucose could produce up to 91 moles of ATP under standard conditions, though under natural cellular conditions this reaction actually produces closer to 38 moles of ATP. This difference in ATP yield occurs because reactions in the cell do not occur under standard conditions. We will see in Section 3.3 that the free energy available under natural conditions differs from the free energy calculated at standard conditions. In addition, chemical reactions release some portion of energy in the form of heat, which cells cannot conserve in the formation of ATP. The heat lost during metabolic reactions is what makes your body warm and what makes a decomposing compost pile become steaming hot.
Anabolic pathways are endergonic processes in which the synthesis of cellular material from simple precursors requires an input of energy (Figure 3.1a). The energy required to fuel anabolic reactions, and to biosynthesize cellular materials, comes from the hydrolysis of ATP. In this way, catabolic and anabolic reactions are fundamentally linked (Figure 3.1a).
Reducing Power
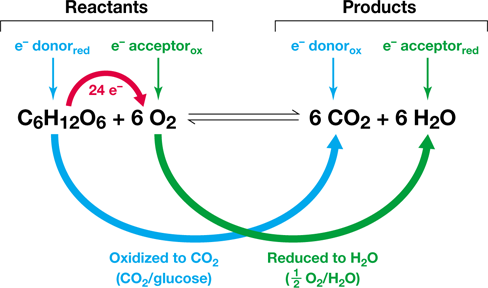
Reducing power is the ability to donate electrons during electron transfer reactions. Electron transfer reactions, called redox reactions, are comprised of two half reactions. During redox reactions, electrons are transferred from an electron donor in one half reaction to an electron acceptor in a second half reaction. When an electron is added to a substance, we say that the substance has been reduced because the addition of an electron reduces the oxidation state of the substance. In contrast, when an electron is removed from a substance we say it has been oxidized (Figure 3.1b). Consider again the aerobic respiration of glucose (Figure 3.2). In this example, which is a redox reaction, glucose is the electron donor and O2 is the electron acceptor. Electrons are transferred from glucose to O2, resulting in the oxidation of glucose to CO2 and the reduction of O2 to H2O (Figure 3.2). Redox reactions are an essential component of catabolism, and we will see that they are integral to energy conservation within the cell.
Figure 3.2 Example of an oxidation–reduction reaction.

The oxidation of glucose to CO2 is coupled with the reduction of O2 to H2O in aerobic chemoorganotrophic organisms. This redox reaction is composed of two half reactions: CO2/glucose and 12 O2/H2O (see Figure 3.4). The overall reaction is balanced, which means that equal amounts of each element (C, H, O) are present in reactants and products. The complete oxidation of one glucose to 6 CO2 donates 24 electrons, which are accepted by 6 O2.
Reducing power is also required by anabolic reactions (Figure 3.1b). The biosynthesis of cellular materials from simple precursors requires both free energy, in the form of ATP, and reducing power, in the form of electron carriers that transfer electrons to anabolic reactions. We will learn more about the metabolic significance of electron carriers and redox reactions in Section 3.2. First, we consider how different organisms obtain the free energy they need to drive ATP synthesis.
Metabolic Classes of Microorganisms
All types of metabolism can be classified based on their source of energy (Figure 3.3). Phototrophs obtain energy for metabolism from light. Plants are one type of phototroph, but we will learn that many different types of phototrophic metabolism exist in the microbial world (see Section 3.11 and Chapters 14 and 15). Chemotrophs obtain energy for metabolism from chemical reactions (Figure 3.3). The aerobic respiration of glucose is an example of chemotrophic metabolism in that free energy comes from the chemical oxidation of glucose to CO2 (Figures 3.2 and 3.3). Chemotrophic reactions are classified as aerobic if they require O2 as an electron acceptor, but they are classified as anaerobic if their electron acceptor is anything other than O2 (see Section 3.10). Chemotrophs can conserve energy from either respiration reactions or fermentation reactions (see Sections 3.7, 3.8 and 3.9). For example, in the presence of O2, yeast can perform aerobic respiration of glucose to CO2, but when O2 is limiting they alter their metabolism to perform anaerobic fermentation of glucose to ethanol and CO2.
Figure 3.3 Classification of metabolic types based on energy sources.

Microorganisms can obtain energy from organic chemicals, inorganic chemicals, or light. Most organisms employ only one type of metabolism, but some microbes can use different energy sources depending on environmental conditions.
The ultimate source of energy for chemotrophic organisms can be organic or inorganic. Organic molecules are those molecules that contain carbon, excluding certain carbon-containing gases and minerals considered inorganic (for example, compounds such as CO2, CO, carbonates, cyanides, diamond, and graphite are considered inorganic). Organisms that obtain their energy and reducing power from organic molecules are called chemoorganotrophs while those that obtain their energy and reducing power from inorganic molecules are called chemolithotrophs (Figure 3.3). Most microorganisms in laboratory culture, and most pathogenic bacteria, use carbohydrates or proteins as their energy source and are thus chemoorganotrophs. Chemolithotrophs, by contrast, are important in many biogeochemical cycles. Electron donors for chemolithotrophs include gaseous hydrogen (H2), hydrogen sulfide (H2S), ammonia (NH3), and ferrous iron (Fe2+). Related groups of chemolithotrophs typically specialize in the oxidation of a group of similar inorganic compounds, and thus we have the “sulfur” bacteria, the “iron” bacteria, the “nitrifying” bacteria, and so on (see Chapters 14 and 15).
Phototrophs contain chlorophylls and other pigments that convert light energy into ATP and thus, unlike chemotrophs, do not require chemicals as a source of energy (Section 3.11). Two forms of phototrophy exist: oxygenic photosynthesis and anoxygenic photosynthesis. Oxygenic photosynthesis, in which O2 is produced, is characteristic of cyanobacteria (Section 15.3) and is also carried out by plants and algae. Anoxygenic photosynthesis (Section 14.5), in which O2 is not produced, predates oxygenic photosynthesis, and it occurs in diverse lineages of Bacteria. We explore the diversity of phototrophs further in Chapters 14 and 15.
Regardless of how a microorganism conserves energy, be it from chemicals or from light, all cells require large amounts of carbon in one form or another to make new cell materials. A heterotroph obtains carbon for biosynthesis from an organic compound; nearly all chemoorganotrophs are also heterotrophs. An autotroph, by contrast, uses carbon dioxide (CO2) as its carbon source, reducing it to cell material at the expense of ATP (Section 3.12). Most chemolithotrophs and phototrophs are autotrophs. Autotrophs are also called primary producers because they synthesize new organic matter from inorganic carbon (CO2). Virtually all organic matter on Earth has been synthesized by primary producers, in particular, by phototrophs.
We now expand on this introduction to metabolism and energy by emphasizing electron transfer reactions, energy-rich compounds, and enzymes in the biochemical workings of a cell.
Check Your Understanding
What is free energy and how is it used by the cell?
What is reducing power and what is its relationship to electron donors and acceptors?
How does a chemoorganotroph differ from a chemolithotroph? A chemotroph from a phototroph?
3.2 Electron Transfer Reactions
Catabolism, and indeed life itself, depends on the directed flow of electrons from an electron donor to an electron acceptor during redox reactions (Figure 3.1b). Redox reactions are also required in many biosynthetic reactions that occur during anabolism (Sections 3.12, 3.13, 3.14 and 3.15). Redox reactions can be understood in terms of reduction potential (Figure 3.4), which measures the affinity of a substance for electrons.
Figure 3.4 The redox tower.
Redox couples are arranged from the strongest electron donors at the top to the strongest electron acceptors at the bottom. The greater the difference in reduction potential between electron donor and electron acceptor, the greater the free energy released. Note the differences in free-energy yield when glucose reacts with either O2 (1) or nitrate (2).
Redox Reactions and Reduction Potentials
Although life requires the transfer of electrons within the cell, electrons cannot exist stably in solution. Therefore, electrons must be transferred directly from one atom or molecule to another during redox reactions. Redox reactions occur in pairs, called half reactions (also called redox couples). A half reaction depicts the conversion of one substance into another caused by adding or removing electrons. By convention, in writing a half reaction, the oxidized form of a substance is always placed on the left (before the forward slash) followed by the reduced form after the forward slash (Figure 3.4). For example, the addition of an electron to nitrate (NO3 −) causes it to be reduced to nitrite (NO2 −) and so we write this redox couple as NO3 −/NO2 − (Figure 3.4).
In a redox reaction the first half reaction produces one or more electrons that are then consumed by the second half reaction. The reactant in the first half reaction donates electrons and is oxidized, whereas the reactant in the second half reaction accepts these electrons and is reduced. In redox reactions of this type, we refer to the reactant that is oxidized as the electron donor, and the reactant that is reduced is the electron acceptor. Consider the redox reaction between glucose and O2 (Figure 3.2). The two half reactions that contribute to this overall reaction are CO2/glucose and 12 O2/H2O (Figure 3.4). In this reaction, glucose is oxidized to CO2, causing the reduction of O2 to H2O. Both glucose and O2 are reactants in the overall reaction with glucose serving as electron donor and O2 serving as electron acceptor. The products of this reaction are CO2 and H2O. Many different redox couples exist in nature (Figure 3.4 and Table 3.1) and can participate in diverse redox reactions within the cell.
Table 3.1 Reduction potentials, E0′ (volts), of some redox half reactions commonly encountered in microbiologya

aEntries are read as: oxidized form/reduced form (E0′ in volts) and number of e− transferred per half reaction. Values are organized from lowest (most electronegative) to highest (most electropositive) reduction potential.
All redox couples have the ability to either donate or accept electrons to another redox couple. Within each redox couple, the reduced substance (which occurs after the forward slash) is the chemical form that donates electrons, and the oxidized substance (which occurs before the forward slash) is the form that accepts electrons. Whether a substance is an electron donor or an electron acceptor in a given redox reaction is determined by the reduction potential (E0′ at standard conditions) of the participating half reactions (Figure 3.4 and Table 3.1). Reduction potentials are measured in volts (V) compared to a reference substance (typically, the reference is H2). Electrons are negatively charged and so the reduced substance in a redox couple will have a strong tendency to donate electrons when the redox couple has a negative reduction potential (for example, glucose in the CO2/glucose couple, E0′=−0.43 V). In contrast, the oxidized substance in a redox couple will have a strong tendency to accept electrons when the redox couple has positive reduction potential (for example, O2 in the 12 O2/H2O couple, E0′=+0.82 V). Ultimately, the tendency of a substance to donate or accept electrons in a redox reaction is determined by the difference in reduction potential between the two participating redox couples. In a given redox couple the substance on the right is the chemical form that serves as an electron donor in a redox reaction, while the substance on the left is the chemical form that serves as an electron acceptor.
By convention, half reactions are written as reductions (as in Figure 3.4 and Table 3.1). However, when two half reactions are combined into a single redox reaction the half reaction that donates electrons (that is, the half reaction with the more negative E0′) proceeds as an oxidation, and its orientation is therefore written in reverse (as an oxidation) in the overall redox reaction. For example, the CO2/glucose couple has a far more negative reduction potential than the 12 O2/H2O couple, a difference of 1.25 V (Figure 3.4). Hence, in the redox reaction that includes these two couples, glucose is the electron donor and its half reaction is written as an oxidation, with the net reaction having the reactants glucose and O2 and the products CO2 and H2O (Figures 3.2 and 3.4).
When aerobic respiration is prevented by the absence of O2, some organisms can carry out anaerobic respiration using an alternative electron acceptor, such as nitrate (see Section 3.10). Nitrate (NO3 −) is the oxidized substance in the NO3 −/NO2 − redox couple (E0′=0.42 V). The CO2/glucose couple has a more negative reduction potential than the NO3 −/NO2 − couple, a difference of 0.85 V (Figure 3.4). Hence, glucose will serve as a favorable electron donor with nitrate as the electron acceptor (see Figure 3.4 for the balanced reaction). The positive difference in reduction potential of the two half reactions indicates that this will be a favorable reaction, though because there is less difference in reduction potential (0.85 V relative to 1.25 V), the anaerobic respiration of glucose using NO3 − as electron acceptor yields less energy than aerobic respiration (Figure 3.4).
In this way, reduction potentials can be used to determine whether any given substance in a half reaction will serve as electron donor or electron acceptor in an overall redox reaction. In addition, the greater the difference in the reduction potentials of the two half reactions in a redox couple, the more energy will be available to the cell. A vast diversity of redox couples exist in nature, allowing for a tremendous diversity of redox reactions (Figure 3.4 and Table 3.1). We will see that microbes have evolved to harness energy from a large number of redox reactions, giving rise to the astounding metabolic diversity present in the microbial world (Chapters 14 and 15). We will learn in Section 3.3 how to use the difference in reduction potential between two half reactions to calculate the overall free-energy yield of a reaction.
Electron Carriers and NAD+/NADH Cycling
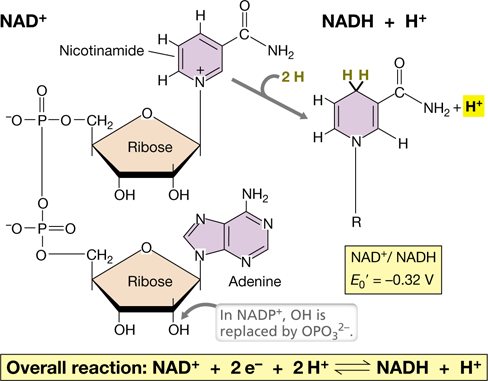
Cells require a net movement of electrons from an electron donor to an electron acceptor, but such reactions are rarely performed in a single step. More often, the movement of electrons from electron donor to electron acceptor proceeds through a series of consecutive reactions at different locations within the cell. Hence, the cell needs soluble electron carriers such as nicotinamide adenine dinucleotide (NAD+/NADH) to carry electrons from one place to another within the cell (Figure 3.5). NAD+/NADH is a redox couple with a reduction potential of −0.32 V, which makes NADH a good electron donor and NAD+ a weak electron acceptor (Figure 3.4). The reduction of NAD+ to NADH requires 2 e− and 1 H+, but the oxidation of electron donors typically results in the production of 2 e− and 2 H+. Therefore, the reduction of NAD+ typically results in the production of NADH+H+ with the extra proton released into solution (Figure 3.5).
Figure 3.5 The redox coenzymes nicotinamide adenine dinucleotide (NAD+) and NADP+.

NAD+ undergoes oxidation–reduction as shown and is freely diffusible. “R” is the adenine dinucleotide portion of NAD+; H is hydrogen (consisting of H++e−).
Electron carriers such as NAD+/NADH are common coenzymes (see Section 3.5) in cells. The use of electron carriers as coenzymes increases the diversity of redox reactions that are possible in a cell by allowing many different electron donors and acceptors to interact. For example, an enzyme can remove electrons from an electron donor and use them to reduce NAD+ to NADH (Figure 3.6). The NADH then diffuses away from the enzyme and attaches to a different enzyme that oxidizes NADH back to NAD+ when it reduces an electron acceptor (Figure 3.6).
Figure 3.6 NAD+/NADH cycling.

A schematic example of redox reactions in which two different enzymes are linked by their requirement for either NAD+ or NADH. One enzyme reduces NAD+ in its reaction while the other enzyme oxidizes NADH in its reaction.
Mastering Microbiology
Electron shuttling mediated by NAD+/NADH is common in microbial catabolism. However, in addition to NAD+/NADH, many other molecules may participate as electron shuttles. For example, nicotinamide adenine dinucleotide phosphate (NADP+) is made from NAD+ by adding a phosphate molecule (Figure 3.5). We will see that NADP+/NADPH participates in many anabolic biosynthetic reactions, whereas NAD+/NADH typically participates in catabolic redox reactions.
Check Your Understanding
In the reaction 2 H2+O2⇌2 H2O, what is the electron donor and what is the electron acceptor?
Which of the following can accept electrons from NADH: CO2, lactate, or pyruvate? Explain your reasoning.
Can both glucose and H2S be used to reduce NAD+? Why or why not?
3.3 Calculating Changes in Free Energy
We have seen that cells conserve energy from chemical reactions or from light, and they transfer electrons from electron donors to electron acceptors by using electron carriers such as NADH. Ultimately, in order to conserve energy, cells need to synthesize energy-rich molecules such as ATP. We will see that numerous (but not all) electron transfer reactions are sufficiently exergonic to produce ATP and thus can support a vast diversity of microbial metabolisms. However, to better understand this diversity in metabolism, we once again return to the concept of reduction potential and this time place it in the context of free-energy change.
The Redox Tower and Its Relationship to ΔG0′
A convenient way of viewing the energy available in electron transfer reactions is to imagine a vertical tower that represents the entire range of reduction potentials possible for redox couples in nature, from those with the most negative E0′ on the top to those with the most positive E0′ at the bottom; this is a redox tower (Figure 3.4). Now imagine electrons from an electron donor near the top of the tower falling and being “caught” by electron acceptors at lower levels. The difference in reduction potential between the donor and acceptor redox couples is expressed as ΔE0′, the change in reduction potential. For example, we have seen that the oxidation of glucose with O2 as electron acceptor yields far more energy than when nitrate (NO3 −) is the electron acceptor (Section 3.2 and Figure 3.4).

The further an electron drops before it is caught by an acceptor, the greater is the ΔE0′ between the two redox couples and the greater is the amount of energy released in the net reaction. That is, ΔE0′ is proportional to ΔG0′ (Figure 3.4). This relationship is expressed more precisely in the equation ΔG0′=−nFΔE0′, where n is the number of electrons transferred and F is the Faraday constant (96.5 kJ/V). Hence, we can see that redox reactions provide a source of energy to the cell and that the greater the difference in reduction potential between electron donor and electron acceptor, the greater the energy available to the cell. This is illustrated for an example reaction, the oxidation of acetate to CO2, in equation 1 of Table 3.2.
Table 3.2 Example of free-energy-change calculations using electrochemical potentials or G f 0 values

aThe reaction is balanced and is an 8-electron oxidation (n=8 in equation 2). Gf 0 values were taken from Table 3.3.
bF is the Faraday constant (96.5 kj/V) and ΔE0′ is calculated from the E0′ values in Figure 3.4 and Table 3.1.
Calculating ΔG0′ from the Free Energy of Formation
There is a second way to calculate ΔG0′ if one knows the free energy of formation (Gf 0) of the reactants and products in the reaction. The free energy of formation is equal to the energy released or required during the formation of a given molecule from its constituent elements. Table 3.3 lists the Gf 0 for a few common substances. By convention, the free energy of formation of the elements in their elemental and electrically neutral form (for instance, C, H2, N2) is zero. For most compounds, Gf 0 is negative. This reflects the fact that compounds tend to form spontaneously (that is, with a free-energy release) from their elements. However, the positive Gf 0 for nitrous oxide (N2O) (Table 3.3) indicates that this compound does not form spontaneously. Instead, over time it decomposes spontaneously to yield N2 and O2. The compounds listed in Table 3.3 are only a small subset of free energy of formation values available from physical chemistry reference sources.
Table 3.3 Free energy of formation (Gf 0, kJ/mol) for some common substancesa

aValues for free energy of formation taken from Speight, J. 2005. Lange’s Handbook of Chemistry, 16th edition, and Thauer, R.K., K. Jungermann, and H. Decker. 1977. Energy conservation in anaerobic chemotrophic bacteria. Bacteriol. Rev. 41: 100–180.
Using free energies of formation, it is possible to calculate ΔG0′ of a reaction. For the reaction A+B⇌C+D, ΔG0′ is calculated by subtracting the sum of the free energies of formation of the reactants (A+B) from that of the products (C+D). Thus ΔG0′=Gf 0[C+D]−Gf 0[A+B]
The value obtained for ΔG0′ tells us whether the reaction is exergonic (and can be a potential energy source for the cell) or endergonic (and requires an energy input to proceed). The phrase “products minus reactants” is a simple way to recall how to calculate changes in free energy during chemical reactions. This is illustrated for an example reaction, the oxidation of acetate to CO2, in Table 3.2, equation 2.
Before free-energy calculations can be made, it is first necessary to balance the reaction. That is, (1) the total number of each kind of atom and ionic charges must be identical on both sides of the reaction, and (2) the oxidation–reduction state must balance such that all of the electrons removed from one substance are transferred to another substance. Once a reaction is balanced, its ΔG0′ can be calculated, and from this, the potential of the reaction as a means of energy conservation for a cell can be assessed.
Calculating Free-Energy Change in Natural Conditions
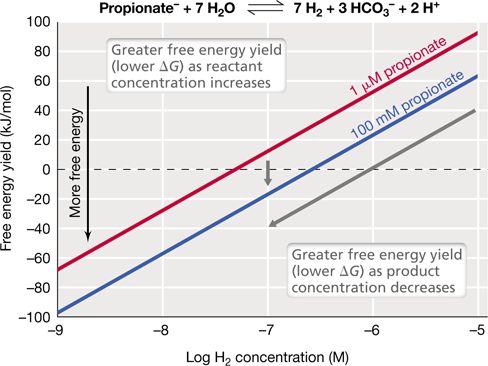
Although calculations of ΔG0′ are often very accurate estimates of actual free-energy changes, in some cases they are not. Consider the oxidation of propionate to H2 and CO2 (Figure 3.7). This reaction is important in wetlands because microorganisms called methanogens in these habitats ultimately consume H2 and CO2 and turn them into methane (CH4), a powerful greenhouse gas (Chapters 14, 17, and 21). Propionate oxidation is quite unfavorable at standard conditions when reactants and products are all present at molar concentration (ΔG0′=+304 kJ/mol). In wetlands, however, methanogens consume H2, lowering its concentration dramatically, and low H2 concentrations allow propionate oxidation to become exergonic (Figure 3.7). We will see when we pick up the bioenergetics theme again in Chapter 14 that the actual concentrations of products and reactants are almost never present at molar concentrations in a microbe’s natural habitat. Hence, the calculation of ΔG0′, while a useful approximation, can misrepresent the actual energy available to cells as they occur in the environment. In this regard, what is most relevant to a bioenergetic calculation is not ΔG0′, but ΔG, the free-energy change that occurs under the actual conditions in which the organism is growing. The equation for ΔG takes into account the actual concentrations of reactants and products in the organism’s habitat and is expressed as
Figure 3.7 Calculating free energy under natural conditions (ΔG).

The example is the oxidation of propionate to H2 and CO2 (as shown at the top of the figure, with CO2 written as HCO3 − [bicarbonate] because this is the form in which it naturally occurs in solution at neutral pH). This reaction is endergonic under standard conditions (ΔG0′=+304 kJ/mol) but can be exergonic under natural conditions in an environment where the H2 produced in the reaction is quickly consumed by other microbes. The Δ G was calculated at different concentrations of reactants (propionate) and products (H2).
Figure 3.7 Full Alternative Text ΔG=ΔG0′+RT ln Keq
where R and T are physical constants and Keq is the equilibrium constant for the reaction. For the reaction aA+bB⇌cC+dD, Keq=[C]c[D]d/[A]a[B]b, A and B are reactants and C and D are products; a, b, c, and d are the number of molecules of each, and the brackets indicate concentrations.
Returning to the example of propionate oxidation (Figure 3.7), we can see that the reaction becomes increasingly exergonic when propionate (the reactant) increases in concentration and when H2 (the product) decreases in concentration. Hence, propionate oxidation to CO2 and H2, though unfavorable at standard conditions, becomes more favorable as reactant concentrations go up and as product concentrations go down. The H2 concentration has a greater effect on ΔG than the propionate concentration because 7 H2 are formed for every propionate molecule oxidized, and hence H2 has a far greater impact on the value of the equilibrium constant for the reaction. While propionate oxidation is endergonic under standard conditions, propionate-oxidizing microbes can thrive in the environment as long as H2-consuming microbes (such as methanogens) coexist with them. Such interdependency is common in the microbial world and we will learn more about metabolic interactions between microbial partners in Chapter 14.
While ΔG0′ and ΔG are rarely identical, at this point in our understanding of bioenergetics the expression ΔG0′ is sufficient to appreciate the general principles of energy flow in microbial systems. The main point to keep in mind is that only exergonic reactions yield energy that can be conserved by the cell, and this will be our focus in the next few sections.
Check Your Understanding
Using Figure 3.4, calculate ΔG0′ for the reaction CH4+2 O2⇌CO2+2 H2O.
Does glucose formation from the elements release or require energy?
Using Table 3.3, calculate Δ G0′ for the reaction 2 CH4+O2⇌2 CH3OH.
3.4 Cellular Energy Conservation
Chemotrophs conserve energy from chemical reactions while phototrophs conserve energy from light. We have just reviewed how to calculate the free-energy change of chemical reactions. Now we introduce mechanisms of energy conservation. Ultimately, cells conserve energy by synthesizing energy-rich compounds. We begin by reviewing the energy-rich compounds that cells commonly use to conserve energy.
Adenosine Triphosphate
The most important energy-rich phosphate compound in cells is adenosine triphosphate (ATP). ATP consists of the ribonucleoside adenosine to which three phosphate molecules are bonded in series. From the structure of ATP (Figure 3.8), it can be seen that only two of the phosphate bonds (ATP⇌ADP+Pi and ADP⇌AMP+Pi) have free energies of hydrolysis more exergonic than −30 kJ/mol. By contrast, the phosphate bond in AMP contains only about half as much energy as those in ADP or ATP (Figure 3.8).
Figure 3.8 Energy-rich bonds in compounds that conserve energy in microbial metabolism.

The range in free energy of hydrolysis values for the phosphate or sulfur bonds highlighted in the compounds is listed in the table. The “R” group of acetyl-CoA is a 3′-phospho ADP group.
Although the energy released in ATP hydrolysis is −31.8 kJ/mol, a caveat must be mentioned here to more precisely state the energy requirements for the synthesis of ATP. In an actively growing Escherichia coli cell, the ratio of ATP to ADP is maintained at about 7:1, and this increases the actual free-energy requirements for ATP synthesis. Thus, in an actively growing cell, the actual energy that must be released (that is, the ΔG, Section 3.3) to drive the synthesis of ATP is closer to −55 to −60 kJ/mol. Nevertheless, for the purposes of learning and applying the basic principles of bioenergetics, we will assume that reactions conform to “standard conditions” (ΔG0′), and therefore we will assume that the energy needed to drive ATP synthesis (from ADP+Pi) is −31.8 kJ/mol. This unit is the fundamental currency of energy conservation within the cell and we will see that a major goal of energy conservation in catabolism is to obtain sufficient energy to form ATP.
Energy-Rich Compounds
ATP is not the only energy-rich compound in the cell; several others exist that, like ATP, have energy-rich phosphate bonds, and still others have energy-rich sulfur bonds. The biosynthesis of these compounds functions to trap free energy, and their hydrolysis releases this energy to drive endergonic reactions such as ATP synthesis.
A range of molecules have bonds consisting of a phosphate linked to an organic compound (Figure 3.8). However, not all phosphate bonds are energy-rich bonds. As seen in this figure, the ΔG0′ of hydrolysis of the phosphate bond in glucose 6-phosphate is −13.8 kJ/mol. By contrast, the ΔG0′ of hydrolysis of the phosphate bond in phosphoenolpyruvate is −61.9 kJ/mol, almost four times that of glucose 6-phosphate. Although the phosphate in either compound could be hydrolyzed to release energy, cells need compounds whose ΔG0′ of phosphate hydrolysis exceeds −31.8 kJ/mol to synthesize ATP.
Some energy-rich compounds have carbon–sulfur bonds. These include, in particular, derivatives of coenzyme A (for example, acetyl-CoA in Figure 3.8). Coenzyme A derivatives contain energy-rich thioester bonds, and hydrolysis of these bonds yields sufficient free energy to couple to the synthesis of an energy-rich phosphate bond. For example, in the coupled reaction Acetyl-S-CoA+H2O+ADP+Pi⇌acetate−+HS-CoA+ATP+H+
the energy released in the hydrolysis of coenzyme A is conserved in the synthesis of ATP. We will return to the importance of coenzyme A derivatives in microbial bioenergetics many times in Chapter 14.
Mechanisms of Energy Conservation
Cells conserve energy by generating ATP through one of three fundamental mechanisms. The first mechanism is substrate-level phosphorylation. In substrate-level phosphorylation the energy-rich bond of a substrate is hydrolyzed to directly drive the formation of ATP. For example, hydrolysis of the phosphate bond in phosphoenolpyruvate is sufficiently exergonic to drive ATP formation (see Figures 3.8 and 3.11). We will see that substrate-level phosphorylation is the dominant mechanism of energy conservation in fermentative organisms (see Section 3.7 and Figure 3.21).
The second mechanism of energy conservation is oxidative phosphorylation. In oxidative phosphorylation the movement of electrons from an electron donor to an electron acceptor (**Figure 3.1*b***) generates a proton motive force. The proton motive force is an electrochemical gradient formed by energy-conserving reactions that transport protons outside the cytoplasmic membrane. This electrochemical gradient creates a force that is ultimately used to synthesize ATP (Section 2.1 and Figure 2.4). Oxidative phosphorylation is the defining feature of respiration reactions and it is performed by diverse chemotrophic organisms (Sections 3.9, 3.10 and 3.11).
The third mechanism of energy conservation is photophosphorylation. In photophosphorylation light energy is used to form the proton motive force that powers ATP synthesis and is the dominant mechanism of energy conservation in phototrophic organisms. Both photophosphorylation and oxidative phosphorylation ultimately rely on electron transfer reactions to drive the formation of the proton motive force (Section 3.11).
We now turn our attention to enzymes—the cell’s premier catalysts—and the importance of catalysis in cellular metabolism.
Check Your Understanding
What are the three fundamental mechanisms of conserving cellular energy and in what types of organisms would these mechanisms be most important?
Explain whether 1,3-bisphosphoglycerate or glucose 6-phosphate (Figure 3.8) could be used to generate ATP through substrate-level phosphorylation.
Which mechanisms of energy conservation rely on the proton motive force?
3.5 Catalysis and Enzymes
Free-energy calculations reveal only whether energy is released or required in a given reaction; they say nothing about the rate of the reaction. If the rate of a reaction is very slow, it may be of no value to a cell. For example, consider the formation of water from O2 and H2. The energetics of this reaction are quite favorable: H2+12 O2→H2O, ΔG0′=−237 kJ. However, if O2 and H2 were mixed in a sealed bottle, no measurable amount of water would form, even after years. This is because the bonding of O2 and H2 to form H2O requires that these two gases become reactive. This requires that their bonds be broken, which requires a small amount of energy. This energy is called activation energy.
Activation energy can be viewed as the minimum energy required for a chemical reaction to begin. For an exergonic reaction, the situation is as shown in Figure 3.9. Although the activation energy barrier is virtually insurmountable in the absence of a catalyst—a substance that facilitates a reaction but is not consumed by it—in the presence of a proper catalyst, this barrier is reduced, allowing the reaction to proceed.
Figure 3.9 Activation energy and catalysis.

Even chemical reactions that release energy may not proceed spontaneously if not activated. Once the reactants are activated, the reaction proceeds spontaneously. Catalysts such as enzymes lower the required activation energy.
Enzymes
Catalysts function by lowering the activation energy of a reaction (Figure 3.9), thereby increasing the reaction rate. Catalysts have no effect on the energetics or the equilibrium of a reaction but only affect the rate at which a reaction proceeds. Most cellular reactions will not proceed at significant rates without catalysis.
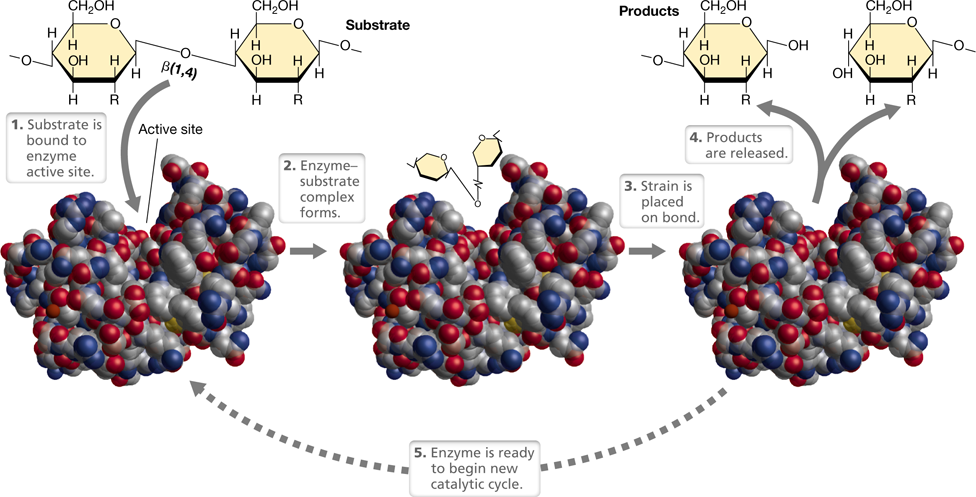
The major catalysts in cells are enzymes, which are proteins (or in a few cases, RNAs) that are highly specific for the reactions they catalyze. This specificity is a function of the precise three-dimensional structure of the enzyme. In an enzyme-catalyzed reaction, the enzyme combines with the reactant, called a substrate, forming an enzyme–substrate complex. Then, as the reaction proceeds, the product is released and the enzyme is returned to its original state, ready to catalyze a new round of the reaction (Figure 3.10). The enzyme is generally much larger than the substrate(s), and the portion of the enzyme to which substrate binds is the enzyme’s active site; the entire enzymatic reaction, from substrate binding to product release, may take only a few milliseconds.
Figure 3.10 The catalytic cycle of an enzyme.

The enzyme depicted here, lysozyme, catalyzes the cleavage of the β-1,4-glycosidic bond in the polysaccharide backbone of peptidoglycan (Section 2.3). Following substrate binding in the enzyme’s active site, strain is placed on the bond, and this favors breakage. Space-filling model of lysozyme courtesy of Richard Feldmann.
Many enzymes contain small nonprotein molecules that participate in catalysis but are not themselves substrates. These small molecules can be divided into two classes based on the way they associate with the enzyme: prosthetic groups and coenzymes. Prosthetic groups bind tightly to their enzymes, usually covalently and permanently. The heme group present in cytochromes such as cytochrome c (Section 3.8) is an example of a prosthetic group. By contrast, coenzymes, with a few exceptions, are loosely and often transiently bound to enzymes; thus, a single coenzyme molecule may associate with a number of different enzymes. NADH is an example of a coenzyme (Figures 3.5 and 3.6), as is ATP. Most coenzymes are derivatives of vitamins (Section 4.1 and Table 4.1).
Enzyme Catalysis
To catalyze a reaction, an enzyme must bind its substrate and position it properly in its active site. The enzyme–substrate complex (Figure 3.10) aligns reactive groups in the substrate(s) and places strain on specific bonds. This reduces the activation energy required to make the reaction proceed (Figure 3.9). This is shown in Figure 3.10 for the enzyme lysozyme, an enzyme whose substrate is the polysaccharide backbone of peptidoglycan, the bacterial cell wall polymer (Section 2.3).
Mastering Microbiology
Art Activity: Figure 3.8 The catalytic cycle of an enzyme
The reaction depicted in Figure 3.9 is exergonic. By contrast, some enzymes catalyze endergonic reactions where they convert energy-poor substrates into energy-rich products. In these cases not only must an activation energy barrier (Figure 3.9) be overcome, but sufficient free energy must also be put into the reaction in order to raise the energy level of the substrates to that of the products. This is done by coupling the energy-requiring reaction to an energy-yielding one, such as the hydrolysis of ATP or dissipation of the proton motive force, so that the overall reaction proceeds with a free-energy change that is either negative in arithmetic sign or zero.
Theoretically, all enzymes are reversible in their activity. However, enzymes that catalyze highly exergonic or highly endergonic reactions typically function in only one direction. If a particularly exergonic or endergonic reaction needs to be reversed, a different enzyme usually catalyzes the reverse reaction.
Check Your Understanding
What is the function of a catalyst? What are the building blocks of an enzyme?
II Catabolism: Chemoorganotrophs
Chemoorganotrophs use organic carbon molecules to fuel metabolism, and glycolysis and the citric acid cycle are central metabolic pathways used in the metabolism of organic carbon molecules.
We have learned in the previous sections that cells harvest energy from redox reactions and they conserve this energy in the synthesis of ATP and other high-energy compounds. We have also learned that there are many different metabolic types of organisms, each defined by its source of energy (Figure 3.3).
Here we examine the metabolic pathways that cells use to transfer electrons and conserve energy. We begin by examining chemoorganotrophs that perform fermentation and respiration. Fermentation is a form of anaerobic catabolism in which organic compounds both donate electrons and accept electrons, and redox balance is achieved without the need for external electron acceptors. By contrast, respiration is a form of aerobic or anaerobic catabolism in which an electron donor, which can be either organic or inorganic, is oxidized using an external electron acceptor such as O2 (in aerobic respiration) or some other compound (in anaerobic respiration). We will revisit the concepts of fermentation and respiration in detail in Chapter 14 and so focus here only on the essentials that define these processes.
3.6 Glycolysis, the Citric Acid Cycle, and the Glyoxylate Cycle
3.6 Glycolysis, the Citric Acid Cycle, and the Glyoxylate Cycle
3.6 Glycolysis, the Citric Acid Cycle, and the Glyoxylate Cycle
Chemoorganotrophs obtain the electrons they need to conserve energy from the oxidation of organic compounds, such as glucose. A nearly universal pathway for the catabolism of glucose is the Embden–Meyerhof–Parnas pathway, better known as glycolysis, a series of reactions in which glucose is oxidized to pyruvate. Many metabolic reactions are modular and glycolysis is a prime example of such a module. Glycolysis can participate in several forms of catabolism including fermentation, aerobic respiration, and anaerobic respiration.
Glycolysis
Glycolysis can be divided into two stages, each consisting of one or more enzymatic reactions. Stage I consists of “preparatory” reactions; these are not redox reactions and do not release energy but instead form a key intermediate of the pathway. In stage II, redox reactions occur, energy is conserved, and two molecules of pyruvate are formed (Figure 3.11).
Figure 3.11 Embden–Meyerhof–Parnas pathway (glycolysis).

The sequence of reactions in the catabolism of glucose to pyruvate. Glycolysis results in the formation of 2 each of pyruvate, ATP, and NADH per glucose molecule.
To begin glycolysis, glucose is phosphorylated to form glucose 6-phosphate. The latter is then isomerized to fructose 6-phosphate, and a second phosphorylation leads to the production of fructose 1,6-bisphosphate. These steps consume, rather than produce, ATP. The enzyme aldolase then splits fructose 1,6-bisphosphate into two 3-carbon molecules, glyceraldehyde 3-phosphate and its isomer, dihydroxyacetone phosphate, which is converted into glyceraldehyde 3-phosphate. To this point, all of the reactions, including the consumption of ATP, have proceeded without any redox changes (Figure 3.11).
The first redox reaction of glycolysis occurs when glyceraldehyde 3-phosphate is oxidized to 1,3-bisphosphoglycerate. In this reaction (which occurs twice, once for each of the two glyceraldehyde 3-phosphates), the enzyme glyceraldehyde-3-phosphate dehydrogenase reduces NAD+ to NADH. Simultaneously, each glyceraldehyde 3-phosphate molecule is phosphorylated by the addition of a molecule of inorganic phosphate. This reaction, in which inorganic phosphate is converted to organic form, sets the stage for energy conservation, since 1,3-bisphosphoglycerate is an energy-rich compound (Figure 3.11; see also Figure 3.8). ATP is then synthesized by substrate-level phosphorylation when (1) each molecule of 1,3-bisphosphoglycerate is converted to 3-phosphoglycerate, and (2) each molecule of phosphoenolpyruvate is converted to pyruvate (Figure 3.11). As a result, glycolysis consumes two ATP molecules in stage I and produces four ATP molecules in stage II. The result is that glycolysis produces a net yield of 2 molecules of ATP, 2 molecules of NADH, and 2 molecules of pyruvate per molecule of glucose.
The glycolytic pathway on its own, while sufficient to make ATP, is insufficient to sustain life because it lacks redox balance. That is, glycolysis produces NADH but it lacks an electron acceptor and so lacks the ability to regenerate the NAD+ needed to oxidize glyceraldehyde-3-P to 1,3-bisphosphoglycerate. Cells performing glycolysis can achieve redox balance by using one of several different mechanisms including fermentation, where NADH oxidation is linked to the reduction of pyruvate (Section 3.7), or respiration, where glycolysis is coupled to the citric acid cycle and respiration (Section 3.8).
The Citric Acid Cycle
The pathway by which pyruvate is oxidized to CO2 is called the citric acid cycle (CAC) (Figure 3.12). Pyruvate from glycolysis is decarboxylated, leading to the production of CO2, NADH, and the energy-rich compound acetyl-CoA, which enters the CAC (Figure 3.12 and see Figure 3.8). The acetyl group of acetyl-CoA combines with the four-carbon CAC intermediate oxaloacetate, forming the six-carbon compound citric acid, for which the CAC is named. A sequence of reactions follows, and two additional CO2 molecules, three more NADH, and one FADH2 are formed per pyruvate oxidized. Ultimately, oxaloacetate is regenerated as the next acetyl acceptor, thus completing the cycle (Figure 3.12). The result is that the citric acid cycle produces a net yield of 1 molecule of ATP (or GTP), 4 molecules of NADH (or NADPH), 1 molecule of FADH2, and 3 molecules of CO2 per molecule of pyruvate.
Figure 3.12 The citric acid cycle.

The citric acid cycle begins when the two-carbon compound acetyl-CoA (derived from pyruvate) condenses with the four-carbon compound oxaloacetate to form the six-carbon compound citrate. Through a series of oxidations and transformations, citrate is converted to two CO2 and the acetyl acceptor molecule, oxaloacetate. For every pyruvate that enters the cycle, 3 CO2, 1 ATP, 4 NADH, and 1 FADH2 are produced.
Linkage of glycolysis with the citric acid cycle allows one molecule of glucose to be completely oxidized to six molecules of CO2, while producing 4 molecules of ATP (2 in glycolysis and 2 in the CAC). However, redox is still out of balance because these pathways have combined to produce 10 molecules of NADH and 2 of FADH2 per molecule of glucose. Redox balance is ultimately achieved in respiration reactions (Section 3.9). However, before we turn our attention to respiration, we consider another important aspect of the citric acid cycle.
Biosynthesis and the Citric Acid Cycle
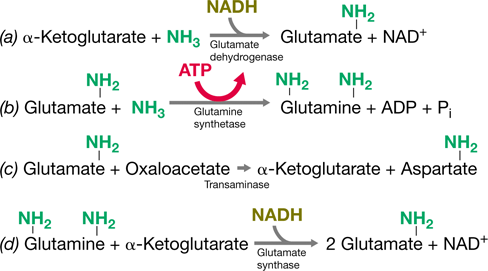
Besides its role in oxidizing pyruvate to CO2, the citric acid cycle also plays a major role in biosynthesis. The cycle is composed of several key organic compounds, small amounts of which are drawn off during growth to produce new cell material. Particularly important in this regard are α-ketoglutarate and oxaloacetate, which are precursors of several amino acids (Section 3.14), and succinyl-CoA, needed to form cytochromes, chlorophyll, and related molecules. Any shortage of oxaloacetate is corrected by the addition of CO2 (carboxylation) to pyruvate or phosphoenolpyruvate (Figure 3.12).
Oxaloacetate is also an important intermediate because it can be converted to phosphoenolpyruvate (a precursor of glucose) if necessary (Section 3.13). In addition, acetate is important because it provides the raw material for fatty acid biosynthesis (Section 3.15). The CAC thus plays two major roles in the cell: oxidation of organic compounds and the biosynthesis of key metabolites. The same can be said about the glycolytic pathway, as certain intermediates from this pathway are also drawn off for biosynthetic needs (Sections 3.13 and 3.14) and then replenished from glucose in the next round of glycolysis.
Other Pathways for Chemoorganotrophy
Chemoorganotrophs can use glycolysis and the CAC to oxidize several compounds including glucose, citrate, malate, fumarate, and succinate (all C4 or C6 compounds). In addition, many unrelated catabolic pathways can be linked to glycolysis and the CAC to enable the oxidation of an even wider diversity of organic compounds. For example, many sugars can be converted to glucose by isomerase enzymes and then metabolized by glycolysis. Likewise, isomerases that yield CAC intermediates can be used to metabolize a wide range of organic compounds.
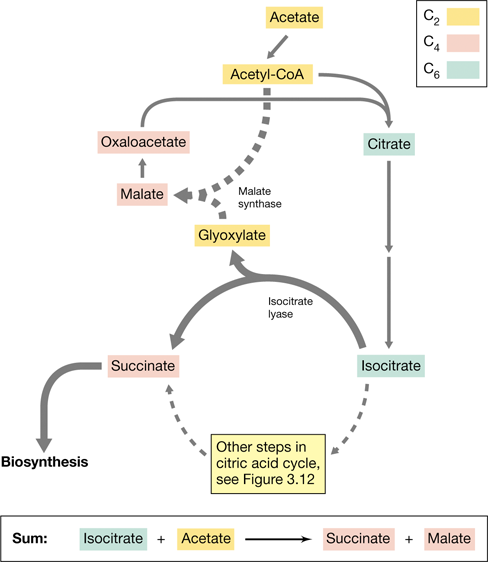
Some chemoorganotrophs can also grow on two-carbon (C2) compounds by using the glyoxylate cycle (Figure 3.13). Microbes cannot grow on C2 compounds such as acetate by using the CAC only. This is because some oxaloacetate must be drawn off from the CAC to biosynthesize glucose and amino acid precursors (Sections 3.13 and 3.14). It is possible to readily synthesize oxaloacetate from a range of C4 and C6 compounds but not from C2 compounds. This inability to regenerate oxaloacetate during growth on acetate means that the CAC would starve for the oxaloacetate needed to accept the acetyl group from acetyl-CoA (Figure 3.12). Hence, cells use the glyoxylate cycle (so named because the C2 compound glyoxylate is a key intermediate) to grow using C2 compounds.
Figure 3.13 The glyoxylate cycle.
These reactions occur in conjunction with the citric acid cycle when cells grow on two-carbon electron donors, such as acetate. The glyoxylate cycle regenerates oxaloacetate (from malate) to maintain an acetyl acceptor for the citric acid cycle.
The glyoxylate cycle is composed of several citric acid cycle enzymatic reactions plus two additional enzymes: isocitrate lyase, which cleaves isocitrate into succinate and glyoxylate, and malate synthase, which converts glyoxylate and acetyl-CoA to malate (Figure 3.13). The succinate formed can be used for biosynthesis, while the glyoxylate combines with acetyl-CoA (C2) to yield malate (C4). From malate, the acceptor molecule oxaloacetate is produced and can enter a new round of acetyl-CoA oxidation in the citric acid cycle (Figure 3.13).
Another example of this principle comes from chemoorganotrophs that grow on C3 compounds. Cells using standard CAC reactions to grow on C3 compounds, such as pyruvate, are also unable to regenerate oxaloacetate. However, carboxylation reactions catalyzed by the enzymes pyruvate carboxylase or phosphoenolpyruvate carboxylase, which convert pyruvate or phosphoenolpyruvate into oxaloacetate (respectively), allow cells growing on C3 compounds to regenerate oxaloacetate (Figure 3.12).
It should thus be clear that metabolic reactions are modular entities and that the reactions of glycolysis, the citric acid cycle, and the glyoxylate cycle allow for a wide diversity of organic electron donors to fuel the energy needs of chemoorganotrophs.
Check Your Understanding
Which reactions in glycolysis are redox reactions?
Why does an organism need the enzyme pyruvate carboxylase to enable growth if pyruvate is its sole source of energy and reducing power?
Why is it not possible for an organism to grow by using only glycolysis?
3.7 Principles of Fermentation
Microbes that ferment glucose achieve energy conservation by substrate-level phosphorylation and achieve redox balance by reducing the pyruvate produced in glycolysis (Section 3.6) and then excreting the reduced molecule to the environment as a waste product (Figure 3.14). These fermentation products are often highly reduced substances such as organic acids or alcohols. However, fermentation products are not waste products to humans. Instead, they are the foundation of the baking and fermented beverage industries, including beer and wine production, and are key ingredients in many other fermented foods, such as the lactic and other acids in fermented dairy products (yogurt, sour cream, buttermilk, and the like), cheeses, pickles, and certain sausages and fish products (Section 1.6). In addition, fermentation products produced by the human microbiome can exert a powerful influence on human health (Chapter 24).
Figure 3.14 The essentials of fermentation.
An organic compound is oxidized, and the electrons are ultimately recycled back to one of the oxidized organic products because an external electron acceptor is lacking. The fermentation product so formed is excreted from the cell and ATP is produced by substrate-level phosphorylation. Most of the fermentable substrate is used for energy generation and only a relatively small amount is used for biosynthesis.
Energy Conservation and Redox Balance in Fermentation
There are a tremendous diversity of fermentation reactions and many employ glycolysis, but in the final analysis, all fermentations must achieve two major results: first, the cell must conserve energy, and second, the cell must achieve a final redox balance. Since fermentative organisms rely on substrate-level phosphorylation to conserve energy, they need to produce compounds that contain energy-rich bonds with which to synthesize ATP (Section 3.4). In addition, since fermentative organisms lack an external electron acceptor they need to find a way to oxidize NADH back to NAD+. Fermentative organisms solve the latter problem by creating an electron acceptor from the organic electron donor they consumed. For example, yeast ferments one molecule of glucose to two molecules of ethanol and two molecules of CO2. During this fermentation, yeast gets all of its ATP from glycolysis and then replenishes NAD+ by donating electrons from NADH to pyruvate, which is the end product of glycolysis (Figure 3.11). In this way, pyruvate is the electron acceptor, and the reduction of pyruvate produces the fermentation products ethanol and CO2, which are excreted from the cell.
Lactic acid bacteria provide an alternative example of this same principle. These bacteria convert one molecule of glucose to two molecules of lactic acid (see Figure 3.21). Like yeast, lactic acid bacteria produce all of their ATP in glycolysis and achieve redox balance by reducing pyruvate. However, unlike yeast, lactic acid bacteria employ different enzymes to reduce pyruvate and as a result, produce lactic acid as their fermentation product instead of ethanol plus CO2. Lactic acid bacteria are a tremendously important group of microorganisms. They are responsible for the production of many important fermented foods (Figure 1.16). Lactic acid bacteria are also the cause of dental caries, and the lactic acid they produce is essential to maintaining the health of the human large intestine and vagina (Chapter 24).
Fermentative Diversity
While sugars can be readily fermented using glycolytic reactions, there is a tremendous diversity of fermentative pathways. Many different organic substrates can be fermented, including amino acids, fatty acids, purines, pyrimidines, and even aromatic compounds. Many fermentative organisms that use these substrates produce acetate or other volatile fatty acids (for example, propionate, buty-rate) as fermentation products. Volatile fatty acid fermentation products contribute to energy conservation because they provide an opportunity to make ATP by substrate-level phosphorylation. The key intermediate in such reactions is the coenzyme-A derivative of each fatty acid, since these are energy-rich compounds. For example, the final step of the butyric acid fermentation performed by the anaerobic bacterium Clostridium butyricum is Butyryl−CoA+ADP+Pi→butyric acid+ATP+CoA
The formation of butyryl-CoA, which has an energy-rich bond, allows C. butyricum to make ATP by substrate-level phosphorylation. In addition, redox balance in many fermentative organisms can be improved by the production of molecular hydrogen (H2). The 2 H+/H2 redox couple has a very low reduction potential (ΔE0′=−0.42) and so only organisms that produce very low reduction potentials in their metabolisms can produce H2 (for example, NADH cannot reduce H+ to H2, Figure 3.4). Often the production of H2 is associated with the activity of ferredoxin, a very low-potential electron carrier (Section 3.8) and catalyzed by the enzyme hydrogenase.
Fermentation reactions support the growth of an enormous diversity of microorganisms, but substrate-level phosphorylation can only conserve a fraction of the energy available in organic substrates. We will see in the next two sections that respiration can conserve far more energy than fermentation and is the favored form of metabolism in oxic environments.
Check Your Understanding
Why is acetate formation energetically beneficial during fermentative metabolism?
What two metabolic events result from any fermentation?
3.8 Principles of Respiration: Electron Carriers
In respiration, electrons are transferred from reduced electron donors such as glucose to external electron acceptors such as O2. During glycolysis and the citric acid cycle, the complete oxidation of glucose to 6 CO2 produces a considerable amount of NADH and FADH2 (Figure 3.12). These molecules must be reoxidized back to NAD+ and FAD in order to achieve redox balance. In glucose respiration, NADH and FADH2 are reoxidized when they donate electrons to electron transport reactions. Electron transport takes place within the cytoplasmic membrane, culminating in the reduction of an external electron acceptor and the formation of an electrochemical gradient across the membrane. The electrochemical gradient is formed when protons (or in some cases, other ions) are pumped across the cytoplasmic membrane, and this electrochemical gradient—the proton motive force—is ultimately used to conserve energy through ATP synthesis. Hence, at a fundamental level, respiration is a process in which chemical energy from redox reactions is conserved by using electron transport reactions to pump protons (or other ions) across a membrane. We now focus on the enzymes, cofactors, and coenzymes that participate in electron transfer reactions.
Mastering Microbiology
Microbiology Animation: Electron Transport Chain: The Process
NADH Dehydrogenases and Flavoproteins
Electron transport reactions occur within membranes, typically in the cytoplasmic membrane of prokaryotic cells or the mitochondrial membrane of eukaryotic cells. Several types of oxidation–reduction enzymes participate in electron transport. These include NADH dehydrogenases, flavoproteins, iron–sulfur proteins, and cytochromes. Also participating are small nonprotein electron carriers called quinones. The carriers are arranged in the membrane in order of increasingly more positive reduction potential, with NADH dehydrogenase first and the cytochromes last (see Figures 3.16 and 3.19).
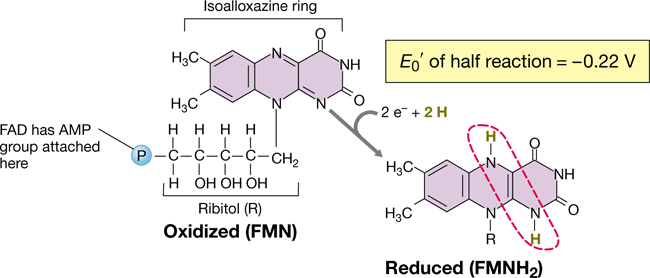
NADH dehydrogenases contain an active site that binds NADH. The 2 e− and the H+ from NADH, and one H+ from the cytoplasm, are transferred by the NADH dehydrogenase to a flavoprotein, the next carrier in the chain. This regenerates NAD+, which can return to participate in glycolysis, the citric acid cycle, or other cellular processes (Sections 3.6 and 3.7). Flavoproteins contain a derivative of the vitamin riboflavin (Figure 3.15). The flavin portion, which is bound to its protein as a prosthetic group, is reduced as it accepts 2 e−+2 H+ and oxidized when 2 e− are passed to the next carrier in the chain (note that flavoproteins accept 2 e−+2 H+ but donate only electrons, the protons are ultimately released to the cytoplasm). Two types of flavins are commonly found in cells, flavin mononucleotide (FMN, Figure 3.15) and flavin adenine dinucleotide (FAD).
Figure 3.15 Flavin mononucleotide (FMN), a hydrogen atom carrier.

The site of oxidation–reduction (dashed red circle) is the same in FMN and the related coenzyme flavin adenine dinucleotide (FAD, not shown). FAD contains an adenosine monophosphate (AMP) group bonded through the phosphate group on FMN.
Cytochromes, Other Iron Proteins, and Quinones
Cytochromes are proteins that contain heme prosthetic groups (Figure 3.16). Cytochromes undergo oxidation and reduction through loss or gain of an electron by the iron atom that exists as either Fe2+ or Fe3+. Several classes of cytochromes exist, differing widely in their reduction potentials (Figure 3.4). Different classes of cytochromes are designated by letters, such as cytochrome a, cytochrome b, cytochrome c, and so on, depending on the type of heme they contain. The redox potential of a cytochrome is ultimately governed by its protein structure and the type of heme present. Occasionally, cytochromes form into complexes with other cytochromes or with iron–sulfur proteins. An important example is the cytochrome bc1 complex, which is a multiprotein enzyme complex that contains two different b-type cytochromes and one c-type cytochrome. The cytochrome bc1 complex plays an important role in energy metabolism, as we will see in the next section.
Figure 3.16 Cytochrome and its structure.

(a) Structure of heme, the iron-containing portion of cytochromes. Hemes are tetrapyrroles, composed of four pyrrole rings. Cytochromes carry electrons only, and the redox site is the iron atom, which can alternate between the Fe2+ and Fe3+ oxidation states. (b) Space-filling model of cytochrome c; heme (light blue) is covalently linked via disulfide bridges to cysteine residues in the protein (dark blue).
In addition to the cytochromes, in which iron is bound to heme, one or more proteins with nonheme iron are also components of electron transport chains. These proteins contain prosthetic groups made up of clusters of iron and sulfur atoms, with Fe2S2 and Fe4S4 clusters being the most common (Figure 3.17). For example, bacterial ferredoxin, a nonheme iron–sulfur protein of low reduction potential (about −0.4 V) (Table 3.1) and an important component in H2 production (Section 3.7), contains an Fe4S4 cluster. The reduction potentials of iron–sulfur proteins vary from −0.2 to about −0.45 V, depending on the iron–sulfur cluster present, the structure of the protein that contains it, and how the cluster is embedded in the protein. Thus, different iron–sulfur proteins can function at different locations in the electron transport chain. Like cytochromes, iron–sulfur proteins carry only electrons.
Figure 3.17 Arrangement of the iron–sulfur centers of nonheme iron–sulfur proteins.

(a) Fe2S2 center. (b) Fe4S4 center. The cysteines (Cys) link the protein to its Fe/S cluster.
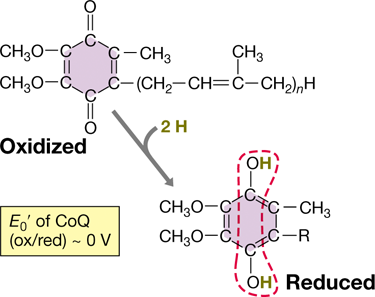
Quinones (Figure 3.18) are small hydrophobic redox molecules that lack a protein component. Because they are small and hydrophobic, quinones can move about within the membrane. Like the flavins (Figure 3.15), quinones accept 2 e−+2 H+ but transfer only 2 e− to the next carrier in the chain. Quinones typically function to link iron–sulfur proteins (Figure 3.17) and the initial cytochrome (Figure 3.16) in the electron transport chain. Several types of quinones exist, but ubiquinone (also called coenzyme Q) and menaquinone are the most common quinones and are widely distributed in species of Bacteria and Archaea.
Figure 3.18 Structure of oxidized and reduced forms of ubiquinone (coenzyme Q, or CoQ).

The five-carbon unit in the side chain (an isoprenoid) occurs in multiples, typically 6–10. Oxidized ubiquinone requires 2 e− and 2 H+ to become fully reduced (dashed red outline).
Check Your Understanding
In what major way do quinones differ from other electron carriers in the membrane?
Which electron carriers described in this section accept 2 e−+2 H+? Which accept electrons only?
Consider an electron transport chain that transports electrons from NADH to O2. In what order would cytochromes a, b, and c need to occur in the electron transport chain? (Hint: Refer to Figure 3.4.)
3.9 Principles of Respiration: Generating a Proton Motive Force
3.9 Principles of Respiration: Generating a Proton Motive Force
3.9 Principles of Respiration: Generating a Proton Motive Force
As we will see later (Section 3.10), respiration can be either aerobic or anaerobic. However, energy conservation in all forms of respiration is linked to an energized state of the membrane—the proton motive force. In addition, we will see that electron transfer reactions and the generation of a proton motive force are also required for energy conservation in phototrophy (Section 3.11). Hence, electron transport and the generation of a proton motive force is a unifying concept that underlies energy conservation in all forms of metabolism except fermentation.
Electron Transport
Understanding the linkage between electron transport and ATP synthesis requires an appreciation for the organization of the electron transport chain in the cytoplasmic membrane (Figure 3.19). While the exact composition of the electron transport chain varies between organisms, and multiple electron transport chains can sometimes function simultaneously in a single organism, the principles of electron transport are nearly universal.
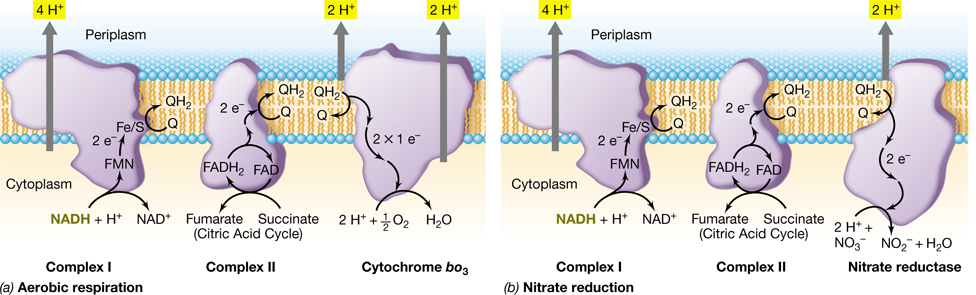
Figure 3.19 Generation of the proton motive force during aerobic respiration.

The orientation of electron carriers in the cytoplasmic membrane of the bacterium Paracoccus denitrificans. The + and − charges at the inner and outer membrane surfaces represent H+ and OH−, respectively. FMN, flavin mononucleotide; FAD, flavin adenine dinucleotide; Q, quinone; Fe/S, iron–sulfur protein; cyt a, b, c, cytochromes (bL and bH, low- and high-potential b-type cytochromes, respectively). The Q-cycle occurs at Complex III (cytochrome bc1). During the Q-cycle, two molecules of QH2 are oxidized and a total of 4 H+ exchanged across the membrane for every 2 e− that pass through the complex. Complex II, the succinate dehydrogenase complex, bypasses Complex I and feeds electrons directly into the quinone pool at a more positive E0′ than NADH (see the redox tower in Figure 3.4).
Electron transport begins when an electron donor adds electrons to the electron transport chain. In chemoorganotrophic organisms, electrons enter electron transport from either NADH or FADH2. By contrast, many different inorganic electron donors can feed into electron transport in various chemolithotrophic organisms (see Section 3.11). However, regardless of the source of electrons, electron carriers within the transport chain are embedded in the membrane in order of their increasingly positive reduction potentials (Figure 3.19; see also Figure 3.4). During electron transport, electrons are passed on from lower-potential (more negative ΔE0′) carriers to higher-potential (more positive ΔE0′) carriers until at the end of the chain they ultimately reduce a terminal electron acceptor, such as O2.
The movements of electrons through the electron transport chain are exergonic reactions that provide the free energy to pump protons to the outer surface of the membrane, thereby generating the proton motive force. Recall that H2O naturally dissociates to form H+ and OH−. Hence, removal of H+ from the cytoplasm results in the accumulation of OH− on the cytoplasmic side of the membrane. Though small, H+ are not free to diffuse across the membrane because they are charged and polar; thus, they must be extruded and this occurs during electron transport reactions. This separation of H+ from OH− creates a difference in pH and an electrochemical potential across the membrane, and this electrochemical potential energizes the membrane (Figure 3.19) much like the potential energy that exists from charge separation in a battery. This electrochemical gradient establishes the proton motive force, which can be used by the cell to do work in the form of ATP synthesis, active transport, motility, and a few other energy-requiring reactions.
The mechanisms of electron transport in a model chemoorganotrophic bacterium widely used for studies of respiration, Paracoccus denitrificans, is shown in Figure 3.19. This bacterium can grow by aerobic respiration of glucose. In so doing, it uses glycolysis and the citric acid cycle to produce NADH and FADH2 destined for the electron transport chain, and it uses O2 as its terminal electron acceptor. We know the electron transport chain of P. denitrificans in detail because it is used as a model system to study mitochondrial function. This resemblance occurs because the mitochondrion is the result of an ancient symbiosis that took place between a bacterial cell and the ancestor of all Eukarya (Section 13.4). Electron transport takes place in the cytoplasmic membrane of P. denitrificans and in the mitochondrial membrane of eukaryotic cells.
Mastering Microbiology
Art Activity: Figure 3.22 Generation of the proton motive force during aerobic respiration
Generation of the Proton Motive Force: Complexes I and II
In P. denitrificans and many aerobic bacteria, electron transport begins at Complex I (Figure 3.19). The term complex refers to the fact that this system is composed of many proteins that function as a unit (for example, in Escherichia coli, Complex I contains 14 different proteins). Complex I is also called NADH:quinone oxidoreductase (or simply NADH dehydrogenase) because the overall reaction is one in which NADH is oxidized and a quinone (Figure 3.18) is reduced. Complex I oxidizes NADH to NAD+ and transfers the resulting 2 e− to a protein containing an FMN coenzyme and from there to a series of Fe/S proteins within the complex, before ultimately reducing ubiquinone (Q) to ubiquinol (QH2). This reduction causes ubiquinone to pick up 2 H+ from the inside of the cell. The QH2 is able to diffuse within the membrane to Complex III, the next step in the electron transport chain. In addition, the electron transfer reactions within Complex I cause a conformational change within the complex resulting in transport of 4 H+ across the membrane (Figure 3.19).
As an alternative entry point to the electron transport chain, electrons from FADH2 enter at Complex II. Complex II is also called the succinate dehydrogenase complex because the oxidation of succinate to fumarate in the citric acid cycle (Figure 3.12) reduces FAD+ to FADH2, ultimately resulting in the reduction of Complex II (Figure 3.19). The 2 e− from FADH2 are transferred through Complex II to ubiquinone (Q), which accepts these 2 e− and 2 H+ from the cytoplasm to become ubiquinol (QH2). In this way Complexes I, II, and III are all linked by the quinone pool. Quinones are reduced by either Complex I or Complex II, and their reduction requires 2 H+, which are obtained from the cytoplasmic side of the membrane; QH2 is then free to diffuse within the membrane to Complex III. However, electrons entering at Complex I result in greater energy conservation than those entering at Complex II because the former translocates 4 additional H+ for every 2 e− transported (Figure 3.19).
Complexes III and IV: bc1 and *c*- and *a*-Type *Cytochromes*
Complex III consists of the cytochrome bc1 complex (Figure 3.19). Electrons enter Complex III from QH2 at the Qo site and they leave when donated to cytochrome c. When QH2 donates its 2 e− to Complex III, it releases its two H+ outside of the cytoplasmic membrane, contributing to the proton motive force. However, QH2 carries 2 e− and cytochrome c only carries 1 e−. This difference creates an opportunity for conserving additional energy through a mechanism known as the Q-cycle. During the Q-cycle, cytochrome bc1 carries out an electron bifurcation in which the two 2 e− from QH2 (E0′=+0.09 V) are sent in different directions. The first e− is transported to cytochrome c (E0′=+0.25 V) in an exergonic reaction whose free-energy release helps drive transfer of the second e− in the slightly endergonic reaction to subunit bL (E0′=−0.06 V). The reduction potential of the bL subunit is sufficient to transfer e− to the bH subunit and then ultimately back to a second molecule of ubiquinone (Q) that is docked at the Qi site of Complex III. It takes two such cycles to reduce fully the ubiquinone at the Qi site to QH2, and the net result of the Q-cycle is that 4 H+ are transferred to the cytoplasmic side of the membrane for every 2 e− that move through Complex III to cytochrome c (Figure 3.19).
Cytochrome c functions as a periplasmic shuttle to transfer e− from Complex III to the high-redox-potential cytochromes a and a3 of Complex IV (Figure 3.19). Complex IV functions as the terminal oxidase and reduces O2 to 2 H2O in the final step of the electron transport chain. It takes 4 e− to reduce O2 to 2 H2O a reaction that also consumes 4 H+ from the cytoplasm. This transfer of electrons causes a conformational change in Complex IV that allows 1 H+ to be pumped to the outer surface of the membrane for every e− transferred through Complex IV (Figure 3.19).
The net result for an organism like P. denitrificans is that for every 2 e− transported from NADH to O2, a total of 10 H+ are transferred to the outside of the membrane (4 at Complex I, 4 at Complex III, and 2 at Complex IV), and another 2 are consumed in the cytoplasm during the formation of water. In contrast, for every 2 e− transported from FADH2 to O2 a total of 6 H+ are transferred to the outside of the membrane (4 at Complex III and 2 at Complex IV), and another 2 are consumed in the cytoplasm during the formation of water.
Thus far in respiration we have disposed of electrons generated in various catabolic reactions and simultaneously generated a proton motive force. The latter drives ATP synthesis through the activity of a remarkable and extremely small cellular motor, and we consider this now.
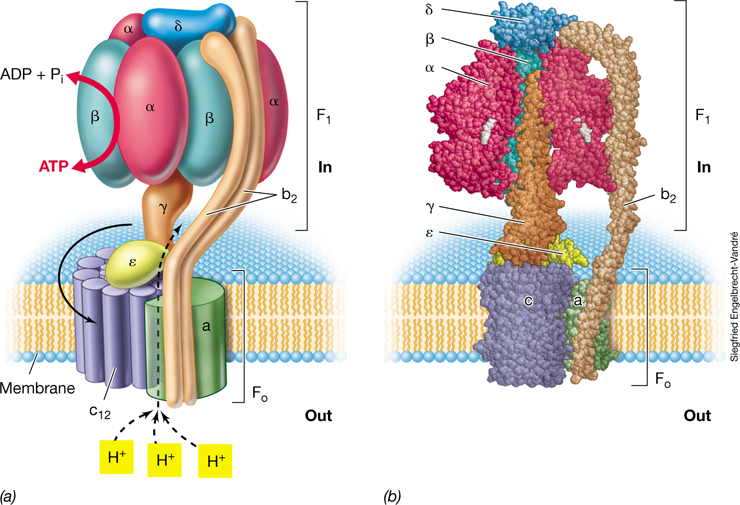
ATP Synthase
How does the proton motive force (pmf), generated from electron transport, actually drive ATP synthesis? Interestingly, a strong parallel exists between the mechanism of ATP synthesis and the mechanism that drives rotation of the flagellar motor in swimming bacteria (Section 2.9). A large protein complex called ATP synthase (ATPase) uses energy from the pmf to catalyze formation of ATP (Figure 3.20). As it does for the flagellar motor, the pmf generates torque within the ATP synthase enzyme complex, and this torque is translated into rotary motion, creating mechanical energy that is used to catalyze the chemical synthesis of ATP from ADP and Pi. ATP formation by ATP synthase is called oxidative phosphorylation when the pmf is generated by respiratory electron flow. In contrast, ATP formation by ATP synthase is called photophosphorylation when the pmf is generated from light energy during phototrophy (Section 3.11).
Figure 3.20 Structure and function of the reversible ATP synthase (ATPase) in *Escherichia coli.*

(a) Schematic. F1, the stator, consists of five different polypeptides forming an α3β3γεΔ complex. F1 is the catalytic complex responsible for the interconversion of ADP+Pi and ATP. Fo, the rotor, is integrated in the membrane and consists of three polypeptides in an ab2c12 complex. As protons enter, the dissipation of the proton motive force drives ATP synthesis (3–5 H+/ATP). (b) Space-filling model. The color-coding corresponds to the art in part a. Since proton translocation from outside the cell to inside the cell leads to ATP synthesis by ATPase, it follows that proton translocation from inside to outside in the electron transport chain (Figure 3.19) represents work done on the system and a source of potential energy. This energy can be used to drive a number of energy-requiring processes in the cell, including in particular motility and some forms of nutrient transport. Nearly all cells of all domains contain ATPases. Also, some bacterial and archaeal ATPases are linked to a sodium (Na+) rather than a proton (H+) gradient.
ATPases consist of two components, a multiprotein complex called F1 that sticks into the cytoplasm and actually catalyzes ATP synthesis, and a membrane-integrated multiprotein complex called Fo that carries out proton translocation across the membrane (Figure 3.20). ATP synthase is found in nearly all cellular organisms, and this enzyme complex is highly conserved, indicating that it is of ancient origin and was likely present in the common ancestor of all cells (Chapter 13).
ATPase catalyzes a reversible reaction between ATP and ADP+Pi, and F1 and Fo are actually two rotary motors. The movement of protons through the ATPase drives the rotation of its c protein subunits, which are organized in a ring within the Fo subunit. Each c protein subunit picks up one H+ from outside the membrane and deposits it in the cytoplasm for every complete revolution. This generates a torque that is transmitted to F1 via the coupled rotation of the γε subunits (Figure 3.20). The rotation causes conformational changes in the β subunits of F1 that allows them to bind ADP+Pi. ATP is synthesized when the β subunits return to their original conformation and the free energy captured in the rotated β subunits is released. For every full rotation of the c ring within the Fo subunit, three ATP are formed by the F1 subunit. The ATPase of E. coli has 10 c protein subunits and as a result, approximately 3.3 H+ are required to synthesize one ATP. However, the number of c protein subunits within an ATPase varies from 8 to 15 in different organisms and thus the number of protons needed to synthesize one ATP also varies.
Oxidative phosphorylation allows the cell to conserve far more energy than is possible in fermentation because the substrate, for example glucose, is completely oxidized (Figure 3.21). A total of 38 ATP are synthesized during the aerobic respiration of glucose to CO2, with 34 of these formed as a result of oxidative phosphorylation (Figure 3.21). By contrast, in lactic acid fermentation, only 2 ATP are produced by substrate-level phosphorylation during the oxidation of glucose in glycolysis. Lactic acid bacteria excrete lactate, which still contains considerable free energy that has not been conserved by the cell.
Figure 3.21 Energetics in fermentation and aerobic respiration.

Fermentation and respiration both oxidize NADH and allow for the regeneration of NAD+. However, respiration conserves far more energy than fermentation as a result of oxidative phosphorylation. Note that reduction of 6 O2 with 24 e− produces 12 H2O while only 6 H2O are produced in the net reaction. The difference between the gross and net production of water occurs because H2O is in equilibrium with H+ and OH− and only the net production of water is written in the balanced reaction. SLP, substrate-level phosphorylation; Ox. Phos., oxidative phosphorylation; CAC, citric acid cycle; ET, electron transport.
ATPases are reversible motors. Hence, ATP hydrolysis can reverse the activity of ATPase and cause protons to be transported out of the cytoplasm. The net result in this case is generation of rather than dissipation of the proton motive force. Reversibility of the ATPase explains why strictly fermentative bacteria still contain ATPases even though they lack electron transport chains and are unable to carry out oxidative phosphorylation. Many important reactions in the cell, such as flagellar rotation and some forms of transport, are coupled to energy from the proton motive force. Thus, in these strictly fermentative organisms, ATPase hydrolyzes ATP formed from substrate-level phosphorylation (Section 3.7) to generate the pmf needed to carry out cellular functions such as motility and transport.
Check Your Understanding
How do electron transport reactions generate the proton motive force?
How much energy is released per NADH oxidized through the electron transport chain of Paracoccus shown in Figure 3.19?
What structure in the cell links the proton motive force to ATP synthesis? How does it function?
III: Catabolism: Electron Transport and Metabolic Diversity
III: Catabolism: Electron Transport and Metabolic Diversity
III Catabolism: Electron Transport and Metabolic Diversity
The respiration of organic or inorganic electron donors and the light-driven reactions of photosynthesis all generate ATP by harvesting energy from the proton motive force that forms from electron transport reactions in the cytoplasmic membrane.
With the exception of fermentation, in which energy is conserved by substrate-level phosphorylation (Section 3.7), all other mechanisms of energy conservation are linked to the proton motive force (or in a few organisms, a gradient of sodium ions, Na+, instead of protons, as we will see in Chapter 14). Whether energy comes from chemical reactions or light, and whether electrons come from the oxidation of organic or inorganic chemicals, nonfermentative energy conservation is always the result of electron transport reactions and the formation of a proton motive force (see Figure 3.22). The proton motive force is then used by ATPase to form ATP (Figure 3.20).
Figure 3.22 Metabolic diversity and its relationship to oxygen.

(a) Chemoorganotrophs can be fermentative or engage in aerobic or anaerobic respiration, whereas (b) chemolithotrophs are typically respiratory. (c) Phototrophs are oxygenic if they generate O2 by oxidizing H2O and anoxygenic if they do not. Pmf, proton motive force.
In earlier parts of this chapter we learned about the fundamental metabolic principles that govern aerobic and fermentative chemoorganotrophs. Now we turn our attention to how these same fundamental principles apply to the vast diversity of metabolic types found in the microbial world. We return to this theme of catabolic diversity and explore its many facets in Chapter 14.
3.10 Anaerobic Respiration and Metabolic Modularity
3.10 Anaerobic Respiration and Metabolic Modularity
3.10 Anaerobic Respiration and Metabolic Modularity
Respiration can occur under both oxic (O2 is present) and anoxic (O2 is absent) conditions. The distinguishing feature between aerobic and anaerobic respiration is that aerobic respiration requires O2 as the terminal electron acceptor, whereas anaerobic respiration uses electron acceptors other than O2 (Figure 3.22). Recall that when life began there was no O2 in the atmosphere, and O2 did not reach current levels until about 600 million years ago (Section 1.5). This means that anaerobic microbes dominated much of the history of life, and these organisms still occur widely today in any environment that lacks O2, such as wetlands, sediments, and in the guts of animals.
Fermentative organisms and microbes that perform anaerobic respiration can both grow under anoxic conditions, and in this sense they are both anaerobes. However, these two forms of metabolism are distinct because all types of respiration require an external electron acceptor and generate ATP primarily by oxidative phosphorylation, whereas all types of fermentation do not require an external electron acceptor and generate ATP by substrate-level phosphorylation. We examine respiratory options in Escherichia coli now.
Respiration in *Escherichia coli E. coli* is a metabolically versatile chemoorganotroph whose electron transport chain is similar to but not identical with that of *Paracoccus denitrificans* (Figure 3.19). *E. coli* can grow by aerobic respiration in the presence of O2, it can carry out fermentation in the absence of external electron acceptors, and it can grow by anaerobic respiration when nitrate is present in the absence of O2. As a result, *E. coli* can optimize its metabolism to grow under a range of conditions. When using an organic carbon source, *E. coli* will grow most rapidly by aerobic respiration, but under anaerobic conditions, as in an animal gut or in sediments, it can grow more rapidly by nitrate respiration than by fermentation, provided that nitrate is available. The energy *E. coli* can conserve during respiration is a function of the organization of its electron transport chain. *E. coli* can insert many different proteins in its electron transport chain, and this versatility allows it to conserve energy through respiration with a range of different electron donors and acceptors.
The basic organization of the E. coli respiratory chain (Figure 3.23) is Complex I followed by Complex II followed by quinones (for example, ubiquinone) and a terminal reductase that contains b-type and o-type cytochromes (in contrast to the a-type cytochromes of Complex IV). Depending on environmental conditions E. coli can swap out these components, employing alternative quinones (for example, ubiquinone or menaquinone), or employing alternative dehydrogenases or terminal reductases (E. coli has at least five sets of each). Note, however, that Complex III is missing from E. coli (Figure 3.23); this has energetic consequences.
Figure 3.23 Respiration and nitrate-based anaerobic respiration in *Escherichia coli*.

Electron transport processes in the membrane of E. coli when (a) O2 or (b) NO3 − is used as terminal electron acceptor. Note the difference in the number of protons translocated with the different terminal electron acceptors and compare these to the difference in the energetics of these reactions when glucose is the electron donor (see Figure 3.4). Q, ubiquinone.
The lack of Complex III means that during aerobic respiration, E. coli conserves less energy than P. denitrificans. For every 2 electrons that pass through its electron transport chain to O2, E. coli exchanges only 8 H+ across the membrane while P. denitrificans exchanges 10 H+ (compare Figures 3.19 and 3.23a). The free energy available to P. denitrificans and E. coli during the aerobic respiration of glucose is identical, but the organization of the electron transport chain in P. denitrificans allows it to conserve more energy than E. coli. This means that P. denitrificans will typically be able to outcompete E. coli for glucose if they are both growing aerobically. Hence, we can see that the amount of energy conserved by a respiratory organism is influenced by both bioenergetics and by the organization of its electron transport chain.
If nitrate is present in the absence of O2, E. coli will use nitrate reductase as its terminal reductase (Figure 3.23b). Nitrate reductase allows E. coli to perform anaerobic respiration with nitrate. However, the NO3 −/NO2 − couple is less electropositive than is the O2/H2O couple (Figure 3.4), and this is why anaerobic respiration provides less energy than aerobic respiration. This bioenergetic difference is reflected in the energy that can be conserved in electron transport, because only 6 H+ are exchanged across the membrane for every 2 electrons passing to nitrate (rather than the 8 H+ extruded aerobically, Figure 3.23). Likewise, for any given electron donor, aerobic organisms will always be able to conserve more energy—and will therefore outcompete—anaerobic organisms. However, oxygen is consumed very rapidly in poorly mixed aqueous environments because it is such a good electron acceptor and because it is poorly soluble in water. Hence, anoxic habitats and anaerobic microbes are widespread in nature and crucial to global biogeochemical cycles (Chapter 21).
Check Your Understanding
If three bacteria were competing for glucose and one was capable only of aerobic respiration, one anaerobic respiration with nitrate ( NO3 −) as electron acceptor, and one anaerobic respiration with sulfate (SO4 2−) as electron acceptor, which do you think would be most competitive in an oxic environment? In an anoxic environment? (Hint: Review Figure 3.4 and Table 3.1.)
Explain why Paracoccus denitrificans would outcompete Escherichia coli if both organisms were competing for glucose in the presence of O2.
If respiratory metabolism always conserves more energy than fermentative metabolism, then why are fermentative organisms so common?
3.11 Chemolithotrophy and Phototrophy
Thus far in this chapter we have focused on chemoorganotrophs, chemotrophic organisms that rely on organic electron donors. Now we will focus on chemolithotrophs, which use inorganic electron donors, and on phototrophs, which get their energy from light (Figure 3.3). Chemolithotrophs can be aerobes or anaerobes and they are found in a wide range of environments, most anywhere that reduced inorganic compounds are present.
Chemolithotrophy
Examples of common inorganic electron donors that fuel chemolithotrophic metabolisms include H2S, H2, Fe2+, and NH4 + (Figure 3.22). Chemolithotrophic metabolisms begin with the oxidation of an inorganic electron donor and with the electrons entering an electron transport chain. For example, the hydrogen bacterium Ralstonia eutropha grows as an aerobic chemolithotroph with H2 as its electron donor (2 H+/H2,−0.42 V). R. eutropha has a membrane-bound hydrogenase that splits H2 in the periplasm and reduces quinones in the electron transport chain (Figure 3.24). Electron transport then generates a proton motive force, as we have already seen for the oxidation of organic electron donors by chemoorganotrophs (Figures 3.19 and 3.23). Cells of R. eutropha actually have two hydrogenases (Figure 3.24). The membrane-bound hydrogenase contributes to energy conservation, as we have described, whereas the soluble hydrogenase, which is present in the cytoplasm, is used to reduce NAD+ to NADH needed for autotrophic growth (Figure 3.24). This reduction is possible because the E0′ of the 2 H+/H2 couple is more electronegative than the E0′ of the NAD+/NADH couple (Figure 3.4).
Figure 3.24 Energy conservation in ***Ralstonia eutropha***, an autotrophic chemolithotroph that oxidizes H2.

(a) In R. eutropha, two hydrogenases are present; the membrane-bound hydrogenase participates in electron transport, whereas the soluble hydrogenase makes NADH for the Calvin cycle (see Section 3.12). Cyt, cytochrome; Q, quinone. For details of the Q-cycle, see Figure 3.19. (b) Transmission electron micrograph of negatively stained cells of R. eutropha. A cell is about 0.6 μm in diameter and contains several flagella.
Respiration by chemoorganotrophs and chemolithotrophs are united by their dependence on oxidative phosphorylation for energy conservation. Just as we saw for anaerobic respiration, the energy available to chemolithotrophs depends on the ΔE0′ between the electron-donating and electron-accepting reactions (Figure 3.4). Like many chemoorganotrophs, many chemolithotrophs can have either aerobic or anaerobic metabolisms (Figure 3.22). Indeed, almost any combination of electron donor and electron acceptor can sustain life if these reactions are coupled to an electron transport chain used in oxidative phosphorylation and if the ΔE0′ of the redox reaction releases sufficient free energy to form ATP.
One major difference between chemoorganotrophs and chemolithotrophs is in their source of cellular carbon. Chemoorganotrophs are heterotrophs and therefore use organic compounds (glucose, acetate, etc.) as carbon sources. These organic compounds are readily converted into cellular materials during biosynthetic reactions. By contrast, chemolithotrophs typically use carbon dioxide (CO2) as a carbon source and are therefore autotrophs (we describe a major autotrophic pathway—the Calvin cycle—in Section 3.12). Autotrophy requires a significant amount of reducing power (Figure 3.1) such as NADH, but the electron donors used by chemolithotrophs often have a more electropositive E0′ than does NADH/NAD+ (Figure 3.4). How do such chemolithotrophs form NADH? In the case of R. eutropha, we saw that a cytoplasmic hydrogenase is used to form NADH directly from NAD+ and H2 (Figure 3.24). However, most chemolithotrophs lack hydrogenases and solve this problem by employing reverse electron transport, an energy-consuming process that we will learn about in the context of phototrophy.
Phototrophy
In the process of photosynthesis, carried out by phototrophs, light energy is used instead of chemical reactions to drive electron flow and generate a proton motive force. During these events, ATP synthase generates ATP by photophosphorylation (**Figure 3.25*a***), the light-driven analog of oxidative phosphorylation. Phototrophs can be either oxygenic (for example cyanobacteria, algae, and plants) or anoxygenic (diverse types of Bacteria), depending on whether or not the process generates O2 as a waste product from the oxidation of H2O (Figure 3.22). Anoxygenic phototrophs evolved long before oxygenic phototrophs (Section 1.5), and there is far more metabolic diversity among the anoxygenic phototrophs than among oxygenic phototrophs. However, light-driven photoreactions in both oxygenic and anoxygenic phototrophs operate on many of the same principles.
Figure 3.25 Electron flow in anoxygenic photosynthesis in a purple bacterium.

(a) Energy conservation in purple bacteria results from cyclic photophosphorylation. The photosynthetic reaction center uses light energy to energize electrons for use in electron transport and the generation of proton motive force. Four protons are translocated for every two electrons that pass from the reaction center to cytochrome bc1. P870 is a form of bacteriochlorophyll a in the ground state and is a fairly electropositive electron donor. P870∗ is formed after light excitation of P870 and is a strongly electronegative electron donor. LH, light-harvesting bacteriochlorophyll complexes; Bph, bacteriopheophytin; Q, quinone; c2, cytochrome c2. For details of the Q-cycle, see Figure 3.19. (b) A bloom of purple bacteria that formed in the Sippewissett Salt Marsh, Cape Cod, Massachusetts, USA. High rates of O2 consumption, and its low solubility in water, can cause anoxic conditions in a water film less than 100 μm thick, and this allows anoxygenic phototrophs to grow on the sediment surface.
Purple bacteria are one type of anoxygenic phototrophs, which are common in anoxic aquatic environments (Figure 3.25b). Like all phototrophic organisms, purple bacteria produce a photosynthetic reaction center that converts light energy into chemical energy (ATP, Figure 3.25a).While we focus here primarily on energy conservation during photophosphorylation, we will learn more about the operation of photosynthetic reaction centers in Chapter 14. All reaction centers contain photopigments such as chlorophylls or bacteriochlorophylls, which are structurally similar to heme molecules (Section 14.3).
Phototrophs use photopigments to absorb light energy and transfer this energy to the photosynthetic reaction center (Figure 3.25a). The reaction center in purple bacteria uses the energy from light to convert a form of bacteriochlorophyll a called P870, which is a relatively weak electron donor, into a very strong electron donor (E0′ near −1.0 V) called P870.The excited P870 then donates two electrons to a quinone within the cytoplasmic membrane. The reduced quinone in turn donates its electrons to the cytochrome bc1 complex, which is similar in structure and function to the Complex III we saw in oxidative phosphorylation (Figure 3.19). The net result is photophosphorylation, which results when light energy is used to generate a proton motive force that is harnessed by ATP synthase to make ATP (Figure 3.25a).
A major difference between respiratory electron transport and photosynthetic electron transport in purple bacteria is that in purple bacteria electrons from cytochrome bc1 are not transferred to an external electron acceptor (such as O2). Instead the electrons are passed to cytochrome c2 and then back to P870, returning it to its original E0′. Hence, purple bacteria reuse the same electrons over and over again to make ATP as long as light is present to fuel the initial P870→P870∗ transition. This process is called cyclic photophosphorylation because electrons move within a closed loop. We consider other aspects of photosynthesis, including the differences between anoxygenic photosynthesis and oxygenic photosynthesis, in Chapter 14.
Generation of Reducing Power
For purple bacteria and chemolithotrophs to grow autotrophically, the formation of ATP is not enough. Reducing power (NADH) is also necessary to produce cellular material. Reducing power can come from many different electron donors. Many purple bacteria and chemolithotrophs use reduced sulfur compounds such as H2S as their electron donors. However, the E0′ of S0/H2S (−0.28 V) is insufficiently electronegative to reduce NAD+ (−0.32 V); hence, these bacteria cannot make NADH directly from H2S. These organisms solve this problem by pushing electrons from the quinone pool backwards (against the electrochemical gradient) towards a dehydrogenase-like protein (similar to Complex I) so that they reduce NAD+ to NADH. This process, called reverse electron transport, requires energy and is driven by dissipation of the proton motive force. We will see this pattern once more when we discuss biosynthetic reactions, in which the reversibility of reactions and pathways is often used to synthesize a great diversity of biomolecules.
Check Your Understanding
What traits are shared between oxidative phosphorylation and photophosphorylation?
Which process do you think evolved first, photosynthesis or respiration? Use your knowledge of metabolism to defend your answer.
What is reverse electron transport, and why is it unnecessary for chemoorganotrophs?
IV Biosynthesis
Biosynthesis encompasses the anabolic reactions that produce cellular materials and requires ATP and building blocks composed of C, N, and other elements obtained from organic or inorganic sources.
We conclude this chapter with an overview of how the monomers that make up the four classes of cellular macromolecules are biosynthesized and also look at how polysaccharides and lipids are biosynthesized in a general way. The biosynthesis of informational macromolecules—proteins and nucleic acids—is the theme of Chapter 6. Collectively, these biosynthetic reactions are that aspect of metabolism we call anabolism.
All cells require a source of carbon and nitrogen to perform biosynthetic reactions. The atmosphere contains a large reservoir of inorganic carbon as CO2, and nitrogen as N2. However, these gases must be chemically reduced before they can be assimilated into cell material. The reductive processes associated with the assimilation of CO2 and N2 are called CO2 fixation and N2 fixation, respectively. Both of these pathways require substantial amounts of energy from the cell in the form of ATP and reducing power. These processes evolved very early in the history of life and they are widely distributed among species of Bacteria and Archaea.
3.12 Autotrophy and Nitrogen Fixation
Many phototrophic and chemolithotrophic microbes get the carbon they need, and in some cases the nitrogen they need, by reducing CO2 and N2, respectively, from air. Because CO2 and N2 are such highly oxidized molecules, their assimilation into cell material requires large amounts of ATP and reducing power (Figure 3.1). Here we consider the assimilation of CO2—autotrophy—only in the context of the Calvin cycle. In Chapter 14 we will see that autotrophy shows considerable metabolic diversity beyond the Calvin cycle.
The Calvin Cycle
In many autotrophic organisms, including all oxygenic phototrophs, the carbon needed for biosynthesis is derived from the reduction of CO2 by the Calvin cycle (Figure 3.26). The Calvin cycle is present in phototrophic purple bacteria (Figure 3.25b), cyanobacteria, algae, green plants, most chemolithotrophic Bacteria, and a few Archaea. The cycle requires CO2, a CO2-acceptor molecule, NADPH, ATP, and two key enzymes, ribulose bisphosphate carboxylase and phosphoribulokinase.
Figure 3.26 Key reactions of the Calvin cycle.

(a) Reaction of the enzyme ribulose bisphosphate carboxylase. (b) Steps in the conversion of 3-phosphoglycerate (PGA) to glyceraldehyde 3-phosphate. Note that both ATP and NADPH are required. (c) Conversion of ribulose 5-phosphate to the CO2 acceptor molecule ribulose 1,5-bisphosphate by the enzyme phosphoribulokinase.
The first step in the Calvin cycle is catalyzed by the enzyme ribulose bisphosphate carboxylase, RuBisCO for short. RuBisCO catalyzes the formation of two molecules of 3-phosphoglycerate (PGA, a C3 compound) from ribulose bisphosphate (a C5 compound) and CO2 as shown in Figure 3.26a. The PGA is then phosphorylated and reduced to a key intermediate of glycolysis, glyceraldehyde 3-phosphate. From this, glucose can be formed by reversal of the early steps in glycolysis (Figure 3.11).
Instead of focusing on the incorporation of a single molecule of CO2, it is easiest to consider Calvin cycle reactions based on the incorporation of 6 molecules of CO2, as this is what is required to make one hexose (C6H12O6). For RuBisCO to incorporate 6 molecules of CO2, 6 molecules of ribulose bisphosphate (total, 30 carbons) are required; carboxylation of these yields 12 molecules of PGA (total, 36 carbons) (Figure 3.27). A series of biochemical rearrangements between various sugars follows, resulting in 6 molecules of ribulose 5-phosphate (30 carbons) plus one hexose (6 carbons) for cell biosynthesis. The final step in the Calvin cycle is the phosphorylation of each ribulose 5-phosphate by the enzyme phosphoribulokinase (Figures 3.26c and 3.27) to regenerate 6 molecules of the acceptor molecule, ribulose bisphosphate. All totaled, 12 NADPH and 18 ATP are required to synthesize one glucose from 6 CO2 by the Calvin cycle.
Figure 3.27 The Calvin cycle.

Shown is the production of one hexose molecule from six CO2. For each six molecules of CO2 incorporated, one fructose 6-phosphate is produced that can be drawn off for biosynthesis. In phototrophs, the energy to produce ATP comes from light; in chemolithotrophs, the energy to produce ATP comes from the oxidation of inorganic electron donors. In oxygenic phototrophs, electrons for NAD(P)H come from water; in anoxygenic phototrophs, the electrons come from reduced substances such as H2S. In chemolithotrophs, NAD(P)H is formed from inorganic electron donors.
Although the Calvin cycle is the most widespread and important pathway of CO2 fixation in the biosphere, many autotrophic Bacteria and Archaea have evolved alternative pathways for fixing CO2. These alternative autotrophic pathways typically reduce CO2 to the level of acetyl-coenzyme A (acetyl-CoA), a central metabolite that feeds into all major biosynthetic pathways (Section 3.6).We will learn about these alternative pathways for CO2 fixation in Chapter 14 when we once again consider metabolic diversity.
Nitrogen Fixation
In addition to carbon, cells need a significant amount of nitrogen to synthesize proteins, nucleic acids, and many other organic molecules. Most microbes obtain this nitrogen from “fixed” forms of nitrogen in their environment, such as ammonia (NH3) or nitrate (NO3 −). However, many Bacteria and Archaea can form ammonia from gaseous dinitrogen (N2), a process called nitrogen fixation (Figure 3.28). The process of nitrogen fixation is of enormous ecological and agricultural importance (Section 1.6) as it provides much of the fixed nitrogen that supports global ecosystems and is important to leguminous crops, such as soybeans, lentils, and alfalfa, which form symbiotic associations with nitrogen-fixing bacteria (Section 23.4).
Figure 3.28 Biological nitrogen fixation by nitrogenase.

The nitrogenase complex is composed of dinitrogenase and dinitrogenase reductase. Electrons from reduced flavodoxin (or ferredoxin) are used to reduce dinitrogenase reductase, and this enzyme in turn reduces dinitrogenase at the expense of ATP. Dinitrogenase ultimately donates electrons to N2 at the active site of the enzyme, resulting in the formation of 2 NH3. H is hydrogen (which consists of H++e−).
Nitrogen fixation is catalyzed by the enzyme complex nitrogenase. Nitrogenase consists of two proteins, dinitrogenase and dinitrogenase reductase (Figure 3.28). Both proteins contain iron, and dinitrogenase contains molybdenum as well. The iron and molybdenum in dinitrogenase are part of the enzyme cofactor called the iron–molybdenum cofactor (FeMo-co), and reduction of N2 occurs at this site. Owing to the stability of the triple bond in N2, its activation and reduction is very energy demanding. Six electrons are needed to reduce N2 to NH3, and the successive reduction steps occur directly on nitrogenase with no free intermediates accumulating. The electrons for N2 reduction are transferred to dinitrogenase reductase from low-potential iron–sulfur proteins such as flavodoxin (or ferredoxin) (Table 3.1). The sequence of electron transfer in nitrogenase is: electron donor→dinitrogenase reductase→dinitrogenase→N2 (Figure 3.28). Although only six electrons are necessary to reduce N2 to two NH3, eight electrons are actually consumed in the process, two electrons being lost as H2 for each molecule of N2 reduced. For unknown reasons, H2 evolution is an obligatory step in nitrogen fixation and occurs in the first step of the nitrogenase reduction cycle.
In addition to electrons, ATP is required for nitrogen fixation. ATP binds to dinitrogenase reductase, and, following its hydrolysis to ADP, lowers the reduction potential of the protein. This allows dinitrogenase reductase to interact with and reduce dinitrogenase. Electrons are transferred from dinitrogenase reductase to dinitrogenase one at a time, and each cycle of reduction requires two ATP. Thus, a total of 16 ATP are required for the reduction of N2 to 2 NH3 (Figure 3.28).
Nitrogenase is inhibited by oxygen (O2), but many nitrogen-fixing bacteria are obligate aerobes. In these organisms, nitrogenase is protected from oxygen inactivation by a combination of the rapid removal of O2 by respiration and the production of O2-retarding slime layers on the outer cell surface (Figure 3.29). In certain cyanobacteria, the nitrogenase enzyme is protected from O2 by its localization in a differentiated cell called a heterocyst (Figure 3.29c; Section 15.3). Inside the heterocyst, conditions are anoxic despite the fact that photosynthesis and O2 production occur in neighboring vegetative cells. Oxygen is not produced in the heterocyst, and this protects nitrogenase so that it can fix N2 into organic nitrogen compounds, which are then shared with adjacent photosynthetic cells (Section 8.9).
Figure 3.29 Two ways of protecting nitrogenase from O2.

Induction of slime formation by O2 is demonstrated by comparing transmission electron micrographs of (a) nitrogen-fixing Azotobacter vinelandii cells grown with 2.5% O2 and showing very little slime with (b) nitrogen-fixing A. vinelandii cells grown in air (21% O2) and showing an extensive, darkly staining slime layer (arrow). The slime retards diffusion of O2 into the cell, thus preventing nitrogenase inactivation by O2. A single cell of A. vinelandii is about 2 μm in diameter. (c) Fluorescence photomicrograph of cells of the filamentous cyanobacterium Anabaena showing a single heterocyst (green). The heterocyst is a thick-walled differentiated cell that does not produce O2 but instead specializes in nitrogen fixation and protects nitrogenase from O2 inactivation.
Check Your Understanding
How many electrons and how much ATP are required to make one hexose molecule by the Calvin cycle?
How many electrons and how much ATP are required to fix one molecule of N2 into two of NH3?
Describe two ways in which N2-fixing organisms can protect their nitrogenase from O2.
3.13 Sugars and Polysaccharides
Polysaccharides are key components of microbial cell walls, and cells often store carbon and energy reserves in the form of the polysaccharides glycogen or starch (Chapter 2). How are such large molecules made?
Polysaccharide Biosyntheses and Gluconeogenesis
Polysaccharides are synthesized from activated forms of glucose, either uridine diphosphoglucose (UDPG; **Figure 3.30*a***) or adenosine diphosphoglucose (ADPG). UDPG is the precursor of several glucose derivatives used in the biosynthesis of important structural polysaccharides, such as N-acetylglucosamine and N-acetylmuramic acid in peptidoglycan or the lipopolysaccharide component of the gram-negative outer membrane (Sections 2.3 and 2.4). Polysaccharides are biosynthesized by adding activated glucose to a preexisting polymer fragment. For example, glycogen is synthesized as ADPG+glycogen→ADP+glycogen-glucose.
Figure 3.30 Sugar metabolism.
(a) Polysaccharides are synthesized from activated forms of hexoses such as UDPG. (b) Gluconeogenesis. When glucose is needed, it can be biosynthesized from other carbon compounds by reversing the steps in glycolysis. (c) Pentoses for nucleic acid synthesis are formed by decarboxylation of hexoses such as glucose 6-phosphate. Note how the precursors of DNA are produced from the precursors of RNA by the enzyme ribonucleotide reductase.
When a cell is growing on a hexose such as glucose, obtaining glucose for polysaccharide synthesis is obviously not a problem. But when the cell is growing on other carbon compounds, glucose must be biosynthesized. This process, called gluconeogenesis, uses phosphoenolpyruvate, one of the intermediates of glycolysis, as a starting material and travels backwards through the glycolytic pathway (Figure 3.11) to form glucose. Phosphoenolpyruvate can also be synthesized from oxaloacetate, a citric acid cycle intermediate (Figure 3.12). An overview of gluconeogenesis is shown in Figure 3.30b.
Pentose Metabolism and the Pentose Phosphate Pathway
Pentoses (C5 sugars) are formed by the removal of one carbon atom from a hexose, typically as CO2. The pentoses needed for nucleic acid synthesis, ribose (in RNA) and deoxyribose (in DNA), are formed as shown in Figure 3.30c. The enzyme ribonucleotide reductase converts ribose into deoxyribose by reduction of the hydroxyl (−OH) group on the 2′ carbon of the 5-carbon pentose ring (Section 6.1 and Figure 6.1b). This reaction occurs after, not before, synthesis of nucleotides. Thus, ribonucleotides are biosynthesized, and some of them are later reduced to deoxyribonucleotides for use as precursors of DNA.
The major pathway for pentose production is the pentose phosphate pathway (Figure 3.31). In this pathway, glucose, a hexose, is oxidized to CO2, NADPH, and the key intermediate, ribulose 5-phosphate; from the latter, several different pentose derivatives can be formed. When pentoses are used as electron donors for energy conservation, they feed directly into the pentose phosphate pathway, typically becoming phosphorylated to form ribose phosphate or a related compound before being further catabolized (Figure 3.31b).
Figure 3.31 Pentose phosphate pathway.

This pathway generates pentoses from other sugars for biosynthesis and also functions to catabolize pentose sugars. (a) Production of the key intermediate, ribulose 5-phosphate. (b) Other reactions in the pentose phosphate pathway.
Besides its importance in pentose metabolism, the pentose phosphate pathway is also responsible for producing many other important sugars in the cell, including those containing 4 to 7 carbons. These sugars can eventually be converted to hexoses for either catabolic or biosynthetic purposes (Figures 3.30 and 3.31). A final important role of the pentose phosphate pathway is that it generates NADPH, a coenzyme used in many biosyntheses and in particular as a reductant for the production of deoxyribonucleotides (Figure 3.30c) and in the biosynthesis of fatty acids (see Figure 3.35). Although most cells have an exchange mechanism for converting NADH into NADPH, the pentose phosphate pathway is the major means by which NADPH is synthesized directly.
Check Your Understanding
What form of activated glucose is used in the biosynthesis of glycogen by bacteria?
What functions does the pentose phosphate pathway play in the cell?
3.14 Amino Acids and Nucleotides
The monomers in proteins and nucleic acids are the amino acids and nucleotides, respectively. Their biosyntheses are typically multistep biochemical pathways that we need not consider in detail here. Instead, we identify the carbon skeletons needed for the biosynthesis of amino acids and nucleotides, look at their origins in pathways we have already considered, and summarize the mechanisms by which they are made.
Monomers of Proteins: Amino Acids
Organisms that cannot obtain some or all of their amino acids preformed from the environment must synthesize them from glucose or other carbon sources. Amino acids are grouped into structurally related families that share several biosynthetic steps. The carbon skeletons for amino acids come almost exclusively from intermediates of glycolysis (Figure 3.11) or the citric acid cycle (Figure 3.12) (Figure 3.32).
Figure 3.32 Amino acid families.

Glycolysis (a) and the citric acid cycle (b) provide the carbon skeletons for most amino acids. Synthesis of the various amino acids in a family may require many steps starting with the parent amino acid (shown in bold as the name of the family).
The amino group (−NH2) of amino acids is typically derived from an inorganic nitrogen source, such as ammonia (NH3). Ammonia is most often incorporated during biosynthesis of the amino acids glutamate or glutamine by the enzymes glutamate dehydrogenase and glutamine synthetase, respectively (Figure 3.33). When NH3 is present at high levels, glutamate dehydrogenase or other amino acid dehydrogenases are used. However, when NH3 is present at low levels, glutamine synthetase, with its energy-consuming reaction mechanism (Figure 3.33b) and correspondingly high affinity for its substrates, is employed. The enzymes glutamate dehydrogenase and glutamine synthetase are present in most Bacteria and Archaea.
Figure 3.33 Ammonia incorporation in bacteria.

Ammonia (NH3) and the amino groups of all amino acids are shown in green. Two major pathways for NH3 assimilation in bacteria are those catalyzed by the enzymes (a) glutamate dehydrogenase and (b) glutamine synthetase. (c) Transaminase reactions transfer an amino group from an amino acid to an organic acid. (d) The enzyme glutamate synthase forms two glutamates from one glutamine and one α-ketoglutarate.
Once ammonia is incorporated into glutamate or glutamine, the amino group of these amino acids can be transferred to other carbon skeletons to form other nitrogenous compounds. For example, glutamate can donate its amino group to oxaloacetate in a transaminase reaction, producing α-ketoglutarate and aspartate (**Figure 3.33*c***). Alternatively, glutamine can react with α-ketoglutarate to form two molecules of glutamate in an aminotransferase reaction (Figure 3.33d). The end result of these types of reactions is the shuttling of ammonia into various carbon skeletons from which further biosynthetic reactions occur to form all of the amino acids needed to make proteins and other nitrogen-containing biomolecules.
Monomers of Nucleic Acids: Nucleotides
The biochemistry behind purine and pyrimidine biosynthesis is quite complex and so only an outline of their biosyntheses is necessary here. Purines are constructed literally atom by atom from several carbon and nitrogen sources, including even CO2 (Figure 3.34). The purine nucleotide skeleton, inosinic acid (Figure 3.34b), is the precursor of the purine nucleotides adenine and guanine. Once these are synthesized (in their triphosphate forms) and attached to ribose, they are ready to be incorporated into DNA (following ribonucleotide reductase activity, Figure 3.30c) or RNA.
Figure 3.34 Biosynthesis of purines and pyrimidines.

(a) Components of the purine skeleton, labeled with their sources. (b) Inosinic acid, the precursor of all purine nucleotides. (c) Components of the pyrimidine skeleton, orotic acid, labeled with their sources. (d) Uridylate, the precursor of all pyrimidine nucleotides. Uridylate is formed from orotate following a decarboxylation and the addition of ribose 5-phosphate.
Like the purine ring, the pyrimidine ring is also constructed from several sources (Figure 3.34c). From the pyrimidine nucleotide skeleton uridylate (Figure 3.34d), all of the pyrimidines—thymine, cytosine, and uracil—are derived.Structures of all the purines and pyrimidines are shown in Chapter 6 (Figure 6.1c).
Check Your Understanding
List the steps required for the cell to incorporate NH3 into amino acids.
Which nitrogen bases are purines and which are pyrimidines?
3.15 Fatty Acids and Lipids
Lipids are major components of the cytoplasmic membrane and of the outer membrane of gram-negative bacteria; lipids can also be carbon and energy reserves. Fatty acids are the backbone of microbial lipids. However, recall that fatty acids, per se, are found only in Bacteria and Eukarya. Archaea do not contain fatty acids in their lipids but instead have hydrophobic isoprenoid side chains that play a similar structural role (Chapter 2). Our focus here is on the biosynthesis of fatty acids in Bacteria.
Fatty Acid Biosynthesis
Fatty acids are biosynthesized two carbon atoms at a time by the activity of a protein called acyl carrier protein (ACP). ACP holds the growing fatty acid as it is being constructed and releases it once it has reached its final length (Figure 3.35). Although fatty acids are constructed two carbons at a time, each C2 (acetyl) unit originates from the three-carbon compound malonate, which is attached to the ACP to form malonyl-ACP. As each malonyl residue is donated, one molecule of CO2 is released (Figure 3.35).
Figure 3.35 The biosynthesis of the C16 fatty acid palmitate.

The condensation of acetyl-ACP (which is formed from acetyl-CoA) and malonyl-ACP forms acetoacetyl-ACP. ACP (acyl carrier protein) is a small protein used to add carbon subunits to a growing fatty acid chain. Each successive addition of an acetyl unit comes from malonyl-ACP.
In addition to saturated, even-carbon-number fatty acids, fatty acids can also be unsaturated, branched, or have an odd number of carbon atoms. Unsaturated fatty acids contain one or more double bonds in the long hydrophobic portion of the molecule. The number and position of these double bonds is often species-specific or group-specific, and the double bonds are formed by desaturation of a saturated fatty acid. Branched-chain fatty acids are biosynthesized using a branched-chain initiating molecule, and odd-carbon-number fatty acids (for example, C13, C15, C17) are biosynthesized using an initiating molecule that contains a propionyl (C3) group instead of acetyl.
The fatty acid composition of cellular lipids varies from species to species and can also vary in a given organism with differences in growth temperature. Cells adjust their fatty acid composition primarily to keep the cytoplasmic membrane from getting either too stiff in the cold or too fluid as temperatures rise. Growth at low temperatures promotes the biosynthesis of shorter-chain and unsaturated fatty acids, whereas growth at higher temperatures promotes the biosynthesis of longer-chain and more saturated fatty acids. The most common fatty acids in lipids of Bacteria are those with chain lengths of C12–C20.
Lipid Biosynthesis
In the assembly of lipids in cells of Bacteria and Eukarya, fatty acids are first added to a molecule of glycerol. For simple triglycerides (fats), all three glycerol carbons are esterified with fatty acids. To form complex lipids, one of the carbon atoms in glycerol is embellished with a molecule of phosphate, ethanolamine, carbohydrate, or some other polar substance (Figure 2.1c). In Archaea, although membrane lipids are constructed from isoprene to form the phytanyl (C15) or biphytanyl (C30) side chains, the glycerol backbone of archaeal membrane lipids contains a polar group (sugar, phosphate, sulfate, or polar organic compound) as for the lipids of Bacteria and Eukarya. Polar groups are important in lipids for forming the canonical membrane architecture: a hydrophobic interior with hydrophilic inner and outer surfaces (Section 2.1 and Figures 2.1 and 2.2).
Check Your Understanding
Explain how fatty acids are constructed two carbon atoms at a time while the immediate donor of these carbons is a three-carbon compound.
What differences exist in lipids from the three domains of life?
What is the result of adding a polar head group to lipids?
Chapter Review
Go to Mastering Microbiology for videos, animations, practice tests, and more.
I Fundamentals of Metabolism
3.1 All cells require water, carbon and other nutrients, free energy, and reducing power. Microorganisms conserve energy either from the oxidation of chemicals or from light. Chemoorganotrophs use organic chemicals as their electron donors, while chemolithotrophs use inorganic chemicals. Phototrophic organisms convert light energy into chemical energy (ATP) and include both oxygenic and anoxygenic species.
**Q To which energy and carbon utilization classes does Escherichia coli belong when it is growing on glucose as both energy and carbon source?**
3.2 Oxidation–reduction reactions require electron donors and electron acceptors. The tendency of a compound to accept or release electrons is expressed by its reduction potential (E0′). Redox reactions in a cell often employ redox coenzymes such as NAD+/NADH as electron shuttles.
Q The following is a series of coupled electron donors and electron acceptors (written as donor:acceptor). Using the data in Table 3.1 and Figure 3.4, order this series from most energy yielding to least energy yielding: H2:Fe3+, NO:Mn4+, H2S:O2, methanol:NO3− (producing NO2−), H2:O2, Fe2+:O2, NO2−:Fe3+ (producing NO3−), and H2S:NO3−.
3.3 Chemical reactions in the cell are accompanied by changes in energy, expressed in kilojoules per mole. Reactions either release or consume free energy. ΔG0′ is a measure of the energy released (if ΔG0′ is negative in arithmetic sign) or consumed (if ΔG0′ is positive in arithmetic sign) in a reaction under standard conditions and reveals which reactions can be used by an organism to conserve energy.
Q Calculate Δ G0′ for the following reaction: glucose+6 O2→6 CO2+6 H2O. Is this reaction exergonic or endergonic? Distinguish between G0′, G, and Gf 0.
3.4 Energy is conserved in compounds that contain energy-rich phosphate or sulfur bonds. The most common of these compounds is ATP, the prime energy carrier in the cell. ATP can be formed by substrate-level phosphorylation, oxidative phosphorylation, or photophosphorylation.
Q Why is acetyl phosphate considered an energy-rich compound while glucose 6-phosphate is not? What are the primary mechanisms of ATP formation in fermentative metabolism and respiratory metabolism, respectively?
3.5 Enzymes are protein catalysts that increase the rate of biochemical reactions by activating the substrates that bind to their active site. Enzymes are highly specific in the reactions they catalyze, and this specificity resides in the three-dimensional structures of the polypeptide(s) that make up the protein(s).
Q What are enzymes composed of, and why are they necessary for the cell?
II Catabolism: Chemoorganotrophs
3.6 The glycolytic pathway is used to break down glucose to pyruvate. The citric acid cycle oxidizes pyruvate to CO2 and releases electrons for the electron transport chain and is also a source of biosynthetic intermediates. The glyoxylate cycle is necessary for the catabolism of two-carbon electron donors, such as acetate.
Q Why are glycolysis and the citric acid cycle, on their own, insufficient to sustain life? What else is needed?
3.7 In the absence of external electron acceptors, organic compounds can be catabolized anaerobically only by fermentation. Fermentations require that an energy-rich organic compound be formed that can yield ATP by substrate-level phosphorylation. Redox balance is achieved by the production of fermentation products.
Q Define the term substrate-level phosphorylation. How does it differ from oxidative phosphorylation? What compound(s) do fermentative bacteria need to synthesize in order to make ATP by substrate-level phosphorylation?
3.8 Electron transport chains are composed of membrane-associated redox proteins that are arranged in order of their increasing E0′ values. The electron transport chain functions in a concerted fashion to carry electrons from the primary electron donor to the terminal electron acceptor, which is O2 in aerobic respiration.
Q What is the value to the electron transport chain of alternating between electron carriers that carry only electrons and those that carry electrons and protons?
3.9 During electron transport, protons are extruded to the outside of the membrane to form the proton motive force. Key electron carriers include flavins, quinones, the cytochrome bc1 complex, and other cytochromes. The cell uses the proton motive force to make ATP through the activity of ATPase.
Q What is the proton motive force and how is it generated? How does the proton motive force result in ATP biosynthesis?
III Catabolism: Electron Transport and Metabolic Diversity
3.10 When conditions are anoxic, several electron acceptors can substitute for O2 in anaerobic respiration. During respiration, energy is conserved by the transport of electrons through the electron transport chain that generates a proton motive force; ATP is synthesized by ATP synthase by dissipating the proton motive force.
**Q E. coli can oxidize glucose to 6 CO2 and 6 H2O by aerobic respiration or by anaerobic respiration with nitrate. How much ATP would it form per glucose oxidized in each circumstance?**
3.11 Chemolithotrophs use inorganic compounds as electron donors to drive electron transport reactions and thereby generate a proton motive force. Phototrophs use light energy to drive electron transport reactions and thereby generate a proton motive force. The proton motive force underlies energy conservation in all forms of respiration and photosynthesis.
Q What is the role of electron transport in cyclic photophosphorylation? If a purple bacterium is performing cyclic photophosphorylation and growing autotrophically, why would it need an electron donor?
IV Biosynthesis
3.12 Autotrophy is supported in most phototrophic and chemolithotrophic bacteria by the Calvin cycle, in which the enzyme RuBisCO plays a key role. The reduction of N2 to NH3 is called nitrogen fixation and is catalyzed by the enzyme nitrogenase.
Q How might the ability to fix CO2 and N2 help a bacterium be more competitive in its environment?
3.13 Polysaccharides are important structural components of cells and are biosynthesized from activated forms of their monomers. Gluconeogenesis is the production of glucose from nonsugar precursors.
Q What is the importance of the enzyme ribonucleotide reductase in the metabolism of sugars? What is the difference between “free” and “activated” glucose?
3.14 Amino acids are formed from carbon skeletons to which ammonia is added from glutamate, glutamine, or a few other amino acids. Nucleotides are biosynthesized using carbon skeletons from several different sources.
Q Name two common enzymes that function to incorporate NH3 into the cell. How do their reaction mechanisms differ?
3.15 Fatty acids are synthesized from the three-carbon precursor malonyl-ACP, and fully formed fatty acids are attached to glycerol to form lipids. Only the lipids of Bacteria and Eukarya contain fatty acids.
Q Describe the process by which a fatty acid such as palmitate (a C16 straight-chain saturated fatty acid) is synthesized in a cell.
Application Questions
For the following reactions identify the electron donor and the electron acceptor, define the type of metabolism and whether the reaction is aerobic or anaerobic, and calculate the free-energy change under standard conditions (see Sections 3.2 and 3.3): 4 H2+2 CO2⇌Acetate−+H++2 H2OAcetate−+SO42−+3 H+⇌H2S+2 CO2+2 H2OH2S+2 O2⇌SO42−+2 H+
Using the data in Figure 3.4, predict the sequence of electron carriers in the electron transport chain of an organism growing aerobically that has the following electron carriers: ubiquinone, cytochrome a, cytochrome b, NADH, cytochrome c, FAD.
Explain the following observation in light of the redox tower: Cells of Escherichia coli fermenting glucose grow faster when NO3 − is supplied to the culture (NO2 − is produced), and then grow even faster (and stop producing NO2 −) when the culture is highly aerated. Calculate the standard free-energy change of the relevant metabolic reactions.
Although physiologically distinct, aerobic chemolithotrophs and chemoorganotrophs share a number of features with respect to the production of ATP. Discuss these common features along with reasons why the growth yield (grams of cells per mole of substrate consumed) of a chemoorganotroph respiring glucose is so much higher than for a chemolithotroph respiring sulfur.
Chapter Glossary
the energy required to bring the substrate of an enzyme to the reactive state Anabolic reactions (anabolism)
the sum total of all biosynthetic reactions in the cell Anaerobic respiration
a form of respiration in which the terminal electron acceptor is not O2 ATP (adenosine triphosphate)
a nucleotide that is the primary form in which chemical energy is conserved and utilized in cells ATPase (ATP synthase)
a multiprotein enzyme complex embedded in the cytoplasmic membrane that catalyzes the synthesis of ATP coupled to dissipation of the proton motive force Autotroph
an organism capable of biosynthesizing all cell material from CO2 as the sole carbon source Calvin cycle
the series of biosynthetic reactions by which most phototrophs and most aerobic chemolithotrophs convert CO2 into organic compounds Catabolic reactions (catabolism)
biochemical reactions leading to energy conservation (usually as ATP) by the cell Chemolithotroph
an organism able to oxidize inorganic compounds (such as H2, Fe2+, S0, or NH4+) as energy sources (electron donors) Chemoorganotroph
an organism that obtains its carbon and reducing power from organic chemicals Chemotroph
an organism that uses chemical reactions as its source of energy Citric acid cycle (CAC)
a cyclical series of reactions resulting in the conversion of acetate to two molecules of CO2 Coenzyme
a small and loosely bound nonprotein molecule that participates in a reaction as part of an enzyme Electron acceptor
a substance that can accept electrons from an electron donor, becoming reduced in the process Electron donor
a substance that can donate electrons to an electron acceptor, becoming oxidized in the process Endergonic
a protein (or in some cases an RNA) catalyst that functions to speed up chemical reactions Exergonic
anaerobic catabolism in which an organic compound is both an electron donor and an electron acceptor and ATP is produced by substrate-level phosphorylation Ferredoxin
an electron carrier with an extremely electronegative reduction potential Free energy (G)
energy available to do work; G0′ is free energy under standard conditions Glycolysis
a biochemical pathway in which glucose is oxidized to pyruvate, which is either used in respiration or fermented (also called the Embden–Meyerhof–Parnas pathway) Glyoxylate cycle
a modification of the citric acid cycle in which isocitrate is cleaved to form succinate and glyoxylate during growth on two-carbon electron donors such as acetate Heterotroph
an organism that uses organic compounds as a carbon source Nicotinamide adenine dinucleotide
(NAD+/NADH) a cytoplasmic coenzyme that shuttles electrons between various enzymes that catalyze redox reactions Nitrogenase
the enzyme complex required to reduce N2 to NH3 in biological nitrogen fixation Nitrogen fixation
the reduction of N2 to NH3 by the enzyme nitrogenase Oxidative phosphorylation
the production of ATP from a proton motive force that is formed by electron transport reactions driven by the oxidation of an electron donor and reduction of an external electron acceptor (see also respiration) Pentose phosphate pathway
a series of reactions in which pentoses are catabolized to generate precursors for nucleotide biosynthesis or to synthesize glucose Photophosphorylation
the production of ATP from a proton motive force formed from light-driven electron transport Phototroph
an organism that uses light as its source of energy Photosynthetic reaction center
the site at which light energy is used to excite an electron to a more electronegative state so that it can be donated to the electron transport chain Proton motive force (pmf)
a source of energy resulting from the separation of protons and hydroxyl ions across the cytoplasmic membrane, generating a membrane electrochemical potential Redox reaction
A chemical reaction in which electrons are transferred between two molecules Reducing power
the ability to donate electrons in metabolic reactions Reduction potential
E0′ the inherent tendency, measured in volts under standard conditions, of a compound to donate or to accept electrons Respiration
the process in which an electron donor is oxidized and electrons are passed through electron transport reactions to an external electron acceptor in order to generate a proton motive force and synthesize ATP by oxidative phosphorylation RuBisCO
ribulose bisphosphate carboxylase, a key enzyme for CO2 fixation by the Calvin cycle Substrate-level phosphorylation
the production of ATP by the direct transfer of an energy-rich phosphate molecule from a phosphorylated organic compound to ADP