When Antibiotics Fail, Bacteriophage Therapy to the Rescue
Acquiring an antibiotic-resistant infection or “superbug” is one of medicine’s biggest nightmares. What can medical practitioners do to treat the patient? Besides drugs, viruses known as bacteriophages have been recruited to specifically target and kill bacteria.
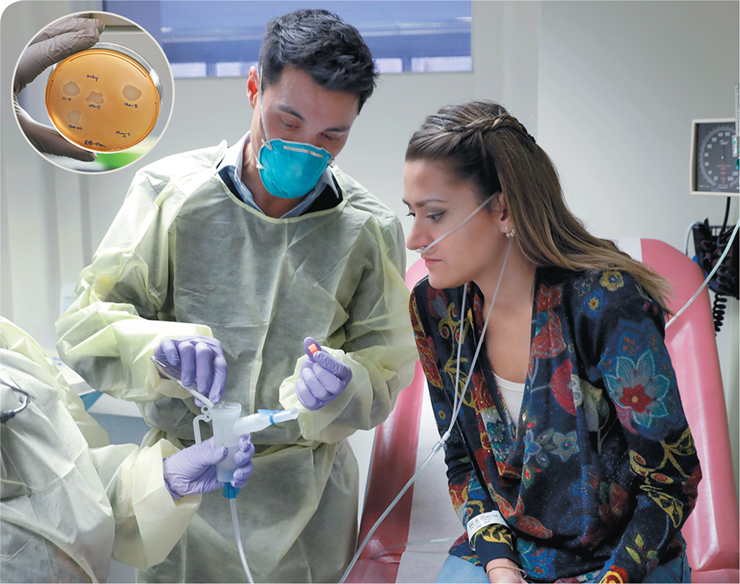
Despite microbiologists’ tinkering with using bacteriophages as antimicrobials for decades, their actual application in medicine has been minimal. However, the emergence of antibiotic resistance has led to renewed focus on using these tiny microbes as therapeutic agents. The photo above shows Ella Balasa (right side of photo), a microbiologist who has cystic fibrosis. Cystic fibrosis is a genetic disease that results in a buildup of thick mucus in the lungs. This mucus allows bacteria to flourish in the lungs, which results in infections and subsequent lung damage that can be fatal. Ella had been treated numerous times with strong antibiotics specific for a respiratory infection caused by the bacterial pathogen Pseudomonas aeruginosa, but the microbial cells had become unresponsive to the drugs. At the time of this photo, the recurrent infection had decreased her lung function to the point where she required constant supplemental oxygen.
As an alternative treatment route, Dr. Benjamin Chan (on the left) took mucus from Ella’s lungs infected with P. aeruginosa and isolated a bacteriophage that specifically killed the pathogen (see zones of clearing on Petri plate). This bacteriophage was propagated and then poured into a device so that Ella could inhale the therapy. The result of her treatment? Amazingly, the bacteriophage therapy along with a mixture of antibiotics resulted in the infection clearing a few weeks later!
While bacteriophage therapy is limited by bacterial host specificity and the ability of the pathogenic bacteria to become resistant to viral infection, this trial and other recent success stories illustrate the future promise of phage therapy when all other options fail.
Source: Kortright, K.E., B.K. Chan, J.L. Koff, and P.E. Turner. 2019. Phage therapy: A renewed approach to combat antibiotic-resistant bacteria. Cell Host Microbe 25(2): 219.
Thus far in Unit 1 we have focused on the fundamentals of the three phylogenetic groups of cells—Bacteria, Archaea, and microbial Eukarya—by highlighting their structure, growth, and overall impact on the biosphere. Here we shift gears and discuss the most abundant microbes on Earth—viruses. With the population of viruses estimated to outnumber bacterial and archaeal cells by a factor of 10, viruses have a huge impact on the biosphere and have a unique biology reminiscent of, yet distinct from, that of the cellular world.
The study of viruses is the science of virology, and this chapter covers the basic principles needed to understand their biology. In Chapter 11 we consider the genomic, replication, and diversity aspects of viruses in detail, and in Unit 7 we consider pathogenic viruses and the extensive role they play in governing human health and disease.
I The Nature of Viruses
Viruses are noncellular microbes that show many of the properties of cells except that viruses absolutely require a growing host cell in order to multiply. Viral diversity in nature is enormous, virus densities can be quantified, and cells in all domains of life are susceptible to viral infection.
5.1 What Is a Virus?
A virus is a genetic element that can multiply only inside a living cell, called the host cell. Not considered living entities, viruses are not found on the tree of life (Figure 1.41b). Viruses rely on the host cell for energy, metabolic intermediates, and protein synthesis, and so they are obligate intracellular parasites. However, viruses possess their own nucleic acid genomes and in this sense are independent of the host’s genome. Viruses infect cells in all three domains of life and are responsible for many infectious diseases of humans, plants, and other organisms.

Although viruses are not cells, their genomes encode those functions needed to multiply and they have a structurally intricate extracellular form, called the virion (Figure 5.1); the virion allows the virus to travel from one host cell to another. Viruses cannot reproduce unless the virion itself, or in some cases its genome only, has gained entry into a suitable growing host cell, a process called infection.
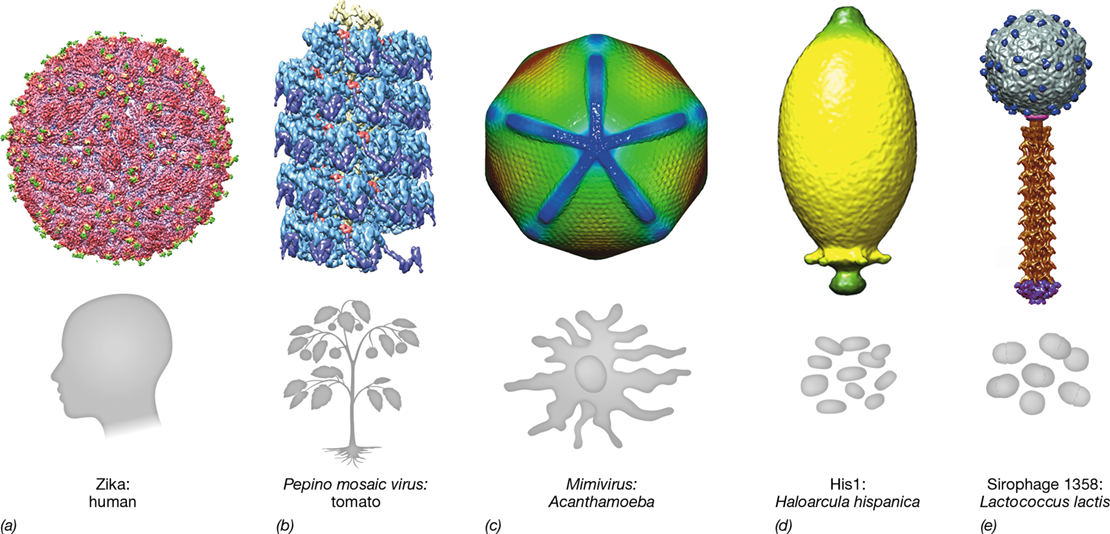
Figure 5.1 Virion morphology and their hosts.

Cryo-electron micrograph (Section 1.10) reconstructions of the following virions: (a) Zika virus (40 nm) that infects humans. (b) Pepino mosaic virus 12.5×510 nm that infects tomatoes. (c) Mimivirus (400 nm) that infects Acanthamoeba, a protozoan. (d) His1 virus (74×44 nm) that infects Haloarcula hispanica (Archaea). (e) Sirophage 1358 (54×103 nm) that infects Lactococcus lactis (Bacteria). Credits for micrographs: (a) Sun, L., Sirohi, D., Chen, Z., Klose, T., Pierson, T.C., Rossmann, M.G., and Kuhn, R.J. (b) Agirrezabala, X., Méndez-López, E., Lasso, G., Sánchez-Pina, M.A., Aranda, M. and Valle, M. 2015. eLife 4: e11795. (c) Xiao, C., Kuznetsov, Y.G., Sun, S., Hafenstein, S.L., Kostyuchenko, V.A., Chipman, P.R., Suzan-Monti, M., Raoult, D., McPherson, A, and Rossmann, M.G. EMDataBank ID: 5039. (d) Hong, C., Pietila, M.K, Fu, C., Schmid, M.F., Bamford, D.H., and Chiu W. EMDataBank ID: 6221. (e) Christian Cambillau.
Viral Components and Activities
Virions are composed of a protein shell, called the capsid, and the virus genome that the capsid contains. Most bacterial and plant viruses are naked, with no further layers, whereas many other types of viruses, especially animal viruses, have an outer layer. If the layer is composed of a phospholipid bilayer taken from the host cell membrane, it is called the envelope (Figure 5.2). In enveloped viruses, the inner structure of nucleic acid plus capsid protein is called the nucleocapsid. The virion protects the viral genome when the virus is outside the host cell, and proteins on the virion surface are important in attaching it to its host cell. The virion may also contain one or more virus-specific enzymes that play a role during infection and replication, as discussed later.
Figure 5.2 Comparison of naked and enveloped virus particles.

The envelope originates from host cytoplasmic membrane.
Once inside the host cell, a viral genome can orchestrate one of two quite different events. The virus may replicate and destroy the host in a virulent infection via a lytic pathway. In a lytic infection, the virus redirects the host cell’s metabolism from growth to support virus multiplication and the assembly of new virions. Eventually, new virions are released, and the process can repeat itself within new host cells. Alternatively, some viruses can cause a lysogenic infection; in this case, the host cell is not destroyed and the viral genome becomes part of the host genome. These types of infection are discussed in detail later in the chapter (Section 5.6).
Viral Diversity and Hosts
While all cells contain double-stranded DNA genomes, viruses often break this rule by having genomes that consist of either DNA or RNA and are further subdivided based on whether the genome is single-stranded or double-stranded. As one might expect, this interesting feature of viral genomes requires special processes for genetic information flow (Chapter 11). Also, while viruses are generally considered to be smaller in both size and gene content than cells, some viruses do not fit into this paradigm. Giant viruses of protozoa are known that possess genomes larger than those of some bacteria. This is in sharp contrast to a common virus such as influenza A virus, which contains only seven genes, and some viruses have even fewer genes than this.

Viruses can be classified on the basis of the hosts they infect as well as by their genome structure (Section 11.1). Thus, we have bacterial viruses, archaeal viruses, animal viruses, plant viruses, protozoan viruses, and so on (Figure 5.1). Bacterial viruses are called bacteriophages (or simply phage for short) and have been intensively studied as model systems for the molecular biology and genetics of virus replication. In this chapter we will use bacteriophages many times to illustrate basic viral principles. Indeed, many of the key tenets of virology were discovered in studies of bacteriophages and subsequently applied to viruses of higher organisms.
Because of their frequent medical and agricultural importance, disease-causing viruses have been extensively studied. However, not all viruses have negative effects on their hosts, and in some cases viruses are even beneficial to their hosts! For example, when Arabidopsis plants are infected with plum pox virus, the plant’s tolerance to drought increases due to virally induced production of the plant growth factor salicylic acid (Figure 5.3). In the rosy apple aphid, infection by an insect densovirus results in both decreased aphid size and offspring numbers. However, despite these negative effects, the infection also results in the formation of wings, structures not produced in uninfected aphids. Wing formation benefits the virus-infected aphid as it allows it to fly to new apple trees to feed on. And finally, in humans, although hepatitis G virus rarely causes symptoms in healthy adults, humans coinfected with both HIV (human immunodeficiency virus) and hepatitis G virus benefit from the hepatitis G infection because it decreases the replication rate and infectivity of HIV.
Figure 5.3 Virus-infected ***Arabidopsis thaliana*** plants and response to drought.

The plants on the left are uninfected, while the plants on the right are infected with plum pox virus. (a) In well-watered plants, the virus affects the plant negatively. (b) Under drought conditions, the virus affects the plant positively. Images adapted from Aguilar, E., Cutrona, C., Del Toro, F.J., Vallarino, J.G., Osorio, S., Perez-Bueno, M.L., Baron, M., Chung, B.M., Canto, T., and Tenllado, F. 2017. Plant, Cell Environ. 40: 2909.
With viruses being the most abundant entity on Earth, numerous other examples of positive host–virus interactions likely exist in nature. But coupled with these, of course, are the countless negative interactions brought about by viral infection, and examples of these will unfold in many places throughout this book.
Check Your Understanding
Compared with cells, what is unusual about viral genomes?
5.2 Structure of the Virion
Virions come in many shapes and sizes (Figure 5.1). Most viruses are smaller than prokaryotic cells, ranging in size from 0.02 to 0.3 μm (20–300 nanometers, nm). While the human smallpox virus is about the size of the smallest known bacterial cells (200 nm in diameter, Section 1.3), giant viruses exist. Viruses such as the Pandoravirus, which has been isolated from protozoa, can rival the size of bacterial cells at over 1 μm in length (Figure 5.4). Poliovirus, one of the smallest viruses at only 28 nm in diameter, is on the opposite end of the size spectrum. This tiny virus is about the size of a ribosome, the cell’s protein-synthesizing machine (Section 1.2).
Figure 5.4 The giant and asymmetrical ***Pandoravirus***.

(a) Scanning electron micrograph of Pandoravirus in a sewage sample from Mergulhão, Brazil. (b) Color-enhanced transmission electron micrograph of Pandoravirus. The virion is ~1.2 μm long and the arrow indicates apical pore.
Virion Structure
The structures of virions are quite diverse, varying widely in size, shape, and chemical composition. The nucleic acid of a virion is always surrounded by its capsid (Figure 5.2). The capsid is composed of a number of individual protein molecules called capsomeres that are often arranged in a precise and highly repetitive pattern around the nucleic acid.
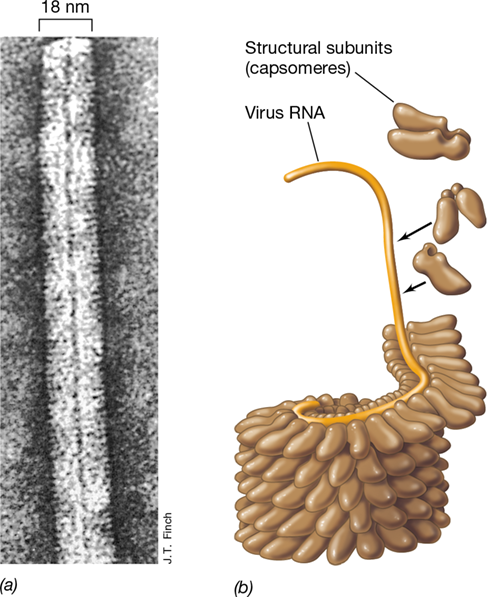
The small size of most viral genomes restricts the number of distinct viral proteins that can be encoded. As a consequence, a few viruses have only a single kind of protein in their capsid. An example is the well-studied tobacco mosaic virus (TMV), which causes disease in tobacco, tomato, and related plants. TMV is a virus in which the 2130 copies of the capsomere protein are arranged in a helix with dimensions of 18×300 nm (Figure 5.5).
Figure 5.5 The arrangement of RNA and protein coat in tobacco mosaic virus, a simple naked virus.

(a) High-resolution electron micrograph of a portion of the tobacco mosaic virus particle. (b) Cutaway showing structure of the virion. The RNA forms a helix surrounded by the protein subunits (capsomeres). The center of the virus particle is hollow.
The information required for the proper folding and assembly of viral proteins into capsomeres and subsequently into capsids is often embedded within the amino acid sequence of the viral proteins themselves. When this is the case, virion assembly is a spontaneous process and is called self-assembly. However, some virus proteins and structures require assistance from the host cell’s folding proteins for proper folding and assembly. For example, the capsid protein of bacteriophage lambda (Section 5.6) requires assistance from an Escherichia coli host cell’s chaperone protein in order to fold into its active conformation.
Virus Symmetry
Many virions are highly symmetric structures. When a symmetric structure is rotated around an axis, the same form is seen again after a certain number of degrees of rotation. Two kinds of symmetry are recognized in viruses, which correspond to the two primary viral shapes, rod and spherical. Rod-shaped viruses have helical symmetry while spherical viruses have icosahedral symmetry. A typical virus with helical symmetry is TMV (Figure 5.5). The lengths of helical viruses are determined by the length of the nucleic acid, and the width of the helical virion is determined by the size and packaging of the capsomeres.
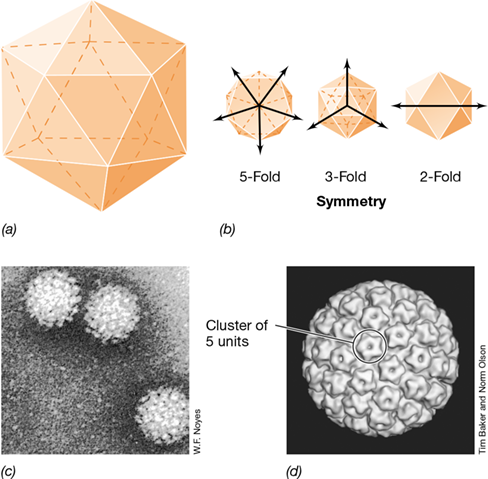
Viruses with icosahedral symmetry contain 20 triangular faces and 12 vertices and are roughly spherical in shape (**Figure 5.6*a***). Axes of symmetry divide the icosahedron into 5, 3, or 2 segments of identical size and shape (Figure 5.6b). Icosahedral symmetry is the most efficient arrangement of subunits in a closed shell because it requires the smallest number of capsomeres to build the shell. The simplest arrangement of capsomeres is 3 per triangular face, for a total of 60 capsomeres per virion. However, most viruses have more nucleic acid than can be packed into a shell made from 60 capsomeres and so viruses with some multiple of 60 capsomeres, such as 180, 240, or 360, are more common. The capsid of the human papillomavirus (Figure 5.6c), for example, consists of 360 capsomeres, with the capsomeres arranged into 72 clusters of 5 each (Figure 5.6d).
Figure 5.6 Icosahedral symmetry.

(a) Model of an icosahedron. (b) Three views of an icosahedron showing 5-, 3-, or 2-fold symmetry. (c) Electron micrograph of human papillomavirus, a virus with 5-fold icosahedral symmetry. The virion is about 55 nm in diameter. (d) Three-dimensional reconstruction of human papillomavirus; a virion contains 360 units arranged in 72 pentacapsomeres, each consisting of 2 separate proteins.
The structure of some viruses is highly complex, with the virion consisting of several parts each displaying its own shape and symmetry. The most structurally complex of all viruses are the head-plus-tail bacteriophages such as phage T4 that infects Escherichia coli. A T4 virion consists of an icosahedral head plus a helical tail (Figure 5.7). Some large viruses that infect eukaryotes, especially the giant viruses of the protozoan Acanthamoeba, are also structurally complex. Quite distinct from the head-plus-tail bacteriophages are Pandoravirus and Mimivirus. Pandoravirus possesses an ovoid morphology and an apical pore (Figure 5.4), while Mimivirus has a five-pronged star feature called stargate (Figure 5.1 and see Figure 5.19b).
Figure 5.7 Structure of T4, a complex bacteriophage.

Transmission electron micrograph of bacteriophage T4 of Escherichia coli. The tail components function in attachment of the virion to the host and injection of the nucleic acid (see Figure 5.15). The T4 head is about 85 nm in diameter.
Enveloped Viruses
Enveloped viruses have a lipoprotein envelope surrounding the nucleocapsid (Figure 5.2). Most enveloped viruses (for example, Ebola, **Figure 5.8a,*b***; see also Figure 5.21) use proteins on the virion’s outer surface to attach to the cytoplasmic membrane and subsequently infect the cell. In contrast to the cells of humans and other animals, plant and bacterial cells are surrounded by a cell wall outside the cytoplasmic membrane, and thus few examples of enveloped viruses are known to infect these organisms. Typically, the entire virion enters an animal cell during infection, with the envelope, if present, assisting in the infection process by fusing with the host membrane. Enveloped viruses also exit more easily from animal cells. As they pass out of the host cell, they are draped in cytoplasmic membrane material. The viral envelope consists primarily of host cytoplasmic membrane, but some viral surface proteins (Figure 5.8a, b) also become embedded in the envelope as the virus passes out of the cell.
Figure 5.8 Enveloped viruses.

(a) Cryo-electron micrograph (Section 1.10) of an Ebola virion. The virions are helical with a diameter of 80 nm. (b) Three-dimensional surface representation of an Ebola tomograph. Color-coding (also applies to arrows in part a) is as follows: red, spikes of envelope surface glycoproteins; orange, lipid envelope; green, membrane-associated proteins; blue/purple, nucleocapsid proteins. (c) Electron micrograph of influenza virus. The virions are about 80 nm in diameter and can have many shapes. (d) Electron micrograph of vaccinia virus, an enveloped icosahedral pox virus about 350 nm wide. The arrows in both c and d point to the envelopes surrounding the nucleocapsids. Parts a and b modified from Beniac, D.R., Melito, P.L., deVarennes, S.L., Hiebert, S.L., Rabb, M.J., Lamboo, L.L., Jones, S.M., Booth, T.F. 2012. PLoS One 7 (1): e29608.
The viral envelope is important in infection, as it is the component of the virion that makes contact with the host cell. The specificity of infection by enveloped viruses and some aspects of their penetration are thus controlled in part by the biochemistry of their envelopes. The virus-specific envelope proteins are critical for both attachment of the virion to the host cell during infection and for release of the virion from the host cell after replication.
Besides an envelope, some viruses possess other structures important for infection. The giant Mimivirus and Tupanvirus that infect Acanthamoeba possess hairlike structures called fibrils on their capsid (Figure 5.9). These fibrils are made of a polymer similar to peptidoglycan (Section 2.3) and are believed to increase infection rates by attracting amoeba hosts that have fed on bacterial cells. Also facilitating infection are the apical pore and stargate features of Pandoravirus and Mimivirus mentioned earlier (Figures 5.4b, 5.1c, and see Figure 5.19b). These unique features act as portals, which open to release the viral genome once the virus is inside the host cell.
Figure 5.9 Giant ***Mimiviridae*** viruses and their surface fibrils.

(a) Transmission electron micrograph of thin sections of the Samba virus. A virion is about 0.57 μm in diameter. Modified from Campos, R.K., et al. 2014. Virol. J. 11: 95. (b) Scanning electron micrograph of Tupanvirus. A virion is about 1.2 μm in length. Both Samba virus and Tupanvirus infect species of amoebae of the genus Acanthamoeba found in Brazil.
Enzymes inside Virions
Viruses do not carry out metabolic processes and are thus metabolically inert. Nonetheless, some viruses carry enzymes in their virions that play important roles in infection. For example, viruses that infect Bacteria and Archaea often only insert their genome into their host. Thus some bacteriophages contain an enzyme that resembles lysozyme (Section 2.3), which is used to make a small hole in the peptidoglycan layer of the bacterial cell to allow nucleic acid from the virion to get into the host cytoplasm. A similar protein is produced in the later stages of infection to lyse the host cell and release new virions. Some animal viruses also contain enzymes that aid in their release from the host. For example, influenza virus (Figure 5.8c) has envelope proteins called neuraminidases that destroy glycoproteins and glycolipids of animal cell connective tissue, thus liberating the virions (Section 11.9).
Viruses with RNA genomes carry enzymes (RNA-dependent RNA polymerases called RNA replicases) in their virion that function to replicate and express the viral RNA genome. Such enzymes are necessary because host cells lack enzymes of any type that can make RNA from an RNA template. Retroviruses, such as HIV, are unusual animal viruses that convert their RNA genome to a DNA intermediate. Because this is another process that host cells cannot do, retroviral virions contain enzymes called reverse transcriptase (Section 11.11).
Although we will see that most viruses do not need to carry special enzymes in their virions, those that do absolutely require them for successful infection and replication. The genes that encode these special enzymes are carried by the virus genome and are expressed in the infection cycle at just the right time to facilitate host infectivity and successful replication of the virus.
Check Your Understanding
Distinguish between a capsid and a capsomere. What is a common symmetry for spherical viruses?
What is the difference between a naked virus and an enveloped virus?
What kinds of enzymes can be found within the virions of RNA viruses? Why are they there?
5.3 Culturing, Detecting, and Counting Viruses
Host cells need to be growing in order for viruses to multiply in them; dead cells cannot produce viruses. While not all viruses kill their host cell, this section focuses on lytic bacteriophages because the culturing and detection of these viruses are easier to describe and visualize.
To culture a bacteriophage such as T4 of Escherichia coli, pure cultures of the bacterial host are either inoculated in liquid medium or spread as “lawns” on the surface of agar plates and then inoculated with a virus suspension. By contrast, animal and plant viruses are cultivated in tissue cultures. For animal viruses, the cells are obtained from an animal organ and grown in sterile glass or plastic vessels containing an appropriate culture medium (see Figure 5.11). Animal tissue culture media are often highly complex, containing a wide assortment of nutrients including blood serum and other highly nutritious substances to feed the animal cells as well as antimicrobial agents to prevent bacterial contamination. For plant tissue cultures, a hairy root-based system is often used to easily propagate viruses, as the infected roots can grow in a liquid medium.
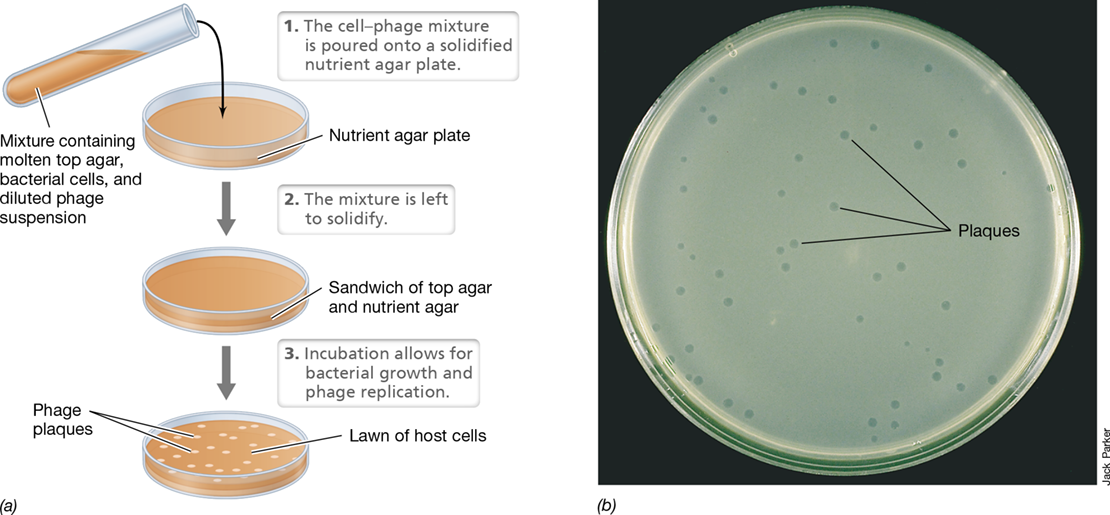
Detecting and Counting Viruses: The Plaque Assay
A viral suspension can be quantified to estimate the number of infectious virions present per volume of fluid, a quantity called the titer. This is typically done using a plaque assay. When a lytic virus infects host cells growing on a flat surface, a zone of cell lysis called a plaque forms and appears as a clear area in the lawn of host cells (Figure 5.10).
Mastering Microbiology
Art Activity: Figure 5.10 Quantification of bacterial virus by plaque assay
Figure 5.10 Quantification of bacterial virus by plaque assay.

(a) “Top agar” (a dilute agar solution) containing a dilution of virions mixed with permissive host bacteria is poured over a plate of “bottom agar” (a more concentrated and thus stiffer agar). Infected cells begin to grow but then are lysed, forming plaques in the lawn. (b) Plaques (about 1–2 mm in diameter) formed by bacteriophage T4.
With bacteriophages, plaques may be obtained when virions are mixed into a small volume of molten agar containing host bacteria that is spread on the surface of an agar medium (Figure 5.10a). During incubation, the bacteria grow and form a turbid layer (lawn) that is visible to the naked eye. However, wherever a successful viral infection has occurred, cells are lysed, forming a plaque (Figure 5.10b). By counting the number of plaques, one can calculate the titer of the virus sample. The titer is typically expressed as the number of “plaque-forming units” per milliliter rather than an absolute viral number because of variations in plating efficiency, to be described next. However, when considering the plaque assay as a method, one should note that quantifying viruses by obtaining their titer is analogous to the counting of bacterial colonies that have formed on the surface of an agar plate from spreading a bacterial cell suspension (“plate count methods, Section 4.4).
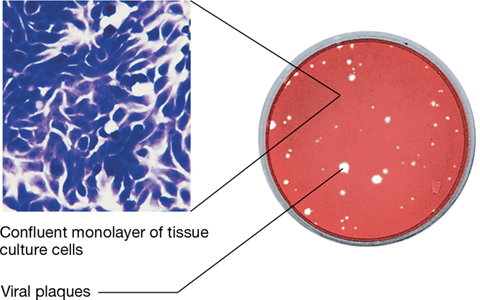
In order to replicate animal viruses that kill their host cell, a tissue culture is grown, and a diluted virus suspension is overlaid on it. As for bacterial viruses, plaques are revealed as cleared zones in the tissue culture cell layer, and from the number of plaques produced, an estimate of the animal virus titer can be made (Figure 5.11). Viral titer is much harder to determine in plants, as plaques do not form in plant tissue culture. Instead viruses must be purified from the plant tissue and enumerated microscopically or through serological or molecular methods that specifically target viral proteins (Chapter 29).
Figure 5.11 Animal cell cultures and viral plaques.

The stained monolayer of animal cells support replication of the virus, and lysed cells result in plaques.
Plating Efficiency in Estimates of Viral Titers
The concept of plating efficiency is important in quantitative virology for all virus types. In any given viral preparation, the number of plaque-forming units is always lower than actual counts of viral particles made microscopically (using an electron microscope). This is because the efficiency with which virions infect host cells is rarely 100% and may often be considerably less. Virions that fail to infect may have assembled incompletely during the maturation process, may contain defective genomes, or may have suffered a spontaneous mutation that prevents them from attaching or otherwise properly replicating. Alternatively, a low plating efficiency may mean that viral growth conditions were not optimal, host viability was low, or that some virions were damaged by handling or storage conditions.
Although plating efficiencies of bacterial viruses can often be higher than 50%, with many animal viruses it may be much lower, 0.1% or 1%. Knowledge of plating efficiency is useful in cultivating viruses because it allows the investigator to estimate what a titer needs to be to yield a certain number of plaques. If the titer is extremely low, the viral suspension may need to be concentrated by centrifugation or filtration before being used to infect host cells. This is especially true of animal viruses, as the costs of growing and maintaining tissue cultures can be significant.
II Overview of the Viral Replication Cycle
Following viral infection of a host cell, new virions are produced by the host as directed by the viral genome. Some aspects of viral infection differ between prokaryotic and eukaryotic cells, but in both cell types, lytic and long-term relationships are possible depending on the virus.
For a virus to replicate, it must induce a living host cell to synthesize all the essential components needed to make new virions. Because of these biosynthetic and energy requirements, dead host cells cannot replicate viruses. During an active viral infection, viral components are assembled into new virions that are released from the cell. Although replication steps are similar in most viruses, a major difference between viral infection of a prokaryotic cell and viral infection of a eukaryotic cell surrounds the initial step in infection. In cells of Bacteria and those Archaea in which the infection process has been studied, only the viral nucleic acid enters the host cell. By contrast, in plant and animal cells the entire virion is taken up. Despite this key difference, the replication of bacterial viruses has been extremely well studied and so we use them here as a model of viral replication events.
5.4 Steps in the Replication Cycle
A cell that supports the complete replication cycle of a virus is said to be permissive for that virus. In a permissive host, the viral replication cycle can be divided into five steps (Figure 5.12):
Attachment (adsorption) of the virion to the host cell
Penetration (entry, injection) of the virion nucleic acid into the host cell
Synthesis of virus nucleic acid and protein by host cell machinery as redirected by the virus
Assembly of capsids and packaging of viral genomes into new virions
Figure 5.12 The replication cycle of a lytic bacterial virus.

The virions and cells are not drawn to scale. The burst size can be a hundred or more virions per cell depending on the virus and the host cell.
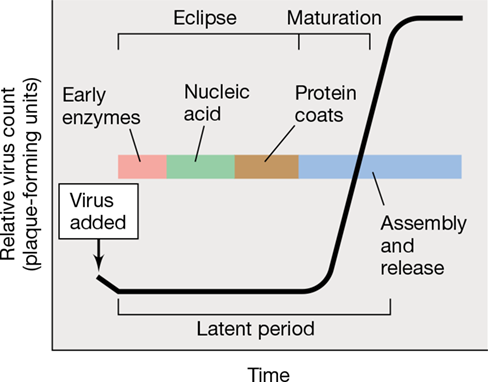
The growth response during virus replication is graphically illustrated in Figure 5.13. The response takes the form of a one-step growth curve, so named because a time course of virion numbers in the culture medium shows essentially no increase during the replication cycle until cells burst and release their newly synthesized virions. In the first few minutes after infection, the virus enters the eclipse phase, during which the viral genome and proteins will be replicated and produced, respectively. Once attached to a permissive host cell, a virion is no longer available to infect another cell. This is followed by the entry of viral nucleic acid into the host cell (Figure 5.12). If the infected cell breaks open at this point, the virion no longer exists as an infectious entity since the viral genome is no longer inside its capsid.
Figure 5.13 One-step growth curve of virus replication.

Following adsorption, infectious virions cannot be detected in the growth medium, a phenomenon called eclipse. During the latent period, which includes the eclipse and early maturation phases, viral nucleic acid replicates and protein synthesis occurs. During the maturation period, virus nucleic acid and protein are assembled into mature virions and then released.
Mastering Microbiology
Art Activity: Figure 5.13 One-step growth curve of virus replication
The maturation phase (Figure 5.13) begins as newly synthesized viral genomes become packaged inside their capsids. During the maturation phase, the number of infectious virions inside the host cell rises dramatically. However, the new virions still cannot be detected in a plaque assay unless the cells are artificially lysed to release them. Because newly assembled virions are not yet present outside the cell, the eclipse and early maturation periods together comprise the latent period of viral infection (Figure 5.13).
At the end of maturation, mature virions are released, either as a result of cell lysis or by budding or excretion, depending on the virus. The number of virions released per cell, called the burst size, varies with the particular virus and the particular host cell, and can range from a few to a few thousand. The duration of the virus replication cycle also varies, from 20–60 min (in many bacterial viruses) to 8–40 h (in most animal viruses).
In Sections 5.5 and 5.6 we use a specific example to revisit these stages of the virus replication cycle and examine each in more detail.
5.5 Bacteriophage T4: A Model Lytic Virus
Much of our understanding of lytic virus replication comes from the study of bacteriophages that infect Escherichia coli. Here we focus on one, bacteriophage T4 (Figure 5.7), as our model for reviewing the individual stages of the lytic virus life cycle in more detail. The study of bacteriophage T4 has contributed to many of the paradigms of modern genetics and molecular biology such as gene structure, the nature of the genetic code, and the discovery of mRNA (Chapter 6).
Attachment and Entry of Bacteriophage T4
The early steps in the life cycle of any bacteriophage are attachment to the surface of its host cell followed by penetration of the host cell outer layer(s) and entry of the viral genome into the cell (Figure 5.12).
A major factor in host specificity of a virus is attachment. The virion itself has one or more proteins on its external surface that interact with specific components called receptors on the host cell surface. In the absence of its specific receptor, the virus cannot attach to the cell and hence cannot infect. Moreover, if the receptor is altered, for example by mutation, the host may become resistant to virus infection. The host range of a given virus is thus to a major extent determined by the presence of a suitable receptor that the virus can recognize and attach to.
Viral receptors are surface components of the host, such as proteins, carbohydrates, glycoproteins, lipids, or lipoproteins, or cell structures made from these macromolecules (Figure 5.14). The receptors carry out normal functions for the cell; for example, the receptor for phage T1 is an iron-uptake protein (Figure 5.14) and that for bacteriophage lambda functions in maltose uptake. Carbohydrates specific to the lipopolysaccharide (LPS) outer membrane of Escherichia coli (Section 2.4) are the receptors recognized by bacteriophage T4 (Figure 5.14). Appendages that project from the cell surface, such as flagella and pili, are also common receptors for bacterial viruses. Small icosahedral viruses often bind to the side of these structures (Figure 2.18), whereas filamentous bacteriophages typically bind at the tip, such as on the pilus (Figure 5.14). Regardless of the receptor used, however, once attachment has occurred, the stage is set for viral infection.
Figure 5.14 Bacteriophage receptors.
Examples of the cell receptor sites used by different bacteriophages that infect Escherichia coli.
Penetration
Attachment of a virus to its host cell causes changes to both the virus and the host cell surface that result in penetration. Bacteriophages abandon their capsid outside the cell and only the viral genome reaches the cytoplasm. However, entry of the viral genome into a host cell only results in virus replication if the viral genome can be read. Consequently, for the replication of some viruses, for example RNA viruses, specific viral proteins must enter the host cell along with the viral genome (Section 5.2).
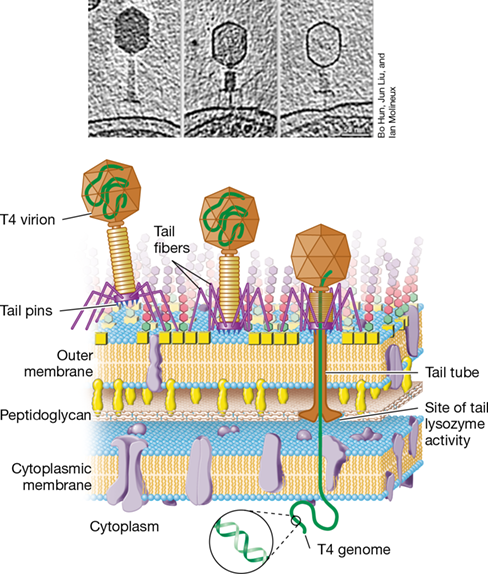
The most intricate viral penetration mechanisms belong to the tailed bacteriophages. Bacteriophage T4 consists of an icosahedral head, within which the viral linear double-stranded DNA is folded, and a long, complex tail, which ends in a series of tail fibers and tail pins that contact the cell surface (Figure 5.7). Phage T4 virions first attach to Escherichia coli cells using their tail fibers (Figure 5.15). The ends of the tail fibers interact specifically with polysaccharides in the cell’s LPS layer and then the tail fibers retract, allowing the tail itself to contact the cell wall via the tail pins. The activity of T4 lysozyme then forms a small pore in the peptidoglycan layer and the tail sheath contracts. When this occurs, the T4 genome enters the cytoplasm of the cell through a tail tube in a fashion resembling that of injection by a syringe. By contrast, the T4 capsid remains outside the cell (Figure 5.15). DNA inside bacteriophage heads is under high pressure, and because the interior of a bacterial cell is also under pressure from osmotic forces, the phage DNA injection process takes several minutes to complete.
Figure 5.15 Attachment and infection of an ***Escherichia coli*** cell by bacteriophage T4.

The three transmission electron tomographs and the art beneath each depict (left to right) the initial attachment of a T4 virion to the cell outer membrane by tail fiber interactions with lipopolysaccharide (LPS); contact of the cell wall by the tail pins; and contraction of the tail sheath and injection of the T4 genome. The tail tube penetrates the outer membrane, and T4 lysozyme digests a small opening through the E. coli cell peptidoglycan layer.
Once a bacteriophage injects its genome into a host cell, a productive infection is not absolutely ensured. Although they lack the immune systems of animals (Chapters 26 and 27), Bacteria and Archaea possess several weapons to protect against viral attack. Toxin–antitoxin modules (Section 8.12) and an antiviral system called CRISPR (Section 9.12) are two of these mechanisms. Additionally, Bacteria and Archaea can destroy double-stranded viral DNA through the activity of restriction endonucleases, enzymes that cleave foreign DNA at specific sites (Section 12.2). Although host restriction systems confer significant protection from viral attack, some DNA viruses have overcome host restriction by modifying their own genome so it is no longer subject to restriction enzyme attack.
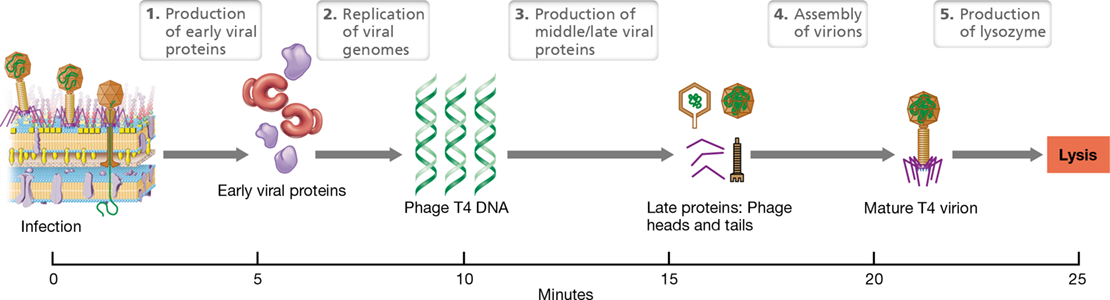
Production of T4 Virions and Release
Once the T4 genome has successfully entered and been retained in a permissive host cell, the process of new virion synthesis begins. In less than half an hour, the process culminates in the release of new virions from the lysed cell. The major events are summarized in Figure 5.16.
Figure 5.16 Time course of events in phage T4 infection.

Following injection of DNA, early and middle mRNAs are produced that encode proteins needed for DNA replication. Following injection of DNA, early and middle proteins needed for DNA replication are produced. Late proteins are virion structural proteins and T4 lysozyme, which is needed to lyse the cell and release new virions.
Within a minute after the T4 genome enters the host cytoplasm, the synthesis of host-specific proteins ceases and production of phage-specific proteins begins, thus inhibiting normal host growth. The T4 genome encodes three major sets of proteins called early proteins, middle proteins, and late proteins, the terms referring to the general order of their appearance in the cell. Early proteins include enzymes for the production of T4 genome copies and proteins that modify host enzymes to express viral genes. This viral takeover of the host cell results in replication of the 170-kilobase T4 genome within 4 minutes of infection (we save the details of T4 genome replication for Chapter 11). By contrast, middle and late proteins include additional proteins that modify host enzymes, and virion structural and release proteins. These include, in particular, viral head and tail proteins and the enzymes required to liberate new virions from the cell (Figure 5.16).
Mastering Microbiology
Art Activity: Figure 5.16 Time course of events in phage T4 infection
Once viral head proteins are made, the bacteriophage T4 DNA genome is forcibly pumped into a preassembled capsid using an energy-linked packaging motor. The motor components are encoded by viral genes, but host cell metabolism is needed to produce the proteins and supply the ATP required for the pumping process. The packaging process can be divided into three stages (Figure 5.17a). First, precursors of the bacteriophage head called proheads are assembled but remain empty of DNA. Proheads contain temporary “scaffolding proteins” as well as head structural proteins. Second, a packaging motor is assembled at the opening to the prohead (**Figure 5.17*b***). The T4 DNA genome is then pumped into the prohead under pressure using ATP as the driving force. The prohead expands when pressurized by the entering DNA, and the scaffolding proteins are simultaneously discarded. Third, the packaging motor itself is discarded and the capsid head is sealed.
Figure 5.17 Packaging of DNA into a T4 phage head.

A T4 virion consists of a head, tail, collar, and various attachment structures (Figure 5.7). Assembly of the T4 virion requires the packaging of viral DNA into the phage head. (a) Proheads are assembled from capsid and portal proteins, both of which remain in the mature virion. As the head fills with DNA (driven by the energy released from ATP hydrolysis), it expands and becomes more angular. Once the head is filled, the packaging motor detaches and the tail components are added. (b) Cryo-electron micrograph reconstruction of the T4 prohead and packaging motor. The blowup shows proteins within the motor and the prohead and their contact with DNA.
Mastering Microbiology
Art Activity: Figure 5.17a Packaging of DNA into a T4 phage head
After the head has been filled, the T4 tail, tail fibers, and the other components of the virion are added, primarily by spontaneous reactions or self-assembly (Figure 5.16). The phage genome encodes a pair of very late enzymes that combine to breach the two major barriers to virion release: the host cytoplasmic membrane and peptidoglycan layer. Once these structures are compromised, the cell breaks open by osmotic lysis and the newly synthesized virions are released. After each replication cycle, which takes only about 25 min (Figure 5.16), over 100 new virions are released from each host cell (the burst size, Section 5.4), and these are now free to infect neighboring host cells.
Check Your Understanding
How does attachment contribute to virus–host specificity?
Why does phage T4 need a lysozyme-like protein in order to infect its host, and what part of T4 enters the host cytoplasm?
What is required to package the T4 genome into its phage head?
5.6 Temperate Bacteriophages and Lysogeny
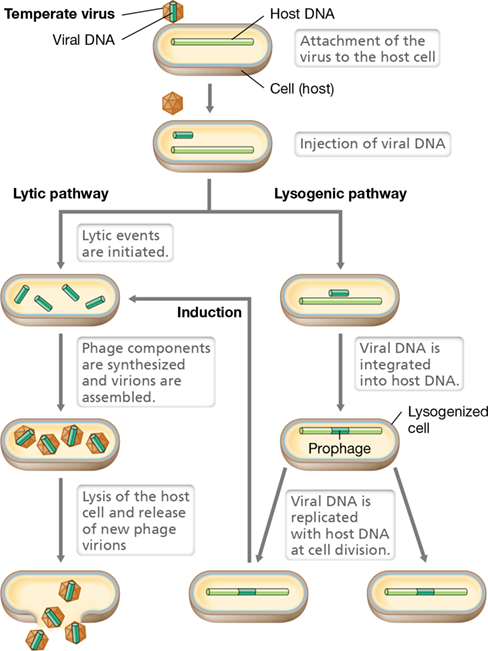
Bacteriophage T4 is a virulent virus and once infection begins, it proceeds to kill its host through lysis. However, some double-stranded DNA bacterial viruses, although capable of a virulent cycle, can also infect their host and establish a long-term stable relationship. Such a virus is called a temperate virus.
Temperate viruses can enter into a state called lysogeny. In this state, very few viral proteins are produced; instead, the virus genome is replicated in synchrony with the host chromosome and passed to daughter cells at cell division. A cell that harbors a temperate virus is therefore called a lysogen. While growth of a lysogen is controlled by its local environment and nutritional profile, the lysogenic state may confer new genetic properties—a condition called lysogenic conversion. We will see several examples in later chapters of pathogenic bacteria whose virulence (ability to cause disease) is at least in part linked to a lysogenic bacteriophage.
Two well-characterized temperate bacteriophages are lambda and P1. The life cycle of a temperate bacteriophage is shown in Figure 5.18. During lysogeny, the temperate virus genome is often integrated into the bacterial chromosome. The viral DNA, now called a prophage, replicates along with the host cell as long as the genes that activate the phage virulent pathway are repressed.
Figure 5.18 Consequences of infection by a temperate bacteriophage.

The alternatives upon infection are replication and release of mature virions (lysis) or lysogeny, often by integration of the virus DNA into the host DNA, as shown here. The lysogen can be induced to produce mature virions and lyse.
Mastering Microbiology
Art Activity: Figure 5.18 Consequences of infection by a temperate bacteriophage
Maintenance of the lysogenic state is due to a phage-encoded repressor protein. Normally, this repressor is maintained at a low level in the host cell. However, if the phage repressor is inactivated or if its synthesis is in some way prevented, the prophage can be induced to switch to the lytic stage. This process is called induction. When induction occurs, the viral DNA is excised and phage early, middle, and late proteins are produced similarly to the process described for T4 phage (Section 5.5 and Figure 5.16). New virions are then produced, and the host cell is lysed (Figure 5.18). Various cell stress conditions, especially damage to host cell DNA, can induce a prophage to enter the lytic pathway. In contrast to this process, the viral “decision” to proceed to lysogeny or to the lytic pathway upon initial viral infection is another matter altogether, and has been particularly well studied in bacteriophage lambda. We explore this rather intricate story in Chapter 11.
We finish our introduction to the viral world by briefly considering viruses that infect eukaryotic hosts, both plant and animal. As we will see in Chapters 29, 30, 31, 32 and 33, many of the most notorious scourges of humankind are of viral origin (see Table 5.1 for some examples).
Check Your Understanding
How do temperate viruses differ from virulent (lytic) viruses?
When considering lysogeny, what occurs during the process of induction?
5.7 An Overview of Viruses of Eukaryotes
The major tenets of virology—presence of a capsid to carry the viral DNA or RNA genome, infection and takeover of host metabolic processes, and assembly and release from the cell—are universal, regardless of the nature of the host. Like bacterial viruses, eukaryotic viruses are classified by their genomes (Table 5.1). However, three key differences between bacterial and eukaryotic viruses are that (1) the entire virion of eukaryotic viruses (rather than just the nucleic acid) enters the host cell, (2) eukaryotic cells contain a nucleus, where many (but not all) viruses replicate, and (3) in some eukaryotic cells, membrane-bound “viral factories,” also known as viroplasms, form to increase the rate of virion assembly and protect the process from host defenses (Figure 5.19). We explore some aspects of animal and plant viruses here.
Table 5.1 Representative viral diseases of humans and plants

bIn bases (ss genomes) or base pairs (ds genomes). These viral genomes have been sequenced and thus their lengths are known precisely. However, the sequence and length often vary slightly among different isolates of the same virus. Hence, the genome sizes listed here have been rounded off in all cases.
Figure 5.19 ***Mimivirus*** viral factory.

(a) Electron micrograph of the cytoplasm of Acanthamoeba castellanii infected with Acanthamoeba polyphaga mimivirus (APMV). Dashed leaders indicate the viral factory or viroplasm where APMV is being replicated and assembled. Modified from Campos, R.K., et al. 2014. Virol. J. 11: 95. (b) Scanning electron micrograph of a viral factory with APMV. Note prominent stargate feature (Figure 5.1c).
Viral Infection of Animal Cells
While a large number of animal viruses exist (Chapter 11), most animal viruses that have been studied in detail are those that can replicate in cell cultures (Section 5.3 and Figure 5.11). To initiate infection, animal viruses also bind to specific host cell receptors. These virus receptors are typically animal cell surface proteins used in cell–cell contact or that function in the immune system. For example, the receptors for poliovirus and for HIV (the virus that causes AIDS) are normally used in intercellular communication between human cells. In multicellular organisms, cells in different tissues or organs often express different proteins on their cell surfaces. Consequently, viruses that infect animals often infect only certain tissues. For example, viruses that cause the common cold infect only cells of the upper respiratory tract.
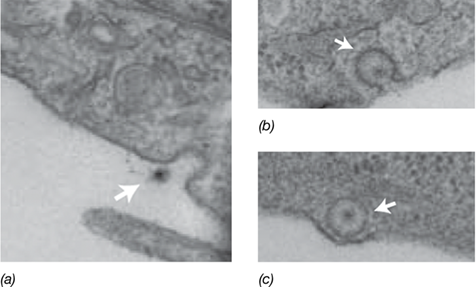
Once an animal virus is bound to a receptor, its entry into a host cell generally occurs by fusion with the cytoplasmic membrane or by endocytosis (Figure 5.20). After entry into the cell, animal viruses must eventually lose their outer coat to deliver their genomes to the cytosol. Some enveloped animal viruses are uncoated at the host cytoplasmic membrane, releasing the nucleocapsid into the cytoplasm, while naked and enveloped animal viruses that enter via endocytosis are uncoated in the host cytoplasm. If the viral genome is DNA, the genome passes through the nuclear membrane to the nucleus for replication. Conversely, the genomes of most RNA-based viruses are replicated or converted to DNA by viral enzymes within the nucleocapsid. We will focus on the unique mode of replication of retroviruses, a special class of highly unusual RNA animal viruses with significant medical implications, in Chapter 11.
Figure 5.20 Rotavirus cell entry.

Electron micrographs of rotavirus virions illustrating the stages of cell entry. The arrows point to the virion. (a) Virion bound to the cell surface. (b) Engulfment of virion by the cytoplasmic membrane. (c) Compartmentalization of the virion. Modified from Abdelhakim, A.H., Salgado, E.N., Fu, X., Pasham, M., Nicastro, D., Kirchhausen, T., and Harrison, S.C. 2014. PLoS Pathog. 10: e1004355.
Mastering Microbiology
Art Activity: Figure 5.22 Possible effects that animal viruses may have on cells they infect
Animal Virion Assembly and Infection Outcomes
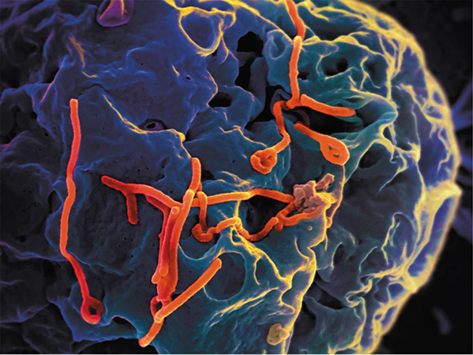
Virion assembly and morphogenesis occurs once viral nucleocapsids have been produced by the host’s machinery. After viral genome copies are packaged within their outer coat, many animal viruses must be enveloped. This typically occurs when the virion exits the animal cell through cell lysis or a process called budding (Figure 5.21). During this process the virus may pick up part of the cell’s cytoplasmic membrane as it exits the cell and use it as part of the viral envelope.
Figure 5.21 Budding of Ebola virus.

Scanning electron micrograph depicting filamentous Ebola virus particles (red) budding from the surface of an infected African monkey kidney epithelial cell (blue-gray).
CDC/PHIL 17778/National Institutes of Allergy and Infectious Disease
Unlike a bacteriophage infection, in which one of only two outcomes—lysis or lysogeny—is possible depending on the virus, other events are possible in an animal virus infection. If an animal virus initially evades the immune system, animal viruses can catalyze at least four different outcomes (Figure 5.22). A virulent infection (or lytic infection) results in lysis of the host cell; this is the most common outcome. By contrast, in a latent infection, the viral DNA exists in the host’s genome as a provirus from a process similar to that described for lysogeny (Section 5.6) and virions are not produced; this leaves the host cell unharmed unless and until an event triggers the virulent pathway. With some enveloped animal viruses, release of virions may be slow, and the host cell may not be lysed (and thus not killed); instead the host cell continues to grow and produce more virions. Such infections are called persistent infections. Finally, certain animal viruses can convert a normal cell into a tumor cell, a process called transformation (Figure 5.22). Some human cancer viruses are capable of such activity (Section 11.7), although the vast majority of cancers are not of viral origin.
Figure 5.22 Possible effects of animal virus infection of host cells.

Most animal viruses are lytic, and only a very few are known to cause cells to transform and become cancerous.
Viral Infection of Plant Cells
Plant viruses share many traits with animal viruses; for example, most have RNA genomes, the complete virion enters the host cell, and viral factories form in infected cells (Figure 5.19). However, three major differences between animal and plant viruses are that (1) many plant viruses have a wide host range (Table 5.1), with viruses such as the cucumber mosaic virus able to infect over 1200 species of plants; (2) most plant viruses are not enveloped; and (3) plant virus transmission to host cells occurs in a different manner due to both the immobility of plants and the rigidity of the cell wall, a structure that is absent from animal cells.
While animal viruses often enter host cells by endocytosis, plant cells contain a protective cell wall preventing such entry. Thus, viruses enter new plant hosts through wounds in the plant tissue or through penetration of the plant cell wall by insects, nematodes, or fungi. In fact, many plant viruses can attach to the mouthparts of plant pests or even propagate in the guts of such pests. Pests that are able to transfer viruses to other types of host cells are called vectors.
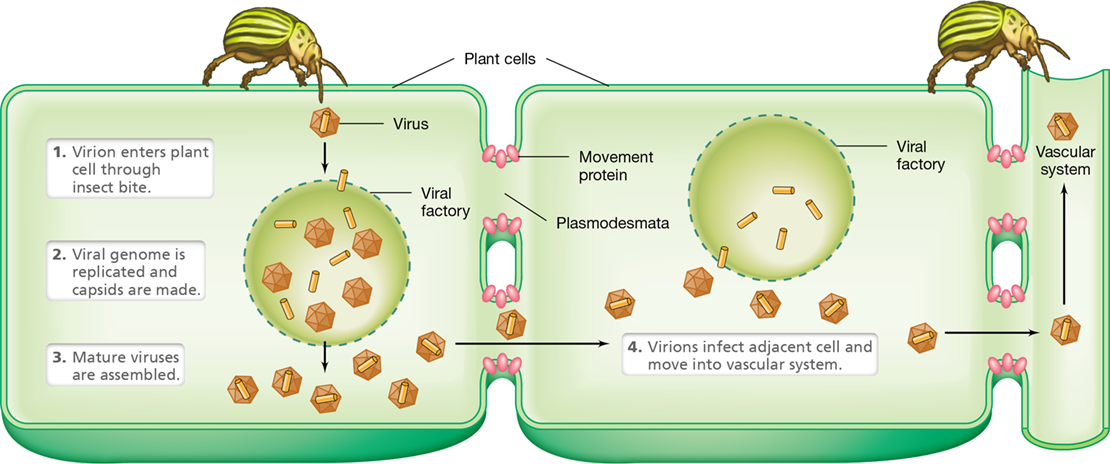
Once the virion has successfully entered a plant cell, the capsid is removed, the genome is replicated in the nucleus or viral factory, and new virions are assembled (Figure 5.23). For infection of other cells within the plant to occur, the viral genome often encodes special movement proteins that help viruses travel through small channels, called plasmodesmata, that connect plant cells. This movement can lead to the viruses entering the vascular system and infecting the entire plant (Figure 5.23). Vectors that feed on this infected plant can then transfer the virus to healthy plants, repeating the infection cycle and causing widespread crop damage.
Figure 5.23 Viral infection of plants.

Unlike infecting an animal, where viruses can readily attach and fuse with the animal cell’s cytoplasmic membrane, to infect a plant, viruses have to traverse the plant cell wall that may be thick. Virions rely on some form of surface damage to the plant, entering a plant cell through a wound or insect bite. Infection leads to viral multiplication and the production of viral-encoded movement proteins, which allow for transfer of viruses to adjacent plant cells through plasmodesmata. The entire plant can be infected once the virus enters the vascular system.
As exemplified by the viruses we have considered in this chapter, viruses are truly fascinating microbes. On the one hand, the multiplication of any virus is completely dependent on the growth and activities of its host cell; on the other hand, viruses can control the growth, genetic properties, and very survival of their hosts. We revisit the viruses in Chapter 11 with a focus on viral genomics as the criterion by which we reveal the enormous genetic diversity of these important microbes. Although our discussion of viruses in this chapter has covered some of their remarkable characteristics, we have only scratched the surface.
Check Your Understanding
Contrast the ways in which animal and bacterial viruses enter their hosts.
What is the difference between a persistent and a latent viral infection?
How does viral infection of plants differ from viral infection of animals?
Chapter Review
Go to Mastering Microbiology for videos, animations, practice tests, and more.
I The Nature of Viruses
5.1 A virus is an obligate intracellular parasite that requires a suitable host cell for replication. A virion is the extracellular form of a virus and contains either an RNA or a DNA genome inside a protein shell. Once the virus is inside the cell, the viral nucleic acid, and sometimes viral enzymes, redirects host metabolism to support virus multiplication. Viruses are classified by the characteristics of their genome and hosts. Bacteriophages infect bacterial cells and are model systems for studying viruses.
Q Once the virion genome is inside a host prokaryotic cell, what are the two types of infection that can be triggered by the virion genome?
5.2 In the virion of a naked virus, only nucleic acid and protein are present; the entire unit is called the nucleocapsid. Enveloped viruses have one or more lipoprotein layers surrounding the nucleocapsid. The nucleocapsid is often arranged in a symmetric fashion, with the icosahedron being a common morphology. Some viruses also possess structures that assist with the infection process. Although virus particles are metabolically inert, some viruses have one or more key enzymes within their virion that carry out processes that the host cannot.
Q Where does the envelope surrounding animal viruses originate?
5.3 Viruses can replicate only in their correct host cells. Bacterial viruses have proved useful as model systems because their host cells are easy to grow and manipulate in culture. Many animal viruses can be grown in cultured animal cells. Viruses can be quantified (titered) by a plaque assay. Plaques are clearings that develop on lawns of host cells, and in analogy to bacterial colonies, they arise from the viral infection of a single cell.
Q Describe the events that occur on an agar plate containing a bacterial lawn when a single bacteriophage particle causes the formation of a bacteriophage plaque.
II Overview of the Viral Replication Cycle
5.4 The virus replication cycle can be divided into five major stages: attachment (adsorption), penetration (uptake of the entire virion or injection of the nucleic acid only), protein and nucleic acid synthesis, assembly and packaging, and virion release.
Q Why does the viral one-step growth curve differ in shape from that of a bacterial growth curve?
5.5 The attachment of a virion to a host cell is a highly specific process. Recognition proteins on the virus recognize specific receptors on the host cell. Sometimes the entire virion enters the host cell, whereas in other cases, as with most bacteriophages, only the viral genome enters. After a T4 virion penetrates a host cell, its genome enters the cytoplasm and viral genes are expressed to redirect the host synthetic machinery to make viral nucleic acid and protein. Early viral genes encode viral genome replication events; middle and late viral genes encode structural proteins and capsid assembly. Once T4 components have been synthesized, new virions are assembled and released after lysis of the host cell.
**Q What is required for a bacteriophage T4 virion to attach to an Escherichia coli cell? What host barriers must be broken before release of T4 virions from the host cell?**
5.6 Some bacteriophages are temperate, meaning that they can initiate lytic events or integrate into the host genome as a prophage. Integration initiates a state called lysogeny in which the virus does not destroy the cell. If the lysogenic state goes through the process of induction, the viral infection switches to virulent (lytic). A well-studied lysogenic virus of Escherichia coli is phage lambda.
Q What kind of protein is required for bacteriophage lambda to maintain a lysogenic state?
5.7 There are eukaryotic viruses with all known modes of viral genome replication. This replication can occur in the nucleus or a compartment called a viroplasm. Many animal viruses are enveloped, picking up portions of host membrane as they leave the cell. Viral infection of animal host cells can result in cell lysis, but latent or persistent infections are also common, and a few animal viruses can cause cancer. In contrast, plant viruses are rarely enveloped and can often infect a wide range of plant, insect, nematode, and fungal species. These pests transfer the virion from one plant to another by puncturing through the rigid plant cell wall.
Q How do virions spread throughout the cells of a plant host? Why do plant viruses require host damage for infectivity?
Application Questions
What causes the viral plaques that appear on a bacterial lawn to stop growing larger?
What would happen if a prophage was not able to undergo the process of induction? How would this affect the virus and its host cell?
Under some conditions, it is possible to obtain nucleic acid–free protein coats (capsids) of certain viruses. Under the electron microscope, these capsids look very similar to complete virions. What does this tell you about the role of the virus nucleic acid in the virus assembly process? Would you expect such particles to be infectious?
Chapter Glossary
the protein shell that surrounds the genome of a virus particle Capsomere
a protein synthesized soon after viral infection and before replication of the viral genome Enveloped
in reference to a virus, having a lipoprotein membrane surrounding the virion Host cell
a cell inside which a virus replicates Late protein
a protein, typically a structural protein, synthesized late in viral infection Lysogen
a state in which the viral genome is replicated in step with the genome of the host Lytic pathway
the type of viral infection that leads to virus replication and destruction of the host cell Middle protein
a protein with either a structural or a catalytic function synthesized after the early proteins in a virus infection Nucleocapsid
the complex of nucleic acid and capsid (shell) proteins of a virus Plaque
a zone of lysis or growth inhibition caused by viral infection of a bacterial lawn or other culture of sensitive host cells Prophage
the lysogenic form of a bacteriophage (see provirus) Provirus
the genome of a temperate, or latent, animal virus when it is replicating in step with the host chromosome Retrovirus
a virus whose RNA genome is replicated via a DNA intermediate Temperate virus
a virus whose genome can replicate along with that of its host without causing cell death, in a state called lysogeny (bacterial viruses) or latency (animal viruses) Titer
the number of individual particles of a substance, such as antibodies or infectious virions, per unit volume of a solution Virion
the infectious virus particle; the viral genome surrounded by a protein coat and sometimes other layers Virulent virus
a virus that lyses, or kills, the host cell after infection Virus
a genetic element that contains either RNA or DNA surrounded by a protein capsid and that replicates only inside host cells
Unit 2 Molecular Biology and Genetics
Chapter 6 Molecular Information Flow and Protein Processing